© 2012 ISZS, Blackwell Publishing and IOZ/CAS 74 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 Use of electrosense in the feeding behavior of sturgeons Xuguang ZHANG, 1,2 Jiakun SONG, 1 Chunxin FAN, 1 Hongyi GUO, 1 Xiaojie WANG 1 and Horst BLECKMANN 3 1 Institute for Marine Biosystem and Neuroscience, Shanghai Ocean University, Shanghai, China, 2 College of Food Science and Technology, Shanghai Ocean University, Shanghai, China and 3 Institute of Zoology, University of Bonn, Bonn, Germany Abstract The electro-receptive lateral line system appeared early in the evolutionary history of fish. Sturgeons, members of the primitive Chondrostei group, are known for their electroreceptors (ampullae of Lorenzini) on the head, which are thought to be sensitive to weak electric fields in aquatic environments and involved in feeding, mating and migration. Here, we report the results of a set of behavioral and electrophysiological experiments designed to determine the function and characteristics of the electrosense in cultured sturgeons. The results showed Sibe- rian sturgeon (Acipenser baerii Brandt, 1869) feeding striking at bio-electric fields produced by living feed-fish enclosed in a gel chamber and at the corrosion field produced by metal rods. With an electric stimulus that mim- ics the bio-electric fields produced by living prey, the relative discharge rate of electrosensory neurons in the dorsal octavolateralis nucleus (DON) was modulated by and phase-locked to sinusoidal stimulus and some units showed selectivity for dipolar direction in white sturgeon (Acipenser transmontanus Richardson,1836). This is the first study to provide the empirical evidence correlating with electrosensory behaviors and electrophysiolog- ical responses in cultured sturgeons, and suggesting that electrosense does play an innate role in feeding behav- ior of sturgeon. We believe this will have important implications for protecting sturgeons in the wild. Key words: behavior, dipole, electrosense, hindbrain, sturgeon Integrative Zoology 2012; 7: 74–82 doi: 10.1111/j.1749-4877.2011.00272.x ORIGINAL ARTICLE Correspondence: Jiakun Song, Institute for Marine Bio-system and Neurosciences, Shanghai Ocean University, 999 Hucheng Road, Shanghai 201306, China. Email: [email protected] In natural habitats, the sturgeon is an opportunistic, non- visual substrate feeder, preying on any living items in its murky environment (Sbikin 1974; Miller 2004). In feeding, they are believed to make use of a combina- tion of sensors, including olfactory sensors, tactile che- mosensory cues on the 4 barbells and electroreceptors (Miller 2004). Over the past 30 years, the electrore- ceptors (ampullae of Lorenzini) in sturgeons, such as Scaphirhynchus platorynchus Rafinesque, 1820 (Tee- ter et al. 1980), Acipenser ruthenus Linnaeus, 1758 (Jørgensen 1980), Acipenser sinensis Gray, 1834 (Li- ang 1999) and Acipenser baerri Brandt, 1869 (W. Song, 2011, unpubl. data) have been characterized as similar INTRODUCTION The sturgeon is an ancient freshwater fish with dis- tinctive electroreceptors on the ventral side of the ros- trum (Bemis et al. 1997; Jørgensen 2005), and has an important evolutionary status in fish (Northcutt 2009).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© 2012 ISZS, Blackwell Publishing and IOZ/CAS74
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Use of electrosense in the feeding behavior of sturgeons
Xuguang ZHANG,1,2 Jiakun SONG,1 Chunxin FAN,1 Hongyi GUO,1 Xiaojie WANG1 and Horst BLECKMANN3
1Institute for Marine Biosystem and Neuroscience, Shanghai Ocean University, Shanghai, China, 2College of Food Science and Technology, Shanghai Ocean University, Shanghai, China and 3Institute of Zoology, University of Bonn, Bonn, Germany
AbstractThe electro-receptive lateral line system appeared early in the evolutionary history of fish. Sturgeons, members of the primitive Chondrostei group, are known for their electroreceptors (ampullae of Lorenzini) on the head, which are thought to be sensitive to weak electric fields in aquatic environments and involved in feeding, mating and migration. Here, we report the results of a set of behavioral and electrophysiological experiments designed to determine the function and characteristics of the electrosense in cultured sturgeons. The results showed Sibe-rian sturgeon (Acipenser baerii Brandt, 1869) feeding striking at bio-electric fields produced by living feed-fish enclosed in a gel chamber and at the corrosion field produced by metal rods. With an electric stimulus that mim-ics the bio-electric fields produced by living prey, the relative discharge rate of electrosensory neurons in the dorsal octavolateralis nucleus (DON) was modulated by and phase-locked to sinusoidal stimulus and some units showed selectivity for dipolar direction in white sturgeon (Acipenser transmontanus Richardson,1836). This is the first study to provide the empirical evidence correlating with electrosensory behaviors and electrophysiolog-ical responses in cultured sturgeons, and suggesting that electrosense does play an innate role in feeding behav-ior of sturgeon. We believe this will have important implications for protecting sturgeons in the wild.
Key words: behavior, dipole, electrosense, hindbrain, sturgeon
Integrative Zoology 2012; 7: 74–82 doi: 10.1111/j.1749-4877.2011.00272.x
ORIGINAL ARTICLE
Correspondence: Jiakun Song, Institute for Marine Bio-system and Neurosciences, Shanghai Ocean University, 999 Hucheng Road, Shanghai 201306, China.Email: [email protected]
In natural habitats, the sturgeon is an opportunistic, non-visual substrate feeder, preying on any living items in its murky environment (Sbikin 1974; Miller 2004). In feeding, they are believed to make use of a combina-tion of sensors, including olfactory sensors, tactile che-mosensory cues on the 4 barbells and electroreceptors (Miller 2004). Over the past 30 years, the electrore-ceptors (ampullae of Lo ren zini) in sturgeons, such as Scaphirhynchus platorynchus Rafinesque, 1820 (Tee-ter et al. 1980), Acipenser ruthenus Linnaeus, 1758 (Jørgensen 1980), Acipenser sinensis Gray, 1834 (Li-ang 1999) and Acipenser baerri Brandt, 1869 (W. Song, 2011, unpubl. data) have been characterized as similar
INTRODUCTIONThe sturgeon is an ancient freshwater fish with dis-
tinc tive electroreceptors on the ventral side of the ros-trum (Bemis et al. 1997; Jørgensen 2005), and has an important evolutionary status in fish (Northcutt 2009).

© 2012 ISZS, Blackwell Publishing and IOZ/CAS 75
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Electrosense in sturgeons
to the electrosensory ampullae in elasmobranches and have been thought to be sensitive to weak electric fields in the aquatic environment and to assist the stu rge on in prey detection (Wilkens & Hofmann 2005). Howev-er, with the blooming of aquaculture, several species of sturgeons have been artificially spawned and raised in captivity, and fed commercial pellets that lack electric cues, generation after generation (Miller 2004; Wei et al. 2004). Whether the sturgeons, especially the cultured sturgeons, use electrosense in feeding behavior?
To address the question, we conducted behavioral and electrophysiological experiments specifically designed to investigate the function and characteristics of electro-sense in sturgeons. We recorded the behavioral pattern of the Siberian sturgeon A. baerii in response to bio-electric fields produced by living prey fish embedded in a gel chamber as well as corrosion fields produced by a metal rod. We then recorded the response proper-ties of the electrosensory neurons in the dorsal octavol-ateralis nucleus (DON) from the hindbrain of the white sturgeon A. transmontanus using simulated sinusoidal electric fields mimicking the characteristics of the bio-electric fields produced by living prey. Our results dem-onstrate that passive electrosense plays a role in cultured sturgeons’ feeding behavior, and provide an experimen-tal explana tion for the interactions between the prey and predator.
MATERIALS AND METHODSA brief description of each of the following experi-
men tal protocols is provided. These protocols were de-signed to determine the role of electrosense in sturgeon feeding behavior. All animal protocols were approved by the Review Committee for the Use of Animal Sub-jects of Shanghai Ocean University.
Feeding behavior
Ten small sturgeons (15–20 cm total length) wer e used in the experiment. These sturgeons ca n prey on the small mos q uito fish, Gambusia affinis Baird and Girard, 1853, which is a common feed-fish in so uth ern Ch ina (about 2 cm in total length) (Fig. 1). The ex pe ri ment was per-form ed i n a hexagonal g la ss tank 100 cm in diameter with 20 cm of water. The bottom of the tank was two layers of glass. The top layer contained two circu lar ho-l es (1 cm in depth and 6 cm in di ame ter) 25 cm apart. These holes were used as the gel cham ber holders. Each living G. affinis was embedded in the gel (1.5%) cham-ber to produce the bio-electric fields that were used as electric stimuli (Kalmijn 1971). A similar gel chamber
filled with a solution of pellets (with a small hole to al-low odors to escape) was placed in the other hole at the bottom of the tank. This was used as an odor stimulus (Fig. 2). Before experiment, the fish was allowed to ha-bituate to the test tank with blank gel chamber (with-out stimuli as a control) for 24 h. Then the gel chamber with electric and odor stimulus were lower into chamber holders randomly. During the experiments, single fish were introduced into the tank to determine their response to both electric and odor stimulus.
All experiments were performed in the dark using in-frared light for illumination. Two IR-sensitive camer-as recorded the movement of the sturgeons, one from above the tank and the other from the side. The swim-ming movements were tracked using a tracking system (EthoVision XT 7.0, Noldus, Wageningen, The Neth-
Figure 1 A Siberian sturgeon (Acipenser baerii) attacking and sucking a mosquito fish (Gambusia affinis) into its mouth.

© 2012 ISZS, Blackwell Publishing and IOZ/CAS76
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
X. Zhang et al.
erlands). To avoid the adaptation, each fish was tested for only 2 min. The sturgeon is a benthic and epibenthic feeder. It uses its powerful suction feeding mechanism to feed on aquatic animals on or within substrate and to prey on small fish (Miller 2004). Feeding striking was here defined as powerful suction on the surface of the gel chamber. Data from 10 fish were analyzed with sta-tistical software SPSS 13.0 (t-test, SPSS Inc, Chicago, IL, USA).
Behavioral response to metal rod
Sturgeons were introduced into the same hexago-nal tank filled with 18 cm of dechlorinated and aerated water at 670 ± 20 μS/cm. A custom-made manipulator lowered the test rod into the water 10 cm from the tank wall, where the fish would swim past between the rod and the wall. The aluminum rod was 2.5 cm in diame-ter and 20 cm long, like the rods used in previous pad-dlefish studies (Gurgens et al. 2000). An aluminum rod covered with an acrylic glass tube (T-aluminum rod), a hollow acrylic glass tube either filled with or empty of pellets (P-glass tubes, both identical in size to the met-al rod), were also lowered into the tank as controls. The odor of the pellets spread via holes in the wall of P-glass tube. Sturgeon behavior patterns were monitored using the same tracking system and recorded using IR-sen-sitive cameras. During the experiment, the fish swam
along the wall of the tank after acclimating for 30–60 minutes. Then the test rod was lowered while the fish was on the opposite side of the tank (Gurgens et al. 2000). Behavioral responses to aluminum, T-aluminum, and P-glass (with and without pellets) rod were record-ed in a series of experimental runs. Each rod was test-ed individually in random order for 5 minutes. Intervals of more than 5 minutes were allowed to elapse between one rod and the next to remove the fish’s memory of the rod. The rod was held perpendicular to the tank bottom and parallel to the barbels of the sturgeon. When the fish swam close to the rod, the barbels could not touch it di-rectly. To compare responses, the numbers of feeding striking from 12 fishes were calibrated and analyzed. Feeding striking was here defined as circling and pow-erfully sucking on the rod.
Bio-electric fields and corrosion fields
The bio-electric fields generating from 20 G. affinis with individual total length ranging from 1.5 cm to 2.5 cm and the corrosion fields from the aluminum rods were mea-sured using Ag/AgCl electrodes in 600 ± 20 μS/cm water. The reference electrodes were positioned on opposite sides of the tank, about 50 cm from the fish and 80 cm from the metal rod. The recording electrodes were po-sitioned at various sites along the axis of the fish’s body at 5 mm intervals to measure the electric fields along the fish’s body. Recording started after the fish was low-ered into the water for 5–10 s. All data were normalized to values of a fish with a 2.5 cm body length. In mea-suring the corrosion field, the recording electrode was positioned 5 mm from the surface of the rod. The re-corded signal was amplified ×500 (DC differential am-plifier, model 5307, NF corporation, Yokohama, Japan) and filtered at DC-30 Hz (wave analysis system, Spike 2, Cambridge Electronic Design, UK), and Fourier analy-sis was performed using Matlab software (13.0).
Electrophysiology
Electrophysiological responses were recorded from the dorsal octavolateralis nucleus (DON), the part of the brain in which electrosensory information is first pro cessed. The brain was exposed by removal of the overlying skull while the fish was under anesthesia (1:10,000v/v, MS-222). The fish was then immobilized by injection of pancuronium (Organon Teknika, Oberschleissheim, Germany) into the back muscles. It was then artificially irrigated with aer-ated water through the mouth. Electric fields simulating live prey fish (sinusoidal currents at 5Hz) were generat-ed using a dipole electrodes connected to a constant cur-
Figure 2 Feeding striking on gel blocks. (a,b) The fish struck a gel block containing Gambusia affinis. (c,d) The fish struck a gel block containing a solution of pellets. The bottom trace shows the changes in swimming velocity. Bold red lines mark fast swimming period. Bold black lines mark the slow swim-ming period.

© 2012 ISZS, Blackwell Publishing and IOZ/CAS 77
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Electrosense in sturgeons
rent source (A395 linear stimulus isolator, WPI, Saraso-ta, FL, USA) driven by a D/A converter (CED system, Micro1401+, Cambridge, UK) at 10 KHz. The dipole was 5 mm away from the target fish, under the rostrum. To stimulate the electrosensory DON units, two dipoles of different orientations were used to examine the neu-rons’ selectivity of field orientation. One dipolar orien-tation paralleled the rostro-caudal axis of the body (0°), and the other was perpendicular to the rostro-caudal axis (90°). Cell responses were recorded with indium-alloy-filled glass electrodes (0.1–5 MW). They were amplified 1000 times, band-pass filtered at 100–10 kHz (DAM80, WPI, USA), and analyzed using Spike 2 software (wave analysis system, Spike 2, Cambridge, UK). The electric fields introduced to the tank were calibrated by measur-ing the voltage drop at 10 mm in the middle of the tank parallel to the field lines (Hofmann et al. 2005).
For responses to 5 Hz fields with amplitudes ranging from 1–100 µV/cm and the different dipoles with 5 Hz at 100 µV/cm sinusoidal wave, two parameters were cal-culated. One was the relative discharge rate, the mean spike rate during the stimulation minus the spontane-ous rate. This was calculated to determine the units’ sen-sitivity (Hofamnn et al. 2008). Another was the degree of phase locking, which was obtained by computing the normalized period histogram of phase angles of spikes relative to the sine wave cycle. Then a value D was cal-culated based on an estimate of entropy of period histo-gram. It was used as a measure of the degree of phase locking (Kajikawa & Hackett 2005). This value is 0 rep-resents random data and 1 represents all spikes in the same bin of the period histogram.
RESULTS
Feeding striking
In this experiment, the fish always swam along the wall of the tank on or close to the bottom in blank con-trols. The gel block containing living G. affinis and that containing pellet solution were lowered into the feed-ing holders when the fish was on the opposite side of the tank. It was observed that the fish swam rapidly to-ward the gel block and sucked at it powerfully, as if it was a prey item. This kind of behavior pattern was here defined as feeding striking. It has an apparent feed-ing character even when no food is obtained. However, there was little or no response to the control gel block. Analysis of the association between swimming veloc-ity and behavior pattern showed that the fish acceler-ated before exerting suction on or near the gel block
(Fig. 2). Figure 2 shows this change in swimming ve-locity between feeding striking on the two kinds of gel blocks. The numbers of feeding strikes were calculated and statistically analyzed (Fig. 3). The results showed no significant difference between the number of feed-ing strikes evoked by bioelectric stimulation and those evoked by odor stimulation (t-test, P = 0.27).
Behavioral response to metal rod
During the acclimation period, the fish swam in larg-er and larger circles until it followed the tank wall. With no interrupting obstacle, the fish seldom deviated from this normal swimming pattern. The lowering of the alu-minum rod into the water disrupted the background be-havior of the fish. Most fish swam around the rod and sucked on the metal rod frequently (Fig. 4). This be-havior pattern should be associated with food searching or feeding behavior. Previous studies on sturgeons have attrib-uted this to the odor of the amino acids (Kasumyan 2002). There were about 100 times (n = 97) as many strikes for the 10 prey fishes. There were fewer strikes at the T-alu-minum rod (n = 8) and P-glass tubes without pellets (n = 13). When the P-glass tube with pellets was provid-ed, the fish showed strong circling and suction responses again (n = 123) (Fig. 4). The numbers of feeding strikes were in this way found to be dependent on the materials of the rods (Kruskal–Wallis test, χ2
2 = 28.754, P < 0.01) .
Bioelectric field from Gambusia affinis and corrosion field from metal rod
The bioelectric field generated by fishes shows typ-ical features, such as a dipole fields with negative po-
Figure 3 Numbers of feeding striking on gel blocks with electric and odor stimuli.

© 2012 ISZS, Blackwell Publishing and IOZ/CAS78
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
X. Zhang et al.
tential up to –24 ± 2.4 µV on the head and positive po-tential up to 21 ± 1.6 µV on the tail (Fig. 5a). An AC modulation superimposed on the DC field with a mean value of 4.2 ± 0.8 µV was observed. The signals from the head and tail were fast Fourier transform analyzed, and the power spectrum showed a large number of DC components in both head and tail. However, the head signal exhibited a large peak near 1 Hz, and almost twice that of the corresponding peak in the tail signals (Fig. 5b). We found the bandwidth of the AC component (1–2.5 Hz) to be consistent with the respiration rhythm (60–170 times/min) of G. affinis.
The electric signals generated by the aluminum rod, T-aluminum rod (2.5 cm diameter), and blank control rod were recorded using a monopolar electrode. Results showed that the amplitude of signals from the T-aluminum rod and control rod were similar, and the peak-to-peak values of both were about 15 µV. However, the wave-form of the signals from aluminum rods was clearly dis-tinct from these. The peak-to-peak value reached 90 µV (Fig. 6a). Power spectra showed that there were two fre-quency peaks in signals generated by aluminum rod, one large peak at 0–1 Hz with power above 2 and one small peak at 1–2 Hz. There was only a small peak in the signal generated by the T-aluminum rod, with power less than 1 (Fig. 6b). This indicated that the aluminum rods generated weak corrosion electric fields ca. 90 μV from peak to peak at 0–2 Hz, whereas the T-aluminum rod fended off the electrochemical reaction on the inter-face between the water and the metal surface.
Electrosensitivity
In the electrophysiological experiments, we dem-onstrated the sensitivity of DON units to weak elec-tric fields with different amplitudes from 1–100 µV/cm in sturgeons. There were spontaneous activities in anes-thetized sturgeons in the absence of electric stimulation, and the spontaneous firing rate ranged from 0.41 Hz to 98.54 Hz in all 17 DON units recorded. The neural activ-ity of DON units was modulated by the external sinusoi-dal electric fields generated from dipole sources on the ventral side of the rostrum. These dipole electrodes sim-ulated the electric field generated by prey and effective-ly evoked responses from most of the electroreceptors. During the positive cycle of the stimulating sine wave, recorded unit activity increased; during the negative cy-cle, firing was suppressed (Fig. 7c). The alternating exci-tation and suppression in the firing showed strong phase coupling with the stimulus. We evaluated the responses by relative discharge rate and degree of phase coupling.
Figure 4 Sturgeon response sequence. (a) Sturgeon response to an aluminum rod. (b) Sturgeon response to a glass tube with pellets inside (P-glass).
Figure 5 Bio-electric fields of Gambusia affinis. (a) Relative po-tential along the fish body, red represents negative potential on the head and yellow represents positive potential on the tail. (b) Power spectra of electric signals from the head and tail.
a
b

© 2012 ISZS, Blackwell Publishing and IOZ/CAS 79
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Electrosense in sturgeons
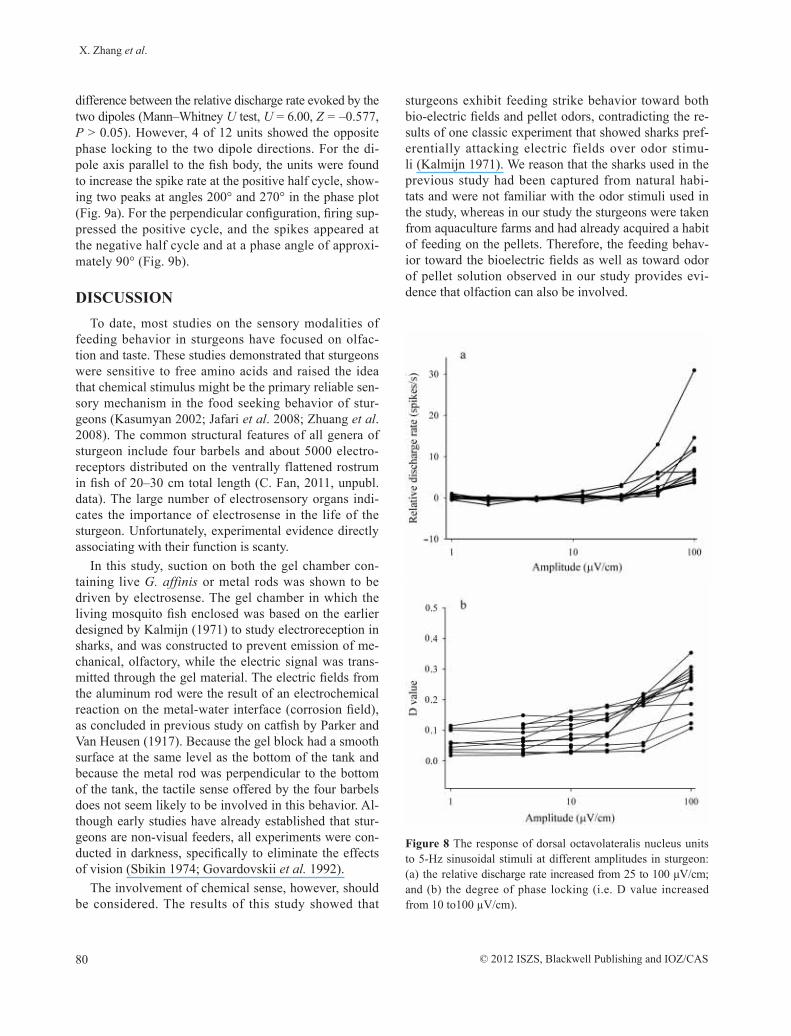
The results showed that response magnitude of DON units (n = 12) were dependent on the amplitude of the stim-ulus. Above 25–100 µV/cm, there was a significant increase in the relative discharge rate (t-test, P = 0.009), to a maxi-mum of 30 spikes/s (Fig. 8a). The degree of phase lock-ing was expressed as D value, which increased slowly as the amplitude of the stimulus increased from 5–100 µV/cm (P = 0.008) (Fig. 8b).
Fish in water generate dipole electric fields and the direction of the dipole field is usually along the axis of the body (Peters et al. 2002; Kalmijn 1972). To estab-lish the neuron’s selectivity for the dipolar direction, two similar dipole electrodes were used with one dipo-lar axis parallel to the rostocaudal axis of the fish body and the other perpendicular to it. For each dipole stim-
Figure 6 Corrosion fields from aluminum rods. (a) Signal from aluminum, T-aluminum and blank control rods in water. (b) Power spectrum of corrosion fields generated by aluminum and T-alumi-num rods.
Figure 7 Recording sites and spikes of 2 exemplar cells. (a) Brain site within the sturgeon and the recording site within the brain. (b) Indi-um electrode inserted into the brain to record single units. (c) Spikes evoked by the sinusoidal stimulation.
a
b
c
ulus, the DON units (n = 12) showed phase locked re-sponses as previous described, and the mean increases of relative discharge rates were 8.07 ± 5.41 spikes/s (0°) and 6.74 ± 3.91 spikes/s (90°). There was no significant
© 2012 ISZS, Blackwell Publishing and IOZ/CAS80
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
X. Zhang et al.
sturgeons exhibit feeding strike behavior toward both bio-electric fields and pellet odors, contradicting the re-sults of one classic experiment that showed sharks pref-erentially attacking electric fields over odor stimu-li (Kalmijn 1971). We reason that the sharks used in the previous study had been captured from natural habi-tats and were not familiar with the odor stimuli used in the study, whereas in our study the sturgeons were taken from aquaculture farms and had already acquired a habit of feeding on the pellets. Therefore, the feeding behav-ior toward the bioelectric fields as well as toward odor of pellet solution observed in our study provides evi-dence that olfaction can also be involved.
Figure 8 The response of dorsal octavolateralis nucleus units to 5-Hz sinusoidal stimuli at different amplitudes in sturgeon: (a) the relative discharge rate increased from 25 to 100 μV/cm; and (b) the degree of phase locking (i.e. D value increased from 10 to100 µV/cm).
difference between the relative discharge rate evoked by the two dipoles (Mann–Whitney U test, U = 6.00, Z = –0.577, P > 0.05). However, 4 of 12 units showed the opposite phase locking to the two dipole directions. For the di-pole axis parallel to the fish body, the units were found to increase the spike rate at the positive half cycle, show-ing two peaks at angles 200° and 270° in the phase plot (Fig. 9a). For the perpendicular configuration, firing sup-pressed the positive cycle, and the spikes appeared at the negative half cycle and at a phase angle of approxi-mately 90° (Fig. 9b).
DISCUSSIONTo date, most studies on the sensory modalities of
feeding behavior in sturgeons have focused on olfac-tion and taste. These studies demonstrated that sturgeons were sensitive to free amino acids and raised the idea that chemical stimulus might be the primary reliable sen-sory mechanism in the food seeking behavior of stur-geons (Kasumyan 2002; Jafari et al. 2008; Zhuang et al. 2008). The common structural features of all genera of sturgeon include four barbels and about 5000 electro-receptors distributed on the ventrally flattened rostrum in fish of 20–30 cm total length (C. Fan, 2011, unpubl. data). The large number of electrosensory organs indi-cates the importance of electrosense in the life of the sturgeon. Unfortunately, experimental evidence directly associating with their function is scanty.
In this study, suction on both the gel chamber con-taining live G. affinis or metal rods was shown to be driven by electrosense. The gel chamber in which the living mosquito fish enclosed was based on the earlier designed by Kalmijn (1971) to study electrorecep tion in sharks, and was constructed to prevent emission of me-chanical, olfactory, while the electric signal was trans-mitted through the gel material. The electric fields from the aluminum rod were the result of an electrochemical reaction on the metal-water interface (corrosion field), as concluded in previous study on catfish by Parker and Van Heusen (1917). Because the gel block had a smooth surface at the same level as the bottom of the tank and because the metal rod was perpendicular to the bottom of the tank, the tactile sense offered by the four barbels does not seem likely to be involved in this behavior. Al-though early studies have already established that stur-geons are non-visual feeders, all experiments were con-ducted in darkness, specifically to eliminate the effects of vision (Sbikin 1974; Govardovskii et al. 1992).
The involvement of chemical sense, however, should be considered. The results of this study showed that

© 2012 ISZS, Blackwell Publishing and IOZ/CAS 81
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051
Electrosense in sturgeons
The use of electrosense in predation by sturgeons was further demonstrated by the electrophysiological prop-erties of DON units. We generated an electric field sim-ulating the bio-electric fields produced by feed-fish G. affinis in order to record the neurons’ response to stim-uli mimicking those produced by natural food. The re-sponses of electrosensory DON units were found to in-crease with the amplitude of the electric stimulus. These results are consistent with the electrosensitivity charac-teristics of other ampullary electrosensory fish (Wilkens & Hofmann 2005). In natural environments, the stron-
ger bioelectric intensities would represent bigger prey or schools of prey (Patullo & Macmillan 2004). Elec-trosensitivity helps fish to find more prey in turbid envi-ronments full of mixed cues. As shown in this and oth-er studies, the bio-electric fields of aquatic animals are dipolar, negative at the head and positive at the tail (Pe-ters et al. 2002; Kalmijn 1972). Some DON units show different phase responses in response to different dipole directions. This may help the sturgeons to distinguish or to chase individuals from schools of prey accord-ing the directions of their head-tail dipoles. The neu-rons with response to object vibration direction was also found in mechanosensory lateral line system and may also be used in orientation of objects (Bleckmann & Zelick 2009). In this study, several lines of evidence re-garding the processing of electrosensory information in sturgeons suggest that electrosense is used in predation. However, it may have other functions, such as in mating or schooling. Our results may help further research into these areas.
To our knowledge, this is not only the first study to provide evidence that sturgeons use electrosense in pre-dation. In addition, because all the sturgeons used in this study were from aquaculture farms, bred artificially and raised in captivity, our results indicate that the habits of pellet feeding in aquaculture have not yet caused stur-geons to lose their natural ability to use electrosense in predation. Thus, the results have important implications for protecting sturgeons and healing sturgeons captured accidentally.
ACKNOWLEDGMENTSWe thank Professors Michael H. Hofmann, Guosheng
Zhang, Lon A. Wilkens and Jizhou Yan for helpful discus-sion and Pengxiang Xu,BinBin Xing for assistance with our experimental equipment. This research was support-ed by the National Natural Science Foundation of China (30970365) and the Science and Technology Commission of Shanghai Municipality of China (073205109), and by hydrobiology funding project S30701 and the Excellent Graduate Students Theses Cultivation Program of Shang-hai Municipality.
REFERENCESBemis WE, Findeis EK, Grande L (1997). An overview
of Acipenseriformes. Environmental Biology of Fishes 48, 25–71.
Bleckmann H, Zelick R (2009). Lateral line system in fish. Intergrative Zoology 4, 13–25.
Figure 9 Responses to different dipolar directions of dorsal oc-tavolateralis nucleus units. (a) Response to dipole fields with direc-tions parallel to the rostro–caudal axis of the body. (b) Response to dipole fields with directions perpendicular to the rostro–caudal axis. The results are displayed as a PSTH (bin width 5 ms)

© 2012 ISZS, Blackwell Publishing and IOZ/CAS82
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051
X. Zhang et al.
Govardovskii V, R Hlich P, Szel A, Zueva L (1992). Im-munocytochemical reactivity of rod and cone visual pigments in the sturgeon retina. Visual Neuroscience 8, 531–37.
Gurgens C, Russell DF, Wilkens LA (2000). Electrosen-sory avoidance of metal obstacles by the paddlefish. Journal of Fish Biology 57, 277–90.
Hofmann MH, Jung SN, Siebenaller U, Preissner M, Chagnaud BP, Wilkens LA (2008). Response proper-ties of electrosensory units in the midbrain tectum of the paddlefish (Polyodon spathula Walbaum). Journal of Experimental Biology 211, 773–79.
Jafari SV, Abtahi B, Kasumyan A, Abedian A, Abta-hi B (2008). Taste attractiveness of free amino acids for juveniles of Persian sturgeon Acipenser persicus. Journal of Ichthyology 48, 124–33.
Jørgensen JM (1980). The morphology of the Lorenzi-nian ampullae of the sturgeon Acipenser ruthenus (Pisces: Chondrostei). Acta Zoologica 61, 87–92.
Jørgensen JM (2005). Morphology of electroreceptive sensory organs. In: Bullock TH, Hopkins CD, Popper AN, Fay RR, eds. Electroreception. Springer, New York, pp. 47–67.
Kajikawa Y, Hackett TA (2005). Entropy analysis of neu ron al spike train synchrony. Journal of Neuroscience Methods 149, 90–93.
Kalmijn AJ (1971). The electric sense of sharks and rays. Journal of Experimental Biology 55, 371–83.
Kalmijn AJ (1972). Bioelectric fields in sea water and the function of the ampullae of Lorenzini in elasmo-branch fishes. Scripps Institution of Oceanography Reference Series 72–83.
Kasumyan AO (2002). Sturgeon food searching behav-iour evoked by chemical stimuli: a reliable sensory mechanism. Journal of Applied Ichthyology 18, 685–90.
Liang XF (1996). The structure and function of the am-pullae on the ventral part of the snout of Chinese sturgeon, Acipenser sinensis Gray. Oceanologia et Limnologia Sinica 27, 1–5.
Miller M (2004). The ecology and functional morphol-ogy of feeding of North American sturgeon and pad-dlefish. Fish and Fisheries Series 27, 87–102.
Northcutt RG (2009). Phylogeny of nucleus medianus of the posterior tubercle in ray-finned fishes. Integrative Zoology 4, 134–51.
Parker GH, Van Heusen AP (1917). The responses of the cat fish, Amiurus nebulosus, to metallic and non-metal lic rods. American Journal of Physiology 44, 405–20.
Patullo BW, Macmillan DL (2004). The relationship be-tween body size and the field potentials generated by swimming crayfish. Comparative Biochemistry and Physiology – Part A 139, 77–81.
Peters RC, Van Wessel T, Van Den Wollenberg BJW, Bretschneider F, Olijslagers AE (2002). The bioelec-tric field of the catfish Ictalurus nebulosus. Journal of Physiology – Paris 96, 397–404.
Sbikin YN (1974). Age related changes in the role of vi-sion in the feeding of various fish. Joural of Ichthyology 14, 133–8.
Teeter JH, Szamier RB, Bennett MVL (1980). Ampul-lary electroreceptors in the sturgeon Scaphirhynchus platorynchus (Rafinesque). Journal of Comparative Physiology – Part A 138, 213–23.
Wei Q, He J, Yang D, Zheng W, Li L (2004). Status of sturgeon aquaculture and sturgeon trade in China: a review based on two recent nationwide surveys. Journal of Applied Ichthyology 20, 321–32.
Wilkens LA, Hofmann MH (2005). Behavior of animals with passive, low-frequency electrosensory systems. In: Bullock TH, Hopkins CD, Popper AN, Fay RR, eds. Electroreception. Springer, New York, pp.229–63.
Zhuang P, Zhang LZ, Luo G, Zhang T, Feng GP, Liu J (2008). Function of sense organ to the feeding beha-vior of juveniles Chinese sturgeon captured from the Yangtze estuary. Acta Hydrobiologica Sinica 32, 475–81.
Related Documents











