L Journal of Experimental Marine Biology and Ecology 247 (2000) 29–49 www.elsevier.nl / locate / jembe Effect of diet and temperature upon muscle metabolic capacities and biochemical composition of gonad and muscle in Argopecten purpuratus Lamarck 1819 a, b a a * ´ Gloria Martınez , Katherina Brokordt , Cristian Aguilera , Viterbo Soto , b Helga Guderley a ´ Departamento de Biologıa Marina, Universidad Catolica del Norte, Casilla 117, Coquimbo, Chile b ´ ´ ´ Departement de Biologie, Universite Laval, Quebec, P .Q., Canada G1K 7P4 Received 2 August 1999; received in revised form 1 November 1999; accepted 15 December 1999 Abstract Recently spawned Argopecten purpuratus broodstock were conditioned at two temperatures and fed three different diets (microalgae, microalgae mixed with lipids and microalgae mixed with carbohydrates) to examine changes in the biochemical composition of gonad and muscle as well as muscle metabolic capacities. During one experiment, scallops were fed at 3% of their dry mass per day whereas during a second experiment, they were fed at 6% of their dry mass per day. During both experiments, total gonadal levels of lipids and protein increased markedly during con- ditioning with the two mixed diets at 168C. These increases were less pronounced at 208C. Carbohydrate gonadal levels only increased during the second experiment at both temperatures and with the three diets. Of the major biochemical components of the adductor muscle, carbohydrate levels changed most during conditioning. Whereas muscle protein levels increased slightly with gonadal maturation, carbohydrate levels dropped considerably. Despite the marked drop in the levels of glycolytic substrates, only the activities of octopine dehydrogenase in the adductor muscle of the scallops conditioned at 168C consistently decreased. Muscle levels of glycogen phosphorylase were higher in mature than in recently spawned (control) scallops, suggesting a role in the transfer of glucose equivalents from the adductor muscle to other tissues. 2000 Elsevier Science B.V. All rights reserved. Keywords: Argopecten purpuratus; Diet; Temperature; Gonads; Muscle; Metabolic capacity; Composition *Corresponding author. Tel.: 1 56-51-209-793; fax: 1 56-51-209-812. ´ E-mail address: [email protected] (G. Martınez) 0022-0981 / 00 / $ – see front matter 2000 Elsevier Science B.V. All rights reserved. PII: S0022-0981(00)00143-X

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

LJournal of Experimental Marine Biology and Ecology247 (2000) 29–49
www.elsevier.nl / locate / jembe
Effect of diet and temperature upon muscle metaboliccapacities and biochemical composition of gonad and muscle
in Argopecten purpuratus Lamarck 1819a , b a a*´Gloria Martınez , Katherina Brokordt , Cristian Aguilera , Viterbo Soto ,
bHelga Guderleya ´Departamento de Biologıa Marina, Universidad Catolica del Norte, Casilla 117, Coquimbo, Chile
b ´ ´ ´Departement de Biologie, Universite Laval, Quebec, P.Q., Canada G1K 7P4
Received 2 August 1999; received in revised form 1 November 1999; accepted 15 December 1999
Abstract
Recently spawned Argopecten purpuratus broodstock were conditioned at two temperatures andfed three different diets (microalgae, microalgae mixed with lipids and microalgae mixed withcarbohydrates) to examine changes in the biochemical composition of gonad and muscle as well asmuscle metabolic capacities. During one experiment, scallops were fed at 3% of their dry mass perday whereas during a second experiment, they were fed at 6% of their dry mass per day. Duringboth experiments, total gonadal levels of lipids and protein increased markedly during con-ditioning with the two mixed diets at 168C. These increases were less pronounced at 208C.Carbohydrate gonadal levels only increased during the second experiment at both temperaturesand with the three diets. Of the major biochemical components of the adductor muscle,carbohydrate levels changed most during conditioning. Whereas muscle protein levels increasedslightly with gonadal maturation, carbohydrate levels dropped considerably. Despite the markeddrop in the levels of glycolytic substrates, only the activities of octopine dehydrogenase in theadductor muscle of the scallops conditioned at 168C consistently decreased. Muscle levels ofglycogen phosphorylase were higher in mature than in recently spawned (control) scallops,suggesting a role in the transfer of glucose equivalents from the adductor muscle to other tissues. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Argopecten purpuratus; Diet; Temperature; Gonads; Muscle; Metabolic capacity; Composition
*Corresponding author. Tel.: 1 56-51-209-793; fax: 1 56-51-209-812.´E-mail address: [email protected] (G. Martınez)
0022-0981/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0022-0981( 00 )00143-X

´30 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
1. Introduction
In most animals, but particularly in broadcast spawners, gonadal maturation is a costlyprocess requiring considerable investment of material and energy. When food availabilityis high, the cost of gonadal maturation can be covered entirely by absorbed materials andenergy, but when food availability is low, gonadal maturation can require mobilizationof macromolecular components from other tissues for their use for gametogenesis.Scallops are broadcast spawners in which the mature gonad can account for more than20% of the body mass. Inverse cycles of the biochemical components in the adductormuscle and gonad occur during the reproductive cycle of various scallop species,supporting the interpretation that materials are mobilized from the muscle to supportgonadal maturation. Such cycles have been observed for Chlamys opercularis (Taylorand Venn, 1979), Chlamys varia (Shafee, 1981), Argopecten irradians concentricus(Barber and Blake, 1981), Argopecten irradians irradians (Epp et al., 1988), Placopec-
´ten magellanicus (Couturier and Newkirk, 1991), Argopecten purpuratus (Martınez,´1991; Martınez and Mettifogo, 1998), Pecten maximus (Pazos et al., 1997) and Euvola
(Pecten) ziczac (Brea, 1986; Boadas et al., 1997). Whereas glycogen and protein are theprincipal macromolecules which are mobilized from muscle (Taylor and Venn, 1979;Barber and Blake, 1981, 1985; Pazos et al., 1997), lipid levels in the adductor musclemay also decline during gonadal maturation (Epp et al., 1988). Clearly, beyond itsobvious contractile role, the adductor muscle fulfills an important role as a site of reservedeposition in scallops.
When muscle macromolecules are mobilized to support the metabolic requirements ofother tissues, both the levels of energetic reserves, such as glycogen and lipid, and thoseof enzymes or contractile proteins can be affected. For example, during their non-feeding spawning migration, sockeye salmon, Oncorhynchus nerka, first metabolize theirlipid deposits in muscle and then break down muscle enzymes and sarcoplasmic proteins(Mommsen et al., 1980). During starvation, cod, Gadus morhua, first mobilise hepaticlipid, then hepatic and muscle glycogen and finally muscle proteins (Black and Love,1986). During the nutritional stress caused by winter conditions, mussels experience ahigh energy loss from body tissues and proteins accounts for 75% of this loss (Gabbottand Bayne, 1973). During gametogenesis in Argopecten irradians irradians, digestivegland lipid reserves are first mobilized followed by muscle glycogen and finally proteinreserves (Barber and Blake, 1981). The sparing of muscle proteins during starvationlikely reflects the fact that proteins play more roles than simply serving as an energeticreserve. Effectively, the mobilisation of muscle proteins during starvation or reproduc-tive maturation decreases muscle metabolic capacities (Guderley et al., 1994, 1996),particularly the glycolytic capacity of white muscle. Studies of relative utilization ofdietary protein for energy and biosynthesis by the mussel Mytilus edulis havedemonstrated a continuous conservation of amino-N over amino-C to satisfy bio-synthetic demands (Kreeger et al., 1995, 1996).
The extensive mobilization of muscle reserves during gametogenesis in scallopssuggests that muscle metabolic capacities change during gonadal maturation. However,little is known of the impact of gonadal maturation upon the metabolic capacities of theadductor muscle. The oxidative capacities and respiratory control ratios of mitochondria

´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 31
isolated from the adductor muscle of Euvola (Pecten) ziczac during their first spawningwere lower than in those isolated at other periods, apparently due to the low foodavailability and high temperatures in the preceding period (Boadas et al., 1997). Whengonadal maturation is induced in recently spawned scallops, muscle metabolic capacitiesmay first rise and then fall if muscle proteins are mobilized for gonadal maturation(Pazos et al., 1997). If muscle enzymes are simply used to provide energy and aminoacids, all enzymes should decrease in parallel during protein mobilization. On the otherhand, enzymes which facilitate reserve mobilization or which are restricted to specificorganelles may be spared. For example, as glycogen phosphorylase liberates glucosemoieties from glycogen, its levels may be maintained.
Reinitiation of gonadal maturation following spawning is of particular interest for theaquaculture industry, which depends upon a ready availability of high quality larvae.The extent and quality of this gonadal recuperation depend upon the available food, withcertain diets leading to faster gonadal maturation, greater fertilization success and
´improved larval survival than others (Martınez et al., 2000). Thus, in Argopectenpurpuratus, gonadal maturation is faster at 168C than at 208C and the best larval survivalis obtained with a microalgal diet supplemented with a lipid emulsion. Duringconditioning of A. purpuratus, considerable time is required for gonadal maturation,
´even with appropriate diets and temperatures (Martınez et al., 2000). Thus, recentlyspawned individuals are likely to replenish tissue reserves before undertaking gonadalmaturation. The levels of biochemical components in muscle would increase before anyimprovement in gonadal status is apparent and then decrease during gonadal maturation,particularly if food availability is insufficient to cover the requirements of gonadalmaturation. Under this scenario, muscle metabolic capacities may first rise and then fallduring broodstock conditioning, although enzymes which play a role in reservemobilisation may be spared.
To examine these possibilities, we evaluated the biochemical composition of muscleand gonad and muscle metabolic capacities in the scallop, A. purpuratus, duringconditioning of recently spawned individuals at 16 and 208C using three diets, oneconsisting of microalgae, another of microalgae supplemented with a lipid emulsion andthe last of microalgae supplemented with carbohydrate. We measured levels ofcarbohydrates, lipids and proteins in muscle and gonad. Since the scallops conditioned at168C showed more complete gonadal maturation than those at 208C and as we wished toascertain the impact of mobilization of muscle reserves in support of gonadal maturationupon muscle metabolic capacities, we determined the activities of glycogen phosphor-ylase (GP), pyruvate kinase (PK), octopine dehydrogenase (ODH) and citrate synthase(CS) only in the muscles of scallops conditioned at 168C. GP is responsible for theliberation of glucosyl units from glycogen, PK catalyses an ATP generating reaction inglycolysis, whereas ODH catalyses the reductive condensation of pyruvate and arginine,thereby regenerating NAD. CS is a component of the Krebs cycle and serves as a markerof mitochondrial abundance. If these enzymes show the same variation duringmobilization of muscle reserves, it would indicate non-selective protein breakdown.Specific changes, in which certain activities were maintained whereas others decline,would indicate targetted protein mobilization. We predicted that the levels of theglycolytic enzymes, PK and ODH, would decrease whereas GP would be spared during

´32 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
mobilization of muscle reserves, given its role in liberating glucose equivalents fromglycogen. We reasoned that the localization of CS in the mitochondrial matrix wouldprotect it during reserve mobilization.
2. Materials and methods
2.1. Feeding and sampling of scallops
Scallops (experiment 1: shell height of 80–90 mm; experiment 2: shell height of70–80 mm) just obtained in mature stage from Culture Centers in Tongoy Bay (308 S)were induced to spawn by increasing the temperature and adding excess microalgaeconsecutive years. The spent animals were distributed among nine tanks (34 animals pertank) at 168C and nine tanks at 208C. Three groups at each temperature were fed with amixture of microalgae (50% of Isochrysis galbana 1 50% of Chaetoceros gracilis),three other groups received 70% of the same mixture of algae plus 30% carbohydrates(commercial potato starch), and the other tanks received 70% microalgae and 30% of alipid emulsion (provided by Artemia Reference Center in Belgium, corresponded to theEmDHA whose composition is described in Caers et al., 1999). The gross biochemicalcomposition of these diets is given in Table 1. More details about diets and theirabsorption efficiency are described in Navarro et al. (2000). The daily food ration,supplied continuously by a peristaltic pump, amounted to 3% of the animals’ dry massduring the first experiment and to 6% during the second experiment. Both experimentswere run from January to March in consecutive years (1998 and 1999).
Before beginning conditioning, five spent scallops were sampled to evaluate theirbiochemical status (this sample was called ‘control’). Periodically, similar analyses wereperformed for three scallops from each experimental treatment (each animal from adifferent tank) to follow the course of gonadal recovery and the levels of the biochemicalcomponents. For this, the scallops were sorted into three groups according to theirvisually determined degree of maturation and one scallop from the largest group in eachtank was sampled. The duration of the experiments was determined according to theprogress of gonadal ripening; when about 40% of the scallops appeared mature in any ofthe tanks, the entire experiment was ended to avoid spontaneous spawning. When thefirst experiment (3% ration) was ended, the gonadal indices in the groups with the bestperformance approached 10% whereas in the second experiment (6% ration), the
´gonadal indices of the best groups were greater than 13% (Martınez et al., 2000).
Table 1aBiochemical composition of the three experimental diets used for conditioning A. purpuratus
Proteins Lipids Carbohydrates
Microalgae 212.99611.02 75.0566.69 62.9564.16Microalgae 1 lipids 191.9568.45 108.8762.92 69.8267.07Microalgae 1 carbohydrates 200.00615.43 80.00616.11 215.00626.26
a 21The concentrations are expressed in mg g . Values are the mean6S.E. (N 5 3).

´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 33
2.2. Biochemical and enzymatic analyses
The scallops were sacrificed and their gonads and muscles were weighed beforefreezing and storage in liquid nitrogen for subsequent biochemical determinations. Themuscle samples used for enzymatic measurements were transported on dry ice to
´Universite Laval where they were stored at 2 708C before analysis. The levels ofproteins, total carbohydrates and lipids in muscle and gonad were analysed following the
21´methods of Martınez (1991). Muscle levels were presented as concentrations (mg gdry mass), whereas gonadal components were presented as total gonadal contents (forthe female and male portions respectively) given the marked changes in gonadal size andno significant changes in muscle size during conditioning (gonadal size increase wasmore than 300%)
For measurements of PK and ODH, portions of the adductor muscle were homogen-ized in 50 mM imidazole–HCl, 2 mM Na -EDTA, 5 mM EGTA, 1 mM dithiothreitol2
and 0.1% Triton X-100, pH 6.6. For GP and CS measurements, samples of adductormuscle were homogenized in this buffer at pH 7.2 supplemented with 150 mM KCl. ThepH of all solutions was adjusted at room temperature. Homogenisation of the musclesamples (1:10 m/v) was carried out on ice using a Polytron (Brinkmann, Rexdale,Ontario) at 50% maximal speed for three, 30-s periods separated by 1-min rest periods.
Spectrophotometric measurements were carried out using a Beckman DU-640 UV-Visspectrophotometer. The cuvette temperature was controlled at 168C by a Haake G8refrigerating circulator. Measurements were made at 340 nm for PK, ODH and GPfollowing the changes in absorbance of NAD(P)H, whereas measurements of CSfollowed the absorbance of 5,59dithio-bis(2-nitro)benzoic acid (DTNB) at 412 nm.Enzyme activities were measured using the following conditions:
Pyruvate kinase: 50 mM imidazole–HCl, 13 mM MgSO , 100 mM KCl, 5 mM ADP,4
0.2 mM NADH, pH 6.6, 5 U lactate dehydrogenase and 5 mM phosphoenolpyruvate(omitted for the control).
Octopine dehydrogenase: 50 mM imidazole–HCl, 2 mM Na EDTA, 5 mM EGTA, 12
mM K-cyanide, 0.2 mM NADH, 6 mM Na-pyruvate and 6 mM arginine (omitted for thecontrol). The control activity, which represents that of lactate dehydrogenase, wasvirtually nil.
Glycogen phosphorylase: 50 mM imidazole–HCl, 80 mM KH PO , 5 mM Mg-2 4
acetate, 2.5 mM Na EDTA, 0.8 mM AMP, 0.5 mM cyclic AMP, 0.6 mM NADP, 0.0042
mM glucose-1,6-diphosphate, 2 U glucose-6-phosphate dehydrogenase, 2.5 U phosphog-21lucomutase, 10 mg ml glycogen, pH 7.5.
Citrate synthase: 75 mM Tris–HCl, 0.25 mM DTNB, 0.35 mM acetyl CoA, 0.05 mMoxaloacetate, pH 8.0. Oxaloacetate was omitted for the control.
The mmolar extinction coefficients were 6.22 and 13.6 for NAD(P)H and DTNB,respectively. Enzyme activities are expressed as international units (mmol of substrate
21 21converted to product min ) 3 g wet mass.
2.3. Calculations and statistical comparisons
The gonadal contents of biochemical components were established by measuring the

´34 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
concentration of the component in a sample of the female and male gonad, and thenextrapolated to the total content of that section by assuming that the gonad was equallydivided into male and female sections (sections never differed by more than 10% andhistological analysis showed a clear separation between them). The effect of diet andduration of conditioning upon the levels of biochemical components in gonad andmuscle, upon enzyme levels in muscle and upon muscle mass and % water content wasexamined using two-way ANOVAs which considered diet and duration of conditioningas the two factors (Systat). When no interactions were observed between the factors,one-way ANOVAs considering only the effect of duration of conditioning were run foreach diet. When significant effects of duration of conditioning were observed, Tukey aposteriori tests were used to identify significant differences (P , 0.05). For eachexperiment, the data obtained under the two thermal regimes were analysed separately.
3. Results
3.1. Biochemical composition of muscle
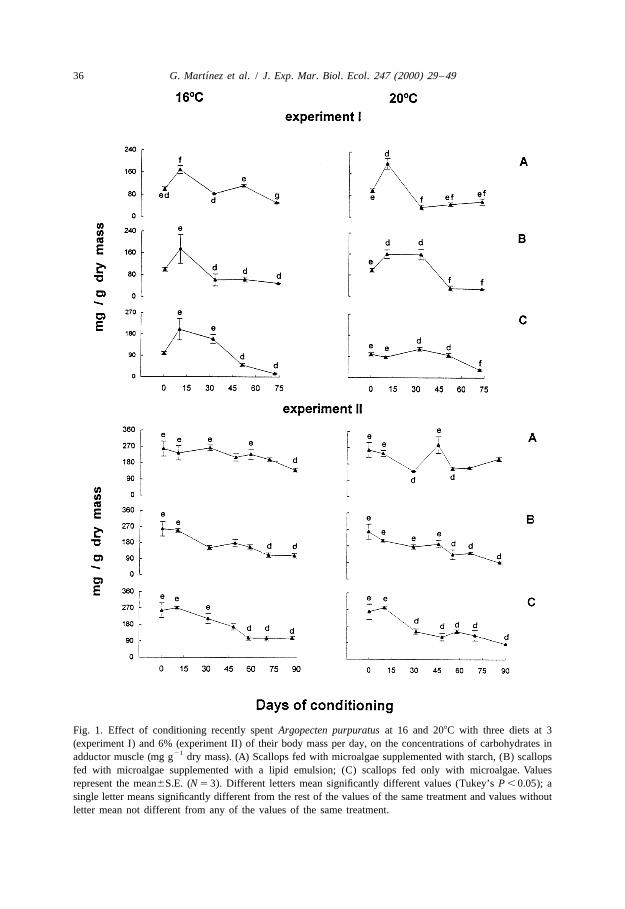
When recently spawned Argopecten purpuratus, maintained at 16 or 208C, weresupplied with the three experimental diets at 3 or 6% of their dry mass per day,carbohydrate levels in the adductor muscle showed the most pronounced fluctuations.During experiment I significant initial increases were followed by marked decreases inall treatments (Table 2, Tukey a posteriori tests, P , 0.05) (Fig. 1). Initial carbohydratelevels were higher in the second experiment than in the first (Student’s test, P , 0.05).Presumably this is why muscle carbohydrates did not rise at the start of the experimentII.
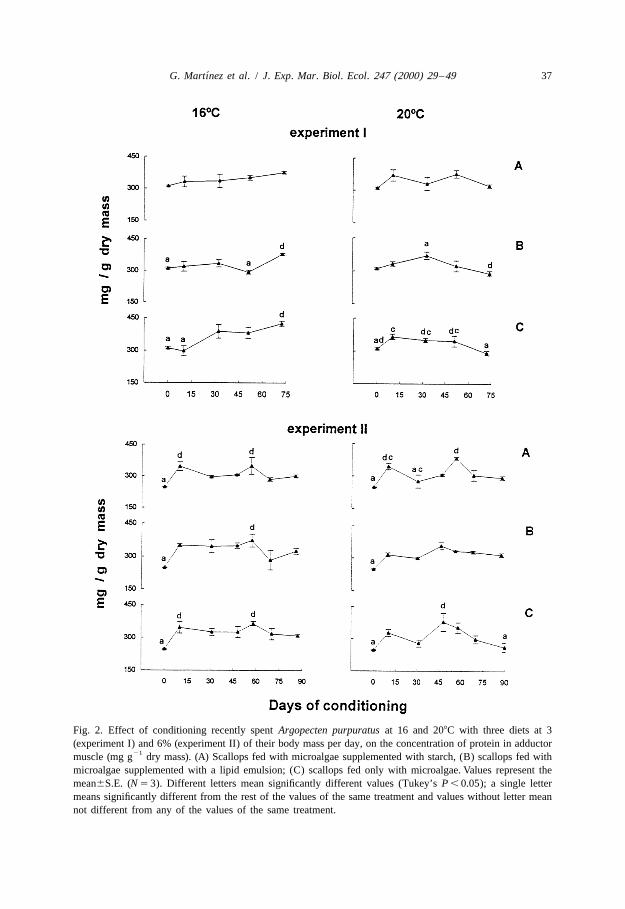
During conditioning at 3%, muscle protein levels showed slight statistical differencesbetween diets at both temperatures (Table 2, Fig. 2); slight increases were detected at theend of conditioning of scallops fed with microalgae plus starch or only microalgae at168C. At the end of conditioning at 208C, muscle protein levels declined in scallopsconditioned with the mixture microalgae–carbohydrate. In contrast to experiment I,muscle protein levels of animals fed at 6% of their dry mass per day rose at the start ofconditioning, but did not increase further nor fall below initial values in scallopsconditioned at either temperature.
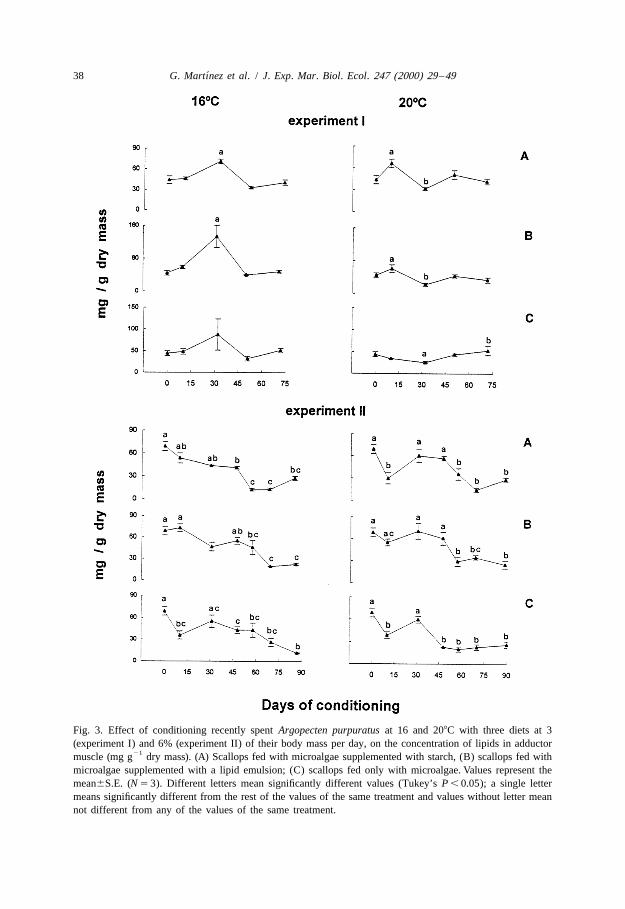
In the first experiment, lipid levels in the adductor muscle were higher, particularly onday 32, in the scallops fed microalgae supplemented with lipids or starch than in themuscle of scallops fed only microalgae (P , 0.05) (Fig. 3). Initial muscle lipid levelswere higher (Student’s test, P , 0.05) in the scallops conditioned at the higher level offood (experiment 2) and declined gradually during their conditioning at 168C; at 208C,except with enriched-lipid diet, there was an initial fall (on day 10) then initial valueswere recovered, followed by a gradual decline during the final maturation of the gonad.
The changes in the biochemical components in muscle during conditioning were notaccompanied by shifts in water content nor wet mass (ANOVA, P . 0.05).
´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 35
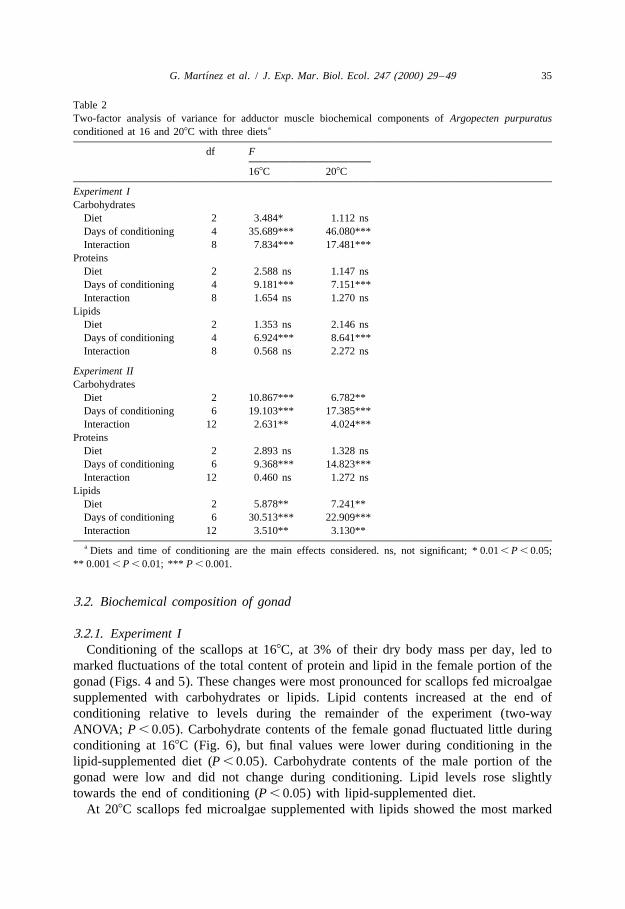
Table 2Two-factor analysis of variance for adductor muscle biochemical components of Argopecten purpuratus
aconditioned at 16 and 208C with three diets
df F
168C 208C
Experiment ICarbohydrates
Diet 2 3.484* 1.112 nsDays of conditioning 4 35.689*** 46.080***Interaction 8 7.834*** 17.481***
ProteinsDiet 2 2.588 ns 1.147 nsDays of conditioning 4 9.181*** 7.151***Interaction 8 1.654 ns 1.270 ns
LipidsDiet 2 1.353 ns 2.146 nsDays of conditioning 4 6.924*** 8.641***Interaction 8 0.568 ns 2.272 ns
Experiment IICarbohydrates
Diet 2 10.867*** 6.782**Days of conditioning 6 19.103*** 17.385***Interaction 12 2.631** 4.024***
ProteinsDiet 2 2.893 ns 1.328 nsDays of conditioning 6 9.368*** 14.823***Interaction 12 0.460 ns 1.272 ns
LipidsDiet 2 5.878** 7.241**Days of conditioning 6 30.513*** 22.909***Interaction 12 3.510** 3.130**a Diets and time of conditioning are the main effects considered. ns, not significant; * 0.01 , P , 0.05;
** 0.001 , P , 0.01; *** P , 0.001.
3.2. Biochemical composition of gonad
3.2.1. Experiment IConditioning of the scallops at 168C, at 3% of their dry body mass per day, led to
marked fluctuations of the total content of protein and lipid in the female portion of thegonad (Figs. 4 and 5). These changes were most pronounced for scallops fed microalgaesupplemented with carbohydrates or lipids. Lipid contents increased at the end ofconditioning relative to levels during the remainder of the experiment (two-wayANOVA; P , 0.05). Carbohydrate contents of the female gonad fluctuated little duringconditioning at 168C (Fig. 6), but final values were lower during conditioning in thelipid-supplemented diet (P , 0.05). Carbohydrate contents of the male portion of thegonad were low and did not change during conditioning. Lipid levels rose slightlytowards the end of conditioning (P , 0.05) with lipid-supplemented diet.
At 208C scallops fed microalgae supplemented with lipids showed the most marked
´36 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Fig. 1. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the concentrations of carbohydrates in
21adductor muscle (mg g dry mass). (A) Scallops fed with microalgae supplemented with starch, (B) scallopsfed with microalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Valuesrepresent the mean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); asingle letter means significantly different from the rest of the values of the same treatment and values withoutletter mean not different from any of the values of the same treatment.
´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 37
Fig. 2. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the concentration of protein in adductor
21muscle (mg g dry mass). (A) Scallops fed with microalgae supplemented with starch, (B) scallops fed withmicroalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Values represent themean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); a single lettermeans significantly different from the rest of the values of the same treatment and values without letter meannot different from any of the values of the same treatment.
´38 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Fig. 3. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the concentration of lipids in adductor
21muscle (mg g dry mass). (A) Scallops fed with microalgae supplemented with starch, (B) scallops fed withmicroalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Values represent themean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); a single lettermeans significantly different from the rest of the values of the same treatment and values without letter meannot different from any of the values of the same treatment.
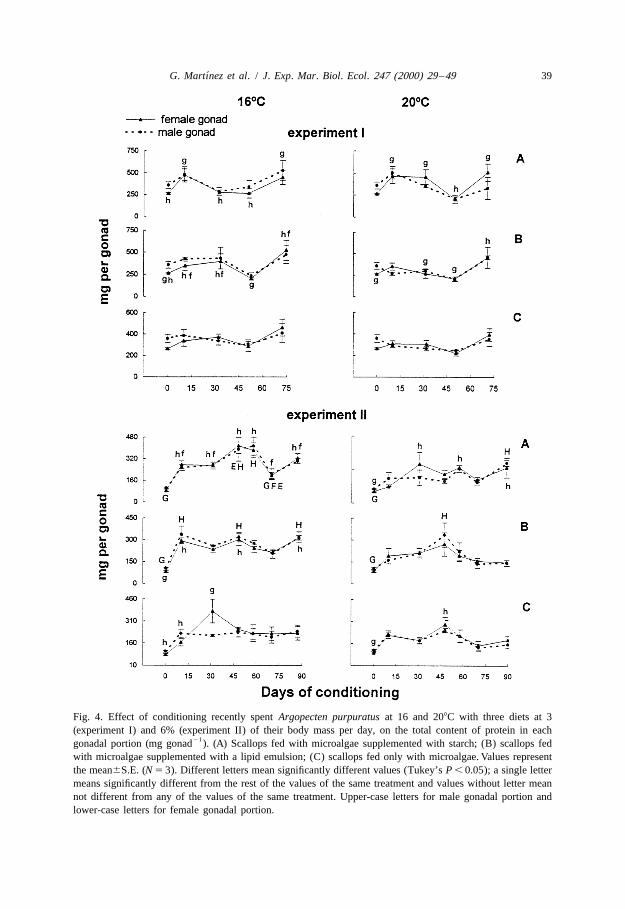
´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 39
Fig. 4. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the total content of protein in each
21gonadal portion (mg gonad ). (A) Scallops fed with microalgae supplemented with starch; (B) scallops fedwith microalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Values representthe mean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); a single lettermeans significantly different from the rest of the values of the same treatment and values without letter meannot different from any of the values of the same treatment. Upper-case letters for male gonadal portion andlower-case letters for female gonadal portion.

´40 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Fig. 5. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the total content of lipids in each
21gonadal portion (mg gonad ). (A) Scallops fed with microalgae supplemented with starch; (B) scallops fedwith microalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Values representthe mean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); a single lettermeans significantly different from the rest of the values of the same treatment and values without letter meannot different from any of the values of the same treatment. Upper-case letters for male gonadal portion andlower-case letters for female gonadal portion.
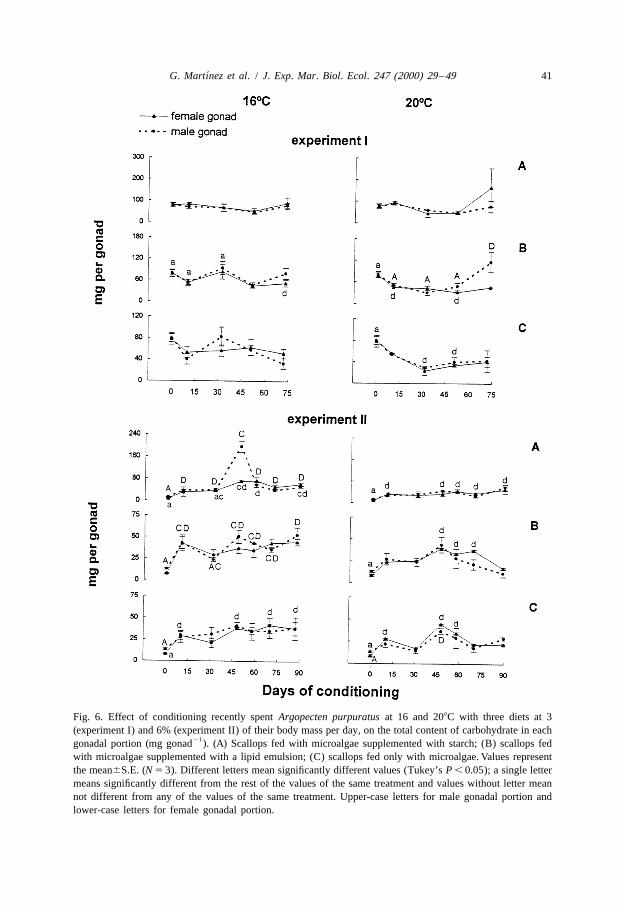
´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 41
Fig. 6. Effect of conditioning recently spent Argopecten purpuratus at 16 and 208C with three diets at 3(experiment I) and 6% (experiment II) of their body mass per day, on the total content of carbohydrate in each
21gonadal portion (mg gonad ). (A) Scallops fed with microalgae supplemented with starch; (B) scallops fedwith microalgae supplemented with a lipid emulsion; (C) scallops fed only with microalgae. Values representthe mean6S.E. (N 5 3). Different letters mean significantly different values (Tukey’s P , 0.05); a single lettermeans significantly different from the rest of the values of the same treatment and values without letter meannot different from any of the values of the same treatment. Upper-case letters for male gonadal portion andlower-case letters for female gonadal portion.

´42 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
changes in gonadal contents of carbohydrates and proteins. Diet had a significant effecton the protein content of the female gonad (Fig. 4), as well as on the lipid andcarbohydrate content of the male gonad (two-way ANOVA; P , 0.05) (Figs. 5 and 6).The fluctuations of the levels of biochemical components led the duration of con-ditioning to significantly affect the contents of proteins and carbohydrates in the femalegonad and on all of the components in the male gonad. Inspection of the overall changesin the gonadal contents of biochemical components indicates that gonadal levels of lipidsand proteins rise during final maturation, whereas increases in proteins which are notaccompanied by increases in lipids may occur well before gonadal maturation.
3.2.2. Experiment IIThe protein content of the female gonad markedly increased at the start of
conditioning with all diets and at both temperatures (Fig. 4). Subsequent fluctuations ofprotein contents occurred under all experimental conditions, but were more pronouncedat 168C. The slight decrease at 70 days was likely to due to a partial spawning. Proteincontents of the female gonad were affected by diet and duration of conditioning at 168Cand only by the duration of conditioning at 208C (Table 3, two-way ANOVA; P , 0.05).At 168C, lipid contents increased considerably during the last 20 days of the experiment.Lipid and carbohydrate contents of the female gonad were affected by duration ofconditioning and diet at 168C (Table 3, Figs. 5 and 6). For each diet, the scallops fed at208C tended to have lower gonadal protein contents than those fed at 168C. The increaseof lipid contents towards the end of conditioning was less pronounced at 20 than at168C.
Protein contents of the male gonad rose considerably at the start of conditioning, andoscillated before showing their final increase in the scallops conditioned at 168C with thetwo supplemented diets (Fig. 4). Again these oscillations may reflect partial spawnings.The carbohydrate contents of the male gonad were affected by both duration ofconditioning and diet at both temperatures (Table 3, two-way ANOVA; P , 0.05). At 16and 208C, lipid and protein contents were only affected by duration of conditioning(Table 3). At 208C, a rise in protein contents was noted only for scallops conditionedwith the microalgae supplemented with carbohydrates. In contrast to the scallopsconditioned in the first experiment, the lipid contents of the male gonad changed littleduring the final stages of conditioning.
3.3. Muscle enzyme levels
21Muscle enzyme levels were expressed as U g wet mass since the water content ofthe muscles did not change during conditioning at 168C (ANOVA, F 5 0.332,3,68
P . 0.05). A two-factor ANOVA with time of conditioning and diet as factors indicatedthat time of conditioning significantly affected GP levels (P , 0.001), but that diet didnot. No interaction was found between time and diet. Both diet (P 5 0.044) andconditioning time (P 5 0.009) significantly affected PK levels. Throughout conditioning,scallops fed microalgae alone maintained lower levels of PK than those fed microalgaewith lipids, whereas those fed microalgae supplemented with carbohydrates showeddecreases followed by an increase at the end of the experiment (Fig. 7). ODH levels
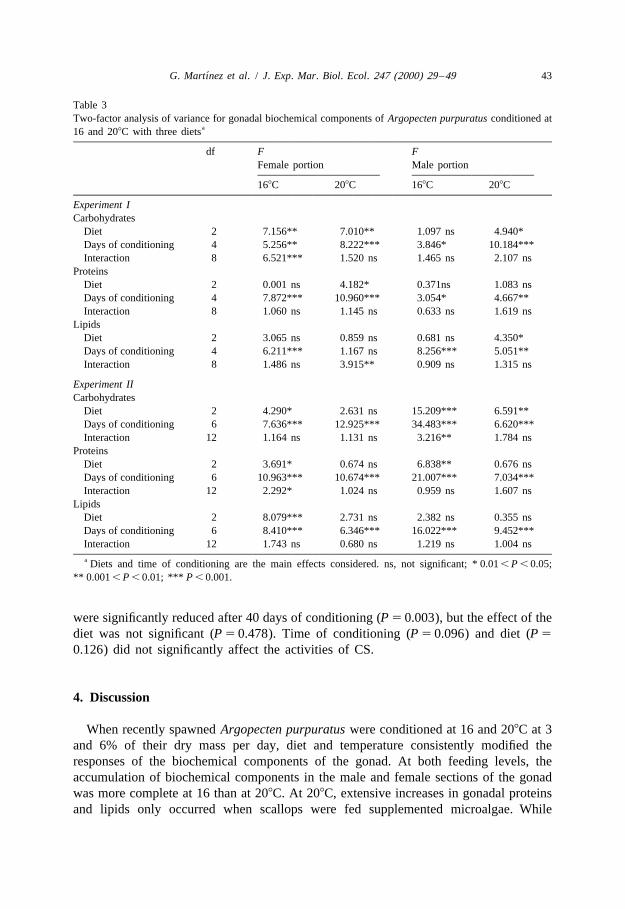
´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 43
Table 3Two-factor analysis of variance for gonadal biochemical components of Argopecten purpuratus conditioned at
a16 and 208C with three diets
df F FFemale portion Male portion
168C 208C 168C 208C
Experiment ICarbohydrates
Diet 2 7.156** 7.010** 1.097 ns 4.940*Days of conditioning 4 5.256** 8.222*** 3.846* 10.184***Interaction 8 6.521*** 1.520 ns 1.465 ns 2.107 ns
ProteinsDiet 2 0.001 ns 4.182* 0.371ns 1.083 nsDays of conditioning 4 7.872*** 10.960*** 3.054* 4.667**Interaction 8 1.060 ns 1.145 ns 0.633 ns 1.619 ns
LipidsDiet 2 3.065 ns 0.859 ns 0.681 ns 4.350*Days of conditioning 4 6.211*** 1.167 ns 8.256*** 5.051**Interaction 8 1.486 ns 3.915** 0.909 ns 1.315 ns
Experiment IICarbohydrates
Diet 2 4.290* 2.631 ns 15.209*** 6.591**Days of conditioning 6 7.636*** 12.925*** 34.483*** 6.620***Interaction 12 1.164 ns 1.131 ns 3.216** 1.784 ns
ProteinsDiet 2 3.691* 0.674 ns 6.838** 0.676 nsDays of conditioning 6 10.963*** 10.674*** 21.007*** 7.034***Interaction 12 2.292* 1.024 ns 0.959 ns 1.607 ns
LipidsDiet 2 8.079*** 2.731 ns 2.382 ns 0.355 nsDays of conditioning 6 8.410*** 6.346*** 16.022*** 9.452***Interaction 12 1.743 ns 0.680 ns 1.219 ns 1.004 nsa Diets and time of conditioning are the main effects considered. ns, not significant; * 0.01 , P , 0.05;
** 0.001 , P , 0.01; *** P , 0.001.
were significantly reduced after 40 days of conditioning (P 5 0.003), but the effect of thediet was not significant (P 5 0.478). Time of conditioning (P 5 0.096) and diet (P 5
0.126) did not significantly affect the activities of CS.
4. Discussion
When recently spawned Argopecten purpuratus were conditioned at 16 and 208C at 3and 6% of their dry mass per day, diet and temperature consistently modified theresponses of the biochemical components of the gonad. At both feeding levels, theaccumulation of biochemical components in the male and female sections of the gonadwas more complete at 16 than at 208C. At 208C, extensive increases in gonadal proteinsand lipids only occurred when scallops were fed supplemented microalgae. While
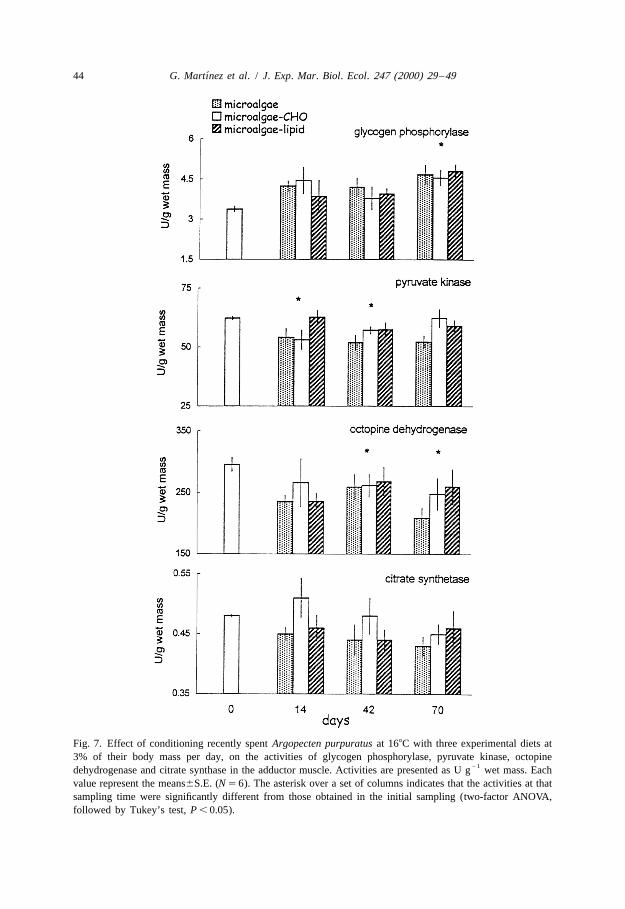
´44 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Fig. 7. Effect of conditioning recently spent Argopecten purpuratus at 168C with three experimental diets at3% of their body mass per day, on the activities of glycogen phosphorylase, pyruvate kinase, octopine
21dehydrogenase and citrate synthase in the adductor muscle. Activities are presented as U g wet mass. Eachvalue represent the means6S.E. (N 5 6). The asterisk over a set of columns indicates that the activities at thatsampling time were significantly different from those obtained in the initial sampling (two-factor ANOVA,followed by Tukey’s test, P , 0.05).

´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 45
gonadal accumulation of biochemical components was more extensive in the scallopsfed at 6% of their dry mass per day, similar responses to the experimental diets andtemperatures were obtained during the two experiments. At the higher temperature, themicroalgal diet must be deficient in some components since even higher feeding levels(6% respect to 3% of the body mass) did not allow sufficient accumulation ofbiochemical components in the gonad. Significantly lower values of scope for growthwere found for the microalgal diet (Navarro et al., 2000). We chose this highertemperature, which is above the range experienced by this population of A. purpuratusto evaluate whether gonadal maturation is accelerated by this fairly small increase intemperature. Instead, our results show that this higher temperature impedes maturation
´(Martınez et al., 2000).In both experiments, muscle carbohydrate levels decreased during accumulation of
protein and lipid in the gonad. According to the status of the control scallops,conditioning led to initial increases in muscle carbohydrate levels (experiment I) or toinitial maintenance of high levels (experiment II). Subsequently, muscle carbohydratelevels declined gradually, particularly towards the end of the experiment when gonadalprotein and lipid contents rose markedly. The scallops fed at 6% of their mass per daydepleted muscle carbohydrates to a lesser extent than those fed at 3% in muscle,maintaining 3–5-fold higher values. Muscle carbohydrate reserves are likely mobilisedto provide energy or precursors for lipid or protein synthesis in the gonad. In Pectenmaximus, gonadal growth in periods of poor food quality has been shown to coincidewith a fall of reserves in muscle (Pazos et al., 1997). Measurements of muscle glycogenlevels and ratios of oxygen consumption to nitrogen excretion (O/N ratios) duringgametogenesis in Argopecten irradians concentricus (Barber and Blake, 1985), indicatethat muscle carbohydrate is converted to lipid which is stored in developing ova.
In agreement with our predictions, the activities of glycogen phosphorylase in theadductor muscle did not decrease during conditioning, but increased during final gonadalmaturation. Glycogen phosphorylase liberates glucose moieties from glycogen andwould be important for transfer of carbohydrate reserves from muscle to other tissues,particularly if glucosidase activities are in the range typical of muscle (Fournier andGuderley, 1993). After phosphorylase produces glucose-1-phosphate, it can then beconverted to glucose-6-phosphate and liberated as glucose after dephosphorylation byglucose-6-phosphatase. The levels of these enzymes should also be maintained duringmobilization of muscle carbohydrate.
Reserve mobilization from muscle was accompanied by decreases in the levels ofODH, fluctuations in those of PK and no changes in CS levels. As protein levels did notdecrease in the scallops conditioned at 168C, a generalized depletion of muscle proteinscannot explain these changes. ODH, one of the enzymes which maintains redox balanceduring anaerobic glycolysis (Fields, 1988), decreased during gonadal maturation inscallops fed the three diets. ODH activities follow carbohydrate levels, suggesting theirco-regulation. The activities of PK only decreased in scallops fed microalgae, whereasthose fed the supplemented diets either showed no changes (lipids) or decreasesfollowed by increases (carbohydrates). PK has a major role in glycolytic regulation inmarine invertebrates (Munday et al., 1980; Gaitanaki et al., 1990), being subject to aninhibitory phosphorylation (Holwerda et al., 1981, 1983; Michaelidis et al., 1988;

´46 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Whitwam and Storey, 1990). This regulatory role may require its protection duringmobilization of muscle reserves. Interestingly, PK levels were maintained best in thescallops conditioned with the most adequate diet (microalgae supplemented with lipids).
The lack of change in CS activity may be due to its mitochondrial localization.Changes in nutritional status bring concomitant modifications of muscle metaboliccapacities in numerous fish species. Ration level affects the activity of lactatedehydrogenase in white muscle of sablefish (Anoplopoma fimbria) (Sullivan andSomero, 1983). Activities of several glycolytic enzymes are positively correlated withgrowth rates in saithe (Pollachius virens) after 2 weeks under differing feedingconditions (Mathers et al., 1992) and in cod (Gadus morhua) after 6–16 weeks exposureto different feeding conditions (Pelletier et al., 1993, 1994; Dutil et al., 1998).Mitochondrial enzymes change less during shifts in growth rates than glycolyticenzymes (Blier et al., 1997).
The impact of changes in reproductive status on muscle metabolic capacities inectotherms has not been systematically examined, but androgen stimulation of chumsalmon (Oncorhynchus keta) decreases the levels of sarcoplasmic proteins (Ando et al.,1986), as does the spawning migration of sockeye salmon (O. nerka) (Mommsen et al.,1980). Since salmon feed little during reproductive maturation, the requirements ofgonadal production are largely covered by reserves in the organism. White musclecomponents thus become prime candidates for deposition in the gonad. Changes inprotein levels should have a more direct impact upon the metabolic capacities of musclethan the depletion of carbohydrate reserves.
In scallops, a central role of carbohydrate reserves of adductor muscle in gonadalproduction has been shown for Chlamys varia (Shafee, 1981), Argopecten concentricus(Barber and Blake, 1981), Argopecten irradians irradians (Epp et al., 1988), Placopec-ten magellanicus (Couturier and Newkirk, 1991) as well as for Argopecten purpuratus
´ ´(Martınez, 1991; Martınez and Mettifogo, 1998). Interestingly, this role remains evenwhen food availability was quite high as in our second experiment. Presumably, at evenhigher food levels, the requirement for the depletion of carbohydrate reserves in musclecould disappear. This argument is supported by the maintenance of higher musclecarbohydrate levels in scallops fed at 6% rather than 3% of their dry mass. Muscleprotein levels declined during conditioning of adult Argopecten purpuratus when foodavailability was relatively low (3%) and temperatures high (208C). In Euvola (Pecten)ziczac, oxidative capacities of mitochondria isolated from the adductor muscle ofscallops which had spawned after a period of high temperatures and low foodavailability (May) decreased relative to those of mitochondria isolated in more favorableperiods (Boadas et al., 1997). Protein levels in the adductor muscle of Euvola werelower in May than in the other periods.
Although muscle protein levels remained stable or increased during conditioning ofArgopecten purpuratus at 168C, muscle metabolic capacities decreased. While this mayreflect the greater sensitivity of enzymatic measurements, this pattern is also suggestiveof modulation of muscle enzyme levels in accordance with their specific roles. Theenzyme that decreased most during reproductive conditioning was ODH, a terminalenzyme of anaerobic glycolysis in molluscs. In gastropods, fish, frogs and mammals,glycolytic enzymes in muscle can be citosolic or attached to structural macromolecules

´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 47
(Clarke et al., 1984; Plaxton and Storey, 1986; Lowery et al., 1987; Brooks and Storey,1991; Guderley et al., 1989; Huber and Guderley, 1993). Enzyme binding is enhancedwhen glycolytic rates increase, such as during burst exercise. Hence, when glycogenlevels decrease, the binding sites for glycolytic enzymes may decrease, accelerating theturnover of these enzymes.
In conclusion, during conditioning of adult Argopecten purpuratus, mobilization ofmuscle carbohydrate reserves appears to play an important role as it consistentlyaccompanied gonadal maturation. Nonetheless, the depletion of muscle carbohydratereserves was less extensive when scallops were fed with a higher ration level. Changesin muscle metabolic capacities occurred despite the maintenance of muscle proteinlevels. The enhanced glycogen phosphorylase activities in muscle towards the end ofconditioning may facilitate this carbohydrate mobilization. The decrease in ODH levelsduring conditioning may reflect their co-regulation with glycogen levels. Finally,conditioning Argopecten purpuratus at 208C did not accelerate gonadal maturation, ifanything the increased metabolic demands brought by this higher temperature con-sistently decreased the accumulation of biochemical components in the gonad and led todepletion of muscle reserves, particularly at the lower ration level.
Acknowledgements
´This study was supported by a grant from the Programa Acuicultura y BiotecnologıaMarina, 1 (97), FONDAP, Chile (Sub-programa Invertebrados). Enzymatic studies werepartially supported by a grant to Helga Guderley from NSERC of Canada. [SS]
References
Ando, S., Hatano, M., Zama, K., 1986. Protein degradation and protease activity of chum salmon(Oncorhynchus keta) muscle during spawning migration. Fish Physiol. Biochem. 1, 17–26.
Barber, B., Blake, N., 1981. Energy storage and utilization in relation to gametogenesis in Argopectenirradians concentricus (Gay). J. Exp. Mar. Biol. Ecol. 52, 121–134.
Barber, B., Blake, N., 1985. Substrate catabolism related to reproduction in the bay scallop Argopectenirradians concentricus, as determined by O/N and RQ physiological indexes. Mar. Biol. 87, 13–18.
Black, D., Love, R.M., 1986. The sequential mobilization and restoration of energy reserves in tissues ofAtlantic cod during starvation and refeeding. J. Comp. Physiol. B. 156, 469–479.
Blier, P.U., Pelletier, D., Dutil, J.D., 1997. Does aerobic capacity set a limit upon fish growth rate? Rev. FishSci. 5, 323–340.
Boadas, M.A., Nusetti, O., Mundarain, F., Lodeiros, C., Guderley, H., 1997. Seasonal variation in theproperties of muscle mitochondria from the tropical scallop Euvola (Pecten) ziczac. Mar. Biol. 28,247–255.
´ ´ ´Brea, J., 1986. Variaciones energeticas estacionales en la composicion quımica de Pecten ziczac (Linne 1758)´ ´en relacion con el metabolismo energetico, reproductivo y crecimiento. Thesis, Biology Department,
´Universidad de Oriente, Cumana, Venezuela.Brooks, S.P.J., Storey, K.B., 1991. Studies on the regulation of enzyme binding during anoxia in isolated
tissues of Busycon canaliculatum. J. Exp. Biol. 156, 193–204.

´48 G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49
Caers, M., Coutteau, P., Cure, K., Morales, V., Gajardo, G., Sorgeloos, P., 1999. The Chilean scallopArgopecten purpuratus (Lamarck, 1819). II. Manipulation of the fatty acid composition and lipid content ofthe eggs via lipid supplementation of the broodstock diet. Comp. Biochem. Physiol. B. 123, 97–103.
Clarke, F.M., Stephan, P., Huxham, G., Hamilton, D., Morton, D.J., 1984. Metabolic dependence of glycolyticenzyme binding in rat and sheep heart. Eur. J. Biochem. 138, 643–649.
Couturier, C.Y., Newkirk, G.F., 1991. Biochemical and gametogenetic cycles in scallops, Placopectenmagellanicus (Gmelin 1791) held in suspension culture. In: Shumway, S.E., Sandiger, P.A. (Eds.), AnInternational Compendium of Scallop Biology and Culture, World Aquaculture Workshops, Vol. 1, WorldAquaculture Society, Baton Rouge, LA, pp. 107–117.
Dutil, J.D., Lambert, Y., Guderley, H., Blier, P.U., Pelletier, D., Desroches, M., 1998. Nucleic acids andenzymes in Atlantic cod (Gadus morhua) differing in condition and growth rate trajectories. Can. J. Fish.Aquat. Sci. 55, 788–795.
Epp, J., Bricelj,V., Malouf, R., 1988. Seasonal partitioning and utilization of energy reserves in two age classesof the bay scallop Argopecten irradians irradians. J. Exp. Mar. Biol. Ecol. 121, 113–136.
Fields, J.H.A., 1988. Fermentative adaptations to the lack of oxygen. Can. J. Zool. 66, 1036–1040.Fournier, P.A., Guderley, H., 1993. Muscle: the predominant glucose-producing organ in the leopard frog
during exercise. Am. J. Physiol. 264 (Regul. Integr. Comp. Physiol. 33), R239–R243.Gabbott, P.A., Bayne, B.L., 1973. Biochemical effects of temperature and nutritive stress on Mytilus edulis L.
J. Mar. Biol. Assoc. UK 53, 269–286.Gaitanaki, C., Papadopoulos, A., Beis, I., 1990. Time-course of covalent modification of pyruvate kinase
during anaerobiosis in the mantle muscle and the hepatopancreas of the limpet Patella caerulea (L). J.Comp. Physiol. B. 160, 529–535.
Guderley, H., Jean, C., Blouin, M., 1989. The effect of fatigue on the binding of glycolytic enzymes in theisolated gastrocnemius muscle of Rana pipiens. Biochim. Biophys. Acta 977, 87–90.
Guderley, H., Lavoie, B.A., Dubois, N., 1994. The influence of age, temperature and growth rate indetermining muscle metabolic capacities and tissue masses in the stickleback. Fish Physiol. Biochem. 13,419–431.
Guderley, H., Dutil, J.D., Pelletier, D., 1996. The physiological status of Atlantic cod, Gadus morhua, in thewild and the laboratory: estimates of growth rates under field conditions. Can. J. Fish Aquat. Sci. 53,550–557.
Holwerda, D.A., Kruitwagen, E.C.J., De Bont, A.M.T.H., 1981. Regulation of pyruvate kinase andphosphoenol-pyruvate carboxykinase activity during anaerobiosis in Mytilus edulis (L.). Mol. Physiol. 1,165–171.
Holwerda, D.A., Veenhof, P.R., Van Heugten, H.A.A., Zandee, D.I., 1983. Modification of mussel pyruvatekinase during anaerobiosis and after temperature acclimation. Mol. Physiol. 3, 225–234.
Huber, M., Guderley, H., 1993. The effect of thermal acclimation and exercise upon the binding of glycolyticenzymes in muscle of the goldfish (Carassius auratus). J. Exp. Biol. 175, 195–209.
Kreeger, D.A., Hawkins, A.J.S., Bayne, B.L., Lowe, D.M., 1995. Seasonal variation in the relative utilizationof dietary protein for energy and biosynthesis by the mussel Mytilus edulis. Mar. Ecol. Prog. Ser. 126,177–184.
Kreeger, D.A., Hawkins, A.J.S., Bayne, B.L., 1996. Use of dual-labeled microcapsules to discern thephysiological fates of assimilated carbohydrates, protein carbon, and protein nitrogen in suspension-feedingorganisms. Limnol. Oceanogr. 41, 208–215.
Lowery, M.S., Roberts, S.J., Somero, G.N., 1987. Effects of starvation on the activities and localization ofglycolytic enzymes in the white muscle of the barred sand bass Paralabrax nebulifer. Physiol. Zool. 60,538–549.
´Martınez, G., 1991. Seasonal variation in biochemical composition of three size classes of the Chilean scallop,Argopecten purpuratus Lamarck 1819. Veliger 34, 335–343.
´Martınez, G., Mettifogo, L., 1998. Mobilization of energy from adductor muscle for gametogenesis of thescallop, Argopecten purpuratus Lamarck. J. Shellfish Res. 17, 113–116.
´Martınez, G., Aguilera, C., Mettifogo, L., 2000. Interactive effects of diet and temperature on reproductiveconditioning of Argopecten purpuratus broodstock Lamarck. Aquaculture 183, 149–159.
Mathers, E.M., Houlihan, D.F., Cunningham, M.J., 1992. Nucleic acids concentrations and enzyme activitiesas correlates of growth rate in the saithe, Pollachius virens: growth rate estimates of open sea fish. Mar.Biol. 112, 363–369.

´G. Martınez et al. / J. Exp. Mar. Biol. Ecol. 247 (2000) 29 –49 49
Michaelidis, B., Gaitanaki, C., Beis, I., 1988. Modification of pyruvate kinase from the foot muscle of Patellacaerulea (L.) during anaerobiosis. J. Exp. Zool. 248, 264–271.
Mommsen, T.P., French, C.J., Hochachka, P.W., 1980. Sites and patterns of protein and amino acid utilizationduring the spawning migration of salmon. Can. J. Zool. 58, 1785–1799.
Munday, K.A., Giles, I.G., Poat, P.C., 1980. Review of the comparative biochemistry of pyruvate kinase.Comp. Biochem. Physiol. B 67, 403–411.
´Navarro, J.M., Leiva, G.E., Martınez, G., Aguilera, C., 2000. Interactive effects of diet and temperature on thescope for growth of the scallop Argopecten purpuratus during reproductive conditioning. J. Exp. Mar. Biol.Ecol. (in press).
´Pazos, A.J., Roman, G., Acosta, C.P., Abad, M., Sanchez, J.L., 1997. Seasonal changes in condition andbiochemical composition of the scallop Pecten maximus L. from suspended culture in the Ria de Arousa(Galicia, N.W. Spain) in relation to environmental conditions. J. Exp. Mar. Biol. Ecol. 211, 169–193.
Pelletier, D., Guderley, H., Dutil, J.D., 1993. Effect of growth rate, temperature and season on glycolyticenzyme activities in white muscle of cod Gadusmorhua. J. Exp. Zool. 265, 477–487.
Pelletier, D., Dutil, J.D., Blier, P.U., Guderley, H., 1994. Relation between growth rate and metabolicorganisation of white muscle, liver and digestive tract in cod, Gadus morhua. J. Comp. Physiol. B 164,179–190.
Plaxton, W.C., Storey, K.B., 1986. Glycolytic enzyme binding and metabolic control in anaerobiosis. J. Comp.Physiol. B156, 635–640.
Shafee, M.S., 1981. Seasonal changes in the biochemical composition and calorific content of the black scallop(Chlamys varia (L.)) from Lanceoc Bay of Brest. Oceanol. Acta 4, 331–341.
Sullivan, K.M., Somero, G.N., 1983. Size- and diet-related variations in enzymatic activity and tissuecomposition in the sablefish Anoplopoma fimbria. Biol. Bull. (Woods Hole) 164, 315–326.
Taylor, A.C., Venn, T.J., 1979. Seasonal variation in weight and biochemical composition of the tissues of thequeen scallop, Chlamys opercularis, from the Clyde Sea Area. J. Mar Biol. Assoc. UK 59, 605–621.
Whitwam, R.E., Storey, K.B., 1990. Organ specific analysis of the time course of covalent modification ofpyruvate kinase during anaerobiosis in a marine whelk. Physiol. Zool. 63, 222–234.
Related Documents











