i 1 E j lundesministerium .. 5 “‘ Cä Q / ä *1L “ ;d V : ; :: ss ; ' :: 2“” ~Er "a“ '“ "9 Beiträ ge zur Zuchtungsf orschung . . 1 Bundesanstalt fi 1r Zü cht ungsf orschung an 1 Kult ur pflanzen Proceedings 8" ‘ Symposium 7 l New Aspects of Resistance Research on Cultivated Plants Bacterial Diseases 15-16, 2001, Aschersleben, Germany Ahrensburg Aschersleben Braunschweig Dresden-Pillnitz Groß Lü sewitz Quedl inburg Siebeldingen 8. Jahrgang, Hef t 3, 2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

f
i
1
Ejlundesministerium..
5“ ‘ CäQ/ä*1L“;dV:;::ss;'::2“”~Er"a“'“"9 Beiträ ge zur
Zuchtungsforschung. .
1Bundesanstalt fi1r Zü chtungsforschung an 1
Kulturpflanzen
Proceedings
8"‘ Symposium 7
l
New Aspects ofResistance Researchon Cultivated PlantsBacterial Diseases
15-16, 2001, Aschersleben, Germany
Ahrensburg Aschersleben Braunschweig Dresden-Pillnitz
Groß Lü sewitz Quedlinburg Siebeldingen
8. Jahrgang, Heft 3, 2002

1
Beiträge zur Zü chtungsforschung
Bundesanstaltfü r Zü chtungsforschung an Kulturpflanzen lDie Zeitschrift „ Beiträ ge zur Zü chtungsforschung — Bundesanstalt fü r Zü chtungsforschung an
N
Kulturpflanzen“ verö ffentlicht Originalbeiträ ge und Ü bersichtsreferate sowie Tagungsberichte _t
aus dem Gesamtbereieh der Zü chtungsforschung.
Mit der Annahme des Manuskriptes und seiner Verö ffentlichung geht das ausschließ liche -
Verlagsrecht an die Zeitschrift „ Beiträ ge zur Zü chtungsforschung — Bundesanstalt fü r
Zü chtungsforschung an Kulturpflanzen“ ü ber.
Die Erscheinungsfolge ist unregelmäß ig, 2—3 Hefte im Jahr sind vorgesehen.
Die Auflagenhöhe ist z. Z. auf300 Exemplare festgelegt.Die Schriftenreihe erscheint in eigener Redaktion im Selbstverlag der BAZ.
'
Die Finanzierung wird durch das BMVEL realisiert.
Impressum
Beiträ ge zur Zü chtungsforschungBundesanstalt fü r Zü chtungsforschung an Kulturpflanzen
Herausgeber: Bundesanstalt fü r Zü chtungsforschung an KulturpflanzenRedaktionsbeirat: Dir. u. Prof. Prof. Dr. W. Flamme, Groß Lü sewitz
Dir. u. Prof. Prof. Dr. J. Grunewaldt, AhrensburgDir. u. Prof. Dr. M. Neumann, Quedlinburg
Schriftleitung: WissDir Dr. K. Peter
Redaktionsleitung und Vertrieb: Bundesanstalt fü r Zü chtungsforschung an KulturpflanzenNeuer Weg 22/2306484 QuedlinburgTel. O 39 46 / 47-244Fax 0 39 46 / 47-202
E-Mail: bafz—[email protected]: www.bafz.de
Diese Zeitschrift wird referiert in: Chemical Abstracts (Columbus)l
AGRIS — FAO_
(International Information System for the Agricultural .
Sciences and Technology)l
Nachdruck, auch auszugsweise, mit Quellenangabe zulässig.ISSN 0948-5538
Druck aufchlnrfrei gebleichtem Papier

Contents
Sé.ssiini?17= f.IiMétli6dsSusanne Jock, Won-Sik Kim, Klaus Geider
Molecular comparison ofErwinias causing fire blight and Asian pear blight I
Susanne Jock, Maja Hildebrand, Won-Sik Kim, Jochen Bogs, Klaus Geider
Markers to distinguish Erwinia amylovara strains and to follow bacterial invasion in plant tissue 4
Klaus Geider, Won-Sik Kim, Heike Salm, Martin SchollmeyerGenetics and biochemistry of viral lysozyme and EPS-depolymerase for control offire blight in plants 7
Athanassios Mavridis, Volker Paul, Klaus RudolphBacterial blight of Camelina sativa caused by Pseudomonas syringae pv. Camelinae„ ..„ .„ .....‚ ......„ .............. l0
Abdel-Rehim, K., Rudolph, K.
Sensitivity to antibiotics and heavy metals and plasmid profiles for characterization of differentraces ofXanthomom1.r campestris pv. malvacearum from different origins 13
B. Venkatesh, M. I. Khan, A. Pant, K. RudolphEnzymatic digestion ofplant pectins from susceptible and resistant cultivars of celery andextraction of a putative carbohydrate binding protein from phytopathogenic pseudomonads 17
Y. Nikolaichik, V. Myamin, I. Limorova, Y. Ignatenko, A. Pesnyakevich, A. Evtushenkov
Detection and characterization ofErwinia Caratovora subsp. Atraseptica genes participatingin the interaction with potato host
Ewa Zimnoch-Guzowska
Research on resistance to Erwinia caratovora ssp. atroseptica identified in diploid potato hybrids................. 24
Klaus Olbriclit, Erika Griesbach
First results ofresistance evaluation ofPeIargum'um to Xanthomonas hortorum pv. pelargom'i...................... 26
M. Hevesi, E. Jambor-Benczur, J. Papp‚ J. Dobranszky, K. Magyar-Tabori, T. Buban
Application of in vitro methods in fire blight resistance programs
V. Zinsou, B. Ahohuendo, Veronique Jorge, Valerie Verdier, Kerstin WydraEvaluation of cassava genotypes, including individuals of the genome mapping population,for resistance against bacterial blight
Kerstin WydraThe concept of resistance, tolerance and latency in bacterial diseases: examples from cassava
and 36
sessiöhlä zsa
Antje Burse, Matthias S. Ullrich
Ecological role ofa thermoregulated multi-drug efflux protein ofErwinia amylovora..‚ .„ .‚ .‚ ..„ ‚ .‚ „ „ „ „ „ ‚ ......... 44
Peter Laux, Wolfgang Zeller
Studies on the biological control offire blight in Egypt

Wolfgang Zeller, Peter Laux
Latest results on fire blight control with natural
V. Sotirova, E. Griesbach
Induction of resistance to Clavibacter michiganensis subsp. michiganensisby pre-inoculation oftomato young plants under field conditions in Bulgaria 52
Gudrun Barchend, Anita Schmidt
Investigations on epidemiological importance of weeds as a source for Ralstonia solanacearurri.................... 55
Harm Lö ptien, Erika Griesbaeh
Breeding of F, hybrids in cabbage with resistance to Xanthomonas campestris pv. campestris 57
C. WegenerTransgenic resistance against Erwinia sofl rot — A four-year field experiment (1997-2000) 61
K. Richter, C. Fischer
Stability offire blight resistance in 64
A. Banito, K.E. Kpémoua, KerstinwydraEcozonal variation in reaction of cassava genotypes from Togo to bacterial blight 67
K. Kass, M.G. Toth, M. Gö ndö r, M. Hevesi
Evaluation oftire blight resistance ofapple cultivars 71
M. Hevesi, M. Gö ndö r, M. G. Tö th, K. Kasa, K. HontySusceptibility of pear cultivars commercially grown in Hungary to the bacterial pathogenErwinia 74
Irmhild Schrö der, Athanassios Mavridis, Klaus.RudolphThe role ofmembrane-vesicle in pathogenesis 77
List of participants

1
Session 1: Methods for detection and identification
Molecular comparison of Erwinias causing fire blight and Asian pear blightSusanne Jock, Won-Sik Kim, Klaus Geider
Max-Planck-Institut fur Zellbiologie, Rosenhof, 68526 Ladenburg, Germany
Keygyordsz Asian pear pathogens, PCR, PFGE analysis, sequence alignments, HR
Abstract
Fire blight affects fruit trees like apple and pear and some ornamentals plants like hawthorn and cotoneaster. Incontrast, Asian pear blight mainly affects pear fruit trees. The disease, caused by Erwinia pyrzfalfae, was firstdocumented 1995 for Korea, and strains resembling E. pyrifoliae were isolated from necrotic pear trees in Japan.Methods to distinguish between these three groups of bacteria were developed. PCR and additional assays suchas sequence analysis of the /1rpN genes showed a close relation of Erwinias from Japan and E. pyrzfoliae, but a
distance to E. amylovortz. Strains were also differentiated by PFGE analysis after the whole genome was
digested with rare cutting enzymes. E. pyrifoliue strains showed three different patterns, and Erwinia strainsfrom Japan were even more diverse. With an Xbal digest E. amylovora strains from Central Europe and theMediterranean region were grouped into mainly four patterns. The PFGE patterns of European andMediterranean isolates can be used to follow the spreading of E. amylovora, they are highly conserved and havebeen constant for many years. The Asian pear pathogens are more divergent, and from a population of diseasedplant tissue, E. pyrifoliae strains were isolated with a mutation in /irpL.Introduction
The necrotic disease fire blight was first described for North America more than 200 years ago by Denning(187). Apart from North America, fire blight was first identified in the early 19th century to occur in Japan. In1919 fire blight has been reported for New Zealand, where the disease still persists. The first occurence inEurope was in England (1956), then in North Africa (Egypt, l962).The spreading to Central and Western Europeand to Eastern Europe happened probably outgoing from these countries. Some of the isolates from thosecountries were previously analyzed by pulsed-field gel electrophoresis (PFGE) (194, 195).In 1997, E. amylovnra was isolated from plant material of the Melbourne Botanic Gardens (189). Aftereradication of all fire blight host plants, the pathogen was not detected anymore. In Korea diseased pear treeswith the same symptoms like fire blight were observed (193). The pear disease has been mostly caused by thenovel pathogen Erwinia pyrifo/iae (190). Another bacterial species, Enterobacterpyrinus, which was describedseveral years ago (188), isolated from leaf spots of Nashi pear trees, may be not a true pear pathogen, because itdoes neither cause a hypersensitive reaction on tobacco nor symptoms on immature pears (S, Jock and K,Geider, unpublished), The recently isolated Erwinia strains fiom Japan (184) show in many regards a highhomology E. pyrifoliae, which is described as a novel species (190).Results and Discussion
PFGE arm/yrir afE. nmylovom strainsfivm Europe am] the Medflerrtzneun region
With an Xbal digest the investigated strains can be grouped into four major pattern types (Ft). The patterns differin a shift or a lack of one band. They are highly conserved for all strains isolated in a large area. The genome ofE. amylovora seems to be rarely subjected to changes, therefore strains can be classified according to their regionof isolation. With the aid of the different patterns we can follow the spread of fire blight in Europe and theMediterranean region. The first occurrence of tire blight in Europe was reported in England (1956-57). There wefound mainly two types of patterns: Ptl and Pt4, Ptl spread later to Central Europe and in the eastern part ofFrance, Pt4 to the western part of France. In the early sixties fire blight affected Egypt, where we find patterntype 2, This pattern was recovered in strains from Israel, Turkey, the Balkans and Hungary, but recently alsofrom the Iran (strains obtained from M. Mohamrnadi, Tehran, Iran). A gradual spread of fire blight to thesecountries can be assumed. The fourth pattern Pt3 was found in strains from Belgium and northern France.
If fire blight is established in a country, we normally find one typical pattern type, although we find some
exceptions - PIS was found for a few E, am_y/ovora strains isolated in Bulgaria and Israel, where strains with Pt2are dornirrating. In England, France, Belgium and the Netherlands several main PFGE pattern types can occurside by side. The novel pattern Pt6 was detected in Italy in strains isolated in 1991 and 1992 in the district ofRavenna, where Pt3 was found for strains isolated later in the same area, This was the only detection of Pt6,

2
indicating that strains with Pt6 did not further expand and might be even extinguished. Pt5 and Pt6 are thereforeminor pattern types.
In Spain and Italy, increasing spread of tire blight was observed during the last years. In Spain, most E.
amylovara strains have Pt4. This pattern can be explained by dissemination of the disease from France, where
we detected Pt4 in the western part. The pattern Pt3 was identified in two nurseries in Spain. We don't find Pt3 in
adjacent regions so probably the import of plant material,which was latently infected with E. amylavora, must beconsidered as the source of these outbreaks.
In Italy, P12 is the typical pattern found in southeastern areas (Apulia region), which may have spreaded from
regions of the Mediterranean basin by means of biotic and/or abiotic vectors (e.g., birds, air currents). For E.
amylavora strains from northern Italy Pt3 is dominating. As in Spain, this pattern type does not exist in adjacentregions with fire blight. Trade with infected plant material may have caused the establishment of Pt3 in
northeastern Italy (Po River Valley). Very recently (Summer 1999), punctifonn foci of fire blight occurred in
Southern Tyrol (close to the Austrian border) and we found Ptl strains besides Pt3 within the same area. Ptl maybe explained by introduction of fire blight from the North.
PFGE analysis ofthe Asian pearpathogens
The PFGE patterns of the Asian pathogens with restriction enzyme Xbal were not overlapping with the EuropeanE. amylovora strains. The patterns of 10 E. pyrzfoliae strains were related to each other. For the Erwinia strainsfrom Japan, a close relationship was found for three isolates and the XbaI pattems diverged in the case of threeother strains, The patterns from the Erwinia strains from Japan did also not fit into the observed PFGE types ofE. pyrzfoliae. It can be assumed that the Erwinia strains from Japan and E. pyrzfoliae are related but diverged in
evolution. Genomic DNA of E. pyrifoliae strains was also digested with restriction enzyme Spel, and
subsequently analyzed by PFGE. The fragment patterns were different from E. amylovora. For E. amylovora we
only have two pattems with an Spel digest. Strains with a Xbal pattern 3 have a different Spel pattern. In the Speldigests of E. pyrifoliae, three different patterns, PtA, PtB, and PtC, were observed, showing heterogeneity withinthe 10 E. pyrifoliue strains assayed.
Molecular detection ofthe Asian pear pathogens
E. amylovora and the Asian pear pathogens can be identified in plate assays (186, 193), Molecular detection ofE. amylovora is usually done by PCR analyses with primers derived from the common plasmid pEA29 or fromthe EPS encoding chromosomal region (185). When primers from the cps region of E. pyrifoliae (192) were
applied to the Erwinia strains from Japan, in all cases a band of the same size as for E. pyrzfoliae was obtained
(191). The signal should be specific for both pathogens, because neither E. amylovora nor other plant—pathogenicbacteria produced the PCR band with this primer pair. When PCR primers from the rRNA genes of E. pyrifoliaewere applied, four out of six Erwinia strains from Japan produced a signal of 0.7 kb, so some divergency occurs
in the intergenic transcribed spacer (ITS) region of these isolates. E. amylovora was negative with the rRNAprimers deduced from E. pyrifoliae.
Variabilty ofhrp genes ofthe Asian pear pathogens and ofE. amylovora
The ability to induce a HR reaction on non host plants is common to all plant pathogens. The responsible genesare ordered in the hrp cluster. E. amylovora is secreting two harpins, encoded by hrpN and hrp W. In order to
compare sequences which could allow conclusions about relationsships of bacteria we cloned and sequenced the
hip N genes of E. pyrifoliae and the Erwinia strains of Japan. We have analyzed the nucleotide sequences of
hrpN genes of six Erwinia strains from Japan, of E. pyrifoliae strain Epl/96 and E. amylavora strain Eal/79.
Alignment of the proteins and the encoding nucleotide sequences revealed conserved and variable regions. Bothends display a high degree of homology, whereas the centers are divergent. Among the six Erwinia strains from
Japan, this divergence continued and from the nucleotide sequences, all strains had characteristic patterns. Onthe other hand, single or clustered changes in the conserved parts were matching more closely for the Erwiniastrains from Japan and E. pyrifoliae, but not with E. amylovora.
Another instability was found for the hrpL gene. From a population in necrotic pear tissue, 13 (out of 24 isolates)E. pyrifaliae strains were detected, which did not produce a hypersensitive reaction on tobacco.
Complementation with genes from the E. amylovora hrp Cluster (together with M.-A. Barny, INRA, Paris)located the defect in hrpL. Cloning and sequencing of the gene from wild type strains and hrp mutants revealed a
change of a single base pair. The HR-deficiency could be advantageous at some of stages of the bacterial lifecycle.
Virulence assays onfire blight hostplrmts with E. pyrlfoliae
Nashi pears and European pears produced disease symptoms after inoculation with several E. pyrrfo/iae strains
(192). On apple seedlings symptoms were only occasionally induced by the pear pathogen from Korea. No
significant virulence symptoms were observed on cotoneaster, hawthorn, raspberry or plum. The bacteria,

3
consistently obtained from diseased Asian pear trees in Korea, have apparently a more narrow host range than E.amylovora. Virulence assays with the Erwinia strains from Japan have not been perfonned to a similar extent as
for E. pyrtfoliae in our labs. A preference of the host range to pear has been reported by others (184).
Conclusions
A novel pathogenic species was isolated from necrotic Nashi pear fruit trees in Korea and produced similar
symptoms as E. amylovora on pear seedlings. It differed from E. amylovora in several microbiological andmolecular criteria and has been classified as the new species Erwinia pyrzfaliue within the genus Elwinia (190).Erwinia strains isolated from necrotic Nashi pear fruit trees in Japan are not identical with pear pathogens from
Korea, but are more related to E. pyrifalfae than to E. amylovora. An important feature is the narrow host rangeof the two Asian pear pathogens compared to the fire blight pathogen E. amylovora. A striking feature of fire
blight outside North America is the close relationship of the PFGE patterns of the isolated strains. By comparingthem with the hetcrogenous patterns of E. (zmylovora strains from Canada, it can be assumed that the pathogendid not repeatedly spread from North America to other countries. The divergent PFGE patterns of isolates of the
two Asian pear pathogens may indicate their long persistence in Korea and Japan.
References
1. BEER, S.V.; KIM, J.-I-I.; ZUMOFF, C.H.; BOGDANOVE, A.J.; LABY, R.J.; GUSTAFSON, H.L.;MOMOL, T.; ALDWINCKLE, H.S.; TANII‚ A.; TAMURA, 0.: Characterization of bacteria that cause
"bacterial shoot blight ofpear" in Japan. Acta Hortic. 411, 1996, 179-1812. BERESWILL, S.; BUGERT, P.; BRUCHMULLER, I.; GEIDER, K.: Identification of Erwinia amylovora
by PCR with chromosomal DNA. Appl. Environ. Microbiol. 61, 1995, 2636-26423. BERESWILL, S.; JOCK, S.; BELLEMANN, P,; GEIDER, K.: Identification of Erwinia amylovora by
growth morphology on agar containing copper sulfate and by capsule staining with lectin. Plant Disease 82,1998, 158-164
4. BONN, W.G.; VAN DER ZWET, T.: Distribution and economic importance of tire blight. In: Vanneste, J.
(Ed.): Fire blight: The disease and its causative agent Erwinia amylavora, CABI Publishing. WallingfordOxon/UK.-New York, 2000, pp. 37-53
5, CHUNG, Y,R.; BRENNER, D.J.; STEIGERWALT, A,G.; KIM, H.T.; CHO K.Y.: Enterabacterpyrinus sp.nov. an organism associated with brown leaf spot disease ofpear tree. Int. J, Syst. Bac. 43, 1993, 157-161
6. JOCK, S,; RODONI, B.; GILLINGS, M.; KIM, W.-S,; COPES, C.; MERRIMAN, P‚ . AND GEIDER K.:
Screening of ornamental plants from the Botanic Gardens of Melbourne and Adelaide for the occurrence ofErwinia amylavora. Australas. Plant Pathol. 29, 2000, 120-128
7. KIM, W.-S.; GARDAN, L.; RHIM, S.-L.; GEIDER, K.: Erwinia pyrtfoliae sp. nov., a novel pathogenaffecting Asian pear trees (Pyrus pyrifolia Nakai). lnt, J. Syst. Bacteriol. 49, 1999, 899-906
8. KIM, W.-S.; HILDEBRAND, M.; JOCK, S.; GEIDER, K.: Molecular comparison of pathogenic bacteriafrom pear trees in Japan and the fire blight pathogen Erwinia amylovora. Microbiology/UK 147, 2001,2951-2959
9. KIM, W.-S.; JOCK, S.; PAULIN, J.-P.; RHIM, S.-L.; GEIDER, K.: Molecular detection and differentiationof Elwinia pyrifaliae and host range analysis of the Asian pear pathogen. Plant Disease 85, 2001, 1 183-1188
10. RHIM, S.-L.; VÖ LKSCH, B,; GARDAN, L.; PAULIN, J.-P.; LANGLOTZ, C.; KIM, W.—S.; GEIDER, K.:Erwinia pyrifoliae, an Erwinia Species different from Erwinia amylovorn, causes a necrotic disease of Asian
pear trees. Plant Pathology 48, 1999, 514-5201 1. ZHANG, Y,; GEIDER, K.: Differentiation of Erwinia amylovora strains by pulsed-field gel electrophoresis.
Appl. Environ. Microbiol. 63, 1997, 4421-442612. ZHANG, Y.; MERIGHI, M.; BAZZI, C.; GEIDER, K.: Genomic analysis by pulsed~field gel electro-
phoresis of Erwinia amylovora strains from the Mediterranean region including Italy. J. Plant Pathol. 80,1998, 225-232

4
Markers to distinguish Erwinia amylovora strains and to follow bacterialinvasion in plant tissue
Susanne Jock, Maja Hildebrand, Won-Sik Kim, Jochen Bogs, Klaus Geider
Max-Planck-Institut fiir Zellbiologie, Rosenhof, 68526 Ladenburg, Germany
Abstract
Several markers were applied to distinguish individual Erwinia amylovora strains during simultaneouscolonization of plants. E. amylovnra strains were labelled with the Green Fluorescent Protein (GFP) andvisualized in plant tissue by fluorescence microscopy. Inoculation of the intercostal region of apple leavescaused movement of the bacteria in the apoplast followed by invasion into the vascular system. After inoculationof apple leaf tips migration of the bacteria was observed through the xylem vessels and outbreaks into the
adjacent intercellular space of the parenchyma. Non-pathogenic mutants did not move from the inoculation site.Plant resistance enhancers caused a decrease in the migration rate. The migration rate in the central vein of appleleaves corresponded to disease ratings from symptom formation on shoots of various apple cultivars. Othermarkers for strain differentiation were the size of short sequence DNA repeats and streptomycin-resistance.Also, these marked strains were also compared in plant tissue colonization. Quantitative PCR was performedwith a light cycler and fluctuation was found for leaf colonization.
Introduction
Fire blight is caused by the Gram-negative bacterium Erwinia amylovora. It is an economically disastrousdisease for apple and pear production. The pathogen is spread by insects, birds, wind and human activities. Entrysites for the pathogen are the nectaries of flowers, stomata or wounded plant tissue.
Diagnosis of fire blight with molecular tools is often performed by PCR with primers derived from plasmid
pEA29, which is common for natural E. amylovora strains (3). They amplify a DNA fragment of about 1 kb,which is specific for E. amylovara. A product of a similar size obtained with pear pathogenic strains from Japanas template has been used as an argument that those isolates are identical with the fire blight pathogen E.amylavara (1), Sequence analysis of this fragment revealed a partial homology with the PCR fragment obtainedfrom E. amylovara, but its repeat sequence is "GGATTCTG" in contrast to "ATTACAGA" for plasmid pEA29
(unpublished). The repeat numbers in the DNA fragment of E. amylovora can vary between 3 to 15 (10). Size
Changes occasionally occur even under laboratory conditions, but they are rare and are often observed after stress
situations of the bacteria. On the other hand, the size is stable enough to differentiate strains from each other,which have been simultaneously inoculated into plants.
In many cases highly conserved genes encode for enzymes or proteins with specific functions in the cellmetabolism and cell structure. These could be used for differentiation of E. amylovora isolates. Nucleotide
sequences of gapDH gene have mainly been applied to distinguish species (5). OmpA is a membrane protein,which may be under pennanent pressure to become adjusted to the need of the cells after environmental changesand seems to be highly variable. Glycerolaldehyde phosphate dehydrogenase is an essential component of
glycolysis. Some changes in the protein structure may be tolerated without changing its catalytic properties.
Streptomycin (Sm) has been applied in many countries for control of fire blight. Resistant strains can arise bytransfer of a transposon, which encodes for a Sm-phosphotransferase or by selection for spontaneouschromosomal resistance (9), In most of these cases, a ribosomal protein, S12 of the small ribosomal subunit isaltered and prevents binding of the antibiotic to the ribosome. Those mutants have a high resistance to Sm andproduce more amylovoran than the corresponding wild type strains (1).
Only differences in the nucleotide sequence without an obvious impact on virulence of strains are thereforeneutral markers to distinguish strains, but these require laborious analyses. With antibiotic resistance markers,cells have to be selected on plates, SSR analysis needs high resolution of PCR fragments on gels, whereas theobservation of fluorescence is easily achieved with an epifluorescence microscope to detect cells and to evaluate
gene expression in intact plant tissue or in tissue extracts.
Numerous studies have been done to examine the colonization of host plants by Erwinia amylovoru. Electronmicroscopy was used to visualize the pathogen inside the plant tissue. Goodman et al. (11) confined bacterialmovement to the xylem after artificial inoculation involving damage of the main vein (13). However, when thebacteria were applied to succulent shoots of the susceptible rootstock M26, E. nmylovora was claimed to
predominantly colonize the cortical parenchyma (8).

5
The Green Fluorescent Protein (GFP) has received attention as useful molecular tool for analysis of cellular geneexpression (6). The GFP protein of the jellyfish Aer/uorea Victoria emits green fluorescence with a maximum at
509 nm when irradiatcd with UV-light (excitation at 488 nm). This chromophore requires no exogenoussubstrates or cofactors except oxygen for fluorescence and labeled cells can be studied without fixation of planttissue or bacteria in real time (7).
This study describes the use of GFP to determine the migration of the pathogen in plants correlated to symptomfonnation and dissection of inoculated mixtures of strains by SSR numbers or distinction of wild type strains andthose with spontaneous streptomycin resistance.
Results and Discussion
Migration ofE. uniylovoru in plant tlirxue
After inoculation of leaf tips with gfjrlabeled Eal/79 cells the bacteria migrated through the xylem at
approximately 4 mm per day against the direction of the transpiration flow in the vessels. Plugging of thevascular water system could partially explain wilting symptoms associated with fire blight. The bacteria alsocolonized the intercellular space of the parenchyma afler disrupting the xylem vessels (4). Movement ofE. amylovonz from the xylem into the apoplast was not restricted to the site of wounding as it was describedbefore (13). Migration in the xylem exceeded the speed of movement in the intercellular space and could be
important for fast colonization of plant tissue by E. amylovorzr. The xylem sap is not devoid of nutrients andcontains amino acids and organic acids (12).
The migration rate of the labeled pathogen inside the middle vein of leaves was measured by epifluorescence
microscopy. The movement rate differed among strains and plant cultivares. The screening was done with leavesof young seedlings. In parallel experiments, several apple cultivars were inoculated at the shoot and progressionof symptoms was evaluated after one or two weeks (with K, Richter, Aschersleben, Germany). The datacorrelated largely with the speed of migration determined via visualization by fluorescent gfp-cells.Ute ofSSRs_fi'am pl(1smiripEA29 (IS a markerfor strain Iflff€ l'€ I1ffllfi0l1Five E. mnylat/om strains from Germany with the PFGE pattern type Ptl and different SSR numbers (4, 5, 6, 7,and l0) were inoculated into shoot tips of various apple cultivars and a plum cultivar. After two weeks, the stem
section between the healthy and necrotic bark were removed and extracted for the residing bacteria. These were
directly assayed by PCR to identify the type of SSR with primers annealing adjacent to the repeats. The positionof the signal (10) and its strength were used to identify the individual strain and to estimate the amount of cellscompared to another strain with a different repeat number. The most virulent strain was Ea286‚ which was alsofound to be more aggressive than other strains, when labelled with a gfiz-plasmid.
Antibiotic resistance am‘ a strain marker
Three strains from Germany were selected for spontaneous streptomycin-resistance on nutrient agar plates withstreptomycin (Sm) (500 pg/ml). Pairs of the resistant mutant and its parent strain were grown over night and 1:1mixtures used for inoculation of apple shoots or leaves of apple seedlings. In case of shoots, bacteria from thetransition zone to healthy bark were extracted and the phenotype of resistance was detennined. Inoculated leaveswere extracted at the petiole after 11 days and a leaf adjacent to the inoculated leaf was also detached andextracted. The pair of Ea7/74 also showed a preference for the wild type strain in the petiole extract of theinoculated leaf, but in the next leaf the streptomycin-resistant mutant comprised two thirds of the population.For the two other strains, the Sm—variant dominated in both leaf types, in one case 99% of the extracted cells
were Sm-resistant.
When the strains were inoculated into slices of immature pears in a 1:1 ratio, no dominance was detected foreither type. This suggests similar growth properties for the wild type and Sm-resistant strains during the strongpropagation during ooze production in pear slices. In apple tissue, the Sm-resistant strains have apparently an
advantage for growth in comparison to the wild type strains. Control of fire blight with streptomycin couldtherefore not only result in a selection for resistant strains, but the resistant variants could be more adapted to
colonize apple tissue than the parental wild type strains.
Quantitative PCR applying (I light cylcer
The concentration of amplicons during PCR was measured with a light cycler (BioRad). Standard curves were
determined, and unkown amounts of E. mnylovoru were calculated. The colonization of leaves was
hetcrogenous, but a trend was seen for low performance of strain PD350 as already found in other assays.

6
Conclusions
The colonization of host plants by E. amylovorzr was studied after artificial inoculation with gfp-labeled bacteria.Inoculation by scissors resulted in fast pathogen migration through the vascular system followed by outbreaks tothe adjacent intercellular space of the parenchyma. The speed ofmigration of labeled E. amylavora in the centralvein of apple leaves was correlated with the virulence of different strains.
Other markers to distinguish E. amylovora strains after inoculation of plants were streptomycin—resistance andthe number of short sequence DNA repeats (SSRs). Both approaches allowed the estimation, of virulence instrain mixtures. Quantitative PCR with a light cycler showed uneven colonization of apple leaves by E.amylovora.
References
1. BEER, S.V.; KIM, J.-H.; ZUMOFF, C.H.; BOGDANOVE, A.J.; LABY, R.J.; GUSTAFSON, H.L,;MOMOL, T.; ALDWINCKLE, H.S.; TANII‚ A.; TAMURA‚ 0,: Characterization of bacteria that cause
"bacterial shoot blight of pear“ in Japan. Acta Hortic. 411, 1996, 179-1812. BELLEMANN P.; BERESWILL S.; BERGER S.; GEIDER K.: Visualization of capsule formation by
Erwinia amylovora and their biochemical characterization, Int. J,Biol. Macromol. 16, 1994, 290-296
3. BERESWILL, S. ; PAHL, A.; BELLEMANN, P.; ZELLER, W.; GEIDER, K.: Sensitive and species-specific detection ofErwim'a amylovora by PCR-analysis. Appl. Environ. Microbiol. 58, 1992, 3522-3526
4. BOGS, J.; BRUCHMÜ LLER, I.; ERBAR, C.; GEIDER, K.: Colonization of host plants by the fire blightpathogen Erwinia amylovora marked with genes for bioluminescence and fluorescence. Phytopathology 88,1998, 416-421
5, BROWN, E.W.; DAVIS, R. M,; GOUK, C.; VAN DER ZWET, T.: Phylogenetic relationships ofnecrogenic Erwinia and Brenneria species as revealed by g1yceraldehyde—3—phosphate dehydrogenase genesequences. Int. J. Syst. Evol. Microbiol. 50, 2000, 2057-2068
6. CHALFIE, M.; TU, Y.; EUSKIRCHEN, G.; WARD, W.W.; PRASHER, D.C.: Green fluorescent protein as
a marker for gene expression. Science 263, 1994, 802-8057. CUBITT, A.B.; HEIM, R.; ADAMS, S.R.; BOYD, A.E.; GROSS, L.A.; TSIEN, R.Y.: Understanding,
improving and using green fluorescent proteins. Trends Biochem. Sci. 20, 1995, 448-4558. EDEN—GREEN, S,J.: studies in fireblight disease of apple, pear and hawthorn (Erwinia amylovora (Burill)
Winslow et al.). PhD thesis, 1972, University of London9. JONES, A.L.; SCHNABEL, E.: The development of streptomycin-resistant strains of Erwinia amylovora.
In: Vanneste, J. (Ed.): Fire blight: The disease and its causative agent Erwinia amylovora, CABI Publishing.Wallingford Oxon/UK.~New York, 2000, pp. 235-251
10. KIM, W.-S,; GEIDER, K.: Analysis of variable short-sequence DNA repeats on the 29 kb plasmid ofErwinia amylavora strains. Eur. ‚I. Plant. Pathol. 108, 1999, 703-713
11. LEWIS, L.N.; GOODMAN, R.N.: Mode of penetration and movement of fire blight bacteria in apple leafand stem tissue. Phytopathology 55, 1965, 719-723
12. PURCELL, A.H.; HOPKINS, D,L.: Fastidious xylem-limited bacterial plant pathogens. Annu. Rev.Phytopathol. 34, 1996, 131-151
13. SUHAYDA‚ C.G.; GOODMAN, R,N.: Early proliferation and migration and subsequent xylem occlusionby Erwinia amylovara and the fate of its extracellular polysaccharide (EPS) in apple shoots. Phytopathology71, 198l, 697-707

7
Genetics and biochemistry of viral lysozyme and EPS-depolymerase forcontrol of fire blight in plants
Klaus Geider, Won-Sik Kim, Heike Salm, Martin SchollmeyerMax-Planck—Institut fiir Zellbiologie, Rosenhof, 68526 Ladenburg, Germany
Abstract
The fire blight pathogen Erwinia amylovora and the Asian pear pathogens synthesize similar capsular
exopolysaccharides (EPS), which are strictly required for pathogenicity. The molecular weight of the EPS is 1 to
5 MDa. The repeating units of EPS synthesized by Erwinia pyrzfoliae from Korea and the Erwinia strains from
Japan do not bear a glucose residue as a second side chain found in the repeating units of amylovoran.Amylovoran expression is controlled by regulatory genes, Alignments of rcsB genes, but also of the nucleotide
sequences upstream of the gene cluster for EPS synthesis show a significant relationship between the two pearpathogens and the evolutionary distance to E. amylovora. From a bacteriophage, a DNA fragment was
sequenced with genes encoding a lysozyme and an EPS depolymerase. The enzyme was used to degradeamylovoran capsules and shows to interfere with colonization of pear slices by E. amylovora. In cooperationwith other labs, the gene was transferred into tobacco, pear and apple and regenerated plants were assayed forexpression of dpo. Lytic effects were demonstrated for the lysozyme in extracts of induced cells, showing thatthe gene expression could also be useful for control of fire blight in affected host plants.
Introduction
Bacteria colonization of fire blight host plants depends on the synthesis of capsular exopolysaccharide (EPS)amylovoran, a virulence factor of Erwinia amylovora (1). Amylovoran consists of repeating units with fourgalactose and one glucuronic acid residues as well as glucose in most of the subunits (12). Its biosynthesisrequires the function of several sugar transferases and of proteins for transport and polymerization encoded bygenes of the ams cluster (2). The precursors for synthesis of the repeating units are presumably UDP-galactose,UDP-glucose and UDP-glucuronic acid. EPS-synthesis in a subcellular system has also been described forSinorhizobium meliloti (13). The molecular weight of amylovoran was determined by analyticalultracentrifugation and SEC/MALLS to be 1.1 x 10° (6). A DNA fragment from phage ¢ Ealh encoded a
lysozyme and an EPS degrading activity (7), which is used by the phage to access the receptors at the surface ofits host cells. Its expression in transgenic plants could expose the uncapsulated bacteria to host defensemechanisms.
Amylovoran synthesis is regulated via a network of proteins. As observed for Escherichia coli and otherbacteria, many regulatory proteins are activators in a two component system (4). A sensor reacts to
environmental signals and phosphorylates the main activator, which is further activated by interaction withanother activator protein. This mode can apply for the synthesis of many polysaccharides including LPS-synthesis (5).
In these studies, two Asian pear pathogens, which are different from E. amylovora, were included. Erwinia
pyrifoliae was isolated from necrotic pear trees in Korea (9) and ordered into a new species based on DNA/DNAhybridization kinetics and Biotype 100 assays (8). Erwinia pathogens from Japan were also isolated from Nashipear and are apparently related to E. pyrifoliae (8). The Asian pathogens also produce an amylovoran-like EPSand seem to be regulated by similar proteins as described for amylovoran synthesis.
Results and Discussion
Precursorsfor amylovoran synthesis
Amylovoran is synthesized by polymerization of repeating units with galactose, glucuronic acid and glucose.The last two genes in the ams-region are connected with synthesis of UDP-sugars. UDP-glucosepyrophosphorylase, encoded by gaIU, is required for the conversion of glucose-I—phosphate to UDP-glucose, a
precursor of UDP-galactose and UDP-glucuronic acid. AmsM is similar to GalF, which has been proposed as a
subunit of GalU. In E. amylovara, am,rM~mutants are deficient in amylovoran production, although their GalU-activity is enhanced. GalE is the epimerase for UDP-glucose/IJDP-galactose conversion (11) and the enzymeactivity is lacking in amsM mutants. This effect can be attributed to a polar effect of resistance cassettes intoamsM preventing transcription ofga/E. Alternatively, AmsM and GalE could interact as proteins.
For Synthesis of a repeating unit of amylovoran, a galactose residue is attached to a lipid carrier in the cellmembrane and then, the sugars galactose, galactose, glucuronic acid and galactose are sequentially connected(3). This unit is transported through the membrane and linked to existing amylovoran at the third galactose

8
molecule. Thereby, the side chain is formed and a glucose residue is attached to the third galactose. E. pyrzfoliaeand the Erwinia strains from Japan like many E. amylovora strains from raspberry lack this glucose residue. The
responsive gene is apparently not part of the ams-cluster, since transfer of genes amsH/ABCDEFKL into E,
pyrtfoliae does not lead to attachment to this glucose residue.
Molecular weight ofamylovoran
Amylovoran was isolated from cells grown on minimal agar. The molecular weight of the EPS preparations was
analyzed on large pore gel permeation columns. Amylovoran eluted as a sharp peak slightly behind levan, used
as a molecular weight marker of 5.5 MDa. The size of approximately 5 MDa is thus often but not alwaysconsiderably higher in preparations from cells grown on agar than for previous preparations derived from
supematants of suspension cultures (6). On the other hand, a more slowly eluting fraction was visible as a
shoulder at a molecular weight of 1-2 MDa. This material could be dominant in several preparations, where highmolecular weight material was almost lacking. Treatment of the amylovoran solution with EPS-depolymerasedegraded the fast eluting material. No major contarninations were thus included in the peak with fast migrationin the gel permeation columns.
Characterization ofEPSfrom E. pyrzfoliae and the Erwinia strainsfrom Japan
The structure of the repeating unit of EPS from the two Asian pear pathogens were elucidated by ESI/MS andalso by NMR spectra in comparison with the repeating units of amylovoran (in cooperation with M. Nimtz andV. Wray, GBF, Braunschweig, Germany). Like for amylovoran, EPS of the Asian pear pathogens consists of a
backbone with three galactose residues and a side chain with glucuronic acid and a terminal galactose, which isdecorated with pynivyl and acetyl groups. The glucose residue at the branched galactose is absent in EPS of bothAsian pear pathogens. It is possible that the lacking glucose residue is one reason among others for their limitedhost range compared to E. amylovora.
Regulation ofEPS synthesis
Amylovoran synthesis is regulated by environmental conditions such as pH, but also by several gene productssuch as the activators RcsA and RcsB and the sensor RcsC. RcsB is conserved for the three pathogens. In
alignments of the proteins and the nucleotide sequences the pear pathogens from Korea and Japan were more
related to each other than to E, amylovora.
In the alignment of the nucleotide sequences upstream of the EPS encoding gene clusters, variable and conservedareas were identified. The conserved sequences could include binding sites for the RcsB/RcsA complex and ofthe RNA polymerase.Characterization ofa viral DNAfragment with the EPS depolymerase gene
From bacteriophage tj)Ea1h, a gene cluster encoding for an EPS depolymerase, a holin and a lysozyme was
characterized (7). The lysozyme gene was cloned on a plasmid under control of the Iac-promoter. Lysozyme was
expressed in E, coli cultures selected with kanamycin. Cell lysates interfered with growth of E. amylovara, butalso with growth ofE. pyrifoliae. The enzyme did not cause growth inhibition of Km-resistant cells.
The fragment with the largest ORF of 1974 bp was also cloned and expressed the EPS cleaving activity incontrast to an EcoRI fragment with part of the ORF. The polypeptide, deduced from its nucleotide sequence,contains 657 amino acids corresponding to a molecular weight of 71 KDa. In an amino acid sequence homologysearch, the gene product of amsF from the amylovoran synthesis encoding region of E. amylovora had thehighest homology (40%). The protein has been postulated to be involved in polymerization of the repeating unitsof amylovoran. The EPS-depolymerase was His-tagged and the purified enzyme used to degrade amylovoranand the EPS of E. pyrzfoliae. No difference was observed for the degradation kinetics. When cell lysates withEPS depolymerase or with lysozyme were applied to slices of immature pears, both inhibited growth of E.amylovora and prevented ooze formation.
The dpo gene was cloned into a binary vector and leaf disks of tobacco (S. Stile, Budapest, Hungary), pear (E.Chevreau, Angers, France), and apple (V. Hanke, Dresden-Pillnitz, Germany; E. Kiss, Gö dö llö , Hungary) were
transformed and plants regenerated. The presence of the gene in transgenic plants was confirmed by Southernblots, and PCR and its expression by Western blots. Preliminary resistance screening showed retardedcolonization of gfp-labelled E. amylovora strains in transgenic apples compared to normal apple leaves (V.Hanke).
Conclusions
The capsular EPS amylovoran of E. amylovora is synthesized as repeating units, which are transported throughthe cell membranes and polymerized into high molecular weight EPS of 1 to 5 MDa. Amylovoran synthesis isregulated by activator proteins, but also by environmental conditions. EPS synthesis is thus responding to manyevents in the bacterial life cycle. Genes encoding an EPS-dependent depolymerase and a viral lysozyme were

9
fused into a His-tag expression vector, and the encoded enzymes were characterized for their biochemicalproperties. Expression in plant cells can expose E. tzmylovora without capsules to plant defense mechanisms, or
the lysozyme could destroy the pathogen when colonizing transgenic plants.
References
1. BELLEMANN, P.; GEIDER, K,: Localization of transposon insertions in pathogenicity mutants of Erwinianmylovora and their biochemical characterization. J. Gen. Microbiol. 138, 1992, 931-940
2. BUGERT, P.; GEIDER, K.: Moleeular analysis of the ams operon required for exopolysaccharide synthesisofErwinia (Imy/ovora. Mol. Microbiol. 15, 1995, 917-933
3. GEIDER, K.: Exopolysaccharides of Erwinia amylovora: Structure, biosynthesis, regulation, role in
pathogenicity of amylovoran and levan. In: Vanneste, J. (Ed.): Fire blight: The disease and its causative
agent Erwinia amylavora, CABI Publishing. Wallingford Oxon/UK.-New York, 2000, pp, 117-1404. GOTTESMAN, S.: Regulation of capsule synthesis: Modification of the two-component paradigm by an
accessory unstable regulator. ln: Hoch, J.A.; Si1havy,T, (Eds.): Two eomponent signal transduction,American Society of Microbiology, Washington, DC, 1995, pp. 253-262
5. JAYARATNE, P.; KEENLYSIDE, W.; MACLACHLAN, P.; DODGSON, C.; WHITFIELD, C.:Characterization of rcsB and rcsC from Escherichia coli O9:K30:H12 and examination of the role of the re:
regulatory system in expression of group I capsular polysaccharides. J.Bacteriol. 175, 1993, 5384-5394
6. JUMEL, K.; GEIDER, K.; HARDING, S,E.: The solution molecular weight and shape of the bacterialexopolysaccharides amylovoran and stewartan. Int. J
.Biol. Macromol, 29, 1997, 251-258
7. KIM, W.-S.; GEIDER K.: Characterization of a viral EPS-depolymerase, a potential tool for control of fire
blight. Phytopathology 90, 2000, 1263-12688. KIM, W.-S.; GARDAN, L.; RHIM, S,-L.; GEIDER, K.: Erwinia pyrtfoliae sp. nov., a novel pathogen that
affects asian pear trees (Pyrus pyrxfo/in Nakai). Int. J. Syst. Bact. 49, 1999, 899-9069. KIM, W.-S.; HILDEBRAND, M.; JOCK, S.; GEIDER, K.: Molecular comparison of pathogenic bacteria
from pear trees in Japan and the fire blight pathogen Erwinia tzmylovora. Microbiology/UK 147, 2001,2951-2959
10. KIM, W.-S.; JOCK, S.; PAULIN‚ J.-P.; RHIM‚ S.-L.; GEIDER, K.: Moleeular detection and differentiationofErwinia pyrifo/iae and host range analysis of the Asian pear pathogen. Plant Disease 85, 2001, 1183-1188
11. METZGER, M.; BELLEMANN, P.; BUGERT, P.; GEIDER, K.: Genetics of galactose metabolism ofErwinia amylovoru and its influence on polysaccharide-synthesis and virulence of the fire blight pathogen. J
.
Bacteriol. 176, 1994,4150-45912. NIMTZ, M.; MORT, A.; DOMKE, T.; WRAY, V.; ZHANG, Y.; QIU, F,; COPLIN, D.; GEIDER, K.:
Structure of amylovoran, the capsular exopolysaccharide from the fire blight pathogen Erwinia amylavora.Carbohyd. Res. 287, 1996, 59-76
13. REUBER, T,L.; WALKER, G.: Biosynthesis of succinoglycan, a symbiotically importantexopolysaccharide ofRhizobium meliloti. Cell 74, 1993, 269-280
14. RHIM, S.-L,; VÖ LKSCH, B.; GARDAN, L.; PAULIN, J.-P.; LANGLOTZ‚ C.; KIM, W.-S,; GEIDER, K.:Erwinia pyrifoliae, an Erwinia species, different from Erwinia amylovora, causes a necrotic disease of Asian
pear trees. Plant Pathology 48, 1999, 514-520

10
Bacteria] blight of Camelina sativa caused by Pseudomonas syringae pv.cam elinae
Athanassios Mavridis], Volker Paulz, Klaus Rudolph‘'Institute ofPlant Pathology and Plant Protection, University of Gö ttingen, Grisebachstr. 6, 37077 Göttingen;
zUniversity ofPaderbom, Division Soest, Lü becker Ring 2, 59494 Soest
Introduction
Wild flax (Camelina Sativa) belongs to the family of Brassicaceae and is a common weed in nearly everyagricultural field, in particular in fields with flax (Linum usitatissimum), In some countries C. sativzz is cultivatedfor seed production containing 30-35% of an oil which can be used for several technical purposes.
In 1996, 1997 and 1998 in the frame of a project supported by the European Union, several attempts were
undertaken to increase the quantity and to improve the quality of the seed oil of C. sativa by breeding measures.The aim was to exploit C. sativa as a crop providing regenerable raw material. During these years an unknowndisease occurred in most of the experimental plots of C. sativa. Disease symptoms appeared on all aerial plantparts and obviously caused yield losses. Preliminary investigations showed that the disease was notcaused by a
fungus. First microscopic tests revealed large masses of bacteria to be extruded from all affected plant tissues. Apreliminary report on the pathogen was published in 1998 (Mavridis et al. 1998), The objective of this study was
to characterize as well as to identify the causal agent of this new disease.
Material and Methods
Isolation ofbacteriaNumerous samples of diseased plants of C. sativa from different localities in Germany were collected over thethree years. Isolations were carried out from spots on leaves, stems and silicles, but also from infected seeds.Cultures of bacteria were obtained by dilution streaking or dilution plating of plant homogenates on Petri disheswith King‘ s medium B (KB). From each isolation representative colonies of the dominant type were selected andtransferred onto slants with yeast-dextrose-carbonate (YDC) agar and maintained at 7 ° C. Prior to furtherinvestigations isolated strains were regrown for 24 h on KB plates at 28 ° C.
PathogenicinrtestPlants of C. sativa were inoculated with 7 representative strains either by pressure-spraying the bacterialsuspension (106 cfu/ml) into the intercellular spaces through the abaxial side of young leaves until tiny water-soaked spots became visible using a glass atomizer and/or by spraying the inoculum (103 cfu/mls) superficiallyonto young and green stems and silicles. For hypersensitivity test bacterial suspension of 10 cfu/ml were
injected in the intercostal fields of tobacco leaves (cv. White Burley). A visible collapse of the infiltrated tissuewithin 24 h was recorded as a positive reaction.
Bacteriological characteristics
Several tests based on the nutritional and physiological properties of bacteria were used for characterizing theisolates from Camelina (Lelliott and Stead, 1987; Klement et al., 1990). These tests included the Gram-reaction,production of fluorescent pigments, and the so-called LOPAT-criteria (levan production, oxidase activity,pectinase activity, arginine dihydrolase and the ability to induce hypersensitivity on tobacco leaves).Additionally, 6 strains were tested with the GN MicroPlate System from Biolog, Inc. (USA) based on thecapability ofthe test organism to utilize 95 carbon sources (Bochner, 1989; Jones et al., 1993).Toxin production
For toxin production 12 bacterial isolates from Camelina were incubated for 48 h in test tubes containing 7 ml ofpotato-dextrose-liquid medium at 28 ° C in a rotary incubator. After pelleting of bacteria by centrifugation thesupernatant was used. Plastic Petri dishes containing 15 ml potato-dextrose—agar were first inoculated with 150pl of a suspension of Geotrichum candidum (ca. 108 conidia/rnl) or Rhodotorula sp. (ca. 10K cells/ml). Prior touse five peripheral wells equidistant from a central well with 6 mm in diameter were cut with a template. Afterremoving of the agar plugs by a suction pipette 30 pl of each supernatant were added per well, and the plateswere incubated at 28 ° C for 3-4 days.Determination ofhost range
Numerous plants of different families and species were inoculated by spraying in order to determine the hostrange of the isolated bacterium. Besides C. sativa five wild Camelina species (C, Iaxa, C. microcarpn, C.

11
rumelica, C. rume/ica, ssp. trzznscaspica, C, hispida var. grandiflora) as well as several other Brassicaceae, such
as Brassica napus var. napux, B. rapa var. rapa, B. juncea var. integrffolia, Capsella bursa-pastoris and Thlaspimvense were included. Additional plant species tested were: Glycine max, Lycopersicon esculentum, Phaseolus
vulgaris, Passwort: edulis, Citrus limon, Citrus paradisi, Syringa vulgaris, and Hibiscus rosa—sinensis.
Results and Discussion
Symptoms
Disease symptoms occurred on all aerial parts of naturally infected C, sativa plants, and cortsisted of small
transparent spots on leaves, which were first watervsoaked and turned brown later on. On the stems elongatebrown spots and sometimes cracked lesions appeared, while on the silicles conspicuous circular brown to dark
brown spots of 1-3 mm diameter were caused. Seeds of severely infected silicles remained small, shrivelled and
showed a brownish discolouration.
Isolation and identification ofthe pathogen
From infected tissue, including seeds, numerous bacterial isolates with rod—shaped, motile cells were obtained.
The predominant bacterial colonies on King‘ s medium B were whitish-grey, circular, raised with an entire edgeand showing blue-green fluorescence. Glasshouse inoculations on C. sativa plants in young and flowering stagewith several bacterial strains of the predominant colony form resulted in symptoms identical to the above
described ones after natural infection. All isolates tested were Gram-negative, strictly aerobic, induced a
hypersensitive reaction on tobacco leaves, produced levan but were negative for oxidase and arginine
dihydrolase and did not rot potato slices.
According to our tests, the bacterial pathogen fulfilled the LOPAT-criteria for the fluorescent Pseudomonas
group Ia and is closely related to Pseudomonas syringae (Lelliott et al., 1966).. Bacterial strains with identical
properties were also isolated from Canadian samples of C. Sativa sent to us by Dr. J. P. Tewari (University of
Alberta, Edmonton, Canada). Tests for toxin production did not result in growth inhibition of G. candidum or
Rhodotorula sp..
Evaluation of results obtained by the database Biolog GN version 3.00 revealed a similarity grade of 50 - 80% to
Pseudomonas syringae pv. pisi (Psp) for five from six strains tested, whereas the sixth strain reacted like
Pseudomonas syringae pv. tomato (Pst; ca. 50% similarity). A comparison of the metabolic patterns of Psp, Pst
and Pseudomonas syringae pv. syringae to those of the Camelina strains revealed at least more than five
differences regularly occurring. Furthermore, reproducible differences between Psp, Pst and Camelina strains
were confirmed by greenhouse inoculations of the bacterial strains in tomato, pea and wild flax plants. Disease
symptoms were induced by each bacterium only on its host plant.
Further investigations on the host range supported the conclusion that this bacterial pathogen is quite distinct
from the hitherto described members of the Pseudomonas syringae group. Besides C, Sativa only the wild
Species C. Inxa, C. microcarpa, C rumelica and C. rumelica ssp. transcaspica reacted susceptible towards the
pathogen (Table 1). All other plants tested developed no symptoms or only few tiny dark necrotic spots.
As far as we know, a bacterial pathogen of the Pseudomonas syringae group has never been described for C.
saliva (Bradbury, 1986). Because our studies revealed a very narrow host range for a few Camelina species the
pathogen appears to represent a yet unnamed pathovar of Pseudomonas syringae. Therefore, the name
Pseudomonas syringae pv. camelinae is proposed for this pathogen indicating that the genus Camelina contains
the main plant species diseased by this pathogen.
12
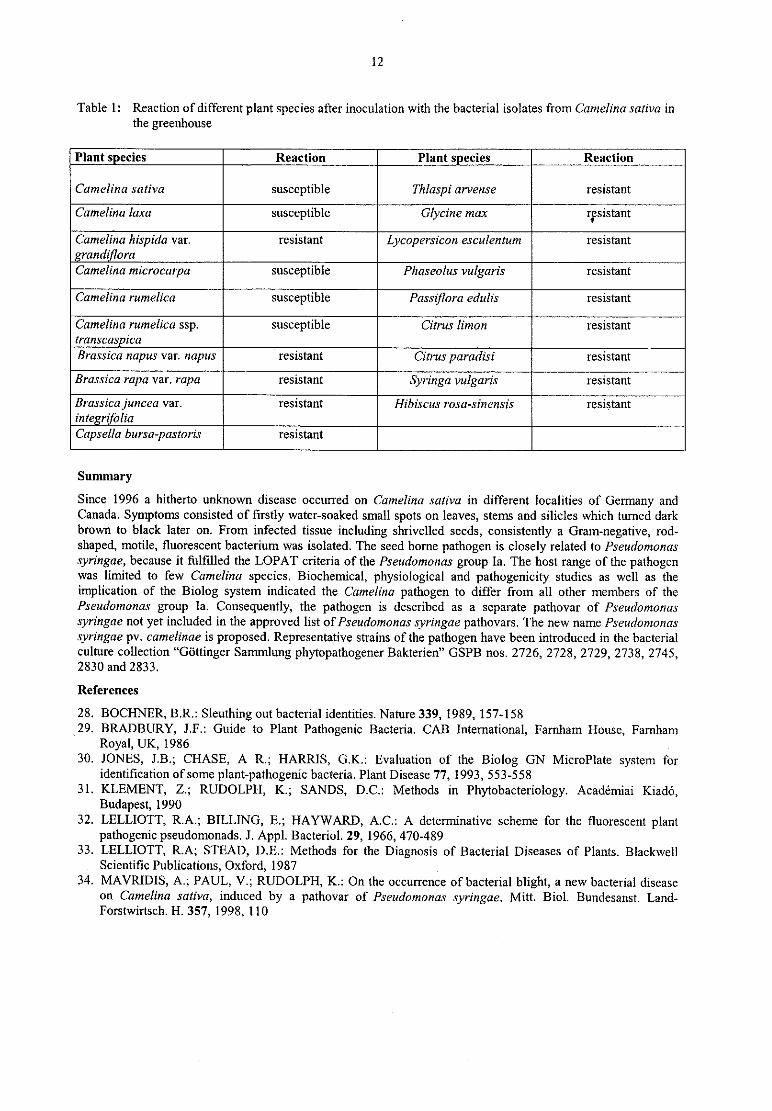
Table 1: Reaction of different plant species after inoculation with the bacterial isolates from Camelina saliva inthe greenhouse
Thlaspi arvense
Camelina hispida var. resistant Lycopersicon esculentum resistantrandi ora
S‘ “ ° ° "‘ “ "° ’ ° "“ ” "’ “ " ” “ ’g“ ” "‘
” “ ” "” ° ’ “ “ M”
Camelina rumelica ssp. susceptible Citrus limon resistanttranscas : ica
SW“ ” “ ’ g“ ""
Brassicajuncea var. resistant Hibiscus rosa-sinensis resistantinteri olia
Summary
Since 1996 a hitherto unknown disease occurred on Camelina Sativa in different localities of Germany andCanada. Symptoms consisted of firstly water-soaked small spots on leaves, stems and silicles which tumed darkbrown to black later on. From infected tissue including shrivelled seeds, consistently a Gram-negative, rod-shaped, motile, fluorescent bacterium was isolated. The seed home pathogen is closely related to Pseudomonassyringae, because it fulfilled the LOPAT criteria of the Pseudomonas group la. The host range of the pathogenwas limited to few Camelina species. Biochemical, physiological and pathogenicity studies as well as theimplication of the Biolog system indicated the Camelina pathogen to differ from all other members of thePseudomonas group la. Consequently, the pathogen is described as a separate pathovar of Pseua’omonassyringae not yet included in the approved list ofPseudomonas syringae pathovars. The new name Pseudomonassyringae pv. camelinae is proposed. Representative strains of the pathogen have been introduced in the bacterialculture collection “ Gö ttinger Sammlung phytopathogener Bakterien” GSPB nos. 2726, 2728, 2729, 2738, Z745,2830 and 2833.
References
28. BOCHNER, B.R.: Sleuthing out bacterial identities. Nature 339, l989, 157-15829. BRADBURY, J,F.: Guide to Plant Pathogenic Bacteria. CAB lntemational, Farnham House, Farnham
Royal, UK, 198630. JONES, J.B.; CHASE, A R.; HARRIS, G.K,: Evaluation of the Biolog GN MicroPlate system for
identification of some plant-pathogenic bacteria. Plant Disease 77, 1993, 553-55831. KLEMENT, Z.; RUDOLPH, K.; SANDS, D.C.: Methods in Phytobacteriology. Acadé miai Kiado,
Budapest, 199032. LELLIOTT, R.A.; BILLING, E.; HAYWARD, A.C.: A determinative scheme for the fluorescent plant
pathogenic pseudomonads. J. Appl. Bacteriol. 29, 1966, 470-489
33. LELLIOTT, R.A; STEAD, D.E.: Methods for the Diagnosis of Bacterial Diseases of Plants. BlackwellScientific Publications, Oxford, 1987
34. MAVRIDIS, A.; PAUL, V.; RUDOLPH, K.: On the occurrence of bacterial blight, a new bacterial diseaseon Camelina sativa, induced by a pathovar of Pseudomonas syringae. Mitt. Biol. Bundesanst. Land-Forstwiitsch. H. 357, 1998, 110

13
Sensitivity to antibiotics and heavy metals and plasmid profiles forcharacterization of different races ofXanthomonas campestris pv.
malvacearum from different originsK. Abdel-Rehim, K. Rudolph
Institute ofPlant Pathology and Plant Protection, University of Gö ttingen, Grisebachstr. 6, D-37077 Gö ttingen,Germany email: [email protected]
lntroduction
Bacterial Blight of cotton caused by Xanthomonas campestris pv. rnalvacearum (Xcm) is an economicallyimportant disease worldwide, resulting in yield losses of l0~30% of seed cotton (Verma 1995; Zachowski andRudolph, 1988), The disease is potentially very destructive in areas where wind driven rain or sprinkler irrigationdisseminate the pathogen (Brinkerhoff, 1963, 1970; Innes, 1983).
Angular leaf spot is the disease term when leaves are infected, whereas the bacteria enter the parenchyrnatousleaf tissue through stomata and grow causing water soaking appearance resulting from the slime containingextracellular polysaccharides which is hydrophilic and fill the intercellular spaces of the plant leaf tissue(Rudolph er al.. 1989),
Indigenous plasmids have have been found in every strain examined of Xanthomonas campesrris pv.malwzcearum isolated from cotton (Lazo and Gabriel, 1987; Chakrabarty, 1992; and Chakrabarty et al., 1992). Itwas found that the majority of the plasmid containing Xanthomonas campestris pv. malvacearum strainscontained only one plasmid, but some carried two or more (Lazo and Gabriel, 1987).
Race 18 which was demonstrated to neutralize five B-genes (B7, B4, B1, BIN & BN) contains five plasmids (Size:60, 40, 10, 5.5 and 2.2 Kb). Moderately virulent race 26 which can neutralize three B-genes (B4, B; & BIN)contains three plasmids (60, 40 & 10 Kb) while weakly virulent race 5 which can neutralize only one B gene(Bm) contains only one plasmid (10 Kb).
Materials and Methods
The bacterial strains originating from different countries (USA, Turkey, Nicaragua, Greece and Sudan) were
taken as lypholyzed samples from the Gö ttingen collection of phytopathogenic bacteria (GSPB) and resuspendedin King's B liquid medium (Bacto—peptone 20 g, KZHPO4 1.5 g, MgSO4X7H;O 1.5 g, glycerol 10 ml/l, pH 7.0)for 30 min, then a few droplets of this suspension were streaked on NGA plates (Nutrient broth 8 gm, Glucose10 gm, Yeast extract 3 gm, Agar 15 gm/L pH. 7.2) and incubated for 3 days.Resistance against antibiotics
The standardized single disk method (Bauer et al., 1966) was used for detennining the antibiotic resistance ofthe Xcm strains.
Heavy metal resistance and MIC determination
According to Ghosh et al., (1997) the Minimum inhibition concentrations (MICs) were deterrnined in triplicateby allowing the bacterial strains to grow on NGA plates containing different concentrations (0.25 mM, 0,5 mM,1 mM, 5 mM, 10 mM and 20 mM) of metal ions (ZnSO.,X7H2O, NiCl;X6H2O, Pb(CzH3O2)2X3H2O andCoSO.,X4H2O)Plasmid isolation
Cultures were grown to rnid-late logarithmic growth phase and extracted by the small-scale alkaline lysisextraction procedure (Kado and Liu, 1981), Extracted DNA was resuspended in TE (10 mM tris base, 1 mM Na;-EDTA, and 20 pg/ml of DNAse-free pancreatic RNAse; pH 7,6). Plasmid DNA fragments were separated bysize using agarose gel electrophoresis (0.7 % agarose, 2-5 V/cm in Tris—acetate buffer (40 mM Tris, 1 mM NazEDTA, adjusted to pH 7.6 with glacial acetic acid). Fragments were visualized by ultraviolet irradiation (302nm) after staining agarose gels in ethedium bromide (0.5 pg/ml).

14
Table 1:
Race
(‘ I...R 7 R l0 R 11z
m voouoocooomwr-— — .-—.....oooco er v- in ac no .-- N ‘n° ° ° ° ° ° Nr'ir’ 1-'
At_b_t.E 22222322221223 2‘ 2 2‘ Q 3 3 ä 2222332,?»
n l 1o ics
SulphamethazoH-Trimetho + + + + + + + + + + + + + + + + + + +
rim 23.75+1.25 ; SXoxaciiiins : OXPenicillin 10 (P)Cefaiexinso z CNCiindam cin 10 z CCOiioxacin 10 z OFX ZIIIIIIIIIIIIII— Z— — — — — IIIIIIII
ZIIIIIIIIIIIIII— — — — — — — IIIIIIIIPol in B 3001-13 PB — IIIlllllIIllIl— — — EE— — IIllIIII
Ell!IIIIIIIIIIIEEZZZZHIIIIIIIITobrom cin 10 z N ElIIIIIIIlIIIlI— E— E— — — llIIIIlI
XIIIIIIIIIIIIIIEHEEHZZIIIIIIII
Chloramhenicol 30 ; C _Gemam ein i0 z Gm — IIllIIIEIIIIIIEEEEEEEEEEEEEEENaiidixic acid 3o z (NA) — llIllllIlIllllE— — — — — ElllIllII
Discussion
' Although the majority of genes involved in the induction and development of disease by xanthomonads are
probably located on the main chromosome, some pathogenicity and virulence genes are present on plasmids,including avr genes, hrp genes and genes that encode the production of toxins (Sigee, 1993). The plasmids curedstrains were avirulent, suggesting strongly the role of individual plasmids in neutralizing respective B-genes(Verma 1995). When the 10 Kb plasmid (common in the three races) was transferred to avirulent pIasmid-curedstrains, the virulence for gene BIN was restored (Sathyanarayana and Verma, 1993; Verrna 1995). The results ofour studies showed that, strains of race 18 and race 4 possess the same plasmid content. Race 18 has the abilityto infect 9 different cotton lines while race 4 infects only three lines. Obviously the higher virulence of race 18 isnot due to plasmids. Also race 12 which developed from race 11 and has the ability to infect one cotton linemore than race 11, possesses the same plasmid profile than race 11. All strains tested showed the same MICsvalues of heavy metals although possessing different plasmid profiles which indicates that the genes of heavymetals resistance are located in the chromosomal DNA. The strains of race 6 and 7 were more resistant to
antibiotics than the most virulent race 18 which, however, possesses more plasmids than race 6 and 7.
Results
All of the strains tested were resistant to SXT, OX, P, CN and CC except race 4 which was sensitive to SXT. Allstrains were sensitive to each of OFX, TE, PB, VA, NN, K, Gm, C and NA, with the exception of race 6 andrace 7 being resistant to C and NA. Thus, strains of race 6 and race 7 are the most resistant ones while race 4showed the highest sensitivity to antibiotics (Table 1) .
The MICs values were the same in all strains tested. Theplasmid profiles revealed four groups: strains of race 18 and race 4 contained four plasmids of 65,0 Kb, 56.5Kb, 47.5 Kb and 30.0 Kb (Fig.1); strains of race 6 and 7 contained three plasmids of 95.0 Kb, 67.0 Kb and 63.0Kb (Fig.2), also strains of race ll and 12 showed three plasmids but of 77.5 Kb, 60.0 Kb and 34.8 Kb (Fig. 3),while the strains of race 10 contained only tow plasmids of 60.0 Kb and 34.0 Kb, (Fig.3).
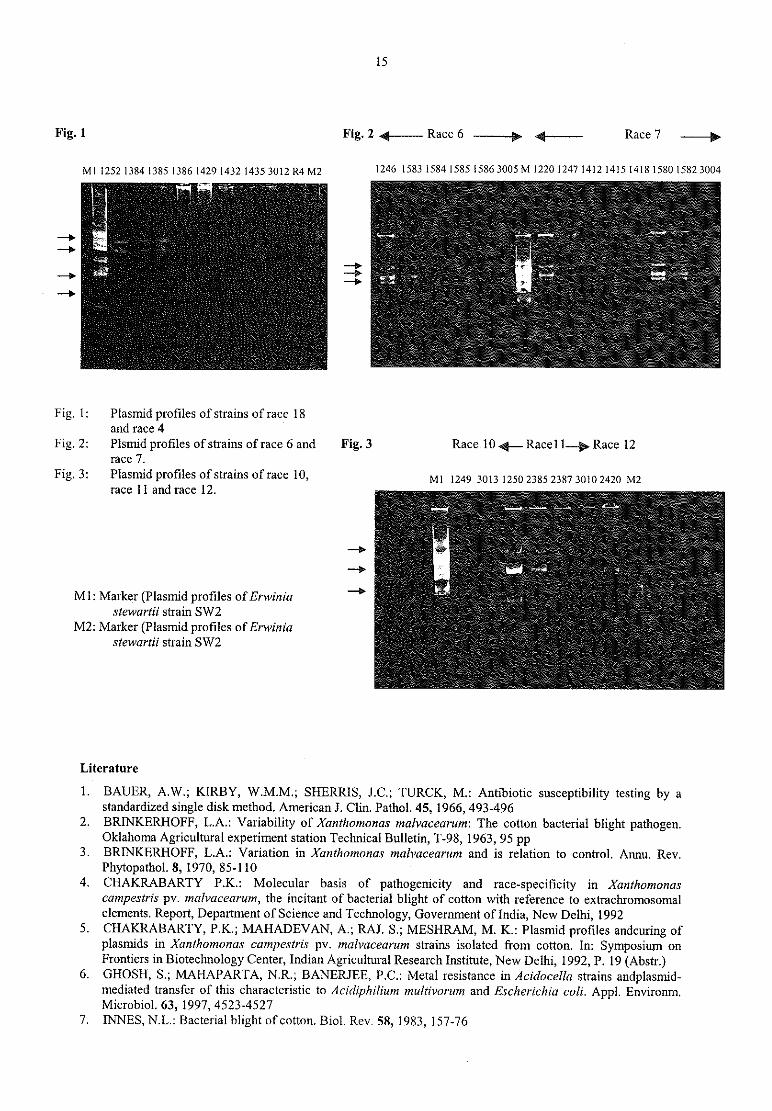
15
Fig. 1 Fig. 2 4? Race 6 — — —.’ {—— — Race 7 __y
M.,2521384,3g51335142914321435 3012 R4 M2 1246 1583 1584 1585 1586 3005 M122012471412 1415141815801582 3004
— )z. „ „ - « <-
— > .. i
— — >V
2.2„
Fig. l: Plasmid profiles of strains of race 18and race 4
Fig. 2: Plsrnid profiles of strains ofrace 6 and Fig. 3 Race 10 4_ Race11__.p Race 12race 7.
Fig 31 Plasmid Profiles Of Strains Of m96 10, Ml 1249 3013 1250 2385 2387 3010 2420 M2race 11 and race 12.
__q
'
‚ __‚ „ ‚_. _.__ ‚ z a 2.:. ' ‘
— » .4 .
...; w’ n51’ z a
M1: Marker (Plasmid profiles ofErwinia -7 9?!v. 1 .
i:
stewartii strain SW2M2: Marker (Plasmid profiles ofErwinia
stewartii strain SW2.
Literature
1. BAUER, A.W.; KIRBY, W.M.M.; SHERRIS, J.C.; TURCK, M.: Antibiotic susceptibility testing by a
standardized single disk method. American J. Clin. Pathol. 45, 1966, 493-4962. BRINKERHOFF, L.A.: Variability of Xanthomonas malvacearum: The cotton bacterial blight pathogen.
Oklahoma Agricultural experiment station Technical Bulletin, T-98, 1963, 95 pp3. BRINKERHOFF, L.A.: Variation in Xanthomonas malvacearum and is relation to control. Annu. Rev.
Phytopathol. 8, 1970, 85-1104. CHAKRABARTY P.K.: Molecular basis of pathogenicity and race~specificity in Xanthomonas
campestris pv. malvacearum, the incitant of bacterial blight of cotton with reference to extrachromosomalelements. Report, Department of Science and Technology, Government of India, New Delhi, 1992
5. CHAKRABARTY, P.K.; MAHADEVAN, A.; RAJ. S.; MESHRAM, M. K.: Plasmid profiles andcuring ofplasmids in Xanthomonas campestris pv. malvacearum strains isolated from cotton. In: Symposium on
Frontiers in Biotechnology Center, Indian Agricultural Research Institute, New Delhi, 1992, P. 19 (Abstr.)6. GHOSH, S.; MAHAPARTA, N.R.; BANERJEE, P.C.; Metal resistance in Acidocella strains andplasmid-
mediated transfer of this characteristic to Acidiphiliurn multivorum and Escherichia coli. Appl. Environm.Microbiol. 63, 1997, 4523-4527
7. INNES, N.L.: Bacterial blight ofcotton. Biol. Rev. 58, 1983, I57-76

16
8. KADO, C.I; LIU, S.~T.: Rapid procedure for detection and isolation of large and small plasmids. J.Bacteriol. 145, 1981, 1365-1373
9, LAZO, G.R.; GABRIEL, D.W.: Conservation of plasmid DNA sequences and pathovar identification ofstrains ofXanthomonas campestris. Phytopathology 77, 1987, 448-453
10. RUDOLPH, K.; GROSS, M,; NEUGEBAUER, M.; ETAL: Extracellular polysaccharides as determinantsofleaf spot diseases caused by pseudomonads and xanthomonads. In. Graniti, A.; Durbin, R.D.; Ballio, A.(Eds.): Phytotoxins and plant pathogenesis, NATO ASI series, Vol. 427, Springer-Verlag, Berlin, 1989, 177-218
,11, SATHYANARAYANA, N,; VERMA, J.P.: Possible role of plasmids in the virulence of Xanthomonas
campesrris pv.. malvacearum. Indian Phytopathology 46, 1993, 165-16612. SIGEE, D.C.: Genetical analysis of plant pathogenic bacteria. In: Bacterial plant pathology, cell and
molecular aspects. Cambridge University Press. 199313. VERMA, J „P.: Advances in bacterial blight of cotton. Indian Phytopathology 48, 1995, 1-1314. ZACHOWSKI, A.; RUDOLPH, K,: Characterization of isolates of bacterial blight of cotton (Xanthomonas
campestris pv. malvacearum) from Nicaragua. J. Phytopathol. 123, 1988, 344-52

17
Session 2: Host/Parasite relationships
Enzymatic digestion of plant pectins from susceptible and resistantcultivars of celery and extraction of a putative carbohydrate binding
protein from phytopathogenic pseudomonads3‘
B. Venkateshi, M.I. Khan*, A. Pant*, K. Rudo1ph§5 Institute for Plant Pathology and Plant Protection, Grisebachstr. 6, D —37077, Gö ttingen, Germany
* Division ofBiological Science, National Chemical Laboratory, Pune, India.
Introduction and Background
Pectins are complex carbohydrates present in the middle lamella of all higher plant cell walls. Structurally, theyare composed of a polygalacturonic acid backbone to which various sugars are attached via glycosidic linkages.Earlier studies from our group have demonstrated that plant pectins interact with bacterial lipopolysaccharides(LPS) and that the nature of the interaction plays a significant role in pathogenesis (Grolms and Rudolph, 1997;Rudolph, 2001). Knowledge of the chemical composition of pectins can help us to better understand the nature
of these interactions. We therefore carried out enzymatic hydrolysis to differentiate pectins from susceptible andresistant cultivars on the basis of their chemical composition. These pectins were also studied for their inhibitiontowards a carbohydrate binding protein from phytopathogenic pseudomonads.
Materials and Methods
Enzymatic hydrolysis ofpectins
We selected the resistant (R) celery (Apium graveolens var. rapaceum) cv. Monarch and the susceptible (S) cv.
Claret (Apium graveolens var. dulce) from which pectins were extracted as described previously‘ (Grolms and
Rudolph, 1996) and subjected to enzymatic hydrolysis with pectate lyase (PL) and polygalacturonase (PG). Thereaction mixture consisted of 5 mg pectins dissolved in 1 ml of 50 mM sodium acetate buffer (pH 5.5) and 17units of either PL or PG. Enzymatic digestion was allowed to proceed for 48 h at 37 ° C, Aliquots of 50 11l were
taken at different time intervals, and digestion was arrested by incubating the sample in a boiling water bath for10 min. The samples were passed through a membrane filter (4 pm) and further analyzed'by HPLC (DEAEglyco column).Extraction ofa carbohydrate bindingprotein (haemagglutinin)Bacterial strains (Pseudomona syringae pv. apii GSPB 2548 and P. s. pv. tomato GSPB 2317) were grown on
nutrient broth at 27 ° C for 48 h. The cells were harvested by centrifugation at 10,000 x g for 20 min andsuspended in 50 mM acetate buffer (pH 6.0) containing 2 M urea, 5 mM EDTA and 1 mM PMSF. Thesuspension was stirred for 12 h at 4 ° C and centrifuged at 10,000 x g for 20 min. The supematant was dialyzedagainst demineralized water for 72 h and lyophilized. The lyophilized powder was then dissolved in double-distilled water and purified on a phenyl—sephadex negative absorption column through which the protein came
unbound. The protein was further purified on DEAE-sephadex and eluted using 0.4 M NaCl and quantified
following the method of Lowry et al., (1951).
Haemagglutination and inhibition assays
The partially purified proteins were serially diluted from a starting concentration of 0.5 pg/ml with Tris bufferedsaline (20 mM Tris—HCl (pH 7.2), 150 mM NaCl) in a microtitre plate and incubated for 1 h at 20 “ C with an
equal volume of 3% (v/v) suspension of rabbit erythrocytes in TBS and examined for agglutination. Inhibition
assays were carried out with pectins from resistant and susceptible cultivars of tomato and celery plants. Somecommercial pectins (apple and citrus) and sugars were also tested. The inhibition assays were performed
essentially as described above. Dilutions of sugars starting from 1 M were incubated with the protein (0.025 pg)suspension and an equal volume of rabbit erythrocytes. The wells were observed after l h for inhibition of
haemagglutination.
Results and Discussion
In the present study, LPS were extracted from two pathovars of Pseudomonas syringae, which were earliershown to possess similar antigenicity towards monoclonal antibodies raised against the O-specific moieties ofthe LPS (Ovod et al., 1997). Pectins were extracted from susceptible and resistant cultivars of their respectivehosts and the nature of interactions between LPS and pectins was characterized rheologically. Earlier, a
18
synergistic interaction was reported only for incompatible combinations but not for incompatible ones (Laux et
al., 1998). Interestingly, LPS from P. s, pv. apii interacted synergistically with pectins from the susceptible
assumed non-host tomato (Venkatesh and Rudolph, 2001 a). Cross-infection of susceptible non—hosts was indeed
confirmed through plant inoculation studies conducted in the green house (Venkatesh et al., unpublished data).
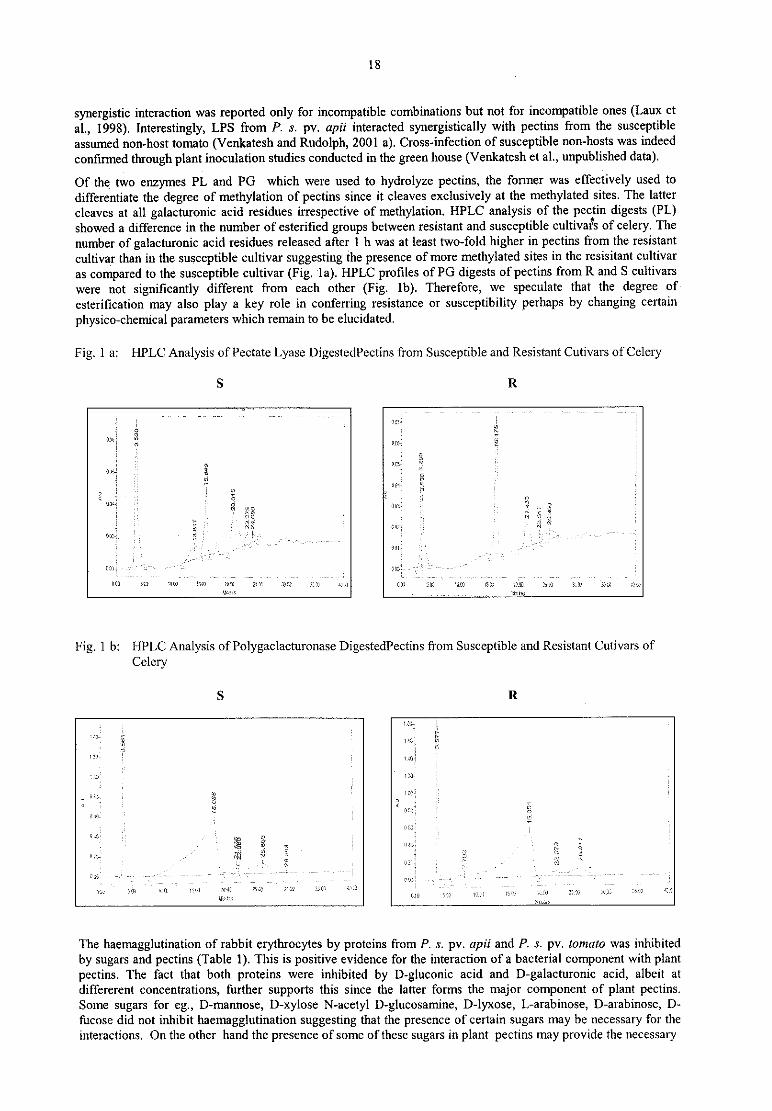
Of the two enzymes PL and PG which were used to hydrolyze pectins, the former was effectively used to
differentiate the degree of methylation of pectins since it cleaves exclusively at the methylated sites. The latter
cleaves at all galacturonic acid residues irrespective of methylation. HPLC analysis of the pectin digests (PL)showed a difference in the number of esterified groups between resistant and susceptible cultivafs of celery. The
number of galacturonic acid residues released after 1 h was at least two-fold higher in pectins from the resistant
cultivar than in the susceptible cultivar suggesting the presence of more methylated sites in the resisitant cultivar
as compared to the susceptible cultivar (Fig. la). HPLC profiles of PG digests of pectins from R and S cultivars
were not significantly different from each other (Fig. lb). Therefore, we speculate that the degree of
esteritication may also play a key role in conferring resistance or susceptibility perhaps by changing certain
physico—chemical parameters which remain to be elucidated.
Fig. 1 a: HPLC Analysis of Pectate Lyase DigestedPectins from Susceptible and Resistant Cutivars ofCele1y
S R
.
‘a’ i" 'T M '
I
32:.I V V 7 V
(H: gweil
2 iii ’ y
‘JC-ll ‘.
2‚ „
7i
,
I s Ev E W zur, il “ ’
ml jy
k
f —.e
als 5:1w’
ism'
1:-so
"
{1I. ‘:111 (so.i
m 2.53’ am m w
111..
Fig. 1 b: HPLC Analysis ofPolygaclacturonase DigestedPectins from Susceptible and Resistant Cutivars of
Celery
S R
.
l185 I
"f tit‘ .11. . „ u;
1»;
"D: '
_
I
V
.
i
,
i
z '— "
'
'
i’ '
1
:1»; 5111 „u. „ .1. 2m Lac: s;m 25.-z .
z
v; M
ilr fizz C13 i321 1‘. ‘.1! 15x71 .« w .‚ ‚u — » \~ z»
The haemagglutination of rabbit erythrocytes by proteins from P, s. pv. apii and P. s. pv. tomato was inhibited
by sugars and pectins (Table 1). This is positive evidence for the interaction of a bacterial component with plant
pectins. The fact that both proteins were inhibited by D-gluconic acid and D-galacturonic acid, albeit at
dlffererent concentrations, further supports this since the latter forms the major component of plant pectins.Some sugars for eg., D-mannose, D—xylose N—acetyl D~glucosamine, D-lyxose, L-arabinose, D-arabinose, D-
fucose did not inhibit haemagglutination suggesting that the presence of certain sugars may be necessary for theinteractions. On the other hand the presence of some ofthese sugars in plant pectins may provide the necessary
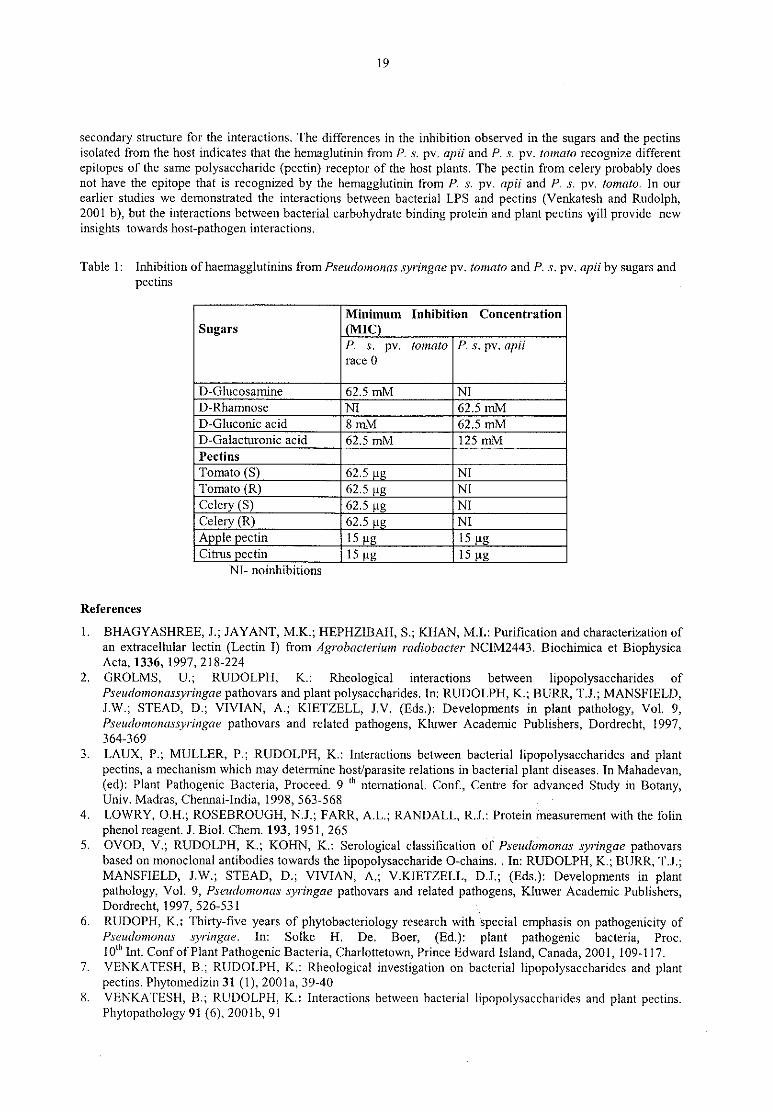
19
secondary structure for the interactions. The differences in the inhibition observed in the sugars and the pectinsisolated from the host indicates that the hemaglutinin from P. s. pv. apii and P. s. pv. tomato recognize differentepitopes of the same polysaccharide (pectin) receptor of the host plants. The pectin from celery probably doesnot have the epitope that is recognized by the hemagglutinin from P. s. pv. apii and P. s. pv. tomato. In our
earlier studies we demonstrated the interactions between bacterial LPS and pectins (Venkatesh and Rudolph,2001 b), but the interactions between bacterial carbohydrate binding protein and plant pectins will provide new
insights towards host—pathogen interactions.
Table 1: Inhibition ofhaemagglutinins from Pseudomonas syringae pv. tomato and P. s. pv. apii by sugars andpectins
Minimum Inhibition Concentration
Sugars MICP. s. pv. tomato P. S. pv. apii
race 0
— —
NI- noinhibitions
References
1. BHAGYASHREE, J .; JAYANT, M.K.; HEPHZIBAH, S.; KHAN, M.I.: Purification and characterization ofan extracellular lectin (Lectiri I) from Agrobacterium radiobacter NCIM2443. Biochirnica et BiophysicaActa, 1336, 1997, 218-224
2. GROLMS, U.; RUDOLPH, K,: Rheological interactions between lipopolysaccharides of
Pseudomormrsyringae pathovars and plant polysaccharides, In: RUDOLPH, K.; BURR, Tl; MANSFIELD,J.W.; STEAD, D.; VIVIAN, A.; KIETZELL, J.V. (Eds.): Developments in plant pathology, Vol. 9,Pseuriomonnrsyringtze pathovars and related pathogens, Kluwer Academic Publishers, Dordrecht, I997,364-369
3. LAUX, P.; MULLER, P.; RUDOLPH, K,: Interactions between bacterial lipopolysaccharides and plantpectins, a mechanism which may determine host/parasite relations in bacterial plant diseases. In Mahadevan,(ed): Plant Pathogenic Bacteria, Proceed. 9 ‘ h ntemational. Conf, Centre for advanced Study in Botany,Univ. Madras, Chennai-India, 1998, 563-568
4. LOWRY, O.H.; ROSEBROUGH, N.J.; FARR, A.L.; RANDALL, R.J.: Protein measurement with the folinphenol reagent. J. Biol. Chem. 193, 1951, 265
5. OVOD, V.; RUDOLPH, K.; KOHN, K.: Serological classification of Pseudomonas syringae pathovarsbased on monoclonal antibodies towards the lipopolysaccharide O-chains. . In: RUDOLPH, K.; BURR, T.J.;MANSFIELD, J,W.; STEAD, D.; VIVIAN, A.; V.KIETZELL, D.J.; (Eds.): Developments in plant
pathology, Vol. 9, Pseudamonas syringae pathovars and related pathogens, Kluwer Academic Publishers,Dordrecht, 1997, 526-531
6. RUDOPH, K.: Thirty-five years of phytobacteriology research with special emphasis on pathogenicity ofPseudomanas syringae. In: Solke H. De, Boer, (Ed.): plant pathogenic bacteria, Proc.10"‘ Int. Conf ofPlant Pathogenic Bacteria, Charlottetown, Prince Edward Island, Canada, 2001, 109-l 17,
7. VENKATESI-l, B.; RUDOLPH, K.: Rheological investigation on bacterial lipopolysaccharides and plantpectins. Phytomedizin 31 (1), 200121, 39-40
8. VENKATESH, B.; RUDOLPH, K.: Interactions between bacterial lipopolysaccharides and plant pectins.
Phytopathology 91 (6), 2001b, 91
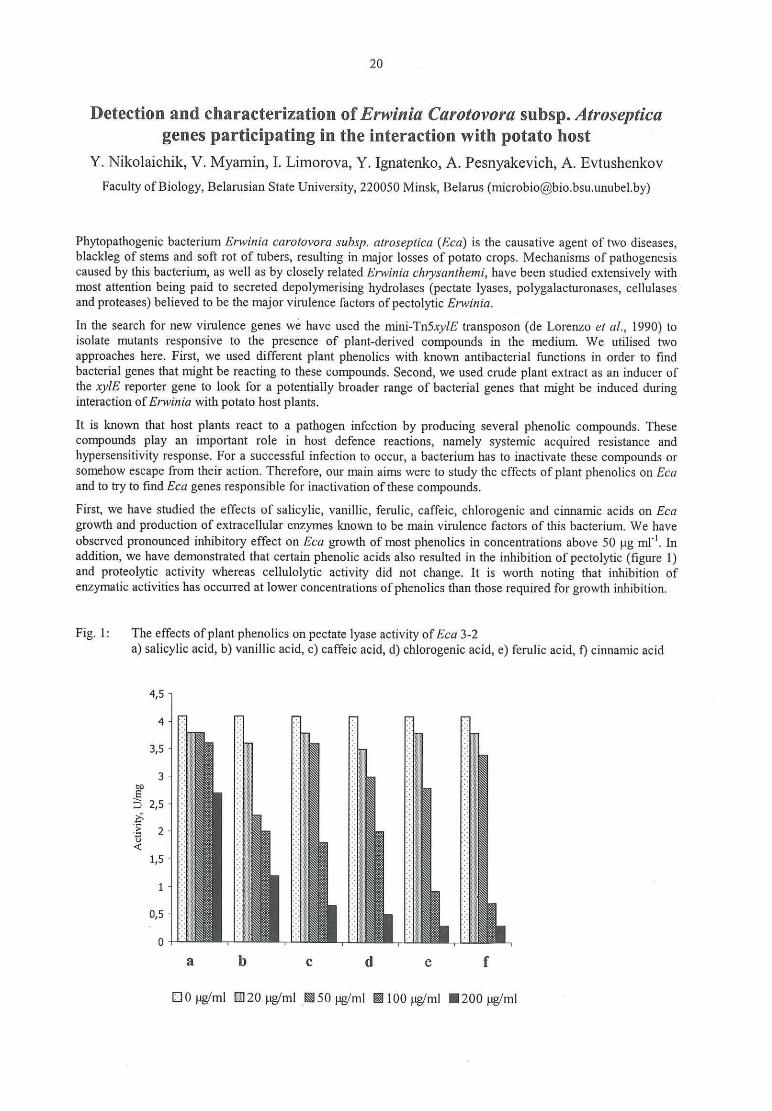
20
Detection and characterization ofErwinia Carotovora subsp. Atrosepticagenes participating in the interaction with potato host
Y. Nikolaichik, V. Myamin, I. Limorova, Y. Ignatenko, A. Pesnyakevich, A. Evtushenkov
Faculty ofBiology, Belarusian State University, 220050 Minsk, Belarus ([email protected])
Phytopathogenic bacterium Erwinia carotovora subsp. atroseptica (Eca) is the causative agent of two diseases,blackleg of stems and soft rot of tubers, resulting in major losses of potato crops. Mechanisms of pathogenesiscaused by this bacterium, as well as by closely related Erwinia c/irysarzthemi, have been studied extensively withmost attention being paid to secreted depolymerising hydrolases (pectate lyases, polygalacturonases, cellulasesand proteases) believed to be the major virulence factors of pectolytic Erwinia.
In the search for new virulence genes we have used the mini—Tn5xylE transposon (de Lorenzo et al., 1990) toisolate mutants responsive to the presence of plant-derived compounds in the medium, We utilised twoapproaches here. First, we used different plant phenolics with known antibacterial functions in order to findbacterial genes that might be reacting to these compounds. Second, we used crude plant extract as an inducer ofthe xyIE reporter gene to look for a potentially broader range of bacterial genes that might be induced duringinteraction of Erwinia with potato host plants.
It is known that host plants react to a pathogen infection by producing several phenolic compounds. Thesecompounds play an important role in host defence reactions, namely systemic acquired resistance and
hypersensitivity response. For a successful infection to occur, a bacterium has to inactivate these compounds or
somehow escape from their action. Therefore, our main aims were to study the effects of plant phenolics on Ecaand to try to find Eca genes responsible for inactivation ofthese compounds.
First, we have studied the effects of salicylic, vanillic, ferulic, caffeic, chlorogenic and cinnamic acids on Ecagrowth and production of extracellular enzymes known to be main virulence factors of this bacterium. We haveobserved pronounced inhibitory effect on Eca growth of most phenolics in concentrations above 50 pg ml". Inaddition, we have demonstrated that certain phenolic acids also resulted in the inhibition of pectolytic (figure 1)and proteolytic activity whereas cellulolytic activity did not change. It is worth noting that inhibition ofenzymatic activities has occurred at lower concentrations of phenolics than those required for growth inhibition.
Fig. 1: The effects of plant phenolics on pectate lyase activity of Eca 3-2a) salicylic acid, b) vanillic acid, c) caffeic acid, d) chlorogenic acid, e) ferulic acid, f) cinnamic acid
4,5
4 "i l i‘ :3 5:e; I 1525;:
.:2 :;
,5 -: \:a:2:s r ‚"1; .;:«=;‘-: : 1:2.. :- es?3
‘ :1 .: :; ‘Er! :; .11‘? :=: 15:81:15 ~ 1:5 2.:. :- n -: 12s
3- § :=;::= -. -;::- r 1-1» 21:; » . at
:': 2 :,j§§ :- f ‘ii; 3:=0 \-:::;: - 1;; ‚ H,;, .-‘.1=1. 11;; v -,;;,.;E v:- § ;i;3:E '- I‘:E: -. -. it-« :- 151i
5 z: :: :111 e11: .211:=2 2:
.2 2 -- :: 151% -: 211 -:ö ;' .: -I I: iii ::< -'
v. ‘St -art? ‘ - :
1.5 E. i: iii} i: it L 5 1:I; 8222:5; I. - 12822222‘ :I 3
5x212. ;- seats: ‘ - asess; I‘ - 12.v. 1181:1. - § s:a2s: : 21:; § a;zs2:- ‚ 821222 „ I sizes: I
a b c d e f
E10 ug/ml E120 uyml 50 pg/ml E100 pg/ml I200 pg/ml

21
In order to find Eca genes responding to the presence of phenolics in the media we have used transposonmutagenesis with the mini-Tn5xyIE transposon. Mutants with random insertions of this transposon in Eca
genome were screened for changes in expression of promoterless xylE reporter gene in the presence of differentphenolic compounds. Out of 3900 clones tested five with the desired phenotype were found. 10-15-folddifferences in the levels of reporter expression were routinely observed for most of the clones when grown in a
medium with and without the corresponding phenolics (figure 2). No noticeable change in the expression of theknown virulence factors of Eca resulting from these insertions was observed (except for moderate increase of
proteolytic activity in strain 4075), nevertheless, virulence of the mutants was somewhat reduced‘ All mutants
were still able to cause blackleg symptoms, but symptom development was delayed and damaged areas were
smaller‘
Fig. 2: Induction of the catechol—2,3~dioxygenase activity in strains 4016 (a) and repression of the catechol-2,3-dioxygenase activity in strain 4104 (b) by plant phenolics vanillic acid (1) and salicylic acid (2)
a) b)
4°3o
F“ i?) ‘E’ 2s
25 1E 203 20 s 15ä 15 1 E l 2z- ; >: 1oE 10 ‘Eg 5 "E 5
0 «+++
<o
b
0 20 50 100 200 o 2o 5o 100 200
C0n€ €l‘ |iF8|i0n» HE m14 Concentration, pg m1’ !
For the 4104 strain catechol-2,3-dioxygenase was inhibited by vanillic and salicylic acids. Increasing theconcentrations of these phenolics from 20 to 200 pg/ml resulted in approximately tenfold decrease of catechol~2,3—dioxygenase activity (figure 2).
Determining the nucleotide sequence of mutated genes has revealed that four out of five mutants have the mini-Tn insertion in one locus with three different mutations (4016, 4006 and 4585) being within one gene and thefourth (4075) located in its neighbour (figure 3). The fifth mutation (4104) is located elsewhere.
Fig. 3: The stmcture of plant phenolics responsive cluster. The positions ofmini-Tn5xyIE insertions(indicated by arrows) and sizes of putative OR.Fs are dravwi to scale.
ORF} ORF2 ORF1
0.5 kl.) 4585 4006 4016 4075
Unfortunately, sequence analysis did not give us definitive clues to the functions of the putative ORFS. ORFlgene is highly homologous (68 per cent identical) to a small gene yeaR from E. coli with unknown function.Both of these genes are homologous to the tehB family of tellurite resistance proteins, but tehB proteins are
larger and clearly contain at least two functional parts‘ The larger C-terminal domain is thought to be requiredfor tellurite resistance, while the function of N-terminal domain (where the homology to ORF1 lies) is unknown,In E. coli those two domains are separated with yeaR representing the N-terminal domain and a separate tehB
gene required for tellurite resistance.
There is a sequence, TTCACA(N),(,TATGAT, with a good homology to the consensus57°
promoter located 68bases upstream of oz;/'/. There is also a sequence, AaTtCACAAACAtTaAAAg ,
with some resemblance to theconsensus MarA binding site (marbox, AYnGCACnnWnn.RYYAAAY) overlapping the -35 region of thisputative promoter. MarA is transcriptional regulator of mar regulon studied in several enterobacteria. Membersof this regulon are responsible for the efflux of many toxic compounds from the cell, including several

22
antibiotics and phenolic compounds. However, right now we have no evidence for the involvement of MarA or
MarR in the regulation of plant phenolics responsive genes. At the moment we can only say that low levels ofantibiotic resistance in Eca3-2 are not induced (as in E. coli) by salicylic acid or any other phenolic acids used inthis study.ORF2 has a high homology (37 per cent identity) to the YdfE protein with unknown function from B. subtilisand a weaker homology to a large family of flavin reductases. These flavin reductases are usually part of two-
component enzyme systems in which a small flavin reductase is providing reduced FMN to a large subunitwhich is usually a mono- or dioxygenase responsible for the degradation of certain toxic organic compounds
including various phenolics.
Another gene (orf3) is located downstream of 0rf2. This gene is transcribed convergently to my‘ ? and codes for a
TetR/AcrR—type transcriptional regulator. The targets of this group of regulators are genes encoding proteinsinvolved in cell envelope permeability. We have cloned the orf3 gene, but have found no evidence for its effecton the expression of orfl or er]? genes (data not shown).The product of last gene repressible by phenols, orf4, which is not linked to the three orfs described above, is
highly homologous (76 per cent identical) to an ORF with unknown functions from E. amylovora. This ORF
(orfl3 in the GenBank entry L258Z8) is the second located afler the last gene (hrcU) of the hrpJ operon, codingfor the components of type III secretion machinery. Both am in E. amylovora following the hrcU gene (0rf12and orf13) are probably cotranscribed from the Hrpl.-dependent promoter located just in front of orfI2 whichsuggests possible involvement of their products in virulence. However, transcriptional organisation in Eva is
clearly different — neither a homologue of orfl2 nor a HrpL—dependent promoter could be identified upstream ofthis gene. Both the orf13 product and the product of its orthologue from Eca 3-2 have rather weak, but clear
homology to a number of proteins, of which with known functions are 4-hydroxybenzoyl-CoA thioesterase and
hydroxymyristoyl-CoA dehydratase.As the sequence analysis gives some hints to the possible involvement of the phenol—responsive genes describedabove in the degradation (or at least detoxification) of phenolic compounds, we have used HPLC analysis to
check if the wild type strain can degrade (or somehow transfonn) several plant phenolics. This analysis hasdemonstrated that salicylic, vanillic, chlorogenic, caffeic, cinnamic and ferulic acids are not degraded by Eczzcells and inactivation of their toxic effect must be due to some other mechanism (for example, their activeefflux).
We have also characterised the effects of phenolic acids on pectate lyase synthesis in the strains Eca3-2 (wildtype) and Eca 1-73 (mutant resistant to glucose catabolite repression). The Ecu1—73 strain was also obtained bymutagenesis with the mini—Tn5xylE transposon. The synthesis of pectate lyases was repressed by phenolic acids
(100 pg/ml ) in Em 3-2 by 70%, whereas the Eca1—73 mutant was resistant to this repression. Glucose repressedthe synthesis of pectate lyase in Eca3-2 by more than 90%, but only by 20 % in Ecal-73. The mutant strain hadmore pronounced decrease of pectate lyase activity when glucose was added together with phenolic acids (30-70%).
Our results indicate that there are common messengers in realisation of effects of Catabolite repression andphenolic acids on pectate lyase synthesis. The molecular cloning ofmutant gene is in progress now.
A search for the genes inducible by potato stem extract has led to the isolation of genes participating in threedifferent processes — iron transport, pectate metabolism and, surprisingly, nitrate reduction. Three mutationsfrom the last two classes are characterised in some detail.
Two independently isolated mutants impaired in pectate metabolism appeared to have mini-Tn5xylE insertion inthe neighbouring genes, kduD, coding for 2,5-diketo~3-deoxygluconate dehydrogenase and pelW, coding for exo
pectate lyase. Both mutants have drastically reduced virulence on potato stems - they are unable to cause
blackleg symptoms. Moreover, both mutants were also unable to cause hypersensitivity response wheninoculated into bean (Viciafaba) leaves. Investigation of the reasons behind the severity of the effects exerted bythose mutations onto virulent properties of Eca has revealed that the mutants accumulate certain products ofpectin degradation in the media and their accumulation leads to significant changes in the production of severalsecreted depolymerising enzymes. The kduD mutation results in the accumulation of 2,5-diketo—3—deoxygluconate (DKII) in the cells and cultural media when the mutant is grown on polygalacturonatecontaining media. We have investigated the influence of different concentrations of DKII onto the majorpathogenicity factors of Eca. The levels of all three virulence factors (pectate lyases (Pel), proteases (Prt) andcellulases (Cel)) were increased by low (l—5 pg/ml) and repressed by high (10—125 pg/ml) DKII concentrations.In Erwinia chrysanthemi (Eck) DKII has increased Pel production with increasing concentrations, caused minoralterations of Cel production and had no effect onto the production of Prts. This implies that, in the contrast to
Ech, Eca has a common regulatory mechanism for virulence factors responding to the presence of
polygalacturonate metabolites. According to the published data of other researchers these metabolites in Erwinia

23
clzrysunr/renzi cells exert their action through the repressor protein KdgR which binds promoter regions of pectatelyase genes. We plan to construct a ka'gR mutant of Em and to compare it with the analogous mutant of Erwiniaclnysanrhemi. At this stage the k(lgR gene ofEca is amplified, cloned and sequenced.
One more mutant with reporter gene expression inducible by potato stem extract was clearly different fi'om bothkduD and pe/W in that it was induced by a soluble fraction of stem extract (rather than insoluble as in case ofkduD and pelW). This induction was most pronounced on minimal media, but still visible on LB.
Sequencing the site of insertion has revealed that the mini-Tn5,rylE transposon is inserted into‘ an open readingframe with high homology to several prokaryotic genes coding for beta subunit of nitrate reductase - 84%identity of its product to major membrane nitrate reductase subunit NarH of E. coli and 74% - to minormembrane nitrate reductase subunit NarY.
The nar mutant displayed markedly reduced virulence and completely failed to induce typical blacklegsymptoms.
References
57. DE LORENZO, V.; HERRERO, M.; JAKUBZIK, U.; TIMMIS, K.: Mini Tn5 transposon derivates forinsertion, mutagenesis, promoter probing and chromosomal insertion of cloned DNA in Gram-negativeEubacteria. J. Bacteriol 172, 1990, 6568-6572
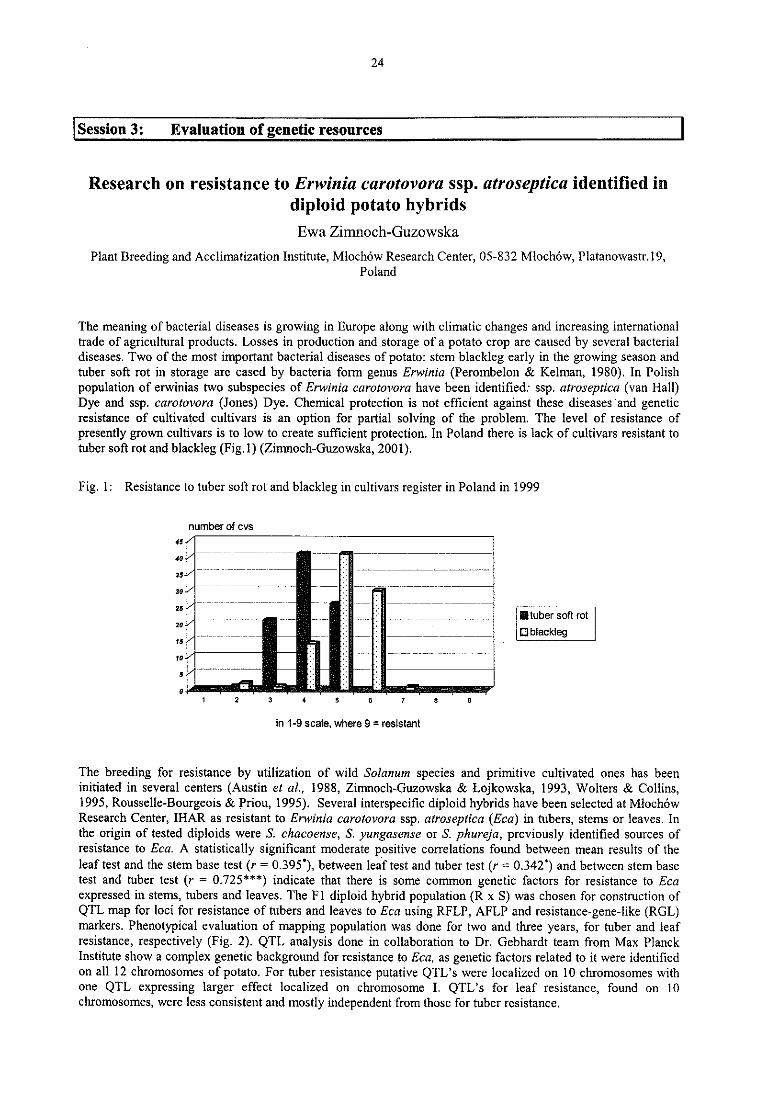
24
Session 3: Evaluation of genetic resources
Research on resistance to Erwinia carotovora ssp. atroseptica identified in
diploid potato hybridsEwa Zimnoch-Guzowska
Plant Breeding and Acclimatization Institute, Mlochow Research Center, 05-832 Mloché w, Platanowastr.l9,Poland
The meaning of bacterial diseases is growing in Europe along with climatic changes and increasing internationaltrade of agricultural products. Losses in production and storage of a potato crop are caused by several bacterialdiseases. Two of the most important bacterial diseases of potato: stern blackleg early in the growing season andtuber sott rot in storage are cased by bacteria form genus Erwinia (Perombelon & Kelman, 1980). In Polishpopulation of erwinias two subspecies of Erwinia carotovora have been identified: ssp. atroseptica (van Hall)Dye and ssp. carotovora (Jones) Dye. Chemical protection is not efficient against these diseases ‘and geneticresistance of cultivated cultivars is an option for partial solving of the problem. The level of resistance of
presently grown cultivars is to low to create sufficient protection. In Poland there is lack of cultivars resistant totuber soft rot and blackleg (Fig.1) (Zimnoch-Guzowska, 2001).
Fig. 1: Resistance to tuber soft rot and blackleg in cultivars register in Poland in 1999
number of cvs
45 ‘4a
7"" TTWWT ‘ WT 7" " T
157 Wu‘ 7 Tut 77“ ’ 7 ’ “ TT7?+7é A§ ” Mi-i
Ja” ” "” ’ "’ "’ ’ ’ §§ ’""" ’ ” "” ” '"'” '” T
u.„ _‚ „ „ _,_, „ __ _„ _„ _„ „ „ „ ‚ _
Iluber soft rot" ‘ „ _„ W
:3g ____"___
Dblackleg1a ' E — —
‘ Z
w._
p E; ._ ..____..._-_
JHVWM‘ -T
_..— ._' '- .2 _.
___ ‚
uI 2 3 4 5 6 7 8 9
in 1-9 scale, where 9 = resistant
The breeding for resistance by utilization of wild Solanum speeies and primitive cultivated ones has beeninitiated in several centers (Austin et al., 1988, Zimnoch-Guzowska & Lojkowska, 1993, Wolters & Collins,1995, Rousselle-Bourgeois & Priou, 1995). Several interspecific diploid hybrids have been selected at MlochowResearch Center, IHAR as resistant to Erwinia carotovora ssp. atroseptica (Eca) in tubers, stems or leaves. Inthe origin of tested diploids were S. chacoense, S. yungasense or S. phureja, previously identified sources ofresistance to Eca. A statistically significant moderate positive correlations found between mean results of theleaf test and the stem base test (r = 0.395’ ), between leaf test and tuber test (r = 0.342’ ) and between stem basetest and tuber test (r = 0.725***) indicate that there is some common genetic factors for resistance to Ecaexpressed in stems, tubers and leaves. The F1 diploid hybrid population (R x S) was chosen for construction ofQTL map for loci for resistance of tubers and leaves to Eca using RFLP, AFLP and resistance-gene~like (RGL)markers. Phenotypical evaluation of mapping population was done for two and three years, for tuber and leafresistance, respectively (Fig. 2). QTL analysis done in collaboration to Dr. Gebhardt team from Max PlanckInstitute show a complex genetic background for resistance to Eca, as genetic factors related to it were identifiedon all 12 chromosomes of potato. For tuber resistance putative QTL’ s were localized on 10 chromosomes withone QTL expressing larger effect localized on chromosome I. QTL’ s for leaf resistance, found on 10chromosomes, were less consistent and mostly independent from those for tuber resistance.

25
Fig. 2: Distribution of genotypes in Fl mapping population DG 83-2025 (R) X DG 81-68 (S) according to
their tuber resistance to Erwinia carotovora ssp atroseptica in two year studies 1994 -1995.
60 ?” ‘ “ "‘ “ “ *“ “ “ ‘ — ‘ ” ‘ *"” ” "'” “ """
50 i6 ‘ “ *
V’ 5 DG81-68zu 59.40 * ’ ’ ’ äb DG83-2025 sO \30 ' \
\‘
u. 20 ä ä ä31: § § Ä \ \‘
° _5I5 It I10 \ \ 5 ä 5\ \ \ \ \I5 It I5 Wm
0 § .- ä ä S s s ä as ..- xx‘ .... ...-
_
..
,3: ‚g ‚f „ b „ Ä 055 „ 9 x59 e," N/:~',‘* G’? N53‘ e‘? G?’Du to ‘bfl/‘ A)‘ ‘ c)‘ ‘ ‘ ‘
q\ N/Qt ~\,\ Ox \”1,\ xbu \<:)\
Diameter of rotted tissue in mm
TR94 UTR95 ITR94/95
Inheritance of resistance to Eca found in diploid hybrids studied in unselected progenies on 2x and 4x levelcontinued their polygenic genetic background and significance of GCA and SCA effects. Study on ffcaresistance transferred on 4x level from 2x hybrids of S. chacoense and S. yungasense has been conducted since
1997. Agronomic performance of 2x and 4x highly resistant to Eca clones, selected in frame of this study, allowto introduce this source in the commercial breeding in near future.
References:
1. AUSTIN, S.; LOJKOWSKA, E.; EHLENFELD, M.K.; KELMAN, A.; HELGESON, J.P.: Fertile
interspecific somatic hybrids of Solanum: A novel source of resistance to Erwinia soft rot. Phytopathology78,1988,1216-1220
2. LEBECKA, R.; DOMANSKI, L.; ZIMNOCH-GUZOWSKA, E.: Combining ability of diploid potatoparents for tuber resistance to Erwinia cararovora subsp. atroseptica. Abstr. Joint EAPR/EUCARPIA
Breeding Section Meeting, 10-12 March 1997, Viterbo, Italy3, LEBECKA, R.; ZIMNOCH-GUZOWSKA, E.: Resistance to tubers soft rot (Erwinia caziotovora ssp.
alraveftica) in potato unseleted progenies of 4x-2x crosses. Abstr. EAPR Pathology Section Meeting,10-15‘ July 2001, Poznan
4. PEROMBELON, M.C.M.; KELMAN, A.: Ecology of the soft rot erwinias. Annu. Rev. Phytopathol. 18,1980, 361-367
5. ROUSSELLE — BOURGEOIS, F.; PRIOU, S.: Screening tuber - bearing Solanum spp. for resistance to softrot caused by Erwinia caratovora spp. atroseptica (van Hall) Dye. Potato Res. 38, 1995, 1 11-118
6. WOLTERS, P.I.; COLLINS, W.W.; Evaluation of diploid potato clones for resistance to tuber soft rot inducedby strains of Erwinia carotovora subsp. (itroseptica, E. carotovora subsp. carotovara and E. clujistinrhemi.Potato Res. 37, 1994, 143-149
.
_ I _7. ZIMNOCH-GUZOWSKA, E.; LOJKOWSKA, E.: Resistance to Erwinia spp. in diploid potato with a highstarch content. Potato Res. 36, 1993, 177-182
8. ZIMNOCH-GUZOWSKA, E.; LEBECKA, R.; PIETRAK, J.: Soft rot and blackleg reactions in diploidpotato hybrids inoculated with Erwinia spp. Amer. J. Potato Res. 76, 1999a, 199-207
9. ZIMNOCH-GUZOWSKA, E.; LEBECKA, R.; SOBKOWIAK, S.: An attempt to evaluate potato resistanceto Elwiltill carotovora ssp. arroseptica by inoculation of detached leaves. Plant Breed. Seed Science 43,1999b,101-112
l0. ZIMNOCH-GUZOWSKA, E.; MARCZEWSKI, W.; LEBECKA, R.; FLIS, B.; SCI-1AFER—PREGL, R.;SALAMINI, F.; GEBHARDT, C.: QTL analysis of new sources of resistance to Erwinia carorovora ssp.arroseprica in potato done by AFLP, RFLP and resistance-gene-like markers. Crop Science 40, 2000, 1156-1167
11. ZIMNOCH-GUZOWSKA, E.: Diploidalne mieszafxce miedzygatunkowe ziemniakajako z'rcid1o odpornoscina mokra zgnilizne bulw i czarna nozke w hodowli ziemniaka. Rozprawa habilitacyjna. Instytut Hodowli 1Akliinatyzacji Roslin. Monografie i Rozprawy Naukowe 8/2001, Radzikow 2001, pp 61

26
First results of resistance evaluation of Pelargonium t0 Xanthomonashortorum pv. pelargonii
Klaus Olbricht 1» Erika Griesbach 2
l Elsner pac® Jungpflanzen, Kipsdorfer Straß e 146, D—01279 Dresden, Germany2 Institut fiir Epidemiologie und Resistenz, Bundesanstalt fiir Zü chtungsforschung an Kulturpflanzen, Theodor-
Roemer-Weg 4, D-06449 Aschersleben, Germany
Abstract
/\'a/it/iomonas /iorrorum pv. pela/‘gonfi (Xhp) is causing the most important epidemic disease in current varietiesof Pelargonium-Zonale- and -Peltatum~Hybrids, the most important bedding plants worldwide. Isolates fromdifferent origins were collected and a system for evaluation of virulence and resistance was established. A groupofresistant species and hybrids are now available for breeding. Abilities of crossing with the hybrids of todayhas to be proved.
Introduction
The bacterium Xunr/iomonux /10l‘I‘ 07'lIm pv. pelurgonfi (Xhp) leads to leaf spot and stem rot (bacterial blight)showing wilting symptoms. The bacterium is specific to Pelargonimn and the highest danger of commercialPelargonium cultivation. That is why expensive cleaning procedures and the holding of certified motherstocksunder permanent testing are necessary for commercial growers. In order to detect resistance in the genusPelargonium and introduce it into the current hydrids, a cooperative project financially supported by theSä chsisches Staatsministerium fur Umwelt und Landwirtschaft began in 1996.
It was a first goal of this investigation to collect isolates of the bacterium of different origins and to select highlyvirulent strains for preparing a mixture of inoculum. Based on that a reliable test system for evaluation ofresistance was developed, Inheritance and crossability of the evaluated resistant species and hybrids are now
under consideration as well as the investigation ofreasons for resistance.
Material and methods
Isolates from different origins have been comparatively tested by inoculation of susceptible Pelargoniuni-Zonale-I-Iybrids in order to find those isolates with the highest degree of virulence. Because of this 5-6 virulentstrains were available for the evaluation of resistance. After cultivation on TS- or YDCa-agar for 48 h thecultures were suspended in water. The test plants (species and hybrids of Pe/urgmiiimi) were inoculated bydropping of 30ul suspension containing 102 cells per shoot for a first screen (applied on section of fresh clippedshoot apex). In following screens higher inoculum pressure of 104 and 10° cells per shoot was used for hightolerant or resistant types which were remained after the first screen. Only for special species with thin stems theinoculation method had to be changed. In such cases 3-5 leaves per shoot were cut by half with contaminatedscissors (ION cells per ml).
Inoculated plants have been cultivated under controlled conditions in an environment chamber with 26° C andartificial light during daytime and 22° C under darkness at night. Disease severity (ds) was rated 2, 3 and 4 weeksafter inoculation using a 1 to 9 scale (1 = symptomless, 2 = slight symptoms, 3 = about one third, 5 = abouthalf, 7 = about two thirds of the plant show symptoms, 9 = plant is dead).Extraction of pressure sap from different parts of inoculated plants were produced for the determination of Xhpcells in the plants (infection density) using dilution plating, plate-count technique and immunofluorescence test
(IF). The dilution plating and plate-count technique were done on TS-agar + 300 ppm streptomycin and theimmuriofluorescence test with MA SF8 (Rabenstein) and Cy 3 marked anti-mouse-conjugat.Results and discussion
Inoculation by dropping is the most reliable method for evaluation ofresistance of Pelargoniuni. In all cases it isnecessary to have frequent tests of virulence of different strains as a basis for reliable results of evaluation. Amixture ofseveral (5-6) virulent strains would suggest.
Current varieties of Pelargonium—Zonale- and -Peltatum-Ilybrids are very susceptible to Xhp. Within 7 daysafter inoculation leafspots appearcd followed by the typical symptoms ofwilting after about 2 weeks.
Scented-lcaved Pelargoniums (some are also commercial varieties) remained mainly symptomless or showedonly local spots of necrosis. Symptoms of wilting could never observed. The susceptibility of scented-leavedPelargoniums after high inoculum pressure can be graded into 3 classes:
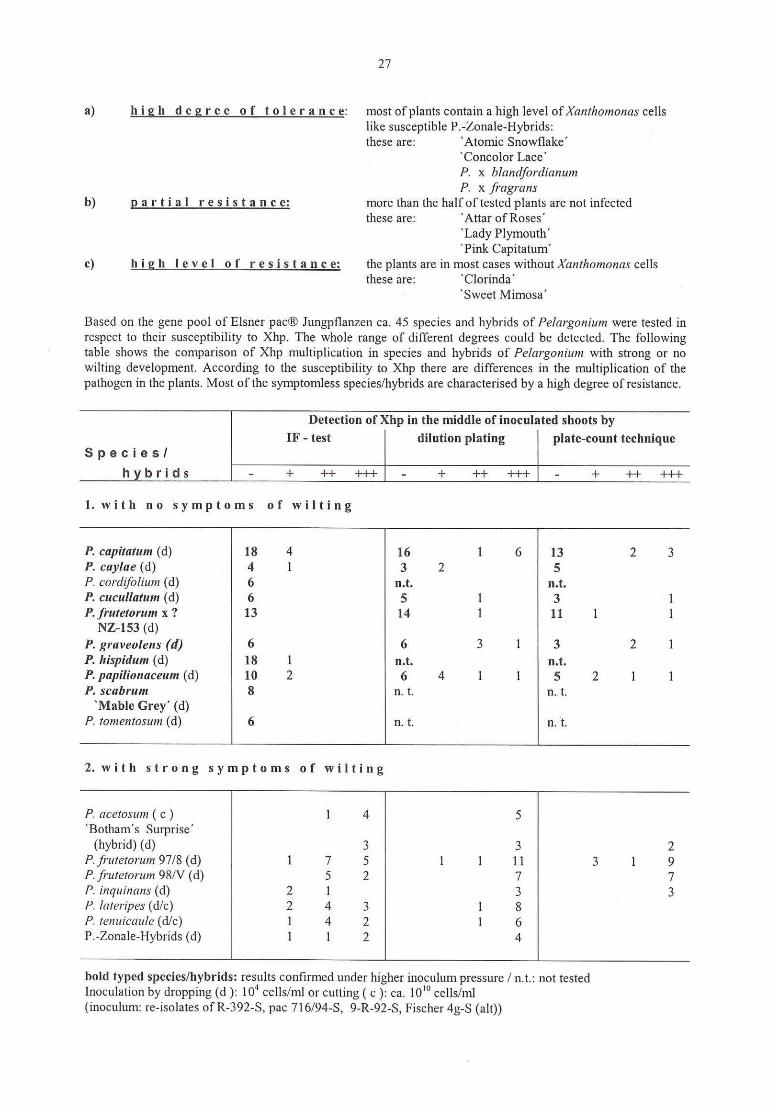
27
a) h i g h d e g r e e o f to l e r a n c e: most ofplants contain a high level ofXamh0mam1s cellslike susceptible P.-Zonale-Hybrids:these are: ‘ Atomic Snowflake’
‘ Concolor Lace’
P. X blannfordianunlP. X fragrans
b) E a r t i a I r e s i s t a n c e: more than the half of tested plants are not infectedthese are: ‘ Attar ofRoses'
‘ Lady Plymouth’‘ Pink Capitatum'
c) h i g h I e v e I o f r e s i s t a n c e: the plants are in most cases without Xanthomonas cellsthese are: ‘ Clorinda'
‘ Sweet Mimosa’
Based on the gene pool of Elsner pac® Jungpflanzen ca. 45 speeies and hybrids of Pe/argomtim were tested in
respect to their susceptibility to Xhp. The whole range of different degrees could be detected. The followingtable shows the comparison of Xhp multiplication in species and hybrids of Pelargariium with strong or no
wilting development. According to the susceptibility to Xhp there are differences in the multiplication of the
pathogen in the plants. Most of the symptomless species/hybrids are characterised by a high degree of resistance.
Detection of Xhp in the middle of inoculated shoots byIF - test dilution plating plate-count technique
S p e c i e s I
h b r i d s - + ++ +++
1. with no symptoms of wilting
P. capimtum (cl) 18 4 l6 l 6 13 2 3P. caylae (d) 4 l 3 2 5P. cordzjblium (d) 6 n.t. n.t.P. cucullarum (cl) 6 5 l 3 1P. frmetorum x ? 13 14 l l1 1 1
NZ—153 (d)P. graveolens (d) 6 6 3 1 3 2 lP. liispidum (d) l8 1 n.t. n.t.P. pupiliomiceum (d) 10 2 6 4 1 1 5 2 I 1P. scabrum 8 n. t. n. t.
‘ Mable Grey’ (d)P. romenrosum (cl) 6 n. t. n, t.
2. with strong symptoms of wilting
P. (Icefosum ( c ) l 4 5‘ Botham's Surprise’
(hybrid) (d) 3 3 2P. frufetorum 97/8 (d) l 7 5 l 1 ll 3 1 9P. fiwemruzn 98/V (d) 5 2 7 7P. irirjuinmrs (d) 2 1 3 3P. Ialeripes (d/c) 2 4 3 l 8P. rem/icuule (d/c) 1 4 2 1 6
P.-Zonale-Hybrids (d) 1 1 2 4
bold typed species/hybrids: results confirmed under higher inoculum pressure / n.t.: not testedInoculation by dropping (d )2 104 cells/ml or cutting ( c ): ca. 10'" cells/ml(inoculum: re-isolates of R-392-S, pac 716/94-S, 9-R-92-S, Fischer 4g-S (alt))

28
In comparison of the three methods used for determination of Xhp in the plants the two microbiological tests
(dilution plating and plate count technique) are more sensitive but have shown the same general results as theimmunofluorescence test (IF).
Evaluations of resistance to Xhp are done in order to get material for resistance breeding. Unfortunately most ofthe resistant types (species and hybrids) are not crossable based on chromosome numbers (see table). However,all crossable types are members of the group with the highest disease severities (not shown in the table).
Degrees of resistance and membership of the sections of the genusP e I a r g 0 n i u m
ß ds < 2,0 Section Crossabilit ß ds 2,1 - 3,0 Section Crossabilit
P. capitatum Pelargium - P. fulgidum Polyactium -
P. caylae Ciconium ? P. glutinosum Pelargium -
P. cardifalium Pelargium - P. grarzdzflorum Glaucophyllum -
P. cucullatum Pelargium - P. hispidum Pelargium -
P. frutetorum x ? Ciconium/7 ? P. tenuicaule 4n Ligularia ?P. graveolens Pelargium —
P. papilionaceum Pelargium -
P. scabrum Pelargium -
‘Mable Grey‘P. tomentosum Pelargium -
Legend: (D Disease severities (ds) from 5 — 6 Screening series; + crossable; - not crossable; '.‘ crossability has to be proved
NZ-153 is one of the chosen types for first steps of breeding because of the determined chromosome number of2n = 18 which corresponds to the chromosome number of Pelargonium-Zonale- and -Peltatum-Hybrids.A first proof of the way of inheritance of resistance by self-pollination showed that there are no differences indisease severity among the progeny in spite of a kind of phenotypical segregation in the progeny (leaf zonationpresent or absent).Further investigation should be focused on evaluation of resistance for other species and hybrids ofPelargonium, investigation of reason for resistance and the proof of crossability of detected resistantPelargonium—Zonale- and -Peltatum-Hybrids.
Supported by the Sä chsisches Staatsministerium fü r Umwelt und mndwirtschaft

29
Application of in vitro methods in tire blight resistance programsM. Hevesi l, E. Jambor-Benczur 2, J. Papp l,
J. Dobranszky 3, K. Magyar-Tébori 3, T. Buban 4
I Department of Fruit Science? Department of Floriculture and Dendrology,3 Research Centre of University ofDebrecen,4Research and Extension Centre for Fruit Growing, Ujfeherto
The first appearance of fire blight (caused by the bacterium Erwinia amylovortz Burill) in Hungary was reportedin 1996 in apple (Hevesi, 1996). Resistance breeding programs for apple and pear started in 1998 by usingseveral international pear and apple cultivars, traditional Hungarian apple cultivars, and breeeding material(hybrids). Experiments have been conducted in a controlled environment, since inoculation of this pathogen infield conditions is not possible due to stringent quarantine regulations. Therefore, evaluation of resistance ofapple and pear cultivars requires application of in vitro methods.
Plant material used in the experiments (leaves, flowers, fruits) was collected from trees of different cultivars in. orchards, seedlings were grown under greenhouse conditions from seeds. Inoculations were carried out either by
spraying with a bacterial suspension (flowers), stabbing (seedlings, fruits), or cutting back leaves (seedlings andmicropropagated plants (Norelli et al
, 1987; Murashige & Skoog, 1962)). Disc were cut from leaves and dippedinto a bacterial suspension (5xlOX cfu/ml). Development ofdisease was monitored continuously by disease ratingDR=(axn+bxn+...xn)/n) (Horsfall and Barrat, 1945; Hevesi et al, 2000), data were processed with the aid of
Statgraph computer software. The effect of several bioregulators (Prohexadion-Ca, Bion 50 WG+Kocide) ofinduced resistance on susceptibility of apple cultivars to fire blight was also tested in leaf discs andmicropropagated plants (Duron and Paulin, 1987).
Fully resistant pear and apple cultivars were not found among those tested (Fig. 1), However, severalinternational pear cultivars (l5—20% of those tested) displayed moderate resistance in our experiments.Furthermore, the first experiments on fire blight resistance of traditional Hungarian apple cultivars and new
breeding material also show promising results. Furthermore, acquired resistance to fire blight was effectivelyinduced in leaf discs of orchard-grown trees (Fig. 2) and rnicropropagated apple seedlings treated with
bioregulators (Prohexadion-Ca, (100g/ha), Bion 50 WG+Kocide) (Fig. 3).We have succesfully applied in vitro methods in screens for apple and pear cultivars resistant to fire blight.However, an important conclusion of our work is that screening for resistance to E. amylovora must be carriedout by the combined application of different inoculation and disease assessment methods.
Fig. 1: Susceptibility of apple cultivars to Erwinia amylovara - infection
Libertyn I
flElstar l*_
E gripiros_— i
[dared l-—Rewenfl
— — — -— — — ..-.
Royal Gala i’
Reafldfl -F101‘ iI|a" -
Freedom —“ m” wet
1,8 2,3 2,8 3,3 3,8Disease rating
.,,, Q30.ADril II2.Mav

30
Fig. 2: Effect ofboiregulators of induced resistance applied to apple trees by spraying in field conditions
2,5 — ---— -~— — ~—
2,077
2
1,51 372
'I I1
I I I I I I I I I i a _ Streptomycin0,5 i (control)
l I I I I I I I I i b _ Prohexadionvcao I
Frezdom Frezd-om Fragen: Jongijold Jangnold Jon:-gold 3:!-Itignfid Jgriggngld J:::ig":ld C_Bi0n.50 WG+Kocide
Fig. 3: Effect of bioregulator of induced resistance applied to micropropagated apple plants (added to culturemedia)
2,5
I. I I I I Is0 s1 s2 s3 s4 s5
Literature cited
1. DOBRANSZKY, J.; ABDUL—KADER, A.; MAGYAR-TABORI, K.; JAMBOR-BENCZUR,E.;BUB1-‘KN,T.; LAZANYI, J.: In vitro shoot multiplication of apple comparative response of three rootstock to
cytokinines and auxin. Int. J. of Hort. Sci. 6 (1), 2000, 36-392. DURON, M.; PAULIN, J.P.: Use of in vitro propagated plant material for rating fire blight susceptibility.
Acta Horticult, 217, 1987, 317-3243. HELLER, R.: Recherches sur la nutrition minerale das tissues vegetaux cultives in vitro. Ann. Sci. Nat. Bot.
Veg. 14, 1953, 1«2234. HEVESI, M.: Az Erwinia amylovora (Burill) Winslow et al. hazai megjelené se almä n. (Appearance of Fire
Blight on Apple in Hungary)’ Niivé nyvédelem 32 (5), 1996, 2252285. HEVESI, M,; PAPP‚ J.; JAMBOR-BENCZUR, E.; KASZANE, CS.K.; POZSGAI, I.; GAZDAG, G,Y‚ ;
BALLA, I.: Int. J. Hortic. Science 6 (4), 2000, 52-556. HORSFALL, J ,G.; BARRAT‚ R,W.: An improved grading system for measuring plant diseases.
Phytophatology 35, 1945, 6557. MURASHIGE, T.; SKOOG, F.: A revised medium for rapid growth and bioassays with tobacco tissue
cultures. Physiol. Plant. 15, 1962, 473-497
8. NORELLI, L.J.; ALDVINKLE, S.H.; BEER, V.S.: An assay for the virulance of Erwinia amylavoru usingMalus tissue culture. Acta Horticult. 217, 1987, 283

31
Evaluation of cassava genotypes, including individuals of the genome
mapping population, for resistance against bacterial blightZinsou, V." 2, Ahohuendo, B. 2, Jorge, Veronique 3, Verdier, Valerie 3, Wydra, Kerstin’
1 Institute ofPlant Diseases and Plant Protection, University ofHannover, Germany, 2 Plant Pathology Depmt.,Fac. of Agriculture, Univ. of Benin, Benin, 3 IRD, France, research group at Centre Intemacional de Agricultura
Tropical, Colombia
Introduction
Cassava is a basic component of the farming systems in most areas of Sub-Saharan Africa (Nweke et al., 1994),Africa produces 48 million tons of cassava roots annually on 7.4 million hectares providing more than 200
calories per day for 200 million people (Dorosh, 1988). The most widely cultivated varieties by farmers are local
ones (Nweke et al., 1994). The crop is attacked by cassava bacterial blight (CBB), caused by Xanthomonas
campestris pv. mtmihotis Bondar (Bondar 1915) (synonym Xanthomonas axonopodis pv. manihotis Vauterin et
al., 1995), one of the most important cassava diseases in Africa (Wydra and Msikita, 1998; Wydra and Verdier
2002). Among the proposed control measures (Wydra and Rudolph, 1999), host plant resistance is one of the
most suitable measures for farmers. Information on the reaction of local cassava varieties in Benin to CBB is not
available. Genetics of resistance to CBB in cassava and genes governing resistance were investigated byinoculation with Colombian and one African strain ofX. campestris pv. manihotis (Jorge et al., 2000; Jorge et
al„ 2001), but analysis had up to now not included more African strains. Therefore, the objectives of the present
studies were (i) to test local genotypes from Benin for their reaction to CBB, (ii) to identify possible pathotypesof X. campestrir pv. manihotis, (iii) to analyse the reaction of individuals of the cassava genome mappingpopulation to inoculation with African strains, and, thus, (iii) to contribute to the identification of CEB-
resistance related markers or involved genes.
Materials and Methods
Screening ofcassava genotyperfor resistance to cassava bacterial blight in different ecozones ofBenin
The studies were conducted in three field sites located in 3 agro—ecological zones, the forest—savanna transition
(lntemational Institute of Tropical Agriculture, IlTA, Cotonou, South Benin), the wet savanna (Save, INRAB
station, Centre Benin), and the dry savanna zones (Ina, INRAB station, North Benin), in the years 1998-1999
and repeated in 2000-2001. An augmented complete randomized block design (RCBD) with 3 blocks and 13
genotypes per block (a total of 33 genotypes was tested, as listed under “ Results” ) was used. The highly vimlent
strains of X. campestris pv. manihotis GSPB 2506, GSPB 2510, Save 10, isolated in Cotonou, Ina and Save,respectively, Benin, were used for inoculation of a suspension of 107 cfu/ml by spraying the lower surface of
cassava leaves. For disease assessment, the number of leaves with spots, blight or the number of wilted leaves
and the number of shoot tips with die—back were recorded on ten plants selected randomly per plot one month
after the first spraying and at a monthly interval over the dry season until the plants had reached 12 months. Each
symptom was evaluated according to a disease index (Wydra et al., 1999). The severity index was calculated
using the weighted mean of the percentage of symptom types. The mean of severity index of each genotype at
each evaluation date was calculated and the area under disease progress curve (AUDPC) was calculated usingthe cumulative disease severity index. The calculus method of integration of area under a curve was used to
compute the AUDPC (Genstat program, 1993). Classes of susceptible (S), medium resistant (MR) and resistant
(R) genotypes were formed using the Genstat program in cases of significant differences (e.g. when differences
between the check genotypes were significant), or were estimated on basis ofthe reaction of the genotypes,
Identification ofgenetic markers testing individuals ofthe cassava mapping population
Hundred-eleven genotypes (mapping population) deriving from a backcross of F1 cross (TMS30572 x
CM2177—2) were grown in a glasshouse at IITA. Three highly virulent strains GSPB2506, GSPB2507 and
GSPB25l1 isolated in Cotonou (Benin), Ibadan and Onne (Nigeria), respectively, were used. The fourth strain,Uganda 12, was isolated in Uganda. All the 4 strains were used for stem inoculation, while GSPB2506,GSPB25l1 and Uganda 12 were also inoculated by leaf infiltration. Stem-inoculated plants were evaluated every5 days over 4 weeks using a 5 point scale of symptom grades: 0 = no symptom, 1 = 1 leaf wilted, 2 = 2-4 leaves
wilted, 3 = more than 4 leaves wilted, 4 = dieback of plant or shoot. The AUDPC was calculated for each
inoculated plant and used to compare the resistance of each plant genotype to the 4 strains and the virulence of
strains on the cassava genotype. Inoculation by leaf infiltration of the eight available genotypes identified as
resistant in the previous stem-puncture test was carried out with suspensions ofX. campestrir pv. mani/mris (100cfu/ml). The area of spots (water-soaked and necrotic spots) was measured each third day and used to calculate
the AUDPC.

32
The association between molecular markers and plant response to X campestris pv. manihotis was determinedby the QTL cartographer program (Basten et al. 1994, 1999). A simple linear regression and interval mappingwere performed. A significant association between DNA marker and CBB resistance was declared if the
probability was equal to or less than 0.05 and the likelihood ratio (LR) statistic was more or equal to 6 in order to
minimize the detection of false positives. The amount of phenotypic variance explained by each marker was
obtained from the regression coefficient.
Results and discussion
Screening ofcassava genotypesfor resistance to cassava bacterial blight in different eco-zones in Benin
The 33 cassava genotypes showed strong genotype x environment interactions with high variations in theirreaction to CBB in the different ecozones. None of the genotypes was found to be resistant in all theenvironments (ecozones, years). Genotypes revealing resistance in one ecozone in the years 1998 and 2000, suchas DANGBO2, RB92l32, BEN860l6, RB92164 (in 1998 and 2000), RB92131, TMS30572 (in 1998 and 2000),CAP92034 in the forest savanna transition zone, RB92033, BEN86018, TMS30572 (in 1998), RB92174,RB92164 (in 2000), RB92l51, CAP92034 in the wet savanna zone and RB92052, CAP94049, BEN86004,RB92132, CAP92034 in the dry savanna zone can only be recommended to farmers in the respective ecozone.
Genotypes RB92164 and TMS30572 showed resistance in two ecozones in yearl998 and 2000, respectively,and CAP92034 revealed resistance in 3 ecozones in year 2000, Medium resistant genotypes were TME1,RB92l51, RB92099, RB92022, BEN86040, RB92l25 in year 1998, while genotypes RB89608, RB92l03,BEN86052 in year 1998 and RB92l82, RB89509, TMEI BEN86052 in year 2000 were classified as susceptiblein all ecozones. Also Fanou (1999) and Otoo et al. (1994) observed high genotype x environment interactionswith other cassava genotypes planted in different ecozones. They concluded that some cultivars are adapted to
specific local environments.
Pathogenic variability, and differential reaction ofgenotypes ofthe mapping population
The AUDPC values obtained after inoculation with four X. campestris pv. manihotis strains revealed variationsnot only between the cassava genotypes deriving from a F1 backcross, but also strain x genotype interactions.Genotypes which could be useful for pathotype identification were differentiated between the 111 genotypes byselecting reverse differential (AUDPC values differ by 20 in the reverse direction ofX. campestris pv. manihotisstrains’ virulence:GSPB2507 > GSPB25l1 > Uganda 12 > GSPB2506) and striking differential reactions(AUDPC values differ by 26) between the strains. Thus, groups of resistant (AUDPC 0-20), medium resistant(AUDPC 20-40) and susceptible (AUDPC 40-60) genotypes were formed, and the strains were classified into 23
groups of differential genotypes according to the reaction to the four strains (Table 1). Genotype groups 4, 7, 9,10, ll, 12, 15, I9 and 21, representing one or more individual genotypes, reacted as resistant and susceptible to
at least one of the strains, respectively. Similar observations were made after leaf-infiltrating strains GSPB 2506,25 l1 and Uganda 12 into ten genotypes identified as resistant to stem-inoculation, revealing 7 different reactions(data not shown). Genotype Ben86052 showed high AUDPC to leaf-inoculation of GSPB 2506, 25011 andUganda 11 (AUDPC 41926, 33261, 8800, respectively), while 3 genotypes showed strong reverse reactions toinoculation of the same three strains (genotype CM-888877-ll: AUDPC 25944, 5314, 8904, respectively;genotype CM8873-64: AUDPC 5140, 6162, 15699, respectively; genotype TMS30572: AUDPC 2530, 2601 and7048, respectively). Genotypes Cm8820—40, followed by Cm8873-69 and Cm8820-30, were resistant to stem-and leaf-inoculation with the four strains.
Strains with higher virulence when stem-inoculated (GSPB 2507, 2511) or leaf-inoculated (2506) were observed.Thus, the results indicate a differentiation of African X. eampestris pv. mariihotis strains into pathotypes, andthat different mechanisms may be involved in the resistance to the pathotypes, and on leaf and stem levels.Pathotypes (Restrepo et al., 2000) and haplotypes of X. campestris pv. manihotis (Restrepo et al., 1998, 1999,2000) were also described for Latin American strains.
The genetic map based on marker segregation RFLP (GY) and rnicrosatellites (SSRY) in parent gametes TMS30572 ( ) and CM 7857-4 ( ) of BC1 populations CM8820+CM8820B revealed linkage groups 2, 7 and 11associated to the female mapping population CM8820, while the linkage group 1 was associated to the maleCM882O (Table 2). The female mapping population CM8873 showed group 14 linked and also non-linkedgroups, while with the male only non-linked groups occurred. Different markers were detected corresponding tothe groups. Groups 2 and 11 of the female of the mapping population are known groups involved in resistance toCBB (Jorge, 2000), Jorge (2000) only used Latin American and one African strains for her analysis. Thus,regions of the cassava genome involved in resistance to X. eampestris pv. maniholis differed according to theinoculated strain, and new markers specific to African strains were identified.
The newly identified markers for CBB resistance can be used to increase the efficiency of selecting resistantgenotypes in Africa by selecting for particular alleles from the linkage groups.
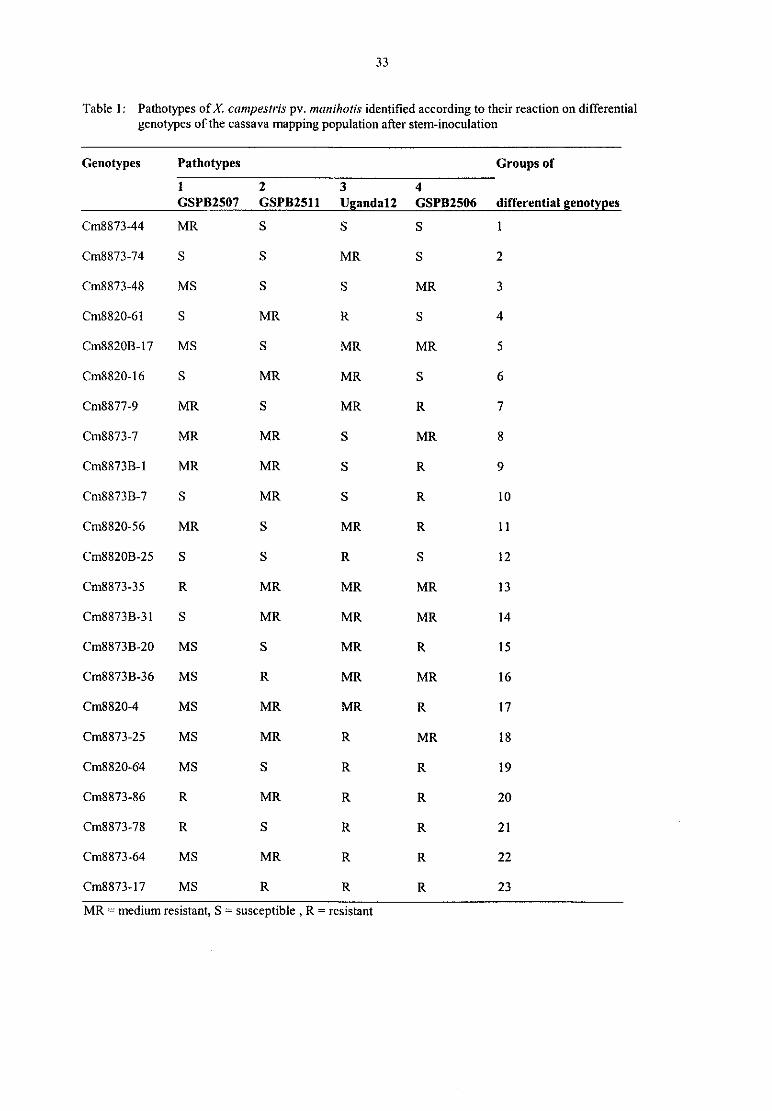
33
Table 1: Pathotypes ofX crzmpestris pv. marzihotis identified according to their reaction on differential
genotypes of the cassava mapping population after stem-inoculation
Genotypes Pathotypes Groups of
1S2_?—‘ T“ S4— “GSPB2507 GSPB2511 Ugandan GSPB2506 differential genotmes
Cm8873-44 MR S S S 1
Cm8873—74 S S MR S 2
Cm8873—48 MS S S MR 3
Cm8820-61 S MR R S 4
Cm8820B—17 MS S MR MR 5
Cm8820-16 S MR MR S 6
Cm8877-9 MR S MR R 7
Cm8873-7 MR MR S MR 8
Cm8873B-1 MR MR S R 9
Cm8873B—7 S MR S R 10
Cm8820-56 MR S MR R 11
Cm882OB-25 S S R S 12
Cm8873-35 R MR MR MR 13
Cm8873B—3 l S MR MR MR 14
Cm8873B-20 MS S MR R 15
Cm8873B»36 MS R MR MR 16
Cm8820-4 MS MR MR R 17
Cm8873—25 MS MR R MR 18
Cm8820-64 MS S R R 19
Cm8873-86 R MR R R 20
Cm8873-78 R S R R 21
Cm8873<64 MS MR R R 22
Cm8873—l7 MS R R R 23

34
Table 2: Markers showing most significant likelihood ratio (LR) values (threshold = 6) after simple linearregression for each map (female- and male- derived) and for each strain inoculated
Mapping Map Strain Linkage Marker LR Significance % variance
population group explainedCM8820 Female GSPB2507 11 SSRY6 7.9 0.010 16
GSPB2511 11 SSRY6 6.7 0.011 18
SSRY170 6.5 0.012 19.7
Uganda 12 2 SSRY55 7.4 0.027 18.17 SSRY93 6.7 0.011 23.2
Male GSPB2507 1 GY12 6.1 0.015 21,8
Uganda 12 1 GY12 6.0 0.015 33.3not linked SSRY17 6.7 0.011 ND
Table 2, ctd.
CM8873 Female GSPB2507 not linked SSRY104 6.1 0.015 ND
GSPB25l1 14 SSRY83 15.4 0.000 19.6
Uganda 12 not linked SSRY104 7.6 0.007 ND
GSPB2506 14 SSRY83 9.5 0.002 18.2not linked SSRY104 8,0 0.005 ND
Male GSPB2507 not linked SSRY7 8,2 0.005 ND
GSPB2511 not linked SSRY84 8.6 0.004 ND
GSPB2506 not linked SSRY157 7.9 0.005 ND
ND = not deterrnined
References
l. BASTEN, C.J.; WEIR, B.S.; ZENG, Z-B.: Zmap-a QTL cartographer. In: Smith, C; Gavora, J.S,; Benkel,BL; Chesnais, ‚I.; Fairfull, W.; Gibson, J.P.; Bumside, E.B. (Eds.): 5th Congress on Genetics Applied toLivestock Production: Computing Strategies and Software, Guelph, Canada, 1994, 65-66
2. BASTEN, C.J.; WEIR, B.S.; ZENG, Z-B.: QTL Cartographer. Version 1.13, Department of Statistics,Raleigh, NC, USA, 1999
3. BONDAR, G,; Molestia bacteriana da mandioca. Boletim de Agricultura. Sao Paulo 16a, 1915, 513-5244. DOROSH, ‘P.: The economics of root and tuber crops in Africa. RCMP Research Monograph No. 1, 1988,
IITA Ibadan, Nigeria5. FANOU, A.: Epidemiological and ecological investigations on cassava bacterial blight and development of
integrated methods for its control in Africa. PhD thesis. University of Gö ttingen, Gennany, 1999, 199 p6. GENSTAT. GENSTAT 5 Release 3 Reference Manual, Oxford Science Publications, Oxford University
Press Inc., New York, 1993, 796 p7. JORGE, V.: Cartographie de la resistance du manioc a 1a bacteriose vasculaire du manioc causee par
Xanthomanas axonopodis pv. manihotis, PhD thesis, 2000, 112 p8. JORGE, V.; FREGENE, M.; DUQUE, M.C.; BONIERBALE, M.W.; TOHME, J.; VERDIER, V.: Genetic
mapping of resistance to bacterial blight disease in cassava (Mzmihot esculenta Crantz). Theor. Appl, Genet.101, 2000, 865-872
9. JORGE, V.; FREGENE, M.; VELEZ, C.M.; DUQUE, M.C.; TOHME, J.; VERDIER, V.: QTL analysis offield resistance to Xanthomonas axonoporlis pv. mlmi/mris in cassava. Theor. Appl. Genet. 102, 2001, 564-571
10. NWEKE, F.I.; DIXON, A.G.O.; ASIEDU, R.; FOLAYAN, S.A.: Attributes of cassava varieties desired byfarmers in sub-Saharan Africa. Proc. 5"’ Symp. lSTRC-AB, 1994, 65-92

35
11. OTOO, J,A.; DIXON, A. G,0.; ASIEDU, R.; OKEKE, J.E.; MAROYA, G.N.; TOUGNON, K.; OKOLI,0,0.; TETTEH, J.P,; HAHN, S.K.: Genotype x environment interaction studies with cassava. Acta. Hort.
(ISHS). 380, 1994, 146-14812. RESTREPO, C.; VELEZ, C.M.; VERDIER, V.: Measuring the genetic diversity of Xanthomonas
trxonapodis pv. mtmihotis within different fields in Colombia. Phytopathology 90, 2000, 683-69013. RESTREPO, S.; DUQUE, M.C.; VERDIER, V.: Characterization of pathotypes among isolates of
Xanthomonas nxariopodis pv. mzmihotis in Colombia. Plant Pathology 49, 2000, 680-68714, RETREPO, S.; VALLE, T.; DUQUE, M.C.; VERDIER, V.: Assessing genetic variability among Brazilian
isolates ofX(mthomam1s axonopadis pv. manihotis in Colombia. Appl. Environ. Microbiol. 63, 1999, 4427-4434
15. VAUTERIN, L.; HOSTE, B.; KERSTERS, K.; SWINGS, J.: Reclassification ofXantham0nas. Int. J. Syst.Bacteriol. 45, 1995, 472-489
16. VERDIER, V.M.; RESTREPO, S.; MOSQUERA, G,; DUQUE, M.C.; GERSTL‚ A.; LABERRY, R.:Genetic and pathogenic variation ofXantham0mzr axonopodis pv. manilmtis in Venezuela. Plant Pathol. 47,1998, 601-608
17. WYDRA, K,; MSIKITA, W.: Ovewiew of the present situation of cassava diseases in West Africa. In:
Akoroda, M.O,; Ekanayake, I.J. (Eds.): Root Crops Poverty Alleviation. Proc. 6"‘ Trienn. Symp. Inter. Soc.
Trop. Root Crops Africa Branch (ISTRC-AB), Lilongwe, Malawi, 22-28 October 1995. ISTRC‚ IITA &Govemment ofMalawi, 1998, 198-206
18. WYDRA, K.; RUDOLPH, K.: Development and implementation of integrated control methods for majordiseases of cassava and cowpea in West-Africa. Gtittinger Beitr. Land- und Forstwirtsch. in den Tropen und
Subtropen, 133, 1999, 174-180‚
19. WYDRA, K.; VERDIER, V.: Occurrence of cassava diseases in relation to environmental, agronomic andplant characteristics. Agr. Ecosyst. Environ. 93 (1-3), 2002, 211-226
20. WYDRA, K.; ZINSOU, V.; FANOU, A.: The expression of resistance against Xanthomonas campeszris pv.manihotis, incitant of cassava bacterial blight, in a resistant variety compared to a susceptible variety. In:
Mahadevan, A, (Ed.): Plant Pathogenic Bacteria. Proc. IX Int. Conf, Madras, India, 1999, 583-592

36
The concept of resistance, tolerance and latency in bacterial diseases:examples from cassava and cowpea
Kerstin WydraInstitute ofPlant Diseases and Plant Protection, University of Hannover, Germany
Introduction
Cassava and cowpea are important staple crops in tropical areas and provide the main carbohydrate and proteinsupplies, respectively, for the poor population in many developing countries. Both crops are attacked bynumerous pests and diseases, with bacterial blights caused by Xanthomonas campestris pv. manihotis andX campestris pv. vignicola, respectively, being major constraints to production (Wydra and Msikita 1998,Sikirou 1999, Wydra and Verdier 2002). Therefore, integrated control systems are being developed for cassava
and cowpea bacterial blight (Wydra and Rudolph 1999, Zinsou 2001). Among various recommended methodssuch as intercropping, sanitary measures and use of clean planting rnaterial/seeds, host plant resistance is themost suitable and preferred method of control (Fanou 1999, Sikirou et al. 2001, Zinsou et al. 2002).Resistance to bacterial blight in cassava and cowpea is polygenic (horizontal, quantitative, partial) (Patel 1987,Kpémoua 1995, Jorge et al. 2000). Although horizontal resistance is the searched trait due to its more durableprotection compared to vertical resistance, it is affected considerably by changes in environmental conditions(Johnson 1984, Agrios 1997). The level of horizontal resistance expression is related to infection pressure whichin turn is influenced by the microclimate. Additionally, varieties with different levels of horizontal resistance are
often difficult to distinguish. These characteristics make a standardized and accurate evaluation and selection ofresistant genotypes difficult. Horizontal resistance is based on many host physiological processes, that contributeto the defense, and which due to their number and often general effects can hardly be overcome by a pathogenstrain. Races were up to now not described for X campestris pv. manihotis nor for X. campestris pv. vignicola(Khatri-Chhetri et a1. 1998, Wydra et al. 1998). Nevertheless, screening for horizontal resistance requires theinoculation with a diverse set of pathogen genotypes (Hammerschmidt 2000) and the determination of degreeand rate of infection. Long-term studies have shown that the ranking of individual varieties generally remainsrelatively constant despite this variability (Walther 1988). Newer concepts of host plant resistance understandhorizontal resistance as a combined effect of vertical resistances.
Although described as resistant, many crop varieties may still harbour the pathogen (Liao et al. 1998). Thislatency — symptomless infection (inapparency) (Bos and Parleviet 1995) -, in which a pathogen is present or
capable of living or developing in a host without producing visible symptoms of disease may be of majorepidemiological importance and contribute to the dissemination of pathogens. Additionally, latent infections maycause reductions in plant growth and yield, as demonstrated e.g. for latent infection with Ralstonia solanacearum(Allen 1998, Liao et al. 1998).Besides resistance, tolerance has proved to be a valuable trait under field conditions. Tolerance is defined as “ thegenetically based ability of a host plant to recover from the attack of a pest and/or from infection by a diseasewithout any or with low reduction of fitness and yield" (Walther et al. 1999). This may be due to specificheritable characteristics of the host plant such as lack of receptor sites for or inactivation of the irritant excretionsor compensation for damage caused by the pathogen (Agrios 1997).
Presently, methods for evaluating the durability of cassava and cowpea resistance include testing at a number ofepidemiologically dissimilar locations and exposing putatively resistant lines to high natural infection pressureacross a number of environments. But, a homogeneous, sufficiently high infection to show differences insymptom expression is not always assured. Effects of tolerance and latency are not taken into account, andbreeding efforts may result in genotypes with high resistance to symptoms, but high yield effect of a remaininglow infection, and/or in selecting plants with high latent infection. To date, varieties with a stable resistance havenot been identified for cassava (Fanou 1999), while some cowpea genotypes revealed multilocational resistance(Sikirou 1999).
The present paper aims at demonstrating the need to critically evaluate and adapt the evaluation of genotypes forresistance to the specific crop and disease, taking into account the effects of tolerance and latency in interactionwith the environment. Background data on effective and suitable methods of selection of cassava and cowpeagenotypes for durable resistance will be provided and contribute to the development of a standardized evaluationsystem for resistance, including effects of tolerance and latency, to bacterial diseases.

37
Materials and Methods
Cassava: Experimental sites for cassava yield loss trials were located in 5 ecozones in Benin and Nigeria: humidforest (Onne, South-East Nigeria), coastal forest savanna transition (Cotonou, South Benin), inland forest
savanna transition (Ibadan, South Nigeria), dry savanna (Ina, North Benin) and Sudan savanna zones (Kano,North Nigeria). The susceptible cultivars Ben 86052 in Benin and Antiota (TME1) in Nigeria, and the ‘ resistant’TMS30572 were used. Highly virulent strains of X. canzpestris pv. manihotis isolated from the locations of theexperimental fields served as inoculum (103 cfu/ml). Symptom data were collected each month during the rainyseason. Fields were harvested each month of the rainy seasons up to 12 months. At each harvest, 5 to 10 plants
were uprooted per plot, but not more than 20% of the total number plants per plot were harvested and root freshand dry weight were recorded. Data of the last harvest in four ecozones are presented.
Cowpea: Experiments were conducted in 1996 and 1997 in the forest savanna transition (FST) zone and in the
dry savanna (DS) zone. Six genotypes (Gtl = IT84E-124, Gt2 = IT84S-2246-4, Gt3 = IT86D—7l5, Gt4 = IT8lD-
1137, Gt5 = IT81D-1228-14, Gt6 : IT86D-719) were tested. At the age of3 weeks, plants were inoculated byspraying X. campestris pv. vignicola strains isolated from the area of the field trials with an inoculumconcentration of 103 cfu/ml. Symptom development was recorded every two weeks starting 0 dpi on 20 plants,selected at random per plot. Seed weight was recorded 10-12 weeks after planting on 10 plants per plot,
Bath craps: Experimental design was a completely randomized, nested arrangement with 4 repetitions per plot
with a non-inoculated and an inoculated variant. Both, inoculated and non-inoculated fields were also naturallyinfected with bacterial blight. For symptom evaluation, the percentage of leaves with spot (S), blight (B), or wilt
symptom (W), including dropped leaves, and the percentage of shoots with dieback (D) were determined.
Severity index was calculated according to the following formula: (1 x S + 2 x B + 3 x W + 4 x D)/10. Statistical
analysis was performed using SAS statistics (Statistical Analysis System (SAS) Institute Inc., 1992). Analysis of
variance was conducted for harvest data comparing inoculated and non-inoculated plots. Area under diseaseprogression curve (AUDPC) was calculated in the SAS program. Further details on materials and methods are
provided in Fanou (1999) and Sikirou (1999).
Results and Discussion
Carsava bacterial blight
Severity of bacterial blight symptoms expressed as area under the disease progress curve (AUDPC) and root
yield loss of two cassava varieties (Ben86052 — susceptible -, TMS30572 — resistant) were compared in naturallyinfected and artificially inoculated plots in four ecozones of West Africa. The resistant variety showed alwaysless symptoms than the susceptible variety. Nevertheless, when increasing the infection pressure by inoculation,the resistant variety also developed higher symptom levels. Although both varieties showed considerable root
yield losses, only for variety TMS30572 in the coastal forest savanna transition zone and for both varieties in the
dry savanna zone the difference in AUDPC between naturally infected and inoculated plots resulted in a
significant root yield loss (Fig. 1). Thus, under favourable growing conditions (e.g. forest savanna zone), theplants seem to compensate the damage caused by the disease, e.g. by additional lateral shoot formation (ownobservations), while under harsh conditions, the plants suffer significant losses (dry savanna zone).

38
Fig. 1: Disease severity (AUDPC of severity index) (A) and root yield (root fresh weight per plant) (B) ofcassava variety Ben86052 (susceptible) and TMS30572 (‘ resistant’ ) in four ecozones of West Africa
A '5“.:.,..
N".
E’ m’ M|3] en mi ml
: 42 „ E..w
E so 2 51 l9 IIJTMS innc.
S 4a 1 35:51< =:=,=;= 5:555: 2252;: n;=;:
o :---:i- E=i:E:5 =E=E.E E‘ 5:£'.
4501|
;- 4000
B E 3500 ‘ O76
E‘ 1000 '.:.:s iii2 :-: 1342
e. :33: 49 :53 * :s=s
g « ooo :5: ..„ ;:;: M»: *
== m 2:1 =E=E0 as:: 2:22 ß
Humid Forest Forest Forest Dry Savanna
Savanna cst. smnnn
* = significantly different from naturally infected plots at p : 0.05; ("‘ ) = significant only for root dry weight; numbersindicate difference between naturally infected and inoculated plots; data from Sudan savanna which showed a non-significantyield increase afler artificial infection compared to natural infection not presented
A tolerance effect could be clearly observed with variety Ben86052, which indeed had generally an overallhigher yield potential, but also showed less yield losses when disease severity increased: an increase of AUDPCby 51 for Ben86052 and of 28 for TMS30572 caused a non-significant yield loss for Ben86052, but a significantloss of 1342 g/plant for TMS30572 (coastal forest savanna transition zone); an increase of AUDPC by 64 forBen86052 compared to an increase of 24 in TMS30572 resulted in a Similar loss for both Varieties in the humidforest zone; Similar observations were made in the forest savanna zone. Nevertheless, it has to be considered,that an increase in AUDPC occurring at higher initial levels may have another effect on yield than an increase atlower disease levels. Further data from trials over three years in two locations also revealed a high variability ofAUDPC and root yield (Wydra et al. 2001). These observations indicate a high influence of the environment on
disease severity as well as on root growth (genotype X enviromnent interaction)‘

39
Fig. 2: Incidence ofblighted leaves (%) of cassava variety Ben860S2 (susceptible) (A) and TMS30572
(resistant) (B) in five ecozones of West Africa over the growing period of 12 months (except drysavanna: 18 months)
(Ecozones: rain forest, coastal (c) forest savanna (FS) transition, forest savanna transition, dry savanna, Sudan savanna)
A9"
l rs...„ .m„ n<ci F namrawan
ä 7° S sun” savanna
f; so
i so’
:
...-~
z“
x,ä an .v
Ä ' jg 2a . . /
:
a L x i— .‘NL’ ! 54511:u— —
'
0 1 2 3 A 5 E 7 B 9 |0 Il l2 I3 |4 |5 Hi 17 15 19 20
Months alter planting
B so
ED
E 7o
ä 50
g ...W
2o_ \ _
Q}.‚
1o '
‚ x ‚
O 1 2 3 4 5 E 7 E 9 |0 |1 l2 I3 |4 I5 |8 |7 1B 19 2D
Monlhs alter planlinq
Analysing the development of symptom types in the two varieties, a similar incidence of leaves with spots was
observed over the growing period (data not shown), while the resistant Variety showed a clear restriction inincidence of leaves with blight symptoms in all the ecozones (Fig. 2). These observations were continued by a
significantly lower number of bacteria in TMS30572 than in Ben86052 (Zinsou 2001). But, although theresistance to symptoms in TMS30572 is higher, the tolerance is not sufficient. Stepwise regression of spot,blight, wilt and dieback symptoms against root weight revealed that among the symptom types, the blightsymptom had the greatest effect on root weight (p-value: combined analysis of both varieties 0,03, analysis forresistant variety 0.01).
Comparing 23 genotypes in naturally infected and artificially inoculated plots in four environments (3 ecozones),Fanou (1999) observed root yield losses of up to 76.8% in susceptible genotypes in thedry savanna zone underfavourable conditions for the disease. Among the tested genotypes, Ben86052 showed high symptomdevelopment, but nevertheless a high yield, and could therefore be classified as tolerant. Thus, some cassava
varieties are able to compensate the negative effect of the disease under favourable growth conditions. Therefore,a symptom threshold for yield loss cannot be determined and loss remains unpredictable.
Latent infection of cassava stems which are used as propagating material was observed for the highly susceptiblevariety Ben86052, while the pathogen could not be found in symptomless stems of the more resistant varietyTMS30572 (Tab. 1). Thus, for selection of planting material from a susceptible variety from infected fields, a
visual inspection ofplants is not sufficient.
Further studies suggested the existence ofpathotypes among African X. campestris pv. manihotis strains (Zinsouet al. 2002, this volume). Therefore, in selection of genotypes, tolerance in terms of low disease effect on root
yield, latent infection and pathogenic variability should be considered as important characteristics. Symptomevaluation alone is not sufficient.
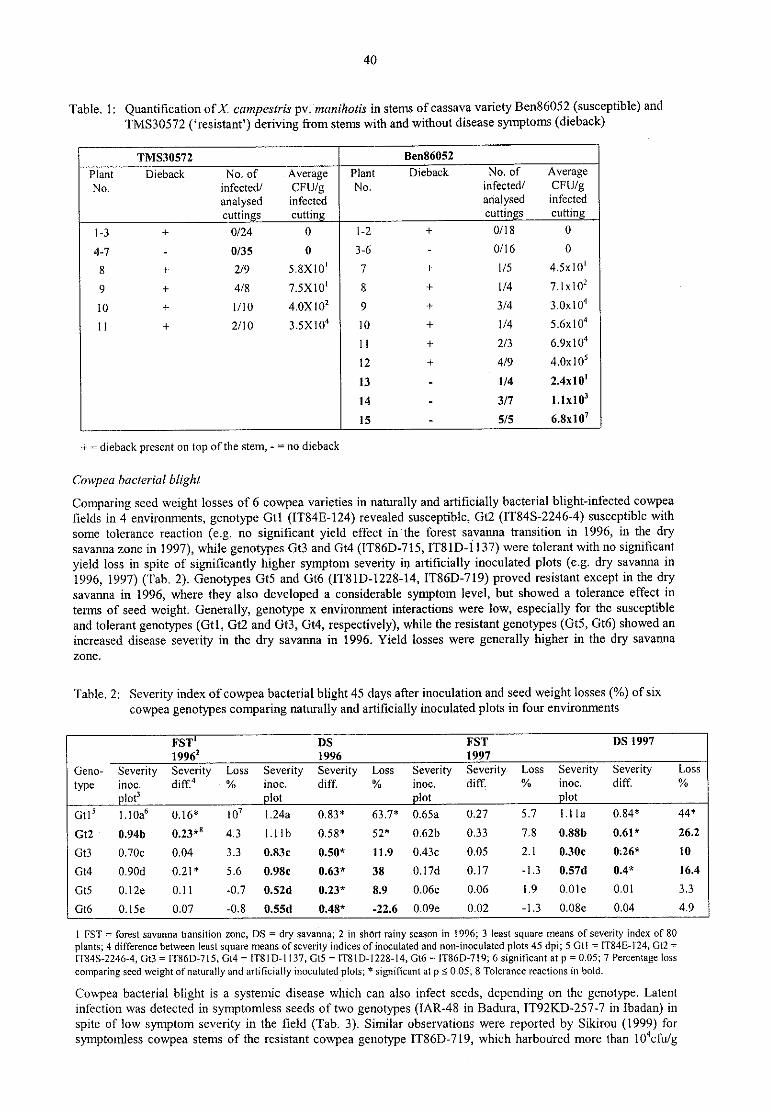
40
Table. 1: Quantification ofX. campestris pv. manihotis in stems of cassava variety Ben86052 (susceptible) and
TMS30572 (‘ resistant’ ) deriving from stems with and without disease symptoms (dieback)
TMS30572 Ben86052
Plant Dieback No. of Average Plant Dieback No. of AverageN0. infectedl CFU/g No. infectedl CFU/g
analysed infected analysed infected
cuttins cuttin cuttins cuttingl-3 + 0/24 0 1-2 + 0/ l8 0
4-7 - 0/35 0 3-6 - 0/16 0
s + 2/9 5.8Xl0' 7 + 1/5 45x10‘
9 + 4/8 7.sx1o' x + 1/4 71x10’
10 + 1/10 40x102 9 + 3/4 30x10"
11 + 2/10 35x10“ 10 + 1/4 56x10“
11 + 2/3 60x10"
12 + 4/9 40x10’
13 - 1/4 2.4x10‘
14 - 3/7 11x10’
1s - 5/5 6.811107
+ 2 dieback present on top of the stem, — = no dieback
Cowpea bacterial blight
Comparing seed weight losses of 6 cowpea varieties in naturally and artificially bacterial blight—infected cowpeafields in 4 environments, genotype Gtl (IT84E-124) revealed susceptible, Gt2 (1T84S-2246-4) susceptible with
some tolerance reaction (e.g. no significant yield effect in’ the forest savanna transition in 1996, in the drysavanna zone in 1997), while genotypes Gt3 and Gt4 (IT86D-715‚ IT81D-1137) were tolerant with n0 significantyield loss in spite of significantly higher symptom severity in artificially inoculated plots (e.g. dry savanna in
1996, 1997) (Tab. 2). Genotypes Gt5 and Gt6 (IT81D-1228-14, IT86D-719) proved resistant except in the drysavanna in 1996, where they also developed a considerable symptom level, but showed a tolerance effect in
terms of seed weight. Generally, genotype x environment interactions were low, especially for the susceptible
and tolerant genotypes (Gtl, Gt2 and Gt3, Gt4, respectively), While the resistant genotypes (Gt5, Gt6) showed an
increased disease severity in the dry savanna in 1996. Yield losses were generally higher in the dry savanna
zone.
Table. 2: Severity index ofcowpea bacterial blight 45 days after inoculation and seed weight losses (%) of six
cowpea genotypes comparing naturally and artificially inoculated plots in four environments
rsr‘ ns rsr 0s 19971996’ 1996 1997
Geno- Severity Severity Loss Severity Severity Loss Severity Severity Loss Severity Severity Loss
type inoc. diff.‘ - % inoc. diff. % inoc. diffi % inoc. diff. %1013 lot lot lot
G115 110a‘ 0.16* 107 1.2421 0.83* 63.7* 065a 0.27 5.7 1.1 la 0.84* 44*
Gt2 0.9413 013*“ 4.3 1.1 lb 0.58* 52* 0.62b 0.33 7.8 0.88b 0.61* 26.2
Gt3 0.70:: 0,04 3.3 0.83c 0.50* 11.9 0.43c 0.05 2.1 0.301: 0126“‘ 10
Gt4 0.90d 0.21 * 5.6 0.981: 0.63* 38 0.l7d 0.17 -1,3 0.570 0.4* 16.4
Gt5 0,l2e 0.11 -0.7 0.5211 0.23* 8.9 0.06e 0.06 1.9 0.01€ 0.01 3.3
G16 0.15€ 0.07 -0.8 0.55d 0.485‘ -22.6 0.09e 0.02 -1.3 0,08e 0.04 4.9
l FST = forest savanna transition zone, DS = dry savanna; 2 in short rainy season in 1996; 3 least square means of severity index of 80
plants; 4 difference between least square means of severity indices of inoculated and non-inoculated plots 45 dpi; 5 Gtl = IT84E-124, Gt2 =
1T84S-2246-4‚ Gt3 =lT86D-715, Gt4 = IT81D-1137, G15 = IT81D-1228-14‚ G16 = 1T86D-7l9; 6 significant at p = 0.05; 7 Percentage loss
comparing seed weight of naturally and artificially inoculated plots; ‘ significant at p S 0,05; 8 Tolerance reactions in bold.
Cowpea bacterial blight is a systemic disease which can also infect seeds, depending on the genotype. Latent
infection was detected in symptomless seeds of two genotypes (IAR-48 in Badura, IT92KD~257~7 in lbadan) in
spite of low symptom severity in the field (Tab. 3). Similar observations were reported by Sikirou (1999) forsymptomless cowpea stems of the resistant cowpea genotype lT86D-719, which harboured more than l0"cfu/g

41
stem. These observations indicate that some genotypes may restrict seed infection. Latent infection of seeds
should therefore be thoroughly considered in selection of genotypes, since seeds are important factors in
dissemination of the disease. Races or pathotypes could not be identified for X. campestris pv. vignicola (Khatri-Chhetri et al. 1999, Wydra et al. 2002), although some genotypes identified as resistant in the field revealed
susceptible to specific X. campestris pv. vignicola strains (Sikirou 1999). Therefore, also for selection for
cowpea bacterial blight resistance, inoculation with virulent strains of various origin is recommended.
Tab. 3: Disease severity (%) under natural infection of three cowpea genotypes in four ecozones in Nigeria(Ibadan — forest savanna transition, Samaru — dry savanna, Kano — Sudan savanna, Badura — highaltitude [944m]) and quantification ofX. campestris pv. vignicola in their seeds
Place Genotype Disease Detected bacteria
severit (%)) cfu/ of seed)lbadan IT93K-l 140 25 0
Samaru IT93K-1 140 5-25 0
Kano IT93K-1 I40 25-75 0
Kano [AR-48 25-50 6x10‘
Badura IAR-48 s 2.3x1o’
Samaru IAR-48 5 O
Kano lT92KD-257~7 50-75 4.2x105
Ibadan IT92KD-257-7 5 3x10‘
l = severity level at flowering to podding stage rated in percentage of leaves with symptoms (D. Florini, IITA. 1997);0%-l0% = low severity; >l0% = high severity
Conclusions
The presented results suggest that besides resistance and susceptibility, tolerance as well as latency play a majorrole in bacterial blight epidemiology in cassava and cowpea. Since various degrees of resistance and tolerance,as well as various levels of latent infections of cassava and cowpea were observed, and the genetic basis of
resistance and tolerance are known to be different (Agrios 1997), a specific evaluation of genotypes for both
traits and an accurate characterization of the relationship between the host and the pathogen are essential to
develop a well standardized and detailed evaluation system. Besides the quantification of symptoms - determined
by counting leaves with symptom types to avoid subjectivity in scoring - and their analysis in form of the area
under the disease progress curve covering the whole growing season, collection ofplant growth and yield data as
well as quantification of bacteria especially in propagating material and seeds should be part of the evaluation
and selection procedure for resistant varieties. In multilocational trials - in different seasons/years and ecozones
in order to identify genotype x environment interactions - genotypes should be grown in comparison to
susceptible and resistant standards. Since the level of disease is often influenced by factors such as the infection
pressure and temporal and spatial fluctuations in environmental conditions, an inoculation is recommended to
achieve a homogeneous infection. Testing with representative pathotypes is proposed to identify genotype x
pathogen interactions, Only the eombination of these selection criteria allows a precise selection of resistant
(including tolerant) genotypes. Parallely, the identification of genetic markers for resistance — for cassava,
markers for resistance to pathotypes ofX. campestris pv. mar/ihotis were recently identified (Jorge et al. 2000,Zinsou et al. 2002, this volume) — should support and accelerate the selection efforts.
Acknowledgements
Part of the presented results were subject of the PhD thesis of R. Sikirou and A, Fanou. The studies were
conducted at the lntemational Institute of Tropical Agriculture, Benin, and financed by BMZ, Germany. I thank
B. Hau, Hannover, for critically reviewing the article.

42
References
1. AGRIOS, G.N.; Plant Pathology. Fourth Edition. Academic Press. SanDiego, USA, 19972. ALLEN, C.: Directions for future research on bacterial pathogenicity. In: Prior, P.; Allen, C.; Elphinstone, J.
(Eds.): Bacterial Wilt Disease. Molecular and Ecological Aspects. Springer Verlag Berlin, 1998, 155-1563. BOS, L.; PARLEVIET, J.E.: Concepts and terminology on plant/pest relationships: toward consensus in
plant pathology and crop protection. Annu. Rev, Phytopathol. 33, 1995, 69-1024. FANOU, A.: Epidemiological and ecological investigations on cassava bacterial blight and development of
integrated methods for its control in Africa. PhD thesis. University of Gö ttingen, Germany, 1999, 199 p5. HAMMERSCHMIDT‚ R.: Changing susceptibility into resistance. Physiol. Mol. Plant Pathol. 57, 2000, 1-36, JOHNSON, R.: A critical analysis of durable resistance. Annu. Rev. Phytopathol. 22, 1984, 309-3307. JORGE, V.; FREGENE, M.A.; DUQUE, M.C.; BONIERBALE, M.W.; TI-IOME, ‚I,; VERDIER, V,:
Genetic mapping of resistance to bacterial blight disease in cassava (Manihot esculenta Crantz). Theor.
Appl. Genet. I01, 2000, 865-8728. KHATRI-CHHETRI, G.; WYDRA, K.; RUDOLPH, K. (): Variability of strains ofXanthomonas campestris
pv. vignicola, incitant of cowpea bacterial blight and bacterial pustule, collected in several African andother countries. In: Mahadevan, A. (Ed.): Plant Pathogenic Bacteria. Centre Adv. Study Botany, Univ.Madras, India, 1998, 296-302
9. KPEMOUA, K.E.: Etude comparative du developpement de Xanthomanax campestris pv. maniholis, chezdes Varieties de manioc sensibles et resistantes; approches histologiques, ultrastructurales et cytochemiquesdes mecanismes de la pathogenese. PhD thesis, University ofNantes, France, 1995
10. LIAO, B.S,; SI-IAN, Z.H.; DUAN, N.X.; TAN, Y.J.; LEI, Y.; LI, D.; MEHAN, V.K.: Relationship betweenlatent infection and groundnut bacterial wilt resistance. In: Prior, P.H.; Allen, C.; Elphinstone, I. (Eds.):Bacterial Wilt Diseases, Molecular and Ecological Aspects, Springer-Verlag, Berlin, Heidelberg, 1998, 294-299
11. PATEL, P.N.: Bacterial pustule disease in Tanzania: pathogenic variability and host resistance. Thirdlntemational Congress ofPlant Pathology, Munich, August 1978, 1978, p 72 (abstr.)
12. SIKIROU, RACHIDATOU: Epidemiological investigations and development of integrated control methodsof bacterial blight of cowpea caused by Xanthomonas campestris pv. vignicola. PhD thesis. University of
Gö ttingen, Germany, 1999, pp 21813. SIKIROU, R.; WYDRA, K.; RUDOLPH, K.: Selection of cowpea genotypes resistant to bacterial blight
caused by Xanthomonas campestris pv. vignicola. In: DeBoer, S. (Ed.): Plant Pathogenic Bacteria, X. Int.Conf., Charlottetown, Canada, 200l,pp 309-314
14. WALTHER, U.: Wie sicher ist „ Feldresistenz“ ? Erlä utert an mehrjähriger Prü fung von Sommergerstengegen Puccinia hordei Ott. Tagungsberichte der AdL der DDR, Nr. 271, Teil II „ Schaderreger in derGetreideproduktion“ , 1988, 243-246
15. WALTHER, U.; HABEKUSS, A,; KOPAHNKE, D.; PROESELER, G,; SCHLIEPHAKE, E.; JAHOOR,A.; KNÜ PFFER, H.; ENNEKING, D,: Methods for the assessment of biotic stress factors — disease and pestresistance. http://barley.igk—gatersleben.de/methods/GENRES-CT98-104-evaluation methods l999.gdf accessed in
January 200216. WYDRA, K.; MSIKITA, W.: Overview of the present situation of cassava diseases in West Africa. In:
Akoroda, M. 0,; Ekanayake, I. J. (Eds.): Root Crops for Poverty Alleviation. Proc. 6th Trienn. Symp. Intem,
Soc. Trop. Root Crops Africa Branch (ISTRC-AB), Lilongwe, Malawi, 22-28 October 1995. ISTRC, IITA& Government of Malawi, 1998, 198-206
17. WYDRA, K.; RUDOLPH, K.: Development and implementation of integrated control methods for majordiseases of cassava and cowpea in West-Africa. Gottinger Beitr. Land- und Forstwirtsch. in den Tropen und
Subtropen 133, 1999, 174-18018. WYDRA, K.; VERDIER, V.: Occurrence of cassava diseases in relation to environmental, agronomic and
plant characteristics. Agr. Ecosyst. Environ. 93 (1-3), 2002, 211-22619. WYDRA, K.; FANOU, A.; DIXON, A.G.O.: Effect of cassava bacterial blight on cassava SP0“ /th
parameters and root yield in different ecozones and influence of the environment on symptom developmentIn: Akoroda, M.C.; Ngeve, IM. (Eds.): 7th Trienn. Symp. Intern. Soc. Trop. Root Crops Africa Branch
(ISTRC-AB), Cotonou, Benin, 1998, 2001, 562-56920. WYDRA, K.; FESSEHAIE, A.; FANOU, A.; SIKIROU, R.; JANSE, J.; RUDOLPH, K.: Variability of
strains of Xanthomonas campestris pv. manihotis, incitant of cassava bacterial blight, from differentgeographic origins in pathological, physiological, biochemical and serological characteristics. In:
Mahadevan, A. (Ed.): Plant Pathogenic Bacteria. Centre Adv. Study Botany, Univ. Madras, India, 1993,317-323
21. WYDRA, K,; FANOU, A.; SIKIROU, R.; ZANDJVANAKOU, M,; ZINSOU, V.; RUDOLPH, K.: Integratedcontrol of bacterial diseases of cassava and cowpea in West Africa. In: DeBoer, S. (Ed.): Plant PmhogenicBacteria, X. Int. Conf„ Charlottetown, Canada, 2001, 280-287

43
22. WYDRA, K.; Kl-IATRI-CHI-IETRI, G.; RUDOLPH, K.: Pathological and physiological characterization of
the incitant of cowpea bacterial blight and bacterial pustule, Xanthomonas campestris pv. vignicola, from
different geographic origin. In: Lalithakumari, D.; Raman, N. (Eds.): Recent Developments in Plant
Pathology, Centre for Advanced Study in Botany, University of Madras, India, 2002 (in press)23. WYDRA, K.; ZINSOU, V,; FANOU, A.: The expression of resistance against Xanthomonas campestris pv.
manihotis, incitant of cassava bacterial blight; in a resistant cassava variety compared to a susceptible
variety. In: Mahadevan, A. (Ed): Plant Pathogenic Bacteria. Centre Adv: Study Botany, Univ. Madras,lndia, 1998, 583-592 -
g
24. ZINSOU, V.: Studies on the expression of resistance of cassava genotypes — including individuals of the
mapping population ~ to cassava bacterial blight. MSc thesis. University of Gö ttingen, Gennany, 2001,
pp 5725. ZINSOU, V., WYDRA, K., AGBICODO, E„ AHOHUENDO, B., AND RUDOLPH, K.: Studies on the
resistance of cassava genotpyes to bacterial blight. In: DeBoer, S. (Ed.): Plant Pathogenic Bacteria, X. Int.
Conf, Charlottetown, Canada, 2001, 301-305
26. ZINSOU, V.; WYDRA, K.; AHOHUENDO, B.; JORGE, V.; VERDIER, V.: Evaluation of cassava
genotypes, including individuals of their genome mapping population, for resistance against bacterial blight.9"‘ Symposium on „ New Aspects of Resistance Research on Cultivated Plants“ : Bacterial Diseases.
November 15 - 16, 2001 Aschersleben, Germany (this volume), 2001

44
Session 4: Ecology, Epidemiology and Prevention of Pathogen Transmission
Ecological role of a thermoregulated multi-drug efflux proteinofErwinia amylovora
Antje Burse, Matthias S. Ullrich
Max-Planck-lnstitute for terrestrial Microbiology, Karl-von—Frisch-Strasse, 35043 Marburg, Germany
Abstract
Fire blight of apples and pears is promoted by particular environmental conditions such as temperature. Analysisof temperature regulated gene expression in E. nmylavom was carried out at 18° C and 28° C. In the course of
these experiments the genetic locus mdeA was identifled. The gene, expressed 2-fold higher at 18° C than at
28° C, codes for a multi-drug efflux protein. This protein belongs to secondary transporters and is integrated in
the inner membrane. As a single-component efflux pump MdeA extrudes its substrate into the periplasm. AmongE, amylovora and related strains the mdeA gene is widely distributed. The transporter mediates resistance to
amphiphilic antibiotics, like norfloxacin, ethidium bromide, and berberine. In the natural environment of E.
amylovora MdeA may function in the defence of the pathogen against toxins produced by antagonisticmicroorganisms, like P. rzgglomerans.
Introduction
Temperature has a profound effect on regulation of many bacterial virulence genes necessary for successful
invasion of a host organism. Analogous to studies of temperature-influenced gene expression in animal
pathogens, virulence genes of plant pathogens could also be affected by temperature. Recently, a mini'1‘ n5-based
promoterless I3-glucuronidase (uidA) reporter gene was used to identify temperature—responsive genetic loci in
the tire blight pathogen E. amylovora [1]. Among mutants with increased uidA expression at lower temperaturea transposon insertion was found in a gene coding for a multi-drug efflux protein. Multi-drug efflux pumps are
recognized as an important component of bacterial resistance to many drugs and toxic compounds. In Gram-
negative bacteria, the majority of these pumps appear to be secondary transporters that utilize the proton motive
force as source of energy [2]. Their broad substrate specificity makes bacteria moderately resistant to various
agents and thus temporarily helps them to either generate mutations or acquire other resistance mechanisms to
adapt to a particular environment. The objective of this project was to characterize the multi-drug efflux protein
of E. amylovora with regard to its temperature-dependence and substrate specificity.
Results and discussion
E. mny/ovora 1189 was mutagenised by random insertion of the miniTnS-uidA transposon. Upon ITIIIIITHS-llid/‘I
insertion, transcriptional fusions could be formed between the promoterless uidA gene and E. amylovora
promoters resulting in production of /J’-glucuronidase (GUS). Quantification of GUS activity in the mutants
revealed that a genetic locus (mrle/4) encoding a multi-drug efflux protein is 2-fold higher expressed at 18° C than
at 28° C.
Complete sequencing of the transporter region from E. amylovara revealed that a single open reading frameencodes MdeA. The transporter is located in the inner membrane and extrudes its substrate into the periplasm.According to sequence comparisons at the protein level with data base entries, MdeA showed high homology to
multi-drug efflux proteins in E. coli and in V. pm'ahaemoIyticus [3]. These proteins are members of a recentlynew identified secondary transporter family, termed the multi-drug and toxic compound extrusion (MATE)family [4]. Proteins were combined in the MATE-family by computer analyses and many of them are
physiologically still uncharacterised.
Investigations of occurrence of mdeA showed that the gene is widely distributed among E. amylovora strains and
its relatives, like E. pyrifolicze, E. r/mpomici, P. agglomerzms, P. carotovorum subsp. carotovarum, and B.
salicis.
To analyse the substrate specificity ofthe transport protein of E. aniylovoru, the mile/l gene was transformed into
a special E. coli mutant KAM3 lacking the major multi-drug efflux system AcrAB. E. coli KAM3 is highlysusceptible to different antibiotics, dyes and other toxic compounds. Minimal inhibition concentration tests of E.
coli‘ KAM} complemented with the mdeA gene of E. amylovora showed substrate specificity of/m/e/1 towardsthe amphiphilic cations norfloxacin, ethidium bromide, and berberine.

45
E. amylavora wild-type and its mde/1 mutant were compared with regard to differences in the resistance towards
toxic compounds. Only in the case of nortloxacin the mutant showed decreased resistance in comparison to the
wild-type strain.
However, norfloxacin is a compound not occurring in the natural environment of E. amylovara. The pathogenhas to cope with different toxins produced on one hand by the host plant and on the other hand by epiphyticmicroorganisms. The epiphyte P. agglomerans is a known antagonist of E. amylovurzz and ofien produces
antibiotics inhibiting growth of the pathogen. Inhibition zone assays were carried out to investigate the role of
MdeA in antibiotic resistance. Seventy P. agglamerzzns strains were tested. Fifieen of them were able to inhibit
growth of the pathogen. To supematants of five P. agglomerans cultures the mdeA mutant was more susceptible
compared to the wild-type (Fig. 1).
Fig. 1. Inhibition zone assays on agar—plates to investigate differences in resistance ofE. amylovora wild-type(A) and mdeA mutant (B) to culture supematants of antibiotic producing P. agglomerans strains.Well 1, P. agglomerans 2668; 2, P. agglomerans 2b/89; 3, P. agglomerans 15b/89; 4, P. agglomerans
16b/89; S, P. agglomerans 1189-3
‚
;:=.->. -
A ‘ "-V
B I’'
_
." V‘ I
.
_‘ ;- r=- i"U
i:
ll
z;e
~ r
7 3V
V T '5‚
'
. „ v,
ii ; ‘ wild-type mdeA mutant
References
1. GOYER, C.; ULLRICH, M.: Thennoregulation of virulence gene expression in the fire blight pathogenErwinia amylovora. Acta Hort. 489, 1999, 321-326
2. NIKAIDO, H.; ZGURSKAYA, H.I.: Antibiotic efflux mechanisms. Curr. Opin. Infect. Dis. 12, 1999, 529-536
3. MORITA, Y.; KODAMA, K.; SHIOTA, S.; MINE, T.; KATAOKA, A.; MIZUSHIMA, T.; TSUCHIYA,T.: NorM, a putative multidrug efflux protein, of Vibria parahaemolyticus and its homolog in Escherichiacoli. Antimicrob. Agents Chemother. 42, 1998, 1778-1782
4. BROWN, M.H.; PAULSEN, I.T.; SKURRAY, R.A.; The multidrug efflux protein NorM is a prototype of a
new family of transporters. Mol. Microbiol. 31, 1999, 393-395

46
Studies on the biological control of fire blight in EgyptPeter Laux, Wolfgang Zeller
Federal Biological Research Centre for Agriculture and Forestry (BBA) Institute for Biological Control,Darmstadt, Germany
Introduction
Two epiphytic bacterial strains, Rzzhnella aquatilis Ra 39 and Pseudomonas spec. R1, were shown to reduce flre
blight symptoms in field trials during 1998, 1999 and 2000 (2, 3, 4).
In studies on the mode of action, we obtained results suggesting that competition for sucrose and induced
resistance might be involved in the mode of action of Rahnellu aquatilis Ra39 against Erwinia amylovora.
Materials and Methods
Efficacy evaluations of antagonistic strains were carried out at experimental orchards in Alexandria (Egypt) and
Darmstadt (Germany). After application ofbactereial antagonists the trees were subjeeted to secondary infection.
Agar diffusion tests were done on Miller—Schroth (MS), Nutrient—Sucrose (NS) and Glucose—Asparagin(GA)-medium, Co-culturing of bacterial antagonists and Ea7/74 was carried out in buffered Nutrient-Sucrose (NS)-medium. The sucrose concentration was varied between 2% and 0,2. For experiments with Co—cultures,spontaneous mutants of Ea7/74 and the antagonistic strains Ra39 and R1 resistant against 100 ppm Streptomycin(Sm) or Rifampicin (Rt) respectively were used. Plate counts of mixed cultures of Ea 7/74Sm and Ra39Rf or
RlRf were done on solid NS-medium containing 100 ppm Streptomycin or Rifampicin respectively.Lipopolysaccharides (LPS) were extracted by the phenol-water method. The aqueous LPS solution was dialysedagainst demineralised water (12000 Da; Serva) until reaching a conductivity below 50 us and lyophilised. For
infiltration of plants with LPS two years old apple trees (Golden Delicious), one year old begonia plants (varietyOslo) and '14 days old bush bean plants were used. LPS were applied at a concentration of 5 mg per ml. After 24
hours treated leafs were checked for the presence of the superoxide anion by staining with
Nitrobluetetrazoliumchloride ( 1 ).
Results
Field experiments
The bacterial strains Ra39 and R1, were shown to reduce fire blight symptoms in field trials in Egypt and
Germany during 1998, 1999 and 2000. The efficacies of both isolates varied between 39 and 53% or 45 and 58%
respectively (Table 1).
Table 1: Field efficacies ofbacterial antagonists against fire blight between 1999-2001
Antaonistic strain Efficac %Rahnella a uatilis Ra39 39-53Pseudomonas sec. R1 45-58
Standard Plantom cin 68-80
Mode ofaction
In the agar diffusion test the antagonistic strain R1 caused the formation of inhibition zones. There was no
difference in antibiotic activity between MS- and GA-medium, Contrastingly Ra39 did not show any antibiotic
activity in this test (Figure 1).
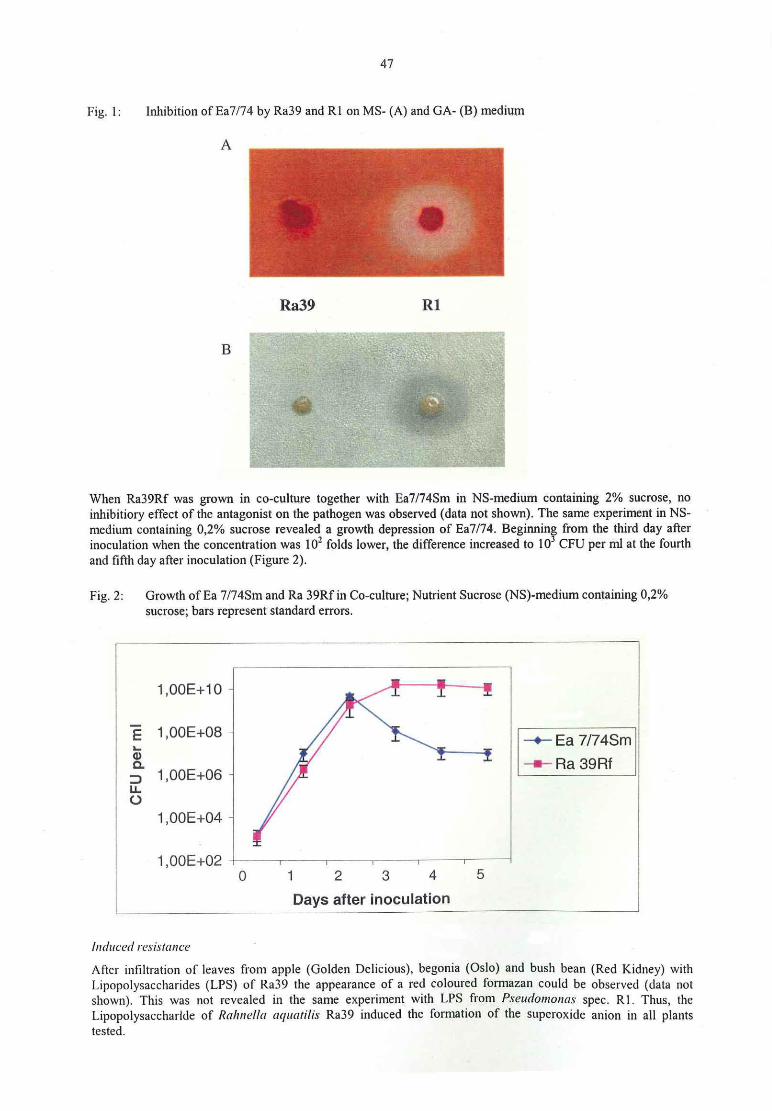
47
Fig. 1: Inhibition of Ea7/74 by Ra39 and R1 on MS- (A) and GA- (B) medium
A
Ra39 R1
When Ra39Rf was grown in co-culture together with Ea7/74Sm in NS—medium containing 2% sucrose, no
inhibitiory effect of the antagonist on the pathogen was observed (data not shown). The same experiment in NS-
medium containing 0,2% sucrose revealed a growth depression of Ea7/74. Beginning from the third day afterinoculation when the concentration was 101 folds lower, the difference increased to 103 CFU per m1 at the fourthand fifth day after inoculation (Figure 2).
Fig. 2: Growth of Ea 7/74Sm and Ra 39Rf in Co-culture; Nutrient Sucrose (NS)-medium containing 0,2%sucrose; bars represent standard errors.
1 ,00E+1 0
_
1 OOE 08E '+
— o— Ea 7/74SmII)D- -l- Fta 39 FIf3 1,00E+06Ll.O
1,00E+O4
l,0OE+020 1 2 3 4 5
Days after inoculation
Induced resistance '
After infiltration of leaves from apple (Golden Delicious), begonia (Oslo) and bush bean (Red Kidney) with
Lipopolysaccharides (LPS) of Ra39 the appearance of a red coloured fonnazan could be observed (data not
shown). This was not revealed in the same experiment with LPS from Pseudomonas spec. R1. Thus, the
Lipopolysaccharide nf Rahnella uquzzrilis Ra39 induced the fomiation of the superoxide anion in all plantstested.

48
Discussion
While the antagonistic strain Pram/omomis spec. R1 was shown to cause the formation of inhibition zones in the
agar diffusion test independent from the amino acid composition of the medium, the strain Ralmella aqmzli/isRa39 did never show any antibiosis in this system. In experiments with co-cultures, a direct inhibition of Ba 7/74
by Ra39 in vitro was observed at a sucrose concentration of0,2% but not at a concentration of 2%. Contrastinglythe strain R1 caused a direct inhibition of Ea7/74 independent from the sucrose concentration. Thereforecompetition for sucrose might be involved in the mode of action of Rahne//a at/untilis Ra39 against fire blight.Until yet, this mechanism has not been demonstrated for antagonistic bacteria of Erwinia anzyfavora. Becausethe strain Pseudomonas R1 caused a direct inhibition of Ea7/74 in the agar diffusion test and in co-culture withthe pathogen, we assume an antibiotic effect against the pathogen for this strain. For many P. z1gglo:neI'm1r-strains the inhibition of bactericide activity by the presence of amino acids in the medium has been described (5).Therefore, amino acid composition of the plant tissue was suggested to affect the efficacy of P. agg/omeransstrains in preventing fire blight symptoms. Contrastingly the antibiotic activity of the strains Rtilmellu uquarilisRa39 and Pseudomonas spec. R1 was not dependent on the amino acid composition of the medium. WhilePseudomonas spec. R1 expressed antibiotic activity on all media tested; Ruhriella (Iqzratilix Ra39 did never showthis effect. Despite these results it cannot be excluded, that specific substrates, only available at the flowersurface of fire blight host plants, are stimulating a direct inhibition of the pathogen by this strain. The result thatLPS of Ra 39 induced the formation of superoxide in leafes of apple, begonia and bush bean indicates theinvolvement of induced resistance in antagonism of Ra 39 against Ea 7/74. Because the same effect was alsoobserved on non-host plants of Erwinia amylovorn, we hypothize that resistance induction by Ra39 is alsoaffecting other pathogens.
References
1. DOKE, N.: Involvement of superoxide anion generation in the hypersensitive response ofpotato tuber tissueto infection with an incompatible race of Phyrophtora iI1fesrm1s' and the hyphal wall components. Physiol.Plant Pathol. 23, 1983, 345-357
2. LAUX, P.; ZELLER, W‚ : Zur Bekaempfung des Feuerbrandes (Erwinia nmylovoru) mit bakteriellen
Antagonisten im Freiland. Mitt, Biol. Bundesanst. Land- Forstwirtsch. 376, 2000, 192-1933. LAUX, P.; HOFER‚ K.; ZELLER, W.: Untersuchungen im Freiland zur Bekaempfung des Feuerbrandes mit
bakteriellen Antagonisten. Abstract. 2. Symposium Phytomedizin und Pflanzenschutz im Gartenbau, 27.-30.09.l999, Wien, 1999, 53-54,
4. STEINBRENNER, B.: Untersuchungen zur Ö kologie des Feuerbrandes (Erwinia umylovora) und
Entwieklung eines Modells zur Prognose der Krankheit. Dissertation Universitä t Heidelberg, 19915. WODZINSKI, R.S.; UMHOLTZ, T.E.; RUNDLE, J.R.; BEER, S.V.: Mechanisms of inhibition of Erwinia
amylnvora by Erw. lierbicola in vitro and in vivo. App. Bacteriol.76‚ 1994, 22-29

49
Latest results on fire blight control with natural products
Wolfgang Zeller, Peter Laux
Federal Biological Research Centre for Agriculture and Forestry (BBA),Institute for Biological Control, Darmstadt, Germany
Introduction
After a severe outbreak of Fire Blight in 1993 the use of streptomycin in the following year was permitted for the
fruit growing practice in Germany (1). Since that time the permission was given under Very restricted conditionsfrom year to year. As in 2000 very low residues of streptomycin could be found in several samples of honey, thepermission of the use of the antibiotic was withdrawn by the ministry of agriculture. So the research onalternatives to the antibiotic for the control of Fire Blight in pome fruits became of main interest. Research inthis direction has been mainly undertaken in Germany by the Federal Biological Research Centre in Darmstadt
(Tab. l). In recent time studies also in cooperation with the plant protection service, the industry and colleaguesof the Universities occurred. In the following mainly studies on the development of biologicals, as for instanceplant extracts, etheric oils and antagonists of the pathogen.Results
Table I: Studies on alternatives to the use of streptomycin at the Biological Research Centre (BBA)
l. Phase: Testing of Fungicidal compounds, Copper-Preparates and other chemical Substances (1975-1985)
Efficient Preparates:
I Copper-Compound COPAC E
c Chinolin-Compound FIRESTOP
I Oxanilic acid
2. Phase: Biological Alternatives (1985-2001)
Development and Testing of Plant Strengtheners0 Plant Extracts with direct Effect0 Plant Extracts with Induced Resistance Effect0 Etheric Oils
A- Plant Extracts
After a screening of more than 100 leaf extracts out of different plant species 24 extracts could be observed withbactericide effects in vitro against E. amylovora (5). The most effective plant extracts could also show underfield conditions a positive percent control, as for instance extracts from Mahonia aquifolium, Berberis vulgariswith an efficacy between 50 to 70% (Tab. 2). Later on another variety ofplant extracts were tested on their plantstrenghtener character, which showed under natural and artificial infection conditions during blossom time an
control efficacy of 50-60% on different apple varieties (5). Actually the experiments are carried out especiallywith the most effective extract of ivy (Hedera helix) in combination with other substance, in order to increase theefficacy (Mosch et al., 2000). In table 3 it is shown that the highest percent control of the ivy extract was
observed by combination with a metal salt or copper preparate Neull40F with 79,0 and 87,8% (4h beforeinoculation), The effect of these mixtures was comparable with the use of Streptomycin after artificial blossominfection. The effect could be continued also for natural infection (Tables 3, 4).
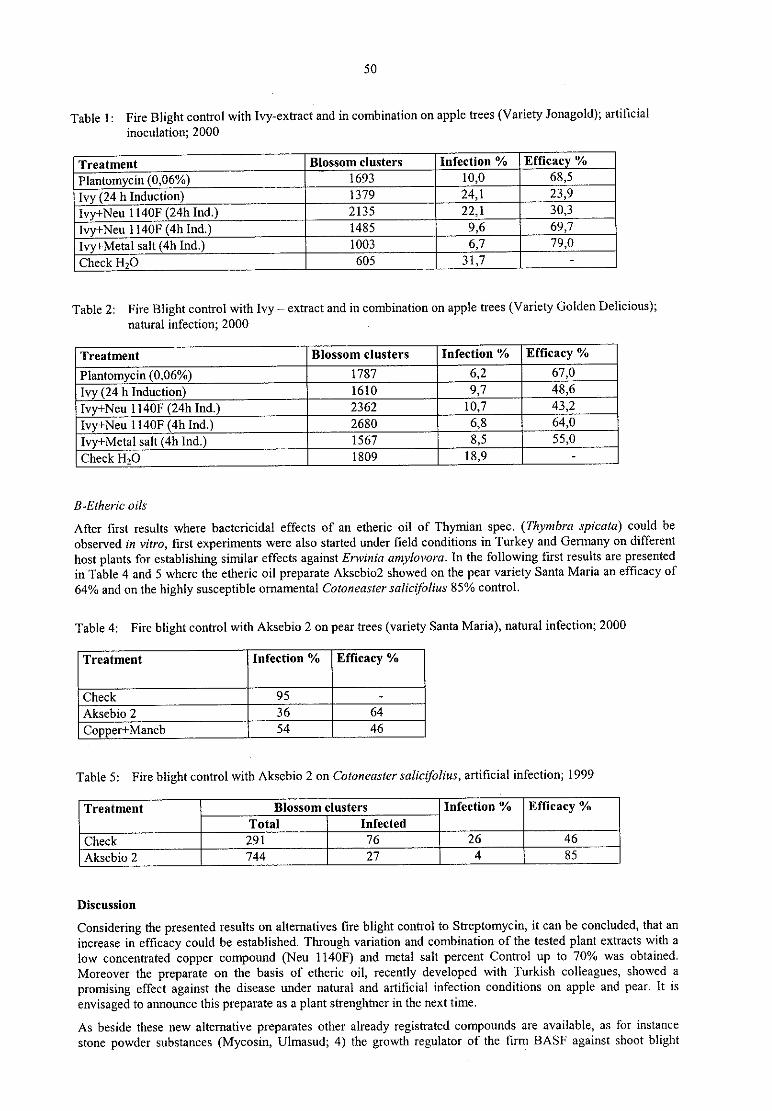
50
Table l: Fire Blight control with Ivy—extract and in combination on apple trees (Variety Jonagold); artificial
inoculation; 2000
Infection % Efficacy %
Plantomycin (0.06%) 1693
Ivy (24 h Induction) I379
lvy+Neu l140F (24h Ind.) 2135
Ivy+Neu n40F (4h 1nd.) 1485 jIvy+Metal salt (4h Ind.) 1003
Check H2O —
Table 2: Fire Blight control with Ivy — extract and in combination on apple trees (Variety Golden Delicious);natural infection; 2000
3'° Ss° m “ um” Effimy "/°
P1antomyc1n<0‚06%) 1787
Ivy<24hInducnon> 1610
Ivy+Neu 1l40F (24h Ind.) 2362
1vy+Neu men (4h Ind.) 2680 1.51Ivy+Metal salt (4h Ind.) 1567
Check H20 I809 —
B~Etheri'c oils
After first results where bactericidal effects of an etheric oil of Thymian spec. (Thymbra spicata) could be
observed in vitro, first experiments were also started under field conditions in Turkey and Germany on different
host plants for establishing similar effects against Erwinia amylovara. In the following first results are presented
in Table 4 and 5 where the etheric oil preparate Aksebio2 showed on the pear variety Santa Maria an efficacy of
64% and on the highly susceptible ornamental Cotaneaster salicifolius 85% control.
Table 4: Fire blight control with Aksebio 2 on pear trees (Variety Santa Maria), natural infection; 2000
—Aksebio 2
Copper+Maneb
Table 5: Fire blight control with Aksebio 2 on Cotoneaster salictfolius, artificial infection; 1999
Treatment Blossom clusters Infection t’/o Efficacy %
Infected
Aksebio 2
Discussion
Considering the presented results on alternatives fire blight control to Streptomycin, it can be concluded, that an
increase in efficacy could be established. Through variation and combination of the tested plant extracts with a
low concentrated copper compound (Neu l140F) and metal salt percent Control up to 70% was obtained.
Moreover the preparate on the basis of etheric oil, recently developed with Turkish colleagues, showed a
promising effect against the disease under natural and artificial infection conditions on apple and pear. It is
envisaged to announce this prcparate as a plant strenghtncr in the next time.
As beside these new alternative preparates other already registrated compounds are available, as for instance
stone powder substances (Mycosin, Ulmasud; 4) the growth regulator of the firm BASF against shoot blight

51
(Prohexadion, BASF) and the plant activator BTH (Bion; Novartis) with a SAR-effect against the disease (2, 3)there are further successful charges for the control of this dangerous bacteriosis in fruit growing, whereStreptomycin could be used with the highest degree of control compared to the alternative preparats (Fried,2001). In consideration of the discussion in the German public moreover of the EEC—adrriinistration to banantibiotics out of the agriculture and the potential occurrence of resistant strains of Erwinia amylovora it seems
necessary, to refer growers on the site of already excisting alternative preparate.
References
1. BACKHAUS, (S.F.; KLINGAUF, F.: Die Feuerbrandkrarikheit und ihre Bekämpfung in der BundesrepublikDeutschland, Nachrichtenbl. Dt. Pflschutzd. 50, 1998, 193499
2. BAYSAL, Ö .; LAUX, P.; ZELLER, W.: Studies on Systemic Acquired Resistance (SAR) effect of BTH
(BION) against fire blight. Abstract of 9"‘ Int. Workshop on Fire Blight, 08.—12.10.2001, Napier Warmemorial Centre, 2001, Abstr. No. p-30
3. BAYSAL, Ö .; LAUX, P.; ZELLER, W.: Further studies on the induced resistance (IR) effect of the plantextract from Hedera helix against fire blight. Abstract of 9"‘ Int. Workshop on Fire Blight, 08.-12.10.2001,Napier War memorial Centre 2001, Abstr. No. P-31
4. KOWALEWSKI, A.: Feuerbrandbekéimpfiing mit alternativen Präparaten. Obstbau 8, 1996, 404~4055. MOSCH, J.; ZELLER, W.: Bekämpfung des Feuerbrandes mit ausgewä hlten Pflanzenextrakten.
Nachrichtenbl. Dt. Pflschutzd. 41, 1989, 149-151

52
Induction of resistance to Clavibacter michiganensis subsp. michiganensisby pre-inoculation of tomato young plants under field conditions in
BulgariaV. Sotirova', E. Griesbachz
‘ lnstitute ofGenetics, BAS, 1 1 13, Sofia, Bulgariallnstitute for Epidemiology and Resistance, Federal Centre for Breeding Research on Cultivated Plants,
Theodor-Roemer-Weg 4, D~O6449 Aschersleben, Germany
Abstract
Pre»inoculation with the avimlent strain NCPP 3123 or injection with its extracellular polysaccharides (EPS)induced a protective effect in young tomato plants against virulent isolates of Clavibncter rnichigazre/1sii' subsp.rnichiganensis (Cmm). This was observed also under field conditions in Bulgaria during 1998 till 2001. The bestprotection was observed when young plants were pre-inociilated respectively the EPS was applied in the axil ofthe topmost full developed leaf or into the stem 14 days before challenge inoculation with virulent Cmm strains.No differences in yields could be observed between these variants and untreated control plants.
Introduction
Bacterial canker or wilt is a very important disease on tomatoes in greenhouse as well as in the field. Thepathogen - CIavi'l7acter michiganensir subsp. niic/1z'ganensi's (Cmm) - occurs world-wide and is sometimes thecause of yield losses up to 60 % or more. Tomato plants are susceptible to Cmm in every stage of development.The reason for the high losses is the early infection through seeds or contaminated soil, the long incubationperiod and the easy spread of the causative agent in the plant population by treatments. At present there are no
possibilities for direct control of the disease. In previous investigations we could find that the avirulent strain ofCmm NCPPB 3123 pre-inoculated in young tomato plants induced resistance to virulent isolates (GRIESBACH etal. 2000a, b). This defensive reaction was mainly dependent on the inoculation sites, the cell concentration usedfor pre~ and challenge inoculation, and the time interval between both inoculations.
in 1998 a bilateral co-operation was started to test this phenomenon under optimal conditions for the pathogen ofbacterial wilt and canker in Bulgarian tomato fields where Cmm has a wide natural distribution.
Material and methods
Following Cmm strains were used: for pre-inoculation the avirulent strain NCPPB 3123 (SÜ LE, Budapest); for
challenge inoculations the highly virulent strain Cmm 8 (Sofia/Bulgaria). Cmm isolates were Cultivated on
p0tato—dextrose-agar (PDA) at 28 ° C for 48 till 72 h, suspended in sterile tap water, and diluted to a density of5x 10g cells/rril for inoculations. For isolation of the EPS the cultivation of NCPPB 3123 was carried out in YDCfluid media. Cmm cells were heat-killed by submerging the bacterial suspension in boiled water (100 DC) for 20min. Viability was checked by plating the suspension on YDC-agar. The freeze dryer LYOVAC GT 2 was usedfor lyophilization of Cmm cells.
The varieties ‘ Ideal’ and ‘Trapezitsa' were cultivated in greenhouse in steamed soil. Generally, young plantswere inoculated before planting out in the field ten or fourteen days later as a rule with 2 or 3 repeats of 15plants/variant. At dryness the tomatoes were irrigated via furrows in the field. Leaf axil injections were carriedout by deep prick with an injection needle (No.16 or 20) into the axil of the uppermost, fully developed leaf of
young plants and filling up this canal with bacterial suspension or dissolved EPS during the drawing of theneedle. Inoculations during the removal of lateral shoots were immediately enforced after breaking out bydropping the suspension of the avirulent isolate on the wounds, and immediately after pulling in of the drop(about 15 min.) ,
suspensions of virulent bacteria were applied. Pre-inoculation of the 3123 or application of itsEPS into the stems of young plants was carried out with the same injection needle using for leaf axil injection byprick into or through the stem and by filling up the canal during the drawing of the needle. For pre-inoculation ofthe avirulent strain or its EPS via leaves the apex or stalk of leaf were cut off with a scissors that has been dippedin the bacterial suspension or EPS solution.
Disease severity (ds) was classified by following scale: 1: no symptoms; 2 : mild wilting of some leaflets; 3 : >
30%; 5 : 30-50%; 7 : 51-75% of leaves wilted; and 9 : > 75 % ofleaves wilted and plants dead. The yields were
calculated on 10 plants per variant.

53
Results
In earlier greenhouse experiments we could find that the resistance inducing effect was optimal when taking a
time interval from 7 and 14 days between pre- and challenge inoculation. Therefore, at first both of these
variants of time were tested under field conditions over a period of two years. The results could been confirmed
only in 1998. One year later an intensive hailstorm heavily damaged the young plants five days after planting.The consequence of this was that only the control plants inoculated with 3123 remained symptomless. Watercontrol plants and those pre—inoculated 14 days before challenge inoculation showed some wilting. But plantspre—inocu1ated 7 days before challenge inoculation had about such strong symptoms as plarfis inoculated onlywith the highly virulent Cmm 8. A repetition of these trials in 2000 using only the optimal time interval from
fourteen days between pre- and challenge inoculation confirmed the results of the previous two years. About60% of pre—inoculated plants did not develop symptoms after challenge inoculation till the end of growingseason, and from the rest only some leaflets or at most two leaves per plant showed wilting. However, controlplants inoculated only with the virulent strain Cmm 8 were extremely damaged.
These results were reflected in the yield level of the different tested variants in 1999 and 2000. Harvest resultswere approximately equal as well in the control variants water and 3123 as at pre-inoculated plants challengedwith highly virulent Cmm 14 days later. In contrast, the yields of tomatoes were very low when the time intervalbetween pre- and challenge inoculation was 7 days or which were infected only with Cmm 8. Based on theseresults for the further investigations on young plants were chose a time interval between pre- and challengeinfection of 14 days.
With regard to a practical use of the resistance induction method, it was necessary to test the influence of
different techniques of preparations of the avirulent strain on its protective effect. For a longer—terrn preservation
as well heat killing and freeze drying as utilisation isolated EPS of bacteria cells was suitable. The wiltingdevelopment after pre-inoculation of those preparations showed that the effectiveness was reduced distinctlyonly by heat killing. In case of the EPS the highest degree of efficiency could be observed after its application in
a concentration of 4 mg/ml. Equally results could also be obtained in the crop of the control variants water and3123 as well as after pre-inoculation of living cells or after application of EPS solution concentrated to 4 mg/ml.From the other pre—treated plants we could reap approximately 20 (pre—inocu1ated with freeze—dried living cells)to 50% (pre—inoculated with heat killed 3123) less. However, harvest of plants inoculated only with Cmm 8 was
reduced to 20%.
Regarding the main mode of spreading of the pathogen via fresh wounds during the removal of lateral shoots, we
tested if the resistance induction in young plants was sufficient to block this way of transmission or whether an
application of the 3123 directly over fresh wounds had a better protective effect. We could observed that the pre-inoculation was somewhat more effective after leaf axil injection of young plants than after removing sideshoots. In 1998, most plants remained symptomless till the end of vegetation or the expression of symptoms was
extremely reduced, irrespective if the challenge infection took place over one, two or three buds. Under theweather conditions in 1999 (strong hailstorm five days after planting outdoors) wilting development started veryearly and was more progressive as one year before, so that the protective effect was lower. But the disease
severity was reduced more than half as well. In contrast for that the pre-inoculation via fresh wounds was not
always so successful and the wilting development increased in dependence on the number of infected wounds.Pre-inoculation carried out at young plants brought about twice the amount of yield as from tomatoes pre-inoculated after removing of lateral shoots.
But under practical conditions the induction of resistance in young tomato plants by leaf axil injection could beto complicated. Therefore the effectiveness of some other methods of pre-inoculation were tested comparativelyas injection into respectively prick trough the stem or cut different parts of leaves with a contaminated scissors.Parallel to living cells of the 3123 were used its EPS too. It turned out that there were no differences betweenboth elicitors. Always, all methods used for pre-inoculation or application via the stem showed no differences inthe wilting development, that is all were equal effective as leaf axil injection. On the other hand the protectiveeffect induced in plants by leaf cutting with bacterial suspension or EPS solution was never so high in all thetested variants.
Discussion
The phenomen of resistance induction by pre-inoculation was tested in the pathosystem tomato/Cmm by otherauthors too. For it were used saprophytic bacteria, pathogens from other host plants (ERCOLANI 1970, SÜ LE
1982, Ts1ANTos 1985), or mutants of Cmm (ERCOLANI 1970). But none of these inductors was so effective as
the avirulent strain NCPP 3123 of Cmm (GRIESBACH et a1. 2000 a, b). In comparison to most other microbialresistance inductors with a wide host plant spectrum (SCHÖNBECK et al. 1993) the defensive reaction after pre-inoculation of this strain or its EPS in young tomato plants was characterised as well by high specificity as
systemic and persistent effect.

54
A comparison between the results received in Germany and under the field conditions in Bulgaria mostlyshowed similar trends. But during the four-years investigation in Bulgaria, where the climatic conditions were
optimal for the pathogen development, the pre-inoculation of NCPPB 3123 or application of its EPS in youngplants always let to a substantial better protective effect till the end of growing season. That was observed
particularly using a time interval of fourteen days between both inoculations. These results are reflected in theyields too, that is, the controls of water or 3123 were just as high-yielding as the variant with a time interval oftwo weeks. On the other hand, if the challenge inoculation took place after seven days, the degree of efficiencyvaried from year to year depending on the weather conditions. This may suggest that the defensive mechanisms
were activated after this time but not yet consolidated optimally. The effect induced by pre-inoculation of youngplants was preserved during a long period of time, so that challenge inoculations via fresh wounds by removingof side shoots could be hindered to a certain degree, depending on weather conditions. Possibly when youngplants were stressed after pre-inoculation to a extreme high degree, the activation of defensive mechanisms couldproceed less intensively than under normal conditions.
All these results indicate that the pre-inoculation of the natural avinilent Cmm strain NCPP 3123 or application
of the EPS in young tomato plants before planting out induced systemic, persistent, and extreme resistance to
virulent Cmm isolates also under a high pressure of infection in Bulgarian fields. Especially the EPS of the 3123could been considered for the development of an elicitor preparation. The handling of numerous young plantswould cause additional work and costs but the growers in afflicted regions would have safer yields using thismethod ofbiological control.
References
1. ERCOLANI, G.L.: Bacterial canker of tomato. IV. The interaction between virulent and avirulent strains of
Carynebacterium michigzmense (E. F, Smith) Jens. in vivo. Phytopathol. Mediterr. IX, 1970, 151-1592. GRIESBACH, E.; EISBEIN, K.; KRAMER, I.; MÜ LLER, J; VÖ LKSCH, B,; Induction of resistance to
bacterial pathogens in the pathosystem tomato/Clavibacter michiganensis subsp. michiganensir I.Characterization of the resistance induction. Z, Pflanzenkr. Pflanzensch. 107, 2000a‚ 449-463
3. GRIESBACH, E.; EISBEIN, K.; KRÄ MER, I.; RAMM, M.; MULLER, J.; VÖ LKSCH, B.: Induction ofresistance to bacterial pathogens in the pathosystem tomato/Clavibacter michigzmensis subsp. michiganensisII. Characterization of the resistance inductor. Z. Pflanzenkr. Pflanzensch. 107, 2000b, 464-483
4. SCHONBECK, F.; STEINER, U.; KRASKA, T.: Induzierte Resistenz: Kriterien, Mechanismen, Anwen»
dung und Bewertung. - Z. Pflanzenkr. Pflanzensch. 100, 1993, 541-557
5. SÜ LE, S.: Induced resistance in tomato to bacterial canker caused by Colynebacterium michiganense. —
Tagungsberichte der AdL der DDR, Nr. 216, Teil II, 1982, 473-4766. TSIANTOS, J.A.: Inhibition of the pathogen Corynebacterium michigahense pv. michiganense by the
epiphytic saprophyte Pseudomonas aeruginosa after spray or systemic inoculation. 3"‘ National Phytopathol.Conf., 16.-18.10.1985, Hellenic Phytopathol. Soc., 1985, 19

55
Investigations on epidemiological importance of weeds as a source forRalstonia solanacearum
Gudrun Barchend, Anita Schmidt
Federal Centre for Breeding Research on Cultivated Plants,Institute for Resistance Research and Pathogen
Diagnostics; Theodor Roemer Weg 4, D~O6449 Aschersleben
Ra/.rta/via soImmccm'u1n is a gram-negative plant-pathogenic bacterium that causes bacterial wilt in a largenumber of different plant species. Since 1976 outbreaks of brown rot or bacterial wilt to potato caused by thebacterium Ralsioniu A'0f(ll1(lC€ (l/‘H711 race 3 biovar 2 have been reported from several Western European countries.Due to its low temperature optimum, race 3 is considered a threat to potato culture in Europe. An infectionsource for R. solanacearmn are latently infected seed potato tubers or contaminated surface water.
The bacterium also attacks various solanaceous weeds. Tusiime et al. (1998) reported on a number of latentlyinfected non-solanaceous weeds in the highland of Uganda. European weeds from several plant families havebeen identified as potential hosts following artificial inoculation (e.g. Eupatorium ca/rnabfum, Tussilugafmfam), although no evidence has been found that they are infected in their natural habitat. In Europe, only twosolanaceous natural weed hosts of R. so/amzcecirum have been described: the perennial bittersweet - Solanumzlulcamaru and the annual black nightshade — Solanum nigrum.
We started to investigate the epidemiology of this bacterium. Of particular interest is the question which of themost common weed species in Germany can serve as sources to maintain the pathogen under field conditions.Weeds often occurring in potato production are Ecliinoc/iloa crus-galli, Elyrrfgia repenr, Gulinsoga p1II'\'I_fl0I‘(I,Lvcopus czu'opaeu.r, Setariu viridis, Salmmm nigrum, Spergula arvensis am! Urticu riioicu.
Plants of several weed species were examined by artificial inoculation to determine whether they can act as hostsfor R. .ro/mmcemunr (race 3 biovar 2). To this aim we artificially infested the soil with a suspension of R.solamrccariun (twice with 200 ml 10 8 cfu/ml ) and then we planted five weed seedlings in the soil. In a furthervariant, R. solanacet/rmn infected plants of S. nigrum or S. me/ongena were used as a source of infection. Percontainer, one infected Solanum plant was placed in the centre and five weed seedlings around it. Theexperiments started in May. The first variants were examined in June/July, Starting with the analysis of thepercolated water: Stalks and roots ofthe weeds were tested 3 weeks later.
During the Vegetation period 900 samples were taken from the test plants. Different parts of these plants were
investigated for occurrence of R. rolanaccarmn. Isolation by semi—selective medium (SMSA),immunofluorescence technique (IF) and bioassays using tobacco and tomato plants were applied as detectionmethods. Table 1 summarises the test results. Regardless ofthe infection method applied it was possible to detectR. rolanttcetzrzzm in all examined weeds. An exception was Spergzrla arvensis. Here we could detected R.salunacenrmn only after planting in infested soil. It seems to be possible that the causal agent may get into thesoil via the roots.
Table 1: Detection ofR. solo/racearzmz in weeds using Solanum plants as inoculation sources
. Presence of R. rolanacearmn inweed Specles weeds planted in soil artificially weeds grown around inoculated
infested with suspension Solanum lants
Gulmsuga lulrwi/[Owl
Spgrgula arwgnsix (/"” CU (ll-01-cu
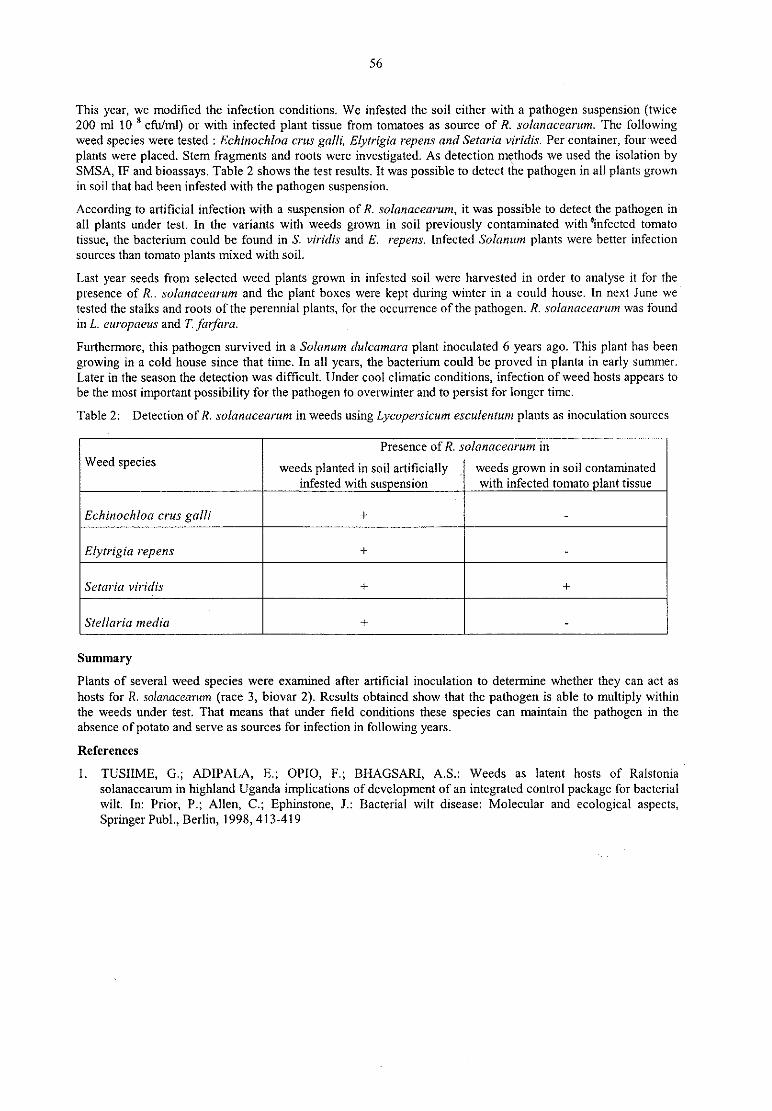
56
This year, we modified the infection conditions. We infested the soil either with a pathogen suspension (twice200 ml 10 8 cfu/ml) or with infected plant tissue from tomatoes as source of R. salrmaceamm. The followingweed species were tested : Echirwchloa crus galli, Elytrigia repens and Setariu viridis. Per Container, four weedplants were placed. Stem fragments and roots were investigated. As detection methods we used the isolation bySMSA, IF and bioassays. Table 2 shows the test results. It was possible to detect the pathogen in all plants grownin soil that had been infested with the pathogen suspension.
According to artificial infection with a suspension of R. solanacenrum, it was possible to detect the pathogen in
all plants under test. In the variants with weeds grown in soil previously contaminated with ‘ infected tomato
tissue, the bacterium could be found in S. viridis and E. repens. Infected Solanum plants were better infection
sources than tomato plants mixed with soil.
Last year seeds from selected weed plants grown in infested soil were harvested in order to analyse it for the
presence of R.. solanucearum and the plant boxes were kept during winter in a could house. In next June we
tested the stalks and roots of the perennial plants, for the occurrence of the pathogen. R. salaziacermun was foundin L. europaeus and T. farfara.
Furthermore, this pathogen survived in a Solanum dulcamara plant inoculated 6 years ago. This plant has been
growing in a cold house since that time. In all years, the bacterium could be proved in planta in early summer.
Later in the season the detection was difficult. Under cool climatic conditions, infection of weed hosts appears to
be the most important possibility for the pathogen to overwinter and to persist for longer time.
Table 2: Detection of R. solanacearum in weeds using Lycopersicum esculentum plants as inoculation sources
Presence of R. salrmacearum inweed Specles weeds planted in soil artificially
_
weeds grown in soil contaminatedinfested with sus ension with infected tomato lant tissue
Echinochloa crus repens
Stellaria media Summary
Plants of several weed species were examined after artificial inoculation to determine whether they can act as
hosts for R. solanacearum (race 3, biovar 2). Results obtained show that the pathogen is able to multiply withinthe weeds under test. That means that under field conditions these species can maintain the pathogen in theabsence of potato and serve as sources for infection in following years.
References
1. TUSIIME, G.; ADIPALA, E.; OPIO, F,; BHAGSARI, A.S.: Weeds as latent hosts of Ralstoniasolanacearum in highland Uganda implications of development of an integrated control package for bacterialwilt. In: Prior, P.; Allen, C.; Ephinstone, J.: Bacterial wilt disease: Molecular and ecological aspects,Springer Publ., Berlin, 1998, 413-419

57
Session 5: Resistance Breeding and Transgenic Resistance
Breeding of F1 hybrids in cabbage with resistance to Xanthomonascampestris pV. campestris
Harm Lö ptien I, Erika Griesbach 2
1 GZG Saaten AG Marne, Konigstr, 47, D-25709 Mame, Germany2 Institut fiir Epidemiologie und Resistenz, Bundesanstalt fiir Zü chtungsforschung an Kulturpflanzen, Theodor-
Roemer-Weg 4, D-06449 Aschersleben, Germany
Abstract
Xanthomonas campestris pv. campestris (Xcc) is one of the most important pathogens in Brassica oleraceavegetables. Several strains of this bacterium were collected and a program was started to develop resistant basic
breeding material. We succeeded in selecting a number of resistant genotypes after spray-inoculation in the
juvenile stage.
Introduction
The bacterium Xanthomonas campestrir pv. campestris (Xcc) is the causal agent of black rot in cabbage and
related Bmssica vegetables. The bacterium is distributed world-wide, it has a wide range of hosts and can
survive in plant refuse for at least two years. Furthemiore Xanthomonas is able to invade seeds and even a verylimited rate of contamination can lead to severe black rot damages in crop production under wet and warm
climatic conditions.
In 1995 a program was started in Germany with the intention to select basic breeding material for the
development of varieties in cabbage and related vegetables which show a good and stable resistance againstXcc. It is the aim of these investigations to collect a number of isolates of the bacterium and to select highlyvirulent strains for preparing the inoculum. Besides it is the intention to develop a method which gives reliableresults in testing plant material for resistance against black rot. It will be another part of the program to identifythe race specifity of the selected Xcc strains and to get insight into the inheritance of resistance in favourableplant genotypes.
Material and methods
The isolates of Xcc were obtained from infected Brassica seed and leaves collected in different parts of Europeand South Africa. Sero-typed strains were received from ALVAREZ (University of Hawaii) and definatedraces from IGNATOV (Russian Academy of Sciences, Moscow). For inoculation Xcc strains were cultivated 48
h on yeast-dextrose-Ca-agar (YDCa) at 28 ° C. Foreign cabbage varieties which are said to have a certain degreeof resistance to Xcc and a wide range of inbred lines were offered by the seed companies which join the project.Another part of the material was received from Cornell University and the Crucifer Genetics Cooperative,University of Wisconsin in the USA. A third part was taken from the collection of Brassica oleracea and relatedspecies present in the German gene bank at Gatersleben.
The plants are inoculated by spraying (Desaga SGI) the whole surface of their leaves and stems until runoff
with a mixture of six to seven highly virulent isolates of Xcc. The concentration of the inoculum is 5 x 10 8
cells/ml. Afterwards the inoculated plants are kept for three days in a humid chamber where they are forced toproduce a lot of guttation droplets at the leaf margins. Then the plants are grown under nomial greenhouse
conditions. The temperature is about 26 0C in the day and about 20 ° C in the night during the whole run of theexperiment. In order to study the influence of the plant age on the development of symptoms, some genotypeswere transplanted into plastic tunnels and inoculated in an adult stage.
About 14 days after inoculation a first check for resistance is done. The plants are scored according to the
appearance and size of water-soaked or necrotic spots on the leaves and typical v-shaped yellow symptomsexpanding from hydathodes. The plants are classified according to a scale ranging from 1 to 9 (1, 2, 3, 5, 7 and
9). Genotypes belonging to disease severity category 1 for instance are completely without any symptoms and a
rating 5 means that about 50 % ofthe surface in a plant is covered by typical symptoms.
Plants which belong to class 1, 2 or sometimes 3 are checked for a second and even a third time after about 7 or
14 additional days. Genotypes belonging to higher ratings are always discarded and finally only plants withpermanent low ratings are selected and raised to flowering. They are self-pollinated or crossed with superior

58
inbred lines. Their progenies are tested again for a further selection cycle and for studies on the inheritance ofresistance to Xcc.
DAS—ELISA (NACHTIGALL 1996) and direct tissue blotting immuno assay (DTBIA) according to the methodof PROLL (1994) were used to detect the bacterium in plant tissue and to analyze the spread of the pathogen insusceptible and resistant plant genotypes.
Results and discussion
It is a prerequisite for a reliable selection of resistant plants to use an inoculum characterized byix high virulence.There are strong differences between Xcc isolates in this respect. In total 94 isolates were tested and about two
third of them proved to have sufficient virulence. It is also necessary to check this character from time to time
again, though we did not find a severe lost of virulence during the growth of the bacterium on YDCa-agar at low
temperatures.
In the Brzzssica/Xantlmnzomzr-system a distinction is made between stern resistance which prevents the plantletsfrom a rapid spread of the pathogen in the vascular system of stem and leaf resistance (IGNATOV et al. 1998,1999) We think that our special procedure for testing against Xcc, especially the scoring of symptomdevelopment after different time intervals makes it possible to check not only leaf but also stem resistance. It
was an extrem rare event to find a plant which was completely free fl'om symptoms in our investigation.Normally the most resistant genotypes were characterized by rating category 2 for at least one time during theselection period. These are plants with a very limited expression of symptoms. Very often they developed smalllocal necrotic symptoms starting from stomata and hydathodes.In Table 1 some results of testing line 3018 are presented. This line is closely related to B. oleracea accession PI436606 selected by DICKSON and HUNTER (1987) as a new source for resistance against Xcc. Most plants inour line 3018 are characterized by a low rating concerning the reaction to Xcc. Two white cabbage hybridvarieties (‘ Se1ma', ‘ Kalorama') are included into this trial. They are susceptible to the pathogen and are
regarded as control genotypes in our resistance evaluation.
Table 1: Disease severity rating in cabbage line 3018 and its progenies from selling (AU 4167, AU 4168) and
crossing with susceptible genotypes (AUK 1256, AUK 1279, AUK 1292, AUK 1296, AUK 1314)
Line Disease severity rating (Number of lants)1 2 3 5 7 9
1997 (20 dp. i.)
3018 1 15 7 2‘ Selma’ 3 7 13 1‘ Kalorama' 4 12 8
1998 (l9 dp. 1.)
AU 4167 5 4AU 4168 2 14 1‘ Selma’ 2 13 1‘ Kalorama' 12 5
2000 (21 dp. i.)
AUK 1256 6 2AUK 1279 8 2AUK 1292 3 19AUK 1296 12 12AUK 1314 17 7‘ Selma’ 10 13 1‘ Kalorama’ 1 17 6
Those individuals from line 3018 showing the lowest rating for reaction to Xcc were self-pollinated. Theprogenies were tested for resistance ( AU 4167, AU 4168) and the rating of disease severity was found to bemainly concentrated on low degrees demonstrating no severe segregation for resistance and susceptibility inthese offsprings.
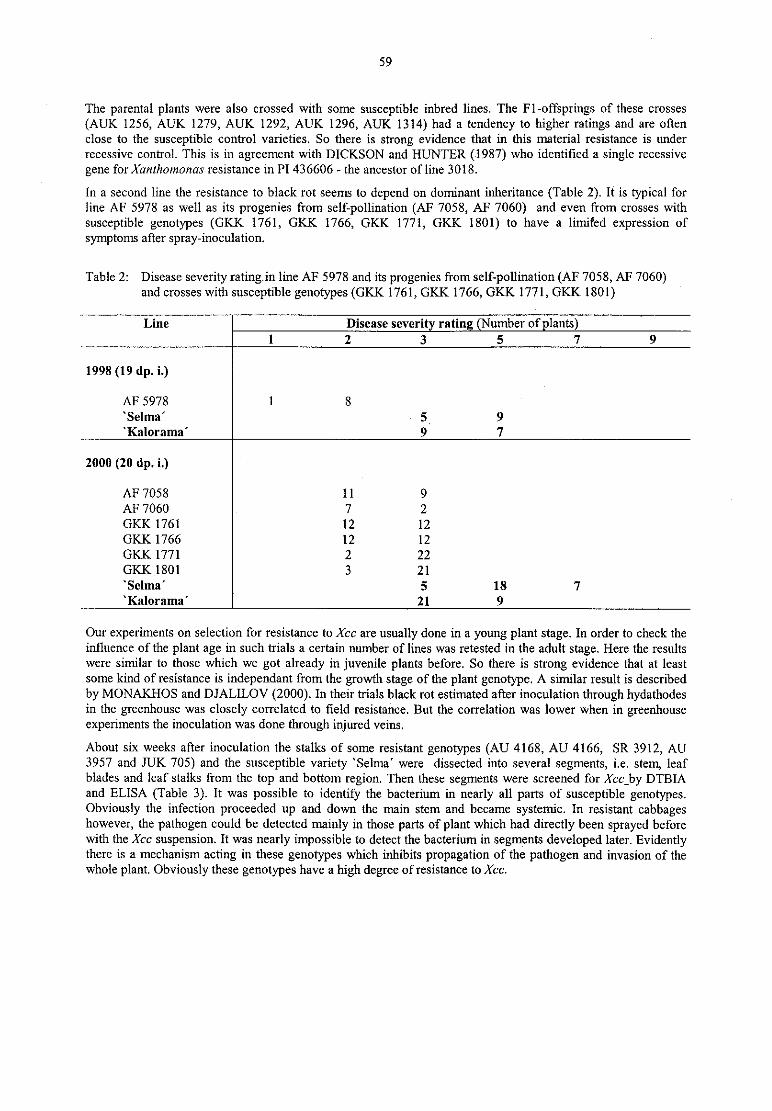
59
The parental plants were also crossed with some susceptible inbred lines. The Fl-offsprings of these crosses
(AUK 1256, AUK 1279, AUK 1292, AUK 1296, AUK 1314) had a tendency to higher ratings and are oftenclose to the susceptible control varieties. So there is strong evidence that in this material resistance is underrecessive control. This is in agreement with DICKSON and HUNTER (1987) who identified a single recessive
gene for Xanthomonas resistance in P1 436606 — the ancestor ofline 3018.
In a second line the resistance to black rot seems to depend on dominant inheritance (Table 2). It is typical forline AF 5978 as well as its progenies from self-pollination (AF 7058, AF 7060) and even from crosses withsusceptible genotypes (GKK 1761, GKK 1766, GKK 1771, GKK 1801) to have a limited expression of
symptoms after spray-inoculation.
Table 2: Disease severity rating in line AF 5978 and its progenies from self-pollination (AF 7058, AF 7060)and crosses with susceptible genotypes (GKK 1761, GKK 1766, GKK 1771, GKK 1801)
Line Disease severit ratin umber of lants)1 2 3 5 7 9
1998 (l9 dp. i.)
AF 5978 1 8‘ Selma’ 5, 9‘ Kalorama’ 9 7
2000 (20 dp. i.)
AF 7058 11 9AF 7060 7 2GKK 1761 12 12GKK 1766 12 12GKK 1771 2 22GK.K 1801 3 21‘ Selma’ 5 18 7'Kalorama' 21 9
Our experiments on selection for resistance to Xcc are usually done in a young plant stage. In order to check theinfluence of the plant age in such trials a certain number of lines was retested in the adult stage. Here the resultswere similar to those which we got already in juvenile plants before. So there is strong evidence that at leastsome kind of resistance is independant from the growth stage of the plant genotype. A similar result is described
by MONAKI-IOS and DJALILOV (2000). In their trials black rot estimated after inoculation through hydathodesin the greenhouse was closely correlated to field resistance. But the correlation was lower when in greenhouseexperiments the inoculation was done through injured veins.
About six weeks after inoculation the stalks of some resistant genotypes (AU 4168, AU 4166, SR 3912, AU3957 and JUK 705) and the susceptible variety ‘ Selma’ were dissected into several segments, i.e. stem, leafblades and leaf stalks from the top and bottom region. Then these segments were screened for Xcc_by DTBIAand ELISA (Table 3). It was possible to identify the bacterium in nearly a11 parts of susceptible genotypes.Obviously the infection proceeded up and down the main stem and became systemic. In resistant cabbagcshowever, the pathogen could be detected mainly in those parts of plant which had directly been sprayed beforewith the Xcc suspension. It was nearly impossible to detect the bacterium in segments developed later. Evidentlythere is a mechanism acting in these genotypes which inhibits propagation of the pathogen and invasion of thewhole plant. Obviously these genotypes have a high degree of resistance to Xcc.

60
Table 3: Detection ofXan!homomzs campestris pv. campestris in different parts of resistant and susceptible
genotypes by DAS—ELISA and direct tissue blotting immuno assay (DTBIA)
NumberLast Tested part of plants (41dp.i.)L i n e of disease Stalk s t e rn leaf blades leaf stalks
tested rate part DTBIA l ELISA ELISA ELISA
lants 0 — + 1 — + — — + - ‘l’
Resistant genotypes gAU 4168 l0 2,0 top 8 2 8 2 9 0 7 3
..
AU 4166 12 2,0 top ll 1 10 1 12 0 12 0
SR 3912 7 2,0 top 7 o 7 0 5 1 6 o
AU 3957 7 2,0 top 7 0 7 0 4 2 7 0
.
JUK 705 l0 2,0 top 10 0 10 O 9 1 10 0
bottom 9 1 9 1 s 3 6 2
1
susceptible cultivar
‘ Selma’ 12 top 0 12 0 12 7 1 9 3
bottom 0 12 5 7 — — — —
These investigations were supported by Dr. M. Nachtigall and Dr. E. Proll
Our experiments show that there is a genetic basis for resistance to Xcc in our plant material. But in this stage of
our project it is not possible to answer questions about the number and action of genes in our selected genotypesbecause most of our results are derived from inoculation with a mixture of highly virulent bacterium isolates ofwhich the race specifity is not clear in all cases up to now. In total Xcc is classified today into six races on thebasis of their reaction to a series of differential Brassica genotypes (VINCENTE et al. 2000). Race 1 and 4 are
said t0 be of major importance in Brassica world-wide and also in Germany these two races seem to beprominent. A part of our future work will concentrate on such an identification of races in our strain collection.In a further step we want to check our resistant plant material with distinct races ofXcc and we intend to identifythe specific genes which are responsible for the resistance reaction.
References
1. DICKSON, M.D.; HUNTER, J .E.: Inheritance of resistance in cabbage seedlings to black rot. Hort. Science
22, 1987, 108-1092. IGNATOV, A.; KUGINUKI, Y.; HIDA, K.: Race-specific reaction of resistance to black rot in Brassiczz
aleracea. Europ. J. Plant Pathol. 104, 1998, 821-8273. IGNATOV, A.; KUGINUKI, Y.; HIDA, K: Vascular stem resistance to black rot in Brassica oleracea. Can.
J. Bot. 77, 1999, 442-4464. MONAKHOS, G.F; DJALILOV, F.S.: Genetic sources and methods for estimation of Brassicas resistance
to black rot. J. Russ. Phytopathol. Soc. 1, 2000, 83-875. NACHTIGALL, M.: Nachweis des Erregers der Ademschwä rze (Xanthomonas campestris pv. campestris)
bei Brassica-Arten. Bundesanstalt fü r Zü chtungsforschung an Kulturpflanzen. Jahresbericht, 1996, 60-616. PROLL, E.: Direct tissue plotting assay — ein einfaches Schnellverfahren zum quantitativen Virusnachweis
in Pflanzen. Nachrichten BAZ 2, 1994, 20-247. VINCENTE, J.G.; CONWAY, J.; ROBERTS, S.J.; TAYLOR, J.D.; Identification and origin of
Xanthomonas campesrris pv. czzmpesrris races and related pathovars. Phytopathology 91, 2001, 492499

61
Transgenic resistance against Erwinia soft rot — A four-year fieldexperiment (1997-2000)
C. WegenerFederal Centre for Breeding Research on Cultivated Plants, Institute for Stress Physiology and Quality of Raw
Materials, D -18190 Gross Liisewitz, Germany ([email protected])
SummaryPectate lyase (PL) enzymes, the virulence determinant of Erwinia bacteria, cause a degradation of tissue cellwalls into unsaturated oligogalacturonates eliciting plant defence responses. Transgenic potato plant linesexpressing an Erwinia PL-enzyme were analysed in a 4-year field experiment. It is shown that the endogenousPL enzyme mediated an enhanced resistance to Ec-soft rot also in field grown tubers. For example, bacterialmaceration of tissue slices cut from tubers of the PL—expressing lines was significantly reduced (P< 0,05)compared to the non-transgenic counterpart. Furthermore, tissue cell walls of PL—transgenic lines were lesssusceptible to Erwinia-derived enzymes. An activation of the plant defence may be indicated by the enhancedPPO-activity in tuber tissue of the PL-active lines as well as by a strengthened formation of necrosis on thewound surface of tubers.
Introduction
Pectate lyase (PL) enzymes are the major virulence determinant of Erwinia carotovora (EC) bacteria. Theydegrade plant cell wall pectin into unsaturated oligogalacturonates (OG) eliciting plant defence responses (Ryan,1988), such as a release of phenols and their oxidation by polyphenol oxidases (PPO), cross linking of cell wallproteins or production of inhibitors, phytoalexins, lignin and suberin. The Erwinia PL-enzymes have a doublefunction in soft rot pathogenesis: 1) The degradation of tissue cell walls leading to soft rot symptoms and 2)formation of QG-signals inducing plant defence mechanisms. The latter was the reason why a DNA sequenceencoding the isoenzyme PL3 of Ec subsp. atroseptica was transformed by means of Agrobacterium tumefaciensinto potatoes of cv. Dé siré e. The aim was that traces of the PL—enzyme produced intracellularly and liberated bywounding the tuber tissue could release OG-elicitors initiating a pre-activation of plant defence leading to anenhancement of resistance to Erwinia soft rot. Greenhouse experiments revealed that plant lines expressing thePL showed indeed a better resistance of tuber tissue to Ec-bacteria (Wegener et al., 1996). In a next step fourPL~transgenic plant lines were grown over a period of four years (1997-2000) on the field in order to investigate:a) How the PL«expressing plants behave under environmental conditions and b) whether the PL-inducedresistance functions also in field grown tubers.
Material and methods
Plant material. Seed tubers of the four PL-transgenic potato lines, the non-transgenic counterpart and theoriginal cv. Dé siré e (Breeders line) were hand planted in April in two plots per line, harvested in September andthen stored at 4 ° C. In lines D4, D19 and D29 enzyme expression was controlled by the potato patatin B33-promoter. In line Cl it was driven by the CaMV 35S~promoter. In case of the C-line the PL-expression was in allplant organs, while in D-lines it was confined to tuber tissue.
Resistance analyses. A) Tuber tissue slices were incubated between filter papers soaked in suspension ofErwinia bacteria or B) infiltrated with a solution containing the Erwinia-derived enzymes before the decrease ofcell viability was measured by neutral-red vital staining (Weber & Wegener, 1986). C) 10 Tubers were coredwith a 10 mm cork-borer, submerged in a suspension of Ec-bacteria for 1 min and then incubated for 5 days at20 ° C in closed PE-bags. After that tubers were cut across the bore hole and the diameter of the rotting zone was
measured. Average values were calculated per tuber half. The data are means of three replicates (SD of the test s5 %). D) Polyphenol oxidase (PPO) activity in tissue extracts was determined according to Jockusch (1966).Statistic analyses were done by t-test.
Results and discussion
Development of the PL-expressing plants on the field was normal as well as tuber production. Plots of the linesD4, D19 and D29 could not be distinguished from that of the non-transformed control line and the original cv.
However, it was noticed in every year that plants of the line C1 bearing a 35S»PL3 gene construct were smallerthan that of the other lines and this correlated with a reduced tuber yield. All four plant lines expressed the PLenzyme which was a good prerequisite for an induction of the plant defence. Consequently, tissue slices cut fromtubers expressing the PL-enzyme showed a significantly enhanced resistance to bacterial maceration comparedto the non-transgenic counterpart (Table 1). Also their cell walls were less susceptible to Erwinia-deriveclenzymes (Table 2), perhaps due to an elicitor induced cross-linking of cell wall proteins resulting a rapid
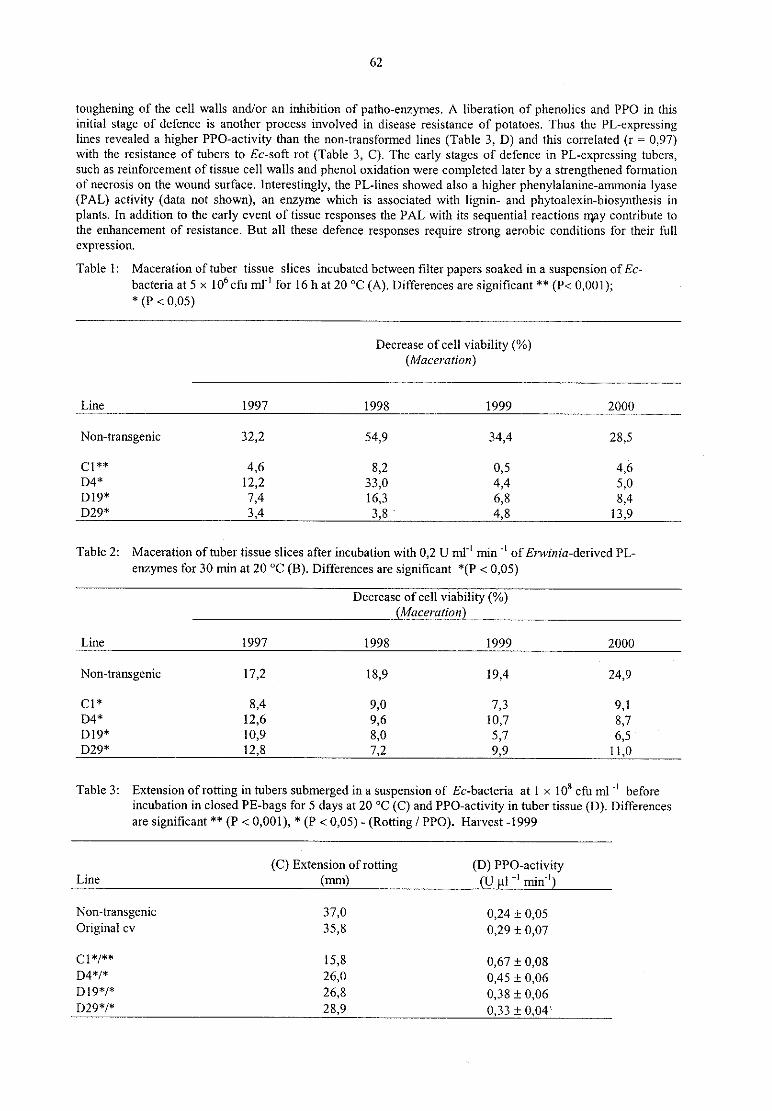
62
toughening of the cell walls and/or an inhibition of patho-enzymes. A liberation of phenolics and PPO in thisinitial stage of defence is another process involved in disease resistance of potatoes. Thus the PL-expressinglines revealed a higher PPO-activity than the non~transformed lines (Table 3, D) and this correlated (r = 0,97)with the resistance of tubers to Ec-soft rot (Table 3, C). The early stages of defence in PL-expressing tubers,such as reinforcement of tissue cell walls and phenol oxidation were completed later by a strengthened formationof necrosis on the wound surface. Interestingly, the PL«lines showed also a higher phenylalanine-ammonia lyase(PAL) activity (data not shown), an enzyme which is associated with lignin- and phytoalexin-biosynthesis inplants. In addition to the early event of tissue responses the PAL with its sequential reactions may contribute to
the enhancement of resistance. But all these defence responses require strong aerobic conditions for their fullexpression.
Table 1: Maceration of tuber tissue slices incubated between filter papers soaked in a suspension of Ec-
bacteria at 5 >< 106 cfu m1" for 16 h at 20 ° C (A). Differences are significant ** (P< 0,001);* (P < 0,05)
Decrease of cell viability (%)(Maceratian)
Line 1997 1998 1999 2000
Non-transgenic 32,2 54,9 34,4 28,5
c1** 4,6 3,2 0,5 4,6D4* 12,2 33,0 4,4 5,0Dl9* 7,4 16,3 6,8 8,4D29* 3,4 3,8 ‘ 4,8 13,9
Table 2: Maceration of tuber tissue slices after incubation with 0,2 U m1" min ‘ l ofErwinia-derived PL-
enzymes for 30 min at 20 ° C (B). Differences are significant *(P < 0,05)
Decrease of cell viability (%)(Macerution)
Line 1997 1998 1999 2000
Non-transgenic 17,2 18,9 19,4 24,9
C1* 8,4 9,0 7,3 9,1D4* 12,6 9,6 10,7 8,7D19* 10,9 8,0 5,7 6,5D29* 12,8 7,2 9,9 11,0
Table 3: Extension of rotting in tubers submerged in a suspension of Ec-bacteria at 1 >< 108 efu ml "l beforeincubation in closed PE-bags for 5 days at 20 ° C (C) and PPO-activity in tuber tissue (D). Differencesare significant ** (P < 0,001), * (P < 0,05) - (Rotting / PPO). Harvest -1999
(C) Extension of rotting (D) PPO—activityLine (mm) (U 1"‘ min")
Non-transgenic 37,0 0,24 i 0,05Original cv 35,8 0,29 1 0,07
C1*/** 15.8 0,67 i 0,08D4*/* 26.0 0,45 i 0,061319*/* 26,3 0,38 i 0,06D29*/* 28,9 0,33 i 0,04‘

63
Due to the fact that Ec-bacteria reveal a tremendous production of cell wall degrading enzymes immediatelyafter invading the tuber tissue rotting must be stopped before it get out of hand. This means, the early events of
tissue responses are decisive to defend the pathogens successfully. The endogenous PL contributes to it by a pre-activation of plant defence mechanisms and the results show that this enables an enhancement of resistance alsoin field grown tubers. But such an approach is always combined with the risk that the PL protein is normally a
pathovenzyrne with the potential to degrade plant cell walls.
The PL-expressing potato lines are an excellent model to study the protein pattern of plants in the stage of an
induced resistance which may led to new strategies in resistance research.
References
1. JOCKUSCI-I, H.: The role of host genes, temperature and polyphenol oxidase in the necrotization of TMVinfected tobacco tissue. Phytopathologische Z. 55, 1966, 185-192
2. RYAN, C.A.: Oligosaccharides as recognition signals for the expression of defensive genes in plants.
Biochemistry 27, 1988, 8879-88983. WEBER, I.; WEGENER, C.: Virulence and enzyme production of Erwinia ssp. atrosepticn on potato tuber
tissue. J. Phytopathol. ll7, 1986, 97-1064. WEGENER, C.; BARTLING, S.; OLSEN, 0.; WEBER, J.; VON WETTSTEIN, D.: Peetate lyase in
transgenicpotatoes confers pre-activation of defence against Erwinia carotnvora. Physiol. Molec. PlantPathol. 49, 1996, 359-376

64
Stability of fire blight resistance in apple
K. Richter‘ , C. Fischerz
Federal Centre for Breeding Research on Cultivated Plants‘ Institute of Epidemiology and Resistance, PO Box 1505, 06435 Aschersleben,
zlnstitute of Fruit Breeding, Pillnitzer Platz 2, 01326 Dresden-Pillnitz
SummaryThe objective of the apple breeding program in the Institute of Fruit Breeding Dresden-Pillnitz is the
development of new varieties with high fruit quality combined with resistance to economically importantdiseases. One of these is fire blight, caused by Erwinia amylovora.
The resistance of the hybrids against fire blight is evaluated at Aschersleben since 1974. New strains of the fire
blight pathogen are collected from infected areas every year. Before resistance evaluation in the greenhouse,three strains with the highest virulence on three different apple varieties are selected.
The hybrids were tested as graftings on the rootstock MM 106. 30 actively growing shoots per accession were
inoculated by a length between 15 and 25 cm with the mixture of three high virulent strains (lxl0° cfu/rnl). Theresistance of the test plants is evaluated by calculating length of necrosis to total shoot length (x 100 %) 4-6
weeks p.i. The level of the evaluation test and the yearly differences caused by different weather conditions anddifferent strains can be assessed by using of standard varieties.
New fire blight resistant cultivars from the Pillnitz breeding program are ‘ Realka’ , ‘ Reanda’ , ‘Rebella‘ , ‘ Regia‘ ,‘Regine’ , ‘Remo’ , ‘ Rene’ , ‘ Resi’ and ‘ Rewena’ . They possess a high level of fire blight resistance in growingshoots and after flower infections. The resistance is stable after repeated inoculations with different E. amylovorastrains in several years.
Introduction
After first outbreak of fire blight at the North— and Baltic Coast in 1971, the scientific research began in theInstitute of Phytopathology at Aschersleben. The screenings of apple collections showed significant differencesin the resistance to Erwinia amylovora. On the basis of these results resistance breeding was started at Dresden-Pillnitz in 1974. The resistance of the hybrids was evaluated in the greenhouse at Aschersleben.
After reunification of Germany the breeding program is continued in the Institute of Fruit Breeding Dresden-Pillnitz. The aim of the apple breeding program is to develop new cultivars with high fruit quality combined withresistance to scab, mildew and fire blight. The control of the fire blight pathogen is very difficult, because of theabsence of available chemicals. Growing resistant or less susceptible apple varieties is the most effective
measure to prevent epidemic outbreaks.
The process of breeding fire blight resistant varieties needs a long time and their introduction into the fruitproduction too. Therefore, the stability of fire blight resistance over a long period is very important.Material and Methods
,
Virulence test. Strains of the fire blight pathogen are collected from infected areas every year. Active growingshoots of the apple variety ‘ Prima’ , the advanced selection Pi-AS-181 and Malus robusta No. 5 (since 2001:
‘ Idared’ ) were inoculated with a suspension of the new isolates (109 cfu/rnl) in comparison to the strains used forthe resistance evaluation in the last years.
Shoots were inoculated by cutting through two unfolding leaves with contaminated scissors. The evaluation of
the symptoms was carried out 2 to 4 weeks p.i.
Resistance evaluation. 35 plants per hybrid had been grafted on the rootstock MM 106. 30 active growing shoots
per accession were inoculated by a length between 15 and 25 cm by cutting through two unfolding leaves withcontaminated scissors, dipped into a mixture of suspensions of three high virulent strains (each 10° cfu/ml). Totest a higher number of hybrids in some cases only 25 shoots per accession were grafted. The tested shoots havebeen evaluated by calculating length of necroses to total shoot length (x 100 °/0) 4 to 6 weeks p.i. These resultswere transformed into a score system (Table 1).

65
Table 1: Score system for the evaluation of fire blight resistance
0- 0.9 9
1.0 - 16.3 816.4 - 31.0 731.1 - 44.2 644.3 - 56.1 556.2 - 66.9 467.0 - 76.5 376.6 - 85.2 2
>86.5 1
*Severity = length ofnecrosis / shoot length x 100 %
Results and Discussion
Virulence mmlysir. Large differences in virulence occur between E. amylovom strains.
In some cases a cultivar specifity of the virulence could be observed, but no genus specifity. Different isolateshave shown the highest virulence on different cultivars. The results of the last years were exemplary for that. In1997 Ea 337 and Ea 344 from Bavaria (host plants: Pyrus and Sorbus), Ea 269 from Baden-Württemberg(Pyrur), Ea 250 from Saxony-Anhalt (Malus) and Ea 222 from the Czech Republic (Cotoneaster) were selectedas most virulent. In 1998 other strains were used in the mixture for the resistance evaluation. Ea 376 and Ea 385had been isolated in Baden-Wü rttemberg (Cotoneaster and Malus), Ea 391 in Bavaria (Craraegus). The strain Ea385 was used in the mixture in 1999 too. The two other strains were replaced by Ea 7 from Saxony-Anhalt(Pyrur) and Ea 454 from Baden-Wü rttemberg (Malus). In 2000 three isolates from South-Germany (Pyrus and
Malus) were selected as most virulent, in 2001 two strains from Baden-Wü rttemberg (Cydonia and Malus) andan isolate from Switzerland (Cydania) were used for the resistance evaluation.
German strains could not infect Malus robustzz No, 5. Therefore, ‘ Idared’ was used as the third cultivar for thevirulence test since 2001. E. amy/ovum strains from all host plants were selected for the mixture in the diverse
years. The most virulent isolates came from Baden-Wiirttemberg and Bavaria. In 2001 strains from Switzerlandcould be involved in the virulence test. Only one isolate was high virulent in relation to those from South
Germany.
The virulence of the strain mixture on the standard varieties is shown in Table 3. Often the isolates with the
highest vimlenee on Pi-AS-181 had not such a high aggressiveness on ‘ Prima’ . Only in some cases the same
isolate was best in two ore more years. Mostly, the new, fresh isolated strains had a higher aggressiveness.
Resistance evaluation. Extensive screenings of apple varieties worldwide showed that the most cultivars are highsusceptible. In a long term program at Dresden-Pillnitz susceptible Varieties with good fruit quality and resistant
genotypes were combined. The evaluation of scab-resistant Pillnitz Revcultivars® ‘ Realka‘ , ‘ Reanda‘ , ‘Rebella‘ ,‘ Regia‘ , ‘ Regine’ , ‘ Remo’ , ‘ Rene’ , ‘ Resi’ and ‘ Rewena’ showed a high level of fire blight resistance (Table 2).
The stability is a very important requirement of resistance. Examinations of stability were carried out in a diallel,crossed in 1986 and screened on one year old seedlings in 1987 (Fischer 1992). The screening was repeated after10 years on adult trees. These results compared with the resistance rate of the first test showed a high level ofcorrespondence (Fischer and Richter 1999). The distribution of the fire blight resistance in the tested progeniesshows that the genetic background seerns to be polygenie (Fischer and Fischer 1996).
To test the stability in the Re-assortment®, plants of these cultivars were evaluated together with the apple
breeding material in the glasshouse. So it was guaranteed that they were inoculated with current E. amylovorastrains from attacked orchards several times. The results are demonstrated in Table 3. The susceptibility of thestandard variety ‘ Idared’ was very high every year. According to different weather conditions differences occur
in some years. In 1997 for example only 58,7 % of the shootlength blighted (score 4,2) because of lowertemperatures in the greenhouse. This indicates that the result of all test plants is shifted in direction of resistancein this year. Malus robusla No. 5, the resistant standard variety, was not infected et all. The resistant varietiesshowed their resistance also after multiple inoculation of high concentrations of the new high virulent strains.The variation in the results was more caused by the differences in weather conditions than in different E.umylovora isolates used.
The variety ‘ Remo’ could be tested since 1991. In 2000 it was more attacked than in the other seasons (seore5,8). Possibly the combination of the different strains in the mixture is responsible for that. The results of theother cultivars are very similar in the years ofcvaluation.
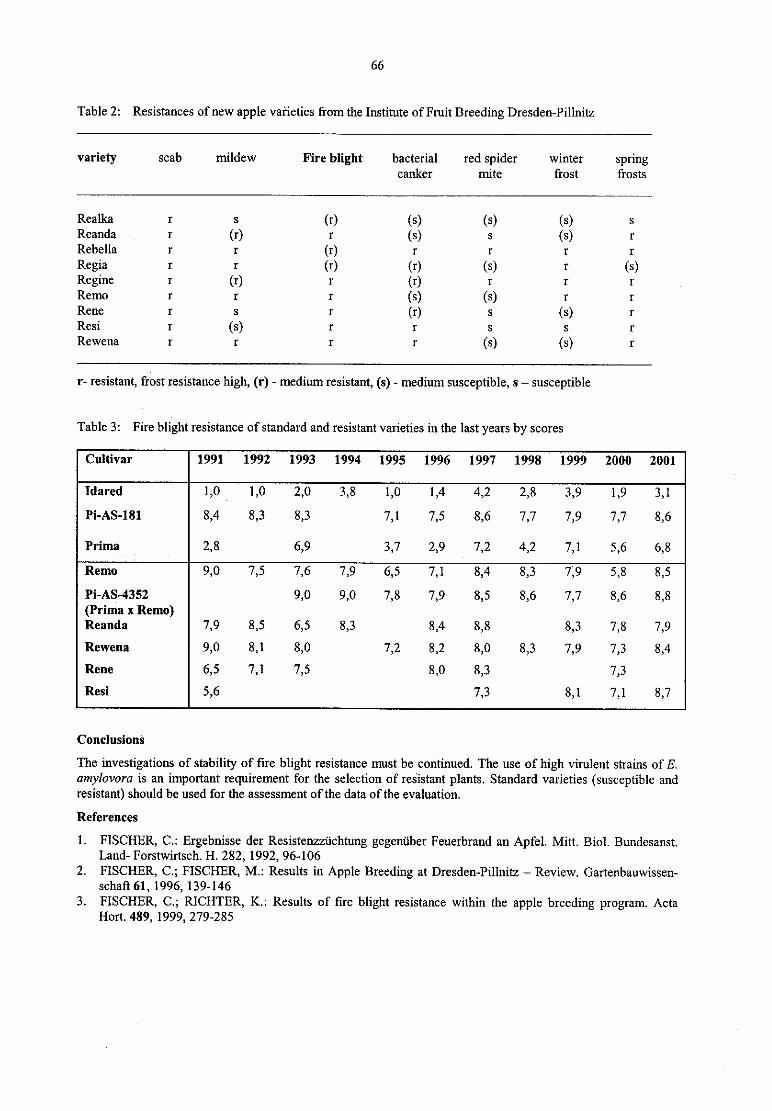
66
Table 2: Resistances of new apple varieties from the Institute of Fruit Breeding Dresden-Pillnitz
variety scab mildew Fire blight bacterial red spider winter springcanker mite frost frosts
Realka r s (r) (s) (s) (s) s
Reanda r (r) r (s) s (s) r
Rebella r r (r) r r r r
Regia r r (r) (r) (s) r (s)Regine r (r) r (r) r r r
Remo r r r (s) (s) r r
Rene r s r (r) s (s) r
Resi r (s) r r s s r
Rewena r r r r (s) (s) r
r- resistant, frost resistance high, (1') - medium resistant, (s) - medium susceptible, s — susceptible
Table 3: Fire blight resistance of standard and resistant Varieties in the last years by scores
1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001
Idared 1,0V
1,0 2,0 3,8 1,0 1,4 4,2 2,8 3,9 1,9 3,1
Pi—AS-181 8,4 8,3 8,3 7,1 7,5 8,6 7,7 7,9 7,7 8,6
Prima 2,8 6,9 3,7 2,9 7,2 4,2 7,1 5,6 6,8
Remo 9,0 7,5 7,6 7,9 6,5 7,1 8,4 8,3 7,9 5,8 8,5
Pi-AS—4352 9,0 9,0 7,8 7,9 8,5 8,6 7,7 8,6 8,8(Prima x Remo)Reanda 7,9 8,5 6,5 8,3 8,4 8,8 8,3 7,8 7,9
Rewena 9,0 8,1 8,0 7,2 8,2 8,0 8,3 7,9 7,3 8,4
Rene 6,5 7,1 7,5 8,0 8,3 7,3
Resi 5,6 7,3 8,1 7,1 8,7
Conclusions
The investigations of stability of fire blight resistance must be continued. The use of high virulent strains of E.amylavora is an important requirement for the selection of resistant plants. Standard varieties (susceptible andresistant) should be used for the assessment of the data of the evaluation.
References
1. FISCHER, C.: Ergebnisse der Resistenzzü chtung gegenü ber Feuerbrand an Apfel, Mitt. Biol. Bundesanst.Land« Forstwirtsch. H. 282, 1992, 96-106
2. FISCHER, C.; FISCHER, M.: Results in Apple Breeding at Dresden-Pillnitz — Review. Gartenbauwissen-schafl 61, 1996, 139-146
3. FISCHER, C.; RICHTER, K.: Results of fire blight resistance within the apple breeding program. ActaHort. 489, 1999, 279-285

67
Ecozonal variation in reaction of cassava genotypes from Togo to bacterial
blightA. Banitol‘2, K.E. Kpémouaz, KerstinWydra‘
1 Institute ofPlant Diseases and Plant Protection, University of Hannover, Germany, 2 Institut Togolais deRecherche Agronomique, ITRA, Lome, Togo
lntroduction
In the framework of a collabo ltiVC EU-financed project on “ Development and implementation of integratedcontrol methods for cassava bz terial blight in Africa” (1998-2001), a follow-up of a BMZ-fmanced researchproject at the lntemational Institute of Tropical Agriculture (IITA) (1994-1999), with partners in Germany,France (IRD, Montpellier), England (University of Bath), Benin (University ofBenin) and Togo (ITRA, Lome) -with the German partner (Universities of Hannover and Göttingen) being the coordinator — field studies were
conducted by ITRA in Togo, and control measures for cassava bacterial blight were developed, combined andtested under various agronomic and ecological conditions in a collaboration of all partners. Supplementarystudies on host plant resistance and detection methods were performed in collaboration between the advancedlaboratories and the African partners. Studies on the reaction of cassava genotypes to bacterial blight are subjectof the present paper.
Cassava (Manihot esculenta Crantz) is the most important root and tuber crop in the tropics where it oftenconstitutes the staple food. Cassava production is largely reduced due to the attack by pests and diseases. Morethan 30 bacterial, fungal, viral, virus-like, mycoplasrna and nematode pathogens are reported. Among thediseases, cassava bacterial blight, caused by Xanthomonas campestris pv. maniholis (Berthet-Bondar) Dye, is -
besides mosaic virus disease - important in different ecozones of West Africa (Wydra and Msikita 1998; Banitoet a1, 2001, Wydra and Verdier 2002), causing losses of more than 50% (Wydra and Rudolph 1999). Typicalsymptoms comprise angular leaf spots, leaf blight, leaf wilt, stem canker and shoot tip dieback. In Togo, thedisease was found - partly with high severity - in all ecozones including the rainforest, an ecozone in which it hasnot beengdescribed in Africa before (Banito et al. 2001). In Benin, Ghana, Cameroon and Nigeria the disease was
most severe in dry savanna zones, but did not occur in the rainforest in 1993-1995 (Wydra and Msikita 1998,Wydra and Verdier 2002).The most appropriate approach for controlling cassava bacterial blight is by growing resistant cultivars.Ecozone—dependent reactions of cassava genotypes to bacterial blight have been reported from Benin andNigeria (Fanou 1999, Wydra et a1. 2001, Zinsou et al. 2001). The present study presents results on theperformance of local and improved genotypes in Togo under various environmental conditions.
Materials and Methods
Four-week old plants from 24 local and improved cassava varieties from Togo (variety names in legend ofFig. 1) were stem-inoculated under controlled conditions in the glasshouse with 4 highly virulent strains ofX. campertris pv. manihotis from various geographic origins (GSPB- Gö ttinger Sammlung PhytopathogenerBakterien - 152 [originz Ibadan, Nigeria], GSPB 246 [On.ne, Nigeria], GSPB 255 [Cotonou, Benin], GSPB 290[Kampala, Uganda]) with 4 repetitions (plants) per strain and variety. The reaction of the genotypes was
evaluated in classes 1-5 with class 1: no symptom, class 2: one wilted leaf, elass 3: two to four wilted leaves,class 4: more than four wilted leaves, class 5: stem dieback. The class data of the four strains were summarizedto give a final evaluation class of the reaction of each variety to four strains.
The field trials were conducted in three eeozones: forest savanna transition and forest zones, characterized byone long rainy season (March — June), one short dry season (July-August), one short rainy season (September —
October) and one long dry season (November ~ March); savanna zone with one long rainy season (April —
September) and one long dry season (October — March). Data from the forest savanna transition and the forestzones are presented. The experimental design was an augmented complete randomized block design with 3blocks of 10 plots. Each plot consisted of 2 rows of 10 cassava plants representing one variety. The three cassavavarieties BEN86052, TMS30572 and Gbazekoute were used as checks and were replicated in each block. Eachbloek consisted of the three checks and 7 other varieties (non-replicated). One-month old plants of 24 local andimproved cassava varieties (Fig. 1) were inoculated withX. campestris pv. manihotis strain X27 from Togo threetimes in three-weekly intervals. Symptorns were evaluated 3 weeks after each inoculation and 6 and 12 monthsafter planting by recording the incidence of leaves with spots (S), blight (B) or wilted leaves (W) and the

68
percentage of shoots with dieback (D). The severity index (S1) was calculated according to the followingformula: (1 x S + 2 x B + 3 x W + 4 x D)/10. The severity indices (SI) for each evaluation date were used to
calculate the area under the disease progress curve (AUDPC) over the whole period (12 months): AUDPC = [(SI2+ S1 1)x21/2 + (Sl3+SI2)x2l/2 + (SI4+SI3)x120/2 + (SI5+SI4)x180/2], with S11, S12, S13, S14 and S15
representing the severity indices at dates 1, 2, 3, 4 and 5. AUDPC values were log-transformed and the analysisof variance was performed using the General Linear Model (~GLM) procedure in the SAS System (SAS, 1990,1997). The Student-Newman-Keuls test was used to compare the means of AUDPC values. Cassava genotypeswere classified into resistant (R), medium resistant (MR) and susceptible (S) groups.
Results and discussion
After stem-inoculation with 4 highly virulent X. campestri: pv. manihotis strains from various geographic origin,varieties Gbazekoute, CVTM4 and TMS91/02316 showed less symptoms than the other varieties (Fig. 1).
Fig. 1: Reaction of 24 cassava varieties to inoculation with four 4 highly virulent X campestris pv. manihotisstrains from different geographic origin (sum of four strains in symptom classes 1-5)(designation of columns according to legend from left to right; from left: column 2: Gbazekoute,column 12: CVTM4, column 14: TMS91/023l6)
after?’7
l'roMA2|9 ICAMEKOUN IMs3os72 ITDM.-us? Imsyz/0343
5312-524 '|'MS91/02316 I'|'r\rIS-ltlpl-13.5 ICVIM-6 IFETDNEGBODJI
I NAKOKO -TMS9HD}lfi UTU/\Kr\ Ü TÜ MA 159 I ANKRA
-'lMS92/lX)(i7 UMAINZ7 "lMS‘Jl/U2.l2Z DTMSDUNZQ
In field trials, reaction of varieties evaluated as the area under the disease progress curve (AUDPC) of severityindices over the growing period varied between the forest savanna transition and forest zones, and between years(Tab. IA, B). In the forest zone varieties CVTM4 (year 1 „ R“ , year 2 ,,MR“ ), Tuaka (.,MR“ , „ R“ ), TMS92/0326(„ MR“ , ,,R“ ) and TMS30572 (,,MR“ , ,,R“ ) revealed most suitable for this ecozone (Tab. 1A, Tab. 2). In theforest savanna transition zone varieties MAIN 27 („ R“ , ,,MR“ ), TMS92/0067 (,,R“ , ,,MR“ ), TMS9l/02316(,.MR“ , ,,R” ) and TOMA159 (,,MR“ , ,,R“ ) showed the lowest severity indices over the two years and can thus berecommended to farmers in this ecozone (Tab. 1B, Tab. 2). Variety Gbazekoute, with considerable resistanceagainst 4 X. campestris pv. manihotis strains (Fig. 1), revealed only medium resistant or susceptible under fieldconditions. Comparing results from 2 ecozones and 2 years (4 environments), no variety was found with a
resistant reaction in more than one environment, except TMS4(2)1425. In both ecozones, TMS4(2)1425 showedthe highest Variability between years, being among the resistant ones in the forest zone (year 1) and the forestsavanna transition zone (year 2), but among the highest susceptible in the forest zone (year 2) and the forestsavanna transition zone (year 1), thus showing highest genotype x environment interactions (Tab. 2). Sevenvarieties revealed resistance in at least one environment, and were medium resistant in the other 3 environments.
The highly variable and environmentally-dependent reaction of cassava to bacterial blight is typical of a
polygenic (horizontal) resistance. In spite of the uniformity of disease pressure achieved by artificial inoculation,varietal reaction generally varied highly from year to year, and from one zone to another. Similar observationson environmental influences on bacterial disease development were reported from other studies (Bayles et al.1985, Fanou 1999, Wydra et al. 2001, Zinsou et al. 2001). Defence reactions such as formation of vasculartylosis, which contribute to limit the progress of the pathogen in the tissue (Kpemoua et al. 1994) and otherfactors involved in the resistance to cassava bacterial blight (Cooper et al. 2001) might be influenced byenvironmental conditions. Studies on these interactions are not available.

69
Conclusions
No resistant variety was found among the 24 mainly grown local and improved cassava varieties in Togo. Due to
high genotype X environment interactions, only varieties suited to one ecozone can be recommended to farmersof the respective ecozone, such as Tuaka, TMS92/0326, TMS30572, CVTM4, MAIN27, TMS91/02316 andTOMA 159, which were generally medium resistant and revealed resistance in at least one environment. Amongthem, CVTM4 and TMS91/02316 were less susceptible to inoculation with 4 X. campestris pv. manihotis strainsfrom various geographic origin. Control strategies using medium resistant varieties should be complemented bycultural and agronomic measures (Fanou 1999, Wydra and Rudolph 1999). Further studies are rieeded in order tounderstand the background of the variability of the environment—dependent reaction of cassava genotypes.Tab. 1: Severity indices (AUDPC over the cassava growing period of 12 months) of cassava bacterial blight
on 24 cassava varieties planted in two ecozones (A: forest zone, B: forest savanna transition zone) intwo years, after inoculation with X. campestrir pv. manihotis strain X27
A: Forest zone
Year I Year 2
Variety AUDPC Ranking Variety AUDPC RankingCVTM4 l 102,0 R TUAKA lO52,0 RTMS4(2)1425 1 184,3 R TMS92/0343 1 136,9 RFETONEGBODJI 1209.9 MR TMS92/0326 1 166,9 RTMs92/0429 1267,2 MR TOMA289 11:30,3 RTMS30572 l30l,8 MR TMS30572 ll90,5 RTMS9l/02322 1323,0 MR TOMA159 l203,6 MR312-524 1340,4 MR TMS92/0429- 1205,9 MRMAIN27 l38l,6 MR TMS91/02316 l207,7 MRTMS92/0057 1389,4 MR MAIN27 l228,4 MRGBAZEKOUTE l393,6 MR CVTM4 1243,5 MRTOMA159 l446,9 MR 3lZ~524 l26S,6 MRTUAKA l458,6 MR TMS92/0067 1328.1 MRCAMEROON 1555.3 MR TMS92/0O57 l343,2 MRTOMA378 1566,2 MR TOMA378 1424/,4 MRTMS91/02316 1572,‘) MR GBAZEKOUTE 1434.2 MRTMS92/0326 1S75,6 MR CAMEROON l437,l MRTMS92/0343 1621,9 S TMS9l/02322 l449,8 MRANKRA 1639,‘) S NAKOKO 14:37,6 MRTOMA289 l768,2 S LAGOS l525,5 MRBEN86052 l805,5 S BEN86052 1533,6 SNAKOKO 1834,] S FETONEGBODJI 1545,5 STMS92/0067 1838,! S BORAM t553,1 SLAGOS 1905,9 s ANKRA I595,9 sBORAM 2638.1 S TMS4(2)1425 2346,2 S
B: Forest savanna transition zone
Year 1 Year 2
Variety AUDPC Ranking Variety AUDPC ‘ RankingMAIN27 827,6 R TMS91/02316 968,8 RTMS92/0067 979,5 R TMS4(2)1425 983,3 RNAKOKO I0l5,2 MR TOMA159 993,0 RFETONEGBODJI 1021,7 MR TMS92/0429 1030,8 MRTOMA159 1024,1 MR TUAKA 1061 ,7 MRBEN86052 1042.2 MR TOMA378 l068,6 MRTUAKA 1053,4 MR NAKOKO |071,0 MRTMS30572 1091,9 MR 312-524 l071,9 MRCAMEROON 1 132,8 MR CVTM4 lO79,5 MRTMS9l/02322 1162,41 MR FETONEGBODJI 1 107,7 MRTMS92/0326 1 194,5 MR TMS92/0343 1 t 14,6 MRTMS91/02316 1202,] MR TMS30572 1l15,5 MRTM S92/0429 l205,2 MR CAMEROON 1125,3 MRTMS92/0343 1214,5 MR MAIN27 1l27,6 MRTOMAZI9 12l8,3 MR TOMAZI9 ll33,0 MRCVTM4 l229,6 MR BEN86052 11359 MR312-524 1256,1 MR TMS92/0J26 t 136,5 MRTMS92/0057 l267,0 MR TMS92/0067 t 171,4 MRANKRA 1270.4 MR TMS91/O2322 l179,5 MRTMS4(2)1425 I 310,6 S GBAZEKOUTE 1189,5 MRTOMA289 l312,3 S TMS92/0057 1223.7 STOMA378 l356,4 S LAGOS 1236,1 SGBAZEKOUTE l460,7 s ANKRA 1307‚7 SLAGOS 1640.8 S TOMAZ89 l323,0 S

70
Tab. 2: Classification of cassava varieties according to their reaction (resistant and/or medium resistant) to
cassava bacterial blight, and identification of genotype x environment (2 ecozones, 2 years)interactions
Variety Forest {year 2) Forest {year 1) FSTgyear I) FST gyear 2)TUAKA R MR MR MR
TMS92/0343 R 5 MR MR
TMS92/0326 R MR MR MR
TOMAZ89 R S S S
TMS30572 R MR MR MR
Forest {year 1) Forest gyear Z) FST [year I ) FST {year 2)CVTM4 R MR MR MR
TMS4(2)l425 R s s R
FST {year I) Forest gyear 1) Forest gyear 2) FST {year 2)MAIN27 R MR MR MR
TMS92/0067 R MR S MR
FSTgyear Z) Forest gyear I) Forest {year 2) FSTgyear 1)TMS9l/02316 R MR MR MR
TOMA 159 R MR MR MR
References
1. BANITO, A.; KPEMOUA, K.; WYDRA, K.; RUDOLPH, K.: Bacterial blight of cassava in Togo: lts
importance, the virulence of the pathogen and the resistance of varieties. In: DeBoer, S. (Ed.): Plant
Pathogenic Bacteria, X. Int. Conf., Charlottetown, Canada, 2001, 259-264
2, BAYLES, M.B.; JOHNSON, W.M.: Growth of homologous and heterologous phytopathogenic bacteria in
cotton lines, resistant, susceptible, or immune to blight. Phytopathology 75, 1985, 515-519
3. COOPER, R.M.; KEMP, B.; GOMEZ-VASQUEZ, R.; BEECHING, J.R.: Pathogenicity and resistance in
Xanthomonas blight of cassava. In: DeBoer, S. (Ed.): Plant Pathogenic Bacteria, X. Int. Conf.,Charlottetown, Canada, 2001, 319-323
4. FANOU, A.: Epidemiological and ecological investigations on cassava bacterial blight and development of
integrated methods for its control in Africa. PhD thesis. University of Gö ttingen, Germany, 1999, pp 199
5. KPEMOUA, K.E.; BOHER, B.; NICOLE, M.; VERDIER, V.; GEIGER, J.P.: Cassava killer tyloses: a lethal
weapon. Colloque Int. de Cytologie ORSTOM/Socié té Francaise de Phytopathologie, Montpellier, 1994
6. WYDRA, K.; MSIKITA, W.: Overview of the present situation of cassava diseases in West Africa. In:
Akoroda, M.O.; Ekanayake, I.J. (Eds.): Root crops for poverty alleviation. Proc. 6"‘ Trienn. Symp. Int. Soc.
Trop. Root Crops Africa Branch (ISTRC-AB), Lilongwe, Malawi, 22.-28.10.1995, ISTRC, IITA &
Government ofMalawi, 1998, 198-206
7. WYDRA, K.; RUDOLPH, K.: Development and implementation of integrated control methods for majordiseases of cassava and cowpea in West-Africa. Gottinger Beitr. Land- und Forstwirtsch. in den Tropen und
Subtropen, 133, 1999, 174-180
8. WYDRA, K.; VERDIER, V.: Occurrence of cassava diseases in relation to environmental, agronomic and
plant characteristics. Agr. Ecosyst. Environ. 93 (1-3), 2002, 211-226
9. WYDRA, K.; FANOU, A.; DIXON, A.G.O. (x): Effect of cassava bacterial blight on cassava growth
parameters and root yield in different ecozones and influence of the environment on symptom development.In: Akoroda, M.O.; Ngeve, J.M. (Eds.): 7"‘ Trienn. Symp. Int. Soc. Trop. Root Crops Africa Branch
(ISTRC-AB), Cotonou, Benin, 1998, 2002, 562-569
10. ZINSOU, V.; WYDRA, K.; AGBICODO, E.; AHOHUENDO, B.; RUDOLPH, K.: Studies on the resistance
of cassava genotpyes to bacterial blight. In: DeBoer, S. (Ed.): Plant Pathogenic Bacteria, X. Int. Conf.,Charlottetown, Canada, 2001, 301-305

71
Evaluation of fire blight resistance of apple cultivars
K. Kä sa, M.G. Toth, M. Gö ndö r‚ M. Hevcsi
Szent Istvan University, Faculty ofHorticultural Science, Department of Fruit Science, Budapest,
Introduction
In Hungary apple is one of the most important fruit crops. Fire blight, a serious and economically importantbacterial disease of apple, is caused by Erwinia amylovora. Application of resistant varieties is the best methodfor controlling the disease. Two years following the appearance of fire blight in Hungary (Hevesi, 1996) we havestarted a research project with the aim to screen domestic gene sources containing traditional old Hungarianvarieties to serve as a basis for future breeding for resistance. In the 1990s an apple breeding program has startedin order to combine resistance to different pathogens e.g. apple scab (Venturia inaeqalis), mildew (Podospheraleucotricha) with high fruit quality in new cultivars grown in Hungary (G. Toth, 1993). During this breedingprogram those hybrids that have displayed considerable resistance to the fungal pathogens mentioned abovewere selected for further experiments on assessment of resistance to fire blight. The relation between pathogenand host is pivotal when evaluating susceptibility/resistance of the host. Evaluation of resistance has beentraditionally done by assessment of disease symptoms. In this study we are the first in Hungary to apply thequantitative method of bacterial multiplication kinetics. In addition, other methods, eg. evaluation of diseaseseverity (disease rating) and speed of disease development were also applied.
Materials and methods
Bacterial strains
For inoculation we used a mixture of three bacterial strains isolated from apple during different seasons andoriginating from different sites:
‚
E. amylovara 2 (Ea 2) Nyärlö rinc, 1996; E. amylovara 60 (Ea 60) Erd, 2000; E. umylovora 67 (Ea 67)Moriostorpalyi, 2000.
The virulence of the strains was checked continuously by either the apple slice test (bacterial suspension drippedonto the core of apple slices) or needle inoculation of immature pear fruits.
Test plants
The plants were grafted on rootstocks with 4 or 8 replicates and potted in spring. Plants were trained.in the
greenhouse with 3-4 shoots per individual produced.
Traditional old Hungarian cultivars:‘ Batul’ , ‘Baumann renet’ , ‘Beregi sovari‘ , ‘ Biirkormos renet’ , ‘ Cserepénya’ , ‘ Kanadai renet’ ,‘Kisasszony’ , ‘Nagy zoldalma’ , ‘ Nemes sovari', ‘ Ré zsa alma‘ , ‘ Sarga szépviragu‘ , ‘ Sikulai’ , ‘ Tafota’ ,‘ Vilmos renet’ , ‘ Zö ld sé vari’ , control] cultivars: ‘ Idared’ (susceptible), ‘ Remo’ , (resistant).
Hybrids:1-12-2/1; 1-139-4/2; 3-03-298; 3-03-408; 4-22-1; 4-22-61; 4-27-26; 139/103/1.
Fruits of international cultivars(/hybrids):‘ Remo’ , ‘ Rewena’ , ‘ Reanda’ , ‘ Reglindis’ , ‘Angold’ , ‘ Selena’ , ‘ Idared’ , ‘Liberty’ , ‘ Prima’ , ‘ Florina’ ,X3184, X3188, X3190, X4981, X6397.
Inoculations
Shoots were inoculated near the tip by a syringe. The bacterial suspension from an overnight culture, was
adjusted to 5 x 109 cfu/rnl with the mixture of strains.
Immature apple fruits were needle inoculated with bacterial suspension. Fruits were maintained in plastic boxes.
Methods of evaluation
Speed ofdisease developmentVisible symptoms (wilting and shoot necrosis) were assessed weekly. The proportion of total shoot length thatbecame necrotic was calculated in % (Gardner et al. 1980).MR — moderately resistant: 0 ~ 30 %; MS — moderately susceptible: 31-70 %; S w susceptible: 70 — . ...%

72
D[.l!3(I.l‘ I3 .revcrity
Data were evaluated by calculation 01' DR(Disease rating), described by Bertrand & Gottwald (1978) modiliedby Hevesi et al. (2000).(Disease rating: f (N1 x 11+ N2 x2) „ (N5 x 5
ZNN1 - 5: Number ofplants in each disease category.
Bacterial multiplicrztian kinetics
A serial tenfold dilution derived from a 1 cm length of stem 3 cm below the inoculation point. was plated.Number of bacterial colonies on agar plates indicated the host response.
MR — moderately resistant: 0-106 cells/1 cm; MS - moderately susceptible: 107 cells/l cm; S — susceptible: I0 3-
cellsll cm
Evaluation offruit infectionWater soaking and necrosis accompanied by the appearance ofbacterial ooze within 21 days after inoculationwas considered as a host response. The size of necrosis and the amount of bacterial ooze served for theevaluation of resistance/susceptibility.We evaluated the data by a three degree scale as follows:
0 — n0 symptom; l — Appearance of water soaked tissue, turned brown (5 mm in diameter) surrounded bya yellowish red halo 20 mm in diameter ; 2 v Area of necrotic tissue more than 40 mm in diameter.
Bacterial exudate on the skin less than 1 mm in diameter ; 3 v Spreading of symptoms to the wholesurface with brownish marbled color. Bacterial exudate on the skin more than 1 mm in diameter,
Statistical analysis of the data was done with Statgraph software.
Results
Among the 15 old Hungarian cultivars ‘Nagy zoldalma’ es ‘ Sikulai’ seemed to be moderately resistant based on
evaluation using the three above mentioned methods (Fig. 1.; Fig. 2.). The ‘ Remo’ cultivar did not show anyVisual symptoms and in host tissues we found minimal number of Erwinia amylavora cells. ‘ Batul’ cultivar did
not display any visual symptoms but host tissues contained high numbers of Erwinia amylovora cells, which
illustrates the importance of the multiplication kinetics method. Based on the results of the one year longexperiment described above we intend to conduct further research in order to confirm our present findings.
F ig. l: Evaluation of susceptibility of old Hungarian variesties to Hungarian Erwinia arnylovora strains.Disease severity .
Disease severity (2001)
5,0 — —— — -»« w—— —e— — — — —w— — — „ 3
.:=
ä ‘ m
E.*, 3,0 l,.,
‘c. l '3>
„ ...ä ä 2 2 ‘.—~1 z 3 - .25:
'9w;
_
i,, 1:1 :f
3
a? 2'» =:.= z’ : 2"»: F: .3 I-s? ‚A.:: a. 1: .= 1 ‚i z «.51
0,0...- as
n x. ' '
_
' w. - K. ‘o 5
_
&¢°Qg~"\‘q,<~°";°,,‘z>‘;°_,*o‘:‘”,’°4*2>‘;ogo“"”&e°‘$§\;_&¢‘¢e°6~°o*&{¢9 $":&\é":96‘:8,<°6° ‘ 0" A9 o" ° e '3’ 0° f’ ‘v9 .15’? 45°
'° -\*° « 4‚° «° 6° ’? s° 4’:¢$"° ~\,,° 4~ „ e: ‘ go we c. o ‚b Q- „ q~l- 61°‘ 6b abe) e
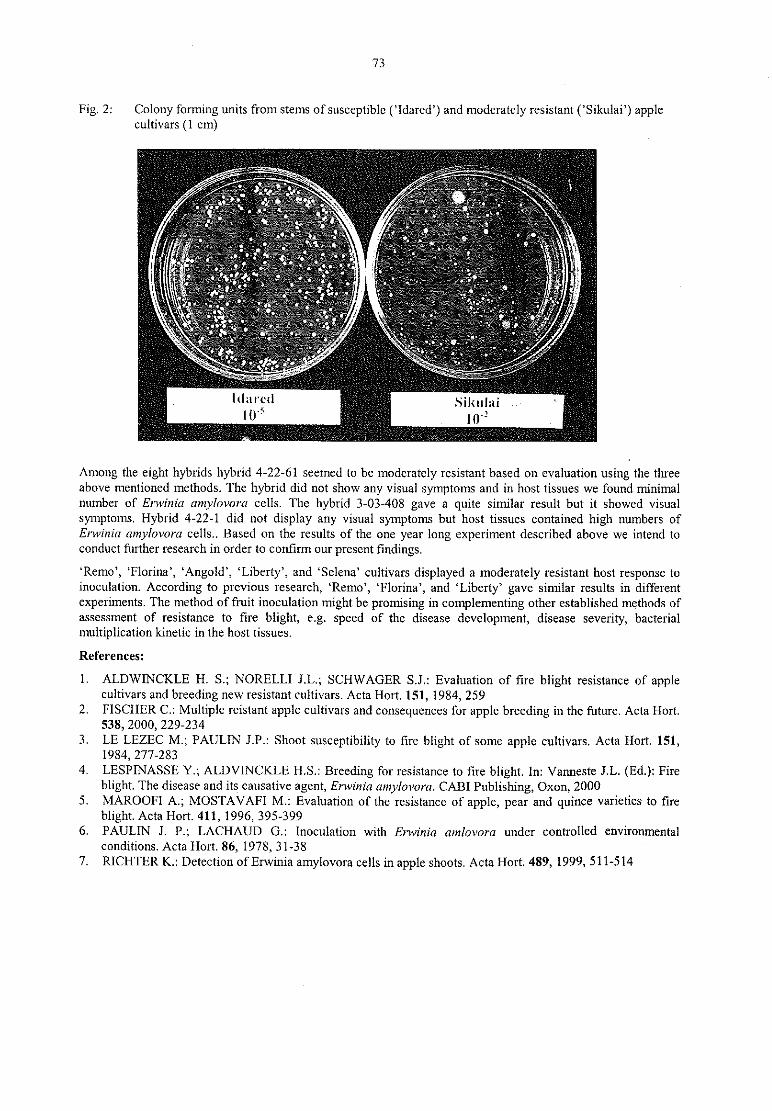
73
Fig. 2: Colony forming units from stems ofsusceptible (’ ldared’ ) and moderately resistant (‘ Sikulai’ ) apple‘
cultivars (1 cm) i
-‚ A
" ’ a:A‘ \:r>- :":{.:-»:-3g
"fix.
‚ v‘ 2-;3""3.'f-7 ' "-'5-,=7l“ ” .. \».
-'1 ‘ E l
C „ v: r:‘.j9.. —
.
‘
„ a. . - ‘z,‚ -:.;=;;.! — »
i
.5'3 .§.95''‘-5'*:-.j‘.7 -33., ‚I Ä .' L‘ “ Q, - 4.. 361%
— — —..-:—-~:. -1-— ‘ t *._ s.Ii „ ä 2“ .‘ 2
.
v
- L:‘ v r 1:1.‘ )! i» , *=-'._ i
‚ ‘.;«;!~~_. .
-
V l‘ E.‘ i. 3J)‘: 3:5! ‚ I; ‘ rA
p ‘ A ‘_-.3." P, .0) „
\ -~.‚ . . : w! .‚ ‚ 7., o ‚. .; - au.» -.
_‚g.
e 2* ‘ :a’s*>‚: '—‘¢'.~* ' o‘ .-"s
‘
--~.
‘ “ -'-
.
‘A‘—~'§\"7;‘-'§ '."',-' V‘ ‘ .19’ , :-.
‘:4 "
a?‘ ' ’" “ W: al.:" '
'
...» '-
' \‘:.,‘ € ;E.." f " '
‚
-.- w ' /
* ‘ z ...’ -.1 s." .‘.:"C'»-' ‘-‘ « v a.
‚ ._y
’ ‚V ‘m;
w”
3
l‘ l“ "f“ l Silurian’ ‘
,
10° 10"
Among the eight hybrids hybrid 4-22-61 seemed to be moderately resistant based on evaluation using the threeabove mentioned methods. The hybrid did not show any visual symptoms and in host tissues we found minimalnumber of Erwinia amylovom cells. The hybrid 3-03-408 gave a quite similar result but it showed visualsymptoms. Hybrid 4-22-l did not display any visual symptoms but host tissues contained high numbers ofEiwinia amylovora cells.. Based on the results of the one year long experiment described above we intend toconduct further research in order to confirm our present findings.
‘ Remo’ , ‘Florina’ , ‘ Angold’ , ‘ Liberty’ , and ‘ Selena’ cultivars displayed a moderately resistant host response toinoculation. According to previous research, ‘ Remo’ , ‘Florina’ , and ‘Liberty’ gave similar results in differentexperiments. The method of fruit inoculation might be promising in complementing other established methods ofassessment of resistance to tire blight, e.g. speed of the disease development, disease severity, bacterialmultiplication kinetic in the host tissues.
References:
1. ALDWINCKLE H. S.; NORELLI J.L.; SCHWAGER S.J.: Evaluation of fire blight resistance of applecultivars and breeding new resistant cultivars. Acta Hort. 151, 1984, 259
2. FISCHER C.: Multiple reistant apple cultivars and consequences for apple breeding in the future. Acta Hort.538, 2000, 229-234
3. LE LEZEC M,; PAULIN J.P.: Shoot susceptibility to tire blight of some apple cultivars. Acta Hort. 151,1984, 277-283
4. LESPINASSE Y.; ALDVINCKLE H.S.: Breeding for resistance to fire blight. ln: Vanneste .l.L. (Ed.): Fireblight. The disease and its causative agent, Erwinia amylovora. CABI Publishing, Oxon, 2000
5. MAROOFI A.; MOSTAVAFI M.: Evaluation of the resistance of apple, pear and quince varieties to fireblight. Acta Hort. 411, 1996, 395-399
6. PAULIN J. P.; LACHAUD G.: Inoculation with Erwinia amlovora under controlled environmentalconditions. Acta Hort. 86, 1978, 31-38
7. RICHTER K.: Detection ofErwinia amylovora cells in apple shoots. Acta Hort. 489, 1999, 511-514
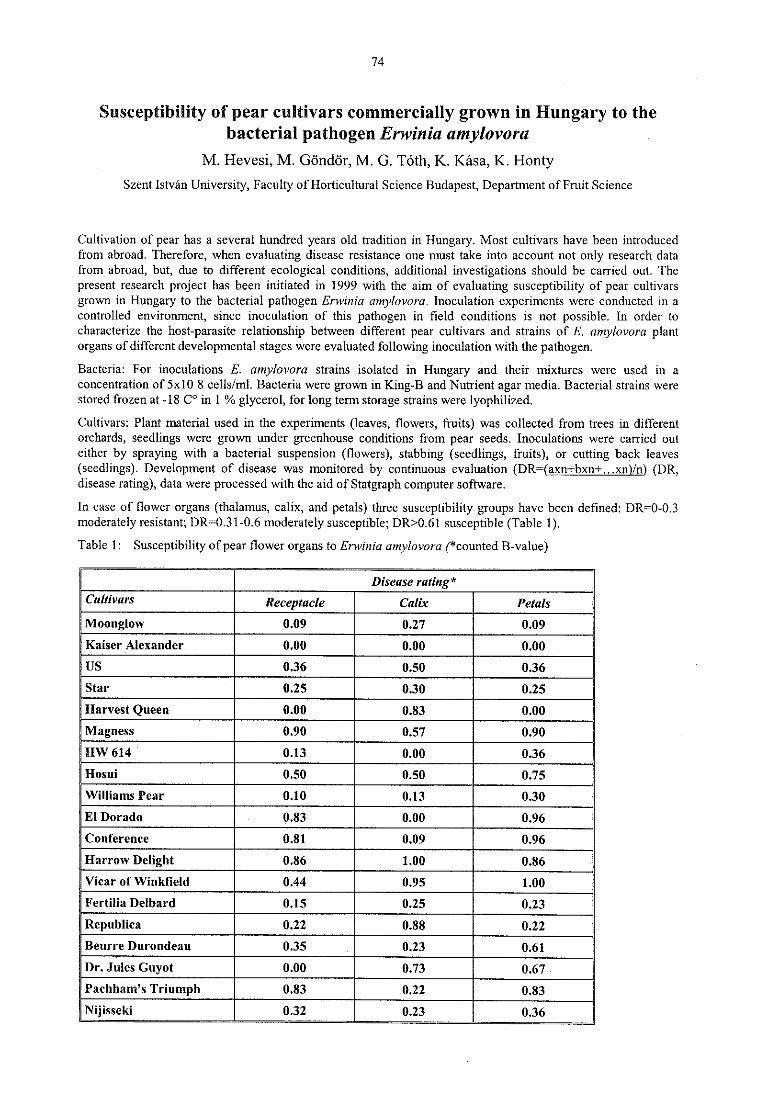
74
Susceptibility of pear cultivars commercially grown in Hungary to thebacterial pathogen Erwinia amylovora
M. Hevesi, M. Gö ndö r, M. G. Tö th, K. Käsa, K. HontySzent Istvan University, Faculty 0fHorti<:ultural Science Budapest, Department of Fruit Science
Cultivation of pear has a several hundred years old tradition in Hungary. Most cultivars have been introducedfrom abroad, Therefore, when evaluating disease resistance one must take into account not only research datafrom abroad, but, due to different ecological conditions, additional investigations should be carried out. Thepresent research project has been initiated in 1999 with the aim of evaluating susceptibility of pear cultivars
grown in Hungary to the bacterial pathogen Erwinia amylovora. Inoculation experiments were conducted in a
controlled environment, since inoculation of this pathogen in field conditions is not possible. In order to
characterize the host-parasite relationship between different pear cultivars and strains of E. amylovora plant
organs of different developmental stages were evaluated following inoculation with the pathogen.Bacteria: For inoculations E. amylovora strains isolated in Hungary and their mixtures were used in a
concentration of 5x10 8 cells/ml. Bacteria were grown in King-B and Nutrient agar media. Bacterial strains were
stored frozen at ~18 C° in 1 % glycerol, for long term storage strains were lyophilized.Cultivars: Plant material used in the experiments (leaves, flowers, fruits) was collected from trees in different
orchards, seedlings were grown under greenhouse conditions from pear seeds. Inoculations were carried outeither by spraying with a bacterial suspension (flowers), stabbing (seedlings, fruits), or cutting back leaves(seedlings). Development of disease was monitored by continuous evaluation (DR=(axn+bxn+...xn)/n) (DR,disease rating), data were processed with the aid of Statgraph computer software.
In case of flower organs (thalamus, calix, and petals) three susceptibility groups have been defined: DR=0-0.3
moderately resistant; DR=0.3l—0.6 moderately susceptible; DR>O,6l susceptible (Table l).Table l: Susceptibility ofpeat flower organs to Erwinia amylovora (*counted B-value)
?i‘J I
ijjjl
JmjJim1@@HW 614 “ J
EI Dorado Emma1mm
Harrow Deiigiiii
Feiiiiiii Deiiiaiii iRepubiica lBeurre Durondeau TDnii-iesciiyoi M.Pi-iiiii=iiii’ s Triumph
l
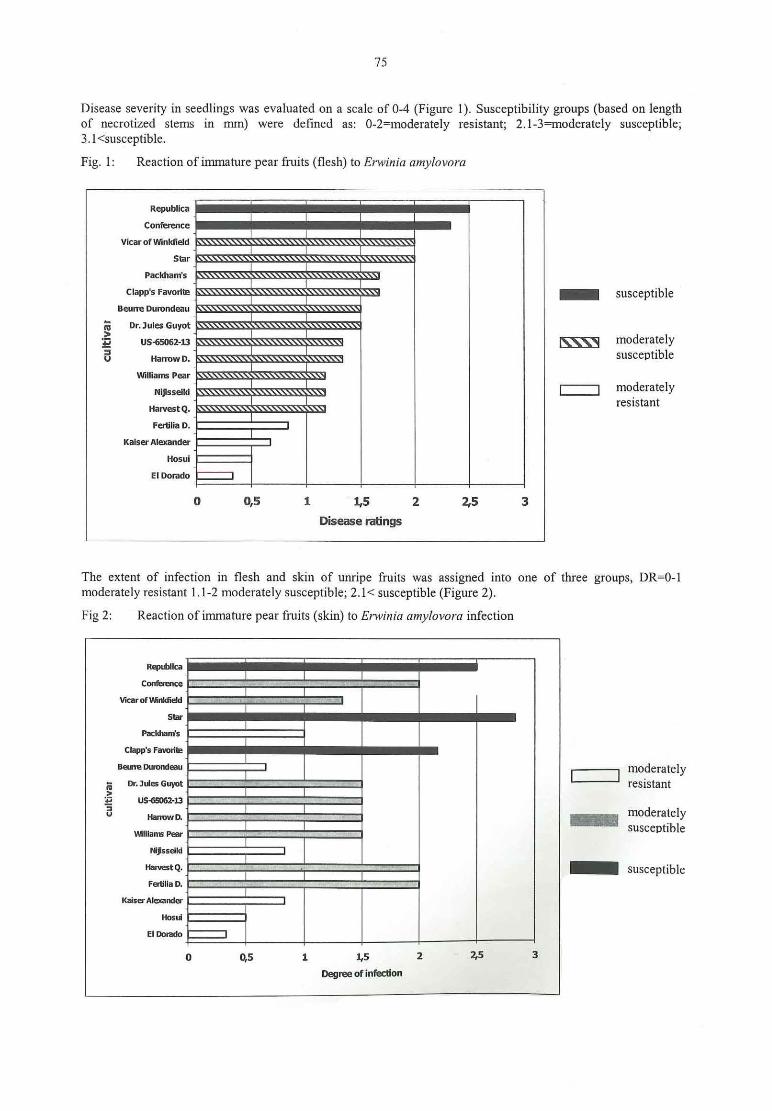
75
Disease severity in seedlings was evaluated on a scale of 0-4 (Figure 1). Susceptibility groups (based on lengthof necrotized stems in mm) were defined as: 0-2=moderately resistant; 2.l—3finoderately susceptible;3.l<susceptible.
Fig. 1: Reaction of immature pear fruits (flesh) to Erwinia amylovora
m
Republica
Confeau:e
_— _— —
Vicaroflmnkfield .\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\v
_-_-SBI‘ \\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\V
— — —Fackham's \\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\V
Cla ' F ‘ In \\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\ {b}uns avon -‘ susce 1 e
jjj Z PBeune Durondaau n\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\V
_-_-
m DY. Jules GING! \\\\\\\\\\\\\\\\\\\\\\\\\\\\\\\V> — -E usssosz-13 \\\\\\\\\\\\\\\\\\\\\\\\\\\V g\\V m0d€ 1’ al€ 1)’:: — —
_ SusceptibleU HanowD. .\\\\\\\\\\\\\\\\\\\\\\\\\\\\
_
-—Willians Pear u\\\\\\\\\\\\\\\\\\\\\\\V
Niissdki \\\\\\\\\\\\\\\\\\\\\\\\\ Z moderately— — resistant
HBNEIQ. .\\\\\\\\\\\\\\\\\\\\\\\\‘
__
-Fenilia D.T
‘KalsyklerandsT
_
—Hosm T
Elnomdo I0 0,5 1 1,5 2 2,5 3
Disease ratings
The extent of infection in flesh and skin of unripe fruits was assigned into one of three groups, DR=0—l
moderately resistant 1.1-2 moderately susceptible; 2.1< susceptible (Figure 2).
Fig 2: Reaction of immature pear fruits (skin) to Erwinia amy/avara infection
.: :jm
Rqublica— ‘ — —
Cunfaluicg %£
— -Vlcarolwiuifidd %
— ‘Star
— -PaclrhanfsT
— -cIapp's Favorit
—BanenuondeauT
I: moderately—.
5 DnJxisGuyut resistant— — —ä usewsz-13 — — —3
Nanvwn. j-zum: E m° de1'ateIY— — — '
\M|lians Pear Susceptible
—Missalri
—Havstq. — susceptible
‘ — — —Fatilla D.
‘Kaisa-AlexandaT
—uosuiT
ElDordo Io 0,5 1 1,5 2 2.5 3
Degneeofinfection
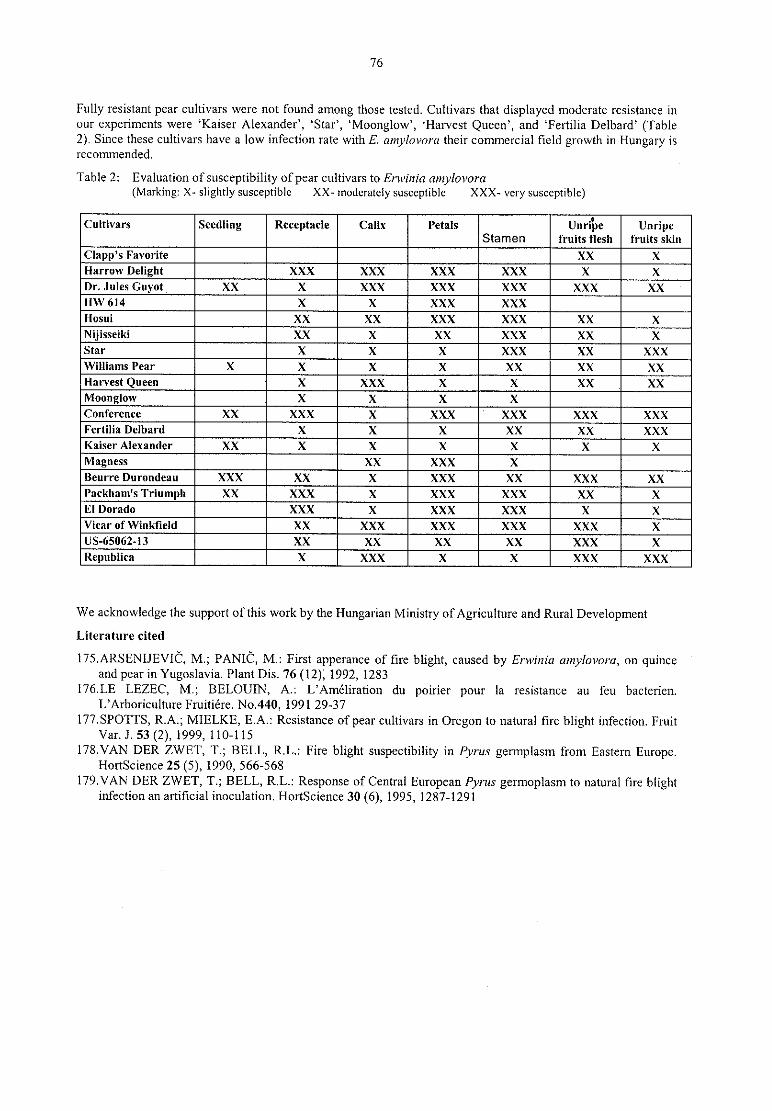
76
Fully resistant pear cultivars were not found among those tested. Cultivars that displayed moderate resistance inour experiments were ‘ Kaiser Alexander’ , ‘ Star’ , ‘Moonglow’ , ‘ Harvest Queen’ , and ‘ Fertilia Delbard’ (Table2). Since these cultivars have a low infection rate with E. amylovora their commercial field growth in Hungary isrecommended,
Table 2: Evaluation ofsusceptibility ofpear cultivars to Erwinia amylovora(Marking: X< slightly susceptible XX- moderately susceptible XXX- very susceptible)
Slamen fruits flesh fruits skin@--— — _——
— — —Em
——
‘— — —
—
-_-‘
—SzC
We acknowledge the support of this work by the Hungarian Ministry ofAgriculture and Rural DevelopmentLiterature cited
l75.ARSENIJEVIC, M.; PANIÖ , M.: First apperance of fire blight, caused by Erwinia amylovorn, on quinceand pear in Yugoslavia. Plant Dis. 76 (l2), 1992, 1283
176.LE LEZEC, M.; BELOUIN, A.: L’ Amé liration du poirier pour la resistance au feu bacterien.L’Arboriculture Fruitié rei No.440, l99l 29-37
17'/SPOTTS, R.A.; MIELKE, E.A.: Resistance of pear cultivars in Oregon to natural fire blight infection, FruitVar. J. 53 (2), 1999, 110-115
l78.VAN DER ZWET, T.; BELL‚ R.L.: Fire blight suspectibility in Pyrus germplasm from Eastern Europe.Hortscience 25(5), 1990, 566-568
l79.VAN DER ZWET, T.; BELL, R.L.: Response of Central European Pyrur germoplasm to natural fire blightinfection an artificial inoculation. Hortscience 30 (6), 1995, l287~l29l

77
The role of membrane-vesicle in pathogenesisIrmhild Schrö der, Athanassios Mavridis, Klaus Rudolph
Institute of Plant Pathology and Plant Protection, University of Gö ttingen, Germany
Introduction
Membrane-vesicles (MV) of many Gram-negative bacteria are released from the outer membrane into the
surrounding medium. MV play an important role in diseases caused by human pathogenic bacteria. The
endotoxins consisting of lipopolysaccharides (LPS) trigger fever, sepsis and the septic shock. In addition,membrane-vesicles transport several substances such as enzymes and nucleic acids which may have a strongimpact on disease development. The role of membranevvesicles of leaf-spot causing bacteria in pathogenesis hasnever been studied. Therefore, we infiltrated tomato leaves with MV from compatible [Pseudomonas syringaepv. tomato (Pst)], incompatible [Pseudomonas syringae pv. glycinea (<Psg)] as well as from saprophytic bacteria
[Pseudomonas fluorescenr (Pf)] to study the effects during the susceptible, resistant and immune reaction,respectively,Material and methods
Isolation of MV : Culture of bacteria up to an ODm= 1.4. Sterile filtration of the supernatant, addition of
sorbitol up to an osmotic pressure of 0.5 osmol/kg, ultracentrifugation 150000 g for l h.
Infiltration of MV into tomato leaves of cultivar "Lyconorma": MV (l0»fold concentrated from the
supernatant) were injected with a blunt Eppendorf combi-tip into tomato leaves until these were totally water-
soaked.
Extraction of the intercellular washing fluid (IWF): Tomato leaves were immersed in water within an
exsiccator and evacuated. By releasing the vacuum the leaves became fully water-soaked. During centrifugationofthe leaves at 3000 g for l0 min in falcon tubes on top ofa sieve the IWF was washed out ofthe leaves.
Detection of MV—LPS in the IWF of tomato leaves of cultivar "Lyconorma": Tomato leaves were infiltratedwith MV, incubated for several hours or days and then infiltrated with water to wash out the IWF. The IWFobtained was separated by SDS-PAGE and electroblotted onto a PVDF-membrane. The MV—LPS were detected
using polyclonal antibodies against purified LPS.
Determination of electrical conductivity: MV were infiltrated into the youngest fully developed tomato leaves
(cultivar “ Lyconorrna“ ). Disks of the treated leaves were cut and immersed in bidist. water. Afterwards theelectrical conductivity of the bathing solution was determined from 2-12 h.
Results
The MV were infiltrated into tomato leaves to study phenotypic as well as physiological reactions. Infiltration ofMV from the phytopathogenic bacteria Prt and Psg in high concentration resulted in a partial collapse of the
leatitissue, MV of the saprophytic Pfdid not show this effect. Analysis of MV constituents by SDS-PAGE and
gas-chromatography revealed the presence of phospholipids and lipopolysaccharides, in addition proteins with a
specific prominent band of 34 kDa. The MV of phytopathogenic bacteria Prt and Psg were regularly associatedwith hydrolytic enzymes such as pectinases and cellulases, but not the MV from the saprophytic bacteria Pj’.Therefore, the MV are regarded as virulence factors.
Infiltration of incompatible MV from Psg as well as from saprophytic Pf in tomato leaves induced a localresistance against a challenge inoculation with compatible strains of Pst. Compatible MV did not induce a
resistance. To quantify the resistance induction by MV, the multiplication of compatible (Pst) and saprophyticbacteria (Pf) in tomato leaves was determined after different pre-treatments. MV (Pst or Pj) were infiltrated into
tomato leaves 24 h before the 2nd inoculum of Pstvbacteria (104 cfu/ml).
Pre-treatment of tomato leaves with MV (Pst or Pf) (Fig.1) reduced the multiplication of Prt—bacteria, for
instance to 1x107 and 3x107 cfix/Cmz after pretreatment with Pst-MV and Pf~MV, respectively, in comparison
to 1x109 cfu/cmz in the water-control 7 days after treatment. Although the inhibition of Pst—bacteria by Pst- andPf-MV was nearly the same, the suppression of symptoms by Pf-MV was more evident.
A different effect of pretreatment with Pst-MV was recorded on the multiplication of Pf-bacteria in tomato
leaves (Fig.2). The bacterial multiplication within the first three days after inoculation was rather similar,However, the drastic decrease of the Pflpopulation in the leaves from 3-5 days after water-treatment did notoccur after pretreatment with Pst-MV,

78
Fig. 1: Multiplication of Pst-bacteria in tomato Fig. 2: Multiplication of Pf-bacteria in tomatoleaves after pretreatment with leaves after pre—treatment with Psf~MV
MV—(Psf or Pf) or water or water
7
cfu/ml '’
cfu7m10000000000 r 7" " r * * '
10000000 . ‚ „ „ ‚ — — n ‚ „ „ „ „ — ‚
100000000 /./*'\.l l° ° ° ° ° ° -41 l
- 100000 /' o l
1000000_
'‘ 10000
l
00010000 — O— water/Pst bacteria g 1100 _._Wa1er/Pfbameria
l
100— I— MV Pst/Pst bacteria l_... My pf/PS‘ bamefia i
10 — I— ' MV Psl/Pf bacteria1 ‚1
O 1 2 3 4 5 6 70 2 dävs e a
V 7
Wgays
The specificity of MV—LPS was tested in the compatible and incompatible interaction. IWF was obtained fromtomato leaves three hours and 1, 2, 3, 4, 6 days after infiltration with Prr- and Psg-MV to study the attachment or
binding of MV. The recovery rate of the LPS from treated leaves differed very much between Pst~MV(compatible pathogen) and Psg-MV (incompatible pathogen). Until 6 days after infiltration the Pst-MV could begained back out of the intercellular space of tomato leaves as shown by the typical ladder-like patterns afterelectrophoresis . Obviously the Psi-MV did not firmly attach to the plant cell walls, whereas the Psg-MV were
only detectable in the IWF at a very early stage (3 h after infiltration). One day after infiltration the Psg-MV—LPS were hardly visible in the IWF. We concluded from these results that the MV of incompatiblepseudomonads were strongly attached by tomato leaf cell walls, so that it was nearly impossible to remove themby flushing the intercellular space with water. This specificity was also detected on the race/cultivar level of PsiR 0 and R l on cultivar "Ontario no. 7710".
Several parameters changed during this "protection effect". One mechanism involved was the ion-efflux (Fig. 3).
Fig. 3: Ion efflux out of tomato leaves after treatment
with MV or water
180 >
E160
-0- P.s.tomat0m 140 I 4I— P.s.g|ycinea l21120 I —fi— P.fluorescens l
E 100 1 — I— contro|— l
ä so g |g eo
.
°40 “20 l
0
O 2 4 6 8 10 12.hours i
Two hours after infiltration of MV from the phytopathogens Pst and Psg an enhanced ion-efflux out of tomatoleaves was observed, but not when MV from Pf or water were infiltrated. The ions in the bathing solution
consisted predominantly of Kl’ and Mg2+.
Discussion
Our investigations indicate a double function of membrane-vesicles of pseudomonads in host! pathogenrelationships. On one side, they may act in the incompatible interaction as elicitor for resistance (SCHRÖ DER,2000) but on the other side, the released MV may increase virulence. In compatible interactions MV appear toincrease virulence of the pathogens, whereas in incompatible interactions MV induce resistance againstsubsequent bacterial inoculations.
By extraction of MV~LPS from treated leaves we could demonstrate, that MV of compatible and incompatiblebacteria were not catabolized but interestingly showed a strong specificity in induction of resistance. MV ofincompatible bacteria (Psg) attached to the cell~walls of tomato leaves where they triggered specific resistantreactions, such as the production of superoxide radicals, ion efflux, synthesis of defense related proteins cm.

79
(SCHRÖ DER e! a], 2000). In contrast, MV of compatible bacteria (Pst) did not attach to plant cell-walls thoughthey are in the same way able to trigger above mentioned reactions. It was shown that the LPS of Pst interact
synergistically with pectins from tomato leaves (GROLMS & RUDOLPH, 1997), so that the resulting gel
supports bacterial multiplication in planta. It is assumed that the low inhibition of Pst-multiplication in planta bycompatible MV (Pst) is an artefact because of too high concentrations of MV and the injection procedure. Thebacterial growth determined in tomato leaves confirmed the apparent symptom development in the differentexperimental variants. But only Pf-MV were able to suppress symptom development of Pst-bacteria. Themechanism by which MV induce resistance is unknown. The LPS—component of the MV appears to be thesubstance responsible for resistance induction, because similar differential effects have been reported for purified
LPS—preparations (LAUX et al., 1998). Interestingly pre—treatrnent of tomato leaves with Pst-MV led to a
supporting effect for Pf—bacteria. The finding that Pf-populations did not decrease after a short multiplicationphase may have been due to the gel formed from Pst-MV with tomato pectins so that Pf-cells were preventedagainst desiccation.
Summary- Treatment of tomato leaves with membrane-vesicles (MV) of phytopathogenic pseudomonads induced an
ion-efflux out of plant cells.- Incompatible MV attached strongly to plant cell walls whereas compatible MV were not bound and could be
regained from the intercellular washing fluid (IWF).- MV of saprophytic bacteria (Pf) or incompatible bacteria (Psg) induced a resistant reaction in tomato leaves
so that symptom induction by P. syringae pv. tomato was suppressed. MV of compatible bacteria (Pst)reduced symptom development only weakly.
— After resistance induction by MV in tomato leaves, multiplication of subsequently inoculated bacteria was
considerably lower than in the water control.- Pre-treatment with compatible MV enabled P. fluorescens to survive more than 4 days in inoculated leaves.
References
l80.GROLMS, U.; RUDOLPH, K.: Rheological interactions between lipopolysaccharides of Pseudomonas
syringae pathovars and plant polysaccharides. In: Rudolph, K.; Burr, T..I.; Mansfield, J.W.; Stead, D.;Vivian, A.; von Kietzell, J
. (Eds): Pseudomonas Syringae pathovars and related pathogens, KluwerAcademic Publ., 1997, 364-369
181.LAUX, P.; MULLER, P.; RUDOLPH, K.: Interactions between bacterial lipopolysaccharides and plantpectins - a mechanism which may determine host/parasite relations in bacterial plant diseases. In:
Mahadevan, A. (Ed.): Plant Pathogenic Bacteria, Proc. 9 "' Int. Conf., Centre for Advanced Study in Botany,University of Madras, India, 1998, 563-568
l82.SCHRODER, I.; Zur Rolle der Membranvesikel von Pseudomonas syringae als Resistenzinduktoren undTransportvehikel fiir Virulenzfaktoren bei Nicoriana tabacum und Lycopersicon esculentum. Ph.D. Thesis,University of Gö ttingen, 2000
l83.SCl-IRODER, I.; KUHLMANN, M.; DRÖ GE—LASER, W.; HOPPERT, M.; RUDOLPH, K.: Membranevesicle mediated transport of virulence-factors and of elicitors for defense-response. In: de Boer, S.H. (Ed):Plant Pathogenic Bacteria, Proc. 10"‘ Int. Conf. on Plant Pathogenic Bacteria. Kluwer Academic Publ., 2000,176-179
AcknowledgementWe thank Dr. Frank Rabenstein, Aschersleben/Germany for generating polyclonal antibodies against LPS of thePi'r- and Psg-bacteria.
80
80' Symposium on „ New Aspects of Resistance Research on Cultivated Plants“
Bacterial Diseases
Novemberl5-16, 2001 Aschersleben, Germany
List 01' Participants
Abdel-Rehim Khalid lnstitute ofPlant Pathology and Plant Protection, [email protected] ofGtittingen, Grisebachstr. 6, D-37077Gtittinen,German
Barchend Gudrun Federal Centre for Breeding Research on Cultivated Plants [email protected] for Resistance Research & Pathogen DiagnosticsPO Box 1505, D-06435 Aschersleben, Germany
Karl-von-Frisch-Strassc, D-35043 Marbur, German deEvtushenkov Anatoli Faculty ofBiology, Bclarusian State University,22005O [email protected]|.by
Minsk, BelarusGeider Klaus Max-Planck-Institut fü r Zellbiologie, [email protected]
Rosenhof, D-68526 Ladenburg, GermanyGriesbach Erika Federal Centre for Breeding Research on Cultivated Plants [email protected]
PO Box 1505, D-06435 Aschersleben, GermanHevesi Maria Szent Istvan University, Faculty of Horticultural [email protected]
Science Budaest, Deartment ofFruit Science'
Jock Susanne Max-Planck-lnsiitut fü r Zellbiologie, [email protected]ü hne Thomas Federal Centre for Breeding Research on Cultivated Plants [email protected]
Institute for Resistance Research & Pathogen DiagnosticsPO Box 1505, D-06435 Aschersleben, Germany
Laux Peter Federal Biological Research Centre for Agriculture and P.Laux.Biocontr0l.bba@t-Forestry (BBA), Institute for Biological Control, online.dcDarmstadt, Germany
Kö nistr. 47, D-25709 Marne, GermanMarthe Frank Federal Centre for Breeding Research on Cultivated Plants t'[email protected]
lnstitute ofHorticultural Crops, Neuer Weg 22/23,D-06484 Quedlinbur, German
Mavridis Athanassios lnstitute ofPlant Pathology and Plant Protection, [email protected] ofGottingcn, Grisebachstr. 6, D-37077Gö ttin en,Gcrmany
Miersch Ulrike GZG Saaten AG Marne, [email protected]“ -Nachtigall Marion Federal Centre for Breeding Research on Cultivated Plants [email protected]
Institute for Resistance Research & Pathogen DiagnosticsPO Box 1505, D-06435 Aschersleben, GermanyAufder Alten Burg 15, 06449 Aschersleben —
Thomas Federal Centre for Breeding Research on Cultivated Plants [email protected] of Horticultural Crops, Neuer Weg 22/23,D-06484 Quedlinbur, German
Klaus Elsner pac® Jungpflanzen, [email protected] Straß e 146, D-0l279 Dresden, German
Proeseler Federal Centre for Breeding Research on Cultivatcd Plants [email protected] Box 1505, D-06435 Aschersleben, German
Rabenstein Frank Federal Centre for Breeding Research on Cultivated Plants f.rabenstein@baf7,.deInstitute for Resistance Research & Pathogen DiagnosticsPO Box 1505, D-06435 Aschersleben, GermanFederal Centre for Breeding Research on Cultivated Plants r.l<laus@baf/..dePO Box 1505, D-06435 Aschersleben, Germany

81
University ofGöttingen, Grisebachstr. 6, D-37077Gö ttin en, German
Schroeder Irmhild lnstitute ofPlant Pathology and Plant Protection, [email protected] ofGottingen, Griscbachstr. 6, D-37077
G6ttinen,German
Warwick CV35 9EF, United Kindom
Venkatesh Balakrishnan Institute ofPlant Pathology and Plant Protection, [email protected] ofGö ttingen, Grisebachstr. 6, D-37077
Gö ttinen, Germany
Graf~Li n e-Str. 1, D-18059 Rostock, German
AA.Enkhuizen, The Netherlands
Institute for Stress Physiology and Quality of Raw
Materials, D ~l8l90 Gross Lü sewitz, German .
Wiedemann Wolfram Sä chsische Landesanstalt fiir Landwirtschaft Fachbereich Wolfram.Wiedemann@
Stü beIaIIee 2, D-01307 Dresden, Gennani:of Hannover, German
i
Zeller Klaus Federal Biological Research Centre for Agriculture and K.Zeller.biocontrol.bba@t—Forestry (BBA), Institute for Biological Control, online.deDarmstadt, German
Zimnoch— Eva Plant Breeding and Acclimatization Institute, Mlochö w E.Zimn0ch-Guz0wska@Guzowska Research Center, 05-832 Mloché w, Platanowa str.l9, ihar.edu.pl
Poland
Zinsou Valerian Institute of Plant Pathology and Plant Protection, [email protected] ofGö ttingen, Grisebachstr. 6, D—37077
Gö ttinen, German

l
Beiträge zur Zü chtungsforschung
Bundesanstaltfü r Zü chtungsforschung an Kulturpflanzen lHinweise fü r die Erarbeitung der Druckvorlagen .
l
Um die Druckvorlagen fü r die Zeitschrift „ Beiträ ge zur Zü chtungsforschung — Bundesanstalt fü r
Zü chtungsforschung an Kulturpflanzen“ mö glichst rationell und in einheitlicher Form fü r den Druck aufbereiten zu
kö nnen, wird um die Beachtung nachfolgender Hinweise gebeten:
Bei Verwendung von PC-Technik bzw. Ü bergabe von Texten (einschließ lich Grafiken und Tabellen) auf Diskette:
o Textverarbeitungsprogramm: Word fü r WindowsBei Verwendung anderer Programme bitte den Text zusä tzlich im ASClLFonnat beilegen.
c Papierformat: A4 (2 I 0 x 297), Hochformat
o Seiteneinrichtung: Seitenrä nder: oben 2,5; unten 2,0; links 2,5, rechts 2,5
v Schriftart: Times New Roman
- Schriftgrö ß e: l0 (auch Ü berschriften etc.)
o Disketten: mö glichst 3,5”
- Ü berschrift vom Text durch eine Leerzeile trennen. Den Text einesjeden Absatzes fortlaufend schreiben
(Kein Enter innerhalb!) und mit einem Absatzende (Enter-Taste) abschließ en.
- Der Text ist formatiert zu speichern.
c Fü r Anstriche im Text die automatische Aufzä hlungsfunktion von Word fü r Windows nutzen, um dieEinheitlichkeit zu gewä hrleisten. Keinesfalls mit Bindestrich und Leerzeichen arbeiten.
I Lateinische Bezeichnungen der Kulturarten und Erreger kursiv fonnatieren.
0 Abbildungen — aufgrund der Vervielfaltigungstechnik bitte vorwiegend Schwarz-weiß -Bilder verwenden.Vorteilhaft ist, die Abzü ge bereits in der endgü ltigen Grö ß e vorzubereiten. Abbildungsunterschriften extra wie
folgt vorbereiten:Abb. 6: Anfalligkeiten abgeschnittener
o Fü r Tabellen die Tabellenfunktion von Word fü r Windows nutzen. Die Tabellen bitte druckfertig formatieren,Tabellenü berschriften wie folgt vorbereiten:
Tab. 3: Untersuchte Sorten
o Das Verzeichnis fü r Publikationen bzw. Literatur sollte wie folgt vorbereitet werden:
Autoren : Titel des Beitrages. Verö ffentlichungsorgan. Jahrgang (fett), Jahr, SeitenzahlenMUSTERFRAU, K.; MUSTERMANN, L.: Die Kartoffel des Nordens.Kartoffelbau 43, 1994, 125-128
Bei der Ü bergabe von bereits mit einer Schreibmaschine geschriebenen Texten (einschließ lich Grafiken und
Tabellen) sind, so weit es mö glich ist, die o.g. Hinweise zu berü cksichtigen.
Seitenzahlen bitte nicht eintragen
Weiß es normales Papier verwenden
Related Documents

![Grundlagen des Agglomerierens - OpenAgrar · 2016. 9. 12. · Agglomerieren z. B. von Instant-Produkten einsetzen [20]. 3.2.3 Kristallisation geloster Stoffe Die Kristallisation geloster](https://static.cupdf.com/doc/110x72/60a8fde0d5507e2608433fa6/grundlagen-des-agglomerierens-openagrar-2016-9-12-agglomerieren-z-b-von.jpg)









