Roux's Arch Dev Biol (1987) 196:391 396 Roux'sArchivesof Developmental motogy Springer-Verlag 1987 Voltage gradients and microtubules both involved in intercellular protein and mitochondria transport in the telotrophic ovariole of Dysdercus intermedius Axel NIiinz and Frank Dittmann Institut fiir Biologic III (Zoologie/Entwicklungsphysiologie), Universit/it Tfibingen, Auf der Morgenstelle 28, D-7400 Tfibingen, Federal Republic of Germany Summary. In the telotrophic ovariole of Dysdercus interme- dius the intercellular transport consists of different subsys- tems. Microinjection of FITC-labeled slowly diffusing pro- teins with opposite electrical net charges and of mitochon- dria was used to study the translocation of macromolecules and organelles, a) By intracellular measurements a voltage gradient of about 4 mV between the tropharium as the more negative side and the previtellogenic oocytes could be dem- onstrated, b) After injection into the tropharium negatively charged proteins migrated according to the electropotential gradient via the trophic cords into the oocytes. Positively charged proteins, however, were retained in the tropharium. c) After injection into previtellogenic oocytes both negative- ly and positively charged proteins moved into the trophic cords. Thus, the effectiveness of the electropotential gra- dient on the distribution of charged proteins is more pro- nounced from the tropharium side. d) Mitochondria micro- injected into the trophic core were probably aligned along microtubules and translocated towards the trophic cords. These results suggest that in the telotrophic bug ovariole a number of intercellular transport subsystems contribute to provide previtellogenic oocytes with nurse cells products. An electrophoretic transport mechanism for soluble pro- teins acting especially within the tropharium and a microtu- bule-associated transport for mitochondria could be evi- denced. Key words: Intercellular transport mechanisms - Voltage gradient electrophoresis Microtubule-associated discharge - Insect telotrophic ovariole - Dysdercus intermedius (He- teroptera) Introduction Growing insect oocytes accumulate large amounts of mito- chondria, ribosomes, tRNA and mRNA for protein synthe- sis during embryogenesis (for review see Berry 1985). In meroistic ovaries RNA is almost exclusively synthesized by the highly endopolyploid trophocytes (Dittmann etal. 1984) and transported into the oocyte. Trophocyte products Offprint requests to." F. Dittmann, LS Entwicklungsphysiologie, Auf der Morgenstelle 28, D-7400 Tfibingen pass through intercellular bridges in polytrophic follicles and through trophic cords in telotrophic ovarioles (Bier 1967; Engels 1969; Telfer 1975; King and Brining 1985). Maternal RNA stored in the ooplasm plays a key role as cytoplasmic determinants for early embryonic development (Sander 1984). A well known example for an ooplasmic factor defined in the posterior pole plasm since late oogene- sis concerns the induction of germ cells (Illmensee et al. 1976). The discussion about the intercellular transport mecha- nisms acting in insect ovaries began more than 20 years ago (Bier 1965) and is still continued rather contradictory (Gutzeit 1986; de Loof 1986). The most fascinating hypoth- esis is the electrophoresis model proposed by Woodruff and Telfer (1973, 1974, 1980) who reported an electrical poten- tial gradient of 6 to 10 mV between trophocytes and oocyte in the polytrophic follicle of HyaIophora cecropia. In telotrophic ovarioles the distance of transport is sub- stantially longer than in polytrophic follicles, up to 1.5 mm in Dysdercus intermedius compared to less than 0.05 mm in H. cecropia (Telfer et al. 1981). Therefore, a steeper volt- age gradient would be required for intercellular electropho- resis. Up to now only extracellular currents were measured in bug ovarioles (Dittmann et aL 1981). Telfer et al. (1981) and Woodruff and Anderson (1984) demonstrated charge dependent translocafion of microinjected proteins in the telotrophic ovarioles of Rhodnius prolixus and Oncopeltus fasciatus, but this was restricted to the tropharium. Further- more, organelle transport has not yet been studied experi- mentally, although a mass flow ofmitochondria originating from nurse cells has been observed passing intercellular bridges (Ramamurty 1977) as well as trophic cords (Mays 1967). This study presents data on the intracellular electrical potential gradient and the tranpsort of microinjected pro- teins and mitochondria in the telotrophic ovariole of Dys- dercus intermedius. Materials and methods Animals. Our laboratory stock of Dysdercus intermedius (Heteroptera: Pyrrhocoridae) was reared at 23•176C and 70% to 80% relative humidity under long day conditions, 14 h L/10 h D. The bugs were fed on freshly swollen cotton seeds. Ovarioles were dissected from females 3 to 5 days

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Roux's Arch Dev Biol (1987) 196:391 396 Roux's Archives of
Developmental motogy �9 Springer-Verlag 1987
Voltage gradients and microtubules both involved in intercellular protein and mitochondria transport in the telotrophic ovariole of Dysdercus intermedius Axel NIiinz and Frank Dittmann Institut fiir Biologic III (Zoologie/Entwicklungsphysiologie), Universit/it Tfibingen, Auf der Morgenstelle 28, D-7400 Tfibingen, Federal Republic of Germany
Summary. In the telotrophic ovariole of Dysdercus interme- dius the intercellular transport consists of different subsys- tems. Microinjection of FITC-labeled slowly diffusing pro- teins with opposite electrical net charges and of mitochon- dria was used to study the translocation of macromolecules and organelles, a) By intracellular measurements a voltage gradient of about 4 mV between the tropharium as the more negative side and the previtellogenic oocytes could be dem- onstrated, b) After injection into the tropharium negatively charged proteins migrated according to the electropotential gradient via the trophic cords into the oocytes. Positively charged proteins, however, were retained in the tropharium. c) After injection into previtellogenic oocytes both negative- ly and positively charged proteins moved into the trophic cords. Thus, the effectiveness of the electropotential gra- dient on the distribution of charged proteins is more pro- nounced from the tropharium side. d) Mitochondria micro- injected into the trophic core were probably aligned along microtubules and translocated towards the trophic cords.
These results suggest that in the telotrophic bug ovariole a number of intercellular transport subsystems contribute to provide previtellogenic oocytes with nurse cells products. An electrophoretic transport mechanism for soluble pro- teins acting especially within the tropharium and a microtu- bule-associated transport for mitochondria could be evi- denced.
Key words: Intercellular transport mechanisms - Voltage gradient electrophoresis Microtubule-associated discharge - Insect telotrophic ovariole - Dysdercus intermedius (He- teroptera)
Introduction
Growing insect oocytes accumulate large amounts of mito- chondria, ribosomes, tRNA and mRNA for protein synthe- sis during embryogenesis (for review see Berry 1985). In meroistic ovaries RNA is almost exclusively synthesized by the highly endopolyploid trophocytes (Dittmann etal. 1984) and transported into the oocyte. Trophocyte products
Offprint requests to." F. Dittmann, LS Entwicklungsphysiologie, Auf der Morgenstelle 28, D-7400 Tfibingen
pass through intercellular bridges in polytrophic follicles and through trophic cords in telotrophic ovarioles (Bier 1967; Engels 1969; Telfer 1975; King and Brining 1985). Maternal RNA stored in the ooplasm plays a key role as cytoplasmic determinants for early embryonic development (Sander 1984). A well known example for an ooplasmic factor defined in the posterior pole plasm since late oogene- sis concerns the induction of germ cells (Illmensee et al. 1976).
The discussion about the intercellular transport mecha- nisms acting in insect ovaries began more than 20 years ago (Bier 1965) and is still continued rather contradictory (Gutzeit 1986; de Loof 1986). The most fascinating hypoth- esis is the electrophoresis model proposed by Woodruff and Telfer (1973, 1974, 1980) who reported an electrical poten- tial gradient of 6 to 10 mV between trophocytes and oocyte in the polytrophic follicle of HyaIophora cecropia.
In telotrophic ovarioles the distance of transport is sub- stantially longer than in polytrophic follicles, up to 1.5 mm in Dysdercus intermedius compared to less than 0.05 mm in H. cecropia (Telfer et al. 1981). Therefore, a steeper volt- age gradient would be required for intercellular electropho- resis. Up to now only extracellular currents were measured in bug ovarioles (Dittmann et aL 1981). Telfer et al. (1981) and Woodruff and Anderson (1984) demonstrated charge dependent translocafion of microinjected proteins in the telotrophic ovarioles of Rhodnius prolixus and Oncopeltus fasciatus, but this was restricted to the tropharium. Further- more, organelle transport has not yet been studied experi- mentally, although a mass flow ofmitochondria originating from nurse cells has been observed passing intercellular bridges (Ramamurty 1977) as well as trophic cords (Mays 1967). This study presents data on the intracellular electrical potential gradient and the tranpsort of microinjected pro- teins and mitochondria in the telotrophic ovariole of Dys- dercus intermedius.
Materials and methods
Animals. Our laboratory stock of Dysdercus intermedius (Heteroptera: Pyrrhocoridae) was reared at 23•176 C and 70% to 80% relative humidity under long day conditions, 14 h L/10 h D. The bugs were fed on freshly swollen cotton seeds. Ovarioles were dissected from females 3 to 5 days

392
after the adult moult in a Ringer medium containing 40 mM NaC1, 2 0 m M KC1, 15 mM MgC12, t mM CaC12 and 1.7 mM Pipes, pH 6.8. Ovarioles cultured in this medium continue to incorporate yolk into vitellogenic oocytes for many hours (Dittmann et al. 1987 a) indicating normal cel- lular functions during the incubation periods used here.
Fluorescent transport probes. Fluorescein-labeled lysozyme (Fly(+)) and methylcarboxylated lysozyme (McFly ( - ) ) were kindly provided by Dr. Woodruff (West Chester, PA, cf. Woodruff et al. 1986). Mitochondria were isolated by differential centrifugation at 4~ from mouse liver (Schnaitman and Greenawalt 1968) and FITC (Sigma, F- 7250) labeled by coupling in 5 mM Na-barbital buffer pH 7.3 for 12 h at 4~ (modified after Goldman 1968). Unbound dye was removed by centrifugation three times at 6500 g for 5 min. The mitochondria were resuspended in isolation medium (220 mM mannitol, 50 mM sucrose, I mM EDTA, 0.1 mM EGTA, 3 mM HEPES). Immediate- ly before injection the mitochondria were centrifuged again and Ringer medium was added until all mitochondria were suspended.
Electrophysiological measurements and microinjection. Intra- cellular measurements were carried out with capillary silver/ silverchloride microelectrodes filled with 3 M KC1 (Wood- ruff 1979). The tip resistance was higher than 10 Ms Ap- proximately 50 pl of fluorescent protein solution were in- jected marginally into the tropharium between the tropho- cytes (Fig. 1 b), and about 10 pl containing 2000-3000 fluo- rescent mitochondria into the trophic core. Microinjections were carried out under microscopic control with an Eppen- doff 4252 unit using pneumatic pressure. All experiments were carried out at room temperature. The number of re- peats is given together with the results.
Sectioning and fluorescence microscopy. After fixation at 4 ~ C in 0.1 mM phosphate buffer pH 7.4 containing 4% formaldehyde the ovarioles were dehydrated in ethanol, em- bedded in glycol-methacrylate (Historesin TM, LKB), and sectioned on a Jung (Heidelberg, FRG) Autocut 1140 mi- crotome. For all observations a Zeiss microscope equipped for epifluorescence using the filter combination for FITC was used.
Results
Electropotential gradient
Intracellular measurements were performed to determine potential differences between different parts of the ovariole and the in vitro Ringer medium. The tropharium potential averaged - 30.07 mV (SD = 1.06 mV, n = 11) and in case of previtellogenic oocytes -26 .40 mV (SD = 1.78 mV, n = 36). The individual tropharium showing the lowest mem- brane potential of -28 .53 mV was connected with 3 previ- tellogenic follicles characterized by membrane potentials of 25.68 mV (SD= 1.92 mV, n=3) . From the differences in membrane potential an average electropotential gradient between tropharium and previtellogenic oocytes of 3.67 mV can be calculated with the tropharium always being more negative.
Protein translocation
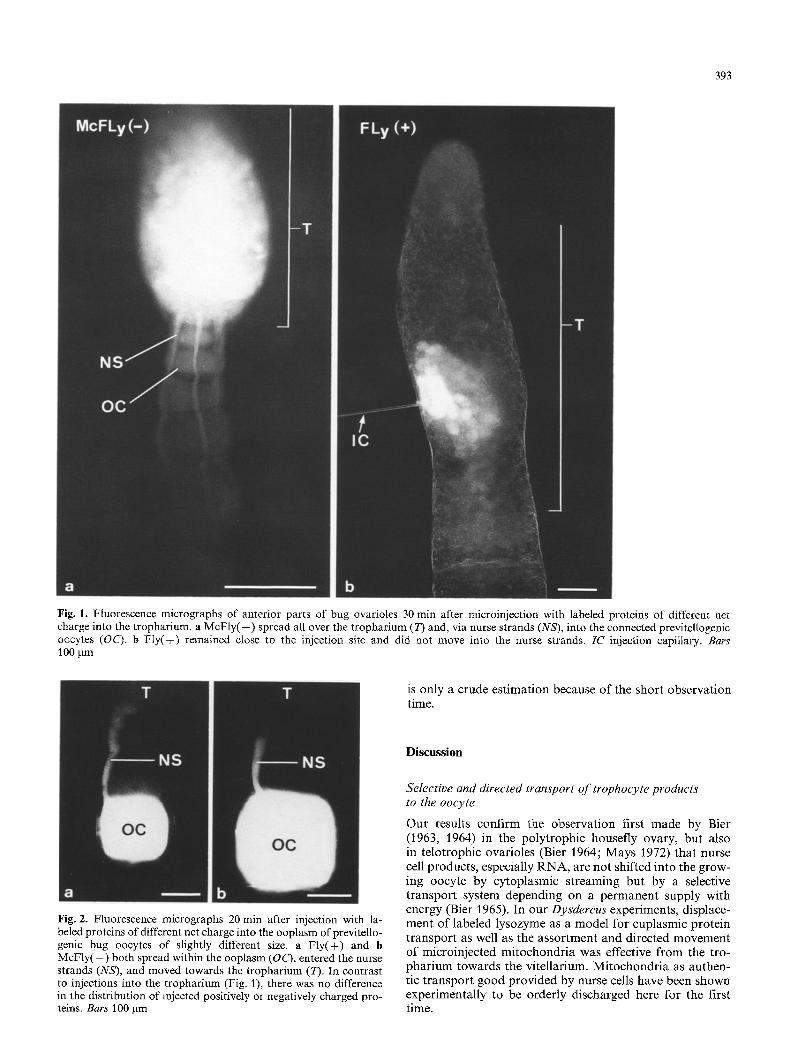
The shift of microinjected fluorescent labeled proteins could be observed in the living ovariole during experimental incu- bation. After injection of M c F l y ( - ) into the tropharium, fluorescence spread throughout the tropharium within 3 to 6 rain and entered the trophic cords in all the 30 experi- ments. Because the labeled proteins moved with a diffuse rather than with a sharp front, we only can give a rough estimation of the transport velocity. Within 15 to 20 min after injection the trophic cords had become visible in a length of 0.2 to 0.3 mm and M c F l y ( - ) had entered the 4 or 5 previtellogenic oocytes closest to the tropharium (Fig. l a). This corresponds to a protein movement of 15-25 gin/rain or 0.9-1.5 mm/h. More distant oocytes did not become fluorescent, even within longer incubations. In contrast to this, after injections of Fly(+ ) into the trophar- ium fluorescence never entered the trophic cords in any of 10 experiments. The label remained close to the injection site and only reached adjacent trophocytes (Fig. 1 b).
Microinjection of previtellogenic oocytes resulted in a uniform displacement of M c F l y ( - ) and Fly( + ). Both posi- tively (Fig. 2a) and negatively (Fig. 2b) charged proteins spread throughout the oocyte and after 10 to 15 rain entered the trophic cord. In the following 15 rain the fluorescence slowly migrated into the nurse strands. When exactly the labeled proteins even reached the tropharium could not be determined because, after 30 to 40 min, the fluorescence of both proteins began to fade, and in 20% of the experi- ments faded away completely, before leaving the nurse strands and entering the trophic core region.
Mitochondria movement
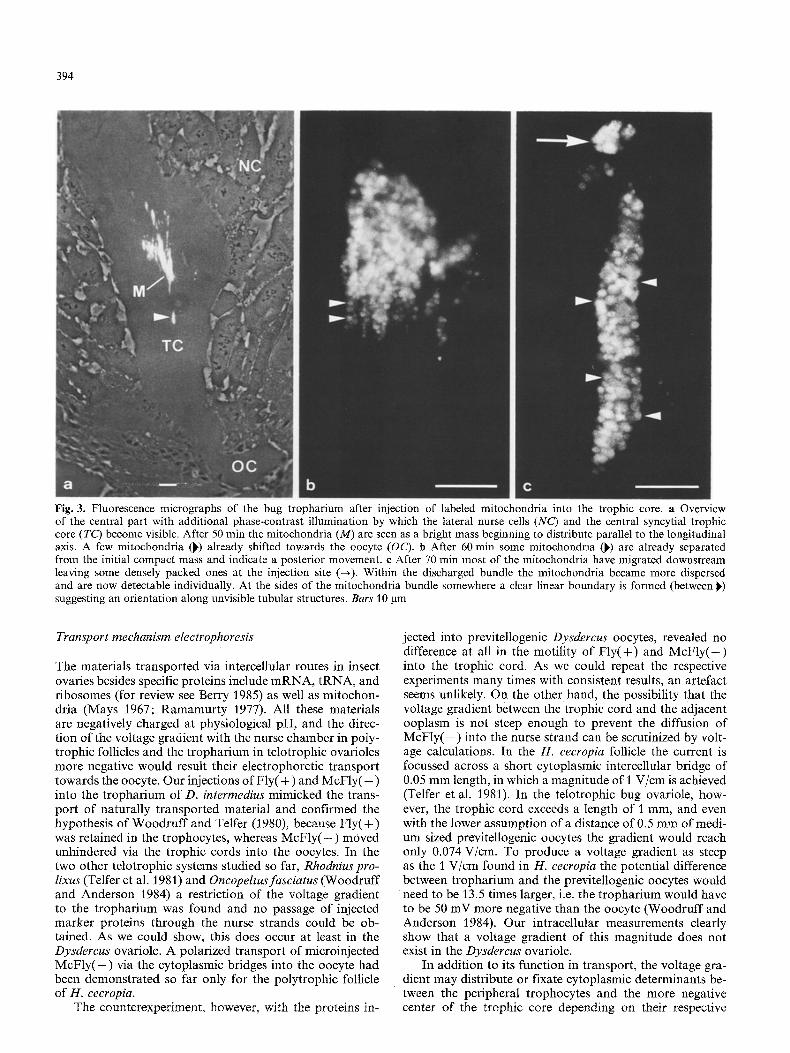
Immediately after injection the mitochondria were densely packed, but later on became more dispersed. Fluorescent labeled mitochondria were not visible in the living ovariole or in whole mounts apparently because the amount of mate- rial was too small. In sections of ovarioles that had been incubated for 30 to 45 rain, however, the fluorescent mito- chondria could be visualized in the tropharium as a disor- ganized cluster. After 50 min of incubation (Fig. 3 a) single mitochondria were found far posterior from the injection site. After 60 min several mitochondria seemed to be orga- nized in parallel longitudinal rows (Fig. 3 b). After 70 min most of the mitochondria formed a bundle (Fig. 3c) ex- tended downstream in the trophic core. In some of the sections the mitochondria were arranged with rather clear outlines along an unvisible elongated structure within the central core (Fig. 3 c). After longer times of incubation the mitochondria appeared separated so that nearly each mito- chondrium was individually distinguishable.
The ovarioles were incubated up to 3 1/2 h to examine large distance transport of the injected mitochondria and to evaluate the transport velocity. However, the mitochon- dria were visible only until about 2 h after injection. This time was not long enough to have entered the nurse strands. The site of injection into the trophic core could be localized even after more than I h by the presence of remains of mitochondria which were not transported and therefore maintained the densely packed configuration (Fig. 3 c). An estimation of the transport velocity within the trophic core was based on serial sections with such localizable injection sites. A value of 1.5 la/s or 0.08 mm/h was calculated which
393
Fig. 1. Fluorescence micrographs of anterior parts of bug ovarioles 30 min after microinjection with labeled proteins of different net charge into the tropharium, a McF ly ( - ) spread all over the tropharium (T) and, via nurse strands (NS), into the connected previteUogenic oocytes (OC). b F ly(+) remained close to the injection site and did not move into the nurse strands. IC injection capillary. Bars 100 ~m
is only a crude est imat ion because of the short observat ion time.
Discus~on
Fig. 2. Fluorescence micrographs 20 rain after injection with la- beled proteins of different net charge into the ooplasm of previtello- genic bug oocytes of slightly different size. a Fly(+) and b McFly(--) both spread within the ooplasm (OC), entered the nurse strands (NS), and moved towards the tropharium (T). In contrast to injections into the tropharium (Fig. 1), there was no difference in the distribution of injected positively or negatively charged pro- teins. Bars 100 Bm
Selective and directed transport o f trophocyte products to the oocyte
Our results confirm the observat ion first made by Bier (1963, 1964) in the poly t rophic housefly ovary, but also in te lotrophic ovarioles (Bier 1964; Mays 1972) that nurse cell products , especially R N A , are not shifted into the grow- ing oocyte by cytoplasmic streaming but by a selective t ranspor t system depending on a permanent supply with energy (Bier 1965). In our Dysdereus experiments, displace- ment o f labeled lysozyme as a model for euplasmic protein t ranspor t as well as the assor tment and directed movement of microinjected mi tochondr ia was effective from the tro- phar ium towards the vitellarium. Mi tochondr ia as authen- tic t ranspor t good provided by nurse cells have been shown experimental ly to be orderly discharged here for the first time.
394
Fig. 3. Fluorescence micrographs of the bug tropharium after injection of labeled mitochondria into the trophic core. a Overview of the central part with additional phase-contrast illumination by which the lateral nurse cells (NC) and the central syncytial trophic core (TC) become visible. After 50 min the mitochondria (M) are seen as a bright mass beginning to distribute parallel to the longitudinal axis. A few mitochondria (~) already shifted towards the oocyte (OC). b After 60 min some mitochondria (~) are already separated from the initial compact mass and indicate a posterior movement, e After 70 min most of the mitochondria have migrated downstream leaving some densely packed ones at the injection site (--,). Within the discharged bundle the mitochondria became more dispersed and are now detectable individually. At the sides of the mitochondria bundle somewhere a clear linear boundary is formed (between ~) suggesting an orientation along unvisible tubular structures. Bars t0 gm
Transport mechanism electrophoresis
The materials transported via intercellular routes in insect ovaries besides specific proteins include mRNA, tRNA, and ribosomes (for review see Berry 1985) as well as mitochon- dria (Mays 1967; Ramamurty 1977). All these materials are negatively charged at physiological pH, and the direc- tion of the voltage gradient with the nurse chamber in poly- trophic follicles and the tropharium in telotrophic ovarioles more negative would result their electrophoretic transport towards the oocyte. Our injections of Fly(+) and M c F l y ( - ) into the tropharium of D. intermedius mimicked the trans- port of naturally transported material and confirmed the hypothesis of Woodruff and Telfer (1980), because Fly(+) was retained in the trophocytes, whereas M c F l y ( - ) mOved unhindered via the trophic cords into the oocytes. In the two other telotrophic systems studied so far, Rhodnius pro- lixus (Telfer et al. 1981) and Oncopeltusfasciatus (Woodruff and Anderson 1984) a restriction of the voltage gradient to the tropharium was found and no passage of injected marker proteins through the nurse strands could be ob- tained. As we could show, this does occur at least in the Dysdercus ovariole. A polarized transport of microinjected M c F l y ( - ) via the cytoplasmic bridges into the oocyte had been demonstrated so far only for the polytrophic follicle of H. cecropia.
The counterexperiment, however, with the proteins in-
jected into previtellogenic Dysdercus oocytes, revealed no difference at all in the motility of Fly(+) and M c F l y ( - ) into the trophic cord. As we could repeat the respective experiments many times with consistent results, an artefact seems unlikely. On the other hand, the possibility that the voltage gradient between the trophic cord and the adjacent ooplasm is not steep enough to prevent the diffusion of M c F l y ( - ) into the nurse strand can be scrutinized by volt- age calculations. In the H. ceeropia follicle the current is focussed across a short cytoplasmic intercellular bridge of 0.05 mm length, in which a magnitude of 1 V/cm is achieved (Telfer et al. 1981). In the telotrophic bug ovariole, how- ever, the trophic cord exceeds a length of 1 mm, and even with the lower assumption of a distance of 0.5 mm of medi- um sized previtellogenic oocytes the gradient would reach only 0.074 V/cm. To produce a voltage gradient as steep as the 1 V/cm found in H. cecropia the potential difference between tropharium and the previtellogenic oocytes would need to be 13.5 times larger, i.e. the tropharium would have to be 50 mV more negative than the oocyte (Woodruff and Anderson 1984). Our intracellular measurements clearly show that a voltage gradient of this magnitude does not exist in the Dysdercus ovariole.
In addition to its function in transport, the voltage gra- dient may distribute or fixate cytoplasmic determinants be- tween the peripheral trophocytes and the more negative center of the trophic core depending on their respective

charge. Electrophoretic restriction of regulatory soluble cy- toplasmic components could enable trophocytes and oo- cytes to develop independently within the syncytium (Telfer et al. 1981; Woodruf f et al. 1986) and also localize other factors within destinct ooplasmic regions during oogenesis (Dittmann et al. 1981) as well as later in early plasmodial embryogenesis (Illmensee 1976).
Our data suggest an effective electrophoretic transport mechanism between the peripheral trophocytes and the center of the trophic core in the Dysdercus t ropharium but raise the question of additional transport mechanisms act- ing inside the trophic cords and extending into the oocytes.
Microtubule-associated transport
The arrangement of microinjected mitochondria within the trophic core of the Dysdercus t ropharium indicates an asso- ciation along tubular structures not visible in the light mi- croscope. This assumption is strongly supported by the ap- parent downstream displacement of the labeled mitochon= dria in long rows with sometimes pronounced sharp aligne- ment. Which types o f tubular elements are known from ultrastructural studies o f bug ovaries? The trophic core and the nurse strands of the Dysdercus ovariole are densely packed with bundles o f parallel microtubules oriented lon- gitudinally (Brunt 1970). In addition the trophic core of the t ropharium is encircled by a filamentous mesh of F- actin (Gutzeit 1986), but the trophic core itself is free of F-actin (Huebner and Gutzeit t986). The involvement of microtubules in the transport of macromolecules or organ- elles has been proved especially in nerve axons (Weiss and Gross 1982). Yet it is not resolved whether microtubule- associated transport is mediated by a kinesin-like ' m o t o r ' (Vale et al. 1985), which requires crossbridging between or- ganelles and microtubules, or by microstreams (Gross 1975; Weiss and Gross 1982), which do not require such specific interactions. Microtubules have already several times con- sidered to play an active role in the cytoplasmic transport in telotrophic ovarioles (Macgregor and Stebbings 1970; Hyams and Stebbings 1977, 1979a, b, c). Stebbings and Hunt (1983) demonstrated by hook-decorat ion in the tro- phic cords o f O. fasciatus an orientation of the microtubules in the same sense with the plus or fast-growing end in the tropharium. In Notonecta, the microtubules form a relative- ly static cytoskeleton, whereas other labeled proteins move independently, suggesting a microtubule-based transport (Stebbings et al. 1985).
Mitochondria injected into the tropharium leave the site of deposition always in direction to the basal part of the trophic core but never in other directions. Unfortunately we were yet unable to follow a translocation of labeled mitochondria right into the long nurse strands. If this does exist, then their movement through the narrow trophic cords might be driven by essentially the same mechanisms as known for axonal transport. We recently began to study this question by means of A V E C - D I C high resolution mi- croscopy. The first results will be published elsewhere (Ditt- mann et al., unpublished work).
Polyfunctional nurse cell-oocyte interactions
The growing insect oocyte in meroistic ovaries is depending on nurse cell products in many respects. The mechanisms involved in short way as well as in long distance intercellular
395
transport according to recent findings also seem to be poly- functional. It may well be that different transport systems are used to move different types of goods like proteins and organelles. Furthermore, there could be some ultrastructur- al and also a functional variation within the various insect orders. This question and the contribution o f the different subsystems evidenced until now has to be clarified by future studies.
Acknowledgements. Support by the DFG (Schwerpunkt Differen- zierung) is greatfuUy appreciated. We wish to thank Drs W Engels, WH Telfer and RI Woodruff for their helpful cooperations, discus- sions and critical comments.
References
Berry SJ (1985) RNA synthesis and storage during insect oogenesis. In : Browder LW (ed) Developmental biology. A comprehensive synthesis, vol l, oogenesis. Plenum Press, New York London, pp 351-384
Bier K (1963) Synthese, interzellul/irer Transport und Abbau yon Ribonukleins/iure im Ovar der Stubenftiege Musca domestica. J Cell Biol 16:436440
Bier K (1964) Die Kern-Plasma-Relation und das Riesenwachstum der Eizellen. Zool Anz [Suppl] 27:84-91
Bier K (1965) Uber den Transport zelleigener Makromolekiile durch die Kernmembran. I. RNS-Synthese und RNS-Transport unter Sauerstoffmangel und bei herabgesetzter Temperatur. Chromosoma 16:58-69
Bier K (1967) Oogenese, das Wachstum yon Riesenzellen. Natur- wissenschaften 54:189-195
Brunt A (1970) Extensive system of microtubules in the ovariole of Dysdercus fasciatus Signoret (Heteroptera: Pyrrhocoridae). Nature 228:80-81
Dittmann F, Ehni R, Engels W (1981) Bioelectric aspects of the hemipteran helotrophic ovariole (Dysdercus intermedius). Wil- helm Roux's Arch 190:221-225
Dittmann F, H6rner R, Engels W (1984) Endoploidization of tro- pharium nuclei during larval development and the first gonocy- cle in Dysdercus intermedius (Heteroptera). Int J Invert Repr Dev 7 : 279-290
Engels W (1969) Geschwindigkeit des RNS-Transportes im Ein/ihr- verband der Dermapteren im Vergleich mit anderen Insekten meroistischen Ovartypes. Zool Anz [Suppl] 33:30-39
Goldman M (1968) Fluorescent antibody methods. Academic Press, New York
Gross GW (1975) The microsteam concept of axoplasmic and den- dritic transport. Adv Neurol 12:283-296
Gutzeit HO (1986) Transport of molecules and organelles in mero- istic ovarioles of insects. Differentiation 31 :t 55-165
Huebner E, Gutzeit HO (1986) Nurse cell-oocyte interaction: a new F-actin mesh associated with the microtubule-rich core of an insect ovariole. Tissue Cell 18:753-764
Hyams JS, Stebbings H (1977) The distribution and function of microtubules in nutritive tubes. Tissue Cell 9 : 537-545
Hyams JS, Stebbings H (1979a) The formation and breakdown of nutritive tubes massive microtubular organelles associated with cytoplasmic transport. J Ultrastruct Res 68:46 57
Hyams JS, Stebbings H (1979b) M~crotubule associated cytoplas- mic transport. In: Roberts K, Hyams JS (eds) Microtubules. Academic Press, New York, pp 487-573
Hyams JS, Stebbings H (1979c) The mechanism of microtubule associated cytoplasmic transport. Isolation and preliminary characterization of a microtubule transport system. Cell Tissue Res 196:103-116
Itlmensee K (1976) Nuclear and cytoplasmic transplantation in Drosophila. In: Lawrence PA (ed) Insect development. Black- well, Oxford, pp 76-96
Illmensee K, Mahowald AP, Loomis MR (1976) The ontogeny

396
of germ plasm during oogenesis in Drosophila. i Dev Biol 49: 40-65
King RC, Brining J (1985) The origin and functioning of insect oocytes and nurse cells. In: Kerkut GA, Gilbert LI (eds) Com- prehensive insect physiology, biochemistry and pharmacology, vol 1. Embryogenesis and reproduction. Pergamon Press, Ox- ford New York, pp 37-82
Loof A de (1986) The electrical dimension of cells: the cell as a miniature electrophoresis chamber. Int Rev Cytol 104:251-352
Macgregor HC, Stebbings H (1970) A massive system of microtu- bules associated with cytoplasmic movement in telotrophic ovarioles. J Cell Sci 6:431-449
Mays U (1967) Parakristallines endoplasmatisches Retikulum im Ovar yon Pyrrhocoris apterus (Heteroptera). Z Naturforsch 22: 459-460
Mays U (1972) Autoradiographische Untersuchungen zum Stoff- transport von den N/ihrzellen zur Oocyte der Feuerwanze Pyr- rhocoris apterus L. (Heteroptera). Z Zellforsch 123:395-410
Ramamurty PS (1977) Trans-fusomal mitochondrial flow in the nurse-chamber of the ovarian follicles in Apis mellifera. J Apic Res 16:27-33
Sander K (1984) Maternal effects on insect development. In: Engels W, Clark WH jr, Fischer A, Oliver PJW, Went DF (eds) Adv invertebr reprod, vol 3. Elsevier Science, Amsterdam, pp 127- 136
Schnaitman C, Greenawalt JW (1986) Enzymatic properties of the inner and outer membranes of rat liver mitochondria. J Cell Biol 38:158 175
Stebbings I-I, Hunt C (1983) Microtubule polarity in the nutritive tubes of insect ovarioles. Cell Tissue Res 233:133-141
Stebbings H, Sharma K, Hunt C (1985) Protein turnover in the cytoplasmic transport system within an insect ovary - a clue
to the mechanism of microtubule-associated transport. FEBS Lett 193:22-26
Telfer WH (1975) Development and physiology of the ooyte-nurse cell syncytium. Adv Insect Physiol 11:223-319
Telfer WH, Woodruff RI, Huebner E (1981) Electrical polarity and cellular differentiation in meroistic ovaries. Am Zool 21 : 675-686
Vale RD, Schnapp BJ, Reese TS, Sheetz MP (1985) Movement of organelles along filaments dissociated from the axoplasm of the giant squid axon. Cell 40:449-454
Weiss DG, Gross GW (1982) The mierostream hypothesis of axo- plasmic transport: Characteristics, predictions and compatibility with data. In: Weiss DG (ed) Axoplasmic transport. Springer, Berlin Heidelberg New York Tokyo, pp 362-383
Woodruff RI (1979) Electrotonic junction in cecropia moth ovaries. Dev Biol 69:281~95
Woodruff RI, Telfer WH (1973) Polarized intercellular bridges in ovarian follicles of the Cecropia moth. J Cell Biol 58:172-188
Woodruff RI, Telfer WH (1974) Electrical properties of ovarian cells linked by intercellular bridges. Ann NY Acad Sci (USA) 238 : 408-419
Woodruff RI, Teller WH (1980) Electrophoresis of proteins in intercellular bridges. Nature 286: 84-86
Woodruff RI, Anderson KL (1984) Nutritive cord connection and dye-coupling of the follicular epithelium to the growing oocytes in the telotrophic ovarioles in Oneopeltus fasciatus, the milk- weed bug. Wilhelm Roux's Arch 193:158-163
Woodruff RI, Huebner E, Telfer WH (1986) Electrical properties of insect ovarian follicles: Some challenges of a multicellular system. In: Nuccitelli R (ed) Ionic currents in development. AR Liss, New York, pp 147-154
Received March 30 / Accepted June 3, 1987
Related Documents











