1 23 Environmental Management ISSN 0364-152X Volume 49 Number 4 Environmental Management (2012) 49:802-815 DOI 10.1007/s00267-012-9815-8 Using a Down-Scaled Bioclimate Envelope Model to Determine Long-Term Temporal Connectivity of Garry oak (Quercus garryana) Habitat in Western North America: Implications for Protected Area Planning Marlow G. Pellatt, Simon J. Goring, Karin M. Bodtker & Alex J. Cannon

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
Environmental Management ISSN 0364-152XVolume 49Number 4 Environmental Management (2012)49:802-815DOI 10.1007/s00267-012-9815-8
Using a Down-Scaled Bioclimate EnvelopeModel to Determine Long-Term TemporalConnectivity of Garry oak (Quercusgarryana) Habitat in Western NorthAmerica: Implications for Protected AreaPlanningMarlow G. Pellatt, Simon J. Goring,Karin M. Bodtker & Alex J. Cannon

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media, LLC. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

RESEARCH
Using a Down-Scaled Bioclimate Envelope Model to DetermineLong-Term Temporal Connectivity of Garry oak (Quercusgarryana) Habitat in Western North America: Implicationsfor Protected Area Planning
Marlow G. Pellatt • Simon J. Goring •
Karin M. Bodtker • Alex J. Cannon
Received: 2 March 2011 / Accepted: 9 January 2012 / Published online: 19 February 2012
� Springer Science+Business Media, LLC 2012
Abstract Under the Canadian Species at Risk Act
(SARA), Garry oak (Quercus garryana) ecosystems are
listed as ‘‘at-risk’’ and act as an umbrella for over one
hundred species that are endangered to some degree.
Understanding Garry oak responses to future climate sce-
narios at scales relevant to protected area managers is
essential to effectively manage existing protected area
networks and to guide the selection of temporally con-
nected migration corridors, additional protected areas, and
to maintain Garry oak populations over the next century.
We present Garry oak distribution scenarios using two
random forest models calibrated with down-scaled biocli-
matic data for British Columbia, Washington, and Oregon
based on 1961–1990 climate normals. The suitability
models are calibrated using either both precipitation and
temperature variables or using only temperature variables.
We compare suitability predictions from four General
Circulation Models (GCMs) and present CGCM2 model
results under two emissions scenarios. For each GCM and
emissions scenario we apply the two Garry oak suitability
models and use the suitability models to determine the
extent and temporal connectivity of climatically suitable
Garry oak habitat within protected areas from 2010 to
2099. The suitability models indicate that while 164 km2 of
the total protected area network in the region (47,990 km2)
contains recorded Garry oak presence, 1635 and 1680 km2
of climatically suitable Garry oak habitat is currently under
some form of protection. Of this suitable protected area,
only between 6.6 and 7.3% will be ‘‘temporally connected’’
between 2010 and 2099 based on the CGCM2 model.
These results highlight the need for public and private
protected area organizations to work cooperatively in the
development of corridors to maintain temporal connectiv-
ity in climatically suitable areas for the future of Garry oak
ecosystems.
Keywords Climate change � Downscaling � Bioclimate
envelope modeling � British Columbia � Garry oak
(Quercus garryana) � Protected area � Western North
America � Temporal connectivity � Pacific Northwest �Washington � Oregon
Introduction
Protected area managers are faced with the daunting task of
trying to retain biodiversity in the face of environmental
and anthropogenic stressors, including global climate
change. Intact ecosystems enhance environmental, social
and economic resilience to the impacts of climate change,
thus ecosystem conservation and restoration activities,
including establishment and effective management of
protected areas, is increasingly seen as an important
mechanism in the mitigation and adaptation of ecosystems
to climate change. In order to manage for ecological
M. G. Pellatt � K. M. Bodtker
Parks Canada, Western and Northern Service Centre, 300-300
West Georgia Street, Vancouver, BC V6B 6B4, Canada
M. G. Pellatt (&)
School of Resource and Environmental Management, Simon
Fraser University, Burnaby, BC, Canada
e-mail: [email protected]
S. J. Goring
Department of Biological Sciences, Simon Fraser University,
Burnaby, BC, Canada
A. J. Cannon
Meteorological Service of Canada, Environment Canada,
Vancouver, BC, Canada
123
Environmental Management (2012) 49:802–815
DOI 10.1007/s00267-012-9815-8
Author's personal copy

resilience, protected area managers need to understand the
potential impacts of climate change on species and eco-
systems at scales relevant to their management plans.
Ecological models using downscaled variables from Gen-
eral Circulation Model (GCM) simulations can help man-
agers and scientists determine climate stressors influencing
ecosystems and the species that they steward. The use of
bioclimatic models to predict potential shifts in the geo-
graphic distributions of plants and animals due to climate
change are becoming more prevalent in the literature
(Araujo 2009; Heikkinen and others 2006). Applications
range from modeling potential shifts of many individual
tree or plant species (e.g., Berry and others 2002; Hamann
and Wang 2006; Rehfeldt and others 2009), to estimating
impacts on biodiversity (Midgley and others 2002; Midgley
and others 2003), extinction risk (Hannah and others 2005),
and conservation planning (Dockerty and Lovett 2003).
More recently the idea of temporal connectivity or tem-
poral corridors for species has been presented in the liter-
ature (Rose and Burton 2009). Temporal connected areas
maintain a suitable climate for a given species or ecosys-
tem over a defined period of time. This tool has much
potential to inform management planning and climate
adaptation strategies for protected areas. Hence bioclimatic
models can be valuable tools for conservation practitioners
planning for individual species responses at local or
national systems planning levels.
When the conservation target is a well defined plant
community, temporal connectivity can be assessed based
on pre-defined community composition (Rose and Burton
2009); however, plant communities that cross political
boundaries are often subject to different classification
schemes, making regional predictions of community con-
nectivity difficult. In cases where communities are defined
largely by the presence of a key species, a single-species
model may be able to capture much of the bioclimatic
range of the community as a whole. This is especially
useful when geo-referenced or an accurate knowledge of
actual species range occurs. Connected areas of climate
suitability are important in shaping the ecosystems present
now and in the future (Davis 1981; Graham and others
2010).
Garry oak (Quercus garryana), is a broadleaved decid-
uous hardwood common inland along the Pacific Coast of
the US and has the longest north–south distribution among
western oaks, occurring from Vancouver Island to southern
California. It is the only native oak in British Columbia and
Washington and the principal one in Oregon (Burns and
Honkala 1990). The range of Garry oak spans more than
15� of latitude from just below the 50th parallel on Van-
couver Island nearly to latitude 34�N. in Los Angeles
County, CA. South of Courtenay, BC, Garry oak occurs in
the eastern and southernmost parts of Vancouver Island and
on adjacent smaller islands from near sea level to
approximately 200 m. In Canada, Garry oak ecosystems
are categorized as a suite of ecosystems within the dry
Coastal Douglas-fir Biogeoclimatic Zone (CDF). This zone
occurs on southeast Vancouver Island, Gulf Islands, and
portions of the British Columbia’s lower mainland. It is
restricted to lower elevation sites characterized as rela-
tively warm and dry, such as slopes with southern expo-
sure, hilltops, rocky outcrops, and coastal bluffs. Many
plant communities within the historic range of Garry oak
ecosystems were dependent on periodic disturbance for
continued persistence across a dynamic landscape. These
sites, usually with deeper soils, were maintained by natural
means, such as annual periods of inundation or wildfire, or
possibly by cultural management practices, including plant
resource harvesting and prescribed burning. In addition,
other threats such as habitat loss and fragmentation, inva-
sive species, and herbivory are affecting the ecological
integrity of Garry oak ecosystems (MacDougall and others
2010). This has lead to the listing of Garry oak ecosystems
under the Canadian Species at Risk Act (SARA). Garry oak
ecosystems are listed as ‘‘at-risk’’ and act as an umbrella
for over one hundred species that are endangered to some
degree (Fuchs 2001).
The decline of Garry oak (and western North American
oak ecosystems as a whole) is attributed to a number of
causes, but changes in disturbance regimes such as fire and
grazing, appear to be two important factors (MacDougall
and others 2010; Pellatt and others 2007). Active man-
agement is being applied in order to successfully maintain
or restore Garry oak ecosystems, (Garry Oak Ecosystem
Recovery Team, Washington State, and Parks Canada
Agency), but site selection and restoration techniques
applied to maintain or restore Garry oak ecosystems tends
to be opportunistic and forethought into the ultimate
adaptation of Garry oak to climate change is limited.
Studies of ecosystem change at the larger landscape level
have been undertaken for many of British Columbia’s
forest species (Hamann and Wang 2006), but a need is still
apparent to examine non-commercial, but ecologically
important species like Garry oak at a scale that is relevant
to protected area managers.
Bioclimatic models should be viewed as an important
source of information in future land-use planning deci-
sions, including the restoration and recovery of endangered
species and protected area network design (Araujo 2009).
The need to translate research into practice and to present
these models as management tools is critical to the
development of management and adaptation strategies.
Once these models are developed, dynamic models can be
created to consider edaphic factors, disturbance regimes,
competition, and other complex ecosystem processes
(Araujo and Rahbek 2006; Heikkinen and others 2006).
Environmental Management (2012) 49:802–815 803
123
Author's personal copy

The bioclimatic models presented in this paper are to be
included in protected area management plans. These plans
will inform species recovery groups about possible climate
change impacts on Garry oak ecosystems and provide
direction for ecosystem restoration projects that use best
practices by including climate change adaptation of Garry
oak into the planning process. This information is partic-
ularly important because the rate of expected climate
change may well be unprecedented during the Holocene
resulting in climates and ecosystems with no modern or
past analogue (Froyd and Willis 2008; Williams and others
2007).
The objectives of this study are to build upon and refine
work by Bodtker and others (2009) and apply it to the
question of protected area planning under changing climate
scenarios that are projected until the end of the 21st century.
Garry oak ecosystems are one of the most endangered
ecosystems in Canada and the Parks Canada Agency has
established the Gulf Island National Park Reserve (GINPR)
in a region where these ecosystems occur. Under Canada’s
Species at Risk Act (SARA), Parks Canada is the lead
federal agency responsible for the recovery of Garry oak
ecosystems, so obvious interest exists in the long term via-
bility of the national park reserve in the protection of these
ecosystems. Fully realizing that international, provincial,
and local cooperation is needed to maintain, restore, and
allow for future adaptation of at-risk species in Garry oak
ecosystems, we evaluate the level of protection of Garry oak
at present and then forecast how well protected areas in the
Pacific Northwest of North America encompass climatically
suitable Garry oak habitat in the future. We then identify the
areas that are predicted to maintain climatic suitability in all
temporal scenarios that we examine. Most importantly we
generate scenarios examining temporally connected areas
that persist throughout the 21st century for Garry oak as well
as how well existing protected areas overlap these tempo-
rally connected regions.
Data and Methods
Study Area
To build a quantitative model of the relationship between a
species or ecosystem and the total range of current climatic
values, the complete global distribution of the species
should ideally be considered (Dockerty and Lovett 2003).
In this case we felt an exception was warranted. Although
Garry oak extends as far south as 35�North (Fig. 1), the
composition of Garry oak-related ecosystems in the region
south of Oregon is significantly different from those to the
north (Fuchs 2001) so we did not include California in the
study area. Based on biogeographic and ecological
characteristics of Garry oak ecosystems in North America
and our knowledge of future climate scenarios, we included
areas to the north and east of the current Garry oak range,
defining a study area extending from 42�–51� North lati-
tude and 119�–129� West longitude, an area encompassing
southwestern British Columbia, Washington, and Oregon
(Fig. 1).
Climate Data
Climate Data for Model Building
In order to develop a bioclimate envelope model at a
suitable spatial scale for our objectives, a resolution
1 9 1 km was used. The 1 9 1 km resolution for the study
area resulted in a raster size of 660 9 1,168 grid cells, of
which 428,592 were on land. To generate the 33 down-
scaled climate variables at the 1 9 1 km resolution used in
this study, a bilinear interpolation/polynomial regression-
based lapse rate upsampling algorithm was applied to a
merged 4 km US/Canada Parameter Regression on Inde-
pendent Slopes Model (PRISM) (Daly and others 1994,
1997) 1961–1990 climate normal dataset following Wang
and others (2006). Using climatologically-aided interpola-
tion (Willmott and Robeson 1995), Global Climate Model
(GCM) scenarios were generated by superimposing future
Fig. 1 Study area and present extent of Garry oak in British
Columbia, Washington, and Oregon. Red represents actual occurrence
of Garry oak. Green represents the location of protected areas
804 Environmental Management (2012) 49:802–815
123
Author's personal copy

GCM climate anomalies relative to the 1961–1990 period
overtop the 1 9 1 km downscaled climate normals
(Table 1).
Climate Data for Scenario Development
In this study we develop a presence/absence (P/A) model
for Garry oak under various future climate scenarios pre-
dicted by four GCMs (CCSRNIES, CSIRO, CGCM2 and
the HADCM3). Each model uses GCM data to predict
conditions driven by two future emissions scenarios: Sce-
nario A2 is driven by a continuously growing world pop-
ulation with regionally oriented economic development
and slow technological advancement. Scenario B2 is driven
by continuous population growth at a lower rate than sce-
nario A2 with emphasis on local solutions to economic,
social and environmental stability, and thus, overall lower
CO2 concentrations than the A2 scenario.
For the development of Garry oak climate suitability
scenarios, we constructed two parallel models using output
from four GCMs. This was done because confidence in
precipitation values modelled using GCMs are generally
lower than for temperature variables. The first model uses a
number of climate parameters selected for their ecological
relevance including temperature and precipitation vari-
ables, the second uses only temperature variables
(Table 2).
We chose to use outputs from four GCMs in an effort to
compare outputs across a range of possible future scenar-
ios. Individual GCM models share some assumptions about
climatological phenomena, but differ in the full suite of
assumptions and also in the mechanics of the model, often
resulting in variable outputs, especially at longer time-
scales (Salathe and others 2007). Prior analysis by Salathe
and others (2007) in the Pacific Northwest indicates that
the models vary both in their expressions of modern cli-
mate normals and in their expressions of future climate
change (Table 3). Since the outcomes from these four
models broadly represent anticipated model outcomes for
the region we believe that they are appropriate for our P/A
modeling. Importance values for the Garry oak models
indicate that P/A is most strongly determined by winter
precipitation, mean summer precipitation and the annual
heat:moisture index (Table 2).
Model Building
The P/A model for Garry oak is generated using random
forest (Breiman 2001), based on downscaled modern cli-
mate normals (1961–1990; Wang and others 2006) and the
geo-referenced distribution of Garry oak in the Pacific
Northwest region of North America (Chappell and others
2003, Ecological Society of America 2006, Harrington
2003, Klinkenberg 2004). Preliminary work on the
assessment and comparison of downscaled climate models,
different GCMs, and statistical models to develop a bio-
climatic envelope model for Garry oak was undertaken by
Bodtker and others (2009) who were able to show that
random forest (Breiman 2001) models provide the best
predictive accuracy of modern Garry oak distributions in
Table 1 Downscaled climate variables generated by applying the Wang and others (2006) algorithm to combined US/Canada PRISM data,
described in Wang and others (2006)
Variable Description and units Variable Range
MAT Mean annual temperature (�C) TAVwt Seasonal Temperatures (�C, wt–Average
winter; sp–Average spring; sm–Average
summer; at–Average autumn)
MWMT Mean warmest month temperature (�C) TAVsp
MCMT Mean coldest month temperature (�C) TAVsm
TD Continentality (MWMT–MCMT, �C) TAVat
MAP Mean annual precipitation (mm) TMXwt Seasonal maximum temperatures (�C)
MSP Mean summer precipitation (mm) TMXsp
AHM Annual heat moisture index (MAT ? 10)/(MAP/1000) TMXsm
SHM Summer heat moisture index (MWMT ? 10)/(MSP/1000) TMXat
DD \ 0 degree-days below freezing (0�C) TMNwt Seasonal minimum temperatures (�C)
DD \ 18 Heating degree days (\18�C) TMNsp
DD [ 18 Cooling degree days ([18�C) TMNsm
bFFP Julian calendar beginning of the frost free period TMNat
eFFP Julian calendar end of the frost free period PPTwt Seasonal precipitation (mm)
FFP Length of the frost free period PPTsp
DD5 [ 100 Budburst, Julian date DD [ 5 reaches 100 PPTsm
EXT_Cold Coldest 30 year temperature (�C) PPTat
Environmental Management (2012) 49:802–815 805
123
Author's personal copy
the Pacific Northwest using several model evaluation
metrics.
Initial model building and selection (Bodtker and others
2009) considered three statistical models; logistic regres-
sion, random forest classification and maximum likelihood.
Preliminary analysis using the Kappa Indices of Agreement
(KIA) and the relative operating characteristic curve
(ROC) in IDRISI (Eastman 2006) indicated that random
forest provided the optimum model for Garry oak climate
suitability (Bodtker and others 2009). For this reason this
paper will focus solely on model output from the random
forest model.
Random forest classification results from the generation
of a large number of classification trees. Each tree is
generated randomly, with nodes split using the best of a
randomly drawn subset of predictor variables from the
original dataset (Breiman 2001). Within the forest each tree
classifies the new data and returns a vote, in this case
presence or absence, thus the ultimate prediction is the
result of hundreds or thousands of votes by randomly
generated trees. By generating a ‘‘forest’’ of classification
trees it is possible to determine which predictor variables
generate the best splits by aggregating votes across all
trees. This method has been used before to predict climate
suitability for trembling aspen (Populus tremuloides) in
western North America under various climate change
scenarios (Rehfeldt and others 2009), and has been shown
to have high predictive success in modelling species ranges
(Lawler and others 2006).
Analysis was carried out in R (R Development Core Team
2011) using the randomForest package (Liaw and Wiener
2002). We selected climate variables for the model based on
both ecological considerations and importance values
returned by a fully parameterized predictive model. Impor-
tance values are returned as a model diagnostic in random
forest classification, and indicate the drop in classification
error as a result of the inclusion of a particular variable in the
splitting criteria within a node in a tree. Simulation has shown
that importance values are acceptable selection criteria for
cases where all variables are continuous or have a similar
numbers of factors (Strobl and others 2007).
Cells with Garry oak presence represent a small pro-
portion of the total cells in the range (3,256 km2 of the total
425,336 km2) which may increase the likelihood of Type II
error in the model, by which sites with potentially suitable
Garry oak habitat are predicted to be unsuitable. Since
modern Garry oak habitat is constrained by both environ-
mental and anthropogenic factors it is possible that
‘absence’ cells are the result of historic human activity
rather than non-climatic influence over Garry oak distri-
bution. Reducing Type II error comes at the expense of
Type I error, however since anthropogenic factors may be
constraining Garry oak distribution, and since preservation
of Garry oak habitat in the future will likely rely on some
form of intensive management we felt that this was an
acceptable trade-off.
Table 3 Climate forecasting features of the four GCMs used to build the presence/absence model for Garry oak in the Pacific Northwest, based
on work by Salathe and others (2007)
Model Model bias Climate variables
Temperature Precipitation Temperature change to 2099 Precipitation percent change to 2099
A2 (�C) B2 (�C) A2 (%) B2 (%)
CGCM Neg. [ 1 SD Pos \ 1 SD 3 1.5 15 0
CSIRO Pos \ 1 SD Pos [ 1 SD 6 2 7 7
CCSM/NIES Neg. \ 1 SD Pos \ 1 SD 2 1 8 0
HADCM3 Neg. \ 1 SD Pos \ 1 SD 4 2 22 12
Table 2 Importance values for climate parameters of interest in the
downscaled CGCM2 model for British Columbia generated from
climate normals for 1961–1990
Climate parameter Importance value
FFP (frost free period) 100.74
MAP (mean annual precipitation) 101.74
TMXsp (maximum summer temperature) 102.79
PPTsm� (summer precipitation) 103.44
MWMT (mean warm month temperature) 107.33
TMXwt (maximum winter temperature)* 114.52
DD.5 (degree days above 5�C) 115.39
TMXsm (maximum summer temperature) 124.48
TAVsp (average spring temperature)* 145.37
PPTsp (total spring precipitation) 149.14
DD.0 (degree days above 0�C)*� 152.68
TAVsm (average summer temperature)*� 153.01
TD (temperature difference–summer to winter)*� 175.13
TMNat (mean autumn temperature)*� 175.85
AHM (annual heat: moisture index)� 185.01
MSP (mean summer precipitation) 186.58
PPTwt (total winter precipitation) � 251.32
The symbols * and � indicate the use of the parameter in the non-
precipitation and precipitation based models, respectively
806 Environmental Management (2012) 49:802–815
123
Author's personal copy

To reduce Type II error we built the model using only
terrestrial cells within 40 km of a cell with known Garry
oak presence rather than the entire study region. Reducing
the total number of cells in this way increased the per-
centage of Garry oak cells to 2.1% of the total dataset.
Since we are more interested in regions that may be suit-
able for Garry oak in the future we wanted to give greater
weight to Garry oak cells. To do this we then constructed
the models by sub-sampling the absence cells, so that they
equalled the number of Garry oak cells. Because the
selection of absence cells was likely to affect the model we
repeated this procedure 100 times, generating 100 separate
P/A predictions for Garry oak, each time re-sampling
the absence cells so that they were equal in number to the
presence cells. We report presence as any site for which the
model predicts presence in more than 50% of the cases.
Protected Area Networks
The ultimate goal of this work is to assess scenarios gen-
erated for Garry oak climate suitability for the network of
protected areas in British Columbia, Washington, and
Oregon over the 21st century. Of particular interest are
areas that maintain continual climate suitability for Garry
oak ecosystems. These results can then assist in regional
planning for Garry oak ecosystem conservation in the
foreseeable future.
A network of protected areas exists through the study
region with a total area of 169,904 km2. Protected areas
range in size from 25 m2 (part of the WA state land trust)
to 50,447 km2 (Oregon, Bureau of Land Management).
The distribution of area values is highly skewed; the
average area is 27 km2 while the median is 0.18 km2. For
the purpose of this study we define protected areas as all
lands protected as parks, various forms of ecological
reserves, experimental forests, or recreation areas by fed-
eral, state or provincial governments. This includes lands
managed by the Parks Canada Agency, the British
Columbia Ministry of the Environment, the United States
Forest Service, the Bureau of Land Management, and the
United States National Parks Service. In practice this land
may currently be under management for resource extrac-
tion, however our objective is to demonstrate the extent of
the landscape upon which management practices may
result in preservation or expansion of suitable habitat and
so our definition of what may constitute protected areas is
broad since we believe that the larger the number of
stakeholders, the greater the possibility that some will
change management practices to accommodate Garry oak
ecosystem adaptation.
The spatial data for the United States was obtained from
the Protected Areas Database (The Conservation Biology
Institute. May 2010. PAD-US 1.1 (CBI Edition). Corvallis,
Oregon) while Canadian data was obtained through
Geogratis (http://geogratis.cgdi.gc.ca). Because of admin-
istrative differences between the United States and Canada,
the extent of managed land in Canada is underreported.
Much of the land base is retained by the Government of
British Columbia and is leased to resource users. This
means that there is land in British Columbia that may have
the potential for the active management of Garry oak that is
currently undergoing resource extraction. Protected areas
in the US are defined as all locations with an IUCN con-
servation ranking based on the PAD-US dataset. Within
British Columbia all protected areas administered by
municipal, provincial, or federal authorities (i.e., National
and Provincial parks, wildlife reserves, conservation areas
and regional parks) jurisdiction are considered as protected.
Much original Garry oak habitat has been lost due to
development and many of the ecological processes neces-
sary for Garry oak persistence have been altered (Lea 2007;
Pellatt and others 2007; Vellend and others 2008; Mac-
Dougall and others 2010). The extent of development
around deep-soiled sites that are presumed to be central to
Garry oak habitat means that changes in habitat suitability
may place serious pressures on the ability of Garry oak to
again reach equilibrium with a changing climate. The
extent of development may also affect our ability to predict
suitable habitat, since much of the original habitat extent
may be lost. This may be offset in part by our grid reso-
lution. The presence of a single Garry oak tree within a
1 km2 grid cell will result in that cell being classified as
indicating presence. However, for this reason we will
principally be investigating the changes in habitat suit-
ability among the region’s protected areas.
Results
Model Output
The fully parameterized (FPm) model using climate nor-
mals from 1961–1990 had a bootstrapped (n = 100) error
of 9.2 ± 0.3%. The smaller model (Pm) including precip-
itation and temperature values (Table 2, parameters indi-
cated by dagger) had an error that was not significantly
different than the model using only temperature-based
variables (NPm; t = -4.01, df = 195.6, P \ 0.001). The
extent of suitable Garry oak sites predicted by the modern
models is broader than the current Garry oak extent, and
Garry oak suitability is predicted in regions that do not
currently support Garry oak populations. The NPm model
predicts a broader extent for Garry oak suitability, specif-
ically indicating broader suitability on the mid-Oregon
coast and along the northern edge of the Olympic Peninsula
in Washington (Fig. 2a, b). The populations in central
Environmental Management (2012) 49:802–815 807
123
Author's personal copy

Washington, scattered suitable sites in central Oregon, and
coastal sites in NW Washington predicted by both models
are unlikely to exist from both a climatic and biogeo-
graphic perspective. It is unlikely that Garry oak will
migrate to some of these sites due to climate related and
other extrinsic factors not testable with this model.
Only 5.0%, or 164 km2 of the current observed Garry
oak extent (3,256 km2) is protected, but it is likely that
most of the existing protected sites have been degraded to
some degree (Fuchs 2001). The Pm model predicts less
regional suitability (26,830 km2 of suitable habitat,
1635 km2 in protected areas) while the NPm model pre-
dicts 27,945 km2 of suitable habitat, of which 1680 km2 is
protected. Based on the four GCMs and the consensus
model, the area covered by suitable Garry oak habitat
within the study area increases over time for both the
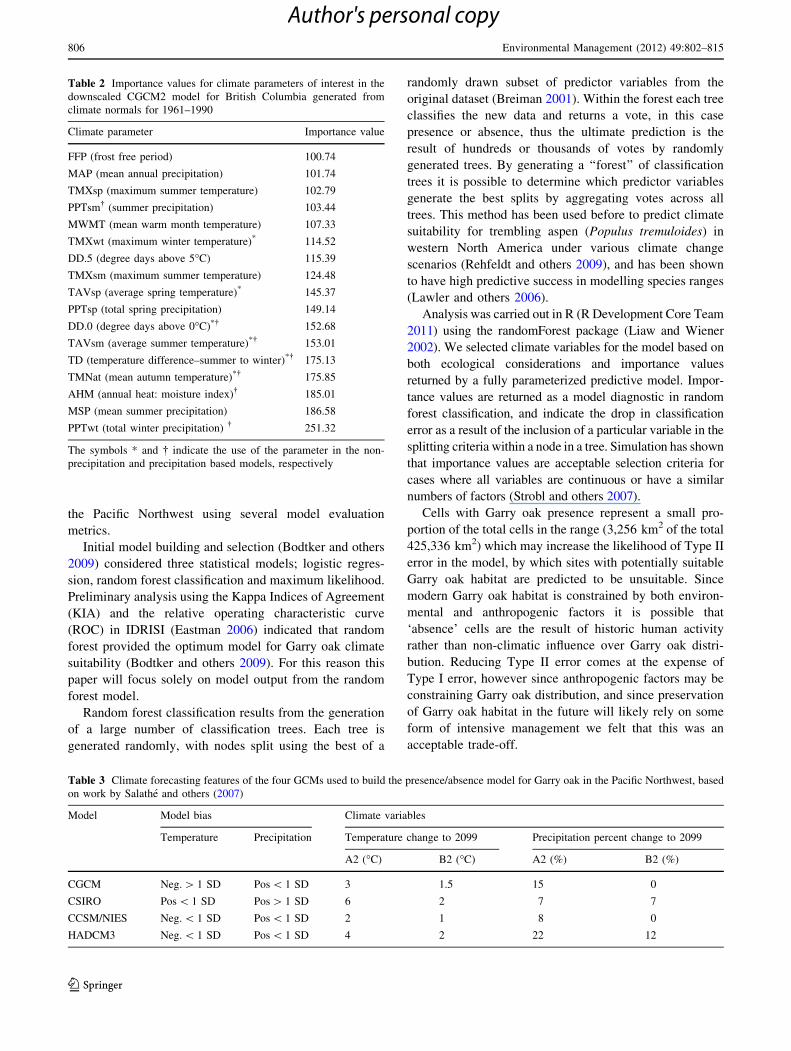
region as a whole and the protected area network (Fig. 3).
Suitability under the A2 scenarios is predicted to be higher
than B2 suitability for all GCMs for both the Pm and the
NPm models (Fig. 3). There appears to be a broad range of
outcomes among GCMs, although predictions using all
four GCMs broadly show increases in suitability. The
CCSR/NIES A2 emissions scenario produces the highest
predictions of Garry oak suitability, except within the NPm
full region model (Fig. 3). Other models appear to show
similar results across ranges (full range or protected areas)
and model types (Pm or NPm). All the GCMs indicate that
climatically suitable Garry oak habitat increases over time
(under A2 and B2 scenarios) in the study region as a whole
and in the protected area network (Fig. 3).
We chose to use the CGCM2 model for further analysis
because the model outputs from the A2 and B2 emissions
scenarios show strong divergence between regions (Figs. 4,
5). Thus the CGCM2 results should illustrate the range of
possible outcomes for Garry oak suitability throughout the
study region. In addition, the Pm model using the A2
scenario (Fig. 4a) generates the lowest predictions of future
Garry oak suitability, providing a worst-case baseline from
which to examine potential management implications of
climate change on Garry oak climate suitability.
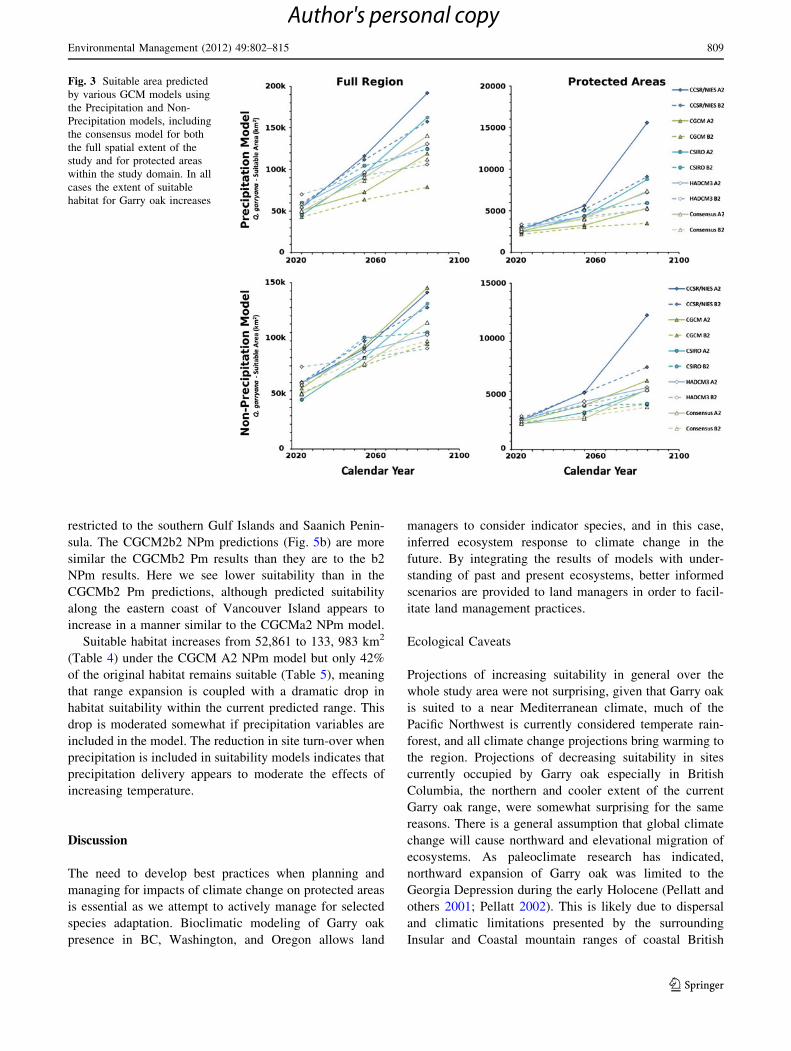
The CGCM2a2 Pm predictions show increases in suit-
ability along the eastern coast of Puget Sound and ulti-
mately along the Pacific coast (Fig. 4a), coupled with
expansion eastward across the Cascade Mountains into the
Klamath Basin. CGCM2b2 Pm predictions (Fig. 4b) indi-
cate less extensive increases in suitability, and lower suit-
ability in the eastern Cascade Range, Klamath Basin and
along the Pacific Coast.
The CGCM2a2 NPm predictions (Fig. 5a) show
increased suitability in the Klamath and eastern Cascade
Range, but interestingly, lower suitability in the Puget
Sound and along the southeastern coast of Vancouver
Island when compared to the CGCM2a2 Pm predictions.
Suitability under the NPm CGCM2a2 model does increase
further north along Vancouver Island, however the current
extent of protected Garry oak in British Columbia is largely
Fig. 2 (a) Study area showing
the predicted distribution of
Garry oak using the random
forest model with precipitation
included. Red represents 100%
suitable habitat grading to
yellow that represents 50%
suitable habitat. Greenrepresents the location of
protected areas; (b) Study area
showing the predicted
distribution of Garry oak using
the random forest model
without precipitation included.
Red represents 100% suitable
habitat grading to yellow that
represents 50% suitable habitat.
Green represents the location of
protected areas
808 Environmental Management (2012) 49:802–815
123
Author's personal copy
restricted to the southern Gulf Islands and Saanich Penin-
sula. The CGCM2b2 NPm predictions (Fig. 5b) are more
similar the CGCMb2 Pm results than they are to the b2
NPm results. Here we see lower suitability than in the
CGCMb2 Pm predictions, although predicted suitability
along the eastern coast of Vancouver Island appears to
increase in a manner similar to the CGCMa2 NPm model.
Suitable habitat increases from 52,861 to 133, 983 km2
(Table 4) under the CGCM A2 NPm model but only 42%
of the original habitat remains suitable (Table 5), meaning
that range expansion is coupled with a dramatic drop in
habitat suitability within the current predicted range. This
drop is moderated somewhat if precipitation variables are
included in the model. The reduction in site turn-over when
precipitation is included in suitability models indicates that
precipitation delivery appears to moderate the effects of
increasing temperature.
Discussion
The need to develop best practices when planning and
managing for impacts of climate change on protected areas
is essential as we attempt to actively manage for selected
species adaptation. Bioclimatic modeling of Garry oak
presence in BC, Washington, and Oregon allows land
managers to consider indicator species, and in this case,
inferred ecosystem response to climate change in the
future. By integrating the results of models with under-
standing of past and present ecosystems, better informed
scenarios are provided to land managers in order to facil-
itate land management practices.
Ecological Caveats
Projections of increasing suitability in general over the
whole study area were not surprising, given that Garry oak
is suited to a near Mediterranean climate, much of the
Pacific Northwest is currently considered temperate rain-
forest, and all climate change projections bring warming to
the region. Projections of decreasing suitability in sites
currently occupied by Garry oak especially in British
Columbia, the northern and cooler extent of the current
Garry oak range, were somewhat surprising for the same
reasons. There is a general assumption that global climate
change will cause northward and elevational migration of
ecosystems. As paleoclimate research has indicated,
northward expansion of Garry oak was limited to the
Georgia Depression during the early Holocene (Pellatt and
others 2001; Pellatt 2002). This is likely due to dispersal
and climatic limitations presented by the surrounding
Insular and Coastal mountain ranges of coastal British
Fig. 3 Suitable area predicted
by various GCM models using
the Precipitation and Non-
Precipitation models, including
the consensus model for both
the full spatial extent of the
study and for protected areas
within the study domain. In all
cases the extent of suitable
habitat for Garry oak increases
Environmental Management (2012) 49:802–815 809
123
Author's personal copy
Columbia. Climate and corresponding ecological change
has been rapid and significant in the region throughout the
Holocene (last 10,000 years), indicating that further work
is needed to understand ecosystem thresholds.
There are several factors that influence the distribution
of Garry oak on the landscape that cannot be taken into
account with this model. These include site factors such as
type of soil, soil properties that vary seasonally like
wetness, and the persistent foggy conditions that occur
seasonally in some coastal areas. Currently Garry oak
morphology (small gnarled trees versus tall and sturdy
trees) and habitat types (deep soil, shallow soil, exposed
rocky outcrops) are varied and likely the result of addi-
tional factors such as successional processes and distur-
bance regimes involving fire and agriculture. These factors
will likely continue to play a role in determining the
Fig. 4 Study area showing the predicted distribution of Garry oak
using the CGCM2 random forest model with precipitation included
for three time scenarios: 2010–2039, 2040–2069, and 2070–2099. a is
the A2 scenario and b is the B2 scenario. Red represents 100%
suitable habitat grading to yellow that represents 50% suitable habitat.
Green represents the location of protected areas
810 Environmental Management (2012) 49:802–815
123
Author's personal copy

distribution of Garry oak at a fine scale in areas where it is
climatically suitable.
Since deglaciation (last *12,000 years) climate has
been the primary driver of Garry oak distribution in the
Pacific Northwest of North America (Pellatt and others
2001, 2007). Even with the ever increasing pressure of
habitat loss, climate change impacts will likely deleteri-
ously affect Garry oak ecosystems; rapid change in eco-
system structure due to climate change, invasive species,
species migration, suppression of natural disturbance
regimes, and habitat fragmentation should be expected to
affect Garry oak and associated ecosystems (Shafer and
others 2001; Pellatt and others 2007).
Protected Areas
Protected areas represent a long-term commitment to the
conservation of species, ecological processes and, in many
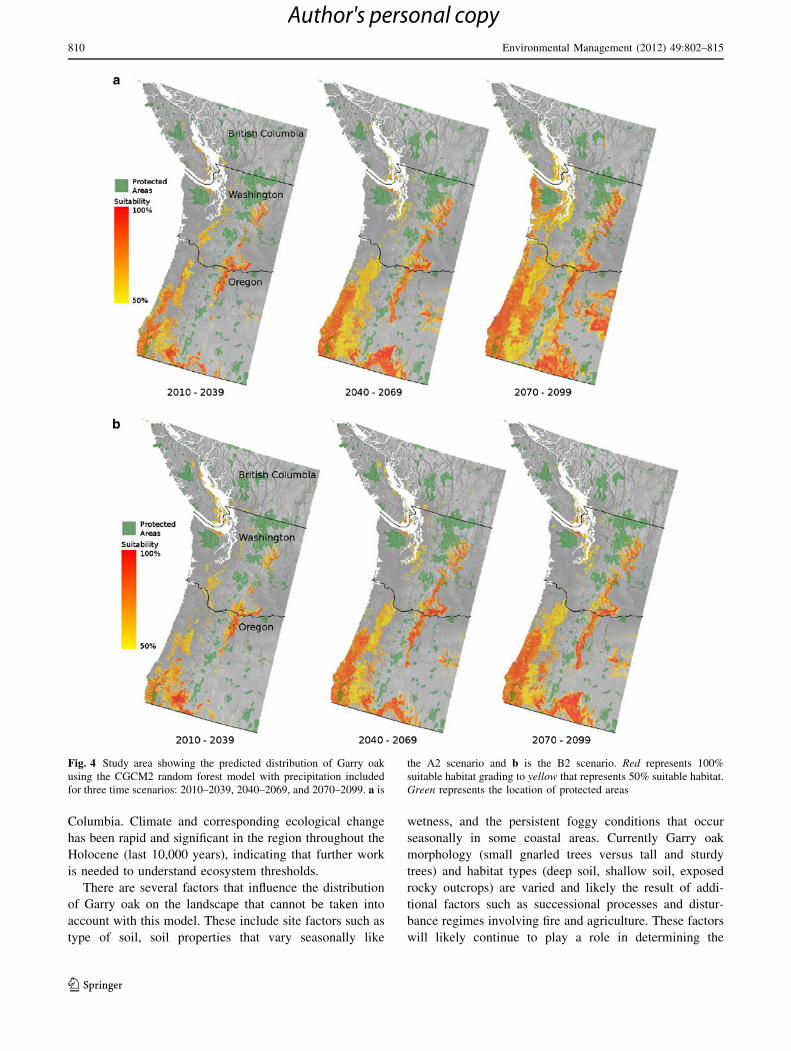
Fig. 5 Study area showing the predicted distribution of Garry oak
using the CGCM2 random forest model without precipitation
included for three time scenarios: 2010–2039, 2040–2069, and
2070–2099. a is the A2 scenario and b is the B2 scenario. Redrepresents 100% suitable habitat grading to yellow that represents
50% suitable habitat. Green represents the location of protected areas
Environmental Management (2012) 49:802–815 811
123
Author's personal copy

cases, associated cultural values and resources. They are
often accorded legal recognition, have agreed-upon man-
agement and governance approaches, and are supported by
management planning and capacity. It is these qualities that
make investments in protected area establishment and
management cost-effective in the context of climate change
adaptation. There will be pressure to reconsider what the
perceived function of a protected area is as the ecosystems
within it begin to change. The need to critically assess the
purpose of a particular area may serve to determine if it
will remain relevant to the public. We have moved from a
single species approach to an ecosystem-based approach to
protected area management, but still we need to consider
ecosystems in the context of a larger temporal framework.
The paucity of protected areas in locations critical for
the protection of Garry oak ecosystems, now, and in future
climatically suitable areas, appears to be the greatest
challenge for conservation of the species when considering
future global change.
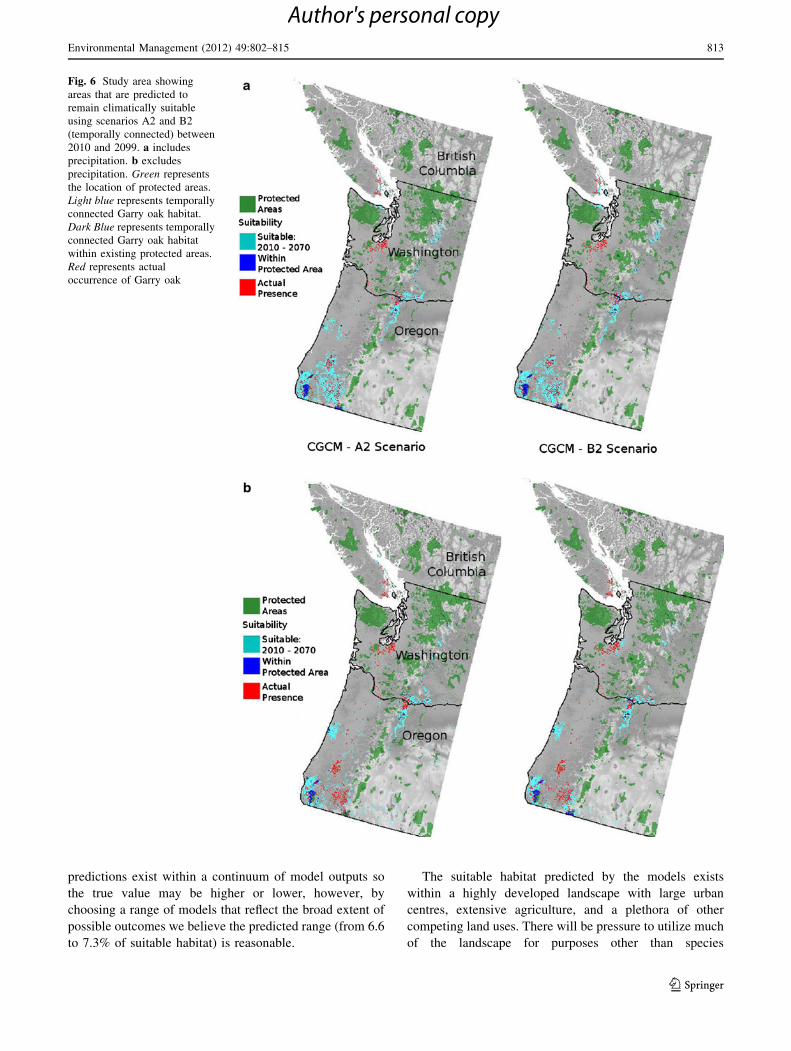
Temporal Connectivity
Although climatically suitable Garry oak habitat will
increase somewhat, primarily in the USA, this habitat will
not be well represented in IUCN Classes I through V
Protected Areas (5.6–7.3%). The distribution of protected
areas in relation to temporally connected climatically
suitable areas between 2010 and 2099 is shown in Fig. 6a
(CGCM2a2), b (CGCM2b2). Of the protected area that
currently contains suitable Garry oak habitat, our models
indicate that between 6.6 and 7.3% will be ‘‘temporally
connected’’ between 2010 and 2099. The CGCM2
Table 4 Change in Garry oak habitat suitability based on the CGCM2–A2 model for climate change. The size of the area of interest is
428,592 km2. Precipitation (PPT)
2010–2039 2040–2069 2070–2099
With PPT
(km2)
Without PPT
(km2)
With PPT
(km2)
Without PPT
(km2)
With PPT
(km2)
Without PPT
(km2)
CGCMa2
Total Region
Predicted suitable habitat 51,947 52,861 84,687 80,737 137,053 133,983
Protected area network 20,548 22,392 37,250 37,255 59,931 60,343
Canada
Predicted suitable habitat 1528 1328 1258 4345 3575 9933
Protected area network 265 108 344 714 985 1435
CGCMb2
Total Region
Predicted suitable habitat 42,376 52,942 69,175 65,398 81,841 83,367
Protected area network 16,057 20,686 29,803 30,733 37,133 38,085
Canada
Predicted suitable habitat 2701 1885 1889 2616 1443 5934
Protected area network 512 166 471 297 380 937
Table 5 Overlap between Garry oak suitable habitat across time-steps using models for the CGCM2a2 and CGCM2b2 climate change models
2010–2039 2040–2069 2070–2099
With PPT (%) Without PPT (%) With PPT (%) Without PPT (%) With PPT (%) Without PPT (%)
CGCMa2
Modern 20.00 20.20 30.40 26.40 45.90 42.20
2010–2039 – – 85.60 62.80 84.40 72.40
2040–2069 – – – – 90.20 79.30
CGCMb2
Modern 17.10 20.20 25.70 22.40 29.70 26.80
2010–2039 – – 75.00 57.10 72.60 56.20
2040–2069 – – – – 90.60 80.60
Each value is calculated as the percentage of the total area predicted by the model at the time indicated by the row heading still covered at the
column heading
812 Environmental Management (2012) 49:802–815
123
Author's personal copy
predictions exist within a continuum of model outputs so
the true value may be higher or lower, however, by
choosing a range of models that reflect the broad extent of
possible outcomes we believe the predicted range (from 6.6
to 7.3% of suitable habitat) is reasonable.
The suitable habitat predicted by the models exists
within a highly developed landscape with large urban
centres, extensive agriculture, and a plethora of other
competing land uses. There will be pressure to utilize much
of the landscape for purposes other than species
Fig. 6 Study area showing
areas that are predicted to
remain climatically suitable
using scenarios A2 and B2
(temporally connected) between
2010 and 2099. a includes
precipitation. b excludes
precipitation. Green represents
the location of protected areas.
Light blue represents temporally
connected Garry oak habitat.
Dark Blue represents temporally
connected Garry oak habitat
within existing protected areas.
Red represents actual
occurrence of Garry oak
Environmental Management (2012) 49:802–815 813
123
Author's personal copy

conservation hence selecting ‘‘critical habitat’’ identified as
climatically and ecologically suitable in the future will be
essential for the adaptation of Garry oak and its associated
ecosystems. There will likely be increased pressure to
reconsider what the perceived function of a protected area
is as the ecosystems within in it begin to change, and best
practices that will help maintain ecosystems that are more
resistant to invasive species and other stressors exacerbated
by climate change will be essential. We believe that
understanding the temporal connectivity of climatically
suitable habitat for species is a component of these best
practices. Temporal connectivity of suitable climate is
critical to the formation of ecosystems. In the past these are
represented by dynamic refugia, for example, glacial
refugia and climatically suitable migration corridors
throughout the Holocene and deep time (Davis 1981;
Graham and others 2010). There can be no question that
these climatically suitable corridors and refugia will be key
factors in the conservation of biodiversity and ecosystems,
now, and in the future.
Acknowledgments The Climate Change Action Fund (Project
A718), Interdepartmental Recovery Fund (Project 733), Parks Can-
ada—Western and Northern Service Centre, and the Natural Science
and Engineering Research Council of Canada (NSERC) provided
financial support for the project to M.G. Pellatt. S. Goring is sup-
ported by a NSERC RC PGS-D3 scholarship. We would like to thank
D. Peter and C. Harrington of the US. Department of Agriculture
Forest Service, C. Chappell of the Washington Department of Natural
Resources, Ted Lea, the British Columbia Ministry of the Environ-
ment, VegBank (NatureServe), E-Flora (UBC), and D.Hrynyk (Parks
Canada) for data contributions. We would also like to extend our
appreciation to three anonymous reviewers whose positive comments
helped improve the manuscript.
References
Araujo MB (2009) Climate change and spatial conservation planning.
In: Moilenen A, Wilson KA, Possingham HP (eds) Spatial
conservation prioritization: quantitative methods and computa-
tional tools. Oxford University Press, New York
Araujo MB, Rahbek C (2006) How does climate change affect
biodiversity? Science 313:1396–1397
Berry PM, Dawson TP, Harrison PA, Pearson RG (2002) Modelling
potential impacts of climate change on the bioclimatic envelope
of species in Britain and Ireland. Global Ecology & Biogeog-
raphy 11:453–462
Bodtker KM, Pellatt MG, Cannon AJ (2009) A bioclimatic model to
assess the impact of climate change on ecosystems at risk and
inform land management decisions. Report for the Climate
Change Impacts and Adaptation Directorate, CCAF Project
A718. Western and Northern Service Centre, Vancouver
Breiman L (2001) Random Forests. Machine Learning 45:5–32
Burns RM, Honkala BH (1990) Silvics of North America: 1. Conifers;
2. Hardwoods Agriculture Handbook 654, vol 2. US Department
of Agriculture, Forest Service, Washington, DC
Chappell CB, Gee MS, Stephens B (2003) A geographic information
system map of existing grasslands and oak woodlands in the
Puget lowland and Willamette valley ecoregions, Washington.
Washington Natural Heritage Program, Washington Department
of Natural Resources, Olympia (http://www.dnr.wa.gov/nhp/
refdesk/gis/oakgrsld.html)
Daly C, Neilson RP, Phillips DL (1994) A statistical-topographic
model for mapping climatological precipitation over mountain-
ous terrain. Journal of Applied Meteorology 33:140–158
Daly C, Taylor G, Gibson W (1997) The PRISM approach to mapping
precipitation and temperature, 10th Conference on Applied
Climatology. American Meteorological Society, Reno NV,
pp 10–12
Davis MB (1981) Quaternary history and the stability of forest
communities. In:West DC, Shugart HH, Botkin DB (eds) Forest
succession concepts and application. Springer Verlag, New
York, pp 132–153
Dockerty T, Lovett A (2003) A location-centred, GIS-based meth-
odology for estimating the potential impacts of climate change
on nature reserves. Transactions in GIS 7:345–370
Eastman JR (2006) IDRISI Andes. Clark University, Worcester
Ecological Society of America (2006) VegBank on-line database.
http://vegbank.org/cite/VB.Ob.17196.INW28670, VB.Ob.18019.
INW29518, VB.Ob.18020.INW29519, VB.Ob.18021.INW29520,
VB.Ob.18023.INW29522, VB.Ob.18032.INW29531, VB.Ob.180
33.INW29532, VB.Ob.18034.INW29533, VB.Ob.18037.INW29
536, VB.Ob.18040.INW29539, VB.Ob.18046.INW29545, VB.O-
b.18052.INW29551, VB.Ob.18053.INW29552, VB.Ob.18055.
INW29554, VB.Ob.18056.INW29555, VB.Ob.18059.INW29558,
VB.Ob.18061.INW29560, VB.Ob.18062.INW29561, VB.Ob.180
67.INW29566, VB.Ob.18069.INW29568, VB.Ob.18081.INW29
580, VB.Ob.18093.INW29592, VB.Ob.18101.INW29600, VB.O-
b.18112.INW29611, VB.Ob.18136.INW29635, VB.Ob.18266.
INW29767, VB.Ob.21620.INW32492, VB.Ob.21621.INW32493,
VB.Ob.21688.INW32567, VB.Ob.21689.INW32568, VB.Ob.216
90.INW32569, VB.Ob.21691.INW32570, VB.Ob.21696.INW32
575, VB.Ob.21703.INW32582, VB.Ob.23237.INW33646, VB.O-
b.23386.INW33800, VB.Ob.23387.INW33801
Froyd CA, Willis KJ (2008) Emerging issues in biodiversity &
conservation management: the need for a palaeoecological
perspective. Quaternary Science Reviews 27:1723–1732
Fuchs MA (2001) Towards a Recovery Strategy for Garry Oak and
Associated Ecosystems in Canada: Ecological Assessment and
Literature Review. Technical Report GBEI/EC-00-030. Environ-
ment Canada, Canadian Wildlife Service, Pacific and Yukon Region
Graham CH, VanDerWal J, Phillips SJ, Moritz C, Williams SE (2010)
Dynamic refugia and species persistence: tracking spatial shifts
in habitat through time. Ecography 33:1062–1069
Hamann A, Wang T (2006) Potential effects of climate change on
ecosystem and tree species distribution in British Columbia.
Ecology 87:2773–2786
Hannah L, Midgley G, Hughes G, Bomhard B (2005) The view from
the Cape: extinction risk, protected areas, and climate change.
Bioscience 55:231–242
Harrington CA (2003) The 1930s survey of forest resources in
Washington and Oregon. General Technical Report, PNW-GTR-
584. Portland, OR. US Department of Agriculture, Forest Service,
Pacific Northwest Research Station. 123 p. [plus CD-ROM]
Heikkinen RK, Luoto M, Araujo MB, Virkkala R, Thuiller W, Sykes
MT (2006) Methods and uncertainties in bioclimatic envelope
modelling under climate change. Progress in Physical Geogra-
phy 30:751–777
Klinkenberg B (2004) E-Flora BC: Electronic Atlas of the plants of
British Columbia [www.eflora.bc.ca]. Lab for Advanced Spatial
Analysis, Department of Geography, University of British
Columbia, Vancouver
Lawler JJ, White D, Neilson RP, Blaustein AR (2006) Predicting
climate-induced range shifts: model differences and model
reliability. Global Change Biology 12:1568–1584
814 Environmental Management (2012) 49:802–815
123
Author's personal copy

Lea T (2007) Historical Garry oak ecosystems of Vancouver Island,
British Columbia, pre-European contact to the present. Davidso-
nia 17:34–50
Liaw A, Wiener M (2002) Classification and regression by random
forest. Rnews 2:18–22
MacDougall AS, Duwyn A, Jones NT (2010) Consumer-based
limitations drive oak recruitment failure. Ecology 9:2092–2099
Midgley GF, Hannah L, Millar D, Rutherford MC, Powrie LW (2002)
Assessing the vulnerability of species richness to anthropogenic
climate change in a biodiversity hotspot. Global Ecology &
Biogeography 11:445–451
Midgley GF, Hannah L, Millar D, Thuiller W, Booth A (2003)
Developing regional and species-level assessments of climate
change impacts on biodiversity in the Cape Floristic Region.
Biological Conservation 112:87–97
Pellatt MG (2002) The role of paleoecology in understanding
ecological integrity: an example from a highly fragmented
landscape in the Strait of Georgia Lowlands. In: Bondrop-
Nielson S, Munro N, Nelson G, Willison JHM, Herman T,
Eagles P (eds) Managing protected areas in a changing world,
Proceedings of the fourth international conference on science
and management of protected areas. SAMPAA, Wolfville,
pp 384–397
Pellatt MG, Hebda RJ, Mathewes RW (2001) High-resolution Holo-
cene vegetation history and climate from Hole 1034B, ODP leg
169S, Saanich Inlet, Canada. Marine Geology 174:211–226
Pellatt MG, Gedalof Z, McCoy M, Bodtker K, Canon A, Smith S,
Beckwith B, Mathewes RW, Smith D (2007) Fire History and
Ecology of Garry Oak and Associated Ecosystems in British
Columbia. Final Report for the Interdepartmental Recovery Fund
Project 733. WNSC Publication. Vancouver, BC
R Development Core Team (2011) R: A language and environment for
statistical computing. R Foundation for Statistical Computing,
Vienna, Austria. ISBN 3-900051-07-0, http://www.R-project.org
Rehfeldt GE, Ferguson DE, Crookston NL (2009) Aspen, climate, and
sudden decline in western USA. Forest Ecology and Manage-
ment 258:2353–2364
Rose NA, Burton PJ (2009) Using bioclimate envelopes to identify
temporal corridors in support of conservation planning in a
changing climate. Forest Ecology and Management 258S:S64–
S74
Salathe EP Jr, Motea PW, Wiley MW (2007) Review of scenario
selection and downscaling methods for the assessment of climate
change impacts on hydrology in the United States pacific
northwest. International Journal of Climatology 27:1611–1621
Shafer SL, Bartlein PJ, Thompson RS (2001) Potential changes in the
distributions of western North America tree and shrub taxa under
future climate scenarios. Ecosystems 4:200–215
Strobl C, Boulesteix A, Zeileis A, Hothorn T (2007) Bias in random
forest variable importance measures: illustrations, sources and a
solution. BMC Bioinformatics 8:25
Vellend M, Bjorkman AD, McConchie A (2008) Environmentally
biased fragmentation of oak savanna habitat on southeastern
Vancouver Island, Canada. Biological Conservation 141:
2576–2584
Wang T, Hamann A, Spittlehouse DL, Aitken SN (2006) Develop-
ment of scale-free climate data for Western Canada for use in
resource management. International Journal of Climatology
26:383–397
Williams JW, Jackson ST, Kutzbach JE (2007) Projected distributions
of novel and disappearing climates by 2100 AD. Proceedings of
the National Academy of Sciences 104:5738–5742
Willmott CJ, Robeson SM (1995) Climatologically aided interpola-
tion (CAI) of terrestrial air temperature. International Journal of
Climatology 15:221–229
Environmental Management (2012) 49:802–815 815
123
Author's personal copy
Related Documents











