Upregulation of osteoblast apoptosis by malignant plasma cells: a role in myeloma bone disease Franco Silvestris, Paola Cafforio, Marco Tucci, Daniela Grinello and Franco Dammacco Department of Internal Medicine and Oncology (DIMO), University of Bari, Bari, Italy Received 8 November 2002; accepted for publication 10 February 2003 Summary. Typical features of multiple myeloma (MM) are osteolytic lesions and severely affected bone regeneration. This study of 53 MM patients demonstrates an enhance- ment of osteoblast cytotoxicity by malignant myeloma cells via the upregulation of apoptogenic receptors, including Fas ligand (Fas-L) and tumour-necrosis-factor-related apoptosis inducing ligand (TRAIL). Both were significantly increased in the marrow myeloma cells of patients with extensive osteolytic lesions in a fashion similar to the highly malig- nant human myeloma cell line MCC-2. Osteoblasts from these subjects over-expressed Fas and death receptor (DR) 4/5 and underwent dramatic apoptosis when co-cultured with either MCC-2 or autologous myeloma cells. In osteo- blast and myeloma cell co-cultures, monocyte chemoat- tractant protein 1 (MCP-1) mRNA was upregulated in osteoblasts from patients with severe bone disease in parallel with increased CC-chemokine receptor R2 (CCR2) expres- sion, the ligand of MCP-1, in the myeloma cells. This chemokine was shown to activate malignant cell migration in vitro. An upregulation of ICAM-1 expression occurred in osteoblasts from patients with active skeleton disease. This upregulation appeared to be an effect of malignant plasma cell contact, as MCC-2 co-culture greatly enhanced ICAM-1 production by resting osteoblasts from patients without skeleton involvement. Our results suggest that osteoblasts in active myeloma are functionally exhausted and promptly undergo apoptosis in the presence of myeloma cells from patients with severe bone disease. It is suggested that this cytotoxic effect plays a pivotal role in the pathogenesis of defective bone repair. Keywords: apoptosis, Fas-L, multiple myeloma, osteoblasts, TRAIL. Multiple myeloma (MM) is characterized by marrow expan- sion of a malignant plasma cell clone, resulting in the gradual replacement of the haematopoietic matrix and dissemination of osteolytic lesions as nests of tumour cells (Hallek et al, 1998; Callander & Roodman, 2001) 1 . Malig- nant plasma cells are apparently engaged in the deregula- tion of bone resorption by inducing stromal cells to secrete a number of osteoclast activating factors, such as interleukin 1b (IL-1b), IL-6 and tumour necrosis factor (TNF)-b (Michigami et al, 2000). These cytokines stimulate osteo- blasts to enhance the production of trans-membranous RANKL [receptor activator of nuclear factor (NF)-kB ligand] (Lacey et al, 1998; Yasuda et al, 1998), a major osteoclas- togenic factor that may also be directly secreted by myeloma cells (Oyajobi et al, 1998). However, its functional expres- sion by malignant plasma cells remains controversial (Croucher et al, 2001; Giuliani et al, 2001; Pearse et al, 2001; Sezer et al, 2002). Loss of bone in MM is also explained by inadequate regeneration, due to the inhibition of osteoblast function, as there is no evidence of new bone formation within the skeleton lesions or in their vicinity. Osteoblasts are differ- entiated from fibroblasts by the activation of specific genes encoding osteocalcin, a secreted molecule that modulates osteoblast function (Ducy et al, 1996). There is much recent evidence to suggest that these cells are highly sensitive to apoptosis in response to treatment with either growth factors or inflammatory cytokines (Jilka et al, 1998). Upregulated osteoblast apoptosis may thus contribute to both the defective repair of resorption cavities and poor bone remodelling that is detected in a number of chronic disorders. This assumption is supported by evidence that IL-6 and TNFa, secreted by the rheumatoid arthritis synovium, suppress new bone apposition while inducing subchondral bone resorption by RANKL overproduction (Miyasaka et al, 1988; Arend & Dayer, 1990). In addition, TNFa is a direct inducer of apoptosis in murine osteoblasts in vitro (Kitajima et al, 1996). Correspondence: Franco Silvestris, MD, DIMO, University of Bari, Section of Internal Medicine and Clinical Oncology, P.za Giulio Cesare, 11, 70124 Bari, Italy. E-mail: [email protected] British Journal of Haematology, 2003, 122, 39–52 Ó 2003 Blackwell Publishing Ltd 39

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Upregulation of osteoblast apoptosis by malignant plasma cells:
a role in myeloma bone disease
Franco Silvestris, Paola Cafforio, Marco Tucci, Daniela Grinello and Franco Dammacco
Department of Internal Medicine and Oncology (DIMO), University of Bari, Bari, Italy
Received 8 November 2002; accepted for publication 10 February 2003
Summary. Typical features of multiple myeloma (MM) areosteolytic lesions and severely affected bone regeneration.This study of 53 MM patients demonstrates an enhance-ment of osteoblast cytotoxicity by malignant myeloma cellsvia the upregulation of apoptogenic receptors, including Fasligand (Fas-L) and tumour-necrosis-factor-related apoptosisinducing ligand (TRAIL). Both were significantly increasedin the marrow myeloma cells of patients with extensiveosteolytic lesions in a fashion similar to the highly malig-nant human myeloma cell line MCC-2. Osteoblasts fromthese subjects over-expressed Fas and death receptor (DR)4/5 and underwent dramatic apoptosis when co-culturedwith either MCC-2 or autologous myeloma cells. In osteo-blast and myeloma cell co-cultures, monocyte chemoat-tractant protein 1 (MCP-1) mRNA was upregulated inosteoblasts from patients with severe bone disease in parallelwith increased CC-chemokine receptor R2 (CCR2) expres-
sion, the ligand of MCP-1, in the myeloma cells. Thischemokine was shown to activate malignant cell migrationin vitro. An upregulation of ICAM-1 expression occurred inosteoblasts from patients with active skeleton disease. Thisupregulation appeared to be an effect of malignant plasmacell contact, as MCC-2 co-culture greatly enhanced ICAM-1production by resting osteoblasts from patients withoutskeleton involvement. Our results suggest that osteoblasts inactive myeloma are functionally exhausted and promptlyundergo apoptosis in the presence of myeloma cells frompatients with severe bone disease. It is suggested that thiscytotoxic effect plays a pivotal role in the pathogenesis ofdefective bone repair.
Keywords: apoptosis, Fas-L, multiple myeloma, osteoblasts,TRAIL.
Multiple myeloma (MM) is characterized by marrow expan-sion of a malignant plasma cell clone, resulting in thegradual replacement of the haematopoietic matrix anddissemination of osteolytic lesions as nests of tumour cells(Hallek et al, 1998; Callander & Roodman, 2001)1 . Malig-nant plasma cells are apparently engaged in the deregula-tion of bone resorption by inducing stromal cells to secrete anumber of osteoclast activating factors, such as interleukin1b (IL-1b), IL-6 and tumour necrosis factor (TNF)-b(Michigami et al, 2000). These cytokines stimulate osteo-blasts to enhance the production of trans-membranousRANKL [receptor activator of nuclear factor (NF)-kB ligand](Lacey et al, 1998; Yasuda et al, 1998), a major osteoclas-togenic factor that may also be directly secreted by myelomacells (Oyajobi et al, 1998). However, its functional expres-sion by malignant plasma cells remains controversial
(Croucher et al, 2001; Giuliani et al, 2001; Pearse et al,2001; Sezer et al, 2002).
Loss of bone in MM is also explained by inadequateregeneration, due to the inhibition of osteoblast function, asthere is no evidence of new bone formation within theskeleton lesions or in their vicinity. Osteoblasts are differ-entiated from fibroblasts by the activation of specific genesencoding osteocalcin, a secreted molecule that modulatesosteoblast function (Ducy et al, 1996). There is much recentevidence to suggest that these cells are highly sensitive toapoptosis in response to treatment with either growthfactors or inflammatory cytokines (Jilka et al, 1998).Upregulated osteoblast apoptosis may thus contribute toboth the defective repair of resorption cavities and poor boneremodelling that is detected in a number of chronicdisorders. This assumption is supported by evidence thatIL-6 and TNFa, secreted by the rheumatoid arthritissynovium, suppress new bone apposition while inducingsubchondral bone resorption by RANKL overproduction(Miyasaka et al, 1988; Arend & Dayer, 1990). In addition,TNFa is a direct inducer of apoptosis in murine osteoblastsin vitro (Kitajima et al, 1996).
Correspondence: Franco Silvestris, MD, DIMO, University of Bari,
Section of Internal Medicine and Clinical Oncology, P.za Giulio
Cesare, 11, 70124 Bari, Italy. E-mail: [email protected]
British Journal of Haematology, 2003, 122, 39–52
� 2003 Blackwell Publishing Ltd 39

The apoptosis of human osteoblasts is efficiently regulatedby the Fas/Fas ligand (Fas-L) pathway (Kawakami et al,1997). This mechanism is physiologically enrolled in thenegative control of cell proliferation during organogenesisand tissue homeostasis, whereas its downregulation ampli-fies the replication of several tumour cells by inhibiting theirapoptosis (Johnstone et al, 2002). Fas-L, a type II trans-membrane protein that clusters Fas to transduce deathsignals by cytoplasmic caspases, including CPP32 (caspase-3)and Fas-associated death domain protein (FADD)-likeinterleukin-1 beta-converting enzyme (FLICE) (caspase-8)(Muzio et al, 1996), is functionally expressed by activatedlymphocytes (CD8 and CD4 Th1 subsets), natural killer(NK) cells, erythroblasts and immune-privileged tissues(Griffith et al, 1995; De Maria et al, 1999). However, anumber of tumour cell types (Hahne et al, 1996; O’Connelet al, 1996), including malignant myeloma cells, alsoupregulate Fas-L expression to counterattack Fas+ effectorcells committed to immune surveillance (Villunger et al,1997).
Fas-L is over-expressed by myeloma cells (Frassanito et al,1998) to promote the tumour expansion in bone marrow,to the detriment of the haematopoietic matrix (Silvestriset al, 2001). The mechanism operates through the inordin-ate presentation of Fas-L and other apoptogenic receptors,including TNF-related apoptosis inducing ligand (TRAIL)which is a member of the TNF family (Wiley et al, 1995), tosensitive erythroblasts, resulting in the chronic impairmentof erythropoiesis and severe anaemia (Silvestris et al, 2002).The present study explored whether a similar mechanism isdetrimental to osteoblast function in invasive MM. Wefound that osteoblasts from the majority of patients withactive bone disease are highly susceptible to apoptogenicstimuli and promptly undergo apoptosis in vitro afterinteraction with malignant plasma cells. This may contrib-ute to the chronic defect of bone repair and loss observed inMM.
MATERIALS AND METHODS
Patients. Twenty-eight patients with invasive stage III(Durie & Salmon, 1975) MM were selected because of theirsevere skeleton involvement, arbitrarily defined as multiplelytic lesions in three or more bone sites (group A). Twenty-five patients with no evidence of osteolysis and initialdisease (stage I) were also enrolled (group B). The total MMpatient population was aged between 49 and 76 years withsimilar median ages (64Æ5 years for group A and 66Æ1 yearsfor group B). Biological samples included osteoblastsobtained from bone biopsy primary cultures and marrowplasma cells derived from needle aspirates. All subjects wereproperly informed and gave their consent to the study,which was approved by the Ethical Committee of theUniversity of Bari.
Plasma cell and osteoblast preparations. Bone marrowplasma cells were enriched from needle aspirates bymagnetic sorting using both anti-CD38 and anti-syndecan-1/CD138 (Dhodapkar et al, 1998) monoclonal antibody(mAb)-conjugated microbeads (Miltenyi Biotec, Bergisch
Gladbach, Germany). This method provided the excellentenrichment of myeloma cells (> 96Æ5% of j/k chain-positivecells), as detected by flow cytometry (FACScan; Becton-Dickinson, Mountain View, CA, USA). The cells were theninvestigated using a double fluorescence assay for theexpression of syndecan-1 (Becton-Dickinson), Fas-L andTRAIL by using the biotinylated immunoglobulin G (IgG)1jfrom the NOK-1 clone (PharMingen, San Diego, CA, USA)treated with phycoerythrin (PE)-conjugated streptavidin forFas-L, and by rabbit antiserum for TRAIL (Alexis). Thesemarkers were also measured in myeloma cell lines, inclu-ding U-266 and RPMI-8226 from the American TypeCulture Collection (ATCC, Rockville, MD, USA), MCC-2 andMCC-4 (Frassanito et al, 1998), which were used infunctional tests as cellular models of human myelomasalong with myeloma clones obtained from several patientsof both groups. Patient primary plasma cells were expandedin vitro in the presence of IL-6, as previously described(Frassanito et al, 1998).
Osteoblasts were derived from iliac crest trabecular bonebiopsies. Bone fragments were digested with 1 mg/ml typeI bacterial collagenase (Sigma-Aldrich, Milan, Italy) ina-minimum essential medium (MEM) (GibcoBRL, LifeTechnologies, Milan, Italy) and cultured in flasks incomplete medium (a-MEM plus 10% fetal calf serum and2 mmol/l glutamine) for up to 4–6 weeks to obtain aconfluent cell monolayer. The production of both osteo-calcin and alkaline phosphatase (Molecular Probes, Eu-gene, OR, USA), as phenotypic markers of osteoblastmaturation, was measured (Robey & Termine, 1985).Next, the osteoblasts were recovered by a 0Æ25% trypsin/EDTA solution (Gibco) and characterized for the expressionof apoptogen receptors, including Fas, and both DR4 andDR5 (Alexis, Vinci, Italy). The latter receptors, the TRAILco-receptors, were also measured after treatment withTNFa and interferon gamma (IFNc) in an attempt toresemble the osteoblast chronic stimulation occurringin vivo in MM due to the increased levels of thoseinflammatory cytokines. In accordance with previousstudies (Fellenberg et al, 1997), osteoblasts were separatelyincubated for 24 h with 500 U/ml IFNc or 100 U/mlTNFa (Peprotech), then washed and further investigatedby flow cytometry to assess their susceptibility to apoptosisand/or in functional tests. Primary osteoblast cultures from11 healthy donors aged between 38 and 67 years (medianage 59Æ2 years) undergoing post-traumatic surgery wereused as normal osteoblast controls. Trabecular bonespecimens from these subjects were treated to produce amonolayer of adherent osteoblasts.
Osteoblast apoptosis and co-cultures with myeloma cells.Osteoblasts were investigated for their susceptibility tospontaneous apoptosis in culture as well as to CH11prototypic agonist anti-Fas mAb (Alexis) at 1 lg/ml afterovernight incubation. In addition, osteoblasts wereco-cultured with myeloma cells to measure their respon-siveness to cellular contact with malignant cells in order toresemble a condition potentially occurring in vivo in MMpatients. Briefly, approximately 2 · 104)5 · 104 adherentosteoblasts were separately incubated overnight with each
40 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

myeloma cell line at 1/1 cellular ratio, and occasionallywith a relative autologous myeloma clone when available.
Osteoblast apoptosis was initially assessed by the detec-tion of subdiploid DNA-containing cell populations bypropidium iodide (PI) and by the TUNEL (TdT-mediateddUTP nick-end labelling) technique, which uses fluorescein-dUTP to label the DNA strand breaks during apoptosis. Thisassay was completed by flow cytometry, according to themanufacturer’s instructions (Boehringer-Mannheim, Milan,Italy). In addition, we evaluated FITC-conjugated annexin Von the membrane of apoptotic osteoblasts (Boehringer-Mannheim). In several co-cultures, the morphologicalchanges of nuclei leading to chromatin condensation weredirectly visualized by fluorescent microscope examinationusing specimens of osteoblasts–myeloma co-cultures treatedwith either 1 lg/ml of 4, 6-diamidino-2-phenylindole(DAPI), or according to the TUNEL technique.
Additional assessment of osteoblast apoptosis includedthe detection of several caspases. We measured the contentof CPP32 (caspase 3) and its active product derived by theproteolytic cleavage of poly(ADP)ribose polymerase (PARP),as well as FLICE (caspase-8). This analysis was completed byflow cytometry, using appropriate mAbs (Biosource, Cama-rillo, CA, USA), as previously described (Silvestris et al,2001).
Chemokine and integrin detection. The expression of mono-cyte chemoattractant protein 1 (MCP-1), ICAM-1 (CD54)and lymphocyte function-associated 1 (LFA-1) by flowcytometry and microscope analysis with specific PE- orFITC-conjugated mAbs was measured to investigate theirreciprocal interaction in co-cultures leading to cell adhesionbetween osteoblasts and plasma cells (Caltag Laboratories,Burlingame, CA, USA; Chemicon, Temecula, CA, USA;Alexis; and Sigma). For this investigation, a number ofosteoblast preparations from both groups of patients (sixfrom group A and five from group B) were selected, whosemyeloma cells were enriched and grown to provide cellclones. In addition, the relative values of the meanfluorescence intensity (MFI) of MCP-1 were evaluated asthe ratio between the mean fluorescence of positive cells andthe corresponding value at the cut-off point (isotypiccontrol).
The RiboQuant� Multi-Probe Rnase Protection Assay(RPA) system (PharMingen, San Diego, CA, USA) was usedfor mRNA quantification of several chemokines and theirreceptors in both osteoblasts and myeloma clones. We useddifferent sets of templates for chemokines [Lymphotactin(Ltn), regulated upon activation, normal T-cell expressed,and presumably secreted (RANTES) interferon-inducibleprotein 10 (IP-10), macrophage inflammatory protein(MIP)-1b, MIP-1a, MCP-1, IL-8, I-309] and receptors(CXCR1, CXCR2, CXCR3, CXCR4, BLR-1, BLR-2, V28) toobtain the T7 polymerase-directed synthesis of [32P]-labelled antisense RNA probes. Briefly, total RNA fromeither osteoblasts or myeloma cells was purified with acommercial kit (InVitrogen, Groningen, the Netherlands),and the probe sets were hybridized in excess to target RNAafter digestion with the RNases of free probes and othersingle-stranded RNA. The remaining RNase-protected
probes were resolved using a denaturing polyacrylamidegel electrophoresis (PAGE) assay and quantified by autora-diography. The intensity signal of each mRNA was corre-lated to the relative protected standard probe and the testwas normalized by using the GAPDH template.
Migration of plasma cells under chemokine stimulationwas evaluated as the percentage of input cells migratingthrough the transwell system, using polycarbonate inserts.Briefly, 1 · 106 cells were located in the upper chamber ofthe transwell system, whereas chemokines includingMCP-1, MIP-1a or MIP-1b were located in the lower one.We used increasing concentrations (0Æ01–1Æ2 lg/ml) inparallel with a positive control stromal-derived factor (SDF)-1.
Statistical analysis. Differences between means of groupsof data were calculated using the Student’s t-test and, inseveral instances, the non-parametric Mann–WhitneyU-test.
RESULTS
Plasma cell phenotyping and osteoblast functional assaysFigure 1 shows the expression of syndecan-1, Fas-L andTRAIL by primary plasma cells along with control cell lines(upper section). These were variably expressed within bothgroups of patients with apparent discrepancy concerningFas-L and TRAIL in particular. While syndecan-1 occurredat slightly higher mean values in plasma cells from patientsof group B compared with group A [mean ± standarddeviation (SD) 59Æ5 ± 9% vs 46Æ7 ± 11%], both apoptogenreceptors were largely expressed by myeloma cells of groupA. We found that the phenotype expression levels of theapoptogen receptors were threefold higher in Group A,compared with an only moderate presence of either Fas-L orTRAIL in plasma cells from group B (P < 0Æ05). Phenotypiccharacterization of myeloma cell lines also revealed differentresults. U-266, RPMI 8226 and MCC-4 showed the prevalentoccurrence of syndecan-1 and a moderate expression of theother molecules in a fashion comparable to that observed inpatients from group B. In particular, U-266 cells wereminimally positive for TRAIL (< 5%), whereas syndecan-1occurred at the highest levels (> 80%) in MCC-4.
This supported the apparent similarity of these cell lineswith plasma cells from MM patients that lacked the clinicalevidence of aggressive disease (group B patients). On thecontrary, MCC-2 expressed very high amounts of both Fas-Land TRAIL when compared with the other cell lines(P < 0Æ02). The parallelism in either the high or lowexpression of both receptors by different myeloma cell linesreflected a putative diversity in their malignancy, assuggested in previous work (Frassanito et al, 1998), descri-bing MCC-2 and MCC-4 as myeloma clones of high and lowmalignancy respectively. Therefore, myeloma cells frompatients of group A were arbitrarily assumed to be highlymalignant plasma cells.
Osteoblast function was also investigated. We primarilyassessed the osteoblast phenotype of cultured cells frombone biopsies by detecting both osteocalcin and alka-line phosphatase production, and found a comparableexpression of both markers between patients and control
Osteoblast Apoptosis in Multiple Myeloma 41
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

Fig 1. The phenotype analysis of myeloma cells and osteoblasts. (Upper panel) The different expression of syndecan-1, Fas-L and TRAIL
molecules by primary marrow plasma cells from patients with active and inactive myeloma with respect to the presence (group A) or absence
(group B) of multiple osteolytic lesions in skeleton (top left) in comparison with human myeloma cell lines, including U-266, RPMI-8226, MCC-2
and MCC-4 (top right). While syndecan-1 was expressed at comparable levels by primary cells and myeloma lines with the exception of MCC-2,
which showed a low presence of the antigen, both Fas-L and TRAIL occurred at significantly higher values in myeloma cells from patients with
bone focal disease than in control subjects (P < 0Æ05). A similar pattern was observed in the highly malignant myeloma cell line MCC-2 with
respect to the other cell lines (P < 0Æ02). (Lower panel) The levels of Fas, DR4 and DR5 in osteoblasts from myeloma patients of both groups A
and B, and from healthy control subjects showing both basal expression (base) and expression after treatment with inflammatory cytokines,
including TNFa and IFNc. The receptors were constitutively upregulated in osteoblasts from patients of group A (P < 0Æ02) and were
minimally affected by incubation with either TNFa or IFNc. By contrast, osteoblasts from the other patients and control subjects were highly
sensitive to both cytokines and significantly increased their receptor expression to a similar extent (P < 0Æ01 in all instances). Values are
mean ± SD of positive cells evaluated by double-fluorescence flow cytometry using appropriate mAbs. Pts ¼ number of patients.
42 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

subjects. Furthermore, a number of osteoblast preparationsfrom both patients and control subjects were measured fortheir expression of unusual apoptosis-related markers, ineither basal conditions or in response to inflammatorycytokines, such as TNFa and IFNc, which are usuallyupregulated in MM (Pisa et al, 1990). Figure 1 (lowersection) shows the mean values of Fas, DR4 and DR5expression by osteoblasts from both patients and controlsubjects. As can be seen, these receptors were variablyupregulated in patients, particularly in those from group A,who showed significantly higher basal levels when com-pared with group B and control subjects (P < 0Æ02 in allinstances). The most remarkable difference was in DR5expression, where the baseline value (82Æ1 ± 9%) wasnearly eightfold higher in group A than group B andapproximately 20-fold that of control subjects. However, werecorded a different expression of these receptors in responseto treatment with both TNFa and IFNc. These cytokineswere unable to induce clear-cut variations in osteoblastsderived from group A patients, in contrast with theconsiderable increases observed in the other culture groups.In particular, the Fas and DR4/DR5 expression of osteo-blasts from healthy donors was dramatically enhanced,resulting in a clear-cut increase of relative basal values(P < 0Æ01 in all instances), whereas a slightly lowerenhancement was recorded in group B (P < 0Æ03). There-fore, the constitutive upregulation of Fas, DR4 and DR5 byosteoblasts from patients with aggressive myeloma wasinterpreted to relate to the chronic in vivo stimulationobserved by those cytokines.
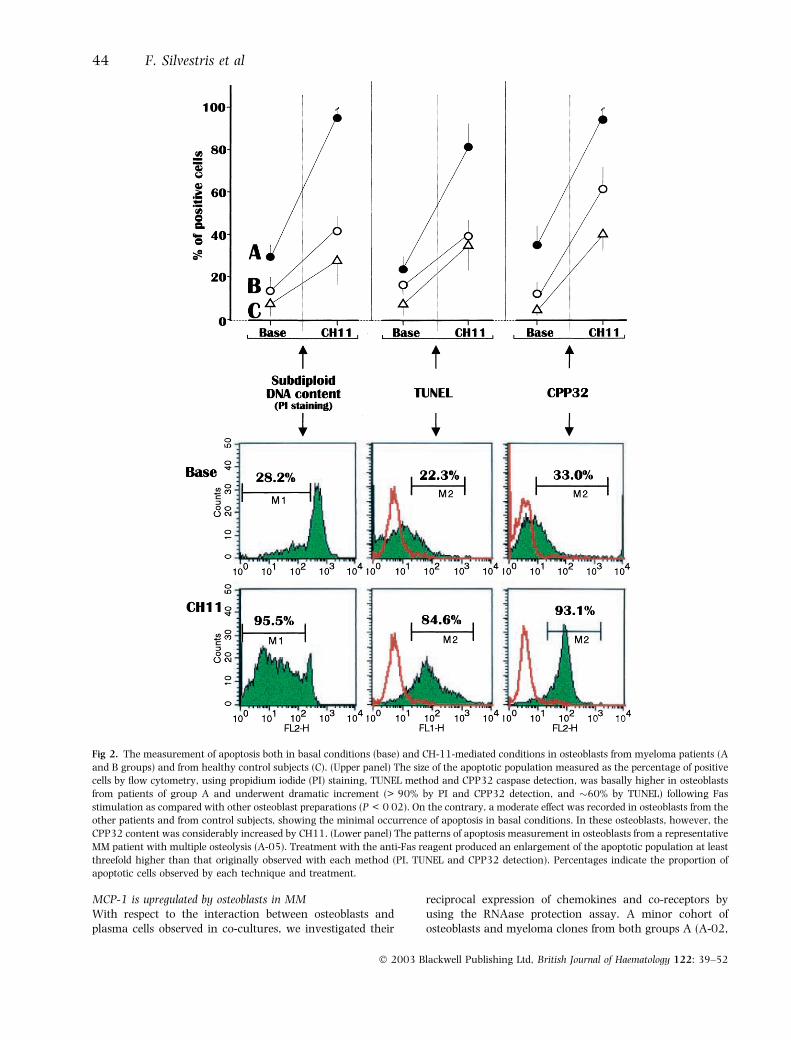
Osteoblasts from patients with active myeloma are proneto apoptosisWe also measured the osteoblast susceptibility to eitherspontaneous apoptosis or in response to stimulation byCH11 anti-Fas in vitro. Figure 2 (upper section) showsboth the basal and CH11-induced apoptosis levels inosteoblast preparations from patients of group A and B,and control subjects. As can be seen, osteoblasts fromgroup A showed the highest levels of both spontaneousand CH11-triggered cell death. The three parameters usedto measure apoptosis dramatically increased (P < 0Æ02) inosteoblasts from these patients after CH11 stimulation,thus suggesting their upregulated Fas sensitivity. Incontrast, the osteoblasts from group B, and to a lowerextent from control subjects, were moderately affected byanti-Fas treatment, with a variable rise in apoptosisobserved. However, the extent of cell death induced byCH11 in osteoblasts from group A was significantly higherthan the corresponding increases in the other groups ofcell preparations (P < 0Æ02 in all instances). The pattern ofvariation from a representative patient of group A isdepicted in Fig 2 (lower section). As shown, the CH11-driven effect was significantly evident on applying thethree methods, with values of apoptosis always more thanthreefold higher following treatment (P < 0Æ02).
These results suggested that osteoblasts from MM patientswith active bone disease are inherently prone to apoptosisin vitro, showing an upregulated responsiveness of Fas
compared with MM patients with no evidence of bone lyticinvolvement.
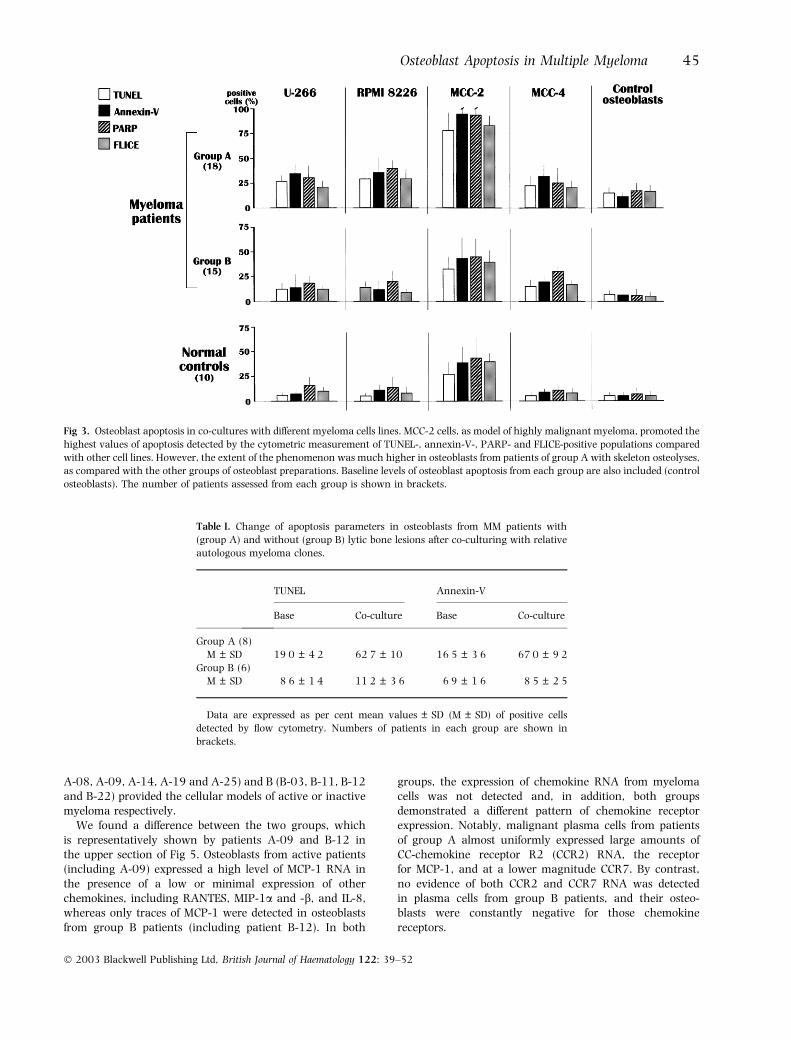
Osteoblasts are attacked in vitro by malignant myeloma cellsIn order to explore the osteoblast susceptibility to apoptosis inthe presence of malignant plasma cells, a number of culturesfrom both patients and control subjects were supplementedwith each myeloma cell line and, in several instances, withautologous myeloma clones. After overnight incubation, theosteoblasts were harvested and analysed by flow cytometry todetect the TUNEL and annexin-V-positive osteoblast popula-tions, along with measurement of the caspase product,PARP, and FLICE. Figure 3 shows that the mean values ofthese parameters in each group of patients and controlsubjects appeared considerably different both between thecell lines and between groups of osteoblast preparations.Indeed, a consistent enhancement of osteoblast apoptosisoccurred in co-cultures including MCC-2 cells when com-pared with other myeloma cell lines. In contrast with themoderate effect of other myeloma cell lines, MCC-2 induceddramatic cell death, in particular in co-cultures with osteo-blasts from group A, with up to 100% of apoptosis observed.A significantly lower effect was observed in cultures fromgroup B and control subjects (P < 0Æ03), which showedapoptosis to a mild extent when assessed by all methods.This finding supports the intrinsic propensity of osteoblastsfrom patients with active MM to undergo apoptosis whenco-cultured with malignant plasma cells overexposing Fas-Land TRAIL, such as MCC-2. However, although no assayswere performed to assess the inhibition of either Fas-L orTRAIL on these cells, the minimal levels of osteoblastapoptosis induced by U-266, RPMI 8226 and MCC-4, asmodels of Fas-L/TRAIL poorly expressing cells, supported thefunctional effect of those receptors exposed by MCC-2.
Table I illustrates the mean values of osteoblast apoptosisin co-cultures with autologous myeloma cells from eightpatients of group A (A-02, A-05, A-08, A-09, A-14, A-19,A-23 and A-25), and six of group B (B-03, B-05, B-11,B-12, B-16 and B-22). Again, the majority of osteoblastcultures from group A underwent the significant enhance-ment of apoptosis above basal levels, detected by bothTUNEL and annexin V (P < 0Æ02 in both instances). On thecontrary, those from group B were either only marginallyaffected or completely refractory to the effects of co-culturewith autologous myeloma clones.
Figure 4 shows the apoptotic patterns of osteoblastsderived from representative patients from groups A and B(patients A-09 and B-12) that were co-cultured with auto-logous plasma cells gated in the flow cytometric analysisaccording to both cell size and side scatter distribution(R1: plasma cells; R2: osteoblasts). As can be seen, theincubation of osteoblasts from patient A-09 with autologousmyeloma cells induced a dramatic enhancement of celldeath by both TUNEL and annexin V measurements. Thisphenomenon was not observed in the co-culture frompatient B-12, whose osteoblasts remained virtually unaffec-ted. These results emphasized the substantial vulnerabilityof osteoblasts to malignant myeloma cells, particularlyduring active bone disease.
Osteoblast Apoptosis in Multiple Myeloma 43
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52
MCP-1 is upregulated by osteoblasts in MMWith respect to the interaction between osteoblasts andplasma cells observed in co-cultures, we investigated their
reciprocal expression of chemokines and co-receptors byusing the RNAase protection assay. A minor cohort ofosteoblasts and myeloma clones from both groups A (A-02,
Fig 2. The measurement of apoptosis both in basal conditions (base) and CH-11-mediated conditions in osteoblasts from myeloma patients (A
and B groups) and from healthy control subjects (C). (Upper panel) The size of the apoptotic population measured as the percentage of positive
cells by flow cytometry, using propidium iodide (PI) staining, TUNEL method and CPP32 caspase detection, was basally higher in osteoblasts
from patients of group A and underwent dramatic increment (> 90% by PI and CPP32 detection, and �60% by TUNEL) following Fas
stimulation as compared with other osteoblast preparations (P < 0Æ02). On the contrary, a moderate effect was recorded in osteoblasts from the
other patients and from control subjects, showing the minimal occurrence of apoptosis in basal conditions. In these osteoblasts, however, the
CPP32 content was considerably increased by CH11. (Lower panel) The patterns of apoptosis measurement in osteoblasts from a representative
MM patient with multiple osteolysis (A-05). Treatment with the anti-Fas reagent produced an enlargement of the apoptotic population at least
threefold higher than that originally observed with each method (PI, TUNEL and CPP32 detection). Percentages indicate the proportion of
apoptotic cells observed by each technique and treatment.
44 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52
A-08, A-09, A-14, A-19 and A-25) and B (B-03, B-11, B-12and B-22) provided the cellular models of active or inactivemyeloma respectively.
We found a difference between the two groups, whichis representatively shown by patients A-09 and B-12 inthe upper section of Fig 5. Osteoblasts from active patients(including A-09) expressed a high level of MCP-1 RNA inthe presence of a low or minimal expression of otherchemokines, including RANTES, MIP-1a and -b, and IL-8,whereas only traces of MCP-1 were detected in osteoblastsfrom group B patients (including patient B-12). In both
groups, the expression of chemokine RNA from myelomacells was not detected and, in addition, both groupsdemonstrated a different pattern of chemokine receptorexpression. Notably, malignant plasma cells from patientsof group A almost uniformly expressed large amounts ofCC-chemokine receptor R2 (CCR2) RNA, the receptorfor MCP-1, and at a lower magnitude CCR7. By contrast,no evidence of both CCR2 and CCR7 RNA was detectedin plasma cells from group B patients, and their osteo-blasts were constantly negative for those chemokinereceptors.
Table I. Change of apoptosis parameters in osteoblasts from MM patients with
(group A) and without (group B) lytic bone lesions after co-culturing with relative
autologous myeloma clones.
TUNEL Annexin-V
Base Co-culture Base Co-culture
Group A (8)
M ± SD 19Æ0 ± 4Æ2 62Æ7 ± 10 16Æ5 ± 3Æ6 67Æ0 ± 9Æ2Group B (6)
M ± SD 8Æ6 ± 1Æ4 11Æ2 ± 3Æ6 6Æ9 ± 1Æ6 8Æ5 ± 2Æ5
Data are expressed as per cent mean values ± SD (M ± SD) of positive cells
detected by flow cytometry. Numbers of patients in each group are shown in
brackets.
Fig 3. Osteoblast apoptosis in co-cultures with different myeloma cells lines. MCC-2 cells, as model of highly malignant myeloma, promoted the
highest values of apoptosis detected by the cytometric measurement of TUNEL-, annexin-V-, PARP- and FLICE-positive populations compared
with other cell lines. However, the extent of the phenomenon was much higher in osteoblasts from patients of group A with skeleton osteolyses,
as compared with the other groups of osteoblast preparations. Baseline levels of osteoblast apoptosis from each group are also included (control
osteoblasts). The number of patients assessed from each group is shown in brackets.
Osteoblast Apoptosis in Multiple Myeloma 45
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

The differing MCP-1 signal in osteoblasts was alsoinvestigated by flow cytometry to evaluate the fluorescenceintensity of its expression. The lower section of Fig 5illustrates the relative values of the MFI. As can be seen, theosteoblast population from patient A-09 was almost totallypositive (95Æ6%) for MCP-1. Although the percentage ofpositive osteoblasts was only slightly lower in patient B-12,the fluorescence intensity value was much lower for patientB-12 compared with patient A-09 (MFI ratio: 4 vs 68)2 . Sucha difference in the expression of MCP-1 occurred in theother cell preparations from both groups A and B, and wasin agreement with the differing RNA signals detected by theprevious assay.
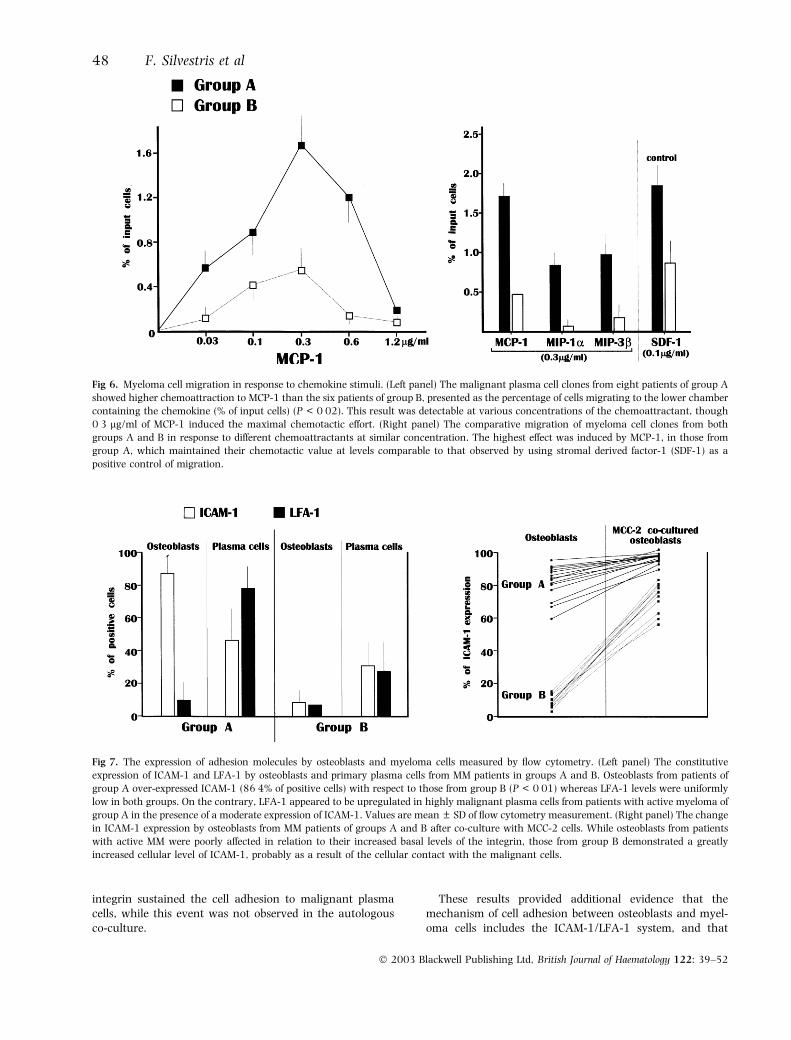
Because of the biological significance of the complement-ary expression of CCR2 and MCP-1 by plasma cells andosteoblasts, respectively, we evaluated the chemotacticability of these plasma cell preparations in response to anumber of chemoattractants, including MCP-1, MIP-1a andMIP-3b. Figure 6 shows mean values of migration bymyeloma cell clones after stimulation with MCP-1 (left side).The effect was dose dependent, and a major stimulation ofmigration was obtained by 0Æ3 lg/ml of MCP-1 in allassays. However, myeloma cells from group A patientsshowed a significantly higher level of migration, expressedas the percentage of input cells, than those of group B(P < 0Æ02). Therefore, we compared the promotion ofmigration elicited by MCP-1 with that of the otherchemokines MIP-1a and MIP-3b that are the ligands ofCCR1 and CCR7 respectively. Data from this assayconfirmed the strong chemotactic effect of MCP-1 for cellsof group A patients, when compared with other chemokines
(P < 0Æ02), which also failed to induce the migration cellsfrom group B patients (Fig 6, right side).
These data supported the reciprocal and functionalexpression of chemokines and ligands by malignant myel-oma cells and osteoblasts from patients with active disease,and provided evidence that these molecules are primarilyinvolved in both cellular attraction and migration in vitro.
Malignant myeloma cells upregulate ICAM-1 expressionin osteoblastsA further investigation explored the cell adhesion betweenmyeloma cells and osteoblasts. Using flow cytometry, theexpression of both ICAM-1 and LFA-1 in osteoblasts beforeand after the co-culture with MCC-2 cells was measured, aswell as in primary plasma cells. Figure 7 (left side) illustratesthe basal expression of these molecules. In particular,osteoblasts from patients of group A over-expressed ICAM-1(86Æ4% of positive cells), in contrast with only moderate tolow levels (P < 0Æ01) observed in group B (11Æ7%). Incontrast, no differences in the expression of LFA-1 wereobserved. A discrepancy was also recorded between thelevels of these molecules in plasma cells from the two patientgroups. Those from group A showed higher levels of LFA-1relative to the expression of ICAM-1, although the amountsof both integrins were greater in group A than group B, inapparent concordance with the postulated difference in themalignancy of these cell preparations.
In addition, co-culture with MCC-2 induced strikingresults (Fig 7, right side). We measured ICAM-1 expressionin 14 osteoblast preparations from group A and 11 fromgroup B, and observed a general trend for the upregulation
Fig 4. Variation of TUNEL- and annexin-V-positive populations in osteoblasts (R2) from example patients of groups A (A-09) and B (B-12)
before (base) and after co-culture with autologous myeloma clones (R1). The osteoblast population from patient A-09 showed a con-
siderable enhancement of apoptosis parameters as compared with B-12 as a control patient with inactive myeloma. The cytometric
analysis was completed by separating osteoblasts from myeloma cells in relation to different physical parameters, including both cell size
and side scatter. Gating of cells is shown in the far left panels. Percentages in the remaining histograms represent the percentages of
apoptotic cells.
46 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

of the integrin in presence of MCC-2 cells. However, basedon their intrinsic high expression of ICAM-1, osteoblastsfrom patients of group A were very little influenced,whereas those from group B demonstrated a dramaticincrease in ICAM-1 expression. Such a great enhancement(�48%) reflected the prompt activating effect of cell
adhesion, induced by the cellular contact between osteo-blasts and highly malignant myeloma cells. Figure 8shows the ultra-violet pattern of ICAM-1 expression byosteoblasts from patient B-05 after incubation with MCC-2cells (upper panel) as well as with autologous plasma cells(lower panel). As shown, a remarkable expression of the
Fig 5. (Upper panel) Chemokine and receptor mRNA quantification in both osteoblasts and myeloma cells from A-09 and B-12 patients as
representative models of MM with and without systemic bone disease, using the RiboQuant� Multi-Probe RPA system, PAGE and autoradi-
ography. Osteoblasts from the majority of patients from group A showed weak mRNA signals for most chemokines, except for MCP-1 which
was greatly upregulated as is shown for patient A-09, with respect to osteoblasts from the other patients, whereas no signals were detected in
plasma cells. By contrast, the mRNA of CCR2, the ligand of MCP-1, was highly expressed by myeloma cells in association with a minor CCR7
signal. No evidence of these receptors was detected in plasma cells from patients of group B, including patient B-12. (Lower panel) The flow
cytometric measurement of MCP-1 expression by cultured osteoblasts from both patients A-09 and B-12. In addition to the higher percentage
of MCP-1-positive cells, osteoblasts from patient A-09 showed a greater MFI than the modest level observed in patient B-12. Pt ¼ patient. The
percentage indicates the proportion of cells expressing MCP-1, and the MFI value is also shown. The red line represents the negative control on
the FL1 fluorescence channel, whereas the green peak includes the MCP-1-positive cells.
Osteoblast Apoptosis in Multiple Myeloma 47
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52
integrin sustained the cell adhesion to malignant plasmacells, while this event was not observed in the autologousco-culture.
These results provided additional evidence that themechanism of cell adhesion between osteoblasts and myel-oma cells includes the ICAM-1/LFA-1 system, and that
Fig 6. Myeloma cell migration in response to chemokine stimuli. (Left panel) The malignant plasma cell clones from eight patients of group A
showed higher chemoattraction to MCP-1 than the six patients of group B, presented as the percentage of cells migrating to the lower chamber
containing the chemokine (% of input cells) (P < 0Æ02). This result was detectable at various concentrations of the chemoattractant, though
0Æ3 lg/ml of MCP-1 induced the maximal chemotactic effort. (Right panel) The comparative migration of myeloma cell clones from both
groups A and B in response to different chemoattractants at similar concentration. The highest effect was induced by MCP-1, in those from
group A, which maintained their chemotactic value at levels comparable to that observed by using stromal derived factor-1 (SDF-1) as a
positive control of migration.
Fig 7. The expression of adhesion molecules by osteoblasts and myeloma cells measured by flow cytometry. (Left panel) The constitutive
expression of ICAM-1 and LFA-1 by osteoblasts and primary plasma cells from MM patients in groups A and B. Osteoblasts from patients of
group A over-expressed ICAM-1 (86Æ4% of positive cells) with respect to those from group B (P < 0Æ01) whereas LFA-1 levels were uniformly
low in both groups. On the contrary, LFA-1 appeared to be upregulated in highly malignant plasma cells from patients with active myeloma of
group A in the presence of a moderate expression of ICAM-1. Values are mean ± SD of flow cytometry measurement. (Right panel) The change
in ICAM-1 expression by osteoblasts from MM patients of groups A and B after co-culture with MCC-2 cells. While osteoblasts from patients
with active MM were poorly affected in relation to their increased basal levels of the integrin, those from group B demonstrated a greatly
increased cellular level of ICAM-1, probably as a result of the cellular contact with the malignant cells.
48 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

malignant plasma cells may elicit and reinforce the integrinexpression in osteoblasts. However, further adhesion mech-anisms cannot be excluded as preliminary experimentsusing MCC-2 cells pretreated with soluble ICAM-1, tosaturate LFA-1, failed to prevent their adhesion to osteo-blasts (data not shown).
DISCUSSION
Multiple osteolytic lesions in MM are associated withexcessive bone resorption, as well as inadequate boneformation and remodelling (Callander & Roodman, 2001)3 .Here, we investigated osteoblast function in relation to theover-expression of apoptogenic receptors by malignantplasma cells and found that the latter greatly enhanceapoptosis in osteoblasts. Therefore, osteoblast cytotoxicityby myeloma cells may account for the defective boneremodelling observed in MM and may contribute tomyeloma bone disease.
Pathophysiological mechanisms of osteolysis focus on thepredominant role of osteoclasts in the formation of myelomaosteolytic lesions (Caligaris-Cappio et al, 1991). The growth
of myeloma cells in the experimental severe combinedimmunodeficiency (SCID) human system, as well as in MMpatients, is associated with the increased differentiation ofosteoclasts (Yacobby et al, 1998) as a result of theirinteraction with stromal cells (Chauhan et al, 1996). Theirreciprocal stimulation by active molecules (Michigami et al,2000) promotes the major secretion of a number ofcytokines, including IL-6 (Nilsson et al, 1999), IL-1 (Lacyet al, 1999), TNFa (Garrett et al, 1987), macrophagecolony-stimulating factor (Bataille et al, 1992), parathyroidhormone-related protein (Suzuki et al, 1994), hepatocytegrowth factor, IL-11 (Hjertner et al, 1999), MIP-1a (Choiet al, 2000) and metalloproteinases (Yacobby et al, 2002).Most of these factors are osteoclastogenic and upregulateRANKL production, resulting in the uncontrolled increase ofosteoclast differentiation by the monocyte–macrophage celllineage and, as recently suggested, also by myeloma clonesas these malignant plasma cells express specific markers ofosteoclasts, namely tartrate-resistant acid phosphatase(TRAP) and vitronectin (Faust et al, 1998).
Bone remodelling is also severely affected in myelomaskeleton disease. A striking histomorphometric difference
Fig 8. The ultra-violet fluorescence pattern of
ICAM-1 expression in osteoblasts from a MM
patient without bone lytic lesions (patient
B-05) after co-culture with MCC-2 cells (upper
panel) or with autologous plasma cells (lower
panel). Arrows highlight the expression of the
ICAM-1 molecule (green), and both MCC-2
and autologous plasma cells (blue) adhesion
to osteoclasts.
Osteoblast Apoptosis in Multiple Myeloma 49
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

between patients with and without lytic lesions is that bothosteoblasts and osteoblast function are defective in thosewith multiple osteolyses (Bataille et al, 1990a, 1991) asbiochemical indicators of osteoid production, such as serumosteocalcin, inversely correlate with the occurrence of lyticlesions (Bataille et al, 1990b; Taube et al, 1992). The failureof bone remodelling is confirmed by evidence showing thatfunctional osteoblasts are usually undetectable within lyticlesions.
Osteoblasts differentiate from fibroblasts and remainentrapped in bone matrix as osteocytes once their bone-forming function is completed. However, they are highlysusceptible to apoptosis by either TNFa (Kitajima et al,1996; Jilka et al, 1998) or Fas/Fas-L (Kawakami et al,1997) pathways, as most of them initially present at theremodelling sites usually die in response to the increasedsecretion of growth factors and cytokines produced withinthe bone microenvironment. We found that the majority ofosteoblasts from myeloma patients with multiple osteolyses(group A) constitutively expressed high levels of both Fasand DR4/DR5 molecules and, moreover, appeared to befunctionally exhausted. In contrast with control osteoblastsfrom either normal donors or patients without bone lesions,those from group A were unable to upregulate theirreceptors in response to cytokine stimulation, and promptlyunderwent Fas-induced apoptosis. These data emphasizedthat osteoblasts in active myeloma are highly sensitive toFas-L-, TNFa- and probably to TRAIL-mediated apoptosis,and that this susceptibility is related to their functionalexhaustion, presumably promoted in vivo by high cytokinelevels (Pisa et al, 1990). In this context, it has been reportedthat normal osteoblasts are also prone to a higher rate ofapoptosis in vitro in response to stimulation by inflamma-tory cytokines (Jilka et al, 1998; Tsuboi et al, 1999).
When culturing osteoblasts with Fas-L+/TRAIL+ MCC-2myeloma cells, we detected the dramatic occurrence ofapoptosis in osteoblasts from patients with severe skeletondisease. In these co-cultures, as in those prepared withautologous myeloma clones, the extent of apoptosis wasremarkable, and the osteoblasts were visibly attacked incultures by the malignant myeloma cells. The evidence thatthis devastating effect was mediated by myeloma cellsthrough Fas-L (and presumably TRAIL) was supported bythe concomitant accumulation of specific caspases and therelated products FLICE and PARP. Therefore, besides thepersistent stimulation of osteoblasts due to chronic cytokinerelease, our data emphasize a direct osteoblast cytotoxicitymediated by malignant plasma cells.
In order to elucidate the molecular events putativelyinvolved in the promotion of the cellular contact betweenmyeloma cells and osteoblasts leading to apoptosis in vivo,we investigated the function of several chemokines andintegrins. MCP-1 is a chemotactic glycoprotein belonging tothe CC chemokine family (Rossi & Zlotnik, 2000), which isvirtually absent in normal bone (Graves et al, 1999) butconsiderably expressed by osteoblasts of patients withrheumatoid arthritis (Lisignoli et al, 1999). We found astriking production of MCP-1 by osteoblasts and, concur-rently, the high accumulation of a CCR2 mRNA signal in
myeloma cells in the majority of patients with lytic bonelesions. As CCR2 is the natural chemokine receptor of MCP-1(Rossi & Zlotnik 2000), it is possible that its increasedexposition by activated osteoblasts strongly attracts theCCR2-positive myeloma cells. This effect was confirmed bythe measurement of chemotaxis, which demonstrated theability of myeloma cells to migrate in response to MCP-1chemoattraction (Fig 4B). In addition, the mild expressionof CCR7 in these cells, a receptor for lymph node T zoneCCL19 and CCL21 chemokines, is suggestive of theirmigration to bone marrow as a secondary lymphoid organ(Wehrli et al, 2001). Whether the upregulation of CCR2chemokine receptor is a peculiar defect of highly malignantmyeloma cells is presently under investigation in our lab,and has been recently proposed by others (Hargreaves et al,2001).
Besides chemoattraction, we investigated the ICAM-1/LFA-1 system as a major molecular mechanism involved inthe adhesion between osteoblasts and plasma cells inco-culture. Osteoblasts from inflamed bone usually expresshigh levels of ICAM-1, vascular cell adhesion molecule 1(VCAM-1), LFA-3 and other integrins that activate theproduction of osteoclastogenic factors, such as IL-1b and IL-6(Tanaka et al, 1995). The upregulation of ICAM-1 byosteoblasts from patients with active myeloma, as observedin the present study, is an expected effect of the chronicrelease of inflammatory cytokines (Pisa et al, 1990;Kurokouchi et al, 1998). However, the tremendous expres-sion of the integrin in osteoblasts from patients withoutbone lesions after co-incubation with MCC-2 myeloma cells,as model of highly malignant plasma cells, supports thehypothesis that the cellular contact itself enables ICAM-1over-expression, leading to cell adhesion. That this trigger-ing mechanism is activated by myeloma cells through LFA-1,namely the ICAM-1 ligand, which appeared upregulated inmyeloma cells from patients with bone lytic lesions, was notassessed in the present study. It is conceivable that othermolecules constitutively upregulated in malignant plasmacells also contribute to enhanced ICAM-1 expression bytheir cell contact with osteoblasts. In this context, the majorexpression of CD56/neural cell adhesion molecule (NCAM)has been described recently in malignant plasma cells frompatients with diffuse skeleton devastation (Ely & Knowles,2002).
Considering the importance of preserved osteoblastfunction in the repair of the hyperactive osteoclastresorption in MM, defective bone remodelling may play acritical role in bone loss and the progression of skeletondevastation. Our results indicate that a primary mechanismof osteoblast failure includes cytotoxic mechanisms pro-moted by the apoptogenic receptors of malignant myelomaclones. A similar mechanism involving Fas-L has beenrecently postulated in bone metastases of breast cancer(Fromigue et al, 2001).
ACKNOWLEDGMENTS
This work was supported in part by a grant from: AIRC(Associazione Italiana per la Ricerca sul Cancro), Milan; the
50 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

Italian Ministry of Education, Universities and Research(MIUR); and the Italian Ministry of Health, ISS (IstitutoSuperiore di Sanita), National Acquired ImmunodeficiencySyndrome (AIDS) Research Project, Rome, Italy.
REFERENCES
Arend, W.P. & Dayer, J.M. (1990) Cytokines and cytokine inhibitors
or antagonists in rheumatoid arthritis. Arthritis and Rheumatism,
33, 305–315.
Bataille, R., Chappard, D., Marcelli, C., Rossi, J.F., Dessauw, P.,
Baldet, P., Sany, J. & Alexandre, C. (1990a) Osteoblast stimula-
tion in multiple myeloma lacking lytic bone lesions. British
Journal of Haematology, 76, 484–487.
Bataille, R., Delmas, P.D., Chappard, D. & Sany, J. (1990b)
Abnormal serum bone Gla protein levels in multiple myeloma.
Crucial role of bone formation and prognostic implications.
Cancer, 66, 167–172.
Bataille, R., Chappard, D., Marcelli, C., Dessauw, P., Baldet, P.,
Sany, J. & Alexandre, C. (1991) Recruitment of new osteoblasts
and osteoclasts is the earliest critical event in the pathogenesis of
human multiple myeloma. Journal of Clinical Investigation, 88,
62–66.
Bataille, R., Chappard, D. & Klein, B. (1992) The critical role of
interleukin-6, interleukin-1B and macrophage colony-stimulat-
ing factor in the pathogenesis of bone lesions in multiple mye-
loma. International Journal of Clinical and Laboratory Research, 21,
283–287.
Caligaris-Cappio, F., Bergui, L., Gregoretti, M.G., Gaidano, G.,
Gaboli, M., Schena, M., Zambonin Zallone, A. & Marchisio, P.C.
(1991) Role of bone marrow stromal cells in the growth of
human multiple myeloma. Blood, 77, 2688–2693.
Callander, N.S. & Roodman, G.D. (2001) Myeloma bone disease.
Seminars in Hematology, 38, 276–285.
Chauhan, D., Uchiyama, H., Akbarali, Y., Urashima, M., Yama-
moto, K., Libermann, T.A. & Anderson, K.C. (1996) Multiple
myeloma cell adhesion-induced interleukin-6 expression in bone
marrow stromal cells involves activation of NF-kappa B. Blood,
87, 1104–1112.
Choi, S.J., Cruz, J.C., Craig, F., Chung, H., Devlin, R.D., Roodman,
G.D. & Alsina, M. (2000) Macrophage inflammatory protein
1-alpha is a potential osteoclast stimulatory factor in multiple
myeloma. Blood, 96, 671–675.
Croucher, P., Shipman, C.M., Lippitt, J., Perry, M., Asosingh, K.,
Hijzen, A., Brabbs, A.C., van Beek, E.J.R., Holen, I., Skerry,
T.M., Dunstan, C.R., Russell, G.R., Camp, B.V. & Vanderker-
ken, K. (2001) Osteoprotegerin inhibits the development of
osteolytic bone disease in multiple myeloma. Blood, 98, 3534–
3540.
De Maria, R., Testa, U., Luchetti, L., Zeuner, A., Stassi, G., Pelosi, E.,
Riccioni, R., Felli, N., Samoggia, P. & Peschle, C. (1999) Apop-
totic role of Fas/Fas ligand system in the regulation of erythro-
poiesis. Blood, 93, 796–803.
Dhodapkar, M.V., Abe, E., Theus, A., Lacy, M., Langford, J.K.,
Barlogie, B. & Sanderson, R.D. (1998) Syndecan-1 is a multi-
functional regulator of myeloma pathobiology: control of tumor
cell survival, growth, and bone cell differentiation. Blood, 91,
2679–2688.
Ducy, P., Desbois, C., Boyce, B., Pinero, G., Story, B., Dunstan, C.,
Smith, E., Bonadio, J., Goldstein, S., Gundberg, C., Bradley, A. &
Karsenty, G. (1996) Increased bone formation in osteoclast-
deficient mice. Nature, 382, 448–452.
Durie, B.M.G. & Salmon, S.E. (1975) A clinical staging system for
multiple myeloma. Cancer, 36, 842–854.
Ely, S. & Knowles, D.M. (2002) Expression of CD56/neural cell
adhesion molecule correlates with the presence of lytic bone
lesions in multiple myeloma and distinguishes myeloma from
monoclonal gammopathy of undetermined significance and
lymphomas with plasmacytoid differentiation. American Journal of
Pathology, 160, 1293–1299.
Faust, J., Hunt, P., Scully, S. & Shalhoub, V. (1998) Multiple
myeloma cells and cells of the human osteoclast lineage share
morphological and cell surface markers. Journal of Cellular Bio-
chemistry, 71, 559–568.
Fellenberg, J., Man, H., Scheverpflug, C., Ewerbeck, V. & Debatin,
K.-M. (1997) Modulation of resistance to anti-APO-1-induced
apoptosis in osteosarcoma cells by cytokines. International Journal
of Cancer, 72, 536–542.
Frassanito, A.M., Silvestris, F., Silvestris, N., Cafforio, P., Camarda, G.,
Iodice, G. & Dammacco, F. (1998) Fas/Fas ligand (Fas-L)-deregu-
lated apoptosis and IL-6 insensitivity in highly malignant myeloma
cells. Clinical and Experimental Immunology, 114, 711–720.
Fromigue, O., Kheddoumi, N., Lomri, A., Marie, P.J. & Body, J.J.
(2001) Breast cancer cells release factors that induce apoptosis in
human bone marrow stromal cells. Journal of Bone and Mineral
Research, 16, 1600–1609.
Garrett, I.R., Durie, B.G., Nedwin, G.E., Gillespie, A., Bringman, T.,
Sabatini, M., Bertolini, D.R. & Mundy, G.R. (1987) Production of
lymphotoxin, a bone-resorbing cytokine, by cultured human
myeloma cells. New England Journal of Medicine, 317, 526–532.
Giuliani, N., Bataille, R., Mancini, C., Lazzaretti, M. & Barille, S.
(2001) Myeloma cells induce imbalance in the osteoprotegerin/
osteoprotegerin ligand system in the human bone marrow
environment. Blood, 98, 3527–3533.
Graves, D.T., Jiang, Y. & Valente, A.J. (1999) The expression of
monocyte chemoattractant protein-1 and other chemokines by
osteoblasts. Frontiers in Bioscience, 4, 571–580.
Griffith, T.S., Brunner, T., Fletcher, S.M., Green, D.R. & Ferguson,
T.S. (1995) Fas-ligand induces apoptosis as a mechanism of
immune privilege. Science, 270, 1189–1192.
Hahne, M., Rimoldi, D., Schroter, M., Romero, P., Schreier, M.,
French, L.E., Scheneider, P., Bornand, T., Fontana, A., Lienard,
D., Cerottini, J. & Tschopp, J. (1996) Melanoma cell expression of
Fas (Apo-1/CD95) ligand: implications for tumor immune escape.
Science, 274, 1363–1366.
Hallek, M., Bergsagel, P.L. & Anderson, K.C. (1998) Multiple mye-
loma: increasing evidence for a multi-step transformation pro-
cess. Blood, 91, 3–21.
Hargreaves, D.C., Hyman, P.L., Lu, T.T., Ngo, V.N., Bidgol, A.,
Suzuki, G., Zou, Y.-R., Littman, D.R. & Cyster, J.G. (2001) A
coordinate change in chemokine responsiveness guides plasma
cell movements. Journal of Experimental Medicine, 194, 45–56.
Hjertner, Ø., Torgersen, M.L., Seidel, C., Hjort-Hansen, H., Waage,
A., Børset, M. & Sundan, A. (1999) Hepatocyte growth factor
(HGF) induces interleukin-11 secretion from osteoblasts: a pos-
sible role for HGF in myeloma-associated osteolytic bone disease.
Blood, 94, 3883–3888.
Jilka, R.L., Weinstein, R.S., Bellido, T., Parfitt, A.M. & Manolagas,
S.C. (1998) Osteoblast programmed cell death (apoptosis): mod-
ulation by growth factors and cytokines. Journal of Bone and
Mineral Research, 13, 793–802.
Johnstone, R.W., Ruefli, A.A. & Lowe, S.W. (2002) Apoptosis: a link
between cancer genetics and chemotherapy. Cell, 108, 153–164.
Kawakami, A., Eguchi, K., Matsuoka, N., Tsuboi, M., Kojit, T.,
Urayama, S., Fujiyama, K., Kiriyama, T., Nakashima, T., Nakane,
P.K. & Nagataki, S. (1997) Fas and Fas ligand interaction is
necessary for human osteoblast apoptosis. Journal of Bone and
Mineral Research, 12, 1637–1646.
Osteoblast Apoptosis in Multiple Myeloma 51
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52

Kitajima, I., Nakajima, T., Imamura, T., Takasaki, I., Kawahara, K.,
Okano, T., Tokioka, T., Soejima, Y., Abeyama, K. & Maruyama, I.
(1996) Induction of apoptosis in murine clonal osteoblasts
expressed by human T-cell leukemia virus type I tax by NF-kB
and TNF-a. Journal of Bone and Mineral Research, 11, 200–210.
Kurokouchi, K., Kambe, F., Yasukawa, K., Izumi, R., Ishiguro, N.,
Iwata, H. & Seo, H. (1998) TNFa increases expression of IL-6 and
ICAM-1 genes through activation of NF-kB in osteoblast-like
ROS17/2.8 cells. Journal of Bone and Mineral Research, 13, 1290–
1299.
Lacey, D.L., Timms, E., Tan, H.L., Kelley, M.J., Dunstan, C.R.,
Burgless, T., Elliott, R., Colombero, A., Elliott, G., Scully, S., Hsu,
H., Sullivan, J., Hawkins, N., Davy, E., Capparelli, C., Eli, A., Qian,
Y.X., Kaufman, S., Sarosi, I., Shalhoub, V., Senaldi, G., Guo, J.,
Delaney, J. & Boyle, W.J. (1998) Osteoprotegerin ligand is a
cytokine that regulates osteoclast differentiation and activation.
Cell, 93, 165–176.
Lacy, M.Q., Donovan, K.A., Heimbach, J.K., Ahmann, G.J. & Lust,
J.A. (1999) Comparison of interleukin-1 beta expression by in
situ hybridization in monoclonal gammopathy of undetermined
significance and multiple myeloma. Blood, 93, 300–305.
Lisignoli, G., Toneguzzi, S., Pozzi, C., Piacentini, M., Riccio, M.,
Ferruzzi, A., Gualtieri, G. & Facchini, A. (1999) Proinflammatory
cytokines and chemokine production and expression by human
osteoblasts isolated from patients with rheumatoid arthritis and
osteoarthritis. Journal of Rheumatology, 26, 791–799.
Michigami, T., Shimizu, N., Williams, P.J., Niewolna, M., Dallas,
S.L., Mundy, G.R. & Yoneda, T. (2000) Cell–cell contact between
marrow stromal cells and myeloma cells via VCAM-1 and a4b1-
integrin enhances production of osteoclast-stimulating activity.
Blood, 96, 1953–1960.
Miyasaka, N., Sato, K., Goto, M., Sasano, M., Natsuyama, M.,
Inonue, K. & Nishioka, K. (1988) Augmented interleukin pro-
duction and HLA-DR expression in the synovium of rheumatoid
arthritis patients: possible involvement in joint destruction.
Arthritis and Rheumatism, 31, 480–486.
Muzio, M., Chinnaiyan, A.M., Kischkel, F.C., O’Rourke, K., Shev-
chenko, A., Ni, J., Scaffidi, C., Bretz, J.D., Zhang, M., Gentz, R.,
Mann, M., Krammer, P.H., Peter, M.E. & Dixit, V.M. (1996)
FLICE, a novel FADD-homologous ICE/CED-3-like protease, is
recruited to CD95 (Fas/APO-I) death-inducing signaling complex.
Cell, 85, 817–827.
Nilsson, K., Georgii-Hemming, P., Spets, H. & Jernberg-Wiklund, H.
(1999) The control of proliferation, survival and apoptosis in
human multiple myeloma cells in vitro. Current Topics in Micro-
biology and Immunology, 246, 325–332.
O’Connell, J., O’Sullivan, J.K. & Shanahan, F. (1996) The Fas
counterattack: Fas-mediated T cell killing by colon cancer cells
expressing Fas ligand. Journal of Experimental Medicine, 184,
1075–1082.
Oyajobi, B.O., Traianedes, K., Yoneda, T. & Mundy, G.R. (1998)
Expression of rank ligand (RankL) by myeloma cells requires
binding to bone marrow stromal cells via an a4b1–VCAM)1
interaction. Bone, 23, s180.
Pearse, R.N., Sordillo, E.M., Yaccoby, S., Wong, B.R., Liau, D.F.,
Colman, N., Michaeli, J., Epstein, J. & Choi, Y. (2001) Multiple
myeloma disrupts the TRANCE/osteoprotegerin cytokine axis to
trigger bone destruction and promote tumor progression. Pro-
ceedings of the National Academy of Sciences of the United States of
America, 98, 11581–11586.
Pisa, P., Stenke, L., Bernell, P., Hansson, M. & Hast, R. (1990)
Tumor necrosis factor-a and interferon-c in serum of multiple
myeloma patients. Anticancer Research, 10, 817–820.
Robey, P.G. & Termine, J.D. (1985) Human bone cells in vitro.
Calcified Tissue International, 37, 453–460.
Rossi, D. & Zlotnik, A. (2000) The biology of chemokines and their
receptors. Annual Review of Immunology, 18, 217–242.
Sezer, O., Heider, U., Jakob, C., Eucker, J. & Possinger, K. (2002)
Human bone marrow myeloma cells express RANKL. Journal of
Clinical Oncology, 20, 353–354.
Silvestris, F., Tucci, M., Cafforio, P. & Dammacco, F. (2001) Fas-L
up-regulation by highly malignant myeloma plasma cells: role in
the pathogenesis of anemia and disease progression. Blood, 97,
1155–1164.
Silvestris, F., Cafforio, P., Tucci, M. & Dammacco, F. (2002) Negative
regulation of erythroblast maturation by Fas-L+/TRAIL+ highly
malignant plasma cells: a major pathogenetic mechanism of ane-
mia in progressing multiple myeloma. Blood, 99, 1305–1313.
Suzuki, A., Takahashi, T., Okuno, Y., Tsuyuoka, R., Sasaki, Y.,
Fukumoto, M., Katakami, H., Kita, K. & Shibuya, T. (1994)
Production of parathyroid hormone-related protein by cultured
human myeloma cells. American Journal of Hematology, 45,
88–90.
Tanaka, Y., Morimoto, I., Nakano, Y., Okada, Y., Hirota, S., Nom-
ura, S., Nakamura, T. & Eto, S. (1995) Osteoblasts are regulated
by their cellular adhesion through ICAM-1 and VCAM-1. Journal
of Bone and Mineral Research, 10, 1462–1469.
Taube, T., Beneton, M.N., McCloskey, E.V., Rogers, S., Greaves, M.
& Kanis, J.A. (1992) Abnormal bone remodeling in patients with
myelomatosis and normal biochemical indices of bone resorption.
European Journal of Haematology, 49, 192–198.
Tsuboi, M., Kawakami, A., Nakashima, T., Matsuoka, N., Urayama,
S., Kawabe, Y., Fujiyama, K., Kiriyama, T., Aoyagi, T., Maeda, K.
& Eguchi, K. (1999) Tumor necrosis factor-a and interleukin-1bincrease the Fas-mediated apoptosis of human osteoblasts. Journal
of Laboratory and Clinical Medicine, 134, 222–231.
Villunger, A., Egle, A., Marschitz, I., Kos, M., Bock, G., Ludwig, H.,
Geley, S., Kofler, R. & Greil, R. (1997) Constitutive expression of
Fas (Apo-1/CD95) ligand on multiple myeloma cells: a potential
mechanism of tumor-induced suppression of immune surveil-
lance. Blood, 90, 12–20.
Wehrli, N., Legler, D.F., Finke, D., Toellner, K.-M., Loetscher, P.,
Baggiolini, M., MacLennan, I.C.M. & Acha-Orbea, H. (2001)
Changing responsiveness to chemokines allows medullary plas-
mablasts to leave lymph nodes. European Journal of Immunology,
31, 609–616.
Wiley, S.R., Schooley, K., Smolak, P.J., Din, W.S., Huang, C.P.,
Nicholl, J.K., Smith, T.D., Rauch, C. & Smith, C.A. (1995) Iden-
tification and characterization of a new member of the TNF
family that induces apoptosis. Immunity, 3, 673–682.
Yacobby, S., Barlogie, B. & Epstein, J. (1998) Primary myeloma cells
growing in SCID-hu mice: a model for studying the biology and
treatment of myeloma and its manifestations. Blood, 92, 2908–
2913.
Yacobby, S., Pearse, R.N., Johnson, C.L., Barlogie, B., Choi, Y. &
Epstein, J. (2002) Myeloma interacts with the bone marrow
microenvironment to induce osteoclastogenesis and is dependent
on osteoclast activity. British Journal of Haematology, 116, 278–
290.
Yasuda, H., Shima, N., Nakagawa, N., Yamaguchi, K., Kinosaki, M.,
Mochizuki, S., Tomoyasu, A., Yano, K., Goto, M., Murakami, A.,
Tsuda, E., Morinaga, T., Higashio, K., Udagawa, N., Takahashi,
N. & Suda, T. (1998) Osteoclast differentiation factor is a ligand
for osteoprotegerin/osteoclastogenesis-inhibitory factor and is
identical to TRANCE/RANKL. Proceedings of the National Academy
of Sciences of the United States of America, 95, 3597–3602.
52 F. Silvestris et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 122: 39–52
Related Documents











