ORIGINAL ARTICLE A naturally occurring rare analog of quercetin promotes peak bone mass achievement and exerts anabolic effect on osteoporotic bone J. A. Siddiqui & G. Swarnkar & K. Sharan & B. Chakravarti & A. K. Gautam & P. Rawat & M. Kumar & V. Gupta & L. Manickavasagam & A. K. Dwivedi & R. Maurya & N. Chattopadhyay # International Osteoporosis Foundation and National Osteoporosis Foundation 2011 Abstract Summary The effect of quercetin C-glucoside (QCG) on osteoblast function in vitro and bone formation in vivo was investigated. QCG supplementation promoted peak bone mass achievement in growing rats and new bone formation in osteopenic rats. QCG has substantial oral bioavailability. Findings suggest a significant bone anabolic effect of QCG. Introduction Recently, we showed that extracts of Ulmus wallichiana promoted peak bone mass achievement in growing rats and preserved trabecular bone mass and cortical bone strength in ovariectomized (OVx) rats. 3,3′,4′,5,7-Pentahydroxyflavone-6-C-β-D-glucopyranoside, a QCG, is the most abundant bioactive compound of U. wallichiana extract. We hypothesize that QCG exerts bone anabolic effects by stimulating osteoblast function. Methods Osteoblast cultures were harvested from rat calvaria and bone marrow (BM) to study differentiation and mineralization. In vivo, growing female Sprague Dawley rats and OVx rats with osteopenia were adminis- tered QCG (5.0 or 10.0 mg kg -1 day -1 ) orally for 12 weeks. Efficacy was evaluated by examining changes in bone microarchitecture using histomorphometric and microcom- puted tomographic analyses and by determination of new bone formation by fluorescent labeling of bone. Plasma and BM levels of QCG were determined by high-performance liquid chromatography. Results QCG was much more potent than quercetin (Q) in stimulating osteoblast differentiation, and the effect of QCG was not mediated by estrogen receptors. In growing rats, QCG increased BM osteoprogenitors, bone mineral density, bone formation rate, and cortical deposition. In osteopenic rats, QCG treatment increased bone formation rate and improved trabecular microarchitecture. Comparison with the sham group (ovary intact) revealed significant restora- tion of trabecular bone in osteopenic rats treated with QCG. QCG levels in the BM were ~50% of that of the plasma levels. Conclusion QCG stimulated modeling-directed bone ac- crual and exerted anabolic effects on osteopenic rats by direct stimulatory effect on osteoprogenitors likely due to substantial QCG delivery at tissue level following oral administration. Keywords Estrogenicity . Flavonoid C-glucoside . Osteogenic . Peak bone mass J. A. Siddiqui and G. Swarnkar contributed equally to this study. Electronic supplementary material The online version of this article (doi:10.1007/s00198-010-1519-4) contains supplementary material, which is available to authorized users. J. A. Siddiqui : G. Swarnkar : K. Sharan : B. Chakravarti : A. K. Gautam : N. Chattopadhyay (*) Division of Endocrinology, Central Drug Research Institute (Council of Scientific and Industrial Research), Chattar Manzil, P.O. Box 173, Lucknow, India e-mail: [email protected] P. Rawat : M. Kumar : R. Maurya Division of Medicinal & Process Chemistry, Central Drug Research Institute (Council of Scientific and Industrial Research), Chattar Manzil, P.O. Box 173, Lucknow, India V. Gupta : A. K. Dwivedi Division of Pharmaceutics, Central Drug Research Institute (Council of Scientific and Industrial Research), Chattar Manzil, P.O. Box 173, Lucknow, India L. Manickavasagam Division of Pharmacokinetics and Metabolism, Central Drug Research Institute (Council of Scientific and Industrial Research), Chattar Manzil, P.O. Box 173, Lucknow, India Osteoporos Int (2011) 22:3013–3027 DOI 10.1007/s00198-010-1519-4 Received: 24 August 2010 / Accepted: 6 December 2010 / Published online: 12 January 2011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
A naturally occurring rare analog of quercetin promotespeak bone mass achievement and exerts anabolic effecton osteoporotic bone
J. A. Siddiqui & G. Swarnkar & K. Sharan & B. Chakravarti & A. K. Gautam & P. Rawat &M. Kumar & V. Gupta & L. Manickavasagam & A. K. Dwivedi & R. Maurya &
N. Chattopadhyay
# International Osteoporosis Foundation and National Osteoporosis Foundation 2011
AbstractSummary The effect of quercetin C-glucoside (QCG) onosteoblast function in vitro and bone formation in vivo wasinvestigated. QCG supplementation promoted peak bonemass achievement in growing rats and new bone formationin osteopenic rats. QCG has substantial oral bioavailability.Findings suggest a significant bone anabolic effect of QCG.Introduction Recently, we showed that extracts of Ulmuswallichiana promoted peak bone mass achievement ingrowing rats and preserved trabecular bone mass andcortical bone strength in ovariectomized (OVx) rats.
3,3′,4′,5,7-Pentahydroxyflavone-6-C-β-D-glucopyranoside,a QCG, is the most abundant bioactive compound of U.wallichiana extract. We hypothesize that QCG exerts boneanabolic effects by stimulating osteoblast function.Methods Osteoblast cultures were harvested from ratcalvaria and bone marrow (BM) to study differentiationand mineralization. In vivo, growing female SpragueDawley rats and OVx rats with osteopenia were adminis-tered QCG (5.0 or 10.0 mg kg−1 day−1) orally for 12 weeks.Efficacy was evaluated by examining changes in bonemicroarchitecture using histomorphometric and microcom-puted tomographic analyses and by determination of newbone formation by fluorescent labeling of bone. Plasma andBM levels of QCG were determined by high-performanceliquid chromatography.Results QCG was much more potent than quercetin (Q) instimulating osteoblast differentiation, and the effect of QCGwas not mediated by estrogen receptors. In growing rats,QCG increased BM osteoprogenitors, bone mineral density,bone formation rate, and cortical deposition. In osteopenicrats, QCG treatment increased bone formation rate andimproved trabecular microarchitecture. Comparison withthe sham group (ovary intact) revealed significant restora-tion of trabecular bone in osteopenic rats treated with QCG.QCG levels in the BM were ~50% of that of the plasmalevels.Conclusion QCG stimulated modeling-directed bone ac-crual and exerted anabolic effects on osteopenic rats bydirect stimulatory effect on osteoprogenitors likely due tosubstantial QCG delivery at tissue level following oraladministration.
Keywords Estrogenicity . Flavonoid C-glucoside .
Osteogenic . Peak bone mass
J. A. Siddiqui and G. Swarnkar contributed equally to this study.
Electronic supplementary material The online version of this article(doi:10.1007/s00198-010-1519-4) contains supplementary material,which is available to authorized users.
J. A. Siddiqui :G. Swarnkar :K. Sharan :B. Chakravarti :A. K. Gautam :N. Chattopadhyay (*)Division of Endocrinology, Central Drug Research Institute(Council of Scientific and Industrial Research),Chattar Manzil, P.O. Box 173, Lucknow, Indiae-mail: [email protected]
P. Rawat :M. Kumar : R. MauryaDivision of Medicinal & Process Chemistry, Central DrugResearch Institute (Council of Scientific and Industrial Research),Chattar Manzil, P.O. Box 173, Lucknow, India
V. Gupta :A. K. DwivediDivision of Pharmaceutics, Central Drug Research Institute(Council of Scientific and Industrial Research),Chattar Manzil, P.O. Box 173, Lucknow, India
L. ManickavasagamDivision of Pharmacokinetics and Metabolism, Central DrugResearch Institute (Council of Scientific and Industrial Research),Chattar Manzil, P.O. Box 173, Lucknow, India
Osteoporos Int (2011) 22:3013–3027DOI 10.1007/s00198-010-1519-4
Received: 24 August 2010 /Accepted: 6 December 2010 /Published online: 12 January 2011

Introduction
In animal models, dietary supplements and nutraceuticalsrich in flavonols including quercetin (Q) and kaempferolhave been reported to counteract the bone deleteriouseffects of estrogen deficiency without uterotrophic effect[1–3]. Although Q is not the most predominant flavonoid inour diet, it is one of the most studied among differentdietary flavonoids [1, 4]. Q and one of its glucosides,quercetin-3-O-rutinose (also called rutin), are present infruits and vegetables. Q [3] and rutin [5] have been reportedto inhibit ovariectomy (OVx)-induced osteopenia in rats.Bone sparing action of Q and rutin is thought to bemediated by anti-oxidant properties that attenuate theproduction of oxidation-derived free radicals from the boneresorbing osteoclasts and their precursors [6–10]. Q andrutin could also act through an estradiol receptor (ER), asphytoestrogens do [4, 11]. Several lines of evidence showthat phytoestrogens can prevent bone loss, which has led totheir increased prophylactic use in postmenopausal women[12]. However, restoration of lost bone requires the use ofbone-forming agents, the so-called bone anabolics orosteogenic agents.
Anabolic therapy or stimulating the function of bone-forming osteoblasts is the preferred pharmacological inter-vention for osteoporosis [13]. Parathyroid hormone (PTH;1–34) remains the only anabolic agent available for clinicaluse that has recently been recommended by FDA to carry ablack-box warning because it is associated with anincreased risk of osteosarcoma in rats. Intermittent PTH(iPTH) increases not only bone mass but also bone qualityand strength by improving microarchitecture and geometryof bone [14]. At the cellular level, iPTH exerts its anaboliceffect by increasing proliferation and differentiation ofosteoprogenitors [15, 16], conversion of bone-lining cells toactive osteoblasts [17, 18], and osteogenic differentiation ofmesenchymal precursor cells at the expense of adipogenesis[19, 20]. Inhibition of osteoblast apoptosis may alsocontribute to the anabolic action of iPTH since PTHtransiently reduces osteoblast apoptosis both in culture[21–24] and in vivo [21, 23].
Flavonols have been reported to promote osteoblastproliferation, differentiation, and mineralization as well asincreasing production of osteoprogenitors [2, 25–31].Reports on the effects of Q on osteoblast function areconflicting. Q has been shown to stimulate differentiationof MG-63 osteoblast “like” cells via extracellular signal-regulated kinases and ER-mediated pathways [27] andincrease production of bone sialoprotein that has beenimplicated in the nucleation of hydroxyapatite crystals. Incontrast, Q has also been reported to induce apoptosis ofMC3T3-E1, murine calvarial osteoblasts [32, 33]. Invari-ably, micromolar concentrations of Q or its analogs were
required for stimulation of osteoblast functions in vitro thatare not likely to be achieved in vivo, particularly at thebone tissue level. From these studies, it appears that Qcould serve as a “lead” pharmacophore for osteogenicactivity if potency and bioavailability of Q are enhanced.
In our search for more potent analog(s) of Q withosteogenic effects, we undertook isolation of bioactivecompounds from a standardized, butanolic fraction (BF)from the stem bark of Ulmus wallichiana (Himalayan Elm).Stem-bark extract of U. wallichiana is known in Indiantraditional medicine to accelerate fracture repair [34, 35].We have recently shown that the BF promoted peak bonemass achievement and prevented OVx-induced bone loss inrats [36]. Among the four compounds of BF that stimulatedosteoblast differentiation in vitro, 3,3′,4′,5,7-pentahydroxy-flavone-6-C-β-D-glucopyranoside or quercetin-6-C-β-D-glucopyranoside (QCG) was the most abundant. In OVxrats, QCG more effectively than Q improved bonebiomechanical quality through positive modifications ofBMD and bone microarchitecture without having hyper-plastic effect on uterus [3]. However, we did not system-atically evaluate the bone anabolic effect of QCG.Accordingly, the present study was designed to assessanabolic effect of QCG in growing as well as osteoporoticbones and its associated cellular mechanisms by usingculture systems and dynamic and static histomorphome-tries. Finally, since bioavailability is required for anycompound to exert biological effects, we determined oralbioavailability of QCG in rats.
Materials and methods
Reagents and chemicals
Cell culture media and supplements were purchased fromInvitrogen (Carlsbad, CA, USA). All fine chemicalsincluding alendronate sodium trihydrate and 17β-estradiol(17β-E2) were purchased from Sigma Aldrich (St. Louis,MO, USA). High-performance liquid chromatography(HPLC) grade acetonitrile was obtained from Merck IndiaLtd (Mumbai, India). Heparin sodium injection(1,000 IU/ml IP) was purchased from Gland Pharma-ceuticals (Hyderabad, India) and human PTH (1–34)from EMD Chemicals (Gibbstown, NJ, USA). QCG waspurified from the total ethanolic extract of the stem barkof U. wallichiana as described before [3].
In vitro and in vivo studies
For harvesting cultures from calvariae and bone marrow(BM) of Sprague Dawley rats, a prior approval from theInstitutional Animal Ethics Committee (IAEC) was
3014 Osteoporos Int (2011) 22:3013–3027

obtained. A separate approval from the IAEC was obtainedfor OVx, husbandry, and treatment of female SpragueDawley rats with various agents described in this study.Euthanasia and disposal of carcass were performed inaccordance with the guidelines laid by IAEC for animalexperimentation.
Culture of calvarial osteoblasts
For each experiment, about 25–30 calvariae were harvestedat room temperature from 1- to 2-day-old rats (both sexes).Briefly, individual calvaria was surgically isolated from theskull, sutures were removed, and adherent tissue materialwas cleaned by gentle scrapping. With the pooled calvariae,a previously described method of repeated digestion(15 min/digestion) with 0.05% trypsin and 0.1% collage-nase P was used to release cells [37]. After discarding thecells from the first two digestions, cells from the next threedigestions were pooled and cultured in α-MEM containing10% FCS and 1% penicillin/streptomycin (complete growthmedium). Cultures of rat calvarial osteoblasts (RCO) wereallowed to reach 80% confluence for other experimentsdescribed below.
ALP assay
For the measurement of alkaline phosphatase (ALP)activity, RCO at ~80% confluence were trypsinized and2×103 cells/well were seeded onto 96-well plates. Cellswere treated with QCG (1, 10, or 100 nM) or vehicle for48 h in α-MEM supplemented with 10% charcoal treatedFCS, 10 mM β-glycerophosphate, 50 μg/ml ascorbic acid,and 1% penicillin/streptomycin (osteoblast differentiationmedium). At the end of incubation period, total ALPactivity was measured using p-nitrophenylphosphate assubstrate and absorbance was read at 405 nm [38].
To study the possible mediation of ER in the QCG-induced ALP production, 17β-E2 (1 nM) and ICI-182780(1 nM), an anti-estrogen, were used. RCO were pre-treatedwith ICI-182780 for 30 min prior to QCG treatment, andALP production was determined as described above [39].
Mineralized nodule formation
Mineralization of RCOs was performed following previ-ously published protocol [2, 40]. Briefly, RCO were seededonto 12-well plates (25,000 cells/well) in osteoblastdifferentiation medium. RCO were cultured with or withoutQCG for 21 days with changing of medium every 48 h. Atthe end of the experiment, cells were washed withphosphate-buffered saline (PBS) and fixed with 4%paraformaldehyde in PBS for 15 min. The fixed cells werestained with alizarin red-S (40 mM, pH 4.5) for 30 min
followed by washing with H2O [41]. Stained cells were firstphotographed under light microscope, and alizarin stainwas then extracted by 10% (v/v) acetic acid with shaking atroom temperature for 30 min [40]. Cells were scrapped outof the culture plates, centrifuged (20,000×g for 15 min),and supernatant was collected. To the supernatant, 10% (v/v)ammonium hydroxide was added to bring the pH of thesolution to 4.5. Absorbance of the solution was read at405 nm [2, 40, 42].
RCO represent osteoblasts from membranous bones thatdo not exhibit osteoporotic bone loss. Therefore, we alsostudied the effect of QCG on the mineralization of BMCfrom one of the bones (femora) that undergo bone lossunder E2 deficiency. BMC from 1-month-old female rats(~40 g) were harvested and cultured as described before [2,43]. Briefly, BMC from the femora was flushed out in20 ml osteoblast differentiation medium containing 10−7 Mdexamethasone (BM differentiation medium). Cells wereseeded (2×106 cells/well) onto 12-well plates in BMdifferentiation medium. BMC were cultured with orwithout QCG for 21 days with change of medium every48 h. At the end of the experiment, mineralized noduleswere stained and mineralization quantified as described incase of RCO.
Quantitative real-time polymerase chain reaction
SYBR green chemistry was used to perform quantitativedetermination of mRNA levels of runt-related transcriptionfactor 2 (Runx-2), bone morphogenetic protein-2 (BMP-2),osteocalcin, collagen type-1 (Col1), and the housekeepinggene, glyceraldehyde-3-phosphate dehydrogenase(GAPDH) from RCO treated with QCG following anoptimized protocol described before [40]. The design ofsense and antisense oligonucleotide primers was based onpublished cDNA sequences using the Universal ProbeLi-brary (Roche Applied Sciences). Primer sequences arelisted in Table 1. cDNA was synthesized with theRevertAid cDNA synthesis kit (Fermentas, Austin, TX,USA) using 2 μg of total RNA in 20 μl reaction volume.For quantitative real-time polymerase chain reaction(qPCR), the cDNA was amplified using Light Cycler 480(Roche Molecular Biochemicals, Indianapolis, IN, USA).
Studies on growing rats
Twenty-one-day-old female rats were treated with 5.0- or10.0-mg kg−1 day−1 dose of QCG or vehicle (gum acacia indistilled water) for 90 consecutive days by oral gavages.Gum acacia with distilled water is routinely used asthickening agent in pharmaceuticals as vehicle [2]. Follow-ing a previously published protocol, each animal receivedintraperitoneal injection of tetracycline (20 mg kg−1) on
Osteoporos Int (2011) 22:3013–3027 3015

days 4 and 54 and calcein (20 mg kg−1) on days 40 and 68,respectively [36]. At autopsy, femora and tibiae weredissected out, cleaned for soft tissue, fixed in 70% ethanol,and stored at 4°C until the measurement of various boneparameters as described below.
Pooled BMC from tibiae and femora of vehicle or QCG-treated rats were harvested, and mineralization was studied asdescribed in the previous section [2]. BMCwere then culturedfor 21 days in BM differentiation medium.Mineralization wasquantified as described in the previous section [40, 44].
Studies on OVx rats
Forty adult female rats (weighing 200±20 g each) werebilaterally OVx and left untreated for 13 weeks prior to thestart of various treatments [45]. For the treatments, OVxrats were randomly divided equally into four groups asfollows: OVx + vehicle (gum acacia in distilled water),OVx+20 μg kg−1 PTH (twice a week by intraperitonialinjection), OVx+3.0 mg kg−1 day−1 alendronate, and OVx+5.0 mg kg−1 day−1 QCG. In addition, ten adult female rats(200±20 g each) were sham-operated (ovary intact group)and given vehicle (control group). Since QCG had bonesparing effect in OVx rats at 5.0 mg kg−1 day−1 dose [3], weused the same dose for the current study. The dose regimenfor PTH used in this study was based on previous studies[46]. Treatments were continued for 12 weeks. Fordynamic histomorphometry, each animal received intra-peritoneal injections of tetracycline (20 mg kg−1) onday 60 (8 weeks) and calcein (20 mg kg−1) on day 90(12 weeks) from the start of the various treatmentsincluding the vehicle-treated control groups. At the endof all treatments, rats were euthanized and autopsied tocollect bones (tibia, femur) for the measurement of boneparameters as described below.
Measurement of bone parameters
Following previously described protocols, BMD measure-ment of isolated bones was performed using dual energy X-
ray absorptiometry (DEXA; Model 4500 Elite, Hologic)fitted with commercially available software (QDR 4500ACCLAIM series) for a fan-beam DXA system that allowsdetermination of global (total) as well as regional BMD ofisolated bones [2, 37].
For dynamic histomorphometric studies, tetracycline andcalcein labeled bones were processed as described before[36, 37]. At the end of the study, isolated bones wereembedded in an acrylic material, and 50-μm sections weremade using Isomet Bone cutter (Buehler, Lake Bluff, IL,USA). Photograph of the sections was taken underfluorescent microscope aided with appropriate filters. Boneformation rate (BFR) and mineral appositional rate (MAR)were then calculated using Leica-Qwin software (LeicaMicrosystems GmbH) according to a previously describedmethod [47].
μCT scanning of excised bones was carried out usingthe Sky Scan 1076 μCT scanner (Aartselaar, Belgium)as described before [3, 45, 48]. The bone samples werescanned in batches of three at a nominal resolution (pixels)of 18 μm. Reconstruction was carried out using amodified Feldkamp algorithm using the Sky Scan Nreconsoftware, which facilitates network distributed reconstruc-tion carried out on four personal computers runningsimultaneously. The X-ray source was set at 70 kV and100 mA, with a pixel size of 18 μm. A hundredprojections were acquired over an angular range of 180°.The image slices were reconstructed using the cone-beamreconstruction software version 2.6 based on the Feld-kamp algorithm (Skyscan). The trabecular bone wasselected by drawing ellipsoid contours with the CTanalyzer (CTAn, Skyscan) software. Trabecular bonevolume, trabecular number, and trabecular separation offemur epiphysis and proximal tibial metaphysis werecalculated by the mean intercept length method. Trabec-ular thickness was calculated according to the method ofHildebrand and Ruegsegger [49]. 3-D parameters werebased on analysis of a Marching cubes-type model with arendered surface. Cortical thickness, cortical area, corticalperimeter, periosteal perimeter, and endosteal perimeter
Gene name Primer sequence Accession number
BMP-2 F—CGGACTGCGGTCTCCTAA NM_007553.2R—GGGGAAGCAGCAACACTAGA
Osteocalcin F—ATAGACTCCGGCGCTACCTC NM_013414R—CCAGGGGATCTGGGTAGG
Collagen type-1 F—CATGTTCAGCTTTGTGGACCT NM_053304R—GCAGCTGACTTCAGGGATGT
Runx-2 F—CCACAGAGCTATTAAAGTGACAGTG NM_053470R—AACAAACTAGGTTTAGAGTCATCAAGC
GAPDH F—TTTGATGTTAGTGGGGTCTCG NM_017008R—AGCTTGTCATCAACGGGAAG
Table 1 Primer sequenceof various genes used for qPCR
301 Osteoporos Int (2011) 22:3013–30276

were calculated by 2-D analysis of cortical bones of femur(mid-diaphysis) and tibiofibula separating point (TFSP).To ensure consistent measurement of cortical parameters,the beginning of the growth plate served as reference pointfrom where 250 serial image slides were discarded toexclude the trabecular region, and the following 200consecutive image slides (representing only cortical bone)were selected for analysis and quantification using CTAnsoftware.
Determination of plasma and bone marrow levels of QCG
Animals, drug administration, and sampling
Adult female rats (200±20 g) were used for this study.The animals were first administered a 5.0-mg kg−1 bolusdose of QCG by oral gavages and sacrificed at 0.25, 0.5,1, 3, 6, 8, 10, 12, and 24 h post-dosing. Three animalswere taken at each time point. Plasma and BM werecollected for the determination of QCG levels. Data thatrepresent the concentration–drug profile at 0 h time pointwere obtained from animals without any previous QCGtreatment.
Sample processing was performed as described before[48]. BM was harvested by repeated flushing of femur andtibia in PBS, as described before [2]. After centrifugationand removal of supernatants, wet weight of BM was notedand 50 mg BM was homogenized in 1 ml PBS. Theresultant samples were centrifuged at approximately9,000×g for 10 min and stored at −20°C until analysisperformed. To 0.5 ml of plasma or BM samples, 1 ml ofacetonitrile was added. The resulting solution was thor-oughly vortex-mixed for 2 min. After centrifugation at3,000×g for 10 min, the supernatant layer was transferredinto a clean test tube, concentrated to dryness undervacuum, and reconstituted in 50 μl of acetonitrile. Fromthe reconstituted samples, aliquots of 20 μl were injectedinto the HPLC system for analysis.
Preparation of standard and quality control samples
Stock solution of QCG was prepared in methanol togive a final concentration of 48 μg ml−1 (103.44 μM).A series of standard solutions with concentration in therange of 96–576 ng ml−1 (206 nM–1.24 μM) wasobtained by serial dilution method with methanol. Toprepare the standard calibration samples, 50 μl ofstandard solutions was added to 250 μl of blank plasmaor 500 μl of BM. The mixture was then treated followingsample extraction procedure described below. The qualitycontrol (QC) samples, which were used in the validation,were prepared in the same way as the standard calibrationsamples.
HPLC conditions
The HPLC system was equipped with 10 ATVP binarygradient pumps (Shimadzu), a Rheodyne 7125 injector with20 μl injecting capacity (Cotati, CA, USA), and 10 ATVPdiode array detector (Shimadzu). HPLC separation wasperformed using a Lichrocart Lichrosphere C18 column(250 mm, 4 mm, 5 μm; Merck). Column effluent wasmonitored at a wavelength of 370 nm. Data were acquiredand processed using Shimadzu software. The mobile phasewas a mixture of potassium dihydrogen phosphate buffer(0.05 M; containing triethyl amine (0.1%) and pH adjustedto 2.5 with phosphoric acid) and acetonitrile, where theratio of buffer to acetonitrile was 65:35. Both the solutionswere filtered and degassed before use. Chromatographywas performed at 25±3°C at a flow rate of 1.5 ml/min.
Statistical analysis
Data are expressed as mean ± SEM. The data obtained inexperiments with multiple treatments were subjected toone-way ANOVA followed by post hoc Newman–Keulsmultiple comparison test of significance using GraphPadPrism 3.02 software. Qualitative observations have beenrepresented following assessments made by three individ-uals blinded to the experimental designs.
Results
Effect of QCG on osteoblast differentiation
Figure 1a shows the chemical structure of QCG (MW 464). At80% confluence, RCO from 1- to 2-day-old rats were treatedwith QCG and as shown in Fig. 1b, QCG stimulated ALPactivity in RCO at 10 and 100 nM compared to control (cellstreated with vehicle; P<0.001). We next studied the effect ofQCG on mineralization and observed that at the highestconcentration (100 nM) used for stimulating ALP activity inRCO, QCG stimulated mineralization of both RCO (P<0.01;Fig. 1c) and BMC (P<0.001; Fig. 1d).
Next, we studied the effect of QCG on the expression ofvarious osteogenic specific genes in RCO. Figure 1e showsthat 100 nM QCG treatment increased Runx-2 (P<0.01)and BMP-2 (P<0.01) mRNA levels over control at as earlyas 4 h. Increases in the mRNA levels of osteocalcin (P<0.05) and Col1 (P<0.05) by QCG treatment were noted at24 h compared to control. Increases in the mRNA levels ofall four osteogenic genes by QCG were found to continueup to 72 h. At 72 h, QCG increased mRNA levels of Runx-2 (P<0.01), BMP-2 (P<0.001), osteocalcin (P<0.001), andCol1 (P<0.01) compared to control. Q had no effect on anyof these osteogenic genes of RCOs at lower than 10 μM
Osteoporos Int (2011) 22:3013–3027 3017

(data not shown). At 10 μM, Q increased mRNA levels ofBMP-2 (P<0.05) at 48 h and Col1 at both 48 h (P<0.01)and 72 h (P<0.001), compared to control. However, Q(10 μM) had no effect on Runx-2 mRNA levels at any ofthe time points studied and inhibited (P<0.05) osteocalcinmRNA levels at 72 h compared to control (Fig. 1e, f).
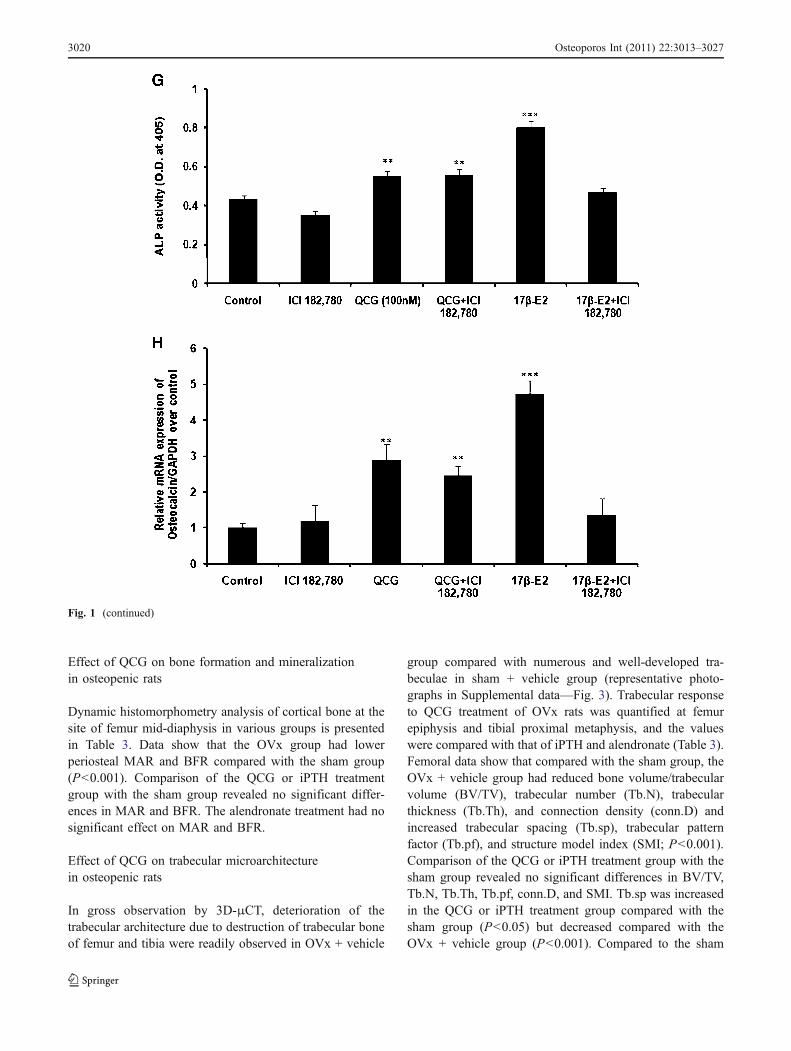
We next tested whether QCG mediated their actions inosteoblasts through ER. Data show that at 48 h, thepresence of anti-estrogen, ICI-182780, did not change theincreases in ALP activity and osteocalcin mRNA levels inRCO by QCG but abolished the effects of 17β-E2 on theseparameters (Fig. 1g, h).
Effect of QCG on body weight of growing rats
QCG (5.0- and 10.0-mg kg−1 day−1 doses) treatments togrowing rats for 12 weeks resulted in no significantdifferences in the initial and final body weights whencompared to the control group (Supplemental data—Fig. 1).
Effect of QCG on mineralization of bone marrow cellsin growing rats
At the end of the 12-week treatment, BMC harvested from thelong bones show more mineralized nodules in the QCG-treated rats compared with the control (rats receiving vehicle)group (Fig. 2a—representative photomicrograph of alizarinred-S stained cells). Quantification of mineralization showsthat the QCG treatment increased mineralization by BMCscompared to the control (P<0.001; Fig. 2b).
Effect of QCG on bone formation and mineralizationin growing rats
Figure 3a shows representative photomicrograph of doublefluorochrome (tetracycline and calcein) labeling at the siteof femur mid-diaphysis during the period of administrationof QCG in growing rats. QCG treatment at either doseincreased MAR (Fig. 3b, c) and BFR (Fig. 3d, e) at theperiosteal and endosteal surfaces compared to the controlgroup (P<0.001).
Effect of QCG on cortical BMD and microarchitectureof long bones in growing rats
The effect of QCG treatment on BMD and statichistomorphometric parameters of cortical bones aredescribed in Table 2. DEXA analyses show that QCG atboth doses increased BMD of global femur and femur atmid-diaphysis compared with control (P<0.01). QCGtreatment at 5-mg kg−1 dose increased BMD of globaltibia (P<0.05) and dose-dependently increased BMD ofTFSP compared to the control.
Static histomorphometry (Table 2) at the site of femurmid-diaphysis showed that the QCG treatment dose-dependently increased cortical mean cross-section area(B.Ar) and endosteal perimeter (B.Pm.–T.Ar) when com-pared with the control group. Further, cortical thickness(Cs.Th), periosteal area (T.Ar), periosteal perimeter (T.Pm), and cortical bone perimeter (B.Pm) were also foundto be significantly higher at both doses of QCG comparedwith the control. Although there were no significantdifferences in T.Ar, T.Pm, and B.Pm between the twodoses of QCG, however, the higher QCG dose(10 mg kg−1) exhibited an increasing trend than the lowerdose (5 mg kg−1).
In TFSP, QCG treatment dose-dependently increased B.Pm.–T.Ar when compared to control (Table 2). B.Ar, Cs.Th, T.Ar, T.Pm, and B.Pm were also found to besignificantly higher at either dose of QCG treatmentcompared to the control group.
Effect of QCG on body weight of OVx rats
Body weight of the OVx + vehicle group was significantlyhigher than the sham group. Body weights of OVx ratstreated with QCG, iPTH, or alendronate were not signifi-cantly different from the OVx + vehicle group (Supple-mental data—Fig. 2).
Fig. 1 QCG stimulates osteoblast differentiation in vitro. a Structureof QCG. b QCG treatment of RCO for 48 in osteoblast differentiationmedium significantly increased ALP production compared to controlcells (receiving vehicle). c QCG treatment of RCO for 21 days inosteoblast differentiation medium significantly increased mineralizednodules compared with control, assessed by alizarin red-S staining(upper panel—representative photomicrograph). Lower panel isquantification of mineralization showing significant increase inmineralization of RCOs by QCG treatment. d QCG treatment ofBMC for 21 days in BM differentiation medium significantlyincreased mineralized nodules compared with control, assessed byalizarin red-S staining (upper panel—representative photomicro-graph). Lower panel is quantification of mineralization showingsignificant increase in mineralization of BMCs by QCG treatment. eQCG (100 nM) treatment of RCO resulted in time-dependent increasein the mRNA levels of various osteogenic genes including runt-relatedtranscription factor 2 (Runx-2), bone morphogenetic protein-2 (BMP-2), osteocalcin, and type I collagen (Col1). f Q (10 μM) treatment ofRCO resulted in increase in the mRNA levels BMP-2 and Col1 at 24and 48 h. Osteocalcin mRNA levels in RCO were significantlyinhibited by Q at 48 h. g RCO were exposed to various treatments asindicated for 48 h in osteoblast differentiation medium. Data show thatthe presence of an anti-estrogen, ICI-182780 (1 nM) did not alterQCG-mediated increase in ALP production in RCO. 17β-E2 (1 nM)served as positive control. h RCO were exposed to various treatmentsas indicated for 48 h in osteoblast differentiation medium. Data showthat the presence of ICI-182780 (1 nM) did not significantly alterQCG-mediated increase in osteocalcin mRNA levels in RCO. Datarepresented as mean ± SEM from three independent experiments. *P<0.05, **P<0.01, and ***P<0.001 compared to control for b–d, g, andh and 0 h for e and f
�
301 Osteoporos Int (2011) 22:3013–30278

Osteoporos Int (2011) 22:3013–3027 3019
Effect of QCG on bone formation and mineralizationin osteopenic rats
Dynamic histomorphometry analysis of cortical bone at thesite of femur mid-diaphysis in various groups is presentedin Table 3. Data show that the OVx group had lowerperiosteal MAR and BFR compared with the sham group(P<0.001). Comparison of the QCG or iPTH treatmentgroup with the sham group revealed no significant differ-ences in MAR and BFR. The alendronate treatment had nosignificant effect on MAR and BFR.
Effect of QCG on trabecular microarchitecturein osteopenic rats
In gross observation by 3D-μCT, deterioration of thetrabecular architecture due to destruction of trabecular boneof femur and tibia were readily observed in OVx + vehicle
group compared with numerous and well-developed tra-beculae in sham + vehicle group (representative photo-graphs in Supplemental data—Fig. 3). Trabecular responseto QCG treatment of OVx rats was quantified at femurepiphysis and tibial proximal metaphysis, and the valueswere compared with that of iPTH and alendronate (Table 3).Femoral data show that compared with the sham group, theOVx + vehicle group had reduced bone volume/trabecularvolume (BV/TV), trabecular number (Tb.N), trabecularthickness (Tb.Th), and connection density (conn.D) andincreased trabecular spacing (Tb.sp), trabecular patternfactor (Tb.pf), and structure model index (SMI; P<0.001).Comparison of the QCG or iPTH treatment group with thesham group revealed no significant differences in BV/TV,Tb.N, Tb.Th, Tb.pf, conn.D, and SMI. Tb.sp was increasedin the QCG or iPTH treatment group compared with thesham group (P<0.05) but decreased compared with theOVx + vehicle group (P<0.001). Compared to the sham
Fig. 1 (continued)
30 Osteoporos Int (2011) 22:3013–302720

group, the alendronate treatment group had no significantdifference in Tb.Th; decreased BV/TV (P<0.05), Tb.N (P<0.05), and conn.D (P<0.001); and increased Tb.sp (P<0.01), Tb.pf (P<0.001), and SMI (P<0.001). However,compared with the OVx + vehicle group, the alendronatetreatment group had increased BV/TV (P<0.001), Tb.N (P<0.05), Tb.Th (P<0.001), and conn.D (P<0.05) and de-creased Tb.sp (P<0.01).
Tibial trabecular data (Table 3) show that compared withthe sham group, the OVx + vehicle group had significantlyreduced BV/TV (P<0.001), Tb.N (P<0.001), Tb.Th (P<0.01), and conn.D (P<0.001) and increased Tb.sp (P<0.001), Tb.pf (P<0.001), and SMI (P<0.001). Compared tothe sham group, the QCG treatment group had nosignificant difference in conn.D; decreased BV/TV (P<0.05), Tb.Th (P<0.05), and Tb.N (P<0.01); and increasedTb.sp (P<0.01), Tb.Pf (P<0.001), and SMI (P<0.01).However, when compared to OVx + vehicle group, QCGtreatment group had increased BV/TV (P<0.001), Tb.N (P<0.001), and conn.D (P<0.001) and decreased Tb.sp (P<0.001), Tb.pf (P<0.001), and SMI (P<0.01). Comparison ofthe QCG treatment group with the OVx group revealed nosignificant difference in Tb.Th. Compared to the shamgroup, the iPTH treatment group had no significant differ-ences in Tb.Th and conn.D, reduced BV/TV (P<0.05) andTb.N (P<0.05), and increased Tb.sp (P<0.05), Tb.Pf (P<0.001), and SMI (P<0.05). When comparison was made
with the OVx + vehicle group, the iPTH treatment groupexhibited increased BV/TV (P<0.001), Tb.Th (P<0.001),Tb.N (P<0.001), and conn.D (P<0.001) and decreased Tb.sp (P<0.001), Tb.pf (P<0.001), and SMI (P<0.01). Com-pared to the sham group, the alendronate treatment grouprevealed decreases in BV/TV (P<0.01), Tb.N (P<0.001),Tb.Th (P<0.01), and conn.D (P<0.01) and increases in Tb.sp (P<0.01), Tb.pf (P<0.001), and SMI (P<0.001). Whencomparison was made with the OVx + vehicle group, thealendronate treatment group exhibited increase in Tb.N (P<0.05) and decrease in SMI (P<0.05).
Serum and bone marrow levels of QCG
The individual plasma and BM pharmacokinetics profile ofQCG were determined over a 24-h period with a limit ofdetection of 48 ng ml−1 (103.4 nM). Figure 4a shows thefirst plasma QCG peak, which is the rapid phase of QCGabsorption during the first 30 min (Tmax) after oraladministration. This initial rapid absorption of QCG wasfollowed by a sharp elimination phase and appearance ofthe second QCG peak after 8 h of oral administration,which could be due to hepatic recirculation. The maximumplasma concentration (Cmax) of QCG was 2.523±0.79 μMand AUC0–24 h was 15,853.09 ng h ml−1.
QCG levels in the BM peaked at 3.0 h (Tmax; Fig. 4b).Cmax and AUC0–24 h of OCG in the BM were 1.172±0.91 μM and 9,131.25 ng h ml−1, respectively. Based onAUC0–24 h, QCG levels in the BM were found to be ~50%of the plasma QCG levels.
Discussion
The present study investigates bone anabolic effect ofQCG. Our data show that QCG is more potent than Q instimulating osteoblast differentiation. QCG facilitatesmodeling-directed bone growth via the increases in boneformation and mineralization. Further, QCG promotes newbone formation and restores trabecular microarchitecture inosteopenic rats, and these effects are comparable to iPTH at20 μg kg−1 (twice a week) dose. Finally, QCG hassubstantial bioavailability in the BM at the effectiveosteogenic dose (5 mg kg−1).
Q is present in fruits and vegetables and occurs inaglycone and O-glucoside forms [50]. QCG, as with C-glucoside forms of flavonoids per se, has much rarerbotanical distribution than aglycone or O-glucoside formsof Q. In QCG, the sugar is directly linked to the flavonoidnucleus via an acid-resistant and largely enzyme-resistantC–C bond. We are not aware of any report concerningosteogenic effect of flavonol C-glucoside (or for that matterC-glucoside of any flavonoid). Our data show that at
Fig. 2 Oral QCG supplementation to growing rats for 12 weeksincreases mineralized nodule formation in the BM. a Representativephotomicrograph showing increased mineralized nodule formation inBMCs obtained from the QCG-treated rats compared to the vehicle(control) group. b Quantification of mineralization showing significantincrease in mineralization of BMC in the QCG-treated rats comparedto the vehicle group. N=10 rats/group; data represented as mean ±SEM; ***P<0.001
Osteoporos Int (2011) 22:3013–3027 3021

100 nM, QCG increased mRNA levels of various osteo-genic genes more robustly than Q at 10 μM, suggestinggreater in vitro osteogenic effect of QCG over Q.Micromolar concentrations of Q in stimulating osteoblastdifferentiation have been reported albeit data are discrepant[27]. Our data demonstrate that despite 100-fold excess, Qwas much less effective than QCG in stimulating osteoblastdifferentiation. Based on these data, C-glucoside conjuga-tion of Q, such as QCG, appears to augment osteogeniceffect of Q. Whether C-glucoside conjugation enhancesosteogenic effect of other flavonoids remains to beinvestigated.
The effect of QCG in peak bone mass achievement wasnext studied in growing female rats because severallongitudinal studies tracking the bone mass and strengththrough childhood and adolescence and mathematicalmodels suggest that modifying peak bone mass has directconsequence on skeletal fragility in old age [51, 52].Growing animals represent bone modeling wherein bone
formation is the dominant event particularly at the corticalsites [53]. QCG, at minimum effective dose of5.0 mg kg−1 day−1, was found to increase corticaldeposition assessed by BMD and dynamic and statichistomorphometric measurements at the sites of femurmid-diaphysis and TFSP compared to control, suggestingthat QCG favored modeling-directed accretion of corticalbone mass. This increase in bone accretion may beattributed by the increase in the differentiation of BMprogenitors to osteogenic lineage in QCG-treated rats. Wehave recently shown that BF from U. wallichiana increasedpeak bone mass in growing rats [36]. As QCG is the mostabundant bioactive constituent of BF, it is reasonable toassume that QCG contributes to the ability of BF topromote peak bone mass achievement.
Next, whether the ability of QCG to promote modeling-directed bone growth translated to osteogenic action underosteopenic condition was studied. Several methoxy deriv-atives of isoflavonoids and plant extracts have been
Fig. 3 Oral QCG supplementation to growing rats for 12 weeksincreases MAR and BFR. a Representative images of transversesections of tetracycline (white arrows) and calcein (black arrows)labeling in the femur mid-diaphyses of QCG-treated rats. Dynamichistomorphometric parameters calculated from the labeling experi-
ments showing increased MAR (b, c) and BFR (d, e) at the periostealand endosteal sites. N=10 rats/group; data represented as mean ±SEM; **P<0.01 and ***P<0.001 compared to vehicle-treated group(control)
30 Osteoporos Int (2011) 22:3013–302722

reported to promote peak bone mass achievement, but theireffects in inducing new bone formation in osteopeniccondition has not been investigated [37, 54]. Here, weused osteopenic rats that had been OVx 13 weeks prior tothe start of the QCG treatment to test if it reversedosteopenia. We also included an osteogenic agent (iPTH)[55] and a potent anti-resorptive compound (alendronate)[56] as controls in this set of study. Dynamic measures ofbone formation at femur mid-diaphysis revealed that theQCG treatment increased MAR and BFR in the OVx-induced osteopenic rats, and these values were similar tothe iPTH treatment, suggesting that QCG stimulatedperiosteal apposition in osteopenic rats. iPTH is known toinduce periosteal apposition in OVx rats [57], and QCGappears to mimic the action of iPTH in osteopenic rats. Asexpected, alendronate, a suppressor of resorption, failed toincrease periosteal apposition in OVx rats.
The preservation of trabecular microarchitecture signif-icantly contributes to bone strength and may reduce fracturerisk beyond BMD [58]. Restoration of microarchitectureparameters is necessary to evaluate the true impact ofanabolic treatment on the quality of trabecular bonebecause trabecular bone is more readily lost because of
OVx in rats [59]. All femoral trabecular parameters exceptTb.sp were comparable between the QCG or iPTHtreatment and the sham group, suggesting substantialrestoration of the lost epiphyseal trabecular bone by theQCG treatment of osteopenic rats. In tibia, none of theparameters except conn.D in the QCG or iPTH treatmentachieved improvements to be comparable to that of thesham group, suggesting incomplete metaphyseal trabecularrestoration by these treatments. A recent report showed thatrecovering treatment (after osteopenia) of PTH had betterresponse in epiphyseal trabecular bone over metaphysealbone [55]. The authors suggested that deterioration of bonemass and structure after OVx in the epiphysis was muchsmaller than in the metaphysis, thus resulting in higherbone deterioration in the metaphysis at the start of PTHtreatment. Similar explanation appears to apply in case ofour study showing better anabolic response of QCG oriPTH treatment in epiphysis over metaphysis. The stabilityof trabecular bone is importantly dependent on structuralparameters determined by Conn.D, SMI, and Tb.pf. Theeffect of QCG or iPTH on conn.D in femur was comparableto the sham group. SMI determines plate- or rod-likegeometry of trabecular structure wherein SMI values of 0
Table 2 DEXA and static bone histomorphometric measurements in growing female rats supplemented with vehicle or QCG for 12 weeks
Parameters Vehicle (gum acacia) QCG (5.0 mg kg−1 day−1) QCG (10.0 mg kg−1 day−1)
DEXA measurements of femur
BMD—global 0.188±0.005 0.217±0.004**** 0.222±0.005****
BMD—mid-diaphysis 0.177±0.005 0.207±0.004**** 0.211±0.006****
DEXA measurements of tibia
BMD—global 0.181±0.003 0.206±0.002*** 0.194±0.004
BMD—TFSP 0.175±0.005 0.193±0.004*** 0.207±0.005*,****
Static cortical bone histomorphometric measurements at mid-diaphysis
B.Ar (mm2) 4.44±0.270 4.73±0.21*** 5.22±0.761**,*****
Cs.Th (μM) 0.321±0.023 0.399±0.006***** 0.373±0.016****
T.Ar (mm2) 10.4±0.366 12.1±0.69*** 12.6±0.81*****
T.Pm (mm) 11.7±0.166 12.8±0.45*** 13.5±0.78*****
B.Pm (mm) 20.6±0.77 28.7±0.366***** 30.6±1.245*****
B.Pm–T.Ar 10.1±0.661 13.1±0.781*** 18.0±0.714**,*****
Static cortical bone histomorphometric measurements at TFSP
B.Ar (mm2) 4.19±0.313 4.75±0.07***** 4.41±0.501***
Cs.Th (μM) 0.254±0.023 0.350±0.019***** 0.322±0.022****
T.Ar (mm2) 7.71±0.365 10.9±1.08**** 9.37±0.38*****
T.Pm (mm) 12.8±0.156 15.0±0.55***** 14.3±0.28*****
B.Pm (mm) 22.2±0.59 27.5±1.95***** 29.6±2.245*****
B.Pm–T.Ar 14.5±0.52 16.6±0.86*** 20.1±1.714**,*****
Values represent mean±SEM (n=10 rats/group). BMD values are presented as grams per square centimeter
B.Ar cortical mean cross-section area, Cs.Th cortical thickness, T.Ar periosteal area, T.Pm periosteal perimeter, B.Pm cortical bone perimeter, B.Pm.–T.Ar endosteal perimeter, TFSP tibiofibular separation point
*P<0.05 compared to QCG (5.0 mg kg−1 day−1 ) treated group; **P<0.01 compared to QCG (5.0 mg kg−1 day−1 ) treated group; ***P<0.05 comparedto vehicle group; ****P<0.01 compared to vehicle group; *****P<0.001 compared to vehicle group
Osteoporos Int (2011) 22:3013–3027 3023

Table 3 Dynamic and static histomorphometric measurements of rats with various treatments
Parameters Sham + vehicle OVx + vehicle OVx + QCG(5.0 mg kg−1 day−1)
OVx + iPTHa
(20 μg kg−1)OVx + Aln(3.0 mg kg−1 day−1)
Dynamic histomorphometric measurements at femur diaphysis
MAR (μm/day) 0.087±0.001****** 0.067±0.002 0.088±0.005****** 0.091±0.002****** 0.066±0.004**
BFR (μm3/μm2/day) 0.086±0.001****** 0.066±0.003 0.082±0.006***** 0.089±0.002****** 0.066±0.004**
Static trabecular bone histomorphometric measurements at femur epiphysis
BV/TV (%) 15.42±0.773****** 6.74±0.334 14.14±0.788****** 14.23±0.853****** 11.62±0.601*,******
Tb.Sp. (mm) 0.77±0.054****** 1.36±0.038 0.99±0512*,****** 1.03±0.034*,****** 1.09±0.039**,*****
Tb.N (1/mm) 1.38±0.043****** 0.64 ±0.058 1.14±0.137****** 1.13±0.077****** 0.94±0.091*,****
Tb.Th (mm) 0.120±0.002****** 0.105±0.001 0.112±0.003***** 0.120±0.001****** 0.115±0.003******
Tb.Pf (1/mm) 2.10±0.875****** 7.26±0.415 3.86±0.726****** 3.60±0.385****** 5.46±0.716***
Conn.D (1/mm3) 59.6±6.276****** 17.7±1.816 44.09±5.165****** 42.78±3.532****** 28.29±1.951***,****
SMI 1.13±0.081****** 1.88±0.030 1.58±0.100****** 1.39±0.041****** 1.70±0.105***
Static trabecular bone histomorphometric measurements at tibia proximal
BV/TV (%) 18.82±1.483 ****** 4.79±0.579 11.00±0.359 *,****** 10.33±0.845 *,****** 9.58±0.395**
Tb.Sp. (mm) 0.524±0.052****** 1.259±0.066 0.931±037**,****** 0.966±0.051*,****** 0.886±0.019**
Tb.N (1/mm) 1.721±0.133****** 0.412±0.050 0.752±0.078**,****** 0.910±0.070*,****** 0.709±0.034***,****
Tb.Th (mm) 0.120±0.003***** 0.109±0.002 0.115±0.002* 0.119±0.001****** 0.112±0.002**
Tb.Pf (1/mm) 3.48±0.556****** 13.56±0.722 10.35±0.98***,****** 9.05±0.392***,****** 10.84±0.421***
Conn.D (1/mm3) 54.54±6.591****** 32.05±6.095 45.24±5.145****** 49.50±10.51****** 35.41±2.17**
SMI 1.433±0.101****** 2.517±0.027 2.03±0.141**,***** 2.05±0.034*,***** 2.37±0.091***,****
Values represent mean ± SEM (n=10 rats/group)
BV/TV bone volume/trabecular volume, Tb.Sp trabecular spacing, Tb.N trabecular number, Tb.Th trabecular thickness, Tb.pf trabecular patternfactor, Conn.D connection density, SMI structure model index, Aln alendronate, vehicle gum acacia
*P<0.05 compared to sham + vehicle group; **P<0.01 compared to sham + vehicle group; ***P<0.001 compared to sham + vehicle group;****P<0.05 compared to OVx + vehicle group; *****P<0.01 compared to OVx + vehicle group; ******P<0.001 compared to OVx + vehiclegroupa iPTH was given twice a week by i.p. route
Fig. 4 Determination of plasmaand BM levels of QCG in adultfemale Sprague Dawley rats. aPlasma concentration–time pro-files of QCG following singleoral dose of 5 mg kg−1. b BMconcentration–time profiles ofQCG following single oral doseof 5 mg kg−1; N=3 rats/timepoint, 24 h study. For data, referto the “Results” section
30 Osteoporos Int (2011) 22:3013–302724

and 3 represent bone that consists purely of plate-like orrod-like structure, respectively. OVx results in higher SMIvalues thus representing more rod-like structures. Inaddition, connectedness of trabecular plates to rods isreflected in Tb.pf, wherein more concave surfaces representa well-connected spongy lattice. The QCG or iPTHtreatment completely reversed the OVx-induced changesin Tb.pf in femur but only partially in tibia.
Our data show that the restoration of trabecular micro-architecture of osteopenic rats that received QCG treatmentwas more or less comparable to those received iPTHtreatment, which suggest anabolic effect of QCG. Onecaveat of the present study is that 20 μg kg−1 iPTHadministered on twice-a-week basis may be lower and lessfrequent than that used in many reports for completerestoration of trabecular bone, and hence, it would bepremature to claim equal efficiency of QCG to iPTHconcerning bone anabolic effect. However, it should benoted that there is no firm experimental evidence thatwould allow one to reliably set the dose and frequency ofPTH to achieve complete trabecular restoration of the lostbone at all skeletal sites even within a given species [46,57, 60–62], and our selection of PTH dose and regimen isreported to be effective in net new bone formation [46].
Alendronate, an anti-resorptive agent, has been shownnot only to prevent bone loss but also maintain BMD inosteopenic rats [63–65]. We recently reported that QCGinhibited osteoclastogenesis, decreased the urinary levelsof the bone resorption marker (deoxypyridinoline) in OVxrats, and mitigated OVx-induced trabecular bone loss,which suggested anti-resorptive mode of QCG action [3].Here, as we investigated possible restorative of effect ofQCG in osteoporotic bones, an alendronate treatmentgroup was included to demonstrate that the restoration oftrabecular bone by QCG was beyond that of thesuppressive effect on bone loss by alendronate. Indeed,trabecular responses in the QCG-treated group weresignificantly better than the alendronate treatment group,indicating osteogenic response to QCG in OVx-inducedosteopenic rats.
Over the past few years, studies on the absorption andmetabolism of flavonoids have increased. From thesestudies, it appears that flavonoids, particularly the aglyconeforms, have low oral bioavailability because of extensiveconjugation of free hydroxyl groups present in theflavonoids [66]. Q is rapidly metabolized [67]. Therefore,ensuring bioavailability of QCG, a flavonoid C-glucoside isimportant to fully exploit its osteogenic effect and realizetherapeutic potential. So far only two studies are availableconcerning the oral bioavailability of flavonoid C-glucosideincluding eriodictyol-C-glucoside from rooibos tea [68] and6-C-β-D-glucopyranosyl-(2S,3S)-(+)-3′,4′,5,7-tetrahydroxy-flavanone (GTDF/ulmoside A) from U. wallichiana [48]. In
both cases, low oral bioavailability was observed [68]. Wefound that QCG had substantial oral bioavailability evidentfrom AUC, Cmax, and Tmax values. Tissue distributionstudies of Q by dietary administration for 11 weeks in ratsrevealed that bone Q levels were 1% to 2% of the plasma Qlevels [67]. In the case of QCG, as calculated from AUC,single oral dose of QCG resulted in BM levels that were >50%of the plasma levels of QCG suggesting that a substantialfraction of circulating QCG reached the BM. The prolongedsystemic and BM concentrations of QCG indicate that once aday dosage regimen of 5.0 mg kg−1 day−1 dose is favorablefor its osteogenic effect in vivo.
In conclusion, the result of the present study demon-strated that 12-week oral QCG treatment promotedmodeling-directed bone growth in growing rats and exertedanabolic effect on osteoporotic bone comparable to that ofiPTH through direct stimulatory effect on osteoblastdifferentiation of osteoprogenitors. Given its lack of uterineestrogenicity, low dose required for bone anabolic effect,and copious oral bioavailability, QCG could be analternative anabolic strategy for the development of newtreatment against postmenopausal osteoporosis.
Acknowledgments This study was supported by grant from theMinistry of Health and Family Welfare, Government of India. Fundingfrom the Indian Council of Medical Research, Government of India(N.C.) is acknowledged. Research fellowship grants from UniversityGrants Commission (J.A.S., A.K.G.), Department of Biotechnology(K.S.), and Council of Scientific and Industrial Research (G.S., B.C.,P.R, M.K.,V.G.,L.M.), Government of India, are also acknowledged.
Conflicts of interest None.
References
1. Tsuji M, Yamamoto H, Sato T, Mizuha Y, Kawai Y, Taketani Y,Kato S, Terao J, Inakuma T, Takeda E (2009) Dietary quercetininhibits bone loss without effect on the uterus in ovariectomizedmice. J Bone Miner Metab 27:673–681
2. Trivedi R, Kumar S, Kumar A, Siddiqui JA, Swarnkar G, GuptaV, Kendurker A, Dwivedi AK, Romero JR, Chattopadhyay N(2008) Kaempferol has osteogenic effect in ovariectomized adultSprague-Dawley rats. Mol Cell Endocrinol 289:85–93
3. Siddiqui JA, Sharan K, Swarnkar G, Rawat P, Kumar M, Manick-avasagam L, Maurya R, Pierroz D, Chattopadhyay N (2010)Quercetin-6-C-beta-d-glucopyranoside isolated from Ulmus wall-ichiana planchon is more potent than quercetin in inhibitingosteoclastogenesis and mitigating ovariectomy-induced bone lossin rats. Menopause. doi:10.1097/gme.0b013e3181e84e67
4. Sharan K, Siddiqui JA, Swarnkar G, Maurya R, Chattopadhyay N(2009) Role of phytochemicals in the prevention of menopausal boneloss: evidence from in vitro and in vivo, human interventional andpharma-cokinetic studies. Curr Med Chem 16:1138–1157
5. Horcajada-Molteni MN, Crespy V, Coxam V, Davicco MJ,Remesy C, Barlet JP (2000) Rutin inhibits ovariectomy-inducedosteopenia in rats. J Bone Miner Res 15:2251–2258
6. Wattel A, Kamel S, Mentaverri R, Lorget F, Prouillet C, Petit JP,Fardelonne P, Brazier M (2003) Potent inhibitory effect of
Osteoporos Int (2011) 22:3013–3027 30 52

naturally occurring flavonoids quercetin and kaempferol on invitro osteoclastic bone resorption. Biochem Pharmacol 65:35–42
7. Jung WJ, Sung MK (2004) Effects of major dietary antioxidantson inflammatory markers of RAW 264.7 macrophages. Biofactors21:113–117
8. Wattel A, Kamel S, Prouillet C, Petit JP, Lorget F, Offord E,Brazier M (2004) Flavonoid quercetin decreases osteoclasticdifferentiation induced by RANKL via a mechanism involvingNF kappa B and AP-1. J Cell Biochem 92:285–295
9. Pang JL, Ricupero DA, Huang S, Fatma N, Singh DP, Romero JR,Chattopadhyay N (2006) Differential activity of kaempferol andquercetin in attenuating tumor necrosis factor receptor familysignaling in bone cells. Biochem Pharmacol 71:818–826
10. Wadsworth TL, Koop DR (1999) Effects of the wine polyphe-nolics quercetin and resveratrol on pro-inflammatory cytokineexpression in RAW 264.7 macrophages. Biochem Pharmacol57:941–949
11. Rassi CM, Lieberherr M, Chaumaz G, Pointillart A, Cournot G(2005) Modulation of osteoclastogenesis in porcine bone marrowcultures by quercetin and rutin. Cell Tissue Res 319:383–393
12. Branca F (2003) Dietary phyto-oestrogens and bone health. ProcNutr Soc 62:877–887
13. Cortet B (2009) Effects of bone anabolic agents on boneultrastructure. Osteoporos Int 20:1097–1100
14. Sato M, Westmore M, Ma YL, Schmidt A, Zeng QQ, Glass EV,Vahle J, Brommage R, Jerome CP, Turner CH (2004) Teriparatide[PTH(1-34)] strengthens the proximal femur of ovariectomizednonhuman primates despite increasing porosity. J Bone Miner Res19:623–629
15. Nishida S, Yamaguchi A, Tanizawa T, Endo N, Mashiba T,Uchiyama Y, Suda T, Yoshiki S, Takahashi HE (1994) Increasedbone formation by intermittent parathyroid hormone administra-tion is due to stimulation of proliferation and differentiation ofosteoprogenitor cells in bone marrow. Bone 15:717–723
16. Kostenuik PJ, Halloran BP, Turner RT, Morey-Holton ER, BikleDD (1999) Skeletal unloading causes resistance of osteoprogeni-tor cells to parathyroid hormone and to insulin-like growth factor-I. J Bone Miner Res 14:21–31
17. Dobnig H, Turner RT (1995) Evidence that intermittenttreatment with parathyroid hormone increases bone formationin adult rates by activation of bone lining cells. Endocrinology136:3632–3638
18. Dobnig H, Turner RT (1997) The effects of programmedadministration of human parathyroid hormone fragment (1–34)on bone histomorphometry and serum chemistry in rats. Endocri-nology 138:4607–4612
19. Rickard DJ, Wang FL, Rodriguez-Rojas AM, Wu Z, Trice WJ,Hoffman SJ, Votta B, Stroup GB, Kumar S, Nuttall ME (2006)Intermittent treatment with parathyroid hormone (PTH) as well asa non-peptide small molecule agonist of the PTH1 receptorinhibits adipocyte differentiation in human bone marrow stromalcells. Bone 39:1361–1372
20. Kulkarni N, Wei T, Kumar A, Dow E, Stewart T, Shou J, N'choM, Sterchi D, Gitter B, Higgs R, Halladay D, Engler T, Martin T,Bryant H, Ma Y, Onyia J (2007) Changes in osteoblast,chondrocyte, and adipocyte lineages mediate the bone anabolicactions of PTH and small molecule GSK-3 inhibitor. J CellBiochem 102:1504–1518
21. Jilka RL, Weinstein RS, Bellido T, Roberson P, Parfitt AM,Manolagas S (1999) Increased bone formation by prevention ofosteoblast apoptosis with parathyroid hormone. J Clin Investig104:439–446
22. Chen HL, Demiralp B, Schneider A, Koh AJ, Silve C, Wang CY,McCauley LK (2002) Parathyroid hormone and parathyroidhormone-related protein exert both pro- and anti-apoptotic effectsin mesenchymal cells. J Biol Chem 277:19374–19381
23. Bellido T, Ali A, Plotkin L, Fu Q, Gubrij I, Roberson P, WeinsteinR, O'Brien C, Manolagas S, Jilka R (2003) Proteasomaldegradation of Runx2 shortens parathyroid hormone-inducedanti-apoptotic signaling in osteoblasts. A putative explanationfor why intermittent administration is needed for bone anabolism.Journal of Biological Chemistry 278:50259–50272
24. Sowa H, Kaji H, Iu MF, Tsukamoto T, Sugimoto T, Chihara K(2003) Parathyroid hormone-Smad3 axis exerts anti-apoptoticaction and augments anabolic action of transforming growthfactor beta in osteoblasts. J Biol Chem 278:52240–52252
25. Yang YJ, Yang ZL, Wang DC, Xiao XC, Li P (2006)[Comparative study on effects of rutin and quercetin onmetabolism in osteoblast cells]. Zhong Yao Cai 29:467–470
26. Miyake M, Arai N, Ushio S, Iwaki K, Ikeda M, Kurimoto M(2003) Promoting effect of kaempferol on the differentiation andmineralization of murine pre-osteoblastic cell line MC3T3-E1.Biosci Biotechnol Biochem 67:1199–1205
27. Prouillet C, Maziere JC, Maziere C, Wattel A, Brazier M, KamelS (2004) Stimulatory effect of naturally occurring flavonolsquercetin and kaempferol on alkaline phosphatase activity inMG-63 human osteoblasts through ERK and estrogen receptorpathway. Biochem Pharmacol 67:1307–1313
28. Yin XX, Chen ZQ, Liu ZJ, Ma QJ, Dang GT (2007) Icariinestimulates proliferation and differentiation of human osteoblastsby increasing production of bone morphogenetic protein 2. ChinMed J (Engl) 120:204–210
29. Chen KM, Ge BF, Ma HP, Liu XY, Bai MH, Wang Y (2005)Icariin, a flavonoid from the herb Epimedium enhances theosteogenic differentiation of rat primary bone marrow stromalcells. Pharmazie 60:939–942
30. Chen KM, Ma HP, Ge BF, Liu XY, Ma LP, Bai MH, Wang Y(2007) Icariin enhances the osteogenic differentiation of bonemarrow stromal cells but has no effects on the differentiation ofnewborn calvarial osteoblasts of rats. Pharmazie 62:785–789
31. Yin XX, Chen ZQ, Dang GT, Ma QJ, Liu ZJ (2005) [Effects ofEpimedium pubescens icariine on proliferation and differentiationof human osteoblasts]. Zhongguo Zhong Yao Za Zhi 30:289–291
32. Son YO, Kook SH, Choi KC, Jang YS, Jeon YM, Kim JG, LeeKY, Kim J, Chung MS, Chung GH, Lee JC (2006) Quercetin, abioflavonoid, accelerates TNF-alpha-induced growth inhibitionand apoptosis in MC3T3-E1 osteoblastic cells. Eur J Pharmacol529:24–32
33. Son YO, Kook SH, Choi KC, Jang YS, Choi YS, Jeon YM, KimJG, Hwang HS, Lee JC (2008) Quercetin accelerates TNF-alpha-induced apoptosis of MC3T3-E1 osteoblastic cells throughcaspase-dependent and JNK-mediated pathways. Eur J Pharmacol579:26–33
34. Gaur RD (1999) Flora of District Garhwal, North West Himalaya.Srinagar, Garhwal, Trans Media: 86
35. Maurya R, Singh G, Yadav PP (2008) Antiosteoporotic agentsfrom natural sources. In: Rahman AU (ed) Studies in naturalproduct chemistry, bioactive natural products. Elsevier, New York,pp 517–548
36. Sharan K, Siddiqui JA, Swarnkar G, Tyagi AM, Kumar A, Rawat P,Kumar M, Nagar GK, Arya KR, Manickavasagam L, Jain GK,Maurya R, Chattopadhyay N (2010) Extract and fraction fromUlmuswallichiana Planchon promote peak bone achievement and have anonestrogenic osteoprotective effect. Menopause 17:393–402
37. Bhargavan B, Gautam AK, Singh D, Kumar A, Chaurasia S,Tyagi AM, Yadav DK, Mishra JS, Singh AB, Sanyal S, Goel A,Maurya R, Chattopadhyay N (2009) Methoxylated isoflavones,cajanin and isoformononetin, have non-estrogenic bone formingeffect via differential mitogen activated protein kinase (MAPK)signaling. J Cell Biochem 108:388–399
38. Ishizuya T, Yokose S, Hori M, Noda T, Suda T, Yoshiki S,Yamaguchi A (1997) Parathyroid hormone exerts disparate effects
30 Osteoporos Int (2011) 22:3013–302726

on osteoblast differentiation depending on exposure time in ratosteoblastic cells. J Clin Invest 99:2961–2970
39. Tyagi AM, Gautam AK, Kumar A, Srivastava K, Bhargavan B,Trivedi R, Saravanan S, Yadav DK, Singh N, Pollet C, Brazier M,Mentaverri R, Maurya R, Chattopadhyay N, Goel A, Singh D(2010) Medicarpin inhibits osteoclastogenesis and has nonestro-genic bone conserving effect in ovariectomized mice. Mol CellEndocrinol 325:101–109
40. Siddiqui JA, Swarnkar G, Sharan K, Chakravarti B, Sharma G,Rawat P, Kumar M, Khan FM, Pierroz D, Maurya R, Chatto-padhyay N (2010) 8, 8″-Biapigeninyl stimulates osteoblastfunctions and inhibits osteoclast and adipocyte functions: osteo-protective action of 8, 8″-biapigeninyl in ovariectomized mice.Mol Cell Endocrinol 323:256–267
41. Prabhakar U, James IE, Dodds RA, Lee-Rykaczewski E, RiemanDJ, Lipshutz D, Trulli S, Jonak Z, Tan KB, Drake FH, Gowen M(1998) A novel human bone marrow stroma-derived cell lineTF274 is highly osteogenic in vitro and in vivo. Calcif Tissue Int63:214–220
42. Gregory CA, Gunn WG, Peister A, Prockop DJ (2004) AnAlizarin red-based assay of mineralization by adherent cells inculture: comparison with cetylpyridinium chloride extraction.Anal Biochem 329:77–84
43. Maniatopoulos C, Sodek J, Melcher AH (1988) Bone formation invitro by stromal cells obtained from bone marrow of young adultrats. Cell Tissue Res 254:317–330
44. Wang L, Zhao R, Shi X, Wei T, Halloran BP, Clark DJ, JacobsCR, Kingery WS (2009) Substance P stimulates bone marrowstromal cell osteogenic activity, osteoclast differentiation, andresorption activity in vitro. Bone 45:309–320
45. Trivedi R, Kumar A, Gupta V, Kumar S, Nagar GK, Romero JR,Dwivedi AK, Chattopadhyay N (2009) Effects of Egb 761 onbone mineral density, bone microstructure, and osteoblast func-tion: possible roles of quercetin and kaempferol. Mol CellEndocrinol 302:86–91
46. Okimoto N, Tsurukami H, Okazaki Y, Nishida S, Sakai A,Ohnishi H, Hori M, Yasukawa K, Nakamura T (1998) Effects of aweekly injection of human parathyroid hormone (1–34) andwithdrawal on bone mass, strength, and turnover in matureovariectomized rats. Bone 22:523–531
47. Hara K, Kobayashi M, Akiyama Y (2002) Vitamin K2 (menate-trenone) inhibits bone loss induced by prednisolone partly throughenhancement of bone formation in rats. Bone 31:575–581
48. Sharan K, Swarnkar G, Siddiqui JA, Kumar A, Rawat P, KumarM, Nagar GK, Manickavasagam L, Singh SP, Mishra G,Wahajuddin JGK, Maurya R, Chattopadhyay N (2010) A novelflavonoid, 6-C-beta-d-glucopyranosyl-(2S, 3S)-(+)-3′, 4′, 5, 7-tetrahydroxyflavanone, isolated from Ulmus wallichiana Planchonmitigates ovariectomy-induced osteoporosis in rats. Menopause17:577–586
49. Hildebrand T, Ruegsegger P (1997) Quantification of bonemicroarchitecture with the structure model index. ComputMethods Biomech Biomed Engin 1:15–23
50. Boyer J, Brown D, Liu RH (2004) Uptake of quercetin andquercetin 3-glucoside from whole onion and apple peel extracts byCaco-2 cell monolayers. J Agric Food Chem 52:7172–7179
51. Theintz G, Buchs B, Rizzoli R, SlosmanD, Clavien H, Sizonenko PC,Bonjour JP (1992) Longitudinal monitoring of bone mass accumula-tion in healthy adolescents: evidence for a marked reduction after16 years of age at the levels of lumbar spine and femoral neck infemale subjects. J Clin Endocrinol Metab 75:1060–1065
52. Fournier PE, Rizzoli R, Slosman DO, Theintz G, Bonjour JP(1997) Asynchrony between the rates of standing height gain and
bone mass accumulation during puberty. Osteoporos Int 7:525–532
53. Ortoft G, Andreassen TT, Oxlund H (1999) Growth hormoneincreases cortical and cancellous bone mass in young growing ratswith glucocorticoid-induced osteopenia. J Bone Miner Res14:710–721
54. Gautam AK, Bhargavan B, Tyagi AM, Srivastava K, YadavDK, Kumar M, Singh A, Mishra JS, Singh AB, Sanyal S,Maurya R, Manickavasagam L, Singh SP, Wahajuddin W, JainGK, Chattopadhyay N, Singh D (2010) Differential effects offormononetin and cladrin on osteoblast function, peak bone massachievement and bioavailability in rats. J Nutr Biochem.doi:10.1016/j.jnutbio.2010.02.010
55. Brouwers JE, van Rietbergen B, Huiskes R, Ito K (2009) Effectsof PTH treatment on tibial bone of ovariectomized rats assessedby in vivo micro-CT. Osteoporos Int 20:1823–1835
56. Iwamoto J, Takeda T, Sato Y (2006) Efficacy and safety ofalendronate and risedronate for postmenopausal osteoporosis.Curr Med Res Opin 22:919–928
57. Sato M, Zeng GQ, Turner CH (1997) Biosynthetic humanparathyroid hormone (1–34) effects on bone quality in agedovariectomized rats. Endocrinology 138:4330–4337
58. Legrand E, Chappard D, Pascaretti C, Duquenne M, Krebs S,Rohmer V, Basle MF, Audran M (2000) Trabecular bone micro-architecture, bone mineral density, and vertebral fractures in maleosteoporosis. J Bone Miner Res 15:13–19
59. Lane NE, Kumer JL, Majumdar S, Khan M, Lotz J, Stevens RE,Klein R, Phelps KV (2002) The effects of synthetic conjugatedestrogens, a (cenestin) on trabecular bone structure and strength inthe ovariectomized rat model. Osteoporos Int 13:816–823
60. Gittens SA, Wohl GR, Zernicke RF, Matyas JR, Morley P, UludagH (2004) Systemic bone formation with weekly PTH administra-tion in ovariectomized rats. J Pharm Pharm Sci 7:27–37
61. Whitfield JF, Morley P, Ross V, Isaacs RJ, Rixon RH (1995)Restoration of severely depleted femoral trabecular bone inovariectomized rats by parathyroid hormone-(1–34). Calcif TissueInt 56:227–231
62. Trivedi R, Mithal A, Chattopadhyay N (2010) Anabolics inosteoporosis: the emerging therapeutic tool. Curr Mol Med 10:14–28
63. Sliwinski L, Janiec W, Pytlik M, Folwarczna J, Kaczmarczyk-Sedlak I, Pytlik W, Cegiela U, Nowinska B (2004) Effect ofadministration of alendronate sodium and retinol on the mechan-ical properties of the femur in ovariectomized rats. Pol JPharmacol 56:817–824
64. Sass DA, Bowman AR, Yuan Z, Ma Y, Jee WS, Epstein S (1997)Alendronate prevents cyclosporin A-induced osteopenia in the rat.Bone 21:65–70
65. Toolan BC, Shea M, Myers ER, Borchers RE, Seedor JG,Quartuccio H, Rodan G, Hayes WC (1992) Effects of 4-amino-1-hydroxybutylidene bisphosphonate on bone biomechanics inrats. J Bone Miner Res 7:1399–1406
66. Scalbert A, Manach C, Morand C, Remesy C, Jimenez L (2005)Dietary polyphenols and the prevention of diseases. Crit Rev FoodSci Nutr 45:287–306
67. de Boer VC, Dihal AA, van der Woude H, Arts IC, WolfframS, Alink GM, Rietjens IM, Keijer J, Hollman PC (2005) Tissuedistribution of quercetin in rats and pigs. J Nutr 135:1718–1725
68. Stalmach A, Mullen W, Pecorari M, Serafini M, Crozier A (2009)Bioavailability of C-linked dihydrochalcone and flavanone gluco-sides in humans following ingestion of unfermented andfermented rooibos teas. J Agric Food Chem 57:7104–7111
Osteoporos Int (2011) 22:3013–3027 3027
Related Documents








![Presence of Sarcopenia in Senescence Accelerated Mouse ...€¦ · induced osteoporotic bone. Bone. 2010;46:1299-305. [2] Guo AY; Leung KS; Qin JH; Chow SK; CheungWH*. Effect of low-magnitude](https://static.cupdf.com/doc/110x72/600eb838bfc6b86d2d3c4425/presence-of-sarcopenia-in-senescence-accelerated-mouse-induced-osteoporotic.jpg)


