ORIGINAL ARTICLE Virus-induced gene silencing in rice using a vector derived from a DNA virus Arunima Purkayastha • Saloni Mathur • Vidhu Verma • Shweta Sharma • Indranil Dasgupta Received: 3 June 2010 / Accepted: 2 September 2010 Ó Springer-Verlag 2010 Abstract Virus-induced gene silencing (VIGS) is a method of rapid and transient gene silencing in plants using viral vectors. A VIGS vector for gene silencing in rice has been developed from Rice tungro bacilliform virus (RTBV), a rice-infecting virus containing DNA as the genetic material. A full-length RTBV DNA cloned as a partial dimer in a binary plasmid accumulated in rice plants when inoculated through Agrobacterium (agroinoculation) within 2 weeks and produced detectable levels of RTBV coat protein. Deletion of two of the four viral ORFs did not compromise the ability of the cloned RTBV DNA to accumulate in rice plants. To modify the cloned RTBV DNA as a VIGS vector (pRTBV-MVIGS), the tissue-spe- cific RTBV promoter was replaced by the constitutively expressed maize ubiquitin promoter, sequences comprising the tRNA-binding site were incorporated to ensure reverse transcription-mediated replication, sequences to ensure optimal context for translation initiation of the viral genes were added and a multi-cloning site for the ease of cloning DNA fragments was included. The silencing ability of pRTBV-MVIGS was tested using the rice phytoene desaturase (pds) gene on rice. More than half of the agroinoculated rice plants showed white streaks in leaves within 21 days post-inoculation (dpi), which continued to appear in all emerging leaves till approximately 60–70 dpi. Compared to control samples, real-time PCR showed only 10–40% accumulation of pds transcripts in the leaves showing the streaks. This is the first report of the con- struction of a VIGS vector for rice which can be introduced by agroinoculation. Keywords Gene silencing Phytoene desaturase Real-time PCR Rice RTBV Virus-induced gene silencing (VIGS) Abbreviations VIGS Virus-induced gene silencing RTBV Rice tungro bacilliform virus PDS Phytoene desaturase RNAi RNA interference GLH Green leafhopper Introduction Virus-induced gene silencing (VIGS) is an emerging technology for rapid and transient gene silencing in plants. VIGS is a manifestation of RNA interference (RNAi), a universal phenomenon playing pivotal roles in defense and cellular regulation at multiple levels in a wide range of biological systems. During viral infection in plants, double- stranded RNA (dsRNA) produced as replication interme- diates of RNA viruses or highly structured regions of viral transcripts can act as efficient inducers of RNAi (Szittya et al. 2002; Moissiard and Voinnet 2006). In VIGS, recombinant viruses carrying plant DNA fragments trigger the silencing of homologous host transcripts by RNAi, enabling this powerful tool to be applied for the analysis of plant gene function (reviewed by Purkayastha and Dasgupta 2009). Electronic supplementary material The online version of this article (doi:10.1007/s00425-010-1273-z) contains supplementary material, which is available to authorized users. A. Purkayastha S. Mathur V. Verma S. Sharma I. Dasgupta (&) Department of Plant Molecular Biology, University of Delhi South Campus, Benito Juarez Road, New Delhi 110021, India e-mail: [email protected] 123 Planta DOI 10.1007/s00425-010-1273-z

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Virus-induced gene silencing in rice using a vector derivedfrom a DNA virus
Arunima Purkayastha • Saloni Mathur •
Vidhu Verma • Shweta Sharma • Indranil Dasgupta
Received: 3 June 2010 / Accepted: 2 September 2010
� Springer-Verlag 2010
Abstract Virus-induced gene silencing (VIGS) is a
method of rapid and transient gene silencing in plants using
viral vectors. A VIGS vector for gene silencing in rice has
been developed from Rice tungro bacilliform virus
(RTBV), a rice-infecting virus containing DNA as the
genetic material. A full-length RTBV DNA cloned as a
partial dimer in a binary plasmid accumulated in rice plants
when inoculated through Agrobacterium (agroinoculation)
within 2 weeks and produced detectable levels of RTBV
coat protein. Deletion of two of the four viral ORFs did not
compromise the ability of the cloned RTBV DNA to
accumulate in rice plants. To modify the cloned RTBV
DNA as a VIGS vector (pRTBV-MVIGS), the tissue-spe-
cific RTBV promoter was replaced by the constitutively
expressed maize ubiquitin promoter, sequences comprising
the tRNA-binding site were incorporated to ensure reverse
transcription-mediated replication, sequences to ensure
optimal context for translation initiation of the viral genes
were added and a multi-cloning site for the ease of cloning
DNA fragments was included. The silencing ability of
pRTBV-MVIGS was tested using the rice phytoene
desaturase (pds) gene on rice. More than half of the
agroinoculated rice plants showed white streaks in leaves
within 21 days post-inoculation (dpi), which continued to
appear in all emerging leaves till approximately 60–70 dpi.
Compared to control samples, real-time PCR showed only
10–40% accumulation of pds transcripts in the leaves
showing the streaks. This is the first report of the con-
struction of a VIGS vector for rice which can be introduced
by agroinoculation.
Keywords Gene silencing � Phytoene desaturase �Real-time PCR � Rice � RTBV � Virus-induced gene
silencing (VIGS)
Abbreviations
VIGS Virus-induced gene silencing
RTBV Rice tungro bacilliform virus
PDS Phytoene desaturase
RNAi RNA interference
GLH Green leafhopper
Introduction
Virus-induced gene silencing (VIGS) is an emerging
technology for rapid and transient gene silencing in plants.
VIGS is a manifestation of RNA interference (RNAi), a
universal phenomenon playing pivotal roles in defense and
cellular regulation at multiple levels in a wide range of
biological systems. During viral infection in plants, double-
stranded RNA (dsRNA) produced as replication interme-
diates of RNA viruses or highly structured regions of viral
transcripts can act as efficient inducers of RNAi (Szittya
et al. 2002; Moissiard and Voinnet 2006). In VIGS,
recombinant viruses carrying plant DNA fragments trigger
the silencing of homologous host transcripts by RNAi,
enabling this powerful tool to be applied for the analysis
of plant gene function (reviewed by Purkayastha and
Dasgupta 2009).
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00425-010-1273-z) contains supplementarymaterial, which is available to authorized users.
A. Purkayastha � S. Mathur � V. Verma � S. Sharma �I. Dasgupta (&)
Department of Plant Molecular Biology, University of Delhi
South Campus, Benito Juarez Road, New Delhi 110021, India
e-mail: [email protected]
123
Planta
DOI 10.1007/s00425-010-1273-z

Although VIGS has been used extensively for dicots, its
use in monocots has been rather limited; only two VIGS
vectors have been reported for monocot species—a Barley
stripe mosaic virus (BSMV)-based vector for use in barley
and wheat (Holzberg et al. 2002) and a Brome mosaic virus
(BMV)-based vector for barley, rice and maize (Ding et al.
2006). Both vectors are based on RNA viruses and their
inoculation involves in vitro transcription, followed by
transcript inoculation to plants. Although there has been a
report of Tobacco rattle virus (RNA virus)-based VIGS
vector being optimized for large-scale functional genomic
screens (Burch-Smith et al. 2006), DNA-based vectors,
rather than RNA-based ones, in general are easier to inoc-
ulate to plants using the methods of biolistics or agroinoc-
ulation, and are potentially useful in such screens (Liu et al.
2002; Lu et al. 2003; Zhang et al. 2009). Thus, developing a
DNA-based VIGS system for monocots would be a useful
tool for deciphering the gene functions of this class of
plants, which includes some of the major food crops.
Rice tungro bacilliform virus (RTBV), genus Tungro-
virus, is a rice-infecting pararetrovirus with dsDNA gen-
ome (Hay et al. 1991; Hull 1996; Fauquet et al. 2005).
RTBV, along with Rice tungro spherical virus (RTSV), a
member of the genus Waikavirus, forms a viral complex
causing the tungro disease of rice in South and Southeast
Asia. The affected plants display stunting and yellow-
orange foliar discoloration. In the viral complex, RTBV
gives rise to most of the symptoms, whereas RTSV is
responsible for the insect vector transmissibility by the
green leafhopper (GLH). Cloned RTBV DNA can be
introduced into rice plants using Agrobacterium-mediated
transfer (agroinoculation), upon which it causes symptoms
of mild stunting (Dasgupta et al. 1991). RTBV has a
genome of around 8 kb encoding four open reading frames
(ORFs I–IV, Hay et al. 1991). Functions of proteins
encoded by ORFs I, II and IV have not yet been elucidated.
ORF III codes for a polyprotein, which is processed to
obtain, among others, coat protein, aspartate protease and
reverse transcriptase–ribonuclease H.
To construct a VIGS vector for rice, a full-length cloned
DNA molecule representing an Indian RTBV isolate (Nath
et al. 2002) was assembled as a partial dimer in a binary
plasmid and tested for infectivity on rice by agroinocula-
tion. To retain the essential ORFs required for the repli-
cation and spread of the molecule in the plant and to make
it more suitable as a VIGS vector, the partial dimer was
appropriately shortened and modified. Here, we describe
the construction of an RTBV-derived VIGS vector and
show that it is capable of accumulation in emerging leaves,
following agroinoculation of rice plants and give evidence
that it silences an endogenous rice gene. This is the first
report of the development of a VIGS vector for rice based
on a DNA virus.
Materials and methods
Plant growth condition
Plants (two varieties of indica rice, PB-1 and TN-1,
obtained from Indian Agricultural Research Institute, New
Delhi) were grown either in glass tubes in Yoshida’s
medium (Yoshida et al. 1976) at 27�C and under 16 h light
and 8 h dark cycle using artificial lighting with 70%
humidity or in soil at 30�C under glasshouse conditions
with 80% relative humidity and supplemented with addi-
tional lighting.
Construction of pRTBV-Inf, pRTBV-Del-Inf,
pRTBV-MVIGS and pRTBV-MVIGS-pds
The transcription start site in pRTBV203, a cloned DNA
representing an Indian RTBV isolate, has been mapped at
position 7392 (Mathur and Dasgupta 2007). To construct
pRTBV-Inf, an approximately 3.9-kb fragment corre-
sponding to position between 6955–7934 and 1–2930 of
pRTBV203 was amplified using the Expand Long Tem-
plate PCR System (Roche, Mannheim, Germany) using
primer P1 (Table S1) and standard M13 reverse primer,
using pRTBV203 as the template. The primer P1 was
designed to include a SalI site at its 50 end, while the M13
reverse primer was complementary to regions of the
pUC18 backbone of pRTBV203. This region of the RTBV
DNA, between position 6955 of pRTBV203, upstream of
the promoter and unique BamHI site at position 2930
within the ORF III was used as the repeat region in the
partial dimer. The amplified fragment was cloned in the
binary vector pBinAR (Hofgen and Willmitzer 1990) using
the enzymes BamHI and SalI. To complete the partial
dimer, the unit-length RTBV genome cloned in the BamHI
site in pRTBV203 was excised and ligated to the BamHI-
linearized pBinAR. The expected partial dimer (pRTBV-
Inf) was selected from transformed colonies following
multiple restriction digestions of the resident plasmids.
The following steps were used to obtain pRTBV-Del-
Inf, the deleted version of pRTBV-Inf. First, an 846-bp
fragment of pRTBV203 from the nucleotide position
7169–7934 and 1–81 was amplified using Pwo polymerase
(Roche) with primers P2 and P3, carrying recognition sites
for EcoRI at their 50 ends (Table S1). The product was
purified, digested with EcoRI and cloned in pCAM-
BIA1380 (Roberts et al. 1998) at the EcoRI site (pRTBV
partial Del-Inf-I). Next, a 761-bp fragment from
pRTBV203 (position 7154–7921) was amplified using Pwo
polymerase (Roche) with primers P4 and P5, carrying
recognition sites for SalI at their 50 ends (Table S1),
purified, digested with SalI and cloned in the SalI site of
the pRTBV partial Del-Inf-I to obtain pRTBV partial
Planta
123

Del-Inf-II. Finally, a 6,124-bp fragment from another
cloned DNA of another Indian RTBV isolate (pRTBV204;
Nath et al. 2002), between the nucleotide residues 999–
7123, was amplified using the Expand High-fidelity PCR
system (Roche) with primers IF1 and IF2, carrying rec-
ognition sites for BamHI at their 50 ends (Table S1). The
product was purified, digested with BamHI and inserted
into the BamHI site of the pRTBV partial Del-Inf-II.
The resulting plasmid (pRTBV-Del-Inf) was checked by
multiple restriction digestions.
A cassette containing MUP, multiple cloning site (MCS)
and nos terminator was obtained from binary vector
pB4NU (Raghuvanshi 2001) by restriction digestion with
EcoRI and HindIII. The cassette was cloned into the binary
vector pCAMBIA2300 (McElroy et al. 1995) digested with
EcoRI and HindIII to obtain an intermediate vector (pINT-
1). The binary vector pB4NU was digested with PstI to
release the MUP fragment of about 2.0 kb. The MUP
fragment was cloned into the PstI site of plasmid pBSK.
The orientation of the insert in pBSK was checked by
restriction digestion with SalI and XbaI. MUP fragment
was obtained by digestion with KpnI and BamHI from the
clone in the sense-orientation and inserted into pINT-1 to
obtain the second intermediate vector, pINT-2, containing
two MUP fragments as direct repeats. Primer pairs
MVIGS-FP and MVIGS-RP (Fig. 1) were designed to
amplify ORF III and ORF IV from pRTBV204 corre-
sponding to nucleotide position 999–7183 of RTBV. The
DNA fragment was amplified using Phusion polymerase
(Finnzymes, Espoo, Finland) and the product was cloned
into TOPO cloning vector (Invitrogen, Carlsbad, CA,
USA). The cloned fragment was subsequently released
from the TOPO vector using BamHI. pINT-2 DNA was
digested with BamHI and ligated to the 6.1-kb amplifica-
tion product from pRTBV204 to obtain pRTBV-MVIGS.
The identity and orientation of the inserts were checked by
digestion with appropriate restriction enzymes.
Total RNA was isolated from rice using TRIzol Reagent
(Invitrogen). A 529-bp cDNA fragment of pds corre-
sponding to nucleotide residues 211–740 on the cDNA
sequence (AF049356) was amplified from the cDNA pool
of rice using primer pairs PDS-FP and PDS-RP (Table S1).
The primers were designed to include MluI and PacI
restriction sites at their respective 50 ends, respectively. The
cDNA fragment thus obtained was cloned into T/A vector
(InsT/A cloning Kit, Fermentas, Ontario, Canada), digested
with MluI and PacI and cloned in a reverse orientation into
the MluI–PacI sites of MCS of pRTBV-MVIGS to give
rise to pRTBV-MVIGS-pds.
Agroinoculation procedures
For agroinoculation, all vectors were transformed into
Agrobacterium tumefaciens strain EHA105 (Hood et al.
1993). Transformation was confirmed by colony PCR.
Agroinoculations were either carried out by the method
described by Dasgupta et al. (1991) or by a modified pro-
cedure. In the former method, the plants were maintained
in soil at 30�C under glasshouse conditions. In the latter
method, a primary culture was initiated from a single
Agrobacterium colony in LB medium supplemented with
appropriate antibiotics. Subsequently, a secondary culture
was grown so as to have an OD600 of 0.6–0.8. The cells
were harvested and resuspended in 10 mM MES, 10 mM
MgCl2, 200 lM acetosyringone to a volume 20-fold less
than the original. Approximately 15-day-old rice plants
grown in Yoshida’s medium were used for agroinoculation,
in which about 50 ll of the bacterial suspension was
injected into the meristematic region located at the crown
of the plants and were then transferred onto sterile What-
man No. 1 filter paper immersed in Yoshida’s medium
placed on a solid support with its ends dipped into a res-
ervoir containing the medium. The plants were covered
with moist tissue paper and transferred to tubes containing
5’GGATCCGTCGACTGGTATCAGAGCAATGTTACCATGACCATGGGGTCTTAGACCG 3’
a
BamHI SalIKozak sequencetRNA binding site
MVIGS-FP
999-1013
5’GGATCCGGGCCCTTAATTAAGACGTGACGTCCACGCGT TTATGCCTCCAGCCT 3’b
MVIGS-RPBamHI PacI MluI
7199-7213
Fig. 1 Primers MVIGS-FP and MVIGS-RP used to amplify 6.1 kb
RTBV ORF III. a Restriction enzyme sites, tRNA-binding site and
Kozak sequences (in bold and underlined) added are indicated in
primer MVIGS-FP. Also indicated are the RTBV nucleotides (boxed)
with their corresponding positions in the viral genome (GenBank Acc.
No. AJ292232). The trinucleotide ACC has been added and a single
nucleotide at position 1002 (G, italicized) has been changed to
introduce the Kozak sequence. b Primer MVIGS-RP indicating the
restriction sites and the RTBV sequences, with their corresponding
positions
Planta
123

Yoshida’s medium 24 h post-inoculation and were main-
tained at 27�C under conditions mentioned earlier.
Assay for the accumulation of pRTBV-Inf, pRTBV-
Del-Inf and pRTBV-MVIGS in inoculated rice plants
PCR was employed to as a primary screen to detect the
presence of all three DNA molecules in the plants at
15 days post-inoculation (dpi). For pRTBV-Inf, ELISA
was performed using antibodies raised against RTBV,
essentially according to the method described by Clark and
Adams (1977). DNA slot-blot analysis was performed for
pRTBV-Del-Inf and pRTBV-MVIGS using methods
described earlier (Borah et al. 2008). For PCR and dot-blot
analysis, total genomic DNA was isolated from the second
leaf of agroinoculated plants at 15 dpi using a method
described by Dellaporta et al. (1983) with modifications.
Briefly, a 5 cm portion of leaf was ground using liquid
nitrogen and the resultant powder was thawed in 600 ll of
extraction buffer (10 mM Tris pH 8.0, 1 mM EDTA pH
8.0, 1% SDS). The mixture was then incubated at 65�C for
20 min followed by cooling on ice for 5 min. Two hundred
microliters of 6 M ammonium acetate was added to the
tube and was mixed thoroughly followed by centrifugation
at 14,000g in a micro-centrifuge for 10 min at 4�C to pellet
the debris. The clear supernatant was mixed with 600 ll of
isopropanol to facilitate precipitation of DNA. The pre-
cipitation was carried out at -20�C for 30 min. The pre-
cipitate was collected by centrifugation at 14,000g for
15 min at 4�C and was washed with 70% ethanol at room
temperature for 10 min. The DNA pellet thus obtained was
air-dried and dissolved in 20 ll of T10E1 buffer. Ten
microliters of RNase (10 mg/ml) was added to the
extraction buffer. The DNA pellet obtained after isopro-
panol precipitation was used for PCR and dot-blot assay.
The DNA was quantified by spectrophotometer.
PCR amplifications were performed with 100 ng of
DNA. Primer pairs IDG-1 and IDG-2 (Table S1) corre-
sponding to nucleotide positions 5444–5465 and
6529–6553, respectively, in pRTBV204 were used to
screen agroinoculated plants for the presence of accumu-
lated pRTBV-Inf. Primer pairs DI-F and DI-R were
designed corresponding to nucleotide positions 7562–7585
and 1027–1051 in pRTBV204 (Table S1) to differentiate
between molecules derived from pRTBV-Del-Inf from a
natural RTBV infection. The forward primer was designed
from the pRTBV203 promoter region and the reverse
primer was designed from the segment of the genome
corresponding to the MP region of pRTBV204. As the
pRTBV-Del-Inf did not include ORF I and ORF II of
RTBV, it was expected that any molecule arising from
pRTBV-Del-Inf would give rise to a shorter amplicon
compared to those arising from a natural RTBV infection.
For DNA slot-blot analysis for both pRTBV-Del-Inf and
pRTBV-MVIGS, 2 lg of DNA was spotted for each of the
inoculated, mock-inoculated and uninoculated samples. As
a positive control, 400 ng of total genomic DNA from
tungro-affected plants at 30 dpi after GLH inoculation and
10 ng of pRTBV-Del-Inf or pRTBV-MVIGS DNA were
used. The probe for hybridization was derived from a DNA
fragment amplified from pRTBV204 using primer pair MP-
FP/MP-RP (Table S1). Megaprime DNA labeling System
(Amersham, Buckinghamshire, UK) was used for prepar-
ing the probe for Southern hybridization of the blots.
RNA isolation and real-time PCR analysis
Total RNA was isolated from plants agroinoculated with
pRTBV-MVIGS-pds or controls with pRTBV-MVIGS at
28 dpi, using TRIzol Reagent (Invitrogen) and treated with
RNase-free DNase-I (Fermentas) prior to cDNA synthesis
following the manufacturer’s recommendations. The first
strand cDNA was generated with 1 lg of total RNA using
high capacity cDNA archive kit (Applied Biosystems,
Carlsbad, CA, USA). Real-time RT-PCR analysis was
carried out with a real-time PCR detection system (7000
Sequence Detection System, Applied Biosystems). Primers
were designed using Primer Express� software (Applied
Biosystems). For SYBR real-time RT-PCR experiments,
primers pdsRT1 and pdsRT2 were used for pds transcripts
and primers ubq5RT1 and ubq5RT2 for ubiquitin (UBQ5)
transcripts (Table S1). The pds primers, indicated above,
were designed to exclude the region of cDNA cloned into
the pRTBV-MVIGS vector to ensure that only the endog-
enous mRNA was amplified. Amplification efficiency of
pds primers was determined by means of a calibration
curve (CT value vs. log of input cDNA) prepared in
duplicate using a tenfold dilution series. The slope of the
log-linear phase of the calibration curve was used to cal-
culate the percentage efficiency using the formula: effi-
ciency (%) = [10(-1/slope) - 1] 9 100. The UBQ5 gene
was used as an internal control. Cycling parameters were as
follows: 1 cycle at 50�C for 2 min, 1 cycle at 95�C for
10 min and 40 cycles, each consisting of 15 s at 95�C and
1 min at 60�C. The real-time RT-PCR experiment was
repeated with two independent sets of agroinoculated
plants showing streaking phenotype. From each set, total
RNA was isolated from two leaves of each plant (two
biological replicates) and from each RNA preparation,
real-time PCR was performed three times to determine
pds transcript levels (three technical replicates). The CT
values obtained for desired genes were normalized with
UBQ5 reference values and the DCT values were thus
obtained. Each DCT value was then compared to that of
the controls which was considered as the calibrator value
and the relative mRNA levels were calculated by the
Planta
123

formula: relative mRNA level ¼ 2�ðDCT sample�DCT calibratorÞ.The results thus obtained were plotted as relative mRNA
levels.
Results
Cloned derivatives of RTBV DNA accumulated
in agroinoculated rice plants
To develop a VIGS vector for rice from RTBV, the ability of
a cloned RTBV DNA present in a partially dimerised form in
a binary plasmid to systemically accumulate in rice plants
was investigated. The strategy described for the construction
of an infectious clone, a Philippine isolate of RTBV,
pRTRB1162 (Dasgupta et al. 1991), was followed. An
Indian isolate of RTBV (pRTBV203) was used as the
starting material for the construction of the VIGS vector. In
pararetroviruses, since a terminally redundant more than
full-length transcript acts as the template for reverse tran-
scription-mediated synthesis of the viral DNA, to check the
infectivity of a viral DNA, it is necessary to have it cloned as
a partial dimer in a binary vector and introduced in the host
plant through Agrobacterium-mediated transfer, a process
named agroinoculation (Grimsley et al. 1986). In such
constructs, the promoter of the viral genome is repeated, so
that a viral transcript with a terminal repeat is generated in
agroinoculated plants (Medberry et al. 1990; Dasgupta et al.
1991; Bouhida et al. 1993; Huang and Hartung 2001). Such a
partial dimer construct from an Indian isolate of RTBV was
constructed and named pRTBV-Inf. The plasmid contained
a unit-viral-length BamHI fragment in an orientation capa-
ble of giving rise to a RTBV transcript with direct terminal
repeats. The integrity of pRTBV-Inf was checked using
appropriate restriction enzymes and the expected digestion
patterns were obtained. A schematic representation of
pRTBV-Inf is shown in Fig. 2a.
The effect of introducing pRTBV-Inf in rice plants by
agroinoculation was tested with an average of 40 plants per
experiment, repeated ten times. The rice variety TN-1,
which displays symptoms of severe stunting and yellow-
orange foliar discoloration, was used for these experiments
and plants were maintained under glasshouse conditions.
PCR performed with primers IDG-1 and IDG-2 (Table S1)
on systemic leaves to amplify RTBV DNA showed
amplification of a band of the expected size in 11–19% of
all the agroinoculated plants by 14 dpi (Fig. 3a). Eighty
percent of the PCR-positive plants displayed symptoms of
stunting and yellow-orange discolouration by 15–20 dpi,
the symptoms getting more pronounced by 30 dpi
(Fig. 3b). None of the mock-inoculated plants (pRTBV-Inf
in E. coli) showed symptoms or amplified RTBV DNA. As
a confirmation for the presence of RTBV, ELISA of nine
randomly selected PCR-positive plants using RTBV anti-
bodies showed average values of 1.76 times that of a mock-
inoculated plant, similar to the values obtained in plants
infected with RTBV and RTSV transmitted using GLH.
pRTBV-Inf, because of its large size (almost 24 kb),
does not lend itself easily as a VIGS vector. Since the
essential functions of replication and systemic spread are
all carried out by the proteins encoded by the product of
ORF III (Hay et al. 1991; Laco and Beachy 1994; Laco
et al. 1995; Marmey et al. 1999), it was reasoned that it
may be possible to have some or all of the ORFs I, II and
IV deleted from pRTBV-Inf, keeping the essential func-
tions intact. Such a molecule was assembled and named
pBinAR backbonea
ORF I ORF II ORF III ORF IVRTBVpromoter
RTBVpromoter
ORF I ORF II ORF III (partial)
pCAMBIA1380 backboneb
pRTBV-Inf
MCS
pCAMBIA1380 backboneb
pRTBV-Del-Inf
MUP
MCS
pCAMBIA 2300 backbonec
MUP
tRNA binding site
pRTBV-MVIGS
Fig. 2 Schematic representation of RTBV-derived vectors used in
this study, pRTBV-Inf (a), pRTBV-Del-Inf (b) and pRTBV-MVIGS
(c). Dashed line in each case represents the binary plasmid, solidblack box represents tRNA-binding site introduced in the primer,
shaded rectangles indicate RTBV ORFs, similar shadings indicate
identical ORFs, shaded arrows indicate promoters. MUP maize
ubiquitin promoter. Position of MCS is indicated in pRTBV-MVIGS
Planta
123

pRTBV-Del-Inf, which contained as an insert, nucleotides
corresponding to positions 7169–7934 and 1–81 from
pRTBV 203, 999–7123 from pRTBV204 and 7154–7921
from pRTBV203, in that order. Such a molecule is
expected to generate, from the internal RTBV promoter
and the transcription-termination site, a terminally redun-
dant transcript containing the complete coding regions of
ORF III and a 50 nucleotide 30-truncated ORF IV, flanked
by a direct terminal repeat of about 500 bp each in rice
plants. A schematic representation of pRTBV-Del-Inf is
shown in Fig. 2b.
Once the infectious nature of pRTBV-Inf was estab-
lished, pRTBV-Del-Inf was tested for accumulation in
systemic leaves of two rice varieties, TN-1, described
earlier and PB-1, a variety which does not show yellow-
orange discolouration on RTBV and RTSV infection, but
shows marked stunting. A total of 121 PB-1 plants were
agroinoculated with pRTBV-Del-Inf in 5 independent
experiments, all being maintained under glasshouse con-
ditions. PCR performed on the plants at 14 dpi showed a
similar proportion amplifying pRTBV-Del-Inf as was
observed for pRTBV-Inf. The forward and reverse primers
to detect the accumulation of pRTBV-Del-Inf inoculated
plants, DI-F and DI-R, were designed from the RTBV
promoter and the 50 end of ORF III, respectively. Since
RTBV ORFs I and II are absent in pRTBV-Del-Inf, an
amplification product of about 0.5 kb was expected as
compared to a 1.4 kb from RTBV DNA. Indeed, all the
amplified products were of 0.5 kb in size (data not shown).
A total of 145 TN-1 plants were agroinoculated in 3
independent experiments to estimate the accumulation of
pRTBV-Del-Inf DNA by dot-blot hybridization, using a
probe derived from RTBV DNA. In contrast to PB-1
plants, TN-1 plants were grown at a lower temperature
(27�C), as compared to the glasshouse (30�C) and in a
liquid medium. Distinct hybridization signals were
obtained in 42 out of 145 plants tested at 15 dpi. A
representative dot blot is shown in Suppl. Fig. S1. Com-
pared to the uninoculated plants (slots E1–E3 and E12), the
hybridization signals in 16 of the 48 pRTBV-Del-Inf
inoculated plants (slots A1–D12) were very bright. Lanes
E5 and E6 show the signals in samples from plants inoc-
ulated with RTBV using GLH.
The promoter of the Indian isolate of RTBV when
expressed transgenically is active mainly in the vascular
bundles in a mature rice plant (Mathur and Dasgupta 2007).
We reasoned that the use of a constitutive promoter, rather
than a tissue-specific one, would be more appropriate for a
VIGS vector. Hence, maize ubiquitin promoter (MUP),
because of its constitutive nature and high levels of
expression in monocots, was chosen to drive the tran-
scription of the viral genes. Apart from replacing the pro-
moter, tRNA-binding site (to enable tRNA-mediated
initiation of reverse transcription, an essential feature of
pararetroviral DNA replication, to take place, Hay et al.
1991) and Kozak sequences (Kozak 1989), to provide an
optimal sequence context for translation initiation of the
viral genes, were introduced immediately downstream of
MUP. The above features were designed to be part of the
primer used to amplify the RTBV genes (MVIGS-FP,
Fig. 1). To introduce the Kozak sequence, three nucleo-
tides ‘‘ACC’’ were introduced and the ‘‘A’’ at nucleotide
position 1002 of the RTBV sequence was replaced with
‘‘G’’ (Fig. 1). The recognition sites of the restriction
enzymes PacI and MluI were introduced in the primer,
MVIGS-RP (Fig. 1). A 6.1-kb DNA fragment representing
ORFs III and IV of the RTBV DNA was amplified from
pRTBV204, using primers MVIGS-FP and MVIGS-RP,
such that the MUP, tRNA-binding site and the Kozak
sequence were positioned at the 50 end of the product and
the PacI–MluI sites at the 30 end, together cloned into the
binary plasmid pCAMBIA 2300. The vector was called
pRTBV-MVIGS, a schematic diagram of which is shown
in Fig. 2c.
a M 1 2 3 4 5 6 7 8 9 10 11 + b c
1.1 kb
6.9
1.1 0.70.70.6
Mock pRTBV-Inf pRTBV-MVIGSMock
Fig. 3 Detection of pRTBV-Inf in rice plants after agroinoculation
with pRTBV-Inf and lack of symptoms following agroinoculation
with pRTBV-MVIGS. a Lanes 1–11 PCR amplification of pRTBV-
Inf in agroinoculated rice plants at 14 dpi; M DNA size marker; ?:
pRTBV-Inf plasmid as positive control, numbers at the side indicate
size markers in kb. b Photograph of plants variety TN-1 mock-
inoculated and agroinoculated by pRTBV-Inf at 30 dpi. c Photograph
of plants variety TN-1 mock-inoculated and agroinoculated by
pRTBV-MVIGS at 20 dpi. The roots have been trimmed for clarity
Planta
123
The ability of pRTBV-MVIGS, the vector with several
new features compared to pRTBV-Del-Inf, to accumulate
in rice plants was checked on the rice variety TN-1. A total
of 110 plants were agroinoculated in 5 independent
experiments and observed over a period of 10 weeks at
27�C maintained in liquid medium. No stunting or yel-
lowing was noticed in plants inoculated with pRTBV-
MVIGS or in mock-inoculated controls (plants inoculated
with pRTBV-MVIGS in E. coli) at 20 dpi (Fig. 3c), a stage
where symptoms appear in case of inoculation with
pRTBV-Inf. The plants were observed till 70 dpi for the
appearance of symptoms but none were apparent. A frac-
tion of plants inoculated with pRTBV-MVIGS was tested
for the accumulation of pRTBV-MVIGS DNA in the sec-
ond leaf at 14 dpi by DNA dot blot. Of the 75 plants tested,
52 showed strong signals indicating accumulation of
pRTBV-MVIGS in the leaf tested.
pRTBV-MVIGS-pds was able to silence the rice pds
gene
To determine the efficacy of pRTBV-MVIGS in gene
silencing, the marker gene phytoene desaturase (pds) was
selected. PDS is an enzyme required for the biosynthesis of
carotenoid pigments that protect chlorophyll from photo-
bleaching and its silencing is associated with the devel-
opment of white streaks in the leaves of monocot plants
(Holzberg et al. 2002). The pRTBV-MVIGS construct
carrying pds were named pRTBV-MVIGS-pds.
Thirty-six TN-1 plants were agroinoculated with
pRTBV-MVIGS-pds in three independent experiments.
These plants were maintained in liquid medium (Yoshida’s
medium) at 27�C. At the same time, 78 TN-1 plants
inoculated with pRTBV-MVIGS-pds were maintained in
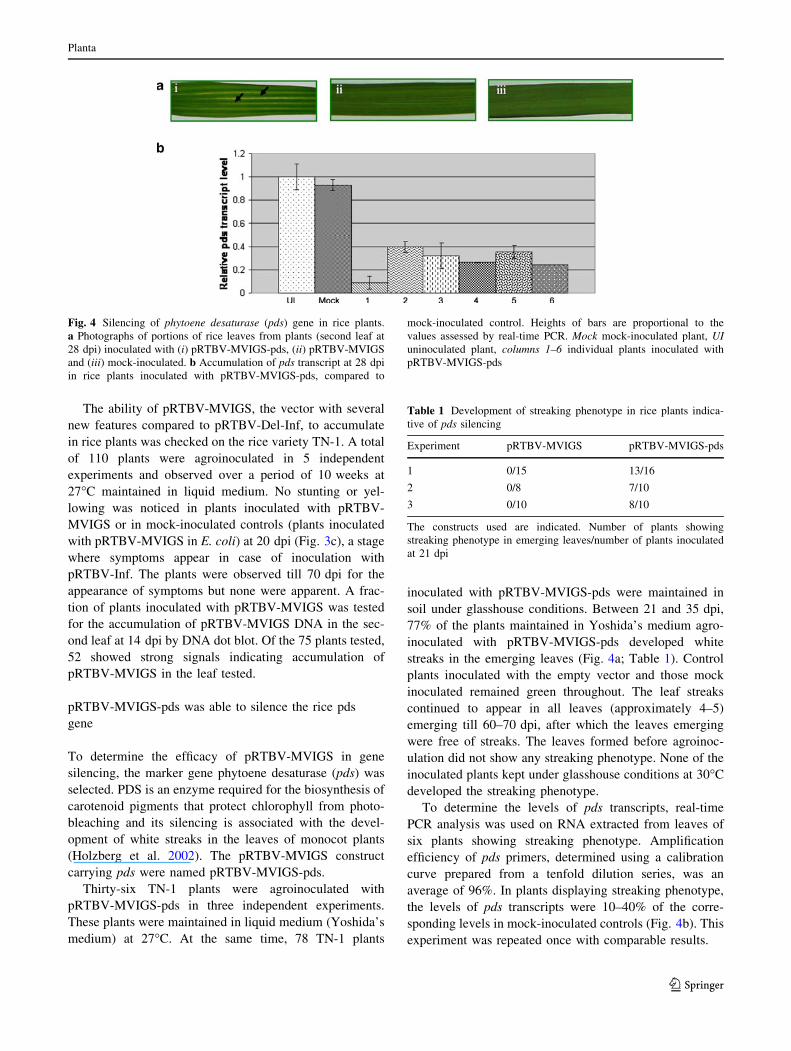
soil under glasshouse conditions. Between 21 and 35 dpi,
77% of the plants maintained in Yoshida’s medium agro-
inoculated with pRTBV-MVIGS-pds developed white
streaks in the emerging leaves (Fig. 4a; Table 1). Control
plants inoculated with the empty vector and those mock
inoculated remained green throughout. The leaf streaks
continued to appear in all leaves (approximately 4–5)
emerging till 60–70 dpi, after which the leaves emerging
were free of streaks. The leaves formed before agroinoc-
ulation did not show any streaking phenotype. None of the
inoculated plants kept under glasshouse conditions at 30�C
developed the streaking phenotype.
To determine the levels of pds transcripts, real-time
PCR analysis was used on RNA extracted from leaves of
six plants showing streaking phenotype. Amplification
efficiency of pds primers, determined using a calibration
curve prepared from a tenfold dilution series, was an
average of 96%. In plants displaying streaking phenotype,
the levels of pds transcripts were 10–40% of the corre-
sponding levels in mock-inoculated controls (Fig. 4b). This
experiment was repeated once with comparable results.
Fig. 4 Silencing of phytoene desaturase (pds) gene in rice plants.
a Photographs of portions of rice leaves from plants (second leaf at
28 dpi) inoculated with (i) pRTBV-MVIGS-pds, (ii) pRTBV-MVIGS
and (iii) mock-inoculated. b Accumulation of pds transcript at 28 dpi
in rice plants inoculated with pRTBV-MVIGS-pds, compared to
mock-inoculated control. Heights of bars are proportional to the
values assessed by real-time PCR. Mock mock-inoculated plant, UIuninoculated plant, columns 1–6 individual plants inoculated with
pRTBV-MVIGS-pds
Table 1 Development of streaking phenotype in rice plants indica-
tive of pds silencing
Experiment pRTBV-MVIGS pRTBV-MVIGS-pds
1 0/15 13/16
2 0/8 7/10
3 0/10 8/10
The constructs used are indicated. Number of plants showing
streaking phenotype in emerging leaves/number of plants inoculated
at 21 dpi
Planta
123

Discussion
Here, we describe, for the first time, the construction of a
DNA-based VIGS vector for rice and demonstrate its use
by silencing the rice pds gene. Reports indicate that DNA-
based VIGS vectors rather than RNA-based ones are the
method of choice especially where high-throughput gene
silencing is required for functional screens of a large
number of genes. For example, the soybean VIGS vector,
based on the RNA virus Bean pod mottle virus (BPMV;
Zhang and Ghabrial 2006), was recently modified to
express the corresponding cDNA under the strong Cauli-
flower mosaic virus (CaMV) 35S promoter and introduced
in the plants by biolistic inoculation (Zhang et al. 2009). In
contrast to the original report of BPMV-based vector, the
DNA vector used a much milder strain of the virus to
achieve comparable levels of silencing in host plant.
Similar modifications have also been reported for the
widely used Tobacco mosaic virus (TMV)- and Potato
virus X-based vectors (Lu et al. 2003; Liu et al. 2002). The
VIGS vector reported in this study, being based on a binary
plasmid, can be easily inoculated into rice plants using
simple agroinoculation procedures, allowing increased
convenience in handling compared to the methods descri-
bed earlier with the BMV-based vectors for VIGS of rice
(Ding et al. 2006). The earlier method required a multi-step
inoculation procedure consisting of an initial in vitro pro-
duction of transcripts and inoculation on barley leaves,
followed by preparation of leaf extracts at 10–14 dpi, with
which to finally inoculate rice plants. In contrast, using
pRTBV-MVIGS, transcript inoculation can be by-passed
and the vector can be directly delivered to the plants in
which gene silencing is desired. A MCS has also been
introduced into the 30 end of the vector to allow easy
cloning of target gene fragments without disrupting the
ORFs.
DNA dot-blot assay indicated accumulation of pRTBV-
Del-Inf molecules in systemic leaves of agroinoculated
plants, despite completely lacking RTBV ORFs I and II
and carrying a truncated ORF IV, which showed that all the
functions required for the replication and spread were
performed by proteins encoded by the ORF III. It has been
reported that ORF II product interacts with Coat protein
and disruption of this interaction affected the infectivity of
cloned RTBV-Phil DNA in agroinoculation experiments
(Herzog et al. 2000). Amino acid residues 56–73 of the
ORF II-encoded protein exhibited coiled structures, similar
to the P3 protein of CaMV, which can form tetramer in
vitro (Leclerc et al. 1998). These two facts taken together
led the authors to propose that the ORF II product may
function in a fashion similar to scaffolding proteins of
dsDNA bacteriophages and certain animal viruses (Hendrix
and Garcea 1994), attributing a role of ORF II product in
capsid assembly of RTBV (Herzog et al. 2000). We did not
attempt to look for virus-like particles in plants agroinoc-
ulated with pRTBV-Del-Inf or pRTBV-MVIGS, something
we plan to do in future. Since we observed efficient rep-
lication of pRTBV-Del-Inf and pRTBV-MVIGS, both
lacking ORF II, we conclude that ORF II product has no
significant role to play in the replication and spread of these
molecules in rice.
The streaking phenotype appearing in newly emerging
leaves is a characteristic feature of pds silencing. The
mock-inoculated and uninoculated plants were indistin-
guishable, indicating that the agroinoculation process, per
se, was not giving rise to any stress response in the inoc-
ulated plant. Just 10–40% levels of pds transcripts in the
leaves displaying steaks compared to the controls were
similar to that reported earlier for pds-silenced barley
(Holzberg et al. 2002; Ding et al. 2006; Bruun-Rasmussen
et al. 2007), using BSMV- and BMV-based VIGS vectors.
Interestingly, it was observed that the silencing phenotype
did not penetrate throughout the entire lamina of the leaves.
One possible explanation for the streaking phenotype,
rather than a uniform bleaching response, may be the
absence of pRTBV-MVIGS-pds in all tissues or the
inability to induce silencing uniformly (Ding et al. 2006).
Development of the photo-bleaching phenotype can be
expected to depend upon the rate of turnover or stability of
PDS. Hence, it is also possible that the silenced phenotype
is not visible in sectors where the carotenoid pigments are
already present in significant quantities or are synthesized
by pre-existing PDS.
One of the challenges of designing a VIGS vector is to
produce symptomless infection or infection with attenuated
symptoms in the host. Both the VIGS vectors available for
monocots (BSMV- and BMV-based) are associated with
viral symptoms, albeit mild (Holzberg et al. 2002; Ding
et al. 2006). In contrast, inoculation of plants with either
the pRTBV-MVIGS or that containing target gene insert
was completely symptomless. It has been reported that
RTBV not RTSV causes most tungro symptoms (Dasgupta
et al. 1991; Encabo et al. 2009), although the identities of
the RTBV genes responsible are unknown. Two rice tran-
scription factors, RF2a and RF2b, interact with and pos-
sibly are sequestered to an element (Box II), present in the
upstream promoter regions (-53 to -39 with respect to the
transcription start site) of RTBV (Dai et al. 2004, 2006).
The above sequestration of the transcription factors has
been proposed to cause perturbation in rice gene expression
patterns leading to the symptoms of tungro. Hence, RF2a-
and RF2b-overexpressing transgenic rice lines do not show
symptoms upon RTBV inoculation (Dai et al. 2008).
Though the Box II element is not completely conserved in
the two groups of RTBV, from Philippines and India
(Mathur and Dasgupta 2007), an interaction of host factors
Planta
123

with elements resembling Box II in the RTBV-WB pro-
moter, causing the tungro symptoms, cannot be ruled out.
In pRTBV-MVIGS, since RTBV promoter is replaced by
MUP, the lack of Box II element or other similar elements,
otherwise interacting with RF2a and RF2b, may contribute
to the lack of symptoms in plants upon agroinoculation
with pRTBV-MVIGS.
White streaks started to appear in the emerging leaves of
the plants agroinoculated with pRTBV-MVIGS-pds from 21
to 35 dpi. All subsequent leaves emerging till 60–70 dpi
continued showing streaks. Thereafter, the emerging leaves
were devoid of streaks. It is interesting to note that similar
streaking phenotype in monocots reported earlier was also
transient in nature (Holzberg et al. 2002; Scofield et al. 2005;
Ding et al. 2006). Stronger levels of silencing might be
achievable using inverted repeat constructs of target genes
(Lacomme et al. 2003), which need to be tested in the future.
Temperature is an important factor affecting the extent
and efficiency of silencing of VIGS vectors. Generally,
lower temperatures enhance the efficiency of VIGS vectors,
such as those based on Tobacco rattle virus (Szittya et al.
2003; Fu et al. 2006; Cai et al. 2007; Nethra et al. 2006;
Tuttle et al. 2008) and Cotton Leaf Crumple Virus
(Chellappan et al. 2005). This probably explains why we, in
this study, observed efficient silencing by pRTBV-MVIGS
only when the plants were grown at a temperature of 27�C in
the culture room and not at 30�C. Lower humidity conditions
have also been shown to be favorable for efficient silencing
by TRV-VIGS (Fu et al. 2006). The humidity conditions in
the culture room were generally lower than that of glass-
house and this factor may also have added to the silencing
phenotype obtained under culture room conditions.
Rice is the single most important monocot plant
responsible for providing dietary calories for a large
number of people, mainly in the developing countries. The
complete genome of rice has been sequenced (International
Rice Genome Sequencing Project 2005) and this major
food crop is poised for functional genomics research
making the development of pRTBV-MVIGS system very
timely. pRTBV-MVIGS lends itself well to high-through-
put methods and has the potential to greatly speed up
functional genomic research of rice.
Acknowledgments This work was funded by Department of Bio-
technology, Government of India, New Delhi, Grant no. BT/AB/03/
FG-I/2003 to ID. AP wishes to thank University Grants Commission,
New Delhi and SM, VV and SS wish to thank Council for Scientific
and Industrial Research, New Delhi for Research Fellowships.
References
Borah BK, Johnson AMA, Sai Gopal DVR, Dasgupta I (2008) A
comparison of four DNA extraction methods for the detection of
citrus yellow mosaic badnavirus from two species of citrus using
PCR and dot-blot hybridisation. J Virol Methods 151:321–324
Bouhida M, Lockhart BEL, Olszewski NE (1993) An analysis of the
complete sequence of a sugarcane bacilliform virus genome
infectious to banana and rice. J Gen Virol 74:15–22
Bruun-Rasmussen M, Madsen CT, Jessing S, Albrechtsen M (2007)
Stability of barley stripe mosaic virus-induced gene silencing in
barley. Mol Plant Microbe Interact 20:1323–1331
Burch-Smith TM, Schiff M, Liu Y, Dinesh-Kumar SP (2006)
Efficient virus-induced gene silencing in Arabidopsis. Plant
Physiol 142:21–27
Cai X, Wang C, Xu Y, Xu Q, Zheng Z, Zhou X (2007) Efficient gene
silencing induction in tomato by a viral satellite DNA vector.
Virus Res 125:169–175
Chellappan P, Vanitharani R, Ogbe F, Fauquet CM (2005) Effect of
temperature on geminivirus-induced gene silencing. Plant Phys-
iol 138:1828–1841
Clark MF, Adams AN (1977) Characteristics of the microplate
method of enzyme-linked immunosorbent assay for the detection
of plant viruses. J Gen Virol 34:475–483
Dai S, Zhang Z, Chen S, Beachy RN (2004) RF2b, a rice bZIP
transcription activator, interacts with RF2a and is involved in
symptom development of rice tungro disease. Proc Natl Acad Sci
USA 101:687–692
Dai S, Zhang Z, Bick J, Beachy RN (2006) Essential role of the Box
II cis element and cognate host factors in regulating the promoter
of rice tungro bacilliform virus. J Gen Virol 87:715–722
Dai S, Wei X, Alfonso AA, Pei L, Duque UG, Zhang Z, Babb GM,
Beachy RN (2008) Transgenic rice plants that over-express
transcription factors RF2a and RF2b are tolerant to rice tungrovirus replication and disease. Proc Natl Acad Sci USA
105:21012–21016
Dasgupta I, Hull R, Eastop S, Poggi-Pollini C, Blakebrough M,
Boulton MI, Davies JW (1991) Rice tungro bacilliform virusDNA independently infects rice after Agrobacterium-mediated
transfer. J Gen Virol 72:1215–1221
Dellaporta SL, Wood J, Hicks JB (1983) A plant DNA mini-
preparation, version II. Plant Mol Biol Rep 1:19–21
Ding XS, Schneider WL, Chaluvadi SR, Rouf Mian M, Nelson RS
(2006) Characterization of a brome mosaic virus strain and its
use as a vector for gene silencing in monocotyledonous hosts.
Mol Plant Microbe Interact 19:1229–1239
Encabo JR, Cabauatan PQ, Cabunagan RC, Satoh K, Lee JH, Kwak
DY, De Leon TB, Macalalad RJ, Kondoh H, Kikuchi S, Choi IR
(2009) Suppression of two tungro viruses in rice by separable
traits originating from cultivar Utri Merah. Mol Plant Microbe
Interact 22:1268–1281
Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA (2005)
Virus taxonomy. Eighth Report of the International Committee
for the Taxonomy of Viruses. Academic Press, New York
Fu DQ, Zhu BZ, Zhu HL, Zhang HX, Xie YH, Jiang WB, Zhao XD,
Luo KB (2006) Enhancement of virus-induced gene silencing in
tomato by low temperature and low humidity. Mol Cells
21:153–160
Grimsley N, Hohn B, Hohn T, Walden R (1986) ‘‘Agroinfection’’ an
alternative route for viral infection of plants by using the Ti
plasmid. Proc Natl Acad Sci USA 83:3282–3286
Hay JM, Jones MC, Blakebrough ML, Dasgupta I, Davies JW, Hull R
(1991) An analysis of an infectious clone of rice tungrobacilliform virus, a plant pararetrovirus. Nucleic Acids Res
19:2615–2621
Hendrix RW, Garcea RL (1994) Capsid assembly of dsDNA viruses.
Semin Virol 5:15–26
Herzog E, Guerra-Peraza O, Hohn T (2000) The rice tungro
bacilliform virus gene II product interacts with the coat protein
domain of the viral gene III polyprotein. J Virol 74:2073–2083
Planta
123

Hofgen R, Willmitzer L (1990) Biochemical and genetic analysis of
different patatin isoforms expressed in various organs of potato
(Solanum tuberosum). Plant Sci 66:221–230
Holzberg S, Brosio P, Gross C, Pogue GP (2002) Barley stripe mosaicvirus-induced gene silencing in a monocot plant. Plant J
30:315–327
Hood EE, Gelvin SB, Melchers LS, Hoekema A (1993) New
Agrobacterium helper plasmids for gene transfer to plants.
Transgenic Res 2:208–218
Huang Q, Hartung JS (2001) Cloning and sequence analysis of an
infectious clone of Citrus yellow mosaic virus that can infect
sweet orange via Agrobacterium-mediated inoculation. J Gen
Virol 82:2549–2558
Hull R (1996) Molecular biology of rice tungro viruses. Annu Rev
Phytopathol 34:275–297
International Rice Genome Sequencing Project (2005) The map-based
sequence of the rice genome. Nature 436:793–800
Kozak M (1989) The scanning model for translation: an update. J Cell
Biol 108:229–241
Laco GS, Beachy RN (1994) Rice tungro bacilliform virus encodes
reverse transcriptase, DNA polymerase, and ribonuclease H
activities. Proc Natl Acad Sci USA 91:2654–2658
Laco GS, Kent SB, Beachy RN (1995) Analysis of the proteolytic
processing and activation of the rice tungro bacilliform virusreverse transcriptase. Virology 208:207–214
Lacomme C, Hrubikova K, Hein I (2003) Enhancement of virus-
induced gene silencing through viral-based production of
inverted-repeats. Plant J 34:543–553
Leclerc D, Burri L, Kajava AV, Mougeot JL, Hess D, Lustig A,
Kleemann G, Hohn T (1998) The open reading frame III product
of cauliflower mosaic virus forms a tetramer through a
N-terminal coiled-coil. J Biol Chem 273:29015–29021
Liu Y, Schiff M, Marathe R, Dinesh-Kumar SP (2002) Tobacco Rar1,
EDS1 and NPR1/NIM1 like genes are required for N-mediated
resistance to tobacco mosaic virus. Plant J 30:415–429
Lu R, Malcuit I, Moffett P, Ruiz MT, Peart J, Wu AJ, Rathjen JP,
Bendahmane A, Day L, Baulcombe DC (2003) High throughput
virus-induced gene silencing implicates heat shock protein 90 in
plant disease resistance. EMBO J 22:5690–5699
Marmey P, Bothner B, Jacquot E, de Kochko A, Ong CA, Yot P,
Siuzdak G, Beachy RN, Fauquet CM (1999) Rice tungrobacilliform virus open reading frame 3 encodes a single
37-kDa coat protein. Virology 253:319–326
Mathur S, Dasgupta I (2007) Downstream promoter sequence of an
Indian isolate of rice tungro bacilliform virus alters tissue-
specific expression in host rice and acts differentially in
heterologous system. Plant Mol Biol 65:259–275
McElroy D, Chamberlain DA, Moon E, Wilson KJ (1995) Develop-
ment of a gusA reporter gene construct for cereal transformation,
availability of plant transformation vectors from CAMBIA
molecular genetic resource service. Mol Breed 1:27–37
Medberry SL, Lockhart BEL, Olszewski NE (1990) Properties of
commelina yellow mottle virus’s complete DNA sequence,
genomic discontinuities and transcript suggest that it is a
pararetrovirus. Nucleic Acids Res 18:5505–5551
Moissiard G, Voinnet O (2006) RNA silencing of host transcripts by
cauliflower mosaic virus requires coordinated action of the four
Arabidopsis Dicer-like proteins. Proc Natl Acad Sci USA
103:19593–19598
Nath N, Mathur S, Dasgupta I (2002) Molecular analysis of two
complete rice tungro bacilliform virus genomic sequences from
India. Arch Virol 147:1173–1187
Nethra P, Nataraja KN, Rama N, Udayakumar M (2006) Standard-
ization of environmental conditions for induction and retention
of post-transcriptional gene silencing using tobacco rattle virusvector. Curr Sci 90:431–435
Purkayastha A, Dasgupta I (2009) Virus-induced gene silencing: a
versatile tool for discovery of gene functions in plants. Plant
Physiol Biochem 47:967–976
Raghuvanshi S (2001) Investigations on chloroplast transformation
and characterization of constitutively photomorphogenic 1(COP1) gene in rice. Ph.D. Thesis, University of Delhi
Roberts CS, Rajagopal S, Smith LA, Nguyen TA, Yang W, Nuroho S,
Ravi KS, Cao M-L, Vijayachandra K, Patell V, Harcourt R,
Dransfield L, Desamero N, Slamet I, Keese P, Kilian A,
Jefferson RA (1998) A comprehensive set of modular vectors for
advanced manipulations and efficient transformation of plants by
both Agrobacterium and direct DNA uptake methods. pCAM-
BIA Vector release manual version 3.05. CAMBIA. Canberra,
Australia
Scofield SR, Huang L, Brandt AS, Gill BS (2005) Development of a
virus-induced gene-silencing system for hexaploid wheat and its
use in functional analysis of the Lr21-mediated leaf rust
resistance pathway. Plant Physiol 138:2165–2173
Szittya G, Molnar A, Silhavy D, Hornyik C, Burgyan J (2002) Short
defective interfering RNAs of tombusviruses are not targeted but
trigger post transcriptional gene silencing against their helper
virus. Plant Cell 14:359–372
Szittya G, Silhavy D, Molnar A, Havelda Z, Lovas A, Lakatos L,
Banfalvi Z, Burgyan J (2003) Low temperature inhibits RNA
silencing mediated defence by the control of siRNA generation.
EMBO J 22:633–640
Tuttle JR, Idris AM, Brown JK, Haigler CH, Robertson D (2008)
Geminivirus-mediated gene silencing from cotton leaf crumplevirus is enhanced by low temperature in cotton. Plant Physiol
148:41–50
Yoshida S, Forno SA, Cock SH, Gomez KA (1976) Routine
procedure for growing rice plants in culture solution. In:
Yoshida S (ed) Laboratory manual for physiological studies of
rice. IRRI, Manila, Philippines, pp 3367–3374
Zhang C, Ghabrial SA (2006) Development of bean pod mottle virus-
based vectors for stable protein expression and sequence-specific
virus-induced gene silencing in soybean. Virology 344:401–411
Zhang C, Yang C, Whitham SA, Hill JH (2009) Development and use
of an efficient DNA-based viral gene silencing vector for
soybean. Mol Plant Microbe Interact 22:123–231
Planta
123
Related Documents











