Vesicular Glutamate Transporter Expression in Supraoptic Neurones Suggests a Glutamatergic Phenotype T. A. Ponzio, Y. Ni, V. Montana, V. Parpura and G. I. Hatton Department of Cell Biology and Neuroscience, University of California, Riverside, CA, USA. Key words: VGLUT, hypothalamus, dendritic release, oxytocin, vasopressin, astrocytes, meninges. OnlineOpen: This article is available free online at www.blackwell-synergy.com Abstract Magnocellular neuroendocrine cells of the supraoptic nucleus (SON) release the peptides oxytocin (OT) and vasopressin (VP) from their dendrites and terminals. In addition to peptide-containing large dense-core vesicles, axon terminals from these cells contain clear microvesicles that have been shown to contain glutamate. Using multilabelling confocal microscopy, we investigated the presence of vesicular glutamate transporters (VGLUTs) in astrocytes as well as VP and OT neurones of the SON. Simultaneous probing of the SON with antibodies against VGLUT isoforms 1–3, OT, VP and glial fibrillary acidic protein (GFAP) revealed the presence of VGLUT-2 in somata and dendrites of SON neurones. Immunoreactivity (-ir) for VGLUT-3 was also detected in both OT and VP neurones as well as in GFAP-ir astrocytes and other cells of the ventral glial lamina. Colocalisation of VGLUT-2 and VGLUT-3 in individual SON neurones was also examined and VGLUT-ir with both antibodies could be detected in both types of SON neurones. Although VGLUT-1-ir was strong lateral to the SON, only sparse labelling was apparent within the nucleus, and no colocalisation with either SON neurones or astrocytes was observed. The SON or the SON plus its surrounding perinuclear zone was probed using the reverse transcriptase-polymerase chain reaction and the presence of mRNA for all three VGLUT isoforms was detected. These results suggest that similar arrangements of transmitters exist in SON neuronal dendrites and their neurohypophysial terminals and that magnocellular neuroendocrine somata and dendrites may be capable of glutamatergic transmission. The supraoptic nucleus (SON) forms a major part of the magnocellular hypothalamic–neurohypophysial neuroendo- crine system. Individual SON neurones primarily synthesise either oxytocin (OT) or vasopressin (VP), which are peptide hormones with well-established roles in water balance, parturition and lactation. Stored in dense-core vesicles (DCVs), OT and VP are released from magnocellular neurone terminals in the pituitary neural lobe (NL) and from dendrites in the SON. Depending upon the stimulus, the peptide hormones are released from dendrites, terminals or both (1). Several neurotransmitters influence this release via activation of receptors found both in the SON and NL (2). In addition to containing DCVs, electron microscopy of the NL has shown that terminals from magnocellular neurones contain clear microvesicles (MVs). These MVs show glutam- ate immunoreactivity (-ir) and appear clustered for release (3). Additionally, the neural lobe has been found to be immuno- reactive for VGLUT-2 (4). The coexistence of glutamate-ir MVs with DCVs in the NL suggests that glutamatergic vesicles might also coexist with the well-documented DCVs in the magnocellular somata and dendrites. Exocytotic release of neuropeptides from SON somata and dendrites, first shown by Pow and Morris (5), accounts for earlier findings descri- bing the presence of OT and VP in the hypothalamus, and their functions in autocrine and retrograde signalling (6). However, anatomical studies addressing the possibility of somatodendritic glutamate release are lacking. Vesicular glutamate transporters (VGLUTs) account for all of the known glutamatergic transmission in the mamma- lian brain (7). Currently, three isoforms of VGLUTs are known to exist. These have been found in both neurones and astrocytes, with VGLUT-3 located in serotonergic, choliner- gic, dopaminergic and even GABAergic neurones (8, 9). A number of studies (4, 10–12) have demonstrated the presence of VGLUT-2-ir and/or mRNA in the magnocellular neuro- endocrine system. This is not surprising because the SON is the destination of myriad glutamate afferents (13). Further- more, the existence of VGLUTs mRNA has long been known in various types of synaptic terminals and protoplasmic astrocytes (which are especially plentiful in the ventral SON) Correspondence to: Dr T. A. Ponzio, NINDS/NIH, Bldg 49/5C68, Bethesda, MD 20892, USA (e-mail: [email protected]). Journal of Neuroendocrinology, 2006, Vol. 18, 253–265 doi:10.1111/j.1365-2826.2006.01410.x ȑ 2006 The Authors. Journal compilation ȑ 2006 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vesicular Glutamate Transporter Expression in SupraopticNeurones Suggests a Glutamatergic Phenotype
T. A. Ponzio, Y. Ni, V. Montana, V. Parpura and G. I. HattonDepartment of Cell Biology and Neuroscience, University of California, Riverside, CA, USA.
Key words: VGLUT, hypothalamus, dendritic release, oxytocin, vasopressin, astrocytes, meninges.
OnlineOpen: This article is available free online at www.blackwell-synergy.com
Abstract
Magnocellular neuroendocrine cells of the supraoptic nucleus (SON) release the peptides oxytocin (OT) and
vasopressin (VP) from their dendrites and terminals. In addition to peptide-containing large dense-core
vesicles, axon terminals from these cells contain clear microvesicles that have been shown to contain
glutamate. Using multilabelling confocal microscopy, we investigated the presence of vesicular glutamate
transporters (VGLUTs) in astrocytes as well as VP and OT neurones of the SON. Simultaneous probing of the
SON with antibodies against VGLUT isoforms 1–3, OT, VP and glial fibrillary acidic protein (GFAP) revealed
the presence of VGLUT-2 in somata and dendrites of SON neurones. Immunoreactivity (-ir) for VGLUT-3 was
also detected in both OT and VP neurones as well as in GFAP-ir astrocytes and other cells of the ventral glial
lamina. Colocalisation of VGLUT-2 and VGLUT-3 in individual SON neurones was also examined and
VGLUT-ir with both antibodies could be detected in both types of SON neurones. Although VGLUT-1-ir was
strong lateral to the SON, only sparse labelling was apparent within the nucleus, and no colocalisation with
either SON neurones or astrocytes was observed. The SON or the SON plus its surrounding perinuclear zone
was probed using the reverse transcriptase-polymerase chain reaction and the presence of mRNA for all
three VGLUT isoforms was detected. These results suggest that similar arrangements of transmitters exist in
SON neuronal dendrites and their neurohypophysial terminals and that magnocellular neuroendocrine
somata and dendrites may be capable of glutamatergic transmission.
The supraoptic nucleus (SON) forms a major part of themagnocellular hypothalamic–neurohypophysial neuroendo-crine system. Individual SON neurones primarily synthesiseeither oxytocin (OT) or vasopressin (VP), which are peptidehormones with well-established roles in water balance,parturition and lactation. Stored in dense-core vesicles(DCVs), OT and VP are released from magnocellular neuroneterminals in the pituitary neural lobe (NL) and from dendritesin the SON. Depending upon the stimulus, the peptidehormones are released from dendrites, terminals or both (1).Several neurotransmitters influence this release via activationof receptors found both in the SON and NL (2).In addition to containing DCVs, electron microscopy of the
NL has shown that terminals from magnocellular neuronescontain clear microvesicles (MVs). These MVs show glutam-ate immunoreactivity (-ir) and appear clustered for release (3).Additionally, the neural lobe has been found to be immuno-reactive for VGLUT-2 (4). The coexistence of glutamate-irMVs with DCVs in the NL suggests that glutamatergicvesicles might also coexist with the well-documented DCVs in
the magnocellular somata and dendrites. Exocytotic release ofneuropeptides from SON somata and dendrites, first shownby Pow and Morris (5), accounts for earlier findings descri-bing the presence of OT and VP in the hypothalamus, andtheir functions in autocrine and retrograde signalling (6).However, anatomical studies addressing the possibility ofsomatodendritic glutamate release are lacking.
Vesicular glutamate transporters (VGLUTs) account forall of the known glutamatergic transmission in the mamma-lian brain (7). Currently, three isoforms of VGLUTs areknown to exist. These have been found in both neurones andastrocytes, with VGLUT-3 located in serotonergic, choliner-gic, dopaminergic and even GABAergic neurones (8, 9). Anumber of studies (4, 10–12) have demonstrated the presenceof VGLUT-2-ir and/or mRNA in the magnocellular neuro-endocrine system. This is not surprising because the SON isthe destination of myriad glutamate afferents (13). Further-more, the existence of VGLUTs mRNA has long been knownin various types of synaptic terminals and protoplasmicastrocytes (which are especially plentiful in the ventral SON)
Correspondence to: Dr T. A. Ponzio, NINDS/NIH, Bldg 49/5C68, Bethesda, MD 20892, USA (e-mail: [email protected]).
Journal of Neuroendocrinology, 2006, Vol. 18, 253–265 doi:10.1111/j.1365-2826.2006.01410.x
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd

contain VGLUT-positive MVs (14). Therefore, in the absenceof positive magnocellular-neurone and glial identifiers, theprecise location of the VGLUT-2-ir and mRNA remainsunclear. This is the first study to specifically address thepresence of VGLUT-3 immunoreactivity in the magnocellularsystem and to build upon earlier observations by investigatingthe presence of different VGLUT isoforms in clearly identi-fied SON neurones and astrocytes.
Earlier ultrastructural analysis of postembedding glutam-ate immunocytochemistry revealed microvesicles in the NLstain for glutamate (3). Additionally, both magnocellularsomata and dendrites are immunoreactive with glutamateantibodies at the light and electron microscopic level (15).However, the amino acid glutamate is present in all cells,functioning in metabolism and incorporated into proteins.Whether or not it is also used as a regulated signallingmolecule depends upon whether the cell contains machineryfor controlled release of the transmitter. To explore thepossibility of glutamate release from neurones and astrocytesof the SON, we studied the presence of the three knownVGLUTs and their mRNA, which function in the storage andconcentration of glutamate into synaptic vesicles.
Materials and methods
Immunocytochemistry
Adult male rats (aged 50–80 days) were deeply anaesthetised with sodium
pentobarbital (100 mg/kg) and transcardially perfused with 4% paraformal-
dehyde in 0.1 M phosphate-buffered saline (PBS). Coronal sections (50–60 lmthick) were cut on a vibratome, treated with 0.3% Triton X-100 in PBS for
30 min and subsequently immunoprocessed (for all antibody identification,
see Table 1). Brain sections were incubated with antibodies for either 3 h at
room temperature or overnight at 4 �C. All secondary antibodies were made
in donkey and were conjugated to AMCA, FITC (Jackson Laboratories, Bar
Harbor, ME, USA) or the Alexa Flour� 350, 488, 546, 555, or 647 (Molecular
Probes, Eugene, OR, USA). A 5-min histochemical treatment with Hoechst
(Sigma, St Louis, MO, USA; 1 : 1000, 1 mg/ml) was also used in several
experiments to stain nuclei. Hoechst, which produces a blue fluorescence in
response to ultraviolet (UV) excitation, is depicted in grey in the confocal
micrographs.
Confocal microscopy
An upright Leica SP2 UV-equipped laser-scanning confocal microscope
(Leica, Heidelberg, Germany) was used in sequential scanning mode with
nonoverlapping photomultiplier tube settings for acquiring all images. All
data were collected with a pinhole set to 1 airy unit corresponding to Z-optical
sections <300 nm. Z-series were collected with a depth difference of
<0.5 lm between successive Z sections.
Cell cultures
We prepared enriched astrocytic cultures using a modification (16) of the
originally described shaking procedure (17). Briefly, visual cortices isolated
from 0- to 2-day-old Sprague-Dawley rats were enzymatically treated (papain,
20 IU/ml; 1 h at 36.8 �C). After mechanical dispersion, cells were initially
plated into tissue culture flasks (25 cm2) and maintained at 36.8 �C in a
humidified 5% CO2 95% air atmosphere in a complete culture medium that
consisted of a-MEM (without phenol red; GibcoTM
, Invitrogen Corp.,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (HyClone,
Logan, UT, USA), L-glutamine (2 mM; Invitrogen Corp.), D-glucose (20 mM;
Sigma-Aldrich, St Louis, MO, USA), sodium pyruvate (1 mM; Invitrogen
Corp.), penicillin (100 IU/ml), streptomycin (100 lg/ml), and sodium bicar-
bonate (14 mM; Invitrogen Corp.) (pH ¼ 7.4). After 6–24 days in culture, the
cells were shaken twice (260 r.p.m. at 36.8 �C), first for 1.5–2 h, and then,
after exchange of complete medium, again for 18–20 h. At that time, the
remaining attached cells were detached from flasks using a cell scraper and
used in preparation of subcellular fractions for Western blotting experiments.
The purity (> 99%) of astrocytic culture was confirmed by anti-glial fibrillary
acidic protein (GFAP) antibody and indirect immunocytochemistry.
Preparation of subcellular fractions and Western blotting
We obtained non-nuclear membranes and vesicular extracts from purified
astrocytes and different brain tissues (occipital lobe, hypothalamus and
posterior pituitary gland) as previously described (18). Preparations were
subjected to 15% SDS-PAGE, followed by transfer to nitrocellulose mem-
branes that were probed with antibodies against VGLUT-1 (rabbit polyclonal,
1 : 1000; provided by Dr R. H. Edwards, University of California, San
Francisco, CA, USA) (19), VGLUT-2 (rabbit polyclonal; 1 : 1000; Synaptic
Systems, Goettingen, Germany, cat no. 135102) or VGLUT-3 (guinea pig
polyclonal, 1 : 5000; Chemicon International, Temecula, CA, USA, cat No.
AB5421). Immunoreactivity of bands was detected using enhanced chemilu-
minescence (ECL; Amersham Biosciences Corp., Piscataway, NJ, USA). We
previously characterised the specificity of these VGLUT antibodies using their
respective antigens in adsorption controls (16).
Reverse transcriptase-polymersase chain reaction (RT-PCR)
Five adult (62 day-old) male rats were used to probe both the SON and SON
plus perinuclear zone for mRNA corresponding to all three VGLUT isoforms.
Total RNA was extracted using TRIzol� Reagent (Invitrogen Corp.) and
protocols provided by the manufacturer as previously described (16). Total
RNA was used for reverse transcription using Oligo(dT)12)18 and super-
scriptTM III reverse transcriptase (Invitrogen Corp.). Two pairs of primers
(each pair 35 cycles) for each of VGLUTs 1, 2 and 3 (GeneBank accession
numbers NM_053859, NM_053427 and NM_153725, respectively) were used
to amplify cDNA using PCR Core System I kit (Promega, Madison, WI,
USA). For VGLUT-1, in the first round we used, forward 5¢-CCG GCA
GGA GGA GTT TCG AGG G-3¢ and reverse 5¢-AGG GAT CAA CAT
ATT TAG GGT GGA GGT AGC-3¢ primers (478 bp product). Amplified
Table 1. Antibody Dilutions and Sources.
Antibody Host Dilution Company or PI
Vesicular glutamate transporter type 1 Rabbit (r) 1 : 500 Synaptic SystemsVesicular glutamate transporter type 1 Rabbit 1 : 1000 R.H. Edwards, UCSFVesicular glutamate transporter type 2 Rabbit 1 : 500 Synaptic SystemsVesicular glutamate transporter type 2 Guinea Pig (gp) 1 : 2500 ChemiconVesicular glutamate transporter type 3 Guinea Pig 1 : 2000 ChemiconOxytocin-Neurophysin Mouse (m) 1 : 400 H. Gainer, NIH, PS38Vasopressin-Neurophysin Mouse 1 : 400 H. Gainer, NIH, PS41Neurophysin II (marker for VP neurones) Rabbit 1 : 500 SigmaNeurophysin II Goat (g) 1 : 20 Santa Cruz BiotechGlial fibrillary acidic protein (marker for astrocytes) Mouse 1 : 500 SigmaGlial fibrillary acidic protein Goat 1 : 20 Santa Cruz BiotechNeurofilament (marker for neurones) Mouse 1 : 100 Calbiochem
254 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

DNA was used as the template for the second round of PCR using nested
primers, forward 5¢-TAC TGG AGA AGC GGC AGG AAG G-3¢ and
reverse 5¢-CCA GAA AAA GGA GCC ATG TAT GAG G-3¢ (311 bp
product). For VGLUT-2 we used in the first round, forward 5¢-AGC AAG
GTT GGC ATG TTG TCT G-3¢ and reverse 5¢-CGG TCC TTA TAG GAG
TAC GCG T-3¢ primers (698 bp product), while in the second round the
nested primers were forward 5¢-TGG TGC AAT GAC GAA GAA CAA G-3¢and reverse 5¢-TCC TTT TTC TCC CAG CCG TT-3¢ (294 bp product). For
VGLUT-3, in the first round the primers were forward 5¢-AGG AGT GAA
GAA TGC CGT GGG AGA T-3¢ and reverse 5¢-ACC CTC CAC CAG ACC
CTG CAA A-3¢ (535 bp product), while the second round nested primers were
forward 5¢-GAT GGG ACC AAC GAG GAG GGA GAT-3¢ and reverse
5¢-TGA AAT GAA GCC ACC GGG AAT TTG T-3¢ (322 bp product).
Results
VGLUT immunocytochemistry positive and negative controlexperiments
Experiments testing the specificity of the VGLUT antibodiesused in this study have been published previously (16).However, further experiments were performed to test theirspecificity in the tissue under study. Expression of VGLUT-2in the adult cerebellum has been described (20) as occurring inthe large mossy fibre terminals in the granule cell layer andalong the thin climbing fibres in the molecular layer. Immu-noreactivity for VGLUT-1 occurs in the molecular layer,labelling parallel fibre terminals (Fig. 1A–D). In situ hybridi-sation and protein expression experiments have shownVGLUT-3 to be present in the hypothalamus and that itsprotein expression is particularly high in the olfactorytubercles (8, 21). Western blot experiments have bands withmolecular weights corresponding to the specific VGLUTbeing immunoprobed (Fig. 1E). In addition to the nucleus ofthe lateral olfactory tract (nLOT), strong VGLUT-3-ir can beseen in the SON (Fig. 1F). Note that the punctate labelling inthe nLOT is different than the cellular labelling observed in theSON and is seen using the same antibody in the same section.Negative control experiments, performed in the absence of
primary antibody exposure, were carried out in parallel withall multilabelling immunocytochemical experiments. Furthercontrol experiments, similar to those previously described(16), in which the antibodies were preincubated with theirrecognised peptide sequences were also performed. Both typesof negative control experiments resulted in the absence ofstained tissue.
VGLUT-3 in the SON
Cellular labelling for VGLUT-3 appeared throughout theSON (Figs 1F and 2). Experiments using double-labelling forVGLUT-3 and OT or VP resulted in apparent colocalisationof peptide- and VGLUT-3-ir. Dorsal to the SON, scatteredcells of the anteriolateral hypothalamic area also displayedVGLUT-3-ir (Fig. 2A). Additionally, lighter VGLUT-3 label-ling was apparent in the ventral glial lamina and in cells of themeninges. SON neuronal VGLUT-3-ir was clustered in theperikarya, while labelling was absent in the nuclei of bothSON neurones and astrocytes, as determined by Hoechststaining.To determine if the VGLUT-3-ir was present intracellular-
ly, Z-series through SON sections immunoprobed for neuro-
physin II, GFAP, OT and VGLUT-3 were taken.Extranuclear puncta representing VGLUT-3 are surroundedby both the VP-specific peptide neurophysin II- (Fig. 2C) andGFAP-ir (Fig. 2D). Shown in all three dimensions in Fig. 2(D,insets 1–3) VGLUT-3-ir is localised within the boundaries ofGFAP-ir, suggesting SON astrocytes express this transporter.OT-positive cells were also seen to contain VGLUT-3-ir(Fig. 2E–G). Interestingly, many of the magnocellular neu-rones that are most immunoreactive for VGLUT-3 displayed2 or 3 nucleoli and regions of ribosome synthesis (Fig. 2C,insets).
The VGLUT-3-ir did not appear to extend into thedendrites or along axons. Rather, it was limited to theperikarya. VGLUT-3-ir was also seen in astrocytic somatabut not processes. Strong labelling was also apparent in theendothelial and meningeal cells at the base of the brain.There, the labelling appears more distributed and notnecessarily restricted to perinuclear areas.
VGLUT-2 in the SON
Two polyclonal antibodies to VGLUT-2 were used to localisethe protein. Punctate labelling by both guinea pig and rabbitantibodies can be seen throughout the SON and was alsoparticularly strong dorsal to the nucleus. Multilabellingexperiments, including probing for the VP cell marker neuro-physin II, revealed colocalisation of VGLUT-2-ir withmany ofthe neurophysin II-positive cell processes (Figs 3 and 4).Examples of neurophysin II-positive dendrites and somatacontaining intracellular VGLUT-2-ir are shown in Fig. 3(A–E).Because these cells are contacted by many glutamatergicterminals, it was necessary to perform Z-series acquisitions tobe sure theVGLUT immunoreactivitywas occurringwithin theboundaries of specific magnocellular neurones as judged bytheir peptide immunoreactivity. Additionally, strong punctatelabelling, appearing just outside much of the neurophysin II-ir,morphologically reveals putative glutamatergic synapses ontothese cells (Fig. 3A,C,E).
Similar multilabelling experiments were performed toinvestigate VGLUT-2 and OT colocalisation. OT-positivecells also appeared to receive glutamatergic input fromVGLUT-2 immunoreactive terminals (Fig. 5D, arrows).Intracellular VGLUT-2-ir was apparent in several OT-posit-ive dendrites (Fig. 5). Unlike VGLUT-3-ir, colocalisation ofVGLUT-2 with GFAP within the VGL was not evident.Furthermore, meningeal and endothelial cells did not showdetectable levels of VGLUT-2-ir with either VGLUT-2antibody.
OT and VP neurones of the SON can express VGLUT-2or VGLUT-3. However, the question remained as towhether or not an individual magnocellular neurone couldexpress both VGLUTs. Multilabelling for the two peptidehormones along with the two VGLUTs revealed VGLUT-2-and VGLUT-3-ir in both types of magnocellular neurones(Fig. 6). Although strong VGLUT-3-ir appeared in virtuallyall labelled neurones (Figs 2, 6 and 7), this was not the casefor VGLUT-2. Some magnocellular neurones were clearlymore immunoreactive than others for VGLUT-2 (Fig. 6D,inset), suggesting that VGLUT expression may be a regu-lated phenomenon.
Vesicular glutamate transporter expression in supraoptic neurones 255
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265
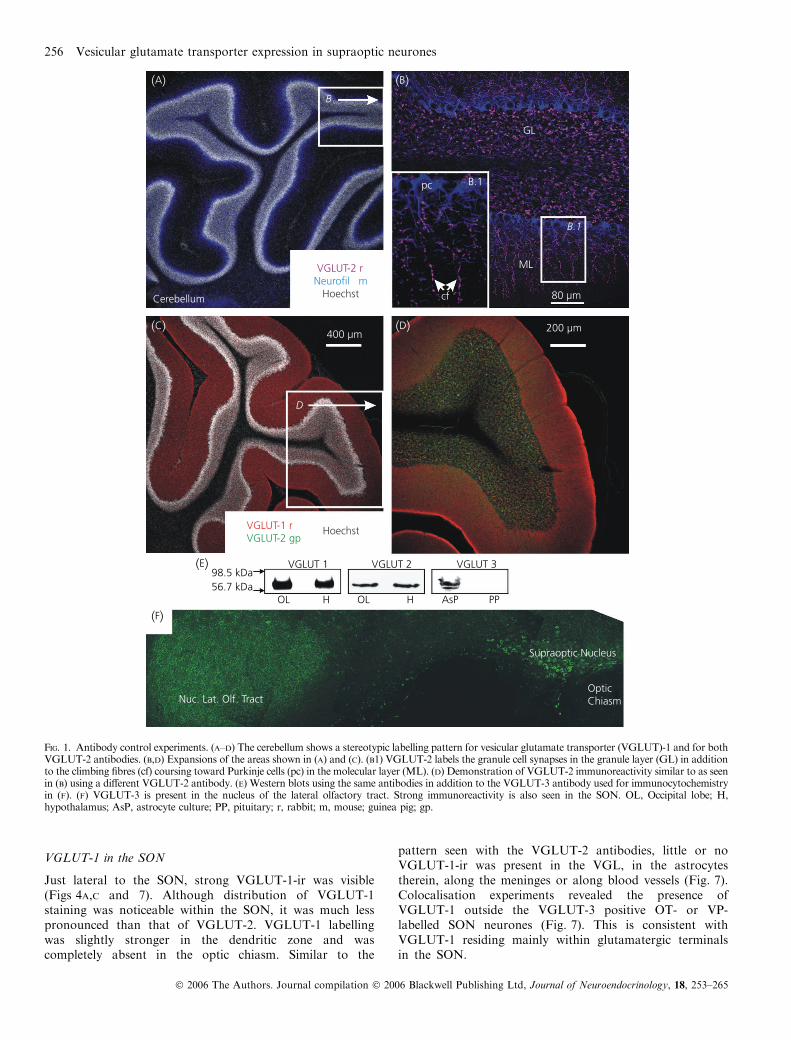
VGLUT-1 in the SON
Just lateral to the SON, strong VGLUT-1-ir was visible(Figs 4A,C and 7). Although distribution of VGLUT-1staining was noticeable within the SON, it was much lesspronounced than that of VGLUT-2. VGLUT-1 labellingwas slightly stronger in the dendritic zone and wascompletely absent in the optic chiasm. Similar to the
pattern seen with the VGLUT-2 antibodies, little or noVGLUT-1-ir was present in the VGL, in the astrocytestherein, along the meninges or along blood vessels (Fig. 7).Colocalisation experiments revealed the presence ofVGLUT-1 outside the VGLUT-3 positive OT- or VP-labelled SON neurones (Fig. 7). This is consistent withVGLUT-1 residing mainly within glutamatergic terminalsin the SON.
(A) (B)
GL
B.1
ML
cf 80 µm
200 µm400 µm
Cerebellum
(C)
(E) VGLUT 198.5 kDa56.7 kDa
OL
(F)
Nuc. Lat. Olf. Tract
H OL H AsP PP
Supraoptic Nucleus
OpticChiasm
VGLUT 2 VGLUT 3
(D)
VGLUT-2 rNeurofil m
Hoechst
VGLUT-1 rVGLUT-2 gp
D
pc
B
B.1
Hoechst
Fig. 1. Antibody control experiments. (A–D) The cerebellum shows a stereotypic labelling pattern for vesicular glutamate transporter (VGLUT)-1 and for bothVGLUT-2 antibodies. (B,D) Expansions of the areas shown in (A) and (C). (B1) VGLUT-2 labels the granule cell synapses in the granule layer (GL) in additionto the climbing fibres (cf) coursing toward Purkinje cells (pc) in the molecular layer (ML). (D) Demonstration of VGLUT-2 immunoreactivity similar to as seenin (B) using a different VGLUT-2 antibody. (E) Western blots using the same antibodies in addition to the VGLUT-3 antibody used for immunocytochemistryin (F). (F) VGLUT-3 is present in the nucleus of the lateral olfactory tract. Strong immunoreactivity is also seen in the SON. OL, Occipital lobe; H,hypothalamus; AsP, astrocyte culture; PP, pituitary; r, rabbit; m, mouse; guinea pig; gp.
256 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

(A) (B)
OC
B,C,D
80 µm
40 µm
BV
D.1
D.2
D.3
X-Y
Y-Z
X-Z
OC
(G)(F)
(D)(C)
C.1 C.2
C.2
C.1
expanded above
40 µm
(E)
OC
150 µm
OC
VGLUT-3
OT
GFAPVGLUT-3
NP-II rHoechst
mgp
gpm
Fig. 2. Vesicular glutamate transporter (VGLUT)-3 expression by neurones, astrocytes, and their underlying meningeal cells. (A) Neurophysin positive cells ofthe supraoptic nucleus (SON) are immunoreactive for VGLUT-3. Scattered cells of the lateral hypothalamic area also show VGLUT-3-immunoreactivity (ir)(arrows). (B) GFAP-ir of the VGL separates the SON from VGLUT-3 immunoreactive meninges and endothelial cells at the base of the brain (arrows). (C)VGLUT-3 is located in the interior of SON neurones immunoprobed with neurophysin II. Several SON neurones with strong VGLUT-3 labelling also havemultiple nucleoli (* and expanded in insets C1 and C2, arrows indicate nucleoli). The scale bar in (C) also applies also to (D). (D) Astrocytes of the VGL expressVGLUT-3 (open arrows). (D1–D3) Expanded X–Y, Y–Z and X–Z projections at the crosshair point inside the red square in (D) show that astrocytic VGLUT-3-ir is completely surrounded by GFAP-ir. Z-thickness is 20 lm in (C) and (D). (E–G) Colocalisation of VGLUT-3 (E,G) with oxytocin (OT) (F,G). Open arrowsin (E) and (G) identify several oxytocin-positive cells colocalised with VGLUT-3 immunoreactive puncta. The scale bar in (E) also applies to (F) and (G). OC,Optic chiasm; BV, blood vessel.
Vesicular glutamate transporter expression in supraoptic neurones 257
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

RT-PCR of VGLUTs in the SON
To corroborate the immunocytochemical experiments, weperformed RT-PCR using mRNA isolated from SON andSON plus the perinuclear zone of five adult rats, the latterserving as a positive control for all three VGLUTs (Fig. 8,top). Whereas PCR products for VGLUTs-2 and -3 werefound in all five of the isolated SONs, PCR products forVGLUT-1 were found in only three of five cases (Fig. 8,bottom). Consistent with the immunocytochemical results,PCR products representing the mRNA for all three VGLUTisoforms were found in SON plus perinuclear zone from allfive rats. These results agree with the idea that VGLUTs 2and 3 are heavily expressed in the SON, whereas VGLUT-1is only sparsely expressed. Additionally, these results arealso consistent with the known presence of mRNAs interminals.
Discussion
For several years, glutamate has been known to be the mainexcitatory transmitter involved in neuroendocrine regulation(22, 23). In addition to general excitation, glutamate-impli-cated phenomena occurring in the magnocellular neuroendo-crine system include osmoresponsiveness (24, 25) andbursting rhythmogenesis (26). Here, we present evidence thatan additional source for that glutamate may be somatoden-dritically released glutamate from magnocellular neuronesthemselves.
A precendent already exists for intranuclear release ofsubstances from magnocellular neurones. The release of OTand VP from SON neuronal somata and dendrites underdifferent conditions can influence presynaptic activity (1, 27).The presence of glutamate-containing MVs in magnocellularneuronal terminals in the NL has also been described (3). The
(A)
(B)
(C)
(D) (E)
Expanded
OC80 µm
40 µm
D
E
A
X-Z projection
B
C
Y-Z
proj
ectio
n
VGLUT-2 rNP-II gHoechst
Fig. 3. Vasopressinergic magnocellular neurone expression of vesicular glutamate transporter (VGLUT)-2-ir. Upper-left inset: area of supraopticnucleus expanded in the figure. Further expansions of outlined areas (A–E) are shown in panels on right. The X–Z and Y–Z projections of neurophysinII-immunoreactivity (ir) completely surrounding that for VGLUT-2 can be seen at the points of the crosshairs (Z-thickness is 12 lm). Expansions of internallylocalised VGLUT-2-ir found at one such point can be seen on the right (circled in A–C). Putative afferent synapses represented by large VGLUT-2 puncta areadjacent to the peptide-labelled cells (arrows in A,C,E). OC, Optic chiasm.
258 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

same group found glutamate-ir both in afferent terminals in theSON and, more relevant here, in magnocellular somata anddendrites (15, 28). This glutamate was postulated to servemainly a metabolic function and the possibility for somato-dendritic exocytosis of glutamate largely remained uninvesti-gated. In the present study, we offer strong evidence that thesomatodendritic compartment of SON neurones contains theprinciple machinery that defines a glutamatergic phenotype.This is consistent with the reported presence of VGLUT-2-ir inthe NL (4). Although earlier investigations of VGLUTs havedescribed a complementary expression pattern of the proteins,we present evidence for both VGLUT-3 and VGLUT-2expression by OT- or VP-positive SON neurones. The possi-bility of multiple VGLUTs in a given neurone presents some
obvious and interesting questions regarding the specific rolesfor the different VGLUTs found within the same neurone.
Potential functions of VGLUT-3 in the SON
Cloned and described by three independent groups in 2002,VGLUT-3 was shown to exist in serotonergic, cholinergic,andGABAergic neurones (8, 9, 21). Unlike VGLUTs-1 and -2,VGLUT-3-ir has been found in somata and dendrites inaddition to synaptic terminals. Here, we report the presenceof VGLUT-3 in the somata of magnocellular neurones,astrocytes, along the endothelium and along meningeal cells.One commonality shared by these various cell types in thisparticular region is a high level of metabolic activity.
(A)
(B)
(C)
80 µm
VGLUT-2 gpVGLUT-1 r
OT mNP-II g
Fig. 4. (A–C) Colocalisation of vesicular glutamate transporter (VGLUT)-2- and peptide hormone-immunoreactivity can be seen with a different VGLUT-2antibody (arrows). Colocalisation occurs in somata and dendrites of magnocellular neurones. OT, Oxytocin; NP-II, neurophysin II.
Vesicular glutamate transporter expression in supraoptic neurones 259
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265
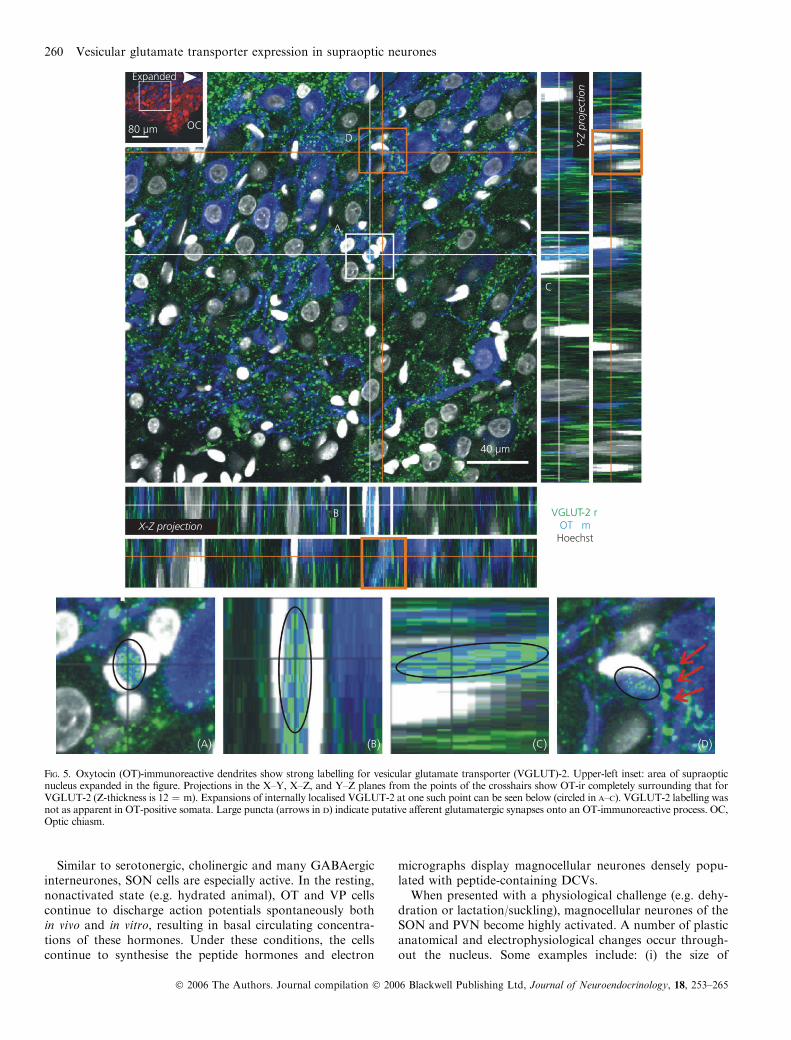
Similar to serotonergic, cholinergic and many GABAergicinterneurones, SON cells are especially active. In the resting,nonactivated state (e.g. hydrated animal), OT and VP cellscontinue to discharge action potentials spontaneously bothin vivo and in vitro, resulting in basal circulating concentra-tions of these hormones. Under these conditions, the cellscontinue to synthesise the peptide hormones and electron
micrographs display magnocellular neurones densely popu-lated with peptide-containing DCVs.
When presented with a physiological challenge (e.g. dehy-dration or lactation/suckling), magnocellular neurones of theSON and PVN become highly activated. A number of plasticanatomical and electrophysiological changes occur through-out the nucleus. Some examples include: (i) the size of
Expanded
OCD
A
B
(A) (B) (C) (D)
C
Y-Z
proj
ectio
n
X-Z projection
80 µm
40 µm
VGLUT-2 rOT mHoechst
Fig. 5. Oxytocin (OT)-immunoreactive dendrites show strong labelling for vesicular glutamate transporter (VGLUT)-2. Upper-left inset: area of supraopticnucleus expanded in the figure. Projections in the X–Y, X–Z, and Y–Z planes from the points of the crosshairs show OT-ir completely surrounding that forVGLUT-2 (Z-thickness is 12 ¼ m). Expansions of internally localised VGLUT-2 at one such point can be seen below (circled in A–C). VGLUT-2 labelling wasnot as apparent in OT-positive somata. Large puncta (arrows in D) indicate putative afferent glutamatergic synapses onto an OT-immunoreactive process. OC,Optic chiasm.
260 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

magnocellular nucleoli increasing significantly (29); (ii) theOT cells bursting synchronously and the VP cells exhibiting astereotypic phasic firing pattern (30); and (iii) astrocyticprocesses in the neural lobe and SON retracting, allowing forincreases in neurone-neurone appositions and synapse for-mation (31). With these changes, elevated levels of circulatinghormones ensue.
The blood supply to the SON is also distinctive. Along withthe PVN, the SON is enrichedwith the highest capillary densityin the brain, at least twice as high as observed in other brainnuclei (32). Doubtless, the cells of this especially active andplastic nucleus require extraordinary levels of factors garneredfrom an enriched blood supply and additionally need a readilyaccessible pool of glutamate to maintain their high metabolic
(A)
(B)
(C)
(D)
14 µm
VGLUT-3 gpVGLUT-2 rOT mNP II g
Fig. 6. Colocalisation of vesicular glutamate transporter (VGLUT)-2 and 3 in magnocellular supraoptic nucleus (SON) neurones. (A) VGLUT-2- and3-immunoreactivity appears in the perikarya of SON cells. Dashed lines in A and B approximate cell boundaries. (B) Oxytocin (OT)- and neurophysin II (NP-II)-ir reveals a heterogeneous population of peptidergic neurones in this section. (C) An OT-positive cell is positive for both VGLUT-2 and -3 (arrowhead). Inthe same field, a VGLUT-3-positive, although only faintly VGLUT-2-positive cell is indicated (asterisk). A few large puncta representing the clustering ofnumerous VGLUT-2 positive vesicles forming a putative glutamatergic synapses are indicated (arrows in C and D). (D) Neurophysin-II-positive cells wereimmunoreactive to both VGLUTs (arrowhead and inset).
Vesicular glutamate transporter expression in supraoptic neurones 261
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

and protein-synthetic activity. In addition to VGLUTinvolvement in exocytosis, the presence of VGLUT-3-ir in thiscase possibly suggests a metabolic function similar to that
proposed byMeeker et al. (15). Consistent with this hypothesisis the strong VGLUT-3-ir observed in neurones with multiplenucleoli (Fig. 2C).
(A)
(B)
(C)
(D)
(E)
A.1 D.1
C.1
SON
A.1 C.1 D.1
100 µm
VGLUT-1 rVGLUT-3 gpOT+VP m
Hoechst
Fig. 7. Vesicular glutamate transporter (VGLUT)-1 expression in the supraoptic nucleus (SON). (A–C) Though faint labelling is seen inside the SON proper,strong VGLUT-1-immunoreactivity consistent with a terminal distribution occurs in the cell sparse region lateral to the nucleus (arrows in A and C).(D) Colocalisation of oxytocin and vasopressin with VGLUT-3 but not VGLUT-1 is seen. (A1,B1,C1) Expansions from boxes in the respective panels. (E) is thesame as (D1) with the addition of the nuclear stain Hoechst.
262 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

VGLUT-2 expression and a glutamatergic phenotype
VGLUT-2 expression is strongly linked to glutamatergictransmission. It is known to exist at many well-characterisedglutamatergic synapses and is restricted to sites of knownrelease. VGLUT-2-ir in OT and VP neurones is in agreementwith a previous study demonstrating VGLUT-2 mRNA inthe magnocellular hypothalamic nuclei (10). However,mRNA expression is not an infallible indication of proteinexpression and such studies reveal no information regardingthe specific location of the molecules of interest. Here, we addto that work by demonstrating that VGLUT-2-ir is not onlyexpressed in local glutamatergic neurones and terminalscontacting SON neurones, but also in OT/VP-positive SONneurones themselves. Colocalisation of the transporter withinthe magnocellular neurones is now established, suggestingthat these cells may release both glutamate and either OT orVP from their somata and dendrites.The suggestion of dendritic glutamate release from mag-
nocellular neurones presents an interesting puzzle concerningits function. The clustering of microvesicles in SON magno-cellular dendrites or somata has not been described, suggest-ing that glutamate is probably not released at specific sites inhigh concentration (i.e. synapses), but is instead released froma more general area (i.e. a somatic or dendritic zone). Becausethese cells and many of their afferent terminals expressglutamate receptors, the released glutamate may function inautocrine, paracrine or retrograde transmission and, in somecases, perhaps contribute to a feed-forward activation system.A similar autocrine feed-forward system exists for OT actingon OT neurones during the episodic firing of milk ejectionbursts. There, somatodendritic released OT results inenhanced local concentrations of OT which lead to anincreased frequency and amplitude of subsequent milk
ejection bursts (33–35). Furthermore, autocrine signalling ofVP-cell derived dynorphin has been shown to affect VPneurone bursting (36, 37).
Whereas activation of pre- or post-synaptic ionotropicglutamate receptors would result in direct membrane depo-larisation and/or release of presynaptic transmitter, theactivation of pre- and post synaptic metabotropic glutamatereceptors (mGluRs) results in more subtle forms of modula-tion. Evidence exists for the presence of mGluRs on magno-cellular neurones, and on glutamatergic and GABAergicterminals in the SON (38, 39). Activation of terminallylocated group III mGluRs causes a reduction in the frequencyof spontaneous glutamatergic and GABAergic release. Alter-natively, activation of postsynaptic group I mGluRs inducesexcitation through activation of inward currents carried byK+ or Ca2+ (38, 39). Therefore, the qualitative result ofsomatodendritic released glutamate will depend upon whichreceptors are subsequently activated.
Consistent with the immunocytochemical evidence pre-sented in a recent study, VGLUT-2 is also probablyinvolved in afferent glutamatergic transmission in the SON(40). Larger VGLUT-2 immunoreactive puncta are seenalong dendrites and somata of SON neurones, which mayrepresent glutamatergic terminals forming synapses withinthe nucleus.
VGLUT-1 immunoreactive input to the SON
Although VGLUT-1-ir in the SON was not as intense as thatseen for either VGLUT-2 or VGLUT-3, strong labelling wasevident lateral to the nucleus. This area is enriched with mitralcell axons and terminals that form the glutamatergic lateralolfactory tract (41–43). Mitral cell and glomerular layers ofthe olfactory bulb are immunopositive for VGLUT-1 (40).
SON + perinuclear zone
VGLUT-1
OCSON
(S + P)
(S + P) SONVGLUT-2
(S + P) SONVGLUT-3
(S + P) SON
5/5 3/5 5/5 5/5 5/5 5/5 Frequency
M
400 bp
300 bp
200 bp
80 µm
(a)
Fig. 8. Detection of vesicular glutamate transporter (VGLUT) mRNA in the supraoptic nucleus (SON). Top: Micrograph showing from where the tissue wascollected, either the SON only (n ¼ 5) or the SON + perinuclear zone (S + P, n ¼ 5). OC, Optic chiasm. Bottom: Reverse transcriptase-polymerase chainreaction analysis from SON and S + P tissue reveals the presence of mRNA for all three VGLUT isoforms: VGLUT-1(311 bp), VGLUT-2(294 bp) andVGLUT-3(322 bp). M, DNA ladder marker. Detection frequencies for the mRNAs are given at the bottom.
Vesicular glutamate transporter expression in supraoptic neurones 263
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

Because mitral cells form dendrodendritic synapses in theglomerular layer, this offers the possibility of VGLUT-1 beingthe transporter used by this cell type.
The detection of VGLUT-1 mRNA-containing SON tissuein three of five cases along with five of five cases using SONtissue plus the surrounding perinuclear region is consistentwith a previous report showing only faint labelling forVGLUT-1 mRNA in the SON and perinuclear area (11).However, neither our study nor theirs addresses the preciselocalisation of VGLUT-1 mRNA within specific cell types ofthe perinuclear zone. A recent anatomical study addressed thepossibility of glutamatergic perinuclear neurones by exploringthe origin of afferent glutamatergic terminals in the SON (13).These results, together with a previous physiological study(44), suggest that many perinuclear neurones are indeedglutamatergic.
Is Dale’s principle upheld?
Magnocellular neurones have two well-documented peptiderelease sites: (i) the NL terminals and (ii) somatodendriticallywithin nuclei. However, somatodendritic release of peptidesdoes not necessarily mirror peripheral release (1). Little isknown concerning MV exocytosis in the NL or from thedendrites. The intranuclear colocalisation of VGLUT-2 withOT and VP agrees with evidence first offered in a recentreview showing that the neural lobe was immunoreactive forVGLUT-2 (4). In agreement with Dale’s principle (45), thissuggests that the somata and dendrites, similar to theterminals, release the same transmitters: peptides and glu-tamate. However, the assumption that the VGLUT-2 label-ling seen in the NL is in magnocellular terminals may not be asafe one. First, it has long been known that there areterminals in the NL originating from neurones other thanmagnocellular neurones (46). Second, neocortical astrocytes,and thus possibly those in the NL (pituicytes), can expressVGLUT-2 (16). In the absence of a positive magnocellularterminal marker, it remains unclear which cell type in the NL(pituicyte, OT, VP, non-neurosecretory neurone, endothelial,etc.) expresses VGLUT-2.
Furthermore, ultrastructural experiments are still lackinginvestigating the subcellular localisation of specific VGLUTisoforms to particular classes of somatic and dendritic MVs.Information addressing these issues could shed further lighton alternative functions of VGLUTs and better clarifywhether SON neurones express VGLUT-1 at levels notdetectible at the light microscopic level. Additionally, phy-siological experiments monitoring VGLUT expression andsomatodendritic release of glutamate under different condi-tions could address whether or not triggers for glutamaterelease are distinctive or the same as those for peptide release.
Summary
In the present study, we investigate the presence of VGLUT-1,2 and 3 protein in the SON. Immunoreactivity for VGLUT-1was observed mainly lateral to the SON, whereas VGLUT-2-irwas apparent in positively identified magnocellular neuronalsomata and dendrites. Expression of VGLUT-3 was seen inSON neurones as well as local SON astrocytes, in addition to
being along endothelial and meningeal cells. Magnocellularneuronal expression of VGLUT-3 was mainly restricted to theperinuclear area of the soma. From these data, we suggest thatmagnocellular SON neurones are both peptidergic and gluta-matergic.
Acknowledgement
This research was supported by NIH grant NS009140.
Accepted 12 January 2006
References
1 Ludwig M. Dendritic release of vasopressin and oxytocin. J Neu-roendocrinol 1998; 10: 881–895.
2 Sladek CD, Kapoor JR. Neurotransmitter/neuropeptide interactionsin the regulation of neurohypophyseal hormone release. Exp Neurol2001; 171: 200–209.
3 Meeker RB, Swanson DJ, Greenwood RS, Hayward JN. Ultra-structural distribution of glutamate immunoreactivity within neu-rosecretory endings and pituicytes of the rat neurohypophysis. BrainRes 1991; 564: 181–193.
4 Hisano S, Nogami H. Transporters in the neurohypophysial neu-roendocrine system, with special reference to vesicular glutamatetransporters (BNPI and DNPI): a review. Microsc Res Tech 2002;56: 122–131.
5 Pow DV, Morris JF. Dendrites of hypothalamic magnocellularneurons release neurohypophysial peptides by exocytosis. Neuro-science 1989; 32: 435–439.
6 Moos F, Gouzenes L, Brown D, Dayanithi G, Sabatier N, Boissin L,Rabie A, Richard P. New aspects of firing pattern autocontrol inoxytocin and vasopressin neurones. Adv Exp Med Biol 1998; 449:153–162.
7 Fremeau RT, Voglmaier S, Seal RP, Edwards, RH. VGLUTs definesubsets of excitatory neurons and suggest novel roles for glutamate.Trends Neurosci 2004; 27: 98–103.
8 Gras C, Herzog E, Bellenchi GC, Bernard V, Ravassard P, Pohl M,Gasnier B, Giros B, El Mestikawy S. A third vesicular glutamatetransporter expressed by cholinergic and serotoninergic neurons.J Neurosci 2002; 22: 5442–5451.
9 Fremeau RT Jr, Burman J, Qureshi T, Tran CH, Proctor J,Johnson J, Zhang H, Sulzer D, Copenhagen DR, Storm-MathisenJ, Reimer RJ, Chaudhry FA, Edwards RH. The identification ofvesicular glutamate transporter 3 suggests novel modes of signa-ling by glutamate. Proc Natl Acad Sci USA 2002; 99: 14488–14493.
10 Hisano S, Hoshi K, Ikeda Y, Maruyama D, Kanemoto M, Ichijo H,Kojima I, Takeda J, Nogami H. Regional expression of a geneencoding a neuron-specific Na(+)-dependent inorganic phosphatecotransporter (DNPI) in the rat forebrain. Brain Res Mol Brain Res2000; 83: 34–43.
11 Ziegler DR, Cullinan WE, Herman JP. Distribution of vesicularglutamate transporter mRNA in rat hypothalamus. J Comp Neurol2002; 448: 217–229.
12 Lin W, McKinney K, Liu L, Lakhlani S, Jennes L. Distribution ofvesicular glutamate transporter-2 messenger ribonucleic acid andprotein in the septum-hypothalamus of the rat. Endocrinology 2003;144: 662–670.
13 Csaki A, Kocsis K, Kiss J, Halasz B. Localization of putativeglutamatergic/aspartatergic neurons projecting to the supraopticnucleus area of the rat hypothalamus. Eur J Neurosci 2002; 16:
55–68.14 Bezzi P, Gundersen V, Galbete JL, Seifert G, Steinhauser C, Pilati E,
Volterra A. Astrocytes contain a vesicular compartment that iscompetent for regulated exocytosis of glutamate. Nat Neurosci 2004;7: 613–620.
15 Meeker RB, Swanson DJ, Hayward JN. Light and electronmicroscopic localization of glutamate immunoreactivity in thesupraoptic nucleus of the rat hypothalamus. Neuroscience 1989;33: 157–167.
264 Vesicular glutamate transporter expression in supraoptic neurones
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265

16 Montana V, Ni Y, Sunjara V, Hua X, Parpura V. Vesicular glu-tamate transporter-dependent glutamate release from astrocytes. JNeurosci 2004; 24: 2633–2642.
17 McCarthy KD, deVellis J, Preparation of separate astroglial andoligodendroglial cell cultures from rat cerebral tissue. J Cell Biol1980; 85: 890–902.
18 Parpura V, Fang Y, Basarsky T, Jahn R, Haydon PG. Expression ofsynaptobrevin II, cellubrevin and syntaxin but not SNAP-25 incultured astrocytes. FEBS Lett 1995; 377: 489–492.
19 Bellocchio EE, Hu H, Pohorille A, Chan J, Pickel VM, EdwardsRH. The localization of the brain-specific inorganic phosphatetransporter suggests a specific presynaptic role in glutamatergictransmission. J Neurosci 1998; 18: 8648–8659.
20 Fremeau RT Jr, Troyer MD, Pahner I, Nygaard GO, Tran CH,Reimer RJ, Bellocchio EE, Fortin D, Storm-Mathisen J, EdwardsRH. The expression of vesicular glutamate transporters defines twoclasses of excitatory synapse. Neuron, 2001; 31: 247–260.
21 Schafer MK, Varoqui H, Defamie N, Weihe E, Erickson JD.Molecular cloning and functional identification of mouse vesicularglutamate transporter 3 and its expression in subsets of novel exci-tatory neurons. J Biol Chem 2002; 277: 50734–50748.
22 van den Pol AN, Wuarin JP, Dudek FE. Glutamate, the dominantexcitatory transmitter in neuroendocrine regulation. Science 1990;250: 1276–1278.
23 Meeker RB, Greenwood RS, Hayward JN. Glutamate is the majorexcitatory transmitter in the supraoptic nuclei. Ann NY Acad Sci1993; 689: 636–639.
24 Inenaga K, Cui LN, Nagatomo T, Honda E, Ueta Y, Yamashita H.Osmotic modulation in glutamatergic excitatory synaptic inputs toneurons in the supraoptic nucleus of rat hypothalamus in vitro.J Neuroendocrinol 1997; 9: 63–68.
25 Dyball RE, McKenzie DN, Thomas GP. Osmoresponsiveness of therat supraoptic nucleus in vivo depends on glutamatergic inputs.Neurobiology (Bp) 1995; 3: 351–362.
26 Hu B, Bourque CW. NMDA receptor-mediated rhythmic burstingactivity in rat supraoptic nucleus neurones in vitro. J Physiol 1992;458: 667–687.
27 Kombian SB, Hirasawa M, Mouginot D, Pittman QJ. Modulationof synaptic transmission by oxytocin and vasopressin in the supra-optic nucleus. Prog Brain Res 2002; 139: 235–246.
28 MeekerRB, SwansonDJ,GreenwoodRS,Hayward JN.Quantitativemapping of glutamate presynaptic terminals in the supraoptic nucleusand surrounding hypothalamus. Brain Res 1993; 600: 112–122.
29 Lafarga M, Andres MA, Berciano MT, Maquiera E. Organizationof nucleoli and nuclear bodies in osmotically stimulated supraopticneurons of the rat. J Comp Neurol 1991; 308: 329–339.
30 Armstrong WE. Morphological and electrophysiological classifica-tion of hypothalamic supraoptic neurons. Prog Neurobiol 1995; 47:291–339.
31 Hatton GI. Function-related plasticity in hypothalamus. Annu RevNeurosci 1997; 20: 375–397.
32 Palkovits M. Micro- and macroscopic structure, innervation, andvasculature of the hypothalamus. In: Conn PM, Freeman ME, eds.Neuroendocrinology in Physiology and Medicine. Totowa, NJ: Hu-mana Press, 2000: 23–40.
33 Freund-Mercier MJ, Richard P. Electrophysiological evidence forfacilitatory control of oxytocin neurones by oxytocin during sucklingin the rat. J Physiol 1984; 352: 447–466.
34 Moos F, Richard P. Paraventricular and supraoptic bursting oxy-tocin cells in rat are locally regulated by oxytocin and functionallyrelated. J Physiol 1989; 408: 1–18.
35 Wang YF, Hatton GI. Burst firing of oxytocin neurons in male rathypothalamic slices. Brain Res 2005; 1032: 36–43.
36 Brown CH, Bourque CW. Autocrine feedback inhibition of plateaupotentials terminates phasic bursts in magnocellular neurosecretorycells of the rat supraoptic nucleus. J Physiol 2004; 557: 949–960.
37 Roper P, Callaway J, Armstrong W. Burst initiation and terminationin phasic vasopressin cells of the rat supraoptic nucleus: a combinedmathematical, electrical, and calcium fluorescence study. J Neurosci2004; 24: 4818–4831.
38 Schrader LA, Tasker JG. Modulation of multiple potassium currentsby metabotropic glutamate receptors in neurons of the hypothalamicsupraoptic nucleus. J Neurophysiol 1997; 78: 3428–3437.
39 Schrader LA, Tasker JG. Presynaptic modulation by metabotropicglutamate receptors of excitatory and inhibitory synaptic inputs tohypothalamic magnocellular neurons. J Neurophysiol 1997; 77: 527–536.
40 Kaneko T, Fujiyama F, Hioki H. Immunohistochemical localizationof candidates for vesicular glutamate transporters in the rat brain.J Comp Neurol 2002; 444: 39–62.
41 Smithson KG, Weiss ML, Hatton GI. Supraoptic nucleus afferentsfrom the main olfactory bulb ) I. Anatomical evidence from ante-rograde and retrograde tracers in rat. Neuroscience 1989; 31: 277–287.
42 Hatton GI, Yang QZ. Supraoptic nucleus afferents from the mainolfactory bulb ) II. Intracellularly recorded responses to lateralolfactory tract stimulation in rat brain slices. Neuroscience 1989; 31:289–297.
43 Smithson KG, Weiss ML, Hatton GI. Supraoptic nucleus afferentsfrom the accessory olfactory bulb. evidence from anterograde andretrograde tract tracing in the rat. Brain Res Bull 1992; 29: 209–220.
44 Boudaba C, Schrader LA, Tasker JG. Physiological evidence forlocal excitatory synaptic circuits in the rat hypothalamus. J Neuro-physiol 1997; 77: 3396–3400.
45 Feldman RS, Meyer JS, Quenzer LF. Principles of Neuropsycho-pharmacology, 1st edn. Sunderland, MA: Sinauer; 1997: 909.
46 Boersma CJ, Van Leeuwen FW. Neuron–glia interactions in therelease of oxytocin and vasopressin from the rat neural lobe: the roleof opioids, other neuropeptides and their receptors. Neuroscience1994; 62: 1003–1020.
Vesicular glutamate transporter expression in supraoptic neurones 265
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 253–265
Related Documents











