479 Unusually severe symptoms are a characteristic of the current epidemic of mosaic virus disease of cassava in Uganda By R W GIBSON. J P LEGG and G W OTIM-NAPE Namulonge Agricultural and Animal Research Institute. PO Box 7084, Kampala, Uganda (Accepted 28 March 1996) Summary Mosaic disease (MD) is more severe in cassava plants infected within the area of the current epidemic in northern and central Uganda than to the south of the affected area. This difference in severity was recorded within a single cultivar as well as amongst the mixtures of cultivars found commonly in farmers' fields. An increase in severity also occurred as the epidemic passed through localities. Varietal or agroecological factors coincident with the area of the epidemic are therefore unlikely to cause the increased severity. The severe disease could also be graft and cutting transmitted and could super-infect mildly diseased plants. Both mildly and severely diseased plants gave positive reactions in ELISA tests to antisera prepared against African cassava mosaic virus (ACMV) and an unusually severe form of ACMV or a closely related geminivirus is likely to be the cause of the severe mosaic disease. The epidemic also involves increased populations of the whitefly vector of ACMV, Bemisia rabaci. and possible hypotheses are presented as to how these phenomena may be related. Key words: Cassava. geminivirus, African cassava mosaic virus. ACMV. virus, severity, Uganda, whitefly Introduction An epidemic of mosaic disease (MD) in cassava has been extending southwards from northern Uganda at some tens of kilometres per aiznum for at least 6 yr, causing major losses of cassava and food shortages, especially when poor rains diminished the yields of other crops. In some of the worst affected areas, the crop has been largely abandoned (Thresh, 0th-Nape & Jennings. 1994). Currently (January 1995). the southern limit of the epidemic is roughly an east/west line just north of Kampala. Jinja and Tororo. but south of Mubende. The epidemic of MD is associated with unusually large numbers of the whitefly, Bemisia tabaci (Gennadius)( Homoptera: Aleyrodidae), vector of African cassava mosaic virus (ACMV; family Geminiuiridae. genus "Subgroup I11 Geminivirus"). The increased occurrences of both MD-affected plants and B. tahaci along two transects crossing the southern border of the epidemic have been documented by Legs (1995). A possible cause of this epidemic is a new strain of B. rabaci. In the Americas and elsewhere. new strains of whiteflies have attained unusually large populations and caused epidemics of other whitefly-borne viruses (Perring er a l . . 1991. 1993: Bedford et al.. 1992). @ 1996 Association of Applied Biologists

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

479
Unusually severe symptoms are a characteristic of the current epidemic of mosaic virus disease of cassava in
Uganda
By R W GIBSON. J P L E G G and G W OTIM-NAPE Namulonge Agricultural and Animal Research Institute. PO Box 7084, Kampala,
Uganda (Accepted 28 March 1996)
Summary Mosaic disease (MD) is more severe in cassava plants infected within the area
of the current epidemic in northern and central Uganda than to the south of the affected area. This difference in severity was recorded within a single cultivar as well as amongst the mixtures of cultivars found commonly in farmers' fields. An increase in severity also occurred as the epidemic passed through localities. Varietal or agroecological factors coincident with the area of the epidemic are therefore unlikely to cause the increased severity. The severe disease could also be graft and cutting transmitted and could super-infect mildly diseased plants. Both mildly and severely diseased plants gave positive reactions in ELISA tests to antisera prepared against African cassava mosaic virus (ACMV) and an unusually severe form of ACMV or a closely related geminivirus is likely to be the cause of the severe mosaic disease. The epidemic also involves increased populations of the whitefly vector of ACMV, Bemisia rabaci. and possible hypotheses are presented as to how these phenomena may be related.
Key words: Cassava. geminivirus, African cassava mosaic virus. ACMV. virus, severity, Uganda, whitefly
Introduction A n epidemic of mosaic disease ( M D ) in cassava has been extending southwards from
northern Uganda at some tens of kilometres per aiznum for at least 6 yr, causing major losses of cassava and food shortages, especially when poor rains diminished the yields of other crops. In some of the worst affected areas, the crop has been largely abandoned (Thresh, 0 t h - N a p e & Jennings. 1994). Currently (January 1995). the southern limit of the epidemic is roughly an east/west line just north of Kampala. Jinja and Tororo. but south of Mubende. The epidemic of M D is associated with unusually large numbers of the whitefly, Bemisia tabaci (Gennadius)( Homoptera: Aleyrodidae), vector of African cassava mosaic virus (ACMV; family Geminiuiridae. genus "Subgroup I11 Geminivirus"). The increased occurrences of both MD-affected plants and B. tahaci along two transects crossing the southern border of the epidemic have been documented by Legs (1995).
A possible cause of this epidemic is a new strain of B. rabaci. In the Americas and elsewhere. new strains of whiteflies have attained unusually large populations and caused epidemics of other whitefly-borne viruses (Perring er al . . 1991. 1993: Bedford et al . . 1992). @ 1996 Association of Applied Biologists

480 R U' GIBSON. J P LEGG AND G W OTIM-NAPE
In Uganda, geographically and host-restricted B . tahaci have been identified (Legg. Gibson & 0 t h - N a p e . 1994).
Severe and mild cassava mosaic diseases have previously been identified in Tanzania (Storey. 1936: Storey 8: Nichols. 1938) and Kenya (Bock. Guthrie & Figueirido. 1981). Cassava mosaic disease in these countries has now been determined t o be caused by two geminiviruses. namely A C M V and East African cassava mosaic virus (EACMV). In Kenya and Tanzania. ACMV occurs largely in the centre and west and EACMV occurs in coastal locations (Swanson & Harrison, 1994). whereas only ACMV has been recorded in Uganda. Observations suggested that M D is unusually severe within the area of the epidemic in Uganda and the present work examined this quantitatively. determined various biological characteristics of the disease and considers how severity of M D might link with other aspects of the epidemic.
Materials and Methods
Locatiori of sites The main area of the epidemic of M D during the period of this study was in northern
Uganda. and transects (Fig. 1) crossing into the epidemic area were made following the lines of the two main roads into this northern area: namely along the road north from Kampala towards Masindi between Kawanda and Nakitoma; and along the road from Tororo north towards Soroti between Tororo and Kumi. During the period of study. the epidemic expanded southwards and details are given in Table 1. Namulonge is about 10 km east of Kawanda, and remained to the south of the epidemic during the period of study.
Scoritig system for sj'mptom seoerity The severity of M D in plants was assessed using a 1-3 (0 = symptomless) scoring system.
1 = mild mosaic present. no leaf distortion; 2 = slight leaf distortion: chlorosis affecting < c. 30% leaf area: 3 = severe leaf distortion: chlorosis affecting < c. 80% leaf area: 4 = leaves severely distorted and stunted. early leaf loss and 8O-lOOV leaf area chlorotic.
Plants growing from infected cuttings were distinguished from those infected during current crop growth by whether or not the oldest leaves present were diseased.
assessing diseased foliage as:
Differences iti severity of symptoms at sites tratisectitig the epicletnic Crops of 3-4 month old cassava were examined along transects following the roads north
from Kampala to Masindi and from Tororo t o Kumi. Crops were examined at 3 month intervals from November 1992 to August 1993 for M D . Nine locations at c. 20 km intervals between Kawanda and Nakitoma were examined on the Kampala-Masindi road and 1 0 at c. 10 km intervals were examined between Tororo and Kumi (Fig. I : Table 1). A t each location, 10 crops. each often comprising a mixture of cultivnrs. were sampled (Tables 2 81 3): scores only o f plants that had grown from infected cuttings were analysed.
Cliutiges it1 M D seoerity as the epidetnic recrched Ioccitiotis During the main period (November 1992 to August 1993) of the transect studies. the
front of the epidemic moved south along the Kampala road and past Kasana. Severity scores when Kasana was ahead of the epidemic (July 1097- to January 1993) and afterwards

Epidemic of severe mosaic disease of cassava
\ Masindi
UGANDA
Nakasongola
Nakitoma
I Kakinzi
t 4 Katikamu . Namulonge
Kawanda
0 Soroti
Tororo
48 1
Lake Victoria
TANZANIA , 50km ,
Fig. 1 . A sketch map showing the locations of the main experimental sites and transects studied.
Table 1. Details of the sites utilised in the transects a ) Transect from Kawanda to Nakitoma h ) Transect from Tororo to Kumi
Km north Km N of Location of Kampala Epidemic arrived Location Tororo Epidemic arrived
Kawanda 20 Bombo 40 Katikamu 60 Kasana 80 Kakinzi I 0 0
Nakasongola 140
Nakitoma 180
Kakoge 120
Migeera 160
1995 1994 199314 1993 1992 1992
Tororo 0 Mukuju 10 Busiu 20 Busoba 30 Mbale 40 Nakaloke 50 Kachumbale 60 Bukedea 70
Kumi 80 Atutur 80
-
I995 1995 I994 I993 1992 1992
* Epidemic present before current study began.
(April 1993 t o July 1993) when it had been engulfed. have been analysed separately (Table 4). Subsequently, the epidemic moved further south and an additional assessment of 10 crops was made at Katikamu in August. 1994. when the front was 20 km further south at Bombo. Similarly, Busiu on the Tororo transect was just ahead of the epidemic during the main period of the transect study but by February 1995 the epidemic front had past it to reach within 10-20 km of Tororo; consequently, a further 10 crops were then scored at Busiu.

482 R W GIBSON. J P LEGG A N D G W OTIM-NAPE
Severity of symptoms in crops of a single cultivar in a transect of the epidemic front In order to exclude varietal effects on symptom severity. an additional transect study was
done examining only plants of the local cv. Kalitunsi. Farmers' fields were again used and the severity of MD was assessed at sites ahead of the epidemic between Namulonge and Kampala. at the front of the epidemic in the area just south of Bombo and behind the front between Kasana and Kakoge. These locations lie on a line running roughly N/S across the front which in August 1994 lay midway between Bombo and Namulonge. At each location, at least six crops were examined. In plants infected during the current season, only the diseased foliage was scored. In each field, up to 30 plants grown from infected cuttings or 30 plants infected during crop growth were scored (Table 5 ) .
Propagation of mild and severe M D through cuttings Six cuttings cv. Kalitunsi were collected from each of six severely diseased plants behind
the front near Bombo and six from each of six mildly diseased plants ahead of the front near Namulonge. They were grown in pots of soil in a screenhouse and the severity of MD and their height recorded 5 wk later. Subsequently, cuttings were collected from a further 26 severely diseased and 26 mildly diseased plants, planted outside and similarly assessed.
Transmission of mild and severe M D by graft inoculation Cuttings from symptomless plants of cassava cv. Ebwanateraka were planted in a field at
Namulonge on 12 May 1994 at 1 m spacing. Plants were top-grafted (late September) in a random sequence with scions obtained from severely diseased plants of cv. Kalitunsi from near Bombo (within the epidemic area), with scions from mildly-diseased plants of cv. Kalitunsi from the vicinity of Namulonge (ahead of the epidemic) or "mock-grafted" by cutting back as if to top-graft. The success of graft unions and the severities of any MD were recorded regularly (Table 6). Ebwanateraka is susceptible to ACMV and was widely grown in northern Uganda before the epidemic.
Cross-protection against severe M D in mildly diseased cassava Cuttings were obtained from symptomless or mildly diseased plants of cv. Kalitunsi
growing near Namulonge. A plot of each was planted on 5 September 1994 both in a field at Namulonge (20 cuttings plot-') and in a farmer's field c. 30 km north at Katikamu (21 cuttings plot-'), which was by then within the epidemic area. The severity of MD was monitored weekly at both sites.
Serological tests using antisera raised against ACMV Triple antibody sandwich enzyme-linked immunosorbent assays (TAS-ELISA) were done
using polyclonal antibodies raised in rabbit as coating antibody, murine monoclonal antibody prepared against ACMV (Mab SCRI No 23) (Swanson & Harrison, 1994) as detecting antibody and anti-mouse alkaline phosphatase conjugated antibody. Shoot tips were sampled in all cases and each sample was tested in duplicate wells. Samples were collected from six mildly diseased plants of cv. Kalitunsi at Namulonge and from six severely diseased plants of cv. Kalitunsi within the epidemic area (Katikamu), and from eight mildly and eight severely diseased plants of cv. Ebwanateraka from the graft-inoculation experiment (Table 6). During this period (1994), the front of the epidemic was approaching Namulonge and a further 20 severely diseased plants of cv. Ebwanateraka were sampled there from a crop which was known to have been planted from undiseased cuttings and in which about 10% of plants were diseased. In all cases, corresponding numbers of symptomless plants were tested as controls.

Epidemic of severe mosaic disease of cassava 483
Effects of MD on whitefly populations Populations of whiteflies were compared on 60 MD-affected and 60 symptomless cassava
plants cv. Kalitunsi from 11 crops near Bombo (just within the epidemic) by counting the adult whiteflies on the top five leaves and the nymphs (instars S 4 ) on the 11-15th leaves down from the first fully expanded leaf. Earlier work had shown that these were the leaves where these instars were most abundant on cassava plants (Legg, 1995). Whole plant counts of nymphs were also done on six diseased and six symptomless plants. Counts were also made of the nymphs and eggs found on chlorotic and dark-green areas of diseased leaves of cvs Bao and Pora.
Statistical analysis Severity scores were compared using chi-squared tests. Means are given % standard
deviation of individual observations
Results
General observations on symptoms of M D
Within the epidemic area Plants infected during the current season within the area of the epidemic often have very
severe symptoms. These include a majority of the leaf area being chlorotic, leaves being twisted and with large indentations to their edges. stunting to perhaps <5% normal leaf area, and premature abscission of diseased leaves. Chlorotic areas of leaves generally comprise a few large patches occupying much of the leaf area rather than a mosaic; chlorosis occurs to the extent that the affected area is almost white or yellow, apparently dependent on the cultivar. Leaves produced before infection had occurred appear undiseased and these often remain on the plants after younger but diseased leaves have fallen. Leaves produced soon after infection commonly have petioles curved in a downward S-shape. This produces an overall appearance of a candle in a leafy candlestick. plants having apparently normal basal leaves, a zone of downward-curled leaves, a zone of bare stem where leaves have fallen prematurely and young stunted and distorted leaves at the top; sometimes the majority of plants in a field have this appearance. Plants produced from infected cuttings are generally very stunted with small distorted and chlorotic leaves. They are also often found growing in poorly kept fields which the farmer had more-or-less abandoned.
Ahead of the epidemic Ahead of the epidemic, infected plants are generally of a similar height to undiseased
plants and are hard to identify from a distance except perhaps by being less bushy than undiseased ones. Leaves are of a similar size to those on unaffected plants although often with a slight downward cupping and the majority of the leaf area is a normal dark green. The remaining chlorotic areas of leaves are scattered in small areas as a mosaic pattern and the chlorosis is generally pale green.
Differences in severity of symptoms at sites transecting the epidemic Analysis of the 1992-3 data for M D severity in crops along the road north from Kampala
(Table 2) showed that ahead of the epidemic (at Kawanda, Bombo and Katikamu). no more than 1-2% of plants had severity score 3 and none scored 4. By contrast, there was

484 R W GIBSON. J P LEGG A N D G W OTIM-NAPE
a much greater proportion ( P < 0.0001) of severely diseased plants at sites within the epidemic area (north of Kakinzi: see Table 1) where 2040% of plants scored 3 and 1-5% scored 4. MD was most severe close to the epidemic front, at Kakinzi and nearby Kakoge, and there was a significant ( P < 0.0001) decrease in scores at sites north of the front.
Analysis of the 1992-3 data for MD severity in crops along the Tororo-Kumi road (Table 3) also showed that only 1-2% of plants scored 3 and none scored 4 at sites ahead of the epidemic front (from Tororo to Busoba). At Mbale, the approximate limit of the epidemic front during this period, c. 5% of plants had scores of 3 and at sites from Nakaloke northwards to Atutur. c. 13% of plants had scores of 3 or 4, significantly ( P < 0.0001) more than at sites ahead of the front.
Table 2. Mean percentages of plants infected as cuttings and with particular severity scores found in farmers’ fields of 3-4 month old cassava crops along a 160 k m trarisect north f rom
Kawarida to Nakitoma“
Site
Kawanda Bomho Katikamu Kasana Kakinzi Kakoge Nakasongola Migeera Nakitoma
“r plants with severity scores of: Total r/r I - 3 4 infected 7
7 - 0 1 I 3 3 4 5 9
12 3
10 9
20 22 29 30 50
7 - 0 1 9
74 1 1 36 24 19
0 0 0 1 5 5 4 3 1
16 3
12 19 53 71 73 62 7x
* Assessments were done at 3 month intervals from July 1992 to July 1993. sampling II total of 15(M plants at each site.
Table 3. Mean percentages of plants infected as cuttings and with particular severity scores found in farmers’ fields of 3 4 month old cassava crops along a 90 km transect north f r o m
Tororo to Kumi* % plants with severity scores of: Total ‘ i
Site 1 2 3 4 infected
Tororo Mukuju Busiu Busoba Mhale Nakaloke Kachumbala Bukedea Atutur Kurni
4 3 3 2
16 8 6 6 8
10
15 9 7
11 25 22 22 36 25 35
3 I 1 2 5
11 13 11 13 16
22 12 11 1.5 46 42 44 53 47 62
* Assessments were done at 3 month intervals from November 1992 to August 1993, sampling a total of 1200 plants at each site.
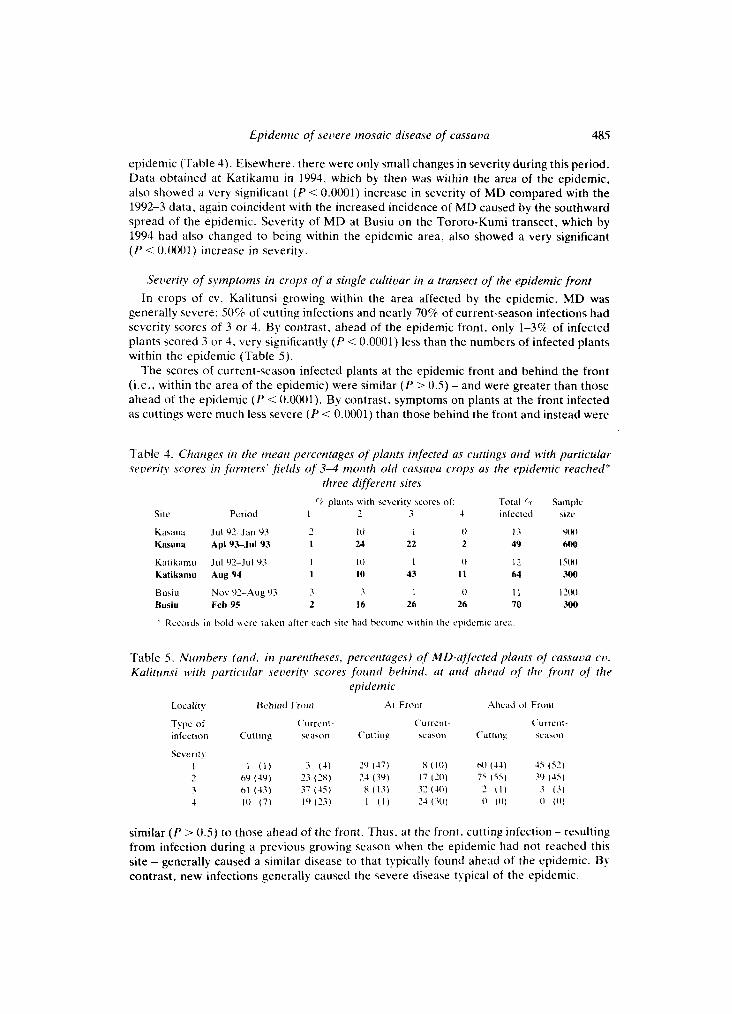
Changes in M D severity as the epidemic reached locutions At Kasana, severity of MD increased very significantly ( P < 0.0001) between July 1992-
January 1993, and April 1993-July 1993. This period coincided with the arrival of the
Epidemic of severe mosaic disease of cassava 485
epidemic (Table 4). Elsewhere, there were only small changes in severity during this period. Data obtained at Katikamu in 1994, which by then was within the area of the epidemic. also showed a very significant ( P < 0.0001) increase in severity of MD compared with the 1992-3 data, again coincident with the increased incidence of MD caused by the southward spread of the epidemic. Severity of M D at Busiu on the Tororo-Kumi transect. which by 1994 had also changed to being within the epidemic area, also showed a very significant ( P < 0.0001) increase in severity.
Seoeriry of symptoms in crops of a sirigle cultiour in a transect o f the epidemic front In crops of cv. Kalitunsi growing within the area affected by the epidemic, MD was
generally severe: 50% of cutting infections and nearly 70% of current-season infections had severity scores of 3 o r 4. By contrast, ahead of the epidemic front, only 1-3% of infected plants scored 3 or 4. very significantly ( P < 0.0001) less than the numbers of infected plants within the epidemic (Table 5 ) .
The scores of current-season infected plants at the epidemic front and behind the front (i.e.. within the area of the epidemic) were similar ( P > 0.5) - and were greater than those ahead of the epidemic ( P < 0.0001). By contrast. symptoms on plants at the front infected as cuttings were much less severe ( P < 0.0001) than those behind the front and instead were
Table 4. Qiariges iri tlie meaii percentages of p l u m infected US cirttirigs urid with particular severity scores iri farrriers' fields of 3 4 morirh old cussuvu crops as the epidemic reached*
three djfererir sires r: plants with sevcrity scores o f : Total r Sample
Site Period 1 7 3 4 inlectcd size
K. . I 7 I 0 1 0 13 Y(H1 ci\rlnd Jul Y2-Jan Y3 Kasana Apl93-Jul 93 I 24 22 2 49 600
Katikarnu Aug 94 1 I0 43 I 1 64 300
Busiu N o \ Y2-Aug Y3 3 3 1 ( 1 I I 121H) Bush Feb 95 2 16 26 26 70 300
Katik;rinu Ju l Y2-Jul Y3 I I 0 I 0 I?. l5(M
Records in bold \\ere tahen alter each 3ite had hecomc sithiti the epidemic area
Table 5. Numbers (arid, iri parentheses. perceritages) of ML)-a[ft~cted p h i i t s of cassuua cu. Kalitunsi N d t h particular seiwrity ,scores foiirid hehirid, at urid uheud o,f the ,frorir of the
epidemic Locality Bchind Front Ar Fronr Ahe;id 01 Front
Type of C urrc i i t - ('urrent- ('urrent infection Cutting seawn Cuttin? w a w n Cut t in s w a w n
Severit! I 1 ( 1 ) 3 (4 ) 29 (47) x (10) h0 (44) 45 ( 5 2 ) - 6') (49) 23 (281 24 (39) 17 (31) 75 ( 5 5 ) .3Y (45) 3 h l (43) 37 (45) x ( 1 3 ) 32 (40 ) 2 ( 1 ) 3 ( 3 ) 4 I 0 ( 7 ) I Y ( 2 3 ) 1 ( I ) 24 ( 3 0 ) 0 ( I l l ( I (0)
1
similar ( P > 0.5) to those ahead of the front. Thus. at the front. cutting infection - resulting from infection during a previous growing season when the epidemic had not reached this site - generally caused a similar disease to that typically found ahead o f the epidemic. By contrast, new infections generally caused the severe disease typical of thc epidemic.

486 R W GIBSON. J P LEGG A N D G W OTIM-NAPE
Propagation of mild arid severe M D through cuttings The cuttings taken from severely diseased plants of cv. Kalitunsi behind the front and
grown in a screenhouse produced severely diseased progeny plants (scores 3 & 4) with a mean height of only 86 2 SO mm. Cuttings from mildly diseased plants ahead of the front produced mildly diseased progeny (scores 1 & 2) with a mean height of 154 * 34mm. When planted in a field, only 16 of the 26 cuttings from the severely diseased plants grew. and 13 (80%) of these were also severely diseased scoring 3 or 4. whereas 24 of the 26 cuttings from mildly diseased plants grew and all surviving plants had mild symptoms. scoring 1 or 2. The height of cuttings from severely diseased plants, 66 * 25 mm. was about half that of cuttings from mildly diseased plants (1 16 2 31 mm).
Transmission of mild arid severe M D by graft inoculation Only two of the 24 plants of cv. Ebwanateraka grafted with severely diseased scions
united successfully whereas 12 of 27 plants grafted with mildly diseased scions united. The two surviving scions from severely diseased plants remained severely diseased with scores of 3 and 4 and had grown only 11 and 13 cm during the two months following grafting. By contrast, the scions from mildly diseased plants had scores of 1 or 2 and grew 30 ? 20 cm.
All plants of Ebwanateraka that united with either mildly or severely diseased scions became diseased. However. despite only two of the severely diseased scions uniting suc- cessfully. 16 (70%) of the 24 Ebwanateraka grafted with them were diseased, all developing severe symptoms typical of plants infected within the epidemic area (Table 6). T h e low frequency of MD amongst ungrafted controls (3%) and the fact that the severity of disease induced corresponded to that of the failed scion both confirm that the graft was the source of infection even in these instances. Leaflets of severely diseased plants were about a third the length and a quarter the width of corresponding symptomless leaflets (Table 7) . Nineteen (70%) of the 27 plants of cv. Ebwanateraka grafted with mildly diseased scions became diseased and, by contrast ( P < 0.001). all had mild symptoms even two months after inoculation, with leaflets only slightly reduced in size compared to controls.
Table 6 . Numbers o f plants cv . Ebwanateraka with particular severity scores, 2 months after graft-inoculation using either severely o r mildly diseased cv . Kalitiinsi
Numbers of plant3 with beverity scores: Scion 0 1 2 3 4
(symptomless)
Severe 8 Mild 8 Ungrafted 90
Table 7. Dimensions in mm of the lippermost fiillv-expatided leuj of disuu.sed plunts cv . E b wanateraka. 2 months after graft-inoculation using severely or mildly diserised co. Kulitirrisi
Leaflet length 1x9 t 6 . K x 1 2 X.1 ’ 236 t 15‘ Leaflet width 41 f 2.4 I6 ? 2.0 h l ? 4.x Petiole length 207 2 7.9 107 2 1 . 1 263 t 2.6 No. of observations I X I9 I 0 1
Mild Severe Unt11st.ased
* 2 Standard Deviation of individual observations

Epidemic of severe mosaic disease of cassava
Cross-protection againsr severe M D in mildly diseased cassava The cuttings of cv. Kalitunsi from either mildly diseased or symptomless plants produced
plants that initially were correspondingly mildly diseased or symptomless at both exper- imental sites. However, at Katikamu, within the epidemic area. MD spread rapidly and by late November, just two months after planting. 19 (90%) of the 21 originally symptomless plants there had MD, and 17 (80%) of these were in the severe categories 3 and 4. At this sampling date, 15 (70%) of the originally mildly diseased plants at Katikamu were also in the severe MD categories, scoring either 3 or 4, demonstrating an inability of mild MD to cross-protect against the severe disease. At Namulonge Farm, just 30 km north of Katikamu, spread of MD was much less and, at the same sampling date. only two (10%) of the originally symptomless plants had developed MD and this was only mild (categories 1 or 2). None of the mildly diseased plants at Namulonge had developed severe symptoms.
487
Serological tests using antisera raised against A CMV The samples from both severely and mildly diseased plants of cv. Kalitunsi and the grafted
cv. Ebwanateraka gave positive reactions in TAS-ELISA assays compared to symptomless controls (Kalitunsi, A405,, = 0.10 f 0.026; Ebwanateraka. A405,, = 0.16 2 0.016), with severely diseased samples generating greater absorbance values (Kalitunsi. A405,, = 0.85 * 0.071; Ebwanateraka, A405,, = 1.03 f 0.46) than those from mildly diseased plants (Kalitunsi, A405,, = 0.32 * 0.13; Ebwanateraka, A405,, = 0.32 * 0.17). The samples from Namulonge of 20 severely diseased but newly infected plants of cv. Ebwanateraka generated widely varying absorbances, with most giving absorbances much greater than symptomless controls. A few initially reacted similarly to controls, but all were positive when retested using further samples.
Effects of M D on whitefly populatioris There were 4.6 f 0.54 adult whiteflies on the top five leaves and 43 * 6.0 nymphs on
leaves 11-16 on symptomless plants which was not significantly different from the 5.0 &
0.38 adults and 46 * 6.4 nymphs on diseased plants. On whole plants, 698 t 191 nymphs were counted on symptomless plants compared with 441 * 55 on diseased plants.
O n chlorotic areas of 15 leaves, 68 nymphs and 12 eggs were counted whereas 741 nymphs and 281 eggs were counted on dark green areas. Chlorotic and dark green areas were eslimated overall as being approximately equal on the leaves sampled so the numbers of nymphs and eggs per unit area were greater on dark green than on chlorotic areas ( P < 0.000l).
Discussion These results demonstrate a marked increase in the severity of MD in Uganda associated
with the epidemic that has been sweeping southwards across the country. The symptoms include severe stunting and chlorosis of leaves and premature leaf loss on current-season infected plants. Symptoms similar to these have also been described from Madagascar (Cours, 1951), presumably caused by EACMV (Swanson & Harrison. 1994). As well as occurring in a range of cultivars. the increase in severity was found in a single cv. Kalitunsi. so it is not the result of especially sensitive cultivars being grown within the area of the epidemic. The increase in severity also occurred at three different sites and during different periods as the epidemic moved into each locality so it is also unlikely to be caused by coincident changes in agroecological or environmental conditions. Furthermore. the dif- ferent symptomatologies were separated by only a few kilometres and could be induced

488 R w GIBSON;. J P LEGG AND G h’ OTIM-NAPE
experimentally at a single site by propagation through cuttings and grafts. A n increase in severity as the epidemic reached a locality was also shown by the generally mild M D on cutting-infected plants (infections presumably having derived from when the locality had been ahead of the epidemic) as compared with new infections. Although the increased severity has been assessed quantitatively along only two transects. observations by the authors indicate that severe M D occurs throughout the area of the epidemic from near Lake Albert in the west to Mbale in the east of Uganda.
Samples from plants with either mild or severe M D all gave positive reactions in TAS- ELISA using antibodies raised against ACMV. However. the antibodies used are not specific to ACMV. cross-reacting to other whitefly-borne geminiviruses in TAS-ELISA (Harrison et al., 1991). T h e samples tested included ones from a crop infected recently and at a relatively low frequency. In such a situation, it is unlikely that the positive reactions were against a virus other than the one associated with the severe disease. In preliminary tests done on severely diseased material sent to the Scottish Crop Research Institute. geminivirus antigen was also detected; three sap-transmissible viruses that occur elsewhere in Africa. uiz. cassava brown streak, cassava Ivorian bacilliform and cassava Q viruses were not detected (B D Harrison, personal communication). More serological and molecular information is needed to confirm the exact aetiology of severe M D , although its most likely cause is a whitefly-borne geminivirus and perhaps just a more severe form of A C M V than was previously common.
As well as an increased incidence of largely severely-diseased cassava. the epidemic of M D in Uganda involves an increased abundance on cassava of the geminivirus vector B. rubaci (Legg, 1995). As the serological evidence suggests that the severe M D is a whitefly- borne geminivirus, it seems likely that there is a common basis for this association. Possible explanations are:-
1. Severe M D could be altering the cassava crop so as to somehow favour the increase of vector populations. Our counts of whitefly adults and nymphs on diseased and symp- tomless plants did not indicate any such effect. and the distributions of nymphs and eggs on individual leaves indicate that diseased areas are avoided. Furthermore. large populations of whiteflies have not been evident when plants have been grown for other experimental purposes at Namulonge (ahead of the front) from cuttings collected from diseased plants located within the area of the epidemic.
A biotype of B. tabaci better adapted to cassava may be responsible for the increased spread of the severe disease. Minor but apparently consistent differences in esterase isoenzymes suggest that the B. [abaci associated with the epidemic may be different and so perhaps better adapted to cassava; this alone could explain the greater abundance of the whiteflies associated with the epidemic (Legg. 1YYS). ACMV has been detected serologically only in chlorotic areas of leaves, which occupy a greater proportion of leaf area in severely diseased plants and its titre is also greater in severely diseased leaves (Fargette, Thouvenel & Fauquet. 1987) as was also indicated by serology in this present study. These attributes can explain how severe M D is transmitted more readily by whiteflies than mild M D (Storey & Nichols. 1938). If the severe M D in the current epidemic in Uganda is also more readily transmitted - especially perhaps by the possible new strain of B. fuhoci - than mild, the epidemic of whiteflies could carry with it an epidemic o f predominantly severe M D . The ability of severe M D t o super-infect mildly diseased plants. as also found by Storey & Nichols (1938), would also assist the severe disease to predominate.
The infrequent occurrence of severely diseased plants ahead of the whitefly epidemic also needs to be considered. It may be that the virus or virus strain causing severe M D is new
2 .

Epidemic of severe mosaic disease of cassava 489
to Uganda and has not yet had time to spread so as to be abundant everywhere. However, Storey & Nichols (1938) reported a whitefly-transmissible severe MD of cassava in neigh- bouring Tanzania several decades ago (although it is uncertain whether their severe strain was as severe as that now found in Uganda) suggesting that the severe disease is endemic. If so, selection by farmers against the use of cuttings from severely diseased plants may previously have maintained it at only a low frequency as, without a large whitefly population, there would be little opportunity for its possibly faster spread to counterbalance this.
Selection of the more tolerant cultivars from amongst those being grown (Thresh er al., 1994) may explain the apparent decline in severity at Nakitoma where the epidemic has been present for several years.
Whatever the explanation, that the current epidemic in Uganda is associated with a more severe MD is itself of extreme importance in the local food economy. Its ability to reduce leaflet area to 10% that of symptomless leaves, to induce chlorosis of much of the remaining leaf area, and to induce premature leaf loss explains how the epidemic has caused such severe damage. It is also a warning to neighbouring countries that their cassava crops may be at risk and of their need to monitor the situation and make contingency plans. There appear to be no practical strategies at hand for controlling the further spread of this epidemic now that it covers such a large area. The epidemic has also proved able to progress through a range of agroecological environments. In developing control strategies we are also hampered by uncertainty of the exact cause of the epidemic, and developing this knowledge is a high priority. In Uganda, the main strategy for combating the problem is to develop and distribute resistant cultivars (Thresh e t a / . , 1994) and, because of the time delay inherent in implementing such a strategy, it is suggested that similar programmes should urgently be begun or expanded elsewhere.
Acknowledgements This work was jointly undertaken under a scientific arrangement between the Ugandan
National Agricultural Research Organisation and the Natural Resources Institute, UK. funded by the Natural Resources Division of the British Overseas Development Adminis- tration. We wish to acknowledge valuable advice from Dr J M Thresh and Professor B D Harrison, and technical support by Mr I Mpembe.
References Bedford I D, Briddon R W, Markham P G , Brown J K, R o d R C. 1992. Bemi.yia rahaci - biotype
characterisation and the threat of this whitefly species to agriculture. Reporr of the 19% Brighron Conference - Pesrs and Diseases, pp. 1235-1240.
Bock K R, Guthrie E J, Figueirido G. 1981. A strain of cassava latent virus occurring in coastal districts of Kenya. Annals of Applied Biology 99:151-159.
Cours G . 1951. Le Manioc a Madagascar. Memoires de I'lnsiiiui Recherche Scienrifique de Madagascar. Serie B , Biologie VegCtal 3:203-400.
Fargette D, Thouvenel J-C, Fauquet C. 1987. Virus content of leaves of cassava infected by African cassava mosaic virus, Annals of Applied Biology 110:65-73.
Harrison B D, Muniyappa V, Swanson M M, Roberts I M, Robinson D J. 1991. Recognition and differentiation of seven whitefly-transmitted gerniniviruses from India. and their relationships to African cassava mosaic and Thailand mung bean yellow mosaic viruses. Annals of Applied Biology 118:299-308.
Legg J P. 1995. The ecology of Bemisia tabaci (Gennadius)(Homoptera: Ale~rodidae). uecror of African cassaua mosaic geniiniuirus in Uganda. Ph.D. Thesis. University of Reading. 183 pp.

490 R W GIBSON. J P LEGG AND G W OTIM-NAPE
Legg J P, Gibson R W, 0 th -Nape G W. 1994. Genetic polymorphism amongst Ugandan populations of Beniisia tabaci (Gennadius)(Homoptera: Aleyrodidae), vector of African cassava mosaic geminivirus. Tropical Science 34:82-Y 1.
Perring T M, Cooper A, Kazmer D J, Shields C, Shields J. 1991. New strain of sweetpotato whitefly invades California vegetables. California Agriculture 45: 10-12.
Perring T M, Cooper A D, Rodriguez R J, Farrar C A, Bellows T S. 1993. Identification of a whitefly species by genomic and behavioural studies. Science 259:74-77.
Storey H H. 1936. Virus diseases of East African plants: VI - A progress report on studies of the disease of cassava. The East African Agricultural Journal 2:34-39.
Storey H H, Nichols R F W. 1938. Studies of the mosaic diseases of cassava. Annals of Applied Biologv
Swanson M M, Harrison B D. 1994. Properties, relationships and distribution of cassava mosaic geminiviruses. Tropical Science 34: 15-25.
Thresh J M, 0 th -Nape G W, Jennings D L. 1994. Exploiting resistance to African cassava mosaic virus. Aspects of Applied Biology39, The lmpact of Genetic Variation on Sustainable Agriculture, pp. 5140 .
251790-806.
(Received 25 January 1996)
Related Documents











