Author's personal copy Why are hatching and emergence success low? Mercury and selenium concentrations in nesting leatherback sea turtles (Dermochelys coriacea) and their young in Florida Justin Perrault a,⇑ , Jeanette Wyneken a , Larry J. Thompson b , Chris Johnson c , Debra L. Miller d,1 a Department of Biological Sciences, Florida Atlantic University, Building 01, Sanson Science, 777 Glades Road, Boca Raton, FL 33431, United States b Nestlé Purina PetCare, 801 Chouteau Ave., St. Louis, MO 63102, United States c Loggerhead Marinelife Center of Juno Beach, 14200 US Highway One, Juno Beach, FL 33408, United States d The University of Georgia, College of Veterinary Medicine, Veterinary Diagnostic and Investigational Laboratory, 43 Brighton Road, Tifton, GA 31793, United States article info Keywords: Dermochelys coriacea Hatching success Emergence success Hatchling Mercury (Hg) Selenium (Se) Marine turtles abstract Leatherback sea turtles (Dermochelys coriacea) have low hatching and emergence success compared to other sea turtle species. Postmortem examinations of hatchlings showed degeneration of heart and skel- etal muscle that was similar to that found in other neonates with selenium deficient mothers. Selenium deficiency can result from elevated concentrations of bodily mercury. Ingested mercury is detoxified by the liver through mercury–selenium compound formation. In animals persistently exposed to mercury, the liver’s ability to detoxify this element may decrease, especially if dietary selenium is insufficient. We measured mercury and selenium concentrations in nesting female leatherbacks and their hatchlings from Florida and compared the levels to hatching and emergence success. Both liver selenium and the liver selenium-to-mercury ratio positively correlated with leatherback hatching and emergence success. This study provides the first evidence for the roles of mercury and selenium in explaining low reproduc- tive success in a globally imperiled species, the leatherback sea turtle. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Mercury (hereafter termed Hg) is found in marine organisms ranging from primary producers to top carnivores and it accumu- lates in the body from both water and food sources (Caurant et al., 1999; Guirlet et al., 2008). While Hg is present naturally in the environment (Campbell et al., 2005), the largest source (75%) of this element is traced to emissions from anthropogenic sources, mainly from the burning of fossil fuels (Pacyna and Pacy- na, 2002). Mercury and Hg compounds (i.e., methylmercury, dimethylmercury, etc.) affect various functional processes includ- ing reproduction, growth, development, vision, and hearing. Mer- cury has no known normal physiological role in the body (EPA, 1985). Miller et al. (2009) conducted postmortem examinations of dead-in-nest leatherback sea turtle (Dermochelys coriacea) hatchlings from Juno Beach, Florida, as well as hatchlings that died in the laboratory shortly after nest emergence and found heart and skeletal muscle pathologies that were similar to those seen in bo- vid neonates whose mothers were selenium (Se) deficient (Orr and Blakely, 1997). They suggested that these changes might reflect Hg toxicity or Se deficiency because, in many animals, ingested Hg is detoxified in the liver (where both Hg and Se are stored) through formation of Hg–Se compounds. Thus, persistently elevated Hg may eventually deplete Se stores (Se deficiency) if dietary levels are not compensatory (Cardellicchio et al., 2002). Selenium defi- ciency may lead to Hg toxicity. Selenium is a trace element and is found naturally in marine food sources that are high in protein (Se is largely associated with proteins), but it has low bioavailabil- ity (Caurant et al., 1996; DRI, 2000). Selenium has known enzy- matic, antioxidant, thyroidal, and immune functions (Rayman, 2000); however, at higher concentrations, Se can produce adverse effects including reduced reproductive fitness, tissue lesions, phys- iological anomalies, and can lead to death (Ohlendorf et al., 1988; Hoffman, 2002). Worldwide, leatherback sea turtles have experienced a substan- tial population decrease (67%) resulting from habitat degrada- tion, poaching, and fisheries bycatch (Pritchard, 1982; Spotila et al., 1996; Sarti-Martinez, 2000). It is difficult for these organisms to rebound from a population decline, as they have delayed 0025-326X/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2011.06.009 ⇑ Corresponding author. Tel.: +1 561 297 0146; fax: +1 561 297 2749. E-mail addresses: [email protected] (J. Perrault), [email protected] (J. Wyneken), [email protected] (L.J. Thompson), [email protected] (C. Johnson), [email protected] (D.L. Miller). 1 Present address: Center for Wildlife Health, Department of Forestry, Wildlife and Fisheries University of Tennessee, Knoxville, TN 37996, United States. Marine Pollution Bulletin 62 (2011) 1671–1682 Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Author's personal copy
Why are hatching and emergence success low? Mercury and seleniumconcentrations in nesting leatherback sea turtles (Dermochelys coriacea)and their young in Florida
Justin Perrault a,⇑, Jeanette Wyneken a, Larry J. Thompson b, Chris Johnson c, Debra L. Miller d,1
a Department of Biological Sciences, Florida Atlantic University, Building 01, Sanson Science, 777 Glades Road, Boca Raton, FL 33431, United Statesb Nestlé Purina PetCare, 801 Chouteau Ave., St. Louis, MO 63102, United Statesc Loggerhead Marinelife Center of Juno Beach, 14200 US Highway One, Juno Beach, FL 33408, United Statesd The University of Georgia, College of Veterinary Medicine, Veterinary Diagnostic and Investigational Laboratory, 43 Brighton Road, Tifton, GA 31793, United States
a r t i c l e i n f o
Keywords:Dermochelys coriaceaHatching successEmergence successHatchlingMercury (Hg)Selenium (Se)Marine turtles
a b s t r a c t
Leatherback sea turtles (Dermochelys coriacea) have low hatching and emergence success compared toother sea turtle species. Postmortem examinations of hatchlings showed degeneration of heart and skel-etal muscle that was similar to that found in other neonates with selenium deficient mothers. Seleniumdeficiency can result from elevated concentrations of bodily mercury. Ingested mercury is detoxified bythe liver through mercury–selenium compound formation. In animals persistently exposed to mercury,the liver’s ability to detoxify this element may decrease, especially if dietary selenium is insufficient.We measured mercury and selenium concentrations in nesting female leatherbacks and their hatchlingsfrom Florida and compared the levels to hatching and emergence success. Both liver selenium and theliver selenium-to-mercury ratio positively correlated with leatherback hatching and emergence success.This study provides the first evidence for the roles of mercury and selenium in explaining low reproduc-tive success in a globally imperiled species, the leatherback sea turtle.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Mercury (hereafter termed Hg) is found in marine organismsranging from primary producers to top carnivores and it accumu-lates in the body from both water and food sources (Caurantet al., 1999; Guirlet et al., 2008). While Hg is present naturally inthe environment (Campbell et al., 2005), the largest source(�75%) of this element is traced to emissions from anthropogenicsources, mainly from the burning of fossil fuels (Pacyna and Pacy-na, 2002). Mercury and Hg compounds (i.e., methylmercury,dimethylmercury, etc.) affect various functional processes includ-ing reproduction, growth, development, vision, and hearing. Mer-cury has no known normal physiological role in the body (EPA,1985). Miller et al. (2009) conducted postmortem examinationsof dead-in-nest leatherback sea turtle (Dermochelys coriacea)hatchlings from Juno Beach, Florida, as well as hatchlings that died
in the laboratory shortly after nest emergence and found heart andskeletal muscle pathologies that were similar to those seen in bo-vid neonates whose mothers were selenium (Se) deficient (Orr andBlakely, 1997). They suggested that these changes might reflect Hgtoxicity or Se deficiency because, in many animals, ingested Hg isdetoxified in the liver (where both Hg and Se are stored) throughformation of Hg–Se compounds. Thus, persistently elevated Hgmay eventually deplete Se stores (Se deficiency) if dietary levelsare not compensatory (Cardellicchio et al., 2002). Selenium defi-ciency may lead to Hg toxicity. Selenium is a trace element andis found naturally in marine food sources that are high in protein(Se is largely associated with proteins), but it has low bioavailabil-ity (Caurant et al., 1996; DRI, 2000). Selenium has known enzy-matic, antioxidant, thyroidal, and immune functions (Rayman,2000); however, at higher concentrations, Se can produce adverseeffects including reduced reproductive fitness, tissue lesions, phys-iological anomalies, and can lead to death (Ohlendorf et al., 1988;Hoffman, 2002).
Worldwide, leatherback sea turtles have experienced a substan-tial population decrease (�67%) resulting from habitat degrada-tion, poaching, and fisheries bycatch (Pritchard, 1982; Spotilaet al., 1996; Sarti-Martinez, 2000). It is difficult for these organismsto rebound from a population decline, as they have delayed
0025-326X/$ - see front matter � 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.marpolbul.2011.06.009
⇑ Corresponding author. Tel.: +1 561 297 0146; fax: +1 561 297 2749.E-mail addresses: [email protected] (J. Perrault), [email protected] (J. Wyneken),
[email protected] (L.J. Thompson), [email protected] (C. Johnson),[email protected] (D.L. Miller).
1 Present address: Center for Wildlife Health, Department of Forestry, Wildlife andFisheries University of Tennessee, Knoxville, TN 37996, United States.
Marine Pollution Bulletin 62 (2011) 1671–1682
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul

Author's personal copy
maturity (Avens et al., 2009) and exhibit low hatching success(25.8–56.0%; Whitmore and Dutton, 1985; Leslie et al., 1996; Bellet al., 2003; Hilterman and Goverse, 2003; Hernández et al.,2007); however, average hatching success at some rookeries is ashigh as 67–73.3% in leatherbacks (Livingstone, 2006; Stewart andJohnson, 2006), which suggests variability within the speciesacross populations. Despite this higher hatching success at somerookeries, the high mortality observed during early life historystages (Zug and Parham, 1996; Davenport, 1997; Bell et al., 2003)limits the potential recovery of this imperiled species.
Although low hatching and emergence success in the leather-back turtle has been explored from several nest-based perspec-tives, the causes remain unidentified. Redfearn (2000) found thatleatherback nest depth did not contribute to mortality in Florida.Bell et al. (2003) documented high early embryonic mortality inan eastern Pacific population and hypothesized that maternalreproductive health, chemical contaminants, or bacterial infectionmay be the cause. Wallace et al. (2004) determined that neitherhypoxia nor high nest temperatures (within the thermal tolerancerange) were correlated with low hatching success at that samerookery. Surprisingly, no hypotheses regarding the roles of toxi-cants (e.g., Hg or persistent organic pollutants [POPs]) on hatchingand emergence success have been tested, despite known and po-tential negative impacts on health and reproductive success ofother vertebrates (loons: Burgess and Meyer, 2008; panthers:USEPA, 2000; marine mammals: Beland et al., 1993).
Leatherback sea turtles make long distance migrations from for-aging grounds to nesting beaches, and then subsequently return toforaging grounds (Eckert et al., 2006; Hays et al., 2006). At thoseforaging grounds, non-essential (e.g., Hg) and essential elements(e.g., Se) can accumulate in the body through food (e.g., pelagicmedusa; Bjorndal, 1997) and water intake (Caurant et al., 1999;Guirlet et al., 2008). Females returning to nesting beaches to lay eggscan transfer these elements to their offspring via egg components(i.e., albumen, yolk), as has previously been shown in leatherbacks(Guirlet et al., 2008) and other reptiles (Roe et al., 2004; Rainwateret al., 2005; Unrine et al., 2006). Elevated concentrations of theseelements could impact leatherback turtle hatching and emergencesuccess. Leatherbacks lay 6–11 nests per season depositing anaverage of 70–80 eggs with a mean interclutch interval of 9–10 days (Miller, 1997; Bell et al., 2003; Stewart and Johnson,2006). In this study, our objectives were to (i) identify or excludeSe deficiency as a factor affecting leatherback turtle nest success,(ii) determine how Hg and Se in hatchling blood, liver, and yolksac are correlated with each other and with maternal blood Hgand Se concentrations, and (iii) characterize Hg and Se concentra-tions in nesting leatherbacks (blood), hatchlings (blood from liveturtles and liver and yolk sac from dead turtles), and egg shellsand shelled albumen globs (SAGs).
2. Materials and methods
2.1. Study period and site description
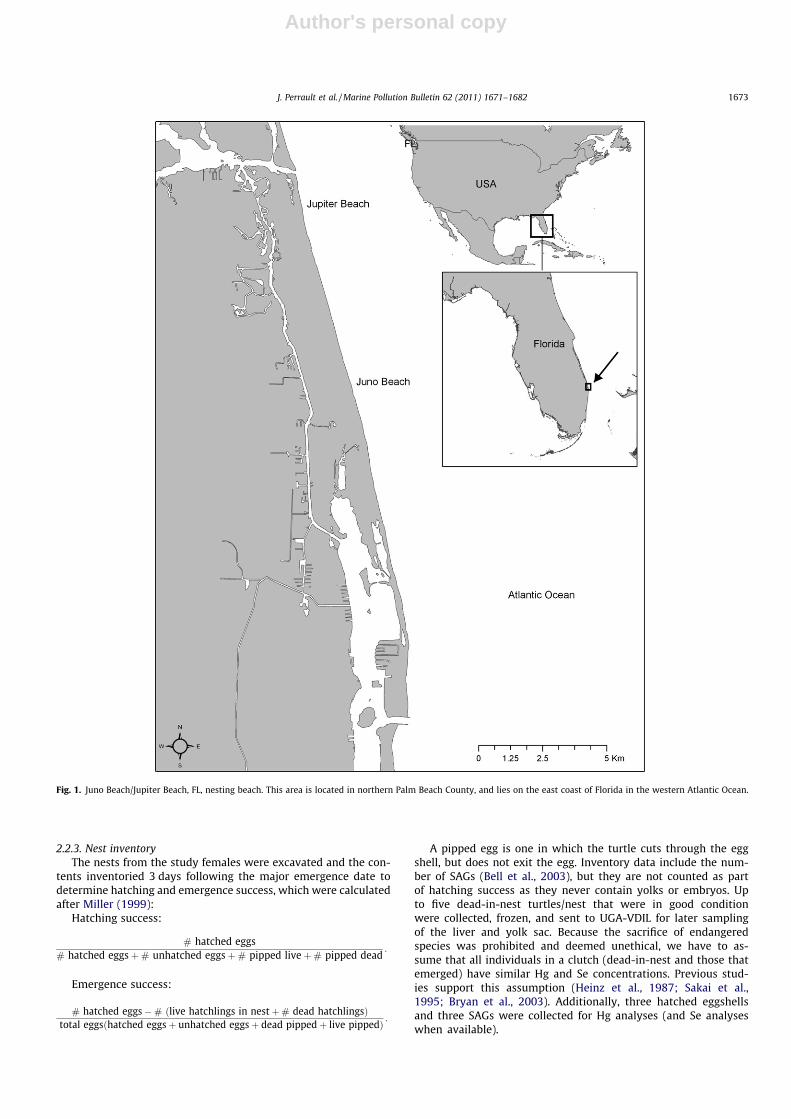
Western Atlantic female leatherbacks and their young weresampled on the nesting beach during the 2007 and 2008 nestingseasons. Samples were collected along Florida’s east coast (JunoBeach and Jupiter Beach, FL), primarily in the 20 km area of peaknesting between Jupiter Inlet (26�5603600N, 80�0401500W) and theLake Worth Inlet (26�4602400N, 80�0105300W, Fig. 1). This rookeryhas been monitored nightly for leatherback nesting activity since2001 (Stewart, 2007). Nesting season runs from March throughearly July in southeastern Florida. Over 400 individual nestingleatherbacks have been identified in Florida since tagging pro-grams began (K. Stewart, personal communication).
2.2. Sample collection
2.2.1. Nesting femalesSampling in Florida was conducted in conjunction with the Log-
gerhead Marinelife Center’s (LMC) leatherback tagging project. Thebeach was patrolled on foot or with all-terrain vehicles each nightto encounter nesting leatherbacks during the nesting season. From10:30 p.m. to 5:30 a.m., night-vision scopes were used to identifyturtle crawls and the stage of nesting before approaching a nestingturtle to minimize the chances of disturbance. Nesting femaleswere approached once they entered a physiological ‘‘trance’’,which begins after oviposition commences (Dutton and Dutton,1994).
Individuals were identified based on their flipper tags and/orinternal PIT (passive integrated transponder) tags. However, if nei-ther of these tags were present, PIT and/or flipper tags were ap-plied by the LMC staff. Blood was collected into 7 mL BD K3EDTAVacutainer� tubes (Becton–Dickinson and Co. Franklin Lakes, NJ,USA) using an 18 gauge venous collection needle fitted in a Vacu-tainer� tube holder. Before insertion of the needle, the entire areawas swabbed with a sterile 70% isopropyl alcohol swab. Approxi-mately 5 mL of blood were taken from the femoral rete system(Dutton, 1996). The location of this vascular network was identi-fied by palpation of the area approximately 10 cm posterior tothe knee. Blood collection ceased when a complete sample was ob-tained or egg-laying terminated (after approximately 7–11 min).The venipuncture site was then disinfected with a new alcoholswab and pressure was applied until the blood site clotted. Thesamples were chilled immediately. After blood collection, the tur-tle’s minimum curved carapace length was recorded (from nuchalnotch, to posterior tip of the caudal peduncle, CCLmin, afterWyneken, 2001). Blood was frozen at �20 �C, and later shippedovernight, on ice, to the Veterinary Diagnostic and InvestigationalLaboratory at the University of Georgia (UGA-VDIL, P.O.Box 1389, 43 Brighton Road, Tifton, GA 31793, USA) for analysis.
2.2.2. HatchlingsWe monitored the progress of all nests and recorded any inci-
dents of predation, sea water inundation, or wash outs due tostorm. On nights when hatchlings were expected to emerge (55–60 days after egg deposition), a 61 cm � 61 cm � 10 cm cage wasplaced on top of the chamber in order to retain the hatchlings sothat blood could be collected. The cages were covered with meshso that predators were excluded if emergence occurred in the ab-sence of the beach patrollers. The cages were monitored every30–60 min.
Upon emergence, up to 10 normal hatchlings were selectedand weighed to the nearest 0.5 g with a PESOLA� 100 g scale.Their straight carapace lengths (SCL), straight carapace widths(SCW), and body depths (BD) were also recorded to the nearest0.05 mm using Vernier calipers. Body condition (mass:SCL ratio)was also calculated. The dorsal and lateral neck of each hatchlingwas swabbed with a sterile 70% isopropyl alcohol pad and bloodwas collected from the external jugular vein (dorsal cervical sinus;Owens and Ruiz, 1980) using a 1 mL BD SafetyGlide™ allergy syr-inge. Approximately 0.1–0.2 mL of blood was collected from eachof the hatchlings and was pooled by clutch into a K3EDTA tube forSe (and Hg if the quantity was sufficient) analysis. This amount(0.1–0.2 mL) is less than 5% blood volume for turtles weighinggreater than 40 g and is considered safe for collection (Jacobson,2007; Strik et al., 2007). Blood was chilled on ice immediatelyafter collection. All samples were frozen at �20 �C until they werelater shipped overnight, on ice, to the UGA-VDIL for analysis.Hatchlings were observed for �1 h and released after bloodcollection.
1672 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682
Author's personal copy
2.2.3. Nest inventoryThe nests from the study females were excavated and the con-
tents inventoried 3 days following the major emergence date todetermine hatching and emergence success, which were calculatedafter Miller (1999):
Hatching success:
# hatched eggs# hatched eggsþ# unhatched eggsþ# pipped liveþ# pipped dead
:
Emergence success:
# hatched eggs�# ðlive hatchlings in nestþ# dead hatchlingsÞtotal eggsðhatched eggsþ unhatched eggs þ dead pipped þ live pippedÞ :
A pipped egg is one in which the turtle cuts through the eggshell, but does not exit the egg. Inventory data include the num-ber of SAGs (Bell et al., 2003), but they are not counted as partof hatching success as they never contain yolks or embryos. Upto five dead-in-nest turtles/nest that were in good conditionwere collected, frozen, and sent to UGA-VDIL for later samplingof the liver and yolk sac. Because the sacrifice of endangeredspecies was prohibited and deemed unethical, we have to as-sume that all individuals in a clutch (dead-in-nest and those thatemerged) have similar Hg and Se concentrations. Previous stud-ies support this assumption (Heinz et al., 1987; Sakai et al.,1995; Bryan et al., 2003). Additionally, three hatched eggshellsand three SAGs were collected for Hg analyses (and Se analyseswhen available).
Fig. 1. Juno Beach/Jupiter Beach, FL, nesting beach. This area is located in northern Palm Beach County, and lies on the east coast of Florida in the western Atlantic Ocean.
J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682 1673

Author's personal copy
2.3. Sample analyses
All sample analyses were standardized by using the same labo-ratory equipment and procedures. Mercury concentrations weredetermined using a flow injection analysis system (FIAS 400, Per-kin-Elmer, Norwalk, CT) by atomic absorption spectrophotometry(AAS, AAnalyst 100, Perkin-Elmer). To quantify total Hg, approxi-mately 1 mL of blood (or 1 g of liver tissue, yolk, albumen, or shell)from each animal was transferred to a Teflon� microwave vesseland mixed with 5 mL of 65% nitric acid (HNO3, Fisher Scientific)and 2 mL of 30% hydrogen peroxide (H2O2, Fisher Scientific). Thesamples then were digested using a laboratory microwave oven(MARS5, CEM Corporation, Matthews, SC, USA) and heated to200 �C under high pressure over 10 min.
The samples were cooled to room temperature (�1 h). Approx-imately 15–20 mL of distilled water was added to the flask so thatthe contents could be easily mixed. Potassium permanganate(KMnO4, 1%) was added until the purple color from the KMnO4 per-sisted for 2–3 s. The sample mixture was diluted to volume withwater (25 mL). The solutions were capped, and inverted 10 timesto mix. The use of HNO3 in AAS can result in artificially high Hgconcentrations; however, the back titration of KMnO4 corrects thiserror. The average of triplicate analyses is reported. Appropriateblanks (deionized water), standards (1, 4, and 5 ppb solutionsmade from Fisher Scientific’s Mercury Reference Standard Solution,1000 ppm ± 1%, Certified), and controls (UTAK Laboratories, Inc.Metals Level 1 Whole Blood Toxicology Control for blood; NIST Bo-vine Liver Standard Reference Material� 1577c for liver, yolk sac,and albumen) were used for each run.
Blood Se was quantified using graphite furnace AAS; 250 lL ofblood was mixed with a prepared diluent (0.2% nitric acid [FisherScientific] and 0.1% Triton X-100 [Acros Organics]). Liver tissue,yolk, and albumen were digested using same procedures as forHg analyses. Appropriate blanks (nitric acid/Triton X-100 diluent),standards (50 and 100 ppb solutions made from Fisher Scientific’sSelenium Reference Standard Solution, 1000 ppm ± 1%, Certified)and controls (UTAK Laboratories, Inc. Metals Level 1 Whole BloodToxicology Control for blood; NIST Bovine Liver Standard ReferenceMaterial� 1577c for liver, yolk sac, albumen, and shell) were em-ployed for each analytical run (Table 1). The average of triplicateanalyses is reported.
2.4. Statistical analyses
Data were tested for normality using the Shapiro–Wilk statistic.Least-squares linear regressions were run with hatching and emer-gence success as the dependent variable and nesting female Hg orSe as the independent variables. A multiple regression analysis wascarried out to determine if Hg and Se were significantly related tohatching and emergence success. Simple regression analysis wascarried out in order to determine if nesting female blood Hg andSe concentrations correlated with body size (CCLmin), clutch size,or hatchling blood, liver, and/or yolk sac Hg and Se concentrations.In order to determine if Hg or Se concentrations were correlated
with hatchling mass, SCL, SCW, BD, or mass:SCL ratio, simpleregressions were performed. Where data were not normally dis-tributed, rank regressions were performed. Nesting female bloodHg and Se were compared to hatchling blood and liver Hg and Seconcentrations using a Mann–Whitney U test. Hatchling blood Hgand liver Hg were compared using a Mann–Whitney U test, in addi-tion to hatchling blood Se and liver Se. Simple linear regressionswere used to determine if a relationship existed between hatchlingblood, liver, and/or yolk sac Hg and Se concentrations and hatchingand/or emergence success. Paired t-tests were carried out to deter-mine if blood Hg or Se concentrations differed significantly in nest-ing females from the first nesting encounter to the second nestingencounter. Similarly, paired t-tests were carried out to comparehatchling blood and liver Hg or Se from subsequent nests fromthe same nesting female. Lastly, SAG albumen Hg concentrationswere compared to yolk sac Hg concentrations using a Mann–Whit-ney U test (Sokal and Rohlf, 1995). Data were analyzed using Systat12 Software (Systat, Inc., Evanston, IL, USA).
3. Results
3.1. Nesting female Hg and Se concentrations and hatching/emergencesuccess
Blood from 60 nesting females was sampled during two nestingseasons, with 39 in 2007, and 21 in 2008, yielding a total of 52 Hgsamples and 71 Se samples. Fewer Hg tests (than Se) were run be-cause an insufficient volume of blood was collected from someindividuals. Eleven females were sampled during more than onenesting event during the nesting season: one turtle was sampledon three occasions for Se, 11 turtles twice for Se, and eight turtleswere sampled twice for Hg. These subsequent nesting events werenot necessarily sequential.
Size, mean CCLmin of the turtles (mean ± SD = 152 ± 8 cm,range = 125–174 cm) did not significantly correlate with eitherblood Hg or Se concentration (p > 0.05). Additionally, the clutchsize of the nest (mean ± SD = 72 ± 14, range = 34–102 eggs, n = 60clutches) did not significantly correlate with either blood Hg orSe concentration (p > 0.05). During excavations in 2007 and 2008,10 nests could not be located (due to washout or inundation)and two were predated. Neither hatching nor emergence successmet the assumptions of normality; therefore the median and rangeare reported (hatching success: median = 59.7%, range 0–93.4%;emergence success: median = 50.8%, range = 0–93.4%).
Maternal blood Hg and Se (Table 2) were not significantly cor-related (p > 0.05). Neither maternal blood Hg nor Se significantlycorrelated with hatching or emergence success (p > 0.05). WhenHg and Se were combined into a multiple regression, no significantcorrelation was observed (p > 0.05).
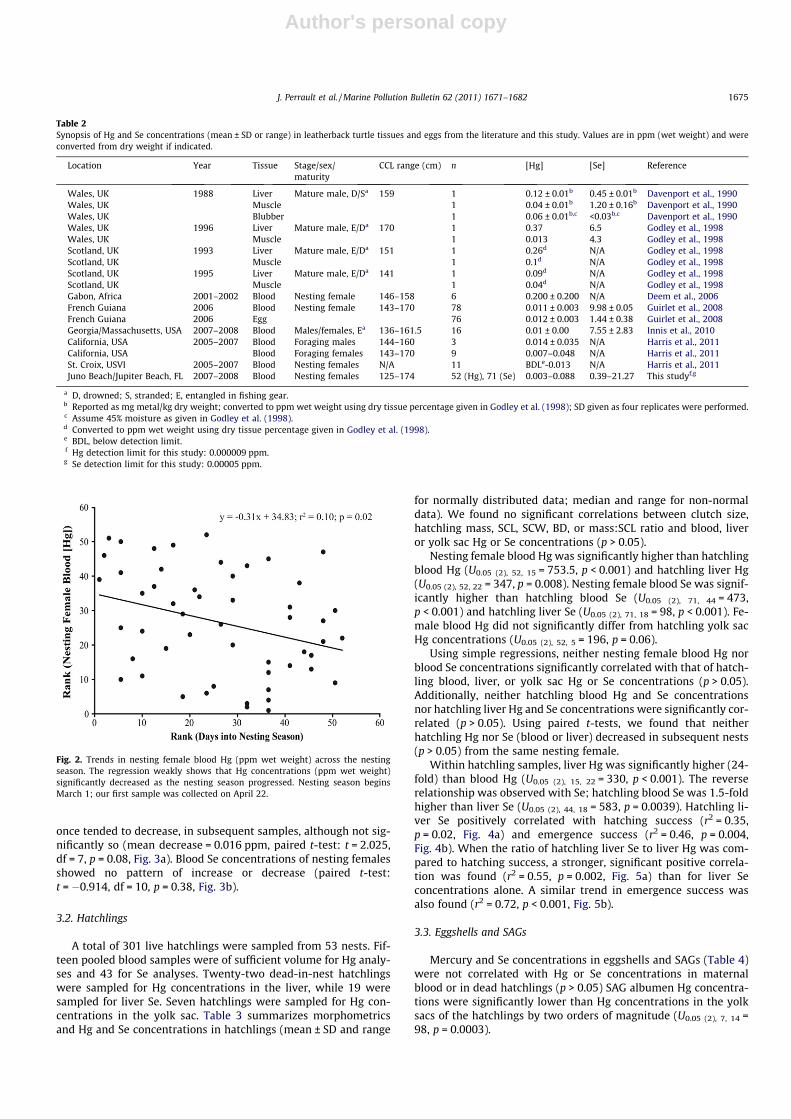
Maternal blood Hg concentrations were negatively correlated(by rank regression) with days into the nesting season (r2 = 0.10,p = 0.02, Fig. 2). Day 1 of the nesting season was set as 1 March.No significant trends in Se concentrations were observed. BloodHg concentrations in each of the eight females sampled more than
Table 1Quality control results (ppm dry weight) attained with reference materials.
Reference material Element Reference value Attained value n
UTAK Metals Level 1 Control Hg 0.0006a 0.0004 ± 0.0002 8Se 0.123b 0.101 ± 0.020 4
NIST Bovine Liver SRM� 1577c Hg 0.00536 ± 0.00017 0.00488 ± 0.00474 9Se 2.031 ± 0.045 2.865 ± 0.153 7
a Expected range: 0.00051–0.00069.b Expected range: 0.105–0.141.
1674 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682
Author's personal copy
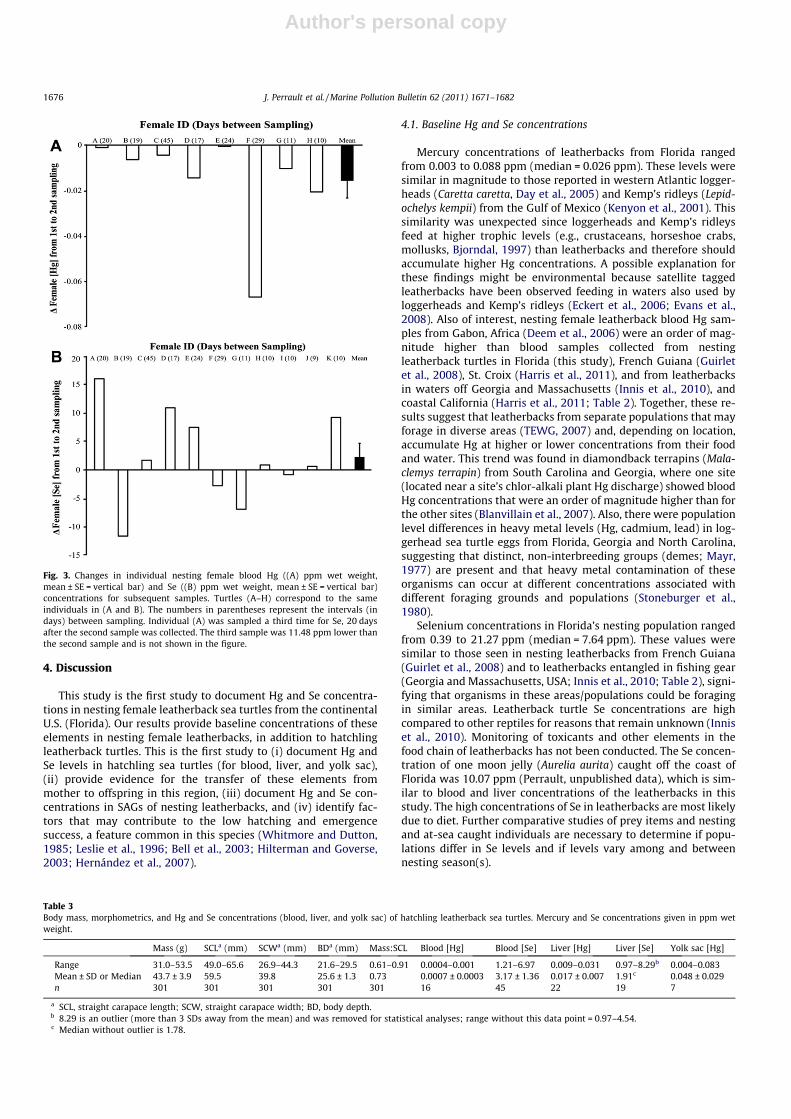
once tended to decrease, in subsequent samples, although not sig-nificantly so (mean decrease = 0.016 ppm, paired t-test: t = 2.025,df = 7, p = 0.08, Fig. 3a). Blood Se concentrations of nesting femalesshowed no pattern of increase or decrease (paired t-test:t = �0.914, df = 10, p = 0.38, Fig. 3b).
3.2. Hatchlings
A total of 301 live hatchlings were sampled from 53 nests. Fif-teen pooled blood samples were of sufficient volume for Hg analy-ses and 43 for Se analyses. Twenty-two dead-in-nest hatchlingswere sampled for Hg concentrations in the liver, while 19 weresampled for liver Se. Seven hatchlings were sampled for Hg con-centrations in the yolk sac. Table 3 summarizes morphometricsand Hg and Se concentrations in hatchlings (mean ± SD and range
for normally distributed data; median and range for non-normaldata). We found no significant correlations between clutch size,hatchling mass, SCL, SCW, BD, or mass:SCL ratio and blood, liveror yolk sac Hg or Se concentrations (p > 0.05).
Nesting female blood Hg was significantly higher than hatchlingblood Hg (U0.05 (2), 52, 15 = 753.5, p < 0.001) and hatchling liver Hg(U0.05 (2), 52, 22 = 347, p = 0.008). Nesting female blood Se was signif-icantly higher than hatchling blood Se (U0.05 (2), 71, 44 = 473,p < 0.001) and hatchling liver Se (U0.05 (2), 71, 18 = 98, p < 0.001). Fe-male blood Hg did not significantly differ from hatchling yolk sacHg concentrations (U0.05 (2), 52, 5 = 196, p = 0.06).
Using simple regressions, neither nesting female blood Hg norblood Se concentrations significantly correlated with that of hatch-ling blood, liver, or yolk sac Hg or Se concentrations (p > 0.05).Additionally, neither hatchling blood Hg and Se concentrationsnor hatchling liver Hg and Se concentrations were significantly cor-related (p > 0.05). Using paired t-tests, we found that neitherhatchling Hg nor Se (blood or liver) decreased in subsequent nests(p > 0.05) from the same nesting female.
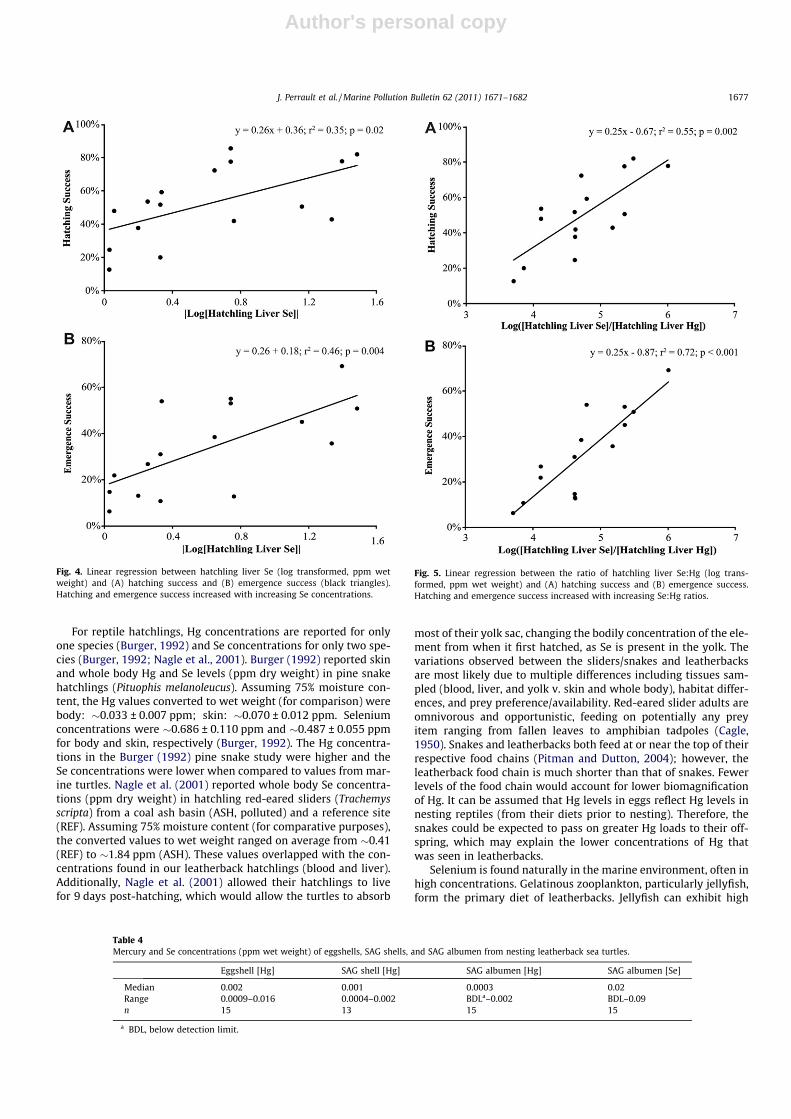
Within hatchling samples, liver Hg was significantly higher (24-fold) than blood Hg (U0.05 (2), 15, 22 = 330, p < 0.001). The reverserelationship was observed with Se; hatchling blood Se was 1.5-foldhigher than liver Se (U0.05 (2), 44, 18 = 583, p = 0.0039). Hatchling li-ver Se positively correlated with hatching success (r2 = 0.35,p = 0.02, Fig. 4a) and emergence success (r2 = 0.46, p = 0.004,Fig. 4b). When the ratio of hatchling liver Se to liver Hg was com-pared to hatching success, a stronger, significant positive correla-tion was found (r2 = 0.55, p = 0.002, Fig. 5a) than for liver Seconcentrations alone. A similar trend in emergence success wasalso found (r2 = 0.72, p < 0.001, Fig. 5b).
3.3. Eggshells and SAGs
Mercury and Se concentrations in eggshells and SAGs (Table 4)were not correlated with Hg or Se concentrations in maternalblood or in dead hatchlings (p > 0.05) SAG albumen Hg concentra-tions were significantly lower than Hg concentrations in the yolksacs of the hatchlings by two orders of magnitude (U0.05 (2), 7, 14 =98, p = 0.0003).
Table 2Synopsis of Hg and Se concentrations (mean ± SD or range) in leatherback turtle tissues and eggs from the literature and this study. Values are in ppm (wet weight) and wereconverted from dry weight if indicated.
Location Year Tissue Stage/sex/maturity
CCL range (cm) n [Hg] [Se] Reference
Wales, UK 1988 Liver Mature male, D/Sa 159 1 0.12 ± 0.01b 0.45 ± 0.01b Davenport et al., 1990Wales, UK Muscle 1 0.04 ± 0.01b 1.20 ± 0.16b Davenport et al., 1990Wales, UK Blubber 1 0.06 ± 0.01b,c <0.03b,c Davenport et al., 1990Wales, UK 1996 Liver Mature male, E/Da 170 1 0.37 6.5 Godley et al., 1998Wales, UK Muscle 1 0.013 4.3 Godley et al., 1998Scotland, UK 1993 Liver Mature male, E/Da 151 1 0.26d N/A Godley et al., 1998Scotland, UK Muscle 1 0.1d N/A Godley et al., 1998Scotland, UK 1995 Liver Mature male, E/Da 141 1 0.09d N/A Godley et al., 1998Scotland, UK Muscle 1 0.04d N/A Godley et al., 1998Gabon, Africa 2001–2002 Blood Nesting female 146–158 6 0.200 ± 0.200 N/A Deem et al., 2006French Guiana 2006 Blood Nesting female 143–170 78 0.011 ± 0.003 9.98 ± 0.05 Guirlet et al., 2008French Guiana 2006 Egg 76 0.012 ± 0.003 1.44 ± 0.38 Guirlet et al., 2008Georgia/Massachusetts, USA 2007–2008 Blood Males/females, Ea 136–161.5 16 0.01 ± 0.00 7.55 ± 2.83 Innis et al., 2010California, USA 2005–2007 Blood Foraging males 144–160 3 0.014 ± 0.035 N/A Harris et al., 2011California, USA Blood Foraging females 143–170 9 0.007–0.048 N/A Harris et al., 2011St. Croix, USVI 2005–2007 Blood Nesting females N/A 11 BDLe-0.013 N/A Harris et al., 2011Juno Beach/Jupiter Beach, FL 2007–2008 Blood Nesting females 125–174 52 (Hg), 71 (Se) 0.003–0.088 0.39–21.27 This studyf,g
a D, drowned; S, stranded; E, entangled in fishing gear.b Reported as mg metal/kg dry weight; converted to ppm wet weight using dry tissue percentage given in Godley et al. (1998); SD given as four replicates were performed.c Assume 45% moisture as given in Godley et al. (1998).d Converted to ppm wet weight using dry tissue percentage given in Godley et al. (1998).e BDL, below detection limit.f Hg detection limit for this study: 0.000009 ppm.g Se detection limit for this study: 0.00005 ppm.
Fig. 2. Trends in nesting female blood Hg (ppm wet weight) across the nestingseason. The regression weakly shows that Hg concentrations (ppm wet weight)significantly decreased as the nesting season progressed. Nesting season beginsMarch 1; our first sample was collected on April 22.
J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682 1675
Author's personal copy
4. Discussion
This study is the first study to document Hg and Se concentra-tions in nesting female leatherback sea turtles from the continentalU.S. (Florida). Our results provide baseline concentrations of theseelements in nesting female leatherbacks, in addition to hatchlingleatherback turtles. This is the first study to (i) document Hg andSe levels in hatchling sea turtles (for blood, liver, and yolk sac),(ii) provide evidence for the transfer of these elements frommother to offspring in this region, (iii) document Hg and Se con-centrations in SAGs of nesting leatherbacks, and (iv) identify fac-tors that may contribute to the low hatching and emergencesuccess, a feature common in this species (Whitmore and Dutton,1985; Leslie et al., 1996; Bell et al., 2003; Hilterman and Goverse,2003; Hernández et al., 2007).
4.1. Baseline Hg and Se concentrations
Mercury concentrations of leatherbacks from Florida rangedfrom 0.003 to 0.088 ppm (median = 0.026 ppm). These levels weresimilar in magnitude to those reported in western Atlantic logger-heads (Caretta caretta, Day et al., 2005) and Kemp’s ridleys (Lepid-ochelys kempii) from the Gulf of Mexico (Kenyon et al., 2001). Thissimilarity was unexpected since loggerheads and Kemp’s ridleysfeed at higher trophic levels (e.g., crustaceans, horseshoe crabs,mollusks, Bjorndal, 1997) than leatherbacks and therefore shouldaccumulate higher Hg concentrations. A possible explanation forthese findings might be environmental because satellite taggedleatherbacks have been observed feeding in waters also used byloggerheads and Kemp’s ridleys (Eckert et al., 2006; Evans et al.,2008). Also of interest, nesting female leatherback blood Hg sam-ples from Gabon, Africa (Deem et al., 2006) were an order of mag-nitude higher than blood samples collected from nestingleatherback turtles in Florida (this study), French Guiana (Guirletet al., 2008), St. Croix (Harris et al., 2011), and from leatherbacksin waters off Georgia and Massachusetts (Innis et al., 2010), andcoastal California (Harris et al., 2011; Table 2). Together, these re-sults suggest that leatherbacks from separate populations that mayforage in diverse areas (TEWG, 2007) and, depending on location,accumulate Hg at higher or lower concentrations from their foodand water. This trend was found in diamondback terrapins (Mala-clemys terrapin) from South Carolina and Georgia, where one site(located near a site’s chlor-alkali plant Hg discharge) showed bloodHg concentrations that were an order of magnitude higher than forthe other sites (Blanvillain et al., 2007). Also, there were populationlevel differences in heavy metal levels (Hg, cadmium, lead) in log-gerhead sea turtle eggs from Florida, Georgia and North Carolina,suggesting that distinct, non-interbreeding groups (demes; Mayr,1977) are present and that heavy metal contamination of theseorganisms can occur at different concentrations associated withdifferent foraging grounds and populations (Stoneburger et al.,1980).
Selenium concentrations in Florida’s nesting population rangedfrom 0.39 to 21.27 ppm (median = 7.64 ppm). These values weresimilar to those seen in nesting leatherbacks from French Guiana(Guirlet et al., 2008) and to leatherbacks entangled in fishing gear(Georgia and Massachusetts, USA; Innis et al., 2010; Table 2), signi-fying that organisms in these areas/populations could be foragingin similar areas. Leatherback turtle Se concentrations are highcompared to other reptiles for reasons that remain unknown (Inniset al., 2010). Monitoring of toxicants and other elements in thefood chain of leatherbacks has not been conducted. The Se concen-tration of one moon jelly (Aurelia aurita) caught off the coast ofFlorida was 10.07 ppm (Perrault, unpublished data), which is sim-ilar to blood and liver concentrations of the leatherbacks in thisstudy. The high concentrations of Se in leatherbacks are most likelydue to diet. Further comparative studies of prey items and nestingand at-sea caught individuals are necessary to determine if popu-lations differ in Se levels and if levels vary among and betweennesting season(s).
Fig. 3. Changes in individual nesting female blood Hg ((A) ppm wet weight,mean ± SE = vertical bar) and Se ((B) ppm wet weight, mean ± SE = vertical bar)concentrations for subsequent samples. Turtles (A–H) correspond to the sameindividuals in (A and B). The numbers in parentheses represent the intervals (indays) between sampling. Individual (A) was sampled a third time for Se, 20 daysafter the second sample was collected. The third sample was 11.48 ppm lower thanthe second sample and is not shown in the figure.
Table 3Body mass, morphometrics, and Hg and Se concentrations (blood, liver, and yolk sac) of hatchling leatherback sea turtles. Mercury and Se concentrations given in ppm wetweight.
Mass (g) SCLa (mm) SCWa (mm) BDa (mm) Mass:SCL Blood [Hg] Blood [Se] Liver [Hg] Liver [Se] Yolk sac [Hg]
Range 31.0–53.5 49.0–65.6 26.9–44.3 21.6–29.5 0.61–0.91 0.0004–0.001 1.21–6.97 0.009–0.031 0.97–8.29b 0.004–0.083Mean ± SD or Median 43.7 ± 3.9 59.5 39.8 25.6 ± 1.3 0.73 0.0007 ± 0.0003 3.17 ± 1.36 0.017 ± 0.007 1.91c 0.048 ± 0.029n 301 301 301 301 301 16 45 22 19 7
a SCL, straight carapace length; SCW, straight carapace width; BD, body depth.b 8.29 is an outlier (more than 3 SDs away from the mean) and was removed for statistical analyses; range without this data point = 0.97–4.54.c Median without outlier is 1.78.
1676 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682
Author's personal copy
For reptile hatchlings, Hg concentrations are reported for onlyone species (Burger, 1992) and Se concentrations for only two spe-cies (Burger, 1992; Nagle et al., 2001). Burger (1992) reported skinand whole body Hg and Se levels (ppm dry weight) in pine snakehatchlings (Pituophis melanoleucus). Assuming 75% moisture con-tent, the Hg values converted to wet weight (for comparison) werebody: �0.033 ± 0.007 ppm; skin: �0.070 ± 0.012 ppm. Seleniumconcentrations were �0.686 ± 0.110 ppm and �0.487 ± 0.055 ppmfor body and skin, respectively (Burger, 1992). The Hg concentra-tions in the Burger (1992) pine snake study were higher and theSe concentrations were lower when compared to values from mar-ine turtles. Nagle et al. (2001) reported whole body Se concentra-tions (ppm dry weight) in hatchling red-eared sliders (Trachemysscripta) from a coal ash basin (ASH, polluted) and a reference site(REF). Assuming 75% moisture content (for comparative purposes),the converted values to wet weight ranged on average from �0.41(REF) to �1.84 ppm (ASH). These values overlapped with the con-centrations found in our leatherback hatchlings (blood and liver).Additionally, Nagle et al. (2001) allowed their hatchlings to livefor 9 days post-hatching, which would allow the turtles to absorb
most of their yolk sac, changing the bodily concentration of the ele-ment from when it first hatched, as Se is present in the yolk. Thevariations observed between the sliders/snakes and leatherbacksare most likely due to multiple differences including tissues sam-pled (blood, liver, and yolk v. skin and whole body), habitat differ-ences, and prey preference/availability. Red-eared slider adults areomnivorous and opportunistic, feeding on potentially any preyitem ranging from fallen leaves to amphibian tadpoles (Cagle,1950). Snakes and leatherbacks both feed at or near the top of theirrespective food chains (Pitman and Dutton, 2004); however, theleatherback food chain is much shorter than that of snakes. Fewerlevels of the food chain would account for lower biomagnificationof Hg. It can be assumed that Hg levels in eggs reflect Hg levels innesting reptiles (from their diets prior to nesting). Therefore, thesnakes could be expected to pass on greater Hg loads to their off-spring, which may explain the lower concentrations of Hg thatwas seen in leatherbacks.
Selenium is found naturally in the marine environment, often inhigh concentrations. Gelatinous zooplankton, particularly jellyfish,form the primary diet of leatherbacks. Jellyfish can exhibit high
Fig. 4. Linear regression between hatchling liver Se (log transformed, ppm wetweight) and (A) hatching success and (B) emergence success (black triangles).Hatching and emergence success increased with increasing Se concentrations.
Fig. 5. Linear regression between the ratio of hatchling liver Se:Hg (log trans-formed, ppm wet weight) and (A) hatching success and (B) emergence success.Hatching and emergence success increased with increasing Se:Hg ratios.
Table 4Mercury and Se concentrations (ppm wet weight) of eggshells, SAG shells, and SAG albumen from nesting leatherback sea turtles.
Eggshell [Hg] SAG shell [Hg] SAG albumen [Hg] SAG albumen [Se]
Median 0.002 0.001 0.0003 0.02Range 0.0009–0.016 0.0004–0.002 BDLa–0.002 BDL–0.09n 15 13 15 15
a BDL, below detection limit.
J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682 1677

Author's personal copy
levels of Se (Perrault, unpublished data). Cnidaria is the only phy-lum that characteristically contains a particular species of Se (SelJselenoprotein homologs) in their tissues (Stillwell and Berry,2005). This finding, along with the large volume of food thatleatherbacks are hypothesized to consume daily (Lutcavage andLutz, 1986; Wallace et al., 2006a), could account for the higher con-centration of Se in leatherback hatchlings than pine snake hatch-lings (through maternal transfer). The reason for high Seconcentrations is unclear; however, it may be that high Se concen-trations in leatherbacks are physiologically necessary to offset thenegative consequences associated with high Hg consumption dueto high prey volume intake. Overall, studies on trace element con-centrations in hatchling sea turtles are lacking and greater atten-tion is needed in this area in order to define baselineconcentrations of these elements.
4.2. Maternal transfer and bodily elimination
Up to 10 months prior to oviposition and when conditions arefavorable, follicular growth occurs in the ovary of sea turtles (Wib-bels et al., 1990; Rostal et al., 1997). In Kemp’s ridleys, vitellogen-esis occurs approximately 4–6 months before the mating period(Rostal et al., 1998); however, in leatherbacks, it is unknown ex-actly how far in advance this occurs (Rostal et al., 1996, 2001).Vitellogenin (VTG) is produced in the liver and is transported tothe oocytes via the plasma (Heck et al., 1997). In the body, Hgand Se are stored in the liver and these elements (along with othersincluding copper, iron, manganese, and zinc) are likely transportedby vitellogenin or other egg proteins (e.g., lipovitellin; Unrine et al.,2006) from maternal liver to the egg (Richards, 1997; Nagle et al.,2001; Roe et al., 2004; Hopkins, 2006). Based on this evidence, ifvitellogenesis occurs while the females are foraging, higher con-centrations of contaminants and/or necessary nutrients may besequestered in the follicles if mothers offload Hg and Se that bioac-cumulate during routine food and water intake (Caurant et al.,1999; Guirlet et al., 2008; Innis et al., 2010). These compoundswould then be passed onto the developing embryos at higher con-centrations than if the females were fasting.
We found that Hg burdens in nesting females tended to pro-gressively decrease with each subsequent nest sampled through-out the season (Figs. 2 and 3a). In contrast, no noticeable trendwas reported for the nesting turtles in the French Guiana popula-tion, even for females laying over six clutches (Guirlet et al.,2008). However, in the same study, they observed a decrease inblood Hg concentrations between the first and the second clutches,which is consistent with our findings. Because the Florida turtlenest over a wide range of available beach sites, just two of oursequential samples represented a single clutch interval. Blood Selevels showed no significant seasonal trends in either Florida orFrench Guiana. Blood Se concentrations, reported here, increasedin 7 of 11 of the nesting turtles sampled more than once, and theaverage showed an increasing trend, although not significantly so(Fig. 3b). Selenium can be stored in blubber of marine organisms(ringed seals, Phoca hispida: Kari and Kauranen, 1978; short-finnedpilot whales, Globicephala macrorhynchus: Stoneburger, 1978;leatherback sea turtles: Davenport et al., 1990), although usuallyin lesser concentrations than those of other tissues. Feeding, drink-ing (Casey et al., 2010), and fat metabolism during the nesting sea-son could mobilize Se, causing a small increase in theconcentrations in the blood (Day et al., 2005).
Deem et al. (2009) observed that blood Hg concentrations innesting loggerheads were greater than foraging loggerheads, indi-cating that nesting females mobilize Hg during the nesting seasonand probably deposit it into the albumen of their eggs. Therefore,hatchlings acquire Hg from both yolk absorption and ingestion ofalbumen. It is likely that, as embryos develop in ovo, Hg and Se
concentrations may change as a result of water and gas exchangebetween the egg and the nest environment (Roe et al., 2004;Guirlet et al., 2008). Therefore, the environment, as well as maternalinput, may influence Hg and Se loads in developing sea turtles.
We did not find significant correlations between maternal bloodHg and Se concentrations and hatchling Hg and Se concentrations.This was unexpected for Se, as Guirlet et al. (2008) found a signif-icant correlation between maternal Se and Se concentration of theegg contents. Theoretically, this lack of correlation may reflectdilution due to uptake of water vapor (and possibly liquid water)from the environment during development (Ackerman et al.,1985; Kam and Ackerman, 1990; Wallace et al., 2006b). However,Nagle et al. (2001) found that there was no difference betweenbodily burdens of trace elements in red-eared slider (Trachemysscripta) hatchlings reared in substrate with elevated concentrationsof trace elements and hatchlings reared in substrate from a refer-ence site. This indicates that parchment-shelled turtle eggs (com-mon to both T. scripta and D. coriacea) and egg membranes mayact as a protective barrier to certain trace elements. If so, maternalinput is the most important source of essential and non-essentialelements to developing turtle embryos. This was verified by Nagleet al. (2001), when they found that hatchlings from mothers thatresided in Se-laden waters had Se concentrations over four timesthat of those with mothers from the reference site.
The lack of correlation between nesting females and hatchlingsdiffers from trends found for other contaminants. For example,persistent organic pollutants (POPs including RDDTs, pp0-DDEand PCB 153 + 105, PCB 180, PCB 138) in blood and eggs were pos-itively correlated nesting leatherbacks from French Guiana (Guirletet al., 2010). Blood sampled from nesting green turtles from Ter-engganu, Malaysia showed concentrations of POPs (RPCBs,RPBDEs, cHCH, and trans-chlordane, mirex) that significantly cor-related with egg contents and blood of hatchling turtles (van deMerwe et al., 2010). The lack of correlation observed in our studymay be explained by the toxicokinetics of Hg and Se in tissueswhen compared to POPs. POP concentrations in the blood at thetime of nesting are thought to represent the contamination ac-crued at foraging grounds prior to nesting (4–10 months prior, Ros-tal et al., 1997; Guirlet et al., 2010), whereas Hg and Se in the bloodmay indicate more recent uptake (days–weeks prior). The lack ofcorrelation between nesting females and hatchlings is consistentwith this hypothesis as the majority of Hg and Se found in hatch-lings comes from the yolk.
Mercury concentrations in the yolk sacs of the hatchlings didnot significantly differ from blood Hg concentrations in the nestingfemales, a finding comparable to that in Guirlet et al. (2008).Maternal blood Hg concentrations were extremely similar(0.001 ppm difference in means) to the concentrations of egg con-tents (yolk and albumen). Because all yolked follicles are formedbefore turtles arrive at the nesting beach (Rostal et al., 1996),leatherbacks may pass on similar concentrations of trace elementsto their offspring in each subsequent clutch (we found no signifi-cant differences in Hg and Se concentrations in hatchlings fromsubsequent clutches). This would ensure that all hatchlings wereequally provisioned with the necessary nutrients in their yolk sacs,and that no single clutch is burdened with a high contaminant load(Sakai et al., 1995), unless high concentrations of these compoundsare deposited into the albumen during the nesting season, which isunlikely. We found significantly lower concentrations (two ordersof magnitude) of Hg in the albumen of the SAGs (we did not test forSe in yolk sac or albumen) when compared to the yolk sacs of thehatchlings, indicating the yolk is the main source of this toxicantfor the hatchlings. Sakai et al. (1995) found little intraclutch andinterclutch variation in Hg concentrations of yolks of loggerheadsea turtle and green sea turtle eggs, respectively. Interclutch Seconcentrations were shown to vary minimally in the eggs of
1678 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682

Author's personal copy
captive bred mallards (Anas platyrhynchos; Heinz et al., 1987) andin eggs from a reference population of common grackles (Quiscalusquiscala; Bryan et al., 2003).
We found significantly higher concentrations of Hg in the liversof hatchlings than in the blood. In other vertebrates, Hg is stored inthe liver where it can be or has been detoxified; assuming hatch-ling leatherbacks process Hg similarly, this may account for thelower concentration in the blood. Interestingly, we found signifi-cantly higher concentrations of Se in the blood than the liver.Magat and Sell (1979) observed that Se binds to the yolk in hens, but,in mallards fed Se as selenomethionine (an amino acid containingSe), a greater proportion of this dietary Se was transferred to theeggs and was bound at greater levels in the albumen. Selenomethi-onine is a natural form of Se most often encountered by animals(Spallholz and Hoffman, 2002). It is not regulated homeostatically.Since embryos of oviparous species consume albumen duringdevelopment (Romanoff, 1967; Palmer and Guillette, 1991), thiscould contribute to their total Hg and Se concentrations. In hatch-lings, absorption of the yolk and ingestion of egg albumen/amni-otic fluid may cause blood Se levels to be elevated over those inthe liver when this form is present due to the lack of bodily regu-lation (Burk and Levander, 1999). Overall, blood Se concentrationsin hatchlings were 4500 times higher than blood Hg concentra-tions; liver Se concentrations were 130 times higher than liverHg concentrations (nesting leatherback blood Se:Hg: 907:1, Guirletet al., 2008; green turtle liver Se:Hg: 12:1, Anan et al., 2001;hawksbill turtle liver Se:Hg: 56:1; diamondback terrapin liverSe:Hg: 1.4:1, Burger, 2002; Black-footed albatross liver Se:Hg:0.78:1, Ikemoto et al., 2004a; common eider ducks liver Se:Hg:7.9:1, Wayland et al., 2000; common eider ducks bloodSe:Hg: 17.8:1, Wayland et al., 2000; ringed and bearded seal liverSe:Hg, <1:1, Smith and Armstrong, 1978, for review; Northern furseal liver Se:Hg, 1:1, Ikemoto et al., 2004a; Dall’s porpose liverSe:Hg: 2.5:1, Ikemoto et al., 2004a; pilot whale blood Se:Hg:3.9:1, Nielsen et al., 2000; sperm whale blood Se:Hg: 0.6:1, Nielsenet al., 2000). Sea turtles have higher Se:Hg ratios than otherbrackish and marine organisms. Therefore, nesting females passon essential elements to their young at greater concentrations thannon-essential and potentially harmful elements.
We found measurable concentrations of Hg in eggshells and Hgand Se in the SAGs of leatherback turtles. Lam et al. (2006) and Bur-ger (1994) reported Hg and Se in green turtle (Chelonia mydas) egg-shells from Hong Kong and in avian eggshells from Long Island,New York, respectively. Egg-laying may provide a means of toxi-cant elimination in reptiles (Burger, 1994), although the amountexcreted may be insignificant (Sakai et al., 1995; Guirlet et al.,2008). SAGs may also reduce toxicant loads slightly in nesting fe-male leatherbacks without causing harm to the offspring. The albu-men and eggshells are produced progressively during the nestingseason. Together, these findings provide a plausible explanationfor the decrease in Hg burden as the nesting season progressed.
4.3. Hatching and emergence success
We found that the concentration of Se (a necessary nutrient) inthe liver of hatchlings as well as the higher ratio of hatchling Se toHg were both positively correlated with leatherback turtle hatch-ing and emergence success at the Florida rookery. While we didnot find a significant correlation between hatching and emergencesuccess and maternal blood Hg and/or Se concentrations, this maynot be surprising. Blood concentrations are more indicative of thecurrent status in the body and of recent dietary intake. Evidence forfeeding by gravid leatherbacks (Casey et al., 2010) suggests preyand seawater intake is low during the nesting season. Mercuryconcentrations are almost always higher in liver than blood, butthis depends on the age, diet, and species being tested (Puls,
1994; Blanvillain et al., 2007). In general, concentrations of theseelements in the liver are better indicators of long term status inthe body, but this sampling technique is not practical for live ani-mals, particularly free-ranging wildlife. Day et al. (2005) found thatHg concentrations in the scutes of loggerheads were accurate pre-dictors of the Hg concentrations in the liver based on strandedindividuals. They also reported that blood and scute concentrationswere highly correlated in loggerheads from the eastern Atlantic;these results provide guidance for sampling other cheloniids, butnot leatherbacks. Because leatherbacks lack thick keratinous scutes(Deraniyagala, 1939), such a comparison cannot be made inleatherbacks.
Miller et al. (2009) documented cardiac and skeletal muscleanomalies in dead-in-nest and captive reared leatherback hatch-lings and post-hatchlings. They noted that the cardiac changeswere similar to those seen in bovine neonates that were deficientin Se (a condition that can result from elevated Hg concentrations;Enjalbert et al., 1999). They hypothesized that maternal or hatch-ling Hg burden/Se status may have contributed to the muscularanomalies and to low hatching and emergence success of the nests.Our findings support the Miller et al. (2009) hypothesis. Both Sestatus and Hg load in leatherback sea turtle hatchlings correlatedwith hatching and emergence success (Figs. 4 and 5). Seleniumdeficiency is linked to muscular degeneration in a variety of ani-mals (Dierenfeld, 1989; Enjalbert et al., 1999; Miller et al., 2009),which may explain decreased hatching/emergence success, ashatchlings must rely on adequate skeletal muscle performance toescape from eggs and the nest.
Selenium is important in detoxifying Hg and other toxicantsincluding cadmium and lead in the body (Naganuma et al., 1983;Sasakura and Suzuki, 1998; Ikemoto et al., 2004a). Mercury concen-trations have been documented in a number of fish, birds, andmammals and its toxic effects include impairment of the nervoussystem, immune system, growth, and development (Zelikoff et al.,1994; Wiener et al., 2003; Day et al., 2007). In marine mammalsand seabirds, methylmercury, the most toxic form, is converted toinorganic Hg (e.g., mercuric chloride, mercuric acetate, and mercu-ric sulfide) by Se (Iwata et al., 1982) and to a lesser extent, othermechanisms (reactive oxygen species: Yasutake and Hirayama,2001; gut bacteria: Rowland, 1988). The inorganic forms mostlikely bind to metallothioneins and subsequently to proteins (highmolecular weight substances, HMWS) in the liver (Ikemoto et al.,2004b). Decomposition of these Hg–Se bound HMWS produces aninsoluble form (Ikemoto et al., 2004a) that can be excreted (Ralstonand Raymond, 2010). While leatherback hatchlings have Hg and Seconcentrations that suggest detoxification is likely, embryos andhatchlings may not be fully protected. Embryonic leatherbacksmay be more sensitive to heavy metal toxicity, as Se may fail to pro-tect the developing embryo during early developmental stages (e.g.,before functional liver formation) as occurs in developing carp(Cyprinus carpio) and chicks (Huckabee and Griffith, 1974; Birgeet al., 1976; Cuvin-Aralar and Furness, 1991). If true in leatherbackhatchlings from Florida, such developmental limitations may ex-plain the decrease in hatching and emergence success when Sewas present in lower ratios when compared to Hg.
In seabirds, methylmercury is the dominant form of Hg in theblood, whereas in the liver and kidney, it is generally present asthe less toxic, inorganic form, suggesting that it has been detoxifiedby Se or metallothioneins (Clarkson, 1994; Thompson et al., 1990).Selenium, which is found at high concentrations in leatherbacks,could act as a protective mechanism against Hg in this species.Additionally, Se concentrations in hatchlings may be at the idealrange for the leatherbacks, allowing for an optimal standard meta-bolic rate for growth and development (Mitchelmore et al., 2006).Selenium in elevated concentrations can cause toxicity; however, fewdata exist on Se and Hg toxicity in reptiles. The Se concentrations
J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682 1679

Author's personal copy
observed in this study (for both liver and blood; adults andhatchlings) fell below or within the range of toxicity thresholdsfor Se predicted for other egg-laying vertebrates (3–16 ppm, fish:Skorupa, 1998; birds: Skorupa and Ohlendorf, 1991; Heinz, 1996;Lemly, 1996; Fairbrother et al., 1999) and were much lower thanSe concentrations that caused anatomical and pathological anom-alies in aquatic birds (eggs: 2.2–110 ppm; livers: 19–130 ppm;Ohlendorf et al., 1986). Mercury concentrations of the hatchlings(blood, liver, and yolk sac) were much lower than those predictedto harm waterbirds (�5 ppm in liver, Zillioux et al., 1993), wildcommon terns (Sterna hirundo, �0.5–1.5 ppm; Fimreite, 1974),and developing captive bred mallard embryos (�1 ppm; Heinzand Hoffman, 2003). Overall, Se’s protective effect against Hg andother toxicants and its importance to growth and development of-fers no surprise that it correlates with the reproductive success ofmarine organisms, including the leatherback sea turtle.
5. Conclusions
This study is the first to correlate both maternal and hatchlingcontaminant loads with reproductive success in sea turtles and isalso the first to document Hg and Se concentrations (blood, liver,and yolk sac) in any hatchling sea turtle species. It is also the firstto provide evidence that SAGs of leatherbacks may help decreasethe bodily burden of toxicants through elimination. Most impor-tantly, we found that Se and the ratio of Se to Hg positively corre-lated with leatherback sea turtle hatching and emergence success.Therefore, the physiological protection of Se against Hg may allowfor more live turtles to hatch and subsequently emerge from thenests. This increase in hatchling production could lead to an in-crease in the leatherback sea turtle population, which is extremelyimportant for this globally imperiled species. Previous studies oftoxicant levels in sea turtles have documented the effects of con-taminants on clinical health parameters in subadult and adultsea turtles (POPs: Keller et al., 2004; Hg: Day et al., 2007; Hg andSe: Innis et al., 2008) and their effects on hatchling body condition(POPs: van de Merwe et al., 2010), but none have focused on nest-ing females and their consequences to reproduction. Further inves-tigations of trace element levels and their toxicokinetics areneeded for hatchling sea turtles to establish baselines, mechanismsof detoxification, and causal relationships.
Acknowledgements
The authors thank the Loggerhead Marinelife Center staff andvolunteers, including S. Bergeron, S. Fournies, K. Garrido, and M.Merrill. K. Stewart provided valuable discussions regarding thisproject. J.E. Knowles provided assistance with maps. The authorsalso thank C.E. Proffitt for statistical advice and D. Scheurle foruse of equipment. Lastly, the authors thank L. Bryan, T. Cook, E.Courtney, E. Dougherty, E. Eads, J. Lasala, M. Martin, A. Merrill, C.Ross, R. Timmons, and J. Yost for sample analyses and field assis-tance. The authors thank the reviewer for his or her thorough re-view and thoughtful comments on this manuscript. This studywas supported, in part, by a Florida Sea Turtle License Plate Grantto D.L.M., The University of Georgia Veterinary Diagnostic andInvestigational Laboratory, the FAU Nelligan Fund, and personalfunds. All procedures were in adherence to Florida Fish and Wild-life Conservation Commission Marine Turtle Permit #073 condi-tions and FAU IACUC approval A07-03.
References
Ackerman, R.A., Seagrave, R.C., Dmi’el, R., Ar, A., 1985. Water and heat exchangebetween parchment-shelled reptile eggs and their surroundings. Copeia 1985,703–711.
Anan, Y., Kunjito, T., Watanabe, I., Sakai, H., Tanabe, S., 2001. Trace elementaccumulation in hawksbill turtles (Eretmochelys imbricata) and green turtles(Chelonia mydas) from Yaeyama Islands. Jpn. Environ. Toxicol. Chem. 20, 2802–2814.
Avens, L., Taylor, J.C., Goshe, L.R., Jones, T.T., Hastings, M., 2009. Use ofskeletochronological analysis to estimate age of leatherback sea turtlesDermochelys coriacea in the western North Atlantic. Endangered Species Res.8, 165–177.
Beland, P., De Guise, S., Girard, C., Lagace, A., Martineau, D., Michaud, R., Muir, D.C.G.,Norstrom, R.J., Pelletier, E., Ray, S., Shugart, L.R., 1993. Toxic compounds andhealth and reproductive effects in St. Lawrence beluga whales. J. Great LakesRes. 19, 766–775.
Bell, B.A., Spotila, J.R., Paladino, F.V., Reina, R.D., 2003. Low reproductive success ofleatherback turtles, Dermochelys coriacea, is due to high embryonic mortality.Biol. Conserv. 115, 131–138.
Birge, W.J., Roberts, O.W., Black, J.A., 1976. Toxicity of metal mixtures to chickembryos. Bull. Environ. Contam. Toxicol. 6, 314–318.
Bjorndal, K., 1997. Foraging ecology and nutrition of sea turtles. In: Lutz, P.L.,Musick, J.A. (Eds.), The Biology of Sea Turtles. CRC Press, Boca Raton, pp. 199–231.
Blanvillain, G., Schwenter, J.A., Day, R.D., Point, D., Christopher, S.J., Roumillat, W.A.,Owens, D.W., 2007. Diamondback terrapins, Malaclemys terrapin, as a sentinelspecies for monitoring mercury pollution of estuarine systems in South Carolinaand Georgia, USA. Environ. Toxicol. Chem. 26, 1441–1450.
Bryan Jr., A.L., Hopkins, W.A., Baionno, J.A., Jackson, B.P., 2003. Maternal transfer ofcontaminants to eggs in common grackles (Quiscalus quiscala) nesting on coalfly ash basins. Arch. Environ. Contam. Toxicol. 45, 273–277.
Burger, J., 1992. Trace element levels in pine snake hatchlings: tissue and temporaldifferences. Arch. Environ. Contam. Toxicol. 22, 209–213.
Burger, J., 1994. Heavy metals in avian eggshells: another excretion method. J.Toxicol. Environ. Health 41, 207–220.
Burger, J., 2002. Metals in tissues of diamondback terrapins from New Jersey.Environ. Monit. Assess. 77, 255–263.
Burgess, N.M., Meyer, M.W., 2008. Methylmercury exposure associated withreduced productivity in common loons. Ecotoxicology 17, 83–91.
Burk, R.F., Levander, O.A., 1999. Selenium. In: Shills, M.E., Olson, J.A., Shike, M., Ross,A.C. (Eds.), Modern Nutrition in Health and Disease, ninth ed. Williams &Wilkins, Baltimore, pp. 561–569.
Cagle, F.R., 1950. Life history of the slider turtle, Pseudemys scripta troostii(Holbrook). Ecol. Monogr. 20, 31–54.
Campbell, L.M., Norstrom, R.J., Hobson, K.A., Muir, D.C., Backus, S., Fisk, A.T., 2005.Mercury and other trace elements in a pelagic Arctic marine food web(Northwater Polynyna, Baffin Bay). Sci. Total Environ. 351–352, 247–263.
Cardellicchio, N., Decataldo, A., Di Leo, A., Misino, A., 2002. Accumulation and tissuedistribution of mercury and selenium in striped dolphins (Stenella coeruleoalba)from the Mediterranean Sea (southern Italy). Environ. Pollut. 116, 265–271.
Casey, J., Garner, J., Garner, S., Williard, A.S., 2010. Diel foraging behavior of gravidleatherback sea turtles in deep waters of the Caribbean Sea. J. Exp. Biol. 213,3961–3971.
Caurant, F., Bustamante, P., Bordes, M., Miramand, P., 1999. Bioaccumulation ofcadmium, copper and zinc in some tissues of three species of marine turtlesstranded along the French Atlantic coasts. Mar. Pollut. Bull. 38, 1085–1091.
Caurant, F., Navarro, M., Amiard, J.-C., 1996. Mercury in pilot whales: possible limitsto the detoxification process. Sci. Total Environ. 186, 95–104.
Clarkson, T.W., 1994. The toxicology of mercury and its compounds. In: Watras, C.J.,Huckabee, J.W. (Eds.), Mercury Pollution – Integration and Synthesis. CRC Press,Boca Raton, pp. 631–641.
Cuvin-Aralar, M.L., Furness, R.W., 1991. Mercury and selenium interaction: areview. Ecotoxicol. Environ. Saf. 21, 348–364.
Davenport, J., 1997. Temperature and the life-history strategies of sea turtles. J.Therm. Biol. 22, 479–488.
Davenport, J., Wrench, J., McEvoy, J., Camacho-Ibar, V., 1990. Metal and PCBconcentrations in the ‘‘Harlech’’ leatherback. Mar. Turtle News 48, 1–6.
Day, R.D., Christopher, S.J., Becker, P.R., Whitaker, D.W., 2005. Monitoring mercuryin the loggerhead sea turtle, Caretta caretta. Environ. Sci. Technol. 39, 437–446.
Day, R.D., Segars, A.L., Arendt, M.D., Lee, A.M., Peden-Adams, M.M., 2007.Relationship of blood mercury levels to health parameters in the loggerheadsea turtle (Caretta caretta). Environ. Health Perspect. 115, 1421–1428.
Deem, S.L., Dierenfeld, E.S., Sounguet, G.P., Alleman, A.R., Cray, C., Poppenga, R.H.,Norton, T.M., Karesh, W.B., 2006. Blood values in free-ranging nestingleatherback sea turtles (Dermochelys coriacea) on the coast of the Republic ofGabon. J. Zoo. Wildl. Med. 37, 464–471.
Deem, S.L., Norton, T.M., Mitchell, M., Segars, A., Alleman, A.R., Cray, C., Poppenga,R.H., Dodd, M., Karesh, W.B., 2009. Comparison of blood values in foraging,nesting and stranded loggerhead turtles (Caretta caretta) along the coast ofGeorgia, USA. J. Wildl. Dis. 45, 41–56.
Deraniyagala, P.E.P., 1939. The Tetrapod Reptiles of Ceylon, vol. I: Testudinates andCrocodilians. Ceylon Journal of Science. Sunil Printers, New Delhi, pp. 1–242.
Dierenfeld, E.S., 1989. Vitamin E deficiency in zoo reptiles, birds, and ungulates. J.Zoo. Wildl. Med. 20, 3–11.
Dietary Reference Intakes (DRI), 2000. National Research Council. NationalAcademy Press, Washington, pp. 284–319.
Dutton, P., 1996. Methods for collection and preservation of samples for sea turtlegenetic studies. In: Bowen, B.W., Witzell, W.N. (Eds.), Proceedings of theInternational Symposium on Sea Turtle Conservation Genetics. NOAA TechnicalMemorandum NMFS-SEFSC-396, 173 pp.
1680 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682

Author's personal copy
Dutton, P., Dutton, D., 1994. Use of PIT tags to identify adult leatherbacks. Mar.Turtle News 67, 13–14.
Eckert, S.A., Bagley, D., Kubis, S., Ehrhart, L., Johnson, C., Stewart, K., DeFreese, D.,2006. Internesting and postnesting movements and foraging habitats ofleatherback sea turtles (Dermochelys coriacea) nesting in Florida. Chel.Conserv. Biol. 5, 239–248.
Enjalbert, F.P., Lebreton, P., Salat, O., Schelcher, F., 1999. Effects of pre- orpostpartum selenium supplementation on selenium status in beef cows andtheir calves. J. Anim. Sci. 77, 223–229.
EPA, 1985. Ambient water quality criteria for mercury – 1984. U.S. EnvironmentalProtection Agency Report 440/5-84-026, 136 pp. Available from Natl. Tech.Infor. Serv., 5285 Port Royal Road, Springfield, Virginia 22161.
Evans, D., Ordoñez, C., Troëng, S., Drews, C., 2008. Satellite tracking of leatherbackturtles from Caribbean Central America reveals unexpected foraging grounds.In: Frick, M., Panagopoulou, A., Rees, A.F., Williams, K. (Eds.), Proceedings of theTwenty-Seventh Annual Symposium on Sea Turtle Biology and Conservation.NOAA Technical Memorandum NMFS-SEFSC-569, 262 p.
Fairbrother, A., Brix, K.V., Toll, J.E., McKay, S., Adams, W.J., 1999. Egg seleniumconcentrations as predictors of avian toxicity. Hum. Ecol. Risk Assess. 5, 1229–1253.
Fimreite, N., 1974. Mercury contamination of aquatic birds in northwestern Ontario.J. Wildl. Manage. 38, 120–131.
Godley, B.J., Gaywood, M.J., Law, R.J., McCarthy, C.J., McKenzie, C., Patterson, I.A.P.,Penrose, R.S., Reid, R.J., Ross, H.M., 1998. Patterns of marine turtle mortality inBritish waters (1992–1996) with reference to tissue contaminant levels. J. Mar.Biol. Assoc. UK 78, 973–984.
Guirlet, E., Das, K., Girondot, M., 2008. Maternal transfer of trace elements ofleatherback turtles of French Guiana. Aquat. Toxicol. 88, 267–276.
Guirlet, E., Das, K., Thomé, J.P., Girondot, M., 2010. Maternal transfer of chlorinatedcontaminants in the leatherback turtles, Dermochelys coriacea, nesting in FrenchGuiana. Chemosphere 79, 720–726.
Harris, H.S., Benson, S.R., Gilardi, K.V., Poppenga, R.H., Dutton, P.H., Work, T.M.,Mazet, J.A.K., 2011. Comparative health assessment of western Pacificleatherback turtles (Dermochelys coriacea) foraging off the coast of California:2005–2007. J. Wildl. Dis. 47, 321–337.
Hays, G.C., Hobson, V.J., Metcalfe, J.D., Righton, D., Sims, D.W., 2006. Flexibleforaging movements of leatherback turtles across the north Atlantic Ocean.Ecology 87, 2647–2656.
Heck, J., MacKenzie, D.S., Rostal, D., Medler, K., Owens, D., 1997. Estrogen inductionof plasma vitellogenin in the Kemp’s ridley sea turtle (Lepidochelys kempii). Gen.Comp. Endocrinol. 107, 280–288.
Heinz, G.H., 1996. Selenium in birds. In: Beyer, W.N., Heinz, G.H., Redmon-Norwood,A.W. (Eds.), Environmental Contaminants in Wildlife: Interpreting TissueConcentrations. CRC Press, Boca Raton, pp. 447–458.
Heinz, G.H., Hoffman, D.J., 2003. Embryotoxic thresholds of mercury:estimates from individual mallard eggs. Arch. Environ. Contam. Toxicol. 44,257–264.
Heinz, G.H., Hoffman, D.J., Krynitsky, A.J., Weller, D.M., 1987. Reproduction inmallards fed selenium. Environ. Toxicol. Chem. 6, 423–433.
Hernández, R., Buitrago, J., Guada, H., Hernández-Hamón, H., Llano, M., 2007.Nesting distribution and hatching success of the leatherback, Dermochelyscoriacea, in relation to human pressures at Playa Parguito, Margarita Island,Venezuela. Chel. Conserv. Biol. 6, 79–86.
Hilterman, M.L., Goverse, E., 2003. Aspects of nesting and nest success of theleatherback turtle (Dermochelys coriacea) in Suriname, 2002. GuianasForests and Environmental Conservation Project (GFECP). Technical Report,World Wildlife Fund Guianas/Biotopic Foundation, Amsterdam, theNetherlands, 31 p.
Hoffman, D.J., 2002. Role of selenium toxicity and oxidative stress in aquatic birds.Aquat. Toxicol. 57, 11–26.
Hopkins, W.A., 2006. Use of tissue residues in reptile ecotoxicology: a call forintegration and experimentalism. In: Gardner, S.C., Oberdörster, E. (Eds.),Toxicology of Reptiles. CRC Press, Boca Raton, pp. 35–62.
Huckabee, J.W., Griffith, N.A., 1974. Toxicity of mercury and selenium to the eggs ofcarp (Cyprinus carpio). Trans. Am. Fish. Soc. 103, 822–825.
Ikemoto, T., Kunito, T., Tanaka, H., Baba, N., Miyazaki, N., Tanabe, S., 2004a.Detoxification mechanisms of heavy metals in marine mammals and seabirds:interaction of selenium with mercury, silver, copper, zinc, and cadmium in liver.Arch. Environ. Contam. Toxicol. 47, 402–413.
Ikemoto, T., Kunito, T., Anan, Y., Tanaka, H., Baba, N., Miyazaki, N., Tanabe, S., 2004b.Association of heavy metals with metallothionein and other proteins in hepaticcytosol of marine mammals and seabirds. Environ. Toxicol. Chem. 23, 2008–2016.
Innis, C., Merigo, C., Dodge, K., Tlusty, M., Dodge, M., Sharp, B., Myers, A., McIntosh,A., Wunn, D., Perkins, C., Herdt, T.H., Norton, T., Lutcavage, M., 2010. Healthevaluation of leatherback turtles (Dermochelys coriacea) in the NorthwesternAtlantic during direct capture and fisheries gear disentanglement. Chel.Conserv. Biol. 9, 205–222.
Innis, C., Tlusty, M., Perkins, C., Holladay, S., Merigo, C., Weber III, E.S., 2008. Tracemetal and organochlorine pesticide concentrations in cold-stunned juvenileKemp’s ridley turtles (Lepidochelys kempii) from Cape Cod, Massachusetts. Chel.Conserv. Biol. 7, 230–239.
Iwata, H., Masukawa, T., Kito, H., Hayashi, M., 1982. Degradation of methylmercuryby selenium. Life Sci. 31, 859–866.
Jacobson, E.R., 2007. Collection biological samples for clinical evaluation. Availablefrom: <http://www.iacuc.ufl.edu/AnimalUseGuides/BiolSamColl.doc>.
Kam, Y.C., Ackerman, R.A., 1990. The effect of incubation media on the waterexchange of snapping turtle (Chelydra serpentine) eggs and hatchlings. J. Comp.Physiol. B 160, 317–324.
Kari, T., Kauranen, P., 1978. Mercury and selenium contents of seals from fresh andbrackish water in Finland. Bull. Environ. Contam. Toxicol. 19, 273–280.
Keller, J.M., Kucklick, J.R., Stamper, M.A., Harms, C.A., McClellan-Green, P.D., 2004.Associations between organochlorine contaminant concentrations and clinicalhealth parameters in loggerhead sea turtles from North Carolina, USA. Environ.Health Perspect. 112, 1074–1079.
Kenyon, L.O., Landry Jr., A.M., Gill, G.A., 2001. Trace metal concentrations in blood ofthe Kemp’s ridley sea turtle (Lepidochelys kempii). Chel. Conserv. Biol. 4, 128–135.
Lam, J.C.W., Tanabe, S., Chan, S.K.F., Lam, M.H.W., Martin, M., Lam, P.K.S., 2006.Levels of trace elements in green turtle eggs collected from Hong Kong:evidence of risks due to selenium and nickel. Environ. Pollut. 144, 790–801.
Lemly, A.D., 1996. Selenium in aquatic organisms. In: Beyer, W.N., Heinz, G.H.,Redmon-Norwood, A.W. (Eds.), Environmental Contaminants in Wildlife:Interpreting Tissue Concentrations. CRC Press, Boca Raton, pp. 427–445.
Leslie, A.J., Penick, D.N., Spotila, J.R., Paladino, F.V., 1996. Leatherback turtle,Dermochelys coriacea, nesting and nest success at Tortuguero, Costa Rica, in1990–1991. Chel. Conserv. Biol. 2, 159–168.
Livingstone, S.R., 2006. Sea turtle ecology and conservation on the north coast ofTrinidad. Ph.D. Dissertation. University of Glasgow, Glasgow, Lanarkshire,Scotland, UK.
Lutcavage, M., Lutz, P.L., 1986. Metabolic rate and food energy requirements of theleatherback sea turtle, Dermochelys coriacea. Copeia 1986, 796–798.
Magat, W., Sell, J.L., 1979. Distribution of mercury and selenium in egg componentsand egg-white proteins. Proc. Soc. Exp. Biol. Med. 161, 458–463.
Mayr, R., 1977. Populations, Species, and Evolution, sixth ed. Harvard UniversityPress, Cambridge.
Miller, J.D., 1997. Reproduction in sea turtles. In: Lutz, P.L., Musick, J.A. (Eds.), TheBiology of Sea Turtles. CRC Press, Boca Raton, pp. 51–82.
Miller, J.D., 1999. Determining clutch size and hatching success. In: Eckert, K.L.,Bjorndal, K.A., Abreu-Grobois, F.A., Donnelly, M. (Eds.), Research andManagement Techniques for the Conservation of Sea Turtles. IUCN/SSCMarine Turtle Specialist Group Publication No. 4.
Miller, D.L., Wyneken, J., Rajeev, S., Perrault, J., Mader, D.R., Weege, J., Baldwin, C.A.,2009. Pathological findings in hatchling and post-hatchling leatherback seaturtles (Dermochelys coriacea) from Florida. J. Wildl. Dis. 45, 962–971.
Mitchelmore, C.L., Rowe, C.L., Place, A.R., 2006. Tools for assessing contaminantexposure and effects in reptiles. In: Gardner, S.C., Oberdörster, E. (Eds.),Toxicology of Reptiles. CRC Press, Boca Raton, pp. 63–122.
Naganuma, A., Tanaka, T., Maeda, K., Matsuda, R., Tabata-Hanyu, J., Imura, N., 1983.The interaction of selenium with various metals in vitro and in vivo. Toxicology29, 77–86.
Nagle, R.D., Rowe, C.L., Congdon, J.D., 2001. Accumulation and selective maternaltransfer of contaminants in the turtle Trachemys scripta associated with coal ashdeposition. Arch. Environ. Contam. Toxicol. 40, 531–536.
Nielsen, J.B., Nielsen, F., Jørgensen, P.-J., Grandjean, P., 2000. Toxic metals andselenium in blood from pilot whales (Globicephala melas) and sperm whales(Physeter catodon). Mar. Pollut. Bull. 40, 348–351.
Ohlendorf, H.M., Hoffman, D.J., Saiki, M.K., Aldrich, T.W., 1986. Embryonic mortalityand abnormalities of aquatic birds: apparent impacts of selenium fromirrigation drainwater. Sci. Total Environ. 52, 49–63.
Ohlendorf, H.M., Kilness, A.W., Simmons, J.L., Stroud, R.K., Hoffman, D.J., Moore, J.F.,1988. Selenium toxicosis in wild aquatic birds. J. Toxicol. Environ. Health 24,67–92.
Orr, J.P., Blakely, B.R., 1997. Investigation of the selenium status of abortedcalves with cardiac failure and myocardial necrosis. J. Vet. Diagn. Invest. 9, 172–179.
Owens, D.W., Ruiz, G.J., 1980. New methods of obtaining blood from cerebrospinalfluid from marine turtles. Herpetologica 36, 17–20.
Pacyna, E.G., Pacyna, J.M., 2002. Global emission of mercury from anthropogenicsources in 1995. Water Air Soil Pollut. 137, 149–165.
Palmer, B.D., Guillette, L.J., 1991. Oviductal proteins and their influence onembryonic development in birds and reptiles. In: Deeming, D.C., Ferguson,M.W.J. (Eds.), Egg Incubation: Its Effects on Embryonic Development in Birdsand Reptiles. Cambridge University Press, Cambridge, pp. 29–46.
Pitman, R.L., Dutton, P.H., 2004. Killer whale predation on a leatherback turtle in thenortheast Pacific. Pac. Sci. 58, 497–498.
Pritchard, P.C.H., 1982. Nesting of the leatherback turtle, Dermochelys coriacea, inPacific Mexico, with a new estimate of the world population status. Copeia1982, 741–747.
Puls, R., 1994. Mineral Levels in Animal Health: Diagnostic Data, second ed. SherpaInternational, Clearbrook, British Columbia.
Rainwater, T.R., Reynolds, K.D., Cañas, J.E., Cobb, G.P., Anderson, T.A., McMurry, S.T.,Smith, P.N., 2005. Organochlorine pesticides and mercury in cottonmouths(Agkistrodon piscivorus) from northeastern Texas, USA. Environ. Toxicol. Chem.24, 665–673.
Ralston, N.V.C., Raymond, L.J., 2010. Dietary selenium’s protective effects againstmethylmercury toxicity. Toxicology 278, 112–123.
Rayman, M.P., 2000. The importance of selenium to human health. Lancet 356, 233–241.
Redfearn, E.C., 2000. A comparative approach to understanding sea turtle hatchlingmetabolism during emergence. Master’s Thesis. Florida Atlantic University,Boca Raton, FL, USA.
J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682 1681

Author's personal copy
Richards, M.P., 1997. Trace mineral metabolism in the avian embryo. Poult. Sci. 76,152–164.
Roe, J.H., Hopkins, W.A., Baionno, J.A., Staub, B.P., Rowe, C.L., Jackson, B.P., 2004.Maternal transfer of selenium in Alligator mississippiensis nesting downstreamfrom a coal-burning power plant. Environ. Toxicol. Chem. 23, 1969–1972.
Romanoff, A.L., 1967. Biochemistry of the Avian Embryo. John Wiley & Sons Inc.,New York.
Rostal, D.C., Grumbles, J.S., Byles, R.A., Márquez, M.R., Owens, D.W., 1997. Nestingphysiology of Kemp’s ridley sea turtles, Lepidochelys kempi, at Rancho Nuevo,Tamaulipas, Mexico, with observations on population estimates. Chel. Conserv.Biol. 2, 538–547.
Rostal, D.C., Grumbles, J.S., Palmer, K.S., Lance, V.A., Spotila, J.R., Paladino, F.V., 2001.Changes in gonadal and adrenal steroid levels in the leatherback sea turtle(Dermochelys coriacea) during the nesting cycle. Gen. Comp. Endocrinol. 122,139–147.
Rostal, D.C., Owens, D.W., Grumbles, J.S., MacKenzie, D.S., Amoss Jr., M.S., 1998.Seasonal reproductive cycle of the Kemp’s ridley sea turtle (Lepidochelyskempii). Gen. Comp. Endocrinol. 109, 232–243.
Rostal, D.C., Paladino, F.V., Patterson, R.M., Spotila, J.R., 1996. Reproductivephysiology of nesting leatherback turtles (Dermochelys coriacea) at Las BaulasNational Park Costa Rica. Chel. Conserv. Biol. 2, 230–236.
Rowland, I.R., 1988. Interactions of the gut microflora and the host in toxicology.Toxicol. Pathol. 16, 147–153.
Sakai, S., Ichihashi, H., Suganuma, H., Tatsukawa, R., 1995. Heavy metal monitoringin sea turtles using eggs. Mar. Pollut. Bull. 30, 347–353.
Sarti-Martinez, A.L., 2000. Dermochelys coriacea. In: IUCN 2011. IUCN Red List ofThreatened Species. Version 2011.1. Available from: <www.iucnredlist.org>.
Sasakura, C., Suzuki, K.T., 1998. Biological interaction between transition metals(Ag, Cd and mercury), selenide/sulfide and selenoprotein P. J. Inorg. Biochem.71, 159–162.
Skorupa, J.P., 1998. Selenium poisoning of fish and wildlife in nature: lessonsfrom twelve real-world examples. In: Frankenberger, W.T., Engberg, R.A.(Eds.), Environmental Toxicology, vol. 2. Elsevier Science, New York, pp. 59–116.
Skorupa, J.P., Ohlendorf, H.M., 1991. Contaminants in drainage water and avian riskthresholds. In: Dinar, A., Zilberman, D. (Eds.), The Economics and Managementof Water and Drainage in Agriculture. Kluwer Academic, Dordrecht, TheNetherlands, pp. 345–368.
Smith, T.G., Armstrong, F.A.J., 1978. Mercury and selenium in ringed and beardedseal tissues from Arctic Canada. Arctic 31, 75–84.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: The Principles and Practice of Statistics inBiological Research, third ed. W.H. Freeman and Company, New York.
Spallholz, J.E., Hoffman, D.J., 2002. Selenium toxicity: cause and effect in aquaticbirds. Aquat. Toxicol. 57, 27–37.
Spotila, J.R., Dunham, A.E., Leslie, A.J., Steyermark, A.C., Plotkin, P.T., Paladino, F.V.,1996. Worldwide population decline of Dermochelys coriacea: are leatherbackturtles going extinct? Chel. Conserv. Biol. 2, 209–222.
Stewart, K., 2007. Establishment and growth of a sea turtle rookery: the populationbiology of the leatherback in Florida. Ph.D. Dissertation. Duke University,Durham, NC, USA.
Stewart, K., Johnson, C., 2006. Dermochelys coriacea – leatherback sea turtle. In:Meylan, P.A. (Ed.), Biology and Conservation of Florida Turtles. ChelonianResearch Monographs, vol. 3, pp. 144–157.
Stillwell, R.J., Berry, M.J., 2005. Expanding the repertoire of the eukaryoticselenoproteome. Proc. Natl. Acad. Sci. USA 102, 16123–16124.
Stoneburger, D.L., 1978. Heavy metals in tissues of stranded short-finned pilotwhales. Sci. Total Environ. 9, 293–297.
Stoneburger, D.L., Nicora, M.N., Blood, E.R., 1980. Heavy metals in loggerhead seaturtle eggs (Caretta caretta): evidence to support the hypothesis that demesexist in the western Atlantic population. J. Herpetol. 14, 171–175.
Strik, N.I., Alleman, A.R., Harr, K.E., 2007. Circulating inflammatory cells. In:Jacobson, E.R. (Ed.), Infectious Disease and Pathology of Reptiles. CRC Press,Boca Raton, pp. 167–218.
Thompson, D.R., Stewart, F.M., Furness, R.W., 1990. Using seabirds to monitormercury in marine environments. Mar. Pollut. Bull. 21, 339–342.
Turtle Expert Working Group (TEWG), 2007. An assessment of the leatherbackturtle population in the Atlantic Ocean. NOAA Technical Memorandum NMFS-SEFSC-555, 116 pp.
United States Environmental Protection Agency (USEPA), 2000. Mercury transportand fate in watersheds. Star Report 10, National Center for EnvironmentalResearch, vol. 4. USEPA, Washington, DC, pp. 1–8.
Unrine, J.M., Jackson, B.P., Hopkins, W.A., Romanek, C., 2006. Isolation and partialcharacterization of proteins involved in maternal transfer of selenium in thewestern fence lizard (Sceloporus occidentalis). Environ. Toxicol. Chem. 25, 1864–1867.
van de Merwe, J.P., Hodge, M., Whittier, J.M., Ibrahim, K., Lee, S.Y., 2010. Persistentorganic pollutants in the green sea turtle, Chelonia mydas: nesting populationvariation, maternal transfer, and effects on development. Mar. Ecol. Prog. Ser.403, 269–278.
Wallace, B.P., Kilham, S.S., Paladino, F.V., Spotila, J.R., 2006a. Energy budgetcalculations indicate resource limitation in Eastern Pacific leatherback turtles.Mar. Ecol. Prog. Ser. 318, 263–270.
Wallace, B.P., Sotherland, P.R., Spotila, J.R., Reinal, R.D., Franks, B.F., Paladino, F.V.,2004. Biotic and abiotic factors affect the nest environment ofembryonicleatherback turtles, Dermochelys coriacea. Physiol. Biochem. Zool.77, 423–432.
Wallace, B.P., Sotherland, P.R., Tomillo, P.S., Bouchard, S.S., Reina, R.D., Spotila, J.R.,Paladino, F.V., 2006b. Egg components, egg size, and hatchling size inleatherback turtles. Comp. Biochem. Physiol. A 145, 524–532.
Wayland, M., Garcia-Fernandez, A.J., Neugebauer, E., Gilchrist, H.G., 2000.Concentrations of cadmium, mercury and selenium in blood, liver and kidneyof common eider ducks from the Canadia Arctic. Environ. Monit. Assess. 71,255–267.
Whitmore, C.P., Dutton, P.H., 1985. Infertility, embryonic mortality and nest-siteselection in leatherback and green sea turtles in Suriname. Biol. Conserv. 34,251–272.
Wibbels, T., Owens, D.Wm., Limpus, C.J., Reed, P.C., Amoss Jr., M.S., 1990. Seasonalchanges in serum gonadal steroids associated with migration, mating, andnesting in the loggerhead sea turtle (Caretta caretta). Gen. Comp. Endocrinol. 79,154–164.
Wiener, J., Krabbenhoft, D., Heinz, G., Scheuhammer, A., 2003. Ecotoxicology ofmercury. In: Hoffman, D., Rattner, B.A., Burton, G.A., Jr., Cairns, J., Jr. (Eds.),Handbook of Ecotoxicology. CRC Press, Boca Raton, pp. 409–463.
Wyneken, J., 2001. The Anatomy of Sea Turtles. U.S. Department of CommerceNOAA Technical Memorandum NMFS-SEFSC-470, pp. 1–172.
Yasutake, A., Hirayama, K., 2001. Evaluation of methylmercury biotransformationusing rat liver slices. Arch. Toxicol. 75, 400–406.
Zelikoff, J.T., Smialowicz, R., Bigazzi, P.E., Goyer, R.A., Lawrence, D.A., Maibach, H.I.,Gardner, D., 1994. Immunomodulation by metals. Fundam. Appl. Toxicol. 22, 1–7.
Zillioux, E.J., Porcella, D.B., Benoit, J.M., 1993. Mercury cycling and effects infreshwater wetland ecosystems. Environ. Toxicol. Chem. 12, 2245–2264.
Zug, G.R., Parham, J.F., 1996. Age and growth in leatherback turtles, Dermochelyscoriacea: a skeletochronolgical analysis. Chel. Conserv. Biol. 2, 244–249.
1682 J. Perrault et al. / Marine Pollution Bulletin 62 (2011) 1671–1682
Related Documents











