Developmental Cell Article Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells Rui Zhao, 1,2,3,4 Timothy R. Fallon, 1 Srinivas Vinod Saladi, 5 Ana Pardo-Saganta, 1,2,3,4 Jorge Villoria, 1 Hongmei Mou, 1,2,3,4 Vladimir Vinarsky, 1,2,3,4 Meryem Gonzalez-Celeiro, 1 Naveen Nunna, 1 Lida P. Hariri, 3,6 Fernando Camargo, 4,7 Leif W. Ellisen, 5 and Jayaraj Rajagopal 1,2,3,4, * 1 Center for Regenerative Medicine, Massachusetts General Hospital, 185 Cambridge Street, Boston, MA 02114, USA 2 Department of Pediatrics, Massachusetts General Hospital, Boston, MA 02114, USA 3 Department of Internal Medicine, Pulmonary and Critical Care Unit, Massachusetts General Hospital, Boston, MA 02114, USA 4 Harvard Stem Cell Institute, Cambridge, MA 02138, USA 5 Massachusetts General Hospital Cancer Center, Boston, MA 02114, USA 6 Department of Pathology, Massachusetts General Hospital, Boston, MA 02114, USA 7 Stem Cell Program, Boston Children’s Hospital, Boston, MA 02115, USA *Correspondence: [email protected] http://dx.doi.org/10.1016/j.devcel.2014.06.004 SUMMARY Our understanding of how stem cells are regulated to maintain appropriate tissue size and architecture is incomplete. We show that Yap (Yes-associated protein 1) is required for the actual maintenance of an adult mammalian stem cell. Without Yap, adult airway basal stem cells are lost through their unre- strained differentiation, resulting in the simplification of a pseudostratified epithelium into a columnar one. Conversely, Yap overexpression increases stem cell self-renewal and blocks terminal differentiation, re- sulting in epithelial hyperplasia and stratification. Yap overexpression in differentiated secretory cells causes them to partially reprogram and adopt a stem cell-like identity. In contrast, Yap knockdown prevents the dedifferentiation of secretory cells into stem cells. We then show that Yap functionally interacts with p63, the cardinal transcription factor associated with myriad epithelial basal stem cells. In aggregate, we show that Yap regulates all of the cardinal behaviors of airway epithelial stem cells and determines epithelial architecture. INTRODUCTION How adult tissues maintain their proper size and architecture is poorly understood. Here we explore how the regulation of adult stem cells is linked to epithelial architecture using the airway epithelium as a model system. Epithelial tissues are generally classified as simple, pseudostratified, or stratified. The murine tracheobronchial airway epithelium represents a model of pseu- dostratified epithelium intermediate between a simple single- layered epithelium and a multilayered stratified epithelium. Airway basal stem cells directly and broadly abut the basement membrane. In contrast, differentiated suprabasal secretory and ciliated cells have smaller zones of contact with the basement membrane and possess extensive luminal surfaces with their nuclei displaced toward the lumen. This arrangement of cells essentially creates a two-layered epithelium (Morrisey and Hogan, 2010; Rock et al., 2009). Theoretically, disturbances in the regulation of basal stem cells could, on the one hand, lead to a hypertrophic epithelium characterized by basal stem cell excess and stratified squamous metaplasia, as is frequently observed in conditions such as chronic obstructive pulmonary disease. Conversely, decreased stem cell numbers would be predicted to result in epithelial hypoplasia, which is thought to play a role in conditions such as bronchiolitis obliterans and airway fibrosis (O’Koren et al., 2013; Rock et al., 2010). Thus, tightly controlled mechanisms to regulate basal stem cell main- tenance, proliferation, and differentiation must exist to properly police epithelial size and architecture. Yap (Yes-associated protein 1) is a transcriptional coactivator in the conserved Hippo kinase cascade that has been shown to be involved in growth control as well as the regulation of stem and progenitors cells (Barry and Camargo, 2013; Halder and Johnson, 2011; Pan, 2007, 2010; Ramos and Camargo, 2012; Zhao et al., 2011). In epithelia, Yap modulation has diverse con- sequences on stem and progenitor cell behaviors (Ramos and Camargo, 2012; Zhao et al., 2011). In the embryonic neuroepi- thelium, Yap loss leads to decreased progenitor cell survival (Cao et al., 2008), whereas in the embryonic epidermis, Yap loss leads to decreased progenitor cell proliferation (Schlegel- milch et al., 2011). In contrast, Yap activation leads to the same phenotype in both of these tissues, namely increased progenitor and stem cell replication (Cao et al., 2008; Schlegel- milch et al., 2011; Zhang et al., 2011). Unexpectedly, Yap loss throughout the intestinal epithelium results in no obvious pheno- type but causes hyperplasia and increased stem cell replication after injury (Barry et al., 2013). Surprisingly, Yap overexpression leads to a loss rather than a gain of intestinal stem cells (Barry et al., 2013). Thus, Yap acts in a tissue-, cell-, and context- dependent manner, even within epithelia. Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 1 Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self- Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
Developmental Cell
Article
Yap Tunes Airway Epithelial Size andArchitecture by Regulating the Identity,Maintenance, and Self-Renewal of Stem CellsRui Zhao,1,2,3,4 Timothy R. Fallon,1 Srinivas Vinod Saladi,5 Ana Pardo-Saganta,1,2,3,4 Jorge Villoria,1 Hongmei Mou,1,2,3,4
Vladimir Vinarsky,1,2,3,4 Meryem Gonzalez-Celeiro,1 Naveen Nunna,1 Lida P. Hariri,3,6 Fernando Camargo,4,7
Leif W. Ellisen,5 and Jayaraj Rajagopal1,2,3,4,*1Center for Regenerative Medicine, Massachusetts General Hospital, 185 Cambridge Street, Boston, MA 02114, USA2Department of Pediatrics, Massachusetts General Hospital, Boston, MA 02114, USA3Department of Internal Medicine, Pulmonary and Critical Care Unit, Massachusetts General Hospital, Boston, MA 02114, USA4Harvard Stem Cell Institute, Cambridge, MA 02138, USA5Massachusetts General Hospital Cancer Center, Boston, MA 02114, USA6Department of Pathology, Massachusetts General Hospital, Boston, MA 02114, USA7Stem Cell Program, Boston Children’s Hospital, Boston, MA 02115, USA
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.devcel.2014.06.004
SUMMARY
Our understanding of how stem cells are regulatedto maintain appropriate tissue size and architectureis incomplete. We show that Yap (Yes-associatedprotein 1) is required for the actual maintenanceof an adult mammalian stem cell. Without Yap, adultairway basal stem cells are lost through their unre-strained differentiation, resulting in the simplificationof a pseudostratified epithelium into a columnar one.Conversely, Yap overexpression increases stem cellself-renewal and blocks terminal differentiation, re-sulting in epithelial hyperplasia and stratification.Yap overexpression in differentiated secretory cellscauses them to partially reprogram and adopt astem cell-like identity. In contrast, Yap knockdownprevents the dedifferentiation of secretory cellsinto stem cells. We then show that Yap functionallyinteracts with p63, the cardinal transcription factorassociated with myriad epithelial basal stem cells.In aggregate, we show that Yap regulates all of thecardinal behaviors of airway epithelial stem cellsand determines epithelial architecture.
INTRODUCTION
How adult tissues maintain their proper size and architecture is
poorly understood. Here we explore how the regulation of adult
stem cells is linked to epithelial architecture using the airway
epithelium as a model system. Epithelial tissues are generally
classified as simple, pseudostratified, or stratified. The murine
tracheobronchial airway epithelium represents a model of pseu-
dostratified epithelium intermediate between a simple single-
layered epithelium and a multilayered stratified epithelium.
Airway basal stem cells directly and broadly abut the basement
membrane. In contrast, differentiated suprabasal secretory and
ciliated cells have smaller zones of contact with the basement
membrane and possess extensive luminal surfaces with their
nuclei displaced toward the lumen. This arrangement of cells
essentially creates a two-layered epithelium (Morrisey and
Hogan, 2010; Rock et al., 2009). Theoretically, disturbances in
the regulation of basal stem cells could, on the one hand, lead
to a hypertrophic epithelium characterized by basal stem cell
excess and stratified squamous metaplasia, as is frequently
observed in conditions such as chronic obstructive pulmonary
disease. Conversely, decreased stem cell numbers would be
predicted to result in epithelial hypoplasia, which is thought to
play a role in conditions such as bronchiolitis obliterans and
airway fibrosis (O’Koren et al., 2013; Rock et al., 2010). Thus,
tightly controlled mechanisms to regulate basal stem cell main-
tenance, proliferation, and differentiation must exist to properly
police epithelial size and architecture.
Yap (Yes-associated protein 1) is a transcriptional coactivator
in the conserved Hippo kinase cascade that has been shown to
be involved in growth control as well as the regulation of stem
and progenitors cells (Barry and Camargo, 2013; Halder and
Johnson, 2011; Pan, 2007, 2010; Ramos and Camargo, 2012;
Zhao et al., 2011). In epithelia, Yap modulation has diverse con-
sequences on stem and progenitor cell behaviors (Ramos and
Camargo, 2012; Zhao et al., 2011). In the embryonic neuroepi-
thelium, Yap loss leads to decreased progenitor cell survival
(Cao et al., 2008), whereas in the embryonic epidermis, Yap
loss leads to decreased progenitor cell proliferation (Schlegel-
milch et al., 2011). In contrast, Yap activation leads to the
same phenotype in both of these tissues, namely increased
progenitor and stem cell replication (Cao et al., 2008; Schlegel-
milch et al., 2011; Zhang et al., 2011). Unexpectedly, Yap loss
throughout the intestinal epithelium results in no obvious pheno-
type but causes hyperplasia and increased stem cell replication
after injury (Barry et al., 2013). Surprisingly, Yap overexpression
leads to a loss rather than a gain of intestinal stem cells (Barry
et al., 2013). Thus, Yap acts in a tissue-, cell-, and context-
dependent manner, even within epithelia.
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 1

Figure 1. Yap Is Required for the Maintenance of Adult Airway Basal Stem Cells and Yap Loss Results in the Simplification of a Pseudostra-
tified Epithelium into a Columnar Epithelium
(A) Expression of Yap mRNA in basal and secretory cells relative to that in ciliated cells.
(B) Immunostaining for Yap (red) and the basal stem cell marker cytokeratin 5 (CK5, green). Yap protein is highly enriched in the nuclei of basal stem cells (white
arrows) as compared to differentiated cells (yellow arrowheads).
(C) A schematic of the strategy and phenotypic outcome of stem cell-specific Yap deletion.
(legend continued on next page)
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
2 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
Here we use the airway epithelia to reveal that Yap, in concert
with the cardinal basal stem cell transcription factor p63, partic-
ipates in themaintenance of an adult stem cell and the regulation
of stem cell identity itself. Furthermore, we demonstrate that
stem cell behaviors including self-renewal and differentiation
can be modulated by Yap, resulting in predictable alterations
in epithelial architecture and size. These findings suggest that al-
terations in Yap activity may be involved in those diseases of the
airways associated with alterations in epithelial architecture,
such as premalignant squamous metaplasia.
RESULTS
Yap Is Required for the Maintenance of Adult AirwayBasal Stem Cells and Yap Loss Results in theSimplification of a Pseudostratified Epithelium into aColumnar EpitheliumWe defined the expression pattern of Yap in the three different
cell types of the adult airway epithelium. Basal, secretory, and
ciliated cells were sorted based upon GSIb4, SSEA1, and
CD24 surface expression, respectively (Figure S1A available on-
line). We verified the cell type-specific exclusive expression of
mRNAs in each sorted cell population (Figure S1B). Yap mRNA
was expressed at higher levels in basal stem cells than in secre-
tory and ciliated cells (Figure 1A). We used three different Yap
antibodies to establish cell type-specific Yap protein expression
patterns using tyramide signal amplification (TSA). Staining
demonstrated that Yap protein is expressed most highly in basal
stem cells (Figure 1B). The nuclear localization of Yap in basal
stem cells (n = 2,893) suggests that Yap is actively performing
its function as a transcriptional co-activator in these stem cells
(Halder and Johnson, 2011; Pan, 2010; Zhao et al., 2011).
To determine the effect of Yap removal specifically from CK5
positive (+) airway basal stem cells, we generated triple trans-
genic mice carrying the Cytokeratin5 (CK5) rtTA (Diamond
et al., 2000), tetO Cre, and floxed Yap (Schlegelmilch et al.,
2011) alleles, referred to as CK5-YapKO throughout the text (Fig-
ure 1C). We first verified that Cre recombinase was expressed
exclusively in basal stem cells using a YFP reporter (Fig-
ure S1C). Then, adult CK5-YapKO animals (age 8–12 weeks)
were treated with both inhaled doxycycline (Tata et al., 2013b)
and doxycycline in their drinking water to induce efficient Yap
deletion. Yap deletion was verified by quantitative RT-PCR anal-
ysis performed on sorted basal stem cells 1 month after doxycy-
cline administration (Figure S1D). Histology of tracheal sections
revealed that the normally pseudostratified airway epithelium
was simplified into a columnar epithelium 3months after doxycy-
cline treatment (Figure 1D). Following Yap loss, we found a sig-
nificant decrease in the total number of airway basal stem cells
as demonstrated by a loss of cells that express CK5 and p63
(Figure 1E). In control animals, 1,174 ± 16 basal stem cells
(D) Hematoxylin and eosin (H&E) staining of tracheal sections from control (left) an
Cre alleles.
(E) Immunofluorescence analysis of basal stem cell markers p63 (red) and CK
doxycycline treatment.
(F) Quantification of basal stem cells marked by p63 in control, CK5-YapKO, and
Data are presented as mean ± SEM. n = 3 for each genotype per each time point.
also Figure S1.
marked by p63 were counted in standardized tracheal sections
covering 12 cartilaginous rings. Following doxycycline treatment
in the CK5-YapKO animals, the number of p63+ basal stem cells
per section dropped to 666 ± 65 in 1 month (p = 0.0017), to 96 ±
14 (p < 0.0001) after 2 months, and to 25 ± 8 (p < 0.0001)
after 3 months (Figures 1E and 1F). These results were further
confirmed using the basal stem cell-specific markers NGFR
and T1a (Figure S1E). Additionally, there was a significant
decrease in the total number of basal stem cells in transgenic an-
imals in which only a single copy of Yapwas removed (referred to
as CK5-YapHet; Figure 1E). As compared to the controls above,
we found only 535 ± 15 basal stem cells marked by p63 per sec-
tion (p < 0.0001) in the trachea of CK5-YapHet animals 3 months
after doxycycline treatment (Figure 1F). In summary, we found
that basal stem cell-specific deletion of Yap led to a dose-depen-
dent loss of stem cells and resulted in a corresponding simplifi-
cation of airway epithelial architecture.
Basal Stem Cells Undergo Unrestrained Differentiationafter Yap LossWe hypothesized that the loss of basal stem cells following
Yap inactivation could have occurred because of their death,
decreased replication, or differentiation. We found no detectable
changes in apoptosis as assessed by activated caspase-3 and
TUNEL staining at 1, 2, and 4 weeks after doxycycline adminis-
tration (Figure S2A and data not shown). With regard to replica-
tion, at homeostasis, cycling basal cells (i.e., positive for both
Ki-67 and p63) accounts for 2.52% ± 0.97% of total basal
stem cells. The percentage of p63+Ki-67+ cells in CK5-YapKO
trachea was unchanged after Yap deletion (2.10% ± 0.81%,
2.94% ± 0.60%, 2.88% ± 1.05% at 1, 2, and 3 months
after Yap deletion, respectively). However, when we assessed
markers of differentiation, we noted the appearance of presump-
tively differentiating stem cells positive for both p63 (basal) and
Scgb3A2 (secretory; Figures 2A and S2B) and cells positive for
both CK5 (basal) and FoxJ1 (ciliated; Figures 2B and S2B) in
CK5-YapKO animals. In controls, these markers were mutually
exclusive (Figures 2A and 2B). We further demonstrated that
such transitional double-positive cells are also present during
the course of normal airway epithelial regeneration following sul-
fur dioxide-induced injury, when stem cells begin to differentiate
into suprabasal cells (Figure S2C).
To definitively confirm that stem cells were being lost through
differentiation following Yap loss, we specifically lineage-traced
the basal stem cells that had lost Yap. Triple transgenic mice
bearing CK5-CreER (Van Keymeulen et al., 2011), floxed Yap,
and Rosa26-LSL-YFP alleles were generated so that a YFP re-
porter could be used to label and trace basal stem cells in which
Yap had been deleted. Two weeks after tamoxifen induction, the
majority of YFP-labeled basal stem cells remained CK5+ basal
stem cells in control mice (98.75%, n = 240 YFP cells counted).
d CK5-YapKO (right) animals. Control animals bear only the CK5 rtTA and tetO
5 (green) in control, CK5-YapKO, and CK5-YapHet tracheal sections after
CK5-YapHet tracheal sections.
DAPI is in blue. Scale bar represents 10 mm in (B) and (E), and 5 mm in (D). See
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 3
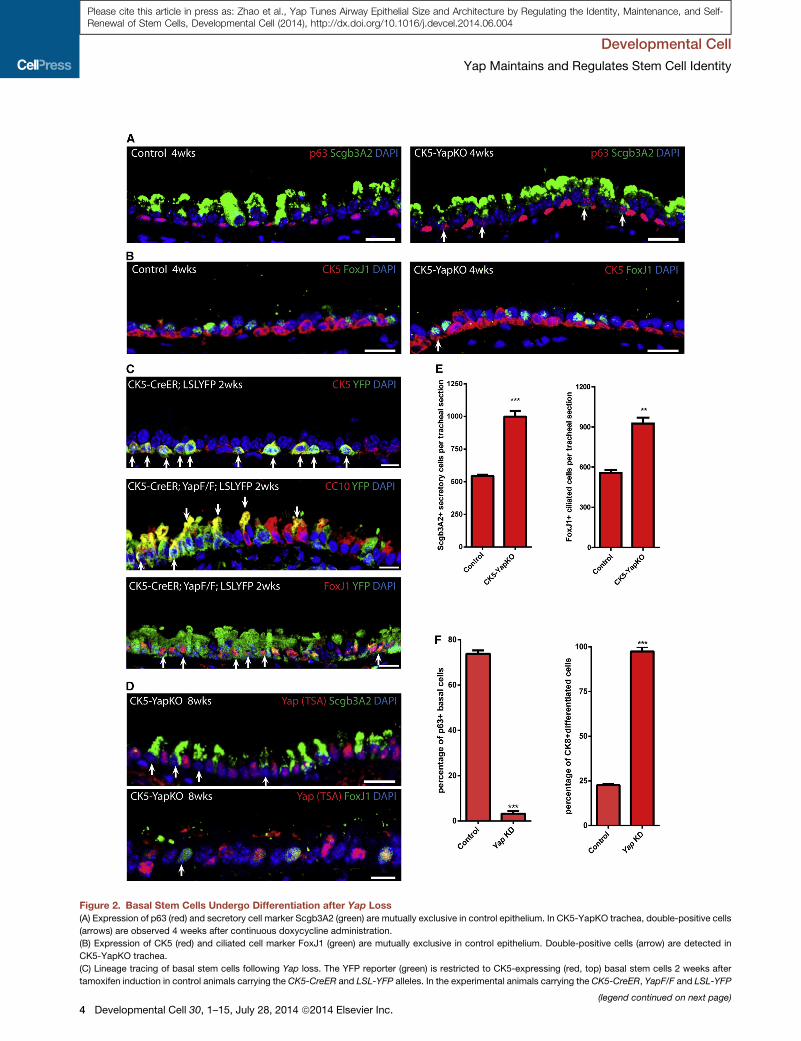
Figure 2. Basal Stem Cells Undergo Differentiation after Yap Loss
(A) Expression of p63 (red) and secretory cell marker Scgb3A2 (green) are mutually exclusive in control epithelium. In CK5-YapKO trachea, double-positive cells
(arrows) are observed 4 weeks after continuous doxycycline administration.
(B) Expression of CK5 (red) and ciliated cell marker FoxJ1 (green) are mutually exclusive in control epithelium. Double-positive cells (arrow) are detected in
CK5-YapKO trachea.
(C) Lineage tracing of basal stem cells following Yap loss. The YFP reporter (green) is restricted to CK5-expressing (red, top) basal stem cells 2 weeks after
tamoxifen induction in control animals carrying the CK5-CreER and LSL-YFP alleles. In the experimental animals carrying the CK5-CreER, YapF/F and LSL-YFP
(legend continued on next page)
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
4 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
However, YFP+ cells that expressed the secretory cell marker
CC10 and the ciliated cell marker FoxJ1 were frequently de-
tected in the experimental animals following Yap deletion (Fig-
ure 2C). Thus, following Yap loss, basal stem cells underwent
unrestrained differentiation. As a further confirmation of these
results, we also traced the lineage of basal stem cells in CK5-
YapKO mice by identifying cells that completely lacked Yap
stainingafter TSAamplification.Whenweusedan increasedanti-
body concentration for Yap staining, every cell normally shows a
strong staining for Yap in control trachea (Figure S2D). Using this
highly sensitive immunohistochemical assay for Yap protein
expression,we foundYap-deleted cells that expressed secretory
(Scgb3A2) and ciliated cell (FoxJ1) markers in CK5-YapKO tra-
chea (Figure 2D). Such cells were never found in control airways.
The complete lack of Yap staining in these differentiated cells
indicated that they originated from basal stem cells in which
Yap had been deleted. These results further confirm that basal
stem cells that lose Yap subsequently differentiate. Consistent
with this notion, we found that there was a significant increase
in secretory and ciliated cells in CK5-YapKO animals (Figure 2E).
In controls, we counted 544 ± 10 Scgb3A2-expressing secretory
cells per standard tracheal section. This number increased signif-
icantly to 998 ± 44 (p < 0.001) in CK5-YapKO tracheal sections
2 months after doxycycline treatment (Figure 2E). Similarly, con-
trol tracheal sections contained 557 ± 22 FoxJ1+ ciliated cells per
tracheal section, whereas there was a significant increase of this
number to 925 ± 44 (p = 0.0017) in CK5-YapKOanimals 2months
after doxycycline treatment (Figure 2E). Of note, at 3 months
following doxycycline treatment, there was a significant increase
in the proportion of ciliated to secretory cells (data not shown). In
sum, because we found no significant changes in basal stem cell
proliferation or cell death after Yap deletion, our results demon-
strate that basal stem cell loss occurs predominantly through
their differentiation.
Finally, we performed ex vivo Yap knockdown experiments us-
ing a culture system that supports basal stem cell expansion and
differentiation. Sorted basal stem cells in culture were infected
with lentiviruses that expressed GFP and short hairpin RNAs
targeting Yap. Costaining for GFP and Yap demonstrated that
infected cells lost Yap protein expression, whereas control cells
showed robust Yap staining that was detectable even without
amplification in culture (Figure S2E). We first examined the effect
of Yap knockdown on cell death and cell proliferation. As we
found in vivo, there was no discernible difference in apoptosis
(as marked by the percentage of activated caspase-3+ cells
within the virally infected and therefore GFP+ cell population)
following Yap knockdown (Figure S2F). However, knockdown
of Yap did decrease cell proliferation as marked by Ki-67 in
GFP+ transfected cells (Figure S2G), although a significant frac-
alleles, YFP+ (green) and CC10-expressing (red, middle) secretory cells and the
detected. White arrows point to double-positive cells.
(D) Scgb3A2 (green, top) and FoxJ1 (green, bottom) are expressed in cells in wh
(arrows) go on to terminally differentiate.
(E) Quantification of Scgb3A2+ secretory cells and FoxJ1+ ciliated cells per trachea
CK5-YapKO animals.
(F) Quantification of the percentage of p63+ cells as a proportion of virally infected
The large majority of these GFP+ infected cells differentiate into CK8+ cells (right
Data are presented as mean ± SEM. Scale bar represents 15 mm in (A), (B), and
tion of the infected cells were still proliferating (13.81% ± 1.68%;
Figure S2G). Presumably, we detected this effect on basal cell
replication in vitro and not following Yap loss in vivo because
our culture conditions mimic injury and are associated with
dramatically increased basal cell replication when compared to
the low degree of basal cell replication during homeostasis
in vivo. We next sought to determine the effect of Yap knock-
down on differentiation. When scrambled virus was used,
73.85% ± 1.43% of infected GFP+ cells continued to express
p63, but this percentage dramatically decreased to 3.26% ±
0.70% after Yap knockdown (p < 0.0001; Figures 2F and S2H).
Although our in vitro platform does not support the terminal
differentiation of secretory or ciliated cells, infected GFP+ cells
went on to express the early differentiation marker cytokeratin
8 (CK8) (Rock et al., 2011; Figure S2H). In the scrambled control,
only 22.62% ± 0.64% of the cells differentiated and expressed
CK8 at day 9 of culture, whereas this fraction increased to
97.41% ± 2.59% following Yap knockdown (p < 0.0001; Figures
2F and S2H). This result further confirms that Yap loss promotes
stem cell differentiation.
Yap Overexpression Promotes Stem Cell Proliferationand Inhibits Terminal Differentiation Resulting inEpithelial Hyperplasia and StratificationTo assess the effect of Yap overexpression on basal stem cells,
we generated double transgenic animals carrying both the CK5
rtTA and tetO Yap (S127A; Camargo et al., 2007) alleles (referred
to as CK5-YapTg). In these animals, the expression of constitu-
tively active Yap (S127A) is induced specifically in CK5+ cells
(Figures 3A and S3). Siblings carrying the CK5rtTA driver alone
were used as controls. Although TSA amplification is required
to detect endogenous Yap, unamplified Yap staining unambigu-
ously identifies only Yap-overexpressing cells in CK5-YapTg
transgenic mice. Using unamplified Yap staining, ectopic Yap
expression was detected only in CK5+ basal stem cells 7 days
after doxycycline administration (Figure S3). Twenty-one days
after doxycycline initiation, a thickened and highly stratified
airway epithelium was evident (Figure 3B).
We then assessed the effect of Yap overexpression specif-
ically on basal stem cells. Immunohistochemical characteriza-
tion of CK5 and p63 in the stratified epithelium at day 21 revealed
that there was a significant increase in the total number of both
CK5+ and p63+ cells (Figures 3C and 3D). As compared to the
controls, which contained 1,082 ± 53 p63+ basal stem cells
per tracheal section, this number increased to 1,975 ± 223
(p = 0.0178) in the CK5-YapTg mice 21 days after continuous
doxycycline administration (Figures 3D and 3F, left graph).
We also examined the effect of Yap overexpression on basal
stem cell proliferation in CK5-YapTg animals. Following Yap
YFP+ (green) and FoxJ1-expressing (red, bottom) ciliated cells are frequently
ich Yap (red) is absent, demonstrating that basal stem cells that have lost Yap
l section reveals a significant increase in differentiated epithelial cell numbers in
GFP+ cells reveals that basal stem cells are lost following Yap knockdown (left).
).
(D, top), and 10 mm in (C) and (D, bottom). See also Figure S2.
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 5
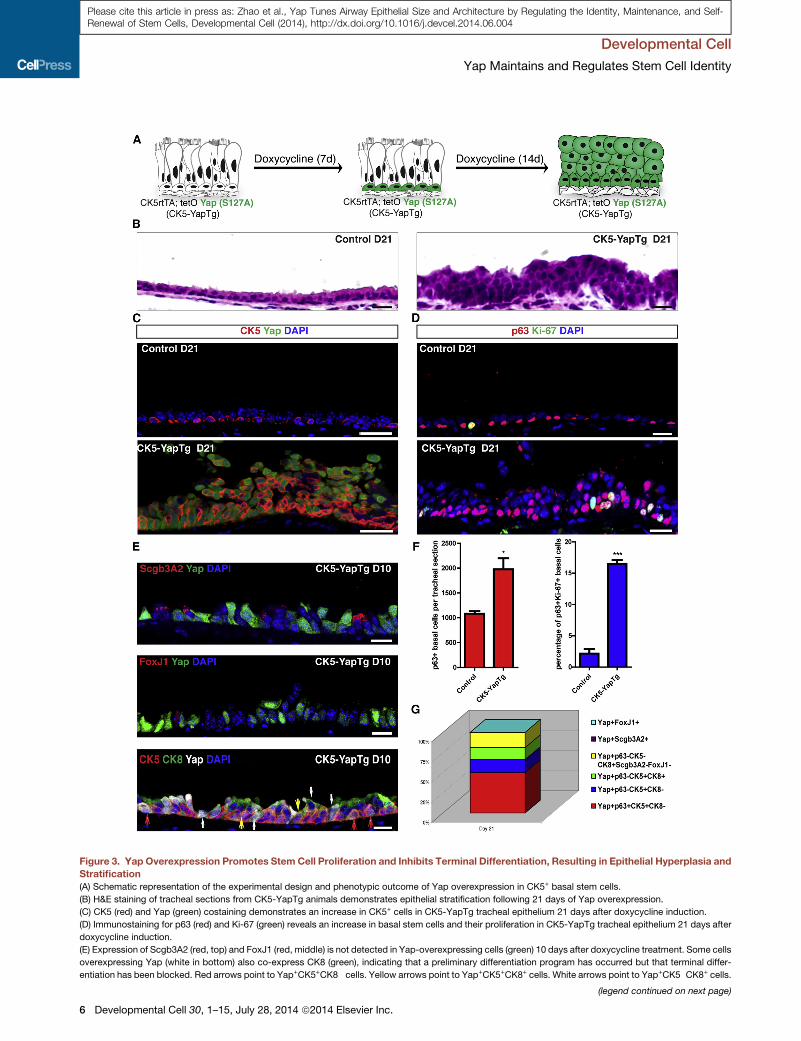
Figure 3. Yap Overexpression Promotes Stem Cell Proliferation and Inhibits Terminal Differentiation, Resulting in Epithelial Hyperplasia and
Stratification(A) Schematic representation of the experimental design and phenotypic outcome of Yap overexpression in CK5+ basal stem cells.
(B) H&E staining of tracheal sections from CK5-YapTg animals demonstrates epithelial stratification following 21 days of Yap overexpression.
(C) CK5 (red) and Yap (green) costaining demonstrates an increase in CK5+ cells in CK5-YapTg tracheal epithelium 21 days after doxycycline induction.
(D) Immunostaining for p63 (red) and Ki-67 (green) reveals an increase in basal stem cells and their proliferation in CK5-YapTg tracheal epithelium 21 days after
doxycycline induction.
(E) Expression of Scgb3A2 (red, top) and FoxJ1 (red, middle) is not detected in Yap-overexpressing cells (green) 10 days after doxycycline treatment. Some cells
overexpressing Yap (white in bottom) also co-express CK8 (green), indicating that a preliminary differentiation program has occurred but that terminal differ-
entiation has been blocked. Red arrows point to Yap+CK5+CK8� cells. Yellow arrows point to Yap+CK5+CK8+ cells. White arrows point to Yap+CK5�CK8+ cells.
(legend continued on next page)
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
6 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
overexpression, the percentage of p63+ basal stem cells that
were positive for Ki-67 increased from 1.23% ± 0.40% in the
control to 16.47% ± 0.60% (p = 0.0001) at day 21 following
doxycycline initiation (Figures 3D and 3F, right graph). Thus,
Yap overexpression promotes stem cell self-renewal.
We then assessed the effect of Yap overexpression on differ-
entiation. Only 0.26% (n = 2,667) of Yap-overexpressing cells
were positive for the secretory cell marker Scgb3A2 at day 10 af-
ter doxycycline administration (Figure 3E, top). At day 21, only a
single Scgb3A2+ cell was found out of 1,299Yap-overexpressing
cells. With respect to ciliated cell differentiation, only 0.22%
(n = 1,349) of Yap-overexpressing cells were colabeled with
FoxJ1 at day 10 (Figure 3E, middle). Not a single FoxJ1+Yap+
cell was detected at day 21 (n = 2,097). Therefore, Yap overex-
pression in CK5-YapTg trachea blocks terminal differentiation.
We also note that Yap overexpression results in an abundance
of CK5+p63�(negative) cells not normally present in the homeo-
static airway epithelium (Figures 3C and 3D). This suggests the
presence of a population of progenitor cells that are not basal
stem cells because they are p63�, but which are blocked from
executing a terminal differentiation program. Quantification re-
vealed that 81.09% (n = 3,229) of Yap overexpressing cells
were CK5+, but only 50.13% were double positive for p63 and
CK5 and thus bona fide basal stem cells. We then assessed
whether the remaining 30.96% of p63�CK5+ Yap-overexpress-
ing cells expressed the luminal progenitor cell marker CK8. Of
the total 30.96% p63�CK5+ progenitor cells, 16.30% were
p63�CK5+CK8� and 14.66% were p63�CK5+CK8+ (Figure 3G).
The remaining CK5� Yap overexpressing cells were exclusively
CK8+ luminal progenitor cells (Figure 3E, bottom and Figure 3G)
that did not initiate a terminal differentiation program. These
results, in aggregate, suggest the presence of a group of stem
cell-derived progenitor cells in varying states of differentiation,
which were prevented from completing terminal differentiation
(Figure 3G). Of note, these progenitors (p63�CK5+CK8�,p63�CK5+CK8+, p63�CK5�CK8+ cells), are exceedingly rare in
the homeostatic tracheal epithelium, but they do occur as orderly
transient intermediates in regenerating airway epithelium (Rock
et al., 2011).
Normalizing Yap Expression Restores PseudostratifiedEpithelial Architecture and a Normally Sized Basal StemCell PoolTo determine whether the maintenance of Yap-induced stem
cell proliferation and epithelial stratification requires persistent
Yap overexpression, we first administered doxycycline to exper-
imental CK5-YapTg animals for 20 days and then withdrew
doxycycline for 20 days. At the end of the chase period, we
observed that the airway epithelium wasmorphologically normal
(Figures 4A and 4B). There was no significant difference between
control and CK5-YapTg animals in the total number of CK5+
basal stem cells per section (Figures 4C and S4A). Additionally,
(F) Graphs showing the total number of p63+ cells (left) and the percentage of pro
numbers and their proliferation rates increase after Yap overexpression.
(G) A column chart showing the relative abundance of p63+CK5+CK8� basal stem
cells (green), p63�CK5�CK8+Scgb3A2�FoxJ1� progenitor cells (yellow), Scgb
expressing epithelial cells 21 days after doxycycline treatment.
Data are presented as mean ± SEM. Scale bar represents 10 mm in (B), (D), and
the proportion of CK5+ cells per total cells present in the epithe-
lium and the percentages of proliferating basal cells marked by
both CK5 and Ki-67 were not statistically different from those
in control animals (Figures 4C, S4B, and S4C). Thus, persistent
Yap overexpression is required tomaintain excess stem cell pro-
liferation and epithelial stratification.
We next examined the effect of Yap normalization on terminal
differentiation using lineage analysis. Triple transgenic mice pos-
sessing CK5 rtTA, tetO Yap (S127A), and tetO histone 2B GFP
(H2BGFP) alleles (referred to as CK5-YapTg-GFP) were gener-
ated. In these animals, the expression of the active Yap allele
and the H2BGFP reporter is induced in concert, under the con-
trol of the CK5rtTA driver. Because the H2BGFP protein has a
very long half-life, GFP labeling persists in the descendants of
basal stem cells. This persistence of the GFP label allows
the use of GFP expression as a lineage-tracing tool to identify
cells that had once overexpressed Yap (Figure 4D). Following
20 days of doxycycline administration, 98.56% (n = 761) of
GFP+ cells overexpressed Yap protein (Figure 4E, left). Although
the GFP lineage label persisted 2 weeks after doxycycline with-
drawal, ectopic Yap expression was lost within 1 week (Fig-
ure 4E, right), confirming that persistent H2BGFP can be used
as an effective lineage tracing tool for marking cells that previ-
ously overexpressed Yap. Two weeks after doxycycline with-
drawal, GFP+ lineage-traced stem cell progeny included cells
positive for mature secretory (Scgb3A2) and ciliated (FoxJ1)
cell markers (Figure 4F). Thus, stem and progenitor cells that pre-
viously expressed ectopic Yap underwent terminal differentia-
tion after Yap withdrawal. We further confirmed this result with
stains for CC10 (secretory cells) and acetylated tubulin (A-tub,
ciliated cells) to ensure that differentiated cells expressed a full
panel of markers associated with terminal differentiation (Fig-
ure S4D). Additionally, immunostaining revealed normally posi-
tioned and shapedCK5+ basal stem cells, CC10+ secretory cells,
and A-tub+ ciliated cells in an architecturally normal pseudostra-
tified epithelium 20 days after doxycycline withdrawal (Fig-
ure S4E). Therefore, when Yap expression was normalized, a
normally sized epithelium was restored with a normally sized
basal stem cell pool and the epithelium was characterized by a
proper pseudostratified architecture.
The Overexpression of Yap in Secretory Cells CausesThem to Adopt a Basal-like ProgramThus far, we have demonstrated that Yap is required for the
maintenance of adult basal stem cells and that Yap overexpres-
sion promotes stem cell proliferation at the expense of terminal
differentiation. These findings suggest the possibility that Yap
overexpression in a differentiated cell could promote a stem
cell-like program. To test this hypothesis, we overexpressed
Yap in secretory cells. We generated triple transgenic mice
carrying the CC10-CreER (Rawlins et al., 2009), Rosa26-LSL-
rtTA-IRES-GFP, and tetO Yap (S127A) alleles (referred to as
liferating basal stem cells (p63+ Ki-67+, right) demonstrate that basal stem cell
cells (red), p63�CK5+CK8� progenitor cells (blue), p63�CK5+CK8+ progenitor3A2+ secretory cells (purple), and FoxJ1+ ciliated cells (cyan) in Yap-over-
(E), and 20 mm in (C). See also Figure S3.
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 7

Figure 4. Normalizing Yap Expression Restores Pseudostratified Epithelial Architecture and a Normally Sized Basal Stem Cell Pool
(A) A schematic of the experimental strategy and phenotypic result of an experiment designed to test whether normalizing Yap signaling in a stratified epithelium
restores normal basal stem cell numbers and pseudostratified epithelial architecture.
(B) H&E staining reveals the restoration of a normal pseudostratified epithelium after the discontinuation of doxycycline-induced Yap overexpression.
(C) Immunostaining for CK5+ basal stem cells (red) and Ki-67 (green) reveals a normalization of stem cell numbers and proliferation rates following doxycycline
withdrawal. The arrow points to a basal stem cell that is double positive for CK5 and Ki-67.
(D) Experimental strategy for using long-lived H2BGFP expression as a lineage tracing tool for tracing the behavior of stem and progenitor cells that have
previously overexpressed Yap.
(E) Expression of GFP (green) is detected in the majority of the cells that overexpress Yap (red, left) following doxycycline treatment. Ectopic Yap expression is no
longer detected following 1 week of doxycycline withdrawal, whereas strong GFP lineage label endures (green, right).
(F) Scgb3A2 (red, left) and FoxJ1 (red, right) occur in H2BGFP+ (green) cells 2 weeks after doxycycline removal (arrows point to double-positive cells).
Scale bar represents 20 mm in (B), (C), (E), and (F). See also Figure S4.
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
8 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Figure 5. The Overexpression of Yap in
Secretory Cells Causes Them to Adopt a
Basal-like Program
(A) Schematic of the strategy to inducibly over-
express Yap in GFP lineage tagged secretory
cells. Following Yap induction, some GFP tagged
secretory cells adopt a basal stem cell-like identity.
(B) Costaining of the secretory cell lineage tag GFP
with the basal stem cell marker T1a (red) shows
that GFP is associated with normal columnar cells
and not basal stem cells in control animals (left). In
experimental animals, GFP+ T1a+ basal-like cells
derived from Yap-overexpressing secretory cells
are observed (right, arrows).
(C) A pyramidal basal-like cell expressing p63 (red)
and ectopic Yap (green) reveals that the stem-
like cell was derived from a Yap-overexpressing
secretory cell (arrow).
(D) Expression of Scgb3A2 (red) is lost in pyrami-
dal Yap-overexpressing (green) secretory-derived
cells (arrow).
(E) Yap knockdown using GFP-expressing lenti-
virus prevents the dedifferentiation of secretory
cells into basal stem cells as demonstrated by the
absence of p63 (red, top) and CK5 (red, bottom)
staining in GFP+ infected cells as opposed to
controls. Arrowheads point to GFP+ cells infected
with control viruses that do dedifferentiate into
p63+ and CK5+ basal stem cells.
(F) Quantification of p63+ and CK5+ basal stem
cells as a fraction of all GFP+ infected cells in
scrambled control and Yap knockdown secretory
cell cultures reveals that inhibition of Yap prevents
secretory cell dedifferentiation into stem cells.
Data are presented as mean ± SEM. Scale bar
represents 20 mm in (B), (C), and (D) and 100 mm in
(E). See also Figure S5.
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
CC10-Yap mice). In these mice, CreER protein is expressed
exclusively in secretory cells. Upon tamoxifen administration, a
multicistronic transcript containing rtTA and GFP is permanently
induced in these cells and doxycycline administration then
induces Yap overexpression specifically in these GFP lineage-
tagged secretory cells (Figure 5A).
Tamoxifen was administered to adult CC10-Yap transgenic
mice (8–10 weeks old) and then followed by doxycycline induc-
tion. Fluorescence-activated cell sorting analysis demonstrated
that GFP+ cells accounted for 3.94% of the total epithelial cells
(Figure S5A). In control animals possessing the CC10-CreER
and Rosa26-LSL-rtTA-IRES-GFP alleles, GFP expression was
appropriately restricted to columnar secretory cells (Figure 5B,
left) and entirely absent in basal stem cells. In experimental ani-
mals, some GFP+ cells exhibited the characteristic pyramidal
morphology of basal stem cells and expressed the basal stem
cell-specific marker T1a (Figure 5B, right). These pyramidal cells
established a broad base in direct contact with the basement
membrane and stained for ectopic Yap, as assessed by Yap
staining without amplification (Figure S5B). Furthermore, the
pyramidal GFP+ Yap-overexpressing cells also expressed the
basal stem cell-specificmarker p63 (Figure 5C). Of all GFP+ cells,
we note that only 79.30% continued to express Scgb3A2
(Figure 5D) and 8.06% became p63+ basal-like cells (n = 589).
Thus, in addition to adopting a basal-like program, secretory
cells that had once overexpressed Yap also suppressed ele-
ments of their secretory cell program.
To test whether the secretory cell-derived partially reprog-
rammed basal-like cells persisted following the normalization
of Yap expression, we administered doxycycline to CC10-Yap
mice for 2 weeks and then withdrew doxycycline for 3 weeks.
After this period, there were no GFP+ basal-like cells (n = 561).
Thus, persistent overexpression of Yap was required to maintain
the partially reprogrammed basal-like state that is transiently
induced by Yap overexpression in secretory cells.
The Inhibition of Yap Blocks the Dedifferentiation ofSecretory Cells into Basal Stem CellsWe recently demonstrated that secretory cells cultured ex vivo
dedifferentiate into p63+ CK5+ basal stem cells (Tata et al.,
2013a). To further examine the role of Yap in regulating stem
cell identity, we performed Yap lentiviral knockdown experi-
ments in secretory cell cultures to assess whether Yap loss
alters the propensity of a secretory cell to dedifferentiate. In
contrast to controls, secretory cells infected with a Yap knock-
down lentivirus failed to dedifferentiate into stem cells and did
not activate p63 expression 9 days after plating (Figures 5E
and 5F). Among the GFP+ cells infected with scrambled virus,
p63+ basal stem cells were easily detected and accounted
for 24.61% ± 0.96% of all GFP+ cells. Among the GFP+ cells
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 9

Figure 6. Yap and p63 Interact in Basal
Stem Cells and Regulate Common Target
Genes
(A) Quantitative-PCR analysis demonstrates that
DNp63a is the major p63 transcript expressed in
sorted basal stem cells.
(B) Immunoprecipitation demonstrates that Yap
interacts with p63 in human basal stem cells.
(C) Quantitative-PCR analysis of p63 target
genes demonstrates that these targets are more
enriched in basal stem cells.
(D) ChIP-PCR analysis demonstrates that Yap
binds to the promoters of p63 target genes.
(E) Decrease in mRNA expression of p63 target
genes following Yap knockdown.
(F) Gene expression analysis shows an increase in
mRNA expression of p63 target genes following
Yap overexpression, but this effect is abolished
when p63 is knocked down.
Data are presented as mean ± SEM. See also
Figure S6.
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
infected with Yap virus, p63+ cells decreased to only 1.18% ±
0.51% of the total population (p < 0.0001; Figure 5F). We
confirmed the finding with another basal stem cell marker
CK5 (Figures 5E and 5F). In cells infected with the scrambled
control virus, 41.33% ± 6.2% of GFP-expressing cells were
CK5+ 9 days after culture. This number dramatically decreased
to 1.23% ± 0.79% (p < 0.0001) when Yap was knocked down
(Figures 5E and 5F). These results suggest that Yap is required
for the dedifferentiation of secretory cells and their adoption of a
stem cell program.
Yap and p63 Interact in Basal Stem Cells and RegulateCommon Target GenesBecause the transcription factor p63 is the most well-estab-
lished regulator of basal stem cells (Koster et al., 2004; Romano
et al., 2012; Su et al., 2013), we sought to determine whether
there was a direct mechanistic interaction between p63 and
Yap. Although p63 has previously been shown to interact with
Yap in cell lines (Chatterjee et al., 2010; Strano et al., 2001;
Yuan et al., 2010), the functional relevance of such an interaction
has not been established in vivo.
10 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
p63 has two isoforms, TA and delta
N (DN), each of which has three splice
variants: a, b, and g (Yang et al., 1998).
Quantitative PCR revealed that TAp63
splice variants were expressed at very
low levels in basal stem cells, and that
DNp63a was by far the most abundant
transcript in basal cells in vivo and in
cultured basal cells (Figures 6A and
S6A). Staining with two DNp63-specific
antibodies showed that the expression
of DNp63 was exclusively restricted
to the basal stem cell population (Fig-
ure S6B). We then verified the previously
reported interaction of Yap and DNp63a
in HEK cells using Myc-tagged Yap and
Flag-tagged DNp63a (Figure S6C). To extend these findings to
our model system, we performed reciprocal immunoprecipita-
tions using primary human basal stem cells and found that Yap
and p63 physically interact with one another specifically in
airway epithelial basal stem cells (Figure 6B). Of note, the size
of the precipitated p63 in the Yap pull-down demonstrates that
the Yap partner is DNp63.
We then examined whether Yap modulation had an effect on
the transcription of p63 target genes in mouse basal stem cells.
In other contexts, p63 has been shown to regulate the transcrip-
tion of CK5, EGFR, FGFR2, Integrin alpha 6 (Igta6), Integrin b4
(Igtb4), and p63 itself (Carroll et al., 2006; McDade et al., 2012;
Romano et al., 2009). All of these p63 target genes were more
enriched in basal stem cells relative to secretory and ciliated
cells (Figure 6C). We verified that p63 binds upstream of the
transcriptional start sites of these genes with chromatin immu-
noprecipitation (ChIP)-PCR (Figure S6D). Additionally, DNp63
knockdown in mouse basal stem cell cultures led to a decrease
in the expression of these target genes after 6 days (Figures
S6E–S6H). Using ChIP-PCR analysis, we found that Yap also
binds upstream of the transcriptional start sites of the

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
aforementioned p63 target genes (Figure 6D). Furthermore, Yap
knockdown resulted in a decrease in the expression of p63
target genes (Figure 6E). Conversely, the transcription of all
p63 target genes was significantly increased 12 hr after doxycy-
cline was administered to induce the expression of Yap in sorted
basal stem cells fromCK5-YapTg trachea, as compared to those
from CK5rtTA controls (Figure 6F). To directly test whether the
transcriptional effect caused by Yap overexpression was medi-
ated by p63, we knocked down p63 while overexpressing Yap.
We harvested and analyzed these cells after 6 days of culture,
because at this time point the majority of infected cells remained
p63+CK5+ basal stem cells, although p63 expression was
decreased given the knockdown (Figure S6E). We found that in-
creases in the transcription of p63 target genes in Yap-overex-
pressing basal cells were significantly suppressed when p63
was inhibited (Figure 6F). In aggregate, these data demonstrate
that Yap and p63 interact in basal stem cells to regulate common
target genes.
p63 and Yap Genetically InteractFinally, we sought to establish whether genetic and phenotypic
data could confirm an in vivo relevance for our biochemical
data demonstrating an interaction of Yap and p63. Specifically,
we asked whether p63 loss would phenocopy Yap loss. Trans-
genic animals carrying the CK5 rtTA, tetO Cre, and floxed p63
alleles (Mills et al., 2002) (referred to as CK5-p63KO) were gener-
ated, and we hypothesized that stem cell loss would occur
following p63 deletion in analogy to the phenotype associated
with Yap deletion (Figure 7A). Using animals bearing only the
CK5 rtTA and tetO Cre alleles as controls, our histological anal-
ysis revealed that p63 deletion resulted in a simplification of
the normal pseudostratified epithelium into a simple columnar
epithelium 3 weeks after doxycycline treatment (Figure 7B).
This was accompanied by a significant decrease in the total
number of p63+ cells (Figure 7C), demonstrating efficient dele-
tion of p63. A loss of CK5+ and T1a+ cells (Figures 7C and
S7A) confirmed the loss of basal stem cells in CK5-p63KO ani-
mals. Controls had 1,184 ± 15 CK5-expressing cells per stan-
dard tracheal section. After doxycycline treatment, this number
decreased to 563 ± 67 (p < 0.001) in 2 weeks and to 126 ± 15
(p < 0.0001) in 3 weeks (Figure 7E). Although we functionally
demonstrated that DNp63 was the predominant isoform of p63
in basal stem cells, to further verify this point, we analyzed the
effect of TAp63 loss on the tracheal epithelium of TAp63 null
animals (Su et al., 2009). No changes in the histology of the
tracheal epithelium and the expression of basal stem cell
markers including DNp63 and CK5 were found in the mutant
TAp63 trachea (Figures S7B and S7C). Furthermore, quantifica-
tion of CK5+ and DNp63+ cells in the mutant and control trachea
revealed no significant differences in the number of basal stem
cells per standard section (Figure S7D), indicating that TAp63
is dispensable for the maintenance of basal stem cells in the tra-
chea during homeostasis. These data together demonstrate that
p63, specifically DNp63, is required for the maintenance of an
adult airway basal stem cell.
We next sought to determine whether basal stem cells under-
went differentiation following p63 loss by assessing whether
transitional cells expressing both basal and differentiated cell
markers were induced. Indeed, a significant increase in cells
double positive for CK5 and CC10 or CK5 and FoxJ1 occurred
(Figure 7D), suggesting that basal stem cells were differentiating
into both secretory and ciliated cells in the absence of p63. Simi-
larly, p63 knockdown in basal stem cell cultures at day 10
decreased the number of cells that were positive for the basal
stem cell marker CK5 (Figure S7E, top) and increased the num-
ber of cells expressing the early differentiation marker CK8 (Fig-
ure S7E, bottom). Taken together, these results demonstrate
that p63, like Yap, is required for the maintenance of adult basal
stem cells.
Finally, we sought to determine whether Yap and p63 geneti-
cally interact. To do this, we examined whether the removal
of one copy of p63 could suppress the phenotype generated
by Yap overexpression. Transgenic animals bearing CK5rtTA,
tetO Yap (S127A), tetO Cre, and a single floxed p63 allele were
generated. Siblings carrying CK5rtTA and tetO Yap (S127A) as
well as animals with CK5rtTA, tetO Cre and floxed p63 were
used as controls. All animals were given doxycycline continu-
ously for 10 days. There was no obvious effect on basal stem
cell maintenance or airway epithelial architecture when one
copy of p63 was deleted at homeostasis (Figure S7F, top). As
we observed previously, epithelial stratification was induced
when Yap was activated in CK5-YapTg animals (Figure S7F,
middle). Epithelial stratification was suppressed when one
copy of p63 was removed (Figure S7F, bottom). Similarly, the
increases in basal stem cell numbers and their proliferation
rates following Yap activation were significantly lessened when
one copy of p63 was removed (Figures 7F and 7G). Ten days
after doxycycline treatment, 1,331 ± 54 basal stem cells marked
by p63 per tracheal section were detected following Yap overex-
pression. This number significantly decreased to 1,143 ± 41
(p = 0.049) when one copy of p63 was removed (Figure 7F).
With regard to basal stem cell proliferation, 8.16% ± 1.00% of
p63+ cells were also Ki-67+ when Yap was overexpressed.
This number significantly decreased to 4.32% ± 0.71% (p =
0.036) when one copy of p63 was removed (Figure 7G). These
data suggest that the effects of Yap overexpression are medi-
ated by p63 and that the two factors genetically interact in the
airway epithelium.
DISCUSSION
Wedemonstrate that the regulation of basal stem cell behavior is
intimately associated with epithelial size and architecture in the
adult airway epithelium. On one hand, basal stem cell loss results
in the simplification of a pseudostratified airway epithelium into a
columnar one. Conversely, Yap overexpression increases stem
cell proliferation, resulting in epithelial hyperplasia and stratifica-
tion. Although one might assume that epithelial size and stratifi-
cation are largely governed by the increased replicative activity
of a stem cell, we note that suprabasal progenitor cells (not nor-
mally present in the homeostatic airway epithelium) also divide,
just as they do in embryonic epidermis (Lechler and Fuchs,
2005). Indeed, after airway injury, stratification is similarly
accompanied by replication in both a stem cell and a newly re-
vealed progenitor cell compartment. The nature of the cell divi-
sions (i.e., symmetric versus asymmetric) occurring in these
two compartments is not understood and is possibly distinct.
Interestingly, in other instances of epithelial stratification such
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 11

Figure 7. p63 and Yap Genetically Interact
(A) A schematic of the strategy for basal stem cell-specific p63 deletion. It is hypothesized that p63 deletion results in a loss of basal stem cells (red), mirroring Yap
deletion.
(B) H&E staining of tracheal sections from control (left) and experimental CK5-p63KO (right) animals reveals a simplification of a normally pseudostratified
epithelium into a columnar epithelium following p63 deletion. Control animals bear only the CK5 rtTA and tetO Cre alleles.
(C) Immunostaining for p63 (red) and CK5 (green) reveals a significant decrease in total basal stem cell numbers in CK5-p63KO tracheal sections as compared to
that in control mice possessing only the CK5 rtTA and tetO Cre alleles.
(legend continued on next page)
Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
12 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
as in the human larynx or esophagus, parabasal cells are the
dominant replicating cell population, and the putative basally
located stem cells are more quiescent (data not shown). Thus,
stem and progenitor cell behaviors may be differentially regu-
lated to produce alternative paths to epithelial stratification. Of
note, premalignant airway squamous metaplasia is also associ-
ated with both stem and progenitor cell replication and excess
(Rock et al., 2010) and our Yap overexpression system may
serve to model some aspects of this pathology.
The size of a steady-state stem cell pool is dictated by
balanced rates of stem cell self-renewal, stem cell survival,
and stem cell differentiation. Previous reports have largely
focused on the role of Yap in promoting stem cell and progen-
itor cell replication and survival (Barry et al., 2013; Cai et al.,
2010; Cao et al., 2008; Schlegelmilch et al., 2011). In contrast,
our loss-of-function study points to a requirement for Yap in re-
straining stem cell differentiation as a mechanism to ensure the
maintenance of stem cells during normal epithelial homeosta-
sis. Indeed, we also demonstrated a dose-dependent effect
of Yap on the maintenance of basal stem cells, suggesting
that the tight control of Yap levels is critical to maintain a nor-
mally sized basal stem cell pool. The notion of a set point for
the airway stem cell pool contingent on particular levels of
Yap expression is further evidenced by the return of a normal
stem cell pool size when Yap expression is normalized after
its overexpression.
Given that Yap overexpression promotes basal stem cell pro-
liferation, it will be of interest to determine whether Yap levels
are transiently increased following airway epithelial injury repair
because the early stratification that accompanies regeneration
is also associated with an increased rate of basal cell prolifera-
tion. This notion would be consistent with our finding that Yap
knockdown inhibits stem cell proliferation in vitro, as dissociation
and culture of basal cells induces a proliferative response similar
to that encountered after airway injury. Analogously, the differen-
tiation associated with late epithelial repair might be associated
with a tuning down of Yap levels.
Furthermore, we revealed a role for Yap in regulating stem cell
identity per se. Previously, we demonstrated that secretory cells
can stably dedifferentiate into stem cells (Tata et al., 2013a).
We now show that Yap loss prevents this dedifferentiation.
Conversely, Yap overexpression in secretory cells promotes a
stem cell-like identity and concomitantly suppresses the secre-
tory cell program. Interestingly, persistent Yap overexpression
is required to maintain the stem cell-like identity despite the
induction of the basal stem cell transcription factor p63. Thus,
the induction of p63 in secretory cells is insufficient to stably
reprogram differentiated cells into stem cells within the time
we assessed. We note that following Yap overexpression in
secretory cells, there aremore cells that suppress their secretory
(D) Detection of transitional cells double positive for CK5 (red) and CC10 (green; le
after doxycycline administration. Arrows point to the double-positive cells.
(E) Quantification of CK5+ basal stem cells per standard tracheal section followin
(F) Quantification of p63+ cell numbers reveals that a loss of one copy of p63 sig
overexpression.
(G) Quantification of p63+ Ki-67+ cells reveals that the loss of one copy of p63 sign
overexpression.
Data are presented as mean ± SEM. Scale bar represents 5 mm in (B), and 10 mm
cell identity than cells that adopt a basal cell-like program,
implying the existence of transitional progenitor cells. Similarly,
Yap overexpression from basal stem cells produces more basal
cells, but Yap overexpression is also associated with an excess
of intermediate progenitor cells, again suggesting that Yap alone
cannot enforce a basal stem cell identity in which differentiation
is fully blocked. It will be of considerable interest to determine
why Yap overexpression partially reprograms secretory cells,
while stem cell ablation induces a permanent conversion to a
stem cell state. Although in aggregate very high levels of Yap
expression seem to drive cells toward a stem cell-like identity,
the low levels of Yap detected in secretory cells, as well as cili-
ated cells, of the homeostatic epithelium may be of some func-
tional relevance that we have not yet assessed.
The interaction of the Yap transcriptional coactivator with
TEAD partners has been well established to have an in vivo phys-
iologic relevance (Yu and Guan, 2013). Other partner transcrip-
tion factors have been shown to interact with Yap in cultured
cells, including Smad 1 (Alarcon et al., 2009), Smad 7 (Ferrigno
et al., 2002), RUNX 1/2 (Yagi et al., 1999), ErbB4 (Komuro
et al., 2003), p73, and p63 (Strano et al., 2001). In this study,
we have now demonstrated that a WW domain (Yap)-PPxY
(p63) interaction is physiologically relevant in an in vivo setting
(Chen and Sudol, 1995; Sudol and Harvey, 2010). Previously, it
had been shown that p63 null mutants die at birth without embry-
onic basal stem cell progenitors in their airway epithelium (Dan-
iely et al., 2004), either because basal stem cells were never
specified or because they could not be maintained after specifi-
cation. We now show that p63, like Yap, is required for the actual
maintenance of adult airway basal stem cells. In addition, both
Yap loss and p63 loss are ultimately associated with an excess
of ciliated cell numbers relative to secretory cell numbers.
Furthermore, p63 overexpression had been shown to promote
epithelial stratification in the murine lung (Koster et al., 2004;
Romano et al., 2009). In aggregate, this suggests a functionally
relevant role for an interaction of Yap with a new transcription
factor partner. This interaction may be relevant in the myriad
epithelia that contain p63+ basal cells. Indeed, prior work in the
epidermis reveals a role for both p63 and Yap in epidermal basal
stem cell proliferation and epithelial stratification (Koster et al.,
2004; Romano et al., 2012; Schlegelmilch et al., 2011; Zhang
et al., 2011). Finally, because Yap and p63 both have roles as tu-
mor suppressors and oncogenes (Overholtzer et al., 2006; Pan,
2010; Su et al., 2013), our results have potential implications for
understanding tumorigenesis.
EXPERIMENTAL PROCEDURES
Detailed protocols are described in the Supplemental Experimental
Procedures.
ft) as well as CK5 (red) and FoxJ1 (green; right) in CK5-p63KO trachea 2 weeks
g p63 loss.
nificantly suppresses the increase in basal stem cells numbers caused by Yap
ificantly suppresses the increased basal stem cell proliferation caused by Yap
in (C) and (D). See also Figure S7.
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 13

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
Animal Models
TheCK5 rtTA (Diamond et al., 2000),CK5-CreER (Van Keymeulen et al., 2011),
CC10-CreER (Rawlins et al., 2009), tetO Yap (S127A; Camargo et al., 2007),
floxed Yap (Schlegelmilch et al., 2011), floxed p63 (Mills et al., 2002), and
TAp63 knockout (Su et al., 2009) mice have been previously described. Trans-
genic mice bearing tetO Cre (006234), tetO H2BGFP (005104), Rosa26-LSL-
YFP (006148), and Rosa26-LSL-rtTA-IRES-GFP (005670) were purchased
from Jackson laboratory. Doxycycline was administered via drinking water
(1 g/l) and inhalation (Tata et al., 2013b). Tamoxifen (2 mg/20 g body weight)
or corn oil was injected intraperitoneally for 3 consecutive days in transgenic
animals carrying CK5-CreER, floxed Yap, and Rosa-LSL-YFP and their
respective control animals, and for 5 consecutive days in CC10-Yap animals
and their respective controls. Mice were killed using CO2. All animal work
was approved by the committee on Research Animal Care of the Massachu-
setts General Hospital according to federal and institutional policies and
regulations.
Cell Dissociation and Sorting of Different Fractions of Epithelial
Cells
Mouse tracheas were dissected, minced manually, and then incubated in
Papain dissociation solution. Cells were sorted using a fluorescence-activated
cell sorting Aria flow cytometer (BD) or a Vantage Cell Sorter (BD). Epithelial
cells were gated as EpCAM+ cells. Basal stem cells were sorted as EpCAM+
GSIb4+ cells (confirmed by staining for p63 after cytospin). Secretory cells
were sorted as EpCAM+ SSEA1+ cells (verified by staining with CC10 after
cytospin). Ciliated cells were sorted as EpCAM+ CD24+ cells (verification
with both Foxj1 and A-tub staining after cytospin).
Yap TSA Amplification
Immunohistochemical staining for Yap protein was performed using a
TSA Biotin Tyramide Amplification system (SAT700001EA, Perkin Elmer).
The following Yap antibodies were used: mouse anti-Yap (101199, Santa
Cruz); mouse anti-Yap (H00010413-M01, Abnova), and rabbit anti-Yap (kindly
provided by Joseph Avruch). When used at a 1:500 dilution, immunohisto-
chemical staining in the case of each antibody revealed a strong nuclear signal
in basal stem cells as compared to weaker staining in differentiated cells. To
confirm that cells were Yap-negative following Yap deletion, we deployed anti-
body staining at a 1:100 dilution so that we could detect even very low levels of
Yap protein expression.
Cell Counting
Cells were manually counted based upon immunofluorescent staining for
markers of specific cell types in at least three different tracheas with three to
five representative regions per tracheal section. For the quantification of total
basal stem cells, p63+ cells were counted along the full length of the tracheal
epithelium extending from cartilage ring 1 to 12 in three to five tracheas with
two to three 6-mm thick sections counted per trachea.
Coimmunoprecipitation and ChIP
Detailed protocols for coimmunoprecipitation and ChIP are provided in the
Supplemental Experimental Procedures.
Statistical Analysis
Data were analyzed and compared between groups using two tailed, unpaired
Student’s t tests (Prism, Graphpad). A p < 0.05 was considered statistically
significant and is presented as * p < 0.05, **p < 0.01, or ***p < 0.001.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures
and seven figures and can be found with this article online at http://dx.doi.
org/10.1016/j.devcel.2014.06.004.
ACKNOWLEDGMENTS
We thank Adam Glick and Alea Mills for providing the CK5rtTA and p63 floxed
mice, Elsa Flores for the TAp63 knockout mice, Brigid Hogan for the CK5-
CreER and CC10-CreER mice, Barry Stripp for providing the goat anti-CC10
14 Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc.
antibody, Shioko Kimura for the rabbit Scgb3A2 antibody, Satrajit Sinha for
the rabbit DNp63 antibody, and Joseph Avruch for the rabbit Yap antibody.
We acknowledge XuWu and Jenna Galloway for their comments on themanu-
script. We thank Nicole Forster, Karin Schlegelmilch, Laura Prickett-Rice, Kat
Folz-Donahue, and David Dombkowski for technical assistance. J.R. is a New
York Stem Cell Foundation-Robertson Investigator. This study was supported
by an NIH-NHLBI Early Career Research New Faculty (P30) award
(5P30HL101287-02), a grant from the NHLBI Progenitor Cell Biology Con-
sortium (PCBC, 5U01HL099997-04; to J.R.), an R01 (1R01HL116756-01a)
from NIH/NHLBI (to J.R.), an NIDCR R01 DE015945 (to L.W.E. and S.V.S.),
and a Harvard Stem Cell Institute Junior Investigator Grant (to J.R.).
Received: January 12, 2014
Revised: April 7, 2014
Accepted: June 6, 2014
Published: July 17, 2014
REFERENCES
Alarcon, C., Zaromytidou, A.I., Xi, Q., Gao, S., Yu, J., Fujisawa, S., Barlas, A.,
Miller, A.N., Manova-Todorova, K., Macias, M.J., et al. (2009). Nuclear CDKs
drive Smad transcriptional activation and turnover in BMP and TGF-beta
pathways. Cell 139, 757–769.
Barry, E.R., and Camargo, F.D. (2013). The Hippo superhighway: signaling
crossroads converging on the Hippo/Yap pathway in stem cells and develop-
ment. Curr. Opin. Cell Biol. 25, 247–253.
Barry, E.R., Morikawa, T., Butler, B.L., Shrestha, K., de la Rosa, R., Yan, K.S.,
Fuchs, C.S., Magness, S.T., Smits, R., Ogino, S., et al. (2013). Restriction of
intestinal stem cell expansion and the regenerative response by YAP. Nature
493, 106–110.
Cai, J., Zhang, N., Zheng, Y., deWilde, R.F., Maitra, A., and Pan, D. (2010). The
Hippo signaling pathway restricts the oncogenic potential of an intestinal
regeneration program. Genes Dev. 24, 2383–2388.
Camargo, F.D., Gokhale, S., Johnnidis, J.B., Fu, D., Bell, G.W., Jaenisch, R.,
and Brummelkamp, T.R. (2007). YAP1 increases organ size and expands
undifferentiated progenitor cells. Curr. Biol. 17, 2054–2060.
Cao, X., Pfaff, S.L., and Gage, F.H. (2008). YAP regulates neural progenitor
cell number via the TEA domain transcription factor. Genes Dev. 22, 3320–
3334.
Carroll, D.K., Carroll, J.S., Leong, C.O., Cheng, F., Brown, M., Mills, A.A.,
Brugge, J.S., and Ellisen, L.W. (2006). p63 regulates an adhesion programme
and cell survival in epithelial cells. Nat. Cell Biol. 8, 551–561.
Chatterjee, A., Sen, T., Chang, X., and Sidransky, D. (2010). Yes-associated
protein 1 regulates the stability of DeltaNp63alpha. Cell Cycle 9, 162–167.
Chen, H.I., and Sudol, M. (1995). The WW domain of Yes-associated protein
binds a proline-rich ligand that differs from the consensus established for
Src homology 3-binding modules. Proc. Natl. Acad. Sci. USA 92, 7819–7823.
Daniely, Y., Liao, G., Dixon, D., Linnoila, R.I., Lori, A., Randell, S.H., Oren, M.,
and Jetten, A.M. (2004). Critical role of p63 in the development of a normal
esophageal and tracheobronchial epithelium. Am. J. Physiol. Cell Physiol.
287, C171–C181.
Diamond, I., Owolabi, T., Marco, M., Lam, C., and Glick, A. (2000). Conditional
gene expression in the epidermis of transgenic mice using the tetracycline-
regulated transactivators tTA and rTA linked to the keratin 5 promoter.
J. Invest. Dermatol. 115, 788–794.
Ferrigno,O., Lallemand, F., Verrecchia, F., L’Hoste, S., Camonis, J., Atfi, A., and
Mauviel, A. (2002). Yes-associated protein (YAP65) interacts with Smad7 and
potentiates its inhibitory activity against TGF-beta/Smad signaling. Oncogene
21, 4879–4884.
Halder, G., and Johnson, R.L. (2011). Hippo signaling: growth control and
beyond. Development 138, 9–22.
Komuro, A., Nagai, M., Navin, N.E., and Sudol, M. (2003). WW domain-con-
taining protein YAP associates with ErbB-4 and acts as a co-transcriptional
activator for the carboxyl-terminal fragment of ErbB-4 that translocates to
the nucleus. J. Biol. Chem. 278, 33334–33341.

Developmental Cell
Yap Maintains and Regulates Stem Cell Identity
Please cite this article in press as: Zhao et al., Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells, Developmental Cell (2014), http://dx.doi.org/10.1016/j.devcel.2014.06.004
Koster, M.I., Kim, S., Mills, A.A., DeMayo, F.J., and Roop, D.R. (2004). p63 is
the molecular switch for initiation of an epithelial stratification program. Genes
Dev. 18, 126–131.
Lechler, T., and Fuchs, E. (2005). Asymmetric cell divisions promote stratifica-
tion and differentiation of mammalian skin. Nature 437, 275–280.
McDade, S.S., Henry, A.E., Pivato, G.P., Kozarewa, I., Mitsopoulos, C.,
Fenwick, K., Assiotis, I., Hakas, J., Zvelebil, M., Orr, N., et al. (2012).
Genome-wide analysis of p63 binding sites identifies AP-2 factors as co-reg-
ulators of epidermal differentiation. Nucleic Acids Res. 40, 7190–7206.
Mills, A.A., Qi, Y., and Bradley, A. (2002). Conditional inactivation of p63 by
Cre-mediated excision. Genesis 32, 138–141.
Morrisey, E.E., and Hogan, B.L. (2010). Preparing for the first breath: genetic
and cellular mechanisms in lung development. Dev. Cell 18, 8–23.
O’Koren, E.G., Hogan, B.L., and Gunn, M.D. (2013). Loss of basal cells pre-
cedes bronchiolitis obliterans-like pathological changes in a murine model of
chlorine gas inhalation. Am. J. Respir. Cell Mol. Biol. 49, 788–797.
Overholtzer, M., Zhang, J., Smolen, G.A., Muir, B., Li, W., Sgroi, D.C., Deng,
C.X., Brugge, J.S., and Haber, D.A. (2006). Transforming properties of YAP,
a candidate oncogene on the chromosome 11q22 amplicon. Proc. Natl.
Acad. Sci. USA 103, 12405–12410.
Pan, D. (2007). Hippo signaling in organ size control. Genes Dev. 21, 886–897.
Pan, D. (2010). The hippo signaling pathway in development and cancer. Dev.
Cell 19, 491–505.
Ramos, A., and Camargo, F.D. (2012). The Hippo signaling pathway and stem
cell biology. Trends Cell Biol. 22, 339–346.
Rawlins, E.L., Okubo, T., Xue, Y., Brass, D.M., Auten, R.L., Hasegawa, H.,
Wang, F., and Hogan, B.L. (2009). The role of Scgb1a1+ Clara cells in the
long-term maintenance and repair of lung airway, but not alveolar, epithelium.
Cell Stem Cell 4, 525–534.
Rock, J.R., Onaitis, M.W., Rawlins, E.L., Lu, Y., Clark, C.P., Xue, Y., Randell,
S.H., and Hogan, B.L. (2009). Basal cells as stem cells of the mouse trachea
and human airway epithelium. Proc. Natl. Acad. Sci. USA 106, 12771–12775.
Rock, J.R., Randell, S.H., and Hogan, B.L. (2010). Airway basal stem cells:
a perspective on their roles in epithelial homeostasis and remodeling. Dis.
Model. Mech. 3, 545–556.
Rock, J.R., Gao, X., Xue, Y., Randell, S.H., Kong, Y.Y., and Hogan, B.L. (2011).
Notch-dependent differentiation of adult airway basal stem cells. Cell Stem
Cell 8, 639–648.
Romano, R.A., Ortt, K., Birkaya, B., Smalley, K., and Sinha, S. (2009). An active
role of the DeltaN isoform of p63 in regulating basal keratin genes K5 and K14
and directing epidermal cell fate. PLoS ONE 4, e5623.
Romano, R.A., Smalley, K., Magraw, C., Serna, V.A., Kurita, T., Raghavan, S.,
and Sinha, S. (2012). DNp63 knockout mice reveal its indispensable role as a
master regulator of epithelial development and differentiation. Development
139, 772–782.
Schlegelmilch, K., Mohseni, M., Kirak, O., Pruszak, J., Rodriguez, J.R., Zhou,
D., Kreger, B.T., Vasioukhin, V., Avruch, J., Brummelkamp, T.R., and
Camargo, F.D. (2011). Yap1 acts downstream of a-catenin to control
epidermal proliferation. Cell 144, 782–795.
Strano, S., Munarriz, E., Rossi, M., Castagnoli, L., Shaul, Y., Sacchi, A., Oren,
M., Sudol, M., Cesareni, G., and Blandino, G. (2001). Physical interaction with
Yes-associated protein enhances p73 transcriptional activity. J. Biol. Chem.
276, 15164–15173.
Su, X., Paris, M., Gi, Y.J., Tsai, K.Y., Cho, M.S., Lin, Y.L., Biernaskie, J.A.,
Sinha, S., Prives, C., Pevny, L.H., et al. (2009). TAp63 prevents premature
aging by promoting adult stem cell maintenance. Cell Stem Cell 5, 64–75.
Su, X., Chakravarti, D., and Flores, E.R. (2013). p63 steps into the limelight:
crucial roles in the suppression of tumorigenesis and metastasis. Nat. Rev.
Cancer 13, 136–143.
Sudol, M., and Harvey, K.F. (2010). Modularity in the Hippo signaling pathway.
Trends Biochem. Sci. 35, 627–633.
Tata, P.R., Mou, H., Pardo-Saganta, A., Zhao, R., Prabhu, M., Law, B.M.,
Vinarsky, V., Cho, J.L., Breton, S., Sahay, A., et al. (2013a). Dedifferentiation
of committed epithelial cells into stem cells in vivo. Nature 503, 218–223.
Tata, P.R., Pardo-Saganta, A., Prabhu, M., Vinarsky, V., Law, B.M., Fontaine,
B.A., Tager, A.M., and Rajagopal, J. (2013b). Airway-specific inducible trans-
gene expression using aerosolized doxycycline. Am. J. Respir. Cell Mol. Biol.
49, 1048–1056.
Van Keymeulen, A., Rocha, A.S., Ousset, M., Beck, B., Bouvencourt, G., Rock,
J., Sharma, N., Dekoninck, S., and Blanpain, C. (2011). Distinct stem cells
contribute to mammary gland development and maintenance. Nature 479,
189–193.
Yagi, R., Chen, L.F., Shigesada, K., Murakami, Y., and Ito, Y. (1999). A WW
domain-containing yes-associated protein (YAP) is a novel transcriptional
co-activator. EMBO J. 18, 2551–2562.
Yang, A., Kaghad, M., Wang, Y., Gillett, E., Fleming, M.D., Dotsch, V.,
Andrews, N.C., Caput, D., and McKeon, F. (1998). p63, a p53 homolog at
3q27-29, encodes multiple products with transactivating, death-inducing,
and dominant-negative activities. Mol. Cell 2, 305–316.
Yu, F.X., and Guan, K.L. (2013). The Hippo pathway: regulators and regula-
tions. Genes Dev. 27, 355–371.
Yuan, M., Luong, P., Hudson, C., Gudmundsdottir, K., and Basu, S. (2010).
c-Abl phosphorylation of DNp63a is critical for cell viability. Cell Death Dis.
1, e16.
Zhang, H., Pasolli, H.A., and Fuchs, E. (2011). Yes-associated protein (YAP)
transcriptional coactivator functions in balancing growth and differentiation
in skin. Proc. Natl. Acad. Sci. USA 108, 2270–2275.
Zhao, B., Tumaneng, K., and Guan, K.L. (2011). The Hippo pathway in organ
size control, tissue regeneration and stem cell self-renewal. Nat. Cell Biol.
13, 877–883.
Developmental Cell 30, 1–15, July 28, 2014 ª2014 Elsevier Inc. 15

Developmental Cell, Volume 30
Supplemental Information
Yap Tunes Airway Epithelial Size and Architecture
by Regulating the Identity, Maintenance,
and Self-Renewal of Stem Cells
Rui Zhao, Timothy R. Fallon, Srinivas Vinod Saladi, Ana Pardo-Saganta, Jorge Villoria,
Hongmei Mou, Vladimir Vinarsky, Meryem Gonzalez-Celeiro, Naveen Nunna, Lida P.
Hariri, Fernando Camargo, Leif W. Ellisen, and Jayaraj Rajagopal






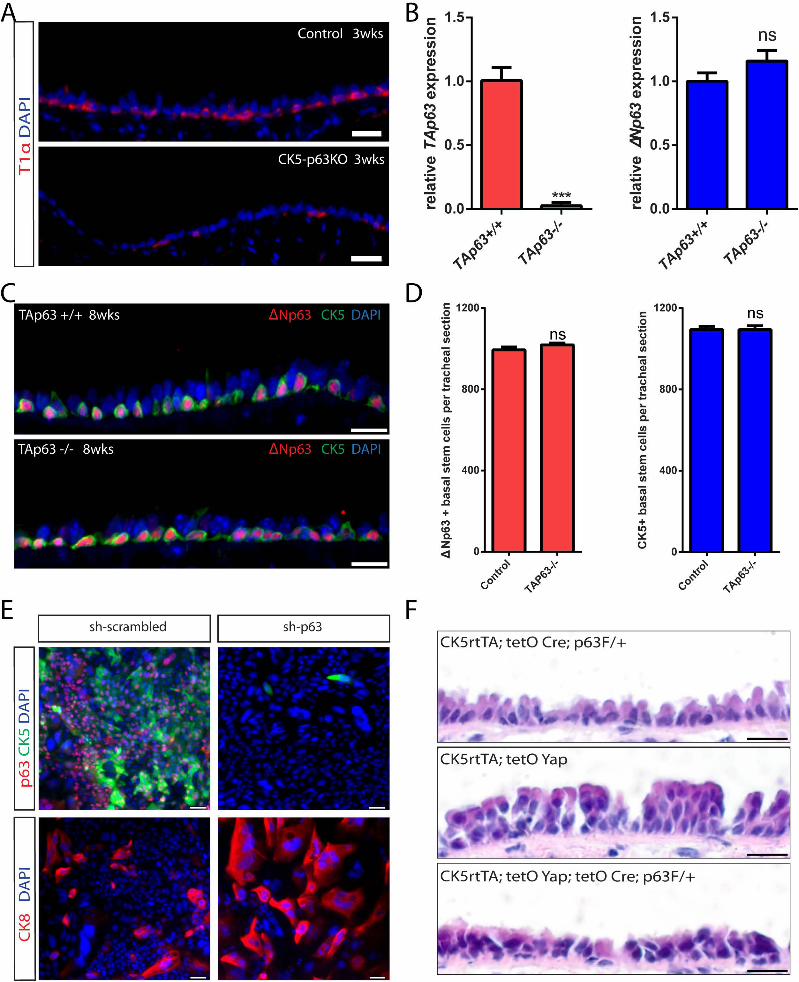

SUPPLEMENTAL INFORMATION
SUPPLEMENTAL FIGURE LEGENDS
Figure S1. Pure Sorts of Airway Epithelial Cell Populations and Stem Cell Loss Following Basal
Stem Cell-Specific Yap Deletion. Related to Figure 1.
(A) Sorting of basal, secretory, and ciliated cells based upon surface expression of GSIβ4,
SSEA1 and CD24 respectively. EpCAM is a marker of all airway epithelial cells. (B)
Quantitative PCR of p63, Scgb1a1 and FoxJ1 transcripts verifies that sorted basal, secretory, and
ciliated cells are pure. Data are represented as mean +/- SEM. (C) Immunostaining for the basal
stem cell-specific marker CK5 (red) and a YFP reporter (green) shows that YFP expression is
induced solely in basal stem cells following doxycycline induction in mice carrying CK5rtTA,
tetO Cre and LSL-YFP alleles. (D) Quantitative PCR using two pairs of primers (Yap-1 and Yap-
2) in control and post-recombination CK5-YapKO sorted basal stem cells one month after
doxycycline treatment. Control mice possess only the CK5 rtTA and tetO Cre alleles. Data are
represented as mean +/- SEM. (E) Immunostaining for T1α (green) and NGFR (red) reveals a
loss of basal stem cells in CK5-YapKO trachea following three months of doxycycline
administration. Scale bars represent 15 μm in C and E.
Figure S2. Basal Stem Cells Undergo Differentiation Upon Yap Loss. Related to Figure 2.
(A) There is no increase in TUNEL (green) or activated Caspase 3 (green) staining in CK5-
YapKO tracheal epithelium 4 weeks after doxycycline administration. (B) Hypothetical schema
showing that, following Yap loss, basal stem cells undergo differentiation transiting through
intermediate cells positive for both basal and differentiated cell markers. (C) Transient
transitional cells double-positive for p63 (red) and Scgb3A2 (green) as well as for CK5 (red) and

FoxJ1 (green) are frequently detected in the tracheal epithelium 3.5 days after sulfur dioxide
(SO2) injury. White arrows point to double-positive cells. (D) Immunostaining for Yap (red)
following tyramide signal amplification (TSA). Yap staining reveals clear signals in both basal
stem cells marked by p63 (green) and suprabasal differentiated cells. Here the Yap antibody
dilution has intentionally been increased to 1:100 to detect even minimal or residual Yap
expression. (E) Immunostaining of GFP (green) and Yap (red) in basal stem cell cultures after
Yap lentiviral transfection demonstrates efficient Yap knockdown (GFP+ cells that are infected
with Yap knockdown virus do not express Yap). (F) Quantification of the percentage of Caspase
3+ cells within the GFP+ virally infected cell population reveals no difference in the degree of
apoptosis following Yap knockdown. (G) Quantification of the percentage of GFP+ virally
infected cells that are also positive for Ki-67 reveals that Yap knockdown decreases cell
proliferation. Data are represented as mean +/- SEM. ns represents not significant. (H)
Immunostaining of GFP (green), basal stem cell marker p63 (red) and differentiation marker
CK8 (red) in basal stem cell cultures after Yap lentiviral knockdown demonstrates that stem cells
differentiate following Yap loss. Scale bar represents 20 μm in A, E and H, and 10 μm in C and
D.
Figure S3. Yap Overexpression in Basal Stem Cells. Related to Figure 3.
Unamplified Yap immunostaining (green) identifies Yap-overexpressing basal cells marked by
CK5 (red) 7 days after doxycycline administration to CK5-YapTg mice. Scale bar represents 30
μm.
Figure S4. Normalizing Yap Expression Restores Basal Stem Cell Proliferation and
Differentiation. Related to Figure 4.

(A) Quantification reveals no significant changes in total p63+ basal stem cell numbers between
control and CK5-YapTg animals 20 days after doxycycline withdrawal (CK5-YapTg Dox Off).
Control animals bearing only the CK5 rtTA driver underwent the same treatment as experimental
animals, namely 20 days of doxycycline followed by a 20-day washout period. (B) The
proportion of basal stem cells (marked by CK5) as a fraction of the total epithelial cells in the
CK5-YapTg animals three weeks after doxycycline withdrawal is not statistically different than
that in control animals. (C) The percentage of proliferating CK5+ basal stem cells in CK5-
YapTg animals after doxycycline withdrawal is not statistically different from that in control
animals. Data are represented as mean +/- SEM in A, B and C. ns indicates a lack of
significance. (D) Co-immunostaining of H2BGFP (green) with the secretory cell marker CC10
(red) and the ciliated cell maker acetylated tubulin (A-tub, white) shows that H2BGFP lineage
labeled stem and progenitor cells that previously overexpressed Yap terminally differentiate after
Yap expression is normalized. Red arrows point to the cells double-positive for H2BGFP and
CC10. White arrows point to the cells double-positive for H2BGFP and A-tub. (E) CK5+ basal
stem cells (red), CC10+ secretory cells (green) and A-tub+ ciliated cells (white) occur in a
normal distribution in a pseudostratified epithelium following Yap normalization. Scale bar = 20
μm in D and E.
Figure S5. Yap Overexpression in Secretory Cells Causes Them to Adopt a Basal-like Identity.
Related to Figure 5.
(A) FACS analysis of control CC10-Yap animals treated with corn oil and PBS (left panel) and
experimental CC10-Yap animals treated with tamoxifen followed by doxycycline (right panel).
No GFP+ cells are detected in the corn oil and PBS treated CC10-Yap animals (left panel),
whereas 3.94% of total epithelial cells are GFP+ in experimental animals. X axis represents GFP

expression and Y axis represents PE, which is used to gate out cells exhibiting autofluoresence.
(B) Staining for ectopic Yap using unamplified Yap staining (red) in CC10-YapTg animals
demonstrates that some Yap-overexpressing secretory cells altered their morphology to resemble
pyramidal basal stem cells (arrows). Scale bar = 10 μm in B.
Figure S6. Yap and p63 Interact and Share Common Target Genes. Related to Figure 6.
(A) Quantitative-PCR analysis shows that ΔNp63α is the major p63 transcript in cultured basal
stem cells. (B) Expression of ΔNp63 protein (red) occurs exclusively in T1α+ (green) basal stem
cells. (C) Yap interacts with ΔNp63α in HEK cells. (D) p63 binds to the promoters of its target
genes in basal stem cells. (E) Immunohistochemical analysis reveals that p63 expression (red) is
decreased at day 6 and is totally abolished at day 10 after p63 knockdown in basal cell cultures.
Immunostaining reveals no significant difference in CK5 (green) between control and p63
knockdown at day 6. However, CK5 protein expression is no longer detectable 10 days after p63
knockdown. (F) Decrease in the mRNA expression of p63 target genes following p63
knockdown in basal stem cells cultured for 6 days. (G) No significant decrease in the mRNA
expression of p63 target genes is detected following TAp63 knockdown. (H) Decreases in mRNA
expression of p63 target genes are observed following ΔNp63 knockdown. Data are represented
as mean +/- SEM. ns represents not significant. Scale bar represents 10 μm in B and E.
Figure S7. p63 and Yap Genetically Interact. Related to Figure 7.
(A) There is a loss of basal stem cells marked by T1α (red) in CK5-p63KO tracheal epithelium
after 3 weeks of doxycycline administration. Transgenic animals bearing only CK5 rtTA and tetO
Cre alleles are used as controls. (B) Quantitative-PCR analysis confirms the lack of TAp63 in
TAp63 knockout mice. No discernible change in the expression of ΔNp63 was observed with

TAp63 deletion. (C) Immunostaining for ΔNp63 and CK5 reveals that basal stem cell
maintenance is not affected by the absence of TAp63. (D) Quantification of cells positive for
ΔNp63 and CK5 reveals that the total number of basal stem cells remains unchanged in the
absence of TAp63, suggesting that TAp63 is dispensable for homeostatic basal stem cell
maintenance in the airway. (E) Immunostaining for p63 (red) confirms that p63 is efficiently
knocked down by lentiviral short hairpin RNAs 10 days after viral infection. A representative
picture showing the lack of CK5+ (green) cells (upper right panel) confirms the loss of basal
stem cells after p63 knockdown. The increase in CK8-expressing cells (red) at day 10 is evidence
for basal stem cell differentiation into suprabasal progenitors following p63 knockdown (bottom
panel). (F) H & E staining of tracheal sections reveals that the epithelial stratification caused by
Yap overexpression is suppressed by removing one copy of p63. Data are presented as mean+/-
SEM in B and D. DAPI is in blue. Scale bar represents 20 μm in A and E, and 10 μm in C and F.
EXTENDED EXPERIMENTAL PROCEDURES
Mouse Tracheal Epithelial Cell Dissociation and Sorting of Different Fractions of
Epithelial Cells
Minced mouse tracheal fragments were incubated in a dissociation solution containing Papain
(20U/mL), EDTA (1.1 mM), 2-Mercaptoethanol (0.067 mM), Cysteine-HCl (5.5 mM) and
DNAse I (100 U/mL) for 1 hour and 30 minutes. The reaction was stopped with Ovomucoid
protease inhibitor (Worthington biochemical Corporation, Cat LK003182) on a rocker at 4ºC for
20 minutes. Cells were centrifuged and resuspended in FACS buffer (2.0% FBS in PBS). Cells
were then stained with EpCAM-PECy7 (1:100; 25-5791-80, eBiosciences), GSIβ4-FITC (1:100;
L2895, Sigma), SSEA1-Alexa Fluor® 647 (1:50; 125608, BioLegend), and CD24-PE (1:75;
553262, BD Pharmingen) for 30 minutes on ice. Cell-specific surface markers for individual cell

populations were identified by screening 40 cell surface markers. Cell specificity was confirmed
by immunostaining of sorted cell populations after cytospin. After selecting the most specific and
reliable markers, basal cells were sorted as EpCAM+ GSIβ4+ cells. Ciliated cells were sorted as
EpCAM+ CD24+ cells. Secretory cells were sorted as EpCAM+ SSEA1+ cells. The purity of the
sorted basal (96.71%), secretory (95.74%), and ciliated cell (96.87%) populations was
established using p63, CC10 (Scgb1A1)/Scgb3A2, and FoxJ1 immunostaining respectively.
Immunofluorescence and Microscopy
Tracheas were dissected and fixed in 4% PFA for 2 hours at 4°C followed by three washes in
PBS. For frozen section preparations, tracheal samples were then placed in 30% sucrose
overnight at 4˚C and embedded in OCT. For paraffin sections, fixed tracheal samples were
dehydrated and embedded in paraffin. Frozen sections or deparaffinized sections (6 μm) were
permeabilized with 0.1% Triton X-100 in PBS, blocked in 1% BSA for 30 minutes at room
temperature, incubated with primary antibodies for 2 hours at room temperature or 4ºC
overnight. The sections were then incubated with secondary antibodies diluted in blocking buffer
for 45 min at room temperature and counterstained with 4', 6-diamidino-2-phenylindole (DAPI,
1μg/mL). Cells stained as whole mounts on a tissue culture dish were fixed with 4% PFA,
washed with PBS, and stained as described above.
The primary antibodies used were: anti-Acetylated tubulin (1:500; T6793, Sigma); rabbit
anti-CK5 (1:1000; ab53121, Abcam); mouse anti-CK5 (1:250; M7237, DAKO); anti-CK8 (1:50;
TROMA 1, Developmental Studies Hybridoma Bank); anti-Caspase3 (1:100; 9661, Cell
Signaling); anti-ΔNp63 (1:50; 619001, BioLegend); rabbit anti-ΔNp63 (1:100, kindly provided
by Satrajit Sinha); anti-FoxJ1 (1:100; 14-9965, eBioscience); chicken anti-GFP (1:500; GFP-
1020, Aves Labs); rabbit anti-GFP (1:100; A11122, Invitrogen); mouse anti-Ki-67 (1:500;

550609, BD Pharmingen); rabbit anti-Ki-67 (1:200; ab15580, Abcam); anti-NGFR (1:100;
ab8875, Abcam); goat anti-CC10 (also named Scgb1A1) (1:5000; kindly provided by Barry
Stripp); rabbit anti-Scgb3A2 (1:3000; kindly provided by Shioko Kimura); anti-SSEA1 (1:100;
125602, BioLegend); anti-T1α (1:50; 8.1.1, Developmental Studies Hybridoma Bank); mouse
anti-p63 (1:100; sc-56188, Santa Cruz); rabbit anti-p63 (1:100; SC8343, Santa Cruz), mouse
anti-Yap (1:100; 101199, Santa Cruz); mouse anti-Yap (1:100; H00010413-M01, Abnova), and
rabbit anti-Yap (1:100; kindly provided by Joseph Avruch). All secondary antibodies were Alexa
Fluor® conjugates (488, 594 and 647) and were used at 1:500 dilutions (Life Technologies).
DeadEnd™ Fluorometric TUNEL System (Promega, G3250) was used to detect apoptosis.
Images were obtained using an Olympus IX81 Inverted microscope (Olympus, Center
Valley, PA). Confocal images were obtained using a Nikon A1R-A1 Confocal Microscope
System (Nikon Instruments Inc., NY).
Yap TSA Amplification
Immunohistochemical staining for Yap protein was performed using a TSA Biotin Tyramide
Amplification system (SAT700001EA, Perkin Elmer). Samples were PFA fixed and prepared as
above. Antigen retrieval was performed in citrate buffer (10mM Citric Acid, 0.1% Tween 20, pH
6.0) using a high pressure controlled retrieval machine. Samples were then treated for 15 minutes
with peroxidase blocking solution (S2003, Dako), and then incubated with primary Yap
antibodies diluted in a blocking buffer containing PBS with 0.1% Triton X-100 (PBST) and 1%
BSA at 4ºC overnight. Following primary antibody staining, samples were washed thoroughly
with PBST and then incubated with anti-primary HRP conjugated secondary antibodies diluted at
1:100 in blocking buffer for 3 hours at room temperature. Following secondary antibody
staining, samples were washed with PBST. Active biotinylated tyramide solution (biotinylated

tyramide reagent dissolved in DMSO) was added. The tyramide deposition reaction was halted
using three PBS washes. Samples were then treated with a 1:250 dilution of streptavidin
conjugated to Alexa Fluor® 594 (Invitrogen) in PBST for 30 minutes at room temperature.
Immunoflourescent stains of additional proteins could then be achieved by sequential application
of the standard immunofluorescence protocol. The following Yap antibodies were used for TSA
amplification: mouse anti-Yap (101199, Santa Cruz); mouse anti-Yap (H00010413-M01,
Abnova), and rabbit anti-Yap (kindly provided by Joseph Avruch).
Mouse Tracheal Epithelial Cell Culture
Sorted basal and secretory cells were plated on transwells with polyester membranes (3470,
Corning) in MTEC medium (You et al., 2002). Rho kinase inhibitor was added for the initial two
days following plating.
Plasmid and Lentiviral Infection
The pLKO.1 backbone vector, with either a GFP reporter or a Puromycin selection cassette, was
used for lentivirally mediated gene knockdown. The sequences of the short hairpins for the
scrambled control and Yap were described previously (Zhang et al., 2011). For p63 knockdown,
the following short hairpins were used: shp63-1 tgatcgatgccgtgcgcttta and shp63-2
gaatgaacagacgtccaattt. For TAp63 knockdown, the following short hairpins were used: shTAp63-
1 tatccgcatgcaagactcaga and shTAp63-2 tgtatccgcatgcaagactca. For ΔNp63 knockdown, the
following short hairpins were used: shΔNp63-1 aacaatgcccagactcaattt and shΔNp63-2
aatgcccagactcaatttagt. Cells infected with shRNA virus were selected with 1 µg/mL puromycin
(Sigma) for 5 days before they were subjected to further analysis.
Quantitative PCR analysis

Total RNA was isolated using Trizol (Invitrogen) and cDNA was produced using the Superscript
III kit (Invitrogen). Quantitative Real Time PCR (qPCR) was done following the instructions in
the Brilliant II SYBR® Green QPCR Master Mix kit (600828, Agilent Technologies), and the
samples were analyzed on a StepOnePlus™ Real-Time PCR System (Applied Biosystems).
Sequence information for qPCR primers is provided below:
Gene name Primer sequence
β-actin Forward: TGGAATCCTGTGGCATCCATGAAAC
Reverse: TAAAACGCAGCTCAGTAACAGTCCG
Scgb1A1 Forward: GATCGCCATCACAATCACTG
Reverse: CTCTTGTGGGAGGGTATCCA
ΔNp63 Forward: GGAAAACAATGCCCAGACTC
Reverse: GATGGAGAGAGGGCATCAAA
EGFR Forward: GAACATCACC TGTACAGGCA
Reverse: GGCATCTGCATACTTCCAGA
FGFR Forward: CCTCGATGTCGTTGAACGGTC
Reverse: CAGCATCCATCTCCGTCACA
FoxJ1 Forward: GAGCTGGAACCACTCAAAGG
Reverse: GGTAGCAGGGCAGTTGATGT
GAPDH Forward: AACTTTGGCATTGTGGAAGG
Reverse: ACACATTGGGGGTAGGAACA
Itgα6 Forward: GCTGTTCTTG CCGGGATTCT
Reverse: AGTATGGATCTCAGCCTTGT

Itgβ4 Forward: ACGATTGCCC CTTTAAAGTC
Reverse: GCAACAGGAGGAAGATGAGC
p63α Forward: GCTGCTCATCATGCCTGGA
Reverse: CAGACTTGCCAAATCATCCA
p63β Forward: GCATGGGAGCCAACATTCCTA
Reverse: TGCCAAATCCTGACAATGCTG
p63γ Forward: CTGCTGTACCTACCAGTGAGA
Reverse: TGAAGCAGGCTGAAAGGAGA
TAp63 Forward: TTACAGATCTGCCATGTCGC
Reverse: CCCAGATATGCTGGAAGACC
Yap-1 Forward: TACATAAACCATAAGAACAAGACCACA
Reverse: GCTTCACTGGAGCACTCTGA
Yap-2 Forward: CATCT TCTGGTCAAA GATACTTC
Reverse: CAGAATTCATCAGCGTCTG
Sulfur Dioxide Injury
The standardization of the sulfur dioxide injury model has been previously described (Kim et al.,
2012; Rock et al., 2011). The exposure time was modified to 3 hours and 40 minutes of 500ppm
SO2 in the strains we used so that the suprabasal epithelium was completely eliminated without
damaging the basal cell layer.
Transfection

For transient transfections, 293T cells were transfected with pLPC-ΔNp63α-Flag and pQCXIH-
Yap-Myc (Zhao et al., 2007) (#33091, Addgene) using Fugene 6 (E2691, Promega) according to
manufacturer’s instructions. Forty-eight hours post transfection, cells were harvested and nuclear
extracts were prepared for co-immunoprecipitations.
Human Basal Cell Culture
This study was approved by the Partners Human Research Committee (IRB Protocol No.
2010P001354/MGH). Primary human airway epithelial cells were isolated from the discarded
trachea and bronchi of donor lungs using the papain dissociation method described above and
cultured in LHC-9 medium (12680-013, Life Technologies). These cultured cells were stained at
each passage. All of the cells expressed the pan epithelial marker EpCAM. Over 96% of the cells
were positive for the basal stem cell markers p63 and CK5.
Co-Immunoprecipitation
Nuclear extracts were prepared by suspending human basal cells in buffer A (10mmol/L Tris-
HCl pH 7.5, 1.5 mmol/L MgCl2, 10 mmol/L KCl) for 20 minutes, followed by douncing.
Pelleted nuclei were first suspended in 1 volume of 20 mM KCl nuclear buffer (20 mM Tris-HCl
pH 7.5, 1.5 mM MgCl2, 0.2 mM EDTA, 25% glycerol). One volume of 1.2 mol/L KCl nuclear
buffer was then added dropwise followed by incubation for 30 minutes at 4°C with rotation.
Nuclear extracts were pre-cleared with equilibrated protein G beads. Cleared nuclear lysates
were incubated with antibody and protein G beads for 4 hours at 4°C, and were then washed with
wash buffer (50 mM Tris-HCl pH 7.5, 5 mM MgCl2, 0.2 mM EDTA, 0.1% NP-40, 10%
glycerol) containing different concentrations of KCl (100 mM, 250 mM, 250 mM, and 100 mM).
The following antibodies were used: anti-c-Myc (9E10, DSHB), anti-Flag (M2 clone, Sigma),

mouse anti-YAP (H00010413-M01, Abnova), rabbit anti-Yap (NB110-58358, Novus), mouse
anti-p63 (4A4, Santa Cruz Biotechnology), rabbit anti-p63 (H129, Santa Cruz Biotechnology),
rabbit anti-TAp63 (618902, Biolegend) and beta-tubulin (D-10, Santa Cruz).
Chromatin Immunoprecipitation
Human basal cells were subjected to additional cross-linking with 2 mM EGS (Ethylene Gycol-
bis (Succinimidylsuccinate)) in PBS for 30 minutes at room temperature followed by washing in
PBS and cross-linking with 1% (v/v) formaldehyde at room temperature for 15 minutes. Cross-
linking was stopped by adding 125 mM glycine. Cells were then lysed in RIPA buffer (10 mM
Tris-HCl pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% (w/v) sodium deoxycholate, 0.1% (w/v)
SDS, 1% (v/v) NP40, proteinase inhibitor cocktail and phosphatase inhibitor cocktail) for 2hr at
40C. Nuclear lysates were sonicated for 5 min 8-10 times in a bioruptor (Diagenode). Sonicated
chromatin was precleared with Protein G sepharose beads (GE healthcare) pre-blocked with BSA
and sonicated salmon sperm DNA (15632-011, Invitrogen). Samples were incubated with 2 μg
antibody overnight at 40C and with beads for an additional 2 hrs. Rabbit anti-Yap (NB110-
58358, Novus) and rabbit anti-p63 (H129, Santa Cruz Biotechnology) were used. Beads were
washed in wash buffer 1 (150 mM NaCl, 20 mM Tris pH 8.1, 2 mM EDTA, 0.1% (w/v) SDS,
1% (v/v) Triton-X-100), wash buffer 2 (500 mM NaCl, 20 mM Tris pH 8.1, 2 mM EDTA,
0.02% (w/v) SDS, 1% (v/v) Triton-X-100), wash buffer 3 (250 mM LiCl ,10 mM Tris pH 8.1, 1
mM EDTA, 1%(w/v) sodium deoxycholate, 1% (v/v) NP40), and 10 mM Tris pH 7.9, 1 mM
EDTA. After the wash, beads were incubated for 3 hours at 550C, then overnight at 650C in 10
mM Tris pH 7.9, 1 mM EDTA, 0.5% (w/v) SDS, 10μg RNase A, and 10 μg Proteinase K. DNA
was purified via QIAquick PCR purification kit (28106, Qiagen) as per manufacturer’s
instructions. PCR was performed using IQ SYBR Green Supermix reagent (Bio-Rad) in a

MX300P machine (Stratagene). Percentage of input from the IPs was calculated by using 10% as
a standard. ChIP primers for FGFR2, EGFR were previously described (McDade et al., 2012).
ChIP-PCR primers are listed below:
ACTB Forward: CCAGGAATTCCGACTTTCAA
Reverse: CAGGACTGGTGAAGTGCTCA
CK5 Forward: AGCCTTGGCCCCCATCCCTTAGATT
Reverse: GGTGTGCCATGAGCAAAACACCTTTCA
ΔNp63 Forward: GCTTGACTACCGTGGAGAGC
Reverse: TTTTCCCAAACTCCAACCTG
EGFR Forward: GGTTAGGGCAGCTCCTCTTT
Reverse: TGGAATTAAGCCTCTCTGCAA
FGFR2 Forward: AAATGAGCGCGCAAGTTAGA
Reverse: CGAACTGGACCGACTTTTTC
Itgα6 Forward: TTATAGGTCCACAAGCGGTGAATGC
Reverse: TCCCCATGACTAAAGCAAGTCCACACC
Itgβ4 Forward: GAGCCGCAGCCCTTTCCG
Reverse: ACGAGGCGGGCAGCGCTTTAT
SUPPLEMENTAL REFERENCES
Kim, J.K., Vinarsky, V., Wain, J., Zhao, R., Jung, K., Choi, J., Lam, A., Pardo-Saganta, A.,
Breton, S., Rajagopal, J., et al. (2012). In vivo imaging of tracheal epithelial cells in mice during
airway regeneration. Am J Respir Cell Mol Biol 47, 864-868.

You, Y., Richer, E.J., Huang, T., and Brody, S.L. (2002). Growth and differentiation of mouse
tracheal epithelial cells: selection of a proliferative population. Am J Physiol Lung Cell Mol
Physiol 283, L1315-1321.
Related Documents











