MARINE MAMMAL SCIENCE, 3(3):242-262 (July 1987) 0 1987 by the Society for Marine Mammalogy USING ORGANOCHLORINE POLLUTANTS TO DISCRIMINATE MARINE MAMMAL POPULATIONS: A REVIEW AND CRITIQUE OF THE METHODS1 ALEX AGUILAR* A-008, Scripps Institution of Oceanography, University of California San Diego, La Jolla, California 92038 ABSTRACT Organochlorine pollutants are potentially useful for identifying discrete pop- ulations of marine mammals that overlap in geographic distribution. However, many factors unrelated to geographical distribution may affect the chemical burden of individual animals or of entire population components even within a homogeneously distributed population. These factors include, among others, nutritional state, sex, age, trophic level, distance of habitat from mainland and pollution source, excretion, metabolism, and tissue composition. Sample storage and analytical methodology may also be an important source of variation. These, and any other factors, must be identified and their effect ascertained before attempting any comparison between populations. This paper critically examines the nature and magnitude of the effects of these factors on organochlorine tissue loads in marine mammals. Pollutant concentrations can be strongly biased if carefully designed sampling regimes are not followed, but they are affected only moderately by sample treatment after collection. Conversely, ratios between concentrations of compounds, such as the DDE/tDDT or the tDDT/PCB ratios, seem less dependent on sampling regime but more affected by storage, analytical procedures and ecological variations such as distance from pollutant source or trophic level. Taking these effects into account, advice is provided about sampling and strategies for selection of variables that will improve the reliability of the comparisons between populations. Key words: marine mammals, cetaceans, pinnipeds, pollutants, organochlorines, techniques, population discreteness, distribution, stock identity. 1 Made while at the Southwest Fisheries Center, National Marine Fisheries Service, NOAA, Ia Jolla, California 92038, USA. 2 Present address: Department of Zoology (Vertebrates), Faculty of Biology, University of Bar- celona, Barcelona 0807 1, Spain. 242 Help Volumes Main Menu

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MARINE MAMMAL SCIENCE, 3(3):242-262 (July 1987) 0 1987 by the Society for Marine Mammalogy
USING ORGANOCHLORINE POLLUTANTS TO DISCRIMINATE MARINE MAMMAL
POPULATIONS: A REVIEW AND CRITIQUE OF THE METHODS1
ALEX AGUILAR*
A-008, Scripps Institution of Oceanography, University of California San Diego, La Jolla, California 92038
ABSTRACT
Organochlorine pollutants are potentially useful for identifying discrete pop- ulations of marine mammals that overlap in geographic distribution. However, many factors unrelated to geographical distribution may affect the chemical burden of individual animals or of entire population components even within a homogeneously distributed population. These factors include, among others, nutritional state, sex, age, trophic level, distance of habitat from mainland and pollution source, excretion, metabolism, and tissue composition. Sample storage and analytical methodology may also be an important source of variation. These, and any other factors, must be identified and their effect ascertained before attempting any comparison between populations. This paper critically examines the nature and magnitude of the effects of these factors on organochlorine tissue loads in marine mammals. Pollutant concentrations can be strongly biased if carefully designed sampling regimes are not followed, but they are affected only moderately by sample treatment after collection. Conversely, ratios between concentrations of compounds, such as the DDE/tDDT or the tDDT/PCB ratios, seem less dependent on sampling regime but more affected by storage, analytical procedures and ecological variations such as distance from pollutant source or trophic level. Taking these effects into account, advice is provided about sampling and strategies for selection of variables that will improve the reliability of the comparisons between populations.
Key words: marine mammals, cetaceans, pinnipeds, pollutants, organochlorines, techniques, population discreteness, distribution, stock identity.
1 Made while at the Southwest Fisheries Center, National Marine Fisheries Service, NOAA, Ia Jolla, California 92038, USA.
2 Present address: Department of Zoology (Vertebrates), Faculty of Biology, University of Bar- celona, Barcelona 0807 1, Spain.
242
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 243
Knowledge of population identity and discreteness is basic for the study and management of animal populations. In marine mammals, most of this knowledge comes from marking, morphological variation and biochemical studies (e.g., Brown 1977, Wada and Numachi 1979, Gaskin 1982, Irvine et ul. 1982, Perrin 1984). However, these methods of population discrimination are often ineffective either because of difficulties in sampling or because the results are inconclusive.
Another method, analysis of pollutant loads in selected tissues, has promise as a discrimination method. This approach has been used to study migratory patterns or population discreteness of fish and birds (Krygier and Pearcy 1977, Dubrawski and Falandysz 1980, Moreau and Barbeau 1982, Dutil et al. 1985), and several authors have suggested that this technique might be useful for discriminating between populations of marine mammals with poorly known dispersion patterns (Frank et al. 19 73, Taruski et al. 1975, Ronald et al. 1984).
Organochlorine pollutants have been reported from the most remote areas and ecosystems of the world. However, the observed pollutant load of an organism living in a given ecosystem results from the action of many factors: the relative quantities of pollutants released in the ecosystem, the transport rate of these compounds from the pollution sources to the ecosystem, and the detoxification and degradation processes experienced by the chemicals during transport and during their residence in the ecosystem prior to their accumulation in the tissues of the organism. Because the relative importance of the different factors varies with compound and environment, and because marine organisms take up pollutants from the environment in which they live, populations in- habiting different geographical areas are likely to have qualitatively and quan- titatively different pollutant burdens.
In the case of marine mammals, numerous reports exist of dissimilarities in tissue pollutant levels between two or more groups of samples, and these dissimilarities are often assumed to indicate geographically discrete populations. (Holden and Marsden 1967, Holden 1972, Frank et al. 1973, Anas 1974, Alzieu and Duguy 1979, Duinker and Hillebrand 1979). Therefore, the uti- lization of organochlorines as population discriminators for marine mammals looks promising.
Nevertheless, many factors unrelated to geographical distribution may affect pollutant burden of a given specimen or even of a given group of specimens, making it extremely easy to find artifactual differences between groups of samples caused more by how and when the samples were collected, preserved or analysed, than by actual variation in the load of chemicals characteristic of the populations.
The objectives of this paper are: 1) to summarize what is known presently about behaviour and dynamics of the most important organochlorine compounds in tissues of marine mammals, 2) to evaluate the adequacy of various pollutant measures and ratios derived from them, 3) to identify sources of variation other than those due to geographic distribution of sampling groups which may alter pollutant burdens, and 4) to suggest, based on this information, sampling strategies and selection criteria for variables to characterize population pollutant loads.
Help Volumes Main Menu

244 MARINE MAMMAL SCIENCE, VOL. 3, N0. 3, 1987
GENERAL LIMITATIONS AND SIGNIFICANCE OF POLLUTANTS AS TRACERS
It is important to keep in mind that tissue pollutant burdens are poorly or not related at all to genetics. This imposes some restrictions in its use for population discrimination.
First, two population units isolated genetically may occupy different geographic areas but have pollutant profiles indistinguishable from each other. Pollutant loads are only useful for discriminating populations when differences between them can be proved to exist, so negative results are inconclusive.
Second, some components of an interbreeding population may be segregated geographically during part of the year or may be exposed to environments with different pollutant loads. This will be especially relevant in species such as large whales, which have extremely large migratory ranges and for which population components are unlikely to be distributed homogeneously within this range or may be bound by different feeding or migratory regimes.
Differences in feeding regime may be especially important because food contributes over 90% of the organochlorine intake by mammals (Campbell et al. 1965). Variation in food composition or in location of feeding grounds can thus produce variations in tissue residue levels. At least two surveys attribute observed differences in pollutant levels to feeding segregation between compo- nents of a single population of marine mammals. Aguilar (1983) found in North Atlantic sperm whales that, contrary to what seems to be the rule in marine mammals (see below), females were carrying higher organochlorine con- centrations in their tissues than males. He suggested that the difference was due to the shallower feeding habits and more southern distribution of females in relation to males. Tanabe et al. (1984a) attributed higher concentrations of DDTs and PCBs found in younger minke whales of the Southern Hemisphere to these whales’ preference for feeding in lower latitudes and also to suspected differences in diet composition between juveniles and adults. It is likely that similar variation in pollutant burden will also be found in many other species which segregate in their distribution range or during their migration, such as fin, blue, humpback, gray or bottlenose whales (Lockyer and Brown 1981). In such situations, segregation patterns must be ascertained and sampling regime designed carefully before accurate comparisons can be made.
.
Third, although pollutant burdens in marine mammals are not susceptible to change on a short time scale, variation in the qualitative and quantitative composition of organochlorine tissue levels can occur over years. For example, the concentration of total DDT in blubber of most North Atlantic marine mammals has decreased in the last two decades, while PCB concentrations and the ratio between DDE and total DDT (tDDT = DDT + TDE + DDE) have increased (Jones et al. 1976, Holden 1978, Donkin et al. 1981, Helle et al. 1983, Aguilar 1984, Addison et al. 1984). In the harbour porpoises of the Bay of Fundy, DDT concentrations declined from 1969 to 1973 but increased during the late 1970s (Gaskin 1982). In the Canadian western Arctic, PCBs and DDTs in ringed seal blubber declined between 1972 and 1981, but PCBs declined more than DDTs (Addison et al. 1986). Therefore, in these cases, no
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 245
strictly valid comparisons can be made between samples collected in different years unless the trend is well known and a correction applied.
VARIABLE TYPES: CONCENTRATIONS AND RATIOS BETWEEN CONCENTRATIONS
Differences in pollutant loads can be investigated using two types of variables: 1) tissue concentrations of compounds, and 2) ratios between tissue concentrations of one compound and another.
Although in theory any ratio between pollutants could be used to discriminate between populations, in practice most research in marine environments has focused on only two of them: the tDDT/PCB ratio (concentration of DDT plus all its derivatives, divided by the concentration of PCBs irrespective of isomeric composition), and the DDE/tDDT ratio (DDE concentration, divided by the concentration of DDT plus all its derivatives). These two ratios have proved useful in identifying distribution patterns and time trends in several marine organisms, and have been tentatively applied to marine mammals (Nelson 1972, Koeman 1973, O’Shea et al. 1980, Aguilar 1984).
Differences in pollutant concentrations between individuals are often large, even within populations with no segregation in feeding habits or in geographical distribution. Comparatively, the intra-population variability of ratios is usually lower than that of the concentrations, because the bioaccumulation processes regulating the rate at which a given organism incorporates pollutants seem to be roughly similar for all lipophilic xenobiotics. This is reflected in the high correlations usually found between levels of different organochlorines in the tissues of marine mammals (Holden 1978, Reijnders 1980, Donkin et al. 1981, Helle et al. 1983, Ronald et al. 1984). Nevertheless, even these ratios tend to be sufficiently variable within single populations to cause problems in comparisons of populations based on small samples.
While some of the sources of variability affecting organochlorine pollutant concentrations and ratios still remain unidentified, some others are known and their effects can be reasonably predicted. It is worthwhile, therefore, to examine the most important of these sources of variation and determine the nature of the biases they may introduce in a marine mammal population study at the population level.
SOURCES OF VARIATION OF POLLUTANT LOADS
a) Fatness State and Selection of Appropriate Units for Expressing Pollutant Concentrations
Since organochlorines are highly apolar compounds and dissolve in neutral lipids, two measurements have been singly or simultaneously used as a basis for expressing tissue concentrations in marine mammals: 1) fresh weight of the sample (fresh weight basis), and 2) quantity of lipids present in the sample
Help Volumes Main Menu

246 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
(lipid basis). Other methods of expression, such as concentration of residues in relation to triglyceride content, have been used in surveys of fish and other organisms and have been proposed for marine mammals (Aguilar 1985). How- ever, they are not used routinely and therefore will not be considered here.
Selecting the appropriate system of expression is important because marine mammals, especially those species which are seasonal feeders, rely strongly on their fat stores. This leads to substantial differences in lipid depot volume between reproductive categories or between seasons in any given reproductive category (Lockyer 1981, Lavigne et al. 1982). Because organochlorines dissolve in lipids, changes in the volume of lipids available, i.e., of solvent available, will affect pollutant tissue concentrations.
Nevertheless, no reliable information exists about dynamics of organochlorine residues as a marine mammal grows thin or fat. When lipids are mobilized, two extreme processes are possible: either 1) pollutants leave the tissues together with the lipids to which they were bound and are then metabolized and excreted from the body, or 2) they remain in the tissue irrespective of variation in lipid content. Pollutant concentrations may or may not be affected by fatness changes depending on which expression system is selected and what the dynamics of organochlorines actually are.
For example, consider a marine mammal with a total blubber fresh weight of 30 Kg, with 70% of that weight made up of neutral lipids, and a total burden of 1 g of a given organochlorine compound. This is a reasonable situation in a small cetacean or seal (Tanabe et al. 1981, Hidaka et al. 1983). When blubber fat is mobilized, organochlorine compounds may 1) remain totally unaffected, or 2) leave the tissue and be excreted in a quantity proportional to the amount of lipids mobilized.
Moreover, it is known that blubber lipids are partially substituted by water after their mobilization (Aguilar 1985), although the magnitude of this sub- stitution is unknown. For modelling purposes, let’s suppose that when a certain amount of lipid leaves the blubber: 1) the reduction in the fresh weight of the tissue is proportional to this loss, and 2) 20% of the weight loss of the blubber is compensated by incorporation of water. This value is arbitrarily chosen to estimate the sensitivity of the model to possible changes in water content.
Assume that the marine mammal uses part of its lipid reserves and reduces the lipid content in the blubber from the initial 70% down to 25%. Such variation occurs normally in wild marine mammal populations (unpublished data).
Changes in concentration, expressed both on a fresh weight basis and on a lipid basis, under the different lipid mobilization regimes are depicted in Figure 1. On a fresh weight basis, organochlorine concentration increases if the pollutant remains unaltered in the tissue, but decreases if it leaves blubber along with the mobilized lipids. If water partially replaces the lost fat, the bias is mitigated. Conversely, when expressing the pollutant concentration on a lipid basis, re- placing fat with water has no effect on the calculation of concentrations. If the organochlorine pollutant is mobilized proportional to the lipids, estimated con- centrations are unbiased. But if it remains unaltered in the tissue, an important
Help Volumes Main Menu
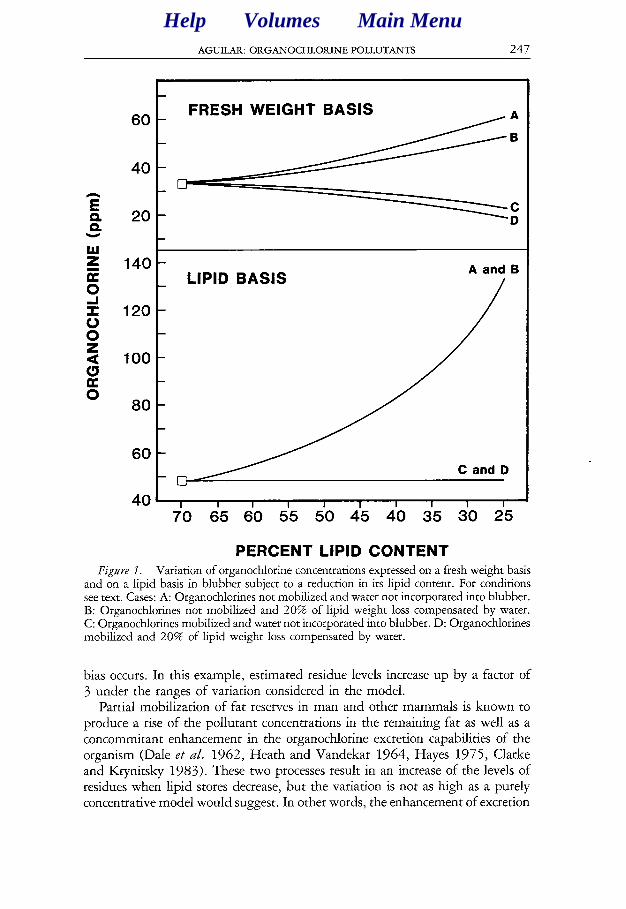
AGUILAR: ORGANOCHLORINE POLLUTANTS 247
60 - FRESH WEIGHT BASIS
40 -
20 -
140 140 - LIPID BASIS
A and B _
120 120 -
100 100 -
80 80 -
60 60 - C and D
40 40 I I I I I I I I I I I I I I I I I I I I 70 70 65 60 55 50 45 40 35 30 25 65 60 55 50 45 40 35 30 25
PERCENT LIPID CONTENT PERCENT LIPID CONTENT Figwe I. Variation of organochlorine concentrations expressed on a fresh weight basis
and on a lipid basis in blubber subject to a reduction in its lipid content. For conditions see text. Cases: A: Organochlorines not mobilized and water not incorporated into blubber. B: Organochlorines not mobilized and 20% of lipid weight loss compensated by water. C: Organochlorines mobilized and water not incorporated into blubber. D: Organochlorines mobilized and 20% of lipid weight loss compensated by water.
bias occurs. In this example, estimated residue levels increase up by a factor of 3 under the ranges of variation considered in the model.
Partial mobilization of fat reserves in man and other mammals is known to produce a rise of the pollutant concentrations in the remaining fat as well as a concommitant enhancement in the organochlorine excretion capabilities of the organism (Dale et ul. 1962, Heath and Vandekar 1964, Hayes 1975, Clarke and Krynitsky 1983). These two processes result in an increase of the levels of residues when lipid stores decrease, but the variation is not as high as a purely concentrative model would suggest. In other words, the enhancement of excretion
Help Volumes Main Menu

248 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
is inadequate to prevent the rising of organochlorine compounds, but it tends to mitigate it.
It is not clear at present what dynamics of changes in organochlorine levels are to be expected during lipid mobilization in the blubber of marine mammals. There are indications that at least some increase in pollutant concentration occurs when lipid reserves decrease in seals (Addison and Smith 1974, Drescher et al. 1977, Donkin et al. 1981, Zande and Ruiter 1983), but it is not known whether or not this increase is proportional to fat loss. However, it is likely that a dual process similar to that found in other mammals (increase both in con- centration and in excretion rate) occurs.
If this is so, expression of residue concentrations on a lipid basis would appear higher in lean animals, the magnitude of the bias depending on how inaccurate is the assumption that the excretion rate of pollutants is proportional to fat mobilization. The fluctuation of residue levels when expressed on a fresh weight basis is impossible to predict at the present time. Residue levels may be over- estimated if the enhancement of excretion is low, and underestimated if it is high.
Furthermore, the parameter values used in the example above are rather conservative. In reality, the problems involved in quantifying pollutant tissue concentrations may be even greater. Marine mammals are often more heavily contaminated than in the case discussed above (see Gaskin 1982). In addition, lipid mobilization can be more intense than the values and ranges considered here. Both greater contamination and greater tendency to mobilize lipids will tend to increase the bias in measurements derived from field samples. Most of these problems could probably be overcome if a more suitable basis for expression of pollutant concentrations were used (Aguilar 1985).
It is difficult to state what the actual effect of fatness changes on ratios will be. If the different organochlorine compounds behave similarly when fat is mobilized, ratios between concentrations of different pollutants will remain unaltered. But if the variation in excretion and degradation rates does not affect all compounds similarly, ratios may change.
d) Sex and Age
Early stages of life-It has been shown above that important changes in residue concentrations occur with changes in fatness state. The most striking variation in lipid content is associated with food availability or reproductive state. However, some age-related trends in lipid content of tissues and total body fat have been observed in fin whales and spotted dolphins, especially during the first stages of life (Aguilar, Borrell and Vetter, unpublished data). These changes are likely to affect pollutant burdens and have not been, but should be, investigated.
No less important are differences in qualitative lipid composition of tissues associated with growth. Not only is lipid content of blubber from foetuses (and probably neonates) about half that of adults, but it is also relatively richer in polar lipids. This explains why blubber from newborn Weddell seals and striped
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 249
dolphins has relatively more lower chlorinated biphenyls than does blubber from adults (Tanabe et al. 1982, Hidaka et al. 1983).
The effects of such developmental changes on pollutant loads have not been investigated. It seems likely, however, that while ratios may be susceptible to change, pollutant concentrations will only be moderately affected, provided that they are expressed on a lipid basis.
Juveniles and adults--Both sex and age have a strong influence on the concentrations of pollutants carried by a given specimen, but the effects are contradictory. Irrespective of sex, pollutant loads tend to increase with age, because uptake usually exceeds excretion. However, they often decrease in sex- ually mature females, because females transfer appreciable quantities of lipophilic xenobiotics to their offspring during pregnancy and lactation, thus reducing their own pollutant burdens (Reijnders 1980, Gaskin 1982, Tanabe et al. 1982, Alzieu et al. 1983).
A female grey seal may reduce her load of organochlorine pollutants by 15- 30% during the time it takes to raise and wean a single pup (Addison and Brodie 1977). This is in addition to what has already been relocated to the foetus during pregnancy. Similar calculations in striped dolphins gave figures ranging between 72% and 98% of the mother’s pollutant load (Fukushima and Kawai 1981, Tanabe et al. 1982).
Relocation of pollutants during pregnancy is not as important as during lactation. In grey seals, the mother transfers to her foetus only about 1% of her pollutant burden (Donkin et al. 1981), in Weddell seals, 2% (Hidaka et al. 1983), in harbour porpoises, 15% (Duinker and Hillebrand 1979), and in striped dolphins between 4% and 9% (Fukushima and Kawai 1981, Tanabe et al. 1982). Logically, on a long time scale, the significance of this translocation will be proportional to the number of times a female reproduces and, therefore, may be highly variable even between females of the same age.
These two processes combine to produce sex- and age-related variation patterns of pollutant levels which differ between species or even between one population and another. Pollutant concentration increases with age in the tissues of most male pinnipeds (Addison et al. 1973, Frank et al. 1973, Addison and Smith 1974, Helle et al. 1983, Zande and Ruiter 1983, Calambokidis et al. 1984), although in some cases individual variability is large and no trends are found (Le Boeuf and Bonnell 197 1, McClurg 1984). In cetaceans, a similar age-related increase has been found in harbour porpoises (Gaskin et al. 1982, 1983), belugas (Martineau et al. 1985), striped dolphins younger than 15 yr (Tanabe et al. 1981), and minke whales older than 20 yr (Tanabe et al. 1984a), but concentrations level off or even decrease with age in some populations of old striped dolphins (Tanabe et al. 1981) and young minke whales (Tanabe et al. 1984a).
Age-related trends of pollutant concentrations in females are even more vari- able than in males. In pinnipeds, reports indicate either an increase with age (Donkin et al. 1981, Helle et al. 1983), an increase in sexually immature animals but a stabilization of levels in mature ones (Frank et al. 1973, Ronald et al. 1984), an absence of any trend (Addison and Smith 1974, Born et al.
Help Volumes Main Menu

250 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
1981), or even a decrease with age (Holden 1978, Born et al. 1981). In cetaceans, the information available is rather limited, but indicates either a decrease with age (Gaskin 1982, Gaskin et al. 1983) an increase with age (Martineau et al. 19SS), or an absence of clear patterns (Fukushima and Kawai 1981).
Therefore, age-related trends, although often occurring, are not consistent among different populations and species. It must be pointed out, moreover, that these dissimilarities in age-related trends between males and females often pro- duce differences between sexes in the mean pollutant concentrations both in pinnipeds (Addison and Smith 1974, Donkin et al. 198 1, Calambokidis et al. 1984) and cetaceans (Forrester et al. 1980, Aguilar and Jover 1982, Aguilar 1983).
Because different compounds probably accumulate differently with age, it is also likely that ratios between them will change with age. Addison et al. (1973) found a positive relationship between the DDE/tDDT ratio and age in harp seals, and a similar trend is expected for this ratio in most marine mammals. However, there is no information available to determine either the nature or the magnitude of changes of other ratios in marine mammals. As discussed below, metabolism seems to be enhanced at higher pollutant concentrations. Therefore, provided that relative excretion rates remain constant, higher pro- portions of metabolites difficult to excrete in relation to their parent compounds should be found in older animals.
c) Differential Metabolism and Excretion
The ability of marine mammals to metabolize and excrete different organo- chlorine compounds or isomers is variable and depends, not only on the molecular structure of the chemical, but also on the individual’s physiological characteristics. This is liable to be an important source of intrapopulation variation. The subject is not well studied, but there are at least four cases in which differential detox- ification rates can be expected:
1) When the specimen sampled is diseased or heavily parasitized. Microsomal enzymatic systems, responsible for catalyzing the metabolism of organochlorines and many other xenobiotics, can be either enhanced or depressed by disease, thus affecting the metabolization rates of pollutants. While this is not important for samples from wild, healthy populations, it could be a critical factor in samples from stranded or captive animals.
2) When interaction between compounds occur; that is, when the concen- tration of one pollutant depends on the concentration of a second. Synergism in induction of microsomal enzymes to higher activity levels by different organ- ochlorines is known to occur in most organisms (Phillips 1980), and this is likely to affect the relative concentrations of parent pollutants and their metab- olites. In general, this effect is thought to be weak, but Borrell and Aguilar (1987) found that DDE/tDDT ratios increased proportionally with the total load of DDT in the blubber of fin and sei whales. Moreover, Zande and Ruiter (1983) found that, because lower chlorinated PCBs are eliminated faster from
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 251
the blubber of harbour seals than higher chlorinated PCBs, the degree of chlorination of PCBs in blubber increased proportional to their tissue concen- tration. Similar relationships probably occur with other organochlorines but have not been studied.
3) When animals grow. Environmental pollutants are metabolized mainly by mixed function oxidases (MFO), an hepatic enzyme system. Recently, Addison and Brodie (1984) found in grey seals that activity of ethoxyresorufin O-de- ethylase, a MFO, increased with age in pups and was greater in adults than in pups. Similarly, Goksoyr et al. (1985, 1986) found in minke whales that activity of cytochrome P-450-system, another MFO, was also much higher in juveniles and adults than in foetuses. This indicates that the ability of an animal to get rid of organochlorines improves with age. However, there is no information available about how this age-related variation may affect organochlorine pollutant concentrations or ratios.
4) When females reproduce. As stated above, transfer of pollutants from the mother to her offspring is a major source of variation for the chemical load of females. The amount transferred seems to differ with different compounds. During pregnancy and lactation of both pinnipeds and cetaceans, the most easily transferred organochlorines are HCH and HCB, followed by DDTs and then PCBs (data from Addison and Brodie 1977, Holden 1978, Fukushima and Kawai 1981, Tanabe et al. 1982). In the case of PCBs the transfer seems to be inversely proportional to the number of chlorine atoms substituted in the biphenyls (Tanabe et al. 1982). It is not clear if the different DDT forms are excreted at similar rates. Addison and Brodie (1977) found that the DDE percentage was virtually constant for milk and blubber samples of grey seals, but Kurtz and Kim (1976) found that blubber of nursing fur seals had a higher DDE/tDDT ratio (90%) than blubber from cows and newborns (60%) of the same population, suggesting that proportionally more DDE is transferred during lactation. As a rule, higher lipophilia of a compound means that transfer rate to offspring is lower. It is important to keep this in mind if populations composed of both sexes are compared, because the ratios between pollutants can be totally transformed in a female with an active reproductive history, in general tending to be richer in highly chlorinated, highly lipophilic compounds.
Therefore, except in extreme situations, such as in heavily diseased animals, the effect of differential metabolism or excretion on the concentration of pollutants is slight. However, ratios, especially those involving metabolites, can be strongly modified.
d) Food Chain Level
The efficiency with which different organochlorine xenobiotics are transferred along food chains depends on the compounds’ physicochemical properties and biological stabilities. Higher lipophilicity usually produces higher transfer rates and bioaccumulation potential, so ratios between concentrations of pollutants vary progressively along food chains.
It is commonly accepted that the DDE/tDDT ratio increases as it progresses
Help Volumes Main Menu

252 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
from lower to higher trophic levels (Jensen et al. 1969, Tanabe et al. 1984b). However, the behaviour of other ratios is not yet well understood, and results of surveys are often contradictory. Jensen et al. (1969), Bowes and Jonkel (1975), Duinker and Hillebrand (1983), Subramanian et al. (1983), and Tanabe et al. (19846) reported that PCB isomers with high chlorine content are more easily transferred than those with a lower content to the following food chain level, but Calambokidis et al. (1984) found the reverse. Similarly, Bowes and Jonkel (1975) reported that the tDDT/PCB ratio decreases along food chains, while Tanabe et al. (198 1, 19846) reported that it increases. It is likely that the finding of one trend or the other in the tDDT/PCB ratio depends essentially on the isomeric composition of the PCBs originally released to the ecosystem, because transfer rates of different PCB isomers probably differ.
Such changes in ratios may be important if the population studied is composed of groups with different feeding habits, a situation which may be rather common in marine mammals. It is known, for example, that male sperm whales feed on a variety of prey, whereas females rely almost entirely on squid. Moreover, food items of males are usually larger in size than those of females (Mizue 195 1, Best 1979). A survey of the food habits of spotted dolphins showed that pregnant, non-lactating females consumed mostly squid, while lactating, non- pregnant females ate mostly fish (Bernard and Hohn 1985). Moreover, several pinnipeds show age-related changes in food habits, shifting from one trophic level to another as they grow older (Spalding 1964, Bonner 1979, King 1983, Doidge and Croxall 1985). Because diving ability is a function of size in at least some species of marine mammals (Tarasevich 1968, Best 1979), it is likely that differences in food habits exist between individuals of different ages or sexes, especially in species with high sexual dimorphism in body size. Doidge and Croxall (1985) found that adult male fur seals from the Antarctic eat more fish and squid than other classes of fur seals, and attributed the difference to the male’s larger body size and hence deeper diving capacity which facilitates feeding on more benthic resources.
.
It is difficult to predict how the pollutant load of marine mammals will change when a shift in trophic level occurs. In general, if the shift is towards a higher food chain level, concentrations will tend to increase and ratios will reflect a higher proportion of the more stable compounds, but the magnitude of this variation is unknown.
e) Distance from Pollutant Source and Mainland
Volatilization, transport and environmental detoxification rates are not iden- tical for all organochlorine compounds or isomers of a given compound. Thus, the availability of each individual chemical to the organisms living in a given water mass is influenced by their distance to the main sources of pollution, usually the mainland. The differences in availability are usually unnoticeable when tissue concentrations are examined, but can appear clearly if ratios are investigated.
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 253
Risebrough et ul. (1968) were the first to report an increase of tDDT/PCB ratios in marine environments correlated with distance of ecosystem location from the mainland. Their results were confirmed later by Risebrough (1969) and Nelson ( 1972). Departing from these findings, Koeman ( 1973) explained that tDDT/PCB ratios of Dutch harbour porpoises were higher than those of common seals because of the more pelagic distribution of the first species.
Other ratios have since been shown to correlate with distance from pollutant source. The DDE/tDDT ratio is probably one of the most sensitive and better studied. DDE is a compound highly accumulative and is difficult to excrete or degrade. Its abundance relative to DDT (the parent compound from which DDE derives) increases with time after release (MacGregor 1974, Barber and Warlen 1979). Because dispersion of DDT compounds is a slow process, typical DDE/tDDT ratios in areas close to the coast and, especially, close to point sources of pollution, are low, while in pelagic or remote areas they are high.
Thus, mean DDE/tDDT ratios in coastal cetaceans from Uruguay, where DDT has been used in recent times, were found to be much lower than those found in Pacific pelagic waters (O’Shea et al. 1980). Aguilar (1984) found that the DDE/tDDT ratio of both coastal and pelagic marine mammals of the North Atlantic had been significantly increasing during the period 1964-198 1, but that DDE/tDDT ratios for pinnipeds (coastal) were higher than values for odontocetes (more pelagic) in any given year.
Therefore, attention should be paid to whether populations studied tend to stratify in relation to the coastline or if spatial segregation of individuals is suspected.
j) Variation in Tissue Sampling
Sampling procedures for comparative studies must be consistent. However, when collecting tissue material from large animals, some variation in body locality at which the tissue is collected is likely to occur. This can strongly affect the organochlorine burden of the samples.
Blubber, the tissue universally used in organochlorine pollutant surveys of marine mammals, is highly heterogeneous both in composition and structure. Neither lipid content, lipid composition, nor lipid polarity are constant from one locality of the body to another. This produces an heterogeneous distribution of pollutants even within a single tissue or body compartment (Aguilar 1985).
This problem is especially relevant if concentrations are to be expressed on a fresh-weight basis, because organochlorines dissolve in lipids and contribution of lipids to tissue weight is highly variable. However, if concentrations are calculated on a lipid basis, pollutant levels will only be affected by changes in the polarity of the lipids constituting the blubber and not by their quantity. On absolute terms, the effect of lipid polarity changes on pollutant concentrations is thought to be weak. On the contrary, ratios, because of their susceptibility to lipid polarity variations, are more likely to be altered by changes in qualitative lipidic composition (Aguilar 1985).
Help Volumes Main Menu

254 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
Since lipid composition of tissues tends to be more variable quantitatively than qualitatively, the effect of changes in body location on ratios or on con- centrations expressed on a lipid basis will be slight, unless major inconsistencies in sampling occur.
g) Tissue Preservation and Analytical Procedures
Tissue Preservation-Despite the early report by Jefferies and Walker (1966) that DDT decomposes into its metabolites in frozen samples and the later confirmation of these findings by a number of researchers (Ecobichon and Saschenbrecker 1967, French and Jefferies 1971, Wiemeyer et al. 1984), many scientists do not seem concerned about the effect storage may have had on their analytical results.
Thus, storage effects are seldom investigated. The only study on storage- related changes in blubber of marine mammals seems to be that of Olsson et al. (1974), who looked at differences in tDDT and PCB concentrations between seal blubber analysed before and after storage for 5 mo at room temperature without any preservative. They concluded that no apparent differences existed between the two sample groups. However, the figures in their paper do show clearly that fresh samples had higher organochlorine levels than those stored unpreserved. Thus, in the two blubber layers examined by them (inner and outer) only the DDT concentration in the inner layer seemed to remain stable. In the remaining cases, concentrations were 20% and 25% higher for PCBs and 12.5% higher for DDTs in the fresh tissues than in the unpreserved ones. No information is available in this paper about changes in the relative abundance of derivatives, but it is likely that substantial decomposition in the unpreserved samples did in fact occur.
.
These results are only indicative. Nevertheless, the information available for other tissues and animals strongly supports the idea that changes in pollutant concentrations and ratios occur in frozen or formalin stored samples (Ecobichon and Saschenbrecker 1967, French and Jefferies 197 1, Wiemeyer et al. 1984). Logically, the magnitude of such changes will be directly proportional to storage time, and inversely proportional to the conditions of preservation (for example, temperature). Therefore, stocks of samples preserved under different conditions are not directly comparable unless the possibility of differential pollutant deri- vation or loss can be reasonably excluded.
Analytical procedures-Intercalibration exercises show that no two laboratories obtain exactly the same results for any given sample, although differences seem to be decreasing progressively as techniques improve with time. This is not surprising given the wide variety of analytical procedures used, but it precludes direct comparisons between samples analyzed in different laboratories. Conclu- sions drawn from different cooperative laboratories may be valid after adequate intercalibration, but ideally all analyses should be carried out by a single lab- oratory and without varying any of the analytical procedures while the survey lasts.
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 255
Table 1. Estimated effect of various factors on pollutant loads of marine mammals. Intensity of effect: H = high, M = moderate, N = none, V = variable, U = unknown.
Factor
Nutritive state Age (early stages) Age (juveniles and adults) Reproductionb Differential metabolism/excretion Food chain level Distance to pollution source Sample location Tissue preservation Analytical procedures
a Especially in males. b Only in females.
DDE/ tDDT
tDDT/ PCB tDDT PCB
H H U U M M M V H H U U H H U M M M H H H H H H M M H H M M M H V V V U M M V V
SELECTION OF VARIABLES AND SURVEY DESIGN
a) Choosing Variable Types: Concentrations or Ratios
The first step in distinguishing between two sample groups on the basis of their pollutant burden is to decide whether the concentration levels of each xenobiotic or the ratios obtained from two or more of these concentrations are the most appropriate parameters to compare. In essence, concentrations and ratios are equally valid and can be used singly or in combination depending on the particular characteristics and goals of each survey. However, apart from their inherent variability, both types of variables are subject to important sources of bias which may render them useless under certain conditions.
The estimated effects that the sources of variation considered above may have on tissue concentrations and ratios are summarized in Table 1. Pollutant con- centrations are plagued with sampling problems (variation in fattening state, age, sex and reproductive state of specimens), and very restrictive criteria should be applied in order to obtain a sample free of bias. On the other hand, particular characteristics of individuals seem to only moderately influence ratios, so these are expected to be less affected by heterogeneities in sample composition. How- ever, it is not clear what will happen to the ratios when fat stores change in volume, when females relocate pollutants into their offspring or when xenobiotics accumulate with age.
If preservation conditions can not be maintained constant for all the samples, attention should be paid to possible variation in ratios between metabolites and their parent compounds (i.e., DDE/tDDT) due to chemical derivation. This is especially important if preservation steps are not taken soon after death.
Because metabolism and excretion abilities can be altered in diseased animals, and because lipid reserves might have been depleted, ratios between chemicals can not be reliably used when the sample includes captive or stranded animals.
Help Volumes Main Menu

256 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
In such surveys, moreover, concentrations of organochlorines can only be con- sidered reliable when lipid reserves of the specimens are within the normal ranges of variation of the population.
In some particular cases, for example when sex- or age-related variations in food habits or distribution are known for the species studied and the samples to be compared have dissimilar age or sex compositions, ratios should also be regarded with distrust.
In general, the behaviour of concentrations is much better known than that of ratios, and the effect of uncontrolled factors may be more accurately predicted in the first than in the second type of variable. At first glance, this suggests that concentrations are more promising when seeking a robust discrimination between populations. Nevertheless, the within-population variance of concentrations tends to be much higher than that of ratios, and it may easily be the case that concentrations in two sampling groups are not distinguishable, while ratios are.
b) Choosing Variable
Concentrations-DDT and PCB levels and their isomers and metabolites are the most abundant organochlorine pollutants found in the tissues of marine mammals. They all have long half-lives, and their concentrations are’usually high enough to allow for accurate quantification. The disadvantage of selecting these chemicals is that they have been so widely used that their geographical variation is often slight.
DDT is probably the best known organochlorine pollutant, and its ecological and physiological behaviour is well documented. The most abundant forms are p,p’-isomers derived from p,p’-DDT; o,p’-isomers are much less abundant, and it is not clear whether their apparently low frequency of detection in marine mammal tissues is due to their non-existence in most populations or to a lack of effort of scientists in determining them. If the first is true, o,p’-DDT and its derivatives may be highly promising for population differentiation.
PCB quantification can be affected adversely by variation in isomeric com- position. However, techniques for identifying and quantifying individual PCB isomers are improving rapidly, and are now making possible much better char- acterizations of PCB tissue loads. Variation in abundance of individual PCB isomers seems one of the most promising tools.
Logically, in areas in which uncommon chemicals have been released, good characterization of pollutant loads can be achieved by searching for these com- pounds. Organochlorines of restricted use, such as Mirex or certain chlordanes may be thus extremely helpful in discriminating populations.
Ratios-The only ratios that have been studied in detail are tDDT/PCB and DDE/tDDT. Because the timing and geographical differences of DDT and PCB introduction vary substantially, the tDDT/PCB ratio is potentially one of the most valuable for population discrimination. It is very sensitive to distance from mainland and trophic level (see above), but its coefficient of variation is usually low, indicating a low intrapopulation variability. Obviously, this ratio
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 257
will be higher in masses of water close to agricultural areas and lower in heavily industrialized basins.
The DDE/tDDT ratio is extremely dependent of the length of time since release of the parent compound (DDT) and is highly indicative when the chronology of pesticide input differs between the areas studied. When using this ratio, problems may arise if large variation in tDDT or in trophic level within the population occur, because of the sensitivity of the DDE/tDDT ratio to the interaction between tDDT and DDE, and because of its food chain related variation (see above). Many other ratios, especially those involving different PCB or DDT isomers, degraded forms of cyclodienes (i.e., Dieldrin) and novel anthropogenic compounds such as Mirex, may be extremely useful, and their dynamics and potentials for population discrimination should be investigated.
c) Survey Design
Ideally, any comparison of pollutant concentrations between different marine mammal populations should use only individuals of the same age, sex, repro- ductive category and fattening state. Also, collection, storage and analytical conditions should be exactly the same for all samples. However, so strict an operative regime is usually not feasible, and some concessions to practicality need to be made. Sampling within a given size range of only one sex (preferably males) on a limited time period would help to overcome many of the problems. However, the particular sampling availability of a survey will suggest which specimens are optimal for collection, which sources of variation can be reasonably ignored, and which type of parameters are likely to be most reliable.
For example, if the species studied is a seasonal feeder and samples are collected at different times of the year, pollutant concentrations are likely to be misleading because of changes in fattening state, and ratios are to be preferred. If the age structure is dissimilar between the populations to be compared, only the portions with reasonably homogeneous age composition should be used.
Therefore, there is a need to know before hand the effects that the different sources of variation are likely to have on the populations concerned. Some of these effects can be guessed from previous studies in other populations or even other species. For example, age-related increases of pollutant concentrations in pinniped males, and higher levels in adult males than in adult females of both pinnipeds and cetaceans are an almost universal result in studies with adequate sample sizes (see section b).
Moreover, the knowledge of the effect of these sources of variation will be useful as a clue not only for deciding which variables may be misleading and therefore avoided, but also to identify which of them can be expected a priori to have a higher power of discrimination.
For example, the Nova Scotia fin whale population feeds mostly on planktonic crustaceans, while Newfoundland fin whales eat capelin (a higher trophic level prey) almost exclusively (Mitchell 1975). In this situation, ratios with known dependence on food chain level can be potentially useful in discriminating between the two populations. Inshore and offshore populations of the same
Help Volumes Main Menu

258 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
species, such as those of Bryde’s whales in South Africa (Best 1977) or bottlenose dolphins in the North Pacific (Duffield et ul. 1983) might be distinguished by ratios known to change with distance from the mainland or from point sources of pollution.
Of course, the reverse approach is also feasible, and pollutant loads from specimens with a well-known population identity can also be investigated to determine ecological divergences. For example, if components of a given pop- ulation known to inhabit a single water mass are thought to feed on different food resources or trophic levels, ratios such as DDE/tDDT can be useful to ascertain whether that feeding segregation is a constant fact or only a transitory event.
ACKNOWLEDGMENTS
I want to express my gratitude to Jay Barlow, Aleta H. Hohn, M. Hall, W. F. Perrin, and E. F. Vetter for their support during the preparation of this paper and for their comments on the original manuscript. Financial support was received from a Fulbright/ Caixa fellowship while doing this work.
LITERATURE CITED
ADDISON, R. F., AND P. F. BRODIE. 1977. Organochlorine residues in maternal blubber, milk, and pup blubber from grey seals (Halichoerus grypus) from Sable Island, Nova Scotia. Journal of the Fisheries Research Board of Canada 34(7):937-941.
ADDISON, R. F., AND P. F. BRODIE. 1984. Characterization of ethoxyresorufin O-de- ethylase in grey seal Halichoerus grypus. Comparative Biochemistry and Physiology 79C(2):261-263.
ADDISON, R. F., AND T. G. SMITH. 1974. Organochlorine residue levels in Arctic ringed seals: variations with age and sex. Oikos 25:335-337.
ADDISON, R. F., P. F. BRODIE, M. E. ZINCK AND D. E. SERGEANT. 1984. DDT has declined more than PCBs in Eastern Canadian seals during the 1970s. Environmental Science and Technology 18(12):935-937.
ADDISON, R. F., S. R. KERR, J. DALE AND D. E. SERGEANT. 1973. Variation of organochlorine residue levels with age in Gulf of St. Lawrence harp seals (Pagophilus groenlandicus). Journal of the Fisheries Research Board of Canada 30:595-600.
ADDISON, R. F., M. E. ZINCK AND T. G. SMITH. 1986. PCBs have declined more than DDT-group residues in Arctic ringed seals (Phoca hispida) between 1972 and 198 1. Environmental Science and Technology 20:253-256.
AGUILAR, A. 1983. Organochlorine pollution in sperm whales, Physeter macrocephalus, from the temperate waters of the Eastern North Atlantic. Marine Pollution Bulletin 14(9):349-352.
AGUILAR, A. 1984. Relationship of DDE/tDDT in marine mammals to the chronology of DDT input into the ecosystem. Canadian Journal of Fisheries and Aquatic Sciences 41(6):840-844.
AGUILAR, A. 1985. Compartmentation and reliability of sampling procedures in or- ganochlorine pollution surveys of cetaceans. Residue Reviews 95:91-l 14.
AGUILAR, A., AND L. JOVER. 1982. DDT and PCB residues in the fin whale, Balaenoptera physalus, of the North Atlantic. Reports of the International Whaling Commission 32:299-301.
ALZIEU, C., AND R. DUGUY. 1979. Teneurs en composes organochlores chez les cetaces et pinnipedes frequentant les cotes francaises. Oceanologica Acta 2( 1): 107-120.
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 259
ALZIEU, C., R. DUGUY AND P. BABIN. 1983. Pathologie des delphinidae: contamination foetale et neo-natale par les PCB. Lesions cutanes ulceratives. Revue et Travaux Institut Peches Maritimes 46(2): 157-166.
ANAS, R. F. 1974. DDT plus PCB’s in blubber of harbor seals. Pesticides Monitoring Journal 8(l): 12-14.
BARBER, R. T., AND S. M. WARLEN. 1979. Organochlorine insecticide residues in deep sea fish from 2 5 00 m in the Atlantic Ocean. Environmental Science and Technology 13:1146-1148.
BERNARD, H. J., AND A. A. HOHN. 1985. Food habits of pregnant and lactating spotted dolphins in the eastern tropical Pacific Ocean: a progress report. Paper presented to the sixth Biennial Conference on the Biology of Marine Mammals, Vancouver, Canada.
BEST, P. B. 1977. Two allopatric forms of Bryde’s whale off South Africa. Reports of the International Whaling Commission (Special Issue 1): 10-38.
BEST, P. B. 1979. Social organization in sperm whales, Physeter macrocephalus. Pages 227-289 in H. E. Winn and B. L. Olla, eds. Behavior of marine animals, vol. 3, cetaceans. Plenum Press, New York, NY.
BONNER, W. N. 1979. Antarctic (Kerguelen) fur seal. In Mammals in the sea. FAO Fisheries Series 5(2):49-5 1.
BORN, E. W., I. KrAUL AND T. KRISTENSEN. 1981. Mercury, DDT and PCB in the Atlantic walrus (Odobenus rosmarus rosmarus) from the Thule District, North Green- land. Arctic 34(3):255-260.
BORRELL, A.., AND A. AGUILAR. 1987. Variations in DDE percentage correlated to total DDT burden in the blubber of fin and sei whales. Marine Pollution Bulletin 18(20): 70-74.
BOWES, G. W., AND C. J. JONKEL. 1975. Presence and distribution of polychlorinated biphenyls (PCB) in Arctic and subarctic marine food chains. Journal of the Fisheries Research Board of Canada 32( 11):2 111-2123.
BROWN, S. G. 1977. Whale marking: a short review. Pages 569-581 in M. Angel, ed. A voyage of discovery. Pergamon Press, New York, NY.
CALAMBOKIDIS, J., J. PEARD, G. H. STEIGER, J. C. CUBBAGE AND R. L. DELONG. 1984. Chemical contaminants in marine mammals from Washington State. U.S. Depart- ment of Commerce, NOAA Technical Memorandum NOS OMS 6, 167 pp.
CAMPBELL, J. E., L. A. RICHARDSON AND M. L. SCHAFER. 1965. Insecticide residues in the human diet. Archives of Environmental Health 10:831-836.
CLARK, D. R., AND A. J. KrYNITSKY. 1983. DDE in brown and white fat of hibernating bats. Environmental Pollution 3 l(A):287-299.
DALE, W. E., T. B. GAINES AND W. J. HAYES. 1962. Storage and excretion of DDT in starved rats. Toxicology and Applied Pharmacology 4:98-106.
DOIDGE, D. W., AND J. P. CROXALL. 1985. Diet and energy budget of the Antarctic fur seal, Arctocephalus gazella, at South Georgia. Pages 543-550 in W. R. Siegfried, P. R. Condy and R. M. Laws, eds. Antarctic nutrient cycles and food webs. Springer- Verlag, New York, NY.
DONKIN, P., S. V. MANN AND E. I. HAMILTON. 198 1. Polychlorinated biphenyl, DDT and dieldrin residues in grey seal (Halicheorus grypus) males, females and mother- foetus pairs sampled at the Farne Islands, England, during the breeding season. Science of the Total Environment 19:12 1-142.
DRESCHER, H. E., U. HARMS AND E. HUSChENBETH. 1977. Organochlorines and heavy metals in the harbour seal Phoca vitulina from the German North Sea Coast. Marine Biology 41:99-106.
DUBRAWSKI, R., AND J. FALANDYSZ. 1980. Chlorinated hydrocarbons in fish-eating birds from the Gdansk Bay, Baltic Sea. Marine Pollution Bulletin ll( 1):15-18.
DUFFIELD, D. A., S. A. RIdgWAY AND L. H. CORNELL. 1983. Hematology distinguishes coastal and offshore forms of dolphins (Tursiops). Canadian Journal of Zoology 61: 930-933.
Help Volumes Main Menu

260 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3 , 1987
DUINKER, J. C., AnD M. T. J. HILLEBRAND. 1979. Mobilization of organochlorines from female lipid tissue and transplacental transfer to fetus in a harbour porpoise (Phocoena phocoena) in a contaminated area. Bulletin of Environmental Contamination and Toxicology 231728-732.
DUInKER, J. C., AND M. T. J. HILLEBRAND. 1983. Composition of PCB mixtures in biotic and abiotic marine compartments (Dutch Wadden Sea). Bulletin of Environ- mental Contamination and Toxicology 3 1:2 5-3 2.
DUTIL, J. D., B. LEGARE AND C. DESJARDINS. 1985. Discrimination d’un stock de poisson, l’anguille (Anguilla rostrata), basee sur la presence d’un produit chimique de synthese, le mirex. Canadian Journal of Fisheries and Aquatic Sciences 42:455- 458.
ECOBICHON, , D. J., AND P. W. SASCHENBRECKER. 1967. Dechlorination of DDT in frozen blood. Science 156:663-665.
FORRESTER, D. J., D. K. ODELL, N. P. THOMPSON AND J. R. WHITE. 1980. Morpho- metrics, parasites, and chlorinated hydrocarbon residues of pygmy killer whales from Florida. Journal of Mammalogy 61(2):356-360.
FRANK, R., K. RONALD AND H. E. BRAUN. 1973. Organochlorine residues in harp seals (Pagophilus groenlandicus) caught in eastern Canadian waters. Journal of the Fisheries Research Board of Canada 30(8):1053-1063.
FRENCH, M. C., AND D. J. JEFFERIES. 197 1. The preservation of biological tissue for organochlorine insecticide analysis. Bulletin of Environmental Contamination and Toxicology 6(5):460-463.
FUKUSHIMA, M., AND S. KAWAI. 1981. Variation of organochlorine residue concentration and burden in striped dolphin (Stenella coeruleoalba) with growth. Pages 97-l 14 in T. Fujiyama, ed. Studies on the levels of organochlorine compounds and heavy metals in the marine organisms. University of the Ryukyus, Ryukyus.
GASKIN, D. E. 1982. The ecology of whales and dolphins. Heinemann, London, 459 pp. GASKIN, D. E., R. FRANK AND M. HOLDRINET. 1983. Polychlorinated biphenyls in
harbor porpoises Phocoena phocoena (L.) from the Bay of Fundy, Canada and adjacent waters, with some information on clordane and hexachlorobenzene levels. Archives of Environmental Contamination and Toxicology 12:2 1 l-2 19.
GASKIN, D. E., M. HOLDRINET AND R. FRANK. 1982. DDT residues in blubber of harbour porpoises, Phocoena phocoena (L.), from eastern Canadian waters during the five-year period 1969-1973. In Mammals in the sea, FAO Fisheries Series 5(4): 135-143.
.
GOKSoYR, A., J. E. SOLBAKKEN, J. TARLEBo AND J. KLUNGSoYR. 1986. Initial charac- terization of the hepatic microsomal cytochrome P-450-system of the piked whale (minke) Balaenoptera acutorostrata. Marine Environmental Research 19: 185-203.
GOKSoYR, A., J. TARLEBo, J. E. SOLBAKKEN AND J. KluNGSoYR. 1985. Characteristics of the hepatic microsomal cytochrome P-450-system of the minke whale (Balaen- optera acutorostrata). Marine Environmental Research 17: 113-l 16.
HAYES, W. J. 1975. Toxicology of pesticides. The Williams and Wilkins Co., Baltimore, MD, 580 pp.
HEATH, D. F., AND M. VANDEKAR. 1964. Toxicity and metabolism of dieldrin in rats. British Journal of Industrial Medicine 2 1:269-279.
HELLE, E., H. HYVARINEN, H. PYYSALO AND K. WICKSTROM. 1983. Levels of organo- chlorine compounds in an inland seal population in eastern Finland. Marine Pollution Bulletin 14(7):256-260.
HIDAKA, H., S. TANABE AND R. TATSUKAWA. 1983. DDT compounds and PCB isomers and congeners in Weddell seals and their fate in the Antarctic Marine ecosystem. Agricultural and Biological Chemistry 47(9):2009-2017.
HOLDEN, A. V. 1972. Monitoring organochlorine contamination of the marine envi- ronment by the analysis of residues in seals. Pages 2-8 in Marine pollution and sea life. Fishing News Ltd., London.
HOLDEN, A. V. 1978. Organochlorine residues in blubber of grey seals (Halichoerus
Help Volumes Main Menu

AGUILAR: ORGANOCHLORINE POLLUTANTS 261
grypus) from the Fame Islands. International Council for the Exploration of the Sea, Document E:41, 6 pp.
HOLDEN, A. V., AND K. MARSDEN. 1967. Organochlorine pesticides in seals and por- poises. Nature (London) 2 16: 1274-1276.
IRVINE, A. B., R. S. WELLS AnD M. D. SCOTT. 1982. An evaluation of techniques for tagging small odontocete cetaceans. Fishery Bulletin 80( 1): 13 5-143.
JEFFERIES, D. J., AND C. H. WALKER. 1966. Uptake of P,P’-DDT and its post-mortem breakdown in the avian liver. Nature (London) 212:533-534.
JENSEN, S., A. G. JOHNELS, M. OLSSOn A~TD G. OTTERLIND. 1969. DDT and PCB in marine animals from Swedish waters. Nature (London) 224:247-250.
JONES D., K. RONALD, D. M. LAVIGNE, R. FRANK, M. HOLDRINET APITD J. F. UTHE. 1976. Organochlorine and mercury residues in the harp seal (Pagophilus groenlandicus). Science of the Total Environment 5:181-195.
KING, J. E. 1983. Seals of the world. 2nd ed., Cornell University Press, New York, NY. 240 pp.
KOEMAN, J. H. 1973. PCB in mammals and birds in the Netherlands. Pages 35-43 in PCB Conference II. National Swedish Environment Protection Board Publications 1973:4E.
KRYGIER, E. E., AND W. G. PEARCY. 1977. The source of cobalt-60 and migrations of albacore off the west coast of North America. Fishery Bulletin 75(4):867-870.
KURTZ, D. A., AND K. C. KIM. 1976. Chlorinated hydrocarbon and PCB residues in tissues and lice of northern fur seals, 1972. Pesticides Monitoring Journal lO(3): 79-83.
LAVIGNE, D. M., W. W. BARCHARD, S. INNES AND N. A. ORISTLAND. 1982. Pinniped bioenergetics. In Mammals in the sea. FAO Fisheries Series 5 (4): 19 l-2 3 5.
LE BOEUF, B. J., AND M. L. BONNELL. 1971. DDT in California sea lions. Nature (London) 234: 108-l 10.
LOCKYER, C. H. 1981. Growth and energy budgets of large baleen whales from the Southern hemisphere. In Mammals in the sea. FAO Fisheries Series 5(3):379-487.
LOCKYER, C. H., AND S. G. BROWN. 1981. The migration of whales. Pages 105-137 in E. Aidley, ed. Animal migration. Society for Experimental Biology, Seminar Series 13.
MACGREGOR, J. S. 1974. Changes in the amount and proportions of DDT and its metabolites, DDE and DDD, in the marine environment off southern California, 1949-72. Fishery Bulletin 72(2):275-293.
MARTINEAU, D., P. BELAND, C. DESJARDINS AND A. VEZINA. 1985. Pathology, toxicology and effects of contaminants on the population of the St. Lawrence beluga (Delphi- napterus leucas), Quebec, Canada. International Council for the Exploration of the Sea, document CM/N:13, 32 pp.
MCCLURG, T. P. 1984. Trace metal and chlorinated hydrocarbons in Ross seals from Antarctica. Marine Pollution Bulletin 15( 10):384-398.
MITCHELL, E. D. 1975. Trophic relationships and competition for food in Northwest Atlantic whales. Proceedings of the Canadian Society of Zoology (1974): 123-l 33.
MIZUE, K. 195 1. Food of whales in the adjacent waters of Japan. Scientific Reports of the Whales Research Institute 5:81-90.
MOREAU, G., AND C. BARBEAU. 1982. Les metaux lourds comme indicateurs d’origine geographique de l’anguille d’Amerique (Anguilla rostrata). Canadian Journal of Fisheries and Aquatic Sciences 39: 1004-1011.
NELSON, N., ED. 1972. Polychlorinated biphenyls-Environmental impact. A review by the Panel on Hazardous Trace Substances. Environmental Research 5:249-362.
OLSSON, M., A. G. JOHNELS AND R. VAZ. 1974. DDT and PCB levels in seals from Swedish waters. The occurrence of aborted seal pups. In Proceedings from the Symposium on the Seal in the Baltic. National Swedish Environmental Protection Board Publication 591:43-54.
O’SHEA, T. J., R. L. BROWNELL, D. R. CLARK, W. A. WALKER, M. L. GAY AND T. G.
Help Volumes Main Menu

262 MARINE MAMMAL SCIENCE, VOL. 3, NO. 3, 1987
LAMONT. 1980. Organochlorine pollutants in small cetaceans from the Pacific and South Atlantic Oceans, November 1968-J une 1976. Pesticides Monitoring Journal 14(2):35-46.
PERRIN, W. F. 1984. Patterns of geographical variation in small cetaceans. Acta Zoologica Fennica 172:137-140.
PHILLIPS, D. J. H. 1980. Quantitative aquatic biological indicators: their use to monitor trace metal and organochlorine pollution. Applied Science, London. 488 pp.
REIJNDERS, P. J. H. 1980. Organochlorine and heavy metal residues in harbour seals from the Wadden Sea and their possible effects on reproduction. Netherlands Journal of Sea Research 14(1):30-65.
RISEBROUGH, R. W. 1969. Chlorinated hydrocarbons in marine ecosystems. Pages 5- 23 in N. W. Miller and G. G. Berg, eds. Chemical fallout. Charles C Thomas Publisher, Springfield, Illinois.
RISEBROUGH, R. W., P. RIECHE, D. B. PEAKALL, S. G. HERMAN AND M. N. KIRVEN. 1968. Polychlorinated biphenyls in the global ecosystem. Nature (London) 220: 1098-1102.
RONALD, K., R. J. FRANK, J. L. DOuGAN, R. FRANK AND H. E. BRAUN. 1984. Pollutants in harp seals (Phoca groenlandica). Organochlorines. Science of the Total Environment 38:133-152.
SPALDING, D. J. 1964. Comparative feeding habits of the fur seal, sea lion and harbour seal on the British Columbia coast. Bulletin of the Fisheries Research Board of Canada 146: l-52.
SUBRAMANIAN, B. R., S. TANABE, H. HIDAkA AND R. TATSUKAWA. 1983. DDTs and PCB isomers and congeners in Antarctic fish. Archives of Environmental Contam- ination and Toxicology 12:62 l-628.
TANABE, S., T. MORI AND R. TATSUKAWA. 1984a. Bioaccumulation of DDTs and PCBs in the southern minke whale (Balaenoptera acutorostrata). Memoirs of the National Institute of Polar Research, Special Issue 32:140-150.
TANABE, S., H. TANAKA AND R. TATSUKAWA. 1981. Ecology and bioaccumulation of Stenella coeruleoalba. Considerations on the several factors related to the variation ’ of tDDT (p,p’-DDE + p,p’-DDD + p,p’-DDT) residue levels with age of striped dolphins (Stenella coeruleoalba). Pages 123-132 in T. Fujiyama, ed. Studies on the levels of organochlorine compounds and heavy metals in the marine organisms. University of the Ryukyus, Ryukyus.
TANABE, S., H. TANAKA AND R. TATSUKAWA. 19846. Polychlorobiphenyls, tDDT, and hexachlorocyclohexane isomers in the Western North Pacific ecosystem. Archives of Environmental Contamination and Toxicology 13 : 7 3 l-7 3 8.
TANABE, S., R. TATSUKAWA, K. MARUYAMA AND N. MIYAZAKI. 1982. Transplacental transfer of PCBs and chlorinated hydrocarbon pesticides from the pregnant striped dolphin (Stenella coeruleoalba) to her fetus. Agricultural and Biological Chemistry 46(5):1249-1254.
TARASEVICH, M. N. 1968. Dependence of distribution of the sperm whale males upon the character of feeding. Zoologischeskii Zhurnal 47: 1683-1688.
TARUSKI, A. G., C. E. OLNEY AND H. E. WINN. 1975. Chlorinated hydrocarbons in cetaceans. Journal of the Fisheries Research Board of Canada 32:2205-2209.
WADA, S., AND K. NUmACHI. 1979. External and biochemical characters as an approach to stock identification for the Antarctic minke whale. Reports of the International Whaling Commission 29:42 l-432.
WIEMEYER, S. N., J. F. MOORE AND B. M. MULHERM. 1984. Formalin preservation of avian blood for metal and DDE analysis. Bulletin of Environmental Contamination and Toxicology 33:525-532.
ZANDE, T. VAN DER, AND E. DE RUITER. 1983. The quantification of technical mixtures of PCBs by microwave plasma detection and the analysis of PCBs in the blubber lipid from harbour seals (Phoca vitulina). The Science of the Total Environment 27:133-147.
Received: September 8, 1986 Accepted: March 5, 1987
Help Volumes Main Menu
Related Documents











