Total number of characters 39998 1 2 3 A NOVEL MECHANISM OF SEQUESTERING FGF-2 BY GLYPICAN IN 4 LIPID RAFTS, ALLOWING SKELETAL MUSCLE DIFFERENTIATION 5 6 7 8 9 10 11 Jaime Gutiérrez and Enrique Brandan 12 13 14 15 Centro de Regulación Celular y Patología (CRCP), Centro de Regeneración y 16 Envejecimiento (CARE), Departamento de Biología Celular y Molecular, MIFAB, 17 Pontificia Universidad Católica de Chile, Santiago, Chile. 18 19 20 21 22 23 24 25 26 Running title: Glypican in raft domains sequesters FGF-2 27 28 29 Author correspondence: Enrique Brandan, Departamento de Biología Celular y 30 Molecular (Department of Cellular and Molecular Biology), Facultad de Ciencias 31 Biológicas (Faculty of Biological Sciences), P. Universidad Católica de Chile, Casilla 32 (P.O. Box) 114-D, Santiago, Chile. Fax 56 2 635 5395. E-MAIL: [email protected] 33 Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Mol. Cell. Biol. doi:10.1128/MCB.01164-09 MCB Accepts, published online ahead of print on 25 January 2010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Total number of characters 39998 1 2 3
A NOVEL MECHANISM OF SEQUESTERING FGF-2 BY GLYPICAN IN 4
LIPID RAFTS, ALLOWING SKELETAL MUSCLE DIFFERENTIATION 5 6
7
8
9
10
11
Jaime Gutiérrez and Enrique Brandan 12
13
14
15
Centro de Regulación Celular y Patología (CRCP), Centro de Regeneración y 16
Envejecimiento (CARE), Departamento de Biología Celular y Molecular, MIFAB, 17
Pontificia Universidad Católica de Chile, Santiago, Chile. 18
19
20
21
22
23
24
25
26
Running title: Glypican in raft domains sequesters FGF-2 27
28
29
Author correspondence: Enrique Brandan, Departamento de Biología Celular y 30
Molecular (Department of Cellular and Molecular Biology), Facultad de Ciencias 31
Biológicas (Faculty of Biological Sciences), P. Universidad Católica de Chile, Casilla 32
(P.O. Box) 114-D, Santiago, Chile. Fax 56 2 635 5395. E-MAIL: [email protected] 33
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Mol. Cell. Biol. doi:10.1128/MCB.01164-09 MCB Accepts, published online ahead of print on 25 January 2010

2
ABSTRACT 34
Heparan sulfate proteoglycans (HSPGs) are critical modulators of growth factor 35
activities. Skeletal muscle differentiation is strongly inhibited by fibroblast growth 36
factor type-2 (FGF-2). We have shown that HSPGs present at the plasma membrane are 37
expressed in myoblasts and are downregulated during muscle differentiation. An 38
exception is glypican-1, which is present throughout the myogenic process. Myoblasts 39
that do not express glypican-1 exhibit a defective differentiation, with an increase in the 40
receptor binding of FGF-2, concomitant to an increased signaling. Glypican-1 deficient 41
myoblasts show a decreased expression of myogenin, the master gene that controls 42
myogenesis, myosin and the myoblast fusion index. These defects were reverted by 43
expression of rat glypican-1. Glypican-1 is the only HSPG localized in lipid raft 44
domains in myoblasts, resulting in the sequestration of FGF-2 away from FGF-2 45
receptors (FGFRs) located in non-raft domains. A chimeric glypican-1, containing 46
syndecan-1 transmembrane and cytoplasmic domains, located in non-raft domains 47
interacting with FGFR-IV and enhanced FGF-2 dependent signaling. Thus glypican-1 48
acts as a positive regulator of muscle differentiation by sequestering FGF-2 in lipid rafts 49
and preventing its binding and dependent signaling. 50
51
52
53
54
55
56
57
58
Key words: Skeletal muscle differentiation / heparan sulfate proteoglycans / glypican-1 59
/ FGF-2 / raft membrane domains / FGF-2 mediated signaling. 60

3
INTRODUCTION 61
Heparan sulfate proteoglycans (HSPGs), key components of cell surfaces and 62
extracellular matrix (ECM), can influence cell growth and differentiation processes by 63
interacting with a large number of macromolecules. One of the most recognized 64
functions of HSPGs is the ability to modulate different growth factor activities. In this 65
context, cell-surface HSPGs bind soluble ligands, increasing their local concentration 66
and modulating ligand-receptor encounters (5). Fibroblast growth factor type-2 (FGF-2) 67
and hepatocyte growth factor (HGF) signaling are markedly enhanced by HSPGs. In 68
particular, FGF-2 completely depends on heparan sulfate to transduce an intracellular 69
signal through its receptors (FGFRs) (49, 66, 87), through the formation of the ternary 70
complex HSPG-FGF-2-FGFR (62). However, if HSPGs are localized at the ECM they 71
can decrease FGF-2 signaling by sequestering it away from the transducing receptors 72
(9). 73
Skeletal muscle formation and regeneration is a complex and regulated process 74
that involves activation, proliferation and differentiation of a muscle precursor, 75
implicating the participation of heparan binding growth factors, such as FGF-2 (13), 76
HGF (2) and transforming growth factor type-β (TGF-β) (50). Skeletal muscle 77
differentiation is regulated by the expression of specific combinations of muscle 78
regulatory transcription factors. Among them, a family of basic helix-loop-helix 79
transcription factors, called muscle regulatory factors (MRFs), is critical for muscle 80
differentiation (20, 73). The activity of MRFs, particularly myogenin, the master gene 81
involved in skeletal muscle differentiation, is highly depressed in the presence of FGF-82
2, HGF or TGF-ß (2, 13, 50). 83
The role of HSPGs in skeletal muscle physiology, as well as in the skeletal 84
muscle differentiation process, has been previously revised (41). In mature skeletal 85

4
muscle tissue, HSPGs also act as co-receptors for the asymmetric form of acetyl-86
cholinesterase, increasing its concentration at the neuromuscular junction (10, 63). 87
Inhibition of proteoglycan sulfation in C2C12 cultures (52, 58), a satellite cell line 88
derived from regenerating adult mouse skeletal muscle undergoing in vitro terminal 89
myogenic differentiation, or from intact myofibers (19), affects the proper progression 90
of the in vitro myogenic program. Syndecans and glypicans are the two families of 91
HSPGs that localize to the plasma membrane. Syndecans are bound to the plasma 92
membrane through a highly conserved transmembrane domain, and are composed of 93
four separate genes in mammals (5, 24, 65), whereas glypicans are bound to the plasma 94
membrane by a glycosyl-phosphatidylinositol (GPI) linkage corresponding to six 95
separate genes (30, 32). 96
We have shown that during C2C12 myogenesis, the expression of all syndecan 97
forms are downregulated (33, 37, 44, 55), whereas the expression of glypican-1, which 98
is the only glypican expressed in myoblasts, remains constant throughout the process (8, 99
37). This differential expression may reflect different functions or macromolecular 100
specificity during myogenesis. Syndecans have been reported to modulate FGF-2 101
activity during in vitro myogenesis (33, 43, 66) and to participate in cell-cell and cell-102
matrix adhesion in development and adult wound repair (65). It has been reported that 103
syndecan-3 and syndecan-4 are expressed during embryonic limb skeletal muscle 104
formation by developing myocytes (19, 55), and that the expression continues in adult 105
muscle tissue restricted to satellite cells (19). Knock out mice for syndecan-3 exhibit a 106
novel form of muscular dystrophy while syndecan-4(-/-) satellite cells fail to 107
reconstitute damaged muscle, which reveals the importance of these macromolecules in 108
the skeletal muscle differentiation process (23). 109
110

5
Little is known about the localization of HSPGs to specific plasma membrane 111
sub-domains. In this context, it can be speculated that glypicans might be associated 112
with lipid raft domains, which corresponds to sphingolipid and cholesterol-rich domains 113
forming phase-separated “lipid rafts” in the membrane (7, 12). Lipid raft domains might 114
be involved in signal transduction processes through specific receptors and proteins 115
anchored by GPI (11, 45). Since all the syndecans are downregulated during 116
myogenesis (33, 37, 44, 55), it is believed that their absence facilitates the silencing of 117
FGF-2 mediated signaling, helping the process of skeletal muscle differentiation and 118
allowing the expression of myogenin. The presence of glypican-1 on myoblast surfaces 119
during the entire process of skeletal muscle differentiation is puzzling, since it is also 120
able to form the ternary complex HSPG-FGF-2-FGFR when glypican-1 and FGFR-I are 121
overexpressed in the same cell (79). This paradoxical situation leads us to study 122
glypican-1 role in the modulation of FGF-2 signaling during the process of skeletal 123
muscle differentiation. We hypothesized that glypican-1, located in lipid raft domains, 124
sequesters FGF-2 avoiding the interaction with its transducing receptors. This, in turn, 125
allows the expression of myogenin, subsequent myoblast fusion and expression of late 126
muscle differentiation markers. In this study, we evaluated glypican-1 localization at the 127
plasma membrane related to FGFRs, and its direct participation in the modulation of 128
FGF-2 activity. Our results strongly suggest that glypican-1 localizes to lipid raft 129
domains where it interacts with FGF-2, sequestering it away from the FGFRs, which 130
prevents signaling and results in a strong positive effect on skeletal muscle 131
differentiation. 132

6
MATERIALS AND METHODS 133
1- Cell culture 134
The mouse skeletal muscle cell line C2C12 (ATCC) (85) was grown and differentiation 135
induced (43). Myoblasts were treated with FGF-2, HGF, TGF-β1 or platelet derived 136
growth factor (PDGF) (R&D, Minneapolis, MN, USA); heparin and methyl-beta-137
cyclodextrin (MβCD) Sigma Chemical, St. Louis, MO, USA); and phosphatidylinositol-138
specific phospholipase C (PI-PLC) Invitrogen, Carlsbad, CA, USA) (14), as indicated in 139
the corresponding figures. MβCD (1mM) or PI-PLC (0.5 U/ml) treatments were done in 140
PBS, containing 0.1 mM CaCl2 and 1 mM MgCl (PBS Ca+2
/Mg+2
), for 1 h at 37 °C in a 141
humidifying chamber. The heparin wash involved the incubation of cells in PBS 142
Ca+2
/Mg+2
, containing 10 µg/ml of heparin, and gentle agitation for 10 min at 4 °C. This 143
was repeated twice. Heparitinase (Hase) myoblast treatment (Seikagaku, Tokyo, Japan) 144
was undertaken as described (37). For the phosphorylation experiments, the cells were 145
serum starved for 6 h and then treated for the indicated times. 146
For the inactivation of FGF-2, 1 or 10 µg/ml of the soluble form of the FGFR-I 147
(rhFGF R1a(IIIc)/Fc Chimera, R&D, Minneapolis, MN, USA) or 1, 5 or 10 ug/ml of a 148
blocking antibody for FGF-2 (anti-FGF-2 neutralizing antibody Millipore #05-117, 149
Bedford. USA) were exogenously added, and their inhibitory effects were analyzed by 150
the FGF-2 dependent activation of ERK 1/2. In both cases the indicated concentrations 151
of FGF-2 were pre-incubated for 30 min at 37° C. In the case of the soluble receptor 0.1 152
µg/ml of heparin was added. The differentiation medium (2% of horse serum) was pre-153
incubated with 10 µg/ml soluble FGFR-I or 10 µg/ml of the neutralizing antibody for 30 154
min at 37° C, prior to being added to the cells. The FGF-2 neutralizing antibody was 155
replaced daily. 156
157

7
158
2- Transient transfection and generation of stable clones: 159
The myogenin reporter plasmid pMyo-Luc (68), shRNA for glypican-1 and its 160
corresponding control (shCtrl), the pEGFP-N1, pRL-Sv40, the pcDNA3.0 empty vector 161
(all from Invitrogen, Carlsbad, CA, USA ) and the pcDNA3.0- rat-glypican-1 (courtesy 162
of Dr. Ralph D. Sanderson, Department of Pathology, University of Alabama at 163
Birmingham, Birmingham, Alabama USA) were transfected using Lipofectamine Plus 164
transfection reagent (Invitrogen, Carlsbad, CA, USA), according to the supplier’s 165
protocol. For the reporter experiments, the cells were plated in 24-well plates. 48 h after 166
transfection, the cells were induced to differentiate for 30 h in the presence of FGF-2. 167
The samples were subsequently assayed for dual luciferase activity (Promega, Madison, 168
WI, USA). 169
shRNA expression vectors were constructed and packaged into recombinant 170
lentiviruses, using the BLOCK-iT Lentiviral RNAi Expression System (Invitrogen, 171
Carlsbad, CA, USA), according to manufacturer’s instructions. The target sequence for 172
the shRNA against mouse glypican-1 (shGly) (NM_016696) was: 5’-173
GACCATCCGCCAGCAGATTATGC-3’. The pU6-shGly was used in transient 174
transfection experiments. From this plasmid we generated the pLenti6-U6-shGly vector, 175
which was co-transfected with the packaging plasmid mixture into 293FT cells to 176
produce recombinant lentiviruses. The conditioned medium of the lentivirus producing 177
cells was harvested and titrated using C2C12 myoblast cells. Transductions were 178
performed at the multiplicity of infection of 50. The cells were maintained in normal 179
growth medium for 48 h after transduction, and supplemented with 30 ug/ml blasticidin 180
(selection antibiotic). The target gene knockdown in transient transfections and in 181

8
selected clones was confirmed by Western blot analysis. A stable cell line expressing a 182
scrambled sequence was made as the control. 183
The chimeric HSPG containing the extracellular domain of rat glypican-1 and 184
the cytoplasmatic domain of mouse syndecan-1 (GlySyn) was donated by Dr. Ralph D. 185
Sanderson (Department of Pathology, University of Alabama at Birmingham, 186
Birmingham, Alabama, USA) (46, 86), and to which a FLAG epitope in the amino 187
terminal of extracellular domain was incorporated. 188
189
3- Isolation of lipid rafts 190
Lipid rafts were prepared as described previously (91), with some modifications. All of 191
the buffers and instruments used in the procedure described below were at 4°C. Briefly, 192
C2C12 myoblasts from a 150 mm dish, either control or treated, were collected in cold 193
PBS and resuspended in 400 ul of lysis buffer, containing 25 mM MES (2-[N-194
morpholino] ethanesulfonic acid), pH 6.5, 150 mM NaCl, with a mixture of protease 195
inhibitors and 1 mM PMSF supplemented with 1% Triton X-100, 1% Lubrol, or 1% 196
Brij 35, as indicated in each case. Cells were homogenized by passing them through a 197
21 gauge needle three times, incubated for 20 min on ice, and then homogenized with 198
ten strokes of a loose-fitting Dounce homogenizer. Homogenates were mixed with 400 199
ul of 90% sucrose (final concentration, 45%), loaded at the bottom of a Sorvall 4 ml 200
centrifuge tube, and overlaid with 1.6 ml of 35% sucrose and 1.6 ml of 5% sucrose, 201
both in the lysis buffer (80). The samples were centrifuged at 45,000 rpm for 18 h at 202
4°C in an AH-650 rotor. Twelve fractions (330 ul each) were collected from top to 203
bottom and designated as fractions 1–12. 204
205
4- SDS-PAGE, Western blot, slot blot and co-immunoprecipitation: 206

9
Aliquots from each gradient fraction were separated on 8% SDS-PAGE (Mini Protean 207
II; Bio-Rad, Richmond CA, USA), and electrotransferred onto Immobilon membranes 208
(Millipore, Bedford. USA). Western blots were probed using various primary 209
antibodies: rabbit anti-mouse FGFR-I (1:500) (Cell Signaling, Danvers, MA, USA); 210
Biotinylated anti-mouse FGFR-IV (1:500) (R&D, Minneapolis, MN, USA); rabbit anti-211
caveolin-1 (Cav-1) (1:500) (Santa Cruz biotechnology, Santa Cruz, CA, USA); anti-212
glypican-1 M95 (1:500) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and mouse 213
anti- Na+/K
+ATPase (1:1,000) (Upstate biotechnology, Lake Placid, NY, USA). To 214
reveal the distribution of the ganglioside GM1 in the gradients, 30 µl of each fraction 215
was spotted onto nitrocellulose membranes using a Bio-Rad Slot Blot apparatus, and 216
probed with horseradish peroxidase-coupled CTX (1:10,000) (Sigma-Aldrich, MO, 217
USA) (39). 218
To identify HSPG core proteins, samples containing equivalent amounts of 219
protein were incubated with Hase and Chondroitinase ABC (Seikagaku, Tokyo, Japan) 220
(79), and analyzed by western blot the anti-∆-heparan sulfate monoclonal antibody 221
(anti-stub) (Seikagaku, Tokyo, Japan), as described previously (17, 37, 55), and 222
visualized by ECL. 223
For immunoblot analysis, myoblasts were lysed in 50 mM Tris-HCl, pH 7.4, 0.1 224
M NaCl, 0.5% Triton X-100 with a mixture of protease inhibitors and 1 mM 225
phenylmethylsulfonyl fluoride. For analysis of phosphorylated proteins, cell extracts 226
were prepared in RIPA buffer (58). Aliquots with equivalent amounts of proteins were 227
subjected to SDS-PAGE gel electrophoresis in 8% polyacrylamide gels, 228
electrophoretically transferred onto immobilon membranes (Millipore, Bedford. USA), 229
and probed with rabbit anti-phospho ERK 1/2 (1:1000); mouse anti-FLAG 1:5000 230
(Stratagene, La Jolla, CA, USA) ; rabbit anti ERK 1/2 (1:1000); rabbit anti-phospho 231

10
AKT (1:1000) (Calbiochem, La Jolla, CA, USA); mouse anti-α-tubulin (1:5000); mouse 232
anti-myosin (1:5,000) (Sigma-Aldrich, MO, USA); mouse anti-glyceraldehyde-3-233
phosphate-dehydrogenase (GADPH, 1:2000) (Chemicon, Temecula, CA, USA); rabbit 234
anti-myogenin (1:500) and mouse anti-caveolin-3 (1:1,000) (Santa Cruz Biotechnology, 235
Santa Cruz, CA, USA). 236
All immunoreactions were visualized by enhanced chemiluminescence (Pierce, 237
IL, USA), using a ChemiDoc-It HR 410 imaging system (Upland, CA, USA). 238
For the co-immunoprecipitation experiment WT and glypican-1 deficient 239
myoblast (C6) were transiently transfected as indicated in the Figure. 48 h after, the 240
cells were incubated whit DMEM 0.1% BSA supplemented with FGF-2 (20 ng/ml) for 241
3 hours on ice with gentle agitation. The cells were lysed in TS buffer (Tris 20mM, 242
pH7.4, NaCl 150mM, MgCl2 1mM, CaCl2 1mM), 0.1% Triton X-100 and pre-cleared by 243
centrifugation. The extracts were immunoprecipitated for 3 hours at 4°C using 10 ug of 244
a mouse anti-FLAG antibody or 10 ug of a rabbit-anti mouse syndecan-4 (nS4ED, 245
kindly donated by Dr. Alan C. Rapraeger, Department of Pathology and Laboratory 246
Medicine, University of Wisconsin, Madison, USA), electrophoresed and analyzed by 247
western blot with rabbit-anti FGFR-IV, rabbit-anti glypican-1, as described above or 248
with the rabbit-anti mouse syndecan-4 (1:1000 from a 1mg/ml stock solution). 249
250
5- Immunofluorescence microscopy 251
Cells to be immunostained were grown on coverslips. The medium was removed and 252
the coverslips rinsed with PBS. Cells were fixed with 3% paraformaldehyde for 15 min 253
at 4°C, incubated for 30 min in PBS containing 3% BSA and incubated with the 254
primary antibody for 1 h in the same buffer. Rabbit anti-glypican-1 (1:300) (courtesy of 255
Dr. David Carey, Sigfried and Janet Weis Center for Research, Danville, PA, USA.); 256

11
mouse anti-FLAG 1:1000; mouse anti-laminin 1:50 (Telios Pharmaceuticals, San 257
Diego, CA) and mouse anti-fibronectin (1:300) were used, as indicated in the 258
corresponding figures. 259
For the detection of myosin the cells were permeabilized with PBS containing 260
0.05% Triton X-100 (2 min at 4 °C), incubated for 30 min in PBS 3% BSA, and 261
subsequently with the primary antibody mouse anti-myosin (1:300) (Sigma-Aldrich, 262
MO, USA). Cells were rinsed with PBS 3% BSA and further incubated for 1 h with 263
Alexa 488 conjugated antibodies (1/1,000) or Alexa 568 conjugated antibodies 264
(1/1,000) (Invitrogen, Carlsbad, CA, USA). 265
Nuclear staining was done using 0.1 µg/ml of Hoechst 33258 in PBS for 10 min 266
(58). After rinsing, the coverslips were mounted and viewed under a Nikon Diaphot 267
inverted microscope equipped for epifluorescence. 268
269
6- FGF-2 affinity labeling and crosslinking assay. 270
Carrier-free FGF-2 was radiolabeled with Na[125
I] using chloramine-T. Binding and 271
crosslinking of 125
I-FGF-2 to cell surfaces were assayed (27). In some experiments, the 272
cells were pre-treated with Hase (37) or competed with 200 molar excess of cold 273
FGF-2. 274
275
7- Protein determination. 276
Protein was determined with the bicinchoninic acid protein assay kit (Pierce) with BSA 277
as standard. 278

12
RESULTS 279
Glypican-1 is required for successful skeletal muscle differentiation. 280
To evaluate the role of glypican-1 during myogenesis, we studied the effect of 281
its absence in this process. By lentiviral infection of a the short-hairpin RNA (shRNA) 282
specific for mouse glypican-1 in C2C12 myoblasts, we produced a stable clone (C6) 283
that expressed low levels of glypican-1, as determined by Western blot analysis of the 284
corresponding HSPG core protein after Hase digestion (Fig. 1A). The core proteins of 285
any proteoglycan that is substituted with heparan sulfate can be traced using the mAb 286
3G10 (anti-stub) (79) that recognizes a neo-epitope generated after the Hase treatment. 287
shRNA-mediated knockdown of glypican-1 has no effect in the protein levels of other 288
HSPGs, since the infection with a lentiviral vector to express a scramble shRNA 289
(shCtrl) in a stable manner results in HSPG core protein levels that are equivalent to 290
those present in WT extracts (Fig. 1A). Figure 1B shows the kinetics expression of 291
myogenin, the master gene that controls skeletal muscle differentiation, and the late 292
skeletal muscle differentiation markers, myosin and caveolin-3 (cav-3) (34) in WT, 293
shCtrl and C6 myoblasts under differentiation conditions. The latter showed a 294
diminished expression of myogenin concomitant with a significant reduction in myosin 295
and cav-3 by day 5 of differentiation, compared to WT and shCtrl . 296
Immunofluorescence analysis (Fig. 1C) confirmed that myoblasts (left panels) not 297
expressing glypican-1 are unable to form myosin-expressing myotubes after 5 days of 298
differentiation (Myotubes D5, right panels). 299
Figure 2A shows that muscle differentiation is modulated specifically by 300
glypican-1 since the deleterious effect over the expression of muscle specific proteins in 301
the C6 myoblasts was restored after re-expressing this HSPG by transient transfection 302
with rat glypican-1, which is not recognized by the shGly, expressing myosin and cav-3 303

13
similar to the WT levels. This Figure also shows that the transiently transfected WT 304
myoblasts with the shGly express very low levels of myosin and cav-3, as in the clone 305
C6. In this sense, the lack of myotube formation observed in the C6 clone (as shown in 306
Fig. 1C) is glypican-1 specific too, since it can be partially restored by re-expressing rat 307
glypican-1, as indicated in Figure 2B. In concordance, myoblasts transiently transfected 308
with the shGly considerably diminished the amount of myotube formation (Fig. 2B). 309
Figure 2C shows the level of glypican-1 core protein in myoblasts under each 310
experimental condition, determined by the anti-stub antibody after Hase treatment. 311
Figure 2D shows the glypican-1 levels of WT, C6 and C6 re-expressing glypican-1 312
myoblasts determined by an antibody specific against glypican-1. 313
The sums of these results indicate that glypican-1 is critical for skeletal muscle 314
differentiation, evaluated by the induction of myogenin, myosin, caveolin-3 and 315
myotube formation. 316
317
The binding of FGF-2 to its receptors and its consequent signaling is augmented in 318
glypican-1 deficient myoblasts. 319
HSPGs are essential for FGFR activation by FGF-2, acting as co-receptors of 320
this muscle differentiation inhibitory growth factor. It is possible that glypican-1, 321
contrary to syndecans, might be sequestering FGF-2 away from its transducing 322
receptors. To determine this, we evaluated the binding of radiolabeled [125
I]-FGF-2 to 323
FGFRs through affinity labeling experiments, in the presence or absence of glypican-1. 324
Figure 3A shows that the crosslinking of [125
I]-FGF-2 to FGFR-I and –IV increased in 325
C6 myoblasts, as well as in wild-type myoblasts transiently transfected with the shGly. 326
Importantly, the levels of FGFRs were unaffected by glypican-1 silencing (Fig. 3B). 327
The observed binding of FGF-2 to its receptors is specific since it is totally competed by 328

14
an excess of cold FGF-2 and dependent of HSPGs since it is abolished when the cells 329
were pre-treated with Hase (87) (Fig. 3C). 330
Subsequently, we evaluated if such increased binding of FGF-2 would result in 331
an augmented FGF-2-dependent signaling. The extent of phosphorylation of the 332
extracellular regulated kinase 1/2 (phospho ERK 1/2) in response to this growth factor 333
was determined (83). Figure 4A left, shows that C6 myoblasts required lower FGF-2 334
concentrations to induce phospho ERK 1/2, compared to WT myoblasts. Figure 4A 335
right shows quantification of two independent experiments. This increased response to 336
FGF-2 in the absence of glypican-1 is specific, since re-expression of rat glypican-1 in 337
C6 myoblasts revert such heightened sensitivity to FGF-2, as shown in Figure 4B left. A 338
quantification of this experiment is shown in Figure 4B (right panel). Similar results 339
were obtained after inducing the phosphorylation of AKT by FGF-2 (26) (data not 340
shown). Then we asked if the absence of glypican-1 could alter the cellular response to 341
other heparin-binding growth factors such as TGF-β1 (48, 67, 76), PDGF (35, 59, 69), 342
or HGF (3, 18, 47). Figure 4C shows that the extent of phosphorylation of Smad-2 in 343
response to TGF-β1, or the phosphorylation of ERK 1/2 in response to PDGF, were 344
unaltered in the C6 glypican-1 deficient myoblast respect to the WT myoblast. 345
Interestingly, the induction of phospho ERK 1/2 in response to HGF was diminished in 346
the glypican-1 deficient myoblast. These results suggest that glypican-1 is not involved 347
with the regulation of TGF-β and PDGF signaling, but they do not exclude the 348
possibility that other signaling pathways, such as HGF, could be regulated directly or 349
indirectly by glypican-1. 350
Since the FGF-2-dependent inhibition of myogenin expression depends on the 351
activation of the MAPK pathway (83), we determined the inhibitory effect of FGF-2 on 352
the expression of myogenin. Figure 5A (upper panel) shows the inhibitory effect of 353

15
FGF-2 on the activity of pMyo-Luc, which is a reporter plasmid containing the 354
promoter region of myogenin coupled to the luciferase gene (68). Exposure of the cells 355
to FGF-2 resulted in significant inhibition of pMyo-Luc activity. However, C6 cells 356
showed a marked shift in the dose-response curve from an IC50 2.0 ng/ml for wild-type 357
myoblasts, to 0.5 ng/ml for non-expressing glypican-1 myoblasts. Re-expression of rat 358
glypican-1 in C6 myoblasts shifted the FGF-2 sensitivity to values closer to WT (~1.3 359
ng/ml). Similar results were obtained when the expression of glypican-1 in wild-type 360
myoblasts was diminished by transient transfection of the shGly (Fig. 5B). A shift in the 361
dose-response curve was observed from an IC50 2.0 ng/ml for wild-type cells to 0.6 362
ng/ml for myoblasts transfected with shGly. When the wild-type myoblasts were 363
transfected with the scrambler shRNA, no effect in the IC50 was observed (Fig. 5B). 364
The above result suggests that the altered muscle differentiation process 365
observed in the glypican-1-deficient myoblasts could be explained by an augmented 366
sensitivity to the inhibitory signaling of FGF-2. To probe this, we decided to block the 367
FGF-2 present in the differentiation medium, which is produced by the myoblast itself 368
(data not shown) (43), through its inactivation with a soluble form of the FGFR-I or by 369
the use of a neutralizing antibody against FGF-2. Figure 5C and 5E shows that the 370
phospho ERK1/2 levels induced by exogenously-added FGF-2 in WT and C6 myoblasts 371
was diminished in the presence of the soluble receptor or the neutralizing antibody, in a 372
dose dependent manner. Figure 5D shows that the myogenin and myosin levels of WT 373
myoblasts after 2 or 4 days in differentiation media are slightly augmented in the 374
presence of the soluble FGFR-I. However, in the C6 myoblasts the presence of the 375
soluble receptor importantly restores the diminished levels. Similar results were 376
obtained with the neutralizing antibody against FGF-2. Under these conditions, 377
myogenin and myosin levels were augmented in the WT and C6 myoblasts respectively, 378

16
when the corresponding cells were treated with the neutralizing antibody as indicated in 379
Figure 5F. 380
Altogether, these results clearly indicate that glypican-1 inhibits the binding of 381
FGF-2 to its transducing receptors thus diminishing the FGF-2-dependent signaling, and 382
that the blockage of endogenous FGF-2 increased the expression of myogenic markers 383
in glypican-1 deficient myoblasts. These results indicate that glypican-1 inhibits FGF-2-384
dependent signaling in myoblasts, modulating the muscle differentiation process. 385
386
Glypican-1 is the only HSPG localized in myoblast lipid raft domains, binding 387
FGF-2 and not co-localizing with FGFRs. 388
The previous results demonstrate that glypican-1, contrary to the syndecans that 389
act like FGF-2 co-receptors, negatively regulate FGF-2-dependent signaling. To act like 390
a co-receptor, HSPGs require a spatial and structural condition to allow the formation of 391
the signaling ternary complex HSPG-FGF2-FGFR (54, 64, 87). In this sense, it is 392
strictly necessary that the HSPGs physically interact with the FGFRs on the plasma 393
membrane. Hence, we decided to evaluate the distribution of HSPGs in myoblast 394
plasma membrane domains and to compare their distribution with the FGFRs. For this, 395
myoblasts were solubilized in Triton X-100 and fractionated in sucrose density 396
gradients. Figure 6A shows that only glypican-1 was enriched in low-density fractions 397
together with classical markers of raft domains, such as GM-1 and caveolin-1, 398
suggesting that glypican-1 localizes in lipid raft domains. On the other hand, all the 399
members of the syndecan family, and some glypican-1, co-migrated at high density 400
fractions together with the sodium potassium ATPase (Na+/K
+ATPase), which is a non-401
raft domain marker. Figure 6B (upper panel) shows, by indirect 402
immunocytolocalization analysis, that glypican-1 presents a punctuated pattern on the 403

17
cell surface, suggesting its association with membrane microdomains (arrows), as well 404
as glypican-1 localized at the ECM with a fibrillar pattern, which is a typical feature of 405
ECM proteins (arrowheads) (8, 58). When lipid rafts were disrupted by MβCD 406
treatment (60), glypican-1 and caveolin-1 were mostly displaced from lipid raft to non-407
raft domains (Fig. 6C). Under this condition, the punctuated staining of glypican-1 408
changed to a more even staining on myoblast cell surfaces (Fig. 6B, middle panel). 409
When myoblasts were incubated with PI-PLC to remove the plasma membrane-410
associated glypican-1 (8), only a remaining stain associated to the ECM was observed 411
(Fig. 6B, lower panel). Then, we asked if the transiently transfected rat glypican-1in 412
the C6 clone was associated with lipid raft domains. For this, C6 myoblasts transfected 413
with rat glypican-1 containing a FLAG epitope, or the empty vector as control, were 414
fractionated as shown in Figure 6A. The fractions were harvested in three groups; I 415
(fractions 1-4), II (fractions 5-8) and III (fractions 9-12), and the distribution of rat 416
glypican-1was determined with an anti-FLAG antibody. Figure 6D (left panel) reveals 417
that rat glypican-1 is mainly associated with the lipid raft domains (group II) as well as 418
some fractionated in non-raft domains (group III). Figure 6D (right panel) shows that 419
rat glypican-1 expressed in C6 myoblasts exhibited a punctuated distribution pattern on 420
the cell surface, which was disrupted after MβCD treatment. All these results strongly 421
suggest that glypican-1 associates with membrane microdomains. 422
If glypican-1 present in lipid rafts regulates the binding and signaling of FGF-2, 423
it must interact with the ligand. WT and C6 myoblasts were incubated with [125
I]-FGF-424
2, then solubilized with Triton X-100 and fractionated in sucrose density gradients. 425
Figure 7A (left panel), shows that [125
I]-FGF-2 co-fractionated in raft and non-raft 426
domains, which is probably evidence of binding to heparan sulfate chains present in 427
glypican-1 and syndecan members respectively. To determine that binding of [125
I]-428

18
FGF-2 to raft domains was indeed to glypican-1, C6 myoblasts were incubated with the 429
radioactive ligand and fractionated like the WT cells. As expected, no [125
I]-FGF-2 430
migrated in raft domains, whereas all detectable [125
I]-FGF-2 migrated together with the 431
non-raft domain markers (Fig. 7A, right panel). These results clearly indicate that 432
glypican-1 is the only HSPG associated with lipid raft domains in myoblast cell 433
surfaces, where it binds and concentrates FGF-2. 434
Next, we determined the plasma membrane localization of the transducing 435
FGFRs. Western blot analysis of fractionated WT myoblasts showed that FGFR-I and 436
FGFR-IV, the main FGF-2 receptors expressed in skeletal muscles (42), were found 437
only in non-raft domains, co-fractionating with syndecans (Fig. 7B). To determine if the 438
plasma membrane FGFRs were located in non-raft membrane microdomains 439
specifically, we analyzed the distribution of the affinity-crosslinked FGFRs to [125
I]-440
FGF-2 followed by Triton X-100 solubilization and sucrose density fractionation. 441
Figure 7C shows that both receptors bound to FGF-2 fractionated only at the higher 442
density fractions, together with non-raft markers. 443
These data indicate that the plasma membrane FGFRs, as the FGF-2 co-444
receptors syndecans, are only present in non-raft domains, where they can interact and 445
facilitate FGF-2 signaling. 446
Since glypican-1 localized in raft domains would be responsible for the 447
sequestering of FGF-2, we expressed a chimeric form of a HSPG containing the 448
extracellular domain of rat glypican-1 and the transmembrane and cytoplasmic domains 449
of mouse syndecan-1 containing a FLAG epitope (F-GlySyn). C6 myoblasts were 450
transfected with F-GlySyn, lysed and subjected to sucrose density fractionation. Figure 451
8A shows that this chimeric HSPG revealed by an anti-FLAG immunoblot migrated 452
only at high-density fractions. The signaling mediated by FGF-2 in C6 myoblasts 453

19
expressing F-GlySyn form was evaluated. Figure 8B shows in F-GlySyn transfected 454
myoblast, that FGF-2 induce phospho ERK 1/2 to levels even higher than observed in 455
the mock transfected or in the glypican-1rescued C6 myoblasts. Consistently, the 456
induction of myogenin and myosin diminished when the chimeric HSPG form was 457
expressed compared to control transfected or glypican-1 rescued C6 myoblasts, as 458
shown in Figure 8C. These results suggest that the F-GlySyn form, present in non-raft 459
domains, would be acting as a presenter of FGF-2 to its transducing receptors. If so, F-460
GlySyn should interacts with the FGFRs. Figure 8D shows that by co-461
immunoprecipitation experiments with anti-FLAG antibodies, that FGFR-IV was co-462
immunoprecipitated with F-GlySyn. As expected, rat glypican-1 containing a FLAG 463
epitope as well as mock myoblasts did not co-immunoprecipitate any FGFR-IV. As 464
positive control syndecan-4 was co-immunopreciptated with FGFR-IV (Fig. 8D). The 465
same Figure shows the expression of rat glypican-1 and the quimeric F-GlySyn 466
determined by immunoreactivity with the anti glypican-1 antibody. Finally the presence 467
of syndecan-4 co-immunoprecipitated from control C& myoblasts is shown. The above 468
results clearly indicate that glypican-1 modulates muscle differentiation processes most 469
likely by sequestering FGF-2 in lipid raft domains, avoiding interaction of the ligand 470
with its receptors. 471
472
Glypican-1, present on the plasma membrane, is required for successful skeletal 473
muscle differentiation, independent from the ECM. 474
Previously it has been shown that glypican-1 is present on the plasma membrane 475
and on the ECM (8). Glypican-1 that is present in non-raft domains (Fig. 6A), as 476
evidenced by its core protein after Hase treatment, likely corresponds to ECM-477
associated glypican-1 since most of it disappears after a cell surface heparin wash. This 478

20
competes for the heparan sulfate chain binding sites without affecting the levels of 479
glypican-1 in the lighter fractions associated to raft domains (Fig. 9A). The heparin 480
wash effectively removes most of the ECM-associated glypican-1, since its fibrillar 481
pattern over the cell disappears after the wash, without affecting the cell-associated 482
glypican-1, as shown by immunofluorescence analysis in Figure 9B. To confirm that 483
part of glypican-1 is associated to the ECM, we determined the co-localization of 484
glypican-1 and the ECM protein laminin. Figure 9C shows that a fraction of glypican-1 485
co-localizes with laminin. All these results suggest that the glypican-1 that fractionates 486
in non-raft domains corresponds to an ECM-associated HSPG. 487
Since glypican-1 was found to be associated with lipid raft domains and also 488
present at the ECM, we raised the question of whether myoblast glypican-1 associated 489
with the plasma membrane, or with the ECM, is required for its negative role over the 490
FGF-2-dependent signaling and skeletal muscle differentiation. Figure 10 shows that in 491
myoblasts transiently transfected with a scrambled shRNA (shCtrl) and an expression 492
vector for E-GFP, myogenin expression was detected in the nuclei (Fig. 10A, D and G). 493
Consequently these cells were also able to fuse with forming myotubes (Fig. 10I). In 494
contrast, WT myoblasts transiently co-transfected with the shGly and an expression 495
vector for E-GFP were unable to express nucleus-associated myogenin after 2 days 496
under differentiation conditions (arrows, Fig. 10B, E and H), confirming that in the 497
absence of glypican-1 this early step in the muscle differentiation process is abolished. 498
These glypican-1 deficient cells, after 4 days under differentiation conditions, were 499
unable to fuse with forming myotubes that express myosin (Fig. 10C, F). Interestingly, 500
these occurred despite the fact that the glypican-1 negative E-GFP-expressing 501
myoblasts (green) were surrounded by glypican-1 present at the ECM network, as 502
evidenced by specific glypican-1 immunostaining (arrowheads). Figure 10J shows 503

21
quantification of this experiment. The left panel indicates that almost 40% of the nuclei 504
of control transfected E-GFP-expressing myoblasts were positive for myogenin, 505
whereas in the shGly-transfected myoblasts, this value was less than 5%. The right 506
panel shows that almost 35% of the control transfected myoblasts were able to fuse with 507
myosin-expressing myotubes, revealed by the co-expression of myosin and E-GFP. On 508
the contrary, less than 10% of the shGly transfected myoblasts were able to fuse with 509
myosin-expressing myotubes. These results strongly suggest that glypican-1 present on 510
the plasma membrane in lipid raft domains is required for successful skeletal muscle 511
differentiation, independent from the ECM-associated glypican-1. 512

22
DISCUSSION 513
In this paper we have shown that glypican-1 is required for a proper skeletal 514
muscle differentiation process. Myoblasts with low levels of glypican-1, either by 515
transient transfection with shGly or a stable clone that constitutively expresses this 516
shRNA (C6 myoblast clone), show low levels of myogenin and myosin with a 517
diminished fusion index when compared to WT myoblasts after having been induced to 518
differentiate. Another marker of skeletal muscle differentiation, namely caveolin-3 (34), 519
presents the same behavior. These defective consequences are glypican-1 dependent, 520
since re-expression of glypican-1 in the C6 myoblasts with rat glypican-1 restores 521
myosin and caveolin-3 expression, as well as myotube formation. The muscle 522
differentiation process depends on the expression levels of glypican-1, since other 523
clones, which express intermediate levels of glypican-1 compared to WT and C6 524
myoblasts, express medium levels of myosin when induced to differentiate (data not 525
shown). 526
It has been well established that FGF-2 a strong myogenesis inhibitor (13, 43, 527
58, 78), diminish the expression of the master gene myogenin (70, 71). Thus, its 528
signaling must be finely controlled. Skeletal muscles mainly express FGFR-I and 529
FGFR-IV (42), which both have a high affinity for FGF-2 (57). FGFs are normally 530
present in muscle tissue and appear to be released upon injury and are expressed at 531
higher levels during regeneration (20, 22, 75). Both receptors have different expression 532
patterns during the muscular differentiation process. FGFR-I is temporally unchanged 533
during the initial days and diminishes later, but it is still present during muscle 534
differentiation (40, 42, 61). In contrast, FGFR-IV is upregulated during this process (42, 535
88), and has been proposed to be essential for muscle regeneration (90). Since the 536
signaling of FGF-2 through its receptor depends on the presence of HSPGs (66, 87), 537

23
regulation by these FGF-2 co-receptors seems to attenuate FGF-2-dependent signaling, 538
thus allowing myogenesis. In different systems, it has been shown that syndecans (6, 25, 539
31, 33, 43, 84, 89) and glypicans (53, 77, 81) have the ability to bind FGF-2, 540
modulating its binding and signaling. 541
We, among others, have shown that HSPGs are essential for FGF-2-mediated 542
signaling in skeletal muscle cells (17, 21, 33, 43, 56, 66). It has been demonstrated that 543
syndecan-1 and-3 are directly involved in this phenomenon, acting as co-receptors of 544
FGF-2 in myoblasts (33, 43). This is crucial, since myoblasts that do not express 545
HSPGs, or are deficient in some of its forms, present an effected process of skeletal 546
muscle formation. 547
The expression of all syndecans is downregulated during the skeletal muscle 548
differentiation process (33, 37, 44) suggesting that might be associated with a 549
diminished sensitivity to the inhibitory effect of FGF-2. In contrast, the expression level 550
of glypican-1 is constant through this process, hence being the main HSPG present 551
during myogenesis (8, 37). 552
Our results unequivocally demonstrate that glypican-1 is required for terminal 553
myogenesis, which raises the question of how glypican-1 regulates the FGF-2-554
dependent signaling during the muscle differentiation process. Our experimental 555
evidence indicates that in the absence of glypican-1, the binding of FGF-2 to its 556
receptors augments, increasing the activation of the MAPK ERK 1/2 and PI3K/AKT 557
pathways and the FGF-2-dependent inhibition of myogenin expression. These effects 558
were directly associated with the absence of glypican-1, since rat glypican-1 re-559
expression restored FGF-2-dependent signaling near to WT levels. The blockage of 560
FGF-2 activity present in the differentiation medium using a soluble form of the FGFR-561
I or a neutralizing antibody against FGF-2, partially restored the altered muscle 562

24
differentiation process in the glypican-1 deficient myoblast, suggesting that the 563
deleterious effect of the absence of glypican-1 over the myogenesis is a consequence of 564
an increased sensitivity to FGF-2. 565
The formation of the ternary signaling complex involving HSPG, FGF2 and 566
FGFR (72) requires that these three components physically interact on the plasma 567
membrane. We show that in myoblasts glypican-1 is the only HSPG found associated 568
with lipid raft membrane domains, away from all the syndecans and FGFRs. The 569
localization in raft domains is sustained by low-density fractionation in the sucrose 570
gradients and co-fractionation with specific lipid raft domain markers. Glypican-1 571
shows a punctuated appearance, which typically characterizes lipid raft domain 572
localization, and disappears after MβCD treatment (4). This pattern corresponds to 573
plasma membrane glypican-1, since totally disappears after treatment with PI-PLC. 574
Glypican-1 localization in raft membrane domains is reinforced by the total abolishment 575
of staining in glypican-1-deficient cells, and the re-appearance of the punctuated 576
staining after re-expression of rat glypican-1. In contrast, all the syndecan forms and 577
FGFRs co-fractionated in non-lipid raft domains, determined by co-fractionation of 578
specific markers at high-density sucrose fractions. Other authors have suggested that 579
clustering of syndecans 1 and 4 with antibodies, or after treatment with FGF-2, induces 580
a re-localization of part of this HSPG from non-raft to raft microdomains in lymphoid 581
and epithelial cells respectively (51, 82). We did not observe any change in the 582
distribution of HSPGs when myoblasts were treated with FGF-2 (data not shown). This 583
indicates that glypican-1 remains the only HSPG associated with lipid rafts under our 584
experimental conditions. 585
These results suggest that gypican-1 could be sequestering FGF-2 in lipid rafts, 586
away from its transducing receptors. Our experiments indicate that FGF-2 binds and co-587

25
migrates with glypican-1 in myoblast isolated membrane raft domains, since no FGF-2 588
was found in lipid rafts of myoblasts deficient for glypican-1. FGF-2 bound to FGFRs 589
located at the cell surface fractionated in non-raft membrane domain fractions, as 590
determined by crosslink assays. Furthermore glypican-1 does not interact with FGFR-591
IV determined by co-immunoprecipitation experiments. This suggests that the FGF-2-592
FGFR complex is formed and maintained in a different spatial localization than 593
glypican-1. The notion that co-localization of HSPG with FGFRs is critical for FGF-2 594
signaling is reinforced by the experiments expressing the F-GlySyn chimeric form. 595
Despite containing a glypican-1 ectodomain, this HSPG was expressed in non-raft 596
domains most likely as a consequence of the presence of syndecan cytoplasmic and 597
transmembrane domains on its structure. This chimeric form increased FGF-2 598
dependent signaling, interacting at least with FGFR-IV. 599
Since glypican-1 is endogenously processed to a soluble form that is 600
incorporated to the ECM (8) co-localizing with laminin, the possibility that glypican-1 601
present in the ECM is sequestering FGF-2 cannot be excluded. Glypican-1 deficient 602
myoblasts did not express myogenin nor fused with control myoblasts, which expressed 603
myogenin and later formed elongated myotubes. These processes occurred in an ECM 604
enriched with glypican-1, synthesized and processed by the control myoblast. This 605
suggests that glypican-1 present in raft membrane domains is the required form for a 606
proper muscle differentiation processes, and is probably responsible for the inhibitory 607
effect on FGF-2 bio-availability. 608
Other functions for glypican-1, besides the inhibitory effect on FGF-2 609
availability, cannot be excluded. HSPGs interact with several ECM constituents (5) and 610
glypican-1 deficient myoblasts present a diminished capability to fuse and form 611
elongated myotubes. This might reflect the possibility of other functions of glypican-1. 612

26
Since HSPG can bind several ligands (28), such as Wnt (15), bone morphogenic protein 613
(BMP), FGF (53, 77, 81), sonic hedgehog (16, 29), distinct members of TGFβ (48, 67), 614
PDGF (35, 36, 59, 69) and HGF (3, 47), it is highly possible that glypican-1 might have 615
other functions in the raft membrane domain. We tested the signaling response to 616
TGFβ-1, PDGF and HGF in the presence or absence of glypican-1. We did not detect 617
any differences between glypican-1 deficient and WT myoblasts to TGF-β and PDGF, 618
suggesting that the response to these growth factors does not depend on glypican-1. 619
However, we detected a decrease in signaling response to HGF in glypican-1 deficient 620
compared to WT myoblats, this might indicate that glypican-1 directly or indirectly is 621
involved in the signaling response to HGF, nevertheless the localization of c-Met, the 622
receptor for HGF, in membrane microdomains is controversial (74). It is worth 623
mentioning that in myoblasts, it has been shown that BMP receptor type II (BMP-RII) is 624
located in lipid raft membrane domains (38) and that the BMP-RII and the BMP-RIA 625
are upregulated during myogenesis (1), although there is no functional evidence for this 626
co-localization. 627
In summary, we have shown that glypican-1 located in membrane raft domains 628
diminishes the bio-availability of FGF-2, sequestering this growth factor away from its 629
transducing receptors. As a consequence, a decrease in FGF-2-dependent signaling 630
occurs, allowing skeletal muscle differentiation to succeed. This novel mechanism of 631
sequestering FGF-2 in lipid rafts, together with the downregulation of the syndecans 632
which co-reside with FGFRs, might be essential to assure successful skeletal muscle 633
differentiation during development and muscle regeneration. 634

27
ACKNOWLEDGEMENTS 635
The authors are indebted to Dr. David J. Carey (Siegfried and Janet Weiss 636
Center for Research, Danville, PA, USA) and to Dr. Ralph D. Sanderson (University of 637
Alabama at Birmingham, Alabama, USA), for providing anti glypican-1 antibody and 638
pcDNA 3.0-rat-glypican-1, respectively. We thank Drs. Juan Larraín and Hugo Olguín 639
(P. Universidad Católica de Chile) for offering encouragement and helpful suggestions. 640
This study was supported by research grants from FONDAP-Biomedicine # 13980001, 641
CARE PFB 12/2007 and the Muscular Dystrophy Association # 89419, as well as the 642
doctoral fellowship granted to Jaime A. Gutierrez by CONYCYT, Chile. 643

28
REFERENCES 644
1. Akiyama, S., T. Katagiri, M. Namiki, N. Yamaji, N. Yamamoto, K. 645
Miyama, H. Shibuya, N. Ueno, J. M. Wozney, and T. Suda. 1997. 646
Constitutively active BMP type I receptors transduce BMP-2 signals without the 647
ligand in C2C12 myoblasts. Exp Cell Res 235:362-9. 648
2. Anastasi, S., S. Giordano, O. Sthandier, G. Gambarotta, R. Maione, P. 649
Comoglio, and P. Amati. 1997. A natural hepatocyte growth factor/scatter 650
factor autocrine loop in myoblast cells and the effect of the constitutive Met 651
kinase activation on myogenic differentiation. J Cell Biol 137:1057-68. 652
3. Ashikari, S., H. Habuchi, and K. Kimata. 1995. Characterization of heparan 653
sulfate oligosaccharides that bind to hepatocyte growth factor. J Biol Chem 654
270:29586-93. 655
4. Beer, C., L. Pedersen, and M. Wirth. 2005. Amphotropic murine leukaemia 656
virus envelope protein is associated with cholesterol-rich microdomains. Virol J 657
2:36. 658
5. Bernfield, M., M. Gotte, P. W. Park, O. Reizes, M. L. Fitzgerald, J. 659
Lincecum, and M. Zako. 1999. Functions of cell surface heparan sulfate 660
proteoglycans. Annu Rev Biochem 68:729-77. 661
6. Bernfield, M., and R. D. Sanderson. 1990. Syndecan, a developmentally 662
regulated cell surface proteoglycan that binds extracellular matrix and growth 663
factors. Philos Trans R Soc Lond B Biol Sci 327:171-86. 664
7. Brady, J. D., T. C. Rich, X. Le, K. Stafford, C. J. Fowler, L. Lynch, J. W. 665
Karpen, R. L. Brown, and J. R. Martens. 2004. Functional role of lipid raft 666
microdomains in cyclic nucleotide-gated channel activation. Mol Pharmacol 667
65:503-11. 668
8. Brandan, E., D. J. Carey, J. Larrain, F. Melo, and A. Campos. 1996. 669
Synthesis and processing of glypican during differentiation of skeletal muscle 670
cells. Eur J Cell Biol 71:170-6. 671
9. Brandan, E., and J. Larraín. 1998. Heparan sulfate proteoglycans during 672
terminal skeletal muscle cell differentiation: Possible functions and regulation of 673
their expression. Basic and Applied Myology 8:107-14. 674
10. Brandan, E., M. Maldonado, J. Garrido, and N. C. Inestrosa. 1985. 675
Anchorage of collagen-tailed acetylcholinesterase to the extracellular matrix is 676
mediated by heparan sulfate proteoglycans. J Cell Biol 101:985-92. 677

29
11. Brown, D. 1994. GPI-anchored proteins and detergent-resistant membrane 678
domains. Braz J Med Biol Res 27:309-15. 679
12. Brown, D. A., and E. London. 1998. Functions of lipid rafts in biological 680
membranes. Annu Rev Cell Dev Biol 14:111-36. 681
13. Brunetti, A., and I. D. Goldfine. 1990. Role of myogenin in myoblast 682
differentiation and its regulation by fibroblast growth factor. J Biol Chem 683
265:5960-3. 684
14. Campos, A., R. Nunez, C. S. Koenig, D. J. Carey, and E. Brandan. 1993. A 685
lipid-anchored heparan sulfate proteoglycan is present in the surface of 686
differentiated skeletal muscle cells. Isolation and biochemical characterization. 687
Eur J Biochem 216:587-95. 688
15. Capurro, M. I., Y. Y. Xiang, C. Lobe, and J. Filmus. 2005. Glypican-3 689
promotes the growth of hepatocellular carcinoma by stimulating canonical Wnt 690
signaling. Cancer Res 65:6245-54. 691
16. Capurro, M. I., P. Xu, W. Shi, F. Li, A. Jia, and J. Filmus. 2008. Glypican-3 692
inhibits Hedgehog signaling during development by competing with patched for 693
Hedgehog binding. Dev Cell 14:700-11. 694
17. Casar, J. C., C. Cabello-Verrugio, H. Olguin, R. Aldunate, N. C. Inestrosa, 695
and E. Brandan. 2004. Heparan sulfate proteoglycans are increased during 696
skeletal muscle regeneration: requirement of syndecan-3 for successful fiber 697
formation. J Cell Sci 117:73-84. 698
18. Catlow, K. R., J. A. Deakin, Z. Wei, M. Delehedde, D. G. Fernig, E. 699
Gherardi, J. T. Gallagher, M. S. Pavao, and M. Lyon. 2008. Interactions of 700
hepatocyte growth factor/scatter factor with various glycosaminoglycans reveal 701
an important interplay between the presence of iduronate and sulfate density. J 702
Biol Chem 283:5235-48. 703
19. Cornelison, D., M. Filla, H. Stanley, A. Rapraeger, and B. Olwin. 2001. 704
Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and 705
are implicated in satellite cell maintenance and muscle regeneration. Dev Biol 706
239:79-94. 707
20. Cornelison, D., and B. Wold. 1997. Single-cell analysis of regulatory gene 708
expression in quiescent and activated mouse skeletal muscle satellite cells. Dev 709
Biol 191:270-83. 710

30
21. Cornelison, D. D., M. S. Filla, H. M. Stanley, A. C. Rapraeger, and B. B. 711
Olwin. 2001. Syndecan-3 and syndecan-4 specifically mark skeletal muscle 712
satellite cells and are implicated in satellite cell maintenance and muscle 713
regeneration. Dev Biol 239:79-94. 714
22. Cornelison, D. D., B. B. Olwin, M. A. Rudnicki, and B. J. Wold. 2000. 715
MyoD(-/-) satellite cells in single-fiber culture are differentiation defective and 716
MRF4 deficient. Dev Biol 224:122-37. 717
23. Cornelison, D. D., S. A. Wilcox-Adelman, P. F. Goetinck, H. Rauvala, A. C. 718
Rapraeger, and B. B. Olwin. 2004. Essential and separable roles for Syndecan-719
3 and Syndecan-4 in skeletal muscle development and regeneration. Genes Dev 720
18:2231-6. 721
24. Couchman, J. R. 2003. Syndecans: proteoglycan regulators of cell-surface 722
microdomains? Nat Rev Mol Cell Biol 4:926-37. 723
25. Chernousov, M. A., and D. J. Carey. 1993. N-syndecan (syndecan 3) from 724
neonatal rat brain binds basic fibroblast growth factor. J Biol Chem 268:16810-725
4. 726
26. Choi, S. C., S. J. Kim, J. H. Choi, C. Y. Park, W. J. Shim, and D. S. Lim. 727
2008. Fibroblast growth factor-2 and -4 promote the proliferation of bone 728
marrow mesenchymal stem cells by the activation of the PI3K-Akt and ERK1/2 729
signaling pathways. Stem Cells Dev 17:725-36. 730
27. Droguett, R., C. Cabello-Verrugio, C. Riquelme, and E. Brandan. 2006. 731
Extracellular proteoglycans modifies TGF-beta bio-availability attenuating its 732
signaling during skeletal muscle differentiation. Matrix Biol 25:332-341. 733
28. Fico, A., F. Maina, and R. Dono. 2007. Fine-tuning of cell signalling by 734
glypicans. Cell Mol Life Sci. 735
29. Filmus, J., and M. Capurro. 2008. The role of glypican-3 in the regulation of 736
body size and cancer. Cell Cycle 7:2787-90. 737
30. Filmus, J., and S. B. Selleck. 2001. Glypicans: proteoglycans with a surprise. J 738
Clin Invest 108:497-501. 739
31. Filla, M. S., P. Dam, and A. C. Rapraeger. 1998. The cell surface 740
proteoglycan syndecan-1 mediates fibroblast growth factor-2 binding and 741
activity. J Cell Physiol 174:310-21. 742
32. Fransson, L. A. 2003. Glypicans. Int J Biochem Cell Biol 35:125-9. 743

31
33. Fuentealba, L., D. J. Carey, and E. Brandan. 1999. Antisense inhibition of 744
syndecan-3 expression during skeletal muscle differentiation accelerates 745
myogenesis through a basic fibroblast growth factor-dependent mechanism. J 746
Biol Chem 274:37876-37884. 747
34. Galbiati, F., D. Volonte, J. A. Engelman, P. E. Scherer, and M. P. Lisanti. 748
1999. Targeted down-regulation of caveolin-3 is sufficient to inhibit myotube 749
formation in differentiating C2C12 myoblasts. Transient activation of p38 750
mitogen-activated protein kinase is required for induction of caveolin-3 751
expression and subsequent myotube formation. J Biol Chem 274:30315-21. 752
35. Garcia-Olivas, R., J. Hoebeke, S. Castel, M. Reina, G. Fager, F. Lustig, and 753
S. Vilaro. 2003. Differential binding of platelet-derived growth factor isoforms 754
to glycosaminoglycans. Histochem Cell Biol 120:371-82. 755
36. Garcia-Olivas, R., S. Vilaro, M. Reina, and S. Castel. 2007. PDGF-stimulated 756
cell proliferation and migration of human arterial smooth muscle cells. 757
Colocalization of PDGF isoforms with glycosaminoglycans. Int J Biochem Cell 758
Biol 39:1915-29. 759
37. Gutierrez, J., N. Osses, and E. Brandan. 2006. Changes in secreted and cell 760
associated proteoglycan synthesis during conversion of myoblasts to osteoblasts 761
in response to bone morphogenetic protein-2: role of decorin in cell response to 762
BMP-2. J Cell Physiol 206:58-67. 763
38. Hartung, A., K. Bitton-Worms, M. M. Rechtman, V. Wenzel, J. H. 764
Boergermann, S. Hassel, Y. I. Henis, and P. Knaus. 2006. Different routes of 765
bone morphogenic protein (BMP) receptor endocytosis influence BMP 766
signaling. Mol Cell Biol 26:7791-805. 767
39. Hering, H., C. C. Lin, and M. Sheng. 2003. Lipid rafts in the maintenance of 768
synapses, dendritic spines, and surface AMPA receptor stability. J Neurosci 769
23:3262-71. 770
40. Itoh, N., T. Mima, and T. Mikawa. 1996. Loss of fibroblast growth factor 771
receptors is necessary for terminal differentiation of embryonic limb muscle. 772
Development 122:291-300. 773
41. Jenniskens, G. J., J. H. Veerkamp, and T. H. van Kuppevelt. 2006. Heparan 774
sulfates in skeletal muscle development and physiology. J Cell Physiol 206:283-775
94. 776

32
42. Kwiatkowski, B. A., I. Kirillova, R. E. Richard, D. Israeli, and Z. Yablonka-777
Reuveni. 2008. FGFR4 and its novel splice form in myogenic cells: Interplay of 778
glycosylation and tyrosine phosphorylation. J Cell Physiol 215:803-17. 779
43. Larrain, J., D. J. Carey, and E. Brandan. 1998. Syndecan-1 expression 780
inhibits myoblast differentiation through a basic fibroblast growth factor-781
dependent mechanism. J Biol Chem 273:32288-96. 782
44. Larrain, J., G. Cizmeci-Smith, V. Troncoso, R. C. Stahl, D. J. Carey, and E. 783
Brandan. 1997. Syndecan-1 expression is down-regulated during myoblast 784
terminal differentiation. Modulation By growth factors and retinoic acid. J Biol 785
Chem 272:18418-24. 786
45. Le Roy, C., and J. L. Wrana. 2005. Clathrin- and non-clathrin-mediated 787
endocytic regulation of cell signalling. Nat Rev Mol Cell Biol 6:112-26. 788
46. Liu, W., E. D. Litwack, M. J. Stanley, J. K. Langford, A. D. Lander, and R. 789
D. Sanderson. 1998. Heparan sulfate proteoglycans as adhesive and anti-790
invasive molecules. Syndecans and glypican have distinct functions. J Biol 791
Chem 273:22825-32. 792
47. Lyon, M., J. A. Deakin, K. Mizuno, T. Nakamura, and J. T. Gallagher. 793
1994. Interaction of hepatocyte growth factor with heparan sulfate. Elucidation 794
of the major heparan sulfate structural determinants. J Biol Chem 269:11216-23. 795
48. Lyon, M., G. Rushton, and J. T. Gallagher. 1997. The interaction of the 796
transforming growth factor-betas with heparin/heparan sulfate is isoform-797
specific. J Biol Chem 272:18000-6. 798
49. Mansukhani, A., P. Dell'Era, D. Moscatelli, S. Kornbluth, H. Hanafusa, and 799
C. Basilico. 1992. Characterization of the murine BEK fibroblast growth factor 800
(FGF) receptor: activation by three members of the FGF family and requirement 801
for heparin. Proc Natl Acad Sci U S A 89:3305-9. 802
50. Massague, J., S. Cheifetz, T. Endo, and B. Nadal-Ginard. 1986. Type beta 803
transforming growth factor is an inhibitor of myogenic differentiation. Proc Natl 804
Acad Sci U S A 83:8206-10. 805
51. McQuade, K. J., and A. C. Rapraeger. 2003. Syndecan-1 transmembrane and 806
extracellular domains have unique and distinct roles in cell spreading. J Biol 807
Chem 278:46607-15. 808

33
52. Melo, F., D. J. Carey, and E. Brandan. 1996. Extracellular matrix is required 809
for skeletal muscle differentiation but not myogenin expression. J Cell Biochem 810
62:227-39. 811
53. Midorikawa, Y., S. Ishikawa, H. Iwanari, T. Imamura, H. Sakamoto, K. 812
Miyazono, T. Kodama, M. Makuuchi, and H. Aburatani. 2003. Glypican-3, 813
overexpressed in hepatocellular carcinoma, modulates FGF2 and BMP-7 814
signaling. Int J Cancer 103:455-65. 815
54. Mohammadi, M., S. K. Olsen, and O. A. Ibrahimi. 2005. Structural basis for 816
fibroblast growth factor receptor activation. Cytokine Growth Factor Rev 817
16:107-37. 818
55. Olguin, H., and E. Brandan. 2001. Expression and localization of 819
proteoglycans during limb myogenic activation. Dev Dyn 221:106-15. 820
56. Olwin, B. B., and A. Rapraeger. 1992. Repression of myogenic differentiation 821
by aFGF, bFGF, and K-FGF is dependent on cellular heparan sulfate. J Cell Biol 822
118:631-9. 823
57. Ornitz, D. M., J. Xu, J. S. Colvin, D. G. McEwen, C. A. MacArthur, F. 824
Coulier, G. Gao, and M. Goldfarb. 1996. Receptor specificity of the fibroblast 825
growth factor family. J Biol Chem 271:15292-7. 826
58. Osses, N., and E. Brandan. 2002. ECM is required for skeletal muscle 827
differentiation independently of muscle regulatory factor expression. Am J 828
Physiol Cell Physiol 282:C383-94. 829
59. Osterholm, C., M. M. Barczyk, M. Busse, M. Gronning, R. K. Reed, and M. 830
Kusche-Gullberg. 2009. Mutation in the heparan sulfate biosynthesis enzyme 831
EXT1 influences growth factor signaling and fibroblast interactions with the 832
extracellular matrix. J Biol Chem 284:34935-43. 833
60. Ostermeyer, A. G., B. T. Beckrich, K. A. Ivarson, K. E. Grove, and D. A. 834
Brown. 1999. Glycosphingolipids are not essential for formation of detergent-835
resistant membrane rafts in melanoma cells. methyl-beta-cyclodextrin does not 836
affect cell surface transport of a GPI-anchored protein. J Biol Chem 274:34459-837
66. 838
61. Patel, S. G., P. E. Funk, and J. X. DiMario. 1999. Regulation of avian 839
fibroblast growth factor receptor 1 (FGFR-1) gene expression during skeletal 840
muscle differentiation. Gene 237:265-76. 841

34
62. Pellegrini, L. 2001. Role of heparan sulfate in fibroblast growth factor 842
signalling: a structural view. Curr Opin Struct Biol 11:629-34. 843
63. Peng, H., H. Xie, S. Rossi, and R. Rotundo. 1999. Acetylcholinesterase 844
clustering at the neuromuscular junction involves perlecan and dystroglycan. J 845
Cell Biol 145:911-921. 846
64. Plotnikov, A. N., J. Schlessinger, S. R. Hubbard, and M. Mohammadi. 1999. 847
Structural basis for FGF receptor dimerization and activation. Cell 98:641-50. 848
65. Rapraeger, A. 2000. Syndecan-regulated receptor signaling. J Cell Biol 849
149:995-8. 850
66. Rapraeger, A. C., A. Krufka, and B. B. Olwin. 1991. Requirement of heparan 851
sulfate for bFGF-mediated fibroblast growth and myoblast differentiation. 852
Science 252:1705-8. 853
67. Rider, C. C. 2006. Heparin/heparan sulphate binding in the TGF-beta cytokine 854
superfamily. Biochem Soc Trans 34:458-60. 855
68. Riquelme, C., J. Larrain, E. Schonherr, J. P. Henriquez, H. Kresse, and E. 856
Brandan. 2001. Antisense inhibition of decorin expression in myoblasts 857
decreases cell responsiveness to transforming growth factor beta and accelerates 858
skeletal muscle differentiation. J Biol Chem 276:3589-96. 859
69. Rolny, C., D. Spillmann, U. Lindahl, and L. Claesson-Welsh. 2002. Heparin 860
amplifies platelet-derived growth factor (PDGF)- BB-induced PDGF alpha -861
receptor but not PDGF beta -receptor tyrosine phosphorylation in heparan 862
sulfate-deficient cells. Effects on signal transduction and biological responses. J 863
Biol Chem 277:19315-21. 864
70. Rudnicki, M. A., and R. Jaenisch. 1995. The MyoD family of transcription 865
factors and skeletal myogenesis. Bioessays 17:203-9. 866
71. Sabourin, L. A., and M. A. Rudnicki. 2000. The molecular regulation of 867
myogenesis. Clin Genet 57:16-25. 868
72. Schlessinger, J., A. N. Plotnikov, O. A. Ibrahimi, A. V. Eliseenkova, B. K. 869
Yeh, A. Yayon, R. J. Linhardt, and M. Mohammadi. 2000. Crystal structure 870
of a ternary FGF-FGFR-heparin complex reveals a dual role for heparin in 871
FGFR binding and dimerization. Mol Cell 6:743-50. 872
73. Seale, P., and M. Rudnicki. 2000. A new look at the origin, function, and 873
"stem-cell" status of muscle satellite cells. Dev Biol 218:115-24. 874

35
74. Seveau, S., H. Bierne, S. Giroux, M. C. Prevost, and P. Cossart. 2004. Role 875
of lipid rafts in E-cadherin-- and HGF-R/Met--mediated entry of Listeria 876
monocytogenes into host cells. J Cell Biol 166:743-53. 877
75. Sheehan, S., and R. Allen. 1999. Skeletal muscle satellite cell proliferation in 878
response to members of the fibroblast growth factor family and hepatocyte 879
growth factor. J Cell Physiol 181:499-506. 880
76. Shi, Y., and J. Massague. 2003. Mechanisms of TGF-beta signaling from cell 881
membrane to the nucleus. Cell 113:685-700. 882
77. Song, H. H., W. Shi, and J. Filmus. 1997. OCI-5/rat glypican-3 binds to 883
fibroblast growth factor-2 but not to insulin-like growth factor-2. J Biol Chem 884
272:7574-7. 885
78. Spizz, G., J. S. Hu, and E. N. Olson. 1987. Inhibition of myogenic 886
differentiation by fibroblast growth factor or type beta transforming growth 887
factor does not require persistent c-myc expression. Dev Biol 123:500-7. 888
79. Steinfeld, R., H. Van Den Berghe, and G. David. 1996. Stimulation of 889
fibroblast growth factor receptor-1 occupancy and signaling by cell surface-890
associated syndecans and glypican. J Cell Biol 133:405-16. 891
80. Stetzkowski-Marden, F., K. Gaus, M. Recouvreur, A. Cartaud, and J. 892
Cartaud. 2006. Agrin elicits membrane lipid condensation at sites of 893
acetylcholine receptor clusters in C2C12 myotubes. J Lipid Res 47:2121-33. 894
81. Su, G., K. Meyer, C. D. Nandini, D. Qiao, S. Salamat, and A. Friedl. 2006. 895
Glypican-1 is frequently overexpressed in human gliomas and enhances FGF-2 896
signaling in glioma cells. Am J Pathol 168:2014-26. 897
82. Tkachenko, E., and M. Simons. 2002. Clustering induces redistribution of 898
syndecan-4 core protein into raft membrane domains. J Biol Chem 277:19946-899
51. 900
83. Tortorella, L. L., D. J. Milasincic, and P. F. Pilch. 2001. Critical proliferation-901
independent window for basic fibroblast growth factor repression of myogenesis 902
via the p42/p44 MAPK signaling pathway. J Biol Chem 276:13709-17. 903
84. Villena, J., C. Berndt, F. Granes, M. Reina, and S. Vilaro. 2003. Syndecan-2 904
expression enhances adhesion and proliferation of stably transfected Swiss 3T3 905
cells. Cell Biol Int 27:1005-10. 906
85. Yaffe, D., and O. Saxel. 1977. Serial passaging and differentiation of myogenic 907
cells isolated from dystrophic mouse muscle. Nature 270:725-7. 908

36
86. Yang, Y., M. Borset, J. K. Langford, and R. D. Sanderson. 2003. Heparan 909
sulfate regulates targeting of syndecan-1 to a functional domain on the cell 910
surface. J Biol Chem 3:3. 911
87. Yayon, A., M. Klagsbrun, J. D. Esko, P. Leder, and D. M. Ornitz. 1991. Cell 912
surface, heparin-like molecules are required for binding of basic fibroblast 913
growth factor to its high affinity receptor. Cell 64:841-8. 914
88. Yu, S., L. Zheng, D. K. Trinh, S. L. Asa, and S. Ezzat. 2004. Distinct 915
transcriptional control and action of fibroblast growth factor receptor 4 in 916
differentiating skeletal muscle cells. Lab Invest 84:1571-80. 917
89. Zhang, Y., J. Li, C. Partovian, F. W. Sellke, and M. Simons. 2003. 918
Syndecan-4 modulates basic fibroblast growth factor 2 signaling in vivo. Am J 919
Physiol Heart Circ Physiol 284:H2078-82. 920
90. Zhao, P., G. Caretti, S. Mitchell, W. L. McKeehan, A. L. Boskey, L. M. 921
Pachman, V. Sartorelli, and E. P. Hoffman. 2006. Fgfr4 is required for 922
effective muscle regeneration in vivo. Delineation of a MyoD-Tead2-Fgfr4 923
transcriptional pathway. J Biol Chem 281:429-38. 924
91. Zhu, D., W. C. Xiong, and L. Mei. 2006. Lipid rafts serve as a signaling 925
platform for nicotinic acetylcholine receptor clustering. J Neurosci 26:4841-51. 926
927

37
FIGURE LEGENDS 928
929
FIGURE 1: Glypican-1 is required for a successful muscular differentiation 930
process. A: C2C12 myoblasts (WT) were infected with a lentiviral vector to generate a 931
stable clone that expresses a shRNA control (shCtrl) or a shRNA specific for mouse 932
glypican-1 (C6). Glypican-1 levels were determined by Western blot analysis using 933
anti-stub antibodies that recognizes a neo-epitope generated in the heparan sulfate 934
chains after digestion with Hase, enabling the core proteins of any HSPG to be 935
visualized. Syn-3, syn-1, syn-2 and syn-4 represent syndecan-3, -1, -2 and -4, 936
respectively. B: WT, shCtrl and C6 myoblasts were induced to differentiate for 0, 2, 4 937
and 6 days in the differentiation medium (Days DM). Cell extracts were analyzed by 938
Western blot for myogenin, myosin and caveolin-3. Tubulin levels are indicated as a 939
loading control. In A and B, the molecular weights are indicated in kD. C: In a parallel 940
experiment WT and C6 myoblasts were fixed and analyzed by Phase contrast or indirect 941
immunofluorescence for glypican-1 (red) and myosin (green) after 5 days of 942
differentiation (Myotubes D5). Nuclei were stained with Hoechst (blue). 943
944
FIGURE 2: Re-expression of glypican-1 restores the impaired muscular 945
differentiation observed in glypican-1 deficient myoblasts. A: WT and C6 myoblasts 946
were transiently transfected with shGly or with rat glypican-1 (Gly), respectively. 48 h 947
after transfection, the myoblasts were induced to differentiate into myotubes for 4 days 948
(Myotubes D4). The extracts were analyzed by immunoblot analysis for the late muscle 949
differentiation markers, myosin and caveolin-3 (Cav-3). GAPDH levels were used as a 950
loading control. B: Phase contrast images of each experimental condition of A, at day 4 951
of differentiation. C: The glypican-1 protein levels of the myoblast transfected as in A 952
were determined after 48 h by immunoblot analysis by the anti-stub, as described in the 953

38
legend of Figure 1A, D: The glypican-1 protein levels of the myoblast transfected as in 954
A were determined after 48 h by immunoblot analysis with a glypican-1 specific 955
antibody. In A, C and D the molecular weights are indicated in kD. 956
957
FIGURE 3: The binding of FGF-2 to its receptors is augmented in glypican-1-958
deficient myoblasts. A: FGF-2 cell surface receptors of WT myoblasts transiently 959
transfected with or without shCtrl and shGly, and C6 myoblasts transiently transfected 960
with or without rat glypican-1 (C6-Gly), were affinity crosslinked to [125
I]-FGF-2 at 961
4ºC. Cell extracts were separated on SDS-PAGE, and then exposed to a phosphorimager 962
(left). On the right, the gel was stained with Coomasie blue as a loading control. B: The 963
same extracts of A, on the left, were analyzed by Western blot to determine the total 964
protein levels of the FGFR-I and FGFR-IV. GAPDH levels were used as a loading 965
control. C: Myoblasts were treated with or without Hase, then FGFRs were affinity 966
crosslinked to [125
I]-FGF-2 at 4ºC in the presence or absence of an excess of cold FGF-967
2. On the right, the gel was stained with Coomasie blue as a loading control. In A, B 968
and C the molecular weight standards are indicated in kD. 969
970
FIGURE 4: Myoblasts deficient in glypican-1 are more sensitive to FGF-2, but not 971
to other heparin binding growth factors. A: WT and C6 myoblasts were treated with 972
the indicated concentration of FGF-2 for 5 min. Cell extracts were analyzed for phospho 973
ERK1/2 by immunoblot analysis. The levels of total ERK 1/2 were used as a loading 974
control. On the right, a quantification of two independent experiments is shown. B: C6 975
myoblasts were transiently transfected with or without rat glypican-1 (C6-Gly), and 48 976
h after transfection the cells were treated as in A. A quantification of this experiment is 977
shown on the right. C: WT and C6 myoblasts were treated with the indicated 978

39
concentrations of TGFβ-1, PDGF for 15 min or HGF for 5 min. The levels of phospho-979
Smad 2 or phospho ERK1/2 were determined by immunoblot analysis. GAPDH or total 980
ERK 1/2 levels were used as a loading control. In A, B and C the molecular weight 981
standards are indicated in kD. 982
983
FIGURE 5: The absence of glypican-1 induces an increase in the FGF-2-dependent 984
inhibition of the muscular differentiation process. A: WT and the C6 myoblasts were 985
transiently co-transfected with pMyo-luc, the transfection control plasmid (pRL-SV40), 986
an empty pcDNA3.1 plasmid as control, or rat glypican-1 to re-express glypican-1 in 987
the C6 myoblasts (C6-Gly). 48 h after transfection, the cells were induced to 988
differentiate for 30 h in the presence of FGF-2 at the indicated concentrations. The 989
values (pMyo-Luc/pRL-SV40 activity) are expressed as percentages of reporter activity 990
in the absence of FGF-2 for each experimental condition. B: WT myoblasts were co-991
transfected with the reporter system described in A, and a shCtrl or shGly. After 30 h in 992
the differentiation medium, pMyo-Luc and pRL-SV40 activities were determined and 993
expressed as in A. The values shown in A and B are the results obtained from three 994
independent experiments performed in triplicate and correspond to the mean and 995
standard deviation. C and E: WT or C6 myoblasts were treated with FGF-2 pre-996
incubated with or without increasing concentrations of a soluble form of the FGFR-I 997
(FGFR (S)) or a neutralizing antibody against FGF-2 (anti-FGF2), respectively. The 998
phospho and total ERK1/2 levels were determined by immunoblot analysis. D and F: 999
WT and C6 myoblasts were induced to differentiate in the presence or absence of the 1000
FGFR (S) or the anti-FGF2, respectively. The anti-FGF2 was replaced daily. Myosin 1001
and myogenin levels were analyzed by immunoblot after 2 or 4 days. Tubulin levels 1002

40
were used as a loading control in both cases. In all figures the molecular weights are 1003
indicated in kD. 1004
1005
FIGURE 6: Glypican-1 is the only HSPG associated with lipid rafts. A: C2C12 1006
myoblasts were lysed and then fractionated in sucrose density gradients (5-45%). The 1007
twelve fractions collected were analyzed by immunoblot for HSPGs, as explained in the 1008
legend of Figure 1A, as well as the lipid raft membrane protein markers, GM-1 1009
(ganglioside GM-1) and Cav-1. Na+/K
+ATPase (ATPase), were used as a non-lipid raft 1010
domain marker. On the left, the molecular weight standards are indicated in kD. B: 1011
Indirect immunocytolocalization analysis for glypican-1 (red) in C2C12 myoblasts 1012
treated with or without MβCD or PI-PLC. The nuclei were stained with Hoechst (blue). 1013
The arrows indicate the punctuated pattern of glypican-1 on the cell surface, and the 1014
arrowheads point at the ECM associated glypican-1. C: C2C12 myoblasts were treated 1015
with or without MβCD, and then fractionated as in A. The fractions were analyzed for 1016
glypican-1 and caveolin-1 distribution by Western blot analysis. D: On the left, C6 1017
clone myoblasts were transiently transfected with rat glypican-1 containing a FLAG 1018
epitope in its amino terminal (F-Gly) or the empty vector. After 48 h, the cells were 1019
fractionated as in A. The twelve fractions were pooled into three groups; I (fractions 1-1020
4), II (fractions 5-8) and III (fractions 9-12), and analyzed for the distribution of rat 1021
glypican-1 using an anti-FLAG antibody. On the right, in a parallel experiment, C6 1022
myoblasts transfected with F-Gly were treated with or without MβCD, then fixed and 1023
analyzed by immunofluorescence for FLAG epitope. Syn-3, syn-1, syn-2 and syn-4 1024
represent syndecan-3, -1, -2 and -4, respectively. 1025
1026

41
FIGURE 7: Glypican-1 concentrates FGF-2 in lipid raft microdomains that 1027
exclude FGF-2 signaling receptors. A: WT and C6 myoblasts were incubated with 1028
[125
I]-FGF-2 for 3 h at 4 ºC, and then fractionated as in Figure 1. The fractions were 1029
analyzed for HSPGs by anti-stub, as in Figure 1A, or exposed to a phosphorimager to 1030
detect the distribution of [125
I]-FGF-2. B: C2C12 myoblasts were fractionated as 1031
explained in Figure 7A. The fractions were analyzed for the distribution of FGFR-I and 1032
FGFR-IV, as well as membrane distribution markers Cav-1, GM-1 and Na+/K
+ATPase 1033
(ATPase). C: FGF-2 cell surface receptors in C2C12 myoblasts were affinity 1034
crosslinked to [125
I]-FGF-2 at 4ºC. The cells were lysed and fractionated, as explained 1035
in Figure 7A. Aliquots of each fraction were separated on SDS-PAGE (4-10%). The gel 1036
was dried and exposed to a phosphorimager (upper panel), or analyzed by Western 1037
immunoblot for Cav-1, GM-1, and ATPase. [125
I]-FGF2-FGFR-I and [125
I]-FGF2-1038
FGFR-IV correspond to [125
I]-FGF2 crosslinked to FGFR-I or FGFR-IV, respectively. 1039
In A, B and C the molecular weight standards are indicated in kD. 1040
1041
FIGURE 8: The expression of glypican-1 outside of the lipid raft acts like a FGF-2 1042
co-receptor. A: C6 myoblasts were transiently transfected with a chimeric HSPG (F-1043
GlySyn) composed of the extracellular domain of rat glypican-1 and the transmembrane 1044
and cytosolic domain of mouse syndecan-1, containing a FLAG epitope. After 48 h the 1045
cells were lysed and fractionated as in Figure 6A. The distribution of F-GlySyn was 1046
evaluated by anti-FLAG analysis. B: C6 myoblasts were transfected with or without rat 1047
glypican (F-Gly) or F-GlySyn and treated with increasing concentrations of FGF-2. The 1048
phospho ERK 1/2, total ERK1/2 and tubulin levels were determined by immunoblot 1049
analysis. C: C6 myoblasts were transfected with or without rat glypican (F-Gly) or F-1050
GlySyn and induced to differentiate for 2 (D2) or 4 (D4) days to determine the 1051

42
myogenin, myosin and tubulin levels by immunoblot. D: WT or C6 myoblasts were 1052
transiently transfected as in C. 48 h after. The cells were lysed and the extracts 1053
immunoprecipitated with an anti-FLAG antibody or an anti mouse syndecan-4. The 1054
immunoprecipitates were analyzed by western blot for the presence of FGFR-IV, F-Gly 1055
and F-GlySyn with an anti-anti glypican-1 or syndecan-4. The molecular weight 1056
standards are shown in kD. 1057
1058
FIGURE 9: Glypican-1 is present in the plasma membrane lipid rafts and in the 1059
extracellular matrix. A: WT myoblasts were fractionated as shown in Figure 7, after 1060
being washed with heparin in PBS Ca2+
/Mg2+
(Heparin Wash) or PBS Ca2+
/Mg2+
alone 1061
(control). The fractions were analyzed for HSPG core proteins and caveolin-1 1062
distribution. B. Indirect immunofluorescence for glypican-1 (red) of cells treated as in 1063
A. C: Indirect immunofluorescence for glypican-1 (red) and laminin (green) in WT 1064
myoblasts. 1065
1066
FIGURE 10: Glypican-1 is required on the plasma membrane for a proper 1067
muscular differentiation process, independent from extracellular matrix-1068
associated glypican-1. C2C12 myoblasts were transiently co-transfected with the 1069
scrambler shRNA (shCtrl) and a plasmid containing the sequence for E-GFP in A, D 1070
and I, or the shGly in B, E, C and F. 48 h after transfection, the myoblasts were induced 1071
to differentiate for 2 (A, B, D and E) or 4 days (C, F and I). The cells were fixed and 1072
analyzed by immunofluorescence for glypican-1 (red) in A, B and C; myogenin (red) in 1073
D and E; or myosin (red) in F and I. G and H are the same images as D and E, 1074
respectively without the E-GFP signal, in order to better visualize myogenin nuclear 1075
staining. The arrowheads indicate the ECM-associated glypican-1. The arrows indicate 1076

43
the nucleus of transfected cells. J: On the left is a quantification of the co-transfected 1077
myoblasts (shCtrl/E-GFP or shGly/E-GFP) containing myogenin positive nuclei, 1078
compared to the total co-transfected cells (E-GFP positive) after 2 days of 1079
differentiation of ten random fields. On the right is a quantification of the E-GFP-1080
myosin positive myotubes compared the total of E-GFP expressing cells, after 4 days of 1081
differentiation. The data correspond to the mean +/- SE of ten random fields. 1082

A Bk D k D 0 2 4 6 0 2 4 6 0 2 4 6W T s h C t r l C 6 M y o g e n i nD a y s D Mg l y p i c a n � 1s y n � 3s y n � 1 1 1 88 5 C a v 33 6 M y o g e n i nM y o s i n2 0 5s y n � 2s y n � 4 4 73 6 5 52 0 T u b u l i nC a v � 3CW T M y o t u b e s D 5M y o b l a s t sC 6 M y o s i n G l y p i c a n1 0 0 u m 5 0 u mG l y p i c a n
F I G U R E 1
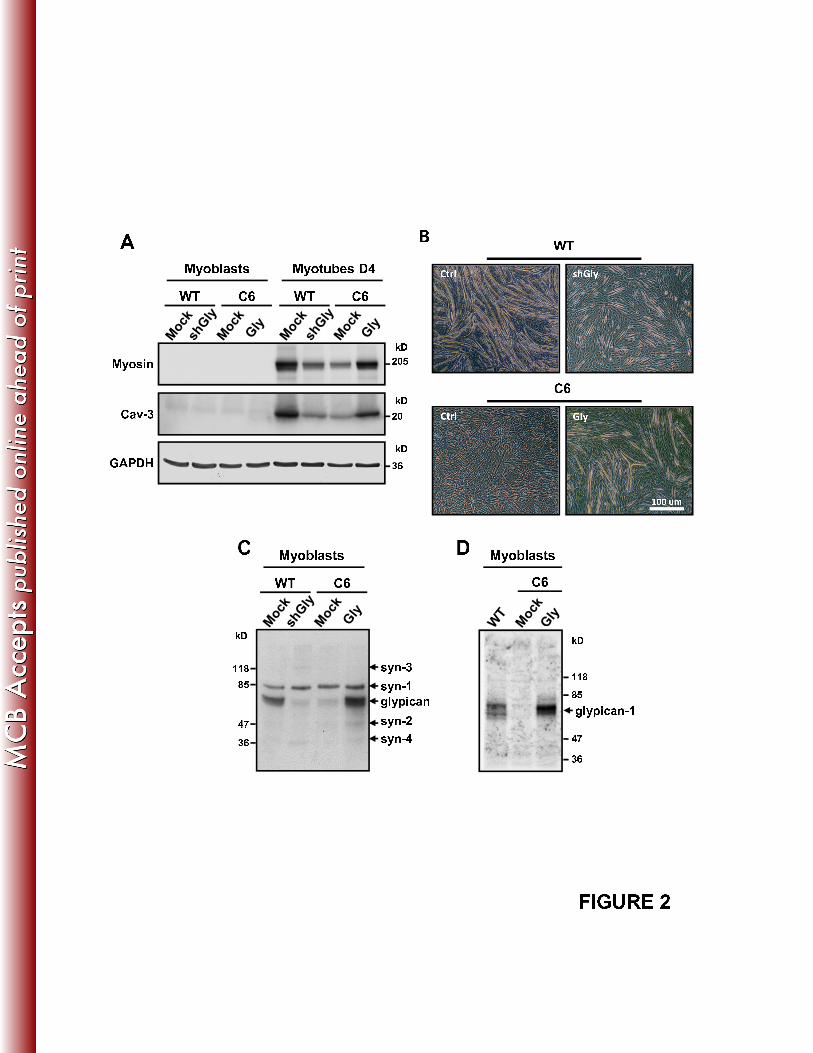
BA C t r l s h G l yW T C 6 W T C 6M y o b l a s t s M y o t u b e s D 4 W TC t r l G l yM y o s i nC a v \ 3 2 02 0 5k Dk D C 6 1 0 0
"u mG A P D H 3 6k DC DW T C 6M y o b l a s t s C 6M y o b l a s t sg l y p i c a ns y n \ 3s y n \ 1s y n \ 21 1 88 54 7k D 1 1 88 5k Dg l y p i c a n \ 1s y n \ 4y4 73 6 4 73 6
F I G U R E 2
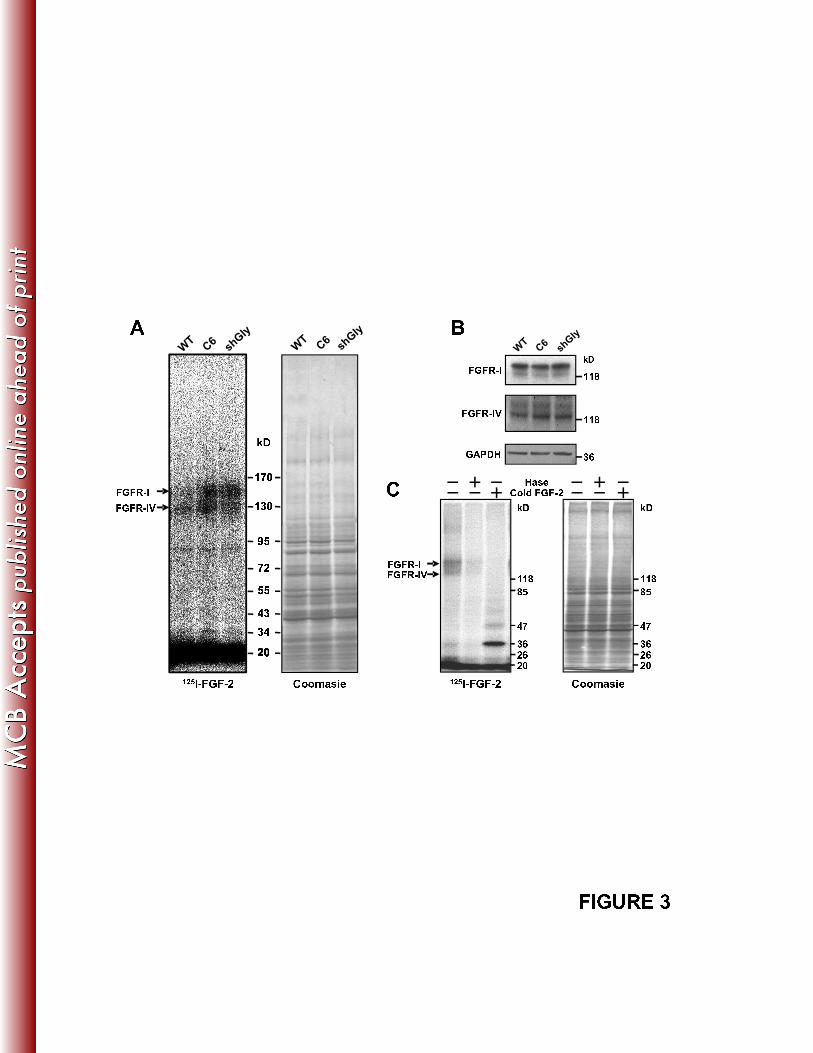
BA B F G F R � I VF G F R � IA 1 1 81 1 8k DG A P D HCF G F R � IF G F R � I V 1 7 01 3 01 7 01 3 0 H a s eC o l d F G F � 2k D k D k D3 69 57 25 54 39 57 25 54 3 F G F R � IF G F R � I V 1 1 88 54 7 1 1 88 54 72 03 42 03 41 2 5 I ¯ F G F ¯ 2 C o o m a s i e 1 2 5 I ¯ F G F ¯ 2 C o o m a s i e2 04 73 62 6 2 04 73 62 6F I G U R E 3

A 68456ot alE rkW T C 6 F G F Ó 2 ( n g / m l )k D 0 , 0 1 0 , 1 1024 W TC 60 0 1 101234Ph osph oE rk/t oP h o s p h oE R K 1 / 24 44 2 T o t a lE R K 1 / 2k D4 44 2B 2323t ot alE rk 0 0 . 1 1F G F Ó 2 ( n g / m l )C 6 C 6 Ó G l y P h o s p h oF G F Ó 2 ( n g / m l )4 4k D 01 C 6 � C t r lC 6 � G l y0 0 . 1 1F G F 2 ( / l )01Ph osph oE rk/pE R K 1 / 24 2 T o t a lE R K 1 / 2k D4 44 2 F G F Ó 2 ( n g / m l )P h o s p h oS m a d � 2 W T C 6 P h o s p h oE R K 1 / 2P D G F( n g / m l )W T C 6C T G F Ó I( n g / m l ) 4 44 2k Dk D6 0 S m a d � 2G A P D H E R K 1 / 2G A P D H4 2k D3 6k D3 6 W T C 6 P h o s p h oE R K 1 / 2H G F( n g / m l )T o t a lE R K 1 / 24 44 2k Dk D4 44 2 F I G U R E 4E R K 1 / 24 2 T u b u l i n

C DE F
F I G U R E 5

BA 1 2 3 4 5 76 98 1 0 1 1 1 2 4 5 %5 % S u c r o s eM y o b l a s t C ont rolg l y p i c a n [ 1k D s y n [ 3s y n [ 1s y n [ 2s y n [ 41 1 88 54 73 6 CD1 2 C a v [ 11 2 3 4 5 76 98 1 0 1 11 2 3 4 5 76 98 1 0 1 1 1 2 G M [ 1s y n [ 42 0k D3 62 6C MCA T P a s eC o n t r o l g l y p i c a n1 2 3 4 5 76 98 1 0 1 1 1 2 4 5 %5 % S u c r o s e1 1 86 6k D PI oPLCM C D C a v [ 1g l y p i c a n1 2 3 4 5 76 98 1 0 1 1 1 2 4 5 %5 % S u c r o s e2 06 6k D C a v [ 1g l y p i c a n 2 0 u m2 06 6 C o n t r o l M C DF [ G l yC o n t r o lD C o n t r o l M C DI I I I I II I I I I I9 57 25 5k D F [ G l y F I G U R E 62 0 u m4 3

A s y n � 3s y n � 1s y n � 3s y n 1 1 2 3 4 5 6 7 8 9 1 0 1 1 1 21 2 3 4 5 6 7 8 9 1 0 1 1 1 2 4 5 %5 % S u c r o s e4 5 %5 % S u c r o s eW T C 61 1 88 5k D 1 1 88 5k D 2 0 k D a1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 s y n � 1s y n � 2g l y � 1s y n � 4s y n � 1s y n � 2g l y � 1s y n � 4G M � 11 2 5 I � F G F � 28 54 73 6 8 54 73 6C k D 1 21 11 0987654321B F G F R I1 21 11 0987654321 4 5 %5 % S u c r o s e 4 5 %5 % S u c r o s ek D 1 1 88 54 7 1 2 5 I È F G F 2 È F G F R È I1 2 5 I È F G F 2 È F G F R È I VF G F R È IG M È 11 2 3 4 5 76 98 1 0 1 1 1 2 F G F R È I V1 1 81 1 8k DG M È 11 2 3 4 5 76 98 1 0 1 1 1 2 C a v È 11 2 3 4 5 76 98 1 0 1 1 1 22 04 7 1 2 5 I È F G F 2C a v È 1A T P a s e1 2 3 4 5 76 98 1 0 1 1 1 21 1 82 0 2 0k Dk D k D A T P a s e1 1 8k D
F I G U R E 7

1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 4 5 %5 % S u c r o s e1 7 0 F í G l y S y nA k D a B C 6 F û G l y S y nF û G l yM o c k P h o p h o4 4k DC 69 57 25 54 31 3 01 7 0 F � G l y S y n P h o p h oE R K 1 / 2T o t a lE R K 1 / 2T u b u l i n4 44 2k D4 44 2G M � 1C 6 DC T u b u l i nC 6M y o g e n i nT u b u l i n5 5k D a3 6 I P P � F L A Gk DI P P � S y n 4 W B : F G F R � I V1 1 8D 2D 0 M y o s i n2 0 5k D a W B : G l y p i c a n � 1W B : S y n � 43 67 24 7T u b u l i n5 5 W B : S y n � 43 6D 4D 0 F I G U R E 8

A 1 2 3 4 5 6 7 8 9 1 0 1 1 1 21 2 3 4 5 6 7 8 9 1 0 1 1 1 2 4 5 %5 % S u c r o s e4 5 %5 % S u c r o s eC o n t r o l H e p a r i n W a s hk D k D C a v l 1s y n l 3s y n l 1s y n l 2s y n l 4g l y p i c a n l 1C a v l 1s y n l 3s y n l 1s y n l 2s y n l 4g l y p i c a n l 1 1 1 88 54 73 61 1 88 54 73 6B C o n t r o l H e p a r i n W a s h 5 0 mL i i MG l i 1C L a m i n i n M e r g eG l y p i c a n � 1 5 0 mF I G U R E 9

BA CB s h G l y + E � G F PA s h C t r l + E � G F P C s h G l y + E � G F PE G L Y P I C A NyD G L Y P I C A Ns h C t r l E G F P F G L Y P I C A NyM Y O G E N I Ns h G l y + E � G F PHM Y O G E N I Ns h C t r l + E � G F PG M Y O S I Ns h G l y + E � G F PIM Y O G E N I Ns h G l y + E � G F PM Y O G E N I Ns h C t r l + E � G F P M Y O S I Ns h C t r l + E � G F P5 0 u mM Y O G E N I NM Y O G E N I N M Y O S I NJ 3 04 05 0positi ve " ve "
cell s positi ve " iti ve "cell s 4 05 001 02 03 0of "
E ど
GFP ど
myogeni n ucl ei/ "
E ど
GFP "
positi of "
E ど
GFP ど
myosi np ot ub es/ "
E ど
GFP "
pos 01 02 03 0F I G U R E 1 0s h G l ys h C t r l s h G l ys h C t r lど
1 0% "
o nu % " myoど1 00
Related Documents











