Vasoactiv ágensek cardioprotectiv szerepe myocardialis ischaemia során Dr. Soós Pál Budapest 2006 Témavezető: Dr. Horkay Ferenc, PhD, DsC Semmelweis Egyetem Elméleti Orvostudományok Doktori Iskola Program: Szív- és érrendszeri betegségek élettana és klinikuma Bírálók: Dr. Mohácsi Attila, PhD Dr. Szokodi István, PhD Szigorlati bizottság: Prof. Dr. Sándor Péter, PhD, DSc Prof. Dr. Székács Béla, PhD Dr. Szokodi István, PhD

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vasoactiv ágensek cardioprotectiv szerepe myocardialis ischaemia során
Dr. Soós Pál
Budapest 2006
Témavezető: Dr. Horkay Ferenc, PhD, DsC
Semmelweis Egyetem Elméleti Orvostudományok Doktori Iskola Program: Szív- és érrendszeri betegségek élettana és klinikuma
Bírálók: Dr. Mohácsi Attila, PhD Dr. Szokodi István, PhD Szigorlati bizottság: Prof. Dr. Sándor Péter, PhD, DSc Prof. Dr. Székács Béla, PhD Dr. Szokodi István, PhD

1
Tartalomjegyzék
Tartalomjegyzék 0
Rövidítések jegyzéke 2
1 Összefoglalás 5
2 Summary 6
3 Bevezetés 7
3.1 Pitvari nátriuretikus peptid (ANP) 9 3.2 A pericardialis folyadéktér 14 3.3 Parathormon (PTH) 18 3.4 L-arginin 21
4 Célkitűzések 25
5 Módszerek 27
5.1 Humán vizsgálatok 27 5.2 Experimentális modellek 28 5.3 Biokémiai vizsgálatok 32 5.4 Kompartmentalizációs matematikai modell 34 5.5 Statisztikai módszerek 36
6 Eredmények 37
6.1 Pericardialis és szöveti ANP szintek elemzése 37 6.2 Az interstitialis és pericardialis folyadéktér dinamikája… 41 6.3 Parathormon hatás a coronariákon 43 6.4 Szisztémás L-arginin hatása a reperfúzióra 47 6.5 L-arginin vascularis hatásai agyhalál állapotában 51
7 Megbeszélés 55
8 Következtetések 71
9 Irodalomjegyzék 73
10 Saját publikációk jegyzéke 91
11 Köszönetnyilvánítás 92

2
Rövidítések jegyzéke
±dP/dt kontraktilitás
ACH acetilkolin
ADO adenozin
AMP adenozin monofoszfát
ANP pitvari (atrialis) nátriuretikus peptid
AoP aortanyomás
AT-II angiotensin II
ATP adenozin trifoszfát
BD agyhalál (brain death)
BNP B-típusú (brain) nátriuretikus peptid
CA szívmegállás (cardiac arrest)
CAD ischaemiás szívbetegség
cAMP ciklikus adenozin monofoszfát
CBF coronariaáramlás
CNP C-típusú nátriuretikus peptid
CO perctérfogat (cardiac output)
CPB cardiopulmonalis bypass
EDRF endothelium-derived relaxing factor
eNOS endothelialis nitrogén monoxid szintáz
ESPVR end-systolic pressure-volume relationship
ET-1 endothelin-1
HR szívfrekvencia
IMP inozin monofoszfát
INO inozin
iNOS indukálható nitrogén monoxid szintáz
KATP ATP-függő kálium csatorna
L-arg L-arginin
MAP artériás középnyomás
NEP neutralis endopeptidáz

3
nNOS neuralis nitrogén monoxid szintáz
NO nitrogén monoxid
NYHA New York Heart Association
PRSW preload recruitable stroke work
PTH parathormon
SAH S-adenozil-homocisztein
SNP nátrium-nitroprussid
VHD szívbillentyű betegség
VP vénás plazma
A matematikai modellben alkalmazott rövidítések: PSg a kapillárisok permeábilis területén átjutó anyag
mennyisége a plazma és az interstitium között
(mL*g-1*min-1)
PSecl az endothelsejtek lumen felőli membránjának permeábilis
területén átjutó anyag mennyisége a plazma és az
endothelsejt cytosol között (mL*g-1*min-1)
PSeca az endothelsejtek lumennel ellentétes oldali (abluminalis)
membránjának permeábilis területén átjutó anyag
mennyisége az interstitialis folyadék és az endothelsejt
cytosol között (mL*g-1*min-1)
PSpc a parenchymasejtek membránjának permeábilis területén
átjutó anyag mennyisége az interstitialis folyadék és a
parenchymasejt cytosolja között (mL*g-1*min-1)
PStransz a transzszudációs folyadék és az interstitialis folyadék
közötti molekulaáramlás nagysága (mL*g-1*min-1)
V’isf az eloszlás során kitöltött tér az interstitiumban (mL/g, az
adott terület-plazma közötti áramlás partíciós koefficiense
szorozva az anatómiai térfogattal (V))
V’ec az eloszlás során kitöltött tér az endothelsejtekben (mL/g,
anatómiai adatok alapján 0.02 mL/g értékben megállapítva)

4
V’pc az eloszlás során kitöltött tér a parenchymasejtekben
(mL/g, kevéssé érzékeny paraméter 0.5-0.7 mg/L között)
Gp az intravasalis metabolizmus során keletkezett anyagok
clearance-e (mL*g-1*min-1)
Gisf az interstitialis metabolizmus során keletkezett anyagok
clearance-e (mL*g-1*min-1)
Gec az intracellularis metabolizmus során keletkezett anyagok
clearance-e az endothelsejtekben (mL*g-1*min-1)
Gpc az intracellularis metabolizmus során keletkezett anyagok
clearance-e a parenchymasejtekben (mL*g-1*min-1)
Gtransz a transzszudációs térben folyó metabolizmus során
keletkezett anyagok clearance-e (mL*g-1*min-1)
Fp áramlás per szövettömeg (mL/g/sec)
C koncentráció (mol/cm3)
D tengely irányú diffúzió, illetve diszperziós koefficiens
(cm2/sec)
L kapilláris hossza (cm; szívben 0.05-0.1 cm)
t idő (sec)
x távolság a kapillárisban (cm)
V egy terület térfogata (mL)
V’ megoszlási térfogat egy adott területen (mL)
α plazmaáramlás sebessége (cm/sec);=Fp*L

5
1 Összefoglalás
Myocardialis ischaemia során egy összetett védekező rendszer
aktiválódik, melynek lokálisan (parakrin) és szisztémásan (endokrin) ható
alkotóelemeit tanulmányoztuk humán és experimentális vizsgálataink
során. Ezek egyik kitüntetett eleme a pericardialis tér volt, melynek a
szívműködés szabályozásában betöltött szerepe az elmúlt évek során
körvonalazódott. Leírtuk a pitvari nátriuretikus hormon (ANP)
koncentrációviszonyait a szívizomszövetben, a plazmában és a
pericardialis folyadékban, továbbá vizsgáltuk az ANP-szintek változásait
különböző klinikai állapotokban. Feltételezésünk szerint a pericardialis
folyadéktér kinetikai szempontból az interstitialis térhez kötött, és a
pericardialis folyadékban mért anyagkoncentrációk talán e nehezen
hozzáférhető területről is indirekt felvilágosítást adhatnak. Elemeztük a
folyadék-kompartmentek közötti anyagcsere-kapcsolatokat, és egy
matematikai modellt készítettünk, mellyel az interstitialis folyadék egyes
alkotóelemeinek koncentrációját becsülhetjük meg a pericardialis
folyadékban és a vérplazmában mért szintjeik függvényében. Az ismert
endokrin tényező, a parathormon coronariadilatatióban játszott szerepét
elemezve megállapítottuk, hogy a cAMP rendszert és az L-típusú Ca
csatorna inaktivációt involváló reakcióutak közötti kapcsolatot a KATP
csatornák jelenthetik a peptid cellularis hatásmechanizmusának kifejtése
során. A NO prekurzor L-arginin cardioprotectiv hatását igazoltuk
szisztémás adás mellett hypothermiás szívmegállást követő reperfúzió
során. Emellett kimutattuk jelentőségét az endothelialis funkció
javításában mind ischaemia-reperfúziós folyamatokat követő, mind az
agyhalál jelenségéhez kapcsolódó endothel diszfunkció kialakulása során.
Vizsgálataink eredményei alapján a vasoactiv ágensek szerepe a
cardioprotectio mechanizmusában jelentős, részletes ismeretek
birtokában, alkalmazásukkal a klinikai terápiás lehetőségek tovább
bővíthetők.

6
2 Summary
The aim of our human and experimental studies was to examine the local
(paracrine) and systemic (endocrine) components of the complex
protective mechanism that activates during myocardial ischaemia.
Recently the role of pericardial fluid in cardiac regulation became certain,
that’s why we have paid special attention on its function. We characterized
the atrial natriuretic peptide (ANP) concentrations of human myocardial
tissues, in blood plasma and in pericardial fluid, moreover we analyzed the
relationship between ANP levels and some special clinical conditions.
Theoretically, pericardial fluid may reflect the composition of interstitial
fluid, thus concentrations measured in pericardial fluid samples may
indirectly provide information about the interstitial space. To present
opportunity even to esteem the composition of interstitial fluid we
composed a mathematical model using blood plasma samples and
pericardial fluid samples to calculate interstitial concentrations. Moreover
we characterized the role of the well known endocrine factor parathyroid
hormone in coronary dilation and concluded that in addition to the
activation of the intracellular cAMP system and inactivation of the L-type
Ca channels described in connection with parathyroid hormone-induced
coronary responses ATP-sensitive potassium channels may play an
integrative role in the intracellular mechanisms of action of the peptide. In
further studies we demonstrated the cardioprotective effect of the NO
precursor L-arginin after reversible deep hypothermic ischaemia and
reperfusion even if applied systemically. In addition, beneficial effect on
endothelial function was proved during ischemic-reperfusion injury, as well
as during endothelial dysfunction following brain death. According to our
results, the role of vasoactive agents in the mechanism of cardioprotection
is considerable, therefore clinical applications based on these molecules
may provide new therapeutic potentialities in the near future.

7
3 Bevezetés
Élettani körülmények között a koszorúerekben a szívizomszövet
metabolikus igényét kielégítő mennyiségű vér áramlik. Ischaemiás
szívbetegségekben a myocardium vérellátása elégtelenné válik, melynek
oka lehet a csökkent véráramlás (coronaria stenosis vagy occlusio), vagy
a megnövekedett perfúziós igény (hypertrophiás myocardium). A
myocardialis ischaemia egy lokális hypoxiás állapot, ami alapvetően
befolyásolja a mitokondriális energiametabolizációs folyamatokat;
csökkenti a kreatin-foszfát és ATP szinteket, fokozza a piruvát-laktát
átalakulást, ami végül intracellularis acidózishoz vezet. Ezeknek az
intracellularis biokémiai változásoknak az eredménye, hogy csökken a
myocardialis kontraktilitás, továbbá elektromos vezetési zavarok jönnek
létre, szívritmuszavarok alakulhatnak ki. A kontraktilitás csökkenése a
perctérfogat romlását okozza, ami elsősorban a terhelhetőség
csökkenését vonja maga után.
A szív pumpafunkcióján túl számos olyan anyagot is termel, melyek
közvetlenül befolyásolják önnön működését vagy részt vesznek
szisztémás szabályozó folyamatokban (pl: vérnyomás illetve a só- és
vízháztartás szabályozásában). Ezek közül legismertebbek a nátriuretikus
peptidek, az endothelinek, a renin-angiotenzin rendszer egyes
komponensei, a nitrogén monoxid (NO), illetve a prostaglandinok. Ezen
ágensek hatásai részben a klasszikus endokrin módon érvényesülnek (pl.
a nátriuretikus peptidek); de feltételezhető, hogy a szív kontraktilitására,
vérellátására, a sejtosztódásra és a növekedés folyamataira a substantiák
többsége közvetlenül, autokrin/parakrin módon is kifejti hatását.
Másrészről, számos, nem a szívben termelődő vasoactiv ágens is
befolyásolhatja a koszorúerek működését. Az endokrin szabályozás ezen
formájára példa a mellékpajzsmirigyben termelődő parathormon (PTH)
hatása, amely elsődlegesen a csontokból történő Ca++ mobilizációval,
illetve foszfátürítés renalis regulációjával a szervezet Ca++ és a

8
foszfátháztartásának szabályozásáért felelős. Emellett azonban
kimutatták, hogy a PTH-nak vasodilatator, ezáltal hypotensiv hatása is
van, illetve növeli a szívfrekvenciát és a bal kamra kontrakciós erejét. A
PTH simaizom sejteken található receptorához kötődve, adenil-cikláz
aktiválásával fejti ki vasodilatator hatását.
A koszorúerek keringésének autoregulációs szabályozásában fontos
szerepet játszik a nitrogén monoxid. A NO erőteljes vasodilatator hatása
mellett gátolja a thrombocyta aggregációt, csökkenti a leukocyta adhéziót
és gátolja az erek simaizom-proliferációját. A NO képződés elsődleges
helyei a coronariák endotheliuma, a cardiomyocyták és az endocardialis
sejtek, ahol L-argininből képződik. Az L-arginin-NO „tengely” fontos
szerepet játszik az ischaemia-reperfúziós folyamatok során; növeli a
postischaemiás coronaria-áramlást, csökkenti az adhéziós molekulák
expresszióját, emellett antioxidáns hatása is feltételezhető.
Ismert, hogy első sorban a cardiomyocyták, az endocardium és a
vascularis endothelium sejtjei képesek cardioactiv faktorok szintézisére.
Míg az endocardium és az endothelium élettani szerepét intenzív
kutatások vizsgálják, ez ideig kevés figyelmet fordítottak az epicardium
élettani jelentőségére. Az epicardium külső felszínét mesothelsejtek
borítják, melyek fejlődéstanilag azonos eredetűek az endotheliummal,
ezért feltételezhető, hogy ezek a sejtek is termelnek cardioactiv faktorokat.
Ezek az anyagok az interstitialis folyadék közvetítésével eljuthatnak a
pericardialis folyadékba és ott feltehetően a lassú eliminációnak
köszönhetően akkumulálódnak. A pericardialis folyadékban így kialakuló
magas cardioactiv szubsztancia koncentráció hatást gyakorolhat a szív
működésére. Az endocardium és a microvascularis endothelium
szívizomra kifejtett hatása, valamint a szívizomsejtek egymás közötti
interakciója mellett, az epi- és pericardialis mesothel sejtek által termelt
ágensek is jelentősen befolyásolhatják a szívműködést. Mindezek alapján
joggal feltételezhető, hogy a szívben több egymással szoros kapcsolatban
lévő autokrin/parakrin mechanizmus létezhet.

9
Ezen megfontolások alapján vizsgálatainkban a myocardialis ischaemia
során cardioprotectiv szerepet játszó vasoactiv ágensek hatását
elemeztük, a coronaria keringés lokális szabályozása mellett kiemelt
figyelmet fordítva a pericardialis tér regulátor funkciójára.
3.1 Pitvari nátriuretikus peptid (ANP)
A nátriuretikus peptid családot három olyan molekula alkotja, melyek
hasonló karakterisztikájú aminosav szekvenciával és hurkolt szerkezettel
rendelkeznek: az atrialis nátriuretikus peptid (ANP), az agyi (brain)
nátriuretikus peptid (BNP) és a C-típusú nátriuretikus peptid (CNP). Az
ANP és a BNP első sorban a szívizomszövetben termelődik, a CNP a
központi idegrendszer neuronjaiban és a vascularis endotheliumban (1). A
BNP élettani hatásai többségükben megegyeznek az ANP hatásaival,
azonban plazmakoncentrációja érzékenyebben reagál a szívelégtelenség
progressziójára (2).
3.1.1 Molekuláris biológia
De Bold és munkatársai 1981-ben megfigyelték, hogy pitvari izomszövet
kivonatot infúzióban juttatva patkányok keringésébe, az állatok
natriuresise és diurézise jelentősen megnőtt (3). Ez a felfedezés vezetett
az ANP izolálásához és klónozásához, mely az első tagja volt egy olyan
peptid-családnak, ami nátriuretikus, diuretikus és vasorelaxans aktivitással
rendelkezik. Az ANP szintézise első sorban a pitvari cardiomyocytákban
történik, azonban a kamrákban is kimutatható kis mértékű ANP képződés
(4). Az ANP molekula prekurzora a 126 aminosavból álló proANP
molekula, ami biológiailag inaktív és a szívizomsejtek granulumaiban
tárolódik. Megfelelő szignál hatására membránhoz kötött proteázok a
proANP-t tovább hasítják egy N-terminális darabra (ANP [1-98]), és egy C-
terminális peptidre (ANP [99-126]) (5). A biológiailag aktív ANP ez utóbbi,
28 aminosavat tartalmazó peptid, mely jellemzője a 17 aminosavat

10
tartalmazó gyűrű, amit egy diszulfid-híd zár két cisztein molekula között (1.
ábra). A humán plazmában meglehetősen magas a proANP N-terminális
szegmentjének koncentrációja (az ANP szintjének több, mint 20-szorosa).
Ez feltehetően a keringésből történő lassabb eliminációjának köszönhető,
ennek ellenére az N- és a C-
terminálisok plazmában mért
koncentrációja erősen korrelál (6).
Emellett a vesékben is kimutatható az
ANP gén expressziója, melynek
során egy alternatív képződési
útvonalon a proANP molekulából egy
32 aminosavból álló peptid keletkezik,
az urodilatin (ANP [95-126]). Ennek
feltehetően a vese lokális nátrium- és
vízháztartás szabályozásában van
szerepe (7).
A normál plazma ANP koncentrációja
10 és 60 pg/mL között van. Az ANP felezési ideje a plazmában 2-5 perc.
Eliminációjában majdnem minden szövet részt vesz; így a tüdők, a vesék,
a gastrointestinalis rendszer, illetve a nagyobb izomtömegek. Az ANP
lebontása két úton történik; az egyik során egy neutrális endoproteáz
(NEP) hasítja, így hozva létre a csökkent aktivitású nyílt láncú formát. A
másik esetben ún. clearance receptorok (NPR-C receptor) kötik meg az
ANP-t a plazmából, a receptor-ligand komplex átjut a sejtmembránon és a
ligandot lizoszómák hidrolizálják (8).
3.1.2 Élettani és patofiziológiás jelentőség
Az ANP szekréció legfontosabb meghatározója a pitvari falfeszülés. Bár a
pitvari nyomás változása is pozitívan korrelál a plazma ANP
koncentrációjával, kimutatták, hogy a pitvari transmuralis nyomás, így a
falfeszülés, az elsődleges meghatározója a keringő ANP mennyiségének
1. ábra: A pitvari nátriuretikus peptid biológiailag aktív formája; ANP [99-126]

11
(9). A pitvari falfeszülés növekedésével különböző endogén faktorok is
felszabadulnak, melyek szintén befolyásolhatják az ANP szekrécióját.
Ilyen az endothelin-1 (ET-1) és az angiotensin II (AT-II), melyek fokozzák
az ANP szekréciót, illetve a NO, ami csökkenti ezt (10).
Az ANP által létrehozott különböző hatások célja akut és krónikus
körülmények között a szisztémás vérnyomás és az intravascularis
folyadék mennyiségének csökkentése.
Az ANP-nek két fő érhatása van; fokozza a plazma redisztribúcióját az
extravasalis térbe, és vasodilatatiót okoz, feltehetően a vascularis
simaizmok intracellularis Ca++ koncentrációjának csökkentésével. Az ANP
emellett csökkenti a perifériás erek szimpatikus tónusát. Az ANP a
vesékben hyperfiltratiót és natriuresist okoz (kapcsolódó kaliuresis nélkül);
a gyűjtőcsatornák Na+ és vízvisszaszívásának gátlásával. Csökkenti a
macula densa renin szekrécióját, csökkenti a keringő AT-II szintjét, és
direkt gátló hatása van a zona glomerulosa aldoszteron szekréciójára (8).
A centrális vénás nyomás emelkedésével járó állapotokban emelkedik a
vér ANP koncentrációja, és ez az intra-atrialis nyomás, illetve pitvari
falfeszülés ANP szekrécióban játszott meghatározó szerepét bizonyítja.
Az emelkedett plazma ANP koncentráció részben a pitvarok fokozott ANP
szintézisének következménye, ugyanakkor kialakulásában a peptid
eliminációja is meghatározó, ami clearance receptorokhoz kötődve vagy a
neutrális endopeptidáz hidrolízisével történhet. Az ANP felezési ideje a
keringésben viszonylag rövid. Hasonlóan az ANP-hez, emelkedett BNP
vérkoncentrációkat is mértek szívbetegekben. A BNP eliminációja az ANP-
hez hasonlóan clearance receptorokon és neutrális endopeptidáz útján
történik, felezési ideje a keringésben szintén rövid. Ezek alapján nem
meglepő, hogy jelentősen emelkedett ANP és BNP plazma koncentrációk
mérhetők jelentős intravasalis volumenterhelés, emelkedett centrális
vénás nyomás, tachycardia és csökkent veseműködés mellett.
A különböző betegség típusokat, illetve az ezeken belüli egyes
stádiumokat vizsgálva komoly különbségeket tapasztalhatunk (1. táblázat).

12
Ezek a különbségek az egyes kórképek eltérő patofiziológiai eredetével
magyarázhatók.
Mindemellett, emelkedett plazma nátriuretikus peptid koncentrációkat
mértek esszenciális hypertoniás betegekben is, ami azért érdekes, mert az
esszenciális hypertonia nem jár jelentős vértérfogat emelkedéssel; bár
ilyen betegekben normális pumpa funkció mellett- a centrális vénás
nyomás és a pulmonalis kapilláris nyomás emelkedését tapasztalták (11).
Az emelkedett praeload a hypertrophiás szívizom ANP és BNP termelését
fokozza; részben ez okozhatja a plazma ANP szintek szignifikánsan
1. táblázat: Plazma ANP és BNP szintek néhány cardiovascularis betegségben (Sagnella 1998, átdolgozva). A jelek a relatív növekedés közelítő értékeit mutatják a megfelelő kontrollokhoz viszonyítva: ▲ kisebb, mint 3-szoros; ▲▲ kb. 3-10-szeres; ▲▲▲ kb. 10-30-szoros; ▲▲▲▲ nagyobb, mint 30-szoros.
ANP BNP
Esszenciális hypertonia ▲ ▲
Tachycardiák ▲▲ ▲
Szívelégtelenség ▲▲▲ ▲▲▲
Izolált diastolés diszfunkció ▲▲ ▲▲
Mitralis stenosis ▲▲ ▲▲
Aortastenosis ▲ ▲▲▲
Dilatatív cardiomyopathia ▲▲▲ ▲▲▲▲
Hypertrophiás cardiomyopathia
nem obstruktív ▲▲ ▲▲▲
obstruktív ▲▲ ▲▲▲▲
Myocardialis infarctus
beteg felvételekor ▲ ▲▲▲
a felépülés során ▲ ▲▲▲▲
Krónikus veseelégtelenség
dialízis kezelést igénylő ▲▲ ▲▲
dialízis kezelést nem igénylő ▲▲▲ ▲▲▲

13
emelkedett koncentrációit. A plazma BNP főként a kamrákból származik.
Fiziológiás körülmények között az ANP plazma koncentrációja sokkal
magasabb, mint a BNP koncentráció; azonban szívelégtelenségben a
plazma BNP szintje eléri, sőt meghaladja az ANP szintjét, ami a
szívelégtelenségben jelentkező kamrai és pitvari túlterhelést mutatja (2).
Ezen túl, a szívelégtelenség különböző stádiumaiban levő betegek plazma
ANP és BNP koncentrációit vizsgálva, korreláció mutatható ki a
koncentrációk és a kamrai diszfunkció között (12). Az ANP hatására a
plazmából folyadék lép ki az extravasalis térbe, ami szívelégtelenség
során szerepet játszik a perifériás oedema képződésben.
A plazma ANP és BNP szintjei mellett a N-terminális ANP is jól jellemzi a
bal kamrai szisztolés funkció csökkenését; hosszú felezési ideje miatt első
sorban a hosszú távú ANP-termelésről ad tájékoztatást (13).
Myocardialis infarctus után emelkedett ANP és BNP koncentrációk
mérhetők a plazmában. A myocardialis infarctus korai fázisában
jelentősen emelkedett (kb. 60-szoros) plazma BNP koncentrációkat
mértek, míg a plazma ANP koncentrációja csak kisebb (kb. 3-szoros)
mértékben emelkedett a kontroll értékekhez képest. Az ANP plazma
szintje az infarctus tüneteinek megjelenése utáni 8 órán belül már a
legmagasabb értéket mutatta, és ez a folyamatosan csökkent a normál
érték eléréséig, amit az első 24 órán belül ért el. A BNP plazma szintjében
gyors emelkedés volt tapasztalható, a csúcsot kb. 20 órával a tünetek első
megjelenése után érte el. A plazma ANP szintekben továbbá második
csúcs is megjelenhet az infarctus subacut fázisában (bifázisos időbeni
lefutás), aminek lehetséges magyarázata, hogy a később gyorsan, nagy
mennyiségben ürített ANP szintézisében és szekréciójában már a kamrák
is részt vesznek (14).
A vérplazma mellett a pericardialis folyadékban is kimutatták az ANP-t, és
szívműtétre kerülő betegek pericardialis folyadékát vizsgálva jelentős
különbség mérhető a plazma, illetve pericardialis folyadék ANP szintjei
között (15).

14
3.2 A pericardialis folyadéktér
A pericardium – a szívet körülölelő zsák – elsődleges szerepe a szív
mellüregben elfoglalt helyének megtartása, izolációja a szomszédos
szervektől (pl. gyulladások esetén) és védelme az esetleges mechanikai
behatásoktól. Határt szab a szívtérfogat akut növekedésének, pl.
papillárisizom ruptura, pulmonalis embolia esetén – mintegy ellentartva az
emelkedett kamrai végdiasztolés nyomásnak. Emellett limitálja a
verőtérfogat nagyságát szívmegnagyobbodás esetén, krónikus
szívelégtelenségben. A pericardialis zsákot kitöltő folyadék, mintegy
„kenőanyag” csökkenti a szív és a pericardium közötti súrlódást, és ezt a
hatást pericardium által termelt foszfolipid surfactant anyagok tovább
fokozzák. A pericardialis folyadék egészen az elmúlt időkig csak csökkent,
illetve kórosan megemelkedett állapotával hívta fel magára a klinikus
orvos figyelmét. Az elmúlt évek élettani kutatásai azonban rámutattak,
hogy a pericardialis folyadék is nagy számban tartalmaz olyan biológiailag
aktív molekulákat, melyek szerepét korábban csak a keringésben tartották
meghatározónak. Ezek az eredmények arra késztették a fiziológusokat,
hogy a pericardialis folyadékot egy önálló folyadéktérként kezeljék és
feltárják kapcsolódási pontjait a szervezet élettanilag fontos
kompartmentjeihez; így a keringő vérhez, pericardiumot alkotó sejtekhez,
szívizomsejtekhez és az interstitialis térhez.
3.2.1 A pericardialis folyadék összetétele
A pericardialis folyadék emberben kb. 15-35 mL serosus folyadék, melyet
a pericardium termel; fehérje- és ionösszetétele nagyjából megfelel egy
plazma ultrafiltrátumnak. A pericardium termeli, de a myocardialis
interstitialis folyadék és a myocardialis nyirokelvezetések alkotói is
megtalálhatóak benne (16). A pericardialis folyadék ozmolaritása
alacsonyabb a plazmáénál; fehérje tartalma mintegy harmada a
plazmában mértnek. Magyar szerzőktől származik az a közel harminc
éves gondolat, hogy a pericardialis térben fellelhető ágensek

15
egyszersmind a biológiai szabályozás hatótényezői is lehetnek (17). A
pericardialis folyadékban mért, a plazmában mértnél magasabb peptid és
más makromolekula koncentrációk azonban azt jelzik, hogy az itt
megtalálható makromolekulák elsődleges forrása nem lehet a vérplazma
(18,15,19,20). Irodalmi adatok alapján, szívműtétre kerülő betegekben a
plazmaszintnél nagyságrendekkel nagyobb pericardialis folyadék ET-1
koncentrációk mérhetők (21,15), és hasonló eredmények születtek adenin
nukleozidokat (22), illetve ferritint (20) vizsgálva is. Bár ezeknek az
anyagoknak sem pontos keletkezési helye, sem a szívműködésben
betöltött szerepe nem eléggé ismert, feltételezhetően hatással vannak a
szívben lejátszódó biológiai folyamatokra (23). A pericardialis folyadékban
jelen levő endothelin-1 szerepet játszhat a koszorúerek véráramlásának
szabályozásában (24), illetve aritmiák kialakulásában (25,26), míg az
adenin nukleozidok (adenin+inozin) az ischaemiás szívizom
vérellátásának kompenzációs szabályozásában vehetnek részt. Ez utóbbi
feltételezést támasztja alá, hogy koszorúér betegek pericardialis
folyadékában szignifikánsan magasabb adenin nukleozid koncentrációk
mérhetők, mint billentyű betegek pericardialis folyadékában (22). Az ANP
esetében a helyzet nem ennyire egyértelmű. Kis betegszámú
vizsgálatokban humán pericardialis folyadékban a plazma szintnél sokkal
alacsonyabb ANP koncentrációkat mértek (27), amit későbbi vizsgálatok
azonban nem tudtak megerősíteni (18,15). A pericardialis folyadékban
jelen levő ANP a koszorúserek tónusának szabályozásában játszhat
szerepet (28).
3.2.2 Anyagcsere a pericardialis folyadéktérben
A pericardialis folyadék nagy számban tartalmaz olyan biológiailag aktív
molekulákat, melyek szerepét korábban csak a keringésben tartottak
meghatározónak. Ezek a megfigyelések adtak alapot arra, hogy a
pericardialis folyadékot egy önálló folyadéktérként kezeljék és feltárják
kapcsolódási pontjait a szervezet élettanilag fontos kompartmentjeihez;

16
így a keringő vérhez, pericardiumot alkotó sejtekhez, szívizomsejtekhez és
az interstitialis térhez. Az interstitialis tér összetevőinek közvetlen
meghatározása jelen ismereteink szerint biztonsággal nem megoldható,
ezért a biológiailag hozzáférhető folyadékterek vizsgálatával és
matematikai eljárások kidolgozásával lehet koncentráció-viszonyait
megbecsülni (29). Az élettani folyamatok matematikai modellezésére
olyan kisméretű molekulák alkalmasak, melyek jól követhetően
(mérhetően) vesznek részt a szabályozó folyamatokban. Ezen
megfontolások alapján az adenin-nukleozid rendszer vizsgálata megfelelő
választásnak tűnik, mert jelentős szerepe van a koszorúerek metabolikus
adaptációjában, emellett a felszabaduló, kis molekulatömegű adenozin
(ADO) és inozin (INO) az epicardialis felszínen keresztül szabadon
diffundál, a pericardialis térben felhalmozódik és mennyisége ezáltal
könnyen meghatározható (30). Feltételezésünk szerint, az epicardialis
felszínen történő szabad molekulaáramlás következtében az interstitialis
és pericardialis folyadék között egyensúly alakul ki a nukleozidok
tekintetében (31). Így, a pericardialis folyadék nukleozid koncentrációja jól
reprezentálhatja az interstitialis viszonyokat.
Az ADO mind a normál, mind az ischaemiás szívizomsejtekben
folyamatosan termelődik, és választódik ki az interstitialis folyadékba. A
szívizom ADO termelését az O2 igény és ellátás közötti egyensúly
megbomlása fokozza, ezáltal nő a sinus coronariusba ürített mennyiség.
Az ADO fő forrása az intracellularis AMP, aminek a koncentrációja függ az
intracellularis ATP mennyiségétől, illetve a sejt energetikai állapotától. Így,
ha fokozódik az ATP hidrolízise vagy csökken az ATP szintézis mértéke, a
sejt energetikai töltöttsége is csökken, és nő az AMP koncentrációja (32).
Az AMP-t az 5’ nukleotidáz enzim defoszforilálja, így képződik az ADO.
Kimutatták, hogy a sejtek energia szintjét csökkentő ingerek (pl. hypoxia,
ischaemia vagy munkavégzés) nagyban fokozzák ezen enzim működését
és ezen keresztül serkentik az AMP→ADO átalakulást. Az 5’ nukleotidáz
enzim megtalálható a cytosolban (cytosol 5’ nukleotidáz) és a
szívizomsejtek felszíni membránján is (ecto 5’ nukleotidáz). Ezért ADO

17
keletkezhet intracellularisan és extracellularisan is az AMP lebontásával. A
cardialis ADO képződés első sorban a szív ereinek endotheliumára és a
cardiomyocytákra lokalizálható. Intracellularisan ADO képződhet
transzmetiláció útján is, S-adenozil-homocisztein (SAH) hidrolízisével,
amely során ADO és homocisztein keletkezik. Ezt a reakciót a SAH
hidroláz enzim katalizálja. Az ezen az úton képződő ADO mennyisége
nagyjából megfelel annak a mennyiségnek, ami a szívben keletkezik
normoxiás állapotban, és úgy tűnik nem változik számottevően hypoxia
során (33). Ezek alapján feltételezhetjük, hogy az 5’ nukleotidáz általi AMP
defoszforiláció a felelős a fokozott ADO képződésért hypoxiás állapotban.
Tehát, a transzmetilációs útvonal a hypoxián kívüli állapotok egyik jelentős
szabályozó tényezője (34). A két folyamat dinamikáját mutatja a 2. ábra.
Az ADO inaktiválása három úton történhet: az ADO kináz AMP-vé
foszforilálja (így kerülhet vissza az ATP raktárba), az ADO deamináz
inozinná alakítja vagy kikerül a keringésbe. Fiziológiás koncentrációban az
ADO első sorban AMP-vé alakul, ennél nagyobb koncentrációnál azonban
Epicardium Parietálispericardium
INTERSTICIÁLISTÉR
HCYS
ADO-deamináz
PERICARDIÁLISTÉR
SZÍVIZOMSEJT
INO
ADO
INTRAVAZÁLISTÉR
ADO-deamináz
diffúzió
diffúzióC ADO
INO
ADO
INO
ADO-deamináz
ADO-deamináz
diffúzió
AMP
ADP
SAH
SAM
5 ' nukleotidáz
ADO
INO
ATP
Hypoxia
MET
Normoxia
C
2. ábra: Adenin nukleozidok képződése a szív pericapillaris szöveteiben

18
deaminálódik, és inozin lesz belőle. Az ADO átalakulhat adeninné is,
amiből utána ugyancsak AMP lehet. Ez a mentő (salvage) út szintén az
intracellularis adenin nukleotid raktárak feltöltődését szolgálja. Az ADO
képződése az 5’ nukleotidáz által történhet a szívizomsejtekben, az
endothel sejtekben, illetve a vörösvértestekben, azonban az ADO
deamináz megtalálható extracellularisan is (34).
INO a szívben az ADO deamináz hatására képződik elsősorban akkor,
amikor az ADO a fiziológiás koncentrációt meghaladó mennyiségben van
jelen. Az INO lebomlása során hypoxantinná alakul, amely foszforilálódik
és IMP-vé alakul. Az IMP-ből transzmetiláció során AMP keletkezik, mely
az intracellularis nukleotid raktár feltöltését szolgálja.
Feltételezésünk szerint az öt legfontosabb paraméter (myocardialis,
endothelialis, pericardialis, vénás keringési és interstitialis ADO
koncentráció) egymással szoros összefüggésben változik. Amennyiben a
köztük fennálló kapcsolatot helyesen, kellő pontossággal le tudjuk írni,
akkor egy ismeretlen érték a többi ismeretében kiszámítható.
3.3 Parathormon (PTH)
A humán parathyreoidea-hormon, (parathormon, PTH) 84 aminosavat
tartalmazó lineáris polipeptid. A
szervezet kalcium homeo-
sztázisának fenntartásában
betöltött szerepe jól ismert.
Ugyanakkor számos kísérlet
kimutatta a PTH, és annak
biológiailag aktív amino-
terminális fragmentjének, a PTH
(1-34)-nak vasodilatator, követ-
kezetesen hypotensiv hatását (35). Crass és munkatársai elsőként
figyelték meg, hogy intracoronariásan adott PTH dózisfüggő vasodilatatiót
hoz létre (36). Később leírták, hogy a peptid nagyobb dózisai növelik a
3. ábra: A parathormon [1-34] szerkezete

19
szívfrekvenciát és a bal kamra kontrakciós erejét is (37). Nyilvánvalóvá
vált, hogy a PTH vasoactiv peptid, mely szignifikáns cardiovascularis
hatásokkal is rendelkezik.
3.3.1 Vascularis hatások
A PTH vascularis hatását jellemző irodalmi adatok szerint a peptid számos
érterületen fejt ki értágító hatást, a különböző artériás szegmentek
válaszkészsége azonban eltérő (38). A peptid gyakorlatilag hatástalannak
bizonyult izolált impedanciaerek (pl. az aorta) relaxálásában, hatása főként
a kisartériákra és precapillaris erekre korlátozódik (39). A vasodilatatiót
nem gátolják sem az alfa- és béta adrenerg, kolinerg-, dopaminerg-, sem a
hisztaminerg-receptor antagonisták, és a hatás közvetítésében az
endogén vasoactiv mediátorok szerepét is kizárták (40,41). Nickols és
munkatársainak sikerült azonosítani az ér simaizomsejteken lévő PTH
receptorokat (39). A hormon vascularis hatása tehát szintén valódi (a PTH
klasszikus célsejtjein lévő receptorokhoz mindenben hasonló) PTH
receptorokon keresztül valósul meg, és PTH receptor antagonistákkal
kompetitíven gátolható. A PTH vascularis simaizomra kifejtett relaxáló
hatása kifejtéséhez nem igényel intakt endotheliumot, és nem függ az
endothel sejtekből felszabaduló EDRF (endothelium-derived relaxing
factor) -tól sem. Hatása tehát az ér simaizomsejtekre kifejtett direkt
hatásnak tulajdonítható (39,40). Mindmáig tisztázatlan azonban a
vasodilatatio sejtszintű hatásmechanizmusa.
Egyes adatok szerint a vasodilatator képesség – legalábbis részben – az
adenil-cikláz stimulálása révén valósul meg. A PTH (1-34) ugyanis mind
simaizom szövetmintában, mind izolált simaizomsejtekben serkenti a
ciklikus AMP (cAMP) képződését (42). Más kutatócsoportok viszont az
intracellularis Ca++ tartalom változásának tulajdonítanak jelentőséget a
peptid indukálta érrelaxáció kialakulásában. A feszültség-függő Ca
csatorna blokkolásának eredményeként létrejövő sejten belüli szabad
kalciumszint csökkenés szintén magyarázhatja a peptid indukálta

20
érrelaxációt (43,44). E kétfajta megközelítés egymással látszatra éppen
ellentétes irányultságú, noha mindkettőre vannak bizonyítékok. Jelen
vizsgálataink kiindulópontjaként a magunk részéről felvetettük, hogy a
hiányzó, a többoldalú hatásmechanizmust integráló elem a peptidnek a
membrán ATP-függő K+ csatornára (KATP) kifejtett effektusa lehet.
3.3.2 KATP csatornák szerepe a coronariakeringés szabályozásában
A KATP csatornák, melyek szerepét a coronaria keringés modulálásában
számos vizsgálati eredmény bizonyítja (45,46,47), univerzális alkotóelemei
a sejtmembránnak, és mivel nem feszültség-, hanem anyagcsere-függő
struktúrák, igen sajátos sejtmembrán elemet képviselnek. Feltételezhetően
a cellularis metabolikus folyamatok és a plazmamembrán elektromos
aktivitása közötti kapocs szerepét töltik be. Közös jellemzőjük, hogy az
intracellularis ATP hiánya esetén nyitva vannak, annak jelenlétében
viszont záródnak. Ezek alapján feltételezhető, hogy a koszorúsereken
hypoxiás hatásra lejátszódó adaptáció folyamatok az ATP szenzitív kálium
csatornákon keresztül valósulnak meg. A mechanizmus – leegyszerűsítve
– a következő:
A koszorúér simaizomsejtek KATP csatornáinak megnyílása – K kiáramlás
révén – hyperpolarizálja a sejtet. A membránpotenciál növekedése a
feszültségfüggő Ca csatornák záródását eredményezi. Ha e csatornákon
keresztül megszűnik a Ca beáramlás, az intracellularis Ca szint csökken,
és ez a coronaria simaizom relaxációjához vezet. A következmény tehát
koszorúér dilatáció, amely véráramlás növekedést eredményez.
A KATP membráncsatorna megnyílásának elindítója az intracellularis ATP
szint csökkenése, és erre a "mutatóra" csatolódik vissza negatív feedback-
ként a koszorúér áramlás növekedése (48).
Vizsgálataink során a PTH által kiváltott coronaria dilatatio változását
vizsgáltuk a KATP csatornát specifikusan blokkoló glibenklamid adása
mellett.

21
3.4 L-arginin
Az L-arginin aminosav a NO prekurzor molekulája. L-argininből és
molekuláris oxigénből a nitrogén-monoxid-szintáz (NOS) enzim hatására
NO képződik (49). A NOS-nak három formája ismert. Az egyik az élettani
körülmények között elsősorban endothelsejtekben, cardiomyocytákban és
endocardialis sejtekben állandóan jelenlevő, kalcium hatására aktiválódó,
konstitucionális forma (eNOS), ami felelős a folyamatos NO termelésért és
kibocsátásért. A másik, a kóros körülmények (pl.: fertőzések, gyulladások,
tumor) hatására expresszálódó, indukálható enzimforma (iNOS), ami első
sorban macrophagokban, lymphocytákban és a vascularis simaizomban
képződik. Az iNOS lényegesen nagyobb mennyiségű NO termelésére
képes és feltehetően közreműködik az immunrendszer védekező
mechanizmusában. Ezek mellett ismert még az idegsejtek NO képzéséért
felelős izoforma, az nNOS (50). A NO lebomlása során oxidálódik és
nitritté alakul, majd oxigenált hemoglobin
hatására gyorsan nitrát lesz belőle (51).
A NO intracellularis jelátvitelben betöltött
szerepét közel 30 évvel ezelőtt írták le
(52), és 1998-ban orvosi és élettani Nobel-
díjjal jutalmazták Robert F. Furchgott,
Louis J. Ignarro és Ferid Murad
gyógyszerkutatókat, akik kimutatták, hogy
a NO az élő szervezet fontos, funkcionális
alkotó eleme, ingerközvetítő szignálja. Az
elmúlt évtizedekben számos vizsgálat
foglalkozott a NO keringés
szabályozásában elfoglalt helyével, majd
az atherosclerosis megelőzésében játszott
szerepével. Kimutatták jelentőségét az
értónus regulációjában; a reaktív
hyperaemia, a myogen vasodilatatio, az 4. ábra: az L-arginin lebontása
és a NO degradációja

22
acetilkolin és a bradykinin hatás mind NO mediálta folyamatok (53).
Emellett bizonyítást nyert, hogy az ischaemiás prekondíció jelenségében
is meghatározó tényező; az első és a második védelmi ablakok (first and
second window of protection) élettani folyamataiban a reperfúziós aritmiák
kialakulásának gátlásával és coronariaáramlás fokozásával (54,55).
Továbbá leírták, hogy a NO-független vasodilatatio (metabolikus vagy
myogen hatás) fokozza a nyíróerőt az erekben, ez eNOS aktivációt
indukál, ennek hatására fokozódik a NO kiválasztás és a továbbiakban ez
tartja fent a vasodilatatiot (56). Emellett a NO fizikai terhelés alatt is fontos
regulátor szerepet játszhat, mert a fokozott nyíróerő és az emelkedett
pulzusnyomás fokozza a NO-szintézist (57). Az L-arginin-NO „tengely”
fontos szerepet játszik az ischaemia-reperfúziós folyamatok során (58),
vazodilatátor hatása mellett gátolja a thrombocyta aggregációt, csökkenti a
leukocyta adhéziót, gátolja az erek simaizom-proliferációját és antioxidáns
hatása is feltételezhető (59,60).
3.4.1 Protektív hatás a reperfúziós károsodás ellen
Az ischaemiás-reperfúziós károsodás ismert jelenség a szívsebészetben.
A myocardium teljesítőképessége a műtétet követő első néhány órában
nem csak a betegek perioperatív állapotát határozza meg, hanem
befolyásolja a hosszú távú kilátásokat is. Az ischaemia és a reperfúzió
során kialakuló myocardialis károsodások és kontraktilitás csökkenés
mellett egyre inkább elfogadott, hogy a szív egészének túlélése a
megfelelő szöveti perfúzió, tehát a kielégítő mikrocirkuláció függvénye.
Több vizsgálat bizonyítja, hogy a microvasculatura védelme mellett
csökkenhet a reperfúziós károsodás (61,62,63). A microvascularis
válaszkészség megőrzése különösen fontos a hosszan tartó hypothermiás
szívműtétek során.
Bár a NO szerepe a cardiovascularis rendszer fiziológiás folyamataiban jól
jellemezhető, patofiziológiás állapotokban betöltött funkciója jórészt
tisztázatlan. Egyrészről fokozza a postischaemiás véráramlást és

23
csökkenti a „no-reflow” fenomént (amikor az újrainduló véráramlás
ellenére sem következik be szöveti reperfúzió), gátolja a leukocyta
adhéziót, emellett (superoxid) szabadgyököket köt meg (64,65).
Másrészről a NO káros hatásairól is beszámoltak ischaemia-reperfúzió
során;pl. negatív inotrop hatás vagy a peroxinitrit képződés gátlása (66).
Összességében úgy tűnik, hogy NO prekurzorok alkalmazása új
cardioprotectiv koncepciót jelenthet az ischaemiás-reperfúziós
károsodások során, melynek lényege, hogy a NO prekurzorok első sorban
a reperfúziós fázisban hatnak, és a (micro)vascularis funkció védelmével
már a reperfúzió során segítik a myocardium megfelelő felépülését.
3.4.2 Protektív hatás agyhalál állapotában
A szívtranszplantációt követő endotheliális diszfunkció gyakran előforduló
jelenség és jelentősen befolyásolja a reperfúziós károsodást, illetve a graft
kilökődését (67). Az endotheliális funkció megőrzése tehát alapvető
fontosságú a graft gyógyulása szempontjából (63). Az ischaemiás-
reperfúziós folyamat során kialakuló endothel károsodás szerepet játszik a
graft ereinek vasculopathiájában, és így nagy mértékben befolyásolja a
szívtranszplantáció sikerét (68,69). A szívtranszplantációt követően
jelentkező endotheliális dysfunctio kialakulásában feltehetően szerepet
játszik a donor állapota, illetve az agyhalál beálltával a donor
szervezetben lezajló cardiovascularis változások (70). A magas
intracranialis nyomással, gyorsan létrehozott agyhalál hyperdinámiás
keringési reakciót okoz, aminek hátterében átmeneti szimpatikus
kisülések, kb. 100-szoros plazma katekolaminszint emelkedés áll (71). Ezt
hemodinamikai instabilitás kíséri jelentős vérnyomáseséssel,
coronariaáramlás csökkenéssel és hormonális változásokkal
(69,70,71,72). A keringési instabilitást feltehetően a katekolaminok
indukálta primer cardialis károsodás okozza, de kiváltó okként felvetődött
a szimpatikus tónus hirtelen elvesztésével kialakuló nagyfokú szisztémás
vascularis rezisztencia csökkenés is (70,73). Továbbá kimutatták, hogy az

24
agyhalál állapota a hemodinamikai megingás mellett csökkenti a
coronariaáramlást és súlyos endotheliális dysfunkciót okoz (69). A direkt
endotheliális szöveti károsodás hátterében katekolamin hatás, illetve a
gyorsan kimerülő hormonális működés állhat (71,72). Agyhalál állapotában
tehát, olyan humoralis változások lépnek fel, melyek során komoly
hemodinamikai instabilitás jelentkezik, és ez a koszorúerek véráramlását,
illetve a coronariák endothelialis funkcióját befolyásolhatja.

25
4 Célkitűzések
Nagy betegszámú vizsgálatban tanulmányoztuk a humán pericardialis
folyadék ANP koncentráció változását:
• összehasonlítottuk a myocardialis ischaemia diagnosztikus jeleit mutató
és nem mutató betegcsoportok plazma, pitvar- és kamraszöveti, illetve
pericardialis folyadékbeli ANP koncentrációs viszonyait.
• vizsgáltuk, milyen hatással van az elégtelen szívműködés súlyossága
az ANP termelődésére és felszabadulására.
• elemeztük a nemek szerepét az ANP cardialis hatásainak
vonatkozásában.
Matematikai modellt készítettünk az interstitialis folyadéktér adenin-
nukleozid összetételének leírására:
• megvizsgáltuk, miként jellemezhetők matematikai modell alapján az
interstitialis tér adenin-nukleozid koncentráció viszonyai.
• vizsgáltuk, meghatározható-e a pericardialis adenin-nukleozid
koncentráció ismeretében az interstitialis térben jelenlevő koncentráció.
Vizsgáltuk parathormon hatására kialakuló koszorúér dilatációt:
• tanulmányoztuk a KATP csatornák szerepét a parathormon vascularis
hatása során.
• jellemeztük a KATP csatornákat specifikusan gátló glibenklamid adására
létrejövő vascularis változásokat.
Tanulmányoztuk a szisztémásan adott NO prekurzor L-arginin protektív
szerepét az ischaemia-reperfúziós károsodások kialakulása során:
• mély hypothermiás ischaemia alatt leírtuk az L-arginin és lebomlási
termékeinek időbeli koncentrációs változásait.

26
• jellemeztük a kapcsolódó cardialis, pulmonalis és vascularis
változásokat.
• vizsgáltuk az agyhalál során jelentkező endothelialis diszfunkciót és
ennek során az L-arginin-NO tengely protektív szerepét a vascularis
funkcióra.

27
5 Módszerek
5.1 Humán vizsgálatok
Vizsgálatainkba 316, a Semmelweis Egyetem Ér-, és Szívsebészeti
Klinikáján szívműtétre kerülő beteget vontunk be. Billentyű-, septum
defektusos, és szívtumoros betegek alkották a nem ischaemiás (VHD;
n=142) kontroll csoportot az ischaemiás szívbetegségben szenvedő (CAD;
n=174) betegekkel szemben. Kiválasztásuk során csak az a szempont
érvényesült, hogy minél szélesebb diagnosztikus skálán mozogjanak. A
betegek átlagéletkora 59±4 év volt. Plazma, pericardialis folyadék,
valamint pitvari és kamrai szövetminták kerültek biokémiai analízisre. A
vérvétel a rutin vizsgálatokkal együtt történt, a szövetminták pedig a rutin
tevékenység során (vénás kanül behelyezése, billentyű eltávolítása)
amúgy is eltávolításra kerülnek, így az elvégzett vizsgálatok a betegek
számára semmilyen többletterhelést nem jelentettek. Mintavételre minden
esetben a betegek felvilágosítása és belegyezése után került sor. A
betegeket csoportosítottuk a New York Heart Association (NYHA) ajánlása
szerint; így a NYHA I stádiumba 4, NYHA II stádiumba 178, a NYHA III
stádiumba 108 és NYHA IV stádiumba 13 beteg került. (Tizenhárom
esetben nem állt rendelkezésre információ a beteg NYHA statusáról.) 111
koszorúér beteg esett át korábban myocardialis infarctuson, de az
esemény minden esetben a műtétnél legalább egy hónappal régebben
történt. A billentyű betegeknél koszorúér betegségre sem klinikai tünetek,
sem EKG jel, illetve coronarographiás elváltozás nem utalt.
A műtét előtt minden beteg részesült a kórállapotának megfelelő
gyógyszeres kezelésben (ACE-gátlók, diuretikumok, digitalis, β-blokkolók,
nitrátok). Az altatás minden betegnél azonos módon, midazolam, fentanyl
és ketalar iv. adásával történt. Az anaesthesia fenntartására diazepam,
fentanyl és droperidol szolgált, az izomrelaxációt pancuronium biztosította.

28
A mellkas feltárása (median sternotomia), majd a pericardiumzsák
megnyitása után 4-8 mL pericardialis folyadékot gyűjtöttünk egy hajlékony
kanül segítségével steril fecskendőbe. Kizárólag vérmentes
folyadékmintákat használtunk fel biokémiai meghatározásra. A műtétek
során – a pericardialis folyadék vételével párhuzamosan – szisztémás
vénás vért is gyűjtöttünk, heparint és dipyridamol+EHNA-t tartalmazó
fecskendőbe. Mind a pericardialis folyadékot, mind a vérmintákat azonnal
centrifugáltuk, és -20 ºC-on tároltuk a biokémiai mérésekig. A pitvari
szövetminták vételére a jobb pitvar kanülálásakor került sor. A kamrai
szövetmintákat a billentyűműtétek során eltávolított kóros mitralis vitorla
papillaris izmából nyertük. A rutin szívsebészeti eljárásoknak
köszönhetően a pitvari és a kamrai szöveti mintavételi helyek betegenként
közel azonos helyzetűek voltak. A szövetmintákat ezt követően folyékony
nitrogénben -80 ºC-on tároltuk a további vizsgálatokig.
5.2 Experimentális modellek
Vizsgálatainkat 37, pentobarbitállal altatott, 25-38 kg közötti, nyitott
mellkasú kutyán végeztük. Valamennyi akut kísérletet a laboratóriumi
állatok tartására vonatkozó nemzetközi szabályoknak megfelelően
végeztük. Na-pentobarbitallal történő altatást követően az állatokat
endotrachealisan intubáltuk, majd gépi lélegeztetést folytattunk. A
szisztémás vérnyomás monitorozásához az arteria femoralist preparáltuk
és kanüláltuk. Rendszeres vérgáz elemzéssel követtük a pulmonalis és
metabolikus státus változását.
5.2.1 Parathormon coronaria hatásának vizsgálata
A kísérleti állatokon haránt thoracotomiát végeztünk az ötödik
bordaközben és a szívet a megnyitott pericardiumban függesztettük fel. A
koszorúér áramlást (CBF) a bal elülső leszálló coronarián mértük. Az
áramlásmérő fejtől distalisan az eret egy vékony tűvel (23G) pungáltuk a

29
szerek intracoronariás adásához, mely manőver a basalis áramlási
paramétereket nem befolyásolta. Vizsgálatainkat két kísérletsorozatban
végeztük, a kísérleteket négy fázisra osztottuk. Valamennyi fázisban
rendre 0.15, 0.44 és 1.33 nmol PTH dózisokat adtunk 1 mL-es
intracoronariás bolusok formájában. Az egyik kísérletsorozatban (n=7), az
első, kontroll fázis után glibenklamidot adtunk fázisonként növekvő
dózisban, 0.1, 0.3, és 1.0 μmol/perces intracoronariás infúzióban. Mind
kontroll fázisban, mind a glibenklamiddal előidézett KATP csatorna blokád
alatt PTH dózis-hatásgörbéket vettünk fel a PTH fenti dózisaival.
A kísérletek jelentős időigénye miatt a PTH esetleges hatásvesztésével is
számolni kellett, ezért egy időkontroll kísérlet sorozatot (n=6) is végeztünk,
melyben glibenklamid helyett fiziológiás sóoldatot infundáltunk. Az első
kísérletsorozathoz hasonlóan itt is PTH dózis-hatásgörbéket vettünk fel
mind a négy fázisban. Az egyes beavatkozások előtt megvártuk a PTH
bolus hatások lecsengését, azaz valamennyi cardiovascularis paraméter
visszatérését a kiindulási szintre.
A kísérletek során szintetikus, szarvasmarha parathyreoid hormon
fragmentet PTH [1-34] (Sigma, USA) alkalmaztunk. A koszorúér relaxáció
mértékének jellemzésére az ér rezisztencia reciprokát, a vascularis
konduktancia paraméterét alkalmaztuk. Értékét a coronariaáramlás és az
artériás nyomás hányadosából képeztük az egyedi adatok alapján. Az így
nyert coronaria konduktancia görbék alatti területet jelöltük coronaria
konduktancia többletnek. A koronária konduktancia többlet az a nyomástól
független virtuális plusz áramlási volumen, amely a vasodilatator válasz
során keletkezik. Értékét a konduktancia görbék integrálásával
számítottuk.
5.2.2 L-arginin hatása hypothermiás ischaemiát követő reperfúzió során
Kísérleteinkben a motoros szívműtétek során létrehozott hypothermiát
modelleztük, ezért minden állaton felfüggesztettük a szívműködést, egy
órán át fenntartottuk az extracorporalis keringést, majd elhagytuk a

30
szívmotort. A beavatkozásokhoz anterolateralis thoracotomiát és
pericardiotomiát követően izoláltuk a nagyereket, majd az artériás kanült a
bal arteria subclaviába, a vénás kanült a jobb pitvarba helyeztük. Az
extracorporalis keringés emellett állt még egy hőcserélőből, egy vénás
reservoir-ból, egy roller pumpából és egy membrán oxigenátorból. A
cardiopulmonalis bypass (CPB) bevezetése után a testhőmérsékletet
28°C-ra hűtöttük. Az aorta lefogása után a szívet 25 mL/kg HTK oldattal
állítottuk meg, melynek összetétele mmolban: 15 NaCl, 9 KCl, 4 MgCl2, 6
H2O, 18 histidin hydroclorid monohydrat, 180 histidin, 2 tryptophan, 30
mannitol, 0.0015 CaCl2, 1 kalium-hydrogen-2-oxopentandioat és H2O. A
szívmegállás alatt a szívmotor áramlása 100 ml/kg/min volt, így a
perfúziós nyomás nem esett 35-40 Hgmm alá a CPB során. Húsz perccel
az aorta felengedése előtt megkezdtük a felmelegítést. A szívmegállás 60.
percében az aortát felengedtük és a keringető rendszer normothermiás
vérével reperfundáltuk a szívet. Szükség esetén DC cardioversiót
végeztünk a szív újraindításához. Hat állatnak az aorta felengedésekor – a
reperfúzió kezdetekor - 40 mg/kg L-arginint (Merck, Németország) adtunk
intravénás bolusban, majd további 20 percig intravénás infúzióban 3
mg/kg fenntartó dózisban. Másik hat állat kontrollként fiziológiás sóoldatot
kapott, ugyanezen protokoll szerint. Minden állat inotrop támogatási igény
nélkül leszoktatható volt a szívmotorról az aorta felengedését követő 20
percen belül. Kísérleti protokollunk szerint minden állat 60 percig volt
extracorporalis keringésen, ebből 40 percen át állt a szív, az aorta
felengedését követően pedig 40 perc reperfúziós szak következett. A bal
és jobb kamrai systolés és diasztolés nyomásokat valamint volumeneket
egy 6F Millar konduktancia katéterrel mértük, amit a bal kamra csúcsán,
illetve az arteria pulmonalis felől vezettünk fel. Vena cava occlusiókat
végezve nyomás-térfogat görbe-sorozatokat készítettünk. A „preload
recruitable stroke work” (PRSW) számításával a kontraktilitás egy
preloadtól független mérőszámát kalkuláltuk. A koszorúér áramlást a bal
elülső leszálló coronarián mértük. Az coronariák endothelium-dependens

31
dilatatioját acetilkolin (ACH, 10-7 M), endothelium-independens dilatatioját
nátrium-nitroprussid (SNP, 10-4 M) intracoronariás adásával vizsgáltuk.
5.2.3 L-arginin hatása agyhalál állapotában
Kísérleteinkben az agyhalál állapotát mesterséges intracranialis
agynyomás-fokozódás létrehozásával értük el. A koponyát átfúrva egy
Foley-katétert helyeztünk a subduralis térbe, majd a katéter ballonját 25
mL fiziológiás sóoldattal gyorsan felfújtuk. Az agyhalál állapotát a cornea
és a pupilla reflexek megszűntével, illetve post mortem patológiai
vizsgálatokkal igazoltuk. Hat állat 40 mg/kg L-arginin kezelést kapott az
agyhalál beálltát követő 60. percben, amit a kísérletek végéig (az agyhalál
indukcióját követő 180 perc) folytattunk. Hat másik állat hasonló protokoll
szerint fiziológiás sóoldatot kapott, ezek szolgáltak kontrollként.
Lateralis thoracotomiát követően és a pericardium megnyitása után a
nagyereket izoláltuk. Perivascularis elektromágneses mérőfejjel mértük a
felszálló aorta véráramlását. A bal kamrai szisztolés és diasztolés
nyomásokat valamint volumeneket egy 6F Millar konduktancia katéterrel
mértük, amit a bal kamra csúcsán vezettünk a szívbe. Emellett
monitoroztuk az aortanyomást és a jobb pitvari nyomást. A kamrai
volumenváltozást folyamatosan regisztráltuk és elemeztük. Vena cava
occlusiókat végezve nyomás-térfogat görbe-sorozatokat készítettünk.
Ezek alapján számítottuk a végszisztolés nyomás-térfogat arányszámot
(end-systolic pressure-volume relationship, ESPVR) és a „preload
recruitable stroke work”-öt, a kontraktilitás egy preloadtól független
mérőszámát. A koszorúér áramlást a bal elülső leszálló coronarián mértük.
Az coronariák endothelium-dependens dilatatióját acetilkolin (ACH, 10-7
M), endothelium-independens dilatatioját nátrium-nitroprossid (SNP, 10-4
M) intracoronariás adásával vizsgáltuk.

32
5.3 Biokémiai vizsgálatok
5.3.1 ANP koncentráció meghatározása
Az ANP extrakcióját és RIA vizsgálatát leíró módszer röviden a következő:
a pericardialis folyadék- és plazma minták tisztítása és – szükség esetén –
koncentrálása SepPak C18-as kromatográfiás oszlopok használatával
történt. A minták vegyhatását 10%-os trifluoroacetáttal pH 4-ig
savasítottuk, és az előzőleg metanollal és trietanolamin-acetát pufferral
(TEA, 20 mM, pH4) aktivált SepPak C18-as cartridge-okra (Waters,
Milford, MA, USA) vittük fel. Az extraktumokat liofilizáltuk és RIA pufferben
újra oldottuk. Az így kapott oldatokkal két párhuzamos radioimmuno-assay
mérés történt. RIA-hoz a mintákat specifikus nyúl antiszérummal (1:25000
végleges hígítás) inkubáltuk. Inkubálás után az immunkomplexeket birka
antiszérummal precipitáltuk és centrifugáltuk. A standard görbe
felvételéhez szintetikus humán ANP-t (Sigma Chemical Co., St. Louis,
MO, USA) használtunk. Az ANP tracer [125I]-el jelzett humán ANP[99-126]
volt (Amersham, UK). Az assay érzékenysége 0.8 pg/kémcső-nek felelt
meg. A standard görbe 50%-os értéke 20 pg/kémcső-nél volt; a változás
intra- és interassay koefficiensei: <10% és <15%.
A szöveti mintákat 4 M-os guanidin tiocyanat-tal (1:10 w/w)
homogenizáltuk. Ezt követően a pitvari minták 1:10000, a kamrai minták
1:100 arányban lettek felhigítva, és RIA-val mérve.
A HPLC analízishez a liofilizált plazma és pericardialis folyadék mintákat
visszaoldottuk 40%-os acetonitrilt és 0.1%-os trifluoroacetátot tartalmazó
vizes oldatba, majd azt egy 7.8x300 mm-es ProteinPak 125 gél-filtrációs
HPLC oszlopra (Waters, Milford, MA, USA) vittük fel, és ugyanezzel az
oldattal oldottuk le. A frakciókat 30 másodpercenként gyűjtöttük,
liofilizáltuk, RIA-pufferben oldottuk, majd RIA-val mértük.

33
5.3.2 L-arginin szint mérése
A plazma L-arginin szintjének meghatározása során a mintákat 0.5 M-os
borsavval oldottuk és kation-cserélő cartridge-on (LiChrolut SCX 200 mg,
Merck, Germany) futtattuk. Internal standarnak 250 μM-os l-
monometilarginin-t (l-NMA) használtunk. A minta futtatása után a
cartridge-ot hatszor 1 mL desztillált vízzel, majd háromszor 1mL 12.5 mM
Na-acetát pufferrel (pH 4.5) átmostuk. A megkötött aminosavakat 1 mL
250 mM Na-acetáttal (pH 8.5) eluáltuk a cartridge-ról. A mintából 160 μL-t
szobahőmérsékleten 2 percig összekevertünk 40 μL o-phthal-dialdehid
(OPA; Sigma, Germany) reagenssel, majd kiegészítettük 1 μL
mercaptoetanollal és 5 percig szobahőmérsékleten inkubáltuk. A
HPLC/fluorescenciás analízis során Merck LiChrospher 100 RP-18
oszlopot használtunk. Az izokratikus eluáció 10 mM KH2PO4 (pH
5.85)/acetonitril/metanol/tetrahidrofuran [80:9.5:9.5:1 (v/v/v/v)] oldattal 1
mL/min áramlás mellett történt. A minták L-arginin szintjének
meghatározása a HPLC görbe alatti területek nagysága alapján, eredeti L-
arginin standardhoz (100 μM) viszonyítva történt.
5.3.3 Nitrit és nitrát szint meghatározások
A plazma nitrit és nitrát koncentrációit ELISA módszerrel határoztuk meg.
A nitrát és nitrit elemek együttes koncentrációját (nitrát&nitrit) a nitrát
reduktáz enzim által katalizált nitrát-nitrit átalakulást követően mértük. A
vizsgált minták 540 nm-en mért optikai denzitását (OD540) standard
nitrát&nitrit és nitrit oldatok OD540-hoz hasonlítottuk. A nitrát
koncentrációkat a nitrát&nitrit és nitrit koncentrációk különbsége alapján
határoztuk meg.

34
5.4 Kompartmentalizációs matematikai modell
A pericardialis folyadéktér dinamikáját jellemző modell az
anyagmegmaradás törvényére épül. A modell leírását egyszerűsítettük, és
a fiziológiáshoz hasonló állapotot közelítettük azzal, hogy a
kapillárisokban egyenletes áramlással számoltunk, és elhanyagoltuk az
egyes szövetterületek közötti radiális koncentráció gradienst. A szövetek
között lineáris molekula transzporttal számoltunk, figyelembe véve az
anyag-metabolizmust, és a regionális áramláskülönbségeket.
A modell a perikapilláris szövetet öt struktúrára osztja: a kapilláris lumene,
endothel sejtek, myocardialis parenchymasejtek, interstitialis folyadék és
epicardialis transzszudátum. Az ezek közötti kapcsolatokat és az egyes
kompartmentekben történő metabolikus folyamatokat mutatja az 5. ábra.
A modell felépítéséhez és a paraméterek beállításához több részkísérlet
elvégzésére volt szükség, amik csak a modell egy-egy paraméteréről,
illetve részfolyamatáról adnak információt. A modell készítésekor ezeket a
korábbi méréseket részben irodalom alapján, részben saját
Parenchima-sejt Endothel-sejt
Endothel-sejt
ISF Kapilláris
FpCpc
Gpc
V’pc
Dpc
PSeca PSecl
PSpc
Cisf
Gisf
V’isf
Disf
PStransz
Ctransz
Gtransz
PSg
Cec
Gec
V’ec
Dec
Cp
Gp
V’pDp
Parenchima-sejt Endothel-sejt
Endothel-sejt
ISF Kapilláris
FpCpc
Gpc
V’pc
Dpc
PSeca PSecl
PSpc
Cisf
Gisf
V’isf
Disf
PStransz
Ctransz
Gtransz
PSg
Cec
Gec
V’ec
Dec
Cp
Gp
V’pDp
5. ábra: A myocardialis szövet alkotóelemei és az ezekben lejátszódó folyamatok

35
eredményeinkből gyűjtöttük össze (29). Modellünk felállításakor az adenin-
nukleozid rendszer alkotóinak metabolizmusát és kinetikáját vettük
figyelembe.
Az 5. ábrán látható paraméterek rövid jellemzése:
PSg A kapillárisok permeábilis területén átjutó anyag mennyisége a
plazma és az interstitium között (mL*g-1*min-1)
PSecl Az endothelsejtek lumen felőli membránjának permeábilis
területén átjutó anyag mennyisége a plazma és az endothelsejt
cytosol között (mL*g-1*min-1)
PSeca Az endothelsejtek lumennel ellentétes oldali (abluminális)
membránjának permeábilis területén átjutó anyag mennyisége az
interstitialis folyadék és az endothelsejt cytosol között
(mL*g-1*min-1)
PSpc A parenchymasejtek membránjának permeábilis területén átjutó
anyag mennyisége az interstitialis folyadék és a parenchymasejt
cytosolja között (mL*g-1*min-1)
PStransz A transzszudációs folyadék és az interstitialis folyadék közötti
molekulaáramlás nagysága (mL*g-1*min-1)
V’isf Az eloszlás során kitöltött tér az interstitiumban (mL/g), az adott
terület-plazma közötti áramlás partíciós koefficiense szorozva az
anatómiai térfogattal (V))
V’ec Az eloszlás során kitöltött tér az endothelsejtekben (mL/g,
anatómiai adatok alapján 0.02 mL/g értékben megállapítva)
V’pc Az eloszlás során kitöltött tér a parenchymasejtekben (mL/g,
kevéssé érzékeny paraméter 0.5-0.7 mg/l között)
Gp Az intravasalis metabolizmus során keletkezett anyagok
clearance-e (mL*g-1*min-1)
Gisf Az interstitialis metabolizmus során keletkezett anyagok
clearance-e (mL*g-1*min-1)
Gec Az intracellularis metabolizmus során keletkezett anyagok
clearance-e az endothelsejtekben (mL*g-1*min-1)

36
Gpc Az intracellularis metabolizmus során keletkezett anyagok
clearance-e a parenchymasejtekben (mL*g-1*min-1)
Gtransz A transzszudációs térben folyó metabolizmus során keletkezett
anyagok clearance-e (mL*g-1*min-1)
Fp áramlás per szövettömeg (mL/g/sec)
C koncentráció (mol/cm3)
D tengely irányú diffúzió, illetve diszperziós koefficiens (cm2/sec)
L kapilláris hossza (cm);(szívben 0.05-0.1 cm)
t idő (sec)
x távolság a kapillárisban (cm)
V egy terület térfogata (mL)
V’ megoszlási térfogat egy adott területen (mL)
α plazma áramlás sebessége (cm/sec);=Fp*L
5.5 Statisztikai módszerek
Parametrikus adatok elemzése során az eredményeket átlag ± átlag
standard hiba (átlag±SEM) formában adtuk meg, az adatcsoportok közötti
összehasonlításokra (egy és kétmintás) T-próbákat és Pearson-analízist
alkalmaztunk. Nem parametrikus adatok esetén az eredményeket medián
és percentilisek formájában adtuk meg, emellett Wilcoxon-próbát, Mann-
Whitney tesztet és Spearman-analízist alkalmaztunk az egyes csoportok
összehasonlítására. Az ismétlődő mérések során történt változások
leírására varianciaanlízist (ANOVA) alkalmaztunk. Szignifikancia szintnek
az általánosan elfogadott p<0.05 értéket választottuk.

37
6 Eredmények
6.1 Pericardialis és szöveti ANP szintek elemzése
A 2. táblázatban láthatók a betegcsoportonként és vizsgált mintánként
csoportosított eredményeink. A szívműtétre került betegek pericardialis
folyadékában mért ANP koncentrációk egy nagyságrenddel (kb. 10-12-
szer) magasabbnak mutatkoztak, mint a megfelelő vénás plazmában
mértek (p<0.001). Emellett a pitvari myocardium ANP koncentrációja 130-
380-szoros különbséget mutatott a kamrai szövetek ANP
koncentrációjához képest. A pitvari és kamrai ANP szintek között
szignifikáns korreláció volt kimutatható (r=0.317; p<0.05). Az ischaemiás
szívbetegek pericardialis folyadék és plazma ANP koncentrációi
alacsonyabbnak bizonyultak a nem-ischaemiás szívbetegekben
mérhetőknél (p<0.05), és ez utóbbi betegcsoportban tudtunk csak
kimutatni szignifikáns korrelációt a plazma és a pericardialis folyadék ANP
koncentrációi között.
Vénás plazma (pg/mL)
Pericardialis folyadék (pg/mL)
Korreláció Pitvari szövet
(ng/mg)
Kamrai szövet
(ng/mg) Korreláció
Billentyű-betegek
(VHD)
28.4
(11.6-61.5)
258.3 * (198.9-342.5)
r = -0.482 • 151.7 #
(78.4-447.6)
0.4
(0.2-1.6) r = 0.261
Koszorúér betegek
(CAD)
19.8 †
(9.4-27.9)
208.1 * †
(153.8-318.9) r = -0.07
129.6 #
(66.5-455.0)
1.0
(0.1-1.8) r = 0.8
2. táblázat ANP koncentrációk összehasonlítása betegcsoportok és vizsgálati minták szerint (medián és 25-75 percentilisek formájában) * p<0.001 vs. megfelelő vénás plazma • p<0.05 # p<0.001 vs. megfelelő kamrai szövet † p<0.05 vs. billentyűbetegek

38
Mindemellett azonban megfigyelhető (6. ábra), hogy az egyedi mérési
eredmények számottevő része - úgy a pericardialis folyadék, mint a
plazma ANP szintjeit figyelembe véve - mindkét betegcsoportban azonos
tartományba esett.
6. ábra A vénás plazma (VP) és a pericardialis folyadék (pericardialis folyadék) ANP koncentrációi a két betegcsoportban. Minden kör egy beteget jelöl. A vízszintes vonal a mediánt mutatja. Az ANP szintek logaritmikus skálán vannak feltüntetve
Az ANP koncentrációkat a betegek NYHA stádiumának függvényében
elemezve (7. ábra) erősen szignifikáns korrelációt találtunk a szív
funkcionális stádiumának romlása és a plazma ANP szintek emelkedése
között (r=0.401; p<0.001). Hasonló, de kevésbé szignifikáns összefüggés
volt kimutatható a NYHA stádiumok és a pitvari (r=0.169; p<0.05), illetve a
kamrai (r=0.313; p<0.05) szöveti ANP koncentrációk között.
Nemek szerint csoportosítva a betegeket, szignifikánsan (p<0.05)
magasabb plazma ANP koncentrációkat mértünk nőkben (25.2 (14.8-60.5)
pg/mL), mint férfiakban (19.8 (9.0-28.4) pg/mL). Ehhez hasonló, de
nagyobb mértékű különbség (p<0.001) mutatkozott a pitvari szöveti ANP
koncentrációkban a nők (414.6 (119.7-734.4)) ng/mg és a férfiak (105.4
(65.3-204.2) ng/mg) között. Nemek szerint csoportosítva az ischaemiás és
nem-ischaemiás betegeket, nők esetében szignifikáns (p<0.05)
1
10
100
1000
1
10
100
1000
AN
P (p
g/m
L)
VP
Billentyű betegek
PFVP
Billentyű betegek
PF VP
Koszorúér betegek
PFVP
Koszorúér betegek
PF
P<0.001 P<0.001
AN
P (p
g/m
L)

39
különbséget találtunk a pericardialis ANP szintekben, azonban férfiaknál
hasonló különbség nem volt kimutatható (3. táblázat).
Nők Férfiak CAD VHD CAD VHD
Vénás plazma (pg/mL)
24.8
(12.8-58.2)
35.9
(19.6-65.1) †
17.3
(9.0-25.8)
28.4
(9.5-54.1)
Pericardialis folyadék (pg/mL)
171.2
(104.0-196.6)
274.2
(226.0-377.5) •
234.5
(164.7-346.6) •
258.3
(191.7-312.7) •
Pitvari szövet (ng/mg)
609.8
(150.2-853.3) * #
240.1
(111.2-706.6) * #
98.4
(49.0-223.2)
113.7
(72.4-198.8)
Kamrai szövet (ng/mg)
1.1
(0.1-1.5)
0.5
(0.2-1.5)
1.0
(0.1-1.9)
0.4
(0.1-12.9)
3. táblázat Ischaemiás és nem-ischaemiás betegek nemek szerinti csoportosítása. † p<0.05 vs. a megfelelő férfi CAD; • p<0.05 vs. a megfelelő női CAD;
* p<0.001 vs. a megfelelő férfi CAD; # p<0.001 vs. a megfelelő VHD
Azoknál a betegeknél, akik átestek myocardialis infarctuson,
szignifikánsan (p<0.05) alacsonyabb pericardialis ANP szintek voltak
mérhetők (202.6 (146.4-293.0) pg/mL), mint azoknál, akik nem estek át
infarctuson (257.1 (177.5-338.0) pg/mL). Ugyanezt az összehasonlítást
elvégezve csak a koszorúér betegcsoporton, nem kaptunk szignifikáns
különbséget az infarctus megtörténte alapján. Hasonlóan nem volt
szignifikáns különbség a plazma és a szöveti ANP koncentrációk esetében
sem.

40
7. ábra NYHA funkcionális stádiumok és ANP koncentrációk összefüggése mintavételi helyek alapján (vénás plazmában, pericardialis folyadékban, pitvari és kamrai szövetekben). (Az ábrán a középső vízszintes vonal a mediánt, a dobozok a 25-75 percentiliseket, a vékony vonalak a szélső értékeket jelölik.)
A minták HPLC analízise
során tisztán elkülöníthető volt
a humán ANP [99-126]
csúcsa, egy másik, a proANP-t
reprezentáló – az össz-
mennyiség 15%-át meg nem
haladó – immunoreaktivitás
mellett (8. ábra).
p<0.05
p<0.05
p<0.001
p<0.05
p<0.05
r=-0.051p=N.S.
r=0.313p<0.05
NYHA IVNYHA IIINYHA IINYHA I
Vén
ás p
lazm
aA
NP
(pg/
ml)
300
200
100
0
NYHA IVNYHA IIINYHA IINYHA I
Per
icar
dial
isfo
lyad
ékA
NP
(pg/
ml)
600
500
400
300
200
100
0NYHA IVNYHA IIINYHA II
Kam
rai s
zöve
tiA
NP
(ng/
mg)
4
3
2
1
0
NYHA IVNYHA IIINYHA II
Pitv
ari s
zöve
tiA
NP
(ng/
mg)
2000
1500
1000
500
0
r=0.401p<0.001
r=0.169p<0.05
p<0.05
p<0.05
p<0.001
p<0.05
p<0.05
r=-0.051p=N.S.
r=0.313p<0.05
NYHA IVNYHA IIINYHA IINYHA I
Vén
ás p
lazm
aA
NP
(pg/
ml)
300
200
100
0
NYHA IVNYHA IIINYHA IINYHA I
Per
icar
dial
isfo
lyad
ékA
NP
(pg/
ml)
600
500
400
300
200
100
0NYHA IVNYHA IIINYHA II
Kam
rai s
zöve
tiA
NP
(ng/
mg)
4
3
2
1
0
NYHA IVNYHA IIINYHA II
Pitv
ari s
zöve
tiA
NP
(ng/
mg)
2000
1500
1000
500
0
r=0.401p<0.001
r=0.169p<0.05
80
60
40
20
ir-AN
P (p
g/fr
actio
n)
10 12 14 16 18 20 22 24Fraction
V0 VtANPp-ANP80
60
40
20
ir-AN
P (p
g/fr
actio
n)
10 12 14 16 18 20 22 24Fraction
V0 VtANPp-ANP
8. ábra Egy ANP minta gél filtrációs HPLC profilja. p-ANP=proANP, ANP= ANP [99-126]

41
6.2 Az interstitialis és pericardialis folyadéktér dinamikája adenin nukleozid koncentrációk alapján
Matematikai modellünk korábban ismertetett paraméterei alapján
meghatározható az egyes szövetalkotók ADO anyagcseréje és a
kompartmentek közötti molekulatranszport.
A plazma ADO koncentráció változása az időben:
( ) ( ) 2p
2
ppp
pecp
p
eclisfp
p
gp
p
pp
xC
DCVG
CCV
PSCC
VPS
xC
VLF
tC
Δ
Δ+−−−−−
Δ
Δ⋅−=
Δ
Δ
ahol az egyenlet jobb oldalán az első tag a plazma ADO koncentráció
változását írja le a kapilláris hosszában. A második tag a plazma és az
interstitialis folyadék közötti, a harmadik tag a plazma és az endothelsejt
közötti ADO diffúziót jelöli (a plazma szemszögéből). A negyedik tag a
plazma saját ADO metabolizmusára vonatkozik. Az utolsó tag a
plazmában levő anyagok tengely irányú diszperzióját jelöli.
Az endothelsejtben az ADO koncentráció változása az időben:
( ) ( ) 2ec
2
ececec
ecisfec
ec
ecapec
ec
eclec
xC
DC'V
GCC
'VPS
CC'V
PSt
CΔΔ
+−−−−−=ΔΔ
ahol az egyenlet jobb oldalán az első tag az endothelsejt és a plazma
közötti, a második tag az endothelsejt és az interstitialis folyadék közötti
ADO diffúziót jelöli (az endothelsejt szemszögéből). A harmadik tag az
endothelsejt saját ADO metabolizmusát jelöli. A negyedik tag az ADO
diszperzióját jelöli a sejten belül.
Az interstitialis folyadékban az ADO koncentráció változása az időben:
( ) ( ) ( ) 2isf
2
isfisfisf
isfpcisf
isf
pcecisf
isf
ecapisf
isf
gisf
xC
DC'V
GCC
'VPS
CC'V
PSCC
'VPS
tC
Δ
Δ+−−−−−−−=
Δ
Δ
ahol az egyenlet jobb oldalán az első tag az interstitialis folyadék és a
plazma közötti, a második tag az interstitialis folyadék és az endothelsejt
közötti, a harmadik tag pedig az interstitialis folyadék és a parenchymasejt
közötti ADO diffúziót jelöli (az interstitium szemszögéből). A negyedik tag

42
az interstitialis folyadék saját metabolizmusát jelöli. Az ötödik tag ismét a
diszperziót jellemzi, az interstitiumon belül.
A parenchymasejtekben az ADO koncentráció változása az időben:
( ) 2pc
2
pcpcpc
pcisfpc
pc
pcpc
xC
DC'V
GCC
'VPS
tC
Δ
Δ+−−−=
Δ
Δ
ahol az egyenlet jobb oldalán az első tag a parenchymasejt és az
interstitialis folyadék közötti ADO diffúziót jelöli (a parenchymasejt
szemszögéből). A második tag a parenchymasejt intracellularis
metabolizmusára utal, míg az utolsó tag a sejten belüli diszperziót jelöli.
Az epicardialis transzszudátumban az ADO koncentráció változása az
időben:
ahol az egyenlet jobb oldalán az első tag a transzszudátum és az
interstitialis folyadék közötti ADO diffúziót jelöli (az epicardialis
transzszudátum szemszögéből). A második tag a transzszudátum ADO
metabolizmusára utal. A transzszudátumból ADO csere csak az
interstitiumba történik. Az interstitiumból viszont az ADO az endothel
sejtekbe, a cardiomyocytákba, kapillárisokba, illetve vissza a
transzszudátumba is juthat.
A megoldás egyszerűsítése végett a további két kitételt tettük: a
modellben a kapillárisokban áramló folyadék sebességprofilját nem
paraboloid alakúnak, hanem minden pontjában azonos sebességűnek
vettük. Ezzel elhanyagolhattuk a tengely irányú diffúzió, illetve diszperzió
mennyiségét (az egyenletekben D); így D értékét nullának vettük. Emellett
tovább egyszerűsítettük a megoldást, azzal, hogy a kapilláris és
környezete között egyensúlyi állapotot feltételeztünk (pl. konstans infúzió
adása mellett). Így az egyenletek bal oldalán nulla állt.
Az egyenleteinkben alkalmazott membránpermeabilitást (PS) és a
metabolikus termékek clearance-ét leíró (G) értékek kísérletes úton
( ) ranszransz
ranszisfransz
transz
transztransz CVGCC
VPS
tC
''−−−=
ΔΔ

43
meghatározhatók, illetve az irodalomban megtalálhatók. A szöveti
áramlásra (FP) szintén található irodalmi adat.
Egységnyi kapillárishosszra tehát, a kapott öt egyenletből az öt változó
nagysága meghatározható, és mivel mind a vérplazma ADO
koncentrációja, mind a pericardialis folyadékban mért ADO koncentráció
klinikailag mérhető paraméter, adatainkból az interstitialis folyadék
adenozin koncentrációját egy adott esetben megbecsülhetjük.
6.3 Parathormon hatás a coronariákon
Az intracoronariásan adott PTH [1-34] önmagában jelentősen,
hatásmaximumán (0.44 nmol dózisban) mintegy 2.5-szeresére képes
fokozni a coronariaáramlást a kiindulási értékhez képest. A bolusban adott
peptid hatása rendkívül elhúzódó.
9. ábra Coronaria áramlás és vérnyomás változás egyidejű ábrázolása PTH adása során, egyedi regisztrátumon. A PTH növekvő dózisainak intracoronariás (ic.) beadását nyilak jelzik. Fent: kontroll fázis. Lent: PTH hatások 1.0 μmol/min dózisú glibenklamid háttérinfúziója mellett.
Parathormon i.c. (nmol)
Glibenklamid infúzió 1.0 μmol/min i.c.
5 min
Coronariaáramlás
(mL/min)
200150
100
50
0
Vérnyomás(mmHg)
150100
50
200150
100
50
0150100
50
0.15 0.44 1.33
Coronariaáramlás
(mL/min)
Vérnyomás(mmHg)
Parathormon i.c. (nmol)
Glibenklamid infúzió 1.0 μmol/min i.c.
5 min
Coronariaáramlás
(mL/min)
200150
100
50
0
Vérnyomás(mmHg)
150100
50
200150
100
50
0150100
50
0.15 0.44 1.33
Coronariaáramlás
(mL/min)
Vérnyomás(mmHg)

44
A peptid által kiváltott típusos koszorúér dilatációt eredeti regisztrátum
szemlélteti a 9. ábrán. Megfigyelhető, hogy a coronaria effektus különösen
a PTH mérsékeltebb - közepes - dózisainál volt szembetűnő, ahol a peptid
szisztémás hypotensiv hatása még nem fékezte az áramlásnövekedést.
Ez utóbbi jelenséget jól szemlélteti a coronariaáramlási görbék
alakváltozása, "letörése". Az áramlási maximumok tekintetében ezért
gyenge a dózisfüggés. Az ábra alsó blokkja mutatja vizsgálataink fő
eredményét. Azt tapasztaltuk, hogy a glibenklamid jelentős mértékben
csökkentette, a bemutatott regisztrátumon csaknem teljesen kiküszöbölte
a PTH áramlásfokozó hatását. A vérnyomás válaszok ugyanakkor csak
kevéssé változtak.
10. ábra Változások a coronariaáramlásban, a vérnyomásban és a szívfrekvenciában PTH 0.15, 0.44 és 1.33 nmol dózisainak intracoronariás (ic.) adása után a kísérlet négy fázisában: kontroll fázisban és glibenklamid 0.1, 0.3 illetve 1.0 μmol/min dózisának ic. adása mellett.
A jelenséget jól jellemzi az összes kísérlet adatainak összegzése (10.
ábra). A hatásmaximumokat értékelve jól megfigyelhető, hogy a növekvő
dózisú PTH valamennyi fázisban fokozta a coronariaáramlást,
csökkentette a szisztémás vérnyomást és növelte a szívfrekvenciát. Az
Glibenklamid i.c. (µmol/min)Kontroll
1.00.30.1
Parathormon ic. (nmol)
0.15 1.33
*
**
***
***
0
50
100
150
200
250p < 0.05 *
0.15 1.33 0.15 1.33 0.15 1.33
200
- 40
0- 20
Coronariaáramlás
(Δ%)
Szívfrekvencia
Artériásvérnyomás
(Δ%)
(Δ%)
Glibenklamid i.c. (µmol/min)Kontroll
1.00.30.1
Parathormon ic. (nmol)
0.15 1.33
*
**
***
***
0
50
100
150
200
250p < 0.05 *
0.15 1.33 0.15 1.33 0.15 1.33
200
- 40
0- 20
Coronariaáramlás
(Δ%)
Szívfrekvencia
Artériásvérnyomás
(Δ%)
(Δ%)
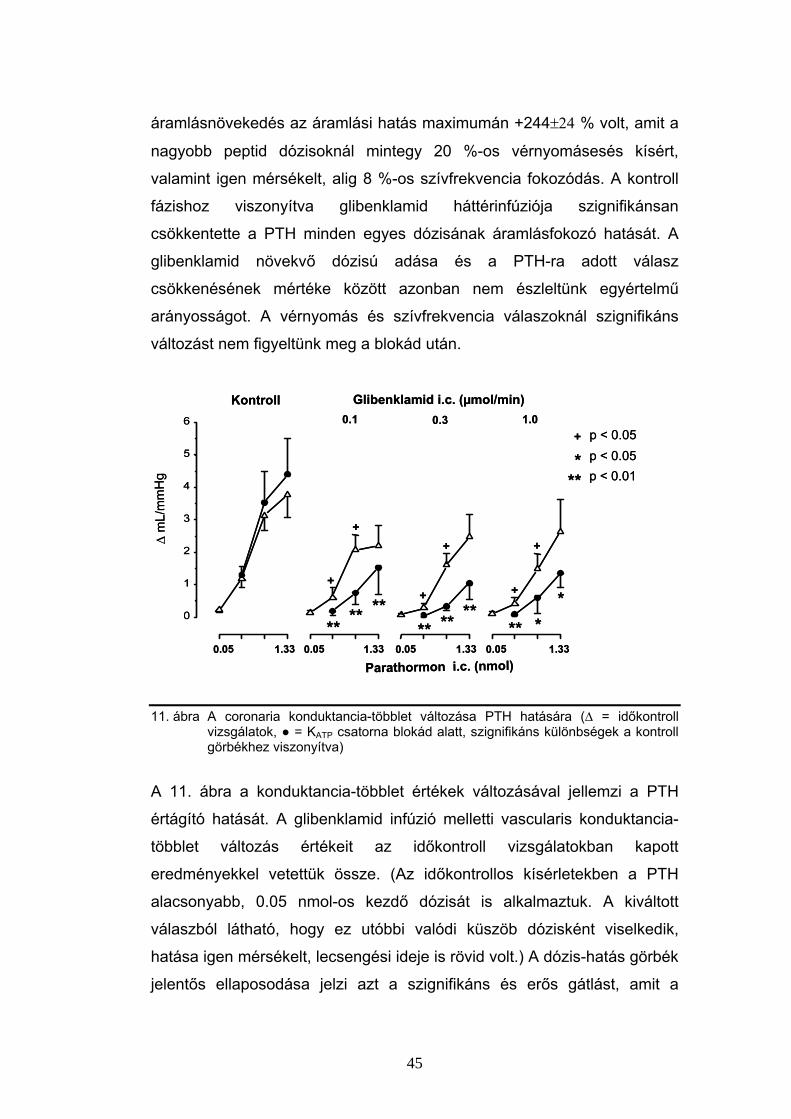
45
áramlásnövekedés az áramlási hatás maximumán +244±24 % volt, amit a
nagyobb peptid dózisoknál mintegy 20 %-os vérnyomásesés kísért,
valamint igen mérsékelt, alig 8 %-os szívfrekvencia fokozódás. A kontroll
fázishoz viszonyítva glibenklamid háttérinfúziója szignifikánsan
csökkentette a PTH minden egyes dózisának áramlásfokozó hatását. A
glibenklamid növekvő dózisú adása és a PTH-ra adott válasz
csökkenésének mértéke között azonban nem észleltünk egyértelmű
arányosságot. A vérnyomás és szívfrekvencia válaszoknál szignifikáns
változást nem figyeltünk meg a blokád után.
11. ábra A coronaria konduktancia-többlet változása PTH hatására (Δ = időkontroll vizsgálatok, ● = KATP csatorna blokád alatt, szignifikáns különbségek a kontroll görbékhez viszonyítva)
A 11. ábra a konduktancia-többlet értékek változásával jellemzi a PTH
értágító hatását. A glibenklamid infúzió melletti vascularis konduktancia-
többlet változás értékeit az időkontroll vizsgálatokban kapott
eredményekkel vetettük össze. (Az időkontrollos kísérletekben a PTH
alacsonyabb, 0.05 nmol-os kezdő dózisát is alkalmaztuk. A kiváltott
válaszból látható, hogy ez utóbbi valódi küszöb dózisként viselkedik,
hatása igen mérsékelt, lecsengési ideje is rövid volt.) A dózis-hatás görbék
jelentős ellaposodása jelzi azt a szignifikáns és erős gátlást, amit a
Δm
L/m
mH
g
0.05 1.33 0.05 1.33
Parathormon i.c. (nmol)0.05 1.33 0.05 1.33
+
**
+
+
+
+
+
****
** **** **
**
p < 0.05 p < 0.01
***
p < 0.05 +
Kontroll1.00.30.1
Glibenklamid i.c. (µmol/min)
Δm
L/m
mH
g
0.05 1.33 0.05 1.33
Parathormon i.c. (nmol)0.05 1.33 0.05 1.33
+
**
+
+
+
+
+
****
** **** **
**
p < 0.05 p < 0.01
***
p < 0.05 +
Kontroll1.00.30.1
Glibenklamid i.c. (µmol/min)
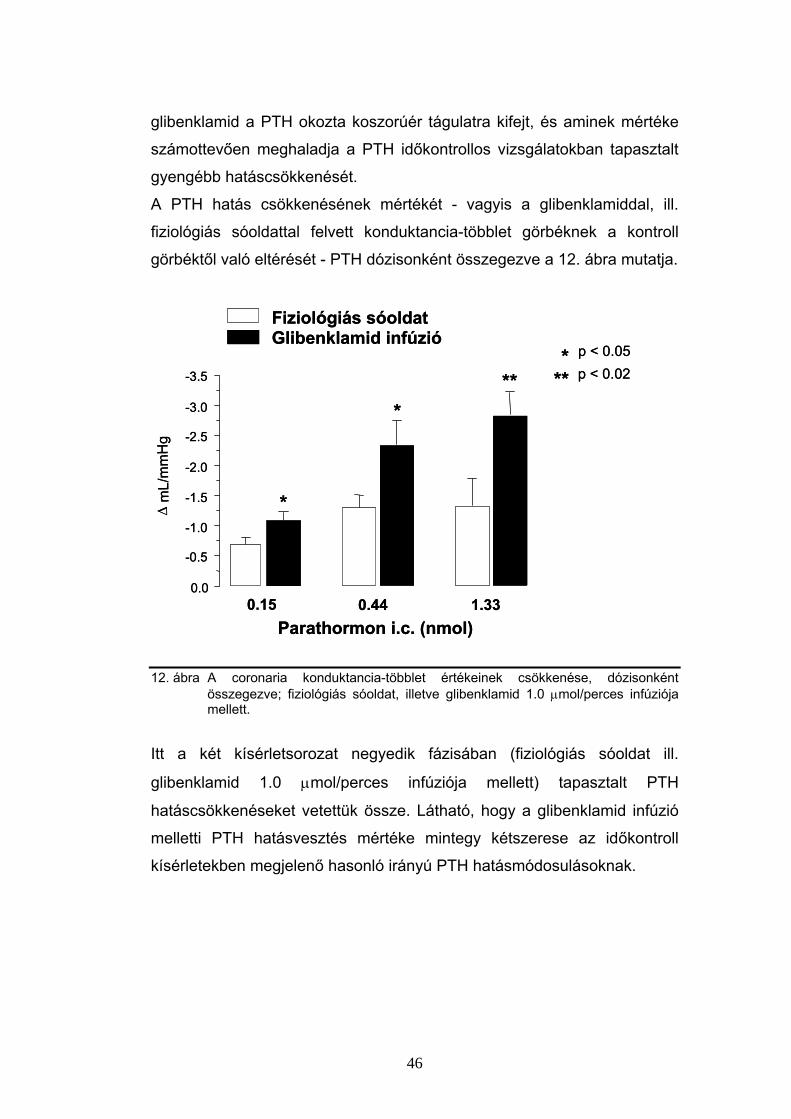
46
glibenklamid a PTH okozta koszorúér tágulatra kifejt, és aminek mértéke
számottevően meghaladja a PTH időkontrollos vizsgálatokban tapasztalt
gyengébb hatáscsökkenését.
A PTH hatás csökkenésének mértékét - vagyis a glibenklamiddal, ill.
fiziológiás sóoldattal felvett konduktancia-többlet görbéknek a kontroll
görbéktől való eltérését - PTH dózisonként összegezve a 12. ábra mutatja.
12. ábra A coronaria konduktancia-többlet értékeinek csökkenése, dózisonként összegezve; fiziológiás sóoldat, illetve glibenklamid 1.0 μmol/perces infúziója mellett.
Itt a két kísérletsorozat negyedik fázisában (fiziológiás sóoldat ill.
glibenklamid 1.0 μmol/perces infúziója mellett) tapasztalt PTH
hatáscsökkenéseket vetettük össze. Látható, hogy a glibenklamid infúzió
melletti PTH hatásvesztés mértéke mintegy kétszerese az időkontroll
kísérletekben megjelenő hasonló irányú PTH hatásmódosulásoknak.
0.15 0.44 1.33Parathormon i.c. (nmol)
Δm
L/m
mH
g
*
***
p < 0.05 p < 0.02
***
0.0
-0.5
-1.0
-1.5
-2.0
-2.5
-3.0
-3.5
Fiziológiás sóoldatGlibenklamid infúzió
0.15 0.44 1.33Parathormon i.c. (nmol)
Δm
L/m
mH
g
*
***
p < 0.05 p < 0.02
***
0.0
-0.5
-1.0
-1.5
-2.0
-2.5
-3.0
-3.5
Fiziológiás sóoldatGlibenklamid infúzió

47
6.4 Szisztémás L-arginin hatása a reperfúzióra
A hemodinamikai paraméterek változását a 4. táblázat mutatja. A
szívfrekvencia nem változott sem a kontroll, sem az L-arginin csoportban.
A CPB alatti artériás középnyomás (MAP) 55.1±7.3 Hgmm volt a kontroll
csoportban és 58.2±3.3 Hgmm az L-arginin csoportban (P=N.S.). Hatvan
perc szívmegállás és 40 perc reperfúziót követően az artériás
középnyomás szignifikánsan lecsökkent a kontroll csoportban. Hasonló
tendenciájú, de nem szignifikáns változás volt látható az L-arginin
csoportban. A perctérfogat (cardiac output, CO) nem változott
szignifikánsan egyik csoportban sem az ischaemiás és reperfúziós
változások során. A coronariaáramlás (CBF) a reperfúzió végére a kontroll
csoportban felére lecsökkent (p<0.05), hasonló CBF csökkenést L-arginin
adása mellett azonban nem lehetett kimutatni.
kiindulás 40 min reperfúzió
Kontroll L-arginin Kontroll L-arginin
szívfrekvencia (ütés/min) 132.1±15.6 127.5±2.1 134.6±4.7 116.4±8.2
art. középnyomás (Hgmm) 104.2±2.4 109.2±2.9 64.3±3.4 * 86.2±13.2 †
CO (L/min) 2.5±0.4 2.7±0.3 2.8±0.4 2.6±0.3
coronariaáramlás (mL/min) 42.3±6.2 39.2±6.3 21.3±3.2 * 45.5±9.3 †
4. táblázat Hemodinamikai paraméterek kiindulási és 60 perc szívmegállást majd 40 perc reperfúziót követő értékei.
(* p<0.05 vs. kiindulás, † p<0.05 vs. kontroll)
A kiindulási PRSW értékek mindkét kamrára vonatkoztatva a fiziológiás
tartományban voltak. A bal kamra kiindulási PRSW értéke 63.1±10.3 kerg
volt a kontroll csoportban, és 48.8±8.5 kerg az L-arginin csoportban. A
jobb kamra kiindulási PRSW értéke 21.1±6.6 kerg volt a kontroll
csoportban, és 26.1±4.8 kerg az L-arginin csoportban. A kontroll csoport
bal kamrai PRSW értéke szignifikánsan csökkent (-25%), míg az L-arginin
csoportban nagyjából változatlan maradt (p=N.S.) a 60 perc ischaemiás és
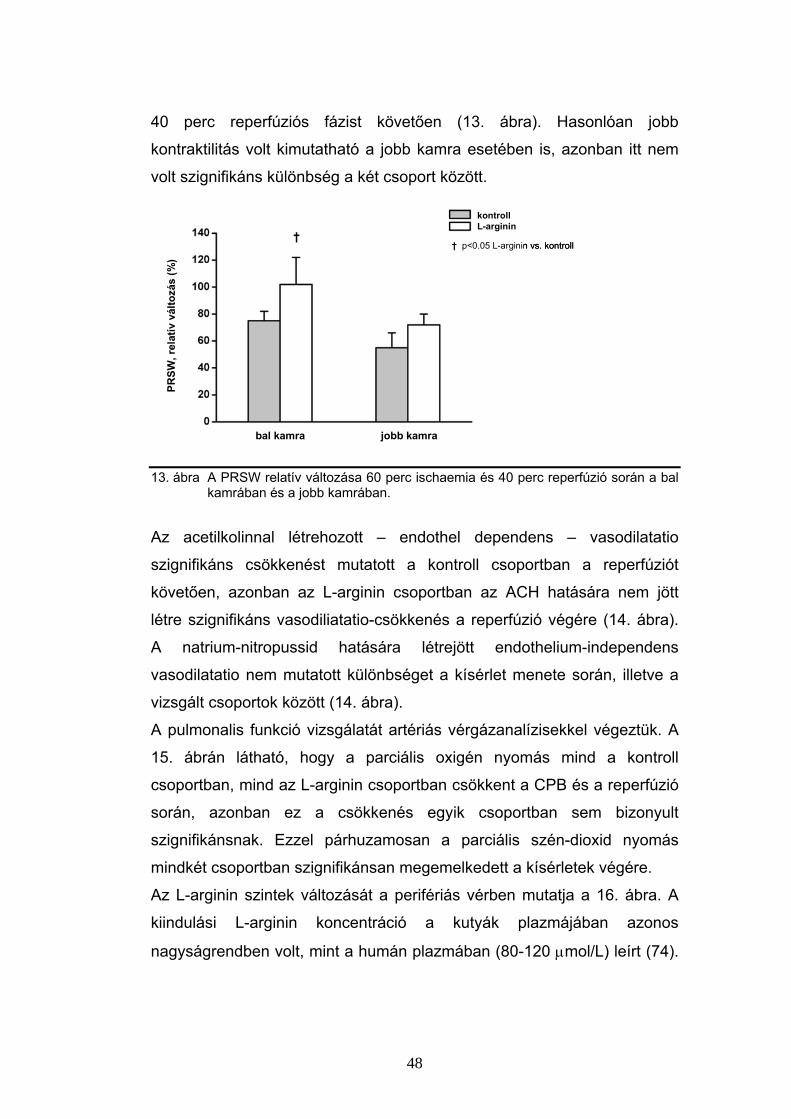
48
40 perc reperfúziós fázist követően (13. ábra). Hasonlóan jobb
kontraktilitás volt kimutatható a jobb kamra esetében is, azonban itt nem
volt szignifikáns különbség a két csoport között.
13. ábra A PRSW relatív változása 60 perc ischaemia és 40 perc reperfúzió során a bal kamrában és a jobb kamrában.
Az acetilkolinnal létrehozott – endothel dependens – vasodilatatio
szignifikáns csökkenést mutatott a kontroll csoportban a reperfúziót
követően, azonban az L-arginin csoportban az ACH hatására nem jött
létre szignifikáns vasodiliatatio-csökkenés a reperfúzió végére (14. ábra).
A natrium-nitropussid hatására létrejött endothelium-independens
vasodilatatio nem mutatott különbséget a kísérlet menete során, illetve a
vizsgált csoportok között (14. ábra).
A pulmonalis funkció vizsgálatát artériás vérgázanalízisekkel végeztük. A
15. ábrán látható, hogy a parciális oxigén nyomás mind a kontroll
csoportban, mind az L-arginin csoportban csökkent a CPB és a reperfúzió
során, azonban ez a csökkenés egyik csoportban sem bizonyult
szignifikánsnak. Ezzel párhuzamosan a parciális szén-dioxid nyomás
mindkét csoportban szignifikánsan megemelkedett a kísérletek végére.
Az L-arginin szintek változását a perifériás vérben mutatja a 16. ábra. A
kiindulási L-arginin koncentráció a kutyák plazmájában azonos
nagyságrendben volt, mint a humán plazmában (80-120 μmol/L) leírt (74).
PRSW
, rel
atív
vál
tozá
s (%
)
kontrollL-arginin
† p<0.05 L-arginin vs. kontroll
bal kamra jobb kamra
PRSW
, rel
atív
vál
tozá
s (%
)
kontrollL-arginin
† p<0.05 L-arginin vs. kontroll
bal kamra jobb kamra

49
A 60 perces hypothermiás szívmegállás alatt az L-arginin szintek mindkét
csoportban közel azonos értéken voltak.
14. ábra Vascularis funkció vizsgálata a coronariákon. Az acetilkolinnal (ACH, 10-7 M) kiváltott endothel-dependens vasodilatatio, illetve a natrium-nitroprussiddal (SNP, 10-4 M) kiváltott endotel-independens vasodilatatio mértéke a coronariák vascularis rezisztenciájának (CVR) százalékos változásában kifejezve látható.
(* p<0.05 vs. kiindulás, † p<0.05 vs. kontroll)
15. ábra A pulmonalis funkció változása. Parciális oxigén (pO2) és szén-dioxid (pCO2) nyomások körlevegőn. (* p<0.05 vs. kiindulás)
kontrollL-arginin
pO2
(Hgm
m)
pCO
2(H
gmm
)
kiindulás 40min rep. kiindulás 40min rep.
kontrollL-arginin
pO2
(Hgm
m)
pCO
2(H
gmm
)
kiindulás 40min rep. kiindulás 40min rep.
acetil-kolin (Ach) Na-nitroprussid (SNP)
kontroll L-arginin
kiindulás 40min rep. kiindulás 40min rep.

50
A reperfúzió 20. percében - ami egyben az L-arginin adásának
befejezését is jelentette - a plazma L-arginin szint szignifikánsan, mintegy
2.5-szeresre emelkedett, majd a 40. percre vissza is tért a kiindulási
szintre. Ezzel szemben a kontroll csoportban az L-arginin szintek közel
változatlanok maradtak a reperfúzió végéig (16. ábra, felső rész).
16. ábra Az NO prekurzor L-arginin (fent), illetve az NO metabolit nitrit (középen) és nitrát (lent) koncentrációváltozásai 60 perces szívmegállás (CA) és 40 perces reperfúziót követően. (* p<0.05 vs. kiindulás, † p<0.05 vs. kontroll)
A NO oxidációja során nitrit és nitrát keletkezik. E két anyag
plazmakoncentrációja megfelelő indikátora az L-arginin lebomlásának. L-
arginin adása után a plazma nitrit koncentrációja szignifikánsan
megemelkedett és a reperfúziós fázis során végig magasabb maradt, mint
kiindulás 20CA 60CA 20rep 40rep
kiindulás 20CA 60CA 20rep 40rep
kiindulás 20CA 60CA 20rep 40rep
L-ar
gini
n(μ
mol
)N
itrit
(μm
ol)
Nitr
át (μ
mol
)
kontrollL-arginin
kontrollL-arginin
kontrollL-arginin
kiindulás 20CA 60CA 20rep 40rep
kiindulás 20CA 60CA 20rep 40rep
kiindulás 20CA 60CA 20rep 40rep
L-ar
gini
n(μ
mol
)N
itrit
(μm
ol)
Nitr
át (μ
mol
)
kontrollL-arginin
kontrollL-arginin
kontrollL-arginin

51
a kontroll csoportban (16. ábra, középső rész). A plazma nitrát szintek
mindkét csoportban közel változatlanok maradtak a szívmegállás során,
de a reperfúzió kezdetén az L-arginin csoport nitrát koncentrációi
szignifikánsan megemelkedtek, és ez a különbség a kísérlet végéig
fennállt (16. ábra alsó rész).
6.5 L-arginin vascularis hatásai agyhalál állapotában
A hemodinamikai paraméterek változását a 5. táblázat mutatja.
Az agyhalál indukciója egy hyperdynam reakciót okozott, melynek során a
legtöbb vizsgált hemodinamikai paraméter nagysága nőtt, a bal kamrai
végdiasztolés nyomás (LVEDP) pedig csökkent. Az akut fázist követően
az aortanyomás szignifikánsan lecsökkent, amit követett a bal kamrai
szisztolés nyomás és a dP/dtmax csökkenése. A perctérfogat (CO) a
kiindulási értékre tért vissza, és közel azonos szinten maradt.
Kiind. t3 t5 t15 t30 t90 t180 HR C 100 ± 3 194±5* 180±13* 150±2* 122±8 98±2 93±3
[min-1] L-arg 92±7 173±7* 174±10* 128±9 103±5 85±5 89±3
AoP C 85±7 139±12* 110±5 59±6* 44±9* 65±3* 61±2*
[Hgmm] L-arg 91±4 125±8* 108±12 54±4* 71±4† 89±7† 87±7†
LVSP C 105±6 158±8* 156±11* 64±2* 72±3* 83±3* 86±2*
[Hgmm] L-arg 105±5 161±6* 128±14* 75±4* 89±2* 101±6† 99±6†
LVEDP C 6±1 3±2 2±0,5 3± 0,7 7±5 5±1 9±5
[Hgmm] L-arg 8±3 3±4 2±2 3±2 5±4 5± 3 7±2
+dP/dt C 1256 ±146
2717 ±164*
2185 ±347*
1135 ±290
818 ±87*
921 ±73
864 ±55*
[Hgmm/s] L-arg 1490 ±122
2462 ±169*
2125 ±226*
1056 ±147
1204 ±98†
1381 ±98†
1269 ±76†
-dP/dt C 889 ±90
1820 ±139*
1370 ±239*
840 ±182*
456 ±39*
663 ±35*
661 ±57*
[Hgmm/s] L-arg 960 ±62
1628 ±195*
1267 ±236*
548 ±81*
699 ±73†
899 ±95†
898 ±72†
CO C 3.4±0.15 6.8±0.3* 6.9±0.4* 4.2±0.8 3.5±0.5 3.1±0.3 2.6*±0.2
[L/min] L-arg 3.3±0.6 5.6±1.2* 5.8±1.1* 3.2±0.5 3.3±0.4 3.5±0.6 3.8±1.1†
5. táblázat Hemodinamikai paraméterek időbeli változása. (t3, t5, … t180 az agyhalál beálltától eltelt időt jelölik percben, * p<0.05 vs. kiindulás, † p<0.05 vs. kontroll)
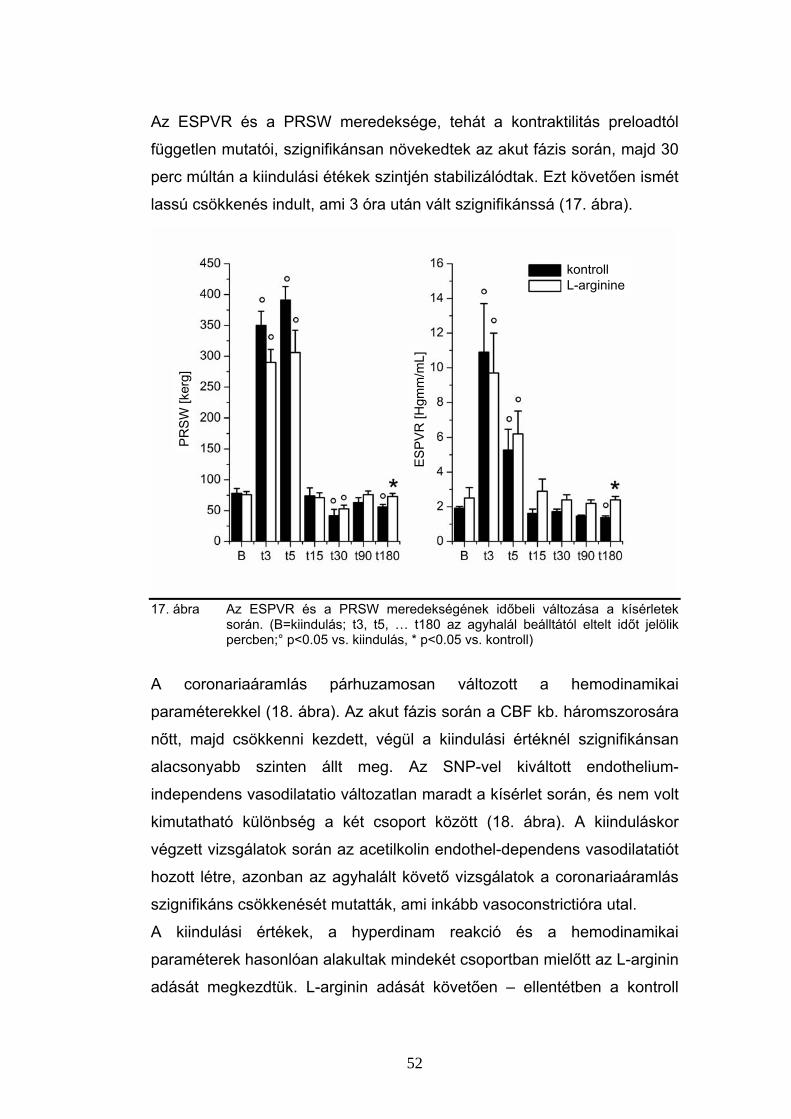
52
Az ESPVR és a PRSW meredeksége, tehát a kontraktilitás preloadtól
független mutatói, szignifikánsan növekedtek az akut fázis során, majd 30
perc múltán a kiindulási étékek szintjén stabilizálódtak. Ezt követően ismét
lassú csökkenés indult, ami 3 óra után vált szignifikánssá (17. ábra).
17. ábra Az ESPVR és a PRSW meredekségének időbeli változása a kísérletek során. (B=kiindulás; t3, t5, … t180 az agyhalál beálltától eltelt időt jelölik percben;° p<0.05 vs. kiindulás, * p<0.05 vs. kontroll)
A coronariaáramlás párhuzamosan változott a hemodinamikai
paraméterekkel (18. ábra). Az akut fázis során a CBF kb. háromszorosára
nőtt, majd csökkenni kezdett, végül a kiindulási értéknél szignifikánsan
alacsonyabb szinten állt meg. Az SNP-vel kiváltott endothelium-
independens vasodilatatio változatlan maradt a kísérlet során, és nem volt
kimutatható különbség a két csoport között (18. ábra). A kiinduláskor
végzett vizsgálatok során az acetilkolin endothel-dependens vasodilatatiót
hozott létre, azonban az agyhalált követő vizsgálatok a coronariaáramlás
szignifikáns csökkenését mutatták, ami inkább vasoconstrictióra utal.
A kiindulási értékek, a hyperdinam reakció és a hemodinamikai
paraméterek hasonlóan alakultak mindekét csoportban mielőtt az L-arginin
adását megkezdtük. L-arginin adását követően – ellentétben a kontroll
ESPV
R [H
gmm
/mL]
PR
SW [k
erg]
kontrollL-arginine
ESPV
R [H
gmm
/mL]
PR
SW [k
erg]
kontrollL-arginine

53
csoporttal – a coronariaáramlás, az aortanyomás, a bal kamrai szisztolés
nyomás, a +dP/dt és a -dP/dt, illetve az ESPVR és a PRSW meredekség
a kiindulási értékre tért vissza (5. táblázat, 17. ábra). L-arginin adása nem
befolyásolta az endothel-independens vasodilatatiót, emellett az endothel-
dependens vasodilatatio sem csökkent – szemben a kontroll kísérletekkel
- a kísérletek végéig (18. ábra, p<0.05 vs. kontroll).
18. ábra Alsó rész: coronariaáramlás (CBF) időbeli változása (B=kiindulás; t3, t5, … t180 az agyhalál beálltától eltelt időt jelölik percben).
Felső rész: a coronariaáramlás százalékos változása acetilkolin (ACH) és natrium-nitroprussid (SNP) adását követően a kísérletek elején és 3 órával az agyhalál beállta után.
(° p<0.05 vs. kiindulás, * p<0.05 vs. kontroll)
A kiindulási nitrit&nitrát szintek nem különböztek a két csoport között. A
kontroll csoportban a kísérletek során időbeli változás sem volt
kimutatható a nitrit&nitrát plazmakoncentrációkban. Ezzel szemben az L-
argininnel kezelt csoportban az infúzió indítását követő azonnali L-arginin
plazmakoncentráció emelkedést egy késleltetett nitrit&nitrát
szintemelkedés követte (19. ábra).
Kiindulás BD után 3 órával Kiindulás BD után 3 órával
kontrollL-arginin
Kiindulás BD után 3 órával Kiindulás BD után 3 órával
kontrollL-arginin

54
19. ábra Plazma L-arginin (balra) és nitrit&nitrát (jobbra) koncentrációk a kísérletek kezdetén (B) és 90, illetve 180 perccel az agyhalál beállta után.
(° p<0.05 vs. kiindulás, * p<0.05 vs. kontroll)
L-ar
gini
n(μ
M)
Nitr
it&ni
trát
(μM
)
kontrollL-arginin
L-ar
gini
n(μ
M)
Nitr
it&ni
trát
(μM
)
kontrollL-arginin

55
7 Megbeszélés
Eredményeink összességükben – az egyes vizsgált paraméterek értékein
túl – a myocardialis ischaemia során aktiválódó lokális regulátor
mechanizmusok heterogén eredetét, ugyanakkor egységes szabályozó
rendszerét vázolják fel. A cardioprotectiv folyamatokban részt vesz a
szöveti struktúrák minden eleme, melyek öt nagy folyadéktér szerint
csoportosíthatók. Ezek a kompartmentek, alkotó substantiáikon és azok
metabolitjain keresztül, egymással folyamatos kapcsolatban állnak, és
tükrözik a szív mindenkori patofiziológiás állapotát. A szív működését
meghatározó biokémiai szabályozásban így részt vesznek a
vérplazmában oldott keringő anyagok, az endothelium (koszorúér,
endocardium, epicardium) által termelt anyagok, a cardiomyocyták által
termelt anyagok, és az interstitialis, illetve a pericardialis térben jelen levő
substantiák. Ezek közül a vérplazma klinikailag könnyen hozzáférhető
folyadéktér, legtöbb alkotóelemének mérése része a rutin laboratóriumi
diagnosztikának, és plazmakoncentrációjuk terápiás irányelveket szabhat
meg. Bár rutindiagnosztikára nem használják, homogén szövetmintákból
szintén meghatározható számos substantia szöveti koncentrációja.
Megfelelően kisméretű molekulák esetében, mint pl. az adenozin, a
szöveti ADO tartalom az intracellularis (endothelialis és parenchymalis
sejtek) és interstitialis ADO tartalom összegét jelentheti. A szöveti ADO
tartalom mérésével az összes ADO képződés változásai jól
jellemezhetőek, mert az intra- és extracellularis szabad ADO
koncentrációk valószínűleg azonosak, illetve az ADO képződés
növekedése mind az intra-, mind az extracellularis ADO szintet növeli. A
nyitott szívműtétek elterjedésével a szervezet egy egzaktul körülhatárolt
folyadéktere, a pericardium is klinikailag hozzáférhetővé vált, lehetővé
téve a szívet teljes egészében körülölelő folyadék-kompartment közvetlen
tanulmányozását.

56
Vizsgálataink során három témát jártunk körül, így a pericardialis tér
szabályozó szerepét a szív patofiziológiás állapotaiban, a parathormon
ismert endokrin hatása mellett jelentkező lokális vasoactiv tulajdonságait,
valamint az NO-prekurzor L-arginin értónust befolyásoló hatásait
myocardialis ischaemia során és agyhalál beálltát követően.
A nátriuretikus peptidek jellemzését szívsebészeti betegekben közel tíz
éve kezdtük el klinikánkon. Azóta az ANP és a BNP plazmaszintjeinek
mérése a kardiológiai diagnosztika első vonalbeli eszköze lett (75,76).
Kihasználva a szívsebészeti beavatkozások adta különleges lehetőséget a
pericardialis folyadék összetételének feltérképezésére, nagy
beteganyagon meghatároztuk a pericardialis tér és a vérplazma ANP
koncentrációs viszonyait. A pericardialis tér ANP tartalmának korábbi
meghatározása során egymásnak ellentmondó eredmények születtek
(15,18,27). Nagy esetszámú méréseink eredménye alapján
megállapítható, hogy a szívműtétre kerülő betegek pericardialis
folyadékában mérhető ANP koncentrációk egy nagyságrenddel
meghaladják a plazmában mérhető koncentrációkat. Emellett a
pitvarokban mért szöveti ANP koncentráció több százszorosa a kamrai
szövetekben mérhetőnek. A pericardialis térben mért magas ANP
immunoaktivitás kromatográfiás méréseink szerint döntően a biológiailag
aktív – ANP [99-126] - molekuláris formától származik. Gyakorlatilag
azonos pericardialis folyadék - vénás plazma koncentráció gradiens
található az ischaemiás és a nem-ischaemiás (billentyű) betegekben; a
koszorúér betegek ANP szintjei azonban némileg alacsonyabbak a másik
betegcsoportban mérteknél. Bár ezek a különbségek a két betegcsoport
között statisztikusan szignifikánsak, az eltérések nagyon kis mértékűek,
így klinikai jelentőségük megkérdőjelezhető. Ezt a feltételezést
támaszthatja alá, hogy a pitvari és kamrai szöveti ANP szintek nem
mutattak különbséget az ischaemiás és a nem-ischaemiás betegcsoportok
között.

57
Általánosan ismert tény, hogy az emlősök perifériás vérében a pitvari
falfeszülés hatására a keringési rendszerbe kiválasztódó ANP a
legfontosabb nátriuretikus peptid (5). Ugyanakkor kevés tudható az
epicardialis ANP szekréció folyamatairól. Az intrapericardialis ANP egyik
forrása a pitvari ANP szekréció lehet, ahogy ezt korábban kifordított nyúl-
pitvaron demonstrálták (77). Szintén ismert, hogy a pericardialis folyadék
protein- és ionösszetételét tekintve nagyjából megfelel egy plazma-
ultrafiltrátumnak. Az ANP magas intrapericardialis tér-plazma irányú
gradiense alapján azonban valószínűbbnek látszik, hogy a pericardialis
folyadékban kimutatható peptid forrása mégsem a vérplazma, hanem a
myocardialis interstitium, és ily módon az intrapericardialis ANP
koncentráció az interstitialis tér viszonyaira utal. Korábbi vizsgálatok
alapján a neutrális endopeptidáz – az ANP lebontásáért felelős
legfontosabb enzim – aktivitása a pericardialis folyadékban és a vénás
plazmában nagyjából azonos mértékűnek bizonyult (78), mutatva, hogy az
intrapericardialisan mért magas ANP szintek nem adódhatnak csökkent
intrapericardialis NEP aktivitásból. Vizsgálatainkban a kamrai ANP szintek
kevesebb, mint 0.5%-át tették ki a pitvari szöveti koncentrációnak.
Eredményeink alapján ezért feltételezhető, hogy a pericardialis ANP
forrásai a pitvari szövetek. Ezt magyarázhatja az a tény, hogy – bár mind a
pitvari, mind a kamrai cardiocyták szintetizálnak és szekretálnak is ANP-t
– a peptidet csak pitvari sejtek képesek szekretoros granulumaikban
tárolni. Míg a kamrai szövetek ANP termelése folyamatos, addig a pitvari
szövetek a megtermelt ANP-t tárolni tudják és szabályozottan ürítik (79).
Mivel azonban a kamrák tömege sokkal nagyobb, mint a pitvaroké, a
pericardialis folyadékban található ANP származhat a pitvarok mellett a
kamrákból is. A pericardialis folyadék kísérleti állatokban mért magas ANP
koncentrációja (19,23,28) alapján az is felvetődik, hogy – legalábbis
néhány fajban – élettani jelenségről és nem kóros állapotról van szó.
Összegezve, az eddigi eredmények felvetik az ANP parakrin hormonként
játszott biológiai szerepének lehetőségét, melynek során, bár az

58
interstitialis és a keringő (endokrin) ANP szabályozó működése
valószínűleg különböző, egymást kiegészítő rendszert képezhetnek.
Mivel lényeges különbség nem volt kimutatható az ischaemiás és nem-
ischaemiás betegek intrapericardialis ANP koncentrációi között, és nem
találtunk szignifikáns korrelációt e betegek plazma és pericardialis ANP
koncentrációi között sem, feltételezhető, hogy az ANP intrapericardialis
térbe történő szekrécióját meghatározó folyamatok működése eltérő más,
az O2 igényt és kínálatot finoman szabályozó rendszerekétől. Az ANP a
coronaria-keringésben meglehetősen gyenge vasodilatator anyag,
nemcsak korlátozott moláris potenciálját, hanem mérsékelt hatékonyságát
tekintve is (80). Az, hogy az intrapericardialis (vagy interstitialis) ANP-nek
van-e számottevő vasoactiv hatása a coronaria mikroerekre, további
vizsgálatokat igényel; jelen ismereteink alapján ez valószínűtlennek tűnik.
Ennek ellenére nem zárható ki, hogy az intrapericardialis ANP-nek
vasoconstrictor mechanizmusokkal szembeni protektív hatása van a nagy
(konduktív) koszorúereken (81), melyeket csak egy vékony serosus
membrán választ el a pericardialis folyadéktól. Az intrapericardialis ANP
által kiváltott más, hosszú távú lokális protektív hatások viszont
összefügghetnek egyes kórfolyamatok (pl. szívizom-hypertrophia vagy
szívelégtelenség) klinikai lefolyásával.
Vizsgálataink szerint a korábban lezajlott myocardialis infarctus (átlagosan
43±6 hónap) nem befolyásolta az ANP szinteket. Irodalmi adatok
lényegében alátámasztják kapott eredményeinket (82), bár ezekben leírtak
olyan al-betegcsoportokat, melyekben emelkedett ANP koncentrációkat
mutattak ki. Ezek, és az általunk kimutatott, MI után inkább csökkenő ANP
szintek (melyek csak a CAD csoportban vizsgálva már nem mutattak
szignifikáns különbséget) azt jelzik, hogy a myocardialis infarctusnak nincs
hosszú távú hatása a plazma ANP koncentrációra. Ez megfelel azoknak a
megfigyeléseknek is, melyek szerint az ANP maximális koncentrációját a
keringésben már az első héten eléri, és azokban a betegekben, akinél
nem lép fel congestiv szívelégtelenség vagy arrhythmia komplikációként,
az infarctust követő második hét során visszatér a normál szintre (83).

59
Emellett figyelembe kell venni, hogy krónikus szívelégtelenség során, a
szervezet adaptációs mechanizmusai hatására a plazma ANP szintje
megnő;elfedheti a myocardialis infarctus okozta ANP emelkedést és a
pericardialis folyadék ANP tartalmának tartós emelkedéséhez vezethet
(83).
A nemi hormonok hatása a szív működésére, illetve biokémiai
folyamataira, ismert. A cardiovascularis megbetegedések incidenciája
fiatal nőbetegekben alacsonyabb, mint férfiakban, azonban ez a
különbség a menopauzát követően megszűnik a nemek között.
Hormonterápiában részesülő nők között kevésbé gyakori a koszorúér
megbetegedés, mint a gyógyszeresen nem kezeltekben, ami felveti a
ösztrogén protektív szerepét a coronaria betegségekkel szemben (84).
Irodalmi adatok szerint a plazma ANP és BNP szintjei szignifikánsan
megemelkednek hormonterápia során, ami felveti a nátriuretikus peptidek
– legalábbis részleges - szerepét az ösztrogén cardioprotectiv hatásának
kifejtése során (85). A szív nátriuretikus peptid termelésére egyrészt
közvetve fejthet ki hatást az ösztrogén;endokrin vagy parakrin úton, a
renin-angiotenzin rendszer stimulálásával, majd a vasoconstrictiv
folyamatokra a nátriuretikus peptidek által adott vasodilatativ válasz során.
Emellett közvetlenül is befolyásolhatja a szív nátriuretikus peptid
termelését;receptor mediálta folyamatokon keresztül fokozhatja a
génexpressziót és a peptidek kiválasztását (86). A myocardiumban
kimutattak ösztrogén receptorokat, azonban ezek pitvari és kamrai
eloszlása még nem tisztázott (87). Eredményeink alapján úgy tűnik, az
ösztrogén direkt és/vagy indirekt hatása a kamrai szöveteken egyformán
érvényesülhet mindkét nemben, azonban felvetődik, hogy az ösztrogén
pitvari szöveteken kifejtett hatása nők esetében erősebb. A pitvari
szövetekben található ösztrogén receptorok részletes leírásával a
továbbiakban tisztázható, hogy az ösztrogén hatására történő nátriuretikus
peptid termelésben mekkora szerepe van a receptor mediálta direkt, illetve
az indirekt úton (renin-angiotenzin rendszer) történő szabályozó
mechanizmusoknak. Továbbá megmagyarázhatja, mi okozza azt a

60
jelenséget, mely szerint a plazma és a pericardialis folyadék esetében
nem volt kimutatható olyan ANP koncentráció különbség, mint a pitvari
szövetek esetében.
A vénás plazma, illetve a pitvari és kamrai ANP szintek szoros
összefüggést mutattak a betegek klinikai állapotával - NYHA stádiumuk
romlásával. A viszonylag magas és nagyjából változatlan mértékű
intrapericardialis ANP koncentráció megerősíti azt az elképzelésünket,
hogy a pericardialis folyadékban felhalmozódó ANP-nek szerepe lehet a
szívműködés szabályozásában, és ez a hatás – legalábbis részben -
parakrin módon érvényesül.
A szív szöveteiben lejátszódó folyamatok még napjainkban sem
figyelhetők meg közvetlenül. A sejtszinten zajló folyamatok leírásában
nagy szükség lenne a közvetlen sejtkörnyezet élettani változásainak
követésére. Ezért az elmúlt évek során számos próbálkozás történt, hogy
a cardialis interstitium összetételét leírják, a benne zajló mechanizmusokat
elemezzék. A vizsgálatok tárgyai – többek között - olyan endogén
ágensek, melyek részt vesznek a myocardium metabolikus
szabályozásában, így koncentrációjuk a myocardium ischaemiás
állapotával együtt változik, ezért utánkövetésük a napjainkban ismert
módszerekkel jó közelítéssel megvalósítható. Myocardialis ischaemia
során az adenin nukleozid rendszer aktiválódik és a felszabaduló
mediátorok – adenozin és inozin - jelentős szerepet játszanak a
myocardium vérellátása és oxigénigénye közötti egyensúly
helyreállításában (88). A klasszikus adenozin hipotézis szerint az
elsődleges mechanizmus, mely a koszorúér áramlást a myocardialis
metabolizmus igényeihez illeszti, az adenozinon keresztül valósul meg.
Emellett az inozin, ami önmagában is vasodilatator hatású, szintén
szerepet játszhat a koszorúerek metabolikus autoregulációjában,
támogatva az adenozin okozta vasodilatatiót stabilizátor és modulátor
hatásával (22,88). Az elmúlt évek során számos kísérlet történt az
interstitialis nukleozid felszabadulás meghatározására hypoxiás

61
myocardiumból. Mérték az adenozin és bomlástermékeinek
koncentrációját szívizom biopsziás mintákban, pericardialis
superfusatumban, szívfelszínre ragasztott epicardialis gyűjtőedényekben,
illetve mikrodialízis módszerrel (22). Ezeket az mérési eredményeket
azonban jelentősen befolyásolhatja az adenozin gyors felhasználása és
lebomlása a vérben (89). Emellett a különböző méréstípusok az eltérő
technikai lehetőségek miatt, akár nagyságrendnyi eltérést is mutathatnak
egymáshoz képest.
Mivel a közvetlen mérési eredmények alapján az interstitialis tér
koncentrációs viszonyait kellő biztonsággal meghatározni nem sikerült,
matematikai eljárásokat dolgoztak ki, melyekkel a klinikailag hozzáférhető
folyadékterek összetételének ismeretében a myocardialis interstitialis tér
koncentrációs viszonyai megbecsülhetők. Ezek a modellek szintén az
adenin nukleozidok élettani szerepére épülnek, mivel mind az adenozin,
mind az inozin metabolikus és transzport folyamatai irodalmi adatok
alapján jól jellemezhetőek (90).
Ismert, hogy a kis molekulatömegű adenozin és inozin az epicardialis
felszínen keresztül szabadon diffundál, és a pericardialis térben
felhalmozódik (31). Munkánk során abból indultunk ki, hogy az epicardialis
felszínen történő molekulaáramlás következtében az interstitialis folyadék
és a pericardialis folyadék között egyensúly alakul ki a nukleozidok
tekintetében, így a pericardialis folyadék nukleozid koncentrációja jól
reprezentálhatja az interstitialis nukleozid viszonyokat. Matematikai
modellünk tartalmazza az öt legfontosabb pericapillaris folyadékteret,
illetve elemzi ezek alapvető metabolikus és transzport folyamatait.
Természetesen ezek az egyenletek nem adhatnak kimerítő képet a
cellularis működésről, és nem írják le az összes biokémiai folyamatot,
melyekben részt vesznek az adenin nukleozidok. Azonban megfontoltan
kiválasztva a legfontosabb alkotóelemeket, és képletesen leírva a köztük
zajló folyamatokat, egy alapszintű összefüggésrendszert alkothatunk,
melyben megfelelő mennyiségű ismert paraméterből, az ismeretlen
értékek kiszámíthatók.

62
Eredményeink alapján egy öt kompartmentből álló matematikai modellel jó
közelítéssel jellemezhetők a perikapilláris szövetek koncentráció-
viszonyai. Az egyes kompartmentekben lezajló metabolikus változások és
a kompartmentek közötti tranzit-folyamatok figyelembevételével egy
egyszerűsített modell hozható létre, ami alapján a vénás plazmában,
illetve a pericardialis folyadékban mérhető ADO koncentrációk alapján a
myocardialis interstitium ADO koncentrációja megbecsülhető. Azonban a
modellezés során kapott eredmények mind az irodalomban, mind korábbi
vizsgálataink alapján, alulról közelítették a kísérletes úton mért értékeket,
felhívva a figyelmet a matematikai leíró modell élettani korlátaira. A
modellezéssel számított értékek a későbbiekben tovább finomíthatók,
amennyiben az élettani folyamatok leírása is tökéletesebbé válik. Egy ilyen
modell alapja lehet más molekulák kompartmentális eloszlásának
vizsgálatára is, azzal a fontos kitétellel, hogy a vizsgált anyag
metabolizációjáról és transzport-folyamatairól legalább közelítő
ismereteink legyenek. Modellünket első sorban "statikus" állapotok
jellemzésére lehet alkalmazni, mert a gyorsan zajló és az egész élő
szervezetet komplexen magába foglaló változásokat az élettani
ismereteink még csak meglehetősen érintőlegesen tudják kezelni.
A parathormon koszorúér keringésre gyakorolt hatásainak experimentalis
vizsgálata során kimutattuk, hogy a parathormon - jól ismert endokrin
hatása mellett - megfelelő koncentrációban a koszorúerek vasodilatatióját
okozza, illetve, hogy a koszorúerek KATP csatornáinak glibenklamiddal
történő gátlása jelentősen és szignifikánsan csökkenti a PTH által kiváltott
coronaria dilatatiót. Ez arra enged következtetni, hogy a KATP csatornák
számottevő szerepet töltenek be a PTH koszorúértágító hatásának
kialakulásában, ami új megvilágításba helyezheti a parathormon
hatásmechanizmusáról alkotott eddigi elképzeléseinket.
Kísérleteinkben a PTH dózisfüggően növelte a coronariaáramlást, ami
egybevág az irodalomban már leírt, PTH által előidézett jelenséggel.
Korábbi vizsgálatok eredményei alapján a parathormon által kiváltott

63
vasodilatatio EC50 értéke nM-os tartományba esik (39). Kísérleteinkben az
alkalmazott parathormon dózisokat ennek megfelelően választottuk meg;
így vizsgálataink alapján az EC50≤0.44 nM. Különböző tanulmányok 2.4 és
100 pM közé teszik a vér parathormon szintjét euparathyreoid
betegekben, azonban a parathormon plazmakoncentrációja akár
többszörösére is emelkedhet patofiziológiás állapotokban (91,92). Így az
általunk végzett kísérletekben, élettani viszonyoknak megfelelő
parathormon [1-34] koncentrációk mellett hoztunk létre coronaria
vascularis rezisztencia csökkenést, ami megerősíti, hogy a parathormon
által kiváltott áramlásnövekedés nem egyszerűen csak gyógyszertani
hatás volt. Alacsony PTH dózisok kísérleteinkben nem befolyásolták a
vérnyomást, ill. a szívfrekvenciát, ugyanakkor a peptid nagyobb dózisainak
alkalmazásakor pozitív chronotrop, és hypotensiv hatás kísérte a
coronaria dilatatiót. Hasonló jelenséget már több esetben leírtak, és ezek
alapján feltételezhető, hogy vérnyomáscsökkentő hatásában a
parathormon érrendszerre kifejtett direkt hatása játszik szerepet
(35,42,93).
Az irodalomban eddig közölt adatok és elképzelések szerint a PTH
vascularis hatásának kifejlődésében részint az intracellularis cAMP
rendszer, részint az L-típusú Ca csatornák játszanak szerepet. Egyes
szerzők szerint ugyanis a PTH az adenil-cikláz rendszer aktivitását
befolyásoló receptorán keresztül növeli a sejten belüli cAMP szintet;ez
aktiválja a cAMP-függő protein kinázt, és ez a miozin könnyű lánc kináz
foszforilálása révén vezet a simaizomsejt csökkent kontraktilitásához
(94,42). Más elképzelések szerint a PTH valamilyen módon blokkolni
képes az L-típusú feszültségfüggő Ca csatornákat, és az ennek
eredményeként létrejövő sejten belüli szabad kalciumszint csökkenésnek
tulajdonítható szerep a peptid-indukálta érrelaxáció kialakulásában
(44,95). Eddigi ismereteink alapján e két felfogás összeegyeztetése
nehézségekbe ütközött. Jelen eredményeink alapján azonban
feltételezhető, hogy a parathormon korábban az irodalomban leírt Ca
csatorna blokkoló hatása - legalábbis részben - a KATP csatornák

64
aktiválódása révén valósul meg. A sejtmembrán KATP csatornái elvileg
több úton kapcsolódhatnak a fenti folyamatba. Ezek közül érdemes
kiemelni a guanil nukleotid regulátorfehérje által stimulált cAMP
képződést, mely a cAMP-függő proteinkináz A aktiváció hatására létrejövő
csatorna foszforiláció következtében a KATP csatorna megnyílásához
vezet. Ezt a szabályozó mechanizmust artériás simaizomszövetekben -
többek között koszorúerekben - írták le, vasodilatatiós működésüket
vizsgálva (96,97). Az intracellularis cAMP rendszert involváló reakcióút
érdekessége lehet, annak potenciális azonossága a PTH klasszikus
célsejtjein (a csont-, ill. a vesesejteken) keresztül kifejtett extracellularis Ca
szint emelő hatásának mechanizmusával. Felmerül annak lehetősége,
hogy a PTH által kiváltott hypercalcaemiás és vascularis válaszreakciók
alapvetően azonos úton, az intracellularis cAMP szint emelkedése által,
valósulnak meg. A PTH koszorúér dilatáló hatásának folyamata, tehát, a
következő hipotézis szerint vázolható fel: A PTH, mint ismeretes, a
koszorúér simaizomsejteken lévő PTH receptorokhoz kötődik. A hormon-
receptor komplex a sejtmembrán GS-regulátorfehérjéjéhez kapcsolódik,
mely aktiválja a membrán belső oldalán lévő adenilciklázt, és ezáltal
megemeli az intracellularis cAMP szintet. A cAMP aktiválja a proteinkináz
A-t, az aktivált enzim pedig foszforilálja az ér simaizomsejtek
membránjának KATP csatornáit. A KATP csatornák aktiválódása és
következményes megnyílása – kálium kiáramlás révén – hyperpolarizálja
a sejtet. A membránpotenciál növekedése az L-típusú feszültségfüggő Ca
csatornák záródását (nyitvatartási valószínűségének csökkenését)
eredményezi, így a csatornákon keresztül csökken a Ca beáramlás, és a
következményes intracellularis Ca szint, ami így a coronaria simaizom
relaxációjához vezet. Az eredmény, tehát koszorúér dilatáció. A PTH
koszorúértágító hatásának általunk feltételezett kialakulásmódja tehát
összeegyeztethető az irodalomban eddig közölt cAMP rendszert, ill. L-
típusú Ca csatornákat érintő hipotézisekkel.
Ma még nem ismert a parathormon élettani szerepe a koszorúértónus
szabályozásában. Az azonban bizonyos, hogy a hormon vascularis

65
hatásának kifejtéséhez lényegesen alacsonyabb dózisok is elegendőek,
mint amennyi a jól ismert hypercalcaemiás válasz kiváltásához szükséges.
A keringésben előforduló parathormon fragmentált szerkezete, a
biológiailag aktív aminoterminális fragment relatív rövid féléletideje (t1/2 =
5-7 perc), és lokális koncentrációinak eltérései mind a peptid élettani
szerepének sokrétűségét tanúsítják. A PTH hatás jelentőségére utaló
elképzeléseket megerősítette, és számottevően kibővítette az az alig több,
mint egy évtizedes felfedezés, hogy számos szerv – köztük a szív és az
érrendszer – maguk is elválasztanak egy, hatásában a PTH-hoz
mindenben hasonló igen nagy molekulatömegű peptidet (PTH-related
protein, PTHrP), mely azonban csak kivételesen (az ún. malignus
humoralis hypercalcaemiában) funkcionál a PTH-hoz hasonló keringő
hormonként, egyébként (fiziológiásan) parakrin működést fejt ki (98).
A NO prekurzor L-arginin vasculaturára kifejtett élettani hatásait két
experimentalis modellen vizsgáltuk. Az első kísérletsorozatban reversibilis
hypothermia és cardiopulmonalis bypass-ot követően a szisztémás
keringésbe adva vizsgáltuk cardiovascularis és pulmonalis hatásait, illetve
pharmakokinetikai tulajdonságait a reperfúzió során. Második
kísérletsorozatunkban vizsgáltuk az agyhalál során jelentkező
endothelialis diszfunkciót, illetve ennek során az L-arginin-NO tengely
protektív szerepét a vascularis funkciókra.
Az L-arginin endothelialis diszfunkciót csökkentő hatását eredetileg
hypercholesterinaemiás nyulakon végzett kísérletek során írták le (99).
Mivel az L-arginin a NO prekurzor molekulája, exogen L-arginin adásával
a NO termelés fokozható, különösen olyan endothel sérüléssel járó
állapotokban, amikor az endothelialis NO termelés alapvetően csökkent
(pl. ischaemia-reperfúzió során). Bár az L-arginin hatásait számos
experimentális és humán vizsgálat során elemezték, a cardialis és
pulmonalis működésre kifejtett hatása még mindig nem egyértelműen
tisztázott (49,59). A legtöbb vizsgálat pozitív cardialis hatásokról számolt
be;úgymint az ischaemiát követően megőrzött szisztolés és diasztolés

66
regionalis szívfunkció, illetve csökkent infarktus-kiterjedés (101,102),
emellett azonban úgy tűnik, az L-arginin pulmonalis rendszerre kifejtett
hatása indifferens (100).
Ischaemia-reperfúziós változásokat modelláló experimentalis
vizsgálatainkban az ACH-ra adott coronaria válasz L-arginin adására a
reperfúzió során jelentős javulást mutatott, továbbá szignifikáns javulás
volt kimutatható a kontraktilitás praeloadtól független mérőszámaiban.
Kísérleteink során a bal kamrai kontraktilitás nem mutatott csökkenést, és
hasonló tendenciát tapasztaltunk a jobb kamrai kontraktilitásokban is, ami
azt mutatja, hogy a reperfúzió során adott L-arginin segíti a szívfunkció
javulását – legalábbis - a reperfúzió korai szakában (első 40 perc).
Adataink alátámasztják azokat a korábbi eredményeket, melyek az L-
arginin által megnövelt NO szintézis ischaemia-reperfúzió során játszott
protektív szerepét hangsúlyozzák (63,103,104). Az L-arginin első sorban a
postischaemiás véráramlásra fejti ki jótékony hatását, mivel az
endothelialis NO termeléséhez szolgáltat további substrat mennyiséget
(63,103). Emellett, a NO gátolja az érfal és a vér keringő elemeinek
köcsönhatását, ami szintén fontos szerepet játszik a reperfúziós
károsodásokban (105). Antioxidáns ágensként továbbá, szabad gyököket
tud megkötni, első sorban superoxid-gyököket, amik az ishcaemia során
keletkeznek (106). Érdekes módon, ezek a hatások első sorban a szíven
jelentkeztek, és a várt pulmonalis hatások elmaradtak, amint ezt a
kísérleteink során szignifikánsan nem változó pulmonalis oxigenizációs
működés mutatta. A két szervrendszer között az ischaemiát követő
felépülésben jelentkező különbség oka egyelőre nem tisztázott, azonban
ebben jelentős szerepet játszat az általunk alkalmazott dózis nagysága,
ahogyan ezt már más kísérletek során is tapasztalták (100). A
szívsebészetben a krisztalloid és a véres cardioplegiát egyaránt
alkalmazzák, azonban mindkettőről ismert, hogy alkalmazásuk mellett
történhetnek reperfúziós károsodások, mint pl. endothelialis diszfunkció
vagy kontraktilitás csökkenés (107). Az elmúlt évek során azonban
mindkettőről kimutatható volt, hogy fokozott myocardialis és endothelialis

67
védő hatása van, ha a kezelést kiegészítik L-argininnel (107,108,109).
Azonban az L-arginin adásánakidőpontja fontos tényező lehet a hatás
kifejlődése szempontjából. Bár a legtöbb munkacsoport jó eredményekkel
alkalmazza az L-arginint cardioplegiás oldatokkal keverve (110), izolált
patkányszíven kimutatták, hogy az L-arginin a reperfúzió során alkalmazva
jobban kifejtheti cardioprotectiv hatását (111). Így, mivel az L-arginin első
sorban a reperfúzió során fejti ki pozitív hatását (63,101), kísérleteinkben a
reperfúziós fázis elején kezdtük el adását. Továbbá feltételeztük, hogy az
L-arginin szisztémás adásával a másodlagos szervkárosodást is
csökkenteni tudjuk. Az L-arginin intravénás adását követően 20-30 perccel
éri el a plazmában csúcskoncentrációját, féléletideje rövidebb, mint 1 óra
(112). Vizsgálataink eredményei azt mutatják, hogy reperfúzió során az L-
arginin farmakokinetikája hasonló lefolyást mutat; kb. 20 perc után éri el a
plazma csúcskoncentrációt, majd 40 perc után visszatér a kiindulási
szintre. A plazmából történő relatív gyors elimináció alapján arra lehet
következtetni, hogy jelentősen nőtt a szervezet L-arginin igénye a
kísérletek során. Az L-arginin lebontása során NO keletkezik, ami először
nitritté oxidálódik, majd oxigenált hemoglobin segítségével gyorsan nitráttá
alakul. Ezért a NO szintézis indikátoraiként gyakran használják ezt a két
NO-metabolitot. Normálisan a plazma nitrit koncentrációja alacsony, a
nitrát koncentrációja ennél kb. 100-szor magasabb (113). Kísérleteinkben
L-arginin adása után a nitrit szintek nagyjából az L-argininével egy időben
érték el csúcskoncentrációjukat, ezzel szemben a nitrát szintek folyamatos
növekedést mutattak és nem tértek vissza a kiindulási szintjükre. Az
ischaemia során mindkét csoportban változatlan szinten maradt L-arginin,
nitrit és nitrát koncentrációk a hypothermia során lecsökkent metabolikus
aktivitás eredményei. Emellett, a reperfúzió során le nem csökkent L-
arginin szintek hátterében a csökkent NOS aktivitás állhat. Korábbi
kísérletek igazolták, hogy a szöveti eNOS aktivitás L-argininnel kezelt
szívben fenntartható ischaemia és reperfúzió során, miközben a
kontrollként szolgáló szívekben szignifikánsan csökken az eNOS aktivitás
(104,114). Összegezve ezeket az adatokat valószínűsíthető, hogy –

68
legalábbis bizonyos körülmények között – az L-arginin jelenléte szerepet
játszik az endothel sejtek NOS aktivitásában. Emellett azonban az
indukálható NOS (iNOS) expressziója is fokozódhat ischaemia-reperfúziós
folyamatok, illetve szisztémás gyulladásos reakciók során (115). Az
ischaemiás-reperfúziós károsodások során a csökkent eNOS aktivitás
csökkent „vascularis NO” termelődéshez vezethet, ezzel szemben a
fokozott iNOS aktivitás emeli a „gyulladásos NO” termelést. E két folyamat
eredményeként a plazma összes NO koncentrációja nem változik; hasonló
jelenséget tapasztaltunk kísérleteink során az L-argininnel nem kezelt
állatokban. Ezek alapján feltételezhető, hogy az L-arginin NO
prekurzorként az eNOS enzimek működését befolyásolja, és ezáltal
fokozza az endothelialis NO szintézist, illetve javítja az erek vasodilatator
képességét. Ahogyan ezt kísérleteink igazolták, exogen L-arginin hatására
tovább nőhet az eNOS aktivitás. Valószínűleg ezeknek a hatásoknak
köszönhető, hogy az L-arginin plazmakoncentrációja gyorsan visszatért
kiindulási értékére a reperfúzió során adott L-arginin kezelést követően.
Eredményeink alapján tehát, a NO prekurzor L-arginin hypothermiás
ischaemia és reperfúzió során csökkenti a reperfúziós károsodásokat és
javítja az endothelialis funkciót. Az L-arginin kedvező hatásait kifejtette úgy
is, hogy csak a reperfúzió során kezdtük adását, és intravénás
alkalmazásával a lokális hatásain túl a szisztémás vasculaturára is
kifejtette hatását.
Az agyhalálhoz kapcsolódó cardiovascularis hatások vizsgálata során
igazoltuk, hogy az L-arginin szignifikánsan csökkenti a cardialis és a
koszorúér diszfunkciót. Vizsgálatainkban az agyhalált kísérő
hemodinamikai jelenségek megfeleltek az irodalomban leírtaknak
(70,116,117); az agyhalál indukcióját követően egy Cushing-reakciónak
megfelelő állapot alakult ki, ami a szimpatikus idegek kisülései, valamint a
szimpatiko-adrenalis rendszer aktivációja és a következményes plazma
katekolamin-szint emelkedéssel magyarázható (116). Ezt az átmeneti
állapotot hemodinamikai instabilitás követte vérnyomáseséssel és
egyidejű hormon deplécióval. Ennek a második fázisnak a patofiziológiai

69
háttere még nem teljesen tisztázott. Számos szerző szerint a katekolamin-
indukálta primer szívkárosodások és az ezt követő hormon-depléció a
felelős a gyakran megfigyelt hemodinamikai összeomlásért (118,119).
Ezzel szemben néhány újabb tanulmányban kimutatható volt, hogy az
agyhalált követően kialakuló hemodinamikai instabilitást inkább a
megváltozott előterhelés okozhatja, nem pedig a primer szívfunkció romlás
(70,116,117,120). Kísérleteinkben az agyhalál indukciója átmeneti
coronariaáramlás emelkedést okozott az akut fázis során, majd az
áramlás a kiindulási értékek alá csökkent. Ez a megfigyelés összhangban
van korábbi vizsgálatok eredményeivel, melyek során az agyhalált követő
myocardialis véráramlást indirekt módon mérték patkány és disznó
modellekben (121,122). Az még azonban nem egyértelmű, mekkora
szerepe lehet a csökkent koszorúér áramlásnak a szívfunkció romlásában.
Korábban, in situ izolált szívmodellen kimutatták, hogy a csökkent
coronaria perfúziós nyomás és coronariaáramlás az ún. „perfúzió-
kontraktilitás összefüggés” alapján lecsökkentheti a myocardialis
kontraktilitást. Így a coronaria perfúziós nyomást követő, falfeszülés-függő
szisztolés funkció változás jöhet létre, amit „locsolócső effektusnak” hívnak
(123). Úgy tűnik viszont, hogy a csökkent coronaria perfúziós nyomás nem
okoz globális ischaemiát agyhalált követően (121,122,123). Korábbi
vizsgálatok bizonyították, hogy az agyhalál súlyos endothelialis
diszfunkciót okoz. Emellett azonban azt is kimutatták, hogy az
endothelium még továbbra is képes volna NO termelésére, de
valószínűleg a NO szintézis lehetséges substratjainak mennyisége is
számottevően lecsökkent (69). Ennek a feltételezésnek igazolására,
kísérleteinkben mértük a NO prekurzor L-arginin és a NO szintézis indirekt
mutatóiként ismert nitrit és nitrát (nitrit&nitrát) koncentrációkat a
plazmában. Várakozásunkkal ellentétben, az agyhalál beállta nem
befolyásolta a NO anyagcsere ezen mutatóit. Ennek alapján úgy tűnik,
hogy nem a csökkent substrat mennyiség (L-arginin) az endothelialis
diszfunkció legfőbb oka. Az agyhalál során kialakuló direkt endothel-
sérülés lehetősége még nem tisztázott (122), azonban a kísérletek során

70
megfigyelt humoralis változások endothelialis diszfunkcióra gyakorolt
hatása sem hagyható figyelmen kívül. Irodalmi adatok alapján pl. a
katekolaminok gátolhatják az endothelialis relaxációban résztvevő
anyagokat (NO) anélkül is, hogy endothelialis termelődésüket vagy az
erek simaizmán kifejtett hatásukat csökkentenék (124). Hormon depléció,
különösen a pajzsmirigy hormonok csökkenése, szintén befolyásolhatja az
endothelium-dependens vasorelaxatiót agyhalál során, amint ezt korábbi
kísérletek igazolták (72). Más vizsgálatok szerint az agyhalál gyulladásos
reakciókat is beindíthat, melyek során a leukocytákban, illetve a
cardiomyocytákban aktiválódhat az iNOS (125). Az iNOS substratja az L-
arginin, ebből NO-t szintetizál, ami viszont a szabad gyök képződés
kiinduló pontja, és így - különösen a peroxynitrit képződés során - további
szöveti károsodásokat okozhat. Eredményeink alapján felvetődik, hogy az
eNOS és az iNOS versenyezhet egymással a felhasználható L-arginin
készletekért. Vizsgálatainkban a változatlan szintű nitrit&nitrát termelés az
összes NO szintézist jellemzi, miközben az endothelialis NO szintézis akár
szignifikánsan csökkent is lehet. Továbbá, a rendelkezésre álló L-arginin
mennyisége befolyásolhatja az eNOS aktivitást is; tehát az L-arginin
mennyiségének csökkenésével tovább csökkenhet az eNOS aktivitás.
Ezek alapján, az agyhalál beállta után a lecsökkent eNOS aktivitás a
továbbiakban csökkent „vascularis NO” termeléshez vezethet,
mindeközben azonban megnőhet az iNOS aktivitása, és így a termelt
„gyulladásos NO” mennyissége is. Vizsgálataink során demonstráltuk,
hogy az agyhalál súlyos, de reverzibilis endothelialis diszfunkcióhoz vezet,
amit L-arginin adásával vissza lehet fordítani, sőt L-arginin hatására a
kontraktilitás javulása is várható. Ezek alapján az L-arginin alkalmazható
lehet szívdonorok esetében az endothelialis funkció javítására, és így
szerepet játszhat a lehetséges szervdonorok számának növelésében,
illetve a transzplantációk rövid- és hosszú távú kimenetelének javításában.

71
8 Következtetések
A myocardialis cardioprotectio lehetőségeit human és experimentalis
modelleken vizsgálva demonstráltuk, hogy vasoactiv ágensek ischaemia
során mind a lokális, mind a szisztémás szintű védekező
mechanizmusokban fontos szerepet játszhatnak.
A pericardialis folyadék ANP koncentrációs viszonyainak nagy
beteganyagon végzett vizsgálata során megállapítottuk, hogy a
pericardialis folyadékban egy nagyságreddel magasabb ANP szintek
mérhetők, mint a plazmában. Ezt a különbséget részben okozhatja
myocardialis ischaemia, azonban a jelenség klinikai jelentősége
megfontolandó. A pitvari és kamrai szöveti ANP koncentrációk között több
mint két nagyságrendnyi különbséget tapasztaltunk. Továbbá kimutattuk,
hogy a nők pitvari szívizomszövetében magasabb ANP koncentrációk
mérhetők, mint a férfiakéban, ami felveti a két nem közötti nátriuretikus
peptid regulációs folyamatok különbözőségét. Ellentétben a plazma ANP
koncentrációval, az intrapericardialis ANP szint (ami jól jellemezheti az
interstitialis ANP koncentrációt) nem korrelál a szívfunkció csökkenésének
súlyosságával, ami felveti az ANP lokálisan eltérő – parakrin - regulációs
szerepét a pericardialis folyadékban. Eredményeink alapján az interstitialis
(parakrin) és a keringésben megtalálható (endokrin) ANP biológiai
funkciója valószínűleg eltérő, azonban a szabályozás e két eleme egy
közös és egymást jól kiegészítő élettani rendszert alkot.
Az interstitialis folyadék adenin nukleozid koncentrációjának
meghatározására matematikai modellt alkottunk, melynek alapja, hogy
kisméretű molekulák esetében a pericardialis folyadék jól tükrözi az
interstitialis kompartment koncentrációviszonyait. Modellünkkel a
plazmában és a pericardialis folyadékban mért koncentrációk alapján
megbecsülhetők az adenin-nukleozidok interstitialis szintjei. Matematikai
modellünk a későbbiek során alapul szolgálhat más molekulák
kompartmentális eloszlásának vizsgálatára is, amennyiben a vizsgált

72
anyag metabolizációjáról és transzport folyamatairól kellő ismeretekkel
rendelkezünk.
Parathormonnal végzett kísérleteinkben a szer hatására elhúzódó,
dózisfüggően növekedő coronariaáramlást tapasztaltunk. Megállapítottuk,
hogy a KATP csatornát specifikusan blokkoló glibenklamid hatására a PTH
által kiváltott koszorúér tágulat jelentősen és szignifikánsan csökkent.
Eredményeink valószínűsítik az ér simaizomsejt KATP csatornáinak
jelentős szerepét a PTH okozta coronaria dilatáció létrejöttében.
Elképzelhetőnek tartjuk azt is, hogy a PTH koszorúértágító hatása során
az irodalomban eddig közölt cAMP rendszert és L-típusú Ca csatorna
ínaktivációt involváló reakcióutak mellett a KATP csatornák integratív
kulcsszerepet kapnak a peptid cellularis hatásmechanizmusának
kibontakozásában.
A NO prekurzor L-arginin cardiovascularis protektív hatásának
experimentalis vizsgálata során demonstráltuk, hogy L-arginin adásával
csökkenhet a reperfúziós károsodás és javulhat az endothelialis funkció
hypothermiás ischaemiát és reperfúziót követően is. A NO prekurzorok
adása új koncepció a cardioprotectio terén, és első sorban a reperfúziós
patofiziológiás folyamatok megelőzésére szolgál. Vizsgálatainkban az L-
arginin reperfúzió során még szisztémásan adva is javította a
szívműködést. Az L-arginin intravénás adásának további előnye lehet,
hogy az érrendszer távolabbi pontjain is javítja az endothelfunkciót. Ezáltal
a NO prekurzor szerepe a cardioprotectio lokális – szervi – szintjéről
globális – egész szervezetet érintő – szintre emelkedhet.
Emellett kimutattuk, hogy az agyhalál jelenségéhez kapcsolódó súlyos
endothelialis diszfunkció kialakulása megakadályozható L-arginin
alkalmazásával, sőt L-arginin hatására nő a kontraktilitás és általános
hemodinamikai javulás érhető el. Megfigyeléseink alapján lehetőség nyílik
a transzplantációs donorok endothelialis funkciójának javítására, és ezáltal
a donorkapacitás növelésére.

73
9 Irodalomjegyzék
(1) Levin ER, Gardner DG, Samson WK: Natriuretic peptides. N Engl
J Med 1998;339:321-328.
(2) Yoshimura M, Yasue H, Okumura K, Ogawa H, Jougasaki M,
Mukoyama M, Nakao K, Imura H: Different secretion patterns of
atrial natriuretic peptide and brain natriuretic peptide in patients
with congestive heart failure. Circulation 1993;87:464-469.
(3) de Bold AJ, Borenstein HB, Veress AT, Sonnenberg H: A rapid
and potent natriuretic response to intravenous injection of atrial
myocardial extract in rats. Life Sci 1981;28:89-94.
(4) Drexler H, Hanze J, Finckh M, Lu W, Just H, Lang RE: Atrial
natriuretic peptide in a rat model of cardiac failure. Atrial and
ventricular mRNA, atrial content, plasma levels, and effect of
volume loading. Circulation 1989;79:620-633.
(5) Ruskoaho H: Atrial natriuretic peptide: synthesis, release, and
metabolism. Pharmacol Rev 1992;44:479-602.
(6) Buckley MG, Sagnella GA, Markandu ND, Singer DR,
MacGregor GA: Concentrations of N-terminal ProANP in human
plasma: evidence for ProANP (1-98) as the circulating form. Clin
Chim Acta 1990;191:1-14.
(7) Schulz-Knappe P, Forssmann K, Herbst F, Hock D, Pipkorn R,
Forssmann WG: Isolation and structural analysis of "urodilatin", a

74
new peptide of the cardiodilatin-(ANP)-family, extracted from
human urine. Klin Wochenschr 1988;66:752-759.
(8) Deutsch A, Frishman WH, Sukenik D, Somer BG, Youssri A:
Atrial natriuretic peptide and its potential role in
pharmacotherapy. J Clin Pharmacol 1994;34:1133-1147.
(9) Edwards BS, Zimmerman RS, Schwab TR, Heublein DM, Burnett
JC, Jr.: Atrial stretch, not pressure, is the principal determinant
controlling the acute release of atrial natriuretic factor. Circ Res
1988;62:191-195.
(10) Ruskoaho H, Leskinen H, Magga J, Taskinen P, Mantymaa P,
Vuolteenaho O, Leppaluoto J: Mechanisms of mechanical load-
induced atrial natriuretic peptide secretion: role of endothelin,
nitric oxide, and angiotensin II. J Mol Med 1997;75:876-885.
(11) Sagnella GA, Markandu ND, Shore AC, MacGregor GA: Raised
circulating levels of atrial natriuretic peptides in essential
hypertension. Lancet 1986;1:179-181.
(12) Burnett JC, Jr., Kao PC, Hu DC, Heser DW, Heublein D, Granger
JP, Opgenorth TJ, Reeder GS: Atrial natriuretic peptide elevation
in congestive heart failure in the human. Science 1986;231:1145-
1147.
(13) Lerman A, Gibbons RJ, Rodeheffer RJ, Bailey KR, McKinley LJ,
Heublein DM, Burnett JC, Jr.: Circulating N-terminal atrial
natriuretic peptide as a marker for symptomless left-ventricular
dysfunction. Lancet 1993;341:1105-1109.
(14) Morita E, Yasue H, Yoshimura M, Ogawa H, Jougasaki M,
Matsumura T, Mukoyama M, Nakao K: Increased plasma levels

75
of brain natriuretic peptide in patients with acute myocardial
infarction. Circulation 1993;88:82-91.
(15) Horkay F, Szokodi I, Selmeci L, Merkely B, Kekesi V, Vecsey T,
Vuolteenaho O, Ruskoaho H, Juhasz-Nagy A, Toth M: Presence
of immunoreactive endothelin-1 and atrial natriuretic peptide in
human pericardial fluid. Life Sci 1998;62:267-274.
(16) Gibson AT, Segal MB: A study of the composition of pericardial
fluid, with special reference to the probable mechanism of fluid
formation. J Physiol 1978;277:367-377.
(17) Szentivanyi M, Magyar Zs, and Juhasz-Nagy A.: The
physiological role of the epicardium. Acta Physiol Hun
1973;44:403-404.
(18) Amano J, Suzuki A, Sunamori M, Shichiri M, Marumo F: Atrial
natriuretic peptide in the pericardial fluid of patients with heart
disease. Clin Sci (Colch ) 1993;85:165-168.
(19) Klemola R, Laine M, Weckstrom M, Vuolteenaho O, Ruskoaho
H, Huttunen P, Leppaluoto J: High concentrations of atrial
natriuretic peptide and brain natriuretic peptide in rat pericardial
fluid and their reduction by reserpine in vivo. Naunyn
Schmiedebergs Arch Pharmacol 1995;352:331-336.
(20) Selmeci L, Antal M, Horkay F, Merkely B, Szokodi I, Biro L,
Szekely M, Jobbagy J, Szepvolgyi J, Toth M: Enhanced
accumulation of pericardial fluid ferritin in patients with coronary
artery disease. Coron Artery Dis 2000;11:53-56.
(21) Horkay F, Laine M, Szokodi I, Leppaluoto J, Vuolteenaho O,
Ruskoaho H, Juhasz-Nagy A, Toth M: Human pericardial fluid

76
contains the highest amount of endothelin-1 of all mammalian
biologic fluids thus far tested. J Cardiovasc Pharmacol 1995;26
Suppl 3:S502-S504.
(22) Fazekas L, Horkay F, Kekesi V, Huszar E, Barat E, Fazekas R,
Szabo T, Juhasz-Nagy A, Naszlady A: Enhanced accumulation
of pericardial fluid adenosine and inosine in patients with
coronary artery disease. Life Sci 1999;65:1005-1012.
(23) Mebazaa A, Wetzel RC, Dodd-o JM, Redmond EM, Shah AM,
Maeda K, Maistre G, Lakatta EG, Robotham JL: Potential
paracrine role of the pericardium in the regulation of cardiac
function. Cardiovasc Res 1998;40:332-342.
(24) Juhasz-Nagy A, Kekesi V, Fazekas L, Merkely B, Toth M:
Uneven flow distribution in the heart induced by endothelin. Adv
Exp Med Biol 1999;471:247-256.
(25) Geller L, Merkely B, Szokodi I, Szabo T, Vecsey T, Juhasz-Nagy
A, Toth M, Horkay F: Electrophysiological effects of
intrapericardial infusion of endothelin- 1. Pacing Clin
Electrophysiol 1998;21:151-156.
(26) Szokodi I, Horkay F, Merkely B, Solti F, Geller L, Kiss P, Selmeci
L, Kekesi V, Vuolteenaho O, Ruskoaho H, Juhasz-Nagy A, Toth
M: Intrapericardial infusion of endothelin-1 induces ventricular
arrhythmias in dogs. Cardiovasc Res 1998;38:356-364.
(27) Kuh JH, Kim KS, Kim SH, Cho KW, Seul KH, Koh GY: Presence
of immunoreactive atrial natriuretic peptides in pericardial fluid of
human subjects with congenital heart diseases. Life Sci
1990;46:1977-1983.

77
(28) Szokodi I, Horkay F, Kiss P, Selmeci L, Merkely B, Kekesi V,
Vuolteenaho O, Leppaluoto J, Ruskoaho H, Juhasz-Nagy A, Toth
M: Characterization and stimuli for production of pericardial fluid
atrial natriuretic peptide in dogs. Life Sci 1997;61:1349-1359.
(29) Kroll K, Deussen A, Sweet IR: Comprehensive model of transport
and metabolism of adenosine and S-adenosylhomocysteine in
the guinea pig heart. Circ Res 1992;71:590-604.
(30) Wangler RD, Gorman MW, Wang CY, DeWitt DF, Chan IS,
Bassingthwaighte JB, Sparks HV: Transcapillary adenosine
transport and interstitial adenosine concentration in guinea pig
hearts. Am J Physiol 1989;257:H89-106.
(31) Hanley F, Messina LM, Baer RW, Uhlig PN, Hoffman JI: Direct
measurement of left ventricular interstitial adenosine. Am J
Physiol 1983;245:H327-H335.
(32) Sparks HV, Jr., Bardenheuer H: Regulation of adenosine
formation by the heart. Circ Res 1986;58:193-201.
(33) Lloyd HG, Schrader J: Adenosine metabolism in the guinea pig
heart: the role of cytosolic S-adenosyl-L-homocysteine
hydrolase, 5'-nucleotidase and adenosine kinase. Eur Heart J
1993;14 Suppl I:27-33.
(34) Mubagwa K, Mullane K, Flameng W: Role of adenosine in the
heart and circulation. Cardiovasc Res 1996;32:797-813.
(35) Crass MF, III, Moore PL, Strickland ML, Pang PK, Citak MS:
Cardiovascular responses to parathyroid hormone. Am J Physiol
1985;249:E187-E194.

78
(36) Crass MF, III, Pang PK: Parathyroid hormone: a coronary artery
vasodilator. Science 1980;207:1087-1089.
(37) Ogino K, Burkhoff D, Bilezikian JP: The hemodynamic basis for
the cardiac effects of parathyroid hormone (PTH) and PTH-
related protein. Endocrinology 1995;136:3024-3030.
(38) Crass MF, III, Jayaseelan CL, Darter TC: Effects of parathyroid
hormone on blood flow in different regional circulations. Am J
Physiol 1987;253:R634-R639.
(39) Nickols GA, Metz MA, Cline WH, Jr.: Endothelium-independent
linkage of parathyroid hormone receptors of rat vascular tissue
with increased adenosine 3',5'-monophosphate and relaxation of
vascular smooth muscle. Endocrinology 1986;119:349-356.
(40) Pang PK, Tenner TE, Jr., Yee JA, Yang M, Janssen HF:
Hypotensive action of parathyroid hormone preparations on rats
and dogs. Proc Natl Acad Sci U S A 1980;77:675-678.
(41) Yang MC, Tenner TE, Jr., Pang PK: Lack of histamine
involvement in parathyroid hormone hypotensive action.
Pharmacology 1981;22:305-310.
(42) Nickols GA: Increased cyclic AMP in cultured vascular smooth
muscle cells and relaxation of aortic strips by parathyroid
hormone. Eur J Pharmacol 1985;116:137-144.
(43) Pang PK, Yang MC, Sham JS: Parathyroid hormone and calcium
entry blockade in a vascular tissue. Life Sci 1988;42:1395-1400.

79
(44) Wang R, Karpinski E, Pang PK: Parathyroid hormone selectively
inhibits L-type calcium channels in single vascular smooth
muscle cells of the rat. J Physiol 1991;441:325-346.
(45) Aversano T, Ouyang P, Silverman H: Blockade of the ATP-
sensitive potassium channel modulates reactive hyperemia in the
canine coronary circulation. Circ Res 1991;69:618-622.
(46) Komaru T, Lamping KG, Eastham CL, Dellsperger KC: Role of
ATP-sensitive potassium channels in coronary microvascular
autoregulatory responses. Circ Res 1991;69:1146-1151.
(47) Mori H, Chujo M, Tanaka E, Yamakawa A, Shinozaki Y,
Mohamed MU, Nakazawa H: Modulation of adrenergic coronary
vasoconstriction via ATP-sensitive potassium channel. Am J
Physiol 1995;268:H1077-H1085.
(48) Fazekas L, Soos P, Fazekas R, Kekesi V, Juhasz-Nagy A: Role
of K+ATP channels in the metabolic adaptation of the coronaries.
Adv Exp Med Biol 1999;471:239-245.
(49) Boger RH, Bode-Boger SM: The clinical pharmacology of L-
arginine. Annu Rev Pharmacol Toxicol 2001;41:79-99.
(50) Andrew PJ, Mayer B: Enzymatic function of nitric oxide
synthases. Cardiovasc Res 1999;43:521-531.
(51) Hishikawa K, Nakaki T, Suzuki H, Kato R, Saruta T: L-arginine as
an antihypertensive agent. J Cardiovasc Pharmacol 1992;20
Suppl 12:S196-S197.

80
(52) Palmer RM, Ferrige AG, Moncada S: Nitric oxide release
accounts for the biological activity of endothelium-derived
relaxing factor. Nature 1987;327:524-526.
(53) Gattullo D, Pagliaro P, Marsh NA, Losano G: New insights into
nitric oxide and coronary circulation. Life Sci 1999;65:2167-2174.
(54) Richard V, Kaeffer N, Tron C, Thuillez C: Ischemic
preconditioning protects against coronary endothelial dysfunction
induced by ischemia and reperfusion. Circulation 1994;89:1254-
1261.
(55) Vegh A, Szekeres L, Parratt J: Preconditioning of the ischaemic
myocardium;involvement of the L-arginine nitric oxide pathway.
Br J Pharmacol 1992;107:648-652.
(56) Gattullo D, Pagliaro P, Linden RJ, Merletti A, Losano G: The role
of nitric oxide in the initiation and in the duration of some
vasodilator responses in the coronary circulation. Pflugers Arch
1995;430:96-104.
(57) Maxwell AJ, Schauble E, Bernstein D, Cooke JP: Limb blood flow
during exercise is dependent on nitric oxide. Circulation
1998;98:369-374.
(58) Moncada S, Higgs A: The L-arginine-nitric oxide pathway. N Engl
J Med 1993;329:2002-2012.
(59) Vallance P, Chan N: Endothelial function and nitric oxide: clinical
relevance. Heart 2001;85:342-350.

81
(60) Gewaltig MT, Kojda G: Vasoprotection by nitric oxide:
mechanisms and therapeutic potential. Cardiovasc Res
2002;55:250-260.
(61) Argano V, Galinanes M, Edmondson S, Hearse DJ: Effects of
cardioplegia on vascular function and the "no-reflow"
phenomenon after ischemia and reperfusion: studies in the
isolated blood-perfused rat heart. J Thorac Cardiovasc Surg
1996;111:432-441.
(62) Murphy CO, Pan C, Gott JP, Guyton RA: Microvascular reactivity
after crystalloid, cold blood, and warm blood cardioplegic arrest.
Ann Thorac Surg 1995;60:1021-1027.
(63) Szabo G, Bahrle S, Batkai S, Stumpf N, Dengler TJ,
Zimmermann R, Vahl CF, Hagl S: L-arginine: effect on
reperfusion injury after heart transplantation. World J Surg
1998;22:791-797.
(64) Pabla R, Buda AJ, Flynn DM, Salzberg DB, Lefer DJ:
Intracoronary nitric oxide improves postischemic coronary blood
flow and myocardial contractile function. Am J Physiol
1995;269:H1113-H1121.
(65) Siegfried MR, Erhardt J, Rider T, Ma XL, Lefer AM:
Cardioprotection and attenuation of endothelial dysfunction by
organic nitric oxide donors in myocardial ischemia-reperfusion. J
Pharmacol Exp Ther 1992;260:668-675.
(66) Schulz R, Wambolt R: Inhibition of nitric oxide synthesis protects
the isolated working rabbit heart from ischaemia-reperfusion
injury. Cardiovasc Res 1995;30:432-439.

82
(67) Treasure CB, Vita JA, Ganz P, Ryan TJ, Jr., Schoen FJ,
Vekshtein VI, Yeung AC, Mudge GH, Alexander RW, Selwyn AP,
.: Loss of the coronary microvascular response to acetylcholine
in cardiac transplant patients. Circulation 1992;86:1156-1164.
(68) Day JD, Rayburn BK, Gaudin PB, Baldwin WM, III, Lowenstein
CJ, Kasper EK, Baughman KL, Baumgartner WA, Hutchins GM,
Hruban RH: Cardiac allograft vasculopathy: the central
pathogenetic role of ischemia-induced endothelial cell injury. J
Heart Lung Transplant 1995;14:S142-S149.
(69) Szabo G, Buhmann V, Bahrle S, Vahl CF, Hagl S: Brain death
impairs coronary endothelial function. Transplantation
2002;73:1846-1848.
(70) Herijgers P, Borgers M, Flameng W: The effect of brain death on
cardiovascular function in rats. Part I. Is the heart damaged?
Cardiovasc Res 1998;38:98-106.
(71) Shanlin RJ, Sole MJ, Rahimifar M, Tator CH, Factor SM:
Increased intracranial pressure elicits hypertension, increased
sympathetic activity, electrocardiographic abnormalities and
myocardial damage in rats. J Am Coll Cardiol 1988;12:727-736.
(72) Vargas F, Fernandez-Rivas A, Garcia EJ, Garcia dR:
Endothelium-dependent and endothelium-independent
vasodilation in hyperthyroid and hypothyroid rats. Pharmacology
1995;51:308-314.
(73) Szabo G, Hackert T, Sebening C, Vahl CF, Hagl S: Modulation of
coronary perfusion pressure can reverse cardiac dysfunction
after brain death. Ann Thorac Surg 1999;67:18-25.

83
(74) Mendes Ribeiro AC, Brunini TM, Ellory JC, Mann GE:
Abnormalities in L-arginine transport and nitric oxide biosynthesis
in chronic renal and heart failure. Cardiovasc Res 2001;49:697-
712.
(75) Morrow DA, Braunwald E: Future of biomarkers in acute
coronary syndromes: moving toward a multimarker strategy.
Circulation 2003;108:250-252.
(76) Ruskoaho H: Cardiac hormones as diagnostic tools in heart
failure. Endocr Rev 2003;24:341-356.
(77) Cho KW, Seul KH, Kim SH, Ryu H, Seul KM, Koh GY: Epicardial
release of immunoreactive atrial natriuretic peptides in inside-out
perfused rabbit atria. Biochem Biophys Res Commun
1988;153:811-817.
(78) Turbucz P, Kiss P, Horkay F, Szokodi I, deChatel R, Selmeci L,
Juhasz-Nagy A, Karadi I, Toth M: High pericardial fluid levels of
endothelin are not caused by altered neutral endopeptidase
activity in cardiac patients. J Cardiovasc Pharmacol 1998;31
Suppl 1:S287-S289.
(79) Bloch KD, Seidman JG, Naftilan JD, Fallon JT, Seidman CE:
Neonatal atria and ventricles secrete atrial natriuretic factor via
tissue-specific secretory pathways. Cell 1986;47:695-702.
(80) Toth P, Toth M, Lang RE, Juhasz-Nagy A: Left atrial distension
induces coronary vasodilation independently of cardiac ANP
release. J Hypertension 1990;3:88A
(81) Supaporn T, Wennberg PW, Wei CM, Kinoshita M, Matsuda Y,
Burnett JC: Role for the endogenous natriuretic peptide system

84
in the control of basal coronary vascular tone in dogs. Clin Sci
(Colch ) 1996;90:357-362.
(82) Klemola R, Tikkanen I, Vuolteenaho O, Toivonen L, Laine M:
Plasma and pericardial fluid natriuretic peptide levels in
postinfarction ventricular dysfunction. Eur J Heart Fail 2001;3:21-
26.
(83) Vesely DL: Atrial natriuretic peptides in pathophysiological
diseases. Cardiovasc Res 2001;51:647-658.
(84) Schwertz DW, Penckofer S: Sex differences and the effects of
sex hormones on hemostasis and vascular reactivity. Heart Lung
2001;30:401-426.
(85) Maffei S, Del Ry S, Prontera C, Clerico A: Increase in circulating
levels of cardiac natriuretic peptides after hormone replacement
therapy in postmenopausal women. Clin Sci (Lond)
2001;101:447-453.
(86) Kuroski de Bold ML: Estrogen, natriuretic peptides and the renin-
angiotensin system. Cardiovasc Res 1999;41:524-531.
(87) Pelzer T, Shamim A, Wolfges S, Schumann M, Neyses L:
Modulation of cardiac hypertrophy by estrogens. Adv Exp Med
Biol 1997;432:83-89.
(88) Juhasz-Nagy A, Papp L: Are coronary vasodilator effects of
inosine and adenosine independent of each other? J Cardiovasc
Pharmacol 1982;4:330-331.

85
(89) Moser GH, Schrader J, Deussen A: Turnover of adenosine in
plasma of human and dog blood. Am J Physiol 1989;256:C799-
C806.
(90) Kroll K, Stepp DW: Adenosine kinetics in canine coronary
circulation. Am J Physiol 1996;270:H1469-H1483.
(91) Gao P, Scheibel S, D'Amour P, John MR, Rao SD, Schmidt-
Gayk H, Cantor TL: Development of a novel immunoradiometric
assay exclusively for biologically active whole parathyroid
hormone 1-84: implications for improvement of accurate
assessment of parathyroid function. J Bone Miner Res
2001;16:605-614.
(92) Quiros RM, Valentin C, DeCresce R, Prinz RA: Intraoperative
total serum calcium levels, unlike intraoperative intact PTH
levels, do not correlate with cure of hyperparathyroidism. J Surg
Res 2003;114:57-63.
(93) Schluter KD, Piper HM: Cardiovascular actions of parathyroid
hormone and parathyroid hormone-related peptide. Cardiovasc
Res 1998;37:34-41.
(94) McDaniel NL, Rembold CM, Murphy RA: Cyclic nucleotide
dependent relaxation in vascular smooth muscle. Can J Physiol
Pharmacol 1994;72:1380-1385.
(95) Pang PK, Wang R, Shan J, Karpinski E, Benishin CG: Specific
inhibition of long-lasting, L-type calcium channels by synthetic
parathyroid hormone. Proc Natl Acad Sci U S A 1990;87:623-
627.

86
(96) Beguin P, Nagashima K, Nishimura M, Gonoi T, Seino S: PKA-
mediated phosphorylation of the human K(ATP) channel:
separate roles of Kir6.2 and SUR1 subunit phosphorylation.
EMBO J 1999;18:4722-4732.
(97) Wellman GC, Quayle JM, Standen NB: ATP-sensitive K+
channel activation by calcitonin gene-related peptide and protein
kinase A in pig coronary arterial smooth muscle. J Physiol
1998;507 ( Pt 1):117-129.
(98) Philbrick WM, Wysolmerski JJ, Galbraith S, Holt E, Orloff JJ,
Yang KH, Vasavada RC, Weir EC, Broadus AE, Stewart AF:
Defining the roles of parathyroid hormone-related protein in
normal physiology. Physiol Rev 1996;76:127-173.
(99) Cooke JP, Andon NA, Girerd XJ, Hirsch AT, Creager MA:
Arginine restores cholinergic relaxation of hypercholesterolemic
rabbit thoracic aorta. Circulation 1991;83:1057-1062.
(100) Angdin M, Settergren G, Liska J, Astudillo R: No effect of L-
arginine supplementation on pulmonary endothelial dysfunction
after cardiopulmonary bypass. Acta Anaesthesiol Scand
2001;45:441-448.
(101) Szabo G, Batkai S, Bahrle S, Dengler TJ, Vahl CF, Zimmermann
R, Hagl S: Effects of nitric oxide synthesis on reperfusion injury
and catecholamine responsiveness in a heterotopic rat heart-
transplantation model. J Cardiovasc Pharmacol 1998;31:221-
230.
(102) Suematsu Y, Ohtsuka T, Hirata Y, Maeda K, Imanaka K,
Takamoto S: L-Arginine given after ischaemic preconditioning

87
can enhance cardioprotection in isolated rat hearts. Eur J
Cardiothorac Surg 2001;19:873-879.
(103) Hatsuoka S, Sakamoto T, Stock UA, Nagashima M, Mayer JE,
Jr.: Effect of L-arginine or nitroglycerine during deep hypothermic
circulatory arrest in neonatal lambs. Ann Thorac Surg
2003;75:197-203.
(104) Wang QD, Morcos E, Wiklund P, Pernow J: L-arginine enhances
functional recovery and Ca(2+)-dependent nitric oxide synthase
activity after ischemia and reperfusion in the rat heart. J
Cardiovasc Pharmacol 1997;29:291-296.
(105) Pabla R, Buda AJ, Flynn DM, Blesse SA, Shin AM, Curtis MJ,
Lefer DJ: Nitric oxide attenuates neutrophil-mediated myocardial
contractile dysfunction after ischemia and reperfusion. Circ Res
1996;78:65-72.
(106) Ferdinandy P, Schulz R: Nitric oxide, superoxide, and
peroxynitrite in myocardial ischaemia-reperfusion injury and
preconditioning. Br J Pharmacol 2003;138:532-543.
(107) Cohen G, Borger MA, Weisel RD, Rao V: Intraoperative
myocardial protection: current trends and future perspectives.
Ann Thorac Surg 1999;68:1995-2001.
(108) Carrier M, Pellerin M, Perrault LP, Bouchard D, Page P, Searle
N, Lavoie J: Cardioplegic arrest with L-arginine improves
myocardial protection: results of a prospective randomized
clinical trial. Ann Thorac Surg 2002;73:837-841.

88
(109) Jiang Z, Kejian H, Lei CY, Gu K: Effects of L-arginine
cardioplegia on myocardium. J Extra Corpor Technol
2001;33:10-14.
(110) Kronon MT, Allen BS, Halldorsson A, Rahman S, Wang T, Ilbawi
M: Dose dependency of L-arginine in neonatal myocardial
protection: the nitric oxide paradox. J Thorac Cardiovasc Surg
1999;118:655-664.
(111) Amrani M, Gray CC, Smolenski RT, Goodwin AT, London A,
Yacoub MH: The effect of L-arginine on myocardial recovery
after cardioplegic arrest and ischemia under moderate and deep
hypothermia. Circulation 1997;96:274-279.
(112) Bode-Boger SM, Boger RH, Creutzig A, Tsikas D, Gutzki FM,
Alexander K, Frolich JC: L-arginine infusion decreases peripheral
arterial resistance and inhibits platelet aggregation in healthy
subjects. Clin Sci (Lond) 1994;87:303-310.
(113) Kelm M: Nitric oxide metabolism and breakdown. Biochim
Biophys Acta 1999;1411:273-289.
(114) Brunner F, Maier R, Andrew P, Wolkart G, Zechner R, Mayer B:
Attenuation of myocardial ischemia/reperfusion injury in mice
with myocyte-specific overexpression of endothelial nitric oxide
synthase. Cardiovasc Res 2003;57:55-62.
(115) Demers P, Elkouri S, Sirois MG, Cartier R: Effects of acute
rejection on L-arginine/iNOS pathway in canine heart
transplantation. Can J Cardiol 2003;19:419-425.
(116) Szabo G: Physiologic changes after brain death. J Heart Lung
Transplant 2004;23:S223-S226.

89
(117) Szabo G, Sebening C, Hackert T, Hagl C, Tochtermann U, Vahl
CF, Hagl S: Effects of brain death on myocardial function and
ischemic tolerance of potential donor hearts. J Heart Lung
Transplant 1998;17:921-930.
(118) Sebening C, Hagl C, Szabo G, Tochtermann U, Strobel G,
Schnabel P, Amann K, Vahl CF, Hagl S: Cardiocirculatory effects
of acutely increased intracranial pressure and subsequent brain
death. Eur J Cardiothorac Surg 1995;9:360-372.
(119) Bittner HB, Kendall SW, Campbell KA, Montine TJ, Van Trigt P:
A valid experimental brain death organ donor model. J Heart
Lung Transplant 1995;14:308-317.
(120) Szabo G, Hackert T, Buhmann V, Graf A, Sebening C, Vahl CF,
Hagl S: Downregulation of myocardial contractility via intact
ventriculo--arterial coupling in the brain dead organ donor. Eur J
Cardiothorac Surg 2001;20:170-176.
(121) Mertes PM, el Abassi K, Jaboin Y, Burtin P, Pinelli G, Carteaux
JP, Burlet C, Boulange M, Villemot JP: Changes in hemodynamic
and metabolic parameters following induced brain death in the
pig. Transplantation 1994;58:414-418.
(122) Herijgers P, Leunens V, Tjandra-Maga TB, Mubagwa K, Flameng
W: Changes in organ perfusion after brain death in the rat and its
relation to circulating catecholamines. Transplantation
1996;62:330-335.
(123) Sunagawa K, Maughan WL, Burkhoff D, Sagawa K: Left
ventricular interaction with arterial load studied in isolated canine
ventricle. Am J Physiol 1983;245:H773-H780.

90
(124) Rubanyi GM, Lorenz RR, Vanhoutte PM: Bioassay of
endothelium-derived relaxing factor(s): inactivation by
catecholamines. Am J Physiol 1985;249:H95-101.
(125) Wilhelm MJ, Pratschke J, Beato F, Taal M, Kusaka M, Hancock
WW, Tilney NL: Activation of the heart by donor brain death
accelerates acute rejection after transplantation. Circulation
2000;102:2426-2433.

91
10 Saját publikációk jegyzéke
Fazekas R, Soos P, Kekesi V, Fazekas L, Juhasz-Nagy S: A parathormon
hatása a koszorúerekre. Cardiologia Hungarica 2000;2:67-72.
Soos P: Emelkedett pitvari nátriuretikus peptid koncentráció a humán
pericardialis folyadéktérben: vizsgálatok szívsebészeti betegekben.
Cardiologia Hungarica 2001;2:147-151.
Soos P, Juhasz-Nagy A, Ruskoaho H, Hartyanszky I, Merkely B, Toth M,
Horkay F: Locally different role of atrial natriuretic peptide (ANP) in the
pericardial fluid. Life Sci 2002;71:2563-2573.
Fazekas R, Soos P, Kekesi V, Fazekas L, Juhasz-Nagy A: The coronary
effects of parathyroid hormone. Horm Res 2004;61:234-241.
Soos, P: Az állati szervek folyadékrendszereinek dinamizmusa:
kompartmentalizációs szívmodell. Magyar Biológiai Társaság
XXV.Vándorgyűlés előadások összefoglalói 2004;87-91.
Soos P, Andrasi T, Buhmann V, Kohl B, Vahl C, Hagl S, Szabo G:
Myocardial protection after systemic application of L-arginine during
reperfusion. J Cardiovasc Pharmacol 2004;43:782-788.
Szabo G, Soos P, Heger U, Mandera S, Buhmann V, Bahrle S, Kohl B,
Hagl S: L-arginine improves endothelial and myocardial function after brain
death. Transplantation 2006;82:108-112.

92
11 Köszönetnyilvánítás
Közel tíz éves kutató és klinikai munkám során nagyon sokan nyújtottak
számomra szakmai és emberi segítséget. Ez úton szeretném
köszönetemet kifejezni mindazoknak, akik lehetőségeik szerint, minden
eszközzel támogattak céljaim elérésében.
Mindenek előtt, köszönettel tartozom a családomnak; feleségemnek
kitartásáért és szeretettel teli támogatásáért, illetve kislányomnak,
Pankának a sok-sok mosolygós, boldog percért, amivel a dolgozatírás
közben lepett meg. Hálás köszönettel tartozom szüleimnek és
testvéremnek, akiktől olyan mindenre kiterjedő figyelmet és támogatást
kaptam, amit csak remélhetek, hogy egyszer viszonozni tudok.
Köszönetemet fejezem ki Dr. Horkay Ferenc professzor úrnak, az MTA
doktorának, témavezetőmnek, aki céljaimat a kezdetektől támogatta, és
gondoskodó figyelmét rajtam tartva irányította kutatói képzésemet, majd
pályaválasztásomat.
Hálásan köszönöm Dr. Juhász-Nagy Sándor professzor úrnak, az MTA
doktorának, hogy kutatói elképzeléseim fő irányát megadta, kezdeti
lépéseimet bátorította, és a világról alkotott képemet élettapasztalatával,
bölcs tanácsaival, filozofikus mélységű gondolataival a klasszikus erkölcsi
normák hagyományainak megfelelően alakította.
Köszönetemet fejezem ki Dr. Tóth Miklósnak, az MTA doktorának, aki
hasznos tanácsaival és a mindennapos munkát összességében átlátva
olyan önzetlen és biztos háttérnek bizonyult, akihez bármikor és bármilyen
kérdéssel fordulhattam segítségért.
Köszönettel tartozom Dr. Merkely Bélának, az MTA doktorának, aki a
kutatómunka során nyújtott hasznos segítsége mellett klinikai
pályafutásom első lépéseit is nagymértékben egyengette.
Köszönöm Dr. Szabó Gábornak, hogy támogatta és ösztönözte kutatói
elképzeléseimet, és értékes tanácsaival nélkülözhetetlen segítséget
nyújtott kutató munkám megvalósításához.

93
Köszönetemet fejezem ki Dr. Nemes Attila és Dr. Acsády György
professzor uraknak, az Ér-és Szívsebészeti Klinika korábbi, illetve jelen
igazgatóinak mind a klinikai, mind a kutatói munkám során nyújtott
támogatásért és segítségért.
Köszönettel tartozom a kutatásban résztvevő kollégáimnak; Dr. Andrási
Teréziának, Dr. Fazekas Rékának, Dr. Fazekas Leventének, Dr.
Hartyánszky Istvánnak, Dr. Kékesi Violettának, Dr. Seres Leilának, illetve
Prof. Dr. Siegfried Haglnak, Dr. Ulrike Hegernek, Dr. Brigitte Kohlnak, Dr.
Susanne Manderának, Prof. Dr. Heikki Ruskoahonak sokrétű, áldozatkész
segítségükért.
Köszönettel tartozom a Semmelweis Egyetem Ér- és Szívsebészeti
Klinikai valamennyi dolgozójának az évek alatt tanúsított segítségért.
Továbbá külön köszönöm közvetlen kollégáimnak; Dr. Apor Asztridnak,
Dr. Bárczi Györgynek, Dr. Bartha Elektrának, Dr. Becker Dávidnak, Dr.
Fülöp Gábornak, Dr. Gellér Lászlónak, Dr. Kiss Orsolyának, Dr. Kutyifa
Valentinának, Dr. Marozsán Ibolyának, Dr. Maurovich Horvat Pálnak, Dr.
Molnár Leventének, Dr. Róka Attilának, Prof. Dr. Selmeci Lászlónak, Dr.
Soltész Ádámnak, Dr. Szabó Györgynek, Dr. Szilágyi Szabolcsnak, Dr.
Szűcs Andreának, Dr. Szűcs Gábornak, Dr. Vágó Hajnalkának, Dr. Zima
Endrének és Sarusi Melindának önzetlen támogatását.
Hálás vagyok néhai Csongorné Rajczi Mártának, Fritz Gábornak, Juhász
Dórának, Juhász Gabriellának, Keresztes Istvánnének, Lantos Mónikának,
Ruska Zoltánnénak, Szabó Henriettnek, Szendrei Eszternek, illetve Karin
Sonnenbergnek, Nicole Stumpfnak a kísérleti munkák során végzett
szakszerű segítségéért.
Related Documents











