Yeasts isolated from three varieties of grapes cultivated in different locations of the Dolenjska vine-growing region, Slovenia Peter Raspor ⁎ , Damjana Miklič Milek, Julijana Polanc, Sonja Smole Možina, Neža Čadež University of Ljubljana, Biotechnical Faculty, Food Science and Technology Department, Jamnikarjeva 101, 1000 Ljubljana, Slovenia Received 11 February 2005; received in revised form 30 November 2005; accepted 4 January 2006 Abstract The number and diversity of yeasts on grape berry surfaces are influenced by several factors, such as grape variety, degree of grape maturity at harvest, climatological conditions, geographic location, physical damage of grapes, the intensity of pest management etc. Cviček is a typical Slovene wine, which has obtained a special protection under the Slovene Wine Law for its geographical origin. This blended red wine is produced from different grape varieties (Vitis vinifera L.), mostly from red grapes of Žametovka and Modra frankinja and from white grapes of Kraljevina. The aim of this study was to evaluate the impact of geographical locations in the Dolenjska vine-growing region and to obtain precise information about the influence of different grape varieties on the composition of yeast community on grape berries. The restriction fragment length polymorphism of PCR-amplified fragments from the rDNA gene cluster (PCR RFLP of rDNA) has been used for the differentiation of yeast species. The standard identification procedure has been performed on representative strains that shared identical RFLP profiles. The number of yeasts and yeast species isolated varied according to different grape varieties, Žametovka, Modra frankinja and Kraljevina (V. vinifera L.) and according to different sampling location. On the surface of grape berries 13 different yeast species have been identified. Saccharomyces cerevisiae has not been found. © 2006 Elsevier B.V. All rights reserved. Keywords: Yeast community; Saccharomyces cerevisiae; Grape berries; Vine variety; PCR-ribotyping 1. Introduction The fermentation of grape must into wine is a complex microbiological process characterized by the presence of a large number of different microorganisms. The process includes ecological and biochemical interactions between yeast species, species of filamentous fungi, lactic acid bacteria, acetic acid bacteria and their viruses (Fleet, 1993). The composition of yeast population on grape berries plays an important role in wine fermentations, as different genera, species and strains with their metabolic activity influence the sensory quality and organoleptic characteristics of wine (reviewed by Pretorius, 2000; Fleet et al., 2002). Yeasts associated with grape/wine ecosystem are usually classified in 15 different yeast genera: Brettanomyces/Dekkera, Candida, Cryptococcus, Debaryo- myces, Hanseniaspora/Kloeckera, Kluyveromyces, Metschniko- wia, Pichia, Rhodotorula, Saccharomyces, Saccharomycodes, Schizosaccharomyces, and Zygosaccharomyces (Kurtzman and Fell, 1998). Various authors isolated yeasts from vineyard ecosystems such as soil, air, vine leaves, grapes and different animal vectors and also from the winery ecosystems such as the equipment, floor, walls, ceilings, vats, utensils (Barnett et al., 1972; Davenport, 1974; Rosini et al., 1982; Longo et al., 1991; Yanagida et al., 1992; Martini et al., 1996; Mortimer and Polsinelli, 1999). The composition and number of indigenous yeasts on grape berries are influenced by several factors, such as grape variety, degree of grape maturity at harvest, climatological conditions, geographic location, physical damage of the grapes and the intensity of pest management (Barnett et al., 1972; Davenport, 1974; Rosini et al., 1982; Longo et al., 1991; Yanagida et al., 1992; Schütz and Gafner, 1994; Martini et al., 1996; De La Torre et al., 1999; Guerra et al., 1999; Mortimer and Polsinelli, 1999). The population density ranges from 10 2 to 10 4 Colony Forming Units (CFU) per ripe berry, depending on the above mentioned factors (Fugelsang, 1997). The yeasts of the Kloeckera and Hanseniaspora genera are the predominant International Journal of Food Microbiology 109 (2006) 97 – 102 www.elsevier.com/locate/ijfoodmicro ⁎ Corresponding author. Tel.: +386 1 423 11 61; fax: +386 1 257 40 92. E-mail address: [email protected] (P. Raspor). 0168-1605/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.ijfoodmicro.2006.01.017

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

biology 109 (2006) 97–102www.elsevier.com/locate/ijfoodmicro
International Journal of Food Micro
Yeasts isolated from three varieties of grapes cultivated in different locationsof the Dolenjska vine-growing region, Slovenia
Peter Raspor ⁎, Damjana Miklič Milek, Julijana Polanc, Sonja Smole Možina, Neža ČadežUniversity of Ljubljana, Biotechnical Faculty, Food Science and Technology Department, Jamnikarjeva 101, 1000 Ljubljana, Slovenia
Received 11 February 2005; received in revised form 30 November 2005; accepted 4 January 2006
Abstract
The number and diversity of yeasts on grape berry surfaces are influenced by several factors, such as grape variety, degree of grape maturity atharvest, climatological conditions, geographic location, physical damage of grapes, the intensity of pest management etc. Cviček is a typicalSlovene wine, which has obtained a special protection under the Slovene Wine Law for its geographical origin. This blended red wine is producedfrom different grape varieties (Vitis vinifera L.), mostly from red grapes of Žametovka and Modra frankinja and from white grapes of Kraljevina.The aim of this study was to evaluate the impact of geographical locations in the Dolenjska vine-growing region and to obtain precise informationabout the influence of different grape varieties on the composition of yeast community on grape berries. The restriction fragment lengthpolymorphism of PCR-amplified fragments from the rDNA gene cluster (PCR RFLP of rDNA) has been used for the differentiation of yeastspecies. The standard identification procedure has been performed on representative strains that shared identical RFLP profiles. The number ofyeasts and yeast species isolated varied according to different grape varieties, Žametovka, Modra frankinja and Kraljevina (V. vinifera L.) andaccording to different sampling location. On the surface of grape berries 13 different yeast species have been identified. Saccharomyces cerevisiaehas not been found.© 2006 Elsevier B.V. All rights reserved.
Keywords: Yeast community; Saccharomyces cerevisiae; Grape berries; Vine variety; PCR-ribotyping
1. Introduction
The fermentation of grape must into wine is a complexmicrobiological process characterized by the presence of a largenumber of different microorganisms. The process includesecological and biochemical interactions between yeast species,species of filamentous fungi, lactic acid bacteria, acetic acidbacteria and their viruses (Fleet, 1993). The composition ofyeast population on grape berries plays an important role inwine fermentations, as different genera, species and strains withtheir metabolic activity influence the sensory quality andorganoleptic characteristics of wine (reviewed by Pretorius,2000; Fleet et al., 2002). Yeasts associated with grape/wineecosystem are usually classified in 15 different yeast genera:Brettanomyces/Dekkera, Candida, Cryptococcus, Debaryo-myces, Hanseniaspora/Kloeckera, Kluyveromyces,Metschniko-wia, Pichia, Rhodotorula, Saccharomyces, Saccharomycodes,
⁎ Corresponding author. Tel.: +386 1 423 11 61; fax: +386 1 257 40 92.E-mail address: [email protected] (P. Raspor).
0168-1605/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ijfoodmicro.2006.01.017
Schizosaccharomyces, and Zygosaccharomyces (Kurtzman andFell, 1998). Various authors isolated yeasts from vineyardecosystems such as soil, air, vine leaves, grapes and differentanimal vectors and also from the winery ecosystems such as theequipment, floor, walls, ceilings, vats, utensils (Barnett et al.,1972; Davenport, 1974; Rosini et al., 1982; Longo et al., 1991;Yanagida et al., 1992; Martini et al., 1996; Mortimer andPolsinelli, 1999).
The composition and number of indigenous yeasts on grapeberries are influenced by several factors, such as grape variety,degree of grape maturity at harvest, climatological conditions,geographic location, physical damage of the grapes and theintensity of pest management (Barnett et al., 1972; Davenport,1974; Rosini et al., 1982; Longo et al., 1991; Yanagida et al.,1992; Schütz and Gafner, 1994; Martini et al., 1996; De LaTorre et al., 1999; Guerra et al., 1999; Mortimer and Polsinelli,1999). The population density ranges from 102 to 104 ColonyForming Units (CFU) per ripe berry, depending on the abovementioned factors (Fugelsang, 1997). The yeasts of theKloeckera and Hanseniaspora genera are the predominant

98 P. Raspor et al. / International Journal of Food Microbiology 109 (2006) 97–102
species on the surface of the grape, accounting for 50–75% ofthe total yeast population (Fleet and Heard, 1993). To a smallerextent, it is also possible to detect the species of Candida,Cryptococcus, Rhodotorula, Pichia, Metschnikowia and Kluy-veromyces (Barnett et al., 1972; Davenport, 1974; Rosini et al.,1982; Heard and Fleet, 1988, Longo et al., 1991; Yanagida etal., 1992; Martini et al., 1996; De La Torre et al., 1999; Guerraet al., 1999; Mills et al., 2002). Recent studies have revealedthat Saccharomyces cerevisiae can be isolated from thevineyard soil or the surface of ripe grapes with extremedifficulty (Yanagida et al., 1992; Martini et al., 1996); accordingto Mortimer and Polsinelli (1999) only about one in a thousandberries is S. cerevisiae positive.
Cviček PTP is a typical Slovene wine; its geographical originis under special protection of the Slovene Wine Law (1997). It isa blended red wine made from different grape varieties, mostlyfrom red grapes of Žametovka (Vitis vinifera L. cv. Žametnačrnina) or Kölner Blauer (40–50%), and Modra frankinja (V.vinifera L. cv. Modra frankinja) or Blaufränkisch (15–20%),and from white grapes of Kraljevina (V. vinifera L. cv.Kraljevina) or Königstraube (10–15%) (Slovene Wine Law,1997; Regulations of production…, 2000).
Various molecular techniques such as electrophoretic karyo-typing, random amplified polymorphic DNA using polymerasechain reaction (RAPD-PCR), PCR-ribotyping have been used bydifferent authors for identifying different wine yeast species. Thelatter method proved to be useful for the differentiation of wineyeasts at species level (Smole Mozina and Raspor, 1997; SmoleMozina et al., 1998;Guillamón et al., 1998; Esteve-Zarzoso et al.,1999; Granchi et al., 1999; Capece et al., 2003).
The aim of this study was to obtain information about theinfluence of grape varieties on the indigenous yeast communitypresent on grape berries, and to evaluate the impact of thegeographical locations in the Dolenjska vine-growing region.
2. Material and methods
2.1. Grape sampling
Grapes were collected from five vineyards: (A) Pleterje, (B)Zavode, (C) Nemška gora, (D) Trška gora and (E) Hom,members of the Dolenjska vine-growing district in the south-eastern part of Slovenia. Only healthy and undamaged grapeswere aseptically collected in plastic autoclavable bags imme-diately before harvest (September, 1999) from the selectedvineyards. Duplicate samples were collected from randomlyselected vine plants of three different vine varieties: Modrafrankinja, Žametovka and Kraljevina, typical constituents of theSlovene wine — Cviček. Immediately after harvesting thesamples of grapes were transported in cold boxes to theBiotechnical Faculty in Ljubljana, located an hour's drive fromthe vine region and analyzed directly.
2.2. Yeast isolation
The content of each bag (100 g) was aseptically homoge-nized on Seward stomacher 400 (BA 7021, England) for 30 s at
normal intensity of homogenization. The homogenized sampleswere plated at different serial decimal dilutions in duplicates onyeast–malt agar (YM), containing 3% yeast extract (Biolife,Milano, Italy), 3% malt extract (Biolife, Milano, Italy), 5%bacteriological peptone (Oxoid, Hampshire, England), 10%glucose (Kemika, Zagreb, Croatia), 20% agar (Biolife, Milano,Italy). After the incubation at 28 °C for 3–5 days the colonieswere counted out in duplicates and differentiated on the basis oftheir morphology. 752 isolates were preserved at −80 °C forfurther identification.
2.3. Identification
DNA isolation was performed according to the protocoldescribed elsewhere (Raspor et al., 2001). The DNA amplifica-tions were performed in Perkin Elmer PCR System 2400. Theywere carried out in a 20 μL reaction volume containing 1×PCRbuffer (Promega, Madison, USA), 0.2 mM of each dNTP(Promega, Madison, USA), 2 mM MgCl2 (Promega, Madison,USA), 0.5 pmol of each primer and 1 U of Taq polymerase(Promega, Madison, USA). The primer pair used for amplifi-cation of 18S-ITS rDNA region was NS1 (5′ GTAGTCATATGCTTGTCTC 3′) and ITS4 (5′ TCCTCCGCTTATTGATATGC 3′) (White et al., 1990). The PCR conditions were asfollows: the initial denaturation cycle of 5 min at 95 °C;followed by 35 cycles of 30 s at 95 °C, 30 s at 60 °C, 3 min at72 °C and the final extension step of 7 min at 72 °C. PCRproducts were digested without further purification with therestriction endonucleases CfoI, HaeIII, MspI and RsaI (Roche,Mannheim, Germany), following the manufacturer’s instruc-tions, and separated by electrophoresis in 1.5% agarose gel(Sigma, St. Louis, USA) and 1×TAE buffer. After electropho-resis, gels were stained with ethidium bromide (0.5 μg/mL), anddocumented by Gel Doc 2000.
2.4. Physiological testing
The physiological characteristics of representatives of strainssharing identical PCR-RFLP profile were studied to confirm theiridentity. Standard procedures for the identification of yeasts, aswell as instructions for the preparation of materials and testingconditions were followed according to Barnett et al. (2000).
3. Results
3.1. Total number of isolated yeasts
Fig. 1 shows the number of yeasts (CFU/mL) isolated fromgrape berry surfaces of three grape cultivars Žametovka, Modrafrankinja and Kraljevina (V. vinifera L.) from five vineyardssituated in the Dolenjska vine-growing region. Within particularsampling location differences in the number of yeasts have beenfound. The number of yeasts isolated from cv. Žametovka variedfrom8.9·103–1.35·106 CFU/mL.On the surface of grape berriescv. Modra frankinja 2.5 ·104–6.05·105 CFU/mL have beenfound. The surface of the white grape cultivar Kraljevina wasinhabited by 3.9 ·104–5.04·105 CFU/mL. For detail see Fig. 1.
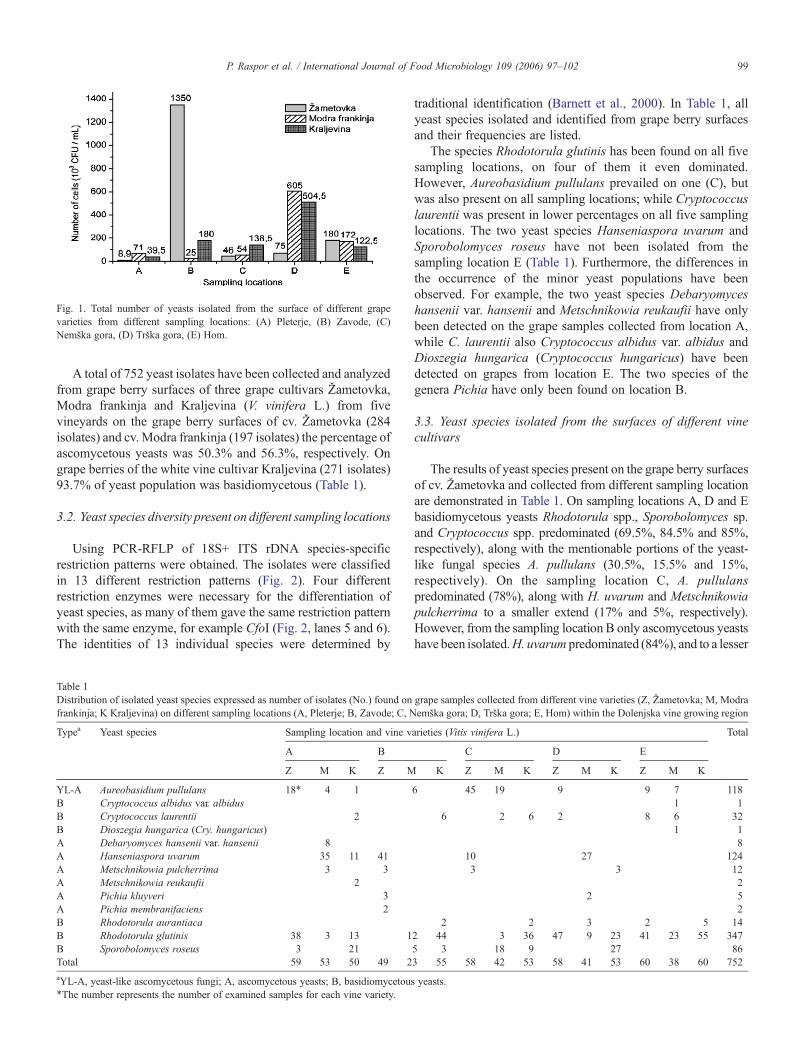
Fig. 1. Total number of yeasts isolated from the surface of different grapevarieties from different sampling locations: (A) Pleterje, (B) Zavode, (C)Nemška gora, (D) Trška gora, (E) Hom.
99P. Raspor et al. / International Journal of Food Microbiology 109 (2006) 97–102
A total of 752 yeast isolates have been collected and analyzedfrom grape berry surfaces of three grape cultivars Žametovka,Modra frankinja and Kraljevina (V. vinifera L.) from fivevineyards on the grape berry surfaces of cv. Žametovka (284isolates) and cv. Modra frankinja (197 isolates) the percentage ofascomycetous yeasts was 50.3% and 56.3%, respectively. Ongrape berries of the white vine cultivar Kraljevina (271 isolates)93.7% of yeast population was basidiomycetous (Table 1).
3.2. Yeast species diversity present on different sampling locations
Using PCR-RFLP of 18S+ ITS rDNA species-specificrestriction patterns were obtained. The isolates were classifiedin 13 different restriction patterns (Fig. 2). Four differentrestriction enzymes were necessary for the differentiation ofyeast species, as many of them gave the same restriction patternwith the same enzyme, for example CfoI (Fig. 2, lanes 5 and 6).The identities of 13 individual species were determined by
Table 1Distribution of isolated yeast species expressed as number of isolates (No.) found onfrankinja; K Kraljevina) on different sampling locations (A, Pleterje; B, Zavode; C, N
Typea Yeast species Sampling location and vine v
A B
Z M K Z M
YL-A Aureobasidium pullulans 18⁎ 4 1B Cryptococcus albidus var. albidusB Cryptococcus laurentii 2B Dioszegia hungarica (Cry. hungaricus)A Debaryomyces hansenii var. hansenii 8A Hanseniaspora uvarum 35 11 41A Metschnikowia pulcherrima 3 3A Metschnikowia reukaufii 2A Pichia kluyveri 3A Pichia membranifaciens 2B Rhodotorula aurantiacaB Rhodotorula glutinis 38 3 13 1B Sporobolomyces roseus 3 21Total 59 53 50 49 2
aYL-A, yeast-like ascomycetous fungi; A, ascomycetous yeasts; B, basidiomycetous⁎The number represents the number of examined samples for each vine variety.
traditional identification (Barnett et al., 2000). In Table 1, allyeast species isolated and identified from grape berry surfacesand their frequencies are listed.
The species Rhodotorula glutinis has been found on all fivesampling locations, on four of them it even dominated.However, Aureobasidium pullulans prevailed on one (C), butwas also present on all sampling locations; while Cryptococcuslaurentii was present in lower percentages on all five samplinglocations. The two yeast species Hanseniaspora uvarum andSporobolomyces roseus have not been isolated from thesampling location E (Table 1). Furthermore, the differences inthe occurrence of the minor yeast populations have beenobserved. For example, the two yeast species Debaryomyceshansenii var. hansenii and Metschnikowia reukaufii have onlybeen detected on the grape samples collected from location A,while C. laurentii also Cryptococcus albidus var. albidus andDioszegia hungarica (Cryptococcus hungaricus) have beendetected on grapes from location E. The two species of thegenera Pichia have only been found on location B.
3.3. Yeast species isolated from the surfaces of different vinecultivars
The results of yeast species present on the grape berry surfacesof cv. Žametovka and collected from different sampling locationare demonstrated in Table 1. On sampling locations A, D and Ebasidiomycetous yeasts Rhodotorula spp., Sporobolomyces sp.and Cryptococcus spp. predominated (69.5%, 84.5% and 85%,respectively), along with the mentionable portions of the yeast-like fungal species A. pullulans (30.5%, 15.5% and 15%,respectively). On the sampling location C, A. pullulanspredominated (78%), along with H. uvarum and Metschnikowiapulcherrima to a smaller extend (17% and 5%, respectively).However, from the sampling location B only ascomycetous yeastshave been isolated.H. uvarum predominated (84%), and to a lesser
grape samples collected from different vine varieties (Z, Žametovka; M, Modraemška gora; D, Trška gora; E, Hom) within the Dolenjska vine growing region
arieties (Vitis vinifera L.) Total
C D E
K Z M K Z M K Z M K
6 45 19 9 9 7 1181 1
6 2 6 2 8 6 321 1
810 27 1243 3 12
22 5
22 2 3 2 5 14
2 44 3 36 47 9 23 41 23 55 3475 3 18 9 27 863 55 58 42 53 58 41 53 60 38 60 752
yeasts.

Fig. 2. Restriction patterns of 13 different wine yeast species obtained with CfoIrestriction enzyme. Lanes: (1 and 15) 1-kb DNA ladder (Invitogen); (2)Hanseniaspora uvarum (ZIM 2022); (3) Debaryomyces hansenii var. hansenii(ZIM 2025); (4) Pichia kluyveri (ZIM 2053); (5) Dioszegia hungarica(Cryptococcus hungaricus) (ZIM 2060); (6) Cryptococcus albidus var. albidus(ZIM 2062); (7) Metschnikowia reukaufii (ZIM 2064); (8) Rhodotorula glutinis(ZIM 2072); (9) Rhodotorula aurantiaca (ZIM 2078); (10) Metschnikowiapulcherrima (ZIM 2082); (11) Cryptococcus laurentii (ZIM 2084); (12) Pichiamembranifaciens (ZIM 2091); (13) Sporobolomyces roseus (ZIM 2095); (14)Aureobasidium pullulans (ZIM 2098), ZIM: Collection of Industrial Micro-organisms, Slovenia.
100 P. Raspor et al. / International Journal of Food Microbiology 109 (2006) 97–102
extend the following yeast species were present: M. pulcherrima(6%), Pichia kluyveri (6%) and P. membranifaciens (4%).
From vine cultivar Modra frankinja 11 different yeast specieshave been isolated. R. glutinis was present on all samplinglocations, but its percentage varied from 6% on sampling locationA to 60% on location E. H. uvarum prevailed on locations A andD (66% on both) but has not been isolated from Modra frankinjagrape surfaces from the other three sampling locations. Also blackyeast, as some authors refers to A. pullulans, was present on foursampling locations. Yeast species such as: M. pulcherrima,P. kluyveri, Cryptococcus spp., S. roseus, D. hansenii var.hansenii and Rhodotorula aurantiaca were present to a lesserextend (Table 1).
From the surface of grapes of the third white vine cultivarKraljevina 8 different yeast species have been isolated (Table 1).The population of yeasts present on sampling locations B, C andE was 100% basidiomycetous yeasts consisting of Rhodothor-ula spp., S. roseus and C. laurentii, while on locations A and Dtheir percentage was 72% and 94%, respectively. However,beside R. glutinis the ascomycetous yeasts H. uvarum, M.reukaufii and A. pullulans have been isolated from grapes fromsampling location A.
4. Discussion
One of the major challenges of wine microbiology is to gainmore information about yeast communities present on grapeberries and their influence on the winemaking process. Thisstudy presents considerable information about yeast communitystructure influenced by geographic location and vine varieties,typical constituents of Cviček.
The large majority of studies on yeast flora of vine grapesreviewed by Fleet and Heard (1993) and Fleet et al. (2002)
pointed out that mature, sound grapes, aseptically crushed willgive must with a total yeast population between 103–105 CFUper mL of grape juice. Comparable yeast population countswere reported by Rosini et al. (1982) and Davenport (1974) forItalian and English vineyards, respectively. The numbers ofyeasts isolated from grape berry surfaces of three different vinevarieties and on five different sampling locations in theDolenjska vine-growing region is generally in agreement withthese numbers (Fig. 1). An exception was the second samplinglocation Zavode where 1.35 ·106 CFU/mL have been isolatedfrom the surface of cv. Žametovka grapes. The higher number ofyeasts could be a result of the rapidly growing ascomycetousyeasts of H. uvarum (84%), and M. pulcherrima (6%). As theyeast species of the genera Hanseniaspora/Kloeckera andMetschnikowia mostly predominate in the early stages of winefermentations, until the rise of ethanol levels up to 3–4%(Rosini et al., 1982; Heard and Fleet, 1988; Schütz and Gafner,1993; Vaughan-Martini and Martini, 1995; Martini et al., 1996;Ramón, 1997; Pretorius, 2000; Ribéreau-Gayon et al., 2000),their presence on grape surfaces is expected.
Several studies have tried to show that the geographiclocation and microclimatological conditions have an impact onthe population density and diversity of yeasts isolated fromgrape surfaces (Barnett et al., 1972; Davenport, 1974; Longo etal., 1991; Yanagida et al., 1992; De La Torre et al., 1999) andconsequently, these factors were also correlated with thecomposition of yeast flora isolated from the early stages ofwine fermentations (Ramón, 1997; Pretorius, 2000; Ribéreau-Gayon et al., 2000). In our case the impact of geographicallocations and microclimatological changes influenced thespecies diversity. For example, on the sampling location Aseven different yeast species have been found but only threedifferent yeast species were present on the surface of Žametovkagrapes, 5 different species on the surface of Modra frankinjagrapes, and 6 different wine yeast species on the surface ofKraljevina grapes have been isolated (Table 1). Similar is truealso for the other four sampling locations. However, for drawingany definite conclusions, further experiments containingecological data reports, identification of yeast species and theirpopulation counts from the same vine variety, vine plant,vineyard during several years should be performed.
The frequency of occurrence of particular yeast speciesshowed their preferences for certain grape varieties. The whitegrape variety Kraljevina mostly attracted red pigmentedbasidiomycetous yeasts belonging to the genera Rhodotorula,Sporobolomyces and Cryptococcus (Table 1) which altogetherdominated on all five sampling locations. On the other hand,yeast populations isolated from the red grape surfaces (cv.Žametovka and cv. Modra frankinja) belonged both toascomycetous and basidiomycetous yeasts in the ratio of 1 :1.From these observations it can be concluded that yeasts havesome preferences for the vine variety but cannot confirm theobservations of Deak and Beuchat (1993) that red vine varietiesoffer an advantage for yeast growth due to higher grape juice pH.
The black yeast-like fungus A. pullulans the taxonomicposition of which is within the ascomycetous order Dothideales(Kurtzman and Fell, 1998) is the most widespread saprophyte in

101P. Raspor et al. / International Journal of Food Microbiology 109 (2006) 97–102
phylosphere and is considered to be a potential biocontrol agentagainst grey mould (Schena et al., 2003). In all considered vine-growing locations A. pullulans was almost non-present onKraljevina grape berries, while on the red vine varietyŽametovka and Modra frankinja, its presence was relativelyhigh. Probably this yeast species has some preferences for thered grape variety but further study should be performed.
Different researchers have published quite contradictoryopinions about the presence of the fermentative yeast species S.cerevisiae in vineyards. Some claimed that the grape berry is thenatural origin of this wine yeast (Török et al., 1996; Pretorius etal., 1999), while others had difficulties isolating S. cerevisiaefrom nature and believe that man-made environment and wineryequipment is the main source of this commercially importantyeast species (Davenport, 1974; Rosini et al., 1982; Yanagida etal., 1992; Vaughan-Martini and Martini, 1995; Martini et al.,1996; Mortimer and Polsinelli, 1999). However, the majority ofstudies suggest that S. cerevisiae does occur on grapes invineyards, but its populations are too low (b10–100 CFU/cm2)to be detected by direct agar plating procedures (reviewed byFleet et al., 2002), therefore enrichment techniques areproposed for isolating this yeast species from nature (Töröket al., 1996). Having used the direct agar plating method mightprovide a possible explanation why S. cerevisiae was foundneither on grape samples of the three different grape cultivarsnor on the five different sampling locations.
The main conclusions of this study can be summarized asfollows: (i) autochthonous yeast flora present on grape surfacesdiffers among vine varieties as well as among differentsampling locations; (ii) thirteen different yeast species belong-ing to 9 different genera have been found on the surface of grapeberries of 3 vine varieties from 5 sampling locations in theSlovene vine-growing region Dolenjska (iii) red or reddishbasidiomycetous yeasts predominated on grapes of the whitevine variety, Kraljevina, while for red vine varieties, Žametovkaand Modra frankinja, equal relation between ascomycetous andbasidiomycetous yeasts has been observed; (iv) A. pullulans, aspotential biocontrol agent probably has some preferences for thered grape variety but further study should be carried out; (v)some yeast species were characteristic for a particularsampling location (e.g., yeast species D. hansenii var. hanseniiand M. reukaufii have been isolated from sampling locationA only; on sampling location B yeast species Pichia membra-nifaciens; and on sampling location E yeast species C. albidusand D. hungarica (C. hungaricus) have been found, exclusive-ly); (vi) S. cerevisiae as a leading yeast of alcoholic fermentationhas not been found.
Acknowledgements
Ministry of Education, Science and Sport of the Republicof Slovenia (Project No L4-8849-98) supported this research.We wish to thank Ursa Debelak, Jana Avbelj and dr. KatjaPovhe Jemec for their kind assistance. We also thank SamoHudoklin for support with grape sample collection. M.M.D.and P.J. express their gratitude to Biotechnical Faculty for theopportunity to work on this project.
References
Barnett, J.A., Delaney, M.A., Jones, E., Magson, A.B., Winch, B., 1972. Thenumbers of yeasts associated with wine grapes of Bordeaux. Archives ofMicrobiology 83, 52–55.
Barnett, J.A., Payne, R.W., Yarrow, D., 2000. Yeasts: Characteristics andIdentification. Cambridge University Press, Cambridge, p. 1139.
Capece, A., Salzano, G., Romano, P., 2003. Molecular typing techniques as atool to differentiate non-Saccharomyces wine species. International Journalof Food Microbiology 84, 33–39.
Davenport, R.R., 1974. Microecology of yeasts and yeast-like organismsassociated with an English vineyard. Vitis 13, 123–130.
De La Torre, M.J., Millan, M.C., Perez-Juan, P., Morales, J., Ortega, J.M., 1999.Indigenous yeasts associated with two Vitis vinifera grape varieties culturedin southern Spain. Microbios 100, 27–40.
Deak, T., Beuchat, L.R., 1993. Yeasts associated with fruit juice concentrates.Journal of Food Protection 56 (9), 777–782.
Esteve-Zarzoso, B., Belloch, C., Uruburu, F., Querol, A., 1999. Identification ofyeasts by RFLP analysis of the 5.8S rRNA gene and two ribosomal internaltranscribed spacers. International Journal of Systematic Bacteriology 49,329–337.
Fleet, G.H., 1993. The microorganisms of winemaking — isolation,enumeration and identification. In: Fleet, G.H. (Ed.), Wine Microbiologyand Biotechnology. Harwood AP, Australia, pp. 1–26.
Fleet, G.H., Heard, G.M., 1993. Yeasts-growth during fermentation. In: Fleet, G.H. (Ed.), Wine Microbiology and Biotechnology. Harwood AP, Australia,pp. 27–54.
Fleet, G.H., Prakitchaiwattana, C., Beh, A.L., Heard, G.M., 2002. The yeastecology of wine grapes. In: Ciani, M. (Ed.), Biodiversity and Biotechnologyof Wine Yeasts. Research Singpost, Kerala, India, pp. 1–17.
Fugelsang, K.C., 1997. Yeasts and molds. In: Fugelsang, K.C. (Ed.), WineMicrobiology. Chapman and Hall, New York, pp. 68–116.
Granchi, L., Bosco, M., Messini, A., Vincenzini, M., 1999. Rapid detection andquantification of yeast species during spontaneous wine fermentation byPCR-RFLP analysis of the rDNA ITS region. Journal of AppliedMicrobiology 87, 949–956.
Guerra, E., Sordi, G., Mannazzu, I., Clementi, F., Fatichenti, F., 1999.Occurrence of wine yeasts on grapes subjected to different pesticidetreatments. Italian Journal of Food Science 3 (11), 221–230.
Guillamón, J.M., Sabaté, J., Barrio, E., Cano, J., Querol, A., 1998. Rapididentification of wine yeast species based on RFLP analysis of the ribosomalinternal transcribed spacer (ITS) region. Archives of Microbiology 169,387–392.
Heard, G.M., Fleet, G.H., 1988. The effects of temperature and pH on thegrowth of yeast species during the fermentation of grape juice. Journal ofApplied Bacteriology 65, 23–28.
Kurtzman, C.P., Fell, J.W., 1998. The Yeasts, a Taxonomic Study, 4th ed.Elsevier, Amsterdam. 1055 pp.
Longo, E., Cansado, J., Agrelo, D., Villa, T.G., 1991. Effect of climaticconditions on yeast diversity in grape musts from northwest Spain.American Journal of Enology and Viticulture 42 (2), 141–144.
Martini, A., Ciani, M., Scorzetti, G., 1996. Direct enumeration and isolation ofwine yeasts from grape surfaces. American Journal of Enology andViticulture 47 (4), 435–440.
Mills, D.A., Johannsen, E.A., Cocolin, L., 2002. Yeast diversity and persistencein Botrytis-affected wine fermentations. Applied and EnvironmentalMicrobiology 68 (10), 4884–4893.
Mortimer, R., Polsinelli, M., 1999. On the origins of wine yeast. Research inMicrobiology 150, 199–204.
Pretorius, I.S., 2000. Tailoring wine yeast for the new millennium: novelapproaches to the ancient art of winemaking. Yeast 16, 675–729.
Pretorius, I.S., Van der Westhuizen, T.J., Augustyn, O.P.H., 1999. Yeastbiodiversity in vineyards and wineries and its importance to the South Africanwine industry. South African Journal of Enology and Viticulture 20, 61–74.
Ramón, D., 1997. Trends in wine microbiology. Microbiología SEM 13,405–412.
Raspor, P., Smole Mozina, S., Čadez, N., 2001. Identification of yeasts fromgrape/must/wine system. In: Spencer, J.F.T., Ragout de Spencer, A.L. (Eds.),

102 P. Raspor et al. / International Journal of Food Microbiology 109 (2006) 97–102
Methods in Biotechnology, vol. 14. Humana Press Inc, Totowa, pp.243–251.
Regulations of production of recognized and traditionally named wine —Cviček, vol. Ur.l. RS 10, 3, pp. 286–287.
Ribéreau-Gayon, P., Dubourdieu, D., Dončche, B., Lonvaud, A., 2000.Cytology, taxonomy and ecology of grape and wine yeasts. Handbook ofEnology. The Microbiology of Wine and Vinifications, vol. 1. John Wileyand Sons, Ltd., pp. 1–49.
Rosini, G., Federici, F., Martini, A., 1982. Yeast flora of grape berries duringripening. Microbial Ecology 8, 83–89.
Schena, L., Nigro, F., Pentimone, I., Ligorio, A., Ippolito, A., 2003. Control ofpostharvest rots of sweet cherries and table grapes with endophytic isolatesof Aureobasidium pullulans. Postharvest Biology and Technology 30,209–220.
Schütz, M., Gafner, J., 1993. Analysis of yeast diversity during spontaneous andinduced alcoholic fermentations. Journal of Applied Bacteriology 75,551–558.
Schütz, M., Gafner, J., 1994. Dynamics of the yeast strain population duringspontaneous alcoholic fermentation determined by CHEF gel electropho-resis. Letters in Applied Microbiology 19, 253–257.
Slovene wine law, 1997. Ur. l. RS, vol. 7, 70, pp. 5483–5492.Smole Mozina, S., Raspor, P., 1997. Molecular techniques for yeast
identification in food processing. Food Technology and Biotechnology 35(1), 55–61.
Smole Mozina, S., Čadez, N., Raspor, P., 1998. rDNA RFLPs and AP-PCRfingerprinting of type strains and grape-must isolates of Hanseniaspora(Kloeckera) yeasts. Food Technology and Biotechnology 36 (1), 37–43.
Török, T., Mortimer, R.K., Romano, P., Suzzi, G., Polsinelli, M., 1996. Quest forwine yeast — an old story revisited. Journal of Industrial Microbiology 17,303–313.
Vaughan-Martini, A., Martini, A., 1995. Facts, myths and legends on the primeindustrial microorganism. Journal of Industrial Microbiology 14, 514–522.
White, T.J., Bruns, T., Lee, S., Taylor, J., 1990. Amplification and directsequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A., Gelfand, D.H., Sinisky, J.J., White, T.J. (Eds.), PCR Protocols. A guideto Methods and Applications. Academic Press Inc., London, pp. 315–322.
Yanagida, F., Ichinose, F., Shinohara, T., Goto, S., 1992. Distribution of wildyeasts in the white grape varieties at Central Japan. Journal of General andApplied Microbiology 38, 501–504.
Related Documents











