Date : 27/03/2012 Page : 45/ 77 Introduction au RET et à ses applications pour étudier les interactions protéine-protéine V – APPLICATION DES TECHNIQUES DE RET A L’ANALYSE DES INTERACTIONS PROTEINE- PROTEINE : CAS DES RECEPTEURS COUPLES AUX PROTEINES G (RCPG) Les RCPG sont des protéines à sept domaines transmembranaires exprimées au niveau de la membrane plasmique des cellules. A l’interface entre le milieu extra- et intracellulaire, ces récepteurs assurent la reconnaissance de stimuli extracellulaires et initient en réponse des cascades d’événements intracellulaires. Les RCPG ont longtemps été considérés comme des récepteurs présents à la membrane uniquement sous forme de monomères. Les avancées techniques de RET et les nombreux travaux réalisés dans le domaine de la réceptologie ont toutefois permis de remettre en question ce dogme. Il a ainsi été proposé que les RCPG pouvaient être aussi exprimés à la membrane plasmique sous forme d’oligomères. La notion d’oligomérisation est un terme général qui décrit l’association de protéines au sein d’un même complexe. La structure minimale d’un oligomère s’organise autour de deux protéines formant un dimère. Une association en homodimères correspond à une interaction entre deux protéines identiques et une association en hétérodimères à deux protéines différentes. Cependant, les techniques d’analyse de RET ne permettent pas toujours de distinguer les formes dimériques des autres formes oligomériques (trimères, tétramères, etc.). L’objet de ce chapitre est de présenter les avantages et les limites des méthodes de RET les plus couramment utilisées pour mettre en évidence des interactions protéine-protéine. De plus, nous discuterons brièvement de l’influence de la taille et du positionnement des sondes fluorescentes par rapport à la taille des complexes protéiques étudiés et par rapport au transfert d’énergie. A/ LES FLUOROPHORES ORGANIQUES EN FRET ET TR-FRET 24 Les fluorophores organiques sont de petites molécules qui nécessitent d’être couplées soit à des groupements réactifs (TétraCystéine FlAsH par exemple) lesquels vont réagir directement avec la protéine cible, soit couplés à des « sondes » (anticorps, SNAP-tag, Halo-tag, etc.). Leur utilisation en RET implique donc d’utiliser des méthodes qui permettent de marquer spécifiquement les protéines d’intérêt avec 24 Voir la revue Cottet M et al., Frontier in Endocrinology 2012.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Date : 27/03/2012 Page : 45/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
V – APPLICATION DES TECHNIQUES DE RET A L’ANALYSE DES INTERACTIONS PROTEINE-PROTEINE : CAS DES RECEPTEURS COUPLES AUX PROTEINES G (RCPG)
Les RCPG sont des protéines à sept domaines transmembranaires exprimées au
niveau de la membrane plasmique des cellules. A l’interface entre le milieu extra- et
intracellulaire, ces récepteurs assurent la reconnaissance de stimuli extracellulaires
et initient en réponse des cascades d’événements intracellulaires. Les RCPG ont
longtemps été considérés comme des récepteurs présents à la membrane
uniquement sous forme de monomères. Les avancées techniques de RET et les
nombreux travaux réalisés dans le domaine de la réceptologie ont toutefois permis
de remettre en question ce dogme. Il a ainsi été proposé que les RCPG pouvaient
être aussi exprimés à la membrane plasmique sous forme d’oligomères. La notion
d’oligomérisation est un terme général qui décrit l’association de protéines au sein
d’un même complexe. La structure minimale d’un oligomère s’organise autour de
deux protéines formant un dimère. Une association en homodimères correspond à
une interaction entre deux protéines identiques et une association en hétérodimères
à deux protéines différentes. Cependant, les techniques d’analyse de RET ne
permettent pas toujours de distinguer les formes dimériques des autres formes
oligomériques (trimères, tétramères, etc.).
L’objet de ce chapitre est de présenter les avantages et les limites des méthodes de
RET les plus couramment utilisées pour mettre en évidence des interactions
protéine-protéine. De plus, nous discuterons brièvement de l’influence de la taille et
du positionnement des sondes fluorescentes par rapport à la taille des complexes
protéiques étudiés et par rapport au transfert d’énergie.
A/ LES FLUOROPHORES ORGANIQUES EN FRET ET TR-FRET24
Les fluorophores organiques sont de petites molécules qui nécessitent d’être
couplées soit à des groupements réactifs (TétraCystéine FlAsH par exemple)
lesquels vont réagir directement avec la protéine cible, soit couplés à des « sondes »
(anticorps, SNAP-tag, Halo-tag, etc.). Leur utilisation en RET implique donc d’utiliser
des méthodes qui permettent de marquer spécifiquement les protéines d’intérêt avec 24 Voir la revue Cottet M et al., Frontier in Endocrinology 2012.

Date : 27/03/2012 Page : 46/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
des fluorophores soit donneur soit accepteur d’énergie25. Nous ne décrirons ici que
les techniques couramment utilisées sur la plateforme ARPEGE.
Les sondes de type anticorps
Les anticorps couplés à des fluorophores (anticorps fluorescents) sont utilisés depuis
de nombreuses années en microscopie de fluorescence et en cytométrie de flux.
Depuis les années 2000, les anticorps sont également utilisés comme sondes
fluorescentes pour étudier en RET l’oligomérisation des Récepteurs Couplés aux
Protéines G (RCPG) à la surface cellulaire26. La grande diversité des anticorps et
leur facilité de marquage avec des molécules fluorescentes en font un outil très
puissant pour ce genre d’analyse.
Principe de la méthode : Cas de l’homodimérisation Des cellules exprimant le récepteur membranaire d’intérêt sont incubées avec deux
anticorps monoclonaux portant respectivement les fluorophores donneur et
accepteur d’énergie. Ces deux anticorps sont soit dirigés contre le même épitope27
(étude d’homodimères) soit dirigés contre deux épitopes différents (étude
d’hétérodimères). Si les deux anticorps marqués d’un fluorophore donneur et
accepteur sont dirigés contre le même épitope, il est nécessaire de les utiliser à la
même concentration afin de ne pas favoriser le marquage du récepteur par un
anticorps plutôt que l’autre28. Sous cette condition, les deux anticorps donneur et
accepteur d’énergie ont la même probabilité de marquer les récepteurs d’intérêt.
Ainsi, statistiquement 50% des homodimères seront marqués avec un anticorps
donneur et accepteur d’énergie alors que 25% seront marqués avec deux anticorps
donneur et 25% avec deux anticorps accepteurs d’énergie (Figure 34).
25 Hinner MJ and Johnsson K, Curr Opin Biotechnol. 2010. 26 Maurel D et al., Anal Biochem. 2004. 27 Un épitope est la partie d’un antigène reconnue par un anticorps. Dans le cas de l’étude de l’oligomérisation des RCPG les épitopes sont souvent de petites étiquettes ou tag (HA, FLAG, etc.) insérées au niveau de la séquence codante du domaine N-terminal des RCPG. 28 On considère que le marquage chimique d’un même anticorps avec un fluorophore donneur ou accepteur d’énergie ne modifie pas l’affinité pour son épitope.

Date : 27/03/2012 Page : 47/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
Donneur Accepteur
Donneur Donneur
Accepteur Accepteur
RET
Pas de RET
Pas de RET
50%
25%
25%
Figure 34. Etude de l’homodimérisation des RCPG avec des anticorps fluorescents en RET. Le signal de RET mesuré représente 50% des homodimères présents à la membrane. 25% des homodimères sont respectivement marqués soit avec deux anticorps donneur soit avec deux anticorps accepteurs d’énergie qui ne permettent pas un transfert d’énergie.
Cas de l’hétérodimérisation Pour étudier des hétérodimères de RCPG, deux anticorps monoclonaux distincts
portant les fluorophores donneur et accepteur d’énergie sont généralement utilisés.
Les affinités des deux anticorps pour leurs épitopes respectifs peuvent être
différentes et doivent donc être déterminées au préalable en réalisant des courbes
de saturation29.
Dans le cas où les deux anticorps ont la même affinité pour leur cible alors 100% des
hétérodimères seront ciblés par un anticorps donneur et un anticorps accepteur
d’énergie (Figure 35). L’efficacité du marquage, correspondant au nombre de
récepteurs sur lesquels se fixe un anticorps, dépend de l’affinité des anticorps pour
leurs épitopes. Cette efficacité peut être optimisée en faisant varier la concentration
des anticorps et/ou le temps d’incubation. 29 Les courbes de saturation consistent à incuber jusqu’à l’équilibre des cellules exprimant le récepteur d’intérêt avec des concentrations croissantes d’anticorps. Des cellules n’exprimant pas le récepteur (cellules Mock) sont utilisées pour déterminer le marquage non spécifique des anticorps sur les cellules. Une fois l’équilibre atteint, les cellules sont lavées et le signal spécifique déterminé. Le Kd (constante de dissociation) de l’anticorps pour sa cible est déterminé en représentant le signal spécifique en fonction de la concentration d’anticorps utilisée. A l’équilibre, les vitesses d’association et de dissociation des anticorps sur leur épitope sont égales.

Date : 27/03/2012 Page : 48/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
Donneur Accepteur
RET
100%
Figure 35. Etude de l’hétérodimérisation des RCPG avec des anticorps fluorescents en RET. Le signal de RET mesuré représente 100% des hétérodimères co-marqués (+/- bruit de fond). N.B. : si les récepteurs étudiés ne forment pas que des hétérodimères (existence de
monomères, homodimères, etc.) toutes les paires d’anticorps ne donneront pas un
signal de RET. D’autre part, si l’affinité de l’anticorps pour son épitope se situe autour
du nanomolaire (nM), il faudra incuber les cellules avec une concentration
d’anticorps autour de 10 nM (équivaut à 1.5 g/ml)30 pour être saturant à l’équilibre31
c’est à dire pour marquer 90% des épitopes (récepteurs). Enfin, la gêne stérique peut
diminuer l’accessibilité des épitopes aux anticorps.
Avantages de cette méthode :
Les anticorps monoclonaux utilisés dans les techniques de RET sont généralement
dirigés contre de petites étiquettes (ou tag) de 6 à 15 résidus (HA, myc, FLAG, 6his,
etc.). L’impact de ces étiquettes sur la conformation des récepteurs est généralement
faible surtout lorsque celles-ci sont insérées au niveau des domaines N-terminaux
des récepteurs de type RCPG. De plus, les anticorps disponibles dans le commerce
présentent de bonnes affinités pour leurs épitopes ce qui permet de les utiliser à de
faibles concentrations (1 – 10 nM) réduisant ainsi les problèmes de marquage non
spécifique des cellules.
Dans le cadre de l’étude des phénomènes d’oligomérisation des RCPG, l’utilisation
d’anticorps fluorescents est un atout. En effet, les anticorps sont incapables de
traverser la membrane plasmique et par conséquent, seuls les récepteurs exprimés 30 La masse molaire d’un anticorps de type IgG est de 150000 g/moles. 31 Il a été montré que l’équilibre est atteint au bout de 8h à 4°C pour un anticorps anti-HA-Alexa647 utilisé à 3 nM sur le récepteur HA-V1a (Albizu L. et al., J. Med. Chem. 2006).

Date : 27/03/2012 Page : 49/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
Noyau
Golgi
à la surface des cellules, sont reconnus. Ainsi, les récepteurs intracellulaires encore
engagés dans les voies de synthèse ne sont pas marqués (Figure 36).
Figure 36. Etude de la dimérisation des RCPG avec des anticorps fluorescents en RET. Le signal de RET mesuré provient uniquement de récepteurs exprimés à la surface des cellules qui ont été marqués avec des anticorps fluorescents.
Cette technique peut être utilisée en FRET classique via l’utilisation de fluorophores
organiques ou en TR-FRET via l’utilisation de cryptates de lanthanide (chapitre IV).
Limites de cette méthode :
Les anticorps sont de grosses molécules de 150 KDa qui peuvent potentiellement
perturber le fonctionnement des récepteurs cibles. Leur taille peut aussi avoir une
influence sur les analyses de RET. Schématiquement un anticorps de type
immunoglobuline G (IgG)32 peut être modélisée sous la forme de cylindres33 (Figure
37).
32 Les IgG sont la classe d’immunoglobuline la plus abondante dans le sang (75 à 80 % des anticorps circulants). 33 Tiré de Snapp EL et al., Current Protocols in Cell Biology 2006.

Date : 27/03/2012 Page : 50/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
a/ b/
c/ Figure 37. Taille des anticorps et répartition des fluorophores. a) volume théorique dans lequel peut se trouver un fluorophore fixé à la surface d’un anticorps. b) amplification du signal par les anticorps. c) structures d’un dimère de RCPG (PDB : 1N3M) et d’une immunoglobuline G (PDB : 1IGT).
En tenant compte de la flexibilité de l’anticorps et de la répartition aléatoire des
fluoropohores sur toute sa surface (un anticorps porte en général de 1 à 8
fluorophores) 34 , on peut schématiquement déterminer le volume théorique dans
lequel peut se trouver un fluorophore. Sa grande taille (3-4 fois celle d’un RCPG) et
la distribution aléatoire des fluorophores sur sa surface, font de l’anticorps une
véritable « antenne » qui permet d’augmenter la probabilité du RET (Figure 37,
anticorps versus protéines fluorescentes). Cette amplification est un atout en terme
de sensibilité du signal mais peut aussi potentiellement représenter une limitation en
favorisant un RET non spécifique entre des récepteurs proches mais non associés.
Enfin, le marquage des récepteurs par les anticorps fluorescents est réversible et le
temps d’incubation permettant d’atteindre l’équilibre est variable d’un anticorps à
l’autre ce qui implique des étapes d’optimisation.
34 Les fluorophores organiques sont attachés aléatoirement et de manière covalente par des groupements amines ou thiols sur les anticorps.
Date : 27/03/2012 Page : 51/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
G
O6-methyl
AGT
AGT methyl
G
AGT dégradation
O6-benzylguanine (BG) portant un fluorophore
Guanine
SNAP SNAPprotéine protéine
La technologie SNAP-tag
- La protéine SNAP-tag : Une autre manière de marquer une protéine d’intérêt est d’utiliser une protéine auto-
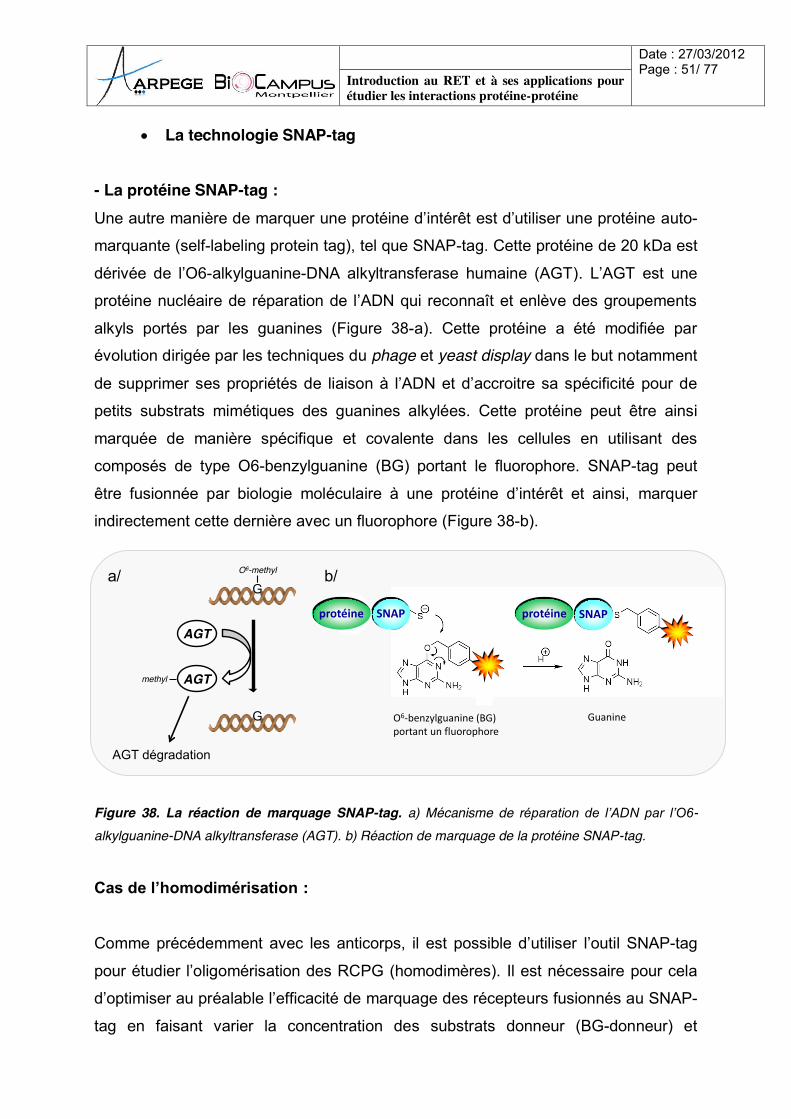
marquante (self-labeling protein tag), tel que SNAP-tag. Cette protéine de 20 kDa est
dérivée de l’O6-alkylguanine-DNA alkyltransferase humaine (AGT). L’AGT est une
protéine nucléaire de réparation de l’ADN qui reconnaît et enlève des groupements
alkyls portés par les guanines (Figure 38-a). Cette protéine a été modifiée par
évolution dirigée par les techniques du phage et yeast display dans le but notamment
de supprimer ses propriétés de liaison à l’ADN et d’accroitre sa spécificité pour de
petits substrats mimétiques des guanines alkylées. Cette protéine peut être ainsi
marquée de manière spécifique et covalente dans les cellules en utilisant des
composés de type O6-benzylguanine (BG) portant le fluorophore. SNAP-tag peut
être fusionnée par biologie moléculaire à une protéine d’intérêt et ainsi, marquer
indirectement cette dernière avec un fluorophore (Figure 38-b).
a/ b/
Figure 38. La réaction de marquage SNAP-tag. a) Mécanisme de réparation de l’ADN par l’O6-alkylguanine-DNA alkyltransferase (AGT). b) Réaction de marquage de la protéine SNAP-tag.
Cas de l’homodimérisation : Comme précédemment avec les anticorps, il est possible d’utiliser l’outil SNAP-tag
pour étudier l’oligomérisation des RCPG (homodimères). Il est nécessaire pour cela
d’optimiser au préalable l’efficacité de marquage des récepteurs fusionnés au SNAP-
tag en faisant varier la concentration des substrats donneur (BG-donneur) et
Date : 27/03/2012 Page : 52/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
-9 -8 -7 -6 -5
0
4000
8000
12000
+ BG-donneur à une concentration fixe
[BG-Accepteur] (log M)
Sign
al d
e FR
ET (u
.a.)
SNAP SNAPSNAP SNAP SNAP SNAP
D AD AAD
Donneur Donneur
Accepteur Accepteur
RET
Pas de RET
Pas de RET
SNAP SNAP
Donneur Accepteur
SNAP SNAP
SNAP SNAP
50%
25%
25%
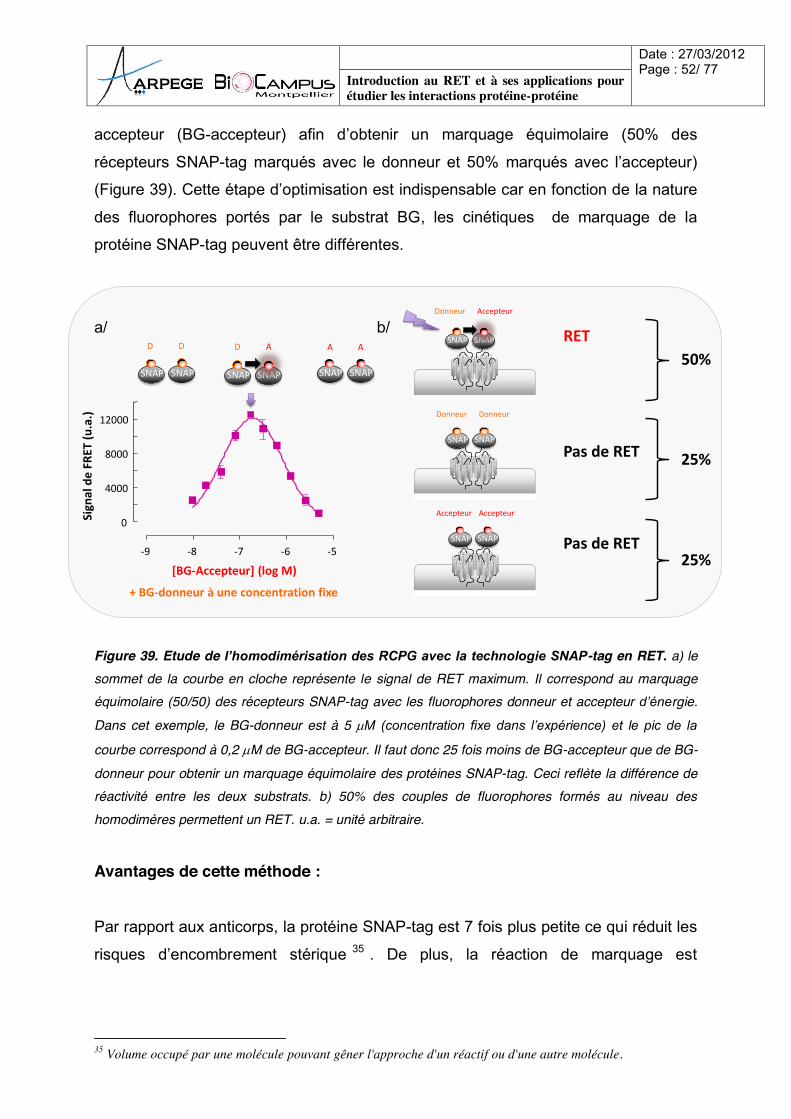
accepteur (BG-accepteur) afin d’obtenir un marquage équimolaire (50% des
récepteurs SNAP-tag marqués avec le donneur et 50% marqués avec l’accepteur)
(Figure 39). Cette étape d’optimisation est indispensable car en fonction de la nature
des fluorophores portés par le substrat BG, les cinétiques de marquage de la
protéine SNAP-tag peuvent être différentes.
a/ b/
Figure 39. Etude de l’homodimérisation des RCPG avec la technologie SNAP-tag en RET. a) le sommet de la courbe en cloche représente le signal de RET maximum. Il correspond au marquage équimolaire (50/50) des récepteurs SNAP-tag avec les fluorophores donneur et accepteur d’énergie. Dans cet exemple, le BG-donneur est à 5 M (concentration fixe dans l’expérience) et le pic de la
courbe correspond à 0,2 M de BG-accepteur. Il faut donc 25 fois moins de BG-accepteur que de BG-donneur pour obtenir un marquage équimolaire des protéines SNAP-tag. Ceci reflète la différence de réactivité entre les deux substrats. b) 50% des couples de fluorophores formés au niveau des homodimères permettent un RET. u.a. = unité arbitraire. Avantages de cette méthode :
Par rapport aux anticorps, la protéine SNAP-tag est 7 fois plus petite ce qui réduit les
risques d’encombrement stérique 35 . De plus, la réaction de marquage est
35 Volume occupé par une molécule pouvant gêner l'approche d'un réactif ou d'une autre molécule.

Date : 27/03/2012 Page : 53/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
irréversible, rapide36 et stœchiométrique (un seul fluorophore par protéine cible) ce
qui permet d’obtenir des résultats quantitatifs (détermination du nombre de
récepteurs marqués). De nombreux substrats fluorescents perméants et non
perméants 37 sont commercialement disponibles ce qui permet d’envisager des
marquages intracellulaires (substrats perméants) et des marquages spécifiquement
de surface (substrats non perméants). Enfin, il est possible de sélectionner très
facilement différents couples de fluorophores pour des analyses de RET en
changeant simplement les couples de substrats fluorescents sans avoir à refaire les
protéines recombinantes.
Limites de cette méthode :
Bien que la protéine SNAP-tag soit plus petite qu’un anticorps, sa taille reste
toutefois suffisamment importante pour potentiellement générer de l’encombrement
stérique. De plus, sa structure (domaines N- et C-terminaux distants), associée à sa
taille, rend difficile l’insertion du SNAP-tag dans une protéine sans que cela n’affecte
son repliement ou sa fonction. Enfin, comme pour les anticorps il est nécessaire
d’optimiser les conditions de marquage.
Pour ne pas interférer avec l’activité des récepteurs étudiés, le SNAP-tag est
généralement inséré au niveau du domaine N-terminal du RCPG. Lorsque l’on
réalise des études de dimérisation par RET il est essentiel de prendre en compte la
taille de ce domaine dans l’analyse des résultats. En effet, les RCPG de classe A
(récepteurs adrénergiques, etc.), ont un petit domaine N-terminal faisant une
trentaine d’acides aminés alors que les récepteurs de classe C (récepteurs
métabotropiques du glutamate, etc.) ont un long domaine N-terminal très bien
structuré de 600 acides aminés environ. Deux récepteurs avec des domaines N-
terminaux différents pourront donc donner des résultats de RET très variables alors
même que les récepteurs forment des dimères (Figure 40).
36 Généralement autour d’une heure d’incubation pour des concentrations oscillant entre 100 nM et 1 M selon les substrats (BG-fluorescéine, BG-Cy5, etc.). 37 Propriété des substrats à traverser ou non la membrane plasmique des cellules.

Date : 27/03/2012 Page : 54/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
SNAP-tag k (M-1s-1)*
BG 100000
BC 30
CLIP-tag k (M-1s-1)*
BG < 1
BC 1000
SNAP SNAP SNAP
SNAPRET
Pas de RET
Figure 40. Analyse de la dimérisation des RCPG avec la méthode SNAP-tag en RET. Panel gauche : deux RCPG avec de petits domaines N-terminaux. Panel droite : deux RCPG avec des domaines N-terminaux très différents.
Cas de l’hétérodimérisation : En 2008, un mutant du SNAP-tag, nommé CLIP-tag, réagissant spécifiquement avec
l’O2-benzylcytosine (BC), un substrat différent de celui du SNAP-tag, a été
développé38. L’introduction du CLIP-tag marque une évolution importante de cette
technologie car il permet de réaliser un marquage spécifique de deux récepteurs
différents chacun avec un ou deux fluorophores distincts. Pour conserver la
spécificité du marquage (orthogonalité) SNAP-tag/CLIP-tag, il est indispensable
d’ajouter simultanément les substrats BG et BC. En effet, SNAP-tag, à l’inverse du
CLIP-tag, n’est pas parfaitement sélectif puisqu’il peut réagir avec le substrat BC du
CLIP-tag (voir tableau ci-dessous).
* La vitesse de réaction k représente la vitesse de transfert du groupement benzyl-fluorophore porté par la guanine (BG) ou la cytosine (BC) au niveau des protéines SNAP-tag et CLIP-tag. Plus la valeur de k est grande plus le marquage est rapide.
N.B. : des mutants plus rapides du SNAP-tag et du CLIP-tag sont commercialisés
par la société New England Biolabs sous la dénomination, SNAPfast et CLIPfast.
38 Gautier et al., Chemestry and Biology 2008.

Date : 27/03/2012 Page : 55/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
RETSNAP CLIP
Donneur Accepteur
100%
Avantages de cette méthode :
Dans le cadre de l’analyse de l’hétérodimérisation des RCPG, l’utilisation du couple
SNAP-tag/CLIP-tag simplifie considérablement l’analyse des résultats (pour
comparaison voir Figures 39 et 41).
Figure 41. Etude de l’hétérodimérisation des RCPG avec la technologie SNAP-tag/CLIP-tag en RET. Le signal de RET mesuré représente 100% des hétérodimères SNAP-tag et CLIP-tag co-marqués avec un fluorophore donneur et un fluorophore accepteur d’énergie.
Limites de cette méthode :
La réactivité du CLIP-tag pour le BC est beaucoup plus faible que celle du SNAP-tag
pour le BG (voir tableau ci-dessus). Par conséquent, il est nécessaire d’utiliser de
plus hautes concentrations de BC pour marquer CLIP-tag ce qui peut poser des
problèmes de marquage non spécifique des cellules (signal sur bruit). De plus, le
marquage SNAP-tag/CLIP-tag ne sera spécifique que si les deux substrats sont
ajoutés simultanément ou si SNAP-tag est d’abord marqué avec le BG puis CLIP-tag
avec le BC.
B/ LE BRET39
Principe de la méthode :
La technique de BRET, décrite dans le chapitre IV, permet d’analyser un grand
nombre d’interactions moléculaires tant pour des protéines exprimées à la surface
cellulaire que pour des protéines intracellulaires. Les avantages et limites de cette
39 Voir la revue Drinovec L. et al., Frontier in Endocrinology 2012.

Date : 27/03/2012 Page : 56/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
Noyau
ReticulumEndoplasmique
Golgi
YFPRluc
méthode qui sont discutés dans cette partie ne concernent que l’étude des dimères
de RCPG.
Avantages de cette méthode : Les fluorophores sont directement synthétisés par les cellules ce qui évite des
étapes de marquage. Par conséquent aucune optimisation du marquage des
protéines d’intérêt n’est nécessaire. D’autre part, la Rluc et la YFP peuvent être
insérées dans différentes régions des récepteurs notamment dans les boucles
intracellulaires (pour analyser par exemple les changements de conformation du
récepteur).
Limites de cette méthode :
Contrairement aux techniques précédentes, il n’est pas possible en BRET de séparer
le signal provenant des récepteurs de surface de celui des récepteurs retenus dans
les compartiments intracellulaires dont la forte proximité peut générer un signal non
spécifique (Figure 42). Le signal de BRET mesuré reflète donc les interactions de
l’ensemble des récepteurs qui sont produits par les cellules.
Figure 42. Etude de la dimérisation des RCPG en BRET. Le signal de RET mesuré provient de l’ensemble des récepteurs exprimés dans les cellules.

Date : 27/03/2012 Page : 57/ 77
Introduction au RET et à ses applications pour étudier les interactions protéine-protéine
Rluc YFP
BRETYFP
Pas de BRET
Rluc
YFP
BRET
Rluc
Pour étudier la dimérisation des RCPG, la Renilla Luciferase (Rluc) et la YFP sont en
général insérées dans la région carboxy-terminale des récepteurs car elles
n’affectent pas leur fonction à cette position. La longueur de ce domaine pour les
RCPG est variable d’un récepteur à l’autre. Elle peut aller de 25 à 150 acides aminés
en moyenne ce qui correspond à une longueur de 80 à 480 angströms (Å) en
considérant une forme linéaire de ce domaine. Par conséquent, deux protéines avec
des domaines C-terminaux de longueurs variables, peuvent donner des signaux de
BRET très différents alors même qu’elles sont en interaction (Figure 42).
Figure 42. Analyse de la dimérisation de RCPG en BRET.
En terme de taille, les protéines fluorescentes font 27 KDa de poids moléculaire et
peuvent être assimilées à des cylindres de 40 Å (angströms) de haut et de 30 Å de
diamètre ce qui correspond approximativement aux dimensions des 7 hélices
transmembranaires d’un RCPG inséré dans la membrane plasmique. La Rluc quand
à elle fait 34 KDa et peut être assimilée à une sphère de 40 Å de diamètre.
Related Documents











