USING QUANTITATIVE PCR TO DETERMINE THE DISTRIBUTION OF A SEMICRYPTIC BENTHIC DIATOM, NAVICULA PHYLLEPTA (BACILLARIOPHYCEAE) 1 Ve´ronique Cre´ach 2 , Anneliese Ernst Netherlands Institute of Ecology, (NIOO-KNAW), PO Box 140, 4400 AC, Yerseke, The Netherlands Koen Sabbe, Bart Vanelslander, Wim Vyverman Laboratory of Protistology & Aquatic Ecology, Department of Biology, Ghent University, Krijgslaan 281-S8, B-9000 Ghent, Belgium and Lucas J. Stal Netherlands Institute of Ecology, (NIOO-KNAW), PO Box 140, 4400 AC, Yerseke, The Netherlands Evidence has accumulated during the last decade showing that many established diatom morpho-spe- cies actually consist of several semicryptic or truly cryptic species. As these species are difficult or even impossible to differentiate by microscopic analysis, there is virtually no information on how they behave in natural environments. In this study, we devel- oped a quantitative real-time PCR (qPCR) assay using TaqMan probes s targeted to the internal tran- scribed spacer 1 (ITS1) to assess the spatial distri- bution and seasonal dynamics of an important component of the microphytobenthos of intertidal sediments. Navicula phyllepta Ku ¨tzing is a brackish- marine morpho-species with a cosmopolitan distri- bution. Axenic clones of this species were isolated from natural assemblages of benthic diatoms at dif- ferent intertidal stations in the Westerschelde estu- ary (The Netherlands). At least two distinct semicryptic species of N. phyllepta were present, as shown by differences in the quantity of DNA per cell, the ITS1 sequences and the copy number of ITS per cell. DNA and chl a concentrations extract- ed from sediment surface samples were closely cor- related, showing that the DNA used for subsequent analysis mostly belonged to the microalgal commu- nity. The results of real-time qPCR from sites throughout the estuary and over several seasons agreed well with microscopic counts. Additionally, the seasonal pattern of the two forms of N. phyllepta showed an overlapping, but unique distribution along the estuary. Key index words: diatoms; estuary; Navicula phylle- pta; niche differentiation; real-time qPCR; semi- cryptic species Benthic diatoms are one of the most important groups of photoautotrophic microorganisms in estua- rine intertidal sediments where they play a major role in ecosystem functioning (Underwood and Kromkamp 1999). Several investigations have shown that benthic diatoms are an essential component of the food web of intertidal mudflats (Middelburg et al. 2000) as well as of the water column after re-suspension (Lucas et al. 2000). Many benthic diatom taxa are well adapted to fluctuating environmental conditions (Admiraal et al. 1984, Colijn and de Jonge 1984), which might explain their cosmopolitan distribution (Round 2004). The growth rate and the rates of carbon fixation of benthic diatoms can equal those of pelagic species (Barranguet and Kromkamp 2000, Brandini et al. 2001). The presence of benthic biofilms may control the rate of nutrient exchange between the sediment and the water column (Su ¨ndback et al. 2000). Moreo- ver, biofilms of benthic diatoms have been shown to contribute to the stabilization of estuarine sediment surfaces by causing an increase of the erosion thresh- old (Tolhurst et al. 2003). Biofilms of microphytobenthos in intertidal mud- flats of the Westerschelde estuary (the Netherlands) are generally dominated by raphid, motile pennate dia- toms (epipelic diatoms) (Muylaert et al. 2002, Forster et al. 2006). These diatoms have been shown to belong to a monophyletic group of organisms (Medlin and Kaczmarska 2004, Sorhannus 2004) that appeared in the middle of the Eocene about 55 million years ago (Medlin et al. 1993). The majority of the epipelic diatoms in temperate intertidal mudflats belong to the order of Naviculales (Montani et al. 2003, Haubois et al. 2005). The microscopic identification of Navicula species is cumbersome because of the morphological plasticity inherent to their life cycle and because some characteristics can only be discerned by scanning elec- tron microscopy (Cox 1997, 1998, Mann 1999). The morpho-species Navicula phyllepta Ku ¨tzing is often a key species in intertidal mudflats. It can reach 60%–75% of the biomass of the total microphytobenthic community 1 Received 23 November 2005. Accepted 3 July 2006. 2 Author for correspondence and present address: Center for En- vironment, Fisheries and Aquaculture Science (Cefas), Pakefield Road, Lowestoft, Suffolk NR33 0HT, UK. e-mail [email protected]. 1142 J. Phycol. 42, 1142–1154 (2006) r 2006 by the Phycological Society of America DOI: 10.1111/j.1529-8817.2006.00268.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

USING QUANTITATIVE PCR TO DETERMINE THE DISTRIBUTION OF ASEMICRYPTIC BENTHIC DIATOM, NAVICULA PHYLLEPTA (BACILLARIOPHYCEAE)1
Veronique Creach2, Anneliese Ernst
Netherlands Institute of Ecology, (NIOO-KNAW), PO Box 140, 4400 AC, Yerseke, The Netherlands
Koen Sabbe, Bart Vanelslander, Wim Vyverman
Laboratory of Protistology & Aquatic Ecology, Department of Biology, Ghent University, Krijgslaan 281-S8, B-9000 Ghent, Belgium
and
Lucas J. Stal
Netherlands Institute of Ecology, (NIOO-KNAW), PO Box 140, 4400 AC, Yerseke, The Netherlands
Evidence has accumulated during the last decadeshowing that many established diatom morpho-spe-cies actually consist of several semicryptic or trulycryptic species. As these species are difficult or evenimpossible to differentiate by microscopic analysis,there is virtually no information on how they behavein natural environments. In this study, we devel-oped a quantitative real-time PCR (qPCR) assayusing TaqMan probess targeted to the internal tran-scribed spacer 1 (ITS1) to assess the spatial distri-bution and seasonal dynamics of an importantcomponent of the microphytobenthos of intertidalsediments. Navicula phyllepta Kutzing is a brackish-marine morpho-species with a cosmopolitan distri-bution. Axenic clones of this species were isolatedfrom natural assemblages of benthic diatoms at dif-ferent intertidal stations in the Westerschelde estu-ary (The Netherlands). At least two distinctsemicryptic species of N. phyllepta were present,as shown by differences in the quantity of DNA percell, the ITS1 sequences and the copy number ofITS per cell. DNA and chl a concentrations extract-ed from sediment surface samples were closely cor-related, showing that the DNA used for subsequentanalysis mostly belonged to the microalgal commu-nity. The results of real-time qPCR from sitesthroughout the estuary and over several seasonsagreed well with microscopic counts. Additionally,the seasonal pattern of the two forms of N. phylleptashowed an overlapping, but unique distributionalong the estuary.
Key index words: diatoms; estuary; Navicula phylle-pta; niche differentiation; real-time qPCR; semi-cryptic species
Benthic diatoms are one of the most importantgroups of photoautotrophic microorganisms in estua-rine intertidal sediments where they play a major rolein ecosystem functioning (Underwood and Kromkamp1999). Several investigations have shown that benthicdiatoms are an essential component of the food web ofintertidal mudflats (Middelburg et al. 2000) as well asof the water column after re-suspension (Lucas et al.2000). Many benthic diatom taxa are well adapted tofluctuating environmental conditions (Admiraal et al.1984, Colijn and de Jonge 1984), which might explaintheir cosmopolitan distribution (Round 2004).
The growth rate and the rates of carbon fixation ofbenthic diatoms can equal those of pelagic species(Barranguet and Kromkamp 2000, Brandini et al.2001). The presence of benthic biofilms may controlthe rate of nutrient exchange between the sedimentand the water column (Sundback et al. 2000). Moreo-ver, biofilms of benthic diatoms have been shown tocontribute to the stabilization of estuarine sedimentsurfaces by causing an increase of the erosion thresh-old (Tolhurst et al. 2003).
Biofilms of microphytobenthos in intertidal mud-flats of the Westerschelde estuary (the Netherlands) aregenerally dominated by raphid, motile pennate dia-toms (epipelic diatoms) (Muylaert et al. 2002, Forsteret al. 2006). These diatoms have been shown to belongto a monophyletic group of organisms (Medlin andKaczmarska 2004, Sorhannus 2004) that appeared inthe middle of the Eocene about 55 million years ago(Medlin et al. 1993). The majority of the epipelicdiatoms in temperate intertidal mudflats belong tothe order of Naviculales (Montani et al. 2003, Hauboiset al. 2005). The microscopic identification of Naviculaspecies is cumbersome because of the morphologicalplasticity inherent to their life cycle and because somecharacteristics can only be discerned by scanning elec-tron microscopy (Cox 1997, 1998, Mann 1999). Themorpho-species Navicula phyllepta Kutzing is often a keyspecies in intertidal mudflats. It can reach 60%–75% ofthe biomass of the total microphytobenthic community
1Received 23 November 2005. Accepted 3 July 2006.2Author for correspondence and present address: Center for En-
vironment, Fisheries and Aquaculture Science (Cefas), Pakefield Road,Lowestoft, Suffolk NR33 0HT, UK. e-mail [email protected].
1142
J. Phycol. 42, 1142–1154 (2006)r 2006 by the Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2006.00268.x
(Thornton et al. 2002, Haubois et al. 2005). N. phylleptahas a widespread distribution, suggesting adaptationacross a range of environmental conditions such as sa-linity, emersion time, and temperature (Krammer andLange-Bertalot 1986, Witkowski et al. 2000, Lange-Be-rtalot 2001, Sabbe et al. 2003).
Ribosomal genes are frequently used for phyloge-netic reconstructions. The nuclear ribosomal operonof diatoms has the structure 18S rDNA–ITS1–5.8SrDNA–ITS2–28S rDNA (Zechman et al. 1994) andmay be present in multiple copies (Armbrust et al.2004). While both 18S and 28S rRNA genes have beenused for the reconstruction of the phylogeny of dia-toms, the ITS region, comprising ITS1–5.8S rRNAgene–ITS2 has been proven to be useful for molecularanalyses at the species level or beyond. Additionally,ITS has been used to resolve intra- and interspecificrelationships of a variety of eukaryotes, including dia-toms (Zechman et al. 1994, Behnke et al. 2004, Orsiniet al. 2004), and the phylogenetic relationships be-tween populations and their biogeographical reparti-tion (Bakker et al. 1992, Kooistra et al. 1992). Thebroad ecological range of N. phyllepta suggests that thisspecies complex may be composed of more than onetaxon. Sequence analysis of the ITS from a variety ofisolated strains demonstrated that the morpho-speciesN. phyllepta actually consists of two well-separated clus-ters which can be regarded as semicryptic taxa as theyshow subtle but stable morphological differences suchas size, difference in valves width and stria density(B. Vanelslander et al. unpublished data).
In this study, a real-time quantitative PCR (real-timeqPCR) assay using primers targeting part of the ITS1region and Taqmans probes was developed in order
to measure the abundance of the two semicrypticforms of N. phyllepta along the salinity gradient in theWesterschelde estuary. We determined their distinctspatial and seasonal distribution patterns along the es-tuary and compared the results with microscopiccounts. Real-time qPCR has been applied in environ-mental studies to quantify the number of copies ofa specific gene and to estimate the number of cells ofa particular microorganism containing the targetedgene. It had been also used to quantify the numberof transcripts of the target gene as a measure ofthe potential metabolic activity coded by that specificgene by using reverse transcriptase PCR in an initialstep. These approaches have been successfully usedin ecological studies on diatoms (Leblanc et al. 1999,Wawrik et al. 2002, Wei et al. 2004), dinoflagellates(Bowers et al. 2000, Galluzzi et al. 2004), cyano-bacteria (Becker et al. 2000, Suzuki et al. 2000), andbacteria (Labrenz et al. 2004, Skovhus et al. 2004).Quantitative PCR offers all the advantages of conven-tional PCR, such as high sensitivity (Becker et al. 2002,Newby et al. 2003), reproducibility, and specificity.
MATERIALS AND METHODS
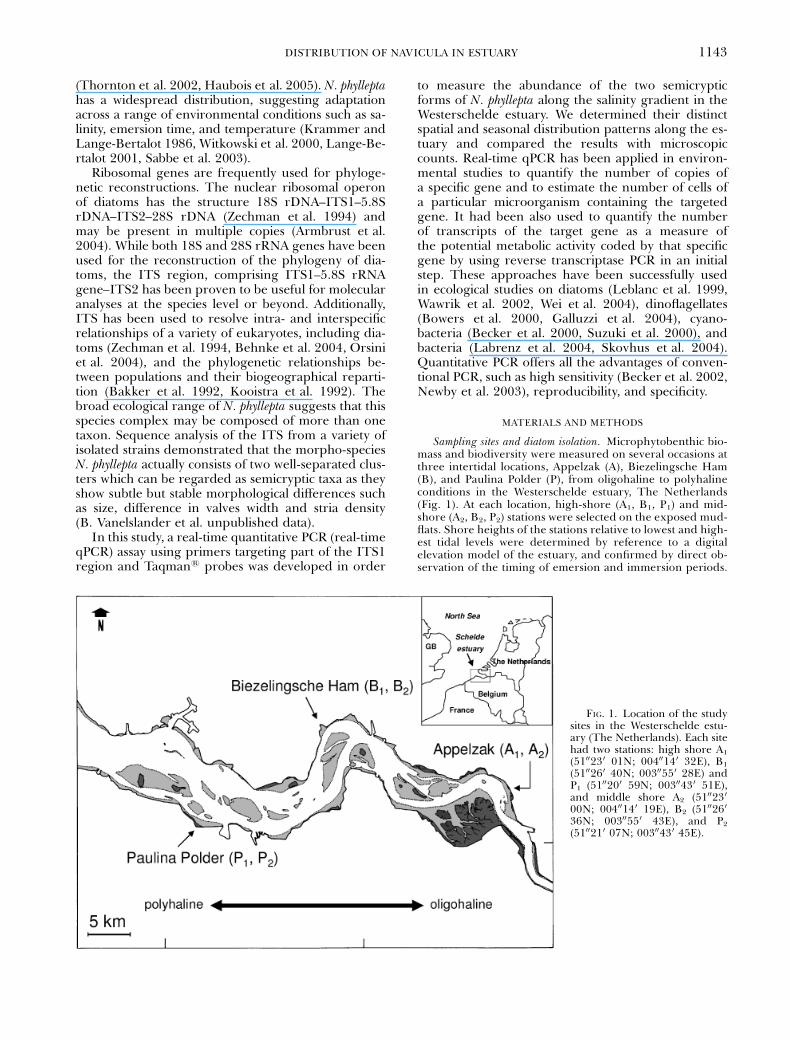
Sampling sites and diatom isolation. Microphytobenthic bio-mass and biodiversity were measured on several occasions atthree intertidal locations, Appelzak (A), Biezelingsche Ham(B), and Paulina Polder (P), from oligohaline to polyhalineconditions in the Westerschelde estuary, The Netherlands(Fig. 1). At each location, high-shore (A1, B1, P1) and mid-shore (A2, B2, P2) stations were selected on the exposed mud-flats. Shore heights of the stations relative to lowest and high-est tidal levels were determined by reference to a digitalelevation model of the estuary, and confirmed by direct ob-servation of the timing of emersion and immersion periods.
FIG. 1. Location of the studysites in the Westerschelde estu-ary (The Netherlands). Each sitehad two stations: high shore A1
(5100230 01N; 00400140 32E), B1
(5100260 40N; 00300550 28E) andP1 (5100200 59N; 00300430 51E),and middle shore A2 (5100230
00N; 00400140 19E), B2 (5100260
36N; 00300550 43E), and P2
(5100210 07N; 00300430 45E).
DISTRIBUTION OF NAVICULA IN ESTUARY 1143
Characteristics of the sites are listed in Table 1. Each stationwas sampled nine times between April 2002 and September2003.
In 2002, diatoms were isolated from all six stations. Biofilmsamples were suspended in seawater from the same locationand this suspension was subsequently spread on 1.5% agaroseplates (Bacto Agar, Brunschwig Chemie, Germany) amendedwith F/2 growth medium (Guillard and Ryther 1962). After 2–4weeks of incubation at 171 C at a 14:10 light:dark (L:D) cycle(light source Cool White 36W compact fluorescent tubes,Philips, The Netherlands) at 80–120mmol photons �m�2 � s�1,individual colonies were picked and sub-cultivated on agaroseplates as well as in liquid medium (f/2 growth medium). Dilu-tion-plating and picking of colonies were repeated until axeniccultures were obtained (Table 2). This was checked by micro-scopic examination, and by the absence of bacterial growth onagarose plates amended with Bacto yeast extract and peptone.
Microscopy. The species composition in the micro-phytobenthos of the Westerschelde was determined from sur-face sediment samples (upper 2 mm, 18 cm2), removed with acontact corer (Ford and Honeywill 2002). At each station, with-in an area of approximately 25 m2, five samples were taken andpooled. In order to visualize the ultrastructural features of thesiliceous cell walls, aliquots of samples from each site wereoxidized with a 1:1 mixture of hydrogen peroxide (30%) andacetic acid (100%). The oxidized material was mounted in Nap-hrax (PhycoTech, St. Joseph, MI, USA). Approximately 300diatom valves (range, 277–336) were identified and countswere made by using a Leitz Diaplan microscope equippedwith Differential Interference Contrast.
Cultivated diatoms were identified as described above. Forquantitative extraction of DNA, cells of three cultures of twoisolates of N. phyllepta (CCY 0222 and CCY 0212) were enu-merated in a Burker chamber. From each culture, four aliquots
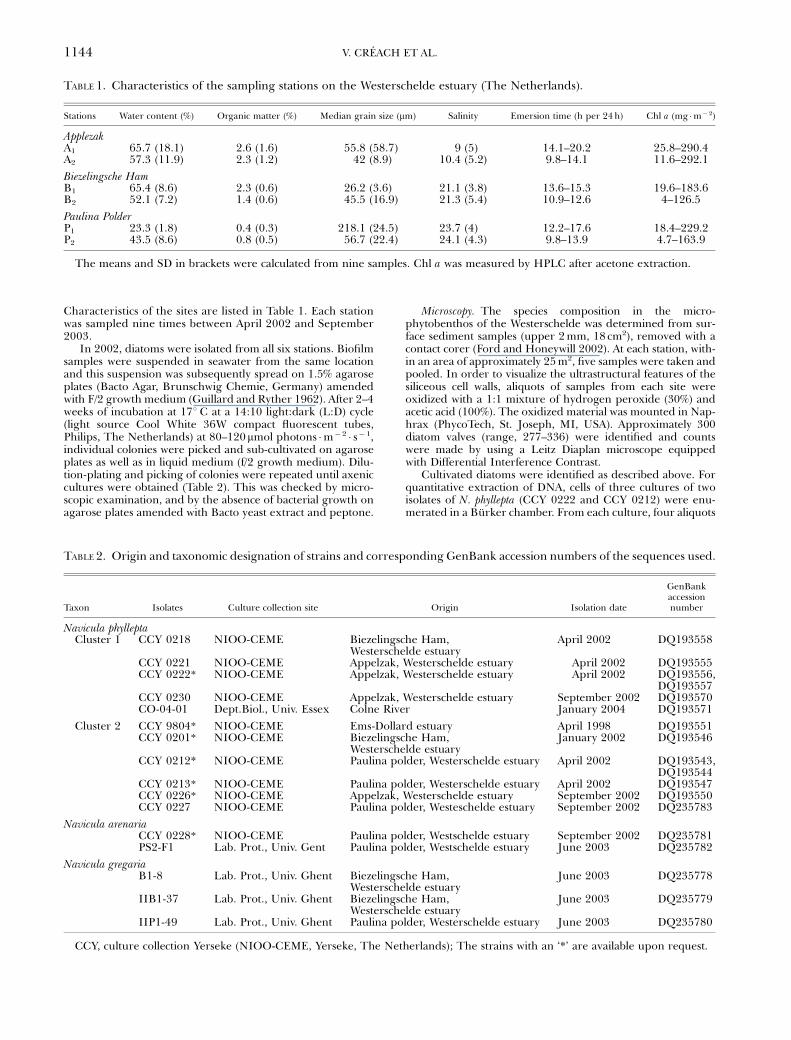
TABLE 1. Characteristics of the sampling stations on the Westerschelde estuary (The Netherlands).
Stations Water content (%) Organic matter (%) Median grain size (mm) Salinity Emersion time (h per 24 h) Chl a (mg �m� 2)
ApplezakA1 65.7 (18.1) 2.6 (1.6) 55.8 (58.7) 9 (5) 14.1–20.2 25.8–290.4A2 57.3 (11.9) 2.3 (1.2) 42 (8.9) 10.4 (5.2) 9.8–14.1 11.6–292.1
Biezelingsche HamB1 65.4 (8.6) 2.3 (0.6) 26.2 (3.6) 21.1 (3.8) 13.6–15.3 19.6–183.6B2 52.1 (7.2) 1.4 (0.6) 45.5 (16.9) 21.3 (5.4) 10.9–12.6 4–126.5
Paulina PolderP1 23.3 (1.8) 0.4 (0.3) 218.1 (24.5) 23.7 (4) 12.2–17.6 18.4–229.2P2 43.5 (8.6) 0.8 (0.5) 56.7 (22.4) 24.1 (4.3) 9.8–13.9 4.7–163.9
The means and SD in brackets were calculated from nine samples. Chl a was measured by HPLC after acetone extraction.
TABLE 2. Origin and taxonomic designation of strains and corresponding GenBank accession numbers of the sequences used.
Taxon Isolates Culture collection site Origin Isolation date
GenBankaccessionnumber
Navicula phylleptaCluster 1 CCY 0218 NIOO-CEME Biezelingsche Ham,
Westerschelde estuaryApril 2002 DQ193558
CCY 0221 NIOO-CEME Appelzak, Westerschelde estuary April 2002 DQ193555CCY 0222* NIOO-CEME Appelzak, Westerschelde estuary April 2002 DQ193556,
DQ193557CCY 0230 NIOO-CEME Appelzak, Westerschelde estuary September 2002 DQ193570CO-04-01 Dept.Biol., Univ. Essex Colne River January 2004 DQ193571
Cluster 2 CCY 9804* NIOO-CEME Ems-Dollard estuary April 1998 DQ193551CCY 0201* NIOO-CEME Biezelingsche Ham,
Westerschelde estuaryJanuary 2002 DQ193546
CCY 0212* NIOO-CEME Paulina polder, Westerschelde estuary April 2002 DQ193543,DQ193544
CCY 0213* NIOO-CEME Paulina polder, Westerschelde estuary April 2002 DQ193547CCY 0226* NIOO-CEME Appelzak, Westerschelde estuary September 2002 DQ193550CCY 0227 NIOO-CEME Paulina polder, Westeschelde estuary September 2002 DQ235783
Navicula arenariaCCY 0228* NIOO-CEME Paulina polder, Westschelde estuary September 2002 DQ235781PS2-F1 Lab. Prot., Univ. Gent Paulina polder, Westschelde estuary June 2003 DQ235782
Navicula gregariaB1-8 Lab. Prot., Univ. Ghent Biezelingsche Ham,
Westerschelde estuaryJune 2003 DQ235778
IIB1-37 Lab. Prot., Univ. Ghent Biezelingsche Ham,Westerschelde estuary
June 2003 DQ235779
IIP1-49 Lab. Prot., Univ. Ghent Paulina polder, Westerschelde estuary June 2003 DQ235780
CCY, culture collection Yerseke (NIOO-CEME, Yerseke, The Netherlands); The strains with an ‘*’ are available upon request.
V. CREACH ET AL.1144

were counted four times and the average number of cells permilliliter were calculated.
Chlorophyll a and DNA extraction. From the same stations asdescribed above, five individual surface sediment samples(upper 2 mm, 18 cm2) were removed with a contact corer.The samples were immediately frozen in liquid nitrogen,freeze-dried in the dark and stored at � 801 C until the anal-ysis. Algal biomass was estimated from measurement of chl a.Pigments were extracted with 90% acetone from aliquots of100 mg of freeze-dried sediment. Mechanical disruptionusing 1 mm beads in a Bead Beater for 20 s ensured theefficient release of pigments. Chlorophyll a was quantifiedfrom the acetone extracts by using HPLC following the pro-cedure described in Rijstenbil (2003) and expressed as thechl content per dry weight of sediment (mg chl a �g�1 dryweight).
Two samples were selected for each station for DNA ex-traction from the freeze-dried contact cores. Nucleic acids wereextracted from approximately 100 to 200 mg dry sediment us-ing the UltraCleant Soil DNA Kit (MoBio Laboratories, Sol-ana Beach, CA, USA). An extra elution step was added to themanufacturer’s instruction to ensure quantitative removal ofDNA from the purification columns (final volume 200mL). Thequality of the nucleic acid was evaluated on a 1% agarose gel(Sigma), run in 1 � TAE buffer and stained by SYBR Gold(Molecular Probes, Leiden, Zwijndrecht, The Netherlands).DNA was quantified with the PicoGreen dsDNA quantificationKit (Molecular Probes) using a microplate reader (FluostarGalaxy, BMG Labtechnologies, Offenburg, Germany).
A similar procedure was used to extract and quantify DNAcontent from N. phyllepta strain CCY 0222 and strain CCY0212. Cells of three cultures of each strain were counted (seeabove), and two aliquots (2 and 4 mL of strain CCY 0222 and 1and 2 mL of strain CCY 0212) of each culture were harvestedby centrifugation (5000g for 10 min). DNA was extracted andquantified as described above.
PCR Primers, TaqMans probes, and real-time qPCR as-says. The entire ITS region was amplified using the forwardprimer in the end of the 18S rDNA (5 0-GGAAGGT-GAAGTCGTAACAAGG-3 0) and reverse primer in the begin-ning of the 28S rDNA (5 0-CTNGTTAGTTTCTTTTCCT-CCG-3 0). The 1021–1084 bp PCR products of ITS werecloned in PCR II-TOPO vector TA cloning kit (Invitrogen,Breda, The Netherlands) according to the manufacturer’sinstructions. Vector primers (T7 forward and T3 reverse)were used as sequencing primers. Sequences were obtainedusing the ABI 3100 prisms BigDyes Terminator CycleSequencing Ready Reaction Kit (Applied Biosystems, Ni-euwerkerk, The Netherlands). The ITS1 sequences of isolat-ed diatoms (accession numbers see Table 2) were alignedusing ClustalX (Thompson et al. 1997). Priming sites andtarget sequences of the probes specific for the two phyloge-netic clusters of N. phyllepta were identified by comparison
with sequences of other Navicula species (Table 2). Primersand probes (Table 3) were designed using the program Prim-er Premier 5 (Premier Biosoft Int., Palo Alto, CA, USA) andthe services of Tib-Molbiol (Berlin, Germany). TaqMans
probes were 5 0-labeled with the fluorescent dye FAMt and3 0-labeled with the quencher TAMRAt (Tib-Molbiol).
Real-time qPCR assays were performed with the Lightcy-clers (Roche Diagnostics, Almere, The Netherlands) using theLightcycler FastStart DNA Masterplus Hybridization Probes andthe Lightcycler FastStart DNA SYBR Green I (Roche Diagnos-tics). The kits were supplemented with our custom primers(final concentration, 0.5mM each) and probes (final concentra-tion, 0.2mM each) and contained 5mL of DNA in a final volumeof 20mL. The PCR program comprised an initial 10 min de-naturation step at 951 C and 40 cycles consisting of a 15 s de-naturation step at 951 C and a 1 min annealing/polymerizationstep either at 601 C (cluster 1) or 631 C (cluster 2) for both assayformat, respectively. The two steps were connected by heatingand cooling speeds of 101 and 201 C/s, respectively. Finally, thePCR products were cooled down to 401 C. Each PCR run in-cluded a standard curve, established with a serial dilution oflinear plasmids from cloned target sequences, ITS1, repre-sentative for cluster 1 (strain CCY 0222) or cluster 2 (strainCCY 0212), respectively. Controls without templates (H2O)and with non-target DNA (105 copies of the plasmid with theinsert from the other cluster) were also included.
Quantification of the amount of target DNA in sedimentsamples was accomplished by measuring the threshold cycle(CT, Heid et al. 1996) and using the standard curve to deter-mine the starting copy number. The entire process of calcu-lating CTs, preparing a standard curve, and determiningstarting copy number for unknown samples was performedby the software of the Lightcycler Software version 3.5 (RocheMolecular Biochemicals, Indianapolis, IN, USA).
RESULTS
Efficiency of the DNA extraction from sediments and itsrelationship with chl a. The efficiency of DNA extrac-tion from mudflat sediments was tested with fivefreeze-dried contact cores collected at different sam-pling sites in the Westerschelde. The samples werechosen according to their silt content (minimum 5%,maximum 87%) and median grain size (21–220 mm).Three to four DNA extractions were processed fromeach sample using from 20 to 200 mg of dry sedi-ment. In all cases, DNA recovery was proportional tothe quantity of sediment treated. The correlation co-efficients were slightly higher in sediments contain-ing 50%–70% of sand (r2 5 0.99, P<0.001, n 5 6) than
TABLE 3. List of primers and probes used for the real-time qPCR assays.
Clusters Sequences (50 ! 30) Tm (1 C)
Cluster 1 (CCY 0222)Primers DITS142F TGGCCTCCTATCTTTGGG 56
DITS1127R* AATACTGGGTTTGGCGTTT 52.4Probe ITS1R* TCGCAAGTGAGCTTGGCAGC 63.8
Cluster 2 (CCY 0212)Primers DITS15F CACACCTTYGATCCAAGATCAAT 58
DITS1113R* CGTTTCAGATTCGCAGCA 52.4Probe ITS1F CCCGTGAACYTGGCCTCCTATC 62.7
Specific primers or probes are specified by an ‘*’.
DISTRIBUTION OF NAVICULA IN ESTUARY 1145

in muddy sediments containing at least 70% of siltand clay (r2 5 0.94, P<0.001, n 5 14).
As a measure for the biomass of the micro-phytobenthos, we determined the chl a content offreeze-dried contact cores collected at the samplingsites during the nine sampling campaigns betweenMay 2002 and September 2003. The DNA and chl acontent expressed on the basis of sediment dry weightvaried between 2.8 and 80 ng � g� 1 dry wt and 1.4–850mg � g� 1 dry wt, respectively. There was a positive
correlation between the quantity of chl a and DNAcontents extracted from the same sediment sample(r2 5 0.78, P<0.001, n 5 95; Fig. 2). The variations inDNA and chl a contents depended on seasons andsediment characteristics. For both DNA and chl a thehighest values were observed in spring and in muddysediments.
Specificity of the primers and efficiency of the real-timeqPCR. Phylogenetic analysis of the 18S rRNA geneand the intergenic transcribed spacer region between18S and 28S rRNA genes showed that N. phylleptaisolates from the Westerschelde formed two phylo-genetic clusters (Fig. 3). The ITS1, which comprised384 bp in cluster 1 and 441–448 bp in cluster 2,exhibited the highest sequence divergence (32%),including insertion/deletion mutations (indels; Fig.4). The PCR-primers for specific detection of eachcluster were designed so that each pair included onehighly specific primer i.e. exhibiting at least 28% se-quence divergence at the target position of the othercluster. Furthermore, they were selected for minimaltendency to form primer dimers. The PCR-primersselected for specific detection of genotypes assignedto clusters 1 and 2 produced 104 and 128 bp frag-ments of the ITS1, respectively. Specificity and effi-ciency of amplification was extensively tested usingendpoint-PCR and real-time qPCR in two detectionformats: SYBR-Green and TaqMan. The specificity ofthe qPCR was assayed using defined mixtures of
FIG. 2. Relationship between DNA and chl a extracted fromsediment (r2 5 0.78, n 5 95, P<0.001). Samples A1, A2, B1, B2 forApril and May 2002–2003 and March 2003 ( � ), samples A1, A2,B1, B2 for October 2002–2003 and all the sampling dates for P1
and P2 (~).
FIG. 3. Maximum likehood (ML) tree for Internal Transcribed Spacer 1 (ITS1) showing the two clusters of Navicula phyllepta.Bootstrap proportion values (450%) for ML (100 replicates) are shown above nodes. Branch lengths are proportional to theamount of substitutions.
V. CREACH ET AL.1146

FIG. 4. Alignment of the ITS1 sequences of the two clusters of Navicula phyllepta. Strains CCY 0222 (DQ193556) and CCY 0212(DQ193543) are in cluster 1 and cluster 2, respectively.
DISTRIBUTION OF NAVICULA IN ESTUARY 1147

reference plasmids, containing the cloned ITS1 frag-ments of N. phyllepta strain CCY 0222 (cluster 1) andstrain CCY 0212 (cluster 2), respectively. The resultsshowed no cross reactivity of the primers with non-target DNA. The two sets of primers were alsoapplied to analyze the genomic DNA of Naviculastrains isolated from the three locations in the Wes-terschelde as well as two reference strains fromthe Ems-Dollard estuary and the Colne River (seeTable 2). An amplification product was only observedfor strains that were identified as N. phyllepta. More-over, in all strains tested only one set of PCR primersgave a positive result, and a fragment of the expectedsize, which allowed unequivocal assignment of allstrains to either one of the clusters.
For reliable quantification, the amplification effi-ciency during PCR must be equal for the recombinantDNA used in the calibration curves and the DNA ex-tracted from the investigated samples, and, for highsensitivity, the amplification efficiency should approach100%. In the exponential phase of the PCR, thenumber of amplicons (Yn) produced after n PCR cy-cles is expressed as
Yn ¼ N�0ð1þ eÞn ð1Þwhere e is the amplification efficiency of one PCR step(0 < e< 1). In calibration curves performed with re-combinant DNA reference plasmids and genomic DNAof N. phyllepta strains CCY 0222 and CCY 0212, e was1.08 � 0.08 (n 5 6, 0.13, 1), and 1.15 � 0.08 (n 5 6,0.122, 1), respectively (in brackets, number of calibra-tion curves, mean squared error, and regression coef-ficient). Because of this high efficiency, the fluorescentsignal produced from 10 target sequences in a singleTaqMan assay passed the threshold value within lessthan 35 cycles. Assays that exhibited lowered amplifi-cation efficiency were excluded from analyses.
DNA content and number of ITS1 sequences percell. We determined the DNA content and the num-ber of ITS1 copies per cell in two isolates of N. phylle-pta, CCY 0212 and CCY 0222. Cells were harvested15 days after inoculation at concentrations of 7.4–8.2 � 105 and 1.5–2.3 � 106 cells �mL� 1, respectively.The mean DNA content per cell was 0.15 � 0.02 pg(n 5 11) for strain CCY 0212 and 0.04 � 0.008 pg(n 5 11) for strain CCY 0222. The DNA samples(n 5 22) were used to estimate the number of ITS1copies per cell in real-time qPCR assays. The numberof copies per cell calculated from the regressionequation (Fig. 5) was 92 copies per cell for CCY0212 and 144 copies per cell for CCY 0222.
Quantification of the two N. phyllepta clusters in theWesterschelde estuary. The number of ITS1 sequencesof each of the two ITS1 genotypes of N. phyllepta wasdetermined by real-time qPCR in the DNA extractedfrom surface sediment samples, which were collectedat three sites (six locations) along the salinity gradientof the Westerschelde estuary (Fig. 1). DNA extractedfrom a high-biomass sediment sample (samples withthe highest chl a content) was analyzed twice and the
average values are shown in Figure 5. Owing to thepatchiness of microphytobenthos in the field, DNAfrom a second sediment sample collected from thesame station at the same date was analyzed. Even ifthe numbers of cells per square meter were differentdue to different biomass, a similar trend for bothsediment samples was observed (data not shown).
The distribution of the two clusters differed signif-icantly (Wilcoxon Matched pairs test, T 5 144, n 5 54,P<0.05) along the estuary (Fig. 6). When both geno-types were present, cluster 1 showed a higher numberof copies per square meter than cluster 2 (2–600 timesmore) except in April and May 2002 in P1 and April2002 and 2003 in P2 (Fig. 6, P1, P2). Assuming that thenumber of ITS sequence copies determined for strainsCCY 0222 (144 copies) and CCY 0212 (92 copies)are representative for all genotypes assigned tothe same cluster, the cell number in surface sedimentsamples was calculated. Cell numbers of cluster 1genotype varied from 8.5 � 106 at station P2 to2.7 � 1010 cells �m� 2 in station A2. No sample was be-low the detection limit. The cell density was signifi-cantly higher in Appelzak than in the BiezelingscheHam and Paulina Polder sediments, and in April 2002compared with the other sampling dates (MANOVA,F(2, 107), (8, 107) 5 11.09 and 4.8, P<0.01). In contrast tocluster 1, the abundance of cluster 2 did not differ be-tween sites (MANOVA, F(2, 107) 5 1.8, P 5 0.155). Thecluster 2 genotype was not detected in February andSeptember 2003 and reached maximum abundance of3.9 � 109 cells �m� 2 in April 2002 at station P1 (Fig.6B). There was no significant difference between highand middle shore sampling sites whatever the cluster(MANOVA, F(1, 107) 5 0.15, P 5 0.15 for cluster1,F(1, 107) 5 2.78, P 5 0.098 for cluster 2).
FIG. 5. Number of copies as a function of DNA amount. Theregression for the compiled data was fitted linearly as Cop-ies 5 6 � 105 ( � 1 � 105) DNA (r2 5 0.9, n 5 11, P<0.01) forCCY 212 and Copies 5 4 � 106 ( � 1 � 106) DNA (r2 5 0.84,n 5 11, P<0.001) CCY 0221, where Copies is the number ofcopies per microliter and DNA is expressed in ng DNA per mi-croliter.
V. CREACH ET AL.1148

Comparison of the number of cells calculated on basis ofreal-time qPCR and the relative abundance of N. phylle-pta from microscope counts. Microscopic counting ofthe cleaned valves of epipelic diatoms showed a high-est percentage of N. phyllepta in the micro-phytobenthos of Appelzak in May 2002: 24% and37% at the sites A1 and A2, respectively. The percent-ages observed at the other locations (BiezelingscheHam and Paulina Polder) varied between 0% and10%, except in September 2002 when 19% of the
valves counted at station P1 were assigned to this spe-cies. As these values represent relative abundanceswithin the total pool of diatoms, they were not di-rectly comparable with cell densities derived fromreal-time qPCR. Using the average chl a per stationand a cellular content of 5.3 pg of chl a per cell (DeJong and Admiraal 1984), we estimated the numberof cells per square meter of N. phyllepta from the mi-croscopic counts (Fig. 7). The values from the real-time qPCR and the microscopic counts were in the
FIG. 6. Temporal and spatial distribution of the two clusters of Navicula phyllepta expressed by number of copies per square meteralong the salinity gradient from oligohaline (A1, A2) to polyhaline (P1, P2) conditions in the Westerschelde estuary.
DISTRIBUTION OF NAVICULA IN ESTUARY 1149

same order of magnitude, 5 � 107–1.7 � 1010 and1.2 � 107–6 � 109 cells �m� 2, respectively. The pop-ulation dynamic of N. phyllepta determined with thetwo methods showed a good agreement (r2 5 0.24,P<0.01, n 5 48), particularly in Appelzak (Fig. 7, A1,A2).
DISCUSSION
Recent investigations have identified epipelic dia-toms as key species in the relationship between biodi-versity and ecosystem function in intertidalenvironments (Forster et al. 2006). However, assessingthe biodiversity of microalgae is not a trivial task. De-termination at the species level is only possible by tax-onomic experts but even then, different genotypesmay reveal the same phenotype and vice versa. Ow-ing to these difficulties molecular genetic approaches
are more informative for following the diversity anddistribution of microalgal populations. In this study,real-time qPCR was used to determine the diversityand distribution of Navicula phyllepta, a key species inintertidal mudflats.
The analysis of ITS sequences demonstrated that N.phyllepta is a semicryptic species that is divided in atleast two clusters that are difficult to identify by mi-croscopy (B. Vanelslander et al. unpublished). Theabundances of the two forms of N. phyllepta were quan-tified in samples obtained from intertidal mudflatsalong a salinity gradient in the Westerschelde estuary.The results showed that the two forms have a differentdistribution. Form 1 (i.e. ITS1 cluster 1) was dominantat the brackish sites and showed a recurrent seasonalpattern with highest densities in late spring. Form 2(i.e. ITS1 cluster 2) was generally less abundant,occurred mainly at higher salinities, and only bloomed
FIG. 7. Comparison betweenthe number of cells per squaremeter of Navicula phyllepta (clus-ter 1þ cluster 2) determined byquantitative real-time PCR (fullline) and microscopic counts(dashed line) for stations A1, A2,B1, B2, P1, and P2.
V. CREACH ET AL.1150

during spring of the first sampling year. A difference ofdistribution patterns of N. phyllepta caused by differentorganic waste and nutrient concentrations has been al-ready described in estuaries (Peletier 1996, Thorntonet al. 2002) but never related to different forms of thespecies. Cryptic or semicryptic species such as N. phylle-pta have been reported for several marine organisms(Knowlton 1993, Darling et al. 2004), which may ormay not be sympatric. The presence of the two formsin the same environment suggests that they occupydifferent niches and thrive under different environ-mental conditions such as temperature and nutrients,sensitivity to pollutants, chemical composition, or lightstress conditions (Mann 1999). The simultaneous pres-ence of semicryptic species in the same environmentcan also be explained by their life cycle strategies andthe control exerted by predators or parasites (Orsiniet al. 2004). In order to investigate the relationships,many clones of each cluster have to be isolated andtested for their properties and behavior under variousconditions (Gallagher 1982, Rynearson and Armbrust2004). Clones of N. phyllepta belonging to the two clus-ters have indeed been shown to differ in their growthresponses to salinity (B. Vanelslander et al. unpub-lished data) in the laboratory. These results confirmthe distribution pattern relative to salinity of the twoforms in the estuary. However a wider survey of pa-rameters is necessary to obtain a complete overview ofthe ecology of the two forms.
Trends in the distribution of N. phyllepta populationsalong the estuary analyzed by real-time qPCR havebeen compared with microscopic counting. In additionto being the only method available to enumerate or-ganisms of cryptic species, this molecular technique ismuch faster compared to traditional taxonomic analy-ses and microscopic counting. It gives an accurateabundance of a certain genotype, which is not biasedby an observer. In this study, the classic and moleculartechniques gave rather similar results for the dynamicand the magnitude of N. phyllepta population. Howev-er, some discrepancies existed. The real-time qPCRshowed a higher number of peaks for the populationparticularly at Biezelingsche Ham (B2) and PaulinaPolder (P1, P2). These discrepancies could be ex-plained not only by the heterogeneity of the samples,but also by using an average of chl a content per cell toconvert the relative to absolute count for the micro-scopic data which does not take into account the var-iability between different species or differentindividuals. A good agreement between data fromreal-time qPCR and other methods has been observedfor picoeukaryotes, Chlorophyta, and Mamiellales(Zhu et al. 2005) and Alexandrium minutum (Galluzziet al. 2004). However, the comparison was not success-ful for Aureococcus anophagefferens (Popels et al. 2003).Although quantitative real-time PCR is attractive dueto its sensitivity, speed, and the possibility of automa-tion, a number of assumptions need to be considered.The various protocols for extracting DNA from micro-organisms in soils or sediments are often not satisfac-
tory. The reasons for this are that somemicroorganisms are difficult to break or that extracel-lular polymeric substances interfere. Co-extraction ofhumic acids is another problem that interferes with thequality of the extracted DNA. In recent years, com-mercial extraction kits for different sample materialshave been significantly improved and are increasinglyused because of their constant quality and hence thepossibility for comparisons between different laborato-ries. The variability in efficiency, reproducibility,phylotype abundance, and composition of the micro-bial community strongly depends on the DNA extrac-tion method applied (Martin-Laurent et al. 2001,Mumy and Findlay 2004). The choice of the DNA ex-traction method is dependent on the type of soil, thecharacteristics of the sediment (Lipthay et al. 2004) orthe type of microorganism under study (Frostegardet al. 1999). In the present study, the extracted DNAwas of high quality and could be directly amplified andquantified. A dilution step to decrease humic acid con-centration was not necessary. Hence, the quantity ofDNA obtained was not the limiting factor and did notaffect the sensitivity of the method. The correlations inthis study between the amount of recovered DNA andthe quantity of extracted sediment, and between DNAand chl a contents in the sample indicated a high re-producibility and DNA extraction efficiency.
There is little known about DNA content per cell(DNA C-value) for diatoms and even less for marinebenthic diatoms. It is also difficult to make any com-parison with the values in the literature for otherorganisms (Vaulot et al. 1994, Veldhuis et al. 1997) asthere is a 5000-fold range in DNA C-values in unicel-lular eukaryotic algae (see Table 1 in Cavalier-Smith1978). The DNA content of N. phyllepta from form 2was found to be in the same range as Navicula pellicu-losa (Holm-Hansen 1969), whereas the DNA contentper algal cell for form 1 was four times lower and ap-peared to be in the bottom end of the known values foralgae (Cavalier-Smith 1978). However, the two formsof N. phyllepta followed the proportional relationshipbetween cell size and DNA C-value observed for algae(Holm-Hansen 1969) and in eukaryotes in general(Gregory 2001).
The division of the two forms of N. phyllepta waseven more obvious in terms of numbers of copies ofITS1 in each cluster. The two forms differed from eachother by 52 copies, but both were close to the regres-sion line established in Zhu et al. (2005), which showeda positive relationship between rRNA gene copy num-ber and cell length. The range of copy numbers is widein unicellular eukaryotic algae from 1 for Nannochloro-psis salina (Zhu et al. 2005) to 1000 in Alexandrium min-utum (Galluzzi et al. 2004). The two forms of N.phyllepta presented copy numbers of 92 and 144, whichare close to that of another Bacillariophyte Nitszchiaclosterium (Zhu et al. 2005). However, environmentalheterogeneity during periods of asexual division, sex-ual recombination, and random mutation can generatediversity in microalgal species. The variability in ITS1
DISTRIBUTION OF NAVICULA IN ESTUARY 1151

between the clusters and even within the cluster can behigh for N. phyllepta and as a result the total number ofcells of this species may be underestimated by a lessefficient detection of target sequence that exhibit a se-quence variation in the target sequences of primers orthe probe. The real-time qPCR assay for N. phylleptahas been developed on the basis of 13 cloned ITS se-quences from 11 strains isolated and cultivated fromthe Westerschelde, one strain from the Ems-Dollardestuary (NL) and one strain from the River Colne(UK). Although this number of strains is limited, theystill probably represent the dominant clones for N.phyllepta, being isolated during a period of 2 years andduring bloom events where the chance to obtain themore abundant genotypes is high. It is difficult to eval-uate the clonal variability in ITS1 because of sexualcrossing events.
In the last few years, the real-time qPCR appears tobe a very powerful method to quantify organisms. It isa sensitive, specific and low time consuming techniquefor processing the analysis. However, the optimizationof the assay can be laborious. For a species complex,one must take into account the number of clusters,their variability and the quantity of targeted gene foreach cluster. This study showed two different clustersfor N. phyllepta. They differed in DNA content and ITSgene copy number and sequences. The structure of thephylogenetic tree supports the conclusion that there isa potential interbreeding between the clones from thesame cluster but not between the two clusters (Mann1999, Behnke et al. 2004). Additionally, the real-timeqPCR showed that their distribution differs along theestuary as well as their abundance. These new sets ofdata on N. phyllepta reinforces the idea (Thornton et al.2002) that this key species for intertidal ecosystemshould be revised, not only in term of genetic varia-tions but also ecological preferences.
We thank Marije Doeleman, Ute Wollenzien, and Dr. VictorChepurnov for their technical assistance and Dr. Graham Un-derwood (University of Essex, UK) for providing a strain ofNavicula phyllepta. We thank Dr. R. M. Forster for providingthe data of chl a. This study was supported by the ResearchCouncil for Earth and Life Science (ALW) in the framework ofthe Flemish-Dutch collaboration on marine research (VLA-NEZO) subsidized by F.W.O.–N.W.O.), contract ALW832.11.003, and by BOF project GOA 01GZ0705 (Ghent Uni-versity). This is Publication 3899 Netherlands Institute ofEcology (NIOO-KNAW).
Admiraal, W., Peletier, H. & Brouwer, T. 1984. The seasonal suc-cession patterns of diatom species on an intertidal Mudflat—an experimental-analysis. Oikos 42:30–40.
Armbrust, E. V., Berges, J. A., Bowler, C., Green, B. R., Martinez,D., Putnam, N. H., Zhou, S. G., Allen, A. E., Apt, K. E.,Bechner, M., Brzezinski, M. A., Chaal, B. K., Chiovitti, A.,Davis, A. K., Demarest, M. S., Detter, J. C., Glavina, T.,Goodstein, D., Hadi, M. Z., Hellsten, U., Hildebrand, M.,Jenkins, B. D., Jurka, J., Kapitonov, V. V., Kroger, N., Lau, W.W. Y., Lane, T. W., Larimer, F. W., Lippmeier, J. C., Lucas, S.,Medina, M., Montsant, A., Obornik, M., Parker, M. S., Palenik,B., Pazour, G. J., Richardson, P. M., Rynearson, T. A., Saito, M.A., Schwartz, D. C., Thamatrakoln, K., Valentin, K., Vardi, A.,
Wilkerson, F. P. & Rokhsar, D. S. 2004. The genome of thediatom Thalassiosira pseudonana: ecology, evolution, and me-tabolism. Science 306:79–86.
Bakker, F. T., Olsen, J. L., Stam, W. T. & Vandenhoek, C. 1992.Nuclear ribosomal DNA internal transcribed spacer regions(ITS1 and ITS2) define discrete biogeographic groups inCladophora-albida (Chlorophyta). J. Phycol. 28:839–45.
Barranguet, C. & Kromkamp, J. 2000. Estimating primaryproduction rates from photosynthetic electron transportin estuarine microphytobenthos. Mar. Ecol. Prog. Ser. 204:39–52.
Becker, S., Boger, P., Oehlmann, R. & Ernst, A. 2000. PCR bias inecological analysis: a case study for quantitative Taq nucleaseassays in analyses of microbial communities. Appl. Environ. Mi-crobiol. 66:4945–53.
Becker, S., Fahrbach, M., Boger, P. & Ernst, A. 2002. Quantitativetracing, by Taq nuclease assays, of a Synechococcus ecotype in ahighly diversified natural population. Appl. Environ. Microbiol.68:4486–94.
Behnke, A., Friedl, T., Chepurnov, V. A. & Mann, D. G. 2004. Re-productive compatibility and rDNA sequence analyses in theSellaphora pupula species complex (Bacillariophyta). J. Phycol.40:193–208.
Bowers, H. A., Tengs, T., Glasgow, H. B., Burkholder, J. M., Rub-lee, P. A. & Oldach, D. W. 2000. Development of real-time PCRassays for rapid detection of Pfiesteria piscicida and relateddinoflagellates. Appl. Environ. Microbiol. 66:4641–8.
Brandini, F. P., da Silva, E. T., Pellizzari, F. M., Fonseca, A. L. O. &Fernandes, L. F. 2001. Production and biomass accumulationof periphytic diatoms growing on glass slides during a 1-yearcycle in a subtropical estuarine environment (Bay of Par-anagua, Southern Brazil). Mar. Biol. 138:163–71.
Cavalier-Smith, T. 1978. Nuclear volume control by nucleoskele-tal DNA, selection for cell volume and cell growth rate, andthe solution of the DNA C-value paradox. J. Cell Sci. 34:247–78.
Colijn, F. & de Jonge, V. N. 1984. Primary production of micro-phytobenthos in the Ems-Dollard estuary. Mar. Ecol. Prog. Ser.14:185–96.
Cox, E. J. 1997. Assessing and designating diatom taxa at or belowthe species level—a consideration of current status and somesuggested guidelines for the future. Nova Hedwigia 65:13–26.
Cox, E. J. 1998. The identity and typification of some naviculoiddiatoms (Bacillariophyta) from freshwater or brackish habitats.Phycologia 37:162–75.
Darling, K. F., Kucera, M., Pudsey, C. J. & Wade, C. M. 2004. Mo-lecular evidence links cryptic diversification in polar plank-tonic protists to Quaternary climate dynamics. Proc. Natl. Acad.Sci. 101:7657–62.
De Jong, L. & Admiraal, W. 1984. Competition between three es-tuarine benthic diatom species in mixed cultures. Mar. Ecol.Prog. Ser. 18:269–75.
Ford, R. B. & Honeywill, C. 2002. Grazing on intertidal micro-phytobenthos by macrofauna: is pheophorbide a useful mark-er? Mar. Ecol. Prog. Ser. 229:33–42.
Forster, R. M., Creach, V., Sabbe, K., Vyverman, W. & Stal, L. J.2006. The biodiversity-ecosystem function relationship in mi-crophytobenthic diatoms of the Westerschelde estuary. Mar.Ecol. Prog. Ser. 229:203–15.
Frostegard, A., Courtois, S., Ramisse, V., Clerc, S., Bernillon, D., LeGall, F., Jeannin, P., Nesme, X. & Simonet, P. 1999. Quantifi-cation of bias related to the extraction of DNA directly fromsoils. Appl. Environ. Microbiol. 65:5409–20.
Gallagher, J. C. 1982. Physiological variation and electrophoresisbanding patterns of genetically different seasonal populationsof Skeletonema costatum (Bacillariophyceae). J. Phycol. 18:148–62.
Galluzzi, L., Penna, A., Bertozzini, E., Vila, M., Garces, E. &Magnani, M. 2004. Development of a real-time PCR assayfor rapid detection and quantification of Alexandrium minutum(a dinoflagellate). Appl. Environ. Microbiol. 70:1199–206.
Gregory, T. R. 2001. Coincidence, coevolution, or causation? DNAcontent, cell size, and C-value enigma. Biol. Rev. 76:65–101.
V. CREACH ET AL.1152

Guillard, R. R. L. & Ryther, J. H. 1962. Studies on marine plank-tonic diatoms. 1. Cyclotella nana (Hustedt) and Detonula con-fervacea (Cleve) Gran. Can. J. Microbiol. 8:229–39.
Haubois, A. G., Sylvestre, F., Guarini, J. M., Richard, P. &Blanchard, G. F. 2005. Spatio-temporal structure of theepipelic diatom assemblage from an intertidal mudflat inMarennes-Oleron Bay, France. Estauries Coast. Shelf Sci. 64:385–94.
Heid, C., Stevens, J., Livak, K. & Williams, P. 1996. Real TimeQuantitative PCR. Genome Methods: Genome Research. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
Holm-Hansen, O. 1969. Algae: amounts of DNA and organic car-bon in single cells. Science 163:87–8.
Knowlton, N. 1993. Sibling Species in the Sea. Annu. Rev. Ecol. Syst.24:189–216.
Kooistra, W. H. C. F., Stam, W. T., Olsen, J. L. & Vandenhoek, C.1992. Biogeography of Cladophoropsis-membranacea (Chloro-phyta) based on comparisons of nuclear rDNA ITS sequenc-es. J. Phycol. 28:660–8.
Krammer, K. & Lange-Bertalot, H. 1986. Bacillariophyceae. Part1—Naviculaceae. In Ettl, H., Gerloff, F., Heynig, H. & Moll-enhauer, D. [Eds.] Su�wasserflora von Mitteleuropa. G. Fischer,Stuttgart, 876 pp.
Labrenz, M., Brettar, I., Christen, R., Flavier, S., Botel, J. & Hofle,M. G. 2004. Development and application of a real-time PCRapproach for quantification of uncultured bacteria in the cen-tral Baltic Sea. Appl. Environ. Microbiol. 70:4971–9.
Lange-Bertalot, H. 2001. Navicula sensu stricto—10 genera separat-ed from Navicula sensu lato—Frustulia. In Lange-Bertalot, H.[Ed.] Diatoms of Europe—Diatoms of European Inland Waters andComparable Habitats. Vol. 2. A. R. G. Gantner Verlag K.G.,Ruggell , 26 pp.
Leblanc, C., Falciatore, A., Watanabe, M. & Bowler, C. 1999. Semi-quantitative RT-PCR analysis of photoregulated gene expres-sion in marine diatoms. Plant Mol. Biol. 40:1031–44.
Lipthay de, J. D., Enzinger, C., Johnsen, K., Aamand, J. & S�ren-sen, S. J. 2004. Impact of DNA extraction method on bacterialcommunity composition measured by denaturing gradient gelelectrophoresis. Soil Biol. Biochem. 36:1607–14.
Lucas, C. H., Widdows, J., Brinsley, M. D., Salkeld, P. N. & Her-man, P. M. J. 2000. Benthic-pelagic exchange of microalgae ata tidal flat. 1. Pigment analysis. Mar. Ecol. Prog. Ser. 196:59–73.
Mann, D. G. 1999. The species concept in diatoms. Phycologia38:437–95.
Martin-Laurent, F., Philippot, L., Hallet, S., Chaussod, R., Ger-mon, J. C., Soulas, G. & Catroux, G. 2001. DNA extractionfrom soils: old bias for new microbial diversity analysis meth-ods. Appl. Environ. Microbiol. 67:2354–9.
Medlin, L. K. & Kaczmarska, I. 2004. Evolution of the diatoms:V. Morphological and cytological support for the major cladesand a taxonomic revision. Phycologia 43:245–70.
Medlin, L. K., Williams, D. M. & Sims, P. A. 1993. The evolution ofthe diatoms (Bacillariophyta). I. Origin of the group and as-sessment of the monophyly of its major divisions. Eur. J. Phycol.28:261–75.
Middelburg, J. J., Barranguet, C., Boschker, H. T. S., Herman,P. M. J., Moens, T. & Heip, C. H. R. 2000. The fate of intertidalmicrophytobenthos carbon: an in situ C-13-labeling study.Limnol. Oceanogr. 45:1224–34.
Montani, S., Magni, P. & Abe, N. 2003. Seasonal and interannualpatterns of intertidal microphytobenthos in combination withlaboratory and areal production estimates. Mar. Ecol. Prog. Ser.249:79–91.
Mumy, K. L. & Findlay, R. H. 2004. Convenient determination ofDNA extraction efficiency using an external DNA recoverystandard and quantitative-competitive PCR. J. Microbiol. Meth-ods 57:259–68.
Muylaert, K., Van Nieuwerburgh, L., Sabbe, K. & Vyverman, W.2002. Microphytobenthos communities in the freshwater tidalto brackish reaches of the Schelde estuary (Belgium). Belg. J.Bot. 135:15–26.
Newby, D. T., Hadfield, T. L. & Roberto, F. F. 2003. Real-time PCRdetection of Brucella abortus: a comparative study of SYBR
green 1,50-exonuclease, and hybridization probe assays. Appl.Environ. Microbiol. 69:4753–9.
Orsini, L., Procaccini, G., Sarno, D. & Montresor, M. 2004.Multiple rDNA ITS-types within the diatom Pseudo-nitzschiadelicatissima (Bacillariophyceae) and their relative abundancesacross a spring bloom in the Gulf of Naples. Mar. Ecol. Prog.Ser. 271:87–98.
Peletier, H. 1996. Long-term changes in intertidal estuarine diatomassemblages related to reduced input of organic waste. Mar.Ecol. Prog. Ser. 137:265–71.
Popels, L. C., Cary, S. C., Hutchins, D. A., Forbes, R., Pustizzi, F.,Gobler, C. J. & Coyne, K. J. 2003. The use of quantitative po-lymerase chain reaction for the detection and enumeration ofthe harmful alga Aureococcus anophagefferens in environmentalsamples along the United States East Coast. Limnol. Oceanogr.:Methods 1:92–102.
Rijstenbil, J. W. 2003. Effects of UVB radiation and salt stresson growth, pigments and antioxidative defence of the marinediatom Cylindrotheca closterium. Mar. Ecol. Prog. Ser. 254:37–47.
Round, F. E. 2004. Vie et milieu of freshwater diatoms. Commenton current issues in diatom ecology and floristics. Vie et Milieu/Life and Environment 54:157–61.
Rynearson, T. A. & Armbrust, E. V. 2004. Genetic differentiationamong populations of the planktonic marine diatom Ditylumbrightwellii (Bacillariophyceae). J. Phycol. 40:34–43.
Sabbe, K., Verleyen, E., Hodgson, D. A., Vanhoutte, K. & Vyver-man, W. 2003. Benthic diatom flora of freshwater and salinelakes in the Larsemann hills and Rauer islands, east Antartica.Antarctic Sci. 15:227–48.
Skovhus, T. L., Ramsing, N. B., Holmstrom, C., Kjelleberg, S. &Dahllof, I. 2004. Real-time quantitative PCR for assessment ofabundance of Pseudoalteromonas species in marine samples.Appl. Environ. Microbiol. 70:2373–82.
Sorhannus, U. 2004. Diatom phylogenetics inferred based on di-rect optimization of nuclear-encoded SSU rRNA sequences.Cladistics 20:487–97.
Sundback, K., Miles, A. & Goransson, E. 2000. Nitrogen fluxes,denitrification and the role of microphytobenthos in micro-tidal shallow-water sediments: an annual study. Mar. Ecol. Prog.Ser. 200:59–76.
Suzuki, M. T., Taylor, L. T. & Delong, E. F. 2000. Quantitativeanalysis of small-subunit rRNA genes in mixed microbial pop-ulations via 50-nuclease assays. Appl. Environ. Microbiol.66:4605–14.
Thornton, D. C. O., Dong, L. F., Underwood, D. J. C. & Nedwell,D. B. 2002. Factors affecting microphytobenthic biomass, spe-cies composition and production in the Colne estuary (UK).Aquat. Microb. Ecol. 27:285–300.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. &Higgins, D. G. 1997. The ClustalX windows interface: flexiblestrategies for multiple sequence alignment aided by qualityanalysis tools. Nucleic Acids Res. 25:4876–82.
Tolhurst, T. J., Jesus, B., Brotas, V. & Paterson, D. M. 2003.Diatom migration and sediment armouring—an examplefrom the Tagus estuary, Portugal. Hydrobiologia 503:183–93.
Underwood, G. J. C. & Kromkamp, J. 1999. Primary productionby phytoplankton and microphytobenthos in estuaries. Adv.Ecol. Res. 29:93–153.
Vaulot, D., Birrien, J-L., Marie, D., Casotti, R., Veldhuis, M. J. W.,Kraay, G. W. & Chretiennot-Dinet, M.-J. 1994. Morphology,ploidy, pigment composition, and genome size of culturedstrains of Phaeocystis (Prymnesiophyceae). J. Phycol. 30:1022–35.
Veldhuis, M. J. W., Cucci, T. L. & Sieracki, M. E. 1997. CellularDNA content of Marine phytoplankton using two new fluoro-chromes: taxonomy and ecological implications. J. Phycol.33:517–44.
Wawrik, B., Paul, J. H. & Tabita, F. R. 2002. Real-time PCR quan-tification of rbcL (ribulose-1,5-bisphosphate carboxylase/oxy-genase) mRNA in diatoms and pelagophytes. Appl. Environ.Microbiol. 68:3771–9.
DISTRIBUTION OF NAVICULA IN ESTUARY 1153

Wei, S. F., Hwang, S. P. L. & Chang, J. 2004. Influence of ultravioletradiation on the expression of proliferating cell nuclear anti-gen and DNA polymerase a in Skeletonema costatum (Bacillar-iophyceae). J. Phycol. 30:655–63.
Witkowski, A., Lange-Bertalot, H. & Metzeltin, D. 2000. Diatomflora of marine coasts I. Iconogr. Diatomol. 7:1–925.
Zechman, F. W., Zimmer, E. A. & Theriot, E. C. 1994. Use of rib-osomal DNA internal transcribed spacers for phylogeneticstudies in diatoms. J. Phycol. 30:507–12.
Zhu, F., Massana, R., Not, F., Marie, D. & Vaulot, D. 2005. mappingof picoeucaryotes in marine ecosystems with quantitative PCRof 18S rRNA gene. FEMS Microbiol. Ecol. 50:231–43.
V. CREACH ET AL.1154
Related Documents











