1 23 !"#"$%&’%(’)$*+’,- "/( !"#"$%$/)’*%/0$/,+ "##$ %&’()%*+, -./012 +3 $01425 , 67/72.48. 67/72.29: ;3<%3= +3>,+()*<* ?@" %<A%<<(BC%3*,+)<%3)<<&,)’ !"#"$%$&%#%’( %* " +,-."/$0*,-$12#(3 .-22#$ 4%&$1$ &%..51-/( *,%. 6",$2%3 7$8"9 "12 " ,$:-$; %* 9/,"/-’,"+<-& %&&5,,$1&$9 %* !"#$%’$1$ #"12 .".."#9 "&,%99 /<$ =5#* >%"9/"# !#"-13 ?@A !"#$% ’( ’$%*+"*$

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
!"#"$%&'%(')$*+',-."/(!"#"$%$/)'*%/0$/,+!"##$!%&'()%*+,-./012!+3$01425!,!67/72.48.!67/72.29:!;3<%3=!+3>,+()*<*?@"!%<A%<<(BC%3*,+)<%3)<<&,)'
!"#"$%$&%#%'()%*)")+,-."/$0*,-$12#(3).-22#$4%&$1$)&%..51-/()*,%.)6",$2%3)7$8"9"12)"),$:-$;)%*)9/,"/-',"+<-&)%&&5,,$1&$9)%*!"#$%'$1$)#"12).".."#9)"&,%99)/<$)=5#*>%"9/"#)!#"-13)?@A!"#$%&'()'$%*+"*$

1 23
Your article is protected by copyright and allrights are held exclusively by SenckenbergGesellschaft für Naturforschung and Springer.This e-offprint is for personal use onlyand shall not be self-archived in electronicrepositories. If you wish to self-archive yourwork, please use the accepted author’sversion for posting to your own website oryour institution’s repository. You may furtherdeposit the accepted author’s version ona funder’s repository at a funder’s request,provided it is not made publicly available until12 months after publication.

ORIGINAL PAPER
Palaeoecology of a primate-friendly, middle Eocene communityfrom Laredo, Texas and a review of stratigraphic occurrencesof Paleogene land mammals across the Gulf Coastal Plain, USA
James W. Westgate
Received: 16 January 2012 /Revised: 14 March 2012 /Accepted: 3 April 2012 /Published online: 11 May 2012# Senckenberg Gesellschaft für Naturforschung and Springer 2012
Abstract Recent analysis of primate remains in the CasaBlanca local fauna from the late middle Eocene LaredoFormation at Laredo, Texas has shown that the fauna’sprimate species diversity was greater than previouslyreported. Recognition ofMahgarita cf.M. stevensi providedthe first record of an Old World cercomoniine species in aUintan community in North America. The presence of threeomomyids indicates that primate species diversity in theparalic Casa Blanca community was similar to that in lateUintan age faunas in both Trans-Pecos, Texas and in theUinta Basin of Utah. Palaeoecologic evidence from associ-ated plant, invertebrate and vertebrate remains indicates thatthe regional community setting was a tropical mangroveswamp fringing a lowland coastal rain forest. This latemiddle Eocene climate was probably similar to the earlymiddle Eocene climate which prevailed in Germany at thetime the Messel community inhabited the region. A reviewof stratigraphic occurrences of Paleogene land mammalremains across the Gulf Coastal Plain indicates that the CasaBlanca fauna is one of only two Paleogene land mammalcommunities known from Gulf Coastal deposits. Thesecoastal land mammal occurrences provide rare opportunitiesto correlate Paleogene land mammal communities known
from the interior of North America with strata bearingmarine species.
Keywords Biostratigraphy . Paleogene . Mammals . GulfCoastal Plain
AbbreviationsAPL Antero-posterior lengthDGF Devil’s Graveyard FormationDP Upper deciduous premolardp Lower deciduous premolarM Upper molarm Lower molarNALMA North American Land Mammal AgeP Upper premolarp Lower premolarppt Parts per thousandTMM Texas Memorial MuseumUi1, Ui2, Ui3 Early, middle, late Uintan, respectivelyUSGS United States Geological SurveyUSNM United States National Museum
The Casa Blanca fossil community
The Casa Blanca fossil community was collected from latemiddle Eocene Laredo Formation outcrops exposed south-east and south of Lake Casa Blanca in Lake Casa BlancaInternational State Park, in Laredo, Texas (see location mapsin Westgate 1988, 1989, 1990, 1994, 1999; Westgate andGee 1990). Most vertebrate remains come from the verte-brate quarry Texas Memorial Museum (TMM) 42486, withindividual specimens from TMM 41871 and 42185. Inver-tebrate fossils were collected from TMM 42182 and 42587.Strata exposed around Lake Casa Blanca belong to the
This article is a contribution to the special issue "Messel and theterrestrial Eocene—Proceedings of the 22nd Senckenberg Conference”
J. W. WestgateDepartment of Earth & Space Sciences, Lamar University,Texas State University System,Beaumont, TX 77710, USA
J. W. Westgate (*)Vertebrate Paleontology Laboratory, Texas Natural Science Center,University of Texas,Austin, TX 78712, USAe-mail: [email protected]
Palaeobio Palaeoenv (2012) 92:497–505DOI 10.1007/s12549-012-0084-6
Author's personal copy

informal “middle” member of the Laredo Formation anddisplay evidence of deposition under estuarine conditions.The diverse suite of 32 mammalian species in the CasaBlanca fauna offers a rare chance for biostratigraphic corre-lation between the mammal-rich deposits of the NorthAmerican interior and those of marine invertebrate-richcoastal plain deposits. The Red Hot fauna from the latestPaleocene and earliest Wasatchian Tuscahoma Formation atMeridian, Mississippi, is the only other diverse land mam-mal community associated with marine Paleogene depositson the Gulf Coastal Plain (Beard 2008; Beard and Dawson2001, 2009; Dawson and Beard 2007). These two faunasprovide a means to better synchronize the North AmericanLand Mammal Age (NALMA) time scale with marine timescales which are used for intra-coastal plain and trans-oceanic correlation.
The Casa Blanca fossil community inhabited a tropicalrain forest/estuarine mangrove swamp which supported bothcercomoniine and omomyid primates. The presence ofMah-garita cf. M. stevensi at Lake Casa Blanca marks the firstrecord of the subfamily Cercomoniinae in Uintan age depos-its in the New World (Westgate et al. 2010). It is significantthat two skulls and a mandible of the cercomoniine Mah-garita stevensi from the upper Devil’s Graveyard Formation(DGF) are the only records of early Duchesnean primates inTexas. Similarly, the skull of the questionable omomyid,Rooneyia viejaensis, from the Vieja region of Trans-Pecos,Texas, is the sole record of a late Duchesnean primate inTexas.
Casa Blanca omomyids are represented by about 50isolated teeth from a small Mytonius (new species descrip-tion in progress by Cope, Westgate and Beard) and a largeundetermined genus. A fourth primate species is representedby an unidentified upper molar and probably belonged to athird omomyid species. This level of omomyid diversityequals that of the Uinta C member of the Uinta Formationin the Uinta Basin of Uintah County, Utah, where Walsh(1996) reported the occurrence of Mytonius and Ourayia.Westgate et al. (2008) also reported the discovery of anOmomys-sized omomyid from Uinta C. However, no cerco-moniines are yet known from the Uinta Formation (Walsh1996; Williams and Kirk 2008). The Uinta Formationyielded the type fauna on which the Uintan NALMA isbased (Wood et al. 1941).
The Casa Blanca primate diversity mirrors that of itsUinta C correlate, the middle portion of the DGF in Trans-Pecos, Texas. Williams and Kirk (2008) listed the occur-rence of three Uinta C omomyids and an adapiform in theDGF. The omomyids are Diablomomys dalquesti, Omomyscarteri and Mytonius hopsoni. The adapiform is the cerco-moniine Mescalerolemur horneri (Kirk and Williams 2011).
Evidence for the palaeoecologic conditions which existedin the Laredo rain forest and estuary comes from plant,
invertebrate and vertebrate remains. Plant specimens includepollen, spores, fruits, leaves, rhizomes and wood. The threemain plant communities were Nypa mangrove swamps, aback mangrove swamp with trees and Acrostichum ferns anda lowland tropical rain forest with woody dicots, palms,ferns and epiphytes (Gee 2007; Gee and Westgate 1990;Westgate and Gee 1990). Nypamangroves indicate that watertemperatures were 24 °C or warmer (Gee 2007; Westgate andGee 1990).
The invertebrate assemblage is dominated by the estuarineoyster Crassostrea amichel in association with mussels pre-served as molds. Nearly all molluscan shells having aragoniticcomposition were destroyed by post-depositional diagenesis.Modern Crassostrea prefer salinities of 10–30 ppt, with mostproductive reefs growing in salinities averaging about 20 ppt.Callianassid ghost shrimp burrows are common in laminatedsandstone facies cropping-out within the park. Mud lobsterburrows may be found below Crassostrea reefs in LaredoFormation outcrops near Lake Casa Blanca. An excellentexample of this was destroyed in 2008 by urban constructionwhich has encroached on the park’s southern border.
The fish and reptilian assemblage provides environmen-tal clues based on modern habitat preferences and geograph-ic distributions of the closest living relatives of the fossiltaxa. The rays Myliobatis, Dasyatis and Rhinoptera preferwater depths of 1–10 m. Both those rays and the sharks,Galeocerdo and Carcharinus, are found in normal salinityand hyposaline estuarine waters today. However, the lowdiversity of the shark fauna (Striatolamia macrota is theonly other selachian present) and a large sample exceeding1,000 teeth suggest that average salinities approached the10 ppt range (Gee 2007; Westgate 1989, 2001).
The presence of a crocodilian suite of genera, including“Allognathosuchus”, Pristichampsus and a much larger,unidentified crocodylian, along with the tortoise Hadrianus,indicates the climate was frost-free and either tropical orsubtropical (Westgate and Gee 1990). Additionally, thepresence of the aquatic turtles cf. Allaeochelys, cf. Baptemysand Xenochelys suggest the climate was fully tropical, basedon the distribution of their modern relatives. The extantcarettochelyid, Carettochelys insculpta, is restricted to trop-ical rivers of southern New Guinea and northern Australia.Modern dermatemyds and staurotypine turtles inhabit trop-ical streams from southern Mexico to northern CentralAmerica. Of interesting note is that the early middle EoceneLake Messel area and the Casa Blanca estuary shared thetaxa Lepisosteus, “Allognathosuchus”, Pristichampsus, cf.Allaeochelys, Apalone and Microparamys, suggesting that asimilar climatic regime existed in both regions. The pres-ence of Nypa mangroves in middle Eocene paralic depositsin both Europe and Texas also supports this idea.
The following non-primate mammals have been rec-ognized in the Casa Blanca community: marsupials
498 Palaeobio Palaeoenv (2012) 92:497–505
Author's personal copy

(Peratherium comstocki and P. marsupium); insectivores(cf. Simidectes magnus and Centetodon pulcher); chi-ropteran undet.; mesonychid (Harpagolestes uintensis);carnivores (cf. Uintacyon and Procynodictis cf. P. vulpi-ceps); Sirenia undet.; perissodactyls (Epihippus gracilisor E. uintensis, Amynodon advenus, cf. Triplopus and Notio-titanops mississippiensis); artiodactyls (Laredochoerusedwardsi, Protoreodon petersoni, P. edwardsi, Leptoreodonpusillus, L. leptolophus and Toromeryx marginensis); rodents(Mytonomys n. sp., Microparamys sp., Pauromys simplex,Laredomys riograndensis, Microeutypomys karenae, Non-omys sp. and cylindrodontid undet. (Westgate 1990, 2001,2009). Remains of Epihippus gracilis or E. uintensis(horse), Amynodon advenus (rhino) and Mytonomys n.sp. (rodent) indicate that the middle Laredo Formationis Uintan in age. Protoreodon petersoni and Procyno-dictis cf. P. vulpiceps suggest it is Ui3 (late Uintan) inage. The presence ofNotiotitanops mississippiensis provides awestward range extension of this coastal brontothere, whichformerly was known only from the marine Cook MountainFormation of Mississippi (Westgate 2009).
Stratigraphic study of the Rio Grande Embayment datesback to the USA–Mexican Boundary Survey of 1848–1855,which was conducted soon after the newly formed countryof Texas joined the USA (Schott 1857). Early 20th centuryworkers considered the marine outcrops of the middle Eo-cene Claiborne Group in the Laredo area to be part of theCook Mountain Formation. The Cook Mountain Formationis now recognised to extend eastward from central Texas toLouisiana and Mississippi. Cook Mountain Formationplankton are placed in the upper half of zone NP 16 andhave been broadly correlated with upper Lutetian/lowerBartonian strata of Western Europe (Westgate 1988), andmore narrowly with the early Bartonian (Dockery 1996;Dockery and Lozouet 2003). Gardner (1938) changed thename of the sandy Cook Mountain Formation correlates inWebb County, Texas and those cropping-out northward tothe Nueces River, to the Laredo Formation. Near Laredo theformation displays a transition from marine conditions in thelower part, to hyposaline in the middle, and back to marinein the upper part. The Laredo Formation overlies the CannelCoal-bearing El Pico Clay. It underlies the latest middleEocene Yegua Formation, which is the youngest formationin the Claiborne Group in the Rio Grande Embayment.
Stenzel (1940) correlated a thin stratum from Mier inTamaulipas, Mexico, through Webb County, Texas to Leonand Houston Counties in east Texas using the large anddistinctive gastropod Turritella cortezi. In east Texas, theT. cortezi zone lies in the Landrum Member, the third lowestof four members of the Cook Mountain Formation. The T.cortezi zone is exposed in outcrops on the Trinity River andin creek beds in Houston County, Texas. Specimens of T.cortezi were collected in the lower drainage of the Lake
Casa Blanca spillway near the confluence with ChaconCreek at TMM locality 42587, approximately 32 m belowthe TMM 42486 vertebrate quarry horizon. Most of theintervening sedimentary beds are sandy, and many containCrassostrea amichel oyster shells and shell fragments.Some outcrops display high-angled cross-bedding indicativeof rapid deposition in inter-tidal environments and suggestthat the 32 m of intervening strata was deposited during arelatively short chronologic interval.
Two bentonite ash layers lie 0.6 m and 5 m abovethe T. cortezi zone in the Cook Mountain Formation atAlabama Ferry on the Trinity River in east Texas.Berggren et al. (1992) and Robinson et al. (2004) indi-cated these bentonites were 41.2–42.8 Ma. Yancey et al.(2003) reported volcanic ash 40Ar/39Ar dates of 41.53and 41.85 Ma for those bentonites. As the T. cortezizone lies only 32 m below the Casa Blanca quarry, thequarry fauna is slightly younger than the ash beds andis probably between 41 and 42 Ma in age.
In 1985 the Bureau of Economic Geology drilled a 33-m-long geologic core from the stratigraphically highest spot inLake Casa Blanca park to a depth 26 m below TMM 42486.The drill site is 250 m north of the Casa Blanca quarry. Dr.Wulf Gose, University of Texas, assisted in the core recov-ery and analysed it for palaeomagnetic data. Numerouspalaeomagnetic changes (“events”) were recorded; however,there appeared to be only one magnetic “reversal” present.The lower core was primarily positive, and the upper core,including the TMM 42486 quarry horizon, was primarilynegative (W. Gose, personal communication). Although asingle reversal is problematic for stratigraphic correlation byitself, the mammalian fauna from the quarry indicates a Ui3age (Robinson et al. 2004; Westgate 2001), which limits thepossibilities to C19r or C18r. Woodburne (2004) correlatedC18r with the early Bartonian and the Ui3, and C19r withthe late Lutetian and Ui3.
Other Gulf Coastal Plain Paleogene land mammalrecords
Paleocene
Although Gulf Coast stratigraphic correlations based onmarine invertebrates date back to the 1830s, Paleogene landmammal information from the entire Gulf Coastal Plain isscanty (Fig. 1). The oldest specimen, USNM V12147, wasdescribed by Simpson (1932), who based Anisonchus for-tunatus on a partial small peryptychid skull recovered froma core drilled into the Paleocene of Caddo Parish, Louisiana.The specimen came from a depth of about 750 m. Accordingto the online database of the United States National Museum(USNM), it is assigned to the Torrejonian NALMA.
Palaeobio Palaeoenv (2012) 92:497–505 499
Author's personal copy
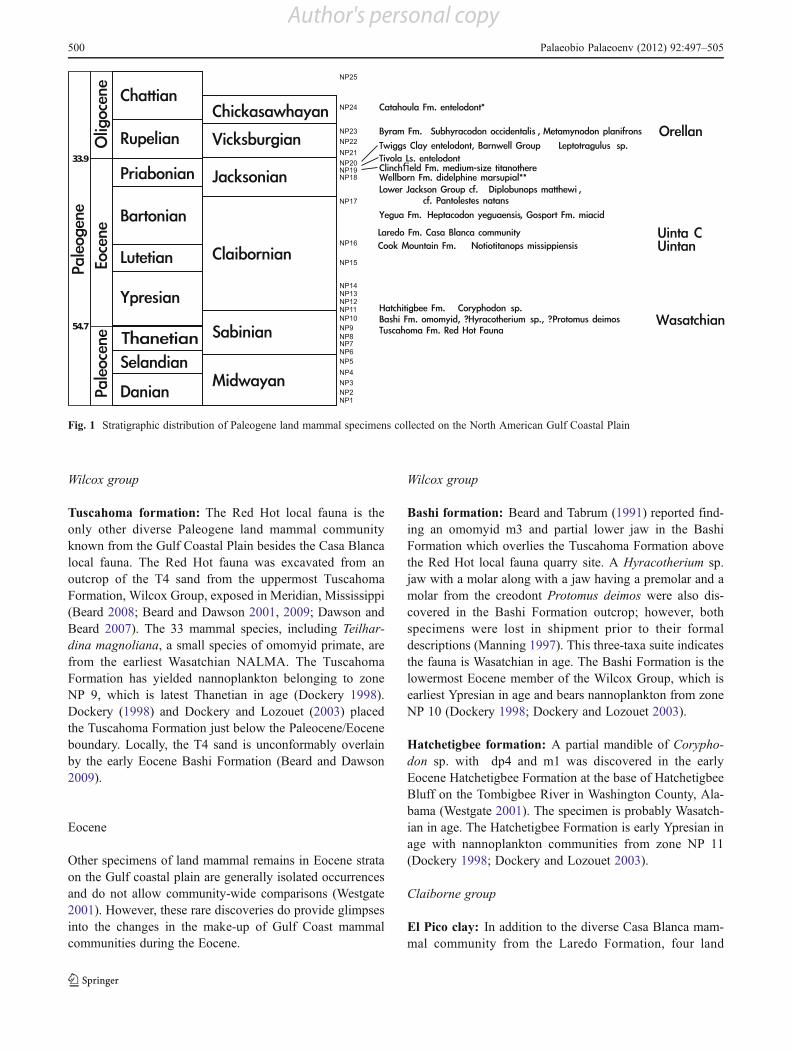
Wilcox group
Tuscahoma formation: The Red Hot local fauna is theonly other diverse Paleogene land mammal communityknown from the Gulf Coastal Plain besides the Casa Blancalocal fauna. The Red Hot fauna was excavated from anoutcrop of the T4 sand from the uppermost TuscahomaFormation, Wilcox Group, exposed in Meridian, Mississippi(Beard 2008; Beard and Dawson 2001, 2009; Dawson andBeard 2007). The 33 mammal species, including Teilhar-dina magnoliana, a small species of omomyid primate, arefrom the earliest Wasatchian NALMA. The TuscahomaFormation has yielded nannoplankton belonging to zoneNP 9, which is latest Thanetian in age (Dockery 1998).Dockery (1998) and Dockery and Lozouet (2003) placedthe Tuscahoma Formation just below the Paleocene/Eoceneboundary. Locally, the T4 sand is unconformably overlainby the early Eocene Bashi Formation (Beard and Dawson2009).
Eocene
Other specimens of land mammal remains in Eocene strataon the Gulf coastal plain are generally isolated occurrencesand do not allow community-wide comparisons (Westgate2001). However, these rare discoveries do provide glimpsesinto the changes in the make-up of Gulf Coast mammalcommunities during the Eocene.
Wilcox group
Bashi formation: Beard and Tabrum (1991) reported find-ing an omomyid m3 and partial lower jaw in the BashiFormation which overlies the Tuscahoma Formation abovethe Red Hot local fauna quarry site. A Hyracotherium sp.jaw with a molar along with a jaw having a premolar and amolar from the creodont Protomus deimos were also dis-covered in the Bashi Formation outcrop; however, bothspecimens were lost in shipment prior to their formaldescriptions (Manning 1997). This three-taxa suite indicatesthe fauna is Wasatchian in age. The Bashi Formation is thelowermost Eocene member of the Wilcox Group, which isearliest Ypresian in age and bears nannoplankton from zoneNP 10 (Dockery 1998; Dockery and Lozouet 2003).
Hatchetigbee formation: A partial mandible of Corypho-don sp. with dp4 and m1 was discovered in the earlyEocene Hatchetigbee Formation at the base of HatchetigbeeBluff on the Tombigbee River in Washington County, Ala-bama (Westgate 2001). The specimen is probably Wasatch-ian in age. The Hatchetigbee Formation is early Ypresian inage with nannoplankton communities from zone NP 11(Dockery 1998; Dockery and Lozouet 2003).
Claiborne group
El Pico clay: In addition to the diverse Casa Blanca mam-mal community from the Laredo Formation, four land
Fig. 1 Stratigraphic distribution of Paleogene land mammal specimens collected on the North American Gulf Coastal Plain
500 Palaeobio Palaeoenv (2012) 92:497–505
Author's personal copy

mammal specimens have been reported from outcrops of themiddle Eocene Claiborne Group. Berry (1924) noted thediscovery of a small mammal femur in Dolores, Texaswhere the El Pico Clay was being mined for Cannel Coalabout 40 km northwest of Laredo. The El Pico Clay under-lies the Laredo Formation. Berry (1924) did not indicatewhat happened to the specimen.
Cook Mountain formation: An unusually complete skulland lower jaw of the titanothere Notiotitanops mississip-piensis was described by Gazin and Sullivan (1942). Thespecimen was discovered in what is now known as theArchusa Marl Member of the Cook Mountain Formationin Clarke County, Mississippi. Gazin and Sullivan (1942)recognised that N. mississippiensis displays a Uintan stageof development. Isolated titanothere teeth, including a bul-bous upper incisor, recovered from the Casa Blanca Quarryin Laredo, Texas are similar in size and morphology to thoseof N. mississippiensis and were referred to that species(Westgate 2009), extending its range from Mississippisouthwest to the Mexican border.
Yegua formation: An anthracothere, Heptacodon yeguaen-sis, was described by Holroyd (2002) based on an M2 andm3 discovered by C. Riley in 1936 at a Yegua Formationoutcrop in Lovelady, Texas. The Yegua Formation is theyoungest unit in the Claiborne Group. Holroyd (2002) notedthat H. yeguaensis displays primitive features for Heptaco-don and is probably older than species of the genus knownfrom the North American interior. These specimens allowonly a broad NALMA age assignment of Ui3 to Chadronianfor the Lovelady fauna. Dockery and Lozouet (2003) indi-cated that the Yegua Formation bears nannoplankton fromthe lower half of zone NP 17 and is middle Bartonian in age.
Holroyd (2002) noted that dermatemyid turtle and ariidcatfish material were associated with H. yeguaensis. TheAmerican Museum on-line catalog also lists a dasyatid, apristid, crocodilians, the estuarine/marine snake Palaeophissp., Baptemys sp., Echmatemys sp. and “Trionyx” (0Apalone) sp. in the suite of fossils collected by Claude Rileyat the AMNH Lovelady Loc. 3 TX site. Although Heptaco-don is not in the Casa Blanca fauna, rays, sawfish, crocodi-lians, and cf. Baptemys sp., Apalone sp. cf. Arius sp.(Westgate 1989) are present and indicate that the two habitatswere similar. The presence of the palaeophid snake Pteros-phenus schucherti in the Casa Blanca fauna is the first evi-dence of the genus in North American middle Eocene strata.The Lovelady paleophid material should be re-examined todetermine whether it is Pterosphenus and not Palaeophis, aspalaeophid vertebrae are not easily identifiable if their ptera-pophyses (aliform processes) are not preserved.
Palynomorphs in the Cook Mountain and Yegua Forma-tions in central and east Texas indicate that tropical
conditions cooled locally through late middle Eocene time(Elsik and Yancey 2000). Oxygen isotope analysis of marinegastropods in the Wheelock Member of the Cook MountainFormation led Andreasson and Schmitz (2000) to concludethat the marine community lived under mean sea surfacewater temperatures of 23–24 °C. That is a tight match withclimatic interpretations for the slightly younger Casa BlancaNypa mangrove community at Laredo which required meantemperatures of at least 24 °C (Gee 2007; Gee and Westgate1990; Westgate 1994). The younger Lovelady communitymay have lived under somewhat cooler paratropical condi-tions than that of the Casa Blanca andWheelock communities.
Gosport sand: A partial m1 of Vulpavus sp. was reportedfrom the Gosport Sand of Clarke County, Alabama byThurmond and Jones (1981). Vulpavus is a well-knownmember of Wasatchian and Bridgerian age faunas in theRocky Mountain Region (early and middle Eocene). Al-though it is possible that Vulpavus survived to the latemiddle Eocene in a Gulf Coast refugium, basing a geologicrange extension of the genus on a partial tooth is question-able. It may be more realistic to identify the specimen asmiacid, with genus and species undetermined. The GosportSand is an eastern Gulf Coastal Plain correlate of the YeguaFormation and contains nannoplankton in the lower part ofNP 17 and is also of middle Bartonian age (Dockery 1998;Dockery and Lozouet 2003).
Jackson group: Only three land mammal specimens areknown from the Jackson Group—two in Arkansas and onein Texas. Westgate and Emry (1985) described a well-preserved M3 and partial maxilla of Diplobunops cf. D.matthewi and an edentulous, but nearly complete, mandib-ular ramus of Pantolestes cf. P. natans from Jackson Groupoutcrops on Crow Creek in St. Francis County, Arkansas.The two taxa fall in the broad time range of the Ui3 toChadronian NALMA. The mammals were found in an es-tuarine community that included the freshwater to hyposa-line gastropod Hemisinus jacksonius, the hyposaline oysterCrassostrea alabamensis, sharks, the giant estuarine snakePterosphenus schucherti, turtles and the archaeocete whalesZygorhiza kochii and Basilosaurus cetoides (Westgate 1982,1984; Westgate and Salazar 1996; Westgate and Ward1981). Harris and Palmer (1947) interpreted the suite of H.jacksonius, C. alabamesis and barnacles to indicate thatdeposition occurred under inter-tidal conditions. TheCrow Creek locality lies approximately 13 km south ofthe northernmost marine Jackson Group outcrops inArkansas (Westgate 2001). The United States GeologicalSurvey (USGS) does not differentiate the Jackson Groupinto formations in Arkansas, but does correlate it with theMoodys Branch and Yazoo Clay Formations of Mississippi.Dockery (1977) described the Crow Creek deposits as being
Palaeobio Palaeoenv (2012) 92:497–505 501
Author's personal copy

part of the lower Jackson Group (Moodys Branch Formation)northern destructional shelf facies. The Moodys Branch For-mation in Mississippi has nannoplankton from the upper halfof NP 17 (Dockery and Lozouet 2003) and planktonic foramsfrom the upper half of P14 (Dockery 1996). If the Jacksoniandeposits on Crow Creek are the same age, they are latestmiddle Eocene and late Bartonian.
Wellborn formation: In the far eastern part of Texas, be-tween Toledo Bend and the Sam Rayburn Reservoir (Sabineand Angelina Rivers, respectively), the Jackson Group isdivided by the Texas Bureau of Economic Geology into theMoodys Branch and Yazoo Formations. West of the SamRayburn Reservoir the Yazoo Formation becomes the Well-born and overlying Manning Formations. Dumble (1918)reported finding small mammal bones and a possible creo-dont jaw in association with shark and ray teeth on RockyCreek in Polk County, Texas. Dumble’s specimens appear tohave been lost. Slaughter (1978) reported the discovery of aDP3 or M1 of a didelphine marsupial in the WellbornFormation on Rocky Creek.
Barnwell group
Clinchfield formation: Deposits of the Barnwell Groupin central Georgia mark the position where the GulfCoastal Plain meets the Atlantic Coastal Plain. Westgate(2001) listed a titanothere ?P3 deuterocone fragment inthe Hardie Mine local fauna. The specimen is in the sizerange of the same cusp on the P3 of the Cook MountainFormation titanothere N. mississippiensis. Parmley andHolman (2002) also noted the presence of unstudied teethfrom a moderately large titianothere from the Hardie Minein the Georgia College and State University collections.The Hardie Mine titanothere material is too fragmentaryto assign a land mammal age; however, Prothero andEmry (2004) noted that titanotheres have not beenfound in post-Chadronian faunas, so the Hardie Minefauna is probably Chadronian or older. The Hardie Minefauna was collected from a 1-m-thick layer of ClinchfieldFormation sands in the Hardie kaolin mine just north ofGordon, Georgia.
Huddlestun and Hetrick (1986) noted that earlier strat-igraphers had often mistakenly correlated the shelly andsandy nearshore Gosport Sand of Alabama with the shellysands of the Clinchfield Formation. Westgate (2001) not-ed that large odontaspid shark teeth are good field indi-cators of Claibornian versus Jacksonian age strata.Striatolamia macrota was the dominant Claibornian odon-taspid, while Carcharias spp. dominated Jacksonianfaunas. S. macrota is common in the Gosport Sandexposures on Little Stave Creek, as is Carcharias spp.in the Clinchfield Formation in the Hardie Mine. The
Clinchfield Formation is the basal nearshore sand left asthe Jackson Sea transgressed across central Georgia(Huddlestun and Hetrick 1986). Huddlestun and Hetrick(1986) correlated the Clinchfield Formation with thelower Moodys Branch Formation of late Bartonian age.However, Parmley and Holman (2002) stated that dino-cysts in the Clinchfield Formation are from zones NP19/20, which are Priabonian and would mean that theClinchfield Formation is a correlate of the Yazoo Clayto the west. If this younger age for the Clinchfield For-mation is correct, it indicates that the Jackson Sea trans-gression into central Georgia occurred during the lateEocene after the central Gulf Coast transgression laiddown the Moodys Branch Formation in Mississippi dur-ing the latest middle Eocene.
Tivola limestone and Twiggs clay: Domning et al. (1982)reported the discovery of a worn M1 or 2 from an entelodontin the Tivola Limestone exposed by kaolin mining in TwiggsCounty, Georgia. Domning et al. (1982) also noted that asubsequent probable entelodont tooth was found in the over-lying Twiggs Clay at the same pit. Huddlestun and Hetrick(1986) correlated the Tivola Limestone with the upper Moo-dys Branch Formation of Alabama and correlated the TwiggsClay with the North Twistwood Creek Clay and Cocoa Sandof Alabama. If the transgression which laid down the Clinch-field Formation reached central Georgia during NP 19/20time, those Barnwell Group units overlying the Clinch-field Formation are no older than the age of NP 20,which is Priabonian in age (Dockery 1998; Dockeryand Lozouet 2003).
Lastly, there is an undescribed jaw in the USNM collec-tions (USNM 244447), identified as Leptotragulus sp.,which was collected from the Barnwell Group in JeffersonCounty, Georgia. Walsh (1996) indicated that Leptotraguluscommonly occurs in Uinta B and C faunas and that it has notbeen reported outside of the Rocky Mountains. Robinson etal. (2004) noted that Leptotragulus is not known from post-Duchesnean faunas. If the specimen actually is Leptotragu-lus, its presence suggests that either part of the BarnwellGroup is Duchesnean or Uintan in age, or Leptotragulussurvived in Georgia past its extinction in the Rocky Moun-tain region. Based on the above discussion, USNM 244447is Priabonian in age.
Oligocene
Vicksburg group
Byram formation: Two biostratigraphically significantspecimens have been collected from Byram Formation out-crops on the Big Black River in Hinds County, Mississsippi(Manning 1997). A skull of Metamynodon planifrons and a
502 Palaeobio Palaeoenv (2012) 92:497–505
Author's personal copy

nearly complete lower jaw of Subhyracodon occidentaliswere found on the same bluff within three vertical metersof each other. Manning et al. (1986) reported the M. plani-frons discovery and suggested that the Byram Formationwas a Chadronian correlate based on the occurrence. In1997, Manning revised that correlation to be Orellan basedon the association with S. occidentalis. The Byram Forma-tion is the second youngest formation in the VicksburgGroup, bears nannoplankton from zone NP 23, and is mid-dle Rupelian and early Oligocene in age (Dockery 1996).
Catahoula formation: Westgate (1993) reported the firstGulf Coastal Plain land mammal found in Oligocene depos-its west of the Mississippi River. The specimen is a nearlycomplete mandible bearing p1–m3 from a large and newgenus of entelodont collected from an outcrop of the Cata-houla Formation by Dr. John A. Wilson near Carmine inFayette County, Texas. Although the jaw length (75 cm) isnearly as long as that of Dinohyus hollandi, the molar seriesAPL is only 75 % of the length of D. hollandi molars. Thelarge but gracile design of the Carmine entelodont suggestsa Whitneyan to early Arikareean stage of evolution.
The Catahoula Formation is of fluvial origin and does nothave marine invertebrates for biostratigraphic correlation.However, Daghlian and Crepet (1983) described a suite offossil Quercus (oak) remains from Catahoula Formationoutcrops near Huntsville, Texas, 130 km northeast of Car-mine. It is notable that Singleton (2001) reported that 50 %of the petrified wood found in the Catahoula Formation ofJasper County, Texas, some 300 km east of Carmine, is fromQuercus sp. Singleton (2001) noted the following common-ly found trees in the Catahoula Formation flora in JasperCounty: Ulmus (elm), Acer (maple), Gleditsia (honey lo-cust) and Ulminium (laurel) and/or legume. This subtropicalforest suite indicates that the fully tropical Texas coastalclimatic conditions which prevailed during the middle Eo-cene had cooled somewhat by mid-Oligocene time.
I-75 fauna
Patton (1969) reported the discovery of a Whitneyan faunain a karst feature overlying the late Eocene Ocala Limestonein Gainesville, Florida. The isolated deposit does not belongto a named formation, but is significant in yielding a diversemammalian and lower vertebrate community. Most of Pat-ton’s (1969) mammals were identified at the family level.Czaplewski and Morgan (2007) provided a detailed list intheir on-line publication on fossil bats of the Americas.Their list includes Herpetotherium sp., Centetodon cf. C.wolffi, seven species of bats, Palaeolagus sp., Daphoenussp., a mustelid, Miohippus sp., a tayassuid, an oreodont,Leptomeryx sp. and four rodents (Eutypomys sp., a hetero-myid, and two eomyids).
Discussion
The record of terrestrial mammals in Gulf Coast Paleogenestrata is important for several reasons. The most important oneis that it allows correlation between marine communities,which are the basis for the classic Lyellian epoch time scaleof 1833, and the NALMAs established byWood et al. (1941).Wood et al. (1941) correlated the Wasatchian, Bridgerian,Uintan and Duchesnean NALMAs with the early, middle, lateand latest Eocene Epoch, respectively. They also correlatedthe Chadronian and Orellan NALMAs with the early andmiddle Oligocene, respectively. In 1941, few land mammalspecimens and no land mammal communities were knownfrom Gulf Coast marine strata to verify that correlation.
The discoveries of terrestrial land mammal communities atthe Red Hot Truck Stop and Lake Casa Blanca, as well as theassociation of two well-known land mammal genera in amarine stratum on the Big Black River allow us to moreprecisely calibrate the early correlations of Wood et al.(1941). Recent discoveries of isolated land mammal taxaacross the Gulf Coastal Plain help fill in details, but are notas useful as assemblages of mammalian species. It is not clearif these isolated occurrences document geographic rangeextensions of Rocky Mountain communities of isochronousage. Instead, they may be chronologic range extensions oftheir lineages which survived local community extinctions inthe Rocky Mountains by inhabiting climatic refugia on theGulf Coast. Although it is possible for whole communities toalso persist in refugia, those communities are also subject tointra- and inter-continental immigration of taxa through time.Therefore, correlations of mammalian community assemb-lages are preferable to single taxon correlations as evidencedbyManning’s (1997) correlation revision of the age of the BigBlack River mammal specimens from Chadronian to Orellan.
Using the Red Hot mammal assemblage, Beard andDawson (2009) correlated the beginning of Wasatchian timewith the latest Paleocene and middle Sabinian. TheWasatchian continued on into the early Eocene and the lateSabinian, as evidenced by the small Tuscahoma Formationmammalian assemblage and the occurrence of Coryphodonin the Hatchetigbee Formation.
No Bridgerian mammals have been recovered from GulfCoastal Plain strata. It cannot yet be determined if thebeginning of Bridgerian time falls in the Sabinian or Clai-bornian age, but it is most likely early Claibornian. Based onthe stratigraphic position of the Casa Blanca community, theBridgerian probably ended during the middle Claibornian.
Ui1 may be middle Claibornian in age; however, with norecord of Bridgerian or middle Uintan (Ui2) mammals fromthe Gulf Coast, no direct correlation may be made. The Ui3Casa Blanca community is late Claibornian in age, as is theNotiotitanops mississippiensis holotype specimen fromMississsippi.
Palaeobio Palaeoenv (2012) 92:497–505 503
Author's personal copy

Although eight land mammal specimens are known fromlatest middle to latest Eocene strata, the boundaries betweenthe Ui3, the Duchesnean and the Chadronian are unresolved.One land mammal specimen is known from the YeguaFormation (latest Claibornian) and three from the JacksonGroup (latest middle Eocene to latest Eocene). However, allfour could be Uintan to Chadronian in age, and none indi-cate when the Uintan ended or the Duchesnean or Chadro-nian began or ended.
Three land mammal specimens from the Barnwell Groupof Georgia provide a little more information. The two ente-lodont specimens are no older than Duchesnean in age.Therefore, the Uintan must have ended and the Duchesneanhave begun prior to the appearance of entelodonts in NP19/20 in the late Jacksonian (later Eocene). The Hardie Minetitanotheres indicate that the Clinchfield Formation (basalBarnwell Group) is probably no younger than Chadronian.
There are no land mammal remains known from thelower Vicksburg Group (early Rupelian, early Oligocene).The Chadronian/Orellan boundary may lie in this intervalrather than coinciding with the Eocene/Oligocene boundary,as indicated by Woodburne (2004), just as the WasatchianNALMA overlaps the Paleocene/Eocene boundary.
The presence of Subhyracodon occidentalis and Metamy-nodon planifrons in the Byram Formation indicates that theOrellan had begun and the Chadronian had ended by mid-Vicksburgian time (middle Rupelian, early Oligocene). Incentral Texas, the Carmine entelodont lacks a marine associ-ation, and it is not clear where it falls on the marine time scale,but it is Orellan to early Arikareean in age. The I-75 Floridafauna is also problematical in being clearly of Whitneyan age,but of indeterminate age on the marine time scale.
Acknowledgements Countless experts have helped to identify andupdate the components of the Casa Blanca community because of itstaxonomic diversity ranging from fully marine invertebrates to terres-trial rain forest species, and the piecemeal preservation and recovery ofits specimens. I thank all of those who have endeavored to better clarifythis unique glimpse of the middle Eocene history of the Texas GulfCoast. I also wish to thank reviewers Drs. Chris Beard and BethTownsend for their helpful suggestions which improved the manu-script. Dr. Jeff Pittman drafted Figure 1.
References
Andreasson FP, Schmitz B (2000) Temperature seasonality in the earlymiddle Eocene North Atlantic region: evidence from stable iso-tope profiles of marine gastropod shells. Geol Soc Am Bull112:628–640
Beard KC (2008) The oldest North American primate and mammalianbiogeography during the Paleocene–Eocene thermal maximum.Proc Natl Acad Sci USA 105:3815–3818
Beard, KC, Dawson MR (2001) Early Wasatchian mammals from theGulf Coastal Plain of Mississsippi: Biostratigraphic and paleobio-logic implications. In: Gunnell G (ed) Eocene biodiversity:
Unusual occurrences and rarely sampled habitats. Kluwer Aca-demic/Plenum Publ, New York, pp 75–94
Beard KC, Dawson MR (2009) Early Wasatchian mammals of the RedHot local fauna, uppermost Tuscahoma Formation, LauderdaleCounty, Mississsippi. Ann Carnegie Mus 78(3):193–243
Beard KC, Tabrum AR (1991) The first early Eocene mammal fromeastern North America: an omomyid primate from the BashiFormation, Lauderdale County, Mississippi. Miss Geol 11:1–6
Berggren WA et al (1992) Toward a revised Paleogene geochronology,pp. 29–45. In: Prothero DR, Berggren WA (eds) Eocene-Oligocene climatic and biotic evolution. Princeton UniversityPress, Princeton, pp 29–45
Berry EW (1924) The middle and upper Eocene floras of southeasternNorth America. U S Geol Surv Prof Pap 92:1–206
Czaplewski NJ, Morgan, GS (2007) Fossil bats of the Americas. Avail-able at: snomnh.ou.edu/collections-research/cr-sub/vertpaleo/fossil_bats. Accessed 3 Jan 2012
Daghlian CP, Crepet WL (1983) Oak catkins, leaves and fruits from theOligocene Catahoula Formation and their evolutionary signifi-cance. Am J Bot 70:639–649
Dawson MR, Beard KC (2007) Rodents of the family Cylindrodonti-dae (Mammalia) from the earliest Eocene of the TuscahomaFormation, Mississippi. Ann Carnegie Mus 76(3):135–144
Dockery DTIII (1977) Mollusca of the Moodys Branch Formation,Mississippi. Miss Geol Surv Bull 120
Dockery DTIII (1996) Toward a revision of the generalized stratigraph-ic column of Mississippi. Miss Geol 17:1–9
Dockery DTIII (1998) Molluscan faunas across the Paleocene/EoceneSeries boundary in the North American Gulf Coastal Plain. In:Aubry M, Lucas S, Berggren W (eds) Late Paleocene–earlyEocene climatic and biotic events in the marine and terrestrialrecords. Columbia University Press, New York, pp 296–322
Dockery DT, Lozouet P (2003) Molluscan faunas across the Eocene/Oligocene boundary in the North American Gulf Coastal Plain,with comparisons to those of the Eocene and Oligocene of France.In: Prothero D, Ivany L, Nesbitt E (eds) From greenhouse toicehouse: The marine Eocene–Oligocene transition. ColumbiaUniversity Press, New York, pp. 303–340
Domning DP, Morgan GS, Ray CE (1982) North American Eocene seacows (Mammalia, Sirenia). Smithsonian Contrib Paleobiol 52:1–69
Dumble ET (1918) The geology of east Texas. Univ Texas Bull1869:1–388
Elsik WC, Yancey TE (2000) Palynomorph biozones in the context ofchanging paleoclimate, middle Eocene to lower Oligocene of theNorthwest Gulf of Mexico. Palynology 24:177–186
Gardner JA (1938) Laredo, a new name for a unit of Cook Mountain agein the Rio Grande Region. Washington Acad Sci J 28(7):297–298
Gazin CL, Sullivan JM (1942) A new titanothere from the Eocene ofMississippi, with notes on the correlation between the marineEocene of the Gulf Coastal Plain and continental Eocene of theRocky Mountain region. Smithsonian Misc Coll 101(13):1–13
Gee CT (2007) The mangrove palm Nypa and its paleoeclogicalimplications for a middle Eocene community from Laredo, Texas,USA. Quat Int 167–168[Suppl]:446–447
Gee CT, Westgate JW (1990) Paleoecological reconstruction of amiddle Eocene mangrove community from southwest Texas usingpalynology. Palynology 14:212
Harris GD, Palmer KV (1947) The mollusca of the Jackson Eocene ofthe Mississippi Embayment (Sabine River to the Alabama River):Part II, Univalves and index. Bull Am Paleontol Paleont Res Inst30(117):207–563
Holroyd PA (2002) New record of anthracotheriidae (artiodactyla:mammalia) from the middle Eocene Yegua Formation (ClaiborneGroup), Houston County, Texas. Texas J Sci 54(4):301–308
Huddlestun PF, Hetrick JH (1986) Upper Eocene stratigraphy of cen-tral and eastern Georgia. Ga Geol Surv Bull 95
504 Palaeobio Palaeoenv (2012) 92:497–505
Author's personal copy

Kirk EC, Williams BA (2011) New adapiform primate of Old Worldaffinities from the Devil’s Graveyard Formation of Texas. J HumEvol 61:156–168
Manning EM (1997) An early Oligocene rhinoceros jaw from themarine Byram Formation of Mississsippi. Miss Geol 18(2):13–31
Manning EM, Dockery DT III, Scheibout JA (1986) Preliminary reportof a Metamynodon skull from the Byram Formation (lower Oli-gocene) in Mississippi. Miss Geol 6(2):1–16
Parmley D, Holman JA (2002) Nebraskophis Holman from the Eoceneof Georgia (USA), the oldest known North American colubridsnake. Acta Zool Cracovien 46(1):1–8
Patton TH (1969) An Oligocene land vertebrate fauna. J Paleontol 43(2):543–546
Prothero DR, Emry RJ (2004) The Chadronian, Orellan and WhitneyanNorth American Land Mammal Ages. In: Woodburne M (ed) LateCretaceous and Cenozoic mammals of North America. ColumbiaUniversity Press, New York, pp 156–168
Robinson PG, Gunnell GF, Walsh SL, Clyde WC, Storer JE, StuckyRK, Froelich DJ, Ferresqilla-Villafranca I, McKenna MC (2004)Wasatchian through Duchesnean biochronology. In: WoodburneMO (ed) Late Cretaceous and Cenozoic mammals of NorthAmerica. Columbia University Press, New York, pp 106–155
Schott A (1857) Geology of the lower Rio Bravo del Norte. MexBound Surv 1(2):28–48
Simpson GG (1932) A new Paleoecene mammal from a deepwell inLouisiana. U S Natl Mus Proc 82(2):1–4
Singleton S (2001) Fossil wood of the Oligocene CatahoulaFormation, Jasper County, Texas. Houston Gem and MineralSociety. Available at: PaleoPetrifiedWoodArticles ([email protected]). Accessed 3 Jan 2012
Slaughter BH (1978) Occurrences of didelphine marsupials from theEocene and Miocene of the Texas Gulf Coastal Plain. J Paleontol52:744–746
Stenzel HB (1940) New zone in the Cook Mountain Formation, theCrassatella texalta Harris-Turritella cortezi Bolwes zone. AmAssoc Petrol Geol Bull 24:1633–1675
Thurmond JT, Jones DE (1981) Fossil vertebrates of Alabama. Uni-versity of Alabama Press, Tuscaloosa
Walsh SL (1996) Middle Eocene mammal faunas of San Diego County,California. In: Prothero DR, Emry RJ (eds) The terrestrial Eocene–Oligocene transition in North America. CambridgeUniversity Press,New York, pp 75–119
Westgate JW (1982) Hemisinus jacksonius (Gastropoda) as an aid indetermining potential of marine deposits for yielding remains ofterrestrial mammals. Mo Acad Sci Trans 16:49–53
Westgate JW (1984) Lower vertebrates from the late Eocene CrowCreek local fauna, St. Francis County, Arkansas. J Vert Paleontol4:536–546
Westgate JW (1988) Biostratigraphic implications of the first Eoceneland-mammal fauna from the North American coastal plain.Geology 16:995–998
Westgate JW (1989) Lower vertebrates from an estuarine facies of themiddle Eocene Laredo Formation (Claiborne Group) of WebbCounty, Texas. J Vert Paleontol 9:282–294
Westgate JW (1990) Uintan land mammals (excluding rodents) froman estuarine facies of the Laredo Formation (middle Eocene,Claiborne Group) of Webb County, Texas. J Paleontol 64:454–468
Westgate JW (1993) A giant entelodont from Oligocene deposits of theTexas Coastal Plain. J Vert Paleontol 13(3):62A
Westgate JW (1994) Eocene forest-swamp. Res Explor Natl Geog Soc10:78–89
Westgate JW (1999) After the dinosaurs: a Texas tropical paradisefound. Texas Parks and Wildlife Press, Austin
Westgate JW (2001) Paleoecology and biostratigraphy of marginalmarine Gulf Coast Eocene vertebrate localities. In: GunnellGF (ed) Eocene biodiversity: unusual occurrences and rarelysampled habitats. Kluwer Academic/Plenum Publ, New York,pp 263–297
Westgate JW (2009) First records of two large land mammals frommiddle Eocene Coastal Plain strata of Texas. Geol Soc Am AnnMeet 41(7):106
Westgate JW, Emry RJ (1985) Land mammals of the Crow Creek localfauna, late Eocene, Jackson Group, St. Francis County, Arkansas.J Paleontol 59:242–248
Westgate JW, Gee CT (1990) Paleoecology of a middle Eocenemangrove biota (vertebrates, plants, and invertebrates) fromsouthwest Texas. Palaeogeogr Palaeoclimatol Palaeoecol 78:163–177
Westgate JW, Salazar AL (1996) Additions to the late Eocene(Jacksonian) cetacean and chondrichthyan faunas of Arkan-sas (abstract). Geol Soc Am South-Central Sect Ann Meet,p 68
Westgate JW, Ward JF (1981) The giant aquatic snake Pterosphenusschucherti (Palaeophidae) in Arkansas and Mississippi. J VertPaleontol 1:161–164
Westgate JW, Townsend KE, Cope DA, Friscia AR (2008) Snapshotfrom the past: a Uintan vertebrate assemblage preserving commu-nity structure. J Vert Paleontol 28(3) [Suppl] 159A
Westgate JW, Cope DA, Beard KC (2010) First occurrence and signif-icance ofMahgarita stevensi in a late middle Eocene (late Uintan)Tethyan-influenced Gulf coast community. J Vert Paleontol SVPProg & Abstr 186A
Williams BA, Kirk EC (2008) New Uintan primates from Texas andtheir implications for North American patterns of species richnessduring the Eocene. J Hum Evol 55:927–941
Wood HE, Chaney R, Clark J, Colbert E, Jepsen E, Reeside W, Stock C(1941) Nomenclature and correlation of the North Americancontinental Tertiary. Geol Soc Am Bull 52:1–46
Woodburne MO (2004) Global events and the North American mam-malian biochronology. In: Woodburne MO (ed) Late Cretaceousand Cenozoic mammals of North America. Columbia UniversityPress, New York, p 391
Yancey TE, Elsik WC, Sancay RH (2003) The palynological record oflate Eocene climate change, northwest Gulf of Mexico. In: Pro-thero DR, Ivany LC, Nesbitt EA (eds) From greenhouse to ice-house: the marine Eocene–Oligocene transition. ColumbiaUniversity Press, New York, pp 252–268
Palaeobio Palaeoenv (2012) 92:497–505 505
Author's personal copy
Related Documents











