Vaccine 24 (2006) 501–509 Vaccination with irradiated Ancylostoma caninum third stage larvae induces a Th2 protective response in dogs Ricardo T. Fujiwara a,∗ , Alex Loukas a,b , Susana Mendez a , Angela L. Williamson a , Lilian L. Bueno a , Yan Wang a , Andre Samuel a , Bin Zhan a , Maria Elena Bottazzi a , Peter J. Hotez a , Jeffrey M. Bethony a,∗ a Department of Microbiology and Tropical Medicine, The George Washington University, 2300 Eye Street, Ross Hall Room 727, Medical Center, Washington, DC 20037, USA b Division of Infectious Diseases and Immunology, Queensland Institute of Medical Research, Qld 4006, Australia Received 28 January 2005; accepted 29 July 2005 Available online 15 August 2005 Abstract While X-irradiated live parasites are not an acceptable proposition for human vaccination, they offer a ready experimental system to explore mechanisms by which immunity against hookworm infection may be induced in humans. As such, we sought to further elucidate the details of this highly protective immune response induced by the irradiated vaccine in canids, with special emphasis on the cellular aspects of the response. Vaccination with irradiated L3 induced high production of antibodies and strong PBMCs proliferation to crude L3 antigen preparation. Elevated IL-4 production was also observed in vaccinated dogs, especially in relation to IFN- production (IL-4/IFN- ratio). Serum from vaccinated animals inhibited penetration of L3 through canine skin in vitro by 60%. Finally, vaccinated animals had a strong antibody response to ASP-2, a promising vaccine antigen that is an excretory–secretory product of L3. These results add further support the idea that the Th2 immune response is required to generate protective immunity against hookworm larvae and that ES molecules released during this developmental stage are likely targets of this response. © 2005 Elsevier Ltd. All rights reserved. Keywords: Vaccination; Hookworm; Irradiated larvae; Immune response; Th2; Dogs 1. Introduction Human hookworm infection is a soil-transmitted helminth infection caused by the nematode parasites Necator amer- icanus and Ancylostoma duodenale. It is one of the most common chronic infections of humans, with an estimated 740 million cases in areas of rural poverty in the tropics and subtropics [1]. Adult hookworms attach to the small intestine inducing blood loss from both direct consumption of blood by the parasite as well as ‘bleeding’ around the attachment site due to secretion of anticoagulants to promote feeding [2]. Hookworm infection is a major cause of intestinal blood ∗ Corresponding authors. Tel.: +1 202 994 2663; fax: +1 202 994 2913. E-mail addresses: [email protected] (R.T. Fujiwara), [email protected] (J.M. Bethony). loss in endemic areas, resulting in iron-deficiency anemia and protein malnutrition among these populations [3]. The major approach to hookworm control currently relies on administra- tion of benzimidazole (BZ) anthelmintic drugs [4]. However, the rapid re-infection after treatment [5,6], the diminishing efficacy of benzimidazoles with repeated use [7], and a grow- ing concern of rapid anthelmintic drug resistance against BZ compounds as seen in veterinary medicine [4] have made the successful development of an anti-hookworm vaccine an important public health priority [3]. A successful X-irradiated larval vaccine that induced up to 90% protection was developed commercially in the 1970s for use against the canine hookworm Ancylostoma caninum [8–13]. The failure of the vaccine to induce sterilizing immu- nity, its short-shelf life, and the cost of production let to its removal from market. While X-irradiated live parasites are 0264-410X/$ – see front matter © 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2005.07.091

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vaccine 24 (2006) 501–509
Vaccination with irradiatedAncylostoma caninum third stagelarvae induces a Th2 protective response in dogs
Ricardo T. Fujiwaraa,∗, Alex Loukasa,b, Susana Mendeza, Angela L. Williamsona,Lilian L. Buenoa, Yan Wanga, Andre Samuela, Bin Zhana, Maria Elena Bottazzia,
Peter J. Hoteza, Jeffrey M. Bethonya,∗a Department of Microbiology and Tropical Medicine, The George Washington University, 2300 Eye Street, Ross Hall Room 727,
Medical Center, Washington, DC 20037, USAb Division of Infectious Diseases and Immunology, Queensland Institute of Medical Research, Qld 4006, Australia
Received 28 January 2005; accepted 29 July 2005Available online 15 August 2005
Abstract
system toe cidate thed lar aspectso 3 antigenpS a stronga support thei les releasedd©
K
1
iic7sibs[
m
andrtra-
,
-t BZ
ee an
up70s
u-itsare
0d
While X-irradiated live parasites are not an acceptable proposition for human vaccination, they offer a ready experimentalxplore mechanisms by which immunity against hookworm infection may be induced in humans. As such, we sought to further eluetails of this highly protective immune response induced by the irradiated vaccine in canids, with special emphasis on the celluf the response. Vaccination with irradiated L3 induced high production of antibodies and strong PBMCs proliferation to crude Lreparation. Elevated IL-4 production was also observed in vaccinated dogs, especially in relation to IFN-� production (IL-4/IFN-� ratio).erum from vaccinated animals inhibited penetration of L3 through canine skin in vitro by 60%. Finally, vaccinated animals hadntibody response to ASP-2, a promising vaccine antigen that is an excretory–secretory product of L3. These results add further
dea that the Th2 immune response is required to generate protective immunity against hookworm larvae and that ES molecuuring this developmental stage are likely targets of this response.2005 Elsevier Ltd. All rights reserved.
eywords: Vaccination; Hookworm; Irradiated larvae; Immune response; Th2; Dogs
. Introduction
Human hookworm infection is a soil-transmitted helminthnfection caused by the nematode parasitesNecator amer-canus and Ancylostoma duodenale. It is one of the mostommon chronic infections of humans, with an estimated40 million cases in areas of rural poverty in the tropics andubtropics[1]. Adult hookworms attach to the small intestinenducing blood loss from both direct consumption of bloody the parasite as well as ‘bleeding’ around the attachmentite due to secretion of anticoagulants to promote feeding2]. Hookworm infection is a major cause of intestinal blood
∗ Corresponding authors. Tel.: +1 202 994 2663; fax: +1 202 994 2913.E-mail addresses: [email protected] (R.T. Fujiwara),
[email protected] (J.M. Bethony).
loss in endemic areas, resulting in iron-deficiency anemiaprotein malnutrition among these populations[3]. The majoapproach to hookworm control currently relies on administion of benzimidazole (BZ) anthelmintic drugs[4]. Howeverthe rapid re-infection after treatment[5,6], the diminishingefficacy of benzimidazoles with repeated use[7], and a growing concern of rapid anthelmintic drug resistance againscompounds as seen in veterinary medicine[4] have madthe successful development of an anti-hookworm vaccinimportant public health priority[3].
A successful X-irradiated larval vaccine that inducedto 90% protection was developed commercially in the 19for use against the canine hookwormAncylostoma caninum[8–13]. The failure of the vaccine to induce sterilizing immnity, its short-shelf life, and the cost of production let toremoval from market. While X-irradiated live parasites
264-410X/$ – see front matter © 2005 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2005.07.091

502 R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509
not an acceptable proposition for human vaccination, theyoffer a ready experimental system to explore mechanismsby which immunity may be induced in humans by vaccina-tion with recombinant vaccines[15]. A. caninum infectionresembles infection with the human hookworms in a numberof ways: (a) third stage infective larvae (L3) are acquired bytheir definitive host through skin penetration (N. americanusandA. duodenale) or oral digestion (A. duodenale), (b) larvaeundergo a short period of arrested development upon entryinto the host followed by a resumption of feeding and devel-opment in host tissues, and (c) the number of blood feedingadult worms in the gut are directly related to host pathol-ogy [14]. More importantly, however, larvae of both speciesrelease antigens (excretory–secretory products) upon entryinto the host that appear to be tightly coupled to the devel-opmental biology of the parasite, especially the transition toparasitism.
Vaccination studies in canine models using larvae atten-uated by ionizing radiation have successfully demonstrateda protective response after subsequent challenge with non-irradiated larvae. Information generated from the develop-ment of attenuated infective larval vaccine has led to thedesign and development of several ES molecules,Ancy-lostoma Secreted Proteins (ASP-1 and ASP-2) as candidatesfor human vaccines[3,15,16]. While the precise function ofthese ASPs remains unknown, the secretion of ASPs alsoa logyo nti-b ion ofl ucht n byt nts)[ orala d tot ine,w esep
2
2
w ain-t archF rec rsityA ina-t bloods
2
lar-v n-
trol (non-vaccinated animals). The vaccinated dogs receivedthree subcutaneous doses of 1000 irradiated L3 at inter-vals of 21 days. L3 were irradiated at 40 krad using a137Cs source (The George Washington University, Wash-ington, DC, USA). The control animals received Alum –AlK(SO4)2·12H2O – prepared as already described[19].The control animals used in this experiment were alsopart of another experiment using vaccines formulated withAlum.
Two weeks after the final vaccination, dogs were chal-lenged with 500A. caninum L3 by footpad infection methodas described elsewhere[19]. Each experimental group con-sisted of five animals.
2.3. Serological analysis
Blood was collected at least once every 21 days, and theserum was separated and stored frozen at−20◦C. Antigen-specific canine IgG1, IgG2 and IgE were measured byindirect enzyme-linked immunosorbent assay (ELISA) per-formed as described elsewhere[20]. Briefly, the antigenswere coated onto 96-well microplates (MaxiSorp
TM, Nalge
Nunc Intl., USA) at a concentration of 1�g/ml for L3 extractantigen and at 5�g/ml for the recombinant antigensNa-ASP-1[3,15], Ac-ASP-2[3,16] andAc-MTP-1 [3,18]. Serawere added at the concentrations 1:100, 1:500 and three-f ddi-t IgE( 00d hro-m A)w LISAm cularD axP en-s e lasts wasc n oft eachg
2
om-b ereb gra-d to an nceI led( s atd anti-d ec-o wasp uck-i reri
ppears to be tightly coupled to the developmental biof the larvae[14]. Previous canine trials suggest that aody responses to these antigens may result in prevent
arval migration through tissue or attenuation of larvae shey do not mature into blood feeding adults as showheir reduced fecundity (i.e., reduction in fecal egg cou17]. We sought to further elucidate the details of the humnd especially the cellular immunological factors relate
he high level of protection induced by irradiated vaccith special attention focused on the role of ASPs in throcesses.
. Material and methods
.1. Study dogs and animal husbandry
Purpose-bred, parasite naı̈ve, male beagles aged 8± 1eeks were purchased, identified by ear tattoo, and m
ained in the George Washington University Animal Reseacility as described elsewhere[18]. The experiments weonducted according to a protocol approved by the Univenimal Care and Use Committee. Before the first vacc
ion and after each subsequent one, a serum and wholeample were obtained from each dog.
.2. Vaccine study design and immunizations
The vaccine trial was designed to test the irradiatedae (L3) ofA. caninum in comparison with a placebo co
old serial dilutions thereafter followed by washes and aion of peroxidase-conjugated anti-dog IgG1, IgG2 orBethyl Laboratories Inc., Montgomery, TX, USA) at 1:10ilution. Wells were then washed and substrate and cogen (O-Phenylenediamine, Sigma–Aldrich Co., USere added and absorbance was read on an automatic Eicroplate reader (SpectraMax 240 PC reader, Moleevices, Sunnyvale, CA, USA) at 492 nm with SOFTMro software (Molecular Devices). The mean optical dity of control canine sera was used as a baseline. Therum dilution greater than three times above baselineonsidered the titration endpoint. The geometric meahese endpoints was calculated for the five dogs fromroup.
.4. Western blot assay
The Western blot was performed using larval recinant antigens (ASP-1, ASP-2 and MTP-1) which woiled in SDS sample buffer, separated on a 4–20%ient SDS-PAGE gel (Invitrogen) and transferred onitrocellulose membrane (Schleicher & Schuell BioScie
nc., Keene, NH, USA). The blot was probed with a poo5�l each) serum from vaccinated and control dogilution 1:1000. Horseradish peroxidase-conjugatedog IgG1, IgG2 and IgA (Bethyl) were used as a sndary antibody at a dilution 1:1000. Developmenterformed with ECL reagent (Amersham Biosciences, B
nghamshire, England) according to the manufactu′snstructions.

R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509 503
2.5. Lymphoproliferation assay
The lymphoproliferation assay was performed using awhole blood microassay as previously described[21]. Briefly,25�l of heparinized blood was diluted in 200�l of RPMI1640 medium (GIBCO, Grand Island, NY, USA) supple-mented with 3% antibiotic/antimycotic solution (GIBCO,Grand Island, NY, USA). All tests were performed in tripli-cate in 96-well flat-bottomed culture plates using L3 extractantigen at a concentration of 25�g/ml and ConcanavalinA (ConA) (Sigma–Aldrich Co., USA) at 80�g/ml. Incu-bation was carried out in a humidified 5% CO2 atmo-sphere at 37◦C for two (ConA-stimulated cultures) andfive (L3-stimulated cultures) days. Cells were pulsed for6 h with 1�Ci of [3H] thymidine (Perkin Elmer LAS,Shelton, CT, USA) and harvested onto glass fibre fil-ters. Radioactive incorporation was determined by liq-uid scintillation spectrometry. Proliferation responses wereexpressed as stimulation index (SI = mean proliferationof stimulated cultures/mean proliferation of unstimulatedcultures).
At the end of the study, mesenteric lymph node andspleen samples were collected at necropsy and maceratedusing glass tissue grinders. Mononuclear cells were sepa-rated using a Ficoll-Hypaque gradient (Histopaque 1.077,Sigma–Aldrich Co., USA) and cultured in RPMI mediums and1 richC ndert tiona
2
larc tiona terso tedw ndI er NJ,U racta rP Co.,U nd1 olsc Theca .,Uf
hio-c in.TS ellsw ple-
mented with 0.5% BSA and 0.1% of Sodium Azide), perme-abilized with a 0.5% saponin solution (Sigma–Aldrich Co,USA) in PBS–BSA buffer before being stained with anti-bovine IFN-�-phycoerythrin (PE) (Serotec Inc.) and anti-bovine IL-4 PE (Serotec Inc.) at room temperature for at least1 h. The cells were washed twice with PBS–BSA buffer andthen fixed with fixative solution (10.0 g/l paraformaldehyde;10.2 g/l cacodylic acid; 6.65 g/l sodium chloride; pH 7.2). Thedata were acquired in a FACScalibur flow cytometer (BectonDickinson, San Jose, CA, USA), selecting the lymphocytepopulation. Thirty thousand events were counted. FITC andPE-labeled Ig control antibodies were included in all experi-ments.
2.7. IL-4, IL-and IFN-γ production by cultured wholeblood
The whole blood was diluted 1:8 in RPMI supplementedwith 3% antibiotic/antimycotic solution (GIBCO, GrandIsland, NY, USA) in a 48-well flat-bottomed culture platewith a final volume of 1.0 ml per well. Cells were stimulatedby the addition of 25�g/ml of L3 extract antigen or 1�g/mlof Phorbol 12-myristate 13-acetate (PMA, Sigma–AldrichCo., USA). After 48 h of incubation at 37◦C, 700�l of super-natant was removed from each well and stored at−20
◦C until
required for the cytokine assay.p-
t po-l s.B withs sub-s
2
wasp indi-c BDF s;u ellsw itha ineC assI c).E oflu-o c.)w
itht with1 ves cid;6 res-c owc lect-i erec
upplemented with 3% antibiotic/antimycotic solution0% of heat inactivated Fetal Bovine Serum (Sigma–Aldo., USA). The cells were cultured and harvested u
he same conditions described for whole blood proliferassays.
.6. Cytokine detection by intracellular immunostaining
Lymphocytes were analyzed for their intracelluytokine expression pattern using whole blood stimuland detection by flow cytometry. Three hundred microlif whole blood was diluted to 1.0 ml in RPMI supplemenith 3% of antibiotic/antimycotic solution (GIBCO, Gra
sland, NY, USA) in sterile 17 mm× 100 mm polypropylenound-bottom tubes (Becton Dickinson, Franklin Lakes,SA). Stimulation was carried out using either L3 extntigen at a concentration of 25�g/ml (6 h of stimulation) ohorbol 12-myristate 13-acetate (PMA, Sigma–AldrichSA) and Ionomycin (Sigma–Aldrich Co., USA) at 25 a�g/ml, respectively (4 h of stimulation). Negative contrultures were performed without antigen or mitogens.ultures were incubated for 2 h at 37◦C in 5% CO2 beforeddition of 10�g/ml of Brefeldin A (Sigma–Aldrich CoSA) followed by an additional incubation at 37◦C for a
urther 4 h.The cells were stained with anti-CD4-fluorescein isot
yanate (FITC) (Serotec Inc., Raleigh, NC, USA) for 30 mhe red blood cells were lysed with BD FACSTM Lysingolution (Becton Dickinson, San Jose, CA, USA). The cere washed with 5 ml of PBS–BSA buffer (PBS sup
IL-4, IL-10 and IFN-� were measured using a caure ELISA assay for dogs (R&D Systems, Minneais, MN, USA) following the manufacturer’s instructioniotin-labeled detection antibodies were used, revealedtreptavidin-HRP (Amersham Biosciences) and OPDtrate system (Sigma–Aldrich Co.).
.8. Cellular profile of peripheral blood
The phenotyping of peripheral blood cell populationserformed using whole blood samples collected at theated days. In short, 1.0 ml of blood was incubated withACSTM Lysing Solution to allow the lysis of erythrocytenlysed cells were washed twice with PBS–BSA buffer. Cere stained in a 96-well round bottomed microplate wnti-canine CD3-FITC, anti-canine CD4-FITC, anti-canD8-FITC, anti-canine B cells-PE, anti-canine MHC Cl
I-FITC and anti-human CD14-FITC (all from Serotec Inosinophils were analyzed by light scatter and by autrescence[22–24]. Ig-FITC and PE controls (Serotec Inere used in all experiments.After incubation of 30 min at room temperature w
he respective antibodies, cells were washed twice50�l of PBS–BSA buffer and then fixed with fixatiolution (10.0 g/l paraformaldehyde; 10.2 g/l cacodylic a.65 g/l sodium chloride; pH 7.2). The data on fluoently labeled cells were acquired in a FACScalibur flytometer (Becton Dickinson, San Jose, CA, USA), seng the lymphocyte population. Thirty thousand events wounted.

504 R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509
2.9. Parasitological evaluation
Quantitative hookworm egg counts (using the McMastertechnique) were performed three times per week beginningtwo weeks after challenge infection. Four weeks after chal-lenge infection, the dogs were killed by intravenous barbitu-rate injection, and the adult hookworms were recovered andcounted from the small and large intestines at necropsy[20].The sex of each of the adult hookworms was determinedand worm lengths were measured as described elsewhere[25]. Approximately 1–2 cm of small intestine was sepa-rated and placed into formalin for future histopathologicalanalysis.
2.10. Tissue penetration assay
The ability of irradiated L3 ofA. caninum to penetratedog skin as well as the inhibitory properties of serum fromdogs immunized with irradiated L3 was evaluated as previ-ously described[26]. Briefly, dog ear skin (obtained from anon-infected animal) was shaved, scrubbed with gauze andwashed with PBS so that the buffer was in contact with theunderside of the skin. In the first experiment, irradiated andnon-irradiatedA. caninum L3 (300 L3/group) were incubatedin 50�l of PBS (pH 7.2) for 30 min at 37◦C. Subsequently,e l ofP ture.L them e L3t xper-i pre-s expei incu-bp gsi trold
2
tp n andn etrict isti-c dentg therew rnovt dis-t st pen-dn thep orm twog
3. Results
3.1. Parasitological
Female hookworms in the vaccinated animals were lessfecund (Fig. 1A) showing a decrease of 87% in egg pro-duction (P = 0.028). The egg counts were calculated frommedians of the last 3 days of the experiment using a non-parametric test for two independent samples. Statisticallysignificant differences in median burdens from adult maleworms between vaccinated and control dogs were found inthe small intestine with reduction of 65.7% (P = 0.032). Atnecropsy, the total number of worms was found to be nor-mally distributed by a Kolmogorov–Smirnoff test and wasanalyzed by Student’st-test resulting in an overall 53% reduc-tion of adult worms (P = 0.011,Fig. 1B). Only small numbersof worms were found in the large intestine (not the normalsite of residence for hookworms).
At necropsy, all worms recovered from the vaccinateddogs were fixed in formalin. The lengths of 100 undamaged
Fig. 1. Parasitological evaluation of irradiated L3 vaccinated (irL3) and con-trol dogs (Alum) after challenge with 500 infective larvae ofA. caninum.(A) Number of eggs per gram of feces at necropsy. Results are expressedas a range of values (box), standard deviation (whiskers), and median (barin box). (B) Number of adult hookworms collected from the small intestineat necropsy. Results are expressed as mean± S.D. Percentage of parasito-logical reduction and statistical significance between vaccinated and controldogs are showed in the graphs.
ach group of L3 was transferred to the dog skin (in 1.0 mBS) and allowed to migrate for 30 min at room tempera3 that migrated through the skin were collected fromedia (PBS) in the collection tube and counted, as wer
hat remained on the outside surface of the skin. Each ement was conducted in triplicate, and the results wereented as the means of these three replicates. A secondment was performed using the same technique but withation of normal (non-irradiated)A. caninum L3 with eitherooled serum (5�l of neat serum from each dog) from do
mmunized with irradiated L3 or pooled serum from conogs.
.11. Statistical analysis
In some cases, the small samples size (n = 10) did norovide enough data to discriminate between Gaussiaon-Gaussian distribution; so we used a non-param
est (Mann–Whitney) to determine if there were statal differences between the medians of two indepenroups (vaccinated and control groups). In cases whereere more values, the one-sample Kolmogorov–Smi
est was used to test that a variable was normallyributed. For normally distributed variables, Student’t-est was used to compare the means from indeent groups. We interpreted theP-value (≤0.05) of bothon-parametric and parametric tests as determiningrobability of random sampling resulting in medianseans as far apart (or more) as observed in theroups.
r-

R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509 505
worms from each group were measured and the mean lengthswere compared statistically. Mean lengths of female and maleworms recovered from vaccinated dogs were 10% (P = 0.309)and 13% (P = 0.031) smaller respectively than worms recov-ered from control dogs.
3.2. Humoral immune response
Dogs vaccinated with irradiatedA. caninum L3 pro-duced IgG1 and IgG2 antibody responses as measured byELISA using the L3 extract antigen (Fig. 2). IgE titers weredetected only after the larval challenge and had decreasedat necropsy. After vaccination, the dogs vaccinated withirradiated L3 presented low antibody titers against recom-binant Na-ASP-1 (IgG1 and IgG2) andAc-MTP-1 (onlyIgG2) (Fig. 3A). Higher titers of IgG1 and IgG2 againstrecombinantAc-ASP-2 (geometric mean of 16,817 and 1204,respectively) were found in sera from the vaccinated dogs(Fig. 3A). Elevated titers of IgA againstAc-ASP-2 were alsofound after vaccination; however, the differences betweenvaccinated and control groups were not statistically sig-nificant (Fig. 3A). In addition, low titers of IgE againstNa-ASP-1,Ac-ASP-2 andAc-MTP-1 (geometric means of
Fig. 2. Geometric mean titers of the IgG1 and IgG2 antibody responsesagainstA. caninum L3 extract in dogs vaccinated with irradiatedA. caninumL3 vaccinated and control dogs. TheX-axis represents the experimental daysafter primary vaccination. The doses of vaccination [1–3] and the challenge[C] were indicated on theX-axis.
16, 138 and 1, respectively) were found. Western blotsconfirmed the presence of specific antibody titers againstAc-ASP-2, mostly for IgG1 but also for IgG2 and IgA(Fig. 3B).
Fr(
ig. 3. Immune recognition of serum from immunized and control dogs. (Aecombinant ASP-1, ASP-2 and MTP-1 in dogs vaccinated with irradiatedA. caninuASP-1, ASP-2 and MTP-1) probed with pooled serum from vaccinated [V] a
) Geometric mean titers of the IgG1, IgG2 and IgA antibody responses againstm L3 and control dogs. (B) Western blots with larval recombinant antigensnd control dogs [C].
506 R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509
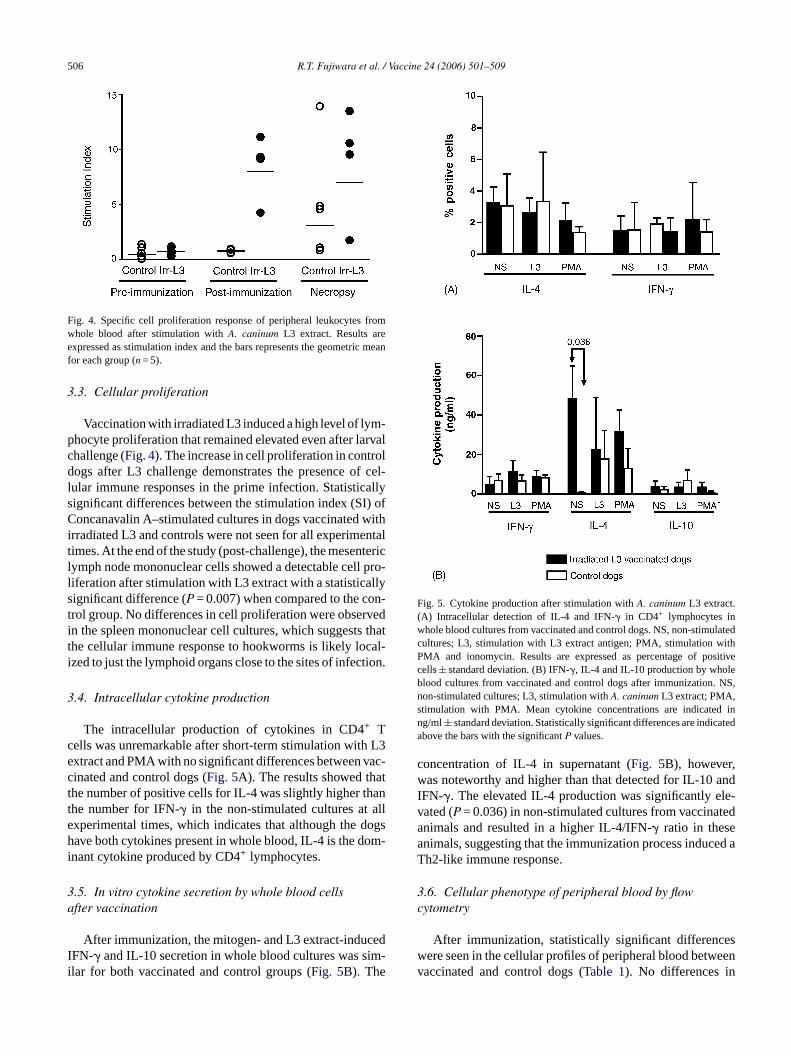
Fig. 4. Specific cell proliferation response of peripheral leukocytes fromwhole blood after stimulation withA. caninum L3 extract. Results areexpressed as stimulation index and the bars represents the geometric meanfor each group (n = 5).
3.3. Cellular proliferation
Vaccination with irradiated L3 induced a high level of lym-phocyte proliferation that remained elevated even after larvalchallenge (Fig. 4). The increase in cell proliferation in controldogs after L3 challenge demonstrates the presence of cel-lular immune responses in the prime infection. Statisticallysignificant differences between the stimulation index (SI) ofConcanavalin A–stimulated cultures in dogs vaccinated withirradiated L3 and controls were not seen for all experimentaltimes. At the end of the study (post-challenge), the mesentericlymph node mononuclear cells showed a detectable cell pro-liferation after stimulation with L3 extract with a statisticallysignificant difference (P = 0.007) when compared to the con-trol group. No differences in cell proliferation were observedin the spleen mononuclear cell cultures, which suggests thatthe cellular immune response to hookworms is likely local-ized to just the lymphoid organs close to the sites of infection.
3.4. Intracellular cytokine production
The intracellular production of cytokines in CD4+ Tcells was unremarkable after short-term stimulation with L3extract and PMA with no significant differences between vac-cinated and control dogs (Fig. 5A). The results showed thatt ant alle dogsh om-i
3a
cedI im-i
Fig. 5. Cytokine production after stimulation withA. caninum L3 extract.(A) Intracellular detection of IL-4 and IFN-� in CD4+ lymphocytes inwhole blood cultures from vaccinated and control dogs. NS, non-stimulatedcultures; L3, stimulation with L3 extract antigen; PMA, stimulation withPMA and ionomycin. Results are expressed as percentage of positivecells± standard deviation. (B) IFN-�, IL-4 and IL-10 production by wholeblood cultures from vaccinated and control dogs after immunization. NS,non-stimulated cultures; L3, stimulation withA. caninum L3 extract; PMA,stimulation with PMA. Mean cytokine concentrations are indicated inng/ml± standard deviation. Statistically significant differences are indicatedabove the bars with the significantP values.
concentration of IL-4 in supernatant (Fig. 5B), however,was noteworthy and higher than that detected for IL-10 andIFN-�. The elevated IL-4 production was significantly ele-vated (P = 0.036) in non-stimulated cultures from vaccinatedanimals and resulted in a higher IL-4/IFN-� ratio in theseanimals, suggesting that the immunization process induced aTh2-like immune response.
3.6. Cellular phenotype of peripheral blood by flowcytometry
After immunization, statistically significant differenceswere seen in the cellular profiles of peripheral blood betweenvaccinated and control dogs (Table 1). No differences in
he number of positive cells for IL-4 was slightly higher thhe number for IFN-� in the non-stimulated cultures atxperimental times, which indicates that although theave both cytokines present in whole blood, IL-4 is the d
nant cytokine produced by CD4+ lymphocytes.
.5. In vitro cytokine secretion by whole blood cellsfter vaccination
After immunization, the mitogen- and L3 extract-induFN-� and IL-10 secretion in whole blood cultures was slar for both vaccinated and control groups (Fig. 5B). The

R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509 507
Table 1Peripheral blood leukocyte phenotype of dogs vaccinated with irradiated L3and control dogs after immunization
Phenotype Groups P
Irradiated L3 (n = 5) Control (n = 5)
LymphocytesCD3 68.89± 11.03 77.77± 4.88 0.160CD4 36.72± 4.35 48.23± 2.45 0.002CD8 16.49± 2.98 16.77± 4.69 0.922B cells 31.32± 5.80 19.25± 6.15 0.019MHC II 37.71 ± 7.48 23.66± 7.84 0.007CD11/18 11.05± 3.27 8.07± 2.62 0.069
Monocytes (CD14) 5.61± 1.87 6.71± 1.40 0.853Eosinophils 14.98± 3.56 7.37± 0.36 0.006
Results are given as percentual mean± standard deviation (statistically sig-nificantP values are given in bold).
the cellular profiles of peripheral blood were seen beforeimmunization or at necropsy (data not shown). Vaccinateddogs had significantly greater numbers of B cells (P = 0.019)and eosinophils (P = 0.006) and elevated cell activation asobserved by the higher expression of MHC class II in lym-phocytes (P = 0.007). Conversely, vaccinated dogs showed
FmPAar
a lower number of CD4+ T cells than the control group(P = 0.002).
3.7. Impaired penetration of irradiated land inhibitionproperty of serum from vaccinated dogs
The larval migration assay showed that 95–100% of irra-diatedA. caninum L3 did not penetrate the skin while just15–20% of normalA. caninum L3 remained on skin afterincubation (Fig. 6A), demonstrating the impaired abilityof irradiated larvae to penetrate skin. When non-irradiatedhealthy L3 were pre-incubated in serum from vaccinateddogs, migration ofA. caninum was inhibited by 60%(Fig. 6B).
4. Discussion
Work on the vaccinology of hookworm first began in the1930s, when infective larvae ofA. caninum were shown tobe antigenic[27] and protective immunity in canines couldbe elicited by administering repeated doses of these larvae[28,29]. Early attempts to understand the mechanisms under-lying the protective effect suggested that both cellular andhumoral responses were in operation[30]. At the time, theability to study cellular immune responses was limited, butiw open-io ivel en-uT tedA ds[ ftercn le bya -c soonr andp pitea -t miao oft le forp thec ing ah tedL une
ig. 6. Inhibition of larval migration. (A) Percentual of inhibition of larvaligration in dog ear skins with irradiated and normalA. caninum L3. (B)ercentual of inhibition of larval migration in dog ear skins with normal. caninum L3 incubated with serum from vaccinated and control dogs,fter the immunization process. The results are expressed as mean of threeeplicates± S.D.
rook-
w ount( s ofa byB iesi Th2
t was determined that the humoral immunity toA. caninumas directed against antigens secreted around the oral
ng and excretory pore of the infective larvae[12,26]. Basedn the success of eliciting protection with living infect
arvae ofA. caninum in canines, efforts were made to attate the hookworm larvae by ionizing radiation[12,31,32].he result was a highly effective vaccine using X-irradia. caninum L3 to control hookworm infections in cani
12], which demonstrated its protective effect in dogs ahallenge with normal, healthy larvae[8–11,13,31]. Immu-ity generated by the irradiated vaccine was transferabcombination of serum and lymphoid cells[32]. The vac
ine was developed commercially in the 1970s, but wasemoved from the market due concerns of veterinarianset-owners that sterilizing immunity was not induced desttaining up to 90% protection[12], which is a level of pro
ection sufficient to prevent serious complications of aner fatal hemorrhage[33]. An understanding of the details
he cellular and humoral factors and antigens responsibrotective immunity in the dog has not been resolved. Inurrent study, our aim was to repeat the studies inducigh level of protection induced in canines using irradia3 and then to characterize the cellular and humoral immesponses to crude and definedA. caninum antigens.
Our results showed a protective response from horm infection characterized by both reduced fecal egg c
85%, P = 0.028) and significantly decreased numberdult worms in the gut (53%,P = 0.011). As suggestedoag et al.[34], the high levels of IgG1 subclass antibod
n dogs protected by the irradiated L3 vaccine implicate a

508 R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509
type response in immunity. This hypothesis receives furthersupport from recent work by Girod et al.[35] in BALB/c mice,where a localized Th2 response was found to be importantfor conferring protection against hookworms. We add to thisgrowing literature several new lines of evidence in supportof the idea that Th2 is the immunological phenotype respon-sible for protection against hookworm infection. First, weobserved increased levels of IL-4, especially in relation to theTh1 cytokine IFN-�; elevated levels of IL-4 were particularlymarked after the immunization. Interestingly, we detected IL-4 only in supernatants from in vitro cultures and not fromCD4+ T cells (as determined by intracellular cytokine stain-ing), indicating that the source of this IL-4 production maybe mast cells, basophils[36], or eosinophils[37]. Our secondline of evidence comes from ex vivo immunophenotyping ofthe peripheral blood leukocytes, where we found elevated lev-els of eosinophils and B cells, likely activated by the elevatedIL-4, along with markedly higher expression of MHC II onlymphocytes. Finally, we observed a strong cellular prolifer-ative response to crude third stage larval antigen extract incanines after immunization with irradiated L3 and then againwhen challenged with non-irradiated healthyA. caninum L3.Interestingly, the hookworm specific proliferative responseperformed with the organ tissues at necropsy was detectedonly in the mesenteric lymph node cells but not in spleencells. This result suggests that the cellular immune responsea arilyl con-s 2r B/cmc itya niza-t imedc cina-t nsew d Bc bod-i lb n oft andg ina-t morea
ess,w d byt etra-t ates owedt theira tedr rvaec diesfw r thep e the
migrating larvae in the host. Finally, our antibody analysisdemonstrated that vaccination with irradiated larvae inducedthe production of antibodies against hookworm recombi-nant antigens, which have been selected as lead candidatesfor further testing by the Human Hookworm Vaccine Ini-tiative [3]. Among these larval stage antigens are MTP-1(astacin-like metalloprotease 1), ASP-1 and ASP-2 (Ancy-lostoma secreted proteins 1 and 2)[3,15,16,18]. The highertiters of IgG1 antibodies against ASP-2 show that this pro-tein is a dominant protein in irradiated larval L3 vaccination.This protein is secreted specifically by larval stages whenstimulated by host-like conditions, and it has been conjec-tured that the ASPs function during the transition of L3 fromtheir free-living form in the soil to a new parasitic existencein their mammalian host[39]. Previous results have shownthat hamsters immunized with recombinant ASP-2 exhibit asignificant reduction in adult hookworm burdens and reduc-tion in length of surviving worms[25]. Similarly, canine trialsusingAc-ASP-2 showed a significant reduction in egg countsand worm load[40]. These animal trials, in conjunction withdata from humans living in hookworm endemic areas show-ing an association between antibodies against ASP-2 and areduced risk of heavy levels ofNecator infection[40], high-light the importance of this molecule in protective immunity.The results of the current study further support the role of thisantigen as well as the Th2 immune response that is requiredt ook-w
A
ac-c ndM y anI ionalC lop-m archC
R
ioliure.
PJ.edSA
ude for
ative.
S.
an1.
gainst the hookworms is not generalized but rather primocalized close to the site of infection; this observation isistent with Girod et al.’s[35] findings that a “localized” Thesponse was important for conferring protection in BALice vaccinated with irradiated hookworm (Necator ameri-
anus). The role of cell proliferation in protective immungainst hookworms clearly demonstrates that the immu
ion process induces a memory response in previously prells. Taken together these data further support that vacion with irradiated L3 induces a cellular immune respoith a Th2 bias, resulting in the activation of T cells anells and concomitant activation of Th2-associated anties. The reduction of the number of T CD4+ in the peripheralood after vaccination may be associated with migratio
hese cells to other points of infections such as skin, lungut, and not with a immunosuppressive effect of the vacc
ion since other cell populations as were increased andctivated (e.g., T cells, B cells and eosinophils).
Among the events triggered by the vaccination proce showed that protective immunity may be establishe
he production of specific antibodies that impair the penion of infective larvae, blocking their ability to penetrkin. The tissue penetration assay used here also shhat the ionizing rays used to irradiated L3 decreasedbility to penetrate through skin. The antibody-mediaesponse elicited by the administration of irradiated laould explain the protection described in previous stuollowing passive transfer of immune sera[32,34,38]. In thisay, the specific antibody responses that clearly impaienetration through skin might also target and damag
o generate protective immunity in vaccines targeting horm larvae.
cknowledgments
This project was supported by the Human Hookworm Vine Initiative of the Sabin Vaccine Institute and by Bill aelinda Gates Foundation. J. Bethony is supported b
nternational Research Scientist Award, Fogarty Internatenter, NIH. A. Loukas is supported by a Career Deveent Award from the National Health and Medical Reseouncil of Australia.
eferences
[1] De Silva N, Brooker S, Hotez P, Montresor A, Engels D, SavL. Soil transmitted helminth infections: updating the global pictTrends Parasitol 2003;12:547–51.
[2] Cappello M, Vlasuk GP, Bergum PW, Huang S, HotezAncylostoma caninum anticoagulant peptide: a hookworm derivinhibitor of human coagulation factor Xa. Proc Natl Acad Sci U1995;92:6152–6.
[3] Hotez PJ, Zhan B, Bethony JM, Loukas A, Williamson A, GoGN, et al. Progress in the development of a recombinant vaccinhuman hookworm disease: the human hookworm vaccine initiInt J Parasitol 2003;33:1245–58.
[4] Hotez PJ, Brooker S, Bethony JM, Bottazzi ME, Loukas A, XiaoHookworm infection. N Engl J Med 2004;351:799–807.
[5] Albonico M, Crompton DW, Savioli L. Control strategies for humintestinal nematodes infections. Adv Parasitol 1999;42:277–34

R.T. Fujiwara et al. / Vaccine 24 (2006) 501–509 509
[6] Albonico M, Smith PG, Ercole E, Hall A, Chwaya HM, AlawiKS, Savioli L. Rate of reinfection with intestinal nematodesafter treatment of children with mebendazole or albendazole in ahighly endemic area. Trans R Soc Med Trop Hyg 1995;89:538–41.
[7] Albonico M, Bickle Q, Ramsam M, Montresor A, Savioli L, TaylorM. Efficacy of mebendazole and levamisole alone or in combi-nantion against intestinal nematode infections after repeated tar-geted mebendazole treatment in Zanzibar. Bull World Health Organ2003;81:343–52.
[8] Miller TA. Effect of X-irradiation upon the infective larvae ofAncy-lostoma caninum and the immunogenic effect in dogs of a singleinfection with 40 Kr irradiation larvae. J Parasitol 1964;50:735–42.
[9] Miller TA. Studies on canine ancylostomiasis – double vaccinationwith X-irradiatedAncylostoma caninum larvae. J Am Vet Med Assoc1965;146:41–4.
[10] Miller TA. Effect of route of administration of vaccine and challengeon the immunogenic efficiency of double vaccination with irradiatedAncylostoma caninum larvae. J Parasitol 1965;51:200–6.
[11] Miller TA. Comparison of immunogenic efficiencies of normaland X-irradiatedAncylostoma caninum larvae in dogs. J Parasitol1966;52:512–9.
[12] Miller TA. Vaccination against the canine hookworm diseases. AdvParasitol 1971;9:153–83.
[13] Miller TA. Industrial development and field use of the canine hook-worm vaccine. Adv Parasitol 1978;16:333–42.
[14] Hotez PJ, Hawdon JM, Cappello M, Jones BF, Ghosh K, VolvovitzF, et al. Molecular approaches to vaccinating against Hookwormdisease. Pediatr Res 1996;40:515–21.
[15] Hawdon JM, Jones BF, Hoffman D, Hotez PJ. Cloning and expres-withBiol
[ nfam-
[ ook-
[ donteinhost-
ar-
[ S,eine
the52–
[ doneins. J
[ sonyte-
[22] Carulli G, Sbrana S, Azzara A, Minnucci S, Angiolini C, MariniA, Ambrogi F. Detection of eosinophils in whole blood samples byflow cytometry. Cytometry 1998;34:272–9.
[23] Vadas MA, Varigos G, Nicola N, Pincus S, Dessein A, Met-calf D, et al. Eosinophil activation by colony-stimulation factorin man: metabolic effects and analysis by flow cytometry. Blood1983;61:1232–41.
[24] Watt SM, Burgess AW, Metcalf D, Battye FL. Isolation of mousebone marrow neutrophils by light scatter and autofluorescence. JHistochem Cytochem 1980;28:934–46.
[25] Goud GN, Zhan B, Ghosh K, Loukas A, Hawdon J, DobardzicA, et al. Cloning, yeast expression, isolation and vaccine testingof recombinantAncylostoma secreted protein 1 (ASP-1) and ASP-2from Ancylostoma ceylanicum. J Infect Dis 2004;189:919–29.
[26] Williamson AL, Brindley PJ, Abbenante G, Datu BJD, Prociv P,Berry C, et al. Hookworm aspartic protease, Na-APS-2, cleaveshuman hemoglobin and serum proteins in a host-specific fashion.J Infect Dis 2003;187:484–94.
[27] Stumberg JE. Precipitin and complement-fixation test on dog serawith antigen from the dog hookwormAncylostoma caninum. Am JHyg 1930;12:657–68.
[28] Foster AO. The immunity of dogs toAncylostoma caninum. Am JHyg 1935;22:65–105.
[29] McCoy OR. Immunity reactions of the dog against hookworm (Ancy-lostoma caninum) under conditions of repeated infection. Am J Hyg1931;14:268–303.
[30] Sarles MP, Taliaferro WH. The local points of defense and the pas-sive transfer of acquired immunity toNippostrongylus muris in rats.J Infect Dis 1936;59:207–20.
[31] Miller TA. Persistence of immunity following double vaccinationof pups with X-irradiatedAncylostoma caninum larvae. J Parasitol
[–41.
[ Clin
[ JL.natedl
[ ion
Para-
[ tion.
[ ronL-5Sci
[ s and
[ the
[ tt J,rvae
acci-
sion of Ancylostoma secreted protein: a polypeptide associatedthe transition to parasitism by infective hookworm larvae. JChem 1996;271:6672–8.
16] Hawdon JM, Narasimhan S, Hotez PJ.Ancylostoma secreted protei2: cloning and characterization of a second member of aily of nematode secreted proteins fromAncylostoma caninum. MolBiochem Parasitol 1999;99:149–65.
17] Ghosh K, Hotez PJ. Antibody-dependent reductions in mouse hworm burden after vaccination withAncylostoma caninum secretedprotein 1. J Infect Dis 1999;180:1674–8.
18] Hotez PJ, Ashcom J, Zhan B, Bethony J, Loukas A, HawJ, et al. Effect of vaccination with a recombinant fusion proencoding an astacinlike metalloprotease (MTP-1) secreted bystimulatedAncylostoma caninum third-stage infective larvae. J Pasitol 2003;89:853–5.
19] Loukas A, Bethony JM, Williamson AL, Goud GN, MendezZhan B, et al. Vaccination of dogs with a recombinant cystprotease from the intestine of canine hookworms diminishesfecundity and growth of worms. J Infect Dis 2004;189:1961.
20] Hotez PJ, Ashcom J, Bin Z, Bethony J, Williamson A, HawJM, et al. Effect of vaccinations with recombinant fusion proton Ancylostoma caninum habitat selection in the canine intestineParasitol 2002;88:684–90.
21] Shifrine M, Taylor NJ, Rosenblatt LS, Wilson FD. Compariof whole blood and purified canine lymphocytes in a lymphocstimulation microassay. Am J Vet Res 1978;39:687–90.
1965;51:705–11.32] Miller TA. Transfer of immunity toAncylostoma caninum infection
in pups by serum and lymphoid cells. Immunology 1967;12:23133] Loukas A, Prociv P. Immune responses in hookworm infections.
Microbiol Rev 2001;14:689–703.34] Boag PR, Parsons JC, Presidente PJA, Spithill TW, Sexton
Characterisation of humoral immune response in dogs vacciwith irradiatedAncylostoma caninum. Vet Immunol Immunopatho2003;92:87–94.
35] Girod N, Brown A, Pritchard DI, Billett EE. Successful vaccinatof BALB/c mice against human hookworm (Necator americanus):the immunological phenotype of the protective response. Int Jsitol 2003;33:71–80.
36] Choi P, Reiser H. IL-4: role in disease and regulation of producClin Exp Immunol 1998;113:317–9.
37] Lamkhioued B, Gounni AS, Aldebert D, Delaporte E, Prin L, CapA, et al. Synthesis of type 1 (IFN gamma) and type 2 (IL-4Iand IL-10) cytokines by human eosinophils. Ann N Y Acad1996;796:203–8.
38] Else KJ, Finkelman FD. Intestinal nematode parasites, cytokineeffector mechanisms. Int J Parasitol 1998;28:1145–58.
39] Hawdon JM, Hotez PJ. Hookworm: developmental biology ofinfectious process. Curr Opin Genet Dev 1996;6:618–23.
40] Bethony J, Loukas A, Smout M, Brooker S, Mendez S, Plieskaet al. Antibodies against a secreted protein from hookworm lareduce the intensity of hookworm infections in humans and vnated laboratory animals. FASEB J, in press.
Related Documents











