Joual of Cerebral Blood Flow and Metabolism 20:207-212 © 2000 The International Society for Cerebral Blood Flow and Metabolism Published by Lippincott Williams & Wilkins, Inc., Philadelphia A New Radiopharmaceutical for the Cerebral Dopaminergic Presynaptic Function: 6-Radioiodinated L-meta-Tyrosine Leo Garcia Flores II, Keiichi Kawai, tMamoru Nakagawa, tNaoto Shikano, *Seishi Jinnouchi, *Shozo Tamura, *Katsushi Watanabe, and tAkiko Kubodera Department of Radiology and *Central Research Laboratories, Miyazaki Medical College, Miyazaki, and tFaculty of Pharmaceutical Science, Science University of Tokyo, Tokyo, Japan Summary: Geometric isomers of radioiodinated L-meta- tyrosine, 6-[1-125]iodo-and 4-[1-125]iodo-L-meta-tyrosine (6-1- L-mTyr, 4-I-L-mTyr) were separated by high-performance liq- uid chromatography . Both 6-1- and 4-I-L-mTyr had high en- ergy-dependent brain accumulation. 6-1- and 4-I-L-mTyr were also metabolically stable and were rapidly excreted through the urine. 6-I-L-mTyr had a predilection for the cerebral aromatic Alteration in the dopamine biosynthesis has been im- plicated in development of neurodegenerative (Tissingh et aI., 1997) and neuropsychiatric diseases and drug abuse (Hornykiewicz, 1982; Volkow and Fowler, 1992). Development of radiopharmaceuticals that would assess dopaminergic function in vivo is beneficial. Radiophar- maceuticals are used to distinguish schizophrenic from normal patients (Dao-Castellana et aI., 1997). In idio- pathic Parkinson's disease, radiopharmaceuticals are used for early detection of disease and to monitor effi- cacy of treatments (Tissingh et aI., 1997). It would not only allow us to develop treatment but also at the same time to directly monitor the result of the management of alterations in the dopamine biosynthesis (Sawle et aI., 1992; Choksi et aI., 1997). At present there are two accepted radiopharmaceuti- cals for the study of the dopaminergic presynaptic ter- minals, 2--carboxymethoxy-3--(4-[I-123]iodophen- yl)tropane (I-CIT) and 6-[F-18]fluoro-3,4,-dihydroxy- Received February 4, 1999; final revision received June 22, 1999; accepted June 22, 1999. This study was supported in part by a Grant-in-Aid for Scientific Research (nos. 09770709,08770750, and 09557071) from the Ministry of Education, Science, Sports and Culture of Japan. Address correspondence and reprint requests to Dr. Leo Garcia Flores II, Department of Radiology, Miyazaki Medical College, 5200 Kihara, Kiyotake-cho, Miyazaki-gun, Miyazaki 889-16, Japan. Abbreviation used: HPLC, high-performance liquid chromatogra- phy. 207 L-amino acid decarboxylase (DOPA decarboxylase), the final enzyme of dopamine biosynthesis. 6-Radioiodinated L-mTyr is a new radiopharmaceutical that can be both useful in assessing cerebral amino acid transport mechanism and quantifying metabolically active DOPA decarboxylase. Key Words: 6-lodo-L-meta-tyrosine-Amino acid transport system- DOPA decarboxylase. L-phenylalanine (6-F-DOPA). I-CIT, a cocaine analog, is for single photon emission-computed tomography and has an affinity for dopamine transporters. Because dopa- mine transporters are localized in presynaptic terminal, it serves as a marker of dopamine neurons (Neumeyer et aI., 1991; Asenbaum et aI., 1997). 6-F-DOPA, an analog of L-DOPA, is the only true radiopharmaceutical for presynaptic dopaminergic bio- synthesis (Garnett et aI., 1978). However, 6-F-DOPA is metabolized by catechol-a-methyl transferase to 3-0- methyl-6-F-DOPA which complicates the specific-to- nonspecific accumulation of radioactivity in the brain (Luxen et aI., 1992). It can only be used for positron emission tomography, which is not widely available and is expensive to maintain. So there was a need to develop a new radiopharma- ceutical for single photon emission-computed tomogra- phy that was easy to prepare and whose uptake pattern was easily interpreted. L-meta-Tyrosine (L-mTyr), also an L-DOPA derivative which is not a substrate of cat- echol-a-methyl transferase (De Jesus et aI., 1990), was selected. It has L-amino acid configuration accounting for amino acid transport and tissue affinity. The phenol moiety provides a site for easy iodination. With the similarity of 3-hydroxy moiety to L-DOPA, L-mTyr re- tained its affinity to an aromatic L-amino acid decarbox- ylase (DOPA decarboxylase) (Srinivasan and Awapara, 1978).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Cerebral Blood Flow and Metabolism 20:207-212 © 2000 The International Society for Cerebral Blood Flow and Metabolism Published by Lippincott Williams & Wilkins, Inc., Philadelphia
A New Radiopharmaceutical for the Cerebral Dopaminergic
Presynaptic Function: 6-Radioiodinated L-meta-Tyrosine
Leo Garcia Flores II, Keiichi Kawai, tMamoru Nakagawa, tNaoto Shikano, *Seishi Jinnouchi,
*Shozo Tamura, *Katsushi Watanabe, and tAkiko Kubodera
Department of Radiology and *Central Research Laboratories, Miyazaki Medical College, Miyazaki, and tFaculty of
Pharmaceutical Science, Science University of Tokyo, Tokyo, Japan
Summary: Geometric isomers of radioiodinated L-metatyrosine, 6-[1-125]iodo-and 4-[1-125]iodo-L-meta-tyrosine (6-1-L-mTyr, 4-I-L-mTyr) were separated by high-performance liquid chromatography . Both 6-1- and 4-I-L-mTyr had high energy-dependent brain accumulation. 6-1- and 4-I-L-mTyr were also metabolically stable and were rapidly excreted through the urine. 6-I-L-mTyr had a predilection for the cerebral aromatic
Alteration in the dopamine biosynthesis has been implicated in development of neurodegenerative (Tissingh et aI., 1997) and neuropsychiatric diseases and drug abuse (Hornykiewicz, 1982; Volkow and Fowler, 1992). Development of radiopharmaceuticals that would assess dopaminergic function in vivo is beneficial. Radiopharmaceuticals are used to distinguish schizophrenic from normal patients (Dao-Castellana et aI., 1997). In idiopathic Parkinson's disease, radiopharmaceuticals are used for early detection of disease and to monitor efficacy of treatments (Tissingh et aI., 1997). It would not only allow us to develop treatment but also at the same time to directly monitor the result of the management of alterations in the dopamine biosynthesis (Sawle et aI., 1992; Choksi et aI., 1997).
At present there are two accepted radiopharmaceuticals for the study of the dopaminergic presynaptic terminals, 213-carboxymethoxy-313-( 4- [I -123 ]iodophenyl)tropane (I-I3CIT) and 6-[F-18]fluoro-3,4,-dihydroxy-
Received February 4, 1999; final revision received June 22, 1999; accepted June 22, 1999.
This study was supported in part by a Grant-in-Aid for Scientific Research (nos. 09770709,08770750, and 09557071) from the Ministry of Education, Science, Sports and Culture of Japan.
Address correspondence and reprint requests to Dr. Leo Garcia Flores II, Department of Radiology, Miyazaki Medical College, 5200 Kihara, Kiyotake-cho, Miyazaki-gun, Miyazaki 889-16, Japan.
Abbreviation used: HPLC, high-performance liquid chromatography.
207
L-amino acid decarboxylase (DOPA decarboxylase), the final enzyme of dopamine biosynthesis. 6-Radioiodinated L-mTyr is a new radiopharmaceutical that can be both useful in assessing cerebral amino acid transport mechanism and quantifying metabolically active DOPA decarboxylase. Key Words: 6-lodo-L-meta-tyrosine-Amino acid transport systemDOPA decarboxylase.
L-phenylalanine (6-F-DOPA). I-I3CIT, a cocaine analog,
is for single photon emission-computed tomography and
has an affinity for dopamine transporters. Because dopamine transporters are localized in presynaptic terminal, it serves as a marker of dopamine neurons (Neumeyer et aI., 1991; Asenbaum et aI., 1997).
6-F-DOPA, an analog of L-DOPA, is the only true radiopharmaceutical for presynaptic dopaminergic biosynthesis (Garnett et aI., 1978). However, 6-F-DOPA is
metabolized by catechol-a-methyl transferase to 3-0-methyl-6-F-DOPA which complicates the specific-tononspecific accumulation of radioactivity in the brain (Luxen et aI., 1992). It can only be used for positron emission tomography, which is not widely available and is expensive to maintain.
So there was a need to develop a new radiopharmaceutical for single photon emission-computed tomography that was easy to prepare and whose uptake pattern
was easily interpreted. L-meta-Tyrosine (L-mTyr), also an L-DOPA derivative which is not a substrate of cat
echol-a-methyl transferase (De Jesus et aI., 1990), was selected. It has L-amino acid configuration accounting for amino acid transport and tissue affinity. The phenol moiety provides a site for easy iodination. With the similarity of 3-hydroxy moiety to L-DOPA, L-mTyr retained its affinity to an aromatic L-amino acid decarboxylase (DOPA decarboxylase) (Srinivasan and Awapara, 1978).

208 L. GARCIA FLORES II ET AL.
In our previous study, the radioiodination of L-mTyr led to the development of radio iodinated L-mTyr (I-LmTyr) which has features that are different from its parent compound. The characteristics of I-L-mTyr were evaluated. It was found that I-L-mTyr had high cerebral uptake. Its accumulation in the brain was stereospecific and energy dependent. It was resistant to deiodination and was rapidly excreted. This was attributed to its being in the L-form which penetrates the brain more rapidly and in the hydroxy group at 3-position which accounts for its stability. Hence, I-L-mTyr was then endorsed as a single photon emission-computed tomography imaging agent for evaluation of cerebral amino acid transport mechanism (Kawai et aI., 1999).
The direct radioiodination of L-mTyr resulted in two major geometric isomers, 6-iodo- and 4-iodo-L-metatyrosine (6-I-L-mTyr, 4-I-L-mTyr). In this study, the two radioiodinated geometric isomers of L-mTyr were separated with high-performance liquid chromatography (HPLC) based on the method of Adam et aI. (1989). In
vitro and in vivo animal studies were carried out to determine the most suitable isomer for evaluation of cerebral amino acid transport mechanism and the dopaminergic presynaptic biosynthesis.
MATERIALS AND METHODS
Preparation of [I-12S]-labeled 6-I-L-mTyr,
4-I-L-mTyr, 6-I-n-mTyr and 4-I-n-mTyr Reagent-grade chemicals were used in this experiment. D,L
mTyr was acquired from Sigma (St. Louis, MO, U.S.A.), and separation was done with HPLC using a chiral column (Crownpak CR(-), 4 x 150 mm, Daicel, Tokyo, Japan). [I-125]-NaI was obtained from Amersham (Tokyo, Japan). Radioiodination was performed using the chloramine-T method. Chloramine-T (Aldrich, Milwaukee, WI, U.S.A.) at a concentration of 1.0 x
10-3 mollL in 10 f.1L of 0.05 mollL phosphate buffer (pH 8.5) was added to a mixture of L- or D-mTyr (100 f.1L of 1.0 x 10-3 mollL) and carrier-free [I-125]-NaI (3.7 to 7.4 MBq) in 35 f.1L of 0.4 mollL phosphate buffer (pH 8.5). The resultant solution was allowed to stand for 30 minutes, and sodium metabisulfite (1.0 x 10-3 mollL in 10 f.1L of 0.05 mollL phosphate buffer, pH 8.5) was then added. A geometric isomer separation was then done with HPLC using Nova-Pak CI8 (3.9 x 300 mm, 0.02 mollL potassium acetate: ethanol = 90: 10, flow rate; 0.5 mLl min, retention time; 6-isomer: 22 to 26 minutes; 4-isomer: 17 to 20 minutes; 1-: 4 to 5 minutes; unlabeled mTyr: 5 to 7 minutes) (Adam et a!., 1989). The isomeric purity was confirmed with reinjection in the same HPLC condition. The labeling efficiency and the radiochemical purity were studied by silica gel thin-layer chromatography (TLC, Merck [Darmstadt, Germany]; Art. 5553) using two solvent systems, methanol to acetic acid = 100: 1 (Rf value: I-mTyr, 0.5; 1-, 0.8) and methanol to 10% ammonium acetate = 10: 1 (Rf value: I-mTyr, 0.5; 1-, 0.8). Solvent exchange was done with saline for animal experiments.
As references, 3-[I-125]-L-tyrosine (I-L-Tyr) was prepared using the above mentioned method except the isomeric purification.
J Cereb Blood Flow Metab, Vol. 20, No. I, 2000
In vivo mouse biodistribution and mouse
excretion studies Three to nine ddY male mice (6 weeks old) received 0.1 mL
saline of radioiodinated tyrosine derivatives (11.1 kBq) via the tail vein (intravenously). The mice were anesthetized with ether and then killed at various time intervals by heart puncture. Organs were then dissected, and their radioactivities were measured by a well-type scintillation counter (Aloka [Tokyo, Japan]; ARC-300).
Different groups of three to five ddY male mice (6 weeks old) injected with 0.1 mL saline containing the I-mTyr (55.5 kBq) via the tail vein were placed in metabolic cages under feeding conditions. Samples of urine and feces were collected at various time intervals, and their radioactivities were measured. Thin-layer chromatography and HPLC analyses of the urine samples were conducted under previously mentioned conditions.
In vitro accumulation studies in rat brain
and pancreas Five Wistar male rats weighing 300 to 350 g were killed by
decapitation, and the brain and the pancreas were quickly dissected. The brain and the pancreas were washed with ice-cold Krebs-Ringer phosphate buffer (pH 7.4) and HEPES buffer (pH 7.4), respectively. Both were sliced with a conventional StadieRiggs slicer. The slices (each weighing 100 ± 5 mg) were next placed into vials containing 1.9 mL of Krebs-Ringer phosphate buffer or HEPES buffer (pH 7.4), respectively, as their incubation medium and were preincubated for 10 minutes for temperature equilibration. Then, 0.1 mL of the buffer containing radioiodinated mTyr (1.85 kBq) was added, and incubation was performed at 37°C for 30 minutes.
Inhibition studies of the membrane-active transport were performed using ouabain inhibition. For the ouabain inhibition study, brain slices were preincubated at 37°C for 10 minutes in medium containing 1.0 x 10-5 mollL ouabain, and the radioactive sample was injected and incubated for 30 minutes. For the inhibition studies on the enzyme of the dopamine biosynthesis, 0.1 mL of 1.0 x 10-3 mollL of 3-hydroxybenzyl hydrazine dihydrochloride (NSD-1015, Aldrich) or 1.0 x 10-3 mollL of 3,4-dihydroxyphenylpropylacetamide (H22/54, Aldrich) was added, and incubation was performed at 37°C for 10 minutes. Solvent exchange was performed, and then 0.1 mL of I-mTyr (1.85 kBq) was added and incubated for 30 minutes.
At the end of the incubation period the medium was removed, and the slices were washed twice with 2.0 mL of icecold buffer. The radioactivities of the slices were measured, and the accumulation percentage of the injected dose per gram of slice was then calculated.
In vivo inhibition studies and autoradiography In a ddY male mouse weighing 30 g, 0.1 mL of 6-I-L-mTyr
(670 kBq) was injected intravenously. For the inhibition studies, NSD-1015 or H22/54 (50 mg/kg body weight) was intraperitoneally preloaded 30 minutes before intravenous injection of 6-I-L-mTyr. Five minutes later, the mouse was anesthetized and killed. The brain was immediately dissected, washed, placed in an embedding medium and then frozen at -15°C for 24 hours. With a cryostat microtome (Handex, Shiraimatsu), 20 f.1m axial slices were generated from top to bottom. Slices were then air-dried at -15°C for another 24 hours. Contact was made between tissue and the imaging plate (Fuji Photo Film; BASTR2040, 20 x 40 cm) for at least 24 hours. Images were processed with Bio-Imaging Analyzer (Fuji Photo Film; BAS-2000).

6-I-L-mTyr FOR DOPA DECARBOXYLASE 209
FIG. 1. Chemical structures of 6-1-125- and 4-1-125-L-metatyrosine.
6-Iodo-L-meta-tyrosine (6-I-L-mTyr)
4-Iodo-L-meta -tyrosine (4-I-L-mTyr)
RESULTS
Preparation of [I-125]-labeled 6-I-L-mTyr,
4-I-L-mTyr, 6-I-n-mTyr, and 4-I-n-mTyr
Figure 1 shows the chemical structures of both 6-1-and 4-I-mTyr. Labeling of mTyr gave [I-125]-L- or DmTyr a labeling efficiency of more than 80%. To simplify the steps for the preparation, the purification was performed in the same manner as the isomeric separation. After the isomeric purification, noncarrier-added 6-[1-125]-L-mTyr, 4-[I-125]-L-mTyr, 6-[I-125]-D-mTyr, and 4- [I-125]-D-mTyr with radiochemical purities greater than 95% were obtained, and separation from unlabeled mTyr which had biological activities was confirmed. Each radioiodinated mTyr in saline was stable for about 3 weeks.
In vivo mouse biodistribution and excretion studies
The brain accumulations of 6-I-L-mTyr and 4-I-LmTyr are tabulated in Table 1. In the brain, there was higher uptake with 4�I-L-mTyr than 6-I-L-mTyr in the early phase. In both I-L-mTyr isomers the highest accumulations were at 2 minutes after intravenous administration which was then followed by a gradual decline. With 6-I-L-mTyr, retention was noted unlike with 4-I-LmTyr. In Table 2, rapid clearance of l-mTyr from the blood was also noted compared to 3-I-L-Tyr which is a metabolic intermediate of thyroid hormones that showed rapid enzymatic deiodination (Kawai et aI., 1999). For both geometric isomers of I-L-mTyr, the uptake in the pancreas was higher than in the liver. 6-I-L-mTyr showed higher uptake in the pancreas than 4-I-L-mTyr (Table 3).
Three hours after administration, more than 70% and less than 5% of I-L-mTyr was found in the urine and the
TABLE 1. Brain accumulation of I-l25-meta-tyrosine in mice (% dose/g tissue)
Time (min) 6-I-L-mTyr 4-I-L-mTyr 6-I-o-mTyr
2 0.79 ± 0.02 0.96 ± 0.04 0.22 ± 0.01 5 0.77 ± 0.10 0.85 ± 0.07 0.15 ± 0.06
10 0.76 ± 0.12 0.67 ±0.04 0.08 ± 0.01 30 0.56 ± 0.06 O.49±0.10 0.05 ± om
All values are mean ± SD.
feces, respectively. The urine analysis by thin-layer chromatography 30 minutes after intravenous administration showed that more than 90% of the radioactivity was attributable to unaltered 6-I-L-mTyr, whereas with HPLC analysis, no in vivo isomeric conversion of 6-I-L-mTyr was noted.
In vitro accumulation studies in rat brain
and pancreas
The differences between in vitro accumulation in the brain and in the pancreas are shown in Table 4. There was higher accumulation both in the brain and the pancreas with L-isomers than with the D-isomer. 6-I-L-mTyr was noted to have a higher uptake in the brain than 6-I-D-mTyr similar to that found in in vivo biodistribution studies.
Table 5 shows the percentage effect of control l-mTyr accumulation with inhibitors. Significant inhibition of 6-I-L-mTyr and 4-I-L-mTyr accumulation in the brain was noted with ouabain. NSD-1015 only had an inhibitory effect on the accumulation of 6-I-L-mTyr in the brain. H22/54 did not have a significant inhibition on the accumulation of radioiodinated mTyr.
In vivo inhibition studies and autoradiography
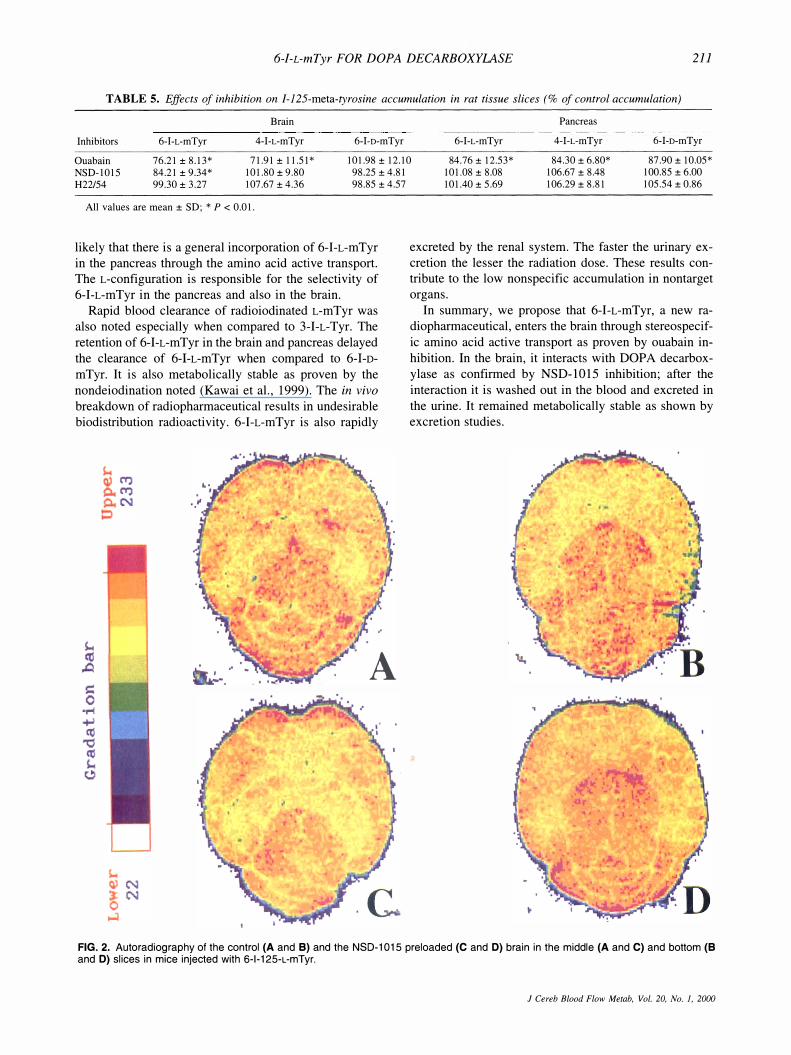
Figures 2A and 2B shows the control uptakes of 6-1-L-mTyr in the midbrain area and in the cerebellum. After preloading with NSD-1015, the uptake in the striatum which is abundant with dopaminergic neurons has decreased (Fig. 2C). In the thalamus and cerebellum, the significant effect of NSD-1015 pretreatment was not observed (Fig. 2D). H22/54 showed no significant effect mentioned above on the same portion (data not shown). These findings coincide with our results in the in vitro
studies.
TABLE 2. Blood clearance of I-l25-tyrosine derivatives in mice (% dose/g tissue)
Time (min) 6-I-L-mTyr 4-I-L-mTyr 6-I-o-mTyr 3-I-L-Tyr
2 8.90 ± 0.83 7.12±0.12 6.53 ± 0.42 11.29 ± 0.55 5 5.66 ± 0.03 4.26 ± 0.35 3.60 ± 0.42 7.29 ± 0.26
10 4.28 ± 0.13 3.50 ± 0.23 2.33 ± 0.22 7.02 ± 2.11 30 2.68 ± 0.16 1.61 ± 0.26 0.68 ± 0.16 4.63 ± 0.83
All values are mean ± SD.
J Cereb Blood Flow Metab. Vol. 20. No.1. 2000

210 L. GARC1A FLORES II ET AL.
TABLE 3. Pancreas and liver accumulation of 1-125-meta-tyrosine in mice (% doselg tissue)
Time Pancreas Liver
(min) 6-I-L-mTyr 4-I-L-mTyr 6-I-D-mTyr 6-I-L-mTyr 4-I-L-mTyr 6-I-D-mTyr
2 14.80 ± 0.61 12.04 ± 2.03 4.53 ± 0.30 9.77 ±0.37 5.83 ± 0.06 7.98 ± 1.08 5 20.02 ± 2.14 13.93 ± 0.84 3.60 ±0.30 8.87 ± 0.93 4.55 ± 0.34 6.42 ± 0.40
10 20.62 ± 2.26 12.43 ± 1.00 2.37 ± 0.27 8.80 ± 0.47 2.93 ± 0.55 5.07 ± 0.56 30 6.29 ± 0.60 4.15 ± 1.07 0.59 ± 0.15 5.80 ± 0.67 1.12 ± 0.09 2.39 ± 0.24
All values are mean ± SD.
DISCUSSION
Radioiodination is used extensively as a means of labeling compounds of medical and biologic interest because iodine radioisotopes have the advantage of availability, low cost, and relatively longer half-lives. In the chloramine-T method, chloramine-T oxidizes the iodine to a reactive iodine species which then labels the compound of interest. High labeling efficiency and very high specific activity of mTyr can be obtained by this method. In our study, the phenol ring of mTyr provided an ample site for labeling.
To produce a suitable radiotracer by the radioiodination method, the radioiodine must be attached in such a way that the physiologic properties of the compound would not be affected. It is also important that the radioiodinated compound is not broken down quickly by general metabolic pathways (Kloss and Leven, 1979).
Oldendorf and Szabo (1976) proposed the presence of specific affinity sites on carrier proteins studded through the plasma of brain capillary endothelial cells that are specific for various amino acids. Betz and Goldstein (1978) confirmed Na+-dependent neutral amino acid transport into isolated brain capillaries. In recent experiments ouabain, an efficient Na+-K+-ATPase inhibitor, has been used to suppress the Na+-dependent concentrative uptake of amino acids (Hughes and Lantos, 1989; Sanchez del Pino et aI., 1995). Because the uptake of 6-I-L-mTyr is inhibited by ouabain, it is transported into the brain by amino acid active transport similar to that of 6-I-L-DOPA (Kawai et aI., 1996). The transport system could be stereospecific because more of the inhibitory effect of ouabain was noted with 6-I-L-mTyr than with 6-I-D- mTyr, similar to the findings of our previous study (Kawai et aI., 1999).
This active amino acid transport system is shared by a radioiodinated tyrosine derivative, 3-iodo-a-methyl-Ltyrosine (I-L-AMT) which is now being clinically used to
TABLE 4. In vitro accumulation of 1-125-meta-tyrosine in the brain and the pancreas (% dose/g slice)
Organs
Brain Pancreas
6-I-L-mTyr
126.89 ± 11.16 126.98 ± 5.22
All values are mean ± SD.
4-I-L-mTyr
122.12 ± 9.07 123.59 ± 4.22
J Cereb Blood Flow Metab, Vol. 20, No. 1, 2000
6-I-D-mTyr
64.00 ± 7.03 64.36 ± 4.61
study brain tumors (Woesler et aI., 1997). Like other amino acids with the L-form, 6-I-L-mTyr also has a high accumulation in the brain and is metabolically stable similar to I-L-AMT (Kawai et aI., 1991). It is the 3-hydroxy moiety of 6-I-L-mTyr which gives it an affinity for DOPA decarboxylase.
DOPA decarboxylase may be rate-limiting for mono
amine synthesis in the human brain (Gjedde et aI., 1993). Because DOPA decarboxylase is not regulated in re
sponse to the intensity of dopaminergic neurotransmission, its activity may be a more precise indicator of the capacity of tissue to synthesize catecholamines (Gjedde et aI., 1991). 6-I-L-mTyr could occupy the active site of DOPA decarboxylase which is likened to affinity labeling (Wold, 1977). Being a marker of DOPA decarboxylase, it could facilitate the quantification of the enzyme. The 3-hydroxy moiety in 6-I-L-mTyr could account for the intracellular interaction with DOPA decarboxylase which could be substantiated by the retention of 6-I-LmTyr in the brain immediately after injection.
NSD-1015, a DOPA decarboxylase inhibitor, is frequently used in dopamine biosynthesis studies (Sved et aI., 1984; Nissbrandt and Carlsson, 1987), and it acts in the active site of enzymes. The decrease in uptake of 6-I-L-mTyr upon inhibition of NSD-lOl5 is an indication of the decrease in availability of active sites to be occupied which could mean less DOPA decarboxylase participating in the dopaminergic presynaptic biosynthesis.
The conversion of L-tyrosine to L-DOPA is catalyzed by the enzyme tyrosine hydroxylase. H22/54 has been used as a tyrosine hydroxylase inhibitor (Maj et aI., 1978; Verhofstad and Jonsson, 1983). Tyrosine hydroxylase and DOPA decarboxylase inhibitors have been used to study dopamine biosynthesis (Demarest and Moore, 1980; Duda and Moore, 1985). The 4-hydroxy group that is present in tyrosine and is lacking in 6-I-LmTyr could explain why there is no effect with H22/54.
6-I-L-mTyr is also noted to have a higher uptake in the pancreas than in the liver. The amino acid is known to accumulate in pancreatic cells and is utilized for synthesis of proteins and enzymes. Although there is dopamine synthesis and release from nonneuronal exocrine cells of the pancreas (Mezey et aI., 1996), the minimal inhibition by NSD-lOl5 could not totally account for the very high accumulation of 6-I-L-mTyr in the pancreas. It is more
6-I-L-mTyr FOR DOPA DECARBOXYLASE 2][
TABLE 5. Effects of inhibition on I-125-meta-tyrosine accumulation in rat tissue slices (% of control accumulation)
Brain Pancreas
Inhibitors 6-I-L-mTyr 4-I-L-mTyr 6-I-D-mTyr 6-I-L-mTyr 4-I-L-mTyr 6-I-D-mTyr
Ouabain 76.21 ± 8.13* 71.91 ± 11.51 * 101.98 ± 12.10 84.76 ± 12.53* 84.30 ± 6.80* 87.90 ± 10.05* NSD-I015 84.21 ± 9.34* 101.80 ± 9.80 98.25 ± 4.81 H22/54 99.30 ± 3.27 107.67 ± 4.36 98.85 ± 4.57
All values are mean ± SD; * P < 0.01.
likely that there is a general incorporation of 6-I-L-mTyr in the pancreas through the amino acid active transport. The L-configuration is responsible for the selectivity of 6-I-L-mTyr in the pancreas and also in the brain.
Rapid blood clearance of radioiodinated L-mTyr was also noted especially when compared to 3-I-L-Tyr. The retention of 6-I-L-mTyr in the brain and pancreas delayed the clearance of 6-I-L-mTyr when compared to 6-1-0-mTyr. It is also metabolically stable as proven by the nondeiodination noted (Kawai et aI., 1999). The in vivo
breakdown of radiopharmaceutical results in undesirable biodistribution radioactivity. 6-I-L-mTyr is also rapidly
A
101.08 ± 8.08 106.67 ± 8.48 100.85 ± 6.00 101.40 ± 5.69 106.29 ± 8.81 105.54 ± 0.86
excreted by the renal system. The faster the urinary excretion the lesser the radiation dose. These results contribute to the low nonspecific accumulation in nontarget organs.
In summary, we propose that 6-I-L-mTyr, a new radiopharmaceutical, enters the brain through stereospecific amino acid active transport as proven by ouabain inhibition. In the brain, it interacts with DOPA decarboxylase as confirmed by NSD-IOI5 inhibition; after the interaction it is washed out in the blood and excreted in the urine. It remained metabolically stable as shown by excretion studies.
FIG. 2. Autoradiography of the control (A and B) and the NSD-1015 preloaded (C and D) brain in the middle (A and C) and bottom (B and D) slices in mice injected with 6-1-125-L-mTyr.
J Cereb Blood Flow Metab, Vol. 20, No.1, 2000

212 L. GARCIA FLORES II ET AL.
CONCLUSION
6-Radioiodinated L-meta-tyrosine is a new radiopharmaceutical for single photon emission-computed tomography that assesses both the amino acid transport system and quantifies the metabolically active DOPA decarboxylase. It provides physiologic information about the functional status of dopaminergic presynaptic neurons.
REFERENCES
Adam MJ, Ponce YZ, Berry JM (1989) Synthesis of L-6-e23Ijiodo-mtyrosine a potential SPECT brain imaging agent. J Labelled Compounds Radiopharmacol 28: 1065-1072
Asenbaum S, Briicke T, Pirker W, Podreka I, Angelberger P, Wenger S, Wober C, Muller C, Deecke L (1997) Imaging of dopamine transporters with iodine-I 23-f3-CIT and SPECT in Parkinson's disease. J Nucl Med 38:1-6
Betz AL, Goldstein GW (1978) Polarity of the blood-brain barrier: neutral amino acid transport into isolated brain capillaries. Science 202:225-227
Choksi NY, Hussain A, Booth RG (1997) 2-Phenylaminoadenosine stimulates dopamine synthesis in rat forebrain in vitro and in vivo
via adenosine A2 receptors. Brain Res 761: 151-155 Dao-Castellana MH, Paillere-Martinot ML, Hantraye P, Attar-Levy D,
Remy P, Crouzel C, Artiges E, Feline A, Syrota A, Martinot JL (1997) Presynaptic dopaminergic function in the striatum of schizophrenic patients. Schizophr Res 23:167-174
De Jesus OT, Sunderland JJ, Nickles JR, Mukherjee J, Appelman EH (1990) Synthesis of radiofluorinated analogs of m-tyrosine as potential L-dopa tracers via direct reaction with acetylhypofluorite. Appl Radiat Isot 41:433-437.
Demarest KT, Moore KE (1980) Accumulation of L-DOPA in the median eminence: an index of tuberoinfundibular dopaminergic nerve activity. Endocrinology 106:463-468
Duda NJ, Moore KE (1985) Simultaneous determination of 5- hydroxytryptophan and 3,4-dihydroxyphenylalanine in rat brain by HPLC with electrochemical detection following electrical stimulation of the dorsal raphe nucleus. J Neurochem 44: 128-133
Garnett ES, Firnau G, <;:han PKH, Sood S, Belbeck LW (1978) [18FjFluoro-dopa, an analogue of dopa, and its use in direct external measurements of storage, degradation and turnover of intracerebral dopamine. Proc Natl Acad Sci USA 75:464-467
Gjedde A, Reith J, Dyve S, Leger G, Guttman M, Diksic M, Evans A, Kuwabara H (1991) Dopa decarboxylase activity of the living human brain. Proc Natl Acad Sci USA 88:2721-2725
Gjedde A, Leger GC, Cumming P, Yasuhara Y, Evans AC, Guttman M, Kuwabara H (1993) Striatal L-DOPA decarboxylase activity in Parkinson's disease in vivo: implication for the regulation of dopamine synthesis. J Neurochem 61:1538-1541
Hornykiewicz 0 (1982) Imbalance of monoamines and clinical disorders. Prog Brain Res 55:419-429
Hughes CCW, Lantos PL (1989) Uptake of leucine and alanine by cultured cerebral capillary endothelial cells. Brain Res 480: 126-132
Kawai K, Fujibayashi Y, Saji H, Yonekura Y, Konishi J, Kubodera A, Yokoyama A (1991) A strategy for the study of cerebral amino acid transport using iodine-123-labeled amino acid radiopharmaceutical: 3-iodo-alpha-methyl-L-tyrosine. J Nucl Med 32:819-824
Kawai K, Ohta H, Kubodera A, Channing MA, Eckelman WC (1996)
J Cereb Blood Flow Metab, Vol. 20, No.1, 2000
Synthesis and evaluation of radioiodinated 6-iodo-L-DOPA as a cerebral L-amino acid transport marker. Nucl Med Bioi 23:251-255
Kawai K, Flores LG II, Nakagawa M, Shikano N, Jinnouchi S, Tamura S, Kubodera A (1999) Brain uptake of iodinated L-meta-tyrosine, a metabolically stable amino acid derivative. Nucl Med Commun
20: 153-157 Kloss G, Leven M (1979) Accumulation of radioiodinated tyrosine
derivatives in the adrenal medulla and in melanomas. Eur J Nucl
Med 4:179-186 Luxen A, Guillaume M, Melega WP, Pike VW, Solin 0, Wagner R
(1992) Production of 6-[ 18Fjfluoro-L-DOPA and its metabolism in
vivo-a critical review. Nucl Med Bioi 19:149-158 Maj J, Mogilnicka E, Klimek V (1978) The influence of mianserin and
danitracen, 5-hydroxytryptamine receptor blockers, on the 5- hydroxytryptamine disappearance induced by H22/54 in the rat brain. Pol J Pharmacol Pharm 30:413-420
Mezey E, Eisenhofer G, Hata G, Hansson S, Gould L, Hunyady B, Hoffman BJ (1996) A novel nonneuronal catecholamine system: exocrine pancreas synthesizes and releases dopamine. Proc Natl
Acad Sci USA 93:10377-10382 Neumeyer JL, Wang S, Milius RA, Baldwin RM, Zhea-Ponce Y, Hof
fer PB, Sybirska E, AI-Tikriti M, Charney DS, Malison RT (1991) [123Ij_2f3_ carboxymethoxy-3 13-( 4-iodophenyl)tropane (f3CIT): high affinity SPECT radiotracer of monoamine reuptake sites in brain. J Med Chem 34:3144-3146.
Nissbrandt H, Carlsson A (1987) Turnover of dopamine and dopamine metabolites in rat brain: comparison between striatum and substantia nigra. J Neurochem 49:959-967
Oldendorf WH, Szabo J (1976) Amino acid assignment to one three blood-brain barrier amino acid carriers. Am J Physiol 230:94-98
Sanchez del Pino MM, Hawkins RA, Peterson DR (1995) Biochemical discrimination between luminal and abluminal enzyme and transport activities of the blood-brain barrier. J Bioi Chem 270: 14907-14912
Sawle GV, Bloomfield PM, Bjorklund A, Brooks DJ, Bmndin P, Leenders KL, Lindvall 0, Marsden CD, Rehncrona S, Widner H, Frackowiak SJ (1992) Transplantation of fetal dopamine neurons in Parkinson's disease: PET [' 8Fj6-L-fluorodopa studies in two patients with putaminal implants. Ann Neurol 31: 166-173
Srinivasan K, Awapara J (1978) Substrate specificity and other properties of DOPA decarboxylase from guinea pig kidneys. Biochim Biophys Acta 526:597-604
Sved AF, Baker HA, Reis DJ (1984) Dopamine synthesis in inbred mouse strains which differ in numbers of dopamine neurons. Brain Res 303:261-266
Tissingh G, Booij J, Winogrodzka A, van Royen EA, Wolters EC (1997) IBZM- and CIT-SPECT of the dopaminergic system in parkinsonism. J Neural Transm 50(suppl):31-37
Verhofstad AA, Jonsson G (1983) Immunohistochemical and neurochemical evidence for the presence of serotonin in the adrenal medulla of the rat. Neuroscience 10:1443-1453
Volkow ND, Fowler JS (1992) Neuropsychiatric disorder's investigation of schizophrenia and substance abuse. Semin Nucl Med 22: 254-267
Wold F (1977) Affinity labeling-an overview. Methods EnzymoI46:3-14
Woesler B, Kuwert T, Morgenroth C, Matheja P, Palkovic S, Schafers M, Vollet B, Schafers K, Lerch H, Brandau W, Samnick S, Wassman H, Schober 0 (1997) Non-invasive grading of primary brain tumours: results of a comparative study between SPET with 1231-alpha-methyl tyrosine and PET with 18F-deoxyglucose. Eur J Nucl
Med 24:428-434.
Related Documents











