ORIGINAL PAPER Up-regulation of cytoskeletal-associated protein 2 in primary human gastric adenocarcinomas Received: 20 March 2003 / Accepted: 4 July 2003 / Published online: 26 August 2003 ȑ Springer-Verlag 2003 Abstract Background and method: We performed dif- ferential-display polymerase chain reaction to find up-regulated sequences in primary human gastric can- cers, and cloned one up-regulated sequence, which was expressed in all the gastric cancer cells that we examined. The cloned sequence was identified as cytoskeletal- associated protein 2 (CKAP2). We also cloned a shorter splice variant, CKAP2-s. The CKAP2 or CKAP2-s protein in HeLa cells was localized to microtubule organizing centers (MTOC) and microtubules. This co- localization pattern was disrupted by nocodazole, a microtubule-destabilizing agent. Results: These obser- vations suggested that CKAP2 might be associated with microtubule networks. CKAP2 protein was detected in neither normal GI tract nor normal gastric mucosa. However, both CKAP2 and CKAP2-s mRNAs were up-regulated in 55% (23 out of 42 samples) of primary human gastric cancers by reverse transcriptase-poly- merase chain reaction (RT-PCR). Moreover, CKAP2 proteins were detected in immunohistochemical staining in all the gastric cancer samples that we examined. CKAP2 protein-expressing cells were also found in gastric adenomas. The average number of CKAP2 protein-positive cells in adenocarcinomas was 48.8%, which was significantly higher than the number in tubular adenomas, 9.1%. Conclusion: When these points were taken together, we concluded that CKAP2 is up-regulated in primary human gastric adenocarcino- mas at high frequency and might be useful for diag- nosing and discriminating adenocarcinomas from tubular adenomas of the stomach. Keywords CKAP2 Up-regulation Gastric cancer DD-PCR Microtubule Introduction The gastric adenocarcinoma is one of the world’s leading cancers, particularly in the far eastern countries, including Korea, Japan and China. So far, many genetic alterations have been discovered in gastric cancers, which can be broadly divided into two groups, namely, hyper-functional and hypo-functional mutations. The J Cancer Res Clin Oncol (2003) 129: 621–630 DOI 10.1007/s00432-003-0484-0 Chang-Dae Bae Yeon-Sun Sung Sang-Min Jeon Yousin Suh Han-Kwan Yang Yong Il Kim Ki-Ho Park Jongsun Choi Geunghwan Ahn Joobae Park The nucleotide sequence data reported in this paper have been deposited with the GenBank database under the accession number AY062261 (CKAP2) and AY062262 (CKAP2-s) C.-D. Bae and Y.-S. Sung contributed equally to this paper C.-D. Bae S.-M. Jeon J. Park (&) Department of Molecular Cell Biology and Samsung Biomedical Research Institute, Sungkyunkwan University School of Medicine, 300 Chunchundong Jangangu, 440-746 Suwon, Korea E-mail: [email protected] Fax: 82-31-2996149 Y.-S. Sung Department of Biochemistry, Dankook University College of Medicine, Cheonan, Korea Y. Suh Department of Biochemistry and Molecular Biology, Seoul National University College of Medicine, Seoul, Korea H.-K. Yang Department of Surgery, and Cancer Research Institute, Seoul National University College of Medicine, Seoul, Korea Y. I. Kim Department of Surgery, Samsung Medical Center, Sungkyunkwan University School of Medicine, Seoul, Korea K.-H. Park Department of Biochemistry, Inje University College of Medicine, Busan, Korea J. Choi G. Ahn Department of Pathology, Samsung Medical Center, Sungkyunkwan University School of Medicine, Seoul, Korea

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Up-regulation of cytoskeletal-associated protein 2 in primaryhuman gastric adenocarcinomas
Received: 20 March 2003 / Accepted: 4 July 2003 / Published online: 26 August 2003� Springer-Verlag 2003
Abstract Background and method: We performed dif-ferential-display polymerase chain reaction to findup-regulated sequences in primary human gastric can-cers, and cloned one up-regulated sequence, which was
expressed in all the gastric cancer cells that we examined.The cloned sequence was identified as cytoskeletal-associated protein 2 (CKAP2). We also cloned a shortersplice variant, CKAP2-s. The CKAP2 or CKAP2-sprotein in HeLa cells was localized to microtubuleorganizing centers (MTOC) and microtubules. This co-localization pattern was disrupted by nocodazole, amicrotubule-destabilizing agent. Results: These obser-vations suggested that CKAP2 might be associated withmicrotubule networks. CKAP2 protein was detected inneither normal GI tract nor normal gastric mucosa.However, both CKAP2 and CKAP2-s mRNAs wereup-regulated in 55% (23 out of 42 samples) of primaryhuman gastric cancers by reverse transcriptase-poly-merase chain reaction (RT-PCR). Moreover, CKAP2proteins were detected in immunohistochemical stainingin all the gastric cancer samples that we examined.CKAP2 protein-expressing cells were also found ingastric adenomas. The average number of CKAP2protein-positive cells in adenocarcinomas was 48.8%,which was significantly higher than the numberin tubular adenomas, 9.1%. Conclusion: When thesepoints were taken together, we concluded that CKAP2 isup-regulated in primary human gastric adenocarcino-mas at high frequency and might be useful for diag-nosing and discriminating adenocarcinomas fromtubular adenomas of the stomach.
Keywords CKAP2 Æ Up-regulation Æ Gastric cancer ÆDD-PCR Æ Microtubule
Introduction
The gastric adenocarcinoma is one of the world’s leadingcancers, particularly in the far eastern countries,including Korea, Japan and China. So far, many geneticalterations have been discovered in gastric cancers,which can be broadly divided into two groups, namely,hyper-functional and hypo-functional mutations. The
J Cancer Res Clin Oncol (2003) 129: 621–630DOI 10.1007/s00432-003-0484-0
Chang-Dae Bae Æ Yeon-Sun Sung Æ Sang-Min Jeon
Yousin Suh Æ Han-Kwan Yang Æ Yong Il Kim
Ki-Ho Park Æ Jongsun Choi Æ Geunghwan Ahn
Joobae Park
The nucleotide sequence data reported in this paper have beendeposited with the GenBank database under the accession numberAY062261 (CKAP2) and AY062262 (CKAP2-s)
C.-D. Bae and Y.-S. Sung contributed equally to this paper
C.-D. Bae Æ S.-M. Jeon Æ J. Park (&)Department of Molecular Cell Biologyand Samsung Biomedical Research Institute,Sungkyunkwan University School of Medicine,300 Chunchundong Jangangu,440-746 Suwon, KoreaE-mail: [email protected]: 82-31-2996149
Y.-S. SungDepartment of Biochemistry,Dankook University College of Medicine,Cheonan, Korea
Y. SuhDepartment of Biochemistry and Molecular Biology,Seoul National University College of Medicine,Seoul, Korea
H.-K. YangDepartment of Surgery, and Cancer Research Institute,Seoul National University College of Medicine,Seoul, Korea
Y. I. KimDepartment of Surgery, Samsung Medical Center,Sungkyunkwan University School of Medicine,Seoul, Korea
K.-H. ParkDepartment of Biochemistry,Inje University College of Medicine,Busan, Korea
J. Choi Æ G. AhnDepartment of Pathology,Samsung Medical Center,Sungkyunkwan University School of Medicine,Seoul, Korea

over-expression of growth-related genes can be classifiedas hyper-functional in nature, while genetic abnormali-ties associated with tumor suppressors can be classifiedas hypo-functional.
Genetic abnormalities in the hypo-functional classare found more frequently in gastric cancers or othercancer types (Block et al. 1997; Werner et al. 2001).Inactivation of adenomatous polyposis coli (APC),resulting from loss of heterozygosity (LOH) or pointmutation, is found in 40�60% of intestinal-type gastriccancer (Sano et al. 1991; Tahara 1995) and in up to 30%of signet-ring cell carcinomas (Nakatsuru et al. 1992).Inactivation of p53 is also the result of LOH or pointmutation and is detected in approximately 30% of gas-tric cancers. Mutations of E-cadherin, a key moleculefor cell–cell adhesion, are found in 50% of the diffusetype but not in intestinal-type gastric cancers (Oda et al.1994; Becker et al. 1994; Berx et al. 1998; Hirohashi1998). However, the genetic alterations found in thehypo-functional class are usually not confined to gastriccancers and are frequently found in other types of can-cer. APC mutations were originally found in coloncancer, and p53 mutations are not related to specificcancer types.
Genetic alterations in the hyper-functional class arefound less frequently and less consistently in primaryhuman gastric cancers. These mutations usually resulteither from the up-regulation, or from mutations leadingto the activation of growth-related genes. Amplificationand/or overexpression of c-erbB-2 is found in ca.20�40% of the intestinal type of gastric cancer (Ranzaniet al. 1992; Werner et al. 2001), and amplification ofc-met or K-sam is found in approximately 30% of thediffuse type of gastric cancer (Kuniyasu et al. 1992;Tahara 1993). Amplification of cyclin E is seen in lessthan 20% and mutation of K-ras in 10% of gastriccancers (Akama et al. 1995; Kihana et al. 1991).Although the genetic alterations in the hyper-functionalclass are found much less frequently than those in thehypo-functional class, they show some relationshipswith certain cancer types, as seen in c-erbB-2 and breastcancers (Bouchard et al. 1989). For this reason wefocused on identifying hyper-functional genetic altera-tions and screened up-regulated sequences in primaryhuman gastric cancers using the differential-displaypolymerase chain reaction technique.
In this study, we reported a cytoskeletal associatedprotein, CKAP2, as a gene up-regulated in primaryhuman gastric cancers in high frequency. CKAP2 pro-tein was expressed only in tumors, and not in normalmucosa in the immunohistochemical staining. More-over, the population of cells expressing CKAP2 proteinwas significantly higher in gastric adenocarcinomas thanin tubular adenomas. We also identified the microtubulenetwork as the cytoskeletal component associated withCKAP2. Microtubules play essential roles in mitosis andintracellular transport. In particular, mitotic spindles arevery dynamic structures and must be properly regulatedduring the cell cycle. Cancer cells may need to adjust
microtubule dynamics to maintain their growth prop-erties. Therefore, it would be of interest to know whetherCKAP2 protein is related to the control of the cell cycleand cell proliferation.
Materials and methods
Cell culture and transfection
SNU-1, 5, 16, 387, 423, 449, 475, 601, 668, 719, and KATO III werepurchased from the Korean Cell Line Bank (Seoul, Korea) andcultivated in RPMI 1640 containing 10% fetal bovine serum(Invitrogen, Carlsbad, Calif., USA) under 5% CO2. HEK293 andHeLa cells were purchased from ATCC (Manassas, Va., USA) andcultivated in DMEM containing 10% fetal bovine serum under 5%CO2. For transfection, 5·105 cells were plated on a 6-well plate24 h prior to transfection. Three micrograms of DNA mixed with2 ll of Lipofectamine 2000 (Invitrogen) was overlaid onto the cells,in accordance with the manufacturer’s instructions. The transfectedcells were fixed or lysed after 24 h for subsequent immunofluores-cence staining or Western blotC.
Differential-display polymerase chain reaction
We isolated total cellular RNA from the mucosa of stomach cancertissues and the adjacent normal mucosa, using the ToTALLYRNA total RNA isolation kit (Ambion, Austin, Tex., USA). Forthe elimination of possible genomic DNA contamination, isolatedtotal RNA was subjected to RNase-free DNase (Invitrogen)digestion in the presence of RNasin (Promega, Madison, Wis.,USA). The absence of genomic DNA contamination was confirmedby the absence of the amplification of the glyceraldehyde 3-phos-phate dehydrogenase (G3PDH) gene by PCR using genomicG3PDH specific primers. Differential-display polymerase chainreaction (DD-PCR) was performed with the RNAimage kit(GeneHunter, Nashville, Tenn., USA) in accordance with themanufacturer’s recommendations. Up-regulated DNA bands wereexcised from the dried gel and re-amplified with the same primer setas was used in DD-PCR. The re-amplified PCR products werecloned into pCR 2.1-TOPO vectors via the TOPO TA Cloningsystem (Invitrogen) and sequenced with the BigDye Terminatorcycle sequencing ready reaction DNA sequencing Kit (Perkin-Elmer, Foster City, Calif., USA).
Rapid amplification of cDNA ends
Both ends of the DD-PCR products were cloned with the SMARTrapid amplification of cDNA ends (RACE) cDNA amplificationkit (Clontech, Palo Alto, Calif., USA) in accordance with themanufacturer’s instruction. Briefly, 1st strand cDNA was synthe-sized from poly A+ RNA from HeLa cells. For 5¢ RACE, a shortsequence in EST clone (GeneBank accession number AA807729),AGAGGGCGCAGTCCGCATTC was used as a CKAP2-specificprimer, and TGTTCCTTGAGTTTTTGTCTTCTTTGCT wasused for 3¢ RACE.
Northern blot
To determine the expression level of CKAP2 in human tissues, weused multiple tissue Northern blot (Clontech), while for stomachcancer cell lines, 20 lg of total cellular RNA was fractionatedthrough 1% agarose containing 2.2 mol/l of formaldehyde andtransferred to Hybond N membrane (Amersham PharmaciaBiotech, Piscataway, N.J., USA). Whole ORF of CKAP2 orb-actin probe (Clontech) was labeled with 32P-dCTP (6,000Ci/mmol, Amersham Pharmacia) via the Megaprime labeling kit
622

(Amersham Pharmacia). The RNA blot was hybridized to the 32P-labeled probe with Hybrisol I (Intergen, Purchase, N.Y., USA) at42�C overnight, and washed three times with 0.1· SSC at 65�C. Thehybridized blot was exposed to Kodak X-Omat AR film overnightat )70�C or exposed to a BAS phosphor storage screen (Fuji Film,Tokyo, Japan).
Reverse transcriptase-polymerase chain reaction of CKAP2and its splice variant, CKAP2-s
Total RNA was extracted from tumor tissues or cultured cells witheither the ToTALLY RNA total RNA isolation kit (Ambion) orTrizol (Invitrogen) and converted into cDNA by thermostablereverse transcriptase (Invitrogen). A 570-bp sequence from CKAP2mRNA was amplified by Taq polymerase (Perkin-Elmer) using anupstream primer (5¢-ATTGAAGAGATGCGACACAC) and adownstream primer (5¢-TTATGTTGTATCAGCCTCATA) for 35to 40 cycles of 95�C for 30 s, 55�C for 45 s and 72�C for 30 s. A348-bp fragment of short-splice variant of CKAP2, CKAP2-smRNA was amplified by Taq polymerase using an upstreamprimer (5¢-GATCTTTACAATTACAGTG) and a downstreamprimer (5¢-TGCGAATTTAATCCCTTTTG) for 50 cycles of 95�Cfor 30 s, 45�C for 45 s and 72�C for 30 s. A 302-bp sequence fromhuman G3PDH mRNA was amplified by Taq polymerase and5¢-CTTCAACAGCGACACCCACTCCTC as an upstream primerand 5¢-GGCCCCTCCCCTCTTCAA as a downstream primer for25 cycles of 95�C for 30 s, 55�C for 45 s and 72�C for 30 s. EachPCR product was subjected to 1% agarose gel electrophoresiscontaining 5 ng/ml ethidium bromide. The fluorescence of eachDNA band was quantified by BAS imaging system (Fuji Film).
Ribonuclease protection assay
A 448-bp CKAP2 fragment was amplified from SNU-5 cDNA byPCR with 5¢-CAACTTGTGCGACCTCCTATTA as an upstreamprimer and 5¢-CTCTTCCTTTGCCAGCTTTCCACT as a down-stream primer and sub-cloned into pGEM3z. The G3PDH PCRfragment, amplified by reverse transcriptase-polymerase chainreaction (RT-PCR), was also sub-cloned into pGEM3z. Theresulting constructs were linearized and transcribed in vitro with T7RNA polymerase (Promega) in the presence of 32P-CTP (6,000 Ci/mmol, Amersham). Each tissue RNA was hybridized simulta-neously with a mixture of 105 cpm of 32P-labeled CKAP2 antisenseprobe and 103 cpm of G3PDH antisense probe. After overnighthybridization the RNA samples were digested with 10 U of RNaseOne (Promega) and fractionated through 8% polyacrylamide gelcontaining 6 mol/l urea.
Polyclonal antibody preparation, Western blot
The whole ORF of CKAP2 was sub-cloned into pGEX-3X(Amersham Pharmacia) and expressed in E. coliDH5a overnight at16�C in the presence of 0.2 mmol/l of IPTG. The bacterial lysatewas obtained after 3 min of sonication in PBS and centrifugationfor 10 min at 4�C. The precipitate was washed five times with PBSand solubilized in 1·SDS sample buffer. The resulting solubilizedinclusion bodies were fractionated through 10% SDS-polyacryl-amide gel and electro-eluted into 50 fractions. Fractions containingGST-CKAP2 protein were pooled and dialyzed against PBS. Sol-uble GST-CKAP2 protein was purified with GSH-Sepharose(Amersham Pharmacia) and pooled with insoluble GST-CKAP2preparation. After rabbits had been treated with four injections ofpurified GST-CKAP2 protein at 2-week intervals, their serum wascollected by heart puncture.
For Western blot, the protein blot was incubated for 1 h atroom temperature with rabbit anti-CKAP2 polyclonal sera at adilution of 1:1,000. After incubation with anti-rabbit IgG-HRPconjugate (Amersham), the protein blot was incubated for 5 min
with WestPico reagent (Pierce, Rockford, Ill., USA). The Westernblot image was obtained via the LAS-1000 chemiluminescenceimaging system (Fuji Film).
Immunofluorescence staining
Cells were fixed with 3.7% formaldehyde for 10 min and permea-bilized with 0.5% Triton X-100 for 30 min. The fixed cells werethen incubated for 1 h in a mixture of anti-CKAP2 polyclonal seraat a dilution of 1:50 and anti-a-tubulin monoclonal Ab (Sigma) at adilution of 1:4000. Cy2-conjugated anti-rabbit IgG Ab (MolecularProbe, Eugene, Ore., USA) or Cy3-conjugated anti-mouse IgG(Rockland, Gilbertsville, Pa., USA) was incubated for 45 min atroom temperature to detect first Ab. The stained image wasobtained with either an AxioCam digital camera (Carl Zeiss,Germany) or an LSM 510 confocal microscope (Carl Zeiss).
Immunohistochemistry
Data from 11 cases of gastric adenocarcinoma and 17 cases oftubular adenoma of the stomach were retrieved from the files of theDepartment of Diagnostic Pathology at the Samsung MedicalCenter. The paraffin was removed from sections for immunohis-tochemical analysis, which were then rehydrated and rinsed in 3%hydrogen peroxide to remove endogenous peroxidase activity. Forantigen retrieval, slides were microwaved in near-boiling citratebuffer (0.01 mol/l, pH 6.0) for 20 min and then held at roomtemperature for 20 min and rinsed three times in citrate buffer(0.05 mol/l, pH 10.0). After being blocked with normal horse ser-um (Vector Laboratories, Burlingame, Calf., USA), the slides wereincubated with anti-CKAP2 Ab (dilution 1:100) overnight at 4�C.After being rinsed with PBS three times, the slides were incubatedfor 35 min in biotinylated horse anti-rabbit IgG (Vector, dilution1:200), then for 45 min in avidin–biotin complex (Dako, dilution1:100). The slides were then rinsed in PBS for 5 min and developedin stable diaminobenzidine (Research Genetics, Huntsville, Ala.,USA) for 2 to 3 min at room temperature and counterstained inhematoxylin for 15 s.
Positive percentages were defined as the percentage of cells thatshowed cytoplasmic staining over total tumor cells in a · 100power field. The staining intensity was divided into three grades(1+ to 3+). Statistical analysis was performed via theMann–Whitney test in SPSS 7.0.
Results
Up-regulated sequences in primary humanstomach cancers
To eliminate individual variations of gene expression wecompared five pairs of adenocarcinoma and matchednormal mucosa from surgically resected stomachs. Wecarefully resected only the mucosal area in gastric cancersamples, to minimize stromal contamination. Similarly,normal mucosa samples were obtained some distancefrom the cancer. In the first round of DD-PCR, wecloned 58 down-regulated (data not shown) and 13 up-regulated sequences, which expressed differentially in atleast three cancer tissues. Four of the up-regulatedsequences were highly homologous with sequencesfrom collagen, RNA helicase, ribosomal protein S24,and mannan-binding lectin (MBL)-associated serineprotease 2. Seven of the cloned DD-PCR products were
623
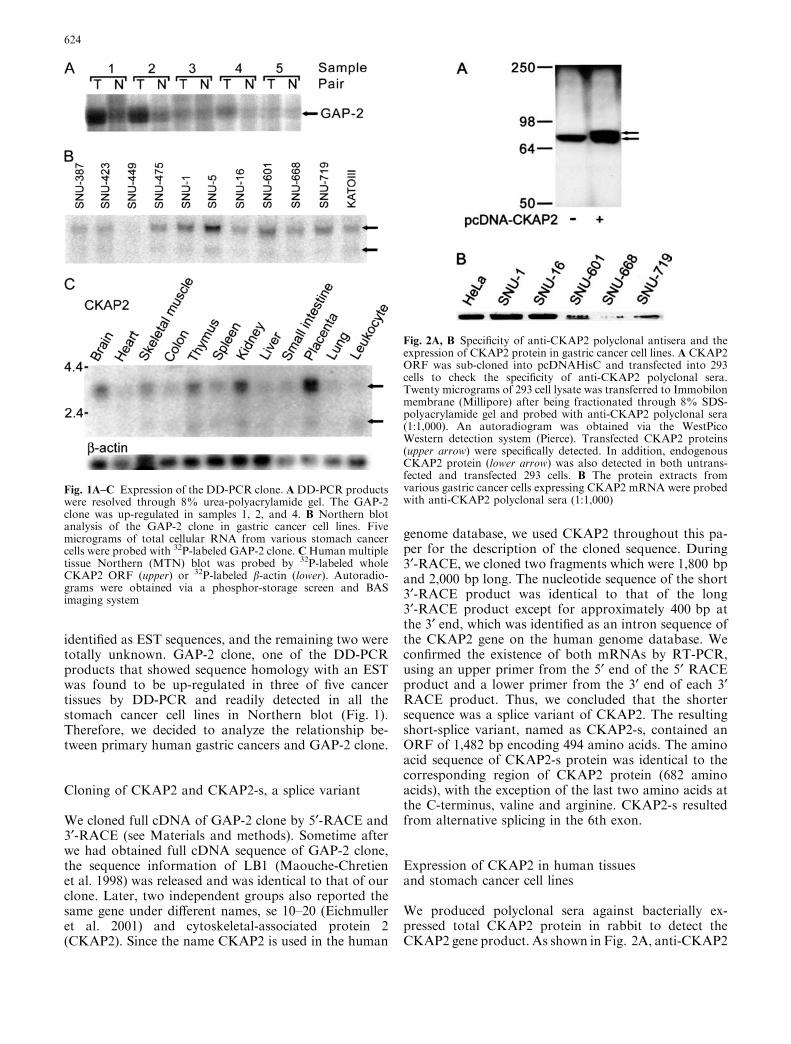
identified as EST sequences, and the remaining two weretotally unknown. GAP-2 clone, one of the DD-PCRproducts that showed sequence homology with an ESTwas found to be up-regulated in three of five cancertissues by DD-PCR and readily detected in all thestomach cancer cell lines in Northern blot (Fig. 1).Therefore, we decided to analyze the relationship be-tween primary human gastric cancers and GAP-2 clone.
Cloning of CKAP2 and CKAP2-s, a splice variant
We cloned full cDNA of GAP-2 clone by 5¢-RACE and3¢-RACE (see Materials and methods). Sometime afterwe had obtained full cDNA sequence of GAP-2 clone,the sequence information of LB1 (Maouche-Chretienet al. 1998) was released and was identical to that of ourclone. Later, two independent groups also reported thesame gene under different names, se 10–20 (Eichmulleret al. 2001) and cytoskeletal-associated protein 2(CKAP2). Since the name CKAP2 is used in the human
genome database, we used CKAP2 throughout this pa-per for the description of the cloned sequence. During3¢-RACE, we cloned two fragments which were 1,800 bpand 2,000 bp long. The nucleotide sequence of the short3¢-RACE product was identical to that of the long3¢-RACE product except for approximately 400 bp atthe 3¢ end, which was identified as an intron sequence ofthe CKAP2 gene on the human genome database. Weconfirmed the existence of both mRNAs by RT-PCR,using an upper primer from the 5¢ end of the 5¢ RACEproduct and a lower primer from the 3¢ end of each 3¢RACE product. Thus, we concluded that the shortersequence was a splice variant of CKAP2. The resultingshort-splice variant, named as CKAP2-s, contained anORF of 1,482 bp encoding 494 amino acids. The aminoacid sequence of CKAP2-s protein was identical to thecorresponding region of CKAP2 protein (682 aminoacids), with the exception of the last two amino acids atthe C-terminus, valine and arginine. CKAP2-s resultedfrom alternative splicing in the 6th exon.
Expression of CKAP2 in human tissuesand stomach cancer cell lines
We produced polyclonal sera against bacterially ex-pressed total CKAP2 protein in rabbit to detect theCKAP2 gene product. As shown in Fig. 2A, anti-CKAP2
Fig. 1A–C Expression of the DD-PCR clone. ADD-PCR productswere resolved through 8% urea-polyacrylamide gel. The GAP-2clone was up-regulated in samples 1, 2, and 4. B Northern blotanalysis of the GAP-2 clone in gastric cancer cell lines. Fivemicrograms of total cellular RNA from various stomach cancercells were probed with 32P-labeled GAP-2 clone. CHuman multipletissue Northern (MTN) blot was probed by 32P-labeled wholeCKAP2 ORF (upper) or 32P-labeled b-actin (lower). Autoradio-grams were obtained via a phosphor-storage screen and BASimaging system
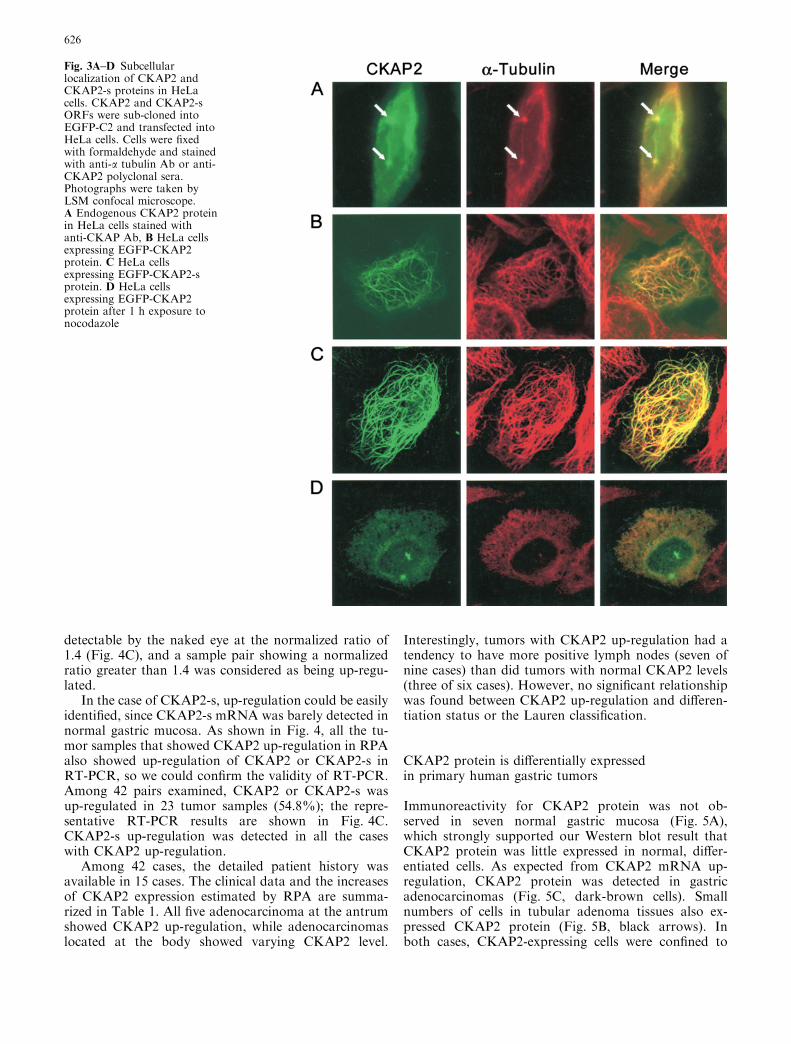
Fig. 2A, B Specificity of anti-CKAP2 polyclonal antisera and theexpression of CKAP2 protein in gastric cancer cell lines. A CKAP2ORF was sub-cloned into pcDNAHisC and transfected into 293cells to check the specificity of anti-CKAP2 polyclonal sera.Twenty micrograms of 293 cell lysate was transferred to Immobilonmembrane (Millipore) after being fractionated through 8% SDS-polyacrylamide gel and probed with anti-CKAP2 polyclonal sera(1:1,000). An autoradiogram was obtained via the WestPicoWestern detection system (Pierce). Transfected CKAP2 proteins(upper arrow) were specifically detected. In addition, endogenousCKAP2 protein (lower arrow) was also detected in both untrans-fected and transfected 293 cells. B The protein extracts fromvarious gastric cancer cells expressing CKAP2 mRNA were probedwith anti-CKAP2 polyclonal sera (1:1,000)
624

polyclonal sera detected the transfected CKAP2 proteinin 293-cells, which migrated at a slower rate due to theN-terminal his and Xpress tag from pcDNA3.1/HisC. Inaddition to the transfected CKAP2 protein, endogenousCKAP2 protein of approximately 75 kDa was detectedin both untransfected and transfected 293-cells. The75-kDa CKAP2 protein was readily detected on Westernblot in all the stomach cancer cell lines expressingCKAP2 mRNA, SNU-1, SNU-16, SNU-601, SNU-668,and SNU-719 (Fig. 2B).
We examined the expression of CKAP2 protein innormal human lung, breast, and gastrointestinal tract,including stomach, colon, and rectum, by Western blotbut were unable to detect CKAP2 protein (data notshown). Owing to limitations in the availability of nor-mal human tissues, we also examined normal mousetissues for proteins reacting with anti-CKAP2 anti-sera,but failed to detect any protein band of 75 kDa (datanot shown). These data suggested that CKAP2 mightexpress at a low level in normal, differentiated cells.
While CKAP protein was not detected in normalhuman tissues, its mRNA was detected weakly in almostall tissues on Northern blot, where CKAP2 mRNA wasexpressed relatively higher in the brain, thymus, kidney,and placenta than in the heart, lung, leukocytes andgastrointestinal tract (Fig. 1C). We also observed an-other very weak message, which was detected in cells ortissues with high CKAP2 expression (Fig. 1B, lowerarrow and Fig. 1C, lower arrow). The size of the shortermessage, 2.2 kbp, suggested that this could be the mes-senger RNA for CKAP2-s.
CKAP2 protein co-localized with microtubules
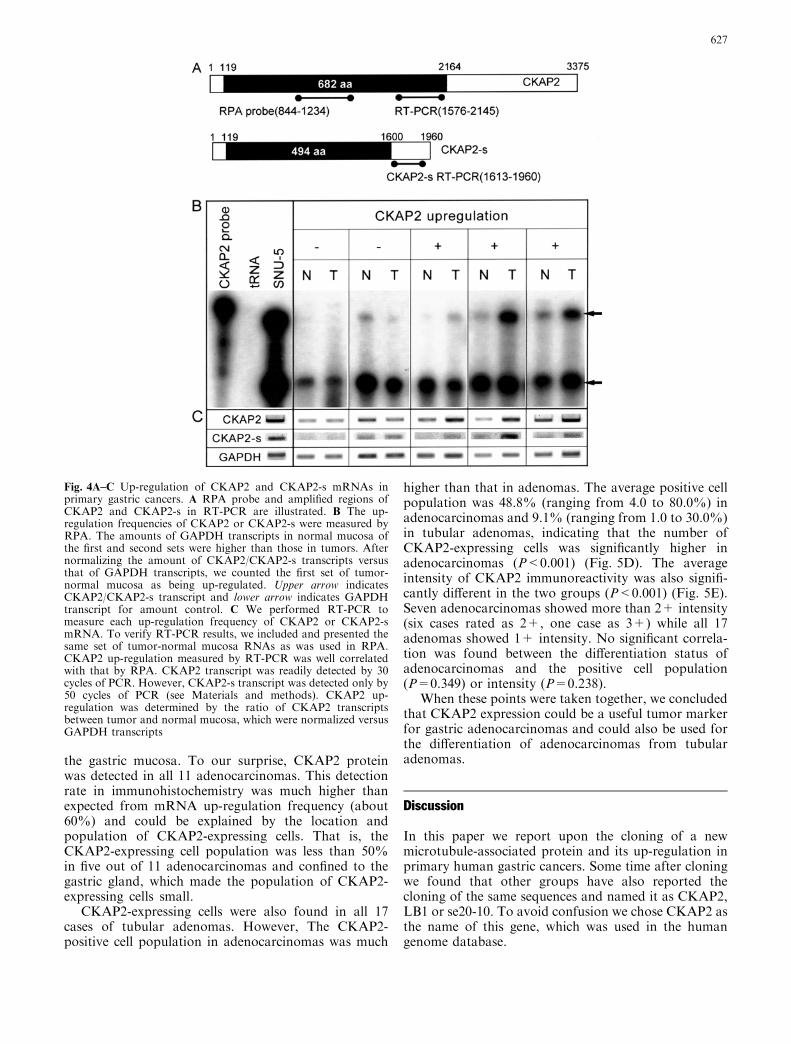
We then examined the subcellular localization ofendogenous CKAP2 protein in HeLa cells, using anti-CKAP2 polyclonal sera. As shown in the left-handcolumn of Fig. 4A, CKAP2 protein was stained mainlyin the cytosol and showed a mixed pattern of small dotsand threads. The striking feature was two dots withradiating thread (white arrows) in the cytosol, whichresembled the microtubule organizing center (MTOC),so we compared localization of CKAP2 protein withthat of microtubules. As shown in the middle and right-hand columns of Fig. 3A, CKAP2 was exactly localizedto the MTOC and radiating microtubules. We alsofound cytosolic threads and dots seen in CKAP2 im-munostaining, also localized to microtubules.
Since polyclonal anti-CKAP2 antisera was made viawhole CKAP2 protein, CKAP2 and CKAP-s could notbe discriminated by this antisera, so we made N-terminalenhanced green fluorescence protein (EGFP) fusionproteins of CKAP2 and CKAP-s. When we expressedEGFP-CKAP2 protein in the HeLa cells, EGFP-CKAP2 in HeLa cells was mainly located in the cytosoland showed a thread-like cytoskeletal distribution pat-tern (Fig. 3B, left-hand column). When both EGFP-CKAP2 image and microtubule image were merged,
EGFP-CKAP2 protein and the microtubules were ex-actly co-localized in the cytosol (Fig. 4B, right-handcolumn). The intracellular localization pattern ofEGFP-CKAP2-s protein was almost indistinguishablefrom that of EGFP-CKAP2 (Fig. 3C).
Both subcellular distribution pattern of EGFP-CKAP2 and immunofluorescence staining pattern ofendogenous CKAP2 protein strongly suggested thatCKAP2 protein was associated with microtubules. Toclarify this point further, we treated HeLa cellsexpressing EGFP-CKAP2 with nocodazole, a microtu-bule destabilizing agent, for 1 h to examine the locali-zation of CKAP2. Under these conditions, microtubuleswere depolymerized and dispersed throughout the cell,as shown in the middle column of Fig. 3D. Similarly,EGFP-CKAP2 also lost its characteristic thread-likepattern and was dispersed throughout the cell (Fig. 3D,left-hand column).
When these points were taken together, we concludedthat CKAP2 protein was associated with the microtu-bule network.
CKAP2 and CKAP2-s mRNAs are up-regulatedin primary human gastric cancers
Since CKAP2 protein was not detected in normal hu-man stomach tissues, we initially determined CKAP2up-regulation frequency, using ribonuclease protectionassay (RPA). We used a common region betweenCKAP2 and CKAP2-s for RPA, since the sequencearound the splice region was AT-rich (Fig. 4A).
As shown in Fig. 4B, a 448-bp ribonuclease-protectedCKAP2 fragment (upper arrow) and a 302-bp GAPDHfragment (lower arrow) were easily detected in SNU-5stomach cancer cells. We selected 15 tumor cases withdetailed patient histories. After normalizing CKAP2density with G3PDH density, we compared the amountof CKAP2 in tumors and matched normal mucosa. Itwas found that nine out of 15 paired samples showedhigher expressions of CKAP2 mRNA in the tumor tis-sues (Fig. 4B). The increase of CKAP2 expression,estimated by RPA, ranged from 1.5-fold to 3.5-fold(Table 1).
Next, we performed RT-PCR, using the primer setspecific for CKAP2 or CKAP2-s to distinguish CKAP2-sfrom CKAP2. The amplified region of CKAP2 orCKAP was designed to contain several exons to elimi-nate possible genomic DNA contamination. As seen inFig. 4C, both a 570-bp CKAP2 and a 348-bp CKAP2-sfragment were detected in SNU-5 cells and gastric can-cer tissues. Various amount of CKAP2 mRNA was alsodetected in all the normal gastric mucosa samples distantfrom the tumor tissues, while CKAP2-s mRNA wasdetected only in several cases. After normalizing theamount of each RT-PCR product versus G3PDH, wecalculated the normalized ratio of the CKAP2 RT-PCRproducts in each pair of tumor and normal mucosa. Thedifferences in CKAP2 RT-PCR products were readily
625
detectable by the naked eye at the normalized ratio of1.4 (Fig. 4C), and a sample pair showing a normalizedratio greater than 1.4 was considered as being up-regu-lated.
In the case of CKAP2-s, up-regulation could be easilyidentified, since CKAP2-s mRNA was barely detected innormal gastric mucosa. As shown in Fig. 4, all the tu-mor samples that showed CKAP2 up-regulation in RPAalso showed up-regulation of CKAP2 or CKAP2-s inRT-PCR, so we could confirm the validity of RT-PCR.Among 42 pairs examined, CKAP2 or CKAP2-s wasup-regulated in 23 tumor samples (54.8%); the repre-sentative RT-PCR results are shown in Fig. 4C.CKAP2-s up-regulation was detected in all the caseswith CKAP2 up-regulation.
Among 42 cases, the detailed patient history wasavailable in 15 cases. The clinical data and the increasesof CKAP2 expression estimated by RPA are summa-rized in Table 1. All five adenocarcinoma at the antrumshowed CKAP2 up-regulation, while adenocarcinomaslocated at the body showed varying CKAP2 level.
Interestingly, tumors with CKAP2 up-regulation had atendency to have more positive lymph nodes (seven ofnine cases) than did tumors with normal CKAP2 levels(three of six cases). However, no significant relationshipwas found between CKAP2 up-regulation and differen-tiation status or the Lauren classification.
CKAP2 protein is differentially expressedin primary human gastric tumors
Immunoreactivity for CKAP2 protein was not ob-served in seven normal gastric mucosa (Fig. 5A),which strongly supported our Western blot result thatCKAP2 protein was little expressed in normal, differ-entiated cells. As expected from CKAP2 mRNA up-regulation, CKAP2 protein was detected in gastricadenocarcinomas (Fig. 5C, dark-brown cells). Smallnumbers of cells in tubular adenoma tissues also ex-pressed CKAP2 protein (Fig. 5B, black arrows). Inboth cases, CKAP2-expressing cells were confined to
Fig. 3A–D Subcellularlocalization of CKAP2 andCKAP2-s proteins in HeLacells. CKAP2 and CKAP2-sORFs were sub-cloned intoEGFP-C2 and transfected intoHeLa cells. Cells were fixedwith formaldehyde and stainedwith anti-a tubulin Ab or anti-CKAP2 polyclonal sera.Photographs were taken byLSM confocal microscope.A Endogenous CKAP2 proteinin HeLa cells stained withanti-CKAP Ab, B HeLa cellsexpressing EGFP-CKAP2protein. C HeLa cellsexpressing EGFP-CKAP2-sprotein. D HeLa cellsexpressing EGFP-CKAP2protein after 1 h exposure tonocodazole
626
the gastric mucosa. To our surprise, CKAP2 proteinwas detected in all 11 adenocarcinomas. This detectionrate in immunohistochemistry was much higher thanexpected from mRNA up-regulation frequency (about60%) and could be explained by the location andpopulation of CKAP2-expressing cells. That is, theCKAP2-expressing cell population was less than 50%in five out of 11 adenocarcinomas and confined to thegastric gland, which made the population of CKAP2-expressing cells small.
CKAP2-expressing cells were also found in all 17cases of tubular adenomas. However, The CKAP2-positive cell population in adenocarcinomas was much
higher than that in adenomas. The average positive cellpopulation was 48.8% (ranging from 4.0 to 80.0%) inadenocarcinomas and 9.1% (ranging from 1.0 to 30.0%)in tubular adenomas, indicating that the number ofCKAP2-expressing cells was significantly higher inadenocarcinomas (P<0.001) (Fig. 5D). The averageintensity of CKAP2 immunoreactivity was also signifi-cantly different in the two groups (P<0.001) (Fig. 5E).Seven adenocarcinomas showed more than 2+ intensity(six cases rated as 2+, one case as 3+) while all 17adenomas showed 1+ intensity. No significant correla-tion was found between the differentiation status ofadenocarcinomas and the positive cell population(P=0.349) or intensity (P=0.238).
When these points were taken together, we concludedthat CKAP2 expression could be a useful tumor markerfor gastric adenocarcinomas and could also be used forthe differentiation of adenocarcinomas from tubularadenomas.
Discussion
In this paper we report upon the cloning of a newmicrotubule-associated protein and its up-regulation inprimary human gastric cancers. Some time after cloningwe found that other groups have also reported thecloning of the same sequences and named it as CKAP2,LB1 or se20-10. To avoid confusion we chose CKAP2 asthe name of this gene, which was used in the humangenome database.
Fig. 4A–C Up-regulation of CKAP2 and CKAP2-s mRNAs inprimary gastric cancers. A RPA probe and amplified regions ofCKAP2 and CKAP2-s in RT-PCR are illustrated. B The up-regulation frequencies of CKAP2 or CKAP2-s were measured byRPA. The amounts of GAPDH transcripts in normal mucosa ofthe first and second sets were higher than those in tumors. Afternormalizing the amount of CKAP2/CKAP2-s transcripts versusthat of GAPDH transcripts, we counted the first set of tumor-normal mucosa as being up-regulated. Upper arrow indicatesCKAP2/CKAP2-s transcript and lower arrow indicates GAPDHtranscript for amount control. C We performed RT-PCR tomeasure each up-regulation frequency of CKAP2 or CKAP2-smRNA. To verify RT-PCR results, we included and presented thesame set of tumor-normal mucosa RNAs as was used in RPA.CKAP2 up-regulation measured by RT-PCR was well correlatedwith that by RPA. CKAP2 transcript was readily detected by 30cycles of PCR. However, CKAP2-s transcript was detected only by50 cycles of PCR (see Materials and methods). CKAP2 up-regulation was determined by the ratio of CKAP2 transcriptsbetween tumor and normal mucosa, which were normalized versusGAPDH transcripts
627
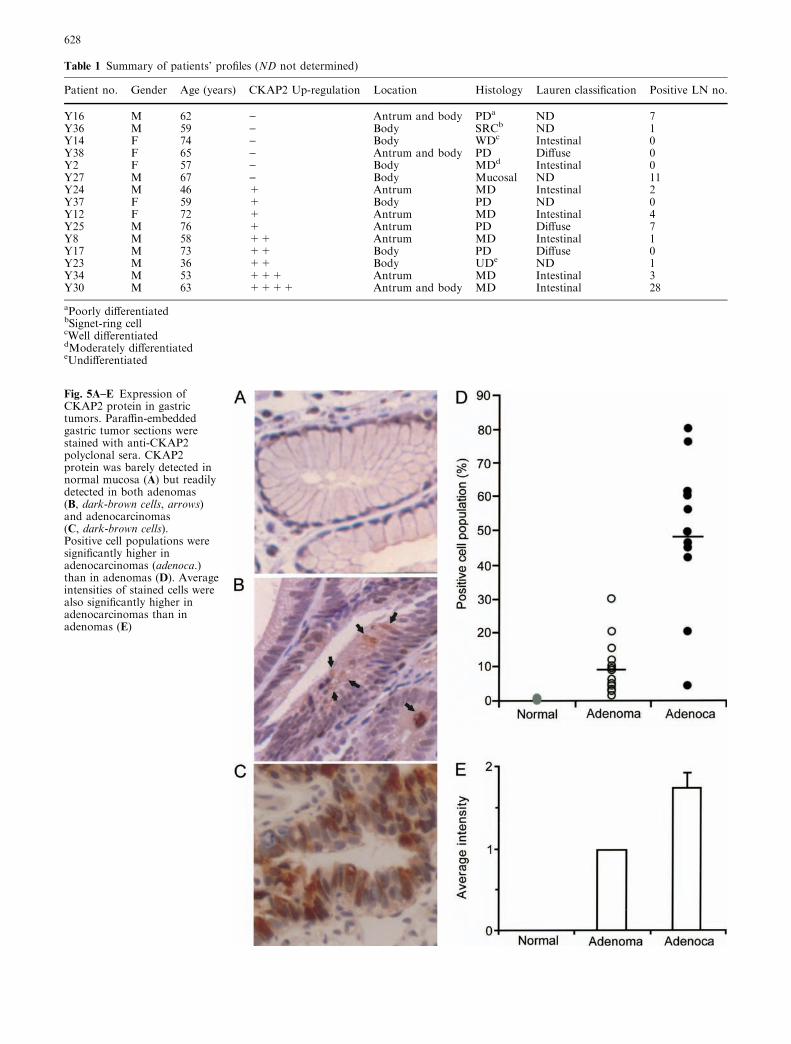
Table 1 Summary of patients’ profiles (ND not determined)
Patient no. Gender Age (years) CKAP2 Up-regulation Location Histology Lauren classification Positive LN no.
Y16 M 62 ) Antrum and body PDa ND 7Y36 M 59 ) Body SRCb ND 1Y14 F 74 ) Body WDc Intestinal 0Y38 F 65 ) Antrum and body PD Diffuse 0Y2 F 57 ) Body MDd Intestinal 0Y27 M 67 ) Body Mucosal ND 11Y24 M 46 + Antrum MD Intestinal 2Y37 F 59 + Body PD ND 0Y12 F 72 + Antrum MD Intestinal 4Y25 M 76 + Antrum PD Diffuse 7Y8 M 58 ++ Antrum MD Intestinal 1Y17 M 73 ++ Body PD Diffuse 0Y23 M 36 ++ Body UDe ND 1Y34 M 53 +++ Antrum MD Intestinal 3Y30 M 63 ++++ Antrum and body MD Intestinal 28
aPoorly differentiatedbSignet-ring cellcWell differentiateddModerately differentiatedeUndifferentiated
Fig. 5A–E Expression ofCKAP2 protein in gastrictumors. Paraffin-embeddedgastric tumor sections werestained with anti-CKAP2polyclonal sera. CKAP2protein was barely detected innormal mucosa (A) but readilydetected in both adenomas(B, dark-brown cells, arrows)and adenocarcinomas(C, dark-brown cells).Positive cell populations weresignificantly higher inadenocarcinomas (adenoca.)than in adenomas (D). Averageintensities of stained cells werealso significantly higher inadenocarcinomas than inadenomas (E)
628

The CKAP2 gene was first cloned as an up-regulatedsequence in two out of six diffuse B-cell lymphoma cases(Maouche-Chretien et al. 1998). Later, CKAP2 up-reg-ulation was also reported from cutaneous T-cell lym-phoma (Eichmuller et al. 2001). These data and highexpression in thymus and spleen gave us the impressionthat CKAP2 alterations might be specific to lymphoma,but our result indicated that up-regulation of CKAP2 ismore frequent in primary human gastric adenocarcino-mas than in lymphomas. The frequency of CKAP2mRNA up-regulation (60%) is also much higher thanthose of other up-regulated genes in gastric adenocar-cinomas, such as ras, c-erbB-2, c-myc (less than 30%).These data suggest the possible usage of CKAP2 up-regulation as a gastric tumor marker.
In this context, a small-splice form of CKAP2,CKAP2-s seems more suitable to detect gastric adeno-carcinomas. CKAP2-s is an alternative spliced form ofCKAP2 and consists of 494 amino acids. Its amino acidsequence is identical to the corresponding part ofCKAP2, except for the last two c-terminal amino acids.The size of its cloned cDNA was 2.2 kbp, which mat-ched the smaller band detected by the Northern blotanalysis of normal tissues and cancer cell lines. TheCKAP2-s mRNA expression was well correlated withCKAP2 mRNA expression. The CKAP-s transcript wasdetected only in tissues or cancer cell lines in whichCKAP2 expression was relatively high. Moreover, theup-regulation of CKAP2-s in cancer tissues was moreobvious than that of CKAP2 by RT-PCR, since theexpression of CKAP2-s in normal tissues was quite low(Fig. 4).
An examination of CKAP2 protein expression ingastric tumors by immunohistochemical staining pro-duced more striking results. CKAP2 protein was de-tected in all the benign and malignant gastric tumorsamples that we examined. CKAP2 protein expressionseemed to be very specific for the tumor, i.e., so far wehave not detected any CKAP2 protein expression innormal gastric mucosa by immunohistochemical stain-ing. As shown in Fig. 5, adenomas and adenocarcino-mas were significantly different with respect to CKAP2expression. The average number of positive cells in ad-enocarcinomas was almost 50%, while that in adenomaswas 9%. Staining intensities were also twice as high inadenocarcinomas than in adenomas. These results sug-gest that the expression of CKAP2 protein may be re-lated to the progression of tumors, and that thedetection of CKAP2 protein in tumor tissues may beuseful in the differential diagnosis of gastric tubularadenoma and adenocarcinoma, especially in smallendoscopic biopsy specimens.
Since we analyzed only 11 cases of adenocarcino-mas by immunohistochemistry, it is too early to besure of any relationship between CKAP2 expression inadenocarcinomas and their clinical pathology. How-ever, CKAP2 mRNA up-regulation tends to be relatedto lymph-node metastasis, based on the analysis of 14cases with patient histories. That is, seven out of ten
cases with lymph-node metastasis showed CKAP2mRNA up-regulation, although we did not get thestatistical significance, due to the small number. Wecould not find any other statistically significant rela-tionship between CKAP2 mRNA up-regulation andthe clinical course of gastric adenocarcinomas, and theanalysis of more cases is needed to clarify the rela-tionship between clinical parameters and CKAP2 up-regulation in human primary gastric cancers.
As the name of the gene indicated, CKAP2 wasthought to be associated with a cytoskeletal component.One report showed that the CKAP2 gene product,exogenously introduced into cells, was associated with acytoskeletal component (Maouche-Chretien et al. 1998).As seen in Fig. 3, we showed that this cytoskeletalcomponent might be the microtubule network. Theimmunofluorescence staining data showed that endoge-nous CKAP2 and the microtubule network are co-localized and that CKAP2 is also localized to theMTOC. The EGFP fusion form of CKAP2 showed analmost identical localization pattern, when compared tomicrotubules. Moreover, this co-localization pattern wasdisrupted by a microtubule-disrupting agent, nocodaz-ole. All these data strongly support the notion thatCKAP2 is related to the microtubule network. However,we did not see the direct association of CKAP2 andtubulin in immuno-precipitation (data not shown), andthe exact molecular nature of the relationship betweenthese two molecules remains unclear. We favored theindirect association between the two molecules, sincethere was no co-localization between CKAP2 anddestabilized microtubules, as seen in the right-handcolumn of Fig. 3D.
The malignant transformation of cells might beaccompanied by a disruption of the cytoskeletalorganization, and actin filament changes correlate withthe decreased expression of cytoskeleton-associatedproteins, suggesting roles of actin filament and cyto-skeleton-associated proteins in malignant transforma-tion (Pawlak and Helfman 2001). Subsequently, theexpression of actin-associated proteins, such as gelso-lin, profilin and vinculin, could alter the characteristicsof the transformed cells. However, the alteration ofmicrotubules during malignant transformation has notbeen well studied. The up-regulation of stathmin, atubulin dimer binding protein, which destabilizesmicrotubules, may correlate with malignant transfor-mation (Belmont and Mitchison 1996). Since CKAP2protein is highly expressed in stomach cancer cell linesand in a large population of malignant cells in gastricadenocarcinomas, there is the possibility that up-reg-ulated CKAP2 protein may affect the stability ofmicrotubules and thus contribute to the malignanttransformation of cells. Certainly, more studies areneeded to elucidate the cellular function of CKAP2protein and its role in cellular malignant transforma-tion.
In conclusion, CKAP2 protein was overexpressed inhuman primary gastric tumors, in high frequency, while
629

not detected in normal gastric mucosa. Moreover,CKAP2 protein is differentially expressed between gas-tric adenomas and adenocarcinomas. These data suggestthat CKAP2 protein overexpression can be used as agastric tumor marker and for differential diagnosis ofgastric adenocarcinomas over adenomas.
Acknowledgement This study was supported by SBRI grant andKOSEF through Cancer Research Center, Seoul NationalUniversity.
References
Akama Y, Yasui W, Yokozaki H, Kuniyasu H, Kitahara K,Ishikawa T, Tahara E (1995) Frequent amplification of thecyclin E gene in human gastric carcinomas. Jpn J Cancer Res86:617–621
Becker KF, Atkinson MJ, Reich U, Becker I, Nekarda H, SiewertJR, Hofler H (1994) E-cadherin gene mutations provide clues todiffuse type gastric carcinomas. Cancer Res 54:3845–3852
Belmont LD, Mitchison TJ (1996) Identification of a protein thatinteracts with tubulin dimmers and increases the catastropherate of microtubules. Cell 84:623–631
Berx G, Becker KF, Hofler H, van Roy F (1998) Mutations of thehuman E-cadherin (CDH1) gene. Hum Mutat 12:226–237
Block P, Craanen ME, Offerhaus GJA, Tytgat GNJ (1997) Gastriccarcinoma: clinical, pathogenic and molecular aspects. Q J Med90:735–749
Bouchard L, Lamarre L, Tremblay PJ, Jolicoeur P (1989)Stochastic appearance of mammary tumours in transgenic micecarrying the mmtv/c-neu oncogene. Cell 57:931–936
Eichmuller S, Usener D, Dummer R, Stein A, Thiel D, Schaden-dorf D (2001) Serological detection of cutaneous T-celllymphoma-associated antigens. Proc Natl Acad Sci U S A 98:629–634
Hirohashi S (1998) Inactivation of the E-cadherin-mediated celladhesion system in human cancers. Am J Pathol 153:333–339
Kihana T, Tsuda H, Hirota T, Shimosato Y, Sakamoto H. TeradaK, Hirohashi S (1991) Point mutations of c-Ki-ras oncogene ingastric adenoma and adenocarcinoma with tubular differentia-tion. Jpn J Cancer Res 82:308–314
Kuniyasu H, Yasui W, Kitadai Y, Yokozaki H, Ito H, Tahara E(1992) Frequent amplification of the c-met gene in scirrhoustype stomach cancer. Biochem Biophys Res Commun 189:227–232
Maouche-Chretien L, Deleu N, Badoual C, Fraissignes P, BergerR, Gaulard P, Romeo PH, Leroy-Viard K (1998) Identificationof a novel cDNA, encoding a cytoskeletal associated protein,differentially expressed in diffuse large B cell lymphomas.Oncogene 17: 1245–1251
Nakatsuru S, Yanagisawa A, Ichii S, Tahara E, Kato Y, Nakam-ura Y, Horii A (1992) Somatic mutation of the APC gene ingastric cancer: frequent mutations in very well-differentiatedadenocarcinoma and signet-ring cell carcinoma. Hum MolGenet 1:559–563
Oda T, Kanai Y, Oyama T, Yoshiura K, Shimoyama Y, Birch-meier W, Sugimura T, Hirohashi S (1994) E-cadherin genemutations in human gastric carcinoma cell lines. Proc NatlAcad Sci U S A 91:1858–1862
Pawlak G, Helfman DM (2001) Cytoskeletal changes in celltransformation and tumorigenesis. Curr Opin Genet Dev11:41–47
Ranzani GN, Pellegata NS, Previdere C, Saragoni A, Vio A,Maltoni M, Amadori D (1992) Heterogeneous proto-oncogeneamplification correlates with tumour progression and presenceof metastases in gastric cancer patients. Cancer Res 50:7811–7814
Sano T, Tsujino T, Yoshida K, Nakayama H, Haruma K, Ito H,Nakamura Y, Kajiyama G, Tahara E (1991) Frequent loss ofheterozygosity on chromosomes 1q, 5q, and 17p in humangastric carcinomas. Cancer Res 51:2926–2931
Tahara E (1993) Molecular mechanism of stomach carcinogenesis.J Cancer Res Clin Oncol 119:265–272
Tahara E (1995) Genetic alterations in human gastrointestinalcancers. Cancer 75:1410–1417
Werner M, Becker KF, Keller G, Hofler H (2001) Gastric adeno-carcinoma: pathomorphology and molecular pathology.J Cancer Res Clin Oncol 127:207–216
630
Related Documents











