Aquatic Toxicology 163 (2015) 102–108 Contents lists available at ScienceDirect Aquatic Toxicology j o ur na l ho me pag e: www.elsevier.com/locate/aquatox Water contaminated with Didymosphenia geminata generates changes in Salmo salar spermatozoa activation times Pamela Olivares a , Paola Orellana a , Guillermo Guerra a , Matías Peredo-Parada b,c , Viviana Chavez d , Alfredo Ramirez e , Jorge Parodi a,∗ a Laboratorio Fisiología de la Reproducción, Escuela de Medicina Veterinaria, Núcleo de Investigación en Producción Alimentaria, Facultad de Recursos Naturales, Universidad Católica de Temuco, Chile b Departamento de Ingeniería en Obras Civiles, Universidad de Santiago de Chile, Chile c Plataforma de Investigación en Ecohidrología y Ecohidráulica, EcoHyd Ltda, Chile d Laboratorio de Investigación y Educación, Tonalli Ltda, Chile e Laboratorio de Criobiología y Análisis de Funcionalidad Espermática. Instituto de Ciencia Animal, Facultad de Ciencias Veterinarias, Universidad Austral de Chile, Valdivia, Chile a r t i c l e i n f o Article history: Received 4 February 2015 Received in revised form 26 March 2015 Accepted 28 March 2015 Available online 8 April 2015 Keywords: Didymo Sperm cells Toxity a b s t r a c t Didimosphenia geminata (“didymo”), has become a powerful and devastating river plague in Chile. A system was developed in D. geminata channels with the purpose evaluating the effects of water polluted with didymo on the activation of Atlantic salmon (Salmo salar) spermatozoa. Results indicate that semen, when activated with uncontaminated river water had an average time of 60 ± 21 s. When using Powermilt, (a commercial activator), times of 240 ± 21 s are achieved, while rivers contaminated with D. geminata achieve a motility time of 30 ± 12 s. Interestingly enough, the kinetic parameters of VSL, VCL and VAP showed no significant changes under all of the conditions. Furthermore, the presence of D. geminata reduces activation time of the samples as the cells age, indicating increased effects in spermatozoa that are conserved for more than 5 days. D. geminata has antioxidant content, represented by polyphenols; 200 ppm of polyphenol were obtained in this study per 10 g of microalgae. Spermatozoa exposed to these extracts showed a reduction in mobility time in a dose dependent manner, showing an IC50 of 15 ppm. The results suggest an effect on spermatozoa activation, possibly due to the release of polyphenols present in contaminated rivers, facilitating the alteration of sperm motility times, without affecting the viability or kinetics of the cells. These findings have important implications for current policy regarding the control of the algae. Current control measures focus on the number of visible species, and not on the compounds that they release, which this study shows, also have a problematic effect on salmon production. © 2015 Elsevier B.V. All rights reserved. 1. Introduction Didymosphenia geminata (D. geminata) is a unicellular benthic diatom. This microalgae known as “didymo” in Chile, has been found in the river waters of southern Chile (Riveraet al., 2013), and has been considered a plague in freshwater sources by the Gov- ernment of Chile and can affect fish population (Reid et al., 2012). International studies indicate that D. geminata alters the microen- vironment, reduces the fish population (Clearwater et al., 2010) and perturbs aquatic macro-vertebrate communities and drinking water system filters (Bergey et al., 2010; Gillis and Chalifour, 2010; ∗ Corresponding author at: Av. Las Mariposas S/N, Campus Dr. Rivas, Universidad Católica de Temuco, Temuco, Chile. Tel.: +56 45205564. E-mail address: [email protected] (J. Parodi). Kilroy et al., 2009). Recently, a toxic effect of microalgae on com- munities in contaminated rivers was described (Larned and Kilroy, 2014). The removal of the microalgae, also introduces damages to rivers (Larned and Kilroy, 2014). In Chile, there are no stud- ies that confirm this hypothesis and we must control these effects with laboratory research to support field observations (Rivera et al., 2013). Fish spermatozoa are immobile in the ejaculate, and it is only after suffering osmotic shock in water that they are activated and begin to move or swim (Alavi et al., 2009; Takei et al., 2012; Vladic and Jarvi, 2001). Spermatozoa are labile cells and are affect by aquatic contamination (Hatef et al., 2013). In the laboratory, one can replicate this effect using water from rivers or commer- cial solutions (Figueroa et al., 2013). Different kinetic parameters can be observed, including mobility time of the masses, either man- ually (Ubilla and Valdebenito, 2011) or by computerized systems (Hu et al., 2013). This is a way of measuring their cellular function http://dx.doi.org/10.1016/j.aquatox.2015.03.022 0166-445X/© 2015 Elsevier B.V. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Wi
PVa
Nb
c
d
e
d
a
ARRAA
KDST
1
dfheIvaw
C
h0
Aquatic Toxicology 163 (2015) 102–108
Contents lists available at ScienceDirect
Aquatic Toxicology
j o ur na l ho me pag e: www.elsev ier .com/ locate /aquatox
ater contaminated with Didymosphenia geminata generates changesn Salmo salar spermatozoa activation times
amela Olivares a, Paola Orellana a, Guillermo Guerra a, Matías Peredo-Parada b,c,iviana Chavez d, Alfredo Ramirez e, Jorge Parodi a,∗
Laboratorio Fisiología de la Reproducción, Escuela de Medicina Veterinaria, Núcleo de Investigación en Producción Alimentaria, Facultad de Recursosaturales, Universidad Católica de Temuco, ChileDepartamento de Ingeniería en Obras Civiles, Universidad de Santiago de Chile, ChilePlataforma de Investigación en Ecohidrología y Ecohidráulica, EcoHyd Ltda, ChileLaboratorio de Investigación y Educación, Tonalli Ltda, ChileLaboratorio de Criobiología y Análisis de Funcionalidad Espermática. Instituto de Ciencia Animal, Facultad de Ciencias Veterinarias, Universidad Australe Chile, Valdivia, Chile
r t i c l e i n f o
rticle history:eceived 4 February 2015eceived in revised form 26 March 2015ccepted 28 March 2015vailable online 8 April 2015
eywords:idymoperm cellsoxity
a b s t r a c t
Didimosphenia geminata (“didymo”), has become a powerful and devastating river plague in Chile. Asystem was developed in D. geminata channels with the purpose evaluating the effects of water pollutedwith didymo on the activation of Atlantic salmon (Salmo salar) spermatozoa. Results indicate that semen,when activated with uncontaminated river water had an average time of 60 ± 21 s. When using Powermilt,(a commercial activator), times of 240 ± 21 s are achieved, while rivers contaminated with D. geminataachieve a motility time of 30 ± 12 s. Interestingly enough, the kinetic parameters of VSL, VCL and VAPshowed no significant changes under all of the conditions. Furthermore, the presence of D. geminatareduces activation time of the samples as the cells age, indicating increased effects in spermatozoa thatare conserved for more than 5 days. D. geminata has antioxidant content, represented by polyphenols;200 ppm of polyphenol were obtained in this study per 10 g of microalgae. Spermatozoa exposed to theseextracts showed a reduction in mobility time in a dose dependent manner, showing an IC50 of 15 ppm. The
results suggest an effect on spermatozoa activation, possibly due to the release of polyphenols present incontaminated rivers, facilitating the alteration of sperm motility times, without affecting the viability orkinetics of the cells. These findings have important implications for current policy regarding the controlof the algae. Current control measures focus on the number of visible species, and not on the compoundsthat they release, which this study shows, also have a problematic effect on salmon production.© 2015 Elsevier B.V. All rights reserved.
. Introduction
Didymosphenia geminata (D. geminata) is a unicellular benthiciatom. This microalgae known as “didymo” in Chile, has been
ound in the river waters of southern Chile (Riveraet al., 2013), andas been considered a plague in freshwater sources by the Gov-rnment of Chile and can affect fish population (Reid et al., 2012).nternational studies indicate that D. geminata alters the microen-
ironment, reduces the fish population (Clearwater et al., 2010)nd perturbs aquatic macro-vertebrate communities and drinkingater system filters (Bergey et al., 2010; Gillis and Chalifour, 2010;∗ Corresponding author at: Av. Las Mariposas S/N, Campus Dr. Rivas, Universidadatólica de Temuco, Temuco, Chile. Tel.: +56 45205564.
E-mail address: [email protected] (J. Parodi).
ttp://dx.doi.org/10.1016/j.aquatox.2015.03.022166-445X/© 2015 Elsevier B.V. All rights reserved.
Kilroy et al., 2009). Recently, a toxic effect of microalgae on com-munities in contaminated rivers was described (Larned and Kilroy,2014). The removal of the microalgae, also introduces damagesto rivers (Larned and Kilroy, 2014). In Chile, there are no stud-ies that confirm this hypothesis and we must control these effectswith laboratory research to support field observations (Rivera et al.,2013). Fish spermatozoa are immobile in the ejaculate, and it isonly after suffering osmotic shock in water that they are activatedand begin to move or swim (Alavi et al., 2009; Takei et al., 2012;Vladic and Jarvi, 2001). Spermatozoa are labile cells and are affectby aquatic contamination (Hatef et al., 2013). In the laboratory,one can replicate this effect using water from rivers or commer-
cial solutions (Figueroa et al., 2013). Different kinetic parameterscan be observed, including mobility time of the masses, either man-ually (Ubilla and Valdebenito, 2011) or by computerized systems(Hu et al., 2013). This is a way of measuring their cellular function
Toxico
avazdpIdotbe
AmTafts
2
2
itac
2
wrtdumti7ttfsw
2
wTaaGpviwiFfv
P. Olivares et al. / Aquatic
nd the effects of molecules within their function, as motility andiability are related (Parodi, 2014) and computer assisted semennalysis (CASA) can be used for evaluated toxic effect on spermato-oa (Kime et al., 1996). Microalgae belonging to the family of browniatom algae are rich in some molecules, such as antioxidants andolyphenols such as diadinoxhantina (Lohr and Wilhelm, 1999).
nterestingly, several researches describe the benefits of antioxi-ants and polyphenols in cellular models. However, its toxic effectsn cancer cells (Korkina et al., 2009) are also described, indicatinghat a certain dose may be lethal to cell groups. It should be remem-ered that the dosage perfectly defines the positive or negativeffect of a molecule in a cell model.
We evaluated the effect of D. geminata in the activation oftlantic salmon sperm. We used water that was contaminated withicroalgae to evaluate motility time, kinetics and cell viability.
he same tests were carried out with uncontaminated river waternd with Powermilt, a commercial activator. Polyphenols extractedrom the didymo were used to evaluate a possible mechanism ofhe effects observed from D. geminata on the spermatozoa functionstudied.
. Methods
.1. Collection of D. geminata samples
D. geminata was collected in the Futaleufu and Biobío riversn winter and spring of 2014. The samples were transported tohe laboratory in plastic, closed dark boxes, at 10 ◦C. River waternd substrate (river rocks) colonized by the microalgae were alsoollected.
.2. Protocol for maintenance of the D. geminata samples
Collected samples were kept in aquariums, closed recirculatingater systems called artificial rivers, for observation in the labo-
atory. The rocks contaminated with D. geminata were arranged inhe artificial rivers, adding 50% of the original river water, plus 50%istilled water, (total volume of 14 l) making sure to leave a col-mn of water of about 15 cm over the rocks. Artificial rivers wereaintained with an expanded polystyrene insulate cover and the
emperature was reduced to an average of 12 ◦C through a cool-ng gel system. The water was kept in constant flux, using a model1009 Plaset-Italy brand motor with 30 W potency. This device cap-ures water from the final part of the artificial river, and is connectedo a 1′′ diameter PVC pipe which returns to the origin of the riveror water discharge, achieving constant flow and aeration. Micro-copic and macroscopic changes in the aquarium with D. geminataere recorded daily.
.3. Mortality studies of D. geminata
The mortality of D. geminata was observed by visual inspectionith clear field microscopy, using an inverted Meijie (VT series,
echno Co., Ltd., Japan) microscope, observing the presence orbsence of granules within the cytoplasm, denominating thems granular forms (Root and O’Reilly, 2013). The numbers of D.eminata cells, through a 40× lens, were counted, recording theercentage of those containing granules within the cytoplasm as aiable form of indication. In order to improve the documentation inmages, a Nomarsky microscope at an objective of 40× (Olympus)
as used for observation of intracellular structures and compar-
son with the viability obtained using modified neutral red dye.or this, the samples were left in a 0.01% neutral red dye solutionor 10 min, which has been reported as a means of assessing theiability of the D. geminata. When a granular red coloration islogy 163 (2015) 102–108 103
observed, it is indicated as a viable form of D. geminata (Jellymanet al., 2010).
2.4. Extraction of polyphenols and confocal images
D. geminata samples were frozen with liquid nitrogen, sub-sequently ground and maintained at 4 ◦C. Soon after, 20 mL ofdeionized water was added and rupture was performed by ultra-sound with a Misonix XL2000 Series sonicator, 10 pulses of 30 sin 1 min intervals until decomposition of the complex for all ofthe samples. They were then incubated for 20 min at 30 ◦C, beingagitated and then filtered by gravity. Samples were then passedthrough a double gauze filter and a No. 2 (125 mm) Wartman asdescribed by Fernández et al. (2013). The final extract was sub-jected to a measurement of polyphenols. Folin & Ciocalteu reagentwas used; following the protocol described by Lowry at 1951 Opti-cal density was measured at 517 nm of the reaction. The sampleswere frozen and passed through HPLC to identify their profile.The protocol described by Lohr and Wilhelm (Lohr and Wilhelm,1999) was modified, using the extracts for measurement with aC18 (Macherey-Nagel, Duren, Germany) column, measuring reten-tion at 440 nm. Then, the presence of diadinoxantina was identifiedin the yellow fraction. The fresh samples were also observed in aspectral confocal Olympus microscope (Olympus Fluoview 1000,USA), the samples were observed from an excitation of 280 nm to680 nm, obtaining images of the stimulated structures. The imagesof the different channels and the DIC images were combined usingthe ImageJ program.
2.5. Collection and handling of semen samples
A selection was made of adult male Atlantic salmon, (Salmosalar) coming from fish farms associated with the Catholic Uni-versity of Temuco. Prior to the extraction of semen, fish wereanesthetized with BZ-20 at 0.015%. Then, through an abdominalmassage or “stripping”, the semen samples were extracted, whichwere maintained with oxygen, placed in hermetically sealed plasticcontainers and refrigerated. In the laboratory, semen was dilutedin a proportion of 1:2 (semen: sperm diluent), composed of 18.8 gNaCl/L CaCl2, 3H2O 2 g/L KCl 72 g/L, NaH2PO4, H2O 4.1 g/L, NaHCO31 g/L, MgSO4, 7H2O 2.3 g/L and glucose 1 g/L. All procedures wereperformed at 4 ◦C. The samples were agitated and aerated dailyby injecting compressed oxygen. For observation, samples weremounted on a slide, adding 10 �L of Atlantic salmon (S. salar) semenand quickly mounted on the NIKON Labophot 2 optical microscope.Once the samples were focused, 20 �L of well water, river water,contaminated river water, and water with the Powermilt activatorwhich was diluted with distilled water (1×), was applied. Protocolwere taken to examine the time from the beginning of sperm move-ment, until all cells in the visual field stopped moving, their kineticswere analyzed through a Computer Assisted Semen Analysis (CASA)system in the ImageJ program, curvilinear velocity (VCL), straightline velocity (VSL) and straight line velocity (VAP) are described.Through this image analysis, the kinetic values were quantifiedunder various conditions (Parodi et al., 2015). A video was madeunder a 10× zoom lens, as to generate a greater contrast with themicroscope in a filter phase, for the Nikon 10× lens and in the short-est possible time after the activation of sperm, in order to capturethe maximum amount of information. All solutions were kept at4 ◦C.
2.6. Statistical analysis
The results are presented as the average ± standard error mean(SEM). An ANOVA variance analysis was performed, comparing allobservations. A post-test was applied and the Bonferroni test was
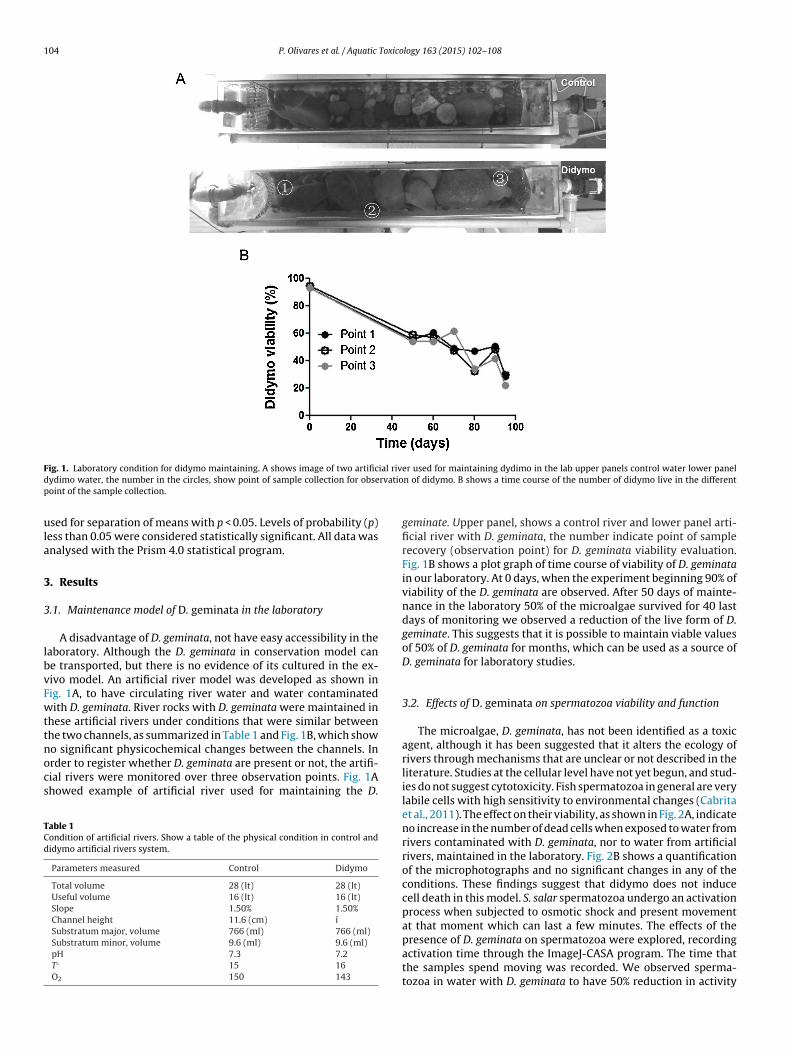
104 P. Olivares et al. / Aquatic Toxicology 163 (2015) 102–108
F ial rivd rvatiop
ula
3
3
lbvFwttnocs
TCd
ig. 1. Laboratory condition for didymo maintaining. A shows image of two artificydimo water, the number in the circles, show point of sample collection for obseoint of the sample collection.
sed for separation of means with p < 0.05. Levels of probability (p)ess than 0.05 were considered statistically significant. All data wasnalysed with the Prism 4.0 statistical program.
. Results
.1. Maintenance model of D. geminata in the laboratory
A disadvantage of D. geminata, not have easy accessibility in theaboratory. Although the D. geminata in conservation model cane transported, but there is no evidence of its cultured in the ex-ivo model. An artificial river model was developed as shown inig. 1A, to have circulating river water and water contaminatedith D. geminata. River rocks with D. geminata were maintained in
hese artificial rivers under conditions that were similar betweenhe two channels, as summarized in Table 1 and Fig. 1B, which show
o significant physicochemical changes between the channels. Inrder to register whether D. geminata are present or not, the artifi-ial rivers were monitored over three observation points. Fig. 1Ahowed example of artificial river used for maintaining the D.able 1ondition of artificial rivers. Show a table of the physical condition in control andidymo artificial rivers system.
Parameters measured Control Didymo
Total volume 28 (lt) 28 (lt)Useful volume 16 (lt) 16 (lt)Slope 1.50% 1.50%Channel height 11.6 (cm) íSubstratum major, volume 766 (ml) 766 (ml)Substratum minor, volume 9.6 (ml) 9.6 (ml)pH 7.3 7.2T◦ 15 16O2 150 143
er used for maintaining dydimo in the lab upper panels control water lower paneln of didymo. B shows a time course of the number of didymo live in the different
geminate. Upper panel, shows a control river and lower panel arti-ficial river with D. geminata, the number indicate point of samplerecovery (observation point) for D. geminata viability evaluation.Fig. 1B shows a plot graph of time course of viability of D. geminatain our laboratory. At 0 days, when the experiment beginning 90% ofviability of the D. geminata are observed. After 50 days of mainte-nance in the laboratory 50% of the microalgae survived for 40 lastdays of monitoring we observed a reduction of the live form of D.geminate. This suggests that it is possible to maintain viable valuesof 50% of D. geminata for months, which can be used as a source ofD. geminata for laboratory studies.
3.2. Effects of D. geminata on spermatozoa viability and function
The microalgae, D. geminata, has not been identified as a toxicagent, although it has been suggested that it alters the ecology ofrivers through mechanisms that are unclear or not described in theliterature. Studies at the cellular level have not yet begun, and stud-ies do not suggest cytotoxicity. Fish spermatozoa in general are verylabile cells with high sensitivity to environmental changes (Cabritaet al., 2011). The effect on their viability, as shown in Fig. 2A, indicateno increase in the number of dead cells when exposed to water fromrivers contaminated with D. geminata, nor to water from artificialrivers, maintained in the laboratory. Fig. 2B shows a quantificationof the microphotographs and no significant changes in any of theconditions. These findings suggest that didymo does not inducecell death in this model. S. salar spermatozoa undergo an activationprocess when subjected to osmotic shock and present movementat that moment which can last a few minutes. The effects of the
presence of D. geminata on spermatozoa were explored, recordingactivation time through the ImageJ-CASA program. The time thatthe samples spend moving was recorded. We observed sperma-tozoa in water with D. geminata to have 50% reduction in activity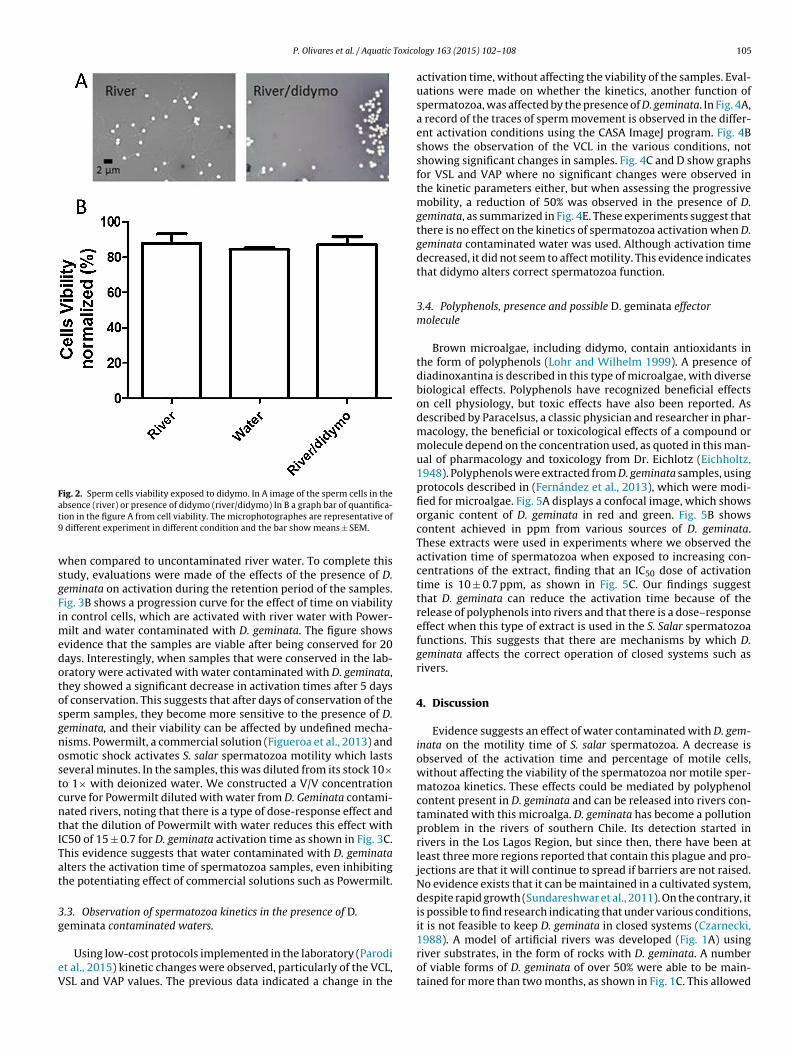
P. Olivares et al. / Aquatic Toxico
Fig. 2. Sperm cells viability exposed to didymo. In A image of the sperm cells in theat9
wsgFimedotosgnostcntITat
3g
eV
bsence (river) or presence of didymo (river/didymo) In B a graph bar of quantifica-ion in the figure A from cell viability. The microphotographes are representative of
different experiment in different condition and the bar show means ± SEM.
hen compared to uncontaminated river water. To complete thistudy, evaluations were made of the effects of the presence of D.eminata on activation during the retention period of the samples.ig. 3B shows a progression curve for the effect of time on viabilityn control cells, which are activated with river water with Power-
ilt and water contaminated with D. geminata. The figure showsvidence that the samples are viable after being conserved for 20ays. Interestingly, when samples that were conserved in the lab-ratory were activated with water contaminated with D. geminata,hey showed a significant decrease in activation times after 5 daysf conservation. This suggests that after days of conservation of theperm samples, they become more sensitive to the presence of D.eminata, and their viability can be affected by undefined mecha-isms. Powermilt, a commercial solution (Figueroa et al., 2013) andsmotic shock activates S. salar spermatozoa motility which lastseveral minutes. In the samples, this was diluted from its stock 10×o 1× with deionized water. We constructed a V/V concentrationurve for Powermilt diluted with water from D. Geminata contami-ated rivers, noting that there is a type of dose-response effect andhat the dilution of Powermilt with water reduces this effect withC50 of 15 ± 0.7 for D. geminata activation time as shown in Fig. 3C.his evidence suggests that water contaminated with D. geminatalters the activation time of spermatozoa samples, even inhibitinghe potentiating effect of commercial solutions such as Powermilt.
.3. Observation of spermatozoa kinetics in the presence of D.eminata contaminated waters.
Using low-cost protocols implemented in the laboratory (Parodit al., 2015) kinetic changes were observed, particularly of the VCL,SL and VAP values. The previous data indicated a change in the
logy 163 (2015) 102–108 105
activation time, without affecting the viability of the samples. Eval-uations were made on whether the kinetics, another function ofspermatozoa, was affected by the presence of D. geminata. In Fig. 4A,a record of the traces of sperm movement is observed in the differ-ent activation conditions using the CASA ImageJ program. Fig. 4Bshows the observation of the VCL in the various conditions, notshowing significant changes in samples. Fig. 4C and D show graphsfor VSL and VAP where no significant changes were observed inthe kinetic parameters either, but when assessing the progressivemobility, a reduction of 50% was observed in the presence of D.geminata, as summarized in Fig. 4E. These experiments suggest thatthere is no effect on the kinetics of spermatozoa activation when D.geminata contaminated water was used. Although activation timedecreased, it did not seem to affect motility. This evidence indicatesthat didymo alters correct spermatozoa function.
3.4. Polyphenols, presence and possible D. geminata effectormolecule
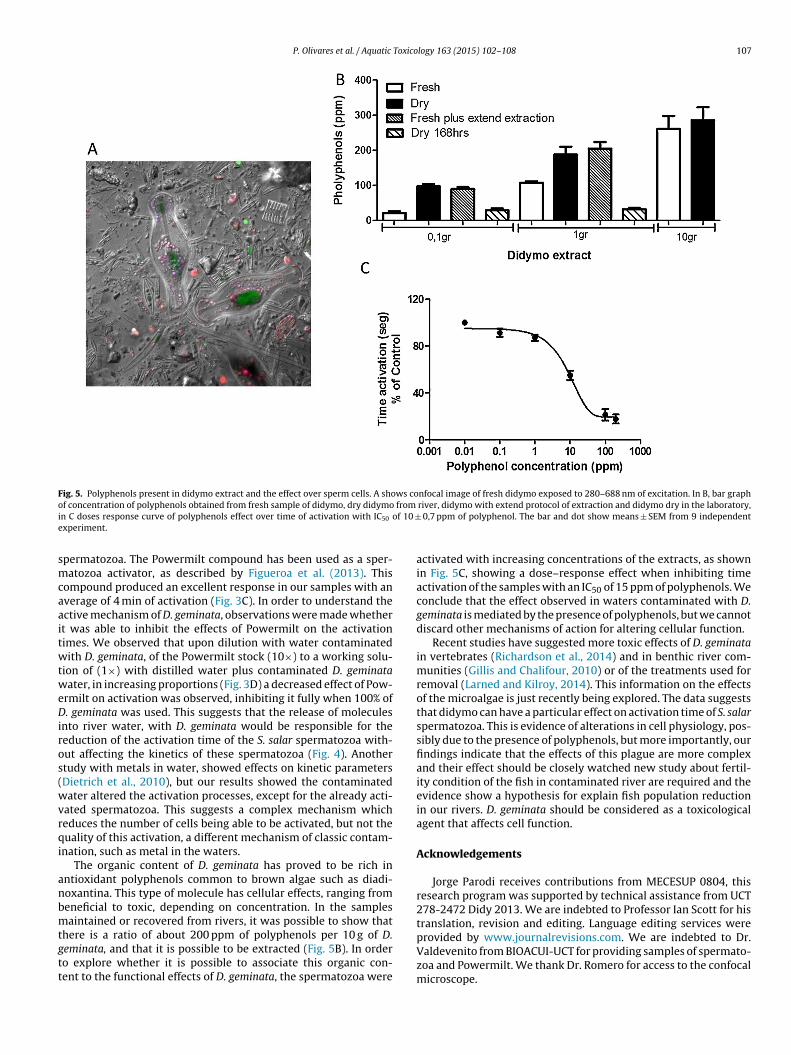
Brown microalgae, including didymo, contain antioxidants inthe form of polyphenols (Lohr and Wilhelm 1999). A presence ofdiadinoxantina is described in this type of microalgae, with diversebiological effects. Polyphenols have recognized beneficial effectson cell physiology, but toxic effects have also been reported. Asdescribed by Paracelsus, a classic physician and researcher in phar-macology, the beneficial or toxicological effects of a compound ormolecule depend on the concentration used, as quoted in this man-ual of pharmacology and toxicology from Dr. Eichlotz (Eichholtz,1948). Polyphenols were extracted from D. geminata samples, usingprotocols described in (Fernández et al., 2013), which were modi-fied for microalgae. Fig. 5A displays a confocal image, which showsorganic content of D. geminata in red and green. Fig. 5B showscontent achieved in ppm from various sources of D. geminata.These extracts were used in experiments where we observed theactivation time of spermatozoa when exposed to increasing con-centrations of the extract, finding that an IC50 dose of activationtime is 10 ± 0.7 ppm, as shown in Fig. 5C. Our findings suggestthat D. geminata can reduce the activation time because of therelease of polyphenols into rivers and that there is a dose–responseeffect when this type of extract is used in the S. Salar spermatozoafunctions. This suggests that there are mechanisms by which D.geminata affects the correct operation of closed systems such asrivers.
4. Discussion
Evidence suggests an effect of water contaminated with D. gem-inata on the motility time of S. salar spermatozoa. A decrease isobserved of the activation time and percentage of motile cells,without affecting the viability of the spermatozoa nor motile sper-matozoa kinetics. These effects could be mediated by polyphenolcontent present in D. geminata and can be released into rivers con-taminated with this microalga. D. geminata has become a pollutionproblem in the rivers of southern Chile. Its detection started inrivers in the Los Lagos Region, but since then, there have been atleast three more regions reported that contain this plague and pro-jections are that it will continue to spread if barriers are not raised.No evidence exists that it can be maintained in a cultivated system,despite rapid growth (Sundareshwar et al., 2011). On the contrary, itis possible to find research indicating that under various conditions,it is not feasible to keep D. geminata in closed systems (Czarnecki,
1988). A model of artificial rivers was developed (Fig. 1A) usingriver substrates, in the form of rocks with D. geminata. A numberof viable forms of D. geminata of over 50% were able to be main-tained for more than two months, as shown in Fig. 1C. This allowed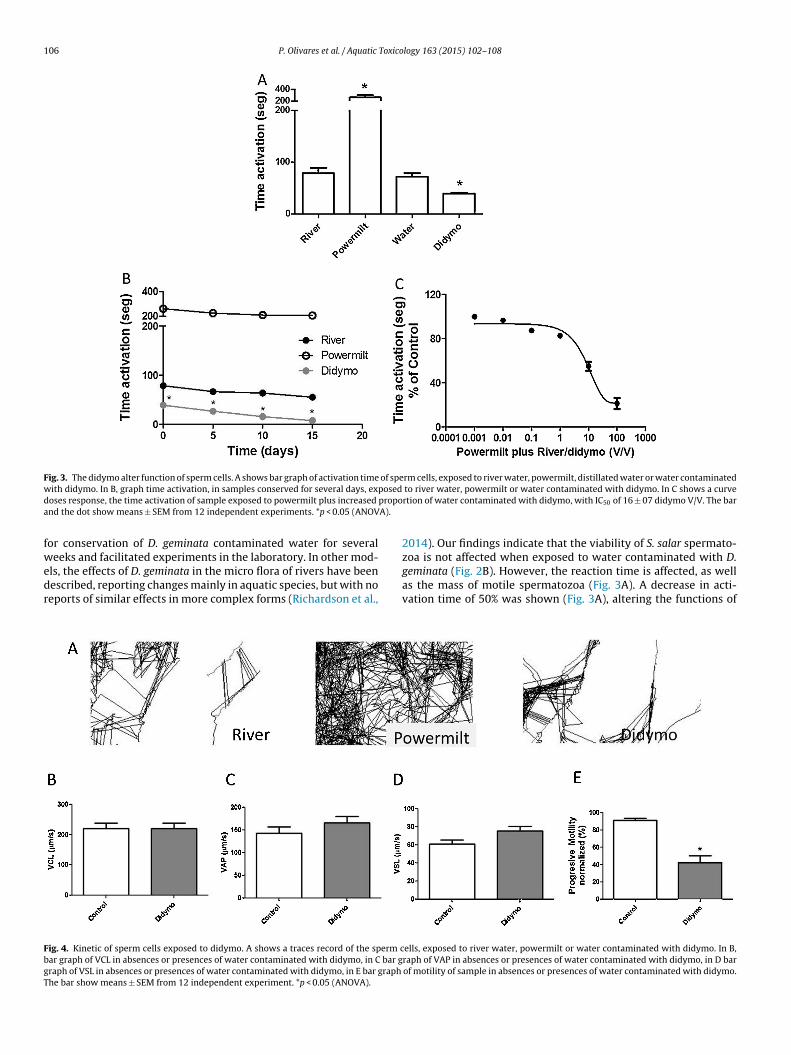
106 P. Olivares et al. / Aquatic Toxicology 163 (2015) 102–108
Fig. 3. The didymo alter function of sperm cells. A shows bar graph of activation time of sperm cells, exposed to river water, powermilt, distillated water or water contaminatedw posedd propoa A).
fwedr
FbgT
ith didymo. In B, graph time activation, in samples conserved for several days, exoses response, the time activation of sample exposed to powermilt plus increased
nd the dot show means ± SEM from 12 independent experiments. *p < 0.05 (ANOV
or conservation of D. geminata contaminated water for severaleeks and facilitated experiments in the laboratory. In other mod-
ls, the effects of D. geminata in the micro flora of rivers have beenescribed, reporting changes mainly in aquatic species, but with noeports of similar effects in more complex forms (Richardson et al.,
ig. 4. Kinetic of sperm cells exposed to didymo. A shows a traces record of the sperm car graph of VCL in absences or presences of water contaminated with didymo, in C bar graph of VSL in absences or presences of water contaminated with didymo, in E bar graph
he bar show means ± SEM from 12 independent experiment. *p < 0.05 (ANOVA).
to river water, powermilt or water contaminated with didymo. In C shows a curvertion of water contaminated with didymo, with IC50 of 16 ± 07 didymo V/V. The bar
2014). Our findings indicate that the viability of S. salar spermato-zoa is not affected when exposed to water contaminated with D.
geminata (Fig. 2B). However, the reaction time is affected, as wellas the mass of motile spermatozoa (Fig. 3A). A decrease in acti-vation time of 50% was shown (Fig. 3A), altering the functions ofells, exposed to river water, powermilt or water contaminated with didymo. In B,raph of VAP in absences or presences of water contaminated with didymo, in D barof motility of sample in absences or presences of water contaminated with didymo.
P. Olivares et al. / Aquatic Toxicology 163 (2015) 102–108 107
Fig. 5. Polyphenols present in didymo extract and the effect over sperm cells. A shows confocal image of fresh didymo exposed to 280–688 nm of excitation. In B, bar grapho from
i of 10 ±e
smcaaitwtweDiros(wvrqi
anbmtgtt
f concentration of polyphenols obtained from fresh sample of didymo, dry didymon C doses response curve of polyphenols effect over time of activation with IC50
xperiment.
permatozoa. The Powermilt compound has been used as a sper-atozoa activator, as described by Figueroa et al. (2013). This
ompound produced an excellent response in our samples with anverage of 4 min of activation (Fig. 3C). In order to understand thective mechanism of D. geminata, observations were made whethert was able to inhibit the effects of Powermilt on the activationimes. We observed that upon dilution with water contaminatedith D. geminata, of the Powermilt stock (10×) to a working solu-
ion of (1×) with distilled water plus contaminated D. geminataater, in increasing proportions (Fig. 3D) a decreased effect of Pow-
rmilt on activation was observed, inhibiting it fully when 100% of. geminata was used. This suggests that the release of molecules
nto river water, with D. geminata would be responsible for theeduction of the activation time of the S. salar spermatozoa with-ut affecting the kinetics of these spermatozoa (Fig. 4). Anothertudy with metals in water, showed effects on kinetic parametersDietrich et al., 2010), but our results showed the contaminatedater altered the activation processes, except for the already acti-
ated spermatozoa. This suggests a complex mechanism whicheduces the number of cells being able to be activated, but not theuality of this activation, a different mechanism of classic contam-
nation, such as metal in the waters.The organic content of D. geminata has proved to be rich in
ntioxidant polyphenols common to brown algae such as diadi-oxantina. This type of molecule has cellular effects, ranging fromeneficial to toxic, depending on concentration. In the samplesaintained or recovered from rivers, it was possible to show that
here is a ratio of about 200 ppm of polyphenols per 10 g of D.eminata, and that it is possible to be extracted (Fig. 5B). In ordero explore whether it is possible to associate this organic con-ent to the functional effects of D. geminata, the spermatozoa were
river, didymo with extend protocol of extraction and didymo dry in the laboratory, 0,7 ppm of polyphenol. The bar and dot show means ± SEM from 9 independent
activated with increasing concentrations of the extracts, as shownin Fig. 5C, showing a dose–response effect when inhibiting timeactivation of the samples with an IC50 of 15 ppm of polyphenols. Weconclude that the effect observed in waters contaminated with D.geminata is mediated by the presence of polyphenols, but we cannotdiscard other mechanisms of action for altering cellular function.
Recent studies have suggested more toxic effects of D. geminatain vertebrates (Richardson et al., 2014) and in benthic river com-munities (Gillis and Chalifour, 2010) or of the treatments used forremoval (Larned and Kilroy, 2014). This information on the effectsof the microalgae is just recently being explored. The data suggeststhat didymo can have a particular effect on activation time of S. salarspermatozoa. This is evidence of alterations in cell physiology, pos-sibly due to the presence of polyphenols, but more importantly, ourfindings indicate that the effects of this plague are more complexand their effect should be closely watched new study about fertil-ity condition of the fish in contaminated river are required and theevidence show a hypothesis for explain fish population reductionin our rivers. D. geminata should be considered as a toxicologicalagent that affects cell function.
Acknowledgements
Jorge Parodi receives contributions from MECESUP 0804, thisresearch program was supported by technical assistance from UCT278-2472 Didy 2013. We are indebted to Professor Ian Scott for histranslation, revision and editing. Language editing services were
provided by www.journalrevisions.com. We are indebted to Dr.Valdevenito from BIOACUI-UCT for providing samples of spermato-zoa and Powermilt. We thank Dr. Romero for access to the confocalmicroscope.
1 Toxico
R
A
B
C
C
C
D
E
F
G
H
H
J
F
08 P. Olivares et al. / Aquatic
eferences
lavi, S.M., Rodina, M., Viveiros, A.T., Cosson, J., Gela, D., Boryshpolets, S., Linhart,O., 2009. Effects of osmolality on sperm morphology: motility and flagellarwave parameters in Northern pike (Esox lucius L.). Theriogenology 72, 32–43.
ergey, E.A., Cooper, J.T., Phillips, B.C., 2010. Substrate characteristics affectcolonization by the bloom-forming diatom Didymosphenia geminata. Aquat.Ecol. 44, 33–40.
abrita, E., Ma, S., Diogo, P., Martinez-Paramo, S., Sarasquete, C., Dinis, M.T., 2011.The influence of certain aminoacids and vitamins on post-thaw fish spermmotility: viability and DNA fragmentation. Anim. Reprod. Sci. 125, 189–195.
learwater, S.J., Jellyman, P.G., Biggs, B.J., Hickey, C.W., Blair, N., Clayton, J.S., 2010.Pulse-dose application of chelated copper to a river for Didymospheniageminata control: effects on macroinvertebrates and fish. Environ. Toxicol.Chem. 30, 181–195.
zarnecki, D.B., 1988. The Loras college non-diatom fresh-water algal culturecollection. Cryptogamie Algol. 9, 203–209.
ietrich, G.J., Dietrich, M., Kowalski, R.K., Dobosz, S., Karol, H., Demianowicz, W.,Glogowski, J., 2010. Exposure of rainbow trout milt to mercury and cadmiumalters sperm motility parameters and reproductive success. Aquat. Toxicol. 97,277–284.
ichholtz, F., 1948. Pharmacology and toxicology. Off. of Military Govt. forGermany, Field Information Agencies Technical, British, French, U.S., n. p.
igueroa, E., Risopatron, J., Sanchez, R., Isachenko, E., Merino, O., Isachenko, V.,Valdebenito, I., 2013. Spermatozoa vitrification of sex-reversed rainbow trout(Oncorhynchus mykiss): effect of seminal plasma on physiological parameters.Aquaculture 372, 119–126.
illis, C.A., Chalifour, M., 2010. Changes in the macrobenthic community structurefollowing the introduction of the invasive algae Didymosphenia geminata in theMatapedia River (Qubec, Canada). Hydrobiologia 647, 63–70.
atef, A., Alavi, S.M.H., Golshan, M., Linhart, O., 2013. Toxicity of environmentalcontaminants to fish spermatozoa function in vitro–a review. Aquat. Toxicol.140–141, 134–144.
u, Y.A., Lu, J.C., Shao, Y., Huang, Y.F., Lu, N.Q., 2013. Comparison of the semenanalysis results obtained from two branded computer-aided sperm analysissystems. Andrologia 45, 315–318.
ellyman, P.G., Clearwater, S.J., Clayton, J.S., Kilroy, C., Blair, N., Hickey, C.W., Biggs,B.J., 2010. Controlling the invasive diatom Didymosphenia geminata: an
ecotoxicity assessment of four potential biocides. Arch. Environ. Contam.Toxicol. 61, 115–127.ernández, I.J., Navarrete, P., Parodi, J., Romero, F., Salvatici, R.S., 2013. Chileancrude extract of Ruta graveolens generates vasodilatation in rat aorta atsubtoxic cellular concentrations. Adv. Biosci. Biotechnol. 04, 29–36.
logy 163 (2015) 102–108
Kilroy, C., Larned, S.T., Biggs, B.J.F., 2009. The non-indigenous diatomDidymosphenia geminata alters benthic communities in New Zealand rivers.Freshwater Biol. 54, 1990–2002.
Kime, D.E., Ebrahimi, M., Nysten, K., Roelants, I., Rurangwa, E., Moore, H.D.M.,Ollevier, F., 1996. Use of computer assisted sperm analysis (CASA) formonitoring the effects of pollution on sperm quality of fish; application to theeffects of heavy metals. Aquat. Toxicol. 36, 223–237.
Korkina, L.G., De Luca, C., Kostyuk, V.A., Pastore, S., 2009. Plant polyphenols andtumors from mechanisms to therapies, prevention, and protection againsttoxicity of anti-cancer treatments. Curr. Med. Chem. 16, 3943–3965.
Larned, S.T., Kilroy, C., 2014. Effects of Didymosphenia geminata removal on rivermacroinvertebrate communities. J. Freshwater Ecol. 29, 345–362.
Lohr, M., Wilhelm, C., 1999. Algae displaying the diadinoxanthin cycle also possessthe violaxanthin cycle. Proc. Natl. Acad. Sci. U. S. A. 96, 8784–8789.
Parodi, J., 2014. Motility viability, and calcium in the sperm cells. Syst. Biol. Reprod.Med. 60, 65–71.
Parodi, J., Ramírez-Reveco, A., Guerra, G., 2015. Example use of low-cost system forcapturing the kinetic parameters of sperm cells in Atlantic Salmon (Salmosalar). Adv. Biosci. Biotechnol. 06, 63–72.
Reid, B.L., Hernandez, K.L., Frangopulos, M., Bauer, G., Lorca, M., Kilroy, C.,Spaulding, S., 2012. The invasion of the freshwater diatom Didymospheniageminata in Patagonia: prospects strategies, and implications for biosecurity ofinvasive microorganisms in continental waters. Conserv. Lett. 5, 432–440.
Richardson, D., Oleksy, I., Hoellein, T., Arscott, D., Gibson, C., Root, S., 2014. Habitatcharacteristics, temporal variability, and macroinvertebrate communitiesassociated with a mat-forming nuisance diatom (Didymosphenia geminata) inCatskill mountain streams. N. Y. Aquat. Sci. 76, 553–564.
Rivera, P., Basualto, S., Cruces, F., 2013. On the diatom Didymosphenia geminata(Lyngbye) M. Schmidt its morphology and distribution in Chile. Gayana Bot. 70,154–158.
Root, S., O’Reilly, C.M., 2013. Didymo control: increasing the effectiveness ofdecontamination strategies and reducing spread. Fisheries 37, 440–448.
Sundareshwar, P.V., Upadhayay, S., Abessa, M., Honomichl, S., Berdanier, B.,Spaulding, S.A., Sandvik, C., Trennepohl, A., 2011. Didymosphenia geminata:algal blooms in oligotrophic streams and rivers. Geophys. Res. Lett. 38, 1–5.
Takei, G.L., Mukai, C., Okuno, M., 2012. Transient Ca2+ mobilization caused byosmotic shock initiates salmonid fish sperm motility. J. Exp. Biol. 215, 630–641.
Ubilla, A., Valdebenito, I.N., 1792. Use of antioxidants on rainbow trout
Oncorhynchus mykiss (Walbaum, sperm diluent: effects on motility andfertilizing capability. Lat. Am. J. Aquat. Res. 39, 338–343.Vladic, T.V., Jarvi, T., 2001. Sperm quality in the alternative reproductive tactics ofAtlantic salmon: the importance of the loaded raffle mechanism. Proc. Biol. Sci.268, 2375–2381.
Related Documents











