Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation Andrew E. Allen* † , Julie LaRoche ‡§ , Uma Maheswari*, Markus Lommer ‡ , Nicolas Schauer ¶ , Pascal J. Lopez*, Giovanni Finazzi , Alisdair R. Fernie ¶ , and Chris Bowler* § ** *Centre National de la Recherche Scientifique Unite Mixte de Recherche 8186, Dept of Biology, Ecole Normale Supe ´ rieure, 46 rue d’Ulm, 75005 Paris, France; ‡ Leibniz-Institut fu ¨ r Meereswissenschaften, 24105 Kiel, Germany; ¶ Max Planck Institute of Molecular Plant Physiology, Am Muhlenberg 1, 14476 Potsdam-Golm, Germany; Centre National de la Recherche Scientifique Unite Mixte de Recherche 7141, Universite ´ Paris 6 Institut de Biologie Physico-Chimique, 13 rue Pierre et Marie Curie, 75005 Paris, France; and **Stazione Zoologica, Villa Comunale, I 80121 Naples, Italy Edited by David M. Karl, University of Hawaii, Honolulu, HI, and approved May 3, 2008 (received for review December 4, 2007) Marine primary productivity is iron (Fe)-limited in vast regions of the contemporary oceans, most notably the high nutrient low chlorophyll (HNLC) regions. Diatoms often form large blooms upon the relief of Fe limitation in HNLC regions despite their prebloom low cell density. Although Fe plays an important role in controlling diatom distribution, the mechanisms of Fe uptake and adaptation to low iron availability are largely unknown. Through a combina- tion of nontargeted transcriptomic and metabolomic approaches, we have explored the biochemical strategies preferred by Phaeo- dactylum tricornutum at growth-limiting levels of dissolved Fe. Processes carried out by components rich in Fe, such as photosyn- thesis, mitochondrial electron transport, and nitrate assimilation, were down-regulated. Our results show that this retrenchment is compensated by nitrogen (N) and carbon (C) reallocation from protein and carbohydrate degradation, adaptations to chlorophyll biosynthesis and pigment metabolism, removal of excess electrons by mitochondrial alternative oxidase (AOX) and non-photochem- ical quenching (NPQ), and augmented Fe-independent oxidative stress responses. Iron limitation leads to the elevated expression of at least three gene clusters absent from the Thalassiosira pseudonana genome that encode for components of iron capture and uptake mechanisms. genome metabalomics photosynthesis transcriptomics nutrients T he oxidizing conditions of contemporary marine ecosystems result in exceedingly low levels of dissolved Fe (1), and the cellular Fe demand of modern phytoplankton is frequently in excess of Fe availability (1, 2). This paradox reflects the likelihood that their biochemical machinery evolved in the Fe-replete, reducing Proterozoic oceans. Fe limitation is a major factor controlling phytoplankton growth in the large, perennially high nutrient low chlorophyll (HNLC) regions in the subarctic Pacific, equatorial Pacific, and Southern Oceans (3–5) and may be important in the North Atlantic (6). Marine diatoms are the most important eukaryotic phytoplank- ton for carbon sequestration, contributing 40% of global oceanic organic carbon production per year. More than 70% of blooms stimulated by mesoscale in situ Fe fertilization of Fe-limited HNLC waters were dominated by diatoms (7), indicating that diatoms persist in chronically Fe-limited environments and resume rapid growth when the limitation pressure is alleviated. The tolerance of diatoms to Fe limitation varies widely between species (8–11). Phaeodactylum tricornutum is highly tolerant to Fe limita- tion and can grow in steady-state laboratory cultures at iron levels 50 times lower than those tolerated by the centric diatom Thalassiosira pseudonana (12). Steady-state growth of P. tri- cornutum is Fe limited in the range of 10–30 pmolliter 1 Fe (Fe is the sum of all unchelated Fe species) (Table 1), similar to T. oceanica and Pseudonitzschia spp., two open ocean diatoms that are commonly found in the most severely Fe limited regions of the world’s oceans (13). The recently completed genome sequence of P. tricornutum (http://genome.jgi-psf.org/Phatr2/Phatr2.home.html) sheds light on some important differences between P. tricornutum and T. pseudonana, which may account for their respective thresholds for Fe limitation. We evaluated the whole-cell response of P. tricornu- tum to Fe limitation with multiple approaches combining gene expression profiling and comparative genomics with gas chroma- tography-mass spectroscopy (GC-MS)-aided nontargeted metabo- lomic analysis and a range of physiological measurements. Genes Author contributions: A.E.A., J.L., P.J.L., and C.B. designed research; A.E.A., J.L., U.M., M.L., N.S., and G.F. performed research; J.L., P.J.L., G.F., A.R.F., and C.B. contributed new reagents/analytic tools; A.E.A., J.L., U.M., M.L., N.S., G.F., and A.R.F. analyzed data; and A.E.A., J.L., and C.B. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Data deposition: The data reported in this paper have been deposited in the Gene Expression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE8675). † Present address: J. Craig Venter Institute, 10355 Science Center Drive, San Diego, CA 92121. § To whom correspondence may be addressed. E-mail: [email protected] or [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0711370105/DCSupplemental. © 2008 by The National Academy of Sciences of the USA Table 1. General cellular, physiological, and biochemical characteristics of Fe-limited P. tricornutum cells and cultures Parameter Fe-limited Fe-replete Fe-limited/ Fe-replete Growth rate 0.18 0.05 0.88 0.01 0.2 0.06 Ferric reductase assay, AU per cell 0.717 0.03 0.020 0.004 35.9 7.3 Ferric reductase assay, AU/m 3 0.01 0.0002 50 Carbon fixation, pmol C per cell hour 0.010 0.006 0.141 0.022 0.07 0.04 Cell diameter, m 2.5 3.5 0.71 Cell volume, m 3 60 100 0.60 Chl per cell, pg per cell 0.22 0.5 0.44 Chl per volume, fg/m 3 3.67 5.0 0.73 Fv/Fm 0.18 0.1 0.5 0.05 0.36 0.2 PSII 1.2 0.14 1 1.2 0.14 PSI 0.43 0.1 1.0 0.43 0.10 Cytochrome b6f 0.75 0.13 1.0 0.75 0.13 Cytochrome c6 0.7 0.05 1.0 0.70 0.05 NPQ 4.8 0.8 3.1 0.51 1.5 0.4 AOX activity 0.6 0.15 0.25 0.18 2.4 1.83 Photosystem and electron carrier concentrations are given in relative units normalized to Fe-replete levels. AOX activity is given as a percentage of total respiration (see SI Methods for details). Fe levels in Fe-limited and Fe-replete cultures corresponded to 13.4 pmolliter 1 Fe and 2.6 nmolliter 1 Fe, respectively. 10438 –10443 PNAS July 29, 2008 vol. 105 no. 30 www.pnas.orgcgidoi10.1073pnas.0711370105

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Whole-cell response of the pennate diatomPhaeodactylum tricornutum to iron starvationAndrew E. Allen*†, Julie LaRoche‡§, Uma Maheswari*, Markus Lommer‡, Nicolas Schauer¶, Pascal J. Lopez*,Giovanni Finazzi�, Alisdair R. Fernie¶, and Chris Bowler*§**
*Centre National de la Recherche Scientifique Unite Mixte de Recherche 8186, Dept of Biology, Ecole Normale Superieure, 46 rue d’Ulm, 75005 Paris, France;‡Leibniz-Institut fur Meereswissenschaften, 24105 Kiel, Germany; ¶Max Planck Institute of Molecular Plant Physiology, Am Muhlenberg 1, 14476Potsdam-Golm, Germany; �Centre National de la Recherche Scientifique Unite Mixte de Recherche 7141, Universite Paris 6 Institut de BiologiePhysico-Chimique, 13 rue Pierre et Marie Curie, 75005 Paris, France; and **Stazione Zoologica, Villa Comunale, I 80121 Naples, Italy
Edited by David M. Karl, University of Hawaii, Honolulu, HI, and approved May 3, 2008 (received for review December 4, 2007)
Marine primary productivity is iron (Fe)-limited in vast regions ofthe contemporary oceans, most notably the high nutrient lowchlorophyll (HNLC) regions. Diatoms often form large blooms uponthe relief of Fe limitation in HNLC regions despite their prebloomlow cell density. Although Fe plays an important role in controllingdiatom distribution, the mechanisms of Fe uptake and adaptationto low iron availability are largely unknown. Through a combina-tion of nontargeted transcriptomic and metabolomic approaches,we have explored the biochemical strategies preferred by Phaeo-dactylum tricornutum at growth-limiting levels of dissolved Fe.Processes carried out by components rich in Fe, such as photosyn-thesis, mitochondrial electron transport, and nitrate assimilation,were down-regulated. Our results show that this retrenchment iscompensated by nitrogen (N) and carbon (C) reallocation fromprotein and carbohydrate degradation, adaptations to chlorophyllbiosynthesis and pigment metabolism, removal of excess electronsby mitochondrial alternative oxidase (AOX) and non-photochem-ical quenching (NPQ), and augmented Fe-independent oxidativestress responses. Iron limitation leads to the elevated expressionof at least three gene clusters absent from the Thalassiosirapseudonana genome that encode for components of iron captureand uptake mechanisms.
genome � metabalomics � photosynthesis � transcriptomics � nutrients
The oxidizing conditions of contemporary marine ecosystemsresult in exceedingly low levels of dissolved Fe (1), and the
cellular Fe demand of modern phytoplankton is frequently in excessof Fe availability (1, 2). This paradox reflects the likelihood thattheir biochemical machinery evolved in the Fe-replete, reducingProterozoic oceans. Fe limitation is a major factor controllingphytoplankton growth in the large, perennially high nutrient lowchlorophyll (HNLC) regions in the subarctic Pacific, equatorialPacific, and Southern Oceans (3–5) and may be important in theNorth Atlantic (6).
Marine diatoms are the most important eukaryotic phytoplank-ton for carbon sequestration, contributing �40% of global oceanicorganic carbon production per year. More than 70% of bloomsstimulated by mesoscale in situ Fe fertilization of Fe-limited HNLCwaters were dominated by diatoms (7), indicating that diatomspersist in chronically Fe-limited environments and resume rapidgrowth when the limitation pressure is alleviated. The tolerance ofdiatoms to Fe limitation varies widely between species (8–11).
Phaeodactylum tricornutum is highly tolerant to Fe limita-tion and can grow in steady-state laboratory cultures at ironlevels 50 times lower than those tolerated by the centric diatomThalassiosira pseudonana (12). Steady-state growth of P. tri-cornutum is Fe limited in the range of 10–30 pmol�liter�1 Fe�(Fe� is the sum of all unchelated Fe species) (Table 1), similarto T. oceanica and Pseudonitzschia spp., two open oceandiatoms that are commonly found in the most severely Felimited regions of the world’s oceans (13).
The recently completed genome sequence of P. tricornutum(http://genome.jgi-psf.org/Phatr2/Phatr2.home.html) sheds light onsome important differences between P. tricornutum and T.pseudonana, which may account for their respective thresholds forFe limitation. We evaluated the whole-cell response of P. tricornu-tum to Fe limitation with multiple approaches combining geneexpression profiling and comparative genomics with gas chroma-tography-mass spectroscopy (GC-MS)-aided nontargeted metabo-lomic analysis and a range of physiological measurements. Genes
Author contributions: A.E.A., J.L., P.J.L., and C.B. designed research; A.E.A., J.L., U.M., M.L.,N.S., and G.F. performed research; J.L., P.J.L., G.F., A.R.F., and C.B. contributed newreagents/analytic tools; A.E.A., J.L., U.M., M.L., N.S., G.F., and A.R.F. analyzed data; andA.E.A., J.L., and C.B. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The data reported in this paper have been deposited in the GeneExpression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE8675).
†Present address: J. Craig Venter Institute, 10355 Science Center Drive, San Diego, CA 92121.
§To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0711370105/DCSupplemental.
© 2008 by The National Academy of Sciences of the USA
Table 1. General cellular, physiological, and biochemicalcharacteristics of Fe-limited P. tricornutum cells and cultures
Parameter Fe-limited Fe-repleteFe-limited/Fe-replete
Growth rate 0.18 � 0.05 0.88 � 0.01 0.2 � 0.06Ferric reductase assay,
AU per cell0.717 � 0.03 0.020 � 0.004 35.9 � 7.3
Ferric reductase assay,AU/�m3
0.01 0.0002 50
Carbon fixation,pmol C percell � hour
0.010 � 0.006 0.141 � 0.022 0.07 � 0.04
Cell diameter, �m 2.5 3.5 0.71Cell volume, �m3 60 100 0.60Chl per cell, pg per cell 0.22 0.5 0.44Chl per volume, fg/�m3 3.67 5.0 0.73Fv/Fm 0.18 � 0.1 0.5 � 0.05 0.36 � 0.2PSII 1.2 � 0.14 1 1.2 � 0.14PSI 0.43 � 0.1 1.0 0.43 � 0.10Cytochrome b6f 0.75 � 0.13 1.0 0.75 � 0.13Cytochrome c6 0.7 � 0.05 1.0 0.70 � 0.05NPQ 4.8 � 0.8 3.1 � 0.51 1.5 � 0.4AOX activity 0.6 � 0.15 0.25 � 0.18 2.4 � 1.83
Photosystem and electron carrier concentrations are given in relative unitsnormalized to Fe-replete levels. AOX activity is given as a percentage of totalrespiration (see SI Methods for details). Fe levels in Fe-limited and Fe-repletecultures corresponded to 13.4 pmol�liter�1 Fe� and 2.6 nmol�liter�1 Fe�,respectively.
10438–10443 � PNAS � July 29, 2008 � vol. 105 � no. 30 www.pnas.org�cgi�doi�10.1073�pnas.0711370105

responsive to Fe limitation were identified through a statistically-verified quantitative comparison (14) of 8,669 expressed sequencedtags (ESTs) derived from Fe-limited P. tricornutum cells with104,783 ESTs derived from cells grown in 11 different (all iron-replete) culture conditions (www.biologie.ens.fr/diatomics/EST). Apartial genome microarray and qRT-PCR provided further assess-ment and verification of differential regulation, leading to theidentification of 212 up-regulated and 26 down-regulated genes.The represented acclimation strategies were grouped into threecategories: down-regulation, compensation, and acquisition.
Results and DiscussionDown-Regulation of Photosynthesis. Low Fe supply leads to cellularenergy limitation in P. tricornutum (15) and causes significantchanges in carbon metabolism. Carbon fixation rates per cell were14-fold lower in Fe-limited P. tricornutum cells compared withFe-replete cells (Table 1). This difference remained significant(8-fold) despite normalization to the smaller cell volume observedin Fe-limited conditions [Table 1 and supporting information (SI)Fig. S1]. Reductions in cell volume, chlorophyll (Chl) per cell,photosynthetic efficiency of PSII (Fv/Fm), and content of Fe-richcomplexes and/or electron carriers of the electron transfer chain areall common responses to Fe limitation (Table 1) (15, 16), reflectingcompromised photosystem reaction centers, reduced photosyn-thetic electron transport rates, decreased reductant production, andan inability to process absorbed photons (4, 17, 18).
Severe down-regulation of genes encoding plastid targeted cop-ies of �-carbonic anhydrase (CA) and phosphoribulokinase (PRK),two enzymes supplying substrate for RuBisCO, and a decrease inexpression of a HCO3
� transporter (Fig. 1 and Table S1) suggest that
carbon fluxes into the cell and toward RuBisCO are adjusted tomatch reductant supply. In accordance, Calvin/Benson cycle-related genes downstream of RuBisCO, such as plastid localizedglyceraldehyde-3-phosphate dehydrogenase (GAPDH) and plastidfructose bisphosphate aldolases (FBA) I and II were up-regulatedunder Fe stress (Fig. 1 and Table S1), likely in response toreductions in substrate abundance.
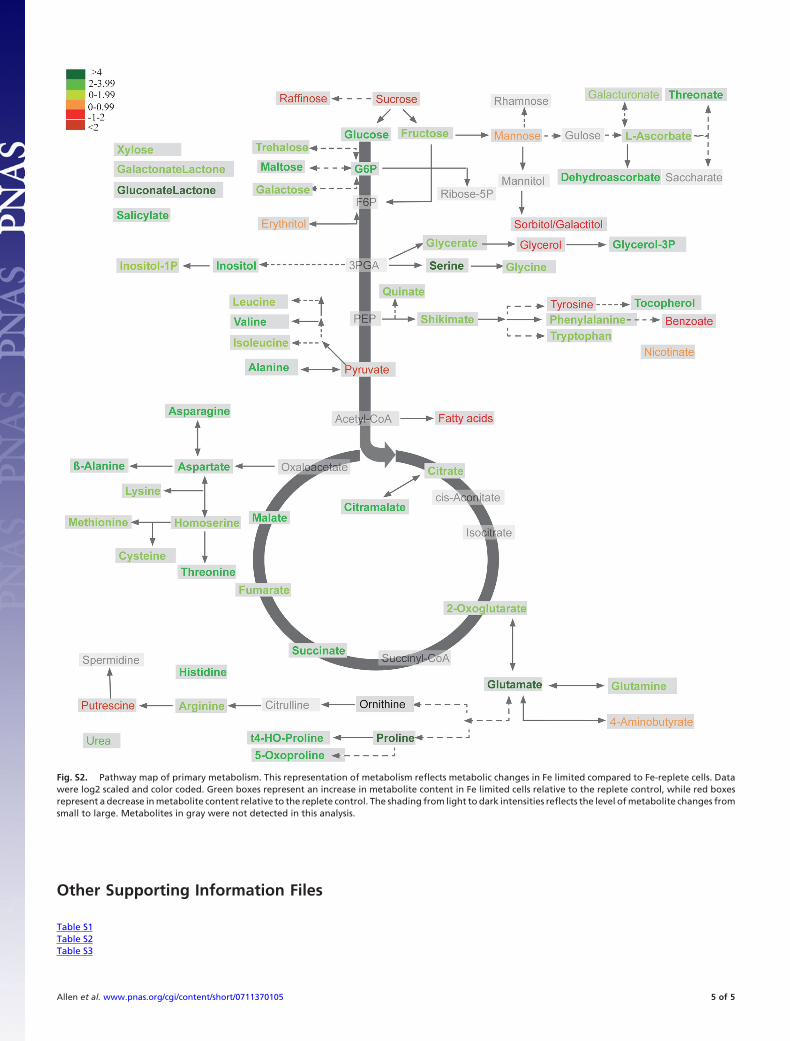
Higher levels of expression of genes encoding galactokinase andendo-1,3-beta-glucanase under Fe limitation (Fig. 1 and Table S1)point to increased degradation of carbohydrates relative to Fe-replete conditions. Significant increases in cellular glucose and inthe intermediate sugars maltose and trehalose (Fig. S2 and TableS2), coupled with the elevated levels of genes encoding the cytosolicenzymes phosphofructokinase, GAPDH, and phosphoglyceratemutase (Fig. 1 and Table S1) are strong indicators that theconversion of polysaccharides to glucose fuels increased glycolyticactivity under Fe limitation (Fig. 1). The altered cell size (Table 1and Fig. S1) and the observed increase in expression of genesputatively encoding polysaccharide-associated cell surface proteins(e.g., integrins, fasciclins and mucins) (Fig. 1 and Table S1) are alsoindicative of the reallocation of carbon from structural molecules.
Chlorophyll Biosynthesis and Pigment Metabolism. Fe limitationinduces chlorosis in photoautotrophs (19), including diatoms (20).Although cellular Chl content decreased by more than half inFe-limited P. tricornutum cells, this difference was much lessstriking when the parallel reduction in cell volume was taken intoaccount (Table 1). However, as observed by the dramatic alter-ations in plastid ultrastructure (Fig. S1), the cell size reduction aloneis not sufficient to account for the changes in physiology observed
Fig. 1. Hypothetical cellular pathways and processes in iron-limited pennate diatom cells. All roman green or red type depicts gene transcripts found to be up- ordown-regulated, respectively. Italicizedandunderlinedgreentype indicatesmetabolites foundtobeenrichedrelative tototalprotein in iron-limitedcells.DF,diffusionfactor; EPS, extracellular polymeric substances; FR, ferric reductase; HMA, heavy metal-associated; PCD, programmed cell death; ROS, reactive oxygen species.
Allen et al. PNAS � July 29, 2008 � vol. 105 � no. 30 � 10439
ECO
LOG
Y

under Fe limitation. Although cellular Chl pools decreased underFe limitation, the expression of 16 genes involved in Chl/hemebiosynthesis pathways were enhanced (Table S1). Conversely, genesencoding proteins that contain heme cofactors, such as thoseinvolved in mitochondrial cytochrome c biogenesis and b5 electrontransport, nitrate reductase, and heme peroxidase, were stronglydown-regulated, indicating that Chl- rather than heme-dependentprocesses were up-regulated.
Glutamate, the precursor for the tetrapyrrole moiety of the Chland heme biosynthesis pathways, was one of the most elevatedmetabolites under Fe limitation (�16-fold) (Table S2). Down-regulation of the gene encoding succinyl-CoA-ligase may accountfor the accumulation of TCA intermediates such as the glutamateprecursor 2-oxoglutarate and thus for glutamate production.
In line with the possibility that high cellular glutamate levels werefueling Chl biosynthesis and the subsequent changes in the chro-mophore composition of pigment-protein complexes, differences inexpression were detected in a variety of other genes involved inpigment metabolism and light harvesting. Genes encoding theenzymes geranylgeranyl reductase, geranylgeranyl pyrophosphatesynthase, phytoene dehydrogenase, and phytoene synthase, whichare involved in the production of carotenoids, were significantlyup-regulated. Geranylgeranyl reductase is also a key regulator ofthe phytol moiety of Chl and is involved in tocopherol production(see below).
Diatoms contain light-harvesting pigment-binding proteins(LHC) that are phylogenetically related to the green/red lineagechlorophyll a/b-binding protein (CAB) family and diatom-specificfucoxanthin-chlorophyll a/c-binding proteins (Fcp). Two genes thatencode the former, one of the latter (FcpD), and a member of thephylogenetically and functionally distinct LI818 family of LHC-likeproteins (21) were significantly up-regulated in Fe-limited condi-tions. Structural modifications to the light-harvesting system andbiochemical adjustment of the Fe stoichiometry of photosystemconstituents under Fe limitation are further supported by the severedown-regulation of genes encoding high light inducible protein(Hlip 2), zeaxanthin epoxidase, photosystem II subunit M (psbM),a ferredoxin subunit, FcpB, and a non-Fcp-type LHC protein. Asexpected, the gene for flavodoxin, a small electron-transfer proteincapable of replacing ferredoxin during Fe deficiency, was signifi-cantly up-regulated (Table S1). The ratio of PSII to PSI complexesand non-photochemical quenching (NPQ) capacity are both ele-vated under Fe deprivation (Table 1).
The onset of Fe deficiency has been shown to induce remodelingof the photosynthetic apparatus in cyanobacteria and green algae;PSI antennas are modified with specialized accessory Chl-proteincomplexes that are thought to play a photoprotective role. Fe stressresponsive LHC gene products, such as CP43� and Tidi, which arefound in cyanobacteria and green algae (22), respectively, functionas peripheral light-harvesting antennas coupled to PSI reactioncenters. Large-scale patterns in open ocean variable fluorescencesuggest that such specialized pigment-protein complexes are typicalin phytoplankton populations found in iron-limited HNLC waters(17). Thus, although levels of Chl a per cell decreased, we proposethat de novo pigment synthesis and changes in the antenna polypep-tide composition may be required to compensate for the relativedecrease of PSI units under periods of Fe limitation (10) and toenhance the NPQ response (Table 1). Although we have not founda direct ortholog of either CP43� or Tidi in diatoms, one of the fourup-regulated Chl binding proteins identified here may fulfill thisrole in P. tricornutum.
Reprogramming of Nitrogen Metabolism. Under optimal growthconditions, nitrate is efficiently used by diatoms. However, under Felimitation genes for nitrate assimilation, such as nitrate reductase(NR), two forms of nitrite reductase, and a plastid-targeted nitritetransporter, were down-regulated, suggesting a reduced capacity forNO3
� assimilation (Fig. 1 and Table S1). This may reflect both the
high Fe costs associated with NO3� assimilation and the reduction
in the supply of organic carbon substrates for amino acid biosyn-thesis associated with the down-regulation of photosynthesis. Tocompensate for the reduced capacity to assimilate nitrate, our datasuggest that amino acids are obtained by reallocation from proteinrecycling and from glycolysis. Supporting this view, the amino acidsisoleucine, leucine, valine, and alanine, which are derived from theglycolytic end product pyruvate, were all significantly elevatedunder low Fe conditions, as was the proteolytically-derived hy-droxyproline (Fig. S2 and Table S2). The apparent increased flowof carbon through glycolysis may therefore be indicative of ademand for amino acids required for the synthesis of a cellularproteome adapted to iron-limited conditions.
In agreement with proteome remodeling, genes encoding pro-teases or proteins linked to proteolytic processes (a proteasomesubunit, a metalloprotease, a cathepsin/cysteine protease, a ubiq-uitin ligase, and two PDZ-domain proteases) were strongly inducedunder Fe limitation. In addition, nine heat shock protein (HSP)genes were preferentially accumulated in response to reduced Felevels, including an HSP containing a Ca2�-binding EF-handdomain (Table S1) (see below).
Multiple Responses to Oxidative Stress. Accumulation of reactiveoxygen species (ROS) under Fe limitation has been suggested tostem from inefficient activity of an Fe-impoverished photosyntheticelectron transport chain, which results in an increased quantity ofelectrons accepted by O2 (23). Typical ROS defense proteins, suchas heme peroxidase, superoxide dismutase (SOD), and aconitase,which is also a key component of the TCA cycle, require iron as acofactor and are of limited value in iron-limited conditions (24), asrevealed by the down-regulation of the genes encoding theseproteins under Fe limitation (Fig. 1 and Table S1). Our resultssuggest alternate biochemical pathways that may compensate forthe decrease in conventional, Fe-rich ROS scavenging machinery.Notably, tocopherol, a powerful antioxidant, increased 5-fold underFe limitation (Table S2), whereas tyrosine, a precursor of tocoph-erol, was reduced twofold. Tryptophan synthase transcript levels,and shikimate, a metabolite involved in the synthesis of aromaticamino acids, were increased. Fe limitation provoked a 12-foldincrease in the glycolytic derivative dehydroascorbate (Table S2),which is also a strong antioxidant, and transcripts encoding a2-phosphoglycolate phosphatase (GPH), involved in repairing aclass of DNA lesions induced by oxidative stress (25), were alsoelevated (Table S1).
Photorespiration is thought to play a role in excess energydissipation under stress conditions, such as high light and lowtemperature (26), and may have a similar function in Fe-limiteddiatoms. Combined with the strong down-regulation of �-plastidcarbonic anhydrase, the large increases observed in the photo-respiratory metabolic intermediates serine, glycine, and glycerate(Fig. S2 and Table S2) may suggest an increased role for photo-respiration under Fe limitation.
Respiratory electron transfer chain activity, one of the mostFe-demanding cellular processes, is restricted during Fe limitation(27). In P. tricornutum, genes encoding mitochondria-targetedproteins involved in cytochrome-based respiration were signifi-cantly down-regulated. A cytochrome pathway restriction wouldpromote ROS accumulation in mitochondria, and our resultsindicate that Fe-limited diatoms may employ at least three mech-anisms effective in reducing mitochondrial ROS production andfacilitating respiratory processes despite impairment of cyto-chrome-based respiration (Fig. 1).
Mitochondrial alternative oxidase (AOX) activity and mRNAlevels were significantly higher under Fe starvation (Table 1 andTable S1). AOX can lower mitochondrial reactive oxygen produc-tion in plant cells and may have a regulatory role in balancingcarbon metabolism, including glycolysis and TCA cycle activity,when the cytochrome pathway is compromised (28).
10440 � www.pnas.org�cgi�doi�10.1073�pnas.0711370105 Allen et al.

Second, the exchange of reducing equivalents between themitochondria and cytosol may be achieved via an alternativeNADH dehydrogenase (NDH)-based shuttle system. NDH, acritical site for superoxide production, is thought to be particularlyimportant during times of intensified glycine and serine metabo-lism. Under Fe limitation, the glycolytic derivative myo-inositol andmRNA levels of a gene encoding mitochondrial myo-inositol de-hydrogenase were both elevated 6-fold (Table S1 and Table S2).The distribution of myo-inositol dehydrogenase among algal classesand the fact that red algal mitochondria are able to use myo-inositolfor respiration suggest that red algae and chromalveolates have theability to use an inositol/inosase shuttle system for reducing equiv-alents (29) in addition to the NDH system common in the greenlineage. The importance of mitigating ROS production and theapparent intensified flux of the photorespiratory metabolites gly-cine and serine under Fe stress could lead to the utilization of AOXand myo-inositol dehydrogenase as respiratory alternatives.
Finally, photosynthetic electrons can be directed into the mito-chondrial electron transport chain through the malate:aspartateshuttle, where AOX functions as a sink for excess reducing equiv-alents (30). Increased levels of malate and AOX activity mayindicate that this shunt is used to transfer excess electrons from thechloroplast toward AOX, further reducing ROS-mediated insultand at the same time generating ATP (Fig. 1).
Intracellular Signaling. The importance of calcium (Ca2�)-basedsecond messenger signaling pathways in P. tricornutum has beenshown for nitric oxide (NO)-mediated death and defense responsesand for cellular perception of bioavailable Fe concentrations (31,32). Diatom AOX proteins contain calcium-binding EF-hand do-mains. This suggests that AOX activation in diatoms is partiallycontrolled by calcium-based signaling mechanisms. One proteinpossibly involved in Ca2�-activated ROS defense is the oxidativestress responsive protein annexin (33). mRNA levels for annexinwere significantly up-regulated in P. tricornutum under Fe limita-tion (Fig. 1 and Table S1). The importance of Ca2� signaling underFe limitation was further reinforced by the up-regulation of Ca2�
responsive genes encoding an HSP and proteins, such as myosin,gelsolin, copine, and scramblase (Fig. 1 and Table S1), involved incytoskeleton and cell membrane regulation and modification.
Metal Assimilation and Homeostasis. Fe-limited P. tricornutum cellsshowed a 20-fold increase in levels of the strong iron chelatingglucose oxidation derivative gluconate (Table S2). Internal chela-tors are important for the intracellular control of Fe, permittingefficient Fe utilization particularly under Fe limitation, and foralleviating oxidative DNA lesions, which may occur through leach-ing of Fe from storage proteins and enzymatic clusters ([4Fe–4S])(34). In marine Synechococcus sp., IscA has recently emergedas a major component in regulating cellular Fe homeostasis anddelivery of iron from organic ligands and chelators to iron–sulfurassembly cluster proteins under conditions of depleted cellular ironpools (35). The observed increase in IscA mRNA in response to Felimitation (Table S1) suggests a similar role in P. tricornutum.Although very little is known about the intracellular handling ofiron in eukaryotes, IscA and gluconate may be important compo-nents for controlling the intracellular availability of Fe duringperiods of limitation. Ferritin is present in the P. tricornutumgenome sequence but absent from the T. pseudonana genome. Thisprompts speculation that fundamental differences in Fe storagecapacity and turnover exist between these two diatom species andmay partly explain the ability of P. tricornutum to grow at lower Feconcentrations than T. pseudonana.
Physiological and molecular evidence from T. pseudonana and P.tricornutum support an important role for high affinity cell surfaceferric-chelate reductases in diatom iron uptake systems (8). P.tricornutum and T. pseudonana each contain two such transmem-brane oxidoreductases (FRE1 and FRE2) with FAD and NADPH-
ribose binding signatures and requisite histidine residues for hemecoordination (12). In P. tricornutum, three additional Fe-responsivegenes encoding cell surface transmembrane proteins may be in-volved in iron uptake. Two DcytB-type ferric reductases belongingto the cytochrome b561 family, FRE3 and FRE4, contain five to sixtransmembrane domains each. Furthermore, an opisthokont-typedecycling heme oxygenase (Table S3) containing two TM domains,if localized to the cell surface, may help destabilize the Fe bound inporphyrins, thus corroborating the report in ref. 36 of heme-basedFe acquisition systems in diatoms. Four gene transcripts encodingproteins for various metal transporters were enriched under Felimitation (Table S1). These included one member of the ZRT,IRT-like protein family (ZIP), one heavy metal ATPase (HMA),and two, putatively organelle targeted, cation diffusion facilitators(CDF). We also observed down-regulation of a gene that encodesthe bacterial chromate efflux protein ChrA (Fig. 1 and Table S1).
Iron-Regulated Gene Clusters. The P. tricornutum genome containsat least three clusters of Fe-responsive genes (Fig. 2). Genes withinthese clusters were among the most highly expressed under Felimitation and, with the exception of ferric reductase (FRE2), areabsent in T. pseudonana. FRE2 is immediately adjacent to a highlyFe-responsive gene encoding a putative ferrichrome-binding pro-tein (FBP1), similar to FhuD found in prokaryotic ferrichromeABC transport systems. In bacteria, FhuD enables the directtransport of Fe(III)-hydroxymate chelates across the plasma mem-brane. The identification of a diatom-type FBP1 is consistent withkinetic measurements of siderophore uptake in P. tricornutum (37)and supports the presence of a coupled FBP1/FRE-dependentsiderophore-based system for the destabilization and subsequentreduction of organically complexed Fe(III)-hydoxymates at the cellsurface. Although standard assays (38) failed to detect the produc-tion of siderophores in P. tricornutum (data not shown), we dis-covered the genetic basis for Fe-uptake mechanisms that are acombination of classical eukaryotic components and those com-monly found in bacteria but not usually present in eukaryotes.
A second Fe-regulated cluster contained genes encoding acopper(Cu)-tyrosinase (TYR1) and a protein of unknown function,ISIP1 (Fig. 2 and Table S1). The role of Cu-nutrition in Fe-limitedphytoplankton has become increasingly apparent (39, 40). Cu-tyrosinase represents an undescribed Fe-sensitive Cu-protein. TheISIP1 encoding gene was the most highly expressed gene on themicroarray of Fe-limited cRNA target and was the most commonlysequenced transcript in the Fe-limited EST library. An orthologous
Fig. 2. Iron-regulated gene clusters in P. tricornutum. With the exception ofFRE2, the iron-regulated cluster genes are absent from the T. pseudonanagenome. Putative components of the Fe uptake system are shown in brown.The number of ESTs retrieved from the Fe limited EST library and the R statisticare provided. R � 12 indicates a statistically significant enrichment of ESTs inthe Fe limited library relative to 11 other Fe-replete EST libraries (14). qRT-PCRdata are given in Table S1.
Allen et al. PNAS � July 29, 2008 � vol. 105 � no. 30 � 10441
ECO
LOG
Y
Fe-responsive gene in Dunaliella salina encodes a protein thoughtto associate with plasma membrane transferrins where it functions,along with a multicopper ferroxidase, as part of a complex thatenhances binding and uptake of ferric ions (41). However, there isno evidence for the occurrence of genes that encode transferrin inthe genome of P. tricornutum.
A third cluster of Fe-regulated genes contains five genes ofunknown function with the exception of a gene encoding anortholog of the cellular repressor of E1A-stimulated genes(CREG). CREG is a secreted glycoprotein that acts as a regulatoryligand for signaling kinases and reduces cell proliferation. InFe-deficient P. tricornutum cells, CREG could be central in con-fining cell growth to match resource availability. Each of the ninegenes found within the Fe-regulated clusters are predicted toencode signal peptides (Fig. 2 and Table S1), suggesting a role forthe gene products in cell surface processes.
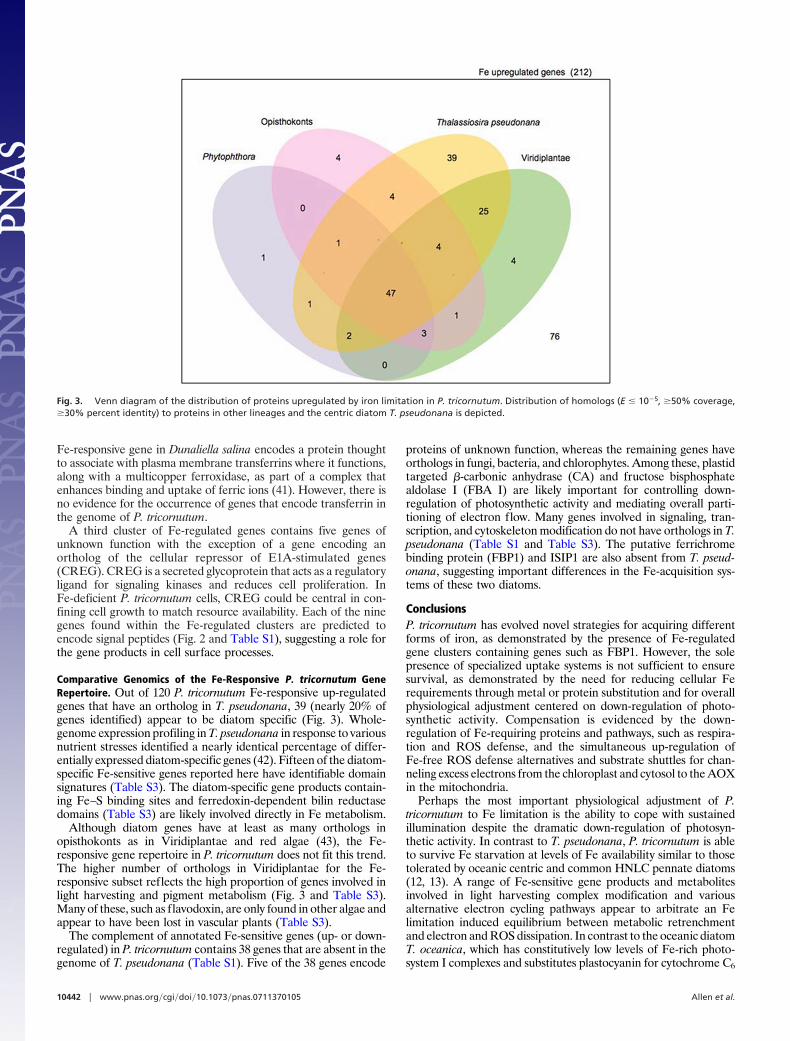
Comparative Genomics of the Fe-Responsive P. tricornutum GeneRepertoire. Out of 120 P. tricornutum Fe-responsive up-regulatedgenes that have an ortholog in T. pseudonana, 39 (nearly 20% ofgenes identified) appear to be diatom specific (Fig. 3). Whole-genome expression profiling in T. pseudonana in response to variousnutrient stresses identified a nearly identical percentage of differ-entially expressed diatom-specific genes (42). Fifteen of the diatom-specific Fe-sensitive genes reported here have identifiable domainsignatures (Table S3). The diatom-specific gene products contain-ing Fe–S binding sites and ferredoxin-dependent bilin reductasedomains (Table S3) are likely involved directly in Fe metabolism.
Although diatom genes have at least as many orthologs inopisthokonts as in Viridiplantae and red algae (43), the Fe-responsive gene repertoire in P. tricornutum does not fit this trend.The higher number of orthologs in Viridiplantae for the Fe-responsive subset reflects the high proportion of genes involved inlight harvesting and pigment metabolism (Fig. 3 and Table S3).Many of these, such as flavodoxin, are only found in other algae andappear to have been lost in vascular plants (Table S3).
The complement of annotated Fe-sensitive genes (up- or down-regulated) in P. tricornutum contains 38 genes that are absent in thegenome of T. pseudonana (Table S1). Five of the 38 genes encode
proteins of unknown function, whereas the remaining genes haveorthologs in fungi, bacteria, and chlorophytes. Among these, plastidtargeted �-carbonic anhydrase (CA) and fructose bisphosphatealdolase I (FBA I) are likely important for controlling down-regulation of photosynthetic activity and mediating overall parti-tioning of electron flow. Many genes involved in signaling, tran-scription, and cytoskeleton modification do not have orthologs in T.pseudonana (Table S1 and Table S3). The putative ferrichromebinding protein (FBP1) and ISIP1 are also absent from T. pseud-onana, suggesting important differences in the Fe-acquisition sys-tems of these two diatoms.
ConclusionsP. tricornutum has evolved novel strategies for acquiring differentforms of iron, as demonstrated by the presence of Fe-regulatedgene clusters containing genes such as FBP1. However, the solepresence of specialized uptake systems is not sufficient to ensuresurvival, as demonstrated by the need for reducing cellular Ferequirements through metal or protein substitution and for overallphysiological adjustment centered on down-regulation of photo-synthetic activity. Compensation is evidenced by the down-regulation of Fe-requiring proteins and pathways, such as respira-tion and ROS defense, and the simultaneous up-regulation ofFe-free ROS defense alternatives and substrate shuttles for chan-neling excess electrons from the chloroplast and cytosol to the AOXin the mitochondria.
Perhaps the most important physiological adjustment of P.tricornutum to Fe limitation is the ability to cope with sustainedillumination despite the dramatic down-regulation of photosyn-thetic activity. In contrast to T. pseudonana, P. tricornutum is ableto survive Fe starvation at levels of Fe availability similar to thosetolerated by oceanic centric and common HNLC pennate diatoms(12, 13). A range of Fe-sensitive gene products and metabolitesinvolved in light harvesting complex modification and variousalternative electron cycling pathways appear to arbitrate an Felimitation induced equilibrium between metabolic retrenchmentand electron and ROS dissipation. In contrast to the oceanic diatomT. oceanica, which has constitutively low levels of Fe-rich photo-system I complexes and substitutes plastocyanin for cytochrome C6
Fig. 3. Venn diagram of the distribution of proteins upregulated by iron limitation in P. tricornutum. Distribution of homologs (E � 10�5, �50% coverage,�30% percent identity) to proteins in other lineages and the centric diatom T. pseudonana is depicted.
10442 � www.pnas.org�cgi�doi�10.1073�pnas.0711370105 Allen et al.

in Fe limiting conditions (10, 39), P. tricornutum appears to usemetabolic reconfigurations to acclimate to low Fe levels. In con-clusion, the cellular strategies identified in P. tricornutum provide afoundation for understanding the constraints on diatom metabo-lism in iron-limited regions of today’s oceans and provide a basis forunderstanding how diatom growth will be affected by nutrientfertilization strategies to enhance CO2 sequestration from theatmosphere.
MethodsGrowth Conditions and Physiological Parameters. Semicontinuous batch cul-tures of Phaeodactylum tricornutum (CCMP2561) were grown in f/2 ASW me-dium modified to contain either 13.4 pmol�liters�1 Fe� (Fe-limited) or 2.6nmol�liters�1 Fe�. Fe� is the sum of all unchelated Fe species. The degree of ironlimitation in steady-state cultures was assessed from various physiological pa-rameters and cellular indicators (Table 1 and Fig. S1) and by following therecovery of high growth rates upon Fe addition to a small aliquot of the cultureat the time of harvest.
Metabolite Extraction and GC-MS Analysis. Approximately 1 � 109 Fe-limited andFe-replete cells were pelleted and used for metabolite analysis. Metabolites wereextracted and derivatization and GC-MS analysis were carried out as described inref. 44. Metabolites were identified compared with database entries of authenticMSRI libraries (45).
Gene Expression. Genes responsive to Fe limitation were evaluated through acombination of statistical analysis of EST libraries, a partial genome microarray,
and qRT-PCR. In the case of EST library analysis, a log-likelihood ratio test statistic,designed specifically for the purpose of statistically evaluating gene expressionlevel across multiple cDNA libraries, was computed (14). The array platform withall of the probe sequences and corresponding genome browser protein IDs andall of the normalized hybridization data are available in Minimum InformationAbout a Microarray Experiment (MIAME) compliant format at the NationalCenter for Biotechnology Information Gene Expression Omnibus (GEO).
Annotation and Comparative Analysis. Two hundred twenty-eight Fe-responsiveEST contigs were mapped to 212 predicted proteins on the P. tricornutumgenome browser. The 212 predicted proteins corresponding to the up-regulatedtranscripts were used for the further comparative analysis against differenttaxonomical lineages (see SI Methods).
ACKNOWLEDGMENTS. We thank Mathangi Thiagarajan for assistance withstatistical analysis of microarray data and Guangzuo Luo for microarray devel-opment. This work was supported by National Science Foundation postdoctoralfellowship in microbial biology DBI-0301626 and Biological Oceanography grantOCE-0727997 (to A.E.A.), Deutches Forschungs Gemeinschaft Grant RO-2138/7-1(awarded to J.L.R. and supporting M.L.), European Union-funded FP6 DiatomicsProject Grant LSHG-CT-2004-512035, EU-FP6 Marine Genomics Network of Excel-lence Grant GOCE-CT-2004-505403, an ATIP ‘‘Blanche’’ grant from Centre Na-tional de la Recherche Scientifique, and the Agence Nationale de la Recherche(France). Diatom genome sequencing at the Joint Genome Institute (WalnutCreek, CA) was performed under the auspices of the U.S. Department of Energy’sOffice of Science, Biological and Environmental Research Program, and by theUniversity of California, Lawrence Berkeley National Laboratory under ContractDE-AC02-05CH11231, Lawrence Livermore National Laboratory under ContractDE-AC52-07NA27344, and Los Alamos National Laboratory under Contract DE-AC02-06NA25396.P. tricornutumESTsweregeneratedatGenoscope(Evry,Paris).
1. Anbar AD, Knoll AH (2002) Proterozoic ocean chemistry and evolution: A bioinorganicbridge? Science 297:1137–1142.
2. Quigg A, et al. (2003) The evolutionary inheritance of elemental stoichiometry inmarine phytoplankton. Nature 425:291–294.
3. Archer DE, Johnson K (2000) A model of the iron cycle in the ocean. Global BiogeochemCy 14:269–279.
4. Behrenfeld MJ, Bale AJ, Kolber ZS, Aiken J, Falkowski PG (1996) Confirmation of ironlimitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature383:508–511.
5. de Baar HJW, et al. (2005) Synthesis of iron fertilization experiments: From the iron agein the age of enlightenment. J Geophys Res Oceans 110:C09S16.
6. Moore C, Hickman A, Poulton A, Seeyave S, Lucas M (2007) Iron-light interactionsduring the CROZet natural iron bloom and EXport experiment (CROZEX): II - Taxonimcresponses and elemental stoichiometry. Deep-Sea Res Pt II 54:2066–2084.
7. Boyd PW, et al. (2007) Mesoscale iron enrichment experiments 1993–2005: Synthesisand future directions. Science 315:612–617.
8. Brand LE, Sunda WG, Guillard RL (1983) Limitation of marine primary production ratesby zinc, manganese, and iron. Limnol Oceanogr 28:1182–1198.
9. McKay RML, Geider RJ, LaRoche J (1997) Physiological and biochemical response of thephotosynthetic apparatus of two marine diatoms to Fe stress. Plant Physiol 114:615–622.
10. Strzepek RF, Harrison PJ (2004) Photosynthetic architecture differs in coastal andoceanic diatoms. Nature 431:689–692.
11. Sunda WG, Huntsman SA (1995) Iron uptake and growth limitation in oceanic andcoastal phytoplankton. Mar Chem 50:189–206.
12. Kustka A, Allen AE, Morel FMM (2007) Sequence analysis and transcriptional regula-tion of Fe acquisition genes in two marine diatoms. J Phycol 43:715–729.
13. Marchetti A, Maldonado MT, Lane ES, Harrison PJ (2006) Iron requirements of thepennate diatom Pseudo-nitzschia: Comparison of oceanic (high-nitrate, low-chlorophyll waters) and coastal species. Limnol Oceanogr 51:2092–2101.
14. Stekel DJ, Git Y, Falciani F (2000) The comparison of gene expression from multiplecDNA libraries. Genome Res 10:2055–2061.
15. Greene RM, Geider RJ, Kolber Z, Falkowski PG (1992) Iron-Induced changes in lightharvesting and photochemical energy conversion processes in eukaryotic marinealgae. Plant Physiol 100:565–575.
16. Geider RJ, LaRoche J, Greene RM, Olaizola M (1993) Response of the photosyntheticapparatus of. Phaeodactylum tricornutum (BACILLARIOPHYCEAE) to nitrate, phos-phate, or iron starvation. J Phycol 29:755–766.
17. Behrenfeld MJ, et al. (2006) Controls on tropical Pacific Ocean productivity revealedthrough nutrient stress diagnostics. Nature 442:1025–1028.
18. Milligan AJ, Harrison PJ (2000) Effects of non-steady-state iron limitation on nitrogenassimilatory enzymes in the marine diatom Thalassiosira weissflogii (Bacillari-ophyceae). J Phycol 36:78–86.
19. Moseley JL, et al. (2002) Adaptation to Fe-deficiency requires remodeling of thephotosynthetic apparatus. EMBO J 21:6709–6720.
20. Greene RM, Geider RJ, Falkowski PG (1991) Effect of iron limitation on photosynthesisin a marine diatom. Limnol Oceanogr 36:1772–1782.
21. Koziol AG, et al. (2007) Tracing the evolution of the light harvesting antennae inchorophyll. a/b-containing organisms Plant Physiol 143:1802–1816.
22. Varsano T, Wolf S, Pick U (2006) A chlorophyll. a/b-binding protein homolog that isinduced by iron deficiency is associated with enlarged photosystem I units in theeukaryotic alga Dunaliella salina. J Biol Chem 281:10305–10315.
23. Niyogi KK (1999) Photoprotection revisited: Genetic and molecular approaches. AnnuRev Plant Physiol Plant Mol Biol 50:333–359.
24. Baxter CJ, et al. (2007) The metabolic eesponse of heterotrophic Arabidopsis cells tooxidative stress. Plant Physiol 143:312–325.
25. Pellicer MT, Nunez MF, Aguilar J, Badia J, Baldoma L (2003) Role of 2-phosphoglycolatephosphatase of Escherichia coli in metabolism of the 2-phosphoglycolate formed inDNA repair. J Bacteriol 185:5815–5821.
26. Parker MS, Armbrust EV (2005) Synergistic effects of light, temperature, and nitrogensource on transcription of genes for carbon and nitrogen metabolism in the centricdiatom Thalassiosira pseudonana (Bacillariophyceae). J Phycol 41:1142–1153.
27. Kudo I, Miyamoto M, Noiri Y, Maita Y (2000) Combined effects of temperature and ironon the growth and physiology of the marine diatom, Phaeodactylum tricornutum(Bacillariophyceae). J Phycol 36:1096–1102.
28. Fernie AR, Carrari F, Sweetlove LJ (2004) Respiratory metabolism: Glycolysis, the TCAcycle and mitochondrial electron transport. Curr Opin Plant Biol 7:254–261.
29. Gross W, Meyer A (2003) Distribution of myo-inositol dehydrogenase in algae. Eur JPhycol 38:191–194.
30. Yoshida K, Terashima I, Noguchi K (2007) Up-regulation of mitochondrial alternativeoxidase concomitant with chloroplast over-reduction by excess light. Plant Cell Physiol48:606–614.
31. Falciatore A, d’Alcala, M. R., Croot P, Bowler C (2000) Perception of environmentalsignal by a marine diatom. Science 288:2363–2366.
32. Vardi A, et al. (2006) A stress surveillance system based on calcium and nitric oxide inmarine diatoms. PLoS Biol 4:411–419.
33. Rhee NJ, Kim GY, Huh JW, Kim SW, Na DS (2000) Annexin I is a stress protein inducedby heat, oxidative stress and a sulfhydryl-reactive agent. Eur J Biochem 267:3220–3225.
34. Keyer K, Imlay JA (1996) Superoxide accelerates DNA damage by elevating free-ironlevels. Proc Natl Acad Sci USA 93:13635–13640.
35. Balasubramanian R, Shen G, Bryant DA, Golbeck JH (2006) Regulatory roles for IscA andSufA in iron homeostasis and redox stress responses in the cyanobacterium Synecho-coccus sp. strain PCC 7002. J Bacteriol 188:3182–3191.
36. Hutchins DA, Witter AE, Butler A, Luther GW (1999) Competition among marinephytoplankton for different chelated iron species. Nature 400:858–861.
37. Soria-Dengg S, Horstmann U (1995) Ferrioxamines B and E as iron sources for themarine diatom Phaeodactylum tricornutum. Mar Ecol Prog Ser 127:269–277.
38. Schwyn B, Neilands J (1987) UNiversal chemical assay for the detection and determi-nation of siderophores. Analyt Biochem 160:47–56.
39. Peers G, Price NM (2006) Copper-containing plastocyanin used for electron transportby an oceanic diatom. Nature 441:341–344.
40. Maldonado MT, et al. (2006) Copper-dependent iron transport in coastal and oceanicdiatoms. Limnol Oceanogr 51:1729–1745.
41. Paz Y, Katz A, Pick U (2007) A multicopper ferroxidase involved in iron binding totransferrins in Dunaliella salina plasma membranes. J Biol Chem 282:8658–8666.
42. Mock T, et al. (2008) Whole-genome expression profiling of the marine diatomThalassiosira pseudonana identifies genes involved in silicon bioprocesses. Proc NatlAcad Sci USA 105:1579–1584.
43. Armbrust EV, et al. (2004) The genome of the diatom Thalassiosira pseudonana:Ecology, evolution, and metabolism. Science 306:79–86.
44. Lisec J, Schauer N, Kopka J, Willmitzer L, Fernie AR (2006) Gas chromatography massspectrometry-based metabolite profiling in plants. Nat Protoc 1:387–396.
45. Schauer N, et al. (2005) GC-MS librraies for the rapid indentification of metabolites incomplex biological samples. FEBS Lett 579:1332–1337.
46. Maheswari U, et al. (2005) The diatom EST database. Nucleic Acids Res 33:344–347.
Allen et al. PNAS � July 29, 2008 � vol. 105 � no. 30 � 10443
ECO
LOG
Y

Supporting InformationAllen et al. 10.1073/pnas.0711370105SI MethodsCulture and Growth Conditions. For the construction of the cDNAlibrary from which ESTs were generated, RNA was extractedfrom cultures acclimated to 5 nM total Fe for several generationsin 1-liter polycarbonate bottles and finally grown in an 8-literbottle to mid-exponential phase at a temperature of 18°C and a16:8 light (150 �E�m�1�sec�1):dark regime. Additional physio-logical parameters were measured in parallel for 1L Fe-limitedand Fe-replete cultures. These included growth rate, cell size,carbon fixation, chlorophyll, f lavodoxin (1), ferric reductaseactivity (2), nonphotochemical quenching (3), respiration andalternative oxidase activity, and spectroscopic determination ofphotosystem and electron carrier concentrations. Fe�, the sum ofall unchelated species, calculated according to the equations of(4) was (Fe� � 0.026 FeT) under our conditions. Therefore, theFe limiting condition had an Fe� level of 13.4 pmol�liters�1 Fe�and the Fe replete condition had an Fe� value of 2.6 nmol�liters�1
Fe�. Cells were grown in semicontinuous batch mode and keptin steady-state with respect to Fe availability. To achieve andmaintain desired Fe levels, Fe was pre-complexed with EDTA(1:1.1 mol:mol) and added to Fe-free media. Thus, the avail-ability of free Fe ions in the culture media was effectivelybuffered to maintain consistent Fe� levels over the course of theexperiments. The concentration of kinetically labile inorganicspecies is known to be proportional to the free ion concentration(5, 6).
Chlorophyll Measurements. Ten to 50 ml of culture were filtered on0.2 �m GF/F filters, chlorophyll was extracted in 10 ml of 90%acetone, using 1:1 2 mm/4 mm glass beads. Chlorophyll concen-tration was measured with a Turner fluorometer.
Variable Fluorescence. Variable fluorescence Fv/Fm was mea-sured after 5 min of dark adaptation with a PhytoPAM(WALTZ) and analyzed with PhytoWin software (7).
Nonphotochemical Quenching (NPQ). NPQ was calculated as (Fm �Fm�)/Fm�, where Fm is the maximum fluorescence yield (i.e., indark adapted samples) in the dark, and Fm� is the maximumfluorescence emission in steady state light conditions. Fm andFm� were determined by exposure of the cell suspension to asaturating light pulse (intensity: 3,000 �E; duration: 150 ms,optical pathway of the cuvette: 1 mm). Actinic light was 1,000 �E.Measurements were performed with a laboratory-built f luoro-meter, as described in ref. 3.
Spectroscopy. Spectroscopic measurements were performed witha LED-based spectrophotometer having a time resolution of 10�s (JTS 10; Biologic). PSI and PSII ratios were calculated fromthe extent of the ECS signal at 520–490 nm upon excitation ofcells with a single turnover saturating flash. This signal isproportional to reaction center photochemistry (8). Thus, thegeneration of a transmembrane potential following a singleturnover flash reflects the turnover of the different photosyn-thetic complexes. The fast phase, completed in �1 �s (phase a)is proportional to the charge separation performed by PSI andPSII. The slower rising signal is attributed to cytochrome b6fturnover, whereas the slow relaxation phase is linked to H� f luxthrough the ATP synthase (9). PSII contribution was deduced asthe difference between the signals measured in the absence andpresence of the PSII inhibitors DCMU and HA (10). The lattercompound was added to destroy the manganese cluster respon-
sible for oxygen evolution and to prevent recombination betweenthe donor and acceptor side of PSII, which would precludecorrect estimation of the PSI/PSII ratio.
The same setup was used to assess the amount of functionalcytochrome c6 and b6f complexes. Cyt c6/P700 ratios wereassessed by comparing the extent of c6 oxidation in continuouslight in the presence of the plastoquinone analogue DBMIB.This compound blocks cyt b6f turnover, allowing full oxidationof the cytochrome c6 pool. This amplitude was normalised on thesignal corresponding to oxidation of P700 under the sameconditions. Cytochrome b6f complex was estimated from thesignal corresponding to the maximum oxidation of cytochromeb6. Cytochrome c6 redox changes were calculated as the differ-ence between the absorption at 554 nm and a baseline drawnbetween 545 and 573 nm. Cytochrome b6 signals at 564 minus thesame baseline. P700 was computed as �I/I P700 � �I/I 820 nm �0.8 � �I/I 870 nm (11). An extinction coefficient of 17.4mM�cm�1 (12) was used to evaluate PSI concentration startingfrom P700 redox change signals.
Cell Volume. Cell volumes of fusiform cells were calculated frommean length and diameter measured under the light microscope,using a double frustum as a simplified model of the fusiform cell.
RNA Extraction for qRT-PCR Analysis. 2–4 � 107 cells were harvestedby 2 � 10 min centrifugations at 4°C. Cell pellets were used forextraction of total RNA, using a Plant RNeasy Kit (Qiagen).RNA was eluted from the columns with 50 �l of RNase-freewater and stored at �20°C. cDNA was synthesized with aQuantitec reverse transcription kit, after treatment with theg-DNA whipeout buffer to remove contaminating genomicDNA. qRTPCR was performed with an ABI 700 instrument,and ABI SyBr Green master mix in 25 �l of total reaction volumecontaining 5 �l of RNA sample, and 10 pmol of each ofgene-specific forward and reverse primers, cycled according tothe manufacturers default program of 10 min at 95°C, followedby 40 cycles between 15 seconds at 95°C and 1 min at 60°Cfollowed by a dissociation curve measurement for each gene-specific primer pair. The efficiency of amplification for eachgene was calculated from a 10-fold dilution series of templatecDNA, and was �95%. RNA and cDNA samples were run inparallel assays with 18S rRNA primers to confirm that the RNAsamples were free of contaminating genomic DNA. Expressionlevels for target genes in each culture were normalized to 18SrRNA expression.
All pairs of qPCR primers are given in Table S1.
14C-Fixation. 14C fixation experiments were carried out with cellsfrom late exponential phase supplied with 1 �M or 5 nM Fe,respectively. Cell densities were 930,000 (1 �M at 5 days) and385,000 (5 nM at 8 days) cells per ml. Culture (40 ml) wasincubated for 7 h at 150 �E with 2 �Curie of NaH14CO3. Culture(8 ml) was filtered on cellulose acetate filters in triplicates, filterswere put in vials and supplied with 5 ml of Lumagel. Vials wereincubated for 1 day and activity was measured with a scintillationcounter.
Dark Respiration and Oxygen Evolution. Respiration and O2-evolution were measured with a Clark Pt/Ag-electrode (Han-saTech). Cells were concentrated 50-fold to 5 � 107 cells perml, resuspended in fresh medium and measured at 20°C. Res-piration was measured with dark adapted cells, and photosyn-
Allen et al. www.pnas.org/cgi/content/short/0711370105 1 of 5

thetic O2-evolution under illumination of 150 �E. The photo-synthetic quotient (moles of O2 evolved per moles of CO2assimilated) for Fe replete and limited cultures was consistentwith theoretical values.
Alternative Oxidase (AOX) Assay. AOX activity was estimated fromrespiration measurements in which the cytochrome inhibitorantimycin A (AA) (1 �M) or the AOX inhibitor salicyl hydrox-amic acid (SHAM) (1 mM) were added independently todifferent sub-samples of the same culture. Subsequently, bothinhibitors were added to the same sample to verify the same levelof inhibition.
Metabolite Extraction and GC-MS Analysis. Approximately 1 � 109
cells were pelleted and used for metabolite analysis. Metaboliteswere extracted by the addition of 600 �L methanol and 15 �L (2mg/ml) ribitol, shock freezing in liquid nitrogen, subsequentvortexing and shock freezing. This was followed by the additionof 250 �L dH2O and 250 �L CHCl3, incubation of the samplesat 70°C for 10 min, and centrifugation for 10 min at 14,000 rpm.50 �L of the supernatant was transferred into a new reactiontube and dried for GC-MS analysis. Derivatization and GC-MSanalysis were carried out as described in ref. 13. The GC-MSsystem comprised a CTC CombiPAL autosampler, an Agilent6890N gas chromatograph and a LECO Pegasus III TOF-MSrunning in EI� mode. Metabolites were identified comparedwith database entries of authentic MSRI libraries (14). Meta-bolites were normalized internally to the added standard Ribitoland quantified relative to total cellular protein levels (15).Recovery and recombination experiments confirmed the validityof this protocol for both qualitative and quantitative analyses.
EST Library Construction, Sequencing, Annotation, and ComparativeAnalysis. cDNA libraries were constructed from poly(A)� RNA,using the CloneMiner cDNA library construction Kit (Invitro-gen) following the supplier’s instructions. Sequencing was per-formed mostly from the 5� end of the insert but for some of thelibraries, an attempt was made to sequence each clone at both the5� and the 3� ends. When both EST reads overlapped, the twosequences were fused into a consensus sequence, using PHRAP(www.phrap.org). Approximately 110,000 ESTs were sequencedfrom cells grown in 12 different growth conditions. ESTs werechecked for ambiguities, vector trimmed, and quality checkedand clustered by using CAP3 software. Approximately 8,000contigs and 5,000 singletons were obtained by this method. Thefrequency of expressed genes under different conditions wascalculated for each contig. In cases where 5� and 3� sequencingwas performed for the same clone, the EST was only countedonce for overall frequency computation. The log-likelihood ratio(R value) was computed to test for statistical significance ofdifferential gene expression by comparing to randomized data-sets to identify transcripts significantly over-represented in aparticular library. This method was designed specifically for thepurpose of statistically evaluating gene expression level acrossmultiple cDNA libraries (16). Log-likelihood ratios calculatedfor the actual data and 1,000 randomized datasets indicated thata log-likelihood ratio �12 for a given contig can be consideredas statistically significant at the 98% confidence level. Contigswith transcript representation in the Fe-limited library wereranked based on decreasing order of EST frequency. Contigswere catalogued as differentially expressed if they met thecriteria of (i) having a log-likelihood ratio �12, (ii) were first orsecond most frequent in the Fe-limited library, and (iii) had aminimum contig size of at least 3 ESTs. 228 contigs wereidentified as being differentially up-regulated in the Fe-limitedtreatment. These contigs were mapped to the genome and the228 contigs were assembled to 212 predicted proteins. Genomebrowser protein IDs for all 212 proteins can be found in Table
S3. R values and EST frequency across all 12 libraries can befound by Blast-searching the PtDB EST database (www.biolo-gie.ens.fr/diatomics/EST). The EST database can also be que-ried with the EST contig IDs found in Table S1. The set of 212proteins identified through analysis of the EST data were usedfor the further comparative analysis against different taxonom-ical lineages.
Representative proteomes were compiled from 14 completedgenomes and the public data repository of National Center forBiotechnology Information and used for comparative analysis.Three major taxonomical groups, heterokonts (3 genomes),plants and algae (5 genomes), and opisthokonts (6 genomes),were represented. Combined proteomes from Thalassiosirapseudonana, Phytophthora sojae, and Phytophthora ramorumwere compiled to represent heterokonts. Complete proteomes ofCyanidioschyzon merolae, Ostreococcus lucimarinus, Ostreococ-cus tauri, Chlamydomonas reinhardtii, and the Chlorophytaprotein sequences from National Center for BiotechnologyInformation were compiled to represent green and red algae.Arabidopsis thaliana and Streptophyta proteins from NationalCenter for Biotechnology Information were combined to form avascular plant dataset. Complete proteomes encoded by thegenomes of Monosiga brevicollis, Ciona intestinalis, Caenorhab-ditis elegans, Saccharomyces cerevisiae, Aspergillus fumigatus, andPichia stipitis were compiled with amino acid sequences fromNational Center for Biotechnology Information representingopisthokonts to form an opisthikont proteome dataset. Theheterokont, green and red algae, and plant proteomes werecompared with the 212 proteins overexpressed in the Fe-limitedcondition, using the Blastp algorithm. Proteins with an E value�10�5, coverage of �50 percent, and �30 percent overallidentity were considered as homologous. P. tricornutum proteinsthat were found to be absent from other genomes were furthercompared against an un-published Pseudo-nitzchia sp. (a pen-nate diatom) EST database. Comparative data is found withinTable S3. The 212 predicted proteins corresponding to theup-regulated transcripts were functionally annotated through acombination of BlastP (Swissprot) results, manual inspection ofalignments, InterPro domain scanning results, and the occur-rence of SignalP, TargetP, ChloroP, and TMHMM targeting andtransmembrane signatures. Proteins determined to have signalpeptides were subsequently examined for plastid targeting byremoval of the signal peptide and evaluation by TargetP, Chlo-roP, and manual inspection (17). Occasionally, the final filteredgene model (ID provided in Table S1) was truncated and theEST extended model was more appropriate for 5� targetingpredictions. BlastP scores, hits against Swissprot, InterPro do-mains, targeting prediction information, EST contig IDs, andgenome browser protein IDs are given in Table S3 for proteinsidentified through EST sequencing and microarray experimentsthat we were able to annotate.
Microarray Fabrication, Hybridization, and Data Processing. The ar-ray platform with all of the probe sequences and correspondinggenome browser protein IDs and all of the normalized hybrid-ization data is available in Minimum Information About aMicroarray Experiment (MIAME) compliant format at theNational Center for Biotechnology Information Gene Expres-sion Omnibus (GEO) website, www.ncbi.nlm.nih.gov/projects/geo. GEO Accession GSE8675 can be viewed at www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token�txohrwusmukyqpw&acc�GSE8675. Oligonucleotetide probes (701) were designed basedprimarily on EST sequence data from a previous P. tricornutumEST sequencing project of 17,000 ESTs (18). A very limitednumber of probes on the microarray were derived from early andpreliminary Fe-limited EST and P. tricornutum genome se-quence data. Spotting of the microarray slides was performed induplicate on each slide by the Genomic Service from the Biology
Allen et al. www.pnas.org/cgi/content/short/0711370105 2 of 5

Department (SGDB) at the Ecole Normale Superieure, Paris.Technical details of the spotting procedure used by the SGDBcan be found at www.transcriptome.ens.fr/sgdb.
Duplicate cultures of Fe-replete and Fe-limited P. tricornutumcells were grown and RNA was isolated as described above. Anon-column digestion of DNA was performed (Qiagen) duringan RNA cleanup step (Qiagen). Purified RNA was preparedseparately for each of the culture replicates. RNA quality andquantity was inspected on a 2100 Agilent Bioanalyzer andNanoDrop (ND-1000), respectively. Using the Ambion AminoAllyl MessageAmp kit (Ambion), 500 ng of total RNA was usedas a template for first strand cDNA synthesis and conversion todouble-stranded DNA (dsDNA). dsDNA was purified and invitro transcribed with amino allyl modified UTPs (aaUTP) togenerate amplified RNA (aRNA). aRNA was purified andquantified, and 10 �g of amino allyl RNA was vacuum-dried andresuspended in 9 �L of dye coupling buffer. Dye couple reac-tions were performed and efficiency was evaluated spectropho-tometrically on a ND-1000. Samples that contained 25–30 in-corporated dye molecules per 1,000 nt were considered suitablefor competitive hybridization.
Before hybridization 5 �g of labeled aRNA made fromFe-replete and Fe-limited samples were mixed together, pre-cipitated, resuspended, and fragmented (Ambion). Each pair ofRNA samples from Fe-replete and Fe-limited duplicate cultures
was competitively hybridized three times for a total of six slides.One slide was a dye-flip hybridization. Microarray slides werescanned by using a GenePix4000B (Axon) two-laser scanner andimage analysis was performed by using GenePix 5.0 (Axon)software. Mean intensities for both RNA populations wereprocessed through minimum and maximum saturation anddiameter filters and normalized by using global lowess followedby a block mean normalization. Image data processing andnormalization was performed by using the SGDB GOULPHARsoftware package http://transcriptome.ens.fr/goulphar (19). Toassign P values and evaluate statistical significance, normalizedlog2ratios were loaded into the Multi experiment Viewer (MeV)software package for analysis of microarray data, www.tm4.org/index.html (20). Normalized log2ratios for each of the duplicateprobes on each slide were averaged. Averaged log2 ratios of thesix slides were converted into two tab delimited multi sample(TDMS) text files with three slides each; each set of threecorresponded to one of the duplicate culture pairs. A one classT test was performed on each group of 3 slides. P values werecalculated based on t distributions with an overall critical valueof 0.05. If the P value of a given gene was less than or equal to0.05 across all of the hybridizations from each of the replicateculture pairs then the change in expression between the twotreatments was considered significant. P values for each set ofthree hybridizations are provided in Table S1.
1. La Roche J, Murray H, Orellana M, Newton J (1995) Flavodoxin expression as anindicator of iron limitation in marine diatoms. J Phycol. 31:520–530.
2. Eckhardt U, Buckhout T (1998) Iron assimilation in Chlamydomonas reinhardtii involvesferric reduction and is similar to Strategy I higher plants. J Exp Bot 49:1219–1226.
3. Finazzi G, et al. (2004) A zeaxanthin-independent nonphotochemical quenchingmechanism localized in the photosystem II core complex. Proc Natl Acad Sci USA101:12375–12380.
4. Sunda W, Huntsman S (2003) Effect of pH, light, and temperature on Fe-EDTA chelationand Fe hydrolysis in seawater. Mar Chem 84:35–47.
5. Hudson RJM, Morel FMM (1990) Iron transport in marine phytoplankton: Kinetics ofcellular and medium coordination reactions. Limnol Oceanogr 35:1002–1020.
6. Hudson RJM, Morel FMM (1993) Trace metal transport by marine microorganisms: Implica-tions of metal coordination kinetics. Deep-Sea Res Part I Oceanogr Res Pap 40:129–151.
7. Schreiber H (1998) in Photosynthesis: Mechanisms and Effects, ed Garab G (KluwerAcademic, Amsterdam), Vol V pp 4253–4258.
8. Witt HT (1979) Energy conversion in the functional membrane of photosynthesis.Analysis by light pulse and electric pulse methods. The central role of the electric field.Biochimm Biophys Acta 505:355–427.
9. Joliot P, Delosme R (1974) Flash-induced 519 nm absorption change in green algae.Biochim Biophys Acta 357:267–284.
10. Bennoun P (1970) Reoxidation of the fluorescence quencher ‘‘Q’’ in the presence of3-(3,4-dichlorophenyl)-1,1-dimethylurea. Biochim Biophys Acta 216:357–363.
11. Joliot P, Joliot A (2005) Quantification of cyclic and linear flows in plants. Proc Natl AcadSci USA 102:4913–4918.
12. Joliot P, Joliot A (1988) The low potential electron transfer chain in the cytochrome b6fcomplex Biochim Biophys Acta 933:319–333.
13. Lisec J, Schauer N, Kopka J, Willmitzer L, Fernie AR (2006) Gas chromatogrephy massspectrometry-based metabolite profiling in plants. Nat Protoc 1:387–396.
14. Schauer N, et al. (2005) GC-MS librraies for the rapid indentification of metabolites incomplex biological samples. FEBS Lett. 579:1332–1337.
15. Bradford M (1976) A rapid and sensitive method for quantitation of microgramquantities of protein utilizing the principle of protein-dye-binding. Anal Biochem72:248–254.
16. Stekel DJ, Git Y, Falciani F (2000) The comparison of gene expression from multiplecDNA libraries. Genome Res 10:2055–2061.
17. Kilian O, Kroth PG (2005) Identification and characterization of a new conserved motifwithin the presequence of proteins targeted into complex diatom plastids. Plant J41:175–183.
18. Maheswari U, et al. (2005) The diatom EST database. Nucleic Acids Res 33:344–347.19. Lemoine S, Combes F, Servant N, Le Crom S (2006) Goulphar: Rapid access and expertise
for standard two-color microarray normalization methods. BMC Bioinformatics. 7:467.20. Saeed AI, et al. (2003) TM4: a free open-source system for microarray data manage-
ment and analysis. BioTechniques. 34:374–378.
Allen et al. www.pnas.org/cgi/content/short/0711370105 3 of 5
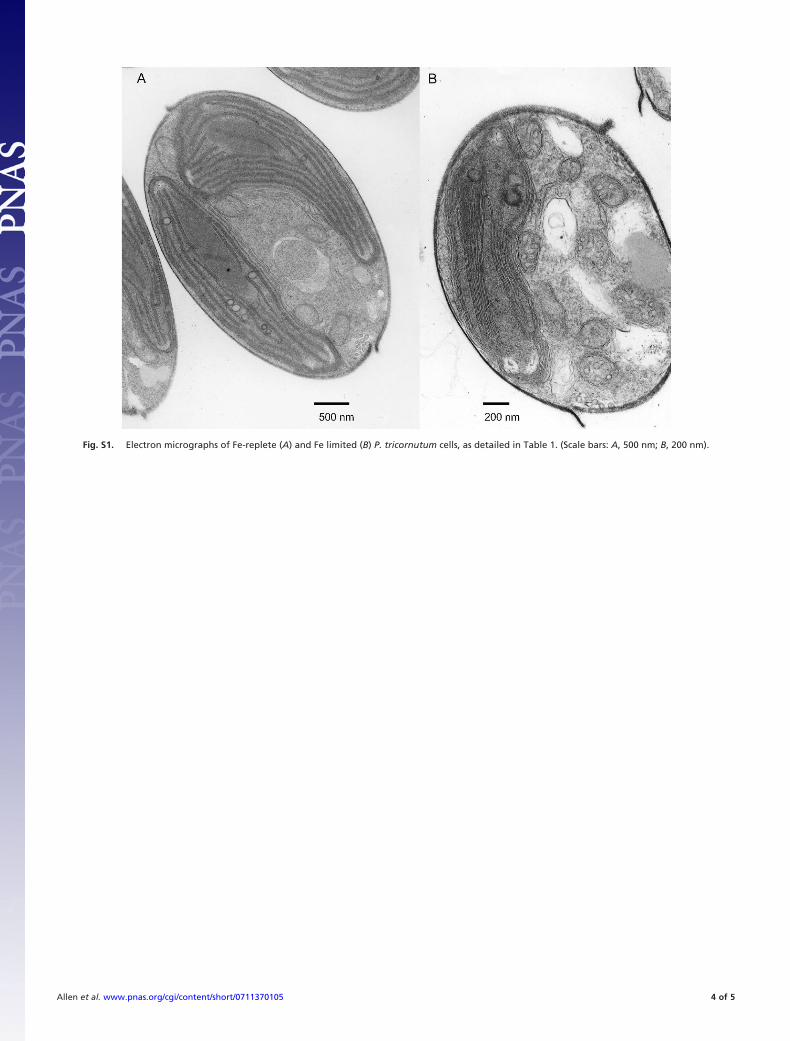
Fig. S1. Electron micrographs of Fe-replete (A) and Fe limited (B) P. tricornutum cells, as detailed in Table 1. (Scale bars: A, 500 nm; B, 200 nm).
Allen et al. www.pnas.org/cgi/content/short/0711370105 4 of 5
Other Supporting Information Files
Table S1Table S2Table S3
Fig. S2. Pathway map of primary metabolism. This representation of metabolism reflects metabolic changes in Fe limited compared to Fe-replete cells. Datawere log2 scaled and color coded. Green boxes represent an increase in metabolite content in Fe limited cells relative to the replete control, while red boxesrepresent a decrease in metabolite content relative to the replete control. The shading from light to dark intensities reflects the level of metabolite changes fromsmall to large. Metabolites in gray were not detected in this analysis.
Allen et al. www.pnas.org/cgi/content/short/0711370105 5 of 5
Related Documents











