THE JOURNAL 0 1985 by The American Society of Biological OF BIOLOGICAL CHEMISTRY Chemists, Inc. Vol. 260, No. 18, Issue of August 25, PP. 10271-10277,1985 Printed in U. S. A. Human 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase CONSERVEDDOMAINSRESPONSIBLEFORCATALYTICACTIVITYANDSTEROL-REGULATED DEGRADATION* (Received for publication, March 22, 1985) Kenneth L. Luskeyg and Bryn Stevens From the Departments of Molecular Genetics and Internal Medicine, University of Texas Health Science Center at Dallas, Dallas, Tex& 75235 A full length cDNA for human 3-hydroxy-3-meth- ylglutaryl coenzyme A reductase, the membrane- bound glycoprotein that regulates cholesterol synthe- sis, was isolated from a human fetal adrenal cDNA library. The nucleotide sequence of this cDNA shows that the human reductase is 888 amino acids long and shares a high degree of homology with the hamster enzyme. The amino-terminal membrane-bound domain is the most conserved region between the two species (7 substitutions out of 339 amino acids). This region, which is predicted to span the endoplasmic reticulum membrane seven times, mediates accelerated degra- dation of reductase in the presence of sterols. The carboxyl-terminal catalytic domain is also highlycon- served (22 substitutions out of 439 amino acids). How- ever, the linker region between these two domains has diverged (32 substitutions out of 110 amino acids). Conservation of the structure of the membrane-bound domain in HMG-CoA reductase supportsthe hypothesis that sterol-regulated degradation is an important mechanism for suppression of reductase activity and for regulation of cholesterol metabolism in humans as well as in hamsters. HMG-CoA reductase’ is the rate-limiting enzyme for cho- lesterol synthesis. Its activity is regulated via a negative feedback mechanism mediated by sterols and non-sterol me- tabolites derived from mevalonate, the product of the reaction catalyzed by reductase (1). Normally in mammaliancells this enzme is suppressed by cholesterol derived from the internal- ization and degradation of low density lipoprotein via the LDL receptor. Competitive inhibitorsof the reductaseinduce the expression of LDL receptors in the liver, which in turn increases the catabolism of plasma LDL and lowers the plasma concentration of cholesterol, an important determi- nant of atherosclerosis (2, 3). Recent studies of the HMG-CoA reductase in hamster cells have shown that this enzyme is encoded by a family of mRNAs of about 4.7 kb, which are transcribed from a 25-kb gene (4, * This research was supported by grants from the National Insti- tutes of Health (HL 20948) and the Clay Weed Memorial Cancer Fund Trust. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solelyto indicate this fact. $ Established Investigator of the American Heart Association. The abbreviations used are: HMG-CoA reductase, 3-hydroxy-3- methylglutaryl coenzyme A reductase; LDL, low density lipoprotein; SDS, sodium dodecyl sulfate; bp, base pairs; kb, kilobase pairs; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid. 5). Full length cDNAclones have been obtained and this has revealed the complete amino acid sequence of the enzyme (6). The hamster reductase is a 97-kDa glycoprotein bound to the membrane of the endoplasmic reticulum by a hydrophobic amino-terminal region that is predicted to span the endo- plasmic reticulum membrane seven times (7,8). The catalytic site of the reductase is in the carboxyl-terminal half of the enzyme, which projects into the cytoplasm of the cell (8). The genomic segments comprising this gene have also been isolated and the sequence of the promoter region has been determined (5). The reductase gene from hamster cells lacks a TATA orCCAAT box; transcription initiatesfrom multiple sites spread out over 100 bp in the genome. The 5’ untrans- lated region contains an intron of about 3.5 kb. Variable splicing at the 5’ end of this intron produces mRNA molecules with 5’ untranslated regions ranging between 68 and 670 nucleotides in length.’ In cultured cells, reductase activity can be suppressed either by cholesterol derived from LDL or by oxygenated derivatives of cholesterol solubilized in solvents and added to the culture medium (1). This suppression is accomplished by two mech- anisms, 1) decreased transcription of the gene (9) and 2) enhanced degradation of the protein (10-13). Inasmuch as both the mRNA and protein turn over with short half-lives, both of these mechanisms are efficient means of regulating reductase activity. Only limited information is available about HMG-CoA reductase in man, primarily due to the lack of availability of tissues that express high levels of the enzyme. Reductase from human liver is immunologically cross-reactive with the rat enzyme (14, 15) and can be inactivated by phosphorylation, just as the rat enzyme (15). Purification of reductase from human liver yields a single species of 52 kDa (14, 16). Purifi- cation of reductase from rat liver by a similar protocol also yields a species of 50-55 kDa. However, subsequent studies have shown that the purified enzyme represents a proteolytic fragment of the native protein whose true molecular weight is 97 kDa (6, 11, 17). In the current studies,we examine the structure and func- tion of human reductase. For this purpose, we have used the cloned hamster cDNA to select a full length cDNA for the human enzyme. The human cDNA was then sequenced to determine the primary structure of the human enzyme. Com- parison of the human amino acid sequence with that of the hamster enzyme has identified two highly conserved regions of the molecule that are separated by a less conserved region. The functional implications of these three domains are dis- cussed. * Reynolds, G. A., Goldstein, J. L., and Brown, M. S., J. Biol. Chem., in press. 10271

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL 0 1985 by The American Society of Biological
OF BIOLOGICAL CHEMISTRY Chemists, Inc.
Vol. 260, No. 18, Issue of August 25, PP. 10271-10277,1985 Printed in U. S. A.
Human 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase CONSERVED DOMAINS RESPONSIBLE FOR CATALYTIC ACTIVITY AND STEROL-REGULATED DEGRADATION*
(Received for publication, March 22, 1985)
Kenneth L. Luskeyg and Bryn Stevens From the Departments of Molecular Genetics and Internal Medicine, University of Texas Health Science Center at Dallas, Dallas, Tex& 75235
A full length cDNA for human 3-hydroxy-3-meth- ylglutaryl coenzyme A reductase, the membrane- bound glycoprotein that regulates cholesterol synthe- sis, was isolated from a human fetal adrenal cDNA library. The nucleotide sequence of this cDNA shows that the human reductase is 888 amino acids long and shares a high degree of homology with the hamster enzyme. The amino-terminal membrane-bound domain is the most conserved region between the two species (7 substitutions out of 339 amino acids). This region, which is predicted to span the endoplasmic reticulum membrane seven times, mediates accelerated degra- dation of reductase in the presence of sterols. The carboxyl-terminal catalytic domain is also highly con- served (22 substitutions out of 439 amino acids). How- ever, the linker region between these two domains has diverged (32 substitutions out of 110 amino acids). Conservation of the structure of the membrane-bound domain in HMG-CoA reductase supports the hypothesis that sterol-regulated degradation is an important mechanism for suppression of reductase activity and for regulation of cholesterol metabolism in humans as well as in hamsters.
HMG-CoA reductase’ is the rate-limiting enzyme for cho- lesterol synthesis. Its activity is regulated via a negative feedback mechanism mediated by sterols and non-sterol me- tabolites derived from mevalonate, the product of the reaction catalyzed by reductase (1). Normally in mammalian cells this enzme is suppressed by cholesterol derived from the internal- ization and degradation of low density lipoprotein via the LDL receptor. Competitive inhibitors of the reductase induce the expression of LDL receptors in the liver, which in turn increases the catabolism of plasma LDL and lowers the plasma concentration of cholesterol, an important determi- nant of atherosclerosis (2, 3).
Recent studies of the HMG-CoA reductase in hamster cells have shown that this enzyme is encoded by a family of mRNAs of about 4.7 kb, which are transcribed from a 25-kb gene (4,
* This research was supported by grants from the National Insti- tutes of Health (HL 20948) and the Clay Weed Memorial Cancer Fund Trust. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Established Investigator of the American Heart Association. The abbreviations used are: HMG-CoA reductase, 3-hydroxy-3-
methylglutaryl coenzyme A reductase; LDL, low density lipoprotein; SDS, sodium dodecyl sulfate; bp, base pairs; kb, kilobase pairs; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid.
5). Full length cDNA clones have been obtained and this has revealed the complete amino acid sequence of the enzyme (6). The hamster reductase is a 97-kDa glycoprotein bound to the membrane of the endoplasmic reticulum by a hydrophobic amino-terminal region that is predicted to span the endo- plasmic reticulum membrane seven times (7,8). The catalytic site of the reductase is in the carboxyl-terminal half of the enzyme, which projects into the cytoplasm of the cell (8).
The genomic segments comprising this gene have also been isolated and the sequence of the promoter region has been determined (5). The reductase gene from hamster cells lacks a TATA or CCAAT box; transcription initiates from multiple sites spread out over 100 bp in the genome. The 5’ untrans- lated region contains an intron of about 3.5 kb. Variable splicing at the 5’ end of this intron produces mRNA molecules with 5’ untranslated regions ranging between 68 and 670 nucleotides in length.’
In cultured cells, reductase activity can be suppressed either by cholesterol derived from LDL or by oxygenated derivatives of cholesterol solubilized in solvents and added to the culture medium (1). This suppression is accomplished by two mech- anisms, 1) decreased transcription of the gene (9) and 2) enhanced degradation of the protein (10-13). Inasmuch as both the mRNA and protein turn over with short half-lives, both of these mechanisms are efficient means of regulating reductase activity.
Only limited information is available about HMG-CoA reductase in man, primarily due to the lack of availability of tissues that express high levels of the enzyme. Reductase from human liver is immunologically cross-reactive with the rat enzyme (14, 15) and can be inactivated by phosphorylation, just as the rat enzyme (15). Purification of reductase from human liver yields a single species of 52 kDa (14, 16). Purifi- cation of reductase from rat liver by a similar protocol also yields a species of 50-55 kDa. However, subsequent studies have shown that the purified enzyme represents a proteolytic fragment of the native protein whose true molecular weight is 97 kDa (6, 11, 17).
In the current studies, we examine the structure and func- tion of human reductase. For this purpose, we have used the cloned hamster cDNA to select a full length cDNA for the human enzyme. The human cDNA was then sequenced to determine the primary structure of the human enzyme. Com- parison of the human amino acid sequence with that of the hamster enzyme has identified two highly conserved regions of the molecule that are separated by a less conserved region. The functional implications of these three domains are dis- cussed.
* Reynolds, G. A., Goldstein, J. L., and Brown, M. S., J. Biol. Chem., in press.
10271

10272 Structural Domains of HMG-CoA Reductase
EXPERIMENTAL PROCEDURES
Materials-A human genomic DNA library (18) was kindly pro- vided by Tom Maniatis of the Department of Biology, Harvard University, Boston, MA. A human fetal adrenal cDNA library (19) was kindly provided by Tokuo Yamamoto of the Department of Molecular Genetics, University of Texas Health Science Center at Dallas, Dallas, TX. Other materials were obtained from previously described sources.
Isolation and Sequencing of Genomic and cDNA Clones-Recom- binant bacteriophage were screened by the method of Benton and Davis (20). Hybridization was performed in buffer A (buffer A = 0.75 M NaC1, 75 mM sodium citrate, 50 mM sodium phosphate, pH 7.0, 0.2% bovine serum albumin, 0.2% Ficoll, 0.2% polyvinyl pyrollidone, 200 pg/ml denatured salmon sperm DNA, and 0.1% SDS) plus 30% formamide with -2 X lo5 cpm/ml of 32P-labeled DNA probe at 42 "C for 16 h. 32P-labeled DNA probes were prepared by random hexanu- cleotide-primed labeling (21). Filters were washed in 0.3 M NaC1, 30 mM sodium citrate (pH 7.0), and 0.1% SDS at 55 "C.
The cDNA library was first enriched for full length cDNAs by linearizing plasmid DNA from the total human fetal adrenal cDNA library with the restriction endonuclease SalI, fractionating the digest
than 6.5 kb in length, religating the DNA, and transforming E. coli on low melting temperature agarose, recovering plasmid DNA greater
HBlOl (22). The resulting clones were screened by colony hybridi- zation as described by Grunstein and Hogness (23). Hybridizations were performed in buffer A plus 50% formamide a t 42 "C for 16 h. Filters were washed in 15 mM NaC1, 1.5 mM sodium citrate (pH 7.0), and 0.1% SDS at 55 "C.
Hybridization-positive genomic and cDNA clones were isolated and phage or plasmid DNA was prepared and characterized by stand- ard techniques (24). DNA sequencing was performed by the dideoxy chain termination method after subcloning DNA fragments into bacteriophage M13 vectors (25, 26). Specific oligonucleotide primers were synthesized by the phosphoramidite method (27) using the Applied Biosystems 380A oligonucleotide synthesizer. Computer analysis of DNA sequences was done on an IBM personal computer using the Beckman Microgenie DNA analysis program (28).
Cell Culture-Cells were grown in monolayer culture at 37 "C in an atmosphere of 5-7% CO,. Stock cultures of Chinese hamster ovary cells were grown in medium A (Ham's F-12 containing 25 mM Hepes (pH 7.4), 100 units/ml of penicillin, 100 pg/ml streptomycin, and 2 mM glutamine) supplemented with 10% (v/v) fetal calf serum. Stock cultures of UT-2 cells (29) were grown in medium A supplemented with 10% fetal calf serum and 0.2 mM mevalonate. Stock cultures of TR-74 cells (see below) were grown in medium A supplemented with 10% lipoprotein-deficient serum. For growth experiments, on day 0 cells from stock flasks were dissociated with trypsin/EDTA and 1.5 X 10' cells were seeded into a 35-mm well in 2 ml of medium A supplemented with 10% fetal calf serum with 0.2 mM mevalonate. On days 1 and 3, the cells were fed medium A supplemented with 10% fetal calf serum either with or without 0.2 mM mevalonate. On day 5, the dishes were fixed with 95% ethanol and stained with 1% crystal violet. For experiments in which reductase activity was measured, on day 0, 3.5 x IO4 Chinese hamster ovary cells or 6 X lo4 TR-74 cells were seeded in 60-mm Petri dishes in 3 ml of medium A supplemented with 10% fetal calf serum. The cells were fed on days 2 and 3 with 3 ml of medium A supplemented with 10% lipoprotein-deficient serum. Additions of sterols were made on days 3 and 4.
Isolation of TR--74 Cells-Plasmid pHRed-102 was introduced in the UT-2 cell line by co-transfection with pSV3-Neo using the cal- cium phosphate precipitation method (30). A co-precipitate of pHRed-102 (9.5 pg) and pSV3-Neo (0.5 pg) was added to 5 X lo5 UT- 2 cells in a 100-mm dish in medium B (Dulbecco's modified Eagle's medium containing 100 units/ml of penicillin, 100 pg/ml streptomy- cin, and 34.5 mg/l of proline) supplemented with 10% fetal calf serum and 0.2 mM mevalonate. After 5 h, the medium was removed, the cells were exposed to medium B plus 10% glycerol for 4 min at 24 "C, and the medium was replaced with medium B supplemented with 10% fetal calf serum plus 0.2 mM mevalonate. The following day the medium was changed to medium B supplemented with 10% fetal calf serum and 700 yg/ml G418 lacking mevalonate. This medium was changed every 3 to 4 days until individual colonies could be picked and expanded in medium B supplemented with 10% fetal calf serum and 350 pg/ml G418. One of these clones, designated TR-74, was subsequently adapted to grow in medium A supplemented with 10% lipoprotein-deficient serum.
Other Assays-Protein was measured by a modified Lowry proce- dure (31). Enzymatic activity of HMG-CoA reductase was measured in detergent-solubilized cell extracts as described (32). One unit of reductase activity represents the formation of 1 nmol of [14C]meva- lonate per min a t 37 "C.
RESULTS
The following strategy was used to isolate a full length cDNA for the human HMG-CoA reductase. First, a human genomic clone was selected with a probe derived from the hamster cDNA. This provided a homologous probe to the 5' end of the human reductase mRNA that was used to screen a cDNA library from the fetal adrenal, a tissue that expresses high levels of HMG-CoA reductase. The use of a human specific probe allowed the screening of the cDNA library to be done at higher stringency with a lower background. Inas- much as the probe was confined to the 5' end of the coding region, only full length cDNA clones would be detected.
A human fetal liver genomic library was screened under reduced stringency with the 1.5-kb PstI fragment from the hamster cDNA for HMG-CoA reductase, pRed-227 (6). This fragment includes 163 bp of 5' untranslated sequence and 1347 bp of the coding region. Southern blots of human ge- nomic DNA indicated that this fragment recognized a unique sequence in the human genome (data not shown). Approxi- mately 1 X IO6 recombinant phage were screened and one clone, designated AHRed-1, was isolated and DNA was pre- pared for further characterization. Studies of AHRed-1 DNA showed that the 0.9-kb BanHI fragment contained sequences that hybridized with the most 5' 500 bp of the hamster cDNA (Fig. 1A). When smaller fragments of this region were se- quenced, it was found that the 149-bp BarnHI/BglII fragment plus 36-bp 3' of the BglII site was 90% homologous with residues -23 to 165 of the hamster cDNA and corresponded to the presumed second exon of the human reductase gene
A. XHRed-1 kilobores l ~ " ' , " ~ ~ , " ' l . 0 5 j0 14
PH H 4
- - 0 200 400 600 bore pairs
B. pHRed-IO2 kilobases
I ~ " . I " " I " " I " " I I 0 1 2 4 43
ME ?g y H P P Bg
'" - " - " - - - - 1
" " " "
- - - d - I " " " I
I#- "
5
- - - 1
r - FIG. 1. Restriction maps and sequencing strategies for ge-
nomic and cDNA clones for human HMG-CoA reductase. Panel A, the genomic clone, XHRed-1, is shown with restriction sites as indicated. The region surrounding the second exon of the reductase gene is expanded beneath the map of XHRed-1 and the sequencing strategy of this region of the genome is shown. The 430-bp EcoRI/ BglII fragment from this region was used to isolate the human reductase cDNA. Panel B, the cDNA clone pHRed-102 is shown with restriction sites as indicated. Restriction fragments that were sub- cloned into MI3 bacteriophage vectors are indicated beneath the cDNA restriction map. The arrows beneath each fragment indicate the direction and extent of sequencing from each oligonucleotide primer. Restriction enzymes are abbreviated as follows: B, BamHI; Bg, BglII; E, EcoRI; H , HindIII; P, PstI.

Struc tura l Domains of HMG-CoA Reductase 10273
(data not shown). The 430-bp EcoRI/BglII fragment that included a portion of the first intron and 150 bp of the second exon was then used to isolate a full length cDNA for the human HMG-CoA reductase.
A size-fractionated human fetal adrenal library (19) was screened using the EcoRI/BglII genomic fragment from XHRed-1. Approximately 3 X lo4 colonies were screened and one positive clone, designated pHRed-102, was identified. This plasmid contained a 4.3-kb insert with the restriction map indicated in Fig. 1B. Restriction fragments of pHRed- 102 were subcloned into M13 bacteriophage vectors and sub- jected to DNA sequencing as indicated in Fig. 1B.
The nucleotide sequence of the human reductase cDNA is shown in Fig. 2. The sequence contains an open reading frame of 2664 nucleotides that would encode an 888-amino acid protein. The first nucleotide of the ATG that begins this open reading frame is designated as position +l. Fifty nucleotides of 5’ untranslated sequence are present in this cDNA clone. The 3‘ untranslated region is approximately 1.6 kb long. Within this region, the sequence of 187 bp 3’ of the coding region was determined.
The nucleotide sequences of the human and hamster reduc- tase mRNAs show a high degree of homology throughout. Within the 5’ untranslated region, the sequences are 74% identical when the sequence of the hamster reductase is aligned so that the donor splice site of the first intron in the hamster gene is located at position -669 (5). Transcripts that splice at this position represent a large number of the reduc- tase transcripts within hamster cells.’ Within the 187 bp of 3’ untranslated sequence that was determined in the human cDNA, there is 85% identity with the hamster sequence with a single nucleotide gap inserted in the hamster sequence to maximize the alignment (Fig. 2).
The coding region of the human reductase also shares a high degree of homology with the hamster sequence both at the nucleotide level (87%) and at the protein level (93%) as deduced from the translated amino acid sequences. Fig. 2 indicates mismatches between the cDNA sequences (aster- isks) and substitutions between the protein sequences (boxes). The coding sequence of the human enzyme is one amino acid longer than that of the hamster, so a gap of three nucleotides has been introduced in the hamster sequence after position 1261 (amino acid residue 420) to maximize the homology.
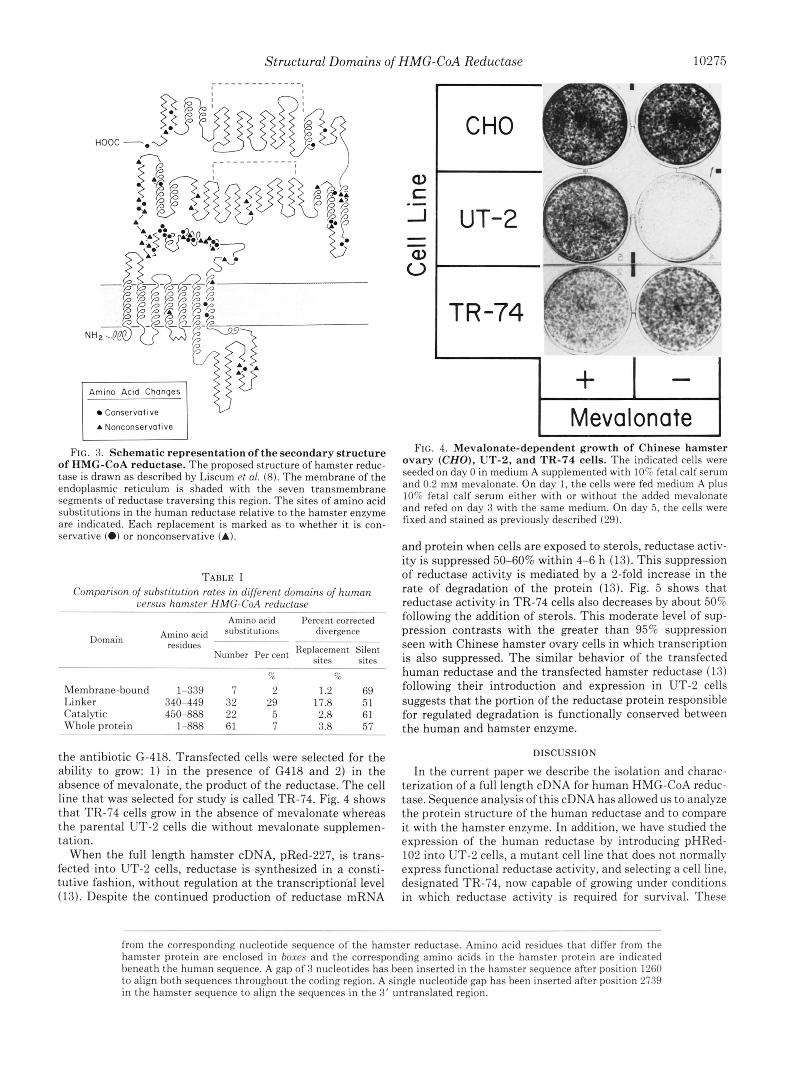
Fig. 3 shows that the distribution of amino acid substitu- tions is nonrandom relative to the predicted protein structure of the reductase enzyme. In this diagram, a model for the protein structure of the hamster reductase derived from pro- tein structural studies and computer modeling is presented (8). Sites of amino acid substitutions in the human enzyme relative to the hamster enzyme are marked. These substitu- tions are coded as to whether they are conservative (i.e. the substitution of a hydrophobic, polar uncharged, acidic, or basic residue for a similar class residue) or nonconservative ( i e . substitution from a dissimilar class) (33).
Within the first 339 residues, 7 amino acid substitutions are noted. This region is extremely hydrophobic and is pre- dicted to span the membrane of the endoplasmic reticulum seven times (8). Only three substitutions are found within the membrane-spanning portions of this sequence. Two of these replacements are conservative substitutions of one hydropho- bic amino acid for another while the third at position 139 is the replacement of an alanine with a threonine, the sixth most common amino acid replacement found in proteins (34). The hydrophilic sequences that connect one transmembrane region with the next are also highly conserved the parts of the protein appearing at both the cytoplasmic and luminal
surfaces are identical with the exception of the large intralu- minal loop between the sixth and seventh transmembrane regions. Four amino acid substitutions occur within this loop. However, they do not affect the site for N-linked glycosylation at position 281, which is the presumed site of the high man- nose carbohydrate chain in the hamster enzyme (7, 8).
Proteolytic experiments have shown that the catalytic ac- tivity of the reductase is confined to a 53,000-Da fragment of the protein (8). Antibodies directed against synthetic peptides that corrspond to segments of the hamster reductase have allowed Liscum et al. (8) to determine that this proteolytic fragment includes the carboxyl terminus of the enzyme and results from a proteolytic clip that occurs around residues 450 to 470. When the human and hamster reductase sequences are compared within this catalytic domain, only 7% of the residues differ between the two species. In particular, the two @-barrel regions predicted in the reductase structure (8) are the most conserved part of this region. Most of the substitu- tions in the catalytic protion of the enzyme occur in sequences that flank the @-barrels and probably represent surfaces of the reductase molecule that interact with the aqueous solvent.
In contrast to the high degree of conservation in the mem- brane-bound and catalytic domains of the protein, the linking portion of the protein, between residues 340 and 449, has diverged substantially. Within this 110-residue segment, 32 amino acid substitutions are found. This region is also the site at which an additional residue has been inserted in the human sequence. The hamster reductase has an unusual sequence, X-Y-Z-Pro, that is repeated contiguously four times (8). This sequence resembles a structure originally observed in the avian pancreatic polypeptide. In the human reductase, the third proline is substituted with a glutamine, but the other three prolines are intact. The most divergent region of the human reductase is found between residues 375 and 430, which is a region that was predicted to have little ordered secondary structure in the hamster protein (8).
The nonrandom distribution of amino acid substitutions is likely attributable to selective constraint on different regions of the protein during evolution. To rule out the alternate possibility ( i x . a difference in the mutation rate at the nu- cleotide level), we compared the rate of silent substitutions with that of replacement substitutions in the three domains of the reductase. The per cent corrected divergence for the hamster and human reductase sequences were calculated ac- cording to the method of Perler et al. (35) and are listed in Table I. The rates of replacement substitutions were 1.2%, 17.8%, and 2.8% in the membrane-bound, linker, and catalytic domains, respectively. In contrast to this large variation, the rates of silent site substitutions were quite similar-70%, 51%, and 61% in the respective domains. These data suggest that the DNA has evolved at similar rates in these three domains except when constrained by replacement amino acid substitutions.
To evaluate the functional behavior of the human reductase cDNA, we introduced the plasmid containing the cDNA into hamster cells by calcium phosphate-mediated transfection. The vector in which the human cDNA was cloned contains the SV40 early promoter region with an intron donor and acceptor splice site upstream from the cDNA insert, which allows expression of the gene in hamster cells (22). As recip- ients we used UT-2 cells, a mutant line of Chinese hamster ovary cells that lacks HMG-CoA reductase activity and re- quires exogenous mevalonate for growth (29). The plasmid containing the human reductase cDNA (pHRed-102) was transfected into UT-2 cells together with pSV3-Neo, a plas- mid that contains a bacterial gene conferring resistance to

10274
- 5 0
*l *I
121 41
241 81
361 121
481 161
601 201
721 241
841 281
961 321
1081 361
1201 401
I321 441
1441 481
I561 521
1681 561
1801 601
1921 641
2041 681
2161 721
2281 761
2401 801
2521 84 1
2641 881
2761
Structural Domains of HMG-CoA Reductase
. . t t t .
* * * *
m . * . t . . GAACCTCGGCCTAATGAAGAATGTCTACAG CAGAGAAAGGTGCAAAA TTCCTTAGTGATGCTGAGATCATCCAGTTA GTCAATGCTAAGCATATCCCA6CCTACAAG GluProArgProAsnGluGluCysLeuGln I a G l u L y r G l y A l a L y s PheLeuSerAspAlaGlullelleGlnLeu YalAsnAlaLysHis l leProAl lTyrLys
... TTGGAAACTCTGATGGAAACTCATGAGCGT GGTGTATCTATTCGCCGACAGTTACTTTCC LeuGluThrLevMetGluThrHisGluArg GlyYalSerIleArgArgGlnLeuLevSer
. t. . . . . *
TTGGTGATGGGAGCTTGTTGTGAGAATGTT ATTGGATATATGCCCATCCCTGTTGGAGTG LeuValWetGlyAlaCysCysGluAsnVal IleGlyTyrMetProlleProValGlyVal
t .
TGTCTTGTGGCCAGCACCAATAGAGGCTGC AGAGCAATAGGTCTTGGTGGAGGTGCCAGC AGCCGAGTCCTTGCAGATGGGATGACTCGT G6CCCAGTTGTGCGTCTTCCACGTGCTTGT CysLeuVaIAlaSerThrAsnArgGlyCys ArgAlalleGlyLeuGlyGlyGlyAlaSer SerArgValLeuAlaAspGlyMetThrArg GlyProVaIValArgLeuProArgAlaCYS
t .
I r t
* * t t t
. . . * . t * t
t t . . t. . * . t .
TACATTGCCTGTGGACAGGATGCA6CACAG AATGTTGGTAGTTCAAACTGTATTACTTTA ATGGAAGCAAGTGGTCCCACAAATGAAGAT TTATATATCAGCTGCACCATGCCATCTATA TyrlleAlaCysGlyGlnAspAlaAlaGln AsnValGlySerSerAsnCyslleThrLev MetGluAlaSerGlyProThrAsnGluAsp LeuTyrlleSerCysThrMetProSerlle
. . t t
. . . . GAGATAGGAACGGTGGGTGGTGGGACCAAC CTACTACCTCAGCAAGCCTGTTTGCAGATG CTAGGTGTTCAAGGAGCATGCAAAGATAAT CCTGGGGAAAATGCCCGGCAGCTTGCCCGA GlulleGlyThrValGlyGlyGlyThrAsn LeuLeuProGlnGlnAlaCysLeuGlnMet LeuGlyValGlnGIyAlaCysLysAspAsn ProGlyGluAsnAlaArgGlnLeuAlaArg
. . . .
ATTGTGTGTGGGACCGTAATGGCTG6GGAA TTGTCACTTATGGCAGCATTGGCAGCAGGA lleValCysGlyThrVallletAlaGlyGlu LeuSerLeuMetAlaAlaLeuAlaAlaGly
. .t . . t
GG C GCACCAAGAA AC CCTGAATA GCCCGACAGTTCTGAACT6GAACATGGGCA TTGGGTTCTAAAGGACTAACctTAAAATCTG TGAATTMAAAAGCTCAATGCATTGTCTTG G l ~ y s T h r L y s L y @ l : * * . " **
. . *. f . .It .. . . . . . . .
TGGAGGATGAATAAATGTGATCACTGAGAC AGCCACTTGGTTTTTGGCTCTTTCAGAGAG GTCTCAGGTTCTTTCCATGCAGACTCCTCA GATC
FIG. 2. Nucleotide sequence and deduced amino acid sequence of pHRed-102 and comparison with the sequence of the hamster reductase. The nucleotide sequence derived from sequencing pHRed-102 as described in Fig. 1 is shown. The ATG that initiates translation of the reductase protein is labeled +l. The 5' untranslated sequence is numbered -50 to -1. The translated 888-amino-acid-residue sequence of the human reductase is listed beneath residues +1 to 2667. Asterisks above the nucleotide sequence indicate bases that differ
120 10
240 BO
360 120
480 160
600 200
720 240
840 280
960 320
360 1080
400 1200
1320 440
480 1440
1560 520
560 1680
1800 600
1920 640
2040 680
2160 720
2280 760
2400 800
2520 840
2640 880
2760 888
2904
Struc tura l Domains of HMG-CoA Reductase 10275
L"J Conservative W
A Nonconservative
FIG. 3. Schematic representation of the secondary structure of HMG-CoA reductase. The proposed structure of hamster reduc- tase is drawn as described by Liscum et a!. (8). The membrane of the endoplasmic reticulum is shaded with the seven transmembrane segments of reductase traversing this region. The sites of amino acid substitutions in the human reductase relative to the hamster enzyme are indicated. Each replacement is marked as to whether it is con- servative (*) or nonconservative (A).
TARLE I Comparison of substitution rates in different domains of human
versus hamster HMG-CoA reductase Amino acid Percent corrected
Amino acid substitutions divergence Domain residues Number Per c e n ~ Replacement Silent
sites sites c. 5
Membrane-bound 1-339 7 2 1.2 69 Linker 340-449 32 29 17.8 51 Catalytic 450-888 22 5 2.8 61 Whole protein 1-888 61 7 3.8 57
the antibiotic G-418. Transfected cells were selected for the ability to grow: 1) in the presence of G418 and 2) in the absence of mevalonate, the product of the reductase. The cell line that was selected for study is called TR-74. Fig. 4 shows that TR-74 cells grow in the absence of mevalonate whereas the parental UT-2 cells die without mevalonate supplemen- tation.
When the full length hamster cDNA, pRed-227, is trans- fected into UT-2 cells, reductase is synthesized in a consti- tutive fashion, without regulation at the transcriptional level (13). Despite the continued production of reductase mRNA
I Mevalonate I FIG. 4. Mevalonate-dependent growth of Chinese hamster
ovary (CHO), UT-2, and TR-74 cells. The indicated cells were seeded on day 0 in medium A supplemented with 10% fetal calf serum and 0.2 mM mevalonate. On day 1, the cells were fed medium A plus 10% fetal calf serum either with or without the added mevalonate and refed on day 3 with the same medium. On day 3, the cells were fixed and stained as previously described (29).
and protein when cells are exposed to sterols, reductase activ- ity is suppressed 50-60% within 4-6 h (13). This suppression of reductase activity is mediated by a 2-fold increase in the rate of degradation of the protein (13). Fig. 5 shows that reductase activity in TR-74 cells also decreases by about 50% following the addition of sterols. This moderate level of sup- pression contrasts with the greater than 95% suppression seen with Chinese hamster ovary cells in which transcription is also suppressed. The similar behavior of the transfected human reductase and the transfected hamster reductase (13) following their introduction and expression in UT-2 cells suggests that the portion of the reductase protein responsible for regulated degradation is functionally conserved between the human and hamster enzyme.
DISCUSSION
In the current paper we describe the isolation and charac- terization of a full length cDNA for human HMG-CoA reduc- tase. Sequence analysis of this cDNA has allowed us to analyze the protein structure of the human reductase and to compare it with the hamster enzyme. In addition, we have studied the expression of the human reductase by introducing pHRed- 102 into UT-2 cells, a mutant cell line that does not normally express functional reductase activity, and selecting a cell line, designated TR-74, now capable of growing under conditions in which reductase activity is required for survival. These
from the corresponding nucleotide sequence of the hamster reductase. Amino acid residues that differ from the hamster protein are enclosed in boxes and the corresponding amino acids in the hamster protein are indicated beneath the human sequence. A gap of 3 nucleotides has been inserted in the hamster sequence after position 1260 to align both sequences throughout the coding region. A single nucleotide gap has heen inserted after position 2739 in the hamster sequence to align the sequences in the 3' untranslated region.

10276 Structural Domains of HMG-CoA Reductase
I \. 2 0 -
, - m , , , , , , /CHo
- 0 6 12 i s 24 Time After Addition of Sterols
(hours)
FIG. 5. Time course of suppression of HMG-CoA reductase activity in Chinese hamster ovary (CHO) (B) and TR-74 (0) cells after incubation with 25-hydroxycholesterol plus cho- lesterol. On day 0, cells were seeded in medium A supplemented with 10% fetal calf serum. On days 2 and 3, cells were fed medium A supplemented with 10% lipoprotein-deficient serum. Additions of 25- hydroxycholesterol (1 pg/ml) and cholesterol (12 pg/ml) were made in a staggered fashion so that all the cells were harvested at the same time on day 4. The 100% of control values for reductase activity (units/mg of protein) were 2.3 (B) and 1.6 (0). Each value represents the mean of triplicate incubations.
studies reveal that the human reductase is functional in hamster cells and moreover that the rate of degradation of the human enzyme appears to be susceptible to regulation by the hamster degradative system.
A comparison of the amino acid sequences of the human and hamster enzymes reveals that many regions are stongly conserved, while others vary. The complex membrane-asso- ciated domain of the reductase is the most highly conserved part of the protein. The catalytic water-soluble domain is also highly conserved although the rate of amino acid substitutions here is about twice the rate seen in the membrane-associated domain. In contrast, the linker region that connects these two domains has diverged at a rate 10-20-fold greater than the rest of the protein.
The regulated degradation of reductase is a unique feature of this membrane-bound enzyme. Under normal metabolic conditions, reductase has a half-life of 2-3 h (36-39). This turnover rate is much faster than that of most cytoplasmic or membrane-bound proteins (37), making the rate of degrada- tion an important mechanism by which sterols can regulate reductase activity in cells. The rate of degradation of reductase is retarded when cells are grown in the absence of exogenous cholesterol (38, 39) and further retarded when the cells are grown in the presence of compactin (10, 13) which prevents endogenous cholesterol synthesis. When compactin-treated cells are supplied with LDL as a source of sterols, the rate of degradation of reductase can increase up to 10-fold (10, 13). This enhanced rate of degradation coupled with a 90% de- crease in the rate of transcription of reductase mRNA (9) accounts for the more than 95% decrease in reductase activity under these conditions.
Recent studies by Gil et al. (46) show that the membrane- bound portion of reductase is responsible for regulated deg- radation of the protein. Expression studies with the hamster cDNA have shown that the nucleotides corresponding to amino acids 10 through 340 can be deleted from the reductase cDNA, without impairing the catalytic activity of the enzyme. However, in contrast to the native membrane-bound form of the enzyme, the shortened protein (60-kDa) now exists as a cytosolic enzyme with a prolonged half-life of greater than 24 h and is no longer subject to accelerated degradation after the addition of sterols (46).
The finding of strong conservation between the membrane domains of the hamster and human reductase supports pre- vious speculation about the functional importance of this region (8). The complex arrangement of seven transmembrane portions with short connecting segments on the luminal and cytoplasmic sides of the membrane of the ER as well as the site for N-linked glycosylation has been maintained in the two species. Even conservative substitutions of one hydropho- bic amino acid for another hydrophobic residue seem not to be tolerated.
How does the conservation within this domain of the re- ductase compare with that of other proteins? Soluble proteins vary widely in their rates of evolution depending on the functions of the proteins and the evolutionary constraints placed on their sequences. Histone IV has had only 2 amino acid substitutions out of 102 residues from plants to mammals (34), whereas the fibrinopeptides have diverged greatly over a short period of evolutionary time (34). Even different parts of the same protein can diverge at different rates depending on the functional importance of the residues. Within hemo- globin, those residues involved in heme binding are conserved much greater than residues on the surface of the protein (40). Membrane proteins display the same diversity of evolutionary rates as soluble proteins. Proteins that span the membrane only once and probably serve primarily just to anchor the protein to the membrane display a high rate of amino acid substitutions within the hydrophobic stretch of the protein. For example, the membrane-spanning region of the LDL receptor is only 68% conserved between the bovine and human sequences (19,41). Similarly the membrane-spanning portion of glycophorin is only 55-70% conserved among the horse, human, and pig sequences (42).
In contrast to proteins anchored to the membrane by a single transmembrane sequence, those proteins that have complex hydrophobic regions that span the membrane several times are believed to perform functions within the membrane. The acetylcholine receptor is such a protein (43). It is a complex of a, p, y, and 6 subunits in the ratio of 2:l:l:l. Each of these subunits contains four stretches of hydrophobic res- idues that span the membrane. The intact receptor binds acetylcholine and forms an ion channel through the mem- brane. Within this complex receptor, different rates of evo- lution are found between the membrane-spanning portions of different subunits and different transmembrane regions of each subunit. In the a subunit, the first three membrane- spanning regions (Ml, M2, and M3) are very strongly con- served among species as diverse as humans and the fish, Torpedo californiu (91-100% conservation) (43). On the other hand, other hydrophobic membrane-spanning regions in the same receptor complex are less conserved. For example, the fourth membrane-spanning region (M4) of the /3 subunit is only 53% conserved between calf and Torpedo (44). Compar- ison of the human and bovine sequence of rhodopsin, a protein in the rod outer segment that spans the rod disk membrane seven times and is responsible for light reception, also illus- trates the high degree of conservation of transmembrane sequences of complex membrane proteins. The seven trans- membrane stretches are conserved 96,91, 100,96,89,92, and 88% (45).
The striking amount of conservation within the membrane- associated portions of proteins that span a membrane several times indicates the degree of evolutionary constraint placed on these regions. Why are conservative substitutions of hy- drophobic amino acids not tolerated in these regions as easily as in proteins that only span the membrane once? Although

Structural Domains of HMG-CoA Reductase 10277
the three-dimensional intramembranous structure of these proteins is not known, it is possible that packing of neighbor- ing helical regions requires that the side chains fit precisely together. Replacement of a small hydrophobic side chain with a longer one might not allow the tight packing necessary for the protein to fold within the membrane. Charged residues are also found in these transmembrane regions and the charges are presumably neutralized by interactions with other charged elements within the lipid bilayer. Substitutions might hinder the interaction that is necessary to neutralize the charge. In addition, the functional role of these membrane segments and the mechanism by which they work is not known. Their sequence might be conserved so that they maintain their function just as the active site of an enzyme is conserved to preserve function.
The membrane-domain of HMG-CoA reductase is one of the most conserved of the membrane regions known to date. Clearly its role in mediating the rapid and regulated degra- dation of the catalytic portion of the enzyme is an important one. It is likely that as more proteins associated with the endoplasmic reticulum are characterized, others will be found in which a complex membrane domain is present that can affect degradation of the protein. Such a mechanism of control could represent a unique mechanism for metabolic regulation of membrane-bound proteins within mammalian cells.
Acknowledgments-We are grateful for the enthusiastic support of Joseph L. Goldstein and Michael S. Brown and to David Russell for assistance with DNA sequencing. Kim Victor, James Cali, Mark Woefle, and Edith Womack provided excellent technical assistance.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
REFERENCES Brown, M. S., and Goldstein, J. L. (1980) J. Lipid Res. 21 , 505-
517 Kovanen, P. T., Bilheimer, D. W., Goldstein, J. L., Jaramillo, J.
J., and Brown, M. S. (1981) Proc. Natl. Acad. Sci. U. S. A. 78,
Bilheimer, D. W., Grundy, S. M., Brown, M. S., and Goldstein, J. L. (1983) Proc. Natl. Acad. Sci. U. S. A. 80,4124-4128
Chin, D. J., Luskey, K. L., Faust, J. R., MacDonald, R. J., Brown, M. S., and Goldstein, J . L. (1982) Proc. Natl. Acad. Sci. U. S. A. 7 9 , 7704-7708
Reynolds, G. A., Basu, S. K., Osborne, T. F., Chin, D. J., Gil, G., Brown, M. S., Goldstein, J . L., and Luskev. K. L. (1984) Cell
1194-1198
38,275-285 " I . ,
Chin, D. J., Gil, G.. Russell, D. W., Liscum. L.. Luskev. K. L.. Basu, S. K., Okayama, H., Berg, P.; Goldstein, J. L., andBrown; M. S. (1984) Nature 308,613-617
Liscum, L., Cummings, R. D., Anderson, R. G. W., DeMartino, G. N., Goldstein, J . L., and Brown, M. S. (1983) Proc. Natl. Acad. Sci. U. S. A. 80, 7165-7169
Liscum, L., Finer-Moore, J., Stroud, R. M., Luskey, K. L., Brown, M. S., and Goldstein, J. L. (1985) J. Biol. Chern. 260 , 522-530
Luskey, K. L., Faust, J. R., Chin, D. J., Brown, M. S., and Goldstein, J. L. (1983) J. Bwl. Chem. 258, 8462-8469
Faust, J. R., Luskey, K. L., Chin, D. J., Goldstein, J. L., and Brown, M. S. (1982) Proc. Natl. Acad. Sci. U. S. A. 7 9 , 5205- 5209
Edwards, P. A., Lan, S.-F., Tanaka, R. D., and Fogelman, A. M. (1983) J. Biol. Chem. 258, 7272-7275
Edwards, P. A., Lan, S.-F., and Fogelman, A. M. (1983) J. Biol. Chem. 258, 10219-10222
Chin, D. J., Gil, G., Faust, J. R., Goldstein, J. L., Brown, M. S., and Luskey, K. L. (1985) Mol. Cell. Biol. 5, 634-641
14. Beg, Z. H., Stonik, J. A., and Brewer, H. B., Jr. (1982) Biochem. Biophys. Res. Commun. 107,1013-1020
15. Angelin, B., Einarsson, K., Liljeqvist, L., Nilsell, K., and Heller, R. A. (1984) J. Lipid Res. 2 5 , 1159-1166
16. Tanaka, R. D., Edwards, P. A., Lan, S.-F. H., Knoppel, E. M., and Fogelman, A. M. (1982) J. Lipid Res. 23,523-530
17. Liscum, L., Luskey, K. L., Chin, D. J., Ho, Y. K., Goldstein, J. L., and Brown, M. S. (1983) J. Biol. Chern. 2 5 8 , 8450-8455
18. Lawn, R. M., Fritsch, E. F., Parker, R. C., Blake, G., and Maniatis, T. (1978) Cell 15 , 1157-1174
19. Yamamoto, T., Davis, C. G., Brown, M. S., Schneider, W. J., Casev. M. L.. Goldstein. J. L., and Russell, D. W. (1984) Cell 39 , i7-38 '
20. Benton, W. D., and Davis, R. W. (1977) Science 196 , 180-182 21. Feinberg, A. P., and Vogelstein, B. (1983) Anal. Biochem. 132 ,
6-1 3 22. Okayama, H., and Berg, P. (1983) Mol. Cell. Biol. 3,280-289 23. Grunstein, M., and Hogness, D. S. (1975) Proc. Natl. Acad. Sci.
24. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Laboratory Manual, pp. 1-545, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
- "
U. S. A. 72,3961-3965
25. Messing, J . (1983) Methods Enzymol. 101,20-78 26. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
Acad. Sci. U. S. A. 74,5463-5467 27. Caruthers, M. H., Beaucage, S. L., Becker, C., Efcavitch, W.,
Fisher, E. F., Galluppi, G., Goldman, R., deHaseth, P., Martin, F., Matteucci, M., and Stabinsky, Y. (1982) Genetic Engineering
28. Queen, C., and Korn, L. J. (1984) Nucleic Acids Res. 12,581-599 29. Mosley, S. T., Brown, M. S., Anderson, R. G. W., and Goldstein
J. L. (1983) J. Biol. Chem. 258, 13875-13881 30. Wigler, M., Sweet, R., Sim, G. K., Wold, B., Pellicer, A., Lacy,
E., Maniatis, T., Silverstein, S., and Axel, R. (1979) Cell 16,
31. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
32. Goldstein, J. L., Basu, S. K., and Brown, M. S. (1983) Methods
33. Lehninger, A. L. (1982) in Principles of Biochemistry, pp. 100-
34. Doolittle, R. F. (1979) The Proteins, 3rd Ed, Vol. 4, pp. 1-118,
35. Perler, F., Efstratiadis, A,, Lomedico, P., Gilbert, W., Kolodner,
36. Edwards, P. A., and Gould, R. G. (1974) J. Biol. Chern. 249 ,
37. Schimke, R. T. (1975) in Methods in Membrane Biology (Korn, E. D., ed) Vol. 3, pp. 201-236, Plenum Press, New York
38. Brown, M. S., Dana, S. E., and Goldstein, J. L. (1974) J. Biol. Chem. 249, 789-796
39. Kirsten, E. S., and Watson, J. A. (1974) J. Biol. Chem. 249,
40. Kimura, M., and Ohta, T. (1973) Genetics Supplement 73, 19-35 41. Russell, D. W., Yamamoto, T., Schneider, W. J., Slaughter, C. J.,
Brown, M. S., and Goldstein, J. L. (1983) Proc. Natl. Acad. Sci.
42. Murayama, J.-I., Tomita, M., and Hamada, A. (1982) J. Membr.
43. Noda, M., Furutani, Y., Takahashi, H., Toyosato, M., Tanabe, T., Shimizu, S., Kikyotani, S., Kayano, T., Hirose, T., Inayama, S., and Numa, S. (1983) Nature 305, 818-823
44. Tanabe, T., Noda, M., Furutani, Y., Takai, T., Takahashi, H., Tanaka, K., Hirose, T., Inayama, S., and Numa, S. (1984) Eur. J. Biochem. 144, 11-17
45. Nathans, J., and Hogness, D. S. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,4851-4855
46. Gil, G., Faust, J. R., Chin, D. J., Goldstein, J. L., and Brown, M. S. (1985) Cell 41,249-258
4 , 1-17
777-785
(1951) J. BWl. Chem. 193,265-275
Enzymol. 98, 241-260
101, Worth Publishers, Inc., New York
Academic Press, New York
R., and Dodgson, J. (1980) Cell 20,555-566
2891-2896
6104-6109
U. S. A. 80, 7501-7505
Biol. 64, 205-215
Related Documents











