This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Use of Enterococcus faecium to improve common sole (Solea solea) larviculture
Matteo A. Avella a, Ike Olivotto a, Stefania Silvi b, Cataldo Ribecco a, Alberto Cresci b, Francesco Palermo b,Alberta Polzonetti b, Oliana Carnevali a,⁎a Department of Marine Sciences, Polytechnic University of Marche, Ancona, Italyb School of Biosciences and Biotechnologies, University of Camerino, Camerino, Italy
a b s t r a c ta r t i c l e i n f o
Article history:Received 22 April 2010Received in revised form 25 February 2011Accepted 28 February 2011Available online 8 March 2011
Keywords:ProbioticMyostatinGrowthAnimal welfareLarvalBacterial
The potential of common sole Solea solea (Linnaeus, 1758) for consistent production in recirculating systemsis limited by the huge losses usually observed during the early stages of larval development, mainly due topathogen infections and insufficient nutritional balance. In the hope that use of a suitable probiotic strain maycounter these difficulties, in this study we isolated Enterococcus faecium IMC 511 from common sole broodstock and utilized it as probiotic candidate during fish feeding. After a daily administration (twice a day) tosole larvae through live feeds, the intestinal presence of E. faecium IMC 511 was estimated on days 10, 30, and50 post hatch, and the effects on intestinal microbial load, fish survival, welfare, and growth were evaluatedby morphometric and molecular approaches by examining relative body weight, total length, and geneexpression of myostatin. In particular, the effect on animal welfare was assessed through analyses of cortisollevels and 70 kDa Heat Shock Protein gene expression.The results showed that while the Vibrio populations of the intestine of S. solea larvae were reduced in asignificant way by the probiotic E. faecium IMC 511, survival was not affected. 70 kDa Heat Shock Protein andmyostatin gene expression were significantly reduced in probiotic fed larvae with respect to the controlgroup. It is of note that E. faecium IMC 511 seemed to be predominant at day 50 post hatching, probably due tostabilization of the intestinal microbiota, a factor that, together with the lower levels of HSP70, could improveanimal well-being and growth.Cortisol levels significantly increased after 10 days of treatment in probiotic fed larvae, while on days 30 and50 no significant differences were observed among all experimental groups. Considered together, themicrobiological data and the molecular and morphometric results suggest that 50 days of probiotic treatmentcan improve common sole larval growth.Also, it is hoped that these results offer a useful starting point for elucidating the pathways involved in S. soleagrowth fostered by E. faecium IMC 511 administration.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
While the finfish aquaculture industry has developed broadly inEurope during the last two decades, this considerable growth hasfocused on just a few species. Scientists and farmers have been mainlyinterested in remunerative species easily reproduced in captivity usingtechniques already available in aquaculture. Among new marinespecies, the common sole Solea solea (Linnaeus, 1758) can beconsidered a valuable candidate for consistent production in recirculat-ing systems. In fact, commercial rearing of common sole and closelyrelated species (Senegalese sole, Solea senegalensis) recently became ofgreat interest among aquaculturists (Overton et al., 2010) due to thehigh value and increasedmarket demand (Howell, 1997; Imsland et al.,2003). Thus different studies with different approaches have been
performed in order to increase quality and productivity of sole species(Lund et al., 2008; Blonk et al., 2010a,b; Lund et al., 2010; Overton et al.,2010). Moreover, some studies dealing with the application of differentprobiotic strains on Senegalese sole have provided encouraging results(de la Banda et al., 2010; Díaz-Rosales et al., 2009;Makridis et al., 2008)concerning growth and survival against pathogens, all of which makethe identification and application of new probiotic strains to the solelarviculture worthwhile. Several different approaches have been usedin order to estimate the beneficial effects of different probiotic strains;some examples are testing feed efficiency, activities of leucineaminopeptidase and alkaline phosphatase and structural changes inthe intestine (Makridis et al., 2008), checking respiratory burst activityof phagocytes, growth and in vivo challenge with pathogens (Díaz-Rosales et al., 2009), or evaluating muscle and liver body compositionand histology of digestive tract and liver (de la Banda et al., 2010).
In order to further maximise prospects for production of captivecommon sole, scientific research needs to focus on how to amelioratethe consistent losses in early stages of larval development (Imsland
Aquaculture 315 (2011) 384–393
⁎ Corresponding author. Tel.: +39 071 2204990; fax: +39 071 2204650.E-mail address: [email protected] (O. Carnevali).
0044-8486/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.aquaculture.2011.02.046
Contents lists available at ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r.com/ locate /aqua-on l ine

Author's personal copy
et al., 2003) caused by pathogen infections and insufficient nutritionalbalance. Given that not even live prey enrichment consistentlyimproves survival (Howell, 1997), the search for alternative dietshas turned to the use of probiotics during the early larval stages.Several studies have reported encouraging results in the application ofprobiotics in aquaculture (Vine et al., 2006; Wang et al., 2008; Swainet al., 2009; Merrifield et al., 2010), and application of probiotics tofish larviculture in particular has yielded to positive effects, mainly insurvival and growth rates (Ringo and Gatesoupe, 1998; Gatesoupe,1999, 2008; Carnevali et al., 2004, 2006; Wang et al., 2008; Avella etal., 2010a,b). In the search for specific probiotic strains that improvefish welfare, the obvious first step for researchers is to determine thatthey pose no danger to fish health; furthermore, the strain should beable to significantly limit the growth of fish pathogens that harmlarval development, enhance the fish larval immune system, andpositively affect fish growth.
On this regard, inadequate aquaculture conditions can be occa-sionally detrimental for fish welfare (Maurilio et al., 2003; Vine et al.,2006; Carnevali et al., 2004, 2006), harming in those instanceslarviculture productiveness; moreover, such conditions yield verypoor quality seafood. Scientists seek to create the best breeding andrearing conditions for producing seafood of a quality comparable tothat of wild caught specimens, and central to this effort is theassessment of larval nutrition needs and development of adequatelarval feeding protocols. In this study, we assessed the usefulness of aprobiotic species isolated from common sole brood stock, the lacticacid bacteria (LAB) E. faecium IMC 511. E. faecium has been recentlyreported as a species exhibiting remarkable probiotic profiles(Hosseini et al., 2009). In this instance, E. faecium IMC 511 potentialprobiotic strain was supplied to sole at different time from hatchingand at different concentrations. Afterwards, upon administration, itseffects on larval sole survival, welfare, and growth were evaluated bymolecular and morphometric approaches. Cortisol levels and quan-titative HSP70 gene expression were assessed in order to evaluate theeffects of E. faecium IMC 511 on common sole larvae well being.Indeed, from amolecular point of view, several factors aremodulated inresponse to probiotics in fish (Suzer et al., 2008; Avella et al., 2010a,b).Cortisol levels areuseful indicators for stress,which cannegatively affectgrowth (Mommsen et al., 1999). In response to stressful rearingconditions, the hypothalamus–pituitary–interrenal (HPI) axisis activated, and the release of pro-opiomelanocortin (POMC)-derivedpeptides from the pituitary gland in turn leads to cortisol release fromthe corticosteroid-producing cells of the head kidney (WendelaarBonga, 1997).
We also assessed HSP70 gene expression because it has been foundto be related to stressful conditions occasionally occurring inaquaculture, such as temperature oscillations, crowding, insufficientwater quality, and improper diet (Barry et al., 1995; Rollo et al., 2006).
Concerning growth, it is well known that fish muscle growth isregulated by a complex set of endo-para-autocrine factors. In fish,insulin-like growth factors (IGFs) and myostatin play a key role ingrowth regulation. The IGF system includes two highly conservedprimary ligands, IGF-I and IGF-II, two trans membrane receptors, andfour binding proteins (Moriyama et al., 2000). The action of the IGFsystem is contrasted by myostatin (MSTN), a member of the TGF-βsuperfamily able to inhibit myoblast proliferation (McPherron et al.,1997). In this study, we used MSTN quantitative gene expression as amarker to check larval muscle growth upon probiotic treatment.While in mammals, cortisol was seen to increase MSTN levels (Maet al., 2001), in fish models the relation between cortisol and MSTN isstill contradictory and seems to depend on fish species (Funkensteinet al., 2009). Thus, in this study, we also investigated the relationbetween cortisol and MSTN gene expression in sole larvae with orwithout probiotic treatment.
To summarize, we sought to determine from the molecular andmorphometric points of view whether E. faecium IMC 511 positively
affects common sole larval growth. In particular, we focused on themolecular changes occurring upon probiotic delivery and the possiblecorrelation among HSP70, MSTN gene expression, cortisol levels, andfish larval growth.
Of note was the lack of available S. solea gene sequences, and theconsequent difficulty in carrying out gene expression studies on thismodel. This study may represent also an initial step in order to obtainsuch important information.
2. Materials and methods
2.1. Larval rearing
Larvae were raised at 18–20 °C in conical-based cylindrical tanks,the water in the 100 l capacity tank was gently replaced twice an hourby a dripping system. Until 28 ph, their diet was exclusively based onlive food, consisting of rotifers enriched with ALGAMAC 3000, Artemianauplii and Artemia metanauplii enriched with ALGAMAC 3000. Thephytoplankton Nannochloropsis oculata was used (50,000 cells/ml) tocondition the tank. Larvae were subjected to a photoperiod of 16 hlight/8 h dark.
2.2. Experimental design
The experimental phase was designed to study the effects onintestine microbial load, survival, stress response, and growth whentwo different concentrations of probiotic E. faecium IMC 511 weresupplied twice a day in the rearing water, starting from the first day ofrotifer feeding (day 2 post hatch — ph) or from the first day of co-feeding (rotifers/Artemia salina nauplia — day 11 ph), and assessedafter 10, 30 and 50 days of treatment. Probiotic suspension wasprepared and administered twice a day to larvae as previouslyreported by Carnevali et al. (2004).
Five experimental groups (2 tanks each) with 500 larvae eachwere fed as follows:
Group 1 Control: standard feeding regime;Group 2: E. faecium IMC 511 supplied from the mouth opening day
(2 ph) at the final concentration of 103colony-forming unit(CFU)/ml;
Group 3: E. faecium IMC 511 supplied from the first day of co-feeding(11 ph) at the final concentration of 103CFU/ml;
Group 4: E. faecium IMC 511 supplied from the mouth opening day(2 ph) at the final concentration of 105CFU/ml;
Group 5: E. faecium IMC 511 supplied from the first day of co-feeding(11 ph) at the final concentration of 105CFU/ml.
At days 10, 30, and 50 ph, 10 larvae or juvenileswere collected fromeach tank for gut microbial analysis and the evaluation of body weight(BW), total length (TL), and biomarker expressions by quantitativepolymerase chain reaction (qPCR). Same number of larvae/juvenileswas pooled from each tank for cortisol level determination.
2.3. Zooplankton cultures
Different species of zooplankton were cultured in order to feedcommon sole larvae during treatments.
Two different rotifers species, Brachionus plicatilis and B. rotundi-formis, with an average size of 239 and 160 μm respectively, werecultured on N. oculata at 30 ppm salinity and 22 °C. Super smallArtemia sp. nauplii (INVE Technologies, Belgium) were introduced inthe larval tank from day 9. All the zooplankton species were enrichedusing Algamac 2000 (Aquafauna Bio-Marine, Inc., USA) following theinstructions provided by the company (0.5 g/million rotifers; at 25 °Cfor 8–12 h).
385M.A. Avella et al. / Aquaculture 315 (2011) 384–393

Author's personal copy
2.4. Probiotic strain
The autochthonous probiotic strain used was E. faecium IMC 511,isolated from adult common sole gut and deposited in the culturecollection Deutsche Sammlung vonMikrooganismen und Zellkulturen(DSMZ, Germany) under the number DSMZ 17263.
The whole intestinal content of healthy adults of S. solea (bodyweight of about 600 g each) was collected after fish transfer to thelaboratory and sacrifice and after cutting the upper intestinal endunder the stomach. The whole intestine was squeezed from the upperend to the anus and the content was collected in sterile vials andcentrifuged at 12,000×g (A-12 Microcentrifugette, rotor 6642, ALC,Milan Italy) for 10 min. The pellet was suspended in 10 ml of MRS(de Man, Rogosa and Sharpe) broth (OXOID, Unipath Limited,Basingstoke, UK) with 2% NaCl and incubated for 5 days at 25±1 °Cin order to achieve growth and proliferation of all LAB strains. Thesestrains were then isolated on MRS agar plates (OXOID) afterpreparation of ten-fold dilutions in sterile saline solution (2% NaCl)of the culture. The plates were incubated for 48–72 h (h) at 25±1 °Cat aerobic conditions. The colonies were counted and identified byGram staining and microscopy observation of cell morphology and byenzymatic tests. Preliminary identification of the ovoid-shaped, non-sporing, gram positive cells was done by the API 20 STREP system(bioMérieux, Marcy l'Etoile, France).
Among LAB isolated from adult common sole gut, the E. faeciumIMC 511 was chosen as the strain to administer during common solelarvae rearing since it was the most represented species of LAB in theintestine of S. solea (22%) and it showed probiotic properties in in vitroassay (unpublished data). It also showed a great ability to survive inaerobiosis at 17–20 °C and the ability to thrive in typical tankconditions.
Molecular characterization of the E. faecium IMC 511 strain wasdone by amplification and sequencing of 16S rDNA using universalprimers P0 and P6 corresponding to positions 27f (forward) and 1495r(reverse) of Escherichia coli 16S rDNA. The strain was culturedaerobically at 37 °C in MRS agar. One colony was suspended in steriledistilled water and utilized for DNA extraction using DNeasy Tissue kits(QIAGEN S.p.A., Milan, Italy). 0.4 μl of DNA extracted was added to19.6 μl of reaction mix containing 0.8 μl distilled water, 18 μl PCRSuperMix (Invitrogen srl, Milan, Italy), 0.4 μl of P0, and P6 (500 ng/μl)(Invitrogen). The reaction mixtures, after incubation at 94 °C for 90 s,were cycled 35 times through the following temperature profile: 94 °Cfor 30 s, 55 °C for 30 s, and 72 °C for 120 s. Finally, the reactions wereincubated at 72 °C for 10 min and at 60 °C for 10 min. The reactionof PCR was conducted in a Tpersonal Thermal Cycler (Biometra,Göttingen, Germany). The PCR products were analyzed by electropho-resis in 2% agarose gel containing 0.5 μg/ml (w/v) of ethidium bromide(GIBCO BRL, Gaithersburg, USA). The PCR products were purified byQIAquick PCR purification kit (Qiagen) and then sequenced. Thesequences obtained were compared to sequence information availableon Genbank database (www.ncbi.nlm.nih.gov/Web/Genbank/index.html) using the BLAST algorithm.
2.5. Microbial analyses
At each sampling 10 larvae were surface-disinfected withbenzalkonium chloride (0.1% w/v) for 30 s, rinsed three times inautoclaved water and homogenized in 3 ml of reducing solution(Carnevali et al., 2004). The homogenate of pooled animals wasobtained using a Stomacher Lab blender (Seward Medical, UK) andwas serially diluted in reducing solution and plated in duplicate. Thetotal counts of aerobic and anaerobic bacteria were performed onMarine agar plates (Difco-Becton, Dickinson and Company, Sparks,USA). MacConkey agar plates (Oxoid) were used to obtain viablecounts of Enterobacteriaceae as reported by Rollo et al. (2006). At thesame time Lactic Acid Bacteria (LAB) counts were conducted on MRS
agar plates, Carnobacterium spp. counts on MRS agar with pHcorrected to 9 (Holzapfel, 1992), and Vibrio spp. counts on ThiosulfateCitrate Bile Sucrose (TCBS) Cholera agar (Oxoid). To all media, withthe exception of Marine agar, 1.5% NaCl was added. The plates wereincubated in aerobic conditions for 48–72 h at 17–20 °C and inanaerobic conditions at the same temperatures inside an anaerobiccabinet (Don Whitley Scientific, Shipley, UK).
Twenty colonies, randomly selected from countable MRS agarplates, were isolated and checked for purity. DNA was extracted usingthe Qiagen Dneasy Tissue kit and analysed using the RAPD technique.RAPD was performed with the random primer M13 minisatellite coresequence (5′-GAGGGTGGCGGTTCT-3′). Reactions were carried out in25 μl amplification mixtures with 12.5 μl of 2× Master Mix (Fermentas,Burlington, Canada), 0.5 μl of primer, 1 μl of total DNA and 11 μl ofwater. The reactionmixtureswithM13 primer, after incubation at 94 °Cfor 2 min, were cycled through the following temperature profile: 30cycles at 94 °C for 60 s, 42 °C for 20 s and 72 °C for 120 s. Final extensionwas carried out at 72 °C for 10 min. The PCR was conducted in aTpersonal Thermal Cycler (Biometra). Amplification products wereanalyzed on a 2% agarose gel, containing 0.5 μg/ml (w/v) of ethidiumbromide (GIBCO BRL).
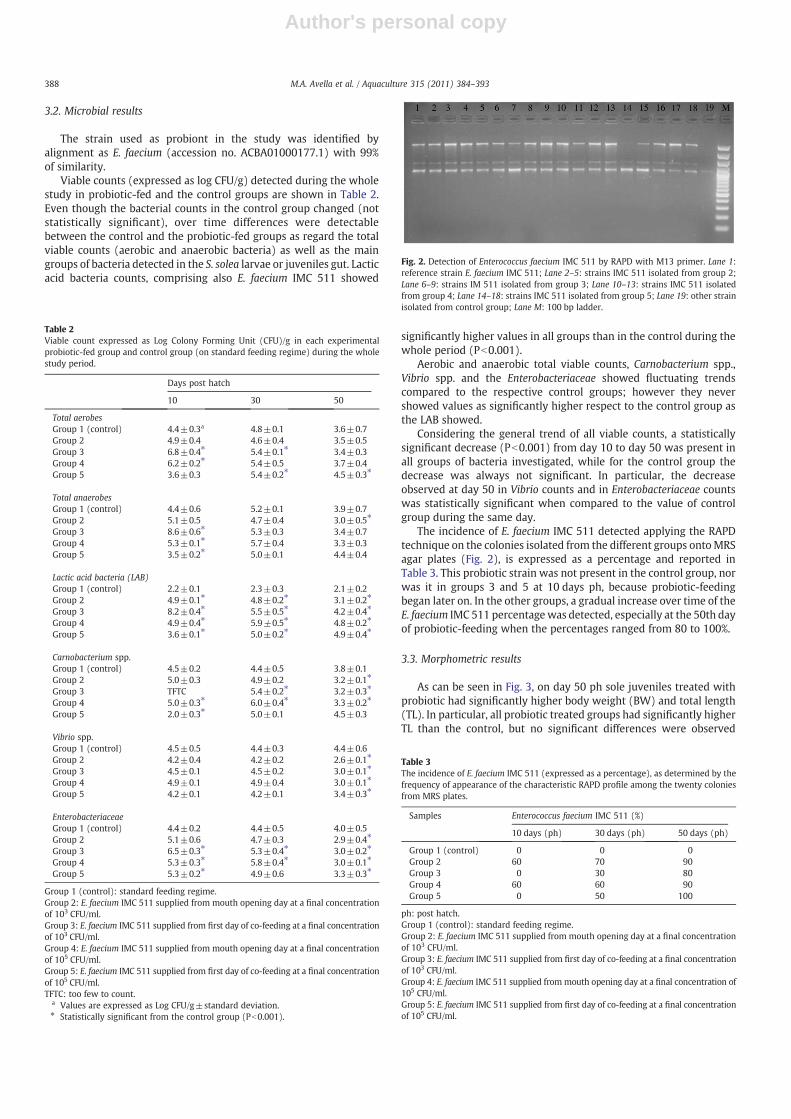
This RAPDmethod based on the use of M13 primer was previouslytested for its reproducibility on the E. faecium IMC 511 demonstratingidentical fingerprint profiles after multiple analysis, moreover RAPDfingerprinting was also able to differentiate genetically unique straintype within very closely related species and species present in solemicrobiota (Fig. 1).
The incidence of E. faecium IMC 511, as determined by thefrequency of appearance of the characteristic RAPD profile amongthe twenty isolated colonies from MRS plates, was expressed as apercentage.
2.6. Morphometric analysis and survival rate
On days 10, 30, and 50 ph, 10 larvae or juveniles were sampled fromeach tank and anesthetized using MS222 (Sigma Aldrich) (1 mg/l) andtotal length (TL) and body weight (BW) were recorded using a Stemi2000 micrometric microscope and OHAUS Explorer E11140 microbal-ance accurate to 0.1 mg. Finally, on day 50 ph, for each experimentalgroup (from each tank), the total numbers of remaining juveniles werecounted and survival rate was calculated.
Fig. 1. RAPD fingerprints of bacterial strain isolated from sole intestine and ofEnterococcus faecium IMC 511 using M13 primer. Lane 0: 100 bp ladder; Lane 1:Pasterella pneumotropica; lane 2: P. pneumotropica; lane 3: P. pneumotropica; lane 4:Vibro vulnificus; lane 5: P. pneumotropica; lane 6: Aeromonas salmonicida, lane 7: V.vulnificus; lane 8: A. salmonicida; lane 9: P. pneumotropica; lane 10: Enterococcus faeciumIMC 511; lane 11: negative control.
386 M.A. Avella et al. / Aquaculture 315 (2011) 384–393

Author's personal copy
2.7. Quantitative Real time PCR analysis
2.7.1. RNA extraction and cDNA synthesisTotal RNA extraction from whole larval body was optimized, the
best recovery from pools (30 mg) of whole larvae using RNAeasy®(Qiagen) extraction kit following the manufacturer's protocol. TotalRNA extracted was eluted in 15 μl of RNAase-free water. Final RNAconcentrations were determined by NanoDrop ND 1000 Spectropho-tometer from NanoDrop Technologies, and the extracted RNAintegrity was verified by ethidium bromide staining of 28S and 18Sribosomal RNA bands on 1% agarose gel. RNA was stored at −80 °Cuntil use.
Total RNAwas treated with DNAase (10 UI at 37 °C for 10 min, MBIFermentas). A total amount of 5 μg of RNA was used for cDNAsynthesis, employing 0.5 μg oligo d(T)+adapter primer, 5′-GACTG-CAGTCGACATCGATTTTTTTTTTTTTTTTTT-3′, in a buffer containing50 mM Tris–HCl (pH 8.3), 75 mM KCl, 3 mM MgCl2, 10 mM DTT,0.5 mM of each dNTP, 40 units RNAase OUT (Invitrogen) and200 units of Superscript II RT (Invitrogen, Life technologies, Milan,Italy). Cycling conditions were: 70 °C for 5 min, 42 °C for 52 min and72 °C for 15 min.
2.7.2. Primer designSequenced regions containing the aforementioned markers for
several species closely related to common sole as well as to outgroupspecies were aligned and the consensus sequences were used todesign primers for qPCR (Table 1). Primer sequences for AcidicRibosomal Phosphoprotein (ARP), MSTN1 and HSP70were used at thefinal concentration of 10 pmol/μl; the percentage identity scores ofthe obtained gene fragments were calculated using NCBI BLAST(www.ncbi.nlm.nih.gov/BLAST/).
2.7.3. qPCRTriplicate PCR reactions were carried out for each sample analysed.
After optimisation of Real time conditions, the qPCRs were performedwith the SYBR green method in an iQ5 multicolor Real time PCRDetection system (Bio-Rad). The reactions were set on a 96-well plateby mixing, for each sample, 1 μl of diluted (1/20) cDNA, 5 μl of 2×concentrated SYBR Green PCR Master Mix (Bio-Rad), containing SYBRGreen as fluorescent intercalating agent, 0.3 μM forward primer and0.3 μM of reverse primer. The thermal profile for all reactions was15 min at 95 °C and then 45 cycles of 20 s at 95 °C, 20 s at 60 °C, and20 s at 72 °C. The fluorescence monitoring occurred at the end of eachcycle. Additional dissociation curve analysis was performed and in allcases showed one single peak.
2.7.4. Quantification of cDNAA relative quantification of cDNAwasmade using Acidic Ribosomal
Phosphoprotein (ARP) as reference gene (housekeeping gene). Pfaffl's
mathematical model (2001) was applied to determine the ratiobetween the different expressions of the aforementioned target genesin Group 2, Group 3, Group 4, Group 5 and Control and the differentexpressions of standard gene (ARP) in Group 2, Group 3, Group 4,Group 5 and Control. The following equation was used to express therelative expression ratio:
ratio =Etarget
� �ΔCP target control−sampleð Þ
Estandardð ÞΔCP standard control−sampleð Þ
where Etarget is the Real time PCR efficiency of target gene transcriptHSP70, MSTN; Estandard is the Real time PCR efficiency of the standardgene transcript (ARP); ΔCPtarget is the Crossing Point deviation ofControl — sample of the target gene transcript; ΔCPstandard is theCrossing Point deviation of Control — sample of standard genetranscript. Modifications of gene expression are represented withrespect to Control, which is assumed to have the value of 1 A.U.(arbitrary unit). Using the Pfaffl's mathematical model, the cDNAlevels of three genes were quantified in each group and at each time ofsampling. To test Real time PCR efficiency, serial dilutions of cDNA,each in triplicate, were amplified by Real time PCR using the specificprimers for target genes (Table 1) and housekeeping gene. Log cDNAdilution was plotted versus ΔCP and the efficiencies were calculatedaccording to E=N10[−1/slope] (Rasmussen, 2001).
2.8. Cortisol analysis
Cortisol extraction was performed in whole-body larvae andjuveniles. Samples from the different groups and at different times ofgrowth were processed following the procedures described byCarnevali et al. (2006).
2.9. Statistical analysis
Results were expressed as the mean±s.d. The significance ofdifferences was determined using a one-way or two-way ANOVA,followed by Tukey's or Bonferroni's test for multigroup comparisons,with a statistical software package, SigmaStat 3.1 (Systat softwareInc.). A probability of 0.05 was utilized to account for the statisticaldifference between the means.
3. Results
3.1. Survival rate
At day 50 ph, survival was not significantly different betweengroups and appeared to conform to normal standards (Survival%:Control 29±2; Group 2: 28±2; Group 3: 30±1; Group 4: 28±1; andGroup 5: 29±1).
Table 1Primer sequences, Genbank access number, % identity, ingroup and outgroup fish species.
Gene Primers sequences Genbankaccession #
Identity Fish sequence Genbankaccession #
Outgroup species Genbankaccession #
Acidic RibosomalPhosphoprotein (ARP)
F 5′-CTGAACATCTCGCCCTTCTC-3′ GU474637 85% withDanio rerio
Danio rerio (BC146738) Rattus norvegicus (BC062028)R 5′-TAGCCGATCTGCAGACACAC-3′ Anguilla Anguilla
(AY763793)70 KDa Heat shock Protein(HSP70)
F 5′-ACGGAGAGTCGATTTCGATG-3′ GU474638 92% with Paralichthysolivaceus
Platichthys flesus(AF187726)
Mus musculus (BC106169)
Paralichthys olivaceus(AF053059)
R 5′-GAAGGACATCAGCGACAACA-3′ Scophthalmus maximus (EU099574)Myostatin (MSTN) F 5′-GGCCTGGACTGTGATGAGAA-3′ GU474639 96% with Paralichthys
adspersusParalichthys adspersus (EU443627) Sus scrofa (AF019623)Scophthalmus maximus (EF683115)
R 5′-GCATGTTGATGGGTGACATC-3′ Dicentrarchus labrax (AY839106)
387M.A. Avella et al. / Aquaculture 315 (2011) 384–393
Author's personal copy
3.2. Microbial results
The strain used as probiont in the study was identified byalignment as E. faecium (accession no. ACBA01000177.1) with 99%of similarity.
Viable counts (expressed as log CFU/g) detected during the wholestudy in probiotic-fed and the control groups are shown in Table 2.Even though the bacterial counts in the control group changed (notstatistically significant), over time differences were detectablebetween the control and the probiotic-fed groups as regard the totalviable counts (aerobic and anaerobic bacteria) as well as the maingroups of bacteria detected in the S. solea larvae or juveniles gut. Lacticacid bacteria counts, comprising also E. faecium IMC 511 showed
significantly higher values in all groups than in the control during thewhole period (Pb0.001).
Aerobic and anaerobic total viable counts, Carnobacterium spp.,Vibrio spp. and the Enterobacteriaceae showed fluctuating trendscompared to the respective control groups; however they nevershowed values as significantly higher respect to the control group asthe LAB showed.
Considering the general trend of all viable counts, a statisticallysignificant decrease (Pb0.001) from day 10 to day 50 was present inall groups of bacteria investigated, while for the control group thedecrease was always not significant. In particular, the decreaseobserved at day 50 in Vibrio counts and in Enterobacteriaceae countswas statistically significant when compared to the value of controlgroup during the same day.
The incidence of E. faecium IMC 511 detected applying the RAPDtechnique on the colonies isolated from the different groups ontoMRSagar plates (Fig. 2), is expressed as a percentage and reported inTable 3. This probiotic strain was not present in the control group, norwas it in groups 3 and 5 at 10 days ph, because probiotic-feedingbegan later on. In the other groups, a gradual increase over time of theE. faecium IMC 511 percentagewas detected, especially at the 50th dayof probiotic-feeding when the percentages ranged from 80 to 100%.
3.3. Morphometric results
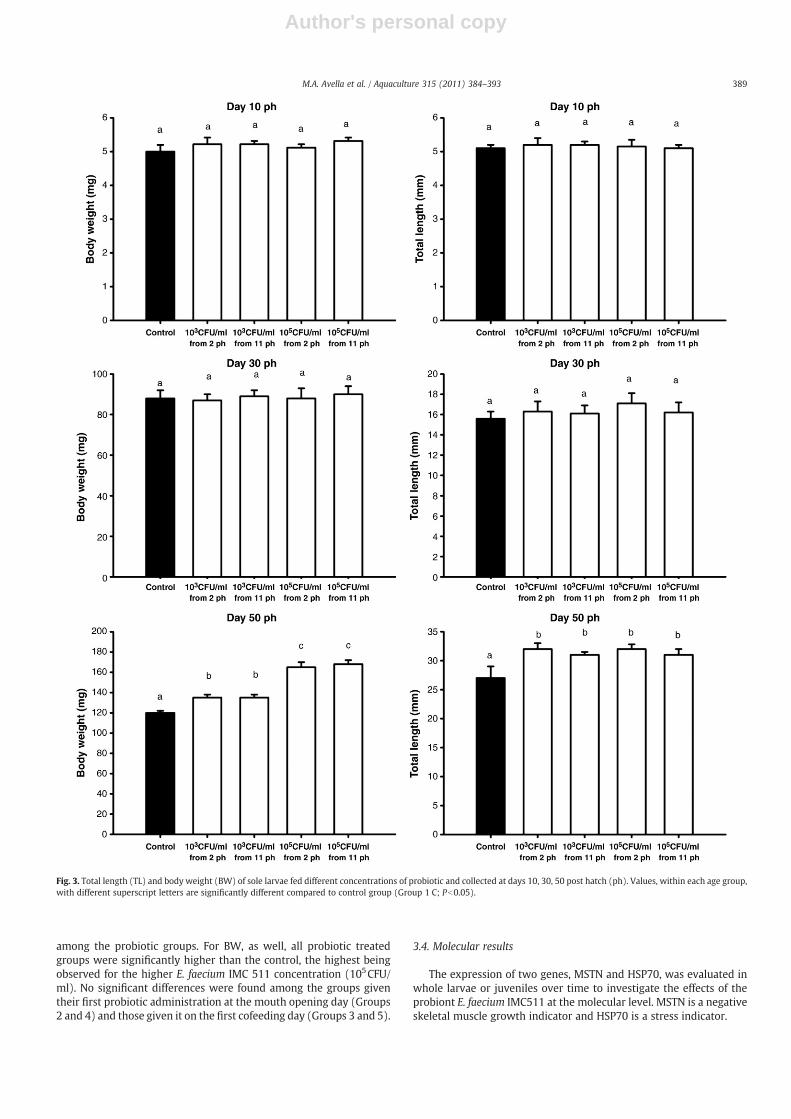
As can be seen in Fig. 3, on day 50 ph sole juveniles treated withprobiotic had significantly higher body weight (BW) and total length(TL). In particular, all probiotic treated groups had significantly higherTL than the control, but no significant differences were observed
Table 2Viable count expressed as Log Colony Forming Unit (CFU)/g in each experimentalprobiotic-fed group and control group (on standard feeding regime) during the wholestudy period.
Days post hatch
10 30 50
Total aerobesGroup 1 (control) 4.4±0.3a 4.8±0.1 3.6±0.7Group 2 4.9±0.4 4.6±0.4 3.5±0.5Group 3 6.8±0.4⁎ 5.4±0.1⁎ 3.4±0.3Group 4 6.2±0.2⁎ 5.4±0.5 3.7±0.4Group 5 3.6±0.3 5.4±0.2⁎ 4.5±0.3⁎
Total anaerobesGroup 1 (control) 4.4±0.6 5.2±0.1 3.9±0.7Group 2 5.1±0.5 4.7±0.4 3.0±0.5⁎
Group 3 8.6±0.6⁎ 5.3±0.3 3.4±0.7Group 4 5.3±0.1⁎ 5.7±0.4 3.3±0.3Group 5 3.5±0.2⁎ 5.0±0.1 4.4±0.4
Lactic acid bacteria (LAB)Group 1 (control) 2.2±0.1 2.3±0.3 2.1±0.2Group 2 4.9±0.1⁎ 4.8±0.2⁎ 3.1±0.2⁎
Group 3 8.2±0.4⁎ 5.5±0.5⁎ 4.2±0.4⁎
Group 4 4.9±0.4⁎ 5.9±0.5⁎ 4.8±0.2⁎
Group 5 3.6±0.1⁎ 5.0±0.2⁎ 4.9±0.4⁎
Carnobacterium spp.Group 1 (control) 4.5±0.2 4.4±0.5 3.8±0.1Group 2 5.0±0.3 4.9±0.2 3.2±0.1⁎
Group 3 TFTC 5.4±0.2⁎ 3.2±0.3⁎
Group 4 5.0±0.3⁎ 6.0±0.4⁎ 3.3±0.2⁎
Group 5 2.0±0.3⁎ 5.0±0.1 4.5±0.3
Vibrio spp.Group 1 (control) 4.5±0.5 4.4±0.3 4.4±0.6Group 2 4.2±0.4 4.2±0.2 2.6±0.1⁎
Group 3 4.5±0.1 4.5±0.2 3.0±0.1⁎
Group 4 4.9±0.1 4.9±0.4 3.0±0.1⁎
Group 5 4.2±0.1 4.2±0.1 3.4±0.3⁎
EnterobacteriaceaeGroup 1 (control) 4.4±0.2 4.4±0.5 4.0±0.5Group 2 5.1±0.6 4.7±0.3 2.9±0.4⁎
Group 3 6.5±0.3⁎ 5.3±0.4⁎ 3.0±0.2⁎
Group 4 5.3±0.3⁎ 5.8±0.4⁎ 3.0±0.1⁎
Group 5 5.3±0.2⁎ 4.9±0.6 3.3±0.3⁎
Group 1 (control): standard feeding regime.Group 2: E. faecium IMC 511 supplied from mouth opening day at a final concentrationof 103 CFU/ml.Group 3: E. faecium IMC 511 supplied from first day of co-feeding at a final concentrationof 103 CFU/ml.Group 4: E. faecium IMC 511 supplied from mouth opening day at a final concentrationof 105 CFU/ml.Group 5: E. faecium IMC 511 supplied from first day of co-feeding at a final concentrationof 105 CFU/ml.TFTC: too few to count.
a Values are expressed as Log CFU/g±standard deviation.⁎ Statistically significant from the control group (Pb0.001).
Fig. 2. Detection of Enterococcus faecium IMC 511 by RAPD with M13 primer. Lane 1:reference strain E. faecium IMC 511; Lane 2–5: strains IMC 511 isolated from group 2;Lane 6–9: strains IM 511 isolated from group 3; Lane 10–13: strains IMC 511 isolatedfrom group 4; Lane 14–18: strains IMC 511 isolated from group 5; Lane 19: other strainisolated from control group; Lane M: 100 bp ladder.
Table 3The incidence of E. faecium IMC 511 (expressed as a percentage), as determined by thefrequency of appearance of the characteristic RAPD profile among the twenty coloniesfrom MRS plates.
Samples Enterococcus faecium IMC 511 (%)
10 days (ph) 30 days (ph) 50 days (ph)
Group 1 (control) 0 0 0Group 2 60 70 90Group 3 0 30 80Group 4 60 60 90Group 5 0 50 100
ph: post hatch.Group 1 (control): standard feeding regime.Group 2: E. faecium IMC 511 supplied from mouth opening day at a final concentrationof 103 CFU/ml.Group 3: E. faecium IMC 511 supplied from first day of co-feeding at a final concentrationof 103 CFU/ml.Group 4: E. faecium IMC 511 supplied frommouth opening day at a final concentration of105 CFU/ml.Group 5: E. faecium IMC 511 supplied from first day of co-feeding at a final concentrationof 105 CFU/ml.
388 M.A. Avella et al. / Aquaculture 315 (2011) 384–393
Author's personal copy
among the probiotic groups. For BW, as well, all probiotic treatedgroups were significantly higher than the control, the highest beingobserved for the higher E. faecium IMC 511 concentration (105CFU/ml). No significant differences were found among the groups giventheir first probiotic administration at the mouth opening day (Groups2 and 4) and those given it on the first cofeeding day (Groups 3 and 5).
3.4. Molecular results
The expression of two genes, MSTN and HSP70, was evaluated inwhole larvae or juveniles over time to investigate the effects of theprobiont E. faecium IMC511 at the molecular level. MSTN is a negativeskeletal muscle growth indicator and HSP70 is a stress indicator.
Fig. 3. Total length (TL) and body weight (BW) of sole larvae fed different concentrations of probiotic and collected at days 10, 30, 50 post hatch (ph). Values, within each age group,with different superscript letters are significantly different compared to control group (Group 1 C; Pb0.05).
389M.A. Avella et al. / Aquaculture 315 (2011) 384–393

Author's personal copy
HSP70 gene expression at day 50 ph was significantly lower in allgroups treated with probiotic compared to the control group; moreover,the lowest levels observed were related to groups 3 and 5, whichreceived E. faecium from the first day of cofeeding (Fig. 4).
In addition, at day 50, significantly lower levels of MSTN wererecorded in groups 4 and 5, which received probiotics at the con-centration of 105CFU/ml (Fig. 5).
3.5. Cortisol levels
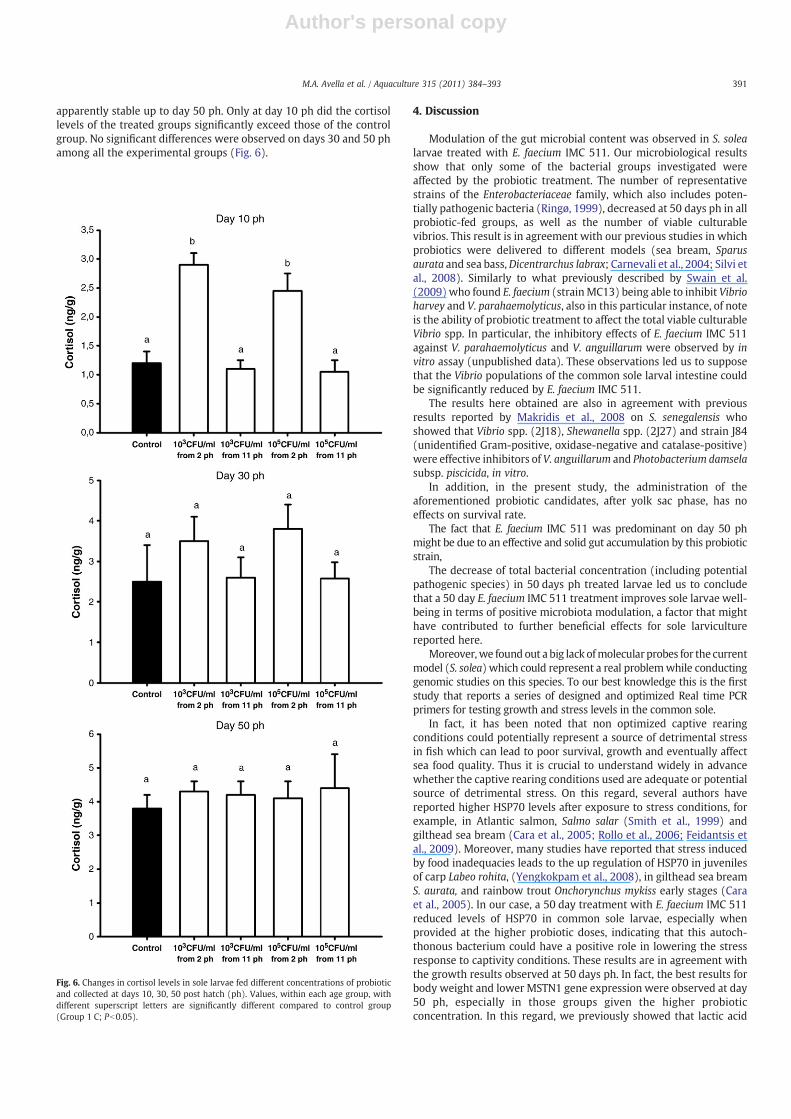
The control group had a significant increase of cortisol levels fromday 10 to day 50 ph, while in the probiotic treated groups, cortisollevels significantly increased from day 10 to 30 ph, and were
Fig. 4. Relative HSP70 gene expression in sole larvae fed different concentrations ofprobiotic and collected at days 10, 30, 50 post hatch (ph). Values, within each agegroup, with different superscript letters are significantly different compared to controlgroup (Group 1 C; Pb0.05).
Fig. 5. Relative MSTN1 gene expression in sole larvae fed different concentrations ofprobiotic and collected at days 10, 30, 50 post hatch (ph). Values, within each agegroup, with different superscript letters are significantly different compared to controlgroup (Group 1 C; Pb0.05).
390 M.A. Avella et al. / Aquaculture 315 (2011) 384–393
Author's personal copy
apparently stable up to day 50 ph. Only at day 10 ph did the cortisollevels of the treated groups significantly exceed those of the controlgroup. No significant differences were observed on days 30 and 50 phamong all the experimental groups (Fig. 6).
4. Discussion
Modulation of the gut microbial content was observed in S. solealarvae treated with E. faecium IMC 511. Our microbiological resultsshow that only some of the bacterial groups investigated wereaffected by the probiotic treatment. The number of representativestrains of the Enterobacteriaceae family, which also includes poten-tially pathogenic bacteria (Ringø, 1999), decreased at 50 days ph in allprobiotic-fed groups, as well as the number of viable culturablevibrios. This result is in agreement with our previous studies in whichprobiotics were delivered to different models (sea bream, Sparusaurata and sea bass,Dicentrarchus labrax; Carnevali et al., 2004; Silvi etal., 2008). Similarly to what previously described by Swain et al.(2009) who found E. faecium (strainMC13) being able to inhibit Vibrioharvey and V. parahaemolyticus, also in this particular instance, of noteis the ability of probiotic treatment to affect the total viable culturableVibrio spp. In particular, the inhibitory effects of E. faecium IMC 511against V. parahaemolyticus and V. anguillarum were observed by invitro assay (unpublished data). These observations led us to supposethat the Vibrio populations of the common sole larval intestine couldbe significantly reduced by E. faecium IMC 511.
The results here obtained are also in agreement with previousresults reported by Makridis et al., 2008 on S. senegalensis whoshowed that Vibrio spp. (2J18), Shewanella spp. (2J27) and strain J84(unidentified Gram-positive, oxidase-negative and catalase-positive)were effective inhibitors of V. anguillarum and Photobacterium damselasubsp. piscicida, in vitro.
In addition, in the present study, the administration of theaforementioned probiotic candidates, after yolk sac phase, has noeffects on survival rate.
The fact that E. faecium IMC 511 was predominant on day 50 phmight be due to an effective and solid gut accumulation by this probioticstrain,
The decrease of total bacterial concentration (including potentialpathogenic species) in 50 days ph treated larvae led us to concludethat a 50 day E. faecium IMC 511 treatment improves sole larvae well-being in terms of positive microbiota modulation, a factor that mighthave contributed to further beneficial effects for sole larviculturereported here.
Moreover,we found out a big lack ofmolecular probes for the currentmodel (S. solea) which could represent a real problemwhile conductinggenomic studies on this species. To our best knowledge this is the firststudy that reports a series of designed and optimized Real time PCRprimers for testing growth and stress levels in the common sole.
In fact, it has been noted that non optimized captive rearingconditions could potentially represent a source of detrimental stressin fish which can lead to poor survival, growth and eventually affectsea food quality. Thus it is crucial to understand widely in advancewhether the captive rearing conditions used are adequate or potentialsource of detrimental stress. On this regard, several authors havereported higher HSP70 levels after exposure to stress conditions, forexample, in Atlantic salmon, Salmo salar (Smith et al., 1999) andgilthead sea bream (Cara et al., 2005; Rollo et al., 2006; Feidantsis etal., 2009). Moreover, many studies have reported that stress inducedby food inadequacies leads to the up regulation of HSP70 in juvenilesof carp Labeo rohita, (Yengkokpam et al., 2008), in gilthead sea breamS. aurata, and rainbow trout Onchorynchus mykiss early stages (Caraet al., 2005). In our case, a 50 day treatment with E. faecium IMC 511reduced levels of HSP70 in common sole larvae, especially whenprovided at the higher probiotic doses, indicating that this autoch-thonous bacterium could have a positive role in lowering the stressresponse to captivity conditions. These results are in agreement withthe growth results observed at 50 days ph. In fact, the best results forbody weight and lower MSTN1 gene expression were observed at day50 ph, especially in those groups given the higher probioticconcentration. In this regard, we previously showed that lactic acid
Fig. 6. Changes in cortisol levels in sole larvae fed different concentrations of probioticand collected at days 10, 30, 50 post hatch (ph). Values, within each age group, withdifferent superscript letters are significantly different compared to control group(Group 1 C; Pb0.05).
391M.A. Avella et al. / Aquaculture 315 (2011) 384–393

Author's personal copy
bacteria lowered stress levels and enhanced larval and juvenile BWwhen delivered during fish larval stages in sea bass, sea bream andclownfish, Amphiprion ocellaris (Carnevali et al., 2006; Rollo et al.,2006; Avella et al., 2010a,b).
In this study, we hypothesise a relationship between the lowerlevels of MSTN and HSP70, and the highest body weight of sole larvaetreated with probiotic. Moreover, on day 50, the significant increase ofBW observed was only found concomitantly with the highest levels ofE. faecium IMC 511 in the intestinal tract.
Given the importance of HSP70 in responses related to conditionsof captivity and the well-established role of MSTN in control of muscledevelopment (McPherron et al., 1997), it is plausible to hypothesisethat E. faecium IMC 511 is involved in improving larval welfare, in thiscase by lowering HSP70 levels. This in turn benefits larval growth, asalso supported by the lower MSTN gene expression.
The cortisol results for the control group indicate an increase ofcortisol levels during larval growth (10–50 dph). In the treatedgroups, cortisol levels appeared significantly higher after 10 days ofprobiotic administration, which could indicate that the treatmentinduces a disturbing effect. Though cortisol is well known to act as animportant negative regulator of growth (Mommsen et al., 1999),apparently, the increase in cortisol levels in this study did not lead todecreased body growth or survival. Cortisol is known to negativelyaffect growth by decreasing food re-absorption, protein digestibility,and food conversion efficiency (Barton, 2002), but on the other hand,lactic acid bacteria are well known to provide nutritional elements(such as fatty acids, amino acids and vitamins) which play essentialroles in fish growth and development (Teusink and Smid, 2006;Walker, 2009). Thus we hypothesise that the beneficial effects ofprobiotic action in the larvae balance out the negative effect of cortisolon larval growth. On days 30 and 50 ph, a sort of adaptation to theprobiotic delivery was observed, since cortisol levels remainedcomparable in all the treated groups while they continued to increasein the control. Concomitantly to the probiotic treatment adaptation,sole larvae showed higher body weight and standard length,suggesting that E. faecium IMC 511, after the above mentionedadaptation period, may work as a valid growth promoter for commonsole larvae.
The relation between cortisol and myostatin remains to beclarified. While it is well established that glucocorticoids upregulatemyostatin expression by inducing myostatin gene transcription inhumans (Ma et al., 2001), information regarding the regulation ofMSTN expression in fish is limited and the relation with cortisolseemed to vary among the different teleost species. For example,chronic stress (Vianello et al., 2003) or exogenous cortisol (Rodgerset al., 2003; Weber et al., 2005) decreased the abundance of MSTNmRNA in some fish species, such as tilapia, where myostatin appearedto be inversely related to cortisol, with the lowest MSTN mRNA inlarval groups with the highest cortisol levels (Rodgers et al., 2003). Onthe contrary, we previously found in D. labrax larvae that lower levelsof cortisol corresponded to lower levels of myostatin together withhigher growth (Carnevali et al., 2006). In this case, cortisol seems tohave no relation with myostatin gene expression, suggesting theinvolvement of other factors in the regulation of myostatin geneexpression.
5. Conclusions
The present study investigated the role of the autochthonousprobiotic bacterial strain E. faecium IMC 511 in common solelarviculture. For the first time, HSP70 and MSTN genes were partiallysequenced and are currently available in Genbank for all researchersinterested in further studies on common sole, S. solea. This work foundno apparent relationship between cortisol and myostatin geneexpression. On the other hand, our results clearly show that E.faecium IMC 511 positively affects sole larval growth when supplied
for 50 days. Moreover, the best results for intestinal microbiota andgrowthwere obtainedmainly with the higher bacterial concentration.
Acknowledgments
Funding for this study was provided by the “SFOP 2007 — RegioneMarche” to Oliana Carnevali.
References
Avella, M.A., Olivotto, I., Silvi, S., Place, A.R., Carnevali, O., 2010a. Effect of dietaryprobiotics on clownfish: a molecular approach to define how lactic acid bacteriamodulate development in amarine fish. Am. J. Physiol. Regul. Integr. Comp. Physiol.298, R359–R371.
Avella, M.A., Gioacchini, G., Decamp, O., Makridis, P., Bracciatelli, C., Carnevali, O., 2010b.Application of multi-species of Bacillus in Sea Bream Larviculture. Aquaculture 305,12–19.
Barry, T.P., Malison, J.A., Held, J.A., Parrish, J.J., 1995. Ontogeny of the cortisol stressresponse in larval rainbow trout. Gen. Comp. Endocrinol. 97, 57–65.
Barton, B.A., 2002. Stress in fishes: a diversity of responses with particular reference tochanges in circulating corticosteroids. Integ. Comp. Biol. 42, 517–525.
Blonk, R.J.W., Komen, J., Kamstra, A., van Arendonk, J.A.M., 2010a. Effects of grading onheritability estimates under commercial conditions: a case study with commonsole, Solea solea. Aquaculture 300, 43–49.
Blonk, R.J.W., Komen, J., Tenghe, A., Kamstra, A., Van Arendonk, J.A.M., 2010b.Heritability of shape in common sole, Solea solea, estimated from image analysisdata. Aquaculture 307, 6–11.
Cara, J.B., Aluru, N., Moyano, F.J., Vijayan, M.M., 2005. Food-deprivation induces HSP 70and HSP 90 protein expression in larval and gilthead sea bream and rainbow trout.Comp. Biochem. Physiol. B 142, 426–431.
Carnevali, O., de Vivo, L., Sulpizio, R., Gioacchini, G., Olivotto, I., Silvi, S., Cresci, A., 2006.Growth improvement by probiotic in European sea bass juveniles (Dicentrarcuslabrax, L.) with particular attention to IGF-1, myostatin and cortisol geneexpression. Aquaculture 258, 430–438.
Carnevali, O., Zamponi, M.C., Sulpizio, R., Rollo, A., Nardi, M., Orpianesi, C., Silvi, S.,Caggiano, M., Polzonetti, A.M., Cresci, A., 2004. Administration of probiotic strain toimprove sea bream wellness during development. Aquacult. Int. 12, 377–386.
de La Banda, I.G., Lobo, C., León-Rubio, J.L., Tapia-Paniagua, S., Balebona, C., Moriñigo, M.A.,Moreno-Ventas, X., Lucas, L.M., Linares, F., Arce, F., Arijo, S., 2010. Influence of twoclosely related probiotics on juvenile Senegalese sole (Solea senegalensis, Kaup 1858)performance and protection against Photobacterium damselae subsp. Piscicida.Aquaculture 306, 281–288.
Díaz-Rosales, P., Arijo, S., Chabrillón, M., Alarcón, F.J., Tapia-Paniagua, S.T., Martínez-Manzanares, E., Balebona, M.C., Moriñigo, M.A., 2009. Effects of two closely relatedprobiotics on respiratory burst activity of Senegalese sole (Solea senegalensis, Kaup)phagocytes, and protection against Photobacterium damselae subsp. Piscicida.Aquaculture 293, 16–21.
Feidantsis, K., Pörtner, H.O., Lazou, A., Kostoglou, B., Michaelidis, B., 2009. Metabolic andmolecular stress responses of the gilthead seabream Sparus aurata during long-term exposure to increasing temperatures. Mar. Biol. 156, 797–809.
Funkenstein, B., Balas, V., Rebhan, Y., Pliatner, A., 2009. Characterization and functionalanalysis of the 5′ flanking region of Sparus aurata myostatin-1 gene. Comp.Biochem. Physiol. 153, 55–62.
Gatesoupe, F.J., 1999. The use of probiotics in aquaculture. Aquaculture 180, 147–165.Gatesoupe, F.J., 2008.Updating the importanceof lactic acidbacteria infish farming:natural
occurrence and probiotic treatments. J. Mol. Microbiol. Biotechnol. 14, 107–114.Holzapfel, W.H., 1992. Culture media for non-sporulating gram-positive food spoilage
bacteria. Int. J. Food Microbiol. 17 (2), 113–133.Hosseini, S.V., Arlindo, S., Böhme, K., Fernández-No, C., Calo-Mata, P., Barros-Velázquez,
J., 2009. Molecular and probiotic characterization of bacteriocin-producingEnterococcus faecium strains isolated from nonfermented animal foods. J applmicrobial 107, 1392–1403.
Howell, B.R., 1997. A re-appraisal of the potential of the sole, Solea solea (L.) forcommercial cultivation. Aquaculture 155, 355–365.
Imsland, A.K., Foss, A., Conceiçao, L.E.C., Dinis, M.T., Delbare, D., Schram, E., Kamstra, A.,Rema, P., White, P., 2003. A review of the culture potential of Solea solea and S.senegalensis. Rev. Fish Biol. Fish. 13, 379–407.
Lund, I., Steenfeldt, S.J., Banta, G., Hansen, B.W., 2008. The influence of dietaryconcentrations of arachidonic acid and eicosapentaenoic acid at various stages oflarval ontogeny on eye migration, pigmentation and prostaglandin content ofcommon sole larvae (Solea solea L.). Aquaculture 276, 143–153.
Lund, I., Steenfeldt, S.J., Hansen, B.W., 2010. Influence of dietary arachidonic acidcombined with light intensity and tank colour on pigmentation of common sole(Solea solea L.) larvae. Aquaculture 308, 159–165.
Ma, K., Mallidis, C., Artaza, J., Taylor, W., Gonzalez-Cadavid, N., Bhasin, S., 2001.Characterization of 51-regulatory region of human myostatin gene: regulation bydexamethasone in vitro. Am. J. Physiol. 281, E1128–E1136.
Maurilio, L., Olvera-Novoa, M.A., Guzmán-Méndez, B.E., López-Madrid, W., 2003. Use ofthe bacteria Streptococcus faecium and Lactobacillus acidophilus, and the yeastSaccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromisniloticus). Aquaculture 216, 193–201.
Makridis, P., Martins, M., Reis, J., Dinis, M.T., 2008. Use of probiotic bacteria in therearing of Senegalese sole (Solea senegalensis) larvae. Aquacult. Res. 39, 627–634.
392 M.A. Avella et al. / Aquaculture 315 (2011) 384–393

Author's personal copy
McPherron, A.C., Lawler, A.M., Lee, S.-J., 1997. Regulation of skeletal muscle mass inmice by a new TGF-β superfamily member. Nature 387, 83–90.
Merrifield, D.L., Bradley, G., Baker, R.T.M., Davies, S.J., 2010. Probiotic applications forrainbow trout (Oncorhynchus mykiss Walbaum) II. Effects on growth performance,feed utilization, intestinal microbiota and related health criteria postantibiotictreatment. Aquacult. Nutr. 16, 496–503.
Mommsen, T.P., Vijayan, M.M., Moon, T.W., 1999. Cortisol in teleosts: dynamics,mechanisms of actions and metabolic regulation. Rev. Fish Biol. Fish. 9, 211–268.
Moriyama, S., Ayson, F.G., Kawauchi, H., 2000. Growth regulation by insulin-like growthfactor-I in fish. Biosci. Biotechnol. Biochem. 64, 1553–1562.
Overton, J.L., Steenfeldt, S.J., Pedersen, P.B., 2010. The effects of grading on the growthand survival of juvenile Dover sole (Solea solea L.). Aquacult. Res. 42, 31–39.
Pfaffl, M.V., 2001. A new mathematical model for relative quantification in real time rtPCR. Nucl. Acids Res. 29 no. 9 00.
Rasmussen, R., 2001. Quantification on the LightCycler. In: Meuer, S., Wittwer, C.,Nakagawara, K. (Eds.), Rapid Cycle Real-time PCR, Methods and Applications.Springer Press, Heidelberg, pp. 21–34.
Ringø, E., 1999. Lactic acid bacteria in fish: antibacterial effect against fish pathogens. In:Krogdahl, Å.,Mathiesen, S.D., Pryme, I. (Eds.), Effects of antinutrients on thenutritionalvalue of legume diets, COST 98. EEC Publication, Luxembourg, pp. 70–75.
Ringo, E., Gatesoupe, F.J., 1998. Lactic acid bacteria in fish: a review. Acquaculture 160,177–202.
Rodgers, B.D., Weber, G.M., Kelley, K.M., Levine, M.A., 2003. Prolonged fasting andcortisol reduce myostatin mRNA levels in tilapia larvae; short-term fastingelevates. Am. J. Physiol. Regul. Integr. Comp. Physiol. 284, R1277–R1286.
Rollo, A., Sulpizio, R., Nardi, M., Silvi, S., Orpianesi, C., Caggiano, M., Cresci, A., Carnevali,O., 2006. Live microbial feed supplement in aquaculture for improvement of stresstolerance. Fish Physiol. Biochem. 32, 167–177.
Silvi, S., Nardi, M., Sulpizio, R., Orpianesi, C., Caggiano, M., Carnevali, O., Cresci, A., 2008.Effect of the addition of Lactobacillus delbrueckii subsp. delbrueckii on the gut
microbiota composition and contribution to the well-being of European sea bass(Dicentrarchus labrax, L.). Microb. Ecol. Health Dis. 20, 53–59.
Smith, T.R., Tremblay, G.C., Bradley, T.M., 1999. Hsp70 and a 54 kDa protein (osp54) areinduced in salmon (Salmo salar) in response to hyperosmotic stress. J. Exp. Zool.284, 286–298.
Suzer, C., Çoban, D., Kamaci, H.O., Saka, S., Firat, K., Otgucuoğlu, Ö., Küçüksari, H., 2008.Lactobacillus spp. bacteria as probiotics in gilthead sea bream (Sparus aurata, L.) larvae:effects on growth performance and digestive enzyme activities. Aquaculture 280,140–145.
Swain, S.M., Singh, C., Arul, V., 2009. Inhibitory activity of probiotics Streptococcusphoecae PI80 and Enterococcus faecium MC13 against Vibriosis in shrimp Penaeusmonodon. World J. Microbiol. Biotechnol. 25, 697–703.
Teusink, B., Smid, E.J., 2006. Modelling strategies for the industrial exploitation of lacticacid bacteria. Nat. Rev. Microbiol. 4, 46–56.
Vianello, S., Brazzoduro, L., Dalla Valle, L., Belvedere, P., 2003. Myostatin expressionduring development and chronic stress in zebrafish (Danio rerio). J. Endocrinol.176, 45–59.
Vine, N.G., Leukes, W.D., Kaiser, H., 2006. Probiotics in marine larviculture. FEMSMicrobiol. Rev. 30, 404–427.
Walker, A., 2009. Milk and two oligosaccharides. Nat. Rev. Microbiol. 7, 483.Wang, Y., Li, J., Lin, J., 2008. Probiotics in aquaculture: challenges and outlook.
Aquaculture 281, 1–4.Weber, T.E., Small, B.C., Bosworth, B.G., 2005. Lipopolysaccharide regulates myostatin
and MyoD independently of an increase in plasma cortisol in channel catfish(Ictalurus punctatus). Domest. Anim. Endocrinol. 28, 64–73.
Wendelaar Bonga, S.E., 1997. The stress response in fish. Physiol. Rev. 77, 591–625.Yengkokpam, S., Pal, A.K., Sahu, N.P., Jain, K.K., Dalvi, R., Misra, S., Debnath, D., 2008.
Metabolic modulation in Labeo rohita fingerlings during starvation: Hsp70expression and oxygen consumption. Aquaculture 285, 234–237.
393M.A. Avella et al. / Aquaculture 315 (2011) 384–393
Related Documents











