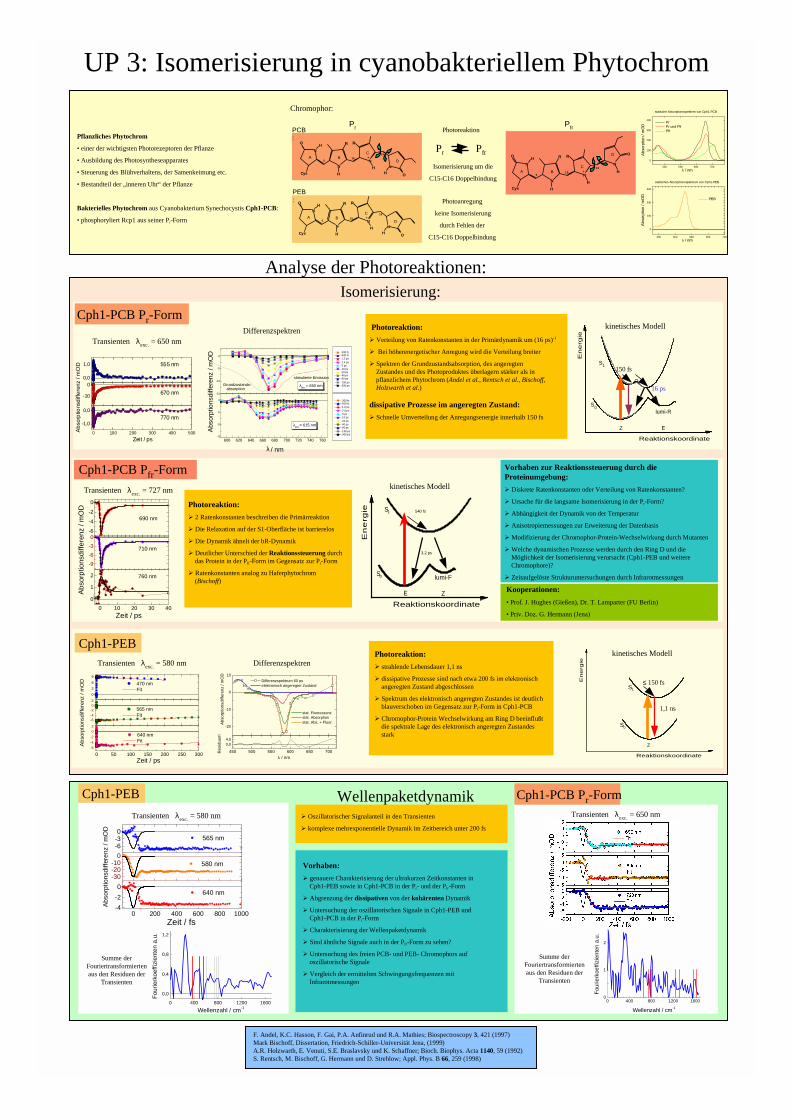
UP 3: Isomerisierung in cyanobakteriellem Phytochrom 400 500 600 700 0 100 200 300 400 statische Absorptionsspektren von Cph1-PCB Pr Pr und Pfr Pfr Absorption / mOD λ / nm 500 550 600 650 700 0 100 200 300 statisches Absorptionsspektrum von Cph1-PEB PEB Absorption / mOD λ / nm 0 100 200 300 400 500 -1,0 0,0 0,0 1,0 -10 0 770 nm Zeit / ps 555 nm 670 nm Absorptionsdifferenz / mOD Pflanzliches Phytochrom • einer der wichtigsten Photorezeptoren der Pflanze • Ausbildung des Photosyntheseapparates • Steuerung des Blühverhaltens, der Samenkeimung etc. • Bestandteil der „inneren Uhr“ der Pflanze Bakterielles Phytochrom aus Cyanobakterium Synechocystis Cph1-PCB: • phosphoryliert Rcp1 aus seiner Pr-Form Photoanregung keine Isomerisierung durch Fehlen der C15-C16 Doppelbindung Cph1-PEB Cph1-PCB P r -Form Isomerisierung: Wellenpaketdynamik Cph1-PCB P fr -Form P r P fr Isomerisierung um die C15-C16 Doppelbindung Photoreaktion Analyse der Photoreaktionen: 0 10 20 30 40 0 1 2 -9 -6 -3 0 -6 -4 -2 0 760 nm Zeit / ps 710 nm 690 nm Absorptionsdifferenz / mOD Transienten λ exc. = 727 nm Reaktionskoordinate E Z lumi-F Energie S 0 S 1 540 fs 3,2 ps kinetisches Modell Transienten λ exc. = 650 nm Differenzspektren -6 -4 -2 0 2 0 50 100 150 200 250 300 -6 -4 -2 0 2 0 3 6 9 565 nm Fit Absorptionsdifferenz / mOD 640 nm Fit Zeit / ps 470 nm Fit Transienten λ exc. = 580 nm 450 500 550 600 650 700 0,0 4,0 -20 -10 0 10 Absorptionsdifferenz / mOD Residuum λ / nm Differenzspektrum 60 ps elektronisch angeregter Zustand stat. Fluoreszenz stat. Absorption stat. Abs. + Fluor. Differenzspektren Reaktionskoordinate E Z lumi-R Energie S 0 S 1 kinetisches Modell 150 fs 16 ps Reaktionskoordinate Z Energie S 0 S 1 1,1 ns ≤ 150 fs kinetisches Modell Transienten λ exc. = 580 nm Transienten λ exc. = 650 nm Cph1-PEB Cph1-PCB P r -Form Vorhaben zur Reaktionssteuerung durch die Proteinumgebung: Ø Diskrete Ratenkonstanten oder Verteilung von Ratenkonstanten? Ø Ursache für die langsame Isomerisierung in der Pr-Form? Ø Abhängigkeit der Dynamik von der Temperatur Ø Anisotropiemessungen zur Erweiterung der Datenbasis Ø Modifizierung der Chromophor-Protein-Wechselwirkung durch Mutanten Ø Welche dynamischen Prozesse werden durch den Ring D und die Möglichkeit der Isomerisierung verursacht (Cph1-PEB und weitere Chromophore)? Ø Zeitaufgelöste Strukturuntersuchungen durch Infrarotmessungen Vorhaben: Ø genauere Charakterisierung der ultrakurzen Zeitkonstanten in Cph1-PEB sowie in Cph1-PCB in der Pr- und der Pfr-Form Ø Abgrenzung der dissipativen von der kohärenten Dynamik Ø Untersuchung der oszillatorischen Signale in Cph1-PEB und Cph1-PCB in der Pr-Form Ø Charakterisierung der Wellenpaketdynamik Ø Sind ähnliche Signale auch in der Pfr-Form zu sehen? Ø Untersuchung des freien PCB- und PEB- Chromophors auf oszillatorische Signale Ø Vergleich der ermittelten Schwingungsfrequenzen mit Infrarotmessungen P fr Chromophor: N H R N O Cys H N H R N H O + A B C D P r 5 10 15 PCB : PEB : 15 10 5 D C B A + N H R N O Cys H N H R O H N N H R N O Cys H N H R N H O + A B C D 5 10 15 Photoreaktion: Ø Verteilung von Ratenkonstanten in der Primärdynamik um (16 ps) -1 Ø Bei höherenergetischer Anregung wird die Verteilung breiter Ø Spektren der Grundzustandsabsorption, des angeregten Zustandes und des Photoproduktes überlagern stärker als in pflanzlichem Phytochrom (Andel et al., Rentsch et al., Bischoff, Holzwarth et al.) dissipative Prozesse im angeregten Zustand: Ø Schnelle Umverteilung der Anregungsenergie innerhalb 150 fs Photoreaktion: Ø 2 Ratenkonstanten beschreiben die Primärreaktion Ø Die Relaxation auf der S1-Oberfläche ist barrierelos Ø Die Dynamik ähnelt der bR-Dynamik Ø Deutlicher Unterschied der Reaktionssteuerung durch das Protein in der Pfr-Form im Gegensatz zur Pr-Form Ø Ratenkonstanten analog zu Haferphytochrom (Bischoff) Kooperationen: • Prof. J. Hughes (Gießen), Dr. T. Lamparter (FU Berlin) • Priv. Doz. G. Hermann (Jena) Photoreaktion: Ø strahlende Lebensdauer 1,1 ns Ø dissipative Prozesse sind nach etwa 200 fs im elektronisch angeregten Zustand abgeschlossen Ø Spektrum des elektronisch angeregten Zustandes ist deutlich blauverschoben im Gegensatz zur Pr-Form in Cph1-PCB Ø Chromophor-Protein Wechselwirkung am Ring D beeinflußt die spektrale Lage des elektronisch angeregten Zustandes stark 0 400 800 1200 1600 0,0 0,4 0,8 1,2 Fourierkoeffizienten a.u. Wellenzahl / cm -1 Summe der Fouriertransformierten aus den Residuen der Transienten Summe der Fouriertransformierten aus den Residuen der Transienten Ø Oszillatorischer Signalanteil in den Transienten Ø komplexe mehrexponentielle Dynamik im Zeitbereich unter 200 fs F. Andel, K.C. Hasson, F. Gai, P.A. Anfinrud und R.A. Mathies; Biospectroscopy 3, 421 (1997) Mark Bischoff, Dissertation, Friedrich-Schiller-Universität Jena, (1999) A.R. Holzwarth, E. Venuti, S.E. Braslavsky und K. Schaffner; Bioch. Biophys. Acta 1140, 59 (1992) S. Rentsch, M. Bischoff, G. Hermann und D. Strehlow; Appl. Phys. B 66, 259 (1998) 600 620 640 660 680 700 720 740 760 -15 -10 -5 0 -15 -10 -5 0 λ anr. = 615 nm Absorptionsdifferenz / mOD λ / nm 300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps Grundzustands- absorption stimulierte Emission λ anr. = 650 nm 300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps 0 400 800 1200 1600 0 1 2 Fourierkoeffizienten a.u. Wellenzahl / cm -1 -30 -20 -10 0 -6 -3 0 0 200 400 600 800 1000 -4 -2 0 Absorptionsdifferenz / mOD 580 nm 565 nm Zeit / fs 640 nm

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
UP 3: Isomerisierung in cyanobakteriellem Phytochrom
400 500 600 700
0
100
200
300
400
statische Absorptionsspektren von Cph1-PCB
Pr Pr und Pfr Pfr
Abso
rptio
n / m
OD
λ / nm
500 550 600 650 700
0
100
200
300
statisches Absorptionsspektrum von Cph1-PEB
PEB
Abso
rptio
n / m
OD
λ / nm
0 100 200 300 400 500
-1,0
0,0
0,0
1,0
-10
0
770 nm
Zeit / ps
555 nm
670 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
Pflanzliches Phytochrom
• einer der wichtigsten Photorezeptoren der Pflanze
• Ausbildung des Photosyntheseapparates
• Steuerung des Blühverhaltens, der Samenkeimung etc.
• Bestandteil der „inneren Uhr“ der Pflanze
Bakterielles Phytochrom aus Cyanobakterium Synechocystis Cph1-PCB:
• phosphoryliert Rcp1 aus seiner Pr-Form
Photoanregung
keine Isomerisierung
durch Fehlen der
C15-C16 Doppelbindung
Cph1-PEB
Cph1-PCB Pr-Form
Isomerisierung:
Wellenpaketdynamik:
Cph1-PCB Pfr-Form
Pr Pfr
Isomerisierung um die
C15-C16 Doppelbindung
Photoreaktion
Analyse der Photoreaktionen:
0 10 20 30 400
1
2
-9-6-30
-6
-4
-2
0
760 nm
Zeit / ps
710 nm
690 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
Transienten λexc. = 727 nm
ReaktionskoordinateE Z
lumi-F
En
erg
ie
S0
S1 540 fs
3,2 ps
kinetisches Modell
Transienten λexc. = 650 nmDifferenzspektren
-6-4-202
0 50 100 150 200 250 300-6-4-202
0369
565 nm Fit
Abso
rptio
nsdi
ffere
nz /
mO
D
640 nm Fit
Zeit / ps
470 nm Fit
Transienten λexc. = 580 nm
450 500 550 600 650 700
0,04,0
-20
-10
0
10
Abso
rptio
nsdi
ffere
nz /
mO
D
Res
iduu
m
λ / nm
Differenzspektrum 60 ps elektronisch angeregter Zustand
stat. Fluoreszenz stat. Absorption stat. Abs. + Fluor.
Differenzspektren
Reaktionskoordinate
EZ
lumi-R
En
erg
ie
S0
S1
kinetisches Modell
150 fs
16 ps
Reaktionskoordinate
Z
Energ
ie
S0
S1
1,1 ns
≤ 150 fs
kinetisches Modell
Transienten λexc. = 580 nm Transienten λexc. = 650 nm
Cph1-PEB Cph1-PCB Pr-Form
Vorhaben zur Reaktionssteuerung durch dieProteinumgebung:� Diskrete Ratenkonstanten oder Verteilung von Ratenkonstanten?
� Ursache für die langsame Isomerisierung in der Pr-Form?
� Abhängigkeit der Dynamik von der Temperatur
� Anisotropiemessungen zur Erweiterung der Datenbasis
� Modifizierung der Chromophor-Protein-Wechselwirkung durch Mutanten
� Welche dynamischen Prozesse werden durch den Ring D und die Möglichkeit der Isomerisierung verursacht (Cph1-PEB und weitere Chromophore)?
� Zeitaufgelöste Strukturuntersuchungen durch Infrarotmessungen
Vorhaben:� genauere Charakterisierung der ultrakurzen Zeitkonstanten in Cph1-PEB sowie in Cph1-PCB in der Pr- und der Pfr-Form
� Abgrenzung der dissipativen von der kohärenten Dynamik
� Untersuchung der oszillatorischen Signale in Cph1-PEB und Cph1-PCB in der Pr-Form
� Charakterisierung der Wellenpaketdynamik
� Sind ähnliche Signale auch in der Pfr-Form zu sehen?
� Untersuchung des freien PCB- und PEB- Chromophors auf oszillatorische Signale
� Vergleich der ermittelten Schwingungsfrequenzen mit Infrarotmessungen
Pfr
Chromophor:
N
H
RN
O
Cys
H
NH
R
NH O
+A BC
D
Pr
510
15
PCB:
PEB:
1510
5 D
CBA +
N
H
RN
O
Cys
H
NH
R
OHN
N
H
R
N
O
Cys
H
N
H
R
N
H
O
+A BC
D
510
15
Photoreaktion:� Verteilung von Ratenkonstanten in der Primärdynamik um (16 ps)-1
� Bei höherenergetischer Anregung wird die Verteilung breiter
� Spektren der Grundzustandsabsorption, des angeregten Zustandes und des Photoproduktes überlagern stärker als in pflanzlichem Phytochrom (Andel et al., Rentsch et al., Bischoff, Holzwarth et al.)
dissipative Prozesse im angeregten Zustand:� Schnelle Umverteilung der Anregungsenergie innerhalb 150 fs
Photoreaktion:� 2 Ratenkonstanten beschreiben die Primärreaktion
� Die Relaxation auf der S1-Oberfläche ist barrierelos
� Die Dynamik ähnelt der bR-Dynamik
� Deutlicher Unterschied der Reaktionssteuerung durch das Protein in der Pfr-Form im Gegensatz zur Pr-Form
� Ratenkonstanten analog zu Haferphytochrom (Bischoff)
Kooperationen:• Prof. J. Hughes (Gießen), Dr. T. Lamparter (FU Berlin)
• Priv. Doz. G. Hermann (Jena)
Photoreaktion:� strahlende Lebensdauer 1,1 ns
� dissipative Prozesse sind nach etwa 200 fs im elektronisch angeregten Zustand abgeschlossen
� Spektrum des elektronisch angeregten Zustandes ist deutlich blauverschoben im Gegensatz zur Pr-Form in Cph1-PCB
� Chromophor-Protein Wechselwirkung am Ring D beeinflußt die spektrale Lage des elektronisch angeregten Zustandes stark
0 400 800 1200 1600
0,0
0,4
0,8
1,2
Four
ierk
oeffi
zien
ten
a.u.
Wellenzahl / cm-1
Summe derFouriertransformiertenaus den Residuen der
Transienten
Summe derFouriertransformiertenaus den Residuen der
Transienten
� Oszillatorischer Signalanteil in den Transienten
� komplexe mehrexponentielle Dynamik im Zeitbereich unter 200 fs
F. Andel, K.C. Hasson, F. Gai, P.A. Anfinrud und R.A. Mathies; Biospectroscopy 3, 421 (1997)Mark Bischoff, Dissertation, Friedrich-Schiller-Universität Jena, (1999)A.R. Holzwarth, E. Venuti, S.E. Braslavsky und K. Schaffner; Bioch. Biophys. Acta 1140, 59 (1992)S. Rentsch, M. Bischoff, G. Hermann und D. Strehlow; Appl. Phys. B 66, 259 (1998)
600 620 640 660 680 700 720 740 760-15
-10
-5
0
-15
-10
-5
0
λanr. = 615 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
λ / nm
300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps
Grundzustands- absorption
stimulierte Emission
λanr. = 650 nm
300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps
0 400 800 1200 16000
1
2
Four
ierk
oeffi
zien
ten
a.u.
Wellenzahl / cm-1
-30-20-10
0-6-30
0 200 400 600 800 1000-4-20
Ab
sorp
tions
diffe
renz
/ m
OD
580 nm
565 nm
Zeit / fs
640 nm
Related Documents


![- 237 - Manz/Saalfrank/Gerber Teilprojekt C1: Theorie zur ...users.physik.fu-berlin.de/~abt/sfb450/1bericht/c1.pdf · Hier erwies sich die von Rabitz et al. [1] entwickelte Methode](https://static.cupdf.com/doc/110x72/5d5416a688c993f52b8b551f/-237-manzsaalfrankgerber-teilprojekt-c1-theorie-zur-users-abtsfb4501berichtc1pdf.jpg)








