A new hominoid species from the middle Miocene site of Pas xalar, Turkey Jay Kelley a, * , Peter Andrews b , Berna Alpagut c a Department of Oral Biology, College of Dentistry, University of Illinois at Chicago, 801 S. Paulina, Chicago, IL 60612, USA b Department of Vertebrate Palaeontology, Natural History Museum, London, UK c Dil ve Tarih Cografya Enstitusu, Ankara Universitesi, Sihhiye/Ankara, Turkey Received 5 July 2001; accepted 16 February 2007 Abstract A new species of fossil hominoid is described from the middle Miocene deposits at Pas xalar, Turkey. It is the less common of the two Pas xalar species discussed by Martin and Andrews (1993), making up approximately 10% of the individuals in the Pas xalar hominoid sample according to analyses of the minimum number of individuals. To the diagnostic features of I 1 described by Alpagut et al. (1990) and Martin and Andrews (1993) can now be added further diagnostic features of all the anterior teeth, as well as both upper premolars and P 3 . These include discrete, nonmetric features and metric differences at all the noted tooth positions. Attempts to distinguish the upper and lower molars of the two species have so far been unsuccessful, with the possible exception of M 3 . The morphology of the new species is similar in most respects to that of Kenyapithecus wickeri from Fort Ternan, especially concerning maxillary morphology. They share robust and moderately deep maxillary alveolar processes, a restricted maxillary sinus with an elevated and uncomplicated floor, lacking the compartmentalization evident to varying degrees in many other taxa, and a zygomatic process that originates and turns laterally fairly high above the alveolar margin. There are also a number of distinctive similarities in the dentition, particularly for I 1 ,C 1 ,P 4 and P 3 . The I 1 morphology in particular, with greatly hypertrophied lingual marginal ridges bounding a uniformly thickened basal crown area, is distinctive among Miocene hominoids. All of these similarities serve to reinforce the differences noted by others between the derived morphology of K. wickeri and the more primitive morphology of Equa- torius africanus from Maboko and Kipsaramon. The new species differs from K. wickeri in morphological details of most of the anterior and premolar teeth that are known for both species, despite the general morphological similarity, and in the size of I 1 versus I 2 . One striking feature of the new species is a relatively large incisive fossa, although it cannot be determined if this is associated with an open palatine fenestra, as in many early Miocene hominoids, or a minimally overlapping palate and nasoalveolar clivus, as in some middle and late Miocene hominoids. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Kenyapithecus; Griphopithecus; Equatorius; Miocene Hominoidea; Taxonomy; Dentition Introduction Hominoid fossils were first uncovered at the site of Pas xalar from a road cut made in the course of a lignite survey. A pre- liminary excavation at the site produced 86 isolated hominoid teeth, which were assigned to two existing species, Sivapithe- cus (now Griphopithecus) darwini and Ramapithecus (now Kenyapithecus) wickeri (Andrews and Tobien, 1977). The two species were differentiated largely by size, S. darwini be- ing the larger of the two. Further excavations commenced in 1983 and have continued to the present, resulting in the recov- ery of hundreds of additional hominoid specimens (Alpagut et al., 1990; Martin and Andrews, 1993). Alpagut et al. (1990) described many of the new specimens, including several partial maxillae and mandibles. They also recognized two spe- cies, one common and one relatively rare, which were differen- tiated by a small number of features of the I 1 and P 3 , but with only the incisor differences being well documented. In contrast * Corresponding author. Tel.: þ1 312 996 6054. E-mail addresses: [email protected] (J. Kelley), [email protected] (P. Andrews), [email protected] (B. Alpagut). 0047-2484/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.jhevol.2007.08.007 Available online at www.sciencedirect.com Journal of Human Evolution 54 (2008) 455e479

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
Journal of Human Evolution 54 (2008) 455e479
A new hominoid species from the middle Miocene site ofPasxalar, Turkey
Jay Kelley a,*, Peter Andrews b, Berna Alpagut c
a Department of Oral Biology, College of Dentistry, University of Illinois at Chicago, 801 S. Paulina, Chicago, IL 60612, USAb Department of Vertebrate Palaeontology, Natural History Museum, London, UK
c Dil ve Tarih Cografya Enstitusu, Ankara Universitesi, Sihhiye/Ankara, Turkey
Received 5 July 2001; accepted 16 February 2007
Abstract
A new species of fossil hominoid is described from the middle Miocene deposits at Pasxalar, Turkey. It is the less common of the two Pasxalarspecies discussed by Martin and Andrews (1993), making up approximately 10% of the individuals in the Pasxalar hominoid sample according toanalyses of the minimum number of individuals. To the diagnostic features of I1 described by Alpagut et al. (1990) and Martin and Andrews(1993) can now be added further diagnostic features of all the anterior teeth, as well as both upper premolars and P3. These include discrete,nonmetric features and metric differences at all the noted tooth positions. Attempts to distinguish the upper and lower molars of the two specieshave so far been unsuccessful, with the possible exception of M3. The morphology of the new species is similar in most respects to that ofKenyapithecus wickeri from Fort Ternan, especially concerning maxillary morphology. They share robust and moderately deep maxillaryalveolar processes, a restricted maxillary sinus with an elevated and uncomplicated floor, lacking the compartmentalization evident to varyingdegrees in many other taxa, and a zygomatic process that originates and turns laterally fairly high above the alveolar margin. There are alsoa number of distinctive similarities in the dentition, particularly for I1, C1, P4 and P3. The I1 morphology in particular, with greatly hypertrophiedlingual marginal ridges bounding a uniformly thickened basal crown area, is distinctive among Miocene hominoids. All of these similaritiesserve to reinforce the differences noted by others between the derived morphology of K. wickeri and the more primitive morphology of Equa-torius africanus from Maboko and Kipsaramon. The new species differs from K. wickeri in morphological details of most of the anterior andpremolar teeth that are known for both species, despite the general morphological similarity, and in the size of I1 versus I2. One striking featureof the new species is a relatively large incisive fossa, although it cannot be determined if this is associated with an open palatine fenestra, as inmany early Miocene hominoids, or a minimally overlapping palate and nasoalveolar clivus, as in some middle and late Miocene hominoids.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Kenyapithecus; Griphopithecus; Equatorius; Miocene Hominoidea; Taxonomy; Dentition
Introduction
Hominoid fossils were first uncovered at the site of Pasxalarfrom a road cut made in the course of a lignite survey. A pre-liminary excavation at the site produced 86 isolated hominoidteeth, which were assigned to two existing species, Sivapithe-cus (now Griphopithecus) darwini and Ramapithecus (now
* Corresponding author. Tel.: þ1 312 996 6054.
E-mail addresses: [email protected] (J. Kelley), [email protected]
(P. Andrews), [email protected] (B. Alpagut).
0047-2484/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jhevol.2007.08.007
Kenyapithecus) wickeri (Andrews and Tobien, 1977). Thetwo species were differentiated largely by size, S. darwini be-ing the larger of the two. Further excavations commenced in1983 and have continued to the present, resulting in the recov-ery of hundreds of additional hominoid specimens (Alpagutet al., 1990; Martin and Andrews, 1993). Alpagut et al.(1990) described many of the new specimens, including severalpartial maxillae and mandibles. They also recognized two spe-cies, one common and one relatively rare, which were differen-tiated by a small number of features of the I1 and P3, but withonly the incisor differences being well documented. In contrast

456 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
to the prior sample sorting, the two species, as defined morpho-logically at these two tooth positions, appeared to be roughlythe same size. Alpagut et al. assigned the more common ofthe two species to S. darwini, while the second remained un-named, but it was also considered a likely member of Sivapithe-cus. Martin and Andrews (1993) later transferred the commonspecies to Griphopithecus alpani, known from the approxi-mately contemporaneous site of Candir in central Turkey (Tek-kaya, 1974; Begun et al., 2003; Gulec and Begun, 2003). Theyagain declined to name a second species because so few spec-imens could be attributed to it, and many of these with less thancomplete confidence, making a useful diagnosis impossible.
The case for two hominoid species at Pasxalar gained furthersupport from sex assignments of some of the lower canines inthe sample (Waddle et al., 1995). Waddle et al. purported toshow two distinct size groups among lower canines identifiedas female and suggested that this could only result from thepresence of two species. Kelley and Alpagut (1999) subse-quently re-evaluated the entire Pasxalar lower canine sampleand demonstrated that, by ignoring extensive wear on somespecimens, Waddle et al. had mistakenly included several prob-able male canines among the females. Reanalysis using onlyunworn or minimally worn canines resulted in single size clus-ters for both the male and female canine samples. Kelley andAlpagut noted, however, that this result does not necessarilydemonstrate that only one species is present in the Pasxalar sam-ple, only that canine sexing cannot be used to argue for two.
Thus, support for the presence of two hominoid species atPasxalar has remained tenuous. Moreover, even if correct, theproposal has been of no practical utility because only a handfulof teeth from the hundreds available have been identified asbelonging to a second species, which is too limited to permita useful species diagnosis or confident assignment of the spe-cies to genus.
The greatest difficulty in documenting the presence of a sec-ond species in the Pasxalar hominoid sample has been that thesample consists almost entirely of isolated teeth. To diagnosea new species and construct a species hypodigm, it is thereforenecessary not only to demonstrate discrete morphologieswithin each tooth type, but also to justify the association ofmorphologies between the different tooth types; that is, todemonstrate why a particular morph for I1, for example, is as-sociated with a particular morph for I2, and so on through thedentition. In the first part of this paper, we report the results ofthese two exercises for the anterior teeth (incisors, canines)and premolars (except for P4), so far the only teeth that wehave been able to sort with confidence. There has been someprogress in sorting the lower third molars as well, but the re-sults are still tentative (discussed further below). Althoughwe have only been able to sort the antemolar teeth, these aresufficient to characterize and diagnose the two hominoid spe-cies present at Pasxalar.
Sorting the antemolar teeth
The following morphological characterizations are not in-tended to be exhaustive descriptions of the anterior and
premolar teeth, which have been described in detail elsewhere(Alpagut et al., 1990; Martin and Andrews, 1993; Alpagutet al. in particular provided photographs that document thedental morphological variation present in the Pasxalar sample).The focus here is rather on aspects of variation by which dis-tinct morphologies can be recognized within each tooth type,and by which the different morphs of each tooth type can beassociated.
All descriptions and identifications are based on observa-tions of original specimens. For each tooth type, the teethwere first segregated according to overall morphology. Formost tooth types, this resulted in two distinct morphs. Fortwo tooth types, however, upper lateral incisors and female up-per canines, sorting was less definitive (see below). The twomorphs at each tooth position were then compared metricallyusing bivariate plots of mesiodistal length and buccolingual orlabiolingual breadth and, for the anterior teeth, histograms ofcrown height. Crown heights were only measured on unwornor minimally worn teeth as maximum labial (incisors) or buc-cal (canines) height. Metric differentiation of the two morphswas checked using two sets of measurements, one made dur-ing this analysis by JK and another made by PA prior to thesorting. In no case did the metric patterning of the two morphsdiffer between the two sets of measurements.
For some tooth types, heavy wear or breakage precluded as-signing some teeth to a particular morphological group. Thus,individual sample sizes of the different morphs, as well asoverall sample sizes, in some cases vary substantially betweenthe different tooth types. Variation in sample size is also due inpart to the expected variation resulting from the vagaries ofpreservation and recovery. For most tooth types, however,wear and breakage presented few problems for reliable assign-ment due to the nature of the particular diagnostic features.Wear and breakage did, nevertheless, sometimes preclude tak-ing measurements from teeth that could be assigned to a mor-phological group. Thus, the numbers of individuals in metricanalyses do not necessarily equal the numbers identified as be-longing to a particular morphology. Measurements of all thePasxalar teeth are available from either JK or PA upon request.
Upper central incisor
Morph I (n¼ 10). The morphology of these incisors is strik-ingly uniform. The lingual surface is characterized by greatlyhypertrophied mesial and distal marginal ridges, especiallythe mesial ridge (Fig. 1; see also Alpagut et al., 1990: Fig. 3[specimen mislabeled as BP32; actual number is BP27]; Mar-tin and Andrews, 1993: Fig. 1). The central area between themarginal ridges thickens cervically, generally following the re-lief of the marginal ridges, and is divided into several wide,more or less longitudinally oriented crenulations. The marginalridges are generally set off from the central area by very narrowfoveae incisallydwhich are only slightly more pronouncedthan the grooves separating the central crenulationsdand bytheir slightly greater relief cervically. On some teeth, one ortwo of the crenulations encroach on the distal marginal ridge,interrupting the continuity of the ridge. The mesial and distal
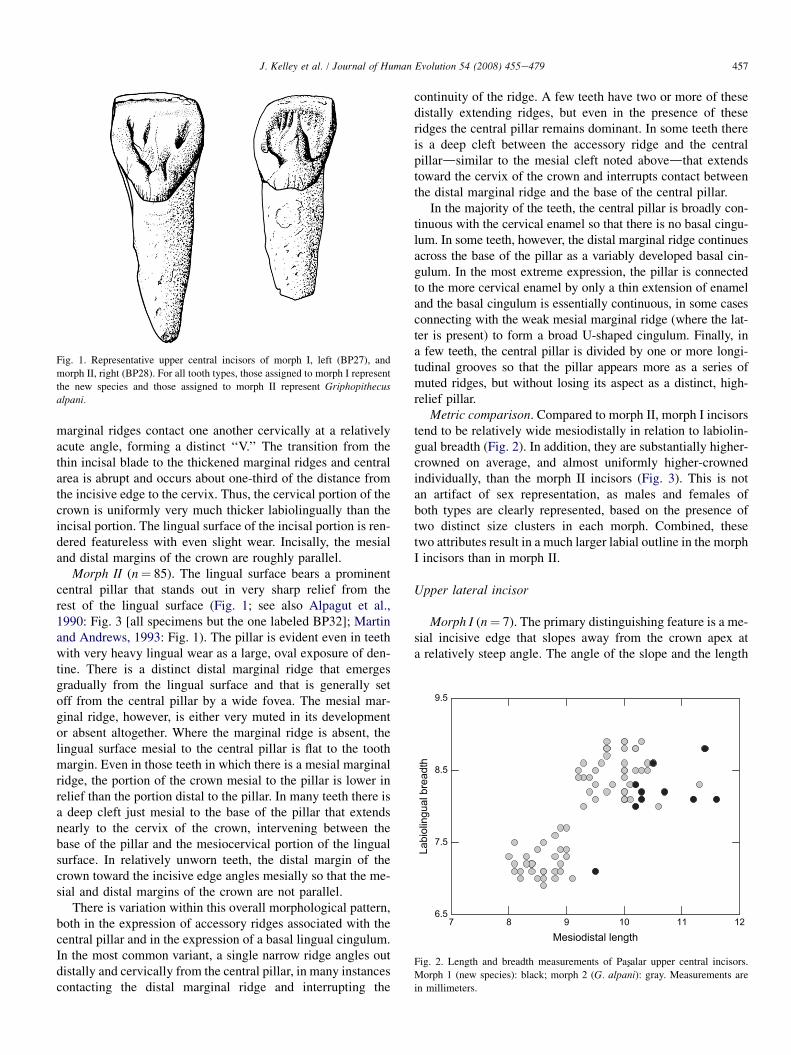
Fig. 1. Representative upper central incisors of morph I, left (BP27), and
morph II, right (BP28). For all tooth types, those assigned to morph I represent
the new species and those assigned to morph II represent Griphopithecus
alpani.
7 8 9 10 11 12Mesiodistal length
6.5
7.5
8.5
9.5
Labi
olin
gual
bre
adth
Fig. 2. Length and breadth measurements of Pasxalar upper central incisors.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are
in millimeters.
457J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
marginal ridges contact one another cervically at a relativelyacute angle, forming a distinct ‘‘V.’’ The transition from thethin incisal blade to the thickened marginal ridges and centralarea is abrupt and occurs about one-third of the distance fromthe incisive edge to the cervix. Thus, the cervical portion of thecrown is uniformly very much thicker labiolingually than theincisal portion. The lingual surface of the incisal portion is ren-dered featureless with even slight wear. Incisally, the mesialand distal margins of the crown are roughly parallel.
Morph II (n¼ 85). The lingual surface bears a prominentcentral pillar that stands out in very sharp relief from therest of the lingual surface (Fig. 1; see also Alpagut et al.,1990: Fig. 3 [all specimens but the one labeled BP32]; Martinand Andrews, 1993: Fig. 1). The pillar is evident even in teethwith very heavy lingual wear as a large, oval exposure of den-tine. There is a distinct distal marginal ridge that emergesgradually from the lingual surface and that is generally setoff from the central pillar by a wide fovea. The mesial mar-ginal ridge, however, is either very muted in its developmentor absent altogether. Where the marginal ridge is absent, thelingual surface mesial to the central pillar is flat to the toothmargin. Even in those teeth in which there is a mesial marginalridge, the portion of the crown mesial to the pillar is lower inrelief than the portion distal to the pillar. In many teeth there isa deep cleft just mesial to the base of the pillar that extendsnearly to the cervix of the crown, intervening between thebase of the pillar and the mesiocervical portion of the lingualsurface. In relatively unworn teeth, the distal margin of thecrown toward the incisive edge angles mesially so that the me-sial and distal margins of the crown are not parallel.
There is variation within this overall morphological pattern,both in the expression of accessory ridges associated with thecentral pillar and in the expression of a basal lingual cingulum.In the most common variant, a single narrow ridge angles outdistally and cervically from the central pillar, in many instancescontacting the distal marginal ridge and interrupting the
continuity of the ridge. A few teeth have two or more of thesedistally extending ridges, but even in the presence of theseridges the central pillar remains dominant. In some teeth thereis a deep cleft between the accessory ridge and the centralpillardsimilar to the mesial cleft noted abovedthat extendstoward the cervix of the crown and interrupts contact betweenthe distal marginal ridge and the base of the central pillar.
In the majority of the teeth, the central pillar is broadly con-tinuous with the cervical enamel so that there is no basal cingu-lum. In some teeth, however, the distal marginal ridge continuesacross the base of the pillar as a variably developed basal cin-gulum. In the most extreme expression, the pillar is connectedto the more cervical enamel by only a thin extension of enameland the basal cingulum is essentially continuous, in some casesconnecting with the weak mesial marginal ridge (where the lat-ter is present) to form a broad U-shaped cingulum. Finally, ina few teeth, the central pillar is divided by one or more longi-tudinal grooves so that the pillar appears more as a series ofmuted ridges, but without losing its aspect as a distinct, high-relief pillar.
Metric comparison. Compared to morph II, morph I incisorstend to be relatively wide mesiodistally in relation to labiolin-gual breadth (Fig. 2). In addition, they are substantially higher-crowned on average, and almost uniformly higher-crownedindividually, than the morph II incisors (Fig. 3). This is notan artifact of sex representation, as males and females ofboth types are clearly represented, based on the presence oftwo distinct size clusters in each morph. Combined, thesetwo attributes result in a much larger labial outline in the morphI incisors than in morph II.
Upper lateral incisor
Morph I (n¼ 7). The primary distinguishing feature is a me-sial incisive edge that slopes away from the crown apex ata relatively steep angle. The angle of the slope and the length
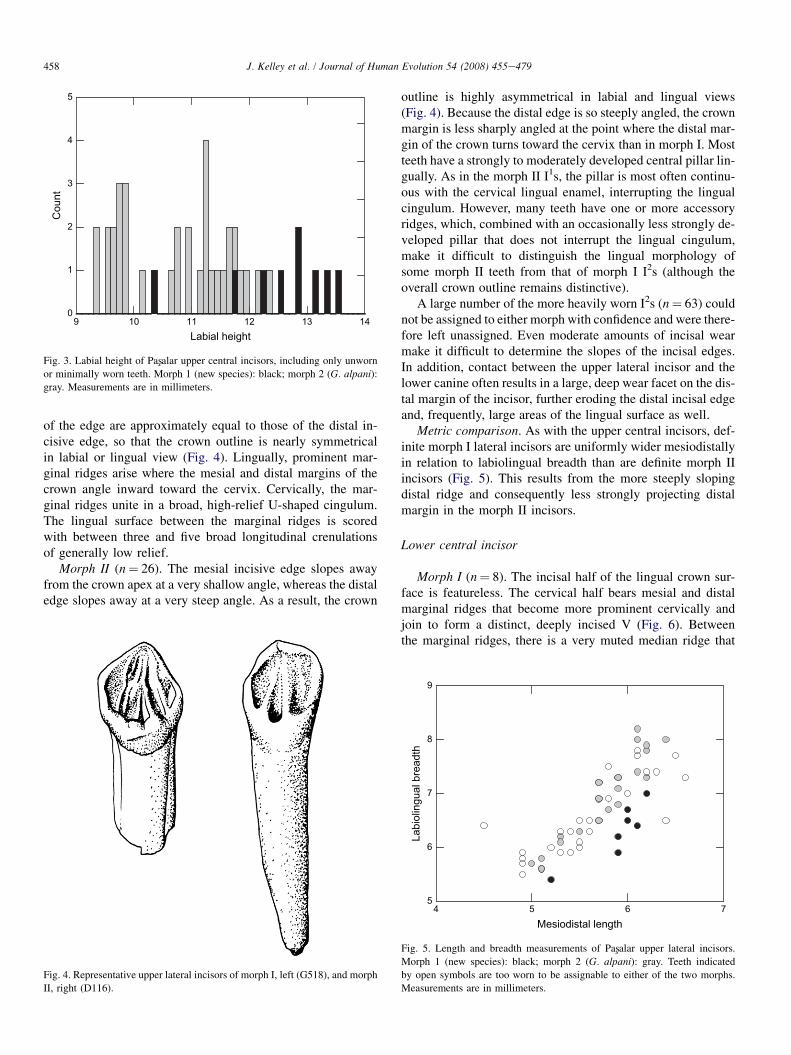
9 10 11 12 13 14Labial height
0
1
2
3
4
5
Cou
nt
Fig. 3. Labial height of Pasxalar upper central incisors, including only unworn
or minimally worn teeth. Morph 1 (new species): black; morph 2 (G. alpani):gray. Measurements are in millimeters.
458 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
of the edge are approximately equal to those of the distal in-cisive edge, so that the crown outline is nearly symmetricalin labial or lingual view (Fig. 4). Lingually, prominent mar-ginal ridges arise where the mesial and distal margins of thecrown angle inward toward the cervix. Cervically, the mar-ginal ridges unite in a broad, high-relief U-shaped cingulum.The lingual surface between the marginal ridges is scoredwith between three and five broad longitudinal crenulationsof generally low relief.
Morph II (n¼ 26). The mesial incisive edge slopes awayfrom the crown apex at a very shallow angle, whereas the distaledge slopes away at a very steep angle. As a result, the crown
Fig. 4. Representative upper lateral incisors of morph I, left (G518), and morph
II, right (D116).
outline is highly asymmetrical in labial and lingual views(Fig. 4). Because the distal edge is so steeply angled, the crownmargin is less sharply angled at the point where the distal mar-gin of the crown turns toward the cervix than in morph I. Mostteeth have a strongly to moderately developed central pillar lin-gually. As in the morph II I1s, the pillar is most often continu-ous with the cervical lingual enamel, interrupting the lingualcingulum. However, many teeth have one or more accessoryridges, which, combined with an occasionally less strongly de-veloped pillar that does not interrupt the lingual cingulum,make it difficult to distinguish the lingual morphology ofsome morph II teeth from that of morph I I2s (although theoverall crown outline remains distinctive).
A large number of the more heavily worn I2s (n¼ 63) couldnot be assigned to either morph with confidence and were there-fore left unassigned. Even moderate amounts of incisal wearmake it difficult to determine the slopes of the incisal edges.In addition, contact between the upper lateral incisor and thelower canine often results in a large, deep wear facet on the dis-tal margin of the incisor, further eroding the distal incisal edgeand, frequently, large areas of the lingual surface as well.
Metric comparison. As with the upper central incisors, def-inite morph I lateral incisors are uniformly wider mesiodistallyin relation to labiolingual breadth than are definite morph IIincisors (Fig. 5). This results from the more steeply slopingdistal ridge and consequently less strongly projecting distalmargin in the morph II incisors.
Lower central incisor
Morph I (n¼ 8). The incisal half of the lingual crown sur-face is featureless. The cervical half bears mesial and distalmarginal ridges that become more prominent cervically andjoin to form a distinct, deeply incised V (Fig. 6). Betweenthe marginal ridges, there is a very muted median ridge that
4 5 6 7Mesiodistal length
5
6
7
8
9
Labi
olin
gual
bre
adth
Fig. 5. Length and breadth measurements of Pasxalar upper lateral incisors.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Teeth indicated
by open symbols are too worn to be assignable to either of the two morphs.
Measurements are in millimeters.
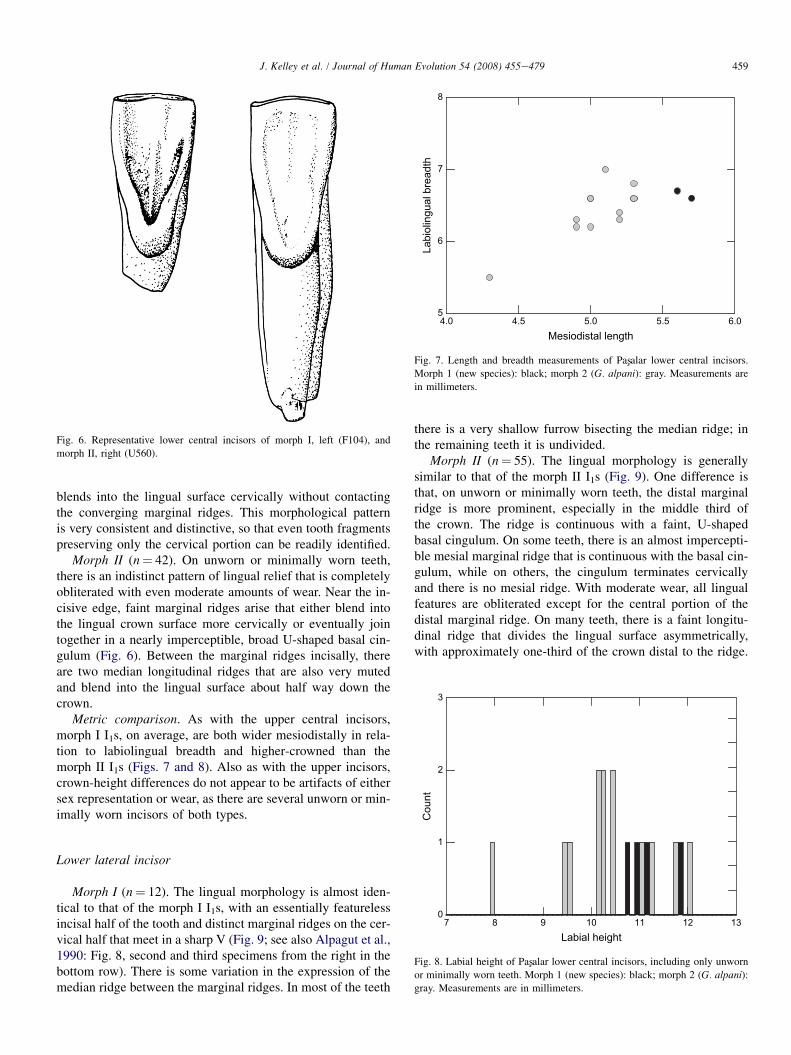
Fig. 6. Representative lower central incisors of morph I, left (F104), and
morph II, right (U560).
4.0 4.5 5.0 5.5 6.0Mesiodistal length
5
6
7
8
Labi
olin
gual
bre
adth
Fig. 7. Length and breadth measurements of Pasxalar lower central incisors.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are
in millimeters.
2
3
Cou
nt
459J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
blends into the lingual surface cervically without contactingthe converging marginal ridges. This morphological patternis very consistent and distinctive, so that even tooth fragmentspreserving only the cervical portion can be readily identified.
Morph II (n¼ 42). On unworn or minimally worn teeth,there is an indistinct pattern of lingual relief that is completelyobliterated with even moderate amounts of wear. Near the in-cisive edge, faint marginal ridges arise that either blend intothe lingual crown surface more cervically or eventually jointogether in a nearly imperceptible, broad U-shaped basal cin-gulum (Fig. 6). Between the marginal ridges incisally, thereare two median longitudinal ridges that are also very mutedand blend into the lingual surface about half way down thecrown.
Metric comparison. As with the upper central incisors,morph I I1s, on average, are both wider mesiodistally in rela-tion to labiolingual breadth and higher-crowned than themorph II I1s (Figs. 7 and 8). Also as with the upper incisors,crown-height differences do not appear to be artifacts of eithersex representation or wear, as there are several unworn or min-imally worn incisors of both types.
1
Lower lateral incisor
7 8 9 10 11 12 13Labial height
0
Fig. 8. Labial height of Pasxalar lower central incisors, including only unworn
or minimally worn teeth. Morph 1 (new species): black; morph 2 (G. alpani):gray. Measurements are in millimeters.
Morph I (n¼ 12). The lingual morphology is almost iden-tical to that of the morph I I1s, with an essentially featurelessincisal half of the tooth and distinct marginal ridges on the cer-vical half that meet in a sharp V (Fig. 9; see also Alpagut et al.,1990: Fig. 8, second and third specimens from the right in thebottom row). There is some variation in the expression of themedian ridge between the marginal ridges. In most of the teeth
there is a very shallow furrow bisecting the median ridge; inthe remaining teeth it is undivided.
Morph II (n¼ 55). The lingual morphology is generallysimilar to that of the morph II I1s (Fig. 9). One difference isthat, on unworn or minimally worn teeth, the distal marginalridge is more prominent, especially in the middle third ofthe crown. The ridge is continuous with a faint, U-shapedbasal cingulum. On some teeth, there is an almost impercepti-ble mesial marginal ridge that is continuous with the basal cin-gulum, while on others, the cingulum terminates cervicallyand there is no mesial ridge. With moderate wear, all lingualfeatures are obliterated except for the central portion of thedistal marginal ridge. On many teeth, there is a faint longitu-dinal ridge that divides the lingual surface asymmetrically,with approximately one-third of the crown distal to the ridge.
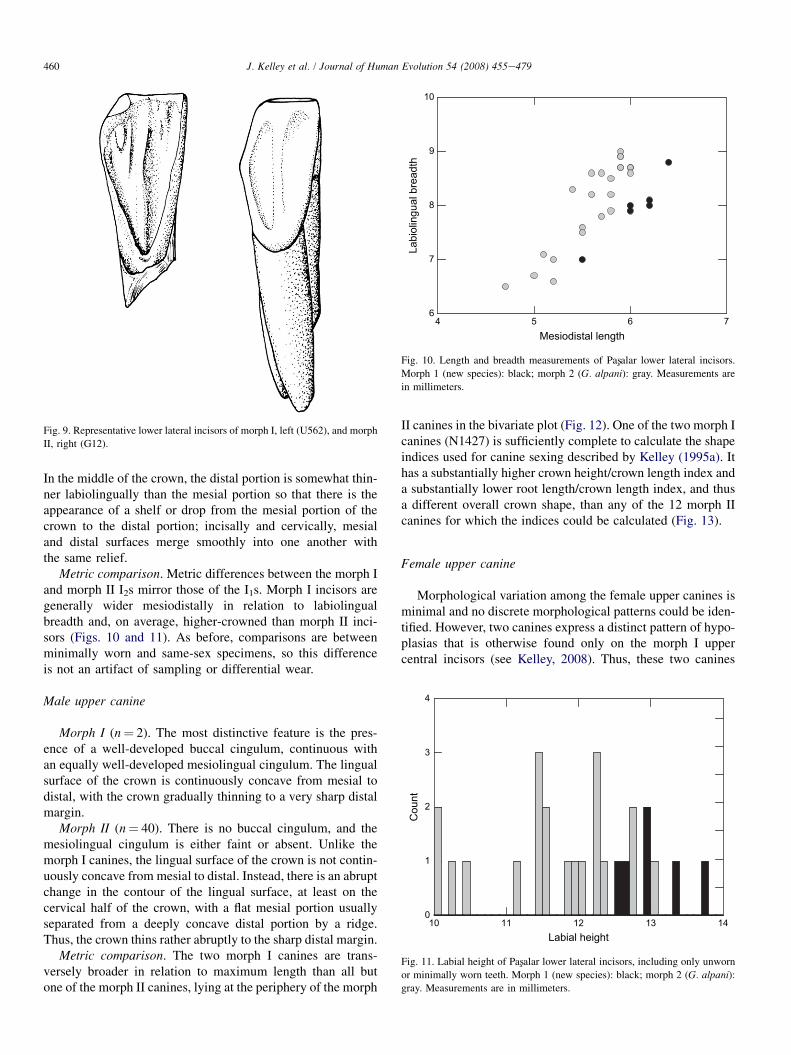
Fig. 9. Representative lower lateral incisors of morph I, left (U562), and morph
II, right (G12).
4 5 6 7Mesiodistal length
6
7
8
9
10
Labi
olin
gual
bre
adth
Fig. 10. Length and breadth measurements of Pasxalar lower lateral incisors.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are
in millimeters.
460 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
In the middle of the crown, the distal portion is somewhat thin-ner labiolingually than the mesial portion so that there is theappearance of a shelf or drop from the mesial portion of thecrown to the distal portion; incisally and cervically, mesialand distal surfaces merge smoothly into one another withthe same relief.
Metric comparison. Metric differences between the morph Iand morph II I2s mirror those of the I1s. Morph I incisors aregenerally wider mesiodistally in relation to labiolingualbreadth and, on average, higher-crowned than morph II inci-sors (Figs. 10 and 11). As before, comparisons are betweenminimally worn and same-sex specimens, so this differenceis not an artifact of sampling or differential wear.
4
Male upper canine10 11 12 13 14Labial height
0
1
2
3
Cou
nt
Fig. 11. Labial height of Pasxalar lower lateral incisors, including only unworn
or minimally worn teeth. Morph 1 (new species): black; morph 2 (G. alpani):gray. Measurements are in millimeters.
Morph I (n¼ 2). The most distinctive feature is the pres-ence of a well-developed buccal cingulum, continuous withan equally well-developed mesiolingual cingulum. The lingualsurface of the crown is continuously concave from mesial todistal, with the crown gradually thinning to a very sharp distalmargin.
Morph II (n¼ 40). There is no buccal cingulum, and themesiolingual cingulum is either faint or absent. Unlike themorph I canines, the lingual surface of the crown is not contin-uously concave from mesial to distal. Instead, there is an abruptchange in the contour of the lingual surface, at least on thecervical half of the crown, with a flat mesial portion usuallyseparated from a deeply concave distal portion by a ridge.Thus, the crown thins rather abruptly to the sharp distal margin.
Metric comparison. The two morph I canines are trans-versely broader in relation to maximum length than all butone of the morph II canines, lying at the periphery of the morph
II canines in the bivariate plot (Fig. 12). One of the two morph Icanines (N1427) is sufficiently complete to calculate the shapeindices used for canine sexing described by Kelley (1995a). Ithas a substantially higher crown height/crown length index anda substantially lower root length/crown length index, and thusa different overall crown shape, than any of the 12 morph IIcanines for which the indices could be calculated (Fig. 13).
Female upper canine
Morphological variation among the female upper canines isminimal and no discrete morphological patterns could be iden-tified. However, two canines express a distinct pattern of hypo-plasias that is otherwise found only on the morph I uppercentral incisors (see Kelley, 2008). Thus, these two canines
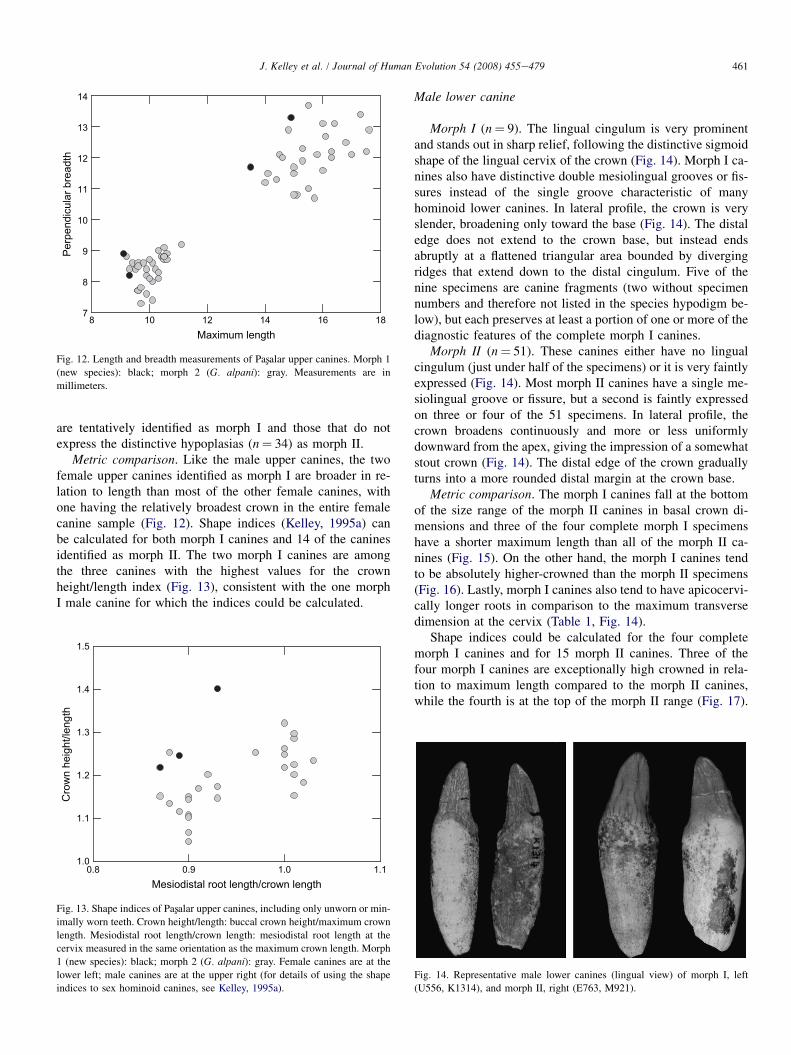
8 10 12 14 16 18Maximum length
7
8
9
10
11
12
13
14
Perp
endi
cula
r bre
adth
Fig. 12. Length and breadth measurements of Pasxalar upper canines. Morph 1
(new species): black; morph 2 (G. alpani): gray. Measurements are in
millimeters.
461J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
are tentatively identified as morph I and those that do notexpress the distinctive hypoplasias (n¼ 34) as morph II.
Metric comparison. Like the male upper canines, the twofemale upper canines identified as morph I are broader in re-lation to length than most of the other female canines, withone having the relatively broadest crown in the entire femalecanine sample (Fig. 12). Shape indices (Kelley, 1995a) canbe calculated for both morph I canines and 14 of the caninesidentified as morph II. The two morph I canines are amongthe three canines with the highest values for the crownheight/length index (Fig. 13), consistent with the one morphI male canine for which the indices could be calculated.
0.8 0.9 1.0 1.1Mesiodistal root length/crown length
1.0
1.1
1.2
1.3
1.4
1.5
Cro
wn
heig
ht/le
ngth
Fig. 13. Shape indices of Pasxalar upper canines, including only unworn or min-
imally worn teeth. Crown height/length: buccal crown height/maximum crown
length. Mesiodistal root length/crown length: mesiodistal root length at the
cervix measured in the same orientation as the maximum crown length. Morph
1 (new species): black; morph 2 (G. alpani): gray. Female canines are at the
lower left; male canines are at the upper right (for details of using the shape
indices to sex hominoid canines, see Kelley, 1995a).
Male lower canine
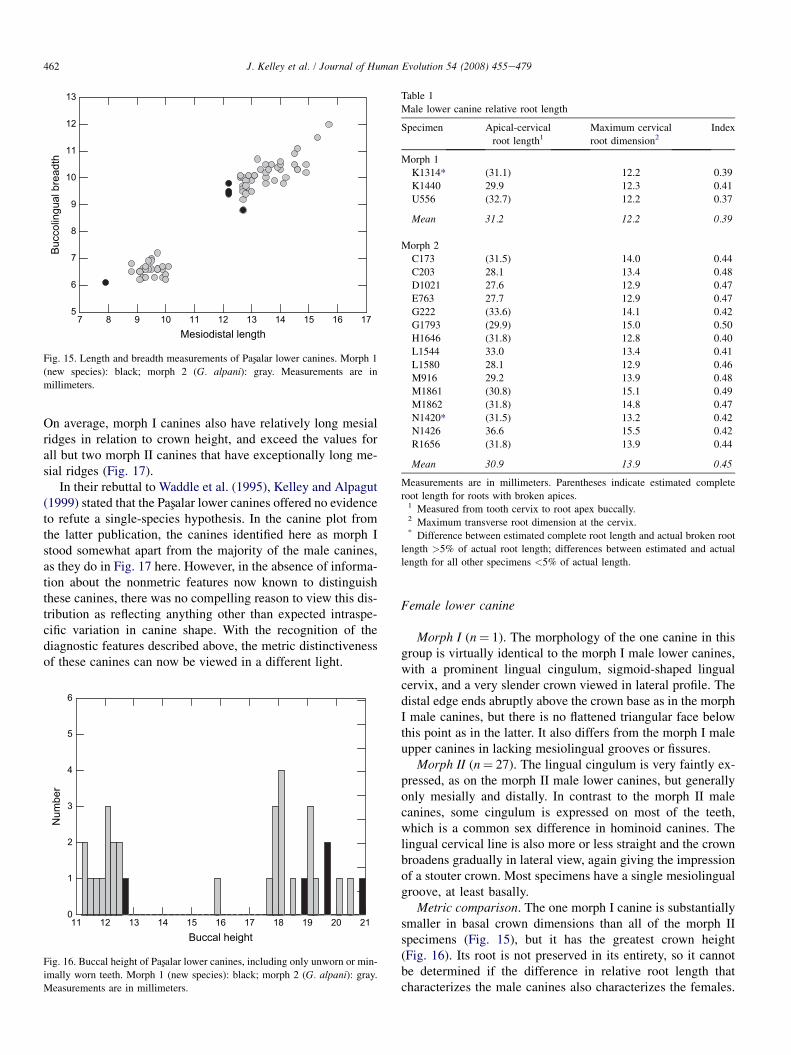
Morph I (n¼ 9). The lingual cingulum is very prominentand stands out in sharp relief, following the distinctive sigmoidshape of the lingual cervix of the crown (Fig. 14). Morph I ca-nines also have distinctive double mesiolingual grooves or fis-sures instead of the single groove characteristic of manyhominoid lower canines. In lateral profile, the crown is veryslender, broadening only toward the base (Fig. 14). The distaledge does not extend to the crown base, but instead endsabruptly at a flattened triangular area bounded by divergingridges that extend down to the distal cingulum. Five of thenine specimens are canine fragments (two without specimennumbers and therefore not listed in the species hypodigm be-low), but each preserves at least a portion of one or more of thediagnostic features of the complete morph I canines.
Morph II (n¼ 51). These canines either have no lingualcingulum (just under half of the specimens) or it is very faintlyexpressed (Fig. 14). Most morph II canines have a single me-siolingual groove or fissure, but a second is faintly expressedon three or four of the 51 specimens. In lateral profile, thecrown broadens continuously and more or less uniformlydownward from the apex, giving the impression of a somewhatstout crown (Fig. 14). The distal edge of the crown graduallyturns into a more rounded distal margin at the crown base.
Metric comparison. The morph I canines fall at the bottomof the size range of the morph II canines in basal crown di-mensions and three of the four complete morph I specimenshave a shorter maximum length than all of the morph II ca-nines (Fig. 15). On the other hand, the morph I canines tendto be absolutely higher-crowned than the morph II specimens(Fig. 16). Lastly, morph I canines also tend to have apicocervi-cally longer roots in comparison to the maximum transversedimension at the cervix (Table 1, Fig. 14).
Shape indices could be calculated for the four completemorph I canines and for 15 morph II canines. Three of thefour morph I canines are exceptionally high crowned in rela-tion to maximum length compared to the morph II canines,while the fourth is at the top of the morph II range (Fig. 17).
Fig. 14. Representative male lower canines (lingual view) of morph I, left
(U556, K1314), and morph II, right (E763, M921).
7 8 9 10 11 12 13 14 15 16 17Mesiodistal length
5
6
7
8
9
10
11
12
13
Bucc
olin
gual
bre
adth
Fig. 15. Length and breadth measurements of Pasxalar lower canines. Morph 1
(new species): black; morph 2 (G. alpani): gray. Measurements are in
millimeters.
Table 1
Male lower canine relative root length
Specimen Apical-cervical
root length1Maximum cervical
root dimension2Index
Morph 1
K1314* (31.1) 12.2 0.39
K1440 29.9 12.3 0.41
U556 (32.7) 12.2 0.37
Mean 31.2 12.2 0.39
Morph 2
C173 (31.5) 14.0 0.44
C203 28.1 13.4 0.48
D1021 27.6 12.9 0.47
E763 27.7 12.9 0.47
G222 (33.6) 14.1 0.42
G1793 (29.9) 15.0 0.50
H1646 (31.8) 12.8 0.40
L1544 33.0 13.4 0.41
L1580 28.1 12.9 0.46
M916 29.2 13.9 0.48
M1861 (30.8) 15.1 0.49
M1862 (31.8) 14.8 0.47
N1420* (31.5) 13.2 0.42
N1426 36.6 15.5 0.42
R1656 (31.8) 13.9 0.44
Mean 30.9 13.9 0.45
Measurements are in millimeters. Parentheses indicate estimated complete
root length for roots with broken apices.1 Measured from tooth cervix to root apex buccally.2 Maximum transverse root dimension at the cervix.* Difference between estimated complete root length and actual broken root
length >5% of actual root length; differences between estimated and actual
length for all other specimens <5% of actual length.
462 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
On average, morph I canines also have relatively long mesialridges in relation to crown height, and exceed the values forall but two morph II canines that have exceptionally long me-sial ridges (Fig. 17).
In their rebuttal to Waddle et al. (1995), Kelley and Alpagut(1999) stated that the Pasxalar lower canines offered no evidenceto refute a single-species hypothesis. In the canine plot fromthe latter publication, the canines identified here as morph Istood somewhat apart from the majority of the male canines,as they do in Fig. 17 here. However, in the absence of informa-tion about the nonmetric features now known to distinguishthese canines, there was no compelling reason to view this dis-tribution as reflecting anything other than expected intraspe-cific variation in canine shape. With the recognition of thediagnostic features described above, the metric distinctivenessof these canines can now be viewed in a different light.
11 12 13 14 15 16 17 18 19 20 21Buccal height
0
1
2
3
4
5
6
Num
ber
Fig. 16. Buccal height of Pasxalar lower canines, including only unworn or min-
imally worn teeth. Morph 1 (new species): black; morph 2 (G. alpani): gray.
Measurements are in millimeters.
Female lower canine
Morph I (n¼ 1). The morphology of the one canine in thisgroup is virtually identical to the morph I male lower canines,with a prominent lingual cingulum, sigmoid-shaped lingualcervix, and a very slender crown viewed in lateral profile. Thedistal edge ends abruptly above the crown base as in the morphI male canines, but there is no flattened triangular face belowthis point as in the latter. It also differs from the morph I maleupper canines in lacking mesiolingual grooves or fissures.
Morph II (n¼ 27). The lingual cingulum is very faintly ex-pressed, as on the morph II male lower canines, but generallyonly mesially and distally. In contrast to the morph II malecanines, some cingulum is expressed on most of the teeth,which is a common sex difference in hominoid canines. Thelingual cervical line is also more or less straight and the crownbroadens gradually in lateral view, again giving the impressionof a stouter crown. Most specimens have a single mesiolingualgroove, at least basally.
Metric comparison. The one morph I canine is substantiallysmaller in basal crown dimensions than all of the morph IIspecimens (Fig. 15), but it has the greatest crown height(Fig. 16). Its root is not preserved in its entirety, so it cannotbe determined if the difference in relative root length thatcharacterizes the male canines also characterizes the females.
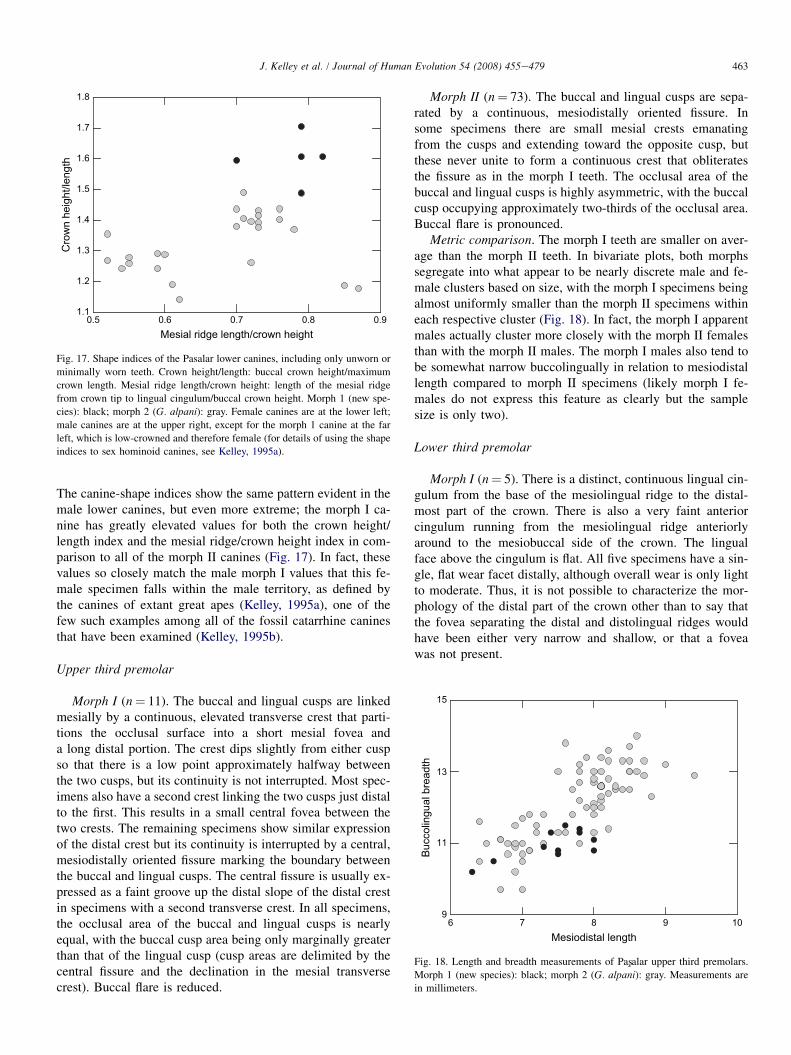
0.5 0.6 0.7 0.8 0.9Mesial ridge length/crown height
1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
Cro
wn
heig
ht/le
ngth
Fig. 17. Shape indices of the Pasalar lower canines, including only unworn or
minimally worn teeth. Crown height/length: buccal crown height/maximum
crown length. Mesial ridge length/crown height: length of the mesial ridge
from crown tip to lingual cingulum/buccal crown height. Morph 1 (new spe-
cies): black; morph 2 (G. alpani): gray. Female canines are at the lower left;
male canines are at the upper right, except for the morph 1 canine at the far
left, which is low-crowned and therefore female (for details of using the shape
indices to sex hominoid canines, see Kelley, 1995a).
463J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
The canine-shape indices show the same pattern evident in themale lower canines, but even more extreme; the morph I ca-nine has greatly elevated values for both the crown height/length index and the mesial ridge/crown height index in com-parison to all of the morph II canines (Fig. 17). In fact, thesevalues so closely match the male morph I values that this fe-male specimen falls within the male territory, as defined bythe canines of extant great apes (Kelley, 1995a), one of thefew such examples among all of the fossil catarrhine caninesthat have been examined (Kelley, 1995b).
Upper third premolar
6 7 8 9 10Mesiodistal length
9
11
13
15
Bucc
olin
gual
bre
adth
Fig. 18. Length and breadth measurements of Pasxalar upper third premolars.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are
in millimeters.
Morph I (n¼ 11). The buccal and lingual cusps are linkedmesially by a continuous, elevated transverse crest that parti-tions the occlusal surface into a short mesial fovea anda long distal portion. The crest dips slightly from either cuspso that there is a low point approximately halfway betweenthe two cusps, but its continuity is not interrupted. Most spec-imens also have a second crest linking the two cusps just distalto the first. This results in a small central fovea between thetwo crests. The remaining specimens show similar expressionof the distal crest but its continuity is interrupted by a central,mesiodistally oriented fissure marking the boundary betweenthe buccal and lingual cusps. The central fissure is usually ex-pressed as a faint groove up the distal slope of the distal crestin specimens with a second transverse crest. In all specimens,the occlusal area of the buccal and lingual cusps is nearlyequal, with the buccal cusp area being only marginally greaterthan that of the lingual cusp (cusp areas are delimited by thecentral fissure and the declination in the mesial transversecrest). Buccal flare is reduced.
Morph II (n¼ 73). The buccal and lingual cusps are sepa-rated by a continuous, mesiodistally oriented fissure. Insome specimens there are small mesial crests emanatingfrom the cusps and extending toward the opposite cusp, butthese never unite to form a continuous crest that obliteratesthe fissure as in the morph I teeth. The occlusal area of thebuccal and lingual cusps is highly asymmetric, with the buccalcusp occupying approximately two-thirds of the occlusal area.Buccal flare is pronounced.
Metric comparison. The morph I teeth are smaller on aver-age than the morph II teeth. In bivariate plots, both morphssegregate into what appear to be nearly discrete male and fe-male clusters based on size, with the morph I specimens beingalmost uniformly smaller than the morph II specimens withineach respective cluster (Fig. 18). In fact, the morph I apparentmales actually cluster more closely with the morph II femalesthan with the morph II males. The morph I males also tend tobe somewhat narrow buccolingually in relation to mesiodistallength compared to morph II specimens (likely morph I fe-males do not express this feature as clearly but the samplesize is only two).
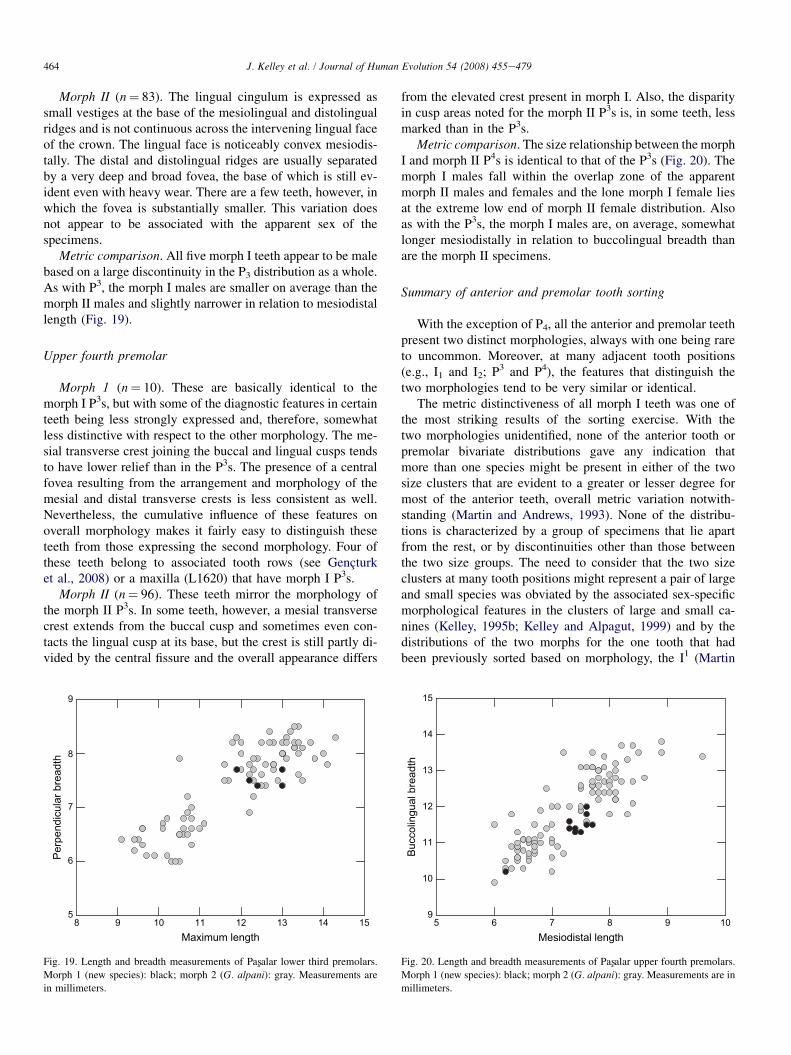
Lower third premolar
Morph I (n¼ 5). There is a distinct, continuous lingual cin-gulum from the base of the mesiolingual ridge to the distal-most part of the crown. There is also a very faint anteriorcingulum running from the mesiolingual ridge anteriorlyaround to the mesiobuccal side of the crown. The lingualface above the cingulum is flat. All five specimens have a sin-gle, flat wear facet distally, although overall wear is only lightto moderate. Thus, it is not possible to characterize the mor-phology of the distal part of the crown other than to say thatthe fovea separating the distal and distolingual ridges wouldhave been either very narrow and shallow, or that a foveawas not present.
464 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
Morph II (n¼ 83). The lingual cingulum is expressed assmall vestiges at the base of the mesiolingual and distolingualridges and is not continuous across the intervening lingual faceof the crown. The lingual face is noticeably convex mesiodis-tally. The distal and distolingual ridges are usually separatedby a very deep and broad fovea, the base of which is still ev-ident even with heavy wear. There are a few teeth, however, inwhich the fovea is substantially smaller. This variation doesnot appear to be associated with the apparent sex of thespecimens.
Metric comparison. All five morph I teeth appear to be malebased on a large discontinuity in the P3 distribution as a whole.As with P3, the morph I males are smaller on average than themorph II males and slightly narrower in relation to mesiodistallength (Fig. 19).
Upper fourth premolar
Morph 1 (n¼ 10). These are basically identical to themorph I P3s, but with some of the diagnostic features in certainteeth being less strongly expressed and, therefore, somewhatless distinctive with respect to the other morphology. The me-sial transverse crest joining the buccal and lingual cusps tendsto have lower relief than in the P3s. The presence of a centralfovea resulting from the arrangement and morphology of themesial and distal transverse crests is less consistent as well.Nevertheless, the cumulative influence of these features onoverall morphology makes it fairly easy to distinguish theseteeth from those expressing the second morphology. Four ofthese teeth belong to associated tooth rows (see Gencturket al., 2008) or a maxilla (L1620) that have morph I P3s.
Morph II (n¼ 96). These teeth mirror the morphology ofthe morph II P3s. In some teeth, however, a mesial transversecrest extends from the buccal cusp and sometimes even con-tacts the lingual cusp at its base, but the crest is still partly di-vided by the central fissure and the overall appearance differs
8 9 10 11 12 13 14 15Maximum length
5
6
7
8
9
Perp
endi
cula
r bre
adth
Fig. 19. Length and breadth measurements of Pasxalar lower third premolars.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are
in millimeters.
from the elevated crest present in morph I. Also, the disparityin cusp areas noted for the morph II P3s is, in some teeth, lessmarked than in the P3s.
Metric comparison. The size relationship between the morphI and morph II P4s is identical to that of the P3s (Fig. 20). Themorph I males fall within the overlap zone of the apparentmorph II males and females and the lone morph I female liesat the extreme low end of morph II female distribution. Alsoas with the P3s, the morph I males are, on average, somewhatlonger mesiodistally in relation to buccolingual breadth thanare the morph II specimens.
Summary of anterior and premolar tooth sorting
With the exception of P4, all the anterior and premolar teethpresent two distinct morphologies, always with one being rareto uncommon. Moreover, at many adjacent tooth positions(e.g., I1 and I2; P3 and P4), the features that distinguish thetwo morphologies tend to be very similar or identical.
The metric distinctiveness of all morph I teeth was one ofthe most striking results of the sorting exercise. With thetwo morphologies unidentified, none of the anterior tooth orpremolar bivariate distributions gave any indication thatmore than one species might be present in either of the twosize clusters that are evident to a greater or lesser degree formost of the anterior teeth, overall metric variation notwith-standing (Martin and Andrews, 1993). None of the distribu-tions is characterized by a group of specimens that lie apartfrom the rest, or by discontinuities other than those betweenthe two size groups. The need to consider that the two sizeclusters at many tooth positions might represent a pair of largeand small species was obviated by the associated sex-specificmorphological features in the clusters of large and small ca-nines (Kelley, 1995b; Kelley and Alpagut, 1999) and by thedistributions of the two morphs for the one tooth that hadbeen previously sorted based on morphology, the I1 (Martin
5 6 7 8 9 10Mesiodistal length
9
10
11
12
13
14
15
Bucc
olin
gual
bre
adth
Fig. 20. Length and breadth measurements of Pasxalar upper fourth premolars.
Morph 1 (new species): black; morph 2 (G. alpani): gray. Measurements are in
millimeters.

465J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
and Andrews, 1993). The less common Pasxalar species wascompletely hidden within the overall distributions, each ofwhich has a spatial patterning indistinguishable from that ofa single, strongly sexually dimorphic species (see Kelley andPlavcan, 1998). However, when the morph I teeth are identi-fied, they are metrically distinctive, each with at least one met-ric attribute that, on average, is substantially and recognizablydifferent from that of the morph II teeth. Again, however, thisonly becomes evident when the teeth are identified by theirdiscrete traits.
Pilbrow (2006) recently voiced the opinion that overall de-grees of I1 trait variation in extant apes suggest that the I1 var-iation in the Pasxalar sample described by Martin and Andrews(1993) should not be interpreted as indicating the presence oftwo species. While there are many debatable issues in Pil-brow’s study concerning the application of her results to fossilsamples, we will only note here that her analysis failed to takeaccount of situations, like that at Pasxalar, where the variationsin individual traits are associated, resulting in discernible, dis-crete morphologies. In such cases, it is the patterning of traitvariation that is most relevant, not just the overall range of var-iation in each particular trait compared to a given referencesample. It is, after all, the patterns of variationdparticularlythe associations of individual traits resulting in discrete mor-phologiesdthat form the basis for all species recognition.Combined with the correlated metric variation, the patternsof discrete-trait variation in the Pasxalar I1s, as in all the Pasxalaranterior and premolar teeth, are compelling evidence for thepresence of two species.
Associations of tooth types
Characterizing the two Pasxalar hominoid species requiresproper association of morphologies between tooth types. Sev-eral criteria were used that, together, permitted unambiguousassociations.
Associations in single specimens. Although the vast major-ity of the Pasxalar hominoid specimens are isolated teeth, thereare several partial maxillae and mandibles as well. Therefore,the first criterion for linking morphologies among tooth typeswas direct association. Two mandibles, R720 and G1313 (Al-pagut et al., 1990), preserve at least I1eP3. There is also a par-tial mandibular dentition (K1356-1369) consisting of CeM3,in which the mandibular bone dissolved during fossilizationbut the teeth were still in position in situ. Both of the mandib-ular specimens and the associated dentition are consistent inhaving anterior and premolar teeth identified as morph II. Un-fortunately, none of the maxillary specimens preserves associ-ations of anterior teeth, nor associations of premolars withanterior teeth. One specimen, L1620, does preserve both upperpremolars, which are very similar and represent morph I.
Commonality of morphology. This attribute was particularlyimportant for establishing associations between upper andlower teeth. The I1s and both the I1s and I2s of morph I haveprominent lingual marginal ridges that meet in a distinctiveV-shape cervically. In addition, the morph I incisors at eachof these three tooth positions are absolutely higher-crowned
on average than those of morph II (as are the morph I lowercanines) and are relatively wider mesiodistally (also character-istic of the morph I I2s). Morph II I1s and I2s both have distinctlingual pillars, which are lacking in the comparable morph Iteeth. Among the lower canines and premolars, morph I teethshare a prominent, continuous lingual cingulum and are smalleron average than those of morph II (the latter is true also of themorph I upper premolars); the morph I upper canines mayalso be smaller on average, but there are too few specimens tobe certain. Both morph I upper premolars are also relativelyelongate mesiodistally compared to the morph II teeth andhave identical morphology. The overall impression from thesevarious commonalities is one of genetically determined dentalfield effects.
Sample sizes. A third criterion, which was important for allteeth but was the only criterion for associating the male uppercanine morphs with other tooth types, is the disparity in sam-ple sizes between the two morphs for all teeth. Aside from theupper lateral incisors, a large number of which could not bereliably assigned, the sample-size ratios of morph II to morphI varied between approximately 5:1 (I1 and I2) to 18.5:1 (uppercanine). It might be thought that the large range in these ratiosis odd given the relatively large samples as a whole at eachtooth position, but with one of the morphs being so uncom-mon, substantial variation in the ratios is to be expected. Giventhe uniformly large disparity in the numbers of the twomorphs, combined with the large sample sizes overall foreach tooth type, it is highly improbable that the ratios mayin some cases be reversed, as would be the case if morph Iat one tooth position associated with morph II at another.
Additional criteria. Two other criteria were important forassociating specific teeth. One, noted above, is the distinctivepattern of hypoplasias present on all the morph I upper centralincisors and on two female upper canines (see Kelley, 2008).This was the only means by which the morph I female uppercanines could be identified initially, although once identified,they, like the morph I male upper canines, could also beseen to be marginalized in the bivariate plot of crown dimen-sions. The other criterion specifically concerned P3 tooth wearand associated morphology. The morph I P3s all have a large,flat wear facet that extends over the entire distal crown surfaceat an early wear stage. This surface occludes with the mesialportion of P3 and the wear facet appears to correspond tothe elevated mesial transverse crest of the morph I P3s.
The molar dentition
To date, no discrete features have been identified that permitthe sorting of the Pasxalar upper molars into distinct morphol-ogies, even among associated specimens that have been as-signed to species based on included anterior or premolar teeth.The same has been true for the lower molars from Pasxalar, al-though there is now some preliminary evidence for the presenceof two distinct morphologies, at least for M3. A strongly devel-oped buccal cingulum is present on the vast majority of Pasxalarlower molars, most often extending around the protoconid to thehypoconid, but frequently extending along the entire buccal
466 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
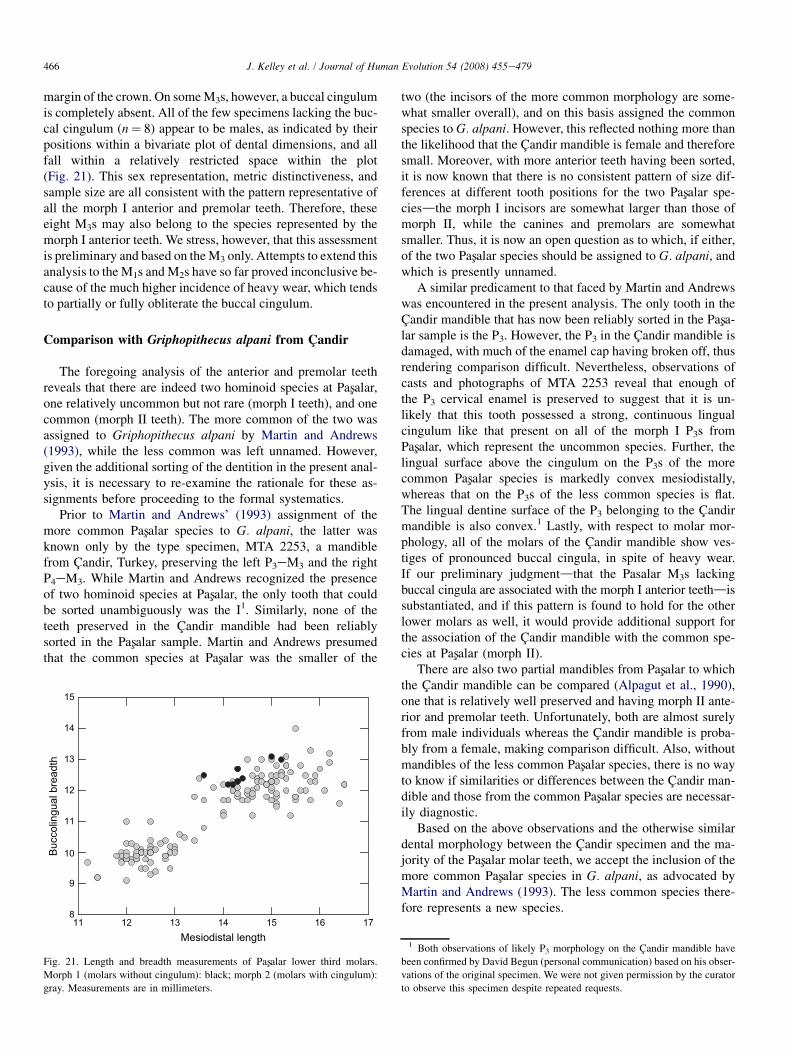
margin of the crown. On some M3s, however, a buccal cingulumis completely absent. All of the few specimens lacking the buc-cal cingulum (n¼ 8) appear to be males, as indicated by theirpositions within a bivariate plot of dental dimensions, and allfall within a relatively restricted space within the plot(Fig. 21). This sex representation, metric distinctiveness, andsample size are all consistent with the pattern representative ofall the morph I anterior and premolar teeth. Therefore, theseeight M3s may also belong to the species represented by themorph I anterior teeth. We stress, however, that this assessmentis preliminary and based on the M3 only. Attempts to extend thisanalysis to the M1s and M2s have so far proved inconclusive be-cause of the much higher incidence of heavy wear, which tendsto partially or fully obliterate the buccal cingulum.
Comparison with Griphopithecus alpani from Candir
The foregoing analysis of the anterior and premolar teethreveals that there are indeed two hominoid species at Pasxalar,one relatively uncommon but not rare (morph I teeth), and onecommon (morph II teeth). The more common of the two wasassigned to Griphopithecus alpani by Martin and Andrews(1993), while the less common was left unnamed. However,given the additional sorting of the dentition in the present anal-ysis, it is necessary to re-examine the rationale for these as-signments before proceeding to the formal systematics.
Prior to Martin and Andrews’ (1993) assignment of themore common Pasxalar species to G. alpani, the latter wasknown only by the type specimen, MTA 2253, a mandiblefrom Candir, Turkey, preserving the left P3eM3 and the rightP4eM3. While Martin and Andrews recognized the presenceof two hominoid species at Pasxalar, the only tooth that couldbe sorted unambiguously was the I1. Similarly, none of theteeth preserved in the Candir mandible had been reliablysorted in the Pasxalar sample. Martin and Andrews presumedthat the common species at Pasxalar was the smaller of the
11 12 13 14 15 16 17Mesiodistal length
8
9
10
11
12
13
14
15
Bucc
olin
gual
bre
adth
Fig. 21. Length and breadth measurements of Pasxalar lower third molars.
Morph 1 (molars without cingulum): black; morph 2 (molars with cingulum):
gray. Measurements are in millimeters.
two (the incisors of the more common morphology are some-what smaller overall), and on this basis assigned the commonspecies to G. alpani. However, this reflected nothing more thanthe likelihood that the Candir mandible is female and thereforesmall. Moreover, with more anterior teeth having been sorted,it is now known that there is no consistent pattern of size dif-ferences at different tooth positions for the two Pasxalar spe-ciesdthe morph I incisors are somewhat larger than those ofmorph II, while the canines and premolars are somewhatsmaller. Thus, it is now an open question as to which, if either,of the two Pasxalar species should be assigned to G. alpani, andwhich is presently unnamed.
A similar predicament to that faced by Martin and Andrewswas encountered in the present analysis. The only tooth in theCandir mandible that has now been reliably sorted in the Pasxa-lar sample is the P3. However, the P3 in the Candir mandible isdamaged, with much of the enamel cap having broken off, thusrendering comparison difficult. Nevertheless, observations ofcasts and photographs of MTA 2253 reveal that enough ofthe P3 cervical enamel is preserved to suggest that it is un-likely that this tooth possessed a strong, continuous lingualcingulum like that present on all of the morph I P3s fromPasxalar, which represent the uncommon species. Further, thelingual surface above the cingulum on the P3s of the morecommon Pasxalar species is markedly convex mesiodistally,whereas that on the P3s of the less common species is flat.The lingual dentine surface of the P3 belonging to the Candirmandible is also convex.1 Lastly, with respect to molar mor-phology, all of the molars of the Candir mandible show ves-tiges of pronounced buccal cingula, in spite of heavy wear.If our preliminary judgmentdthat the Pasalar M3s lackingbuccal cingula are associated with the morph I anterior teethdissubstantiated, and if this pattern is found to hold for the otherlower molars as well, it would provide additional support forthe association of the Candir mandible with the common spe-cies at Pasxalar (morph II).
There are also two partial mandibles from Pasxalar to whichthe Candir mandible can be compared (Alpagut et al., 1990),one that is relatively well preserved and having morph II ante-rior and premolar teeth. Unfortunately, both are almost surelyfrom male individuals whereas the Candir mandible is proba-bly from a female, making comparison difficult. Also, withoutmandibles of the less common Pasxalar species, there is no wayto know if similarities or differences between the Candir man-dible and those from the common Pasxalar species are necessar-ily diagnostic.
Based on the above observations and the otherwise similardental morphology between the Candir specimen and the ma-jority of the Pasxalar molar teeth, we accept the inclusion of themore common Pasxalar species in G. alpani, as advocated byMartin and Andrews (1993). The less common species there-fore represents a new species.
1 Both observations of likely P3 morphology on the Candir mandible have
been confirmed by David Begun (personal communication) based on his obser-
vations of the original specimen. We were not given permission by the curator
to observe this specimen despite repeated requests.

467J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
Maxillary morphology
There are two partial maxillae assignable to the new Pasxalarspecies based on associated anterior teeth, G1996 (associatedI2) and L1620 (associated P3eP4). Because these are theonly relatively complete specimens that can presently be in-cluded in the new species, and because maxillary morphologyis crucial for supraspecific taxonomy of hominoids, these twospecimens will be described in detail.
Specimen L1620
Fig. 22. Type specimen of Kenyapithecus kizili, L1620, occlusal (top) and
lateral (bottom) views.
Specimen L1620 is a partial left maxilla preserving thecrowns of P3eM2 and portions of the roots of M3; the distalsurface of the canine alveolus is also preserved (Table 2;Fig. 22). Judging by the size of the canine alveolus and thesizes of the other teeth, this specimen is male. The maxillarybody is well preserved, with the palatal process extending tothe intermaxillary suture from opposite P4 to the mesial halfof M2. The floor of the maxillary sinus is preserved from itsanterior extent to the M3 alveolus. A small portion of the lat-eral margin of the incisive fossa is preserved on the anterioredge of the palatal process. The buccal surface of the alveolarprocess is rather eroded, exposing the roots of the premolars;all but the very base of the root of the zygomatic process isbroken and eroded.
The palatal process is approximately 7.5 mm thick overmuch of its length, thinning anteriorly and just slightly at itsmost posterior extent in the vicinity of M2 (Fig. 23). At its bro-ken anterior edge opposite P3, it is still approximately 2 mmthick in the midline. The break angles anteriorly toward the al-veolar process and it therefore gradually thins as it nears the
Table 2
Measurements (in mm) of L1620, the holotype maxilla of Kenyapithecus kizili
Dimension Value
Depth of palate at P4 8.2
Breadth of palate at P4 27.2*
Depth of palate at M1 9.3
Breadth of palate at M1 26.8*
Depth of palate at M2 9.4
Breadth of palate at M2 24.0*
Maximum palatal thickness 7.5
Height of maxillary sinus floor above alveolar margin at M2 13.5
P3
MD 7.6
BL 11.5
P4
MD 7.6
BL 12.0
M1
MD 11.1
BL 12.5
M2
MD 12.0
BL 13.9
* Palatal process of L1620 is preserved to the intermaxillary suture.
alveolar process in the vicinity of the C-P3 interproximalzone. The palate would have been somewhat broader anteriorlythan posteriorly, which is not unexpected since L1620 is almostsurely male. Mirroring the left side, reconstructed palatalbreadth at P4 is 27.2 mm, which decreases to 24.0 mm at M2
(Fig. 24; Table 2). The palate is only moderately deep,
Fig. 23. Type specimen of Kenyapithecus kizili, L1620, lingual view.
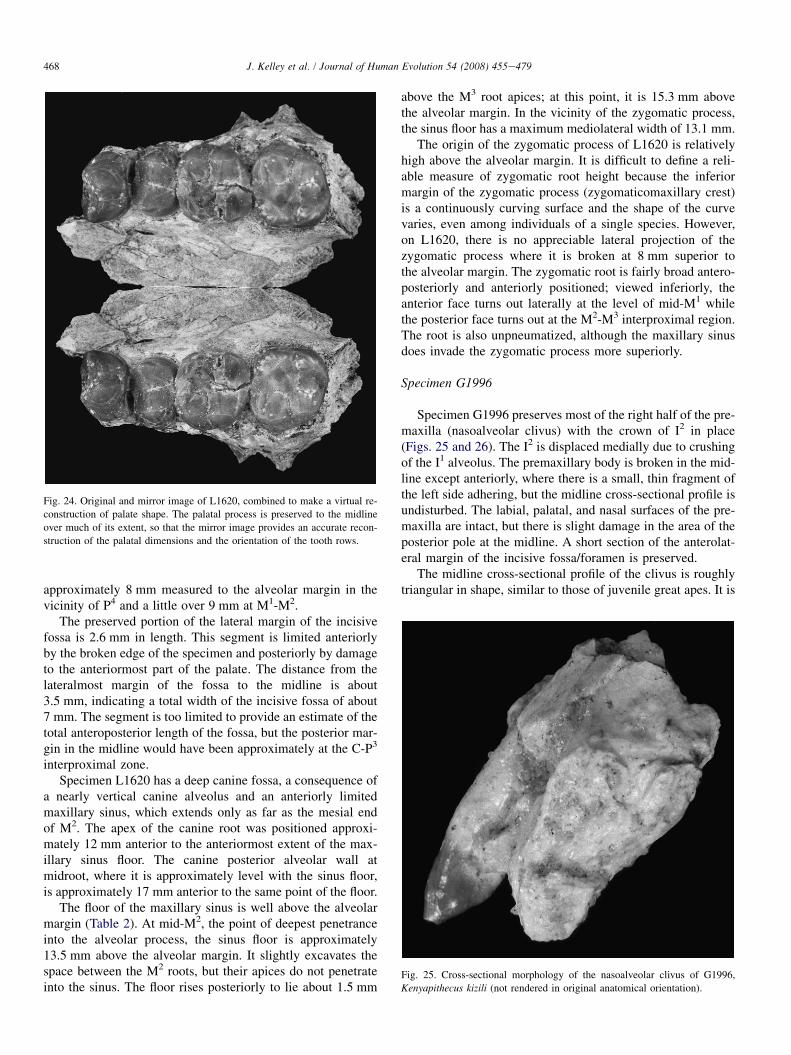
Fig. 24. Original and mirror image of L1620, combined to make a virtual re-
construction of palate shape. The palatal process is preserved to the midline
over much of its extent, so that the mirror image provides an accurate recon-
struction of the palatal dimensions and the orientation of the tooth rows.
Fig. 25. Cross-sectional morphology of the nasoalveolar clivus of G1996,
Kenyapithecus kizili (not rendered in original anatomical orientation).
468 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
approximately 8 mm measured to the alveolar margin in thevicinity of P4 and a little over 9 mm at M1-M2.
The preserved portion of the lateral margin of the incisivefossa is 2.6 mm in length. This segment is limited anteriorlyby the broken edge of the specimen and posteriorly by damageto the anteriormost part of the palate. The distance from thelateralmost margin of the fossa to the midline is about3.5 mm, indicating a total width of the incisive fossa of about7 mm. The segment is too limited to provide an estimate of thetotal anteroposterior length of the fossa, but the posterior mar-gin in the midline would have been approximately at the C-P3
interproximal zone.Specimen L1620 has a deep canine fossa, a consequence of
a nearly vertical canine alveolus and an anteriorly limitedmaxillary sinus, which extends only as far as the mesial endof M2. The apex of the canine root was positioned approxi-mately 12 mm anterior to the anteriormost extent of the max-illary sinus floor. The canine posterior alveolar wall atmidroot, where it is approximately level with the sinus floor,is approximately 17 mm anterior to the same point of the floor.
The floor of the maxillary sinus is well above the alveolarmargin (Table 2). At mid-M2, the point of deepest penetranceinto the alveolar process, the sinus floor is approximately13.5 mm above the alveolar margin. It slightly excavates thespace between the M2 roots, but their apices do not penetrateinto the sinus. The floor rises posteriorly to lie about 1.5 mm
above the M3 root apices; at this point, it is 15.3 mm abovethe alveolar margin. In the vicinity of the zygomatic process,the sinus floor has a maximum mediolateral width of 13.1 mm.
The origin of the zygomatic process of L1620 is relativelyhigh above the alveolar margin. It is difficult to define a reli-able measure of zygomatic root height because the inferiormargin of the zygomatic process (zygomaticomaxillary crest)is a continuously curving surface and the shape of the curvevaries, even among individuals of a single species. However,on L1620, there is no appreciable lateral projection of thezygomatic process where it is broken at 8 mm superior tothe alveolar margin. The zygomatic root is fairly broad antero-posteriorly and anteriorly positioned; viewed inferiorly, theanterior face turns out laterally at the level of mid-M1 whilethe posterior face turns out at the M2-M3 interproximal region.The root is also unpneumatized, although the maxillary sinusdoes invade the zygomatic process more superiorly.
Specimen G1996
Specimen G1996 preserves most of the right half of the pre-maxilla (nasoalveolar clivus) with the crown of I2 in place(Figs. 25 and 26). The I2 is displaced medially due to crushingof the I1 alveolus. The premaxillary body is broken in the mid-line except anteriorly, where there is a small, thin fragment ofthe left side adhering, but the midline cross-sectional profile isundisturbed. The labial, palatal, and nasal surfaces of the pre-maxilla are intact, but there is slight damage in the area of theposterior pole at the midline. A short section of the anterolat-eral margin of the incisive fossa/foramen is preserved.
The midline cross-sectional profile of the clivus is roughlytriangular in shape, similar to those of juvenile great apes. It is

Fig. 26. Superior (left) and medial (right) views of G1996. Arrows indicate
the preserved portion of the margin of the incisive fossa.
469J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
relatively thick labiolingually (maximum length measuredalong the palatal surface: 18.3 mm; maximum perpendicularbreadth: 9.0 mm). The angle between the nasal and palatal sur-faces is approximately 55�. There is no firm reference for ori-enting G1996, but we estimate that the angle of the long axisof the clivus relative to the horizontal (approximate occlusal oralveolar plane) was fairly steep, between approximately 35�
and 50�. This orientation is consistent with the position ofthe I2, which, although slightly displaced medially, appearsto have been minimally affected otherwise. Regardless ofthe precise orientation of the premaxilla, the nasoalveolar cli-vus appears to have been anteroposteriorly short. It also seemslikely that the posterior margin of the clivus at nasospinale waselevated above the nasal floor (‘‘stepped’’), regardless of theprecise relationship between the premaxilla and the portionsof the maxilla contributing to the nasal cavity entrance (seeMcCollum and Ward, 2000).
The preserved segment of the margin of the incisive fossa isindicated by a continuous cortical surface from the nasal to thepalatal surfaces (Fig. 26). There is a tiny chip of bone missingfrom this margin adjacent to the midline, but the damage isminimal and the margin to the midline can be reconstructedbased on the preserved portion. The mediolateral breadth ofthe fossa can be estimated by doubling the linear distancefrom the midline to the lateralmost point of the preserved mar-gin (4.2 mm), which equals a minimum breadth for the inci-sive fossa of about 8.4 mm. However, the curvature of thepreserved segment of the margin reveals that the very lateral-most portion is broken away, so the fossa would have beenslightly wider (Fig. 26). This is compatible with the estimatederived from L1620 (see above).
Based on the preserved morphology of both G1996 andL1620, it appears, therefore, that the incisive fossa/foramenin this species was relatively broad and fairly large overall.While the precise conformation of the subnasal region is un-known, in particular the topographic relationship of the pre-maxilla to the palate, judging by the large size of theincisive fossa and the shortness and probable orientation ofthe premaxilla, we do not consider it likely that there was ex-tensive overlap between the palate and premaxilla such aswould produce an elongate incisive canal as in extant greatapes (Andrews and Cronin, 1982; Ward and Kimbel, 1983)
or late Miocene apes such as Sivapithecus (Ward and Kimbel,1983), Ankarapithecus (Andrews and Tekkaya, 1980; Begunand Gulec, 1998), or Ouranopithecus (Graecopithecus) (deBonis and Koufos, 1993). Beyond this, it is impossible tobe more definitive. There could have been either a broad sep-aration between the premaxilla and palate, resulting in a size-able palatine fenestra as in early Miocene apes or gibbons, noseparation such that the anterior margin of the palate waslocated more or less directly beneath the posterior pole ofthe premaxilla as in Dryopithecus brancoi (Begun, 1994;Ward and Kimbel, 1983), or slight overlap producing a short,broad incisive canal as in Nacholapithecus (Kunimatsu et al.,2004). In each of these possibilities, it is likely that therewould have been a large incisive foramen palatally.
Systematics
Order PRIMATES Linnaeus, 1758Superfamily HOMINOIDEA Gray, 1825Family INCERTAE SEDISGenus KENYAPITHECUS Leakey, 1962
Diagnosis (replaces the emended diagnosis of Leakey,1967). A genus of hominoid combining: robust upper centralincisors that are both mesiodistally broad and labiolinguallythick, with hypertrophied lingual marginal ridges and lackinga distinct lingual pillar or well-defined tubercle; slender lowercanine crowns that are tall relative to mesiodistal length andthat have a distinct and continuous lingual cingulum; upperpremolars with approximately equal cusp areas and well-de-veloped transverse crests linking the paracone and protocone;lower third premolars with distinct cingulum or cingular rem-nants; upper molars with low, bunodont cusps, thick enamelwith low dentine penetrance, and insubstantial cingular rem-nants restricted to the mesial portion of the protocone; maxillawith a moderately deep, robust alveolar process, an anteriorlyrestricted maxillary sinus that does not extend over the premo-lars, a relatively flat sinus floor positioned mostly superior tothe molar root apices, an anteriorly positioned zygomatic pro-cess with a moderately high origin above the alveolar marginand a zygomaticoalveolar crest orientation that becomes morelateral than superior well above the alveolar margin, and a pro-nounced canine jugum and fossa; nasoalveolar clivus short andtriangular in cross section; large incisive fossa and foramen;distal humerus in which the trochlea is broader than the capit-ulum and has a well-developed but bulbous lateral lip delimit-ing a distinct zona conoidea, but in which both the trochleaand the zona conoidea are shallower than in extant greatapes, the medial epicondyle is short and strongly posteriorlyoriented, the lateral epicondyle is anteroposteriorly deep, thecoronoid fossa is larger than the radial fossa, and the lateralarticular surface of the olecranon fossa extends posteriorlyas a well-developed flange.
Kenyapithecus differs from all other hominoid genera inhaving upper central incisors that lack a lingual tubercle or pil-lar and that have greatly hypertrophied lingual marginal ridges;differs from all other hominoids except perhaps Afropithecus in

470 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
having upper premolars with equally sized protocone and para-cone that are joined by a continuous, elevated mesial crest; dif-fers from other Miocene hominoids except Equatorius andGriphopithecus in having a strongly proclined mandibular sym-physis and a shallowly sloping sublingual planum with respectto the alveolar margins of the corpora; differs from early Mio-cene hominoids such as Proconsul, Afropithecus, andMorotopithecus in having upper molars with low, bunodontcusps with low dentine penetrance and only small vestiges ofcingula; differs from all early Miocene hominoids, as well asmiddle Miocene Equatorius, Nacholapithecus, and Griphopi-thecus, in having slender lower canines that are high-crownedin relation to maximum crown length at the cervix; lacks the hy-pertrophied upper premolars of Morotopithecus and Afropithe-cus; differs from Equatorius and Nacholapithecus in havinga restricted, elevated maxillary sinus, a higher origin of the zy-gomatic root and a zygomaticoalveolar crest that turns laterallyhigher above the alveolar margin, lower third premolars witha prominent, continuous lingual cingulum, upper molars witha less extensive protocone cingulum, and (compared to Equa-torius only) less posteriorly inclined upper canine roots; differsfrom Otavipithecus in having molars with thick enamel and lowdentine penetrance of the cusps; differs from late Miocene gen-era such as Sivapithecus, Ankarapithecus, and Ouranopithecusin having a short nasoalveolar clivus with a large incisive fossa/foramen rather than a long, narrow incisive canal, and fromthese and other late Miocene hominoids in retaining a continu-ous cingulum on the lower third premolar; lacks the distinctive,intricately crenulated molars with peripheralized cusps ofLufengpithecus and, to a slightly lesser extent, Khoratpithecus;also differs from Khoratpithecus in having a more slender andrelatively higher-crowned male lower canine and molar cuspswith lower dentine penetrance; differs from Dryopithecus inhaving upper molars with thick enamel and low dentine pene-trance, and a smaller, more restricted maxillary sinus, andfrom both Dryopithecus and Pierolapithecus in having a lowerorigin of the zygomatic root with a zygomaticoalveolar crestthat becomes laterally oriented closer to the alveolar margin;also differs from Pierolapithecus in having upper premolarsthat are much smaller than the molars and less mesiodistallyelongate molars with less peripheralized cusps; differs fromSamburupithecus in having upper molars with a distinct, contin-uous crista obliqua and cusps with lower dentine penetrance,and upper premolars that are much smaller in relation to themolars.
Type species. Kenyapithecus wickeri.
KENYAPITHECUS KIZILI, sp. nov.
Sivapithecus darwini (Abel), Andrews and Tobien, 1977(in part)
cf. Sivapithecus (Pilgrim), Alpagut et al., 1990
Diagnosis. A species of Kenyapithecus distinguished fromK. wickeri by the following features: I1 higher-crowned in
relation to mesiodistal length, with lingual marginal ridgesthat meet cervically in a sharp V; I2 much narrower mesiodis-tally in comparison to I1, with nearly symmetrical mesial anddistal incisal edges; male upper canine with a distinct buccalcingulum; lower canine with a very prominent lingual cingu-lum and, in males, a double mesiolingual groove; P3 and P4
with a continuous, elevated transverse crest linking the proto-cone and paracone mesially and separating the mesial foveafrom the rest of the crown; P3 with a shallow, restricted foveabetween the distal and distolingual ridges. Other salient fea-tures of K. kizili that are unknown for K. wickeri include: I1
and I2 with distinct mesial and distal marginal ridges confinedto the lower half of the crown that join cervically in a sharp V;male lower canine root very long in relation to root cervicaldimensions.
Type specimen. Ankara University L1620, a partial leftmaxilla with P3 to M2 (Fig. 22; Table 2).
Referred material. Maxilla: G1996 (with I2); I1: BP27,B454, C153, D298, D655, H1641, K1356, N1390, R1678,Y826; I1: BP1303, D1164, F104, H1819, K1387, L1606,M1858, S1116; I2: C148, C152, D869, G518, K1334,N1394; I2: BP565, B622, C162, C631, D983, G1477,G1545, L1605, L1619, R1658, R1674, U562; C1: B446,M913 (females), N1427, S1112 (males); C1: T582 (female),E598, F547, G1352, K1314, K1440, N1515, U556 (males);P3: BP43, D772, D1161, K1413, K1456, L1594, L1623,L1624, M1876, U517; P3: G1141, G1292, L1593, M947,S1111; P4: C182, D178, H1893, L1629, L1630, M954,R1682, V761, V763.
Horizon. Gray sand unit in the middle Miocene deposits atPasxalar, Turkey.
Etymology. Kizil means red in Turkish and the name istaken from Kizil Tepe, meaning red hill, which is the hill over-looking the site of Pasxalar.
Discussion
There is presently little agreement, and certainly no consen-sus, regarding family-level systematics among Miocene fossilcatarrhines (e.g., various contributions in Hartwig, 2002). Theplacement of Kenyapithecus is particularly problematic be-cause it is represented by so little morphology beyond the den-tition. For these reasons, we have chosen to refrain fromassigning Kenyapithecus to any existing family, or to namea new family that is either monotypic for the genus or thatmight contain other genera as well.
Generic assignment of K. kizili
The type species of Kenyapithecus, K. wickeri, was de-scribed by Leakey (1962) based on left and right partial max-illae from the site of Fort Ternan, Kenya. Fort Ternan has beendated to ca.14 Ma (Bishop et al., 1969; Shipman et al., 1981)or perhaps slightly younger (Pickford et al., 2006). Leakey(1967) later described an additional specimen from FortTernan, an upper central incisor (KNM-FT 49) and alsoamended the diagnosis of the genus. A mandible thought to

Table 3
Male relative lower canine height in Miocene hominoids
n HT/L
Early Miocene
Afropithecus turkanensis 3 1.11e1.25
Proconsul nyanzae/heseloni 7 1.41e1.50
Proconsul major 1 1.41
Middle Miocene
Nacholapithecus kerioi 1 1.23
Griphopithecus alpani 15 1.24e1.50
Equatorius africanus 1 1.35
KNM-MB 32348 (Maboko) 1 1.67
Kenyapithecus kizili 4 1.49e1.70
Kenyapithecus wickeri 1 1.63
471J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
be from the same individual as the type specimen was subse-quently described by Andrews (1971), and some isolated teethwere added to the hypodigm by Andrews and Walker (1976).Further specimens were added by Pickford (1985) and Harri-son (1992), including some that had previously been attributedto Proconsul nyanzae.
In the 1967 paper, Leakey named a second species of Ken-yapithecus, K. africanus (Leakey, 1967). The type specimen,a maxilla with P3eM1 (BMNH M16649)doriginally madethe holotype of Sivapithecus africanus by Le Gros Clark andLeakey (1951)dwas considered by Leakey to have comefrom the early Miocene site of Rusinga. It was subsequentlyshown to be unlike other material from Rusinga in featuresof preservation (Andrews and Molleson, 1979) and is moreconsistent with material from the middle Miocene site of Ma-boko Island, a provenance that is now widely accepted. Theactual early Miocene specimens (e.g., Leakey, 1968) wereeventually reassigned to species of Proconsul (Andrews,1978).
It eventually became clear to many workers (e.g., Pickford,1985; Harrison, 1992) that K. africanus differs from K. wickeriin preserved parts, particularly maxillary morphology. Thisdifference has now been formally recognized with the transferof K. africanus to a new genus, Equatorius (Ward et al., 1999).Other specimens from Mabokodas well as the nearby sites ofMajiwa and Kalomadthat had been included in Kenyapithe-cus (Pickford, 1982, 1985; McCrossin and Benefit, 1993)were also transferred to Equatorius. This left K. wickeri,known only by the small number of specimens from FortTernan, as the sole representative of the genus. Kenyapithecuskizili is, therefore, only the second species attributable to Ken-yapithecus and greatly expands our knowledge of the genus.
In almost every respect, the type specimen of K. kizili isnearly identical to the comparable elements of the type spec-imen(s) of K. wickeri, KNM-FT 46a.2 Comparison is facili-tated by the fact that the two maxillae are almost identicallypreserved (L1620 preserves the P3 and more of the palatal pro-cess, while KNM-FT 46a preserves more of the zygomaticprocess and slightly more of the anterior alveolar process, in-cluding more of the posterior surface of the canine alveolus).Both specimens display a combination of morphological fea-tures that, collectively, distinguish them from comparable re-mains of other hominoid genera. These features, in additionto those enumerated in the diagnosis of the genus, includethe following: a large separation between the maxillary sinusand the canine alveolus, which contributes to the deep caninefossa; a zygomatic process that originates superior to M1-M2
with a root that is not excavated by the maxillary sinus (al-though the sinus does invade the process more superiorly);and a palate of only moderate depth. Kenyapithecus kizilithus shares the morphology by which K. wickeri can be distin-guished from other Miocene hominoid genera.
2 Leakey (1962) designated four separate specimens as the type of K. wick-
eri, only two of which, the left and right partial maxillae, KNM-FT 46a and
KNM-FT 47, are generally accepted as coming from the same individual
(see Pickford, 1985; Harrison, 1992).
Not surprisingly, many genera, and even species, are vari-able in the expression of some of the above features. For ex-ample, both Proconsul and Sivapithecus show variable sinusdevelopment (Andrews, 1978; Teaford et al., 1988; Wardand Pilbeam, 1983), which changes ontogenetically and mayshow sexual dimorphism in some species. Variation in sinusdevelopment also results in a certain amount of correspondingvariation in the expression of the canine fossa in Proconsul(Teaford et al., 1988) and in the robusticity of the zygomaticroot in Sivapithecus (Ward and Pilbeam, 1983; JK, personalobservation). Such variation notwithstanding, it is possible todistinguish Kenyapithecus from all other hominoid genera bythe features described above.
Early Miocene genera. Kenyapithecus displays a number ofderived features, particularly of the dentition, with respect tothe early Miocene genera Proconsul, Rangwapithecus, Moro-topithecus, and Afropithecus (Le Gros Clark and Leakey,1951; Pilbeam, 1969; Andrews, 1974, 1978; Leakey et al.,1988; Gebo et al., 1997). Upper molars in all of these generahave more discrete, conical cusps, more pronounced and ex-tensive lingual cingula (plus mesial and buccal cingula insome), and extensive enamel wrinkling. Upper premolar cuspsare more heteromorphic and separated by a deep fissure exceptin Afropithecus, which has well-developed transverse crestsbetween the paracone and protocone, but greatly enlarged up-per premolars (Leakey et al., 1988). Lower canines in Procon-sul and Afropithecus are low-crowned in relation to mesiodistallength and are blunt in appearance (Table 3); a lone lowercanine of Rangwapithecus is relatively high-crowned but themorphology is highly derived (Kelley, 1986; Kelley, 1995b;Ward et al., 1999). Upper central incisors are known for all gen-era but Morotopithecus, and all have distinct lingual pillars ortubercles and lack hypertrophied marginal ridges.
Despite the variation in sinus development in Proconsuland Rangwapithecus noted by Andrews (1978) and Teafordet al. (1988), the early Miocene genera generally have moreextensive maxillary sinuses then Kenyapithecus, particularly
Late Miocene
Lufengpithecus lufengensis 7 1.56e1.85
Dryopithecus fontani 2 1.57e1.63
Dryopithecus laietanus 2 1.54e1.57
Ouranopithecus macedoniensis 2 1.43e1.55
HT/L¼ buccal crown height/maximum crown length at the cervix.

472 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
with respect to the degree of anterior projection (except Rang-wapithecus, which, however, has the greatest penetration of thealveolar process [Andrews, 1978]). Afropithecus is the leastdissimilar, at least in terms of alveolar process penetration,but the sinus still extends into the premolar region and nearlycontacts the apex of the canine root (Leakey et al., 1988; Lea-key and Walker, 1997; Ward et al., 1999). As a consequence,the early Miocene taxa also generally have less deeply incisedcanine fossae. The latter might also be a reflection in part ofthe primitively elongate snout of the early Miocene genera.While the morphology of the lower anterior face is mostly un-known for Kenyapithecus, the shape and likely orientation ofthe K. kizili premaxilla (G1996) is a good indication that thelower face was less projecting than in the early Miocene gen-era. Early Miocene taxa such as Proconsul and Morotopithe-cus also have relatively thin maxillary palatine processes(Ward and Pilbeam, 1983; Begun, 1994), whereas it is rela-tively quite thick in K. kizili (Table 2). Finally, the origin ofthe zygomatic process, when scaled according to tooth size,appears to be somewhat higher in Kenyapithecus than in theearly Miocene genera, especially since the value used for Ken-yapithecus is a minimum due to zygomatic breakage (Table 4;Fig. 27; see also Kunimatsu et al., 2004). Kunimatsu et al.(2004) noted that the orientation of the inferior border of the
Table 4
Comparative maxillary metrics of Miocene hominoids
Taxon Specimens Maxillary sinus/ca
root distance1
Proconsul nyanzae RU 1600 (_) w9
Proconsul heseloni RU 2036 (\) dRU 7290 (\) d
Afropithecus turkanensis WK 16999 d
Kenyapithecus kizili L1620 (_) 17
Kenyapithecus wickeri FT 46/47 (\) 11
Griphopithecus alpani G432 (\?) d
Equatorius africanus M16649 (_) 4
Nacholapithecus kerioi BG 14708 (\) d
BG 17835 (\?) dBG 14700 (_) d
BG 17840 (_) d
Pierolapithecus catalaunicus IPS 21350 (_) dAnkarapithecus meteai AS95-500 (\) d
MTA 2125 (_) 17
Sivapithecus sivalensis GSP 9977 (_) 15
GSP 11704 (_) dGSP 15000 (_) 17
Dryopithecus brancoi RUD 12 (\) d
RUD 200 (\) 8
RUD 7 (_) dDryopithecus laietanus CLl 18000 (_) 9
M1 and some maxillary data are from Alpagut et al. (1996), Andrews (1978), Begun
et al. (1988), Moya-Sola and Kohler (1995), Pickford (1985), and Teaford et al. (19
with these types of measurements is unwarranted.1 Measured from the anteriormost extent of the maxillary sinus floor to the poste
canine root).2 Measured from the lowest recess of the maxillary sinus to the alveolar margin3 Measured from the alveolar margin to the point where the inferior surface of th
imens in which the crest does not achieve a completely lateral orientation, height w
oriented (see text for further explanation).
process (zygomaticoalveolar crest) also differs between K.wickeri and all of the early Miocene taxa, being directed later-ally in the latter and superolaterally in the former. This obser-vation is addressed further below.
The mandibular symphysis of K wickeri, preserved inKNM-FT 45, has a strongly developed and posteriorly projec-ting inferior transverse torus (Andrews, 1971), which contrib-utes to the strongly proclined symphyseal orientation. Thissymphyseal morphology contrasts with that of most early Mio-cene taxa, in which the inferior torus is either absent or onlyweakly expressed (Andrews, 1978; Brown, 1997); in Afropi-thecus, the inferior torus may be robust but it does not projectfar posteriorly, in keeping with the more vertical orientation ofthe symphysis (Brown, 1997).
The K. wickeri distal humerus, KNM-FT 2751, differs fromthat of Proconsul species in a number of respects (Senut,1986; Rose, 1997). The lateral epicondyle is more developed(especially in terms of its thickness anteroposteriorly), the me-dial epicondyle is more posteriorly oriented, the coronoid fossais larger than the radial fossa, and the trochlea is deeper, result-ing in a more symmetrical shape of the lateral trochlear lip.
Middle Miocene genera. Kenyapithecus can be distin-guished from Griphopithecus by the derived features of I1,C1, and P4 described above in the comparison of K. kizili
nine Maxillary sinus
floor to alveolar
margin2
Zygomatic process
height3M1 area
d 11 85
d 6 76
d 7 74
d 14 150
14 >8 139
12e14 >15 110
14 d 107
10 9 118
d 7 80
d 8 dd 10 102
d 10 97
d 19 113
d 16 133
d 26 162
8 19 157
17 27 146
6 24 163
13 >15 94
d 17 88
d >20 118
7 24 107
and Gulec (1998), Kordos and Begun (2001), Kunimatsu et al. (2004), Leakey
88). All measurements rounded to nearest millimeter because higher resolution
rior surface of the canine alveolus, except in MTA 2125 (measured to apex of
.
e zygomaticoalveolar crest becomes laterally (horizontally) oriented. For spec-
as measured at the point where the crest became more laterally than superiorly
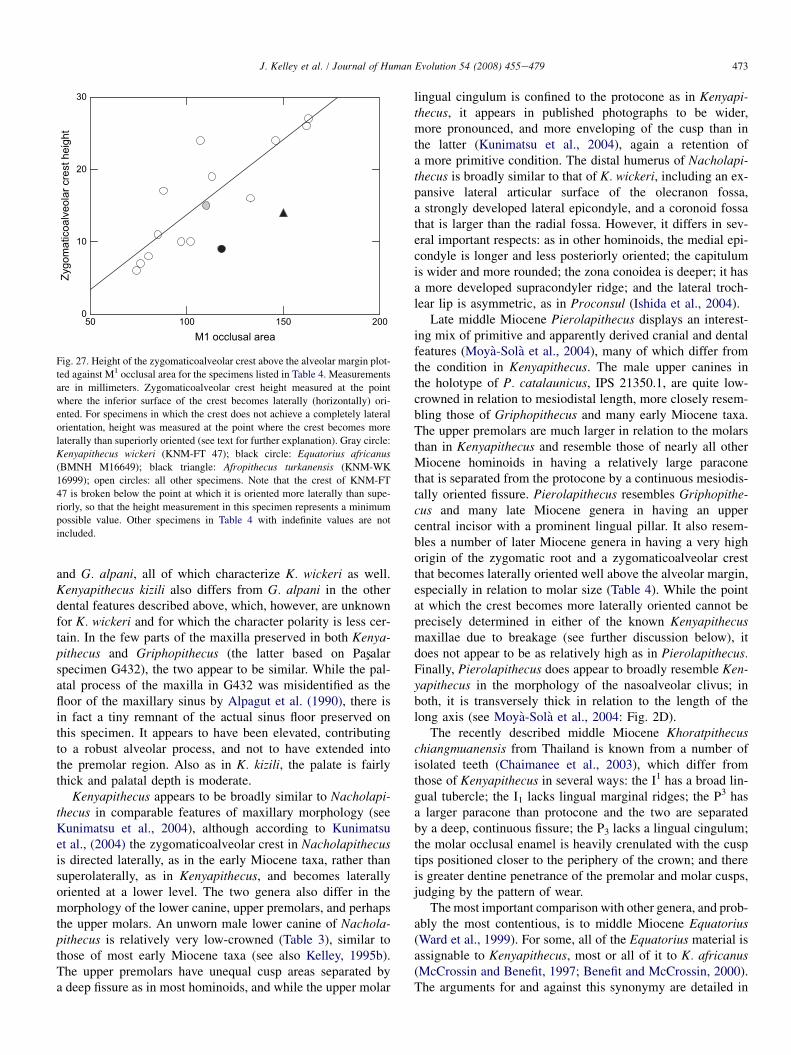
50 100 150 200M1 occlusal area
0
10
20
30
Zygo
mat
icoa
lveo
lar c
rest
hei
ght
Fig. 27. Height of the zygomaticoalveolar crest above the alveolar margin plot-
ted against M1 occlusal area for the specimens listed in Table 4. Measurements
are in millimeters. Zygomaticoalveolar crest height measured at the point
where the inferior surface of the crest becomes laterally (horizontally) ori-
ented. For specimens in which the crest does not achieve a completely lateral
orientation, height was measured at the point where the crest becomes more
laterally than superiorly oriented (see text for further explanation). Gray circle:
Kenyapithecus wickeri (KNM-FT 47); black circle: Equatorius africanus(BMNH M16649); black triangle: Afropithecus turkanensis (KNM-WK
16999); open circles: all other specimens. Note that the crest of KNM-FT
47 is broken below the point at which it is oriented more laterally than supe-
riorly, so that the height measurement in this specimen represents a minimum
possible value. Other specimens in Table 4 with indefinite values are not
included.
473J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
and G. alpani, all of which characterize K. wickeri as well.Kenyapithecus kizili also differs from G. alpani in the otherdental features described above, which, however, are unknownfor K. wickeri and for which the character polarity is less cer-tain. In the few parts of the maxilla preserved in both Kenya-pithecus and Griphopithecus (the latter based on Pasxalarspecimen G432), the two appear to be similar. While the pal-atal process of the maxilla in G432 was misidentified as thefloor of the maxillary sinus by Alpagut et al. (1990), there isin fact a tiny remnant of the actual sinus floor preserved onthis specimen. It appears to have been elevated, contributingto a robust alveolar process, and not to have extended intothe premolar region. Also as in K. kizili, the palate is fairlythick and palatal depth is moderate.
Kenyapithecus appears to be broadly similar to Nacholapi-thecus in comparable features of maxillary morphology (seeKunimatsu et al., 2004), although according to Kunimatsuet al., (2004) the zygomaticoalveolar crest in Nacholapithecusis directed laterally, as in the early Miocene taxa, rather thansuperolaterally, as in Kenyapithecus, and becomes laterallyoriented at a lower level. The two genera also differ in themorphology of the lower canine, upper premolars, and perhapsthe upper molars. An unworn male lower canine of Nachola-pithecus is relatively very low-crowned (Table 3), similar tothose of most early Miocene taxa (see also Kelley, 1995b).The upper premolars have unequal cusp areas separated bya deep fissure as in most hominoids, and while the upper molar
lingual cingulum is confined to the protocone as in Kenyapi-thecus, it appears in published photographs to be wider,more pronounced, and more enveloping of the cusp than inthe latter (Kunimatsu et al., 2004), again a retention ofa more primitive condition. The distal humerus of Nacholapi-thecus is broadly similar to that of K. wickeri, including an ex-pansive lateral articular surface of the olecranon fossa,a strongly developed lateral epicondyle, and a coronoid fossathat is larger than the radial fossa. However, it differs in sev-eral important respects: as in other hominoids, the medial epi-condyle is longer and less posteriorly oriented; the capitulumis wider and more rounded; the zona conoidea is deeper; it hasa more developed supracondyler ridge; and the lateral troch-lear lip is asymmetric, as in Proconsul (Ishida et al., 2004).
Late middle Miocene Pierolapithecus displays an interest-ing mix of primitive and apparently derived cranial and dentalfeatures (Moya-Sola et al., 2004), many of which differ fromthe condition in Kenyapithecus. The male upper canines inthe holotype of P. catalaunicus, IPS 21350.1, are quite low-crowned in relation to mesiodistal length, more closely resem-bling those of Griphopithecus and many early Miocene taxa.The upper premolars are much larger in relation to the molarsthan in Kenyapithecus and resemble those of nearly all otherMiocene hominoids in having a relatively large paraconethat is separated from the protocone by a continuous mesiodis-tally oriented fissure. Pierolapithecus resembles Griphopithe-cus and many late Miocene genera in having an uppercentral incisor with a prominent lingual pillar. It also resem-bles a number of later Miocene genera in having a very highorigin of the zygomatic root and a zygomaticoalveolar crestthat becomes laterally oriented well above the alveolar margin,especially in relation to molar size (Table 4). While the pointat which the crest becomes more laterally oriented cannot beprecisely determined in either of the known Kenyapithecusmaxillae due to breakage (see further discussion below), itdoes not appear to be as relatively high as in Pierolapithecus.Finally, Pierolapithecus does appear to broadly resemble Ken-yapithecus in the morphology of the nasoalveolar clivus; inboth, it is transversely thick in relation to the length of thelong axis (see Moya-Sola et al., 2004: Fig. 2D).
The recently described middle Miocene Khoratpithecuschiangmuanensis from Thailand is known from a number ofisolated teeth (Chaimanee et al., 2003), which differ fromthose of Kenyapithecus in several ways: the I1 has a broad lin-gual tubercle; the I1 lacks lingual marginal ridges; the P3 hasa larger paracone than protocone and the two are separatedby a deep, continuous fissure; the P3 lacks a lingual cingulum;the molar occlusal enamel is heavily crenulated with the cusptips positioned closer to the periphery of the crown; and thereis greater dentine penetrance of the premolar and molar cusps,judging by the pattern of wear.
The most important comparison with other genera, and prob-ably the most contentious, is to middle Miocene Equatorius(Ward et al., 1999). For some, all of the Equatorius material isassignable to Kenyapithecus, most or all of it to K. africanus(McCrossin and Benefit, 1997; Benefit and McCrossin, 2000).The arguments for and against this synonymy are detailed in

474 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
Ward et al. (1999), Benefit and McCrossin (2000), and Kelleyet al. (2000). Briefly, Benefit and McCrossin contended thatwhatever differences there are between the K. wickeri and E.africanus hypodigms, particularly in maxillary morphology,they are appropriately viewed as expected variation within a sin-gle genus (see also McCrossin and Benefit, 1997), a position dis-puted by most other researchers (e.g., Pickford, 1985; Harrison,1992; Ward et al., 1999; Kelley et al., 2000; Begun, 2000; Ku-nimatsu et al., 2004).
The maxillary morphology of Equatorius is known fromthe type specimen of E. africanus (BMNH M16649). Thisspecimen has an expansive maxillary sinus that extends ante-riorly over the P4, into the root of the zygomatic process, anddeeply into the alveolar process between the molar roots. Theheight of the lowest recess of the sinus floor above the alveolarmargin in M16649 is much less than in the type specimens ofK. kizili, L1620, or K. wickeri, KNM-FT 46a/47, in spite of atleast the Fort Ternan specimens being somewhat smaller thanM16649 (Table 4). These differences cannot be attributed tosexual dimorphism, as suggested for the Maboko and FortTernan specimens by McCrossin and Benefit (1994) and Ben-efit and McCrossin (2000), because L1620, like M16649, isprobably male and KNM-FT 46/47 is probably female. Theyare also unlikely to be explainable as ontogenetic differences,as L1620 and M16649 are both young adults with minimalpremolar and molar wear; KNM-FT 46a is somewhat olderbased on tooth wear. Since sinus volume tends to increasewith age, the variation in maxillary sinus development in thesethree specimens is opposite of what would be expected froman ontogenetic perspective.
Opinions also differ regarding the origin of the zygomaticprocess in relation to the alveolar margin. Although the heightof the process in M16649 is unquestionably somewhat lowerthan in KNM-FT 46a, as has been frequently noted, whenscaled against tooth size, the difference does not exceed thevariation observable in a single species (McCrossin and Ben-efit, 1997; Kunimatsu et al., 2004). However, the zygomaticroot presents a continuously curving surface that graduallyemerges from the alveolar process or the maxillary body andit is not clear that attempts of different workers to measurea precise point of origin above the alveolar margin can be con-sidered comparable (compare, for example, values in Andrews[1978], Teaford et al. [1988], and Kunimatsu et al. [2004] forProconsul ) or that any attempt is particularly meaningful.This issue was recognized by Kunimatsu et al., (2004) who fo-cused more on the orientation of the zygomaticoalveolar crest(inferior border of the process). They correctly noted that theorientation in M16649 is lateral, as in early Miocene genera,whereas it is superolateral in KNM-FT 46a, a condition theyliken to that of extant great apes. In fact, the extant greatapes vary in this respect and both orientations can be observedwithin Pan. A more accurate, or at least more useful, perspec-tive is perhaps the height above the alveolar margin at whichthe lateral orientation is achieved rather than whether the ori-entation is lateral or superolateral, because in most Miocenetaxa, the crest eventually becomes more or less horizontal (lat-erally oriented) or more laterally than superiorly oriented. In
this respect, the K. wickeri and K. kizili type specimens differgreatly from M16649, with the latter clearly having an extremeexpression of the more primitive morphology expressed byearly Miocene species, as noted by Kunimatsu et al., (2004)and the two Kenyapithecus specimens more closely resemblinglater Miocene genera and extant apes (Table 4; Fig. 27; see fur-ther discussion below). In M16649, the zygomaticoalveolarcrest is fully horizontal only 9.0 mm superior to the alveolarmargin. At the same height in the K. kizili maxilla, the point atwhich the zygomatic process is broken, the crest has barelydiverged from the alveolar process. In KNM-FT 46a, the crestis still more superiorly than laterally oriented at the break,12.5 mm superior to the alveolar margin; in KNM-FT 47, thecrest is broken approximately 15 mm superior to the alveolarmargin and it is still more superiorly oriented.
The Kenyapithecus and Equatorius maxillae also differgreatly in the orientation of the canine alveolus and the spatialproximity of the alveolus to the maxillary sinus, with a directinfluence on the development of the canine fossa. In L1620,the anterior margin of the sinus floor is separated from the ca-nine alveolus by about 17 mm. In KNM-FT 46a, this distanceis just over 11 mm. The equivalent distance in M16649 is onlyabout 4 mm; it would have been even less above the level ofthe sinus floor given the strong posterior inclination of the ca-nine alveolus. In the two Kenyapithecus specimens, the canineroot is more nearly vertical in lateral view and inclined medi-ally. These spatial relationships in large part account for thevery deep canine fossae in the latter, whereas the Equatoriusmaxilla appears to have either a very shallow fossa or tolack a fossa altogether (breakage makes it difficult to ascertainwhich), again resembling most early Miocene forms. Lastly,the palatine process of the maxilla in M16649, at just over4 mm, is quite thin, as in early Miocene taxa; that of L1620is nearly twice as thick, relatively somewhat thicker eventhan in some late Miocene taxa such as Dryopithecus andSivapithecus.
Kenyapithecus and Equatorius also differ dentally (Wardet al., 1999; Kelley et al., 2002). Equatorius I1s, while mesio-distally broad, have slender marginal ridges offset from a low,rounded lingual tubercle. The lingual morphology of the lowerincisors of Equatorius is strikingly similar to that of Griphopi-thecus alpani, with very muted central ridges and equallymuted marginal ridges that join in a U-shaped cingulum (Pick-ford, 1985; Ward et al., 1999; Kelley et al., 2000). A lower ca-nine of Equatorius from Kipsaramon is relatively low-crownedand stout like those of Proconsul, and especially Afropithecusand Nacholapithecus (Table 3). In contrast, Benefit andMcCrossin (2000) provided measurements for one male lowercanine from Maboko, KNM-MB 32348, that has a height/length index similar to those of Kenyapithecus (Table 3). How-ever, until the Maboko material is fully accessible and themorphology and taxonomy can be independently evaluated,the significance of this canine is unclear (see Kelley et al.,2002). While molar morphology is broadly similar in thetwo genera, Pickford and Ishida (1998) noted that the majorityof Equatorius upper molars from Maboko have a more exten-sive protocone cingulum. Finally, Equatorius P3s, like those of

475J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
most other taxa, have a disproportionately large paracone interms of occlusal area that is separated from the protoconeby a deep fissure, while the P3s lack the extensive lingual cin-gulum present in Kenyapithecus (Kelley et al., 2002).
The morphology of the mandibular symphysis is very sim-ilar in Kenyapithecus (KNM-FT 45) and Equatorius (KNM-MJ 5; KNM-MB 20573); in both, the symphysis is stronglyproclined and the inferior torus is robust, although it projectsmore posteriorly in the Kenyapithecus mandible (Andrews,1971; Pickford, 1982; McCrossin and Benefit, 1993, 1997).Benefit and McCrossin (2000) claimed that this morphologyis unique to these specimens among Miocene apes, whichforms part of their justification for rejecting the validity ofEquatorius. However, a mandible of Griphopithecus fromPasxalar, G1313 (Alpagut et al., 1990), has a symphysis thatis intermediate in orientation (long-axis orientation of 36� rel-ative to the alveolar margin) between that of KNM-FT 45(32�) and KNM-MB 20573 (38�), a robust inferior torus,and a sublingual plane that is even somewhat shallower thanthat of the latter. This general morphology therefore appearsto be common among middle Miocene apes.
A partial skeleton of Equatorius from Kipsaramon pre-serves a humerus with a distal end that has suffered some dam-age and distortion (Ward et al., 1999; Sherwood et al., 2002).In spite of this, it is clear that Equatorius shares with Kenya-pithecus a truncated and posteriorly oriented medial epicon-dyle. They differ, however, in the depth of the coronoidfossa, with the Equatorius specimen having a shallow fossaas in many early Miocene taxa, in contrast to the deeper fossain KNM-FT 2751 (Rose, 1997).
It has been argued that the dental and gnathic differencesbetween K. wickeri and E. africanus can be accommodatedby the levels of interspecific variation typically expressed inindividual hominoid genera (McCrossin and Benefit, 1997;Benefit and McCrossin, 2000), which might be true in termsof a simple feature-by-feature comparison. What this ignores,however, is that one of these species, E. africanus, expressesa consistent suite of features that is more typical of primitiveearly Miocene genera while the other does not. More impor-tantly, this argument is no longer confined to just these twospecies. There is now a second species, K. kizili, that is nearlyidentical to, or very similar to, K. wickeri in every aspect of itsmore derived morphology, indicating cladistic affinity thatdoes not extend to Equatorius.
Late Miocene genera. Most, perhaps all, late Miocene hom-inoids uniformly differ from Kenyapithecus in upper centralincisor and upper premolar morphology. No late Miocene spe-cies for which I1 morphology is known shows the greatly in-flated lingual marginal ridges typical of Kenyapithecus. Inaddition, Lufengpithecus, Dryopithecus (except D. laietanus),Ouranopithecus, and Oreopithecus all have prominent, high-relief lingual pillars similar to those of G. alpani. Themorphology of the I1 lingual surface is unknown in Ankarapi-thecus. The upper premolars in most late Miocene hominoidsare basically like those of most early/middle Miocene taxa,with an expanded paracone area separated from the protoconeby a deep fissure; none of these taxa has the subequal cusp
areas and continuous, elevated transverse crest that bisectsthe crown’s occlusal surface buccolingually as in K. kizili (es-pecially in P3), and, in a less extreme fashion, in the one upperpremolar of K. wickeri. Lufengpithecus also differs from Ken-yapithecus in having a highly derived molar morphology, withmuted cusps, expansive occlusal basins, and complexly crenu-lated enamel. Oreopithecus has a highly derived dentition thatdiffers from that of Kenyapithecus in a number of featuresother than I1 morphology. The upper premolars have tall,closely approximated cups separated by a deep fissure anda distinct lingual cingulum. The upper molars have tall, volu-minous cusps, accessory crests and conules, and a prominentmesiolingual cingulum (Harrison and Rook, 1997).
Two late Miocene species are known only by their holo-types, Samburupithecus kiptalami from Kenya, representedby a partial maxilla (Ishida and Pickford, 1997; Pickford andIshida, 1998), and Khoratpithecus piriyai from Thailand, repre-sented by a relatively complete mandible (Chaimanee et al.,2004). Samburupithecus differs from Kenyapithecus in a num-ber of features of gnathic and dental morphology. It has a rela-tively low origin of the zygomatic process, as in early Miocenetaxa (although it cannot be determined at what level the zygo-maticoalveolar crest would have become more laterally thansuperiorly oriented), and the zygomatic root is excavated bythe maxillary sinus. The upper molars are characterized bygreatly inflated cusps, a deep fissure that effectively eliminatesthe crista obliqua (giving the teeth an almost suidlike appear-ance), high dentine penetrance, and a prominent cingulumthat extends around the protocone. Khoratpithecus piriyai,has a male canine that is relatively low-crowned in relationto maximum cervical length and only a faint lingual cingulum(Chaimanee et al., 2004), a P3 that lacks a cingulum, and lowermolars with higher dentine penetrance than in Kenyapithecus.
Distal humeral morphology is known for Sivapithecus, Dry-opithecus brancoi from Hungary, and Oreopithecus. All sharea more derived morphology than that of Kenyapithecus, witha deeper coronoid fossa, a more incised zona conoidea, a troch-leiform trochlea with the medial lip more prominent in distalview, and a more sharply defined lateral trochlear lip (Morbeck,1983; Rose, 1997). In addition, all of these taxa have a medialepicondyle that is more medially than posteriorly oriented, incontrast to the highly derived, more posteriorly directedepicondyle of Kenyapithecus, although only in Oreopithecusis it as medially oriented as in extant great apes (Rose, 1997;McCrossin et al., 1998). Dryopithecus brancoi also differs inhaving a shallow olecranon fossa with minimal developmentof articular surface on the lateral wall of the fossa, and a medi-olaterally narrow region lateral to the olecranon fossa (Rose,1997).
All late Miocene genera differ from Kenyapithecus in a vari-ety of other features as well. Sivapithecus maxillary morphologydiffers from that of Kenyapithecus in a number of ways. Sivapi-thecus has an elongate nasoalveolar clivus with a greatly re-duced incisive fossa leading into a long, narrow incisive canal(Ward and Kimbel, 1983). While the height above the alveolarmargin at which the zygomaticoalveolar crest becomes horizon-tal is not precisely known for either species of Kenyapithecus, it

Table 5
Index of upper central incisor crown height/crown length for K. kizili and
K. wickeri
Specimen Index*
K. kizili
BP27 1.30
B454 1.17
D298 1.25
D655 1.25
H1641 1.21
K1356 1.08
N1390 1.14
R1678 1.21
K. wickeri
KNM-FT 49 1.03
Index¼mesiodistal length/labial crown height.
476 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
is unlikely to have been nearly as high as in Sivapithecusd19 to27 mm in the three specimens in which the zygomatic process iscomplete to this point (Table 4). In several other specimens, theprocess is broken anywhere between 18 and 27 mm superior tothe alveolar margin without the crest having attained a mostlyhorizontal orientation. The palate of Sivapithecus is very deep,as in extant apes (Pilbeam, 1982). The maxillary sinus typicallyextends into the premolar area. Likewise, invasion of the alveo-lar process by the sinus is extensive in most Sivapithecus males(contra Begun and Gulec, 1998), with the formation of complexpatterns of septa and loculi (Ward and Pilbeam, 1983); the lowestrecess of the sinus floor is typically less than 10 mm superior tothe alveolar margin. In two specimens, however, one female(YPM 13799; Ward and Pilbeam, 1983) and one male (GSP11704), the sinus floor is quite flat and consistently elevatedabove the molar root apicesdapproximately 15 and 17 mm su-perior to the alveolar margin, respectively. Expansion of the si-nus into the zygomatic root is also variable. In some specimens,the zygomatic root is a solid strut of bone as in Kenyapithecus. Inothers, the sinus either completely occupies the root so that thezygomaticoalveolar crest consists of a relatively thin plate ofbone (GSP 15000; Ward and Brown, 1986), or it only partlydoes so (GSP 9977; JK, personal observation).
Ouranopithecus and Ankarapithecus also have a more de-rived maxillary morphology than Kenyapithecus. Both havea relatively labiolingually thin, elongate and procumbent na-soalveolar clivus (de Bonis and Melentis, 1987; Begun andGulec, 1998). Ankarapithecus also has an elevated zygomaticroot and zygomaticoalveolar crest that becomes horizontalwell above the alveolar margin (Table 4), as well as an exten-sive maxillary sinus that extends into the posterior premolarregion, but with an uncomplicated floor positioned above themolar roots (Andrews and Tekkaya, 1980; Begun and Gulec,1998). It also has a highly procumbent midfacial area, result-ing in a substantial distance between the maxillary sinus andthe canine alveolus, 17 mm to the apex of the canine rootand greater at the level of the sinus floor (Table 4; Begunand Gulec, 1998).
Dryopithecus species from western and central Europe havedistinctly different maxillary morphologies. The maxilla of D.brancoi is strikingly similar to that of Kenyapithecus in mostrespects (Kretzoi, 1975; Kelley and Pilbeam, 1986; Begun,1994), except that the maxillary sinus extends well into thepremolar region (Kordos and Begun, 2001). Also, it appearsto be similar to other late Miocene hominoids in having a zy-gomaticoalveolar crest that becomes laterally directed (hori-zontal) well above the alveolar margin (Table 4). In RUD12, the crest is only just beginning to turn out laterally9 mm superior to the alveolar margin and is still not nearinga horizontal orientation approximately 15 mm superior at itsbroken end. The same measurements for the larger, presum-ably male RUD 7 are 17 and 20 mm. In another female,RUD 200, the crest becomes horizontal at about 20 mm abovethe alveolar margin.
Maxillary morphology in D. laietanus, as representedby CLl 18000 from Can Llobateres, is more similar to thatof extant African apes in many respects, and it differs from
that of Kenyapithecus in several features. The sinus floor pen-etrates somewhat between the molar roots, creating low septa.In the vicinity of M2, the floor of the sinus is a mere 7 mm su-perior to the alveolar margin. The floor does not extend ante-rior to M1, but it is nevertheless no more than 9 mm posteriorto the canine alveolus (Table 4). However, the sinus penetratesinto the premolar region more superiorly (Moya-Sola andKohler, 1995) and is only a few millimeters posterior to the ca-nine root at its apex. It also invades the zygomatic root. Asa consequence of these relationships, and combined with a pos-teriorly inclined canine root, there is little development of a ca-nine fossa. Finally, the origin of the zygomatic process is highabove the alveolar margin and the zygomaticoalveolar crestapproaches a horizontal orientation approximately 24 mmabove the margin (Table 4).
Comparison of K. kizili and K.wickeri
Specimens of K. wickeri preserve the morphology of themandibular body and symphysis, the maxillary body, and allteeth except the lower incisors, female lower canine, P3, M1,and M3, with all the teeth known from only one or two spec-imens. There are no mandibular specimens known for K. kizili.Moreover, as noted, it has not been possible so far to confi-dently distinguish the molars from those of G. alpani, althoughM1 and M2 morphology can be characterized for K. kizili, asthey are present in the type specimen. Thus, comparisons be-tween the two species of Kenyapithecus are limited to themaxilla (in which they are virtually identical, as described),M1 and M2, and the anterior and premolar teeth known fromboth.
The central incisors of the two Kenyapithecus species differin a number of details. Those of K. kizili are higher-crowned inrelation to mesiodistal length, in spite of all being slightly worn,whereas the one K. wickeri I1 is an unworn germ (Table 5).Kenyapithecus kizili I1s also have a distinct V-shaped junctionof the marginal ridges lingually, which contrasts with thebroad, U-shaped marginal ridges and cingulum of the K. wick-eri I1. Finally, in the K. wickeri incisor, the lingual marginalridges have higher relief than the central area that they bound,
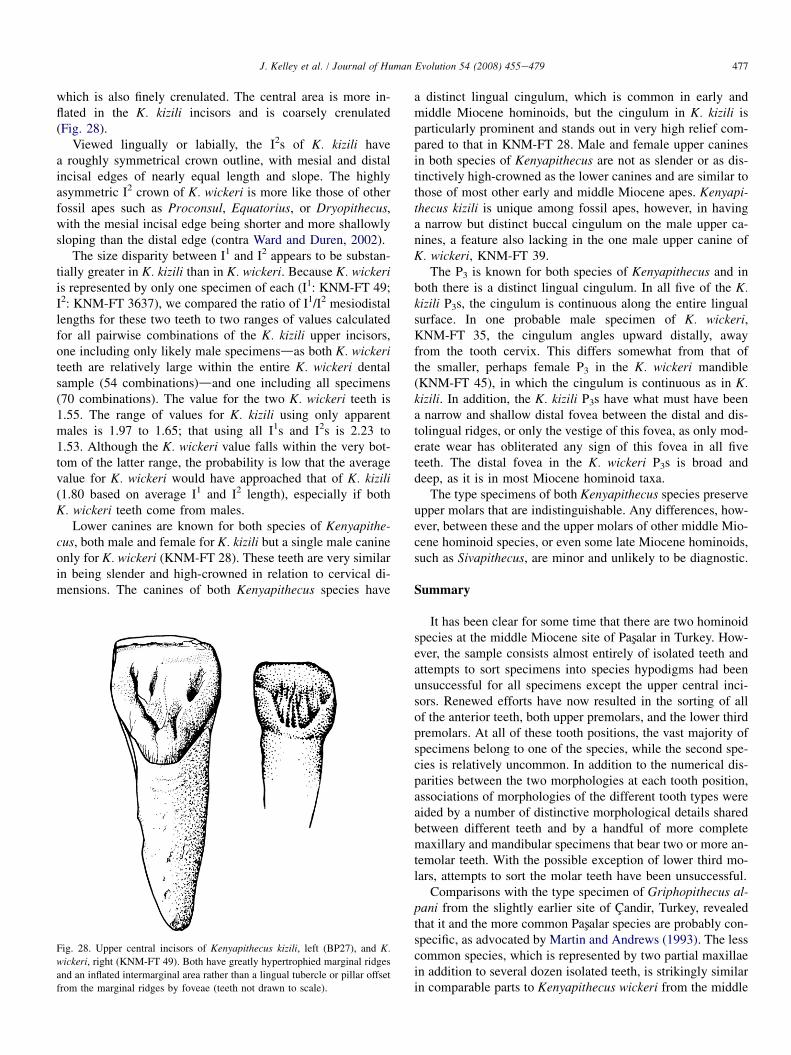
477J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
which is also finely crenulated. The central area is more in-flated in the K. kizili incisors and is coarsely crenulated(Fig. 28).
Viewed lingually or labially, the I2s of K. kizili havea roughly symmetrical crown outline, with mesial and distalincisal edges of nearly equal length and slope. The highlyasymmetric I2 crown of K. wickeri is more like those of otherfossil apes such as Proconsul, Equatorius, or Dryopithecus,with the mesial incisal edge being shorter and more shallowlysloping than the distal edge (contra Ward and Duren, 2002).
The size disparity between I1 and I2 appears to be substan-tially greater in K. kizili than in K. wickeri. Because K. wickeriis represented by only one specimen of each (I1: KNM-FT 49;I2: KNM-FT 3637), we compared the ratio of I1/I2 mesiodistallengths for these two teeth to two ranges of values calculatedfor all pairwise combinations of the K. kizili upper incisors,one including only likely male specimensdas both K. wickeriteeth are relatively large within the entire K. wickeri dentalsample (54 combinations)dand one including all specimens(70 combinations). The value for the two K. wickeri teeth is1.55. The range of values for K. kizili using only apparentmales is 1.97 to 1.65; that using all I1s and I2s is 2.23 to1.53. Although the K. wickeri value falls within the very bot-tom of the latter range, the probability is low that the averagevalue for K. wickeri would have approached that of K. kizili(1.80 based on average I1 and I2 length), especially if bothK. wickeri teeth come from males.
Lower canines are known for both species of Kenyapithe-cus, both male and female for K. kizili but a single male canineonly for K. wickeri (KNM-FT 28). These teeth are very similarin being slender and high-crowned in relation to cervical di-mensions. The canines of both Kenyapithecus species have
Fig. 28. Upper central incisors of Kenyapithecus kizili, left (BP27), and K.
wickeri, right (KNM-FT 49). Both have greatly hypertrophied marginal ridges
and an inflated intermarginal area rather than a lingual tubercle or pillar offset
from the marginal ridges by foveae (teeth not drawn to scale).
a distinct lingual cingulum, which is common in early andmiddle Miocene hominoids, but the cingulum in K. kizili isparticularly prominent and stands out in very high relief com-pared to that in KNM-FT 28. Male and female upper caninesin both species of Kenyapithecus are not as slender or as dis-tinctively high-crowned as the lower canines and are similar tothose of most other early and middle Miocene apes. Kenyapi-thecus kizili is unique among fossil apes, however, in havinga narrow but distinct buccal cingulum on the male upper ca-nines, a feature also lacking in the one male upper canine ofK. wickeri, KNM-FT 39.
The P3 is known for both species of Kenyapithecus and inboth there is a distinct lingual cingulum. In all five of the K.kizili P3s, the cingulum is continuous along the entire lingualsurface. In one probable male specimen of K. wickeri,KNM-FT 35, the cingulum angles upward distally, awayfrom the tooth cervix. This differs somewhat from that ofthe smaller, perhaps female P3 in the K. wickeri mandible(KNM-FT 45), in which the cingulum is continuous as in K.kizili. In addition, the K. kizili P3s have what must have beena narrow and shallow distal fovea between the distal and dis-tolingual ridges, or only the vestige of this fovea, as only mod-erate wear has obliterated any sign of this fovea in all fiveteeth. The distal fovea in the K. wickeri P3s is broad anddeep, as it is in most Miocene hominoid taxa.
The type specimens of both Kenyapithecus species preserveupper molars that are indistinguishable. Any differences, how-ever, between these and the upper molars of other middle Mio-cene hominoid species, or even some late Miocene hominoids,such as Sivapithecus, are minor and unlikely to be diagnostic.
Summary
It has been clear for some time that there are two hominoidspecies at the middle Miocene site of Pasxalar in Turkey. How-ever, the sample consists almost entirely of isolated teeth andattempts to sort specimens into species hypodigms had beenunsuccessful for all specimens except the upper central inci-sors. Renewed efforts have now resulted in the sorting of allof the anterior teeth, both upper premolars, and the lower thirdpremolars. At all of these tooth positions, the vast majority ofspecimens belong to one of the species, while the second spe-cies is relatively uncommon. In addition to the numerical dis-parities between the two morphologies at each tooth position,associations of morphologies of the different tooth types wereaided by a number of distinctive morphological details sharedbetween different teeth and by a handful of more completemaxillary and mandibular specimens that bear two or more an-temolar teeth. With the possible exception of lower third mo-lars, attempts to sort the molar teeth have been unsuccessful.
Comparisons with the type specimen of Griphopithecus al-pani from the slightly earlier site of Candir, Turkey, revealedthat it and the more common Pasxalar species are probably con-specific, as advocated by Martin and Andrews (1993). The lesscommon species, which is represented by two partial maxillaein addition to several dozen isolated teeth, is strikingly similarin comparable parts to Kenyapithecus wickeri from the middle

478 J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
Miocene site of Fort Ternan in Kenya. The new species istherefore identified as a new species of Kenyapithecus, K. ki-zili. In our present state of knowledge, and given current dis-agreements regarding the suprageneric taxonomy of Miocenehominoids, it is not possible to assign this genus to any family-group taxon.
The presence of a second species that is similar in almostall respects to the type species of Kenyapithecus, K. wickeri,affirms the distinctiveness of the genus. More particularly, itprovides additional support for removing from Kenyapithecusfossils from Maboko and nearby sites that display an overallmore primitive morphology, and assigning them instead toEquatorius (Ward et al., 1999; Kelley et al., 2000).
Acknowledgements
We express our gratitude to all of those who have workedat the Pasxalar site over the years, in particular the many Turk-ish students who have participated. We also acknowledge theconsiderable efforts of Lawrence Martin, both in the field andfor his earlier contributions toward documenting the existenceof a second hominoid species at Pasxalar. We thank Terry Har-rison and especially Eric Delson for conversations regardingthe Pasxalar material and for comments on an earlier draft ofthis manuscript. The Pasxalar excavations were carried out onbehalf of the T.C. Ministry of Culture and Tourism, Director-ate of Antiquities, and the University of Ankara, Faculty ofLanguage, History and Geography. Excavations were fundedby the Directorate of Antiquities and the L.S.B. Leakey Foun-dation for Anthropological Research. JK was supported by theNational Science Foundation (SBR 9408664).
References
Alpagut, B., Andrews, P., Fortelius, M., Kappelman, J., Temizsoy, I.,
Celebi, H., Lindsay, W., 1996. A new specimen of Ankarapithecus meteai
from the Sinap Formation of central Anatolia. Nature 382, 349e351.
Alpagut, B., Andrews, P., Martin, L., 1990. New hominoid specimens from the
middle Miocene site at Pasxalar, Turkey. J. Hum. Evol. 19, 397e422.
Andrews, P., 1971. Ramapithecus wickeri mandible from Fort Ternan, Kenya.
Nature 231, 192e194.
Andrews, P., 1974. New species of Dryopithecus from Kenya. Nature 249,
188e190.
Andrews, P., 1978. A revision of the Miocene Hominoidea of East Africa.
Bull. Br. Mus. Nat. Hist. (Geol.) 30, 85e224.
Andrews, P., Cronin, J., 1982. The relationships of Sivapithecus and Ramapi-
thecus and the evolution of the orang-utan. Nature 297, 541e546.
Andrews, P., Molleson, T., 1979. The provenance of Sivapithecus africanus.
Bull. Br. Mus. Nat. Hist. (Geol.) 32, 19e23.
Andrews, P., Tekkaya, I., 1980. A revision of the Turkish Miocene hominoid
Sivapithecus meteai. Palaeontology 23, 85e95.
Andrews, P., Tobien, H., 1977. New Miocene locality in Turkey with evidence
on the origin of Ramapithecus and Sivapithecus. Nature 268, 699e701.
Andrews, P., Walker, A., 1976. The primate and other fauna from Fort Ternan,
Kenya. In: Isaac, G.L., McCown, E.R. (Eds.), Human Origins: Louis
Leakey and the East African Evidence. W.A. Benjamin, Menlo Park, pp.
279e304.
Begun, D.R., 1994. Relations among the great apes and humans: new interpre-
tations based on the fossil great ape Dryopithecus. Yearb. Phys. Anthropol.
37, 11e63.
Begun, D.R., 2000. Middle Miocene hominoid origins. Science 287, 2375a.
http://www.sciencemag.com/cgi/content/full/287/5462/2375a.
Begun, D.R., Gulec, E., 1998. Restoration of the type and palate of Ankarapi-
thecus meteai: Taxonomic and phylogenetic implications. Am. J. Phys.
Anthropol. 105, 279e314.
Begun, D.R., Geraads, D., Gulec, E., 2003. The Candir hominoid locality: im-
plications for the timing and pattern of hominoid dispersal events. Cour.
Forsch. Inst. Senckenberg 240, 251e265.
Benefit, B.R., McCrossin, M.L., 2000. Middle Miocene hominoid origins.
Science 287, 2375a. http://www.sciencemag.com/cgi/content/full/287/
5462/2375a.
Bishop, W.W., Miller, J.A., Fitch, F.J., 1969. New potassium-argon age deter-
minations relevant to the Miocene fossil mammal sequence in East Africa.
Am. J. Sci. 267, 669e699.
Brown, B., 1997. Miocene hominoid mandibles: Functional and phylogenetic
perspectives. In: Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function,
Phylogeny, and Fossils: Miocene Hominoid Evolution and Adaptations.
Plenum Press, New York, pp. 153e171.
Chaimanee, Y., Jolly, D., Benammi, M., Tafforeau, P., Duzer, D., Moussa, I.,
Jaeger, J.-J., 2003. A middle Miocene hominoid from Thailand and orang-
utan origins. Nature 422, 61e65.
Chaimanee, Y., Suteethorn, V., Jintasakul, P., Vidthayanon, C., Marandat, B.,
Jaeger, J.-J., 2004. A new orang-utan relative from the late Miocene of
Thailand. Nature 427, 439e441.
de Bonis, L., Koufos, G., 1993. The face and mandible of Ouranopithecus
macedoniensis: description of new specimens and comparisons. J. Hum.
Evol. 24, 469e491.
de Bonis, L., Melentis, J., 1987. Interet de l’anatomie naso-maxillaire pour
l’phylogenie de Hominoidea. C.R. Acad. Sci. Paris 304, 767e769.
Gebo, D.L., MacLatchy, L., Kityo, R., Deino, A., Kingston, J., Pilbeam, D.,
1997. A hominoid genus from the early Miocene of Uganda. Science
276, 401e404.
Gencturk, I., Alpagut, B., Andrews, P., 2008. Interproximal wear facets and
tooth associations in the Pasxalar hominoid sample. J. Hum. Evol. 54 (4),
480e493.
Gulec, E., Begun, D.R., 2003. Functional morphology and affinities of the hom-
inoid mandible from Candir. Cour. Forsch.-Inst. Senckenberg 240, 89e111.
Harrison, T., 1992. A reassessment of the taxonomic and phylogenetic
affinities of the fossil catarrhines from Fort Ternan, Kenya. Primates 33,
501e522.
Harrison, T., Rook, L., 1997. Enigmatic anthropoid or misunderstood ape? The
phylogenetic status of Oreopithecus bambolii reconsidered. In:
Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function, Phylogeny, and
Fossils: Miocene Hominoid Evolution and Adaptations. Plenum Press,
New York, pp. 327e362.
Hartwig, W.C. (Ed.), 2002. The Primate Fossil Record. Cambridge University
Press, Cambridge.
Ishida, H., Kunimatsu, Y., Takano, T., Nakano, Y., Nakatsukasa, M., 2004.
Nacholapithecus skeleton from the middle Miocene of Kenya. J. Hum.
Evol. 46, 69e103.
Ishida, H., Pickford, M., 1997. A new late Miocene hominoid from Kenya:
Samburupithecus kiptalami gen. et sp. nov. C.R. Acad. Sci., Paris 325,
823e829.
Kelley, J., 1986. Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus. J. Hum. Evol. 15, 461e495.
Kelley, J., 1995a. Sexual dimorphism in canine shape among extant great apes.
Am. J. Phys. Anthropol. 96, 365e389.
Kelley, J., 1995b. Sex determination in Miocene catarrhine primates. Am. J.
Phys. Anthropol. 96, 390e417.
Kelley, J., 2008. Identification of a single birth cohort in Kenyapithecus kiziliand the nature of sympatry between K. kizili and Griphopithecus alpani at
Pasxalar. J. Hum. Evol. 54 (4), 530e537.
Kelley, J., Alpagut, B., 1999. Canine sexing and species number in the Pasxalar
large hominoid sample. J. Hum. Evol. 36, 335e341.
Kelley, J., Pilbeam, D., 1986. The dryopithecines: taxonomy, comparative
anatomy and phylogeny of Miocene large hominoids. In: Swindler, D.R.,
Erwin, J. (Eds.), Comparative Primate Biology. Systematics, Evolution
and Anatomy, vol. 1. Alan R. Liss, New York, pp. 361e411.

479J. Kelley et al. / Journal of Human Evolution 54 (2008) 455e479
Kelley, J., Plavcan, J.M., 1998. A simulation test of hominoid species number
at Lufeng, China: Implications for the use of the coefficient of variation in
paleotaxonomy. J. Hum. Evol. 35, 577e596.
Kelley, J., Ward, S., Brown, B., Hill, A., Downs, W., 2000. Middle Miocene
hominoid origins: response. Science 287, 2375a. http://www.sciencemag.
com/cgi/content/full/287/5462/2375a.
Kelley, J., Ward, S., Brown, B., Hill, A., Duren, D., 2002. Dental remains of
Equatorius africanus from Kipsaramon, Tugen Hills, Baringo District,
Kenya. J. Hum. Evol. 42, 39e62.
Kordos, L., Begun, D.R., 2001. A new cranium of Dryopithecus from Rudaba-
nya, Hungary. J. Hum. Evol. 41, 689e700.
Kretzoi, M., 1975. New ramapithecines and Pliopithecus from the lower
Pliocene of Rudabanya in northeastern Hungary. Nature 257, 578e581.
Kunimatsu, Y., Ishida, H., Nakatsukasa, M., Nakano, Y., Sawada, Y.,
Nakayama, K., 2004. Maxillae and associated gnathodental specimens of
Nacholapithecus kerioi, a large-bodied hominoid from Nachola, northern
Kenya. J. Hum. Evol. 46, 365e400.
Leakey, L.S.B., 1962. A new lower Pliocene fossil primate from Kenya. Ann.
Mag. Nat. Hist. 4, 689e696.
Leakey, L.S.B., 1967. An early Miocene member of Hominidae. Nature 213,
155e163.
Leakey, L.S.B., 1968. Lower dentition of Kenyapithecus africanus. Nature
217, 827e830.
Leakey, R.E., Leakey, M.G., Walker, A.C., 1988. Morphology of Afropithecus
turkanensis from Kenya. Am. J. Phys. Anthropol. 76, 289e307.
Leakey, M.G., Walker, A.C., 1997. Afropithecus: function and phylogeny. In:
Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function, Phylogeny, and
Fossils: Miocene Hominoid Evolution and Adaptations. Plenum Press,
New York, pp. 225e239.
Le Gros Clark, W.E., Leakey, L.S.B., 1951. Fossil Mammals of Africa, Vol. 1:
The Miocene Hominoidea of East Africa. British Museum (Natural
History), London.
Martin, L., Andrews, P., 1993. Species recognition in middle Miocene homi-
noids. In: Kimbel, W.H., Martin, L.B. (Eds.), Species, Species Concepts,
and Primate Evolution. Plenum Press, New York, pp. 393e427.
McCollum, M.A., Ward, S.C., 2000. Subnasoalveolar anatomy and hominoid
phylogeny: evidence from comparative ontogeny. Am. J. Phys. Anthropol.
102, 377e405.
McCrossin, M.L., Benefit, B.R., 1993. Recently recovered Kenyapithecus
mandible and its implications for great ape and human origins. Proc.
Natl. Acad. Sci. U. S. A. 90, 1962e1966.
McCrossin, M.L., Benefit, B.R., 1994. Maboko Island and the evolutionary his-
tory of Old World monkeys and apes. In: Corruccini, R.S., Ciochon, R.L.
(Eds.), Integrative Paths to the Past: Paleoanthropological Advances in
Honor of F.C. Howell. Prentice-Hall, Englewood Cliffs, pp. 95e122.
McCrossin, M.L., Benefit, B.R., 1997. On the relationships and adaptations of
Kenyapithecus, a large-bodied hominoid from the middle Miocene of east-
ern Africa. In: Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function, Phy-
logeny, and Fossils: Miocene Hominoid Evolution and Adaptations.
Plenum Press, New York, pp. 241e267.
McCrossin, M.L., Benefit, B.R., Gitau, S.N., Palmer, A.K., Blue, K.T., 1998.
Fossil evidence for the origins of terrestriality among Old World higher
primates. In: Strasser, E., Fleagle, J., Rosenberger, A., McHenry, H.
(Eds.), Primate Locomotion: Recent Advances. Plenum Press, New
York, pp. 353e396.
Morbeck, M.E., 1983. Miocene hominoid discoveries from Rudabanya: Impli-
cations from the postcranial skeleton. In: Ciochon, R.L., Corruccini, R.S.
(Eds.), New Interpretations of Ape and Human Ancestry. Plenum Press,
New York, pp. 369e404.
Moya-Sola, S., Kohler, M., 1995. New partial cranium of Dryopithecus Lartet
1863 (Hominoidea, Primates) from the upper Miocene of Can Llobateres,
Barcelona, Spain. J. Hum. Evol. 29, 109e139.
Moya-Sola, S., Kohler, M., Alba, D.M., Casanovas-Vilar, I., Galindo, J., 2004.
Pierolapithecus catalaunicus, a new middle Miocene great ape from Spain.
Science 306, 1339e1344.
Pickford, M., 1982. New higher primate fossils from the middle Miocene
deposits at Majiwa and Kaloma, western Kenya. Am. J. Phys. Anthropol.
58, 1e19.
Pickford, M., 1985. A new look at Kenyapithecus based on recent discoveries
in western Kenya. J. Hum. Evol. 14, 113e143.
Pickford, M., Ishida, H., 1998. Interpretation of Samburupithecus, an
upper Miocene hominoid from Kenya. C.R. Acad. Sci. Paris 326,
299e306.
Pickford, M., Sawada, Y., Tayama, R., Matsuda, Y., Itaya, T., Hyodo, H.,
Senut, B., 2006. Refinement of the age of the middle Miocene Fort Ternan
Beds, western Kenya, and its implications for Old World biochronology.
C.R. Geoscience 338, 545e555.
Pilbeam, D.R., 1969. Tertiary Pongidae of East Africa: evolutionary relation-
ships and taxonomy. Bull. Peabody Mus. Nat. Hist. 31, 1e185.
Pilbeam, D., 1982. New hominoid skull material from the Miocene of Paki-
stan. Nature 295, 232e234.
Pilbrow, V., 2006. Lingual incisor traits in modern hominoids and an assess-
ment of their utility for fossil hominoid taxonomy. Am. J. Phys. Anthropol.
129, 323e338.
Rose, M.D., 1997. Functional and phylogenetic features of the forelimb in
Miocene hominoids. In: Begun, D.R., Ward, C.V., Rose, M.D. (Eds.),
Function, Phylogeny, and Fossils: Miocene Hominoid Evolution and Ad-
aptations. Plenum Press, New York, pp. 79e100.
Senut, B., 1986. New data on Miocene hominoid humeri from Pakistan and
Kenya. In: Else, J.G., Lee, P.C. (Eds.), Proceedings of the 10th Congress
of the International Primatological Society, vol. 1. Cambridge University
Press, Cambridge, pp. 151e161.
Sherwood, R.J., Ward, S., Hill, A., Duren, D.L., Brown, B., Downs, W., 2002.
Preliminary description of the Equatorius africanus partial skeleton
(KNM-TH 28860) from Kipsaramon, Tugen Hills, Baringo District,
Kenya. J. Hum. Evol. 42, 63e73.
Shipman, P., Walker, A., Van Couvering, J.A.H., Hooker, P.J., Miller, J.A.,
1981. The Fort Ternan hominoid site, Kenya: Geology, age, taphonomy
and palaeoecology. J. Hum. Evol. 10, 49e72.
Teaford, M.F., Beard, K.C., Leakey, R.E., Walker, A.C., 1988. New hominoid
facial skeleton from the early Miocene of Rusinga Island, Kenya, and its
bearing on the relationship between Proconsul nyanzae and Proconsul afri-
canus. J. Hum. Evol. 17, 461e477.
Tekkaya, I., 1974. A new species of Tortonian anthropoid (Primates, Mammalia)
from Anatolia. Bull. Miner. Res. Explor. Inst. Ankara 83, 148e165.
Waddle, D., Martin, L., Stock, D., 1995. Sexing isolated hominoid canines
with special reference to the middle Miocene specimens from Pasxalar, Tur-
key. J. Hum. Evol. 28, 385e403.
Ward, S.C., Brown, B., 1986. The facial skeleton of Sivapithecus indicus.
In: Swindler, D.R., Erwin, J. (Eds.), Comparative Primate Biology. Sys-
tematics, Evolution and Anatomy, vol. 1. Alan R. Liss, New York, pp.
413e452.
Ward, S.C., Brown, B., Hill, A., Kelley, J., Downs, W., 1999. Equatorius: A
new hominoid genus from the middle Miocene of Kenya. Science 285,
1382e1386.
Ward, S.C., Duren, D.L., 2002. Middle and late Miocene African hominoids.
In: Hartwig, W. (Ed.), The Primate Fossil Record. Cambridge University
Press, Cambridge, pp. 385e397.
Ward, S.C., Pilbeam, D., 1983. Maxillofacial morphology of Miocene homi-
noids from Africa and Indo-Pakistan. In: Ciochon, R.L., Corruccini, R.S.
(Eds.), New Interpretations of Ape and Human Ancestry. Plenum Press,
New York, pp. 211e238.
Ward, S.C., Kimbel, W.H., 1983. Subnasal morphology and the systematic
position of Sivapithecus. Am. J. Phys. Anthropol. 61, 157e171.
Related Documents











