This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Wild pollinator communities are negatively affected by invasion of aliengoldenrods in grassland landscapes
Dawid Moron a,*, Magdalena Lenda a, Piotr Skórka b, Hajnalka Szentgyörgyi a, Josef Settele c,Michal Woyciechowski a
a Institute of Environmental Sciences, Jagiellonian University, Gronostajowa 7, 30-387 Kraków, Polandb Institute of Nature Conservation, Polish Academy of Sciences, Mickiewicza 33, 31-120 Kraków, Polandc UFZ, Helmholtz Centre for Environmental Research, Department of Community Ecology, Theodor-Lieser-Str. 4, D-06120 Halle, Germany
a r t i c l e i n f o
Article history:Received 14 June 2008Received in revised form 19 December 2008Accepted 24 December 2008Available online 6 March 2009
Keywords:Wild beesBiodiversityButterfliesHoverfliesPlant–insect relationshipsSolidago spp.
a b s t r a c t
The increasing spread of invasive alien plants has changed biodiversity throughout the world. To dateresearch in this area has focused on how invasive plant species affect pollinator behaviour, but there isa lack of data on the impact that alien plant species have on wild pollinator populations. Since their intro-duction in the 19th century, and rapid spread after the 1950s, alien goldenrods (Solidago canadensis, Sol-idago gigantea) have been among the most successful invasive plant species in Europe. We studied theeffects of goldenrods on wild pollinator communities in SE Poland. The abundance, species richnessand diversity of wild bees, hoverflies and butterflies were compared between wet meadows invadedby goldenrod (10 transects) and non-invaded controls (10 transects). Furthermore, we compared theplant diversity and average cover between the two groups of sites. Invasion of goldenrods had a verystrong negative effect on wild pollinator diversity as well as abundance. Plant diversity and average coverwere also negatively affected by goldenrod invasion. Wild pollinators were grouped according to theirnesting and food specialization, but none were resistant to the invasion, indicating that introduced gold-enrod may affect the entire wild pollinator community. Our study emphasises the urgent need to developspecific protection plans for wild pollinators in habitats threatened by foreign plants and we call for theintroduction of programs to stop the invasion of goldenrod not only in Poland, but also on a continentalscale.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Invasions of non-native plant species have led to changes in bio-diversity and ecosystem functioning (McKinney and Lockwood,1999). Although most invader plants are not successful in colonis-ing new areas, some are very competitive and able to spread rap-idly, affecting interactions between wild pollinators (hereafterpollinators) and native plants (Bjerknes et al., 2007). Although aconsiderable amount of research has examined how alien invasionaffects pollinator behaviour (e.g. flower visitation rates, switches toalien flowers), the data concerning impacts on pollinator popula-tions are scarce (for a review see Bjerknes et al., 2007). Alien plantinvasions can alter pollinator populations in a positive as well as in
a negative manner (Graves and Shapiro, 2003). Alien floweringcrops may increase the geographic ranges of pollinators and theduration of their flight period (e.g. butterflies: Graves and Shapiro,2003) and boost pollinator population sizes by increasing their re-source availability (e.g. bees: Tepedino et al., 2008; butterflies:Karowe, 1990; Bowers et al., 1992; Courant et al., 1994; hoverflies:de Groot et al., 2007). On the other hand, alien plant species mayhave a negative effect on native plant diversity (Hejda and Pyšek,2006; Valtonen et al., 2006; Gerber et al., 2008), and subsequentlydecrease pollinator populations (e.g. butterflies: Valtonen et al.,2006; de Groot et al., 2007; Skórka et al., 2007). Also, the toxicityof plant invaders can reduce pollinator population sizes (butter-flies: Porter, 1994; Graves and Shapiro, 2003).
Pollinators play a key role in the maintenance of diversity ofwild plant communities (Roubik, 1995; Kearns et al., 1998; Wil-liams et al., 2001) and provide pollination services worth billionsof Euro per year on many continents (Williams, 1994; Kremenet al., 2002; Klein et al., 2003; Gallai et al., in press). Three maininsect pollinator groups occur in temperate climates: wild bees(Apoidea), hoverflies (Syrphidae) and butterflies (Lepidoptera)
0006-3207/$ - see front matter � 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocon.2008.12.036
* Corresponding author. Address: Institute of Systematics and Evolution ofAnimals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, Poland. Tel.:+48 431 19 63; fax: +48 422 42 94.
E-mail addresses: [email protected] (D. Moron), [email protected](M. Lenda), [email protected] (P. Skórka), [email protected] (H.Szentgyörgyi), [email protected] (J. Settele), [email protected](M. Woyciechowski).
Biological Conservation 142 (2009) 1322–1332
Contents lists available at ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/locate /biocon

Author's personal copy
(Kevan, 1999). These groups differ markedly in life strategies andecology. Many wild bees are closely associated with plant speciesand need suitable nesting space (Spira, 2001; Biesmeijer et al.,2006). Therefore these include many of the most valuable pollina-tors in natural ecosystems as well as agricultural landscapes (Pottset al., 2006; Inouye, 2007). Many butterfly species are also con-nected with one or a few plant species both as adults and larvae(Thompson and Pelmyr, 1991; Renwick and Chew, 1994). In con-trast, all hoverflies are generalist pollinators feeding on nectarand pollen of flowers from a wide range of plant species (Biesmei-jer et al., 2006; Kaczorowska, 2006).
The exotic goldenrods Solidago spp. is among the most success-ful invasive plant species in the world (Weber, 1998; Weber et al.,2008). It is generally understood that these species were intro-duced into Europe from their original ranges in eastern NorthAmerica during the first half of the 19th century as a desirable hor-ticultural plant (Weber, 1998). Following its introduction into Po-land in the 1960s, a rapid increase in the number of localitieswas observed (Guzikowa and Maycock, 1986). Solidago spp. inSouthern Poland often occurs in dense monospecific stands, eachcovering tens or even hundreds of hectares (personal observation).Goldenrod has a high growth rate, an allelopathic impact on otherplants, vegetative proliferation and light, wind-dispersed seedsthat readily germinate on a wide range of soils (Guzikowa andMaycock, 1986; Butcko and Jensen, 2002). Hence it successfullyoutcompetes native plant species and has the potential to affectmany insects by reducing their floral resources (de Groot et al.,2007; Skórka et al., 2007). Insects which are food specialists areespecially vulnerable to native plant decline as they cannot rapidlyswitch to other plant species (de Groot et al., 2007). Moreover, Sol-idago spp. may change local biotic factors through dead biomassaccumulation and its significant height (up to 2 m) (Chapuis-Lardyet al., 2006; personal observations). This may disturb insects whichrequire special habitat features, such as partially open ground toestablish nests (e.g. the majority of wild bees; Michener, 2000).Goldenrod patches may be a physical barrier for butterflies flyinginto sites with these plants (Wratten et al., 2003; authors’ unpub-lished data). Furthermore, a high density of goldenrod and theirovergrowing of other plant species may physically deter access ofpollinators to the usually smaller species of other plants. On theother hand, goldenrod blooms in late summer when most of thenative European species have already flowered, potentially provid-ing a resource and hence having possible positive effects on pollin-ators emerging in late summer.
The aim of this study was to determine whether highly dense,monospecific stands of alien goldenrod are a potential threat towild pollinator communities. We compared wild bee, hoverflyand butterfly species richness, abundance and diversity betweenwet meadows either invaded or not invaded by goldenrod. Wetested whether wild bee and butterfly species belonging to differ-ent ecological groups (nesting and food specialist vs. generalists)are particularly sensitive to goldenrod invasion. We also comparedoverall and flowering plant diversity (suitable for adult pollina-tors) and cover between the two types of sites. Moreover, the pro-portion of low growing plant species was assessed between theinvaded and non-invaded sites. We excluded Apis mellifera as adomesticated species from these analyses, as its abundance de-pends mostly on local beekeeping practices. However, to controlfor the potential effect of interspecific competition between hon-eybee and wild pollinator groups, which could alter our conclusion(we found more honeybee individuals on invaded sites), we corre-lated wild bee, hoverfly and butterfly abundance with A. melliferaabundance. Specifically, we expected: (1) lower plant (overalland flowering) diversity on invaded wet meadows, (2) a lower pro-portion of low growing plants on invaded sites, (3) lower speciesrichness, abundance and diversity of pollinators on sites with gold-
enrod before flowering (March–July), and higher values duringflowering (August–September), (4) a smaller proportion of foodspecialized bee and butterfly species on meadows with Solidagospp., and (5) a smaller proportion of ground nesting bee specieson sites invaded by goldenrod, and (6) no negative correlation(lack of competition) between A. mellifera and wild pollinatorabundance.
The study was conducted within the framework of the EU Inte-grated Project ALARM, which assesses large scale risks to biodiver-sity (www.alarmproject.net, see also Settele et al., 2005).
2. Materials and methods
2.1. Study area
The study area is located in a vast complex of wet grasslands inthe flat bed of the Vistula River valley near the city of Kraków (50�030 N/19� 560 E; �206 m above sea level). All of the grasslands aresimilar in geological history, microclimate, vegetation, and soiltype (Gradzinski, 1974; Kornas and Medwecka-Kornas, 1974; Lan-ger and Szczepanowicz, 1996). The main plant community at thestudy sites is the Molinietum caeruleae Koch 1926 association.Molinietum meadows at the sites consist of a characteristic com-position of species which results in a floristically rich plant com-munity with many rare species such as Epipactis palustris,Gentiana pneumonanthe, Gladiolus imbricatus, Iris sibirica, Orchislatifolia, Sanguisorba officinalis and Trollius europaeus (Dubiel,1996, 2003). Initially, goldenrod occupied ruderal sites and shrub-by habitats close to Vistula River, but recently (since around1990), it has also invaded wet meadows in the area followingthe abandonment of agricultural fields. At present, the vulnerablegrasslands are threatened by goldenrod which already covers over10% of the area.
Using historical and present vegetation data (Kornas and Med-wecka-Kornas, 1974; Dubiel, 1995, 1996; Kudłek, 2002; Pepkows-ka, 2002) we located wet meadow patches of two types: invadedand non-invaded by goldenrod. Then, we randomly chose 10 sitesin each type, ensuring that all had a similar size (�10 ha) andmanagement and were interspersed geographically (Appendix E).Because the areas of the sites were large enough to prevent thecapture of pollinators from adjacent patches (e.g. Bink, 1992;Gathmann and Tscharntke, 2002), we consider it as a reasonableassumption that pollinators occupying invaded and non-invadedsites are representative for those sites. This assumption is alsosupported to some degree by a study on butterfly movements.This study showed that some butterflies species (e.g. Phengaristele-ius, Phengaris nausithous, Brenthis ino, Melanargia galathea) coversmall distances and avoid crossing the edges of habitat patches(Skórka et al., 2007, authors’ unpublished data). Invaded sites weredominated by goldenrod which covered between 90% and 100% ofthe soil surface (mix of S. canadensis L. and S. gigantea A.). All siteswere 1268 ± 45 m (mean ± SE, range: 1020–1722 m) apart whichwas larger than the expected foraging distances of most speciesbelonging to the investigated groups (e.g. Nowicki et al., 2005;Greenleaf et al., 2007). The proportion of permanent grasslandwithin a 500 m radius from the sites’ boundaries was similar be-tween non-invaded (mean ± SE = 35 ± 4%) and invaded sites(mean ± SE = 38 ± 5%, t = 0.482, df = 18, P = 0.636). Also, the meandistance to nearest watercourse (non-invaded sites: 484 ± 67 m,invaded sites: 680 ± 161 m, t = 0.952, df = 18, P = 0.354), meandistance to nearest forest (non-invaded sites: 628 ± 133 m, in-vaded sites: 503 ± 86 m, t = 0.884, df = 18, P = 0.388) and distanceto human settlements (non-invaded sites: 390 ± 64 m, invadedsites: 296 ± 46 m, t = 1.272, df = 18, P = 0.220) were similar onaverage.
D. Moron et al. / Biological Conservation 142 (2009) 1322–1332 1323

Author's personal copy
2.2. Plant counts
At each site, five circular plots of 1 m in diameter (0.79 m2)were established with a distance of 40 m between the plots. Totalplant species diversity was assessed in each plot through the esti-mation of cover (percentage scale: 0–100%). Surveys were donetwice during the study: first in the pre-blooming period (beginningof May) and second at the beginning of the goldenrod floweringperiod (in the middle of July).
2.3. Pollinator surveys
A 200 m transect was established at each site for pollinator sur-veys (Pollard and Yates, 1993). In total, there were 10 transects forinvaded sites and 10 for non-invaded sites. Each transect waslocated in the middle of a given patch, at least 50 m from the patchboundary, in order to restrict edge effects and the impact of sur-rounding areas.
Bees and hoverflies were swept on each transect from April2007 to the beginning of September 2007. During seven transectwalks on each site, the collectors walked at a slow pace making ex-actly 500 sweeps to standardise sweeping effort at floristically dif-ferent patches (invaded vs. non-invaded). Sweeps encompassed allflowering plants at transects of both habitat patches. Individualswere sorted, pinned and dried, prior to species identification, ex-cept for bumblebees protected by Polish law which were identifiedunder field conditions. Butterflies were also counted on each tran-sect from the beginning of May to the beginning of September onseven occasions. A single collector surveyed one systematic groupthroughout the entire season. Pollinators were surveyed duringclear, warm and calm weather conditions on all sampling days(maximum wind at three in the Beaufort scale, cloud cover up to25%). Surveys commenced at 9 a.m. at the earliest, and were fin-ished before 4 p.m. (in peak summer). In spring the surveys startedsomewhat later and ended earlier. The order in which the transectswere sampled was random. Each transect was visited during differ-ent parts of the day throughout the season both in the invaded andnon-invaded sites.
2.4. Data analysis and statistics
To examine how plant diversity and cover are affected by Soli-dago spp. invasion, we calculated the average number of all plantspecies and flowering plant species and their average percent ofcover per survey per site as a response variable. Then, we used gen-eralized linear models (GENMOD procedure in SAS, 2004), employ-ing a normal distribution. The model included the factor ‘‘solidagopresence” and the repeated measure of ‘‘site”. We recognized flow-ering species as plants suitable as food source for adult pollinators(excluding e.g. grasses and sedges; Rothmaler, 2000). We also di-vided the noted plant species into two groups: low growing (aver-age height 620 cm; Rothmaler, 2000) and tall species (>20 cm;Rothmaler, 2000; for comparison see Valtonen et al., 2006). In or-der to examine which plant group is more vulnerable to goldenrodinvasion, we employed a generalized linear mixed model (GLMM)implemented in SAS (2004), that assigned the binomial error vari-ance with logit link. Sites were included in this analysis as a ran-dom effect.
Comparisons of abundance and species richnesses of bees(excluding Apis mellifera), butterflies and hoverflies between in-vaded and non-invaded sites were performed on all 140 counts(20 sites � 7 occasions). Datasets for bees, butterflies and hover-flies were analyzed using a GENMOD procedure with repeatedobservations (surveys at each site), employing a Poisson distribu-tion. The models included the factors ‘‘solidago presence” (pres-ence/absence of goldenrod), ‘‘period” (before/after the start of the
goldenrod bloom, i.e. goldenrod was in bloom during the last threetransect walks), and their interactions term. Moreover, to controlpotential interspecific competition between A. mellifera and wildpollinator groups, we correlated wild bee, hoverfly and butterflyabundance per transect with honeybee abundance per transectfor invaded and non-invaded sites separately. We used Spearmancorrelation implemented in SAS (2004; NPAR1WAY procedure).
We could not calculate the Shannon index for all the surveysseparately because bees, butterflies or hoverflies were not ob-served during some of the visits. Moreover, the absence of individ-uals from some pollinator groups during surveys was not randomlydistributed (a low number of individuals was recorded mostly forsites with Solidago spp.), especially during the pre-blooming peri-od. During the blooming period of Solidago spp., all groups of poll-inators were noted during most surveys and in every transect. Wethen used the average index per blooming period per site (mean atleast from two surveys) excluding surveys with no species found,and applied non-parametric (NPAR1WAY) procedures in SAS(2004). First, we compared the average Shannon index per bloom-ing period between invaded and non-invaded sites using a Wilco-xon signed-rank test. Next, because there were no differences inthe index between flowering periods for all pollinator groups, wecompared the index between invaded and non-invaded plots forthe overall data (periods combined) using a Mann-Whitney U-test.We also conducted an analysis that included surveys with no spe-cies recorded. This lead to the same results as analysis where sur-veys with no species found were excluded. Therefore, we presentthe more conservative results in which surveys with no species re-corded are excluded from the analyses.
In order to determine which pollinator ecological groups areparticularly vulnerable to goldenrod invasion, we divided the re-corded bee and butterfly species into two food specialization cate-gories: polylectic species (gathering pollen from a variety ofunrelated plant species) and oligolectic species (utilising pollenfrom a particular family or genus of plants; Westrich, 1990; Setteleet al., 2000; Celary and Wisniowski, 2003; Dylewska and Wisniow-ski, 2003; Celary, 2005). We also divided bee species according topreferences for nest building below or above ground (Westrich,1990; Celary and Wisniowski, 2003; Dylewska and Wisniowski,2003; Celary, 2005). A generalized linear mixed model (GLMM)was used to analyze this dataset. The models included the factor‘‘solidago presence” (presence or absence of goldenrod) and thenumber of species belonging to a particular ecological group (poly-lectic or oligolectic and ground or above ground nesters) as a re-sponse variable. Sites were included in the analysis as a randomeffect.
3. Results
A total of 102 plant species were recorded in both meadowtypes (Appendix A). The cover of goldenrod on invaded sites rangedfrom 90% to 100%, but on non-invaded meadows it was neverabove 1%. The species richness of plants was significantly loweron sites covered with goldenrod compared to those without gold-enrod (Fig. 1a; GENMOD, overall species: v2
1 ¼ 106:01, P < 0.0001;flowering plants: v2
1 ¼ 57:26, P < 0.0001). Average plant coverwas higher on wet meadows without the invader compared to in-vaded ones (Fig. 1b; GENMOD, overall species: v2
1 ¼ 71:97,P < 0.0001; flowering plants: v2
1 ¼ 10:77, P < 0.0001). However, acomparison of the proportion of low growing plants showed nosignificant differences between invaded and non-invaded wetmeadows (GLMM; F1, 342 = 0.45 P = 0.5009).
We recorded a total of 78 bee species and 583 individuals alongthe transects (Appendix B). Six species made up nearly 50% of thetotal number of noted individuals, these included Bombus lapidari-
1324 D. Moron et al. / Biological Conservation 142 (2009) 1322–1332
Author's personal copy
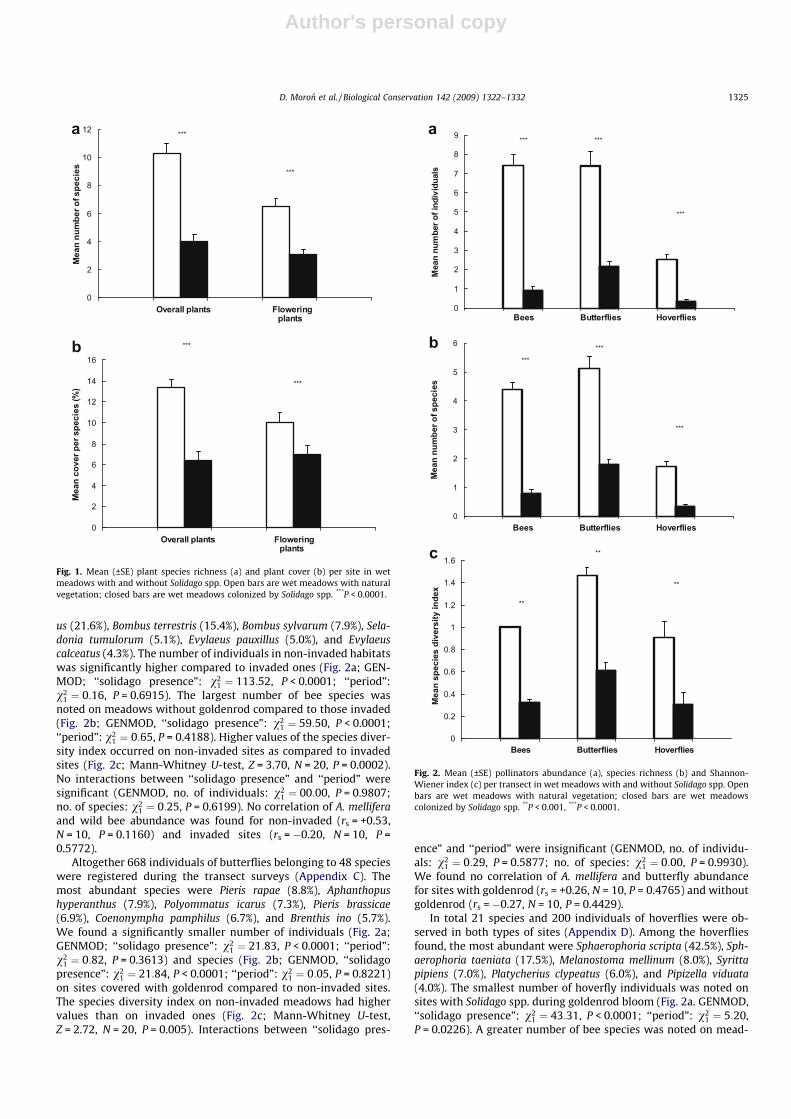
us (21.6%), Bombus terrestris (15.4%), Bombus sylvarum (7.9%), Sela-donia tumulorum (5.1%), Evylaeus pauxillus (5.0%), and Evylaeuscalceatus (4.3%). The number of individuals in non-invaded habitatswas significantly higher compared to invaded ones (Fig. 2a; GEN-MOD; ‘‘solidago presence”: v2
1 ¼ 113:52, P < 0.0001; ‘‘period”:v2
1 ¼ 0:16, P = 0.6915). The largest number of bee species wasnoted on meadows without goldenrod compared to those invaded(Fig. 2b; GENMOD, ‘‘solidago presence”: v2
1 ¼ 59:50, P < 0.0001;‘‘period”: v2
1 ¼ 0:65, P = 0.4188). Higher values of the species diver-sity index occurred on non-invaded sites as compared to invadedsites (Fig. 2c; Mann-Whitney U-test, Z = 3.70, N = 20, P = 0.0002).No interactions between ‘‘solidago presence” and ‘‘period” weresignificant (GENMOD, no. of individuals: v2
1 ¼ 00:00, P = 0.9807;no. of species: v2
1 ¼ 0:25, P = 0.6199). No correlation of A. melliferaand wild bee abundance was found for non-invaded (rs = +0.53,N = 10, P = 0.1160) and invaded sites (rs = �0.20, N = 10, P =0.5772).
Altogether 668 individuals of butterflies belonging to 48 specieswere registered during the transect surveys (Appendix C). Themost abundant species were Pieris rapae (8.8%), Aphanthopushyperanthus (7.9%), Polyommatus icarus (7.3%), Pieris brassicae(6.9%), Coenonympha pamphilus (6.7%), and Brenthis ino (5.7%).We found a significantly smaller number of individuals (Fig. 2a;GENMOD; ‘‘solidago presence”: v2
1 ¼ 21:83, P < 0.0001; ‘‘period”:v2
1 ¼ 0:82, P = 0.3613) and species (Fig. 2b; GENMOD, ‘‘solidagopresence”: v2
1 ¼ 21:84, P < 0.0001; ‘‘period”: v21 ¼ 0:05, P = 0.8221)
on sites covered with goldenrod compared to non-invaded sites.The species diversity index on non-invaded meadows had highervalues than on invaded ones (Fig. 2c; Mann-Whitney U-test,Z = 2.72, N = 20, P = 0.005). Interactions between ‘‘solidago pres-
ence” and ‘‘period” were insignificant (GENMOD, no. of individu-als: v2
1 ¼ 0:29, P = 0.5877; no. of species: v21 ¼ 0:00, P = 0.9930).
We found no correlation of A. mellifera and butterfly abundancefor sites with goldenrod (rs = +0.26, N = 10, P = 0.4765) and withoutgoldenrod (rs = �0.27, N = 10, P = 0.4429).
In total 21 species and 200 individuals of hoverflies were ob-served in both types of sites (Appendix D). Among the hoverfliesfound, the most abundant were Sphaerophoria scripta (42.5%), Sph-aerophoria taeniata (17.5%), Melanostoma mellinum (8.0%), Syrittapipiens (7.0%), Platycherius clypeatus (6.0%), and Pipizella viduata(4.0%). The smallest number of hoverfly individuals was noted onsites with Solidago spp. during goldenrod bloom (Fig. 2a. GENMOD,‘‘solidago presence”: v2
1 ¼ 43:31, P < 0.0001; ‘‘period”: v21 ¼ 5:20,
P = 0.0226). A greater number of bee species was noted on mead-
0
2
4
6
8
10
12
Overall plants Floweringplants
Mea
n nu
mbe
r of s
peci
es
***
***
0
2
4
6
8
10
12
14
16
Overall plants Floweringplants
Mea
n co
ver p
er s
peci
es (%
)
***
***
a
b
Fig. 1. Mean (±SE) plant species richness (a) and plant cover (b) per site in wetmeadows with and without Solidago spp. Open bars are wet meadows with naturalvegetation; closed bars are wet meadows colonized by Solidago spp. ***P < 0.0001.
0
1
2
3
4
5
6
Bees Butterflies Hoverflies
Mea
n nu
mbe
r of s
peci
es
***
***
***
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Bees Butterflies Hoverflies
Mea
n sp
ecie
s di
vers
ity in
dex
**
**
**
b
c
0
1
2
3
4
5
6
7
8
9
Bees Butterflies Hoverflies
Mea
n nu
mbe
r of i
ndiv
idua
ls
*** ***
***
a
Fig. 2. Mean (±SE) pollinators abundance (a), species richness (b) and Shannon-Wiener index (c) per transect in wet meadows with and without Solidago spp. Openbars are wet meadows with natural vegetation; closed bars are wet meadowscolonized by Solidago spp. **P < 0.001, ***P < 0.0001.
D. Moron et al. / Biological Conservation 142 (2009) 1322–1332 1325

Author's personal copy
ows without goldenrod compared to those with the invader(Fig. 2b. GENMOD, ‘‘solidago presence”: v2
1 ¼ 26:94, P < 0.0001;‘‘period”: v2
1 ¼ 1:71, P = 0.1905). Also, the species diversity indexin invaded sites had lower values than non-invaded sites (Fig. 2c;Mann-Whitney U-test, Z = 2.72, N = 20, P = 0.005). Interactions be-tween ‘‘solidago presence” and ‘‘period” were insignificant (GEN-MOD, no. of individuals: v2
1 ¼ 0:48, P = 0.4882; no. of species:v2
1 ¼ 0:22, P = 0.6424). Honeybee abundance was not correlatedwith hoverfly abundance on invaded (rs = �0.04, N = 10,P = 0.9201) and non-invaded sites (rs = �0.17, N = 10, P = 0.6297).
A comparison of the frequency of food specialization of bee andbutterfly species showed no significant differences between in-vaded and non-invaded wet meadows (GLMM; bees: F1, 219 = 1.55P = 0.2147 butterflies: F1, 273 = 0.70 P = 0.4029). The proportion ofground nesting bees was similar in both types of sites. (GLMM;F1, 30.5 = 0.00 P = 0.9899).
4. Discussion
4.1. Solidago spp. negatively affect plants and pollinators
The invasion of goldenrod has a strong negative effect on spe-cies richness of other plants and their average cover (Fig. 1a),which is consistent with previous findings (Meiners et al., 2002;Jakobs et al., 2004; de Groot et al., 2007). The invader replacesnative flowering plants (Fig. 1a) and simplifies trophic relation-ships for many pollinator species. Goldenrod prevents the estab-lishment of native plant species due to allelopathy (Butcko andJensen, 2002) and possibly pollen pollution, shown for other plantspecies (Bartomeus et al., 2008; Jakobsson et al., 2008). The consid-erable height of Solidago spp. in invaded sites (often up to 2 m) sug-gests that goldenrod may outcompete native vegetation also due toshading. However, both low and high growing species have arather low diversity on the investigated meadows regardless ofthe invasion status. Shading may not seriously affect the nativeflora during most of the season in the case of Solidago spp., whichstarts growing in mid summer.
Invaded wet meadows support a significantly lower number ofindividuals and species than non-invaded meadows (Fig. 2a–c).The importance of goldenrod invasion for species groups occupy-ing different trophic levels (including butterflies and hoverflies)was recognized by de Groot et al. (2007). Our results only partlyconfirm their observation on changes in butterfly and hoverflyabundance and diversity caused by goldenrod invasion. Similarto our findings, de Groot et al. (2007) showed that butterflies wereconsistently and negatively affected by Solidago spp. However,hoverflies were only adversely affected in the pre-blooming sam-pling period (July; de Groot et al., 2007), whereas in our investiga-tion they were strongly and negatively affected by the invasionboth in non-flowering and flowering periods. The difference in re-sults between the surveys could be an effect of large patch size(�10 ha) in our study, which should have minimised wild pollina-tor dispersal events from adjacent areas. Our study is the firstshowing that wild bee (the most important pollinator group)abundance and diversity is negatively affected by goldenrod.Although it is believed that invasive alien species may affect polli-nator populations negatively as well as positively (Bowers et al.,1992; Graves and Shapiro, 2003; Nielsen et al., 2008; Tepedinoet al., 2008), we have found a one-sided, strong negative effect ofthe introduction of goldenrod on pollinators. The similarity ofthe response across three taxonomic groups suggests that ingeneral, goldenrod invasion has a negative effect on wild pollina-tor populations.
Interestingly, we noticed a larger number of honeybees on in-vaded sites during the goldenrod blooming season (on average
12.4 per site) than during the pre-blooming season (2.5 individualsper site). This suggests that Solidago spp. may positively affectthese domesticated pollinators as an additional source of pollenand nectar. However, because of beekeeping practices and the abil-ity to fly many kilometres (more than 12 km; Ratnieks, 2000) tosearch for food, it seems that honeybees are not negatively affectedby goldenrod. We expect that species with a high dispersal and/orflying ability should be less prone to the negative effects of golden-rod invasion. On the other hand, our unpublished experimentsshowed that butterflies and bumblebees avoided crossing edgesbetween meadows without goldenrod and meadows invaded bygoldenrod. Also, when the butterflies were experimentally trans-ferred into invaded meadows, they left the meadows and soughtpatches without goldenrod (authors unpublished data).
Competition for floral resources between wild pollinators and A.mellifera could be an alternative potential explanation for the ob-served pattern of wild pollinator species decrease on invaded sites(Westrich, 1990; Evertz, 1995). However, we showed no negativecorrelation between honeybees and wild pollinators abundanceindicating a lack of competition between the groups on both in-vaded and non-invaded sites. These findings are not surprising, be-cause most examples of competition between wild bees andhoneybees are from regions in which A. mellifera is an introducedspecies (Steffan-Dewenter and Tscharntke, 2000).
4.2. No effect of food and nesting specialization on vulnerability toSolidago spp. invasion
Many species of bees and butterflies are highly selective as poll-inators, choosing a particular taxonomic order or even species ofplant with specialized flower structures as foraging sites (Michen-er, 2000; Settele et al., 2000; but see Waser et al., 1996; Waser andOllerton, 2006). Thus, mutually-dependent pollinator and plantspecies can drive each other to extinction (Spira, 2001; Goulson,2003), contributing to the decline of oligolectic species in manydisturbed habitats (Steffan-Dewenter and Tscharntke, 1999). Theinvasion should, due to the elimination of many flowering plantspecies (Fig. 1a and b), firstly decrease the diversity and abundanceof pollinators closely associated with the declining plant species.Contrary to the expectations, our results do not confirm this. Theproportion of both polylectic and oligolectic species did not differbetween invaded and non-invaded sites. Moreover, hoverflies – agroup including exclusively polylectic species, were also negativelyaffected by the invasion (but see de Groot et al., 2007). The lack ofhigher sensitivity of food specialists to Solidago spp. invasion sug-gests that goldenrod does not provide sufficient food for any of theecological groups of wild pollinators. Another possible explanationis that some of the wild pollinators do not recognize this non-na-tive plant as a potentially-to-be-preferred food plant (Knightet al., 2005). This is surprising because goldenrod produces largeamounts of nectar (up to 385 kg/ha) and pollen (up to 90 kg/ha)during the flowering period (Strzałkowska, 2005). A possible expla-nation is that the flowering period may be too late in the season(August through September in Poland) and/or too short to sustainlarge pollinator populations. On the other hand, goldenrod flowersmay provide food only for the imago of the pollinators, whereasmany butterfly and bee larvae have disparate food specializationhabits than imagoes (Michener, 2000; Dennis et al., 2003, 2006;Dover and Settele, in press). Also hoverfly larvae, known to bephytophages, saprophages, coprophages, predators–saprophages–parasites and predators (Kaczorowska, 2006), do not feed onpollen. These factors may explain why the three differentpollinator groups responded identically to Solidago spp. invasionin this study.
Contrary to butterflies and hoverflies, wild bees need a suitablenesting site. We have accounted for two types of bees: ground
1326 D. Moron et al. / Biological Conservation 142 (2009) 1322–1332

Author's personal copy
(mining) and above ground nesting species. Goldenrod may im-pede nest building in the ground, due to changing co-occurrenceof plant species, significant vegetation height, and accumulationof dead biomass (personal observations). Moreover, the invasiondoes not disturb plants with structures useful for above groundnesting species (e.g. shrubs; personal observations). Both miningand ground nesting species declined equally. The invaded sitesoccasionally had small gaps with goldenrod lying on the ground(broken by strong wind), which probably created a refuge withconditions which may have impeded the decline of mining bees.In general we have found evidence for a very strong and similarvulnerability to Solidago spp. invasion in all investigated ecologicalgroups.
4.3. Implications for conservation
Wet grasslands support many species of insects, includingpollinators (Skórka et al., 2007; Moron et al., 2008), thereforemany researchers stress that they should be protected (e.g. Den-isiuk et al., 1991; Sporek and Rombel-Bryzek, 2005). Moreover,these wet meadows represent mainly Molinietalia associations,belonging to the most endangered habitats in Europe (Denisiuket al., 1991; Schnittler and Günther, 1999). Therefore conserva-tion priorities for both plants and pollinators should be created.This could be achieved by rotational management, often citedfor such situations (Cremene et al., 2005; Ogden and Rejmánek,2005; Russell et al., 2005; Hopwood, 2008). Thus, greater publicawareness of the effects of goldenrod and other invasive speciesis needed.
It must be emphasized that goldenrods are the most wide-spread alien plants in Poland. Due to political changes in the1990s, large areas of agricultural land (arable fields, meadows)have been abandoned (Nowicki et al., 2007). A lack of managementleads to natural succession and unfortunately promotes invasion.Our data indicate that in many areas over 80% of abandoned fieldsand meadows have been invaded by goldenrod (unpublished data).This constitutes a great possible threat for pollinator biodiversity inagricultural landscapes. We believe that special action plansshould be implemented to control and stop the invasion of golden-rod. First, within invaded areas goldenrod should be mown at leasttwice a year, before blooming in the spring (May) and during flow-ering in August (Weber, 2000; Rebele and Lehmann, 2002). Thismay involve long-term management due to the presumably largeseed bank of goldenrods. Second, governmental agencies shouldprohibit planting this species in gardens and fields. In Poland gold-enrods are still often planted as decorative plants and food sourcesfor honeybees; these plants may probably act as sources of seedsfor this alien species.
Acknowledgements
Four anonymous referees greatly improved the quality of thispaper with their comments. We wish to thank Damian Chmurafor helpful comments on the initial manuscript. This study wassupported by the 6th EU Framework Programme within theALARM Integrated Project (GOCE-CT-2003-506675; Settele et al.,2005) and the research Grant of the Ministry of Science and HigherEducation SPUB 3053 and DS/WBINOZ/INOS/756/2007.
Appendix A
List of 102 plant species recorded in wet meadows located nearKraków, SE Poland. Meadows were invaded (S) or non-invaded (C)by goldenrod.
Plant species Site type
Achillea millefolium CAchillea pannonica CAegopodium podagraria C, SAgrimonia eupatoria CAllium rotundum CAlopecurus geniculatus CAlopecurus pratensis C, SAngelica sylvestris CAnthoxanthum odoratum CArrhenatherum elatius C, SArtemisia vulgaris C, SCalamagrostis canescens CCalamagrostis epigejos CCalystegia sepium CCampanula patula C, SCampanula trachelium CCardamine pratensis C, SCarex hirta C, SCarex nigra C, SCentaurea cyanus CCentaurea jacea CCirsium arvense C, SCirsium oleraceum CCirsium rivulare CDactylis glomerata CDaucus carota C, SDeschampsia caespitosa CElymus repens C, SEpilobium parviflorum CEquisetum arvense C, SErigeron annuus C, SEuphorbia esula CFestuca pratensis C, SFilipendula ulmaria CFragaria vesca CGaleopsis tetrahit CGalium boreale CGalium mollugo CGalium uliginosum CGalium verum CGentiana pneumonanthe CGeranium palustre CGeranium pratense CGeum rivale CHolcus lanatus C, SHolcus mollis CHypericum maculatum CHypericum perforatum C, SIris pseudacorus CJuncus compressus CJuncus conglomeratus CJuncus effusus CLathyrus pratensis CLathyrus tuberosus CLeucanthemum praecox CLeucanthemum vulgare C, SLotus corniculatus CLotus uliginosus CLychnis flos-cuculi CLysimachia vulgaris C, SLythrum salicaria CMelilotus officinalis CMolinia caerulea C
D. Moron et al. / Biological Conservation 142 (2009) 1322–1332 1327

Author's personal copy
Appendix B
List of 78 wild bee species surveyed in wet meadows locatednear Kraków, SE Poland. Meadows were invaded (S) or non-in-
vaded (C) by goldenrod. Wild bee classification as food specialist(O – oligolectic species) or non-specialist (P – polylectic species)and ground (G) or above ground (A) nesting species based on liter-ature cited in References column. (–) lack of data or category notmatching a species.
Appendix A (continued)
Plant species Site type
Myosotis arvensis COenothera sp. CPhleum pratense CPhragmites communis CPlantago lanceolata CPlantago major CPoa annua CPoa pratensis C, SPoa trivialis CPolygonum bistorta CPotentilla anserina CPotentilla erecta CPotentilla reptans C, SRanunculus acris C, SRubus caesius C, SRumex acetosa CRumex acetosella CRumex crispus CSanguisorba officinalis C, SSerratula tinctoria C
(continued on next column)
Appendix A (continued)
Plant species Site type
Silene pratensis CSolidago canadensis CSolidago gigantea CSymphytum officinale CTanacetum vulgare C, STaraxacum officinale C, STorilis japonica C, STrifolium hybridum CTrifolium pratense CTrifolium repens CTussilago farfara C, SUrtica dioica CValeriana officinalis CVeronica chamaedrys CVeronica persica CVicia cracca C, SVicia grandiflora C, SVicia sepium C, SVicia tetrasperma C
Wild bee species Number of individuals Site type Nesting preferences Floral relationship References
Andrena chrysosceles 3 C G P Westrich (1990)Andrena coitana 2 S G P Westrich (1990)Andrena denticulata 1 S G O Westrich (1990)Andrena dorsata 7 C, S G P Westrich (1990)Andrena falsifica 2 C G P Westrich (1990)Andrena flavipes 2 C G P Westrich (1990)Andrena fulva 4 C G P Westrich (1990)Andrena fulvago 1 C G O Westrich (1990)Andrena gelriae 2 C, S G O Westrich (1990)Andrena gravida 1 C G P Westrich (1990)Andrena haemorrhoa 1 C G P Westrich (1990)Andrena hattorfiana 3 C G O Westrich (1990)Andrena minutula 4 C G P Westrich (1990)Andrena minutuloides 14 C G P Westrich (1990)Andrena mitis 5 C, S G O Westrich (1990)Andrena nitida 1 C G P Westrich (1990)Andrena pontica 3 C G P Celary and Wisniowski (2003)Andrena praecox 2 C G O Westrich (1990)Andrena rosae 4 C G O Westrich (1990)Andrena subopaca 2 C G P Westrich (1990)Andrena vaga 5 C G O Westrich (1990)Andrena viridescens 1 C G O Westrich (1990)Anthidiellum strigatum 1 C A O Dylewska and Wisniowski (2003)Anthophora aestivalis 1 C G P Westrich (1990)Anthophora bimaculata 1 C G P Westrich (1990)Bombus lapidarius 126 C, S G P Westrich (1990)Bombus muscorum 15 C, S G P Westrich (1990)Bombus pascuorum 5 C G P Westrich (1990)Bombus sylvarum 45 C, S G P Westrich (1990)Bombus terrestris 90 C, S G P Westrich (1990)Ceratina cyanea 2 C A P Westrich (1990)
1328 D. Moron et al. / Biological Conservation 142 (2009) 1322–1332

Author's personal copy
Appendix C
List of 48 butterfly species surveyed in wet meadows locatednear Kraków, SE Poland. Meadows were invaded (S) or non-in-
vaded (C) by goldenrod. Butterfly classification as food specialist(O – oligolectic species) or non-specialist (P – polylectic species)based on literature cited in References column.
Appendix B (continued)
Wild bee species Number of individuals Site type Nesting preferences Floral relationship References
Chelostoma rapunculi 2 S A O Dylewska andWisniowski (2003)
Coelioxys conoidea 1 C – P Westrich (1990)Colletes fodiens 6 C, S G O Westrich (1990)Colletes similis 2 C, S G O Dylewska and
Wisniowski (2003)Dasypoda altercator 8 C G O Celary (2005)Evylaeus brevicornis 1 C G O Westrich (1990)Evylaeus calceatus 25 C, S G P Westrich (1990)Evylaeus fulvicornis 8 C G P Westrich (1990)Evylaeus laticeps 1 S G P Westrich (1990)Evylaeus leucopus 1 C G P Westrich (1990)Evylaeus morio 4 C G P Westrich (1990)Evylaeus pauxillus 29 C G P Westrich (1990)Evylaeus quadinotatulus 2 C G P Westrich (1990)Evylaeus villosulus 9 C, S G P Westrich (1990)Halictus maculatus 1 C G P Westrich (1990)Halictus rubicundus 1 C G P Westrich (1990)Halictus sexcinctus 6 C G P Westrich (1990)Heriades truncorum 2 C A O Westrich (1990)Hoplitis spinulosa 3 C A O Westrich (1990)Hylaeus annularis 1 C A P Westrich (1990)Hylaeus confusus 1 C A P Westrich (1990)Hylaeus cornutus 2 C A P Westrich (1990)Hylaeus gracillicornis 2 C A P Westrich (1990)Hylaeus hyalinatus 2 C A P Westrich (1990)Hylaeus nigritus 2 C A O Westrich (1990)Hylaeus pectoralis 1 S A P Westrich (1990)Hylaeus rinki 1 S A P Westrich (1990)Lasioglossum leucozonium 17 C, S G P Westrich (1990)Lasioglossum sexnotatum 1 C G P Westrich (1990)Lasioglossum zonulum 5 C, S G P Westrich (1990)Macropis europaea 8 C, S G O Celary (2005)Megachile alpicola 2 C A P Westrich (1990)Megachile centuncularis 1 C A P Westrich (1990)Melitta leporina 3 C G O Celary (2005)Melitta nigricans 1 C G O Celary (2005)Melitta tricincta 9 C G O Celary (2005)Nomada fabriciana 2 C – – –Nomada ruficornis 1 C – – –Nomada zonata 1 C – – –Osmia fulviventris 2 C A O Westrich (1990)Osmia rufa 3 C A P Westrich (1990)Seladonia subaurata 13 C, S G P Westrich (1990)Seladonia tumulorum 30 C, S G P Westrich (1990)Sphecodes hyalinatus 1 C – P Westrich (1990)Sphecodes miniatus 1 C – P Westrich (1990)Sphecodes monilicornis 1 C – P Westrich (1990)Sphecodes puncticeps 1 C – P Westrich (1990)
D. Moron et al. / Biological Conservation 142 (2009) 1322–1332 1329

Author's personal copy
Appendix D
List of 21 hoverfly species surveyed in wet meadows locatednear Kraków, SE Poland. Meadows were invaded (S) or non-in-vaded (C) by goldenrod.
Hoverfly species Number of individuals Site type
Chrysotoxum bicinctum 1 CEpisyrphus balteatus 2 C, SEristalinus sepulchralis 3 CEristalis arbustorum 3 CEristalis tenax 4 CHelophilus trivittatus 3 C
Hoverfly species Number of individuals Site type
Melanostoma scalare 6 CMelanostoma mellinum 16 C, SMyathropa florea 2 CParagus albifrons 1 CPipizella viduata 7 CPlatycheirus angustatus 3 CPlatycheirus clypeatus 12 C, SPlatycheirus scambus 2 C, SSpazigaster ambulans 1 CSphaerophoria scripta 84 C, S
Butterfly species Number of individuals Site type Floral relationship References
Anthocharis cardamines 23 C, S O Settele et al. (2000)Aphanthopus hyperanthus 55 C, S P Settele et al. (2000)Araschnia levana 12 C, S O Settele et al. (2000)Argynnis aglaja 1 S O Settele et al. (2000)Boloria dia 2 C O Settele et al. (2000)Boloria selene 6 C O Settele et al. (2000)Brenthis daphne 2 C O Settele et al. (2000)Brenthis ino 38 C O Settele et al. (2000)Callophrys rubi 4 C P Settele et al. (2000)Celastrina argiolus 10 C P Settele et al. (2000)Coenonympha glycerion 11 C O Settele et al. (2000)Coenonympha pamphilus 46 C, S P Settele et al. (2000)Colias hyale 3 C, S O Settele et al. (2000)Cupido argiades 4 C O Settele et al. (2000)Erynnis tages 4 C, S O Settele et al. (2000)Gonepteryx rhamni 28 C, S O Settele et al. (2000)Phengaris nausithous 16 C O Settele et al. (2000)Phengaris teleius 19 C O Settele et al. (2000)Hesperia comma 7 C, S O Settele et al. (2000)Issoria lathonia 1 C O Settele et al. (2000)Leptidea sinapis 9 C, S O Settele et al. (2000)Lycaena helle 20 C O Settele et al. (2000)Lycaena tityrus 9 C, S O Settele et al. (2000)Lycaena dispar 7 C O Settele et al. (2000)Lycaena hippothoe 2 C O Settele et al. (2000)Lycaena phlaeas 9 C, S O Settele et al. (2000)Lycaena virgaureae 1 C O Settele et al. (2000)Maniola jurtina 36 C, S P Settele et al. (2000)Melanargia galathea 22 C, S O Settele et al. (2000)Nymphalis antiopa 2 C, S P Settele et al. (2000)Nymphalis io 14 C, S P Settele et al. (2000)Nymphalis urticae 7 C, S O Settele et al. (2000)Ochlodes sylvanus 7 C, S P Settele et al. (2000)Papilio machaon 4 C, S P Settele et al. (2000)Pararge aegeria 2 C O Settele et al. (2000)Pieris brassicae 43 C, S O Settele et al. (2000)Pieris napi 24 C, S P Settele et al. (2000)Pieris rapae 56 C, S P Settele et al. (2000)Plebeius argus 1 C P Settele et al. (2000)Plebeius argyrognomon 1 C O Settele et al. (2000)Polyommatus icarus 49 C, S O Settele et al. (2000)Polyommatus semiargus 7 C O Settele et al. (2000)Pyrgus malvae 2 C, S O Settele et al. (2000)Thecla betulae 5 C, S O Settele et al. (2000)Thymelicus lineola 19 C, S O Settele et al. (2000)Thymelicus sylvestris 10 C, S O Settele et al. (2000)Vanessa atalanta 4 C, S O Settele et al. (2000)Vanessa cardui 4 C, S P Settele et al. (2000)
1330 D. Moron et al. / Biological Conservation 142 (2009) 1322–1332

Author's personal copy
Appendix D (continued)
Hoverfly species Number of individuals Site type
Sphaerophoria taeniata 33 C, SSyritta pipiens 14 C, SSyrphus ribesii 1 CXanthogramma pedissequum 1 CXylota segnis 1 C
Appendix E
Location of study sites in the Kraków region, SE Poland.
References
Bartomeus, I., Bosch, J., Vilà, M., 2008. High invasive pollen transfer, yet lowdeposition on native stigmas in Carpobrotus-invaded community. Annals ofBotany 102, 417–424.
Biesmeijer, J.C., Roberts, S.P.M., Reemer, M., Ohlemüller, R., Edwards, M., Peeters, T.,Schaffers, A.P., Potts, S.G., Kleukers, R., Thomas, C.D., Settele, J., Kunin, W.E.,2006. Parallel declines in pollinators and insect-pollinated plants in Britain andthe Netherlands. Science 313, 351–354.
Bink, F.A., 1992. Ecologische atlas van de dagvlinders van Nordwest-Europa. Schuyt,Haarlem, The Netherlands.
Bjerknes, A.-L., Totland, Ø., Hegland, S.J., Nielsen, A., 2007. Do alien plant invasionsreally affect pollination success in native plant species? Biological Conservation138, 1–12.
Bowers, M.D., Stamp, N.E., Collinge, S.K., 1992. Early stage of host race expansion bya specialist herbivore, Euphydryas phaeton (Nymphalidae). Ecology 73, 526–536.
Butcko, V.M., Jensen, R.J., 2002. Evidence of tissue-specific allelopathic activity inEuthamia graminifolia and Solidago canadensis (Asteraceae). American MidlandNaturalist 148, 253–262.
Celary, W., Wisniowski, B., 2003. Contribution to the bee fauna (Hymenoptera:Apoidea) of Poland. II. Acta Zoologica Cracoviensia 46, 359–364.
Celary, W., 2005. Melittidae of Poland (Hymenoptera: Apoidea: Anthophila) theirBiodiversity and Biology. ISEZ PAS Press, Kraków, Poland.
Chapuis-Lardy, L., Vanderhoeven, S., Dassonville, N., Koutika, L.-S., Meerts, P., 2006.Effect of the exotic invasive plant Solidago gigantea on soil phosphorus status.Biology and Fertility of Soils 42, 481–489.
Courant, A.V., Holbrook, A.E., Van Der Riejden, E.D., Chew, F.S., 1994. Native pierinebutterfly (Pieridae) adapting to naturalized crucifer? Journal of theLepidopterists’ Society 48, 168–170.
Cremene, C., Groza, G., Rakosy, L., Schileyko, A.A., Baur, A., Erhardt, A., Baur, B., 2005.Alterations of steppe-like grasslands in Eastern Europe: a threat to regionalbiodiversity hotspots. Conservation Biology 19, 1606–1618.
Denisiuk, Z., Kalemba, A., Zajac, T., Ostrowska, A., Gawlinski, S., Sienkiewicz, J.,Rejman-Czajkowska, M., 1991. Interaction Between Agriculture and NatureConservation in Poland. Environmental Research Series, IUCN.
Dennis, R.L.H., Shreeve, T.G., van Dyck, H., 2003. Towards a functional resource-based concept for habitat: a butterfly biology viewpoint. Oikos 102, 417–426.
Dennis, R.L.H., Shreeve, T.G., van Dyck, H., 2006. Habitats and resources: the needfor a resource-based definition to conserve butterflies. Biodiversity andConservation 15, 1943–1966.
Dover, J., Settele, J., in press. The influences of landscape structure on butterflydistribution and movement: a review. Journal of Insect Conservation.doi:10.1007/s10841-008-9135-8.
Dubiel, E., 1995. Kierunki antropogenicznych przemian szaty roslinnej doliny Wislyw Krakowie. Folia Geographica Series Geographica–Physica 26–27,139–148.
Dubiel, E., 1996. Łaki Krakowa. Czesc I. Klasa Molinio-Arrhenatheretea. StudiaOsrodka Dokumentacji Fizjograficznej 24, 145–171.
Dubiel, E., 2003. Łaki Krakowa – ginacy element krajobrazu. Aura 11, 27–29.Dylewska, M., Wisniowski, B., 2003. _Zadłówki (Hymenoptera, Aculeata)
Ojcowskiego Parku Narodowego. Ojcowski Park Narodowy, Ojców, Poland.Evertz, S., 1995. Inter-specific competition amongst honey-bees (Apis mellifera) and
solitary wild bees (Hymenoptera Apoidea). Natur und Landschaft 70, 165–172.Gallai, N., Salles, J.-M., Settele, J., Vaissière, B.E., in press. Economic valuation of the
vulnerability of world agriculture confronted to pollinator decline. EcologicalEconomics. doi:10.1016/j.ecolecon.2008.06.014.
Gathmann, A., Tscharntke, T., 2002. Foraging ranges of solitary bees. Journal ofAnimal Ecology 71, 757–764.
Gerber, E., Krebs, Ch., Murrell, A., Moretti, M., Rocklin, R., Schaffner, U., 2008. Exoticinvasive knotweeds (Fallopia spp.) negatively affect native plants andinvertebrate assemblages in European riparian habitats. BiologicalConservation 141, 646–654.
Goulson, D., 2003. Bumblebees, Behaviour and Ecology. Oxford University Press,New York, USA.
Gradzinski, R., 1974. Budowa geologiczna terytorium Krakowa. Folia GeographicaSeries Geographica–Physica 8, 11–17.
Graves, S.D., Shapiro, A.M., 2003. Exotics as host plants of the California butterflyfauna. Biological Conservation 110, 413–433.
Greenleaf, S.S., Williams, N.M., Winfree, R., Kremen, C., 2007. Bee foraging rangesand their relationship to body size. Oecologia 153, 589–596.
de Groot, M., Kleijn, D., Jogan, N., 2007. Species groups occupying different tropiclevels respond differently to the invasion of semi-natural vegetation by Solidagocanadensis. Biological Conservation 136, 612–617.
Guzikowa, M., Maycock, P.F., 1986. The invasion and expansion of three NorthAmerica species of goldenrod (Solidago canadensis L. sensu lato, S. gigantean Ati.and S. graminifolia (L.) Salisb.) in Poland. Acta Societatis Botanicorum Polonae55, 367–384.
Hejda, M., Pyšek, P., 2006. What is the impact of Impatiens glandulifera on speciesdiversity of invaded riparian vegetation? Biological Conservation 132,143–152.
Hopwood, J.L., 2008. The contribution of roadside grassland restorations to nativebee conservation. Biological Conservation 141, 2632–2640.
Inouye, D.W., 2007. The value of bees. Biological Conservation 140, 198–199.Jakobs, G., Weber, E., Edwards, P.J., 2004. Introduced plants of the invasive Solidago
gigantean (Asteracae) are larger and grow denser than cospecifics in the nativerange. Diversity and Distributions 10, 11–19.
Jakobsson, A., Padrón, B., Traveset, A., 2008. Pollen transfer from invasiveCarpobrotus spp. to natives – a study of pollinator behaviour andreproduction success. Biological Conservation 141, 136–145.
Kaczorowska, E., 2006. Hover flies (Diptera: Syrphidae) of the coastal and marinehabitats of Poland. Part II – ecological characteristics. Polish Journal ofEntomology 75, 471–490.
Karowe, D.N., 1990. Predicting host range evolution: colonization of Coronilla variaby Colias philodice (Lepidoptera: Pieridae). Evolution 44, 1637–1647.
Kearns, C.A., Inouye, D.W., Waser, N.M., 1998. Endangered mutualisms: theconservation of plant–pollinator interactions. Annual Review of Ecology andSystematics 29, 83–112.
Kevan, P.G., 1999. Pollinators as bioindicators of the state of the environment:species, activity and diversity. Agriculture, Ecosystems and Environment 74,373–393.
Klein, A., Steffan-Dewenter, I., Tscharntke, T., 2003. Fruit set of highland coffeeincreases with the diversity of pollinating bees. Proceedings of the Royal Societyof London, Series B 270, 955–961.
Knight, T.M., Steets, J.A., Vamosi, J.C., Mazer, S.J., Burd, M., Campbell, D.R., Dudash,M.R., Johnston, M.O., Mitchell, R.J., Ashman, T.-L., 2005. Pollen limitation ofplant reproduction: pattern and process. Annual Review of Ecology, Evolution,and Systematics 36, 467–497.
Kornas, J., Medwecka-Kornas, A., 1974. Szata roslinna Krakowa. Folia GeographicaSeries Geographica–Physica 8, 153–169.
D. Moron et al. / Biological Conservation 142 (2009) 1322–1332 1331

Author's personal copy
Kremen, C., Williams, N.M., Thorp, R.W., 2002. Crop pollination from native bees atrisk from agricultural intensification. Proceedings of the National Academy ofSciences USA 99, 16812–16816.
Kudłek, J., 2002. Plan ochrony i zagospodarowania dydaktyczno - turystycznegoterenów przyległych do Kampusu 600-lecia Odnowienia UniwersytetuJagiellonskiego. MSc Thesis, Jagiellonian University.
Langer, M., Szczepanowicz, B., 1996 Warunki glebowe typowych zbiorowiskłakowych klasy Molinio-Arrhenatheretea na obszarze Kakowa. Studia OsrodkaDokumentacji Fizjograficznej 24, 173–190.
McKinney, M.L., Lockwood, J.L., 1999. Biotic homogenization: a few winnersreplacing many losers in the next mass extinction. Trends in Ecology andEvolution 14, 450–453.
Meiners, S.J., Pickett, S.T.A., Cadenasso, M.L., 2002. Exotic plant invasions over 40years of old field successions: community patterns and associations. Ecography25, 215–223.
Michener, C.D., 2000. The Bees of the World. The Johns Hopkins University Press,Baltimore, MD.
Moron, D., Szentgyörgyi, H., Wantuch, M., Celary, W., Westphal, C., Settele, J.,Woyciechowski, M., 2008. Diversity of wild bees in wet meadows: implicationsfor conservation. Wetlands 28, 975–983.
Nielsen, Ch., Heimes, Ch., Kollmann, J., 2008. Little evidence for negative effects ofan invasive alien plant on pollinator services. Biological Invasions. doi:10.1007/s10530-007-9210-1.
Nowicki, P., Settele, J., Thomas, J.A., Woyciechowski, M., 2005. A review ofpopulation structure of Maculinea butterflies. In: Settele, J., Kuhn, E., Thomas,J.A. (Eds.), Studies in the Ecology and Conservation of Butterflies in Europe. vol.2. Species Ecology along a European Gradient: Maculinea Butterflies as a Model.Pensoft Publishers, Sofia-Moscow, pp. 144–149.
Nowicki, P., Pepkowska, A., Kudłek, J., Skórka, P., Witek, M., Settele, J.,Woyciechowski, M., 2007. From metapopulation theory to conservationrecommendations: lessons from spatial occurrence and abundance patterns ofMaculinea butterflies. Biological Conservation 140, 119–129.
Ogden, J.A.E., Rejmánek, M., 2005. Recovery of native plant communities after thecontrol of a dominant invasive plant species, Foeniculum vulgare: Implicationsfor management. Biological Conservation 125, 427–439.
Pepkowska, A., 2002. Inwentaryzacja i waloryzacja krajobrazowo – przyrodniczaterenów przyległych do Kampusu 600-lecia Odnowienia UniwersytetuJagiellonskiego. MSc Thesis, Jagiellonian University.
Pollard, E., Yates, T.J., 1993. Monitoring Butterflies for Ecology and Conservation.Chapman & Hall, London.
Porter, A., 1994. Implications of introduced garlic mustard (Alliaria petiolata) in thehabitat of Pieris virginiensis (Pieridae). Journal of the Lepidopterists’ Society 48,171–172.
Potts, S.G., Petanidou, T., Roberts, S., O’Toole, C., Hulbert, A., Willmer, P., 2006. Plant-pollinator biodiversity and pollination services in a complex Mediterraneanlandscape. Biological Conservation 129, 519–529.
Ratnieks, F.L.W., 2000. How far do bees forage. Bee Improvement 6, 10–11.Rebele, F., Lehmann, C., 2002. Restoration of a landfill site in Berlin, Germany by
spontaneous and directed succession. Restoration Ecology 10, 340–347.Renwick, J.A., Chew, F.S., 1994. Oviposition behavior in Lepidoptera. Annual Review
of Entomology 39, 377–400.Rothmaler, W., 2000. Exkursionsflora von Deutschland, t. 3. Gefäßpflanzen:
Atlasband. Spektrum Akademischer Verlag, Heidelberg, Berlin, Germany.Roubik, D.W., 1995. Pollination of Cultivated Plants in the Tropics. Food and
Agricultural Organisation Service Bulletin 118. Food and AgriculturalOrganisation, Rome, Italy.
Russell, K.N., Ikerd, H., Droege, S., 2005. The potential conservation value of unmovedpowerline strips for native bees. Biological Conservation 124, 133–148.
SAS Institute. 2004. SAS User’s Guide. Version 9.1.3. SAS Institute, Cary, NorthCarolina.
Schnittler, M., Günther, K.F., 1999. Central European vascular plants requiringpriority conservation measures – an analysis from national Red Lists anddistribution maps. Biodiversity and Conservation 8, 891–925.
Settele, J., Feldmann, R., Reinhardt, R., 2000. Die Tagfalter Deutschlands. Ulmer,Stuttgart, Germany.
Settele, J., Hammen, V., Hulme, P., Karlson, U., Klotz, S., Kotarac, M., Kunin, W.,Marion, G., O’Connor, M., Petanidou, T., Peterson, K., Potts, S., Pritchard, H.,Pysek, P., Rounsevell, M., Spangenberg, J., Steffan-Dewenter, I., Sykes, M., Vighi,M., Zobel, M., Kühn, I., 2005. ALARM: assessing Large-scale environmental risksfor biodiversity with tested methods: the concept, objectives, structure andmanagement of a large Integrated Project within the 6th frameworkprogramme of the European Commission. Gaia 14, 69–72.
Skórka, P., Settele, J., Woyciechowski, M., 2007. Effects of management cessation ongrassland butterflies in Southern Poland. Agriculture, Ecosystems andEnvironment 121, 319–324.
Spira, T.P., 2001. Plant–pollinator interactions: a threatened mutualism withimplications for the ecology and management of rare plants. Natural AreasJournal 21, 78–88.
Sporek, M., Rombel-Bryzek, A., 2005. Wetland restoration enhances thedevelopment of protected species (Iris siberica L.) – a case study. PolishJournal of Ecology 53, 591–595.
Steffan-Dewenter, I., Tscharntke, T., 1999. Effects of habitat isolation on pollinatorcommunities and seed set. Oecologia 121, 432–440.
Steffan-Dewenter, I., Tscharntke, T., 2000. Resource overlap and possiblecompetition between honey bees and wild bees in Central Europe. Oecologia122, 288–296.
Strzałkowska, M., 2005. Nawłoc – cenna roslina po _zytkowa. Abstracts of XLIINaukowa Konferencja Pszczelarska, Puławy, Poland, pp. 116–117.
Tepedino, V.J., Bradley, B.A., Griswold, T.L., 2008. Might flowers of invasive plantsincrease native bee carrying capacity? Intimations from Capitol Reef NationalPark, Utah. Natural Areas Journal 28, 44–50.
Thompson, J.N., Pelmyr, O., 1991. Evolution of oviposition behavior and hostpreference in Lepidoptera. Annual Review of Entomology 36, 65–89.
Valtonen, A., Jantunen, J., Saarinen, K., 2006. Flora and lepidoptera fauna adverselyaffected by invasive Lupinus polyphyllus along road verges. BiologicalConservation 133, 389–396.
Waser, N.M., Chittka, L., Price, M.V., Williams, N.M., Ollerton, J., 1996. Generalizationin pollination systems, and why it matters. Ecology 77, 1043–1060.
Waser, N.M., Ollerton, J., 2006. Plant–Pollinator Interactions: From Specialization toGeneralization. University of Chicago Press, Chicago, USA.
Weber, E., 1998. The dynamics of plant invasions: a case study of three exoticgoldenrod species (Solidago L.) in Europe. Journal of Biogeography 25, 147–154.
Weber, E., 2000. Biological flora of central Europe: Solidago altissima L. Flora 195,123–134.
Weber, E., Sun, S.-H., Li, B., 2008. Invasive alien plants in China: diversity andecological insights. Biological Invasions. doi:10.1007/s10530-008-9216-3.
Westrich, P., 1990. Die Wildbienen Baden-Württembergs. Ulmer, Stuttgart,Germany.
Williams, I.H., 1994. The dependence of crop pollination within the European Unionon pollination by honey bees. Agricultural Zoology Reviews 6, 229–257.
Williams, N.M., Minckley, R.L., Silveira, F.A., 2001. Variation in native bee faunas andits implications for detecting community changes. Conservation Ecology 5 (1),7.
Wratten, S.D., Bowie, M.H., Hickman, J.M., Evans, A.M., Sedcole, J.R., Tylianakis, J.M.,2003. Field boundaries as barriers to movement of hover flies (Diptera:Syrphidae) in cultivated land. Oecologia 134, 605–611.
1332 D. Moron et al. / Biological Conservation 142 (2009) 1322–1332
Related Documents











