ACTA BOTANICA UNIVERSITATIS COMENIANAE 57/2021 COMENIUS UNIVERSITY IN BRATISLAVA

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A C T A B O T A N I C A
UNIVERSITATISCOMENIANAE
57/202 1
COMENIUS UNIVERSITY IN BRATISLAVA

Scope of JournalThe journal Acta Botanica Universitatis Comenianae publishes papers dealing with phytocoenology, plant biology and morphology, phytogeography, environmental botany, ecology and taxonomy of non-vascular and vascular plants. It is published twice per year. All published papers are reviewed; list of reviewers of each volume is published on the inside cover.
Zameranie časopisuČasopis Acta Botanica Universitatis Comenianae publikuje príspevky z problematiky fytocenológie, biológie a morfológie rastlín, fytogeografie, environmentálnej botaniky, ekológie a taxonómie bezcievnych a cievnatých rastlín. Vychádza dvakrát ročne. Všetky príspevky sú recenzované; zoznam recenzentov každého čísla je uverejnený na vnútornej strane obálky.
Reviewers / RecenzentiMgr. Silvia Kubalová, PhD. (Comenius University in Bratislava, Faculty of Natural Sciences)RNDr. Ivona Kautmanová, PhD. (Slovak National Museum, Museum of Natural History, Bratislava)Mgr. Katarína Godovičová (Comenius University in Bratislava, Faculty of Natural Sciences) Ing. Mgr. Eva Zahradníková, PhD. (Comenius University in Bratislava, Faculty of Natural Sciences)RNDr. Hubert Žarnovičan, PhD. (Comenius University in Bratislava, Faculty of Natural Sciences)

ACTABOTANICA
UNIVERSITATISCOMENIANAE
Volume 57
2021COMENIUS UNIVERSITY IN BRATISLAVA

The journal was edited with the title / Časopis bol vydávaný pod názvom Acta Facultatis rerum naturalium Universitatis Comenianae, Botanica
Editor in Chief / Predseda redakčnej radyKarol Mičieta; [email protected]
Executive Editor / Výkonný redaktorSoňa Jančovičová, [email protected]
Editorial Board / Členovia redakčnej radyDana Bernátová, Michal Hrabovský, Katarína Mišíková, Jana Ščevková, Eva Zahradníková
Editor Ship / Adresa redakcieRedakcia Acta Botanica Universitatis Comenianae, Révová 39, SK-811 02 Bratislava 1Tel. ++421 2 54411541Fax ++421 2 54415603
Cover picture: freepik.com
Published by / Vydavateľ© Comenius University in Bratislava, 2021© Univerzita Komenského v Bratislave, 2021
ISSN 2729-7330

5
Acta Botanica Universitatis Comenianae Vol. 57, 2021
Peľová sezóna 2017 na území BratislavyPollen season 2017 in tHe Bratislava area
Jozef Dušička, Jana Ščevková*
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 811 02 Bratislava, Slovakia
Received 17 Febuary 2021; Received in revised form 18 February 2021; Accepted 19 February 2021
Abstract
From March to October 2017, we collected pollen grains of seed plants in the atmosphere of Bratislava using the Hirst-type pollen trap for their identification and quantification. During the reporting period, we registered 36,101 pollen grains, which belonged to 30 plant taxa. The taxa contributing to the highest concentration of pollen grains were Betula (13.97 %), Cupressaceae/Taxaceae (13.62 %), Urticaceae (12.84 %), Pinaceae (10.88 %), Quercus (9.46 %), Populus (9.25 %), Fraxinus (8.22 %) and Ambrosia (5.25 %). During the analysed period, we observed significant oscillations in the number of pollen grains in the air. The maximum daily pollen concentrations were recorded during the spring (March−May) and in summer (August). The knowledge of the seasonal pattern of the type and number of pollen grains in the air will provide clinicians and sufferers of pollen allergy with valuable information on the prophylaxis of respiratory allergic diseases
Key words: aeropalynology, pollen grains, pollinosis
úvod
Peľové zrná anemofilných rastlín sú jednou z hlavných príčin vzniku alergických respiračných ochorení. V ovzduší Bratislavy sa alergénne peľové zrná vyskytujú najmä počas vegetačného obdobia, pričom v dôsledku diaľkových atmosférických prenosov nie sú peľové zrná v ovzduší vzácnosťou ani počas zimných mesiacov, v čase vegetačného pokoja. Tieto peľové zrná sú na naše územie transportované z južnejšie položených geografických oblastí s teplejšou klímou. Nemenej dôležitou je skutočnosť, že v dôsledku globálneho otepľovania dochádza tiež k zmenám fenofáz rastlín, čím sa posúva aj obdobie ich polinácie. Z uvedeného vyplýva, že atmosféra predstavuje potenciálne riziko vzniku polinózy v ktoromkoľvek ročnom období. Na druhej strane, navodenie alergických symptómov u senzitívnych jedincov závisí od miery expozície, a to ako z kvantitatívneho, tak aj z časového hľadiska.
Kvitnutie rastlín, ktoré produkujú alergénne peľové zrná, je na našom území rozdelené na celé vegetačné obdobie. Stromy kvitnú na jar, najmä v marci a apríli. Trávy kvitnú koncom jari, od druhej polovice mája do druhej polovice júna. Neskoré leto – skorá jeseň (august, september) je obdobím, kedy kvitnú ostatné byliny (Jurko 1990). Informácie
* Corresponding author: [email protected]

6
o výskyte alergénnych peľových zŕn v ovzduší v priebehu kalendárneho roka sú z hľadiska liečby či prevencie polinóznych ochorení veľmi dôležité. Od roku 2002 je na Katedre botaniky Prírodovedeckej fakulty UK v Bratislave zriadená aerobiologická monitorovacia stanica, ktorá je súčasťou slovenskej Peľovej informačnej služby (PIS), ako aj Európskej aeroalergénnej siete (European Aeroallergen Network – EAN). Od uvedeného roku takmer nepretržite, s výnimkou zimných mesiacov so snehovými zrážkami, sledujeme zastúpenie peľových zŕn a ich kvantitu v ovzduší hlavného mesta.
Cieľom tejto práce je prezentácia výsledkov aeropalynologickej analýzy na území Bratislavy v roku 2017 so zameraním na druhové spektrum a kvantitu alergénnych peľových zŕn v ovzduší.
materiál a metódy
Aeropalynologická analýza sa uskutočnila v Bratislave, v meste s teplým a suchým kontinentálnym podnebím, s priemernými teplotami v rozmedzí od -1 do -4 °C v januári a od 19,5 do 20,5 °C v júli. Ročné zrážky sa pohybujú v priemere od 530 do 650 mm (Hrvoľ 2014).
Priemerné denné koncentrácie peľu sme monitorovali od marca do októbra 2017 pomocou Burkardovho volumetrického peľového lapača Hirstovho typu (Hirst 1952). Lapač bol umiestnený na streche budovy Katedry botaniky Prírodovedeckej fakulty UK (48°08′46″ S, 17°04′43″ V), vo výške 10 m nad zemou v nadmorskej výške 183 m. Použité metodické postupy odberu vzoriek, prípravy mikroskopických preparátov, ich analýzy a finálnej korekcie nameraných údajov sú v súlade s požiadavkami European Aerobiology Society (EAS) (Galán et al. 2014). Na odčítanie peľových zŕn zo vzorky sme použili metódu 12 zvislých transektov pri zväčšení × 400 podľa metodiky publikovanej v práci Ščevková et al. (2010). Výsledkom aeropalynologickej analýzy boli údaje o priemernej dennej koncentrácii peľových zŕn v metri kubickom vzduchu (PZ/m3). Hlavnú peľovú sezónu vybraných taxónov sme vymedzili podľa metódy autorov Nilsson, Persson (1981), ktorí definujú začiatok/koniec peľovej sezóny ako deň, kedy kumulatívna suma peľu dosahuje hodnotu 5 %, resp. 95 % zo sumárnej ročnej koncentrácie peľu.
V práci sme vyhodnotili vplyv teploty vzduchu (ºC), relatívnej vlhkosti vzduchu (%) a zrážok (mm) na koncentráciu dominantne zastúpených typov peľových zŕn v ovzduší s použitím Spearmanovej korelačnej analýzy. Meteorologické údaje boli zaznamenané na meteorologickej stanici Katedry astronómie, fyziky Zeme a meteorológie FMFI UK vzdialenej 1 km SZ od monitorovacej stanice.

7
výsledky a diskusia
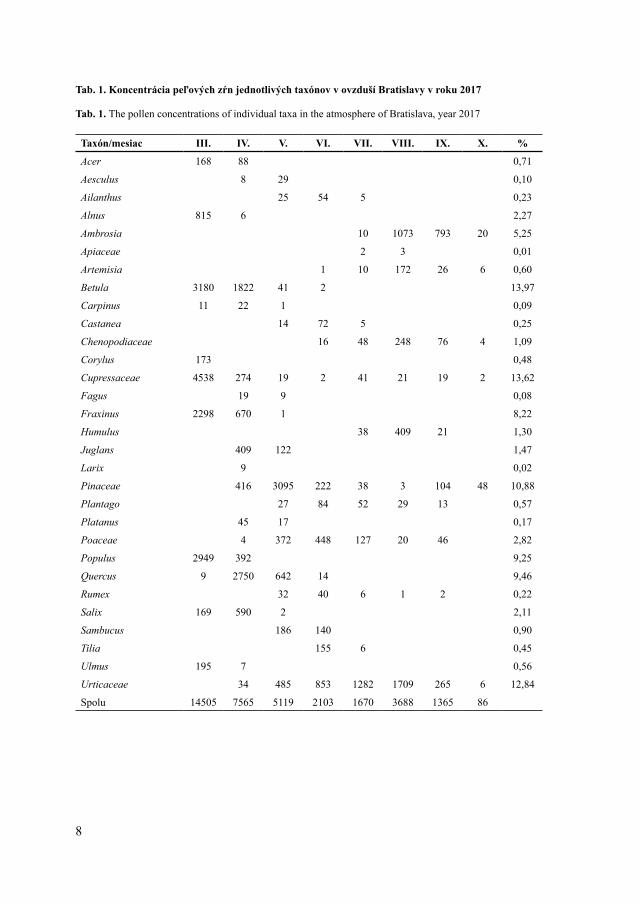
V roku 2017 sme v ovzduší Bratislavy zaznamenali peľové zrná 30 taxónov cievnatých rastlín (z toho 20 drevín a 10 bylín), sumárne 36 101 peľových zŕn. Medzi dominantne zastúpené taxóny patrili Betula (13,97 % z celkového počtu všetkých peľových zŕn), Cupressaceae/Taxaceae (ďalej uvádzané ako Cupressaceae, 13,62 %), Urticaceae (12,84 %), Pinaceae (10,88 %), Quercus (9,46 %), Populus (9,25 %), Fraxinus (8,22 %) a Ambrosia (5,25 %) (Tab. 1).
Najviac peľových zŕn sme zaznamenali v jarných mesiacoch (marec ‒ máj), s dominantným zastúpením peľových zŕn drevín, hlavne taxónov Betula, Populus, Fraxinus, Cupressaceae a Pinaceae (Tab. 1), čo je v súlade s inými štúdiami z územia Bratislavy (Ščevková et al. 2010, 2015). Množstvo peľu v ovzduší opätovne vzrástlo v auguste, v dôsledku kvitnutia invázneho druhu Ambrosia artemisiifolia, ktorý je známy vysokou produkciou peľu (Sikoparija et al. 2017).
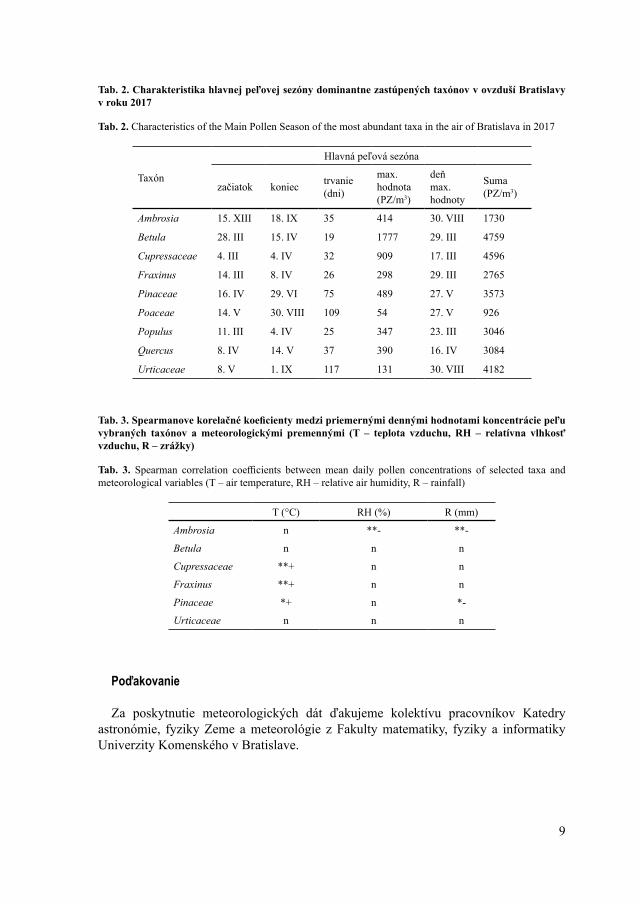
Najdlhšiu peľovú sezónu sme zaznamenali pri čeľadi Urticaceae (117 dní) a najkratšiu pri rode Betula (19 dní) (Tab. 2). Dlhú peľovú sezónu sme pozorovali aj u tráv (109 dní), čo súvisí s veľkou druhovou pestrosťou v rámci tejto skupiny rastlín a ich odlišnou fenológiou. Nakoľko sú peľové zrná tráv morfologicky podobné a nedajú sa diferencovať na úroveň druhov, vyčleňujeme peľovú sezónu pre všetkých zástupcov čeľade spolu, čo prirodzene ich sezónu predlžuje. Pre taxóny, najmä zo skupiny drevín, ktorých peľová sezóna je krátka, sú typické vysoké denné koncentrácie peľu v ovzduší (Ščevková et al. 2010). Aj v rámci tejto štúdie sme spomedzi všetkých analyzovaných taxónov zaznamenali najvyššiu maximálnu dennú koncentráciu (1 777 PZ/m3) u brezy a to 29. marca (Tab. 2).
Meteorologické faktory, najmä teplota vzduchu, relatívna vlhkosť vzduchu a zrážky priamo ovplyvňujú uvoľňovanie peľových zŕn z mikrosporangií a ich šírenie v atmosfére. Slnečné a teplé počasie všeobecne korešponduje s vyššou produkciou peľu, zatiaľ čo chladné a vlhké počasie zabraňuje uvoľňovaniu peľových zŕn z peľníc a zrážky dokonca priamo eliminujú peľové zrná z atmosféry (Bartková-Ščevková 2003).
Z výsledkov Spearmanovej korelačnej analýzy vyplýva, že so zvyšujúcou sa teplotou vzduchu množstvo peľových zŕn taxónov Cupressaceae, Fraxinus a Pinaceae v ovzduší narastá. So zvyšujúcou sa vlhkosťou ovzdušia naopak klesá množstvo peľových zŕn taxónov Ambrosia a Pinaceae (Tab. 3). Na rozdiel od predchádzajúcich štúdií z územia Bratislavy (Ščevková et al. 2010, 2015), ktoré poukazujú na signifikantnú korelačnú závislosť aj medzi koncentráciou peľu taxónov Betula a Urticaceae a teplotou vzduchu a/alebo vlhkosťou vzduchu, výsledky našich analýz vyšli pri týchto taxónoch ako nepreukazné (Tab. 3).
8
Tab. 1. Koncentrácia peľových zŕn jednotlivých taxónov v ovzduší Bratislavy v roku 2017
Tab. 1. The pollen concentrations of individual taxa in the atmosphere of Bratislava, year 2017
Taxón/mesiac III. IV. V. VI. VII. VIII. IX. X. %
Acer 168 88 0,71
Aesculus 8 29 0,10
Ailanthus 25 54 5 0,23
Alnus 815 6 2,27
Ambrosia 10 1073 793 20 5,25
Apiaceae 2 3 0,01
Artemisia 1 10 172 26 6 0,60
Betula 3180 1822 41 2 13,97
Carpinus 11 22 1 0,09
Castanea 14 72 5 0,25
Chenopodiaceae 16 48 248 76 4 1,09
Corylus 173 0,48
Cupressaceae 4538 274 19 2 41 21 19 2 13,62
Fagus 19 9 0,08
Fraxinus 2298 670 1 8,22
Humulus 38 409 21 1,30
Juglans 409 122 1,47
Larix 9 0,02
Pinaceae 416 3095 222 38 3 104 48 10,88
Plantago 27 84 52 29 13 0,57
Platanus 45 17 0,17
Poaceae 4 372 448 127 20 46 2,82
Populus 2949 392 9,25
Quercus 9 2750 642 14 9,46
Rumex 32 40 6 1 2 0,22
Salix 169 590 2 2,11
Sambucus 186 140 0,90
Tilia 155 6 0,45
Ulmus 195 7 0,56
Urticaceae 34 485 853 1282 1709 265 6 12,84
Spolu 14505 7565 5119 2103 1670 3688 1365 86
9
Tab. 2. Charakteristika hlavnej peľovej sezóny dominantne zastúpených taxónov v ovzduší Bratislavy v roku 2017
Tab. 2. Characteristics of the Main Pollen Season of the most abundant taxa in the air of Bratislava in 2017
Taxón
Hlavná peľová sezóna
začiatok koniec trvanie(dni)
max. hodnota(PZ/m3)
deň max.hodnoty
Suma(PZ/m3)
Ambrosia 15. XIII 18. IX 35 414 30. VIII 1730
Betula 28. III 15. IV 19 1777 29. III 4759
Cupressaceae 4. III 4. IV 32 909 17. III 4596
Fraxinus 14. III 8. IV 26 298 29. III 2765
Pinaceae 16. IV 29. VI 75 489 27. V 3573
Poaceae 14. V 30. VIII 109 54 27. V 926
Populus 11. III 4. IV 25 347 23. III 3046
Quercus 8. IV 14. V 37 390 16. IV 3084
Urticaceae 8. V 1. IX 117 131 30. VIII 4182
Tab. 3. Spearmanove korelačné koeficienty medzi priemernými dennými hodnotami koncentrácie peľu vybraných taxónov a meteorologickými premennými (T – teplota vzduchu, RH – relatívna vlhkosť vzduchu, R – zrážky)
Tab. 3. Spearman correlation coefficients between mean daily pollen concentrations of selected taxa and meteorological variables (T – air temperature, RH – relative air humidity, R – rainfall)
T (°C) RH (%) R (mm)
Ambrosia n **- **-
Betula n n n
Cupressaceae **+ n n
Fraxinus **+ n n
Pinaceae *+ n *-
Urticaceae n n n
Poďakovanie
Za poskytnutie meteorologických dát ďakujeme kolektívu pracovníkov Katedry astronómie, fyziky Zeme a meteorológie z Fakulty matematiky, fyziky a informatiky Univerzity Komenského v Bratislave.

10
literatúra
Bartková-Ščevková, J., 2003: The influence of temperature, relative humidity and rainfall on the occurrence of pollen allergens (Betula, Poaceae, Ambrosia artemisiifolia) in the atmosphere of Bratislava (Slovakia). Int J Biometeorol, 48: 1-5.
Galán, C., Smith, M., Thibaudon, M., Frenguelli, G., Oteros, J. A., Gehrig, R., et al., 2014: Pollen monitoring: minimum requirements and reproducibility of analysis. Aerobiologia, 30: 385-395.
Hirst, J. M., 1952: An automatic volumetric spore trap. Ann Appl Biology, 39: 257-265.Hrvoľ, J., 2014: Extrémne teploty vzduchu na stanici Bratislava, Mlynská dolina za obdobie 1983-2012. In:
Čelková, A. (ed.), 21st international poster day transport of water, chemicals and energy in the soil-plant-atmosphere system, p. 93-101, Bratislava.
Jurko, A., 1990: Sezonalita kvitnutia rastlín a peľové alergény v našej vegetácii. Biológia, Bratislava, 45: 367-374.
Nilsson, S., Persson, S., 1981: Tree pollen spectra in the Stockholm region (Sweden), 1973-1980. Grana, 20: 179-182.
Ščevková, J., Dušička, J., Chrenová, J., Mičieta, K., 2010: Annual pollen spectrum variations in the air of Bratislava (Slovakia): years 2002–2009. Aerobiologia, 26: 277-287.
Ščevková, J., Dušička, J., Mičieta, K., 2015: Aerobiological analysis of airborne pollen and fungal spore fall in Bratislava in 2015. Acta Botanica Universitatis Comenianae, 50: 3-8.
Šikoparija, B., Skjøth, C. A., Celenk, S., Testoni, C., Abramidze, T., Scevkova, J., et al., 2017: Spatial and temporal variations in airborne Ambrosia pollen in Europe. Aerobiologia, 33: 181-189.
Abstrakt
V ovzduší Bratislavy sme od marca do októbra 2017 zachytávali peľové zrná semenných rastlín pomocou peľového lapača Hirstovho typu za účelom ich identifikácie a kvantifikácie. Počas sledovaného obdobia sme sumárne zaznamenali 36 101 peľových zŕn, ktoré patrili 30 rastlinným taxónom. Dominovali peľové zrná taxónov Betula (13,97 %), Cupressaceae/Taxaceae (13,62 %), Urticaceae (12,84 %), Pinaceae (10,88 %), Quercus (9,46 %), Populus (9,25 %), Fraxinus (8,22 %) a Ambrosia (5,25 %). Počas analyzovaného obdobia sme sledovali výrazné oscilácie v kvantite peľových zŕn v ovzduší. Maximálne denné koncentrácie peľových zŕn sme zaznamenali počas jarných mesiacov (marec − máj) a v lete (august). Poznatky týkajúce sa sezónnej variability druhového spektra a koncentrácie jednotlivých typov peľových zŕn v ovzduší poskytnú lekárom a osobám trpiacim peľovou alergiou cenné informácie o profylaxii respiračných alergických ochorení.
Kľúčové slová: aeropalynológia, peľové zrná, polinóza

11
Acta Botanica Universitatis Comenianae Vol. 57, 2021
let’s go to tHe field. Botanical excursion 4Poďme sPolu do terénu. Botanická exkurzia 4
Katarína Mišíková*, Jozef Dušička, Ján Miškovic, Martina Michalová, Soňa Jančovičová
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 811 02 Bratislava, Slovakia
Received 8 March 2021, Received in revised form 17 March 2021, Accepted 18 March 2021
Abstract
The 4th botanical excursion within the series „Let‘s go to the field″ took place in the Malé Karpaty Mts. in the wine-growing town of Svätý Jur and its surroundings. The field trip leads through the deciduous mixed forests and habitats of the cultural landscape, such as vineyards, ruins of the Biely Kameň Castle, anthropogenic and ruderal sites. On five localities, we focused on basic floristic research, as well as the monitoring the occurrence and distribution of archaeophytes, neophytes and invasive species. Overall, we recorded 63 species of vascular plants, 41 bryophytes (three species of liverworts and 38 species of mosses), 12 lichens and 44 taxa of macroscopic fungi. Valuable findings include confirmation of the occurrence of critically endangered moss Rhynchostegium rotundifolium on the ruins of the Biely Kameň Castle.
Key words: Slovakia, Malé Karpaty Mts., vineyards, castle ruins, vascular plants, bryophytes, lichenised fungi, non-lichenised macroscopic fungi
introduction
The 4th botanical excursion was intended for students of the second semester of the master´s study; the course is called Botany field practice. The excursion took place in the town Svätý Jur, along the route of the vineyard educational trail to the Biely Kameň Castle ruins. The field trip leads through the deciduous mixed forests (alliance Carpinion betuli Issler 1931 em. Meier 1937) and habitats of the cultural landscape, such as vineyards, Castle ruins, anthropogenic and ruderal sites.
During the excursion, we visited a historical and cultural monument, the ruins of the Biely Kameň Castle, and focused on the photoautotrophic and heterotrophic organisms inhabiting them (bryophytes, algae, vascular plants, lichenised and non-lichenised fungi), as well as plant communities in anthropogenic and ruderal habitats. In the vineyards, we investigated the occurrence of archaeophytes, neophytes and invasive plant species, and cultivation methods and fungal diseases of the vine. As a part of the history of botanical research, we reminded some important scientists of Slovakia who botanized here, such as J. Ľ. Holuby, J. Baumgartner and A. Zahlbruckner.
* Corresponding author: [email protected]

12
The town Svätý Jur is located northeast of Bratislava on the slopes of the Malé Karpaty Mts. It is one of the architecturally best-preserved wine-growing towns in Slovakia; the first written mention is from 1209. In 1647, it was promoted to a free royal town. Important historical and cultural monuments include, for example, the Gothic church of St. George from the end of the 13th century and the wooden bell tower next to it (17th century), the Great Moravian fortified settlement above the town part Neštich and the ruins of the Biely Kameň Castle from the 13th century that was destroyed in 1663 (Anonymus 1, Mereďa 2009). On the Prostredná street, there is a memorial plaque to the world-famous native – lichenologist Alexander Zahlbruckner (* 31 May 1860 Svätý Jur; † 8 May 1938 Vienna). His most important work is a comprehensive catalogue “Catalogus lichenum universalis (Vols. I-X)” that presents a list of all in that time known lichens including complete bibliographic sources. The Catalogus has become a permanent source for the study of lichens (Pišút 2002).
Vineyards, together with orchards, form a vine-growing cultural region, which represents an important refuge for many organisms in the cultural landscape (Eliáš 2009). The Small Carpathian region is the biggest vineyard region in Slovakia with a vineyard area of 5 000 ha (Anonymus 3). Old stone walls, in Slovak called „kamenice″ or „rúny″ are a remarkable part of the vineyards in the Malé Karpaty Mts. They were formed by removing rocks and stones from vineyards on weathered bedrock, which create granodiorites, granites and gneisses. The old stone walls were usually laid across the contour lines, sometimes forming boundaries or terraces between individual vineyards. In the forest stands, the presence of old stone walls indicates the extinct vineyards that were here in the past (Anonymus 1).
Tab. 1. Localities of the field excursion to the Biely Kameň Castle ruins, 10th September 2020.A – locality code; elevation (m a. s. l.); GPS coordinates (DMS); B – position, specifics; C – vegetation, bold – dominant species, (%) – percentage of the most abundant tree species (according to Anonymus 2).
Tab. 1. Lokality exkurzie na ruinu hradu Biely Kameň, 10. september 2020. A – číslo lokality; nadmorská výška (m n. m.); GPS súradnice (DMS); B – poloha, špecifiká lokality; C – vegetácia, tučným písmom dominantné druhy, (%) – percento najviac zastúpených druhov drevín (podľa Anonymus 2).
A B C
1 Mariánska cesta street; 220; 48°15′25.4″N, 17°12′21.7″E
Acer campestre, Euonymus europaeus, Prunus cerasifera, Rubus caesius agg., Syringa vulgaris
2 Vineyards; 240–300; 48°15′21.2″N, 17°12′15.1″E
Acer campestre, Castanea sativa, Prunus cerasifera, Robinia pseudoaccacia, Rosa canina, Rubus fruticosus agg., Syringa vulgaris, Vitis vinifera
3 Old stone walls in vineyards and forests; 330; 48°15′12.8″N, 17°11′35.3″E
Acer campestre, Acer pseudoplatanus, Carpinus betulus, Castanea sativa, Pinus sylvestris, Quercus petraea agg., Robinia pseudoaccacia, Rubus fruticosus agg., Syringa vulgaris, Vitis vinifera
4 Mixed deciduous forests; 330–350; 48°15′31.1″N, 17°11′19.1″E
Acer pseudoplatanus, Carpinus betulus (10–20%), Fagus sylvatica (10%),, Quercus petraea agg. (70–80%)
5 Ruins of Biely Kameň Castle and surrounding forests; 300; 48°15′39.8″N, 17°11′47.8″E
Acer pseudoplatanus (10%), Carpinus betulus (50–70%), Fagus sylvatica (10%), Quercus petraea agg. (30–40%), Tilia cordata (5%)

13
material and methods
Localities of the field work. The excursion took place on September 10, 2020 in the town Svätý Jur, within the phytogeographical area Malé Karpaty Mts. (Tab. 1, Fig. 1).
Vascular plants. Vascular plants were identified using the indentification key by Kaplan et al. (2019). The nomenclature of vascular plants follows Marhold et al. (1998), threat categories are according to Eliáš et al. (2015) and non-native taxa are according to Medvecká et al. (2012).
Bryophytes. Bryophytes were recorded on various types of substrates, e.g. rotten wood, bark of trees, soil and stones. The most of bryophytes were identified in the field based on the macromorphological characteristics. The micromorphological characteristics were observed (under the microscope) to identify the collections of the genera Plagiothecium and Pseudoleskeella. The publication by Smith (2004) was mostly used for species identification.The nomenclature of mosses and threat categories follows the study by Mišíková et al. (2020) and of liverworts by Mišíková et al. (2021).
Lichenised fungi. As we lacked experts on these groups of organisms, we focused our field research only on macroscopic, easily recognizable species. Nomenclature of lichenised fungi follows Guttová et al. (2013).
Fig. 1. Localities of the field excursion to the Biely Kameň Castle ruins: 1 – Mariánska cesta street, 2 – Vineyards, 3 – Old stone walls in vineyards and forests, 4 – Forests, 5 – Castle ruin Biely kameň and surrounding forests; for more information about localities see Tab. 1. (Source: https://sk.mapy.cz/; modified by K. Mišíková).
Obr. 1. Lokality exkurzie na ruinu hradu Biely Kameň: 1 – Mariánska cesta, 2 – vinice, 3 – rúny vo vinohradoch a lesoch, 4 – lesy, 5 – ruina hradu Biely kameň a okolité lesy; pre viac informácií k lokalitám pozri Tab. 1. (Zdroj: https://sk.mapy.cz/; upravila K. Mišíková).

14
Non lichenised macroscopic fungi. Fruiting bodies visible to the naked eye of Ascomycota and Basidiomycota (randomly of Myxogastria) were collected, identified and processed by standard mycological techniques (e.g. Holec et al. 2012). Ecological entries were given for each collection following Ripková et al. (2007). The publications by Holec et al. (2012), Læssøe, Petersen (2019) and Knudsen, Vesterholt (2012) were mostly used for species identification. The nomenclature of the recorded fungi follows the Index Fungorum database (Kirk 2021). The specimens are deposited in the herbarium SLO (the herbarium of the Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany).
results and discussion
Vascular plants During the field excursion to the Biely Kameň Castle Ruins, we recorded in total
63 taxa of vascular plants (20 woody plants and 43 herbaceous plants) (Tab. 2). The highest number of species we observed at locality 5 – Ruins of Biely Kameň Castle and surrounding forests with 27 plant species.
From the important species of vascular plants, two critically endangered taxa are mentioned from the area of St. George. The archaeophyte Polycarpon tetraphyllum belongs to the Mediterranean elements of the Slovak flora, occurring mainly in vineyards and gardens in the southwestern part of the state (Mereďa 2012). Recent localities are recorded from Bratislava (Mereďa 2012, Májeková et al. 2020), from Sv. Jur (Mereďa 2012) and from Modra (leg. K. Mišíková 2018, unpublished, herbarium SLO). Ruscus hypoglossum is an endangered species (EN) from the family Liliaceae. in Slovakia, it represents a tertiary relic on the northern border of its distribution. It grows mainly in oak-hornbeam forests in the southeastern part of the Malé Karpaty Mts. For its protection, the Protected Area Svätojurské hradisko was established in 2002 (Mereďa 2009). During the excursion, we have not found either of these two species. On the other hand, we observed several non-native taxa. Medvecká et al. (2012) use categories for the invasion status (IS): cas – casual, nat – naturalized, inv – invasive; and residence time (RT): arch – archaeophyte, neo – neophyte. We found naturalized archaeophytes Berteroa incana, Cichorium intybus, Chelidonium majus, Setaria pumila, Solanum nigrum; naturalized neophytes Syringa vulgaris and invasive neophytes Ambrosia artemisiifolia, Erigeron annuus and Impatiens parviflora. The invasive taxa pose threat to the local biodiversity.
Locality 1. Synanthropic and ruderal vegetation was typical for this locality represented by taxa Cichorium intybus, Clematis vitalba, Geum urbanum, Humulus lupulus, Chelidonium majus, Plantago major, Polygonum aviculare agg., Rubus caesius agg., Taraxacum sect. Ruderalia, Urtica dioica and invasive species Impatiens parviflora.
Locality 2. Invasive neophytes (Ambrosia artemisiifolia, Erigeron annuus) occurred significantly on the locality Vineyards. There were also naturalized archaeophytes Berteroa incana and Solanum nigrum.
Locality 3. Locality Old stone walls in vineyards and forests was dominated by Quercus petraea agg. Herbaceous plants of the forest such as Galium odoratum, Geranium robertianum, Ligustrum vulgare, Alliaria petiolata and invasive taxon

15
Impatiens parviflora were also present. Locality 4. Vascular plants in this locality were dominated by woody plants such as
Quercus petraea agg., Fagus sylvatica and Carpinus betulus. Herbaceous plants of the forest Melica uniflora, Convallaria majalis, Lamium maculatum and Vinca minor were also present.
Locality 5. The highest number of vascular plant species we observed on the Ruins of Biely Kameň Castle and surrounding forests represented especially by Carpinus betulus and also by Quercus petraea agg. and herbaceous plants of the forest such as Galium odoratum, Melica uniflora, Hieracium murorum, Mycelis muralis, Pulmonaria officinalis, Polygonatum multiflorum and Aegopodium podagraria.
Bryophytes and LichensOverall, 12 lichen and 41 bryophyte species were found in the studied area, including
three liverworts (Frullania dilatata, Porella platyphylla, Radula complanata) and 38 mosses (Tab. 3). Up to date, there is a lack of data from the area of the Svätý Jur. The epilithic bryoflora of Biely Kameň astle was processed by Uhereková Šmelková et al. (2011), bryophytes on cemeteries were studied by Mišíková et al. (2018).
Tab. 2. The vascular plant list. A – locality code: 1 – Mariánska cesta street, 2 – Vineyards, 3 – Old stone walls in vineyards and forests, 4 – Forests, 5 – Castle ruin Biely Kameň and surrounding forests; for more information about localities see Tab. 1; B – the list of recorded taxa. The taxa are in alphabetical order.
Tab. 2. Zoznam vyšších rastlín. A – číslo lokality: 1 – Mariánska cesta, 2 – vinice, 3 – rúny vo vinohradoch a lesoch, 4 – lesy, 5 – ruina hradu Biely Kameň a okolité lesy, pre viac informácií k lokalitám pozri Tab. 1; B – zoznam zaznamenaných taxónov. Taxóny sú zoradené v abecednom poradí.
A B
1 Acer campestre, Berberis vulgaris, Cichorium intybus, Clematis vitalba, Euonymus verrucosus, Geum urbanum, Hedera helix, Humulus lupulus, Chelidonium majus, Impatiens parviflora, Syringa vulgaris, Lamium maculatum, Lotus corniculatus, Plantago major, Polygonum aviculare agg., Prunus cerasife-ra, Pyrus sp., Rubus caesius agg., Sambucus nigra, Taraxacum sect. Ruderalia, Tilia cordata, Ulmus carpinifolia, Urtica dioica.
2 Acer campestre, Asplenium trichomanes, Ambrosia artemisiifolia, Berteroa incana, Castanea sativa, Erigeron annuus, Lactuca serriola, Prunus cerasifera, Rosa canina agg., Setaria pumila, Solanum nigrum, Syringa vulgaris, Robinia pseudoaccacia, Rosa canina, Rubus fruticosus agg., Tithymalus cyparissias, Trifolium repens, Vitis vinifera.
3 Acer campestre, A. pseudoplatanus, Alliaria petiolata, Carpinus betulus, Castanea sativa, Galium mollugo agg., Galium odoratum, Geranium robertianum, Hedera helix, Hieracium laciniatum, Im-patiens parviflora, Ligustrum vulgare, Melampyrum nemorosum, Pinus sylvestris, Quercus petraea agg., Robinia pseudoaccacia, Rosa canina agg., Rubus fruticosus agg., Stellaria media, Tilia cordata, Syringa vulgaris, Vitis vinifera.
4 Acer pseudoplatanus, Carpinus betulus, Convallaria majalis, Dryopteris filix-mas, Fagus sylvatica, Melica uniflora, Quercus petraea agg., Vinca minor.
5 Acer platanoides, A. pseudoplatanus, Aegopodium podagraria, Cardamine impatiens, Carpinus betu-lus, Clematis vitalba, Corylus avellana, Dryopteris filix-mas, Euonymus verrucosus, Fagus sylvatica, Galium odoratum, Geranium robertianum, Chelidonium majus, Hieracium murorum, Hylotelephium maximum, Impatiens parviflora, Juglans regia, Lamium maculatum, Lunaria rediviva, Hedera helix, Melica uniflora, Mycelis muralis, Pulmonaria officinalis, Polygonatum multiflorum, Quercus petraea agg., Sambucus nigra, Solidago virgaurea, Taraxacum sect. Ruderalia, Tilia cordata.

16
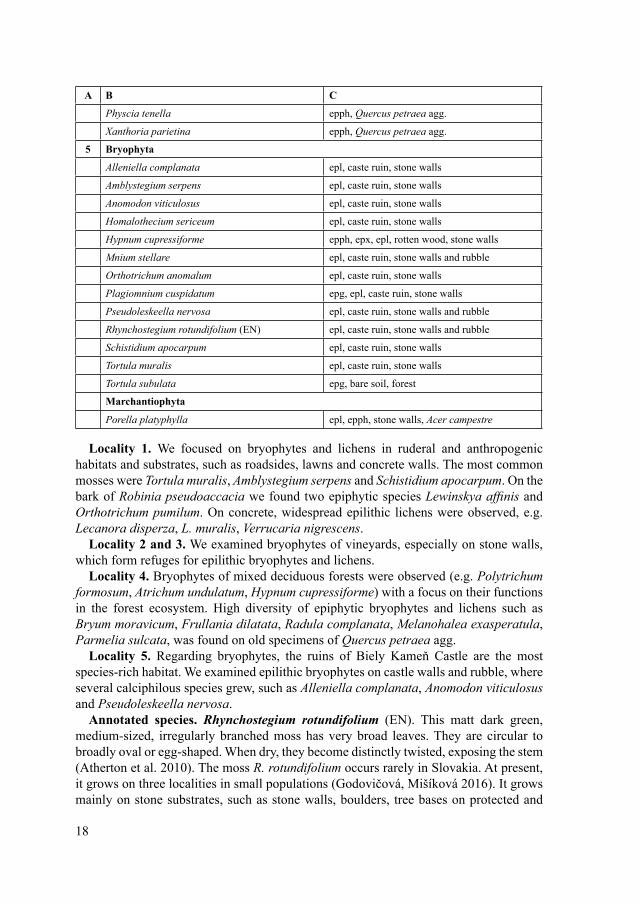
Tab. 3. The list of bryophytes and lichens recorded during the field excursion on the Biely Kameň Castle ruins. A – locality code: 1 – Mariánska cesta street, 2 – Vineyards, 3 – Old stone walls in vineyards and forests, 4 – Forests, 5 – Castle ruin Biely Kameň and surrounding forests; for more information about localities see Tab. 1; B – taxon; threat category; C – substrate: epg – epigeic species, epph – epiphytic species, epl – epilithic species, epx – epixylic species.
Tab. 3. Zoznam machorastov a lišajníkov zistených počas terénnej exkurzie na hrad Biely Kameň. A – číslo lokality: 1 – Mariánska cesta, 2 – vinice, 3 – rúny vo vinohradoch a lesoch, 4 – lesy, 5 – ruina hradu Biely Kameň a okolité lesy; pre viac informácií k lokalitám pozri Tab. 1; B – taxón; kategória ohrozenosti; C – substrát: epg – epigeický druh, epph – epifytický druh, epl – epilitický druh, epx – epixylický druh.
A B C
1 Bryophyta
Amblystegium serpens epg, epl, anthropogenic sites
Barbula unguiculata
Brachythecium albicans epg, soil among grass
Brachythecium rutabulum epg, soil among grass
Ceratodon purpureus epg, epl, anthropogenic sites
Encalypta streptocarpa epl, concrete
Grimmia pulvinata epl, concrete
Hypnum cupressiforme epph, Robinia pseudoaccacia
Lewinskya affinis epph, Robinia pseudoaccacia
Orthotrichum anomalum epl, concrete
Orthotrichum pumilum epph, Robinia pseudoaccacia
Oxyrrhynchium hians epg, bare soil
Rhynchostegium murale epl, concrete
Schistidium apocarpum epl, concrete
Syntrichia ruralis epg, epl, anthropogenic sites
Tortula muralis epl, concrete
Lichenes
Lecanora disperza epl, concrete
Lecanora muralis epl, concrete
Lecidella stigmatea epl, concrete
Physcia caesia epl, concrete
Physcia adcendens epph, Robinia pseudoaccacia
Verrucaria nigrescens epl, concrete
Xanthoria parietina epph, Robinia pseudoaccacia
2 Bryophyta
Amblystegium serpens epl, concrete
Anomodon viticulosus epl, old stone wall
Brachythecium albicans epg, dry soil among herb vegetation
Bryum argenteum epg, dry soil in vineyards
Bryum capillare epg, dry soil in vineyards

17
A B C
Ceratodon purpureus epg, dry soil in vineyards
Grimmia pulvinata epl, concrete, old stone walls
Homalothecium sericeum epl, old stone wall
Hypnum cupressiforme epl, old stone wall, epph Castanea sativa
Orthotrichum anomalum epl, concrete, old stone walls
Orthotrichum diaphanum epph, Robinia pseudoaccacia
Orthotrichum pumilum epph, Robinia pseudoaccacia
Osculatia caespiticia epg, dry soil in vineyards
Tortula muralis epl, concrete, old stone walls
Marchantiophyta
Porella platyphylla epph, Castanea sativa
3 Bryophyta
Ceratodon purpureus epl, old stone walls
Dicranum scoparium epl, old stone walls
Hedwigia ciliata epl, old stone walls
Hypnum cupressiforme epl, old stone walls
Plagiomnium cuspidatum epl, old stone walls
Lichenes
Parmelia saxatilis epl, old stone walls
4 Bryophyta
Atrichum undulatum epg, forests
Brachytheciastrum velutinum epg, forests
Brachythecium rutabulum epx, rotten wood in forests
Bryum moravicum epph, Quercus petraea agg.
Dicranella heteromalla epg, forests
Hypnum cupressiforme epg, epph, epx, rotten wood in forests
Plagiomnium cuspidatum epx, epg, forests
Plagiothecium nemorale epg, forests
Plagiothecium succulentum epg, forests
Platygyrium repens epph, Quercus petraea agg.
Polytrichum formosum epg, forests
Marchantiophyta
Frullania dilatata epph, Quercus petraea agg.
Radula complanata epph, Quercus petraea agg.
Lichenes
Hypogymnia physodes epph, Quercus petraea agg.
Melanohalea exasperatula epph, Quercus petraea agg.
Parmelia sulcata epph, Quercus petraea agg.
Physcia adcendens epph, Quercus petraea agg.
18
A B C
Physcia tenella epph, Quercus petraea agg.
Xanthoria parietina epph, Quercus petraea agg.
5 Bryophyta
Alleniella complanata epl, caste ruin, stone walls
Amblystegium serpens epl, caste ruin, stone walls
Anomodon viticulosus epl, caste ruin, stone walls
Homalothecium sericeum epl, caste ruin, stone walls
Hypnum cupressiforme epph, epx, epl, rotten wood, stone walls
Mnium stellare epl, caste ruin, stone walls and rubble
Orthotrichum anomalum epl, caste ruin, stone walls
Plagiomnium cuspidatum epg, epl, caste ruin, stone walls
Pseudoleskeella nervosa epl, caste ruin, stone walls and rubble
Rhynchostegium rotundifolium (EN) epl, caste ruin, stone walls and rubble
Schistidium apocarpum epl, caste ruin, stone walls
Tortula muralis epl, caste ruin, stone walls
Tortula subulata epg, bare soil, forest
Marchantiophyta
Porella platyphylla epl, epph, stone walls, Acer campestre
Locality 1. We focused on bryophytes and lichens in ruderal and anthropogenic habitats and substrates, such as roadsides, lawns and concrete walls. The most common mosses were Tortula muralis, Amblystegium serpens and Schistidium apocarpum. On the bark of Robinia pseudoaccacia we found two epiphytic species Lewinskya affinis and Orthotrichum pumilum. On concrete, widespread epilithic lichens were observed, e.g. Lecanora disperza, L. muralis, Verrucaria nigrescens.
Locality 2 and 3. We examined bryophytes of vineyards, especially on stone walls, which form refuges for epilithic bryophytes and lichens.
Locality 4. Bryophytes of mixed deciduous forests were observed (e.g. Polytrichum formosum, Atrichum undulatum, Hypnum cupressiforme) with a focus on their functions in the forest ecosystem. High diversity of epiphytic bryophytes and lichens such as Bryum moravicum, Frullania dilatata, Radula complanata, Melanohalea exasperatula, Parmelia sulcata, was found on old specimens of Quercus petraea agg.
Locality 5. Regarding bryophytes, the ruins of Biely Kameň Castle are the most species-rich habitat. We examined epilithic bryophytes on castle walls and rubble, where several calciphilous species grew, such as Alleniella complanata, Anomodon viticulosus and Pseudoleskeella nervosa.
Annotated species. Rhynchostegium rotundifolium (EN). This matt dark green, medium-sized, irregularly branched moss has very broad leaves. They are circular to broadly oval or egg-shaped. When dry, they become distinctly twisted, exposing the stem (Atherton et al. 2010). The moss R. rotundifolium occurs rarely in Slovakia. At present, it grows on three localities in small populations (Godovičová, Mišíková 2016). It grows mainly on stone substrates, such as stone walls, boulders, tree bases on protected and

19
shaded places. According to some authors (Hill et al. 1994), its occurrence in Central Europe in habitats such as castle ruins, walls and rubble points to an introduced origin. From the locality Biely Kameň Castle, which has been known for more than 100 years, R. rotundifolium was mentioned for the first time in 1901 (Baumgartner 1901). The species occurs here scattered in several small vital populations and regularly forms sporophytes. The ruins of the castle are forested with trees and the shading of the site contributes to the favorable condition of the populations. In the case of reconstruction works and deforestation of the ruins, this critically endangered species would not survive on the Biely Kameň Castle.
Hedwigia ciliata. Gray-green to brown-green moss growing in tufts or cushions. It is a xerophilous taxon on which we can demonstrate macro- and microscopic adaptations of poikilohydric bryophytes to a dry environment – leaves with a hyaline tip, markedly papillose cells, capsules with a short seta immersed in leaves. It grows mainly on silicate or weakly basic rocks and boulders exposed or slightly protected from the sun. During this field trip, we found it on old stone walls and granite boulders in oak forests above the vineyards.
Porella platyphylla. The foliose liverwort P. platyphylla is familiar to many botanists because it is larger than most others. Plants are up to 1.5–4 mm wide, and several centimeters long. It grows in loose patches or wefts of pinnate shoots (Atherton et al. 2010). Porella platyphylla is an abundant plant on calcareous boulders. It can also grow on old walls, in villages and towns. As epiphyte, it is often found on Fraxinus excelsior. From the educational point of view, due to its size, this species is suitable for demonstrating the structure of the foliose liverworts, as well as differences between a moss and a liverwort.
Non lichenised macroscopic fungi On 10 September 2020, we have recorded 44 macroscopic fungi during the botanical
excursion on the vineyard educational trail to the Biely Kameň Castle ruins (Tab. 4). Locality 1. Along the road at the beginning of our route, some microscopic fungi
can be demonstrated here in the spring. Distinctly elongated, flattened, hollow and light green coloured fruits of Prunus trees are characteristic symptoms of the ascomycetous pathogen Taphrina pruni (Fuckel) Tul. The presence of another pathogen, basidiomycete Uromyces pisi-sativi (Pers.) Liro, reveals morphologically changed habitus of Euphorbia sp. plants growing on open sites near the abandoned vineyards. Infected leaves on their bottom-side display distinct orange aecia producing aeciospores (Pfunder, Roy 2000).
On the day of our excursion we found a yellow staining and ink-like smelling Agaricus moelleri on soil on the side of the road under Rosa sp., Syringa vulgaris, Hedera helix and Acer campestre. It has brown-grey adpressed fine scales on the cap, smooth white stem broadening towards the base and double ring. The spores are ellipsoid, measuring 4.5–6.5 × 3–4 μm (cf. Knudsen et al. 2012).
Locality 3. We found basidiomata of only three species on the stand 3. Fomitoporia robusta, the large, hoof-shaped, perennial polypore grew on trunks of living Quercus tree. Gymnopus dryophilus, a yellow-brown collybioid, and Lycoperdon molle, a pale puffball with small grains and spines, occurred on mossy soil among stones.
20
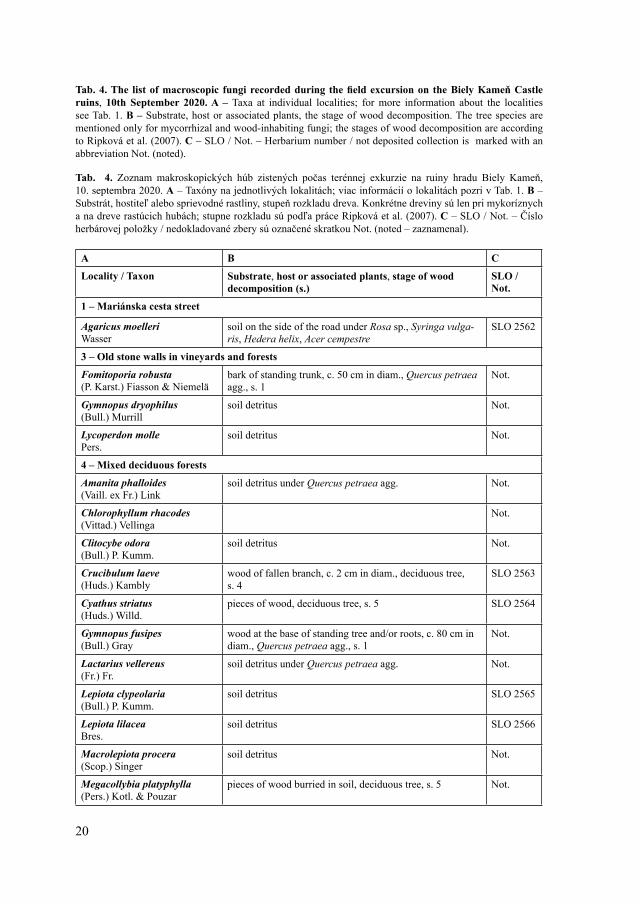
Tab. 4. The list of macroscopic fungi recorded during the field excursion on the Biely Kameň Castle ruins, 10th September 2020. A – Taxa at individual localities; for more information about the localities see Tab. 1. B – Substrate, host or associated plants, the stage of wood decomposition. The tree species are mentioned only for mycorrhizal and wood-inhabiting fungi; the stages of wood decomposition are according to Ripková et al. (2007). C – SLO / Not. – Herbarium number / not deposited collection is marked with an abbreviation Not. (noted).
Tab. 4. Zoznam makroskopických húb zistených počas terénnej exkurzie na ruiny hradu Biely Kameň, 10. septembra 2020. A – Taxóny na jednotlivých lokalitách; viac informácií o lokalitách pozri v Tab. 1. B – Substrát, hostiteľ alebo sprievodné rastliny, stupeň rozkladu dreva. Konkrétne dreviny sú len pri mykoríznych a na dreve rastúcich hubách; stupne rozkladu sú podľa práce Ripková et al. (2007). C – SLO / Not. – Číslo herbárovej položky / nedokladované zbery sú označené skratkou Not. (noted – zaznamenal).
A B C
Locality / Taxon Substrate, host or associated plants, stage of wood decomposition (s.)
SLO / Not.
1 – Mariánska cesta street
Agaricus moelleriWasser
soil on the side of the road under Rosa sp., Syringa vulga-ris, Hedera helix, Acer cempestre
SLO 2562
3 – Old stone walls in vineyards and forests
Fomitoporia robusta(P. Karst.) Fiasson & Niemelä
bark of standing trunk, c. 50 cm in diam., Quercus petraea agg., s. 1
Not.
Gymnopus dryophilus(Bull.) Murrill
soil detritus Not.
Lycoperdon mollePers.
soil detritus Not.
4 – Mixed deciduous forests
Amanita phalloides(Vaill. ex Fr.) Link
soil detritus under Quercus petraea agg. Not.
Chlorophyllum rhacodes(Vittad.) Vellinga
Not.
Clitocybe odora(Bull.) P. Kumm.
soil detritus Not.
Crucibulum laeve(Huds.) Kambly
wood of fallen branch, c. 2 cm in diam., deciduous tree, s. 4
SLO 2563
Cyathus striatus (Huds.) Willd.
pieces of wood, deciduous tree, s. 5 SLO 2564
Gymnopus fusipes(Bull.) Gray
wood at the base of standing tree and/or roots, c. 80 cm in diam., Quercus petraea agg., s. 1
Not.
Lactarius vellereus(Fr.) Fr.
soil detritus under Quercus petraea agg. Not.
Lepiota clypeolaria (Bull.) P. Kumm.
soil detritus SLO 2565
Lepiota lilaceaBres.
soil detritus SLO 2566
Macrolepiota procera(Scop.) Singer
soil detritus Not.
Megacollybia platyphylla(Pers.) Kotl. & Pouzar
pieces of wood burried in soil, deciduous tree, s. 5 Not.
21
A B C
Meripilus giganteus (Pers.) P. Karst.
base of standing tree, c. 90 cm in diam., Fagus sylvatica, s. 1
SLO 2567
Kuehneromyces mutabilis(Schaeff.) Singer & A. H. Sm.
wood of fallen trunk, c. 30 cm in diam., Fagus sylvatica, s. 3
Not.
Rickenella fibula(Bull.) Raithelh.
thallus of living Polytrichum formosum Not.
Schizophyllum communeFr.
bark of fallen branch, c. 10 cm in diam., deciduous tree, s. 3.
Not.
Scleroderma citrinumPers.
soil detritus under Quercus petraea agg., Robinia pseudoa-ccacia
Not.
Trametes versicolor(L.) Lloyd
wood and bark of fallen trunk, c. 40 cm in diam., Cerasus avium, s. 3
Not.
5 – Ruins of Biely Kameň Castle and surrounding forests
Amanita phalloides(Vaill. ex Fr.) Link
soil detritus under Carpinus betulus, Quercus petraea agg. Not.
Amanita rubescensPers.
soil detritus under Carpinus betulus, Quercus petraea agg. Not.
Bjerkandera adusta(Willd.) P. Karst.
wood on the cut surface of fallen trunk, c. 20 cm in diam., Fagus sylvatica, s. 3
Not.
Entoloma jahnii Wölfel & Winterh.
wood of fallen trunk, c. 20 cm in diam., deciduous tree, s. 4 SLO 2568
Cyathus olla(Batsch) Pers.
soil detritus Not.
Exidia glandulosa(Bull.) Fr.
bark of fallen branch, c. 10 cm in diam., Fagus sylvatica, s. 3
Not.
Geastrum triplex Jungh.
soil detritus (in thick layer of fallen decaying leaves) SLO 2569
Gymnopus brassicolens (Romagn.) Antonín & Noordel.
soil detritus SLO 2570
Gymnopus dryophilus(Bull.) Murrill
soil detritus Not.
Hymenopellis radicata(Relhan) R. H. Petersen
base of stump of Fagus sylvatica, c. 30 cm in diam., s. 3 Not.
Lycoperdon perlatumPers.
soil detritus Not.
Macrolepiota procera(Scop.) Singer
soil detritus Not.
Mycena galericulata(Scop.) Gray
pieces of wood near Acer campestre, Aesculus hippocasta-num, Euonymus europaeus, s. 5
Not.
Mycena roseaGramberg
soil detritus Not.
Mycena renatiQuél.
bark of fallen branch, c. 10 cm in diam., Carpinus betulus, s. 3
Not.
Phloeomana speirea (Fr.) Redhead
wood of fallen trunk, c. 20 cm in diam., deciduous tree, s. 4 SLO 2571
22
A B C
Picipes badius(Pers.) Zmitr. & Kovalenko
timber of stairs leading to the Castle ruins, deciduous tree, s. 2
Not.
Parasola conopilea(Fr.) Örstadius & E. Larss.
pieces of wood burried in soil, deciduous tree, s. 5 Not.
Psathyrella noli-tangere(Fr.) A. Pearson & Dennis
soil detritus SLO 2572
Radulomyces molaris(Chaillet ex Fr.) M. P. Christ.
bark of fallen trunk, c. 20 in cm diam., Quercus petraea agg., s. 3
Not.
Rickenella fibula(Bull.) Raithelh.
thalli of living Bryum capillare, Hypnum cupressiforme Not.
Russula pectinatoides Peck
soil detritus under Quercus sp. SLO 2573
Russula ochroleuca Fr.
soil detritus under Fagus sylvatica SLO 2574
Schizophyllum communeFr.
bark of fallen branch, c. 10 cm in diam., Carpinus betulus, s. 3
Not.
Simocybe sumptuosa.(P.D. Orton) Singer
wood of fallen trunk, c. 20 cm in diam., deciduous tree, s. 4 SLO 2575
Steccherinum bourdotii Saliba & A. David
bark of fallen branch, c. 1 cm in diam., deciduous tree, s. 3 SLO 2576
Stereum hirsutum(Willd.) Pers.
bark of fallen branch, c. 5 cm in diam., deciduous tree, s. 3 Not.
Trichaptum biforme(Fr.) Ryvarden
bark of fallen trunk, c. 30 cm in diam., Quercus petraea agg., s. 3
Not.
Xerocomellus chrysenteron (Bull.) Šutara
soil detritus under Fagus sylvatica, Carpinus betulus SLO 2577
Myxogastria:Ceratiomyxa fruticulosa(O. F. Müll.) T. Macbr.
wood of fallen trunk, c. 20 cm in diam., deciduous tree, s. 4 Not.
Myxogastria:Stemonitis sp.
wood of fallen trunk, c. 20 cm in diam., Fagus sylvatica, s. 4
Not.
Localities 4 and 5. These stands, rich in dead wood and soil organic matter, had the highest diversity of fungi. We were especially careful when digging out basidiomata of Gymnopus fusipes and Hymenopellis radicata from the soil. These both collybioids are typically deeply rooting; G. fusipes occurred at the base of Quercus tree and H. radicata around the Fagus sylvatica stump – they probably grew out of the tree roots. From a distance, we could spot a large annual polypore Meripilus giganteus at the base of standing Fagus sylvatica tree; it probably also grew out of the roots.
Amanita rubescens may in some way resemble Chlorophyllum rhacodes. Both agarics are similar in general appearance and stain red when bruised. These two unrelated species can be easily distinguished by several characters; we focused on the striate ring of A. rubescens that is clearly different from the muff-like and double ring of C. rhacodes. During our excursion, we also found a very poisonous Amanita phalloides.
They differ in size, which is also indicated by their names: Macrolepiota and Lepiota.

23
We found several times larger basidiomata of Macrolepiota procera than those of L. clypeolaria and L. lilacea. Lepiota clypeolaria is characteristic by the whitish woolly-tomentose girdles below the woolly ring zone on the stem and fusiform spores measuring 11–18 × 4.5–6 μm. Lepiota lilacea has a muff-like ring with a greyish-violet underside and ellipsoid spores of size 4–6.5 × 2.5–4 μm (Lange 2012).
The students could compare the spiny hymenophore of two species: Steccherinum bourdotii and Radulomyces molaris. Spines of S. bourdotii are apricot orange, cylindrical or slightly flattened, and up to 2.5 mm long; microscopically, it is characteristic by its thick-walled, crystal-covered cystidia and spores 3–4.5 μm wide. Spines of R. molaris are vinaceous-brown, coarsely flat, and 3–8 mm long; it lacks cystidia and spores are 6–8 μm wide (Læssøe, Petersen 2019).
During our excursion, we collected also fungi with angiocarpic basidiomata: Crucibulum laeve, Cyathus olla and Cyathus striatus. All three are so-called Bird’s nest fungi as producing basidia and spores within special egg-like structures called peridioles. While peridioles of Crucibulum laeve are creamy in colour, peridioles of Cyathus species are grey. Inner side of bell-shaped basidiomata of C. olla is smooth, but distinctly furrowed-striate of cup-shaped basidiomata of C. striatus (Læssøe, Petersen 2019, Holec et al. 2012).
During the excursion, we also tested our sense of smell which is an important diagnostic tool to help identify some species. We collected anise-smelling Clitocybe odora, radish-smelling Mycena rosea and Russula pectinatoides with a smell of fish. We convinced ourselves that Lepiota clypeolaria has an unpleasant, Scleroderma-like odour, because we managed to find both of these species.
At first glance, they looked like sporophytes of mosses, but on closer observation we recognised basidiomata of Rickenella fibula. They are subtle, but with their orange colouring contrast markedly with green mosses. They grew on the thali of Bryum capillare, Hypnum cupressiforme and Polytrichum formosum.
We did not overlook a delicate plasmodial slime mould Ceratiomyxa fruticulosa (O. F. Müll.) T. Macbr. (Myxogastria) whose white pilar sporangia covered wood of strongly decayed fallen trunk of deciduous tree. It shared the same substrate with two small fungal species: Entoloma jahnii and Phloeomana speirea (syn. Mycena speirea). Entoloma jahnii, a rare species in Europe (Nita, Stefaniak 2010), was first reported from Slovakia only in 2012 (Jančovičová, Adamčík 2012). Macroscopically, it is interchangeable with other Entoloma species with crepidotoid basidiomata, as well as with some species of the genera Crepidotus and Clitopilus. Microscopically, however, it is distinct by its angled spores and capitate pileocystidia and caulocystidia (Jančovičová, Adamčík 2012, Noordeloos 2012). Phloeomana speirea is usually easily determined by its habitat, the yellowish colours and decurrent gills (Aronsen, Læssøe 2016). Stemonitis sp. was another striking slime mould that we saw on the nearby rotten trunk of Fagus sylvatica; it is characterised by the cylindrical brown sporangia supported on slender, hair-like stalks growing in clusters.

24
acknowledgements
We would like to thank Mgr. Michal Hrabovský, PhD. for determinating some vascular plant taxa. The research was funded by the Operational Programme Research and Innovation for programme ″Advancing University Capacity and Competence in Research, Development and Innovation, ITMS2014+: 313021X329”.
references
Anonymus 1: http://naucnechodniky.eu/svatojursky-naucny-chodnik/ [accessed 17 February 2021]Anonymus 2: ISLHP (nlcsk.org) [accessed 18 February 2021]Anonymus 3: http://www.vino.sk/encyklopedia-vina/vinohradnicke-oblasti/slovensko-malokarpatska-
vinohradnicka-oblast/ [accessed 19 February 2021]Aronsen, A., Læssøe, T., 2016: The genus Mycena s.l. Fungi of Northern Europe, 5: 1-376.Atherton, I., Bosanquet, S., Lawley, M. (eds.), 2010: Mosses and liverworts of Britain and Ireland – a field
guide. British Bryological Society. Available at http://www.britishbryologicalsociety.org.uk Baumgartner, J., 1901: Bryologische Exkursionen in das Gebiet der Pressburger Karpathen. Verh. Vereins
Natur-Heilk. Pressburg, 13: 17-23.Eliáš, P. jun., Dítě, D., Kliment, J., Hrivnák, R., Feráková, V., 2015: Red list of ferns and flowering plants
of Slovakia, 5th edition (October 2014). Biologia, 70(2): 218-228.Eliáš, P., 2009: Opustené vinohrady: vznik, biodiverzita a význam. Životné Prostredie, Bratislava, 43(1): 24-
28. Godovičová, K., Mišíková, K., 2016: Rhynchostegium rotundifolium (Scop. ex Brid.) Schimp. (Bryophyta,
Bryopsida) na Slovensku. In: Galamboš, M., Džugasová, V., Ševčovičová, A., Bobovská, A., (eds.), p. 224-228, Študentská vedecká konferencia PriF UK konaná 26. apríla 2016.
Guttová, A., Lackovičová, A., Pišút, I., 2013: Revised and updated checklist of lichens of Slovakia (May 2013). Biologia, 68: 845-850.
Hill, M. O., Preston, C. D., Smith, A. J. E., 1994: Atlas of the bryophytes of Britain and Ireland. Volume 3. Mosses (Diplolepideae). Harley Books, Essex.
Holec, J., Bielich, A., Beran, M., 2012: Přehled hub střední Evropy. Academia, Praha.Jančovičová, S., Adamčík, S., 2012: Entoloma jahnii (Fungi, Agaricales) reported from Slovakia and notes on
differences with E. byssisedum. Czech Mycology, 64: 209-222.Jančovičová, S., Godovičová, K., Trojanovičová, L., Vašková, Z., Michalová, M., Dušička, J., Hrabovský,
M., Miškovic, J., Mišíková, K., 2019: Let’s go to the field. Botanical excursion 3. Acta Botanica Universitatis Comenianae, 54: 39-56.
Kaplan, Z., Danihelka, J., Chrtek, J. jun., Kirschner, J., Kubát, K., Štech, M., Štěpánek, J. (eds.), 2019: Klíč ke květeně České republiky [Key to the flora of the Czech Republic]. Ed. 2., Academia, Praha.
Kirk, P., 2021: Continuously updated: Index Fungorum. http://www.indexfungorum.org [accessed 18 February 2021].
Knudsen, H., Christian, L., Knutsson, T., 2012: Agaricus L.: Fr. In: Knudsen, H., Vesterholt, J. (eds.), Funga Nordica, p. 597-608, Nordsvamp, Copenhagen.
Læssøe, T., Petersen, J. H., 2019: Fungi of Temperate Europe. Princeton University Press, New Jersey, Oxfordshire.
Lange, C., 2012. Lepiota (Pers.: Fr.) Gray. In: Knudsen, H., Vesterholt, J. (eds.), Funga Nordica, p. 627-637, Nordsvamp, Copenhagen.
Májeková, J., Letz, D. R., Mereďa, P. jun., Zaliberová, M., Hodálová, I., 2020: Výsledky Floristického minikurzu konaného v roku 2019 v Bratislave, v mestských častiach Rača a Vajnory. Bull. Slov. Bot. Spoločn., 42(1): 75-92.
Marhold, K., Goliašová, K., Hegedüšová, K., Hodálová, I., Jurkovičová, V., Kmeťová, E., Letz, R., Michalková, E., Mráz, P., Peniašteková, M., Šípošová, H., Ťavoda, O., 1998: Papraďorasty a semenné rastliny. In: Marhold, K., Hindák, F. (eds.), Zoznam nižších a vyšších rastlín Slovenska, p. 333-687, Veda, Bratislava.

25
Medvecká, J., Kliment, J., Májeková, J., Halada, Ľ., Zaliberová, M., Gojdičová, E., Feráková, V., Jarolímek, I., 2012: Inventory of the alien flora of Slovakia. Preslia, 84: 257-309.
Mereďa, P. jun., 2009: Botanické a historické zaujímavosti Svätého Jura a okolia. Informačné materiály pre účastníkov Exkurzie na veľkomoravské hradisko pri Svätom Jure konanej 3. 9. 2009 pri príležitosti životných jubileí RNDr. Kamily Zahradníkovej, CSc. a RNDr. Magdalény Peniažtekovej, CSc. https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwiEur6Hm_DuAhUByIUKHYrxCaoQFjAAegQIAxAC&url=http%3A%2F%2Fsbs.sav.sk%2Fdocs%2Fexkurzia_nestich_info09.pdf&usg=AOvVaw3au47gZS4Ho4MOtMDbttdT [accessed 17 February 2021]
Mereďa, P. jun., 2012: Polycarpon Loefl. ex L. In: Goliašová, K., Michalková, E. (eds.), Flóra Slovenska. VI/3, p. 99-105, Veda, Bratislava.
Mišíková, K., Godovičová, K., Šoltés, R., Širka, P., 2020: Checklist and Red List of mosses (Bryophyta) of Slovakia. Biologia, 75(1):21-37.
Mišíková, K., Godovičová, K., Šoltés, R., Širka, P., 2021: Checklist and red list of hornworts (Anthocerotophyta) and liverworts (Marchantiophyta) of Slovakia. Biologia, https://doi.org/10.2478/s11756-020-00670-0
Mišíková, K., Orbánová, M., Godovičová, K., 2018: Bryophytes in cemeteries in the Small Carpathian region (Slovakia). Acta Botanica Universitatis Comenianae, 53: 45-53.
Nita, J., Stefaniak, M., 2010: Entoloma jahnii, a rare species in Europe. Acta Mycologica, 45: 157-162. Noordeloos, M., 2012: Entoloma (Fr.) P. Kumm. In: Knudsen, H., Vesterholt, J. (eds.), Funga Nordica, p. 517-
576, Nordsvamp, Copenhagen.Pfunder, M., Roy, B. A., 2000: Pollinator-mediated interacions between a pathogenic fungus, Uromyces pisi
(Pucciniales), and its host plant, Euphorbia cyparissias (Euphorbiaceae). Amercan Journal of Botany, 87(1): 48-55.
Pišút, I., 2002: Alexander Zahlbruckner (1860–1938): The Author of Two Historical Milestones in Lichenology. The Bryologist, 105(2): 243-245.
Ripková, S., Adamčík, S., Kučera, V., Palko, L., 2007: Fungi of the Protected Landscape Area of Vihorlat / Huby chránenej krajinnej oblasti Vihorlat. Botanický ústav SAV, Bratislava.
Smith, A. J. E., 2004: The moss flora of Britain and Ireland, Ed. 2. Cambridge University Press, Cambridge.Uhereková Šmelková, D., Mišíková, K., Kubinská, A., 2011: Bryophytes of medieval ruins in the Protected
Landscape Area Malé Karpaty Mts. Acta Botanica Universitatis Comenianae, 46: 35-39.
Abstrakt
Štvrtá botanická exkurzia v rámci cyklu „Poďme spolu do terénu″ [Lets go to the field] sa konala v Malých Karpatoch vo vinohradníckom meste Svätý Jur a jeho okolí. Trasa terénu viedla cez listnaté zmiešané lesy a biotopy kultúrnej krajiny, ako vinohrady, ruina hradu Biely Kameň, antropogénne a ruderálne stanovištia. Na piatich lokalitách sme sa zamerali na základný floristický výskum, ako aj monitoring výskytu a rozšírenia archeofytov, neofytov a inváznych druhov. Celkovo sme zaznamenali 63 druhov cievnatých rastlín, 41 machorastov (tri druhy pečeňoviek a 38 druhov machov), 12 lišajníkov a 44 taxónov makroskopických húb. Medzi hodnotné nálezy patrí potvrdenie výskytu kriticky ohrozeného machu Rhynchostegium rotundifolium na zrúcanine hradu Biely Kameň.
Kľúčové slová: Slovensko, Malé Karpaty, vinohrady, zrúcanina hradu, cievnaté rastliny, machorasty, lichenizované huby, nelichenizované makroskopické huby


27
Acta Botanica Universitatis Comenianae Vol. 57, 2021
BryoPHyte flora of tHe stuPava town and its surroundings (western slovakia) 2. BryoPHytes
on meadows in agricultural landscaPeBryoflóra stuPavy a jej okolia (záPadné slovensko)
2. macHorasty lúk v agrikultúrnej krajine
Katarína Mišíková1*, Miroslav Mišík2
1 Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 811 02 Bratislava, Slovakia
2 Medical University in Vienna, Cancer Research Institut, Borschkegasse 8a, AT-1090 Wien
Received 23 March 2021; Received in revised form 6 April 2021; Accepted 8 April 2021
Abstract
The paper provides a basic overview of the bryophyte diversity on two sites of mowed meadows in southwestern Slovakia in the cadastre of the town Stupava. Overall, 42 bryophyte species were found, out of them, three liverworts (Marchantia polymorpha, Radula complanata and Riccia sorocarpa) and 39 mosses. On both sites, terrestrial species and epilithic species growing on concrete and stone substrata predominate, while bryophytes on rotten wood being sparsely represented. On mowed meadows, the number of species increases with the higher number of substrata, e.g. bark of solitary trees, concrete and rocks. The occurrence of the endangered moss Rhynchostegium rotundifolium is associated with piles of granite stones in an alder forest patch. Based on Ellenberg´s indicator values, the ecological conditions are very similar on both sites. Colonists predominate in forest-free habitats, as bare soil, bark, and concrete are ideal substrata for bryophytes with this life strategy. The forests create a stable environment for perennial species, therefore this life strategy predominates in the alder stand.
Key words: Bryophyta, pleurocarpous, acrocarpous, land use, grasslands, biodiversity
introduction
The paper follows the work of Mišíková (2020), who first performed a bryofloristic survey of the town Stupava and its surroundings according to habitat types. The present article evaluated two sites of semi-dry meadows, first on the Záhorská nížina lowland, second in the Malé Karpaty Mts. These two locations are indicated on old maps (the end of the 19th and the first half of the 20th century) (Anonymus 1) as forest-free, probably already at that time used as pastures or meadows.
In Slovakia, semi-dry and dry meadows are bryologically insufficiently studied. Published data are almost exclusively a part of phytosociological studies, however, they are primarily focused mainly on vascular plants. The main purpose of the present study
* Corresponding author: [email protected]

28
is to provide an overview concerning species diversity and ecological requirements of bryophytes on mowed meadows in southwestern Slovakia.
methods
Field research took place in March, November and December 2020 and in January and February 2021. Species were collected from all substrata found. The nomenclature and threat categories of liverworts are based on the work of Mišíková et al. (2021), that of mosses on Mišíková et al. (2020). The names of vascular plants are according to Marhold et al. (1998). The phytogeographical division of Slovakia follows Futák (1980). The life strategies of bryophytes are based on the work of Dierßen (2001).
Ellenberg´s indicator values are listed according to Düll (1992), the median function was used in the processing of ecological factors. The N value is given according to Simmel et al. (2020), who considered it as indicators of the general nutrient status of the sites in which the species was found. The specimens of selected bryophytes are deposited in the herbarium SLO (the herbarium of the Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany).
Sites (Fig. 1).1. Záhorská nížina lowland, Stupava, site Malý Háj, meadows, c. 150 m a. s. l., area
size c. 23.8 ha, 48°17‘12.5“N 16°59‘16.1“E. The meadows are unfertilized, mowed once a year, the mown biomass is weighed. A water channel runs across the meadows, which was probably dug in the 1960s and 1970s (Anonymus 2, 3). Solitary trees and shrubs (Acer campestre, Robinia pseudoaccacia, Prunus cerasifera, P. spinosa) grow on the meadow margins, along the channel there is a narrow strip of trees (Populus sp., Robinia pseudoaccacia, Prunus cerasifera). In addition to mowing, the grass cover is intensively grazed and the soil is disturbed by numerous forest animals such as roe deers, wild boars and mouflons. A small part of the area (approx. 10x20 m) is plowed by regional hunters. The meadows neighbors with abandoned vineyards and forest.
Localities: 1a – mowed meadows, 1b – plowed part of meadow.2. Malé Karpaty Mts., Stupava, site Nová hora, c. 200-220 m a. s. l., area size
c. 33 ha, 48°17‘04“N 17°02‘30“E. The meadows are unfertilized, mowed once a year, the mown biomass is weighed. The meadows divide narrow hedgerows with trees and shrubs (Prunus domestica, Cerasus vulgaris, Robinia pseudoaccacia, Sambucus nigra, Crataegus sp.). On the western margin, an alder forest patch with periodic wetlands and a stream separates the mowed and unmowed part of the meadows, species as Alnus glutinosa, Robinia pseudoaccacia and Sambucus nigra grow here. The meadows neighbors with residential town parts, gardens, forests, paddocks, and pastures for horses.
Localities: 2a – mowed meadows, 2b – alder forest patch, 2c – non mowed meadow.

29
results and disccusion
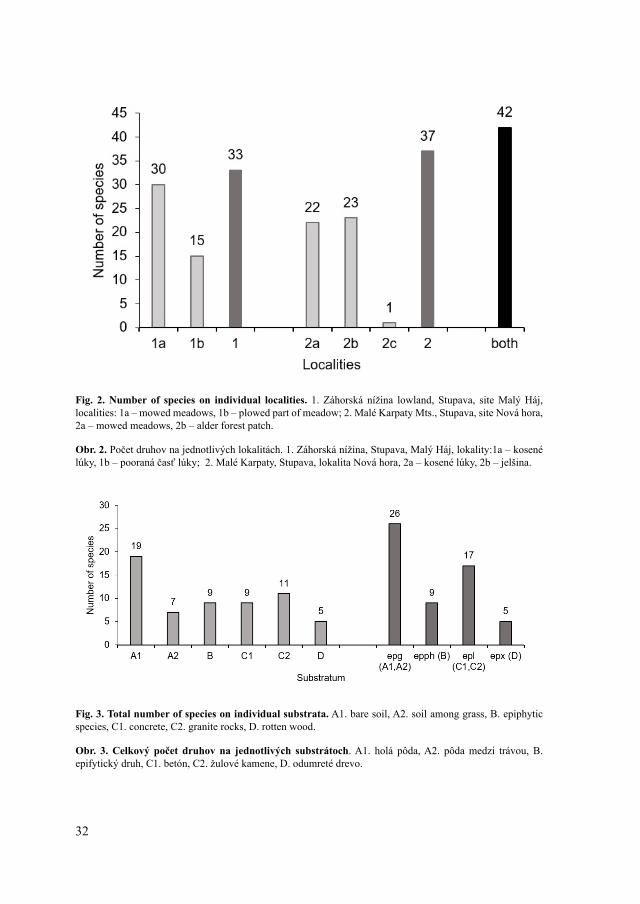
Species diversityOn both sites, overall 42 bryophytes were identified, out of them 39 mosses and three
liverworts (Marchantia polymorpha, Radula complanata and Riccia sorocarpa) (Tab. 1, Fig. 2). On the site Malý Háj, 33 bryophytes were found and 37 species on the site Nová hora. Within the individual localities, the lowest number of species was found on the non-mowed meadow (locality 2c), only the moss Brachythecium rutabulum grows there sporadically. This locality is overgrown with vascular plants such as Calamagrostis epigeios, Solidago gigantea and Tanacetum vulgare. The thick layer of plant biomass prevents the growth of competitively weaker bryophytes. Most bryophytes were found on the locality 1a (mowed meadow), where several different types of substratum (e.g. solitary trees, water channel banks, concrete) contribute to the species diversity.
The most common pleurocarpous mosses on the mowed meadows are Brachythecium albicans, B. rutabulum and Rhynchostegium megapolitanum, of the acrocarpous ones the species Ceratodon purpureus, Bryum argenteum and Osculatia caespiticia predominated. These species are typical for dry, sunny sites with well-drained sandy soil. Dry conditions are not suitable for many foliose liverworts, therefore they were not found on investigated localities. Marchantia polymorpha and Riccia sorocarpa were the only epigeic frondose liverworts found. They grow on the plowed soil (locality 1b) and are typical for agrocoenoses.
The moss Rhynchostegium megapolitanum was in the past evaluated in Slovakia as a rare species, however, in the last two decades, several new localities have been found, especially on dry and semi-dry meadows. It seems the climatic conditions of the recent warm years are suitable for this species. Zechmeister (2005) mentioned this species as
Fig. 1. Map of localities. 1. Záhorská nížina lowland, Stupava, site Malý Háj, localities: 1a – mowed meadows, 1b –plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, 2a – mowed meadows, 2b – alder forest patch, 2c – non mowed meadow (Source: www.mapy.hiking.sk, modified by K. Mišíková)
Obr. 1. Mapa lokalít. 1. Záhorská nížina, Stupava, lokalita Malý Háj, stanovištia: 1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, 2a – kosené lúky, 2b – jelšina. (Zdroj: www.mapy.hiking.sk, upravila K. Mišíková)
30
locally abundant in slightly saline, dry habitats in Austria.Rhynchostegium rotundifolium (EN), the only red-listed species was found in the alder
forest patch (locality 2b) in an extremely small population (population size c. 3x4 cm). The population was probably larger in the past, however, due to dry and warm years, this sciophytic and hygrophilous moss has declined. In Slovakia, this species has the center of its distribution in the Malé Karpaty Mts. (Godovičová, Mišíková 2016), it grows mainly on secondary sites (old stone walls, rubble).
On both meadow localities (1a, 2a), the fungus Bovistella utriformis (Bull.) Demoulin et Rebriev (Basidiomycota) occurs frequently.
Species diversity of dry meadow bryophytes is not high due to the strong competition of vascular plants (Jerram 2001, Zechmeister et al. 2002). Zechmeister et al. (2003a) observed four to fifty-eight species from moderately intensive meadows in the Austrian agricultural landscape outside the alpine and subalpine areas. From the dry meadows in xerothermic habitats, Zechmeister et al. (2002) reported as the most common species Barbula convoluta, Thuidium abietinum, Ceratodon purpureus, Homalothecium lutescens and Pseudoscleropodium purum. Gálusová et al. (2014) point out Homalothecium lutescens and Hypnum cupressiforme as the species with the greatest cover on the xerothermic grasslands in the limestone area of Slovenský Kras Mts.
Müller et al. (2012) found out the occurrence of 44 terrestrial species on 84 sites of managed meadows in Germany, the most common were pleurocarpous mosses Brachythecium rutabulum, Eurhynchium hians, Amblystegium serpens, and the acrocarpous Phascum cuspidatum, Barbula unguiculata and Osculatia rubens.
Tab. 1. The list of recorded bryophytes on meadows in Stupava. With asterisk (*) – Marchantiophyta; without any mark – Bryophyta; threat category. 1. Záhorská nížina lowland, Stupava, site Malý Háj, 1a – mowed meadows, 1b – plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, localities: 2a – mowed meadows, 2b – alder forest patch. Substratum: A1. bare soil, A2. soil among grass, B. epiphytic species (host tree), C1. concrete, C2. granite rocks, D. rotten wood. LS – life strategy: a – annual shuttle species, c – colonists, ce – ephemeral colonists, s – short lived shuttle species, l – long lived shuttle species, p – perennials.
Tab. 1. Zoznam zistených druhov machorastov na kosených lúkach v Stupave. So znakom (*) – Marchantiophyta; bez označenia – Bryophyta; kategória ohozenosti. 1. Záhorská nížina, Stupava, lokalita Malý Háj, 1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, stanovištia: 2a – kosené lúky, 2b – jelšina. Substrát: A1. holá pôda, A2. pôda medzi trávou, B. epifytický druh (hostiteľská drevina), C1. betón, C2. žulové kamene, D. odumreté drevo. LS – životná stratégia: a – jednoročný druh, c – kolonisti, ce – efemérni kolonisti, s – druhy „short lived shuttle species″, l – druhy„long lived shuttle species″, p – trváce druhy.
Species 1 2 Substratum LS
1a 1b 2a 2b
*Marchantia polymorpha + A1 c
*Radula complanata + C2 l
*Riccia sorocarpa + A1 a
Amblystegium serpens + + + B, C1, C2, D p
Atrichum undulatum + A1 s
Barbula convoluta + + + A1 c
Barbula unguiculata + + A1 c
31
Species 1 2 Substratum LS
1a 1b 2a 2b
Brachytheciastrum velutinum + A1, C2 p
Brachythecium albicans + + A2 p
Brachythecium populeum + C2 p
Brachythecium rutabulum + + + + A2, D p
Brachythecium salebrosum + A2, C2 p
Bryum argenteum + + + A1, C1 c
Bryum capillare + + A1, C2 c
Bryum moravicum + + B, D, Robinia pseudoacacia c
Ceratodon purpureus + + + A1, C1, D c
Dicranella staphylina + + A1 ce
Grimmia pulvinata + + C1 c
Hypnum cupressiforme + + + A1, B, C1, C2, D p
Leskea polycarpa + + + B, Robinia pseudoacacia p
Lewinskya affinis + + + B, Robinia pseudoacacia c
Orthotrichum anomalum + + C1, C2 c
Orthotrichum cupulatum + C2 c
Orthotrichum diaphanum + + B, C1, Robinia pseudoacacia c
Orthotrichum pumilum + + + B, Robinia pseudoacacia c
Osculatia bicolor + + + A1 c
Osculatia caespiticia + + + A1 c
Osculatia rubens + + + A1 c
Oxyrrhynchium hians + + + A1, A2 p
Plagiomnium cuspidatum + + A2, C2 p
Platygyrium repens + + + B, Robinia pseudoacacia p
Pseudosleropodium purum + A2 p
Pylaisia polyantha + + B, Robinia pseudoacacia p
Rhynchostegium megapolitanum + + A2 p
Rhynchostegium rotundifolium (EN) + C2 p
Rhytidiadelphus squarrosus + A2 p
Schistidium apocarpum + + C1 c
Syntrichia ruralis + + A1 c
Tortula acaulon + + + A1 a
Tortula caucasica + + A1 ce
Tortula muralis + + + C1 c
Tortula truncata + + A1 a
32
Fig. 2. Number of species on individual localities. 1. Záhorská nížina lowland, Stupava, site Malý Háj, localities: 1a – mowed meadows, 1b – plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, 2a – mowed meadows, 2b – alder forest patch.
Obr. 2. Počet druhov na jednotlivých lokalitách. 1. Záhorská nížina, Stupava, Malý Háj, lokality:1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, 2a – kosené lúky, 2b – jelšina.
Fig. 3. Total number of species on individual substrata. A1. bare soil, A2. soil among grass, B. epiphytic species, C1. concrete, C2. granite rocks, D. rotten wood.
Obr. 3. Celkový počet druhov na jednotlivých substrátoch. A1. holá pôda, A2. pôda medzi trávou, B. epifytický druh, C1. betón, C2. žulové kamene, D. odumreté drevo.
33
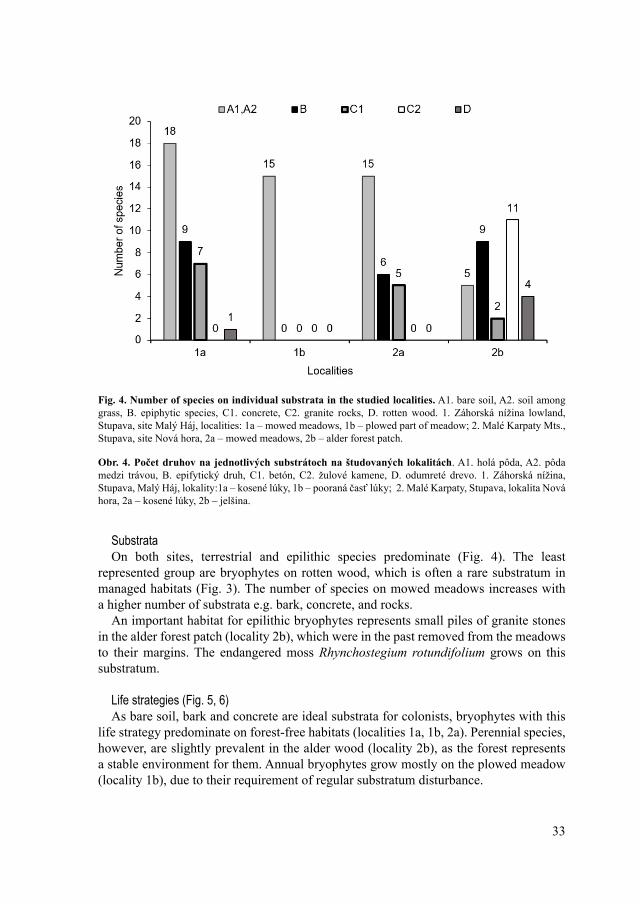
SubstrataOn both sites, terrestrial and epilithic species predominate (Fig. 4). The least
represented group are bryophytes on rotten wood, which is often a rare substratum in managed habitats (Fig. 3). The number of species on mowed meadows increases with a higher number of substrata e.g. bark, concrete, and rocks.
An important habitat for epilithic bryophytes represents small piles of granite stones in the alder forest patch (locality 2b), which were in the past removed from the meadows to their margins. The endangered moss Rhynchostegium rotundifolium grows on this substratum.
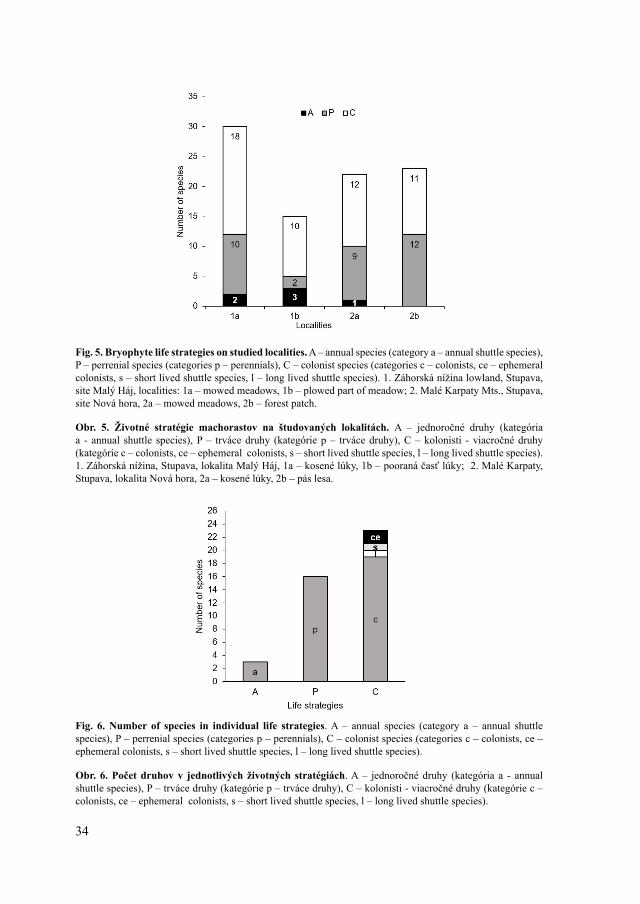
Life strategies (Fig. 5, 6)As bare soil, bark and concrete are ideal substrata for colonists, bryophytes with this
life strategy predominate on forest-free habitats (localities 1a, 1b, 2a). Perennial species, however, are slightly prevalent in the alder wood (locality 2b), as the forest represents a stable environment for them. Annual bryophytes grow mostly on the plowed meadow (locality 1b), due to their requirement of regular substratum disturbance.
Fig. 4. Number of species on individual substrata in the studied localities. A1. bare soil, A2. soil among grass, B. epiphytic species, C1. concrete, C2. granite rocks, D. rotten wood. 1. Záhorská nížina lowland, Stupava, site Malý Háj, localities: 1a – mowed meadows, 1b – plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, 2a – mowed meadows, 2b – alder forest patch.
Obr. 4. Počet druhov na jednotlivých substrátoch na študovaných lokalitách. A1. holá pôda, A2. pôda medzi trávou, B. epifytický druh, C1. betón, C2. žulové kamene, D. odumreté drevo. 1. Záhorská nížina, Stupava, Malý Háj, lokality:1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, 2a – kosené lúky, 2b – jelšina.
34
Fig. 5. Bryophyte life strategies on studied localities. A – annual species (category a – annual shuttle species), P – perrenial species (categories p – perennials), C – colonist species (categories c – colonists, ce – ephemeral colonists, s – short lived shuttle species, l – long lived shuttle species). 1. Záhorská nížina lowland, Stupava, site Malý Háj, localities: 1a – mowed meadows, 1b – plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, 2a – mowed meadows, 2b – forest patch.
Obr. 5. Životné stratégie machorastov na študovaných lokalitách. A – jednoročné druhy (kategória a - annual shuttle species), P – trváce druhy (kategórie p – trváce druhy), C – kolonisti - viacročné druhy (kategórie c – colonists, ce – ephemeral colonists, s – short lived shuttle species, l – long lived shuttle species). 1. Záhorská nížina, Stupava, lokalita Malý Háj, 1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, 2a – kosené lúky, 2b – pás lesa.
Fig. 6. Number of species in individual life strategies. A – annual species (category a – annual shuttle species), P – perrenial species (categories p – perennials), C – colonist species (categories c – colonists, ce – ephemeral colonists, s – short lived shuttle species, l – long lived shuttle species).
Obr. 6. Počet druhov v jednotlivých životných stratégiách. A – jednoročné druhy (kategória a - annual shuttle species), P – trváce druhy (kategórie p – trváce druhy), C – kolonisti - viacročné druhy (kategórie c – colonists, ce – ephemeral colonists, s – short lived shuttle species, l – long lived shuttle species).
35
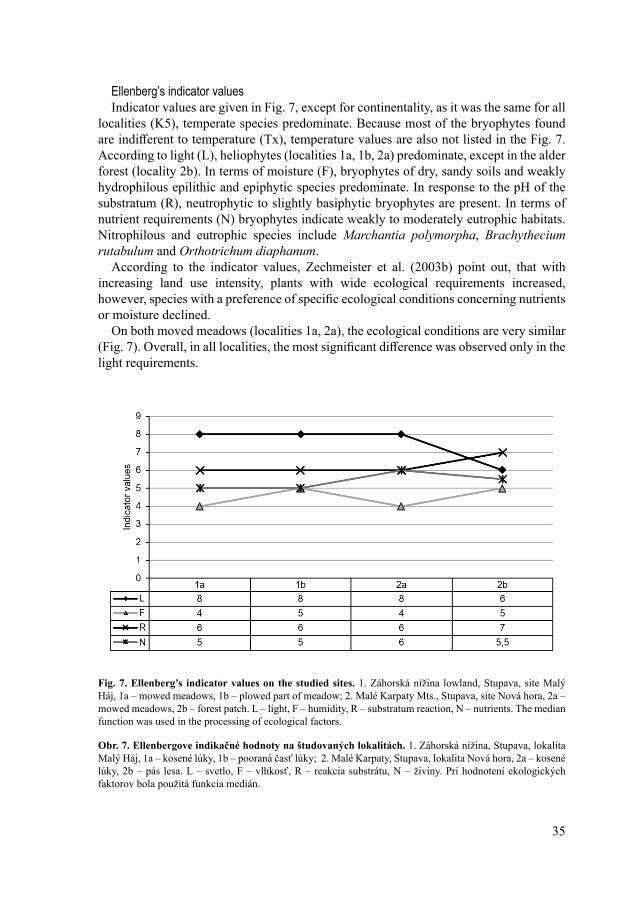
Ellenberg’s indicator valuesIndicator values are given in Fig. 7, except for continentality, as it was the same for all
localities (K5), temperate species predominate. Because most of the bryophytes found are indifferent to temperature (Tx), temperature values are also not listed in the Fig. 7. According to light (L), heliophytes (localities 1a, 1b, 2a) predominate, except in the alder forest (locality 2b). In terms of moisture (F), bryophytes of dry, sandy soils and weakly hydrophilous epilithic and epiphytic species predominate. In response to the pH of the substratum (R), neutrophytic to slightly basiphytic bryophytes are present. In terms of nutrient requirements (N) bryophytes indicate weakly to moderately eutrophic habitats. Nitrophilous and eutrophic species include Marchantia polymorpha, Brachythecium rutabulum and Orthotrichum diaphanum.
According to the indicator values, Zechmeister et al. (2003b) point out, that with increasing land use intensity, plants with wide ecological requirements increased, however, species with a preference of specific ecological conditions concerning nutrients or moisture declined.
On both moved meadows (localities 1a, 2a), the ecological conditions are very similar (Fig. 7). Overall, in all localities, the most significant difference was observed only in the light requirements.
Fig. 7. Ellenberg’s indicator values on the studied sites. 1. Záhorská nížina lowland, Stupava, site Malý Háj, 1a – mowed meadows, 1b – plowed part of meadow; 2. Malé Karpaty Mts., Stupava, site Nová hora, 2a – mowed meadows, 2b – forest patch. L – light, F – humidity, R – substratum reaction, N – nutrients. The median function was used in the processing of ecological factors.
Obr. 7. Ellenbergove indikačné hodnoty na študovaných lokalitách. 1. Záhorská nížina, Stupava, lokalita Malý Háj, 1a – kosené lúky, 1b – pooraná časť lúky; 2. Malé Karpaty, Stupava, lokalita Nová hora, 2a – kosené lúky, 2b – pás lesa. L – svetlo, F – vlhkosť, R – reakcia substrátu, N – živiny. Pri hodnotení ekologických faktorov bola použitá funkcia medián.

36
Several authors focused on the function, significance, diversity and threat of bryophytes on mowed meadows. Zechmeister et al. (2003b) evaluated the impact of land use parameters, such as the number of mowings and fertilizer input, and found out a constant decrease in bryophyte species richness, with the increasing quantity of fertilizer usage.
Klaus, Müller (2014) reported that bryophytes significantly contribute to grassland diversity and fundamental ecosystem functions. The bryophyte layer increases water retention capacity and interacts with vascular plants by germination. While soil disturbance by grazing and mowing can increase bryophyte diversity, high input of fertilizers and intensive land use leads to a reduction of bryophyte diversity.
The studied meadows in Stupava are not fertilized for at least four years and the mowing intensity is low, due to this, a relatively high number of bryophyte species grow there. In addition to their agricultural function, both meadows are also important habitats and refuges for many organisms in the agricultural landscape, as well as for recreation, as they are frequent visited by the inhabitants of Stupava. In the cultural landscape of Central Europe, moderately managed grasslands contribute to the stable species richness and diversity of different groups of organisms.
acknowledgements
We would like to thank Anna Mišíková for accompanying in the field. The research was funded by the Operational Programme Research and Innovation for programme “Advancing University Capacity and Competence in Research, Development and Innovation, ITMS2014+: 313021X329”.
references
Anonymus 1: StareMapy.sk, https://www.staremapy.sk/ [accessed Mar 10 2021]Anonymus 2: Kronika obce Stupava, rok 1980, https://www.stupava.sk/modules/file_storage/download.
php?file=d5f4a825%7C112 [accessed Sep 07 2020]Anonymus 3: Kronika obce Stupava, rok 1981, https://www.stupava.sk/modules/file_storage/download.
php?file=07de765e%7C113 [accessed Sep 07 2020]Dierßen, K., 2001: Distribution, Ecological Amplitude and Phytosociological Characterization of European
Bryophytes. Bryophytorum Bibliotheca, 56: 1-289.Düll, R., 1992: Zeigerwerte von Laub- und Lebermoosen. In: Ellenberg, H., Weber, H. E., Düll, R., Wirth, V.,
Werner, W., Paulissen D., Zeigerwerte von Pflanzen in Mitteleuropa, Scripta Geobotanica, 18: 175-214.Futák, J., 1980: Fytogeografické členenie. In: Mazúr, E., (ed.), Atlas Slovenskej socialistickej republiky,
Bratislava.Gálusová, T., Pružinová, L., Petrášová, A., Šuvada, R., Kuna, R., 2014: Xerothermic bryoflora in selected
areas of Slovak Karst National Park. Thaiszia – Journal of Botany, Košice, 24(1): 23-34.Godovičová, K., Mišíková, K., 2016: Rhynchostegium rotundifolium (Scop. ex Brid.) Schimp. (Bryophyta,
Bryopsida) na Slovensku. In: Galamboš, M., Džugasová, V., Ševčovičová, A., Bobovská, A., (eds.), p. 224-228, Študentská vedecká konferencia PriF UK konaná 26. apríla 2016.
Jerram, R., 2001: The upland management handbook. Chapter 7: Meadows and enclosed pasture. Aavailable at: http://publications.naturalengland.org.uk/file/86044 [accessed Feb 28 2021].
Klaus, V. H., Müller, J., 2014: The role of bryophytes in Central European grasslands. In: Mariotte, P., Kardol, P., (eds.), Grassland Biodiversity and Conservation in a Changing World, Nova Science Publishers.

37
Marhold, K., Goliašová, K., Hegedüšová, K., Hodálová, I., Jurkovičová, V., Kmeťová, E., Letz, R., Michalková, E., Mráz, P., Peniašteková, M., Šípošová, H., Ťavoda, O., 1998: Papraďorasty a semenné rastliny. In: Marhold, K., Hindák, F., (eds.), Zoznam nižších a vyšších rastlín Slovenska, Veda, Bratislava.
Mišíková, K., 2020. Bryoflóra Stupavy a jej okolia (západné Slovensko) 1. Machorasty v opustených vinohradoch a sadoch. Acta Botanica 56: 13-20.
Mišíková, K., Godovičová, K., Šoltés, R., Širka, P., 2020: Checklist and Red List of mosses (Bryophyta) of Slovakia. Biologia, 75(1): 21-37.
Mišíková, K., Godovičová, K., Šoltés, R., Širka, P., 2021: Checklist and red list of hornworts (Anthocerotophyta) and liverworts (Marchantiophyta) of Slovakia. Biologia, https://doi.org/10.2478/s11756-020-00670-0 [accessed Feb 27 2021].
Müller, J., Klaus, V. H., Kleinebecker, T., Prati, D., Hölzel, N., Fischer, M., 2012: Impact of land-use intensity and productivity on bryophyte diversity in agricultural grasslands. PLoS ONE 7(12): e51520. doi:10.1371/journal.pone.0051520 [accessed Feb 28 2021].
Simmel, J., Ahrens, M., Poschlod, P., 2020: Ellenberg N values of bryophytes in Central Europe. Journal of Vegetation Science, https://doi.org/10.1111/jvs.12957 [accessed Feb 28 2021].
Zechmeister, H. G., 2005. Bryophytes of continental salt meadows in Austria. Journal of Bryology, 27: 297-302.
Zechmeister, H. G., Tribsch, A., Hohenwallner, D., 2002: Die Moosflora von Linz und ihre Bedeutung für die Bioindikation. In: Schwarz F., Leister G. (eds), Naturkundliches Jahrbuch der Stadt Linz. Magistrat des Landeshauptstadt Linz, p. 111-191, http://www.linz.at/umwelt/4031.asp [accessed Feb 27 2021].
Zechmeister, H. G., Tribsch, A., Moser, D., Peterseil, J., Wrbka, T., 2003a: Biodiversity ′hot-spots’ for bryophytes in landscapes dominated by agriculture in Austria. Agriculture, Ecosystems and Environment, 94: 159-167.
Zechmeister, H.G., Schmitzberger, I., Steurer, B., Peterseil, J., Wrbka, T., 2003b: The influence of land-use practices and economics on plant species richness in meadows. Biological Conservation, 114: 165-177.
Abstrakt
Príspevok prináša základný prehľad druhovej diverzity machorastov dvoch lokalít kosených lúk na juhozápade Slovenska v katastri mesta Stupava. Celkovo bolo zistených 42 druhov machorastov, z toho tri pečeňovky (Marchantia polymorpha, Radula complanata, Riccia sorocarpa) a 39 machov. Na oboch lokalitách prevládajú terestrické druhy a epilitické druhy rastúce na betóne a kamenných substrátoch, najmenej sú zastúpené machorasty na odumretom dreve. Na kosených lúkach druhovú diverzitu pozitívne ovplyvňuje vyšší počet substrátov, ako borka drevín, betón a kamene. Významný biotop pre epilitické machorasty vytvárajú v lesnom poraste nízke kopy žulových kameňov. Na tieto plochy je viazaný výskyt ohrozeného machu Rhynchostegium rotundifolium. Vychádzajúc z Ellenbergových indikačných hodnôť sú ekologické podmienky na oboch lokalitách veľmi podobné. Z hľadiska životných stratégií na bezlesých stanovištiach prevládajú kolonisti, nakoľko holá pôda, kôra drevín a betón sú ideálne substráty pre tieto machorasty. Trváce druhy mierne prevládajú v lesnom poraste, pretože lesný porast tvorí pre viacročné machorasty stabilné životné prostredie.
Kľúčové slová: Bryophyta, pleurokarpný, akrokarpný, využitie pôdy, lúčne porasty, biodiverzita


39
Acta Botanica Universitatis Comenianae Vol. 57, 2021
leaf evolution and classification 3. gymnosPermoPsidamorfogenéza a klasifikácia listov 3. naHosemenné rastliny
Michal Hrabovský*
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 81102 Bratislava, Slovakia
Received 31 March 2021; Received in revised form 24 May 2020; Accepted 25 May 2020
Abstract
The previous theory about the origin of microphyllous and megaphyllous leaves can also be applied to seed plants. The existence of a microphyll of archaeophyll type offers new perspectives on the ovule origin. There are various phylogenetic classifications of gymnosperms. This paper brings new morphological insights on these classifications and recognizes three morphological groups of gymnosperms – primary cupulate and primary acupulate megaphyllous lineages and strobilate microphyllous lineage. Evolution from these three groups could continue in various directions, therefore there are many morphological similarities among them leading to many phylogenetic interpretations. The taxonomic groups based on the antophyte theory would be polyphyletic according to new morphological interpretations because all types of gymnosperm strobili could lead to formation of flowers.
Key words: ovule origin, conifers, pteridosperms
introduction
From the evolutionary perspective, gymnosperms are a very interesting plant group to which many researchers paid attention. All current knowledge about fossil ancestors is summarized in several monographs (Němejc 1968; Meyen 1987; Stewart, Rothwell 1993; Anderson, Anderson 2003; Anderson et al. 2007; Taylor et al. 2009), so I will focus on the ambiguous aspects which could be explained by my new theory of the possible existence of an archaeophyll.
A widely accepted view of evolution sees the origin of seed plants in the progymnosperms (Beck 1960; Stewart, Rothwell 1993; Taylor et al. 2009). Therefore, I will also proceed from this assumption and from the type of leaves in progymnosperms, regardless of their placement in the phylogram (Hrabovský 2020, Fig. 1). Based on this placement, the possible origin of seed plants is from lycophytes, monilophytes or their hybridisation. The origin of the ovule was described by three possible theories – axial, foliolar and sui generis (Worsdell 1904). Later, the theory of “units” proposed by De Haan (1920) and telomic theory (Zimmermann 1930) corresponding with the axial theory of Worsdell (1904) have become widely accepted (Herr 1995). I redescribed the
* Corresponding author: [email protected]

40
axes of progymnosperms as branches bearing archaeophylls or appendages formed by archaeophylls and not by telomes (Hrabovský 2020), so in this paper I will return to the foliar theory of the ovule origin, because one archaeophyll seems to be homologous to a single ovule as is evident from a comparison with sporophylls of progymnosperms.
Before the arising of many phylogenetic systems based on the anthophyte hypothesis (Cronquist 1960, Doyle at al.1986, Donoghue et al. 2000, Rothwell et al. 2009), there were several traditional classifications of gymnosperms. Meyen (1984) divided all known gymnosperms into three classes Ginkgoopsida, Cycadopsida and Pinopsida based especially on ovule anatomy and morphology. But this system was criticized by Miller (1985), stating a lack of knowledge of homologies among fossil plants. Anderson et al. (2007) extended the number of classes by exclusion of Lyginopteridopsida, Ottokariopsida, Bennettitopsida and Gnetopsida. This paper will revaluate homologies among orders classified in these excluded classes and the three Meyen’s classes Ginkgoopsida, Cycadopsida and Pinopsida, which will be in the next text described as subclasses Ginkgoidae, Cycadidae and Coniferidae (syn. Pinidae).
leaf and ovule homology
The main criteria of the homology are similarity, conjunction and congruence (Patterson 1982). These criteria combined with the relationships between the leaf types demonstrated in previous two parts of this series (Hrabovský 2020) reveal the leaf evolution in the gymnosperms (Fig. 1). The ancestral type of leaf in the gymnosperms is an appendage (syn. meiophyll), which could reduce or metamorphose to a pteridophyll and an euphyll (a pteridophyll with the reticulate venation)1. The evolutionary circle shown in the Fig. 1 demonstrates two possible ways of the appendage reduction. It is this reduction that can cause a misinterpretation of leaf origin and lead to a wrong phylogenetic conclusion. The reduced leaves of Dicranophyllales (Fig. 5.5–7) can be interpreted as axial because of fertile leaves having several lateral ovules. The leaves of conifers are interpreted in the same way (Benson 1921) and the result of this is that the conifers are considered as related to Dicranophyllales (Anderson et al. 2007). But I
1 I followed the traditional simplified classification of leaves for a better understanding. This classification discerns microphyllous and megaphyllous leaves. Megaphyllous leaves can be primitive without leaf blade (appendages) or pteridophylls and euphylls with leaf blade. But there is a much more natural alternative classification partially corresponding with the Benson’s leaf systems (1921), only the terms can be taken from Klotz (1985). The progress of foliation is taken into account in this natural classification. I use the term foliation for the leaf formation process in this article. The definition of microphylls remains unchanged, but the megaphyllous leaves are divided into macrophylls and megaphylls. The macrophylls can be defined as foliated terminal axes with or without microphylls, while the megaphylls in the narrow sense are whole foliated fronds bearing macrophylls, So, the appendages are macrophylls and most of pteridophylls and euphylls are megaphylls. But some appendages with leaf blade (e.g. Sphenophyllum) or secondary simplified megaphylls (e.g. Ginkgo) previously classified as pteridophylls are also macrophylls. Also some euphylls of appendage origin (e.g. Glossopteris) should be reclassified as macrophylls. Then, if microphylls can be divided into lycophylls and archaeophylls and megaphylls can be divided into pteridophylls and euphylls, the macrophylls (appendages) could be divided into meiophylls (macrophylls without leaf blade) and sphenophylls (macrophylls with leaf blade). The classification I used in the article is almost the same, only does not use the term sphenophylls for primitive pteridophylls without compound structure.

41
disagree with this opinion, because leaves of Cordaites, Voltziales or Agathis could be interpreted as archaeophylls (branched microphylls) and the needles of recent conifers as lycophylls (unbranched microphylls). One such leaf is homologous with one terminal ovule, which is the main difference from Dicranophyllum sporophyll with lateral ovules. However, this does not contradict the theory of megaphyllous origin of needles (Benson 1921), only the final product of the reduction is not the appendage axis but the terminal microphyll from the original appendage. The megaphyllous origin of the needle could be supported by the seed stalks (megasporophylls) of Cordaitanthus (Procordaiconus)
Fig. 1. Sterile leaves in Gymnospermopsida. The evolutionary cycle of megaphyll (appendage) transforming into single microphylls (grey) or pteridophyll, euphyll or losing the microphylls. Author: M. Hrabovský.

42
pseudofluitans (Fig. 11.1), which bear more than one ovule (Ignatiev, Meyen 1989). The origin of ovule was demonstrated from telomes (Taylor et al. 2009) and this theory
would be quite acceptable if the telomes were substituted by segments of dissected archaeophylls found in psilophytes and progymnosperms (Fig. 2). The comparison of sterile pinnules of lyginopteroid pteridosperms with their ovulate cupules and sporangia suggests that one segment of pinnule corresponding with one archaeophyll is homologous to one ovule. The whole original ovule consists from two parts – the sterile segments and the ovule itself. The sterile part in various groups of progymnosperms can form the cupule (Ginkgoidae), part of leaf blade (Cycadidae), seed scale (Coniferidae) or be absent in more advanced members of all previous groups (Ginkgo, Cycas, Ephedra). The ovulate part is formed by one sporangium and a few (most often two) basally or fully fused segments forming the integument. As Herr (1995) proposed, the sporangium is embedded into tissue of sporangiophore (analogy with floral hypanthium), what explains the origin of the lagenostome and the pollination chamber. The male archaeophyll bears several terminal sporangia which tend to form a synangium. So, in most cases one synangium is homologous to one ovule. But synangia of medullosalean pteridosperms are most likely of whole pinnule origin (Fig. 8.5).
Fig. 2. Male sporangia and ovule evolution in gymnosperms. 1.–2. Formation of male synangia from the archaeophyll (Ginkgoidae, Coniferidae). 3.–4. Formation of the sori with leaf blade equivalent to one archaeophyll (Cycadidae). 5.–8. Formation of the ovule through nesting of megasporangium into the sporangiophore and creation of the integument from segments of the archaeophyll. One half of the archaeophyll is sterile. 9.–10. The sterile segments of archaeophyll form a cupule or scale under the ovule or disappears (Ginkgoidae, Coniferidae). 11.–12. The sterile segments of archaeophyll form a leaf blade or absent (Cycadidae). Author: M. Hrabovský.

43
leaf evolution in ginkgoid (cupulate) pteridosperms (Ginkgoidae Wettst.2, Fig. 3–7)
It is difficult to determine the morphological boundary between ginkgoid and cycadoid pteridosperms like the boundary between magnoliids and eudicots. However, we should talk about evolutionary trends in morphology rather than strict boundaries. The strict boundaries led to morphological systems which were not widely accepted. Meyen (1984) included gymnosperms with bicornute ovules among the ginkgoid pteridosperms. Thus, orders such as Calamopityales and Callistophytales were included there. Anderson et al. (2007) excluded glossopterids and two orders mentioned above from ginkgopsids. Based on leaf evolutionary trend of simplification of bipartite compound leaves it is obvious that ginkgoaleans (Fig. 7) as well as glossopterids (Fig. 5.11–15) could have evolved from a common ancestor with dicranophyllaleans (Fig. 5.5–7) (Meyen 1987, Naugolnykh 2007). Dicranophyllaleans, Dichophyllum (Fig. 5.1–2) and peltasperms (Fig. 6) could be traceable to the order Calamopityales based on foliage, and calamopityaleans (Fig. 4) to the order Lyginopteridales (Fig. 3) (Anderson et al. 2007). Lyginopteridaleans were classified in Meyen’s system (1984, 1987) among cycadoid pteridosperms due to radiospermic ovules. The cycases and their relatives evolved most likely form medullosaleans (Anderson et al. 2007) which are morphologically related to Callistophytales. However, the morphology of these two orders and other cycadophytes differs from cupulate Lyginopteridales by the missing cupules. It leads me to conclusion that Devonian cupulate pteridosperms led to ginkgoid radiation, while acupulate pteridosperms evolved into cycadoid forms. However, the presence or the absence of the cupule cannot be the determining characteristic between current ginkgopsids and cycadopsids, since simplification led to a secondary loss of cupule in ginkgopsids. The terminology for cupules was reviewed by Doweld (1998), but it cannot always be accepted in the view of my theory.
The order Lyginopteridales (Fig. 3) includes traditionally cupulate and acupulate families (Anderson et al. 2007). But recent findings of Genomosperma (Fig. 3.1) cupules suggest, that all lyginopteridaleans including Physostoma (Fig. 3.2) could be cupulate (Hetherington 2020). Sometimes cupules of genera as Lagenospermum (Fig. 3.3) or Eonotosperma (Fig. 3.4) are interpreted as integuments (Césari 2014), but this could
2 The author citations used in series of publications (Doweld 2001; Chase, Reveal 2009; Christenhusz et al. 2011) have to be revised. The authorship of subclass Ginkgoidae is attributed to H.G.A. Engler (Engl. in H.G.A. Engler et K.A.E. Prantl, Nat. Planzenfam. Nacht.: 341, 1897), but this is not true. Engler (1897) used the same suffix -ales for both class and order, so he called this class as Ginkgoales. The name Ginkgoales is attributed to I.N. Gorozhankin (Gorozh., Lekts. Morf. Sist. Archegon.: 73 1904). In the fact, the name Ginkgoinae was published for the first time by Wettstein (1903, p. 132). The suffix -inae was the regular way of naming a class or a subclass at that time. This was spontaneously changed to the endings -psida or -idae around 1950 and recommended by International Code in 1961 (Cronquist 1960, Zabnikova 1964). Cronquist et al. (1966) did not accept the ending -psida for the class and used -atae to create Ginkgoatae. The name Ginkgoinae was replaced by Ginkgoidae around 1966–1971 by an unknown author. On this occasion, I would like to correct the author citations stated in Lycopodiopsida and Polypodiopsida, which I have uncritically taken over from the peer-reviewed publications as well. The name Lycopodiidae (Bek., Kurs Bot. 1: 115. 1863.) is incorrect. The name Lycopodinae was used for the first time by Link (1824, p. 355). He is also the author of the name Equisetinae. The names Ophioglossidae, Marattidae and Polypodinae were published by Burnett (1835). Wiggers (1840) corrected the previous two names to Ophioglossinae and Marattinae. Marattidae and Polypodinae were changed to Marattiidae and Polypodiidae by Novák (1954, p. 37), not by Cronquist et al. (1966). Finally, the authors of the name Archaeopterididae (in the original as Archaeopteridiidae) are Cronquist et al. (1966).

44
Fig. 3. The leaf evolution in the subclass Ginkgoidae – Lyginopteridales. 1. Genomosperma, an ovule and its longitudinal section; 2. Physostoma, a longitudinal section of ovule; 3. Lagenospermum, a pinna with cupulate ovules homologous to Palmatopteris pinna (8); 4. Eonotosperma, two cupulate ovules and pinnules from different parts of the same plant, homologous to one appendage; 5. Elkinsia, a cupule and its sterile equivalent – the pinnule homologous to one appendage; 6. Telangiopsis, a sterile pinnule and its equivalent with synangia; 7. Telangium, a segment from the pinnule with synangia homologous to one archaeophyll; 8. Palmatopteris, pinna homologous to a branch with appendages; 9. Eosphenopteris, pinnae, E. obtusiloba left, E. triginoloba right; 10. Mariopteris, a metamorphosed pinna of Palmatopteris type; 11. Lyginopteris, a pinna; 12. Cosmosperma, pinnule with archaeophylls and its female and male parallels; 13. Lagenostoma, a longitudinal section of ovule; 14. Moresnetia, a female pinnule forming the cupule; 15. Calathospermum, a cupule homologous to Elkinsia and Moresnetia cupule; 16. Parkvillia; a pinna (?) with synangia. Modified after Anderson et al. (2007) (1–2,7–11, 13–15); Arnold (1939) (3); Césari (2014) (4); Rothwell et al. (1989) (5); Wang et al. (2016) (6); Wang et al. (2014) and Liu et al. (2017) (12); Serbet et al. (2016) (16). Author: M. Hrabovský.

45
be disputed by the theory of the fusion of true integument with megasporangium as it is known in Lagenostoma. Lyginopteridaleans are the best model for studying the evolution of the gymnosperm leaves. The most primitive genera as Elkinsia (Fig. 3.5), Telangiopsis (Fig. 3.6) or Telangium (Fig. 3.7) are well comparable with psilophytes. The Palmatopteris (Fig. 3.8) or Eusphenopteris pinnule (Fig. 3.9) is homologous to one appendage (a brachyblast with few archaeophylls). From pinnae of previous genera, the foliage of Mariopteris (Fig. 3.10) or Lyginopteris (Fig. 3.11) is derived. Sterile segments of archaeophyll can either form a single cupule as in Cosmosperma (Fig. 3.12), Eonotosperma, Lagenospermum or Lagenostoma (Fig. 3.13), or whole fertile pinnule of Palmatopteris type is transformed into compound cupule as in Moresnetia (Fig. 3.14), Elkinsia, Calathospermum (Fig. 3.15) or Archaeosperma (Arnold 1939, Rothwell et al. 1989, Anderson et al. 2007, Césari 2014, Wang et al. 2014). The pollen organs are well comparable with sporangiate appendages of psilophytes with the difference that they tend to form synangia. Pollen organ of Parkvillia (Fig. 3.16) also had an affinity with Lyginopteridales (Serbet et al. 2016). The lyginopteridalean (called also hydrasperman) ovule had an exceptional structure. The lagenostome or salpinx took over the function of a still dysfunctional integument. The terminology for this structure is different, but in the Fig. 3 I follow the terminology of Long (1944) where the salpinx is a structure above the lagenostome and the lagenostome surrounds the central column and pollen chamber. The microsporophyll of Crossotheca is associated with the Lyginopteris foliage but according to its morphology, described in detail by Brouschmiche (1982), it is very similar to the order Callistophytales. However, if the pinnule had no blade and the sporangia sat on an extended receptacle, then the affinity with lyginopteridaleans is permissible.
Calamopityaleans (Calamopityales) had mostly bicornute seeds (Fig. 4.1) borne in multiovular cupules (Anderson et al. 2007). Radiospemic Eurystoma (Fig. 4.2) had cupules homologous to Sphenopteridium pinnae (Fig. 4.3) as it is clear from branching of vascular bundles in cupules described by Long (1965). The unplaced family Austrocalycaceae with genera Triphyllopteris (Fig. 4.4), Fedekurtzia (Fig. 4.5) and Austrocalyx (Fig. 4.6) have modified leaves of Sphenopteridium origin which tend to simplify into a pinna with entire leaf blade (Vega, Archangelsky 1996; Coturel, Césari 2017). They also have unique cupules homologous to simplified Sphenopteridium pinnae, which could suffice for placement of this family into the order Calamopityales.
Another unplaced family Cornucarpaceae represented by Cornucarpus ovules and Eremopteris foliage (Fig. 4.7) lost sterile segments of archaeophyll under bicornute ovules, therefore genera placed here are secondarily missing the cupules. I do not agree with the placement of this family into Callistophytales (Fig. 8.6–10) (Meyen 1987, Doweld 2001) due to a missing sterile segment, which forms a leaf blade in this order. Its placement as a satellite family to Medullosales is more likely (Cleal et al. 2009), but I rather suggest its placement into Angaridiales because the ovules of Medulosalles are radiospermic, not bicornute. The family Angaranthaceae (Fig. 4.8–10), very similar to Cornucarpaceae, was also placed within Callistophytales (Naugolnykh 2012a), but Ignatiev, Moseychik (2015) placed it into the order Angaridiales (in original paper as Angaridales from type Angaridium). This order as well as the family Cornucarpaceae could have evolved from calamopityaleans or medullosaleans. If the placement of the family Cornucarpaceae into the order Angardidiales would be considered, thus the redefined order is more ginkgoid than cycadoid in nature, because it is more reminiscent

46
Fig. 4. The leaf evolution in the subclass Ginkgoidae – Calamopityales (1–6) and Angaridiales (7–10). 1. Lyrasperma, a longitudinal section of ovule; 2. Eurystoma, a cupule homologous to Sphenopteridium pinna; 3. Sphenopteridium, a pinna homologous to a branch with appendages; 4. Triphyllopteris, a pinna and a male pinnule with archaeophyllous segments; 5. Fedekurtzia, a pinna (a), a fertile leaf with cupules and reduced basal pinnae (b), a seed with adjacent part of cupule (c); 6. Austrocalyx, a fertile leaf; 7. Eremopteris, a pinnule; 8. Angaridium, pinnules of different shape (a,b); 9. Angaranthus, a male pinnule; 10. Gondwanotheca, a female pinnule. Modified after Anderson et al. (2007) (1,3); Long (1965) (2); Skog, Gensel (1980) (4); Coturel, Césari (2017) (5); Vega, Archangelsky (1996) (6); Cleal et al. (2009) (7); Naugolnykh (2012a) (8). Author: M. Hrabovský.

47
of the calamopityaleans or peltasperms than medullosaleans or callistophytaleans with their pollen organs.
The following group is directly related to the previous two orders Calamopityales and Angaridiales. Genus Dichophyllum is traditionally classified into Trichopityaceae (Meyen 1987) or rarely into its own family Dichophyllaceae (Naugolnykh 2007). Its bipartite fronds (Fig. 5.1–2) morphologically represent the reduced fronds of lyginopteridaleans and calamopityaleans and their acupulate seeds of Samaropteris type are related to the angaridialeans. From this genus, orders Dicranophyllales, Peltaspermales, Psygmophyllales, core ginkgopsids – Ginkgoales, Hamshawviales, Iraniales, Leptostrobales (Andrews 1941, Meyen 1987, Naugolnykh 2007) as well as another unplaced Permian fossil taxa as Kladistamuos (Fig. 5.3), Nystroemiaceae (Fig. 5.4) or Glossopteridales should be derived. Therefore, Dichophyllum should be classified into its own order, which would have basal phylogenetic position to the above mentioned plant groups.
The putatively coniferous order Dicranophyllales consists recently of two families Dicranophyllaceae and Trichopityaceae (Anderson et al. 2007). Meyen (1987) classified the family Dicranophyllaceae into conifers and Trichopityaceae into ginkgoes. The original frond of Dichophyllum type is reduced to needle-like dichotomized leaf (Fig. 5.5–7) in this order, what led to the concept that dicranophyllaleans were conifers. But the less reduced megasporophylls of Dicranophyllum (Fig. 5.5) and Polyspermophyllum (Fig. 5.6) (Archangelsky, Cúneo 1990) are comparable with polysperms of Glossopteridales (Fig. 5.13) and Psygmophyllales (Fig. 5.8) (Naugolnykh 2007) and not with single seed scales of Cordaitales, Voltziales or other conifers. On the other side, the family Trichopityaceae is almost indistinguishable from conifers due to presence of axillar strobili, only the leaf anatomy and morphology of Trichopitys (Fig. 5.7) proves that it belongs to Dicranophyllales. The axillar position of its fertile organ suggests that it is a true strobilus and not a single polysperm known from Dicranophyllaceae which bears megasporophylls mixed with sterile leaves on macroblasts. But the presence of a single strobilus does not immediately mean the affinity with conifers, because a single strobilus is a feature known for many core ginkgopsids. In the genus Kandyria, the axillar strobilus is reduced to one sporophyll which is reduced to an ovule (Naugolnykh 2007). Such reduction to one sporophyll is a quite common feature in the subclass Ginkgoidae (e.g. Arberia, Nystroemia).
The families Psygmophyllaceae and Cheirocladaceae represent a transition from Dichophyllum to ginkgoaleans (Fig. 5.8–10). Naugolnykh (2007) classified them within the order Ginkgoales. But accepting the concept of Ginkopsida sensu Anderson et al. (2007), the Psygmophyllum (Fig. 5.8) would be an ancestor form not only for Ginkgoales, but also for Leptostrobales and other orders. Therefore, the order Psygmophyllales could be used in systematics. Polysperms of Psygmophyllales are homologous to sterile leaves which could be described as metamorphosed bipartite fronds bearing simplified pinnae formed by archaeophylls.
The order Glossopteridales (Fig. 5.11–15) has unclear phylogenetic position. It could be evolved from preginkopsids (Meyen 1987) or from coniferous Cordaitales (Anderson et al. 2007; Mcloughlin, Prevec 2019). There are two keys to solve this dilemma. Are the Glossopteris leaves microphylls like in the conifers or megaphylls like in the ginkgopsids? Are pollen and ovuliferous reproductive organs (fertiligers) compound

48
Fig. 5. The leaf evolution in the subclass Ginkgoidae – Dichophyllaceae (1–2), Kladistamuos (3), Nystroemiaceae (4), Dicranophyllales (5–7), Psygmophyllales (8–10) and Glossopteridales (11–15). 1. Dichophyllum moorei, a bipartite leaf; 2. Dichophyllum flabellifera, a pinna; 3. Kladistamuos, a male pinna (above) and equivalent sterile pinna (bellow); 4. Nystroemia, an ovulate leaf (a), a male pinna (b) and associated Chiropteris foliage (c); 5. Dicranophyllum, a reduced megasporophyll; 6. Polyspermophyllum, a megasporophyll; 7. Trichopitys, a reduced megasporophyll and a trophophyll; 8. Psygmophyllum, a leaf with associated Biarmopteris polysperm; 9. Alternopsis, a leaf and a megasporophyll; 10. Cheirocladus, a megasporophyll; 11. Gangamopteris; 12. Glossopteris; 13. Arberia minasica, a polysperm; 14. Arberia madagascariensis, a polysperm; 15. Rygbia, a flabellate polysperm. Modified after Andrews (1941) (1); Carrizo, Archangelsky (2013) (3); Wang, Pfefferkorn (2009) (4); Meyen (1987) (5,8); Archangelsky, Cúneo (1990) (6); Read (1933) and Florin (1949) (7); Naugolnykh (2012b) (8); Naugolnykh (2007) (9–10); Schwarz (1912) (11); Prevec (2011) (12); Cariglino (2015) (14); Mcloughlin, Prevec (2019) (13,15). Author: M. Hrabovský.

49
strobili like in the gnetophytes or simple strobili like in the ginkgopsids? The venation in glossopterid polysperms is comparable with sterile leaves, so one sterile leaf could be homologous to one polysperm. Then one fertiliger could be defined as a sterile leaf (often) fused with axillar strobilus bearing one or few polysperms (megasporophylls) as claims one of the hypotheses of Mcloughlin, Prevec (2019). If the sterile leaf would be an archaeophyll, the polysperm had to be defined as a fertile cladode. Then genera with more polysperms in a fertiliger (e.g. Lidgettonia, Eretmonia) had to have compound strobili related to gnetophytes. However, the compound strobili require bracts under partial strobili and this feature is not known in glossopterids. Therefore, the fertiligers are simple strobili and the leaves are megaphyllous. The flange segments above the ovules may be equivalent to seed scales of conifers (Rothwell 1982; Mcloughlin, Prevec 2019), but such segments are also known in ginkgopsids (e.g. Matatiellales, Leptostrobales) and all such forms could be homologous to the sterile segments of archaeophylls bearing ovules. In conclusion, the ginkgoid origin of Glossopteridales is more likely. The presence of polysperms suggests that glossopterids are more related to Psygmophyllales and other core ginkgopsids than to the Peltaspermales. Their leaves are in principle very similar to leaves of Ginkgo, Chiropteris (Fig. 5.4c), Kannaskoppifolia (Fig. 6.12) or Sagenopteris (Fig. 6.16), so it is possible that they are derived from a Dichophyllum-like ancestor.
The order Peltaspermales is characterised by peltate cupules (Fig. 6.1–2). One cupule seems to be analogous with a sporangiophore of Equisetum, so it could be defined as a metamorphosed appendage or a Sphenopteris pinnule. The cupule itself is formed by sterile segments of ovule bearing archaeophylls, which should be equivalent with flanges of Glossopteridales or cupules of Lyginopteridales. The leaves of peltasperms are compound or simplified pteridophylls (Fig. 6.3–5). And here, based on the criteria of homology between sterile and fertile leaves, a problem arises in the traditional understanding of the fertile organs of peltasperms and related orders as Caytoniales, Hlatimbiales, Matatiellales and Umkomasiales. If the sterile leaf is compound, the fertile leaf is also expected to be compound. So, the cupules traditionally defined as megasporophylls (Anderson et al. 2007) are not sporophylls but only pinnules forming the peltate structures. The structure traditionally called strobilus is the true megasporophyll. The missing bract under this structure is evidence of it being a megasporophyll and not a strobilus. The true microsporophylls are equivalent to the megasporophylls and have also the appearance of a strobilus. One ovule is homologous to a fascicle of male sporangia (e.g. Antevsia; Anderson, Anderson 2003).
The related order Matatiellales has the same problem in the interpretation of strobili. What is called a strobilus is really a compound megasporophyll or microsporophyll. The palmate cupules (Fig. 6.6) are homologous to Sphenopteris pinnules and the flanges under the seeds have the same origin as those of glossopterids or peltasperms.
The corystosperms (Umkomasiales) are cupulate ginkopsid plant with compound pteridophylls (Fig. 6.7–10). Their cupules are homologous to the order Peltaspermales with the difference that only one archaeophyll from the original pinnule is fertile (Fig. 6.11). The cupules are arranged on the strobilus-like megasporophyll in small numbers. Some basal cupules are reduced to sterile leaflets. The whole megasporophyll is subtended by a bract (Anderson, Anderson 2003; Anderson et al. 2007). There are two possible interpretations of this bract. It could be the reduced basalmost cupule, but the

50
Fig. 6. The leaf evolution in the subclass Ginkgoidae – Peltaspermales (1–5), Matatiellales (6), Umkomasiales (7–11), Petriellales (12–14), Caytoniales (15–17), Hlatimbiales (18), Perezlaria and Vladimariales. 1. Peltaspermum monodiscum, a cupule homologous to a single pinnule (an appendage); 2. Peltaspermum turbanatum; a cupule; 3. Lepidopteris scassoi, a part of a pinna with pinnules; 4. Lepidopteris stormbergensis, a part of a pinna with pinnules; 5. Tatarina, a leaf; 6. Matatiella rosetta; a pinna and a cupule; 7. Dicroidium irnense, a part of a pinna with pinnules; 8. Dicroidium hughesii, a pinna; 9. Dicroidium jordanense, a pinna; 10. Jordaniopteris robustum, a pinna; 11. Umkomasia, a cupule; 12. Kannaskoppifolia, a leaf; 13. Kannaskoppianthus, a male cupule (above) and Kannaskoppia, a female cupule (below); 14. Petriellaea, a pair of cupules (a megasporophyll); 15. Caytonia, a cupule; 16. Sagenopteris, a single pinna (a) and a leaf with pinnae (b); 17. Caytonanthus, a male pinna; 18. Hlatimbia, a bipartite cupule. Modified after Anderson et al. (2007) (1,2,6,13–15,17); Elgorriaga et al. (2019a) (3); Anderson, Anderson (2003) (4,12,18); Meyen (1969) (5); Blomenkemper et al. (2020) (7–9); Anderson et al. (2019) (10); Shi et al. (2016) (11); Elgorriaga et al. (2019b) (16); Delevoryas, Gould (1971) (19); Gordenko (2010) (20); Herrera et al. (2017) (21,22). Author: M. Hrabovský.

51
interpretation that the bract is a sterile leaf subtending the axillar stobilus, which is reduced to one megasporophyll and has the appearance of a strobilus, is more likely. As it was said, this feature is common in ginkgoid pteridosperms (e.g. Nystroemia, Arberia) and this hypothesis could be supported by a presence of bracts under the male microsporophylls of Pteruchus (Anderson, Anderson 2003). The Pteruchus microsporophyll could be evolved by a simplification of Zuberia bipartite leaf (Frenguelli 1946). The sterile equivalent to fertile structures of corystosperms is the Dicroidium foliage (Fig. 6.7–9) (Escapa et al. 2011; Blomenkemper et al. 2020).
The order Petriellales is characterized by flabellate leaves like Ginkgo, which tend to anastomose (Fig. 6.12). These leaves are of compound origin, so the fertile leaves should be compound as well. Therefore, the interpretation of Anderson et al. (2007) should be altered as it was for peltasperms. The cupules are not megasporophylls, but the true megasporophyll is a whole bipartite strobilus-like frond bearing single seed cupules and the true microsporophyll similarly bears male pinnules (Fig. 6.13–14). Several of these sporophylls grow axillary on one brachyblast, subtended by a bract. Such arrangement could lead to the evolution of corystosperm fertile organs by reduction to one sporophyll.
The same problem with terminological interpretation is in the order Caytoniales. The cupules (Fig. 6.15) homologous to Sphenopteris pinnules are called megasporophylls (Anderson et al. 2007) but based on the comparison with the compound sterile Sagenopteris leaves (Fig. 6.16b), the megasporophylls should be also compound. Therefore, strobili of Caytonia or Caytonanthus are not true strobili but the megasporophylls bearing cupules or pollen pinnae (Fig. 6.17). This claim is supported by the absence of bracts under such megasporophylls.
Similarly, the strobili of unplaced Hlatimbiales (Fig. 6.18) are true megasporophylls bearing bivalved cupules, which may be equivalent to two Sphenopteris pinnules. The strobilus-like microsporophyll of unplaced Perezlaria bearing lateral pollen pinnules (Fig. 6.19) can be interpreted similarly (Delevoryas, Gould 1971). The systematically disputed order Dordrechtitales (Anderson et al. 2007, Bergene et al. 2017, Beltrán et al. 2021) seems to be evolved rather from corystosperms than from conifers and its megasporophylls could be interpreted like previous ginkgophytes.
Another cupulate ginkgopsids related to Peltaspermales and Umkomasiales belong to the order Vladimariales. The older name for this order is Umaltolepidales (Doweld 2001), but Anderson et al. (2007) did not accept the erection of a new order and placed the family Umaltolepidaceae into the order Ginkgoales as was proposed by Zhou (1997). The new fossil genus Vladimaria led to a reassessment of older concepts. The cupules of Vladimaria (Fig. 6.20) and Umaltolepis (Fig. 6.21) are homologous to the cupules of peltasperms and corystosperms (Gordenko 2010, Herrera et al. 2017). So, they can be defined as a metamorphosed pinnules from a compound ancestral megasporophyll. The unique feature is the presence of a collar under the cupule of some species. The sterile leaves (e.g. Pseudotorellia, Fig. 6.22) are also advanced in simplification and resemble the leaves of Cordaitales. They could represent one or few archeophylls preserved form the compound ancestral leaf.
Countless studies have been written about the order Ginkgoales (consisting of families Karkeniaceae, Yimaiaceae and Ginkgoaceae), but the theory of archaeophyll can still bring new insights to homology. The sterile fan-shaped leaf (Fig. 7.1) can be derived from the compound leaf of Dichophyllum or Psygmophyllum (Meyen 1987;

52
Fig. 7. The leaf evolution in the subclass Ginkgoidae – Ginkgoales (1–6), Leptostrobales (7,9–11), Hamshawviales (8,13) and Iraniales (12). 1 Ginkgo biloba, a leaf; 2. G. biloba, an unreduced megasporophyll; 3. G. biloba, a megasporophyll; 4. G. biloba, a mikrosporophyll; 5. Karkenia, a megasporophyll; 6. a fossil aberrant ginkgoalean leaf; 7. Ginkgoites, a leaf; 8. Sphenobaiera, a leaf; 9. Avatia, a megasporophyll (left) and microsporophyll (right); 10. Leptostrobus megasporophyll with Czekanowskia leaf (above) and Ixostrobus microsporophyll (bellow); 11. Schmeissneria, a leaf and a megasporophyll with associated Stachyopitys preslii microsporophyll; 12. Irania, a megasporophyll and a microsporophyll; 13. Hamshawvia megasporophyll with associated Stachyopitys matatilongus microsporophyll. Modified after Fuji (1896) (2); Anderson et al. (2007) (5,9,10,11,13); Fischer et al. (2010) (6); Anderson, Anderson (2003) (7); Herman, Spicer (2010) (8), Harris (1951) (10); Kimura, Ohana (1978) (10) and Meyen (1987) (12). Author: M. Hrabovský.
Naugolnykh 2007). Based on the criteria of homology, the fertile leaf should be also derived from the compound leaf, but the simplification must be taken into account. The female ovuliferous organ could be interpreted as a single leaf (Fig. 7.2) or a strobilus (Douglas et al. 2007). Its position in the axil of a bract leaf (Wettstein 1899; Karstens 1945; Douglas et al. 2007) indicates the stobilar nature of this organ. Even if some ginkgopsids (Glossopteridales, Hamshawviales, Trichopityales, Nystroemiaceae) have strobilus reduced to one megasporophyll, anomalous forms of Ginkgo with a terminal bud between ovules (Fuji 1896; Wettstein 1899) or fossil relatives bearing axillar ovulate strobili (e.g. Karkenia, Yimaia) testify the axial origin of the female organ. Two types of aberrant forms indicate that the female structure of Ginkgo is a simplified strobilus bearing reduced megasporophylls (Fuji 1896), quite like Trichopitys strobilus. So, one megasporophyll consists of one ovule and a collar (Sakisaka 1927, Fig. 7.3). In Karkenia

53
the collar lacks (Fig. 7.5). The megasporophyll is homologous to the sterile leaf and its nature is megaphyllous as it is supported by fossil nonreduced specimens (Fischer et al. 2010, Fig. 7.6). As for the collar, already Douglas et al. (2007) assumed that it is not homologous to a cupule, and the findings of fossil Vladimariales confirmed it (Gordenko 2010, Herrera et al. 2017). It was defined as a sui generis structure. The marginal outgrowths on the collar in Nagrenia could lead back to the cupulate origin theory, but they could be rather interpreted as remnants of a reduced leaf blade (Gordenko, Broushkin 2013). This is supported by the morphology of aberrant leaves of ginkgo (Fuji 1896), which simultanously bear a remnant of leaf blade, collars, and a few ovules. Indeed, all facts indicate that the collar is a sui generis organ and I suppose that this mechanism is analogous with floral hypanthium. After all, the ovule was formed in the same way in this subclass. The occasional presence of a flap between the collar and the ovule led to an interpretation of the flap as a structure sui generis (Douglas et al. (2007). But if the whole part above the collar is homologous to one cupule of Vladimariales, the flap could be either homologous to a sterile scale, a sterile segment of an archaeophyll subtending the ovule or to another reduced ovule. Male strobilus is also axillar and subtended by a bract leaf. The microsporophylls are reduced to a stalk with most often two sporangia (Fig. 7.4), only the basal most ones are in the form of sterile bracts (Mundry, Stützel 2004a). This simplification of fertile and sterile organs of Ginkgo led to the assumption that this genus is related to conifers (Benson 1921; Mundry, Stützel 2004a). However, the aberrant forms (Fuji 1896) and the prefoliation are the evidence that the leaf of Ginkgo is morphologically similar to a leaf of Sphenophyllum and therefore, it could be interpreted as a bipartite compound appendage of archaeophylls.
Last group of ginkgopsids with fan-shaped leaves includes several families or orders. The fossil leaves of Ginkgoites (Fig. 7.7) and Sphenobaiera (Fig. 7.8) appear clearly like compound structures, which connects them with the leaves of Psygmophyllum. Fertile genera connected with these two foliage genera differ from ginkgoaleans by the presence of sterile segments of archaeophyll under the ovule fused into palmate blade (Fig. 7.9). The morphological interpretation of ovulate structures presented in this article will be not quite identical with the traditional explanation (Anderson et al. 2007). The ovulate structures of Avatiaceae (Fig. 7.9), Leptostrobaceae (Fig. 7.10) or Schmeissneriaceae (Fig. 7.11) are interpreted as strobili bearing pairs of megasporophylls. I agree that these strobili are true axillar strobili, but the pair of megasporophylls should be interpreted not as a pair but as single bipartite megasporophyll. Such bipartite megasporophyll is fully homologous to a sterile fan-shaped leaf and so its interpretation should be established as a strobilus reduced to a single simplified compound bipartite leaf. Since the ovules are subtended by remnant sterile segments of archaeophylls, these tend to form heads or cupules as it was in previous lineages of ginkgoid pteridosperms. The family Avatiaceae was placed within the order Ginkgoales (Anderson et al. 2007), but just its megasporophylls (specifically the morphology of fused female archaeophylls) as well as microporophylls support the placement of this family into the order Leptostrobales. The family Schmeissneriaceae had cupules derived from leptostrobaleans megasporophylls. It was placed within Ginkgoales (Anderson et al. 2007). But there was an attempt to classify it as an angiosperm by Wang et al. (2007) who interpreted putative resin bodies as seeds (Herendeen et al. 2017). Although Wang (2017) fought for his hypothesis, there are still several ambiguities. The male strobili and leaves associated with the genus

54
Schmeissneria (Fig. 7.11) are typical ginkgoid (van Konijnenburg-van Cittert 2010). The single winged seed per one cupule is the main diagnosis for this genus (Kirchner, van Konijnenburg-van Cittert 1994) and this aspect is quite different from the interpretation of Wang (2010), who interpreted the original cupule as tepals, the single seed as ovary and probably two cotyledons as two chambers of the ovary. If the angiosperm origin were admitted, the two locular syncarpous ovary with basally fused tepals would connect this genus with asterids, which would be a very interesting, just incredible discovery for the Early Jurassic. Therefore, I take the original diagnosis and the geological time into account and I propose to include this family into the order Leptostrobales, where two parts of one megasporophyll form a capsule-like fruit consisting of two adjacent opposite heads with several seeds. One pair of Schmeissneria cupules may be equivalent to one leptostrobalean megasporophyll. Irania hermaphroditica (Fig. 7.12) is included into its own order Iraniales due to hermaphroditic fertile brachyblasts. Otherwise, it is morphologically almost identical with the leptostrobaleans. Both Leptostrobales and Iraniales may have arisen from a common ancestor with the order Hamshawviales and may represent a single order. The bipartite megasporophylls of Hamshawviales (Fig. 7.13) are homologous to those of leptostrobaleans. The female strobilus is reduced to one megasporophyll as in Nystroemia or Arberia. The completely preserved microsporophyll without any simplification shows what the ancestral ginkopsids could look like. It is very similar to microsporophyll of Eretmonia (Glossopteridales), which only supports the placement of glossopterids among ginkgoid pteridosperms.
leaf evolution in cycadoid (acupulate) pteridosperms (Cycadidae Engl. in H.G.A. Engler et K.A.E. Prantl3, Fig. 8–10)
For the understanding of this subclass, the origin of the gymnosperms should be revealed. As it was stated, early gymnosperms arose from progymnosperms (Beck 1960; Stewart, Rothwell 1993; Taylor et al. 2009), most likely from the subclass Psilophytidae in the sense of my previous publications (Hrabovský 2020). But I deny a simple cladogenesis: although the early gymnosperms described here as ginkgoid pteridospems are morphologically more similar to psilophytes, the cycadoid pteridosperms resemble ferns. a hypothesis about the hybridogenic origin of some subclasses may clarify this. So, the morphology of early cycadophytes is well comparable by ferns. Although female sori are replaced by ovules, the male sori are most frequently on abaxial surface of the leaf blade. Even if the leaf blade is reduced (e.g. Medullosales), the clusters of microsporangia are quite different from the microsporophylls of ginkgophytes. The leaf origin of cycadophytes is entirely homologous to the leaf of ginkgophytes. The sterile segments of archaeophylls bearing ovules are generally preserved, but never form cupules and rather fuse to create a leaf blade as in true ferns. If these segments are reduced, the
3 Here too there is a problem with the author’s citation. Widespread “Cycadidae Pax in K.A.E.Prantl 1894” should not be a correct name. Pax (1894) named this class as Cycadaceae with the ending for a family. The form Cycadinae was published for the first time by Engler (1897), therefore the form Cycadidae H.G.A. Engler (Engl. in H.G.A. Engler et K.A.E. Prantl, Nat. Planzenfam. Nacht.: 341 (1897) could be used. The name Cycadinae was changed to Cycadidae by Tahktajan (1956, nomen nudum, p. 257). The subclass Cycadidae was accepted by Němejc (1963).

55
plants could be very similar to ginkgophytes except of sori-like microsporophylls, which are decisive in that case.
The order Medullosales was morphologically derived from the lyginopterids in the past. The medullosalean ovule was thought to be two integumentary in its nature due to the vascularised nucellus (Meyen 1984, 1987). But there is no indication of the
Fig. 8. The leaf evolution in the subclass Cycadidae – Medullosales (1–5), Callistophytales (6–10) and Gigantopteridales (11–12). 1. Pachytesta, a longitudinal section of ovule; 2. Alethopteris, a pinnule homologous to a single appendage; 3. Neuropteris, a pinnule; 4. Reticulopteris, a pinnule; 5. Whittleseya, a synangium equivalent to one pinnule; 6. Callospermarion, a longitudinal section of ovule; 7. Callospermarion, a pinnule homologous to medullosalean pinna with abaxial ovules; 8. Dicksonites, a sterile pinna (a) and a pinna with abaxial sori (b); 9. Emplectopteris, pinnules with abaxial ovules; 10. Gigantonoclea, a pinna of fused pinnules; 11. Gigantopteris, a part of a leaf showing fused pinnules and its male equivalents (Gigantotheca); 12. Gigantonomia, a bipartite megasporophyll of taeniopteroid type with abaxial ovules. Modified after Anderson et al. (2007) (1–7,9–12) and Galtier, Béthoux (2002) (8). Author: M. Hrabovský.

56
veracity of this hypothesis. The single integument of Pachytesta (Fig. 8.1) identified with lyginopterid cupule consists of three layers while nucellus has no indication of this anatomical structure (Raymond, McCarty 2009). The vascularisation of nucellus does not immediately mean its fusion with an inner integument. Then, integument of medullosaleans is a true integument and cupule is absent in this order. Sterile leaves are compound with ultimate pinnules homologous to the pinnules of marattialean ferns. The structure of Alethopteris (Fig. 8.2) or Paripteris pinules is arisen from one pinnate appendage of several archaeophylls. By shortening the main vein of the Paripteris pinnule the Neuropteris pinnule (Fig. 8.3) arises. By anastomosing the archaeophyll veins the Lonchopteris and Reticulopteris pinnules are formed (Fig. 8.4). The evolution of male sporophylls was demonstrated by several authors (Stidd 1981; Pacyna, Zdebska 2010). It is difficult to delineate the homology between sterile leaves and compound microsporophylls, since the original synangia homologous to archaeophylls are fused into compound synangial structures (Fig. 8.5) homologous to pinnules or whole pinnae (e.g. the synangium of Whittleseya could be equivalent to single Neuropteris pinnule).
The next fossil order Callistophytales had bicornute ovules (Fig. 8.6) almost identical with ginkgoid calamopityaleans, supposedly as a consequence of parallelism. However, they were borne on abaxial side of leaves, not in cupules (Fig. 8.7). The sterile segments of archaeophylls bearing ovules are laterally fused into leaf blade. The Dicksonites foliage (Fig. 8.8) can be compared with marattialean Pecopteris. The lower ultimate pinnules are reduced Sphenopteris pinnae, distal pinnules are at the level of Sphenopteris pinnules. Emplectopteris pinnules (Fig. 8.9) are morphologically equivalent to Sphenopteris pinnules although they most likely arose from simplification of whole Sphenopteris pinnae. Related genus Gigantonoclea (Fig. 8.10) had an euphyll homologous to compound Emplectopteris pteridophyll (Anderson et al. 2007).
The leaves of Gigantopteridales had the same origin. Sterile and male fertile leaves were euphylls (Fig. 8.11). The megasprophylls had ovules embedded within the leaf blade of taeniopteroid pteridophyll (Fig. 8.12). The homology of microsporophylls is not well determinable for the vague knowledge of abaxial coenosori.
Phasmatocycadales are claerly related to previous orders and their taeniopteroid leaves are morphologically equivalent to an appendage, although this type of leaf results from reduction of lateral appandages of compound leaves to one vein and their fusion. This is the case of almost all Mesozoic cycadophytes with Taeniopteris foliage (e.g. Zamiaceae, cycadeoids). The phasmatocycadalean megasporophyll bore abaxial ovules (Fig. 9.1). a simplification of a bipartite megasporophyll of Taeniopteris type with abaxial ovules can lead to the evolution of the family Dirhopalostachyaceae (Fig. 9.2).
The order Cycadales is known by reduced sporophylls forming strobili or cones, while sterile leaves are always compound. The compound origin of microsporophylls (Fig. 9.3a) is visible from their vascularisation (Mundry, Stützel 2008). One ovule (Fig. 9.3b) as well one sorus with adjacent part of blade is homologous to one archaeophyll. Pinnae of Zamia or Bowenia seem to be archaeophylls (Fig. 9.3c) similar to the leaves of coniferous genera Cordaites or Agathis, but they certainly originate from simplification of compound pinnae. It is possible to interpret the ultimate leaflets of pinnate to multipinnate leaves of Cycas as equivalent with archaeophylls, but I think this is not so due to the decurrent leaflet blade and circinate prefoliation (Fig. 9.4). They may be equivalent to the appendage with lost archaeophylls.

57
Fig. 9. The leaf evolution in the subclass Cycadidae – Phasmatocycadales (1–2), Cycadales (3–4), Fredlindiales and Bennettitales (5–10), Pentoxylales (11–13); Bernettiales (14–15); Nataligmales (16) and Fraxinopsidales (17). 1. Archaeocycas, an abaxial view of megasporophyll; 2. Dirhopalostachys, an abaxial view of bipartite megasporophyll; 3. Zamia, a microsporophyll (a), a longitudinal section of ovule (b) and a pinna of archaeophyll type (c); 4. Cycas, a megasporophyll and circinate pinna of appendage type; 5. Schematical structure of cycadeoid sporophyll with ovules and sori (a common ancestor); 6. Fredlindia, an adaxial view of megasporophyll (’gynoecium’); 7. Bennettistemon ovatum, a microsporophyll with adaxial sporangia; 8. a schematical lateral view of Fredlindia megasporophyll; 9. a bennettitalean gynoecium of fused whorl of Fredlindia megasporophylls; 10. Halleyoctenis brachypinnata, a compound leaf; 11. Carnoconites, a simplified interpretation of one megasporophyll with two pinnae (gynoecia); 12. Lindtheca, a multiovulate unit (right) and whole megasporophyll (left); 13. Taeniopteris homerifolius; 14. Bernettia inopinata, a subsidiary megasporophyll with abaxial ovules; 15. Piroconites kuespertii, a subsidiary microsporophyll with adaxial synangia; 16. Nataligma, a megasporophyll (gynoecium) with pinnae (ovules) in whorls; 17. Fraxinopsis, a megasporophyll with one adaxial ovule (bellow) and its transverse section (above). Modified after Taylor and Millay 1979 (1), Krassilov (1975) (2), Mundry, Stützel 2008 (3a); Mcloughlin et al. (2018) (6), Anderson et al. (2007) (7–13); Kustatscher et al. (2016) (14); Van Konijnenburg-van Cittert (1992) (15) and Anderson, Anderson (2003) (16-17). Author: M. Hrabovský.

58
Many scientists would rather see the following plant groups among gnetophytes, antophytes or conifers, but I will try to prove their undeniable affinity with cycadophytes. Traditional paleobotany classified cycadeoids as cycadophytes (Seward 1917, Němejc 1968, Meyen 1987). Statistical morphological analysis showed that cycadeoids should be relative to Gnetales and angiosperms and not to Cycadales (Crane 1985). I fundamentally disagree with such “phylogenetic analyses” among remote plant groups based on many morphological characteristics. Of course, they are an awesome tool for revealing the phylogenesis among relative groups but using the misinterpreted morphological characteristics, they become completely misguided. The main characteristics supporting the kinship with Gnetales are syndetocheilic stomata, pollen grains and seed anatomy (Rothwell et al. 2009). The most authoritative structure of these morphological characters is the seed. Although some interpretations tried to compare bennettitalean seeds with those of Gnetales and interpreted the bennettitalean sarcotesta as a gnetalean envelope (Friis et al. 2007), but this was not supported by other analyses (Stockey, Rothwell 2003; Rothwell et al. 2009; Pott 2016). The absence of cupules has already been proposed by Harris (1962) who redefined pentoxylalean and bennettitalean ‘cupules’ (including the cupules of Vardecloeftia) as the sarcotesta.
The origin of the cycadeoids lies in the answer to the question: what is the male and female sporophyll in the cycadeoid flower. The possible origin of the microsporphyll can be through reduction as it is in Cycadales; the microsporophyll can be at the same level as sterile leaf; it can be less simplified and more branched than sterile leaf or it can be a part of a phylloclade. The criteria of homology indicate that both sterile and fertile leaves should be homologous. So, are the pinnately compound sterile leaves (Fig. 9.10) of multiaxilar origin like Gnetum leaves, phylloclades like Phyllocladus (Fig. 11.17) or are they compound leaves like those of Cycadales? The microsporophyll is in general an ultimate fertile leaf arranged in strobilus. Since the fertile leaves should be homologous to sterile, the male pinnate structures (Fig. 9.5) could similarly be either compound strobili, phylloclades or single microsporophylls. The first option of interpretation of the leaf is homologous to a strobilus of Gnetales, which was described as compound by Mundry, Stützel (2004b). This option is impossible because of the absence of bracts under the ultimate axes in male pinnate structures. The second option is that both sterile and fertile leaves are phylloclades. In this case they would be subtended by bracts as all axillar shoots. Indeed, the bracts of Cycadolepis seem to subtend the leaves (Crane, Herendeen 2009). The same position of male pinnate structures seemingly fused with bracts in Weltrichia or Lunzia (Pott et al. 2017; Popa 2019) and sterile pinnate leaves seemingly fused with midvein of Cycadolepis bract is direct evidence of homology between them. However, this structure may not be a result of the fusion of phylloclade with bract as it seems at first glance. It could be an analogy with the leaf of ophioglossids, with the fertile part subtended by the sterile part. However, there is a new morphological feature sui generis, occurring in Triassic-Jurassic plants. It can be found in the order Bernettiales (Kustatscher et al. 2016). This new feature of one leaf subtending another, called leaf subsidiarity, was found in some aberrant extant Cycadales (Srivastava 1952). It could be an important diagnostic feature in systematics. The bract-leaf complex is de facto one leaf with a subsidiary rachis bearing subsidiary pinnae or leaf blade. In aberrant Cycas these rachises are fused, but e.g. in Bernettiales they are free (Fig. 9.14). All plant groups with leaf subsidiarity have characteristic winged leaf scars. If the microsporophylls of

59
cycadeoids were interpreted as subsidiary structures, this feature should be sufficient to accept cycadeoids as cycadophytes, therefore interpreting their leaves as compound (the third option). The male fertile leaves are true microsporophylls but non-reduced unlike the microsporophylls of Cycadales.
All following groups with the same morphological structure of sporophyll (Fig. 9.5) should be considered as cycadophytes including some putative gnetophytes sensu Anderson et al. (2007). The cycadeoids consist of several orders and families (Anderson et al. 2007). The order Fredlindiales is crucial for morphological explanation of more advanced groups. Megasporophylles (called gynoecia) with abaxial ovules (Fig. 9.6) are arranged in whorls. I cannot agree with the traditional morphological terminology (Rothwell, Stockey 2002; Anderson et al. 2007) that the ovuliferous unit (one ovule) is a megasporophyll. If we compare the whorls of Bennettistemon (Fig. 9.7), Weltrichia or Leguminanthus microsporophylls with the whorls of Fredlindia gynoecia, these gynoecia are true megasporophylls and ovuliferous units are homologous with male synangia. One pinnate sterile leaf is also homologous to one sporophyll, although the reduction of sterile leaves is more advanced than those of sporophylls. The order Bennettitales differs with radially symmetrical gynoecia and presence of interseminal scales. Micropsporophylls are arranged in a single whorl. The traditional explanation of radial gynoecium describes it as an axis with many fertile sporophylls reduced to one ovule and sterile sporophylls (interseminal scales). But such reduction of compound megasporophyll to one scale or ovuliferous unit is very unlikely if the microsporophylls are not reduced in the same way. In morphology, it is usually common that the simplest possible explanation is true. If the microsporophylls in a whorl are fused, the whorl of the bilateral Fredlindia megasporophylls is also expected to be fused in advanced morphological groups. So, I interpret the single radial gynoecium as a whorl of bilateral gynoecia (megasporophylls) fused together with central axis (Fig. 9.8–9). Similar interpretation claims that the radial gynoecium is analogous with one whorl of Gnetum (Sharma 1974, Endress 1996), but in Gnetum the structure is formed by fused lateral compound strobili, while here it represents a simple strobilus. Some of the species of this group have gynoecia arranged in lax strobili. If the radial gynoecium is a simple strobilus, the lax strobilus of Westerheimia with several pinnately attached gynoecia should be a compound strobilus. The ancestral microsporophyll in this group is a compound leaf in its nature (e.g. Weltrichia). There is a visible trend to form entire leaf blade (e.g. Leguminanthus). The synangia on these microsporophylls could be compared with synangia of Medullosales or Marattiales (Meyen 1987). One such synangium as well one ovuliferous unit are homologous to one archaeophyll based on my theory. Each adaxial synangium tends to be reduced to one sporangium (e.g. Bennettistemon p.p.) (Fig. 9.7). If the microsporophyll of Bennettistemon is compared with the megasporophyll of Fredlindia, two conclusions are deductible. Firstly, the megasporophylls precede the microsporophylls in the evolution. Secondly, the synangia are adaxial, while the ovules are abaxial. The pinnate leaves (Fig. 9.10) tend to form taeniopterid leaves. Based on the leaf venation (Moisan et al. 2011), the pinnae may be equivalent to the of Zamia.
Pentoxylales is an enigmatic group with uncertain systematic position. Němejc (1950) or Anderson et al. (2007) considered it as related to cycadeoids. The gynoecia formed by ovuliferous units were considered to be rather megasporophylls than cones (Němejc 1968). Anderson et al. (2007) called the ovuliferous units as megasporophylls and

60
gynoecia as cones, but I follow the older terminology of Němejc (1968) for the absence of any bracts subtending the gynoecia. If the pedicels with female Carnoconites gynoecia (Fig. 9.11) are compared with Sahnia microsporophylls (Bose et al. 1985; Suthar, Sharma 1989; Osborn et al. 1991, Srivastava, Banerji 2001), their homology could be revealed. Megasporophylls were advanced in evolution and formed by compact heads on peduncles while microsporophylls bore sparsely arranged clusters of sporangia. It is not clear whether the gynoecium with one pedicle is one megasporophyll or only its pinnule. The peduncle bearing pedicels with heads could be a stobilus or a megasporophyll. This is also the case of the microsporophyll, but I incline to the compound megasporophyll interpretation with the gynoecium representing a pinuule. The gynoecia of Lindtheca differed slightly from those of Carnoconites by multiovulate segments (incorrectly called megasporophylls) and an enclosing sheath (Anderson, Anderson 2003). Again, it is difficult to say whether the multiovulate segment of Lindtheca (Fig. 9.12) is homologous with the whole Carnoconites gynoecium or whether the Carnoconites gynoecium with uniovulate segments is reduced Lindtheca gynoecium. But the gynoecia are certainly not strobili, due to the absence of bracts, expected in the case of strobilar nature of these structures. The order Pentoxylales could be therefore morphologically derived only from some of cycadophytes or ginkgophytes. The taeniopteroid foliage (Fig. 9.13) and a similarity with medullosaleans (Srivastava, Banerji 2001) support its cycadoid origin.
Other orders are considered to be gnetophytes (Anderson et al. 2007), but these fossil groups bore reproductive organs not of compound strobilar origin as it is expected in Gnetidae clade (Mundry, Stützel 2004b). The extant gnetophytes are mostly Carboniferous groups, while the orders Bernettiales, Dinophytonales, Fraxinopsidales (originally published as Fraxinopsiales from the type genus Fraxinopsis) or Nataligmales are known form Triassic to Jurassic. The cycadeoid origin of these orders is more likely than their placement among coniferous gnetophytes. Everything depends on the interpretation of the origin of sporophyll. If the sporophylls were phylloclades, there is no problem with gnetophyte hypothesis, but there is no proof of their strobilar nature. On the contrary, the leaf subsidiarity supports their placement into the subclass Cycadidae. Bernettia (Fig. 9.14) had reproductive structures in cones. One reproductive structure consisted of a bract and megasporophyll with abaxial ovules (Kustatscher et al. 2016). The male representative of the order Bernettiales is a genus Piroconites (Fig. 9.15), which differs by a bract subtending a micropsorophyll with adaxial three-locular synangia (Van Konijnenburg-van Cittert 1992). Although many morphological characteristics are common with Benettitales, the pollen and synangia seems to be gnetaceous, megasporophylls are reminding on the reproductive structures of glossopterids, and cone arrangement is coniferous (Kustatscher et al. 2016). From the point of view of leaf subsidiarity, the megasporophyll and microsporophylls are not glossopteroid but subsidiary cycadoid structures. Abaxial ovules and adaxial synangia is the common feature with Bennettitales. These sporophylls may be equivalent to the cycadoid sporophylls. Cycadeoid flowers are de facto strobili like strobili of Bernettiales. Strobilar organization is common in all gymnosperms, not only conifers. The pollen type variability is large across all gymnosperms (e.g. the presence of pollen with two air sacs among the ginkgoid glossopterids, the cycadoid callistophytaleans or many conifers; Anderson et al. 2007), therefore the Ephedripites pollen does not immediately mean kinship with gnetophytes.

61
The order Nataligmales (Fig. 9.16) is, according to Anderson, Anderson (2003), a putative gnetopsid. Their “strobili” are reminiscent of those of Carnoconites (Pentoxylales), but they are placed in gnetophytes for the whorled structure and anatomical similarities with Fraxinopsidales. I cannot agree with this placement of both the order Nataligmales and Fraxinopsidales. The lateral “strobili” in the “compound” strobilus lack subtending bract, namely. a much more likely explanation is the megasporophyllous nature of these so-called lateral strobili or cones. They seem to be homologous with gynoecia of Carnoconites. The whorled organisation of ovules does not contradict this claim. The whorls are a very common feature for Bennettitales. Since the leaves of cycadoid pteridosperms are megaphylls and therefore homologous to metamorphosed lateral axes, the progress in whorl organisation of branches could be also shifted into the axes of megaphyllous leaves. Finally, if they are true strobili, why do not any leaves subtend them? Therefore, there is no obstacle to define the lateral “strobili” as megasporophylls and the “compound” strobilus as simple. The affiliated Gontriglossa foliage with reticulate venation resembles glossopterid leaves but is rather homologous with the fraxinopsidalean Yabeiella foliage of taeniopteroid origin (Anderson et al. 2007).
Regarding the placement of the order Fraxinopsidales, the seeds of Fraxinopsis (Fig. 9.17) are indeed similar to voltzialean Cycadocarpidium seed scales and to think of them as conifers is in place, but their placement among gnetophytes was based on the cuticular structure (Anderson, Anderson 2003). The venation of associated Yabeiella leaf is taeniopteroid and not gnetaceous. The clear midvein excludes the possibility of microphyllous leaf origin. Since the organ bearing Fraxinopsis seeds is not known, it is difficult to describe the homology. The gnetaceous origin of Fraxinopsis would require strobilar nature of one seed. The wing could be a bract and strobilus would be reduced to one seed. The cycadoid hypothesis would explain the bract with seed as a single megasporophyll. The leaf subsidiarity cannot be also excluded for the possible interpretation of the seed as reduced fertile part of megasporophyll to single seed. For the similarity with Nataligma and the occurrence in Triassic period the explanation of Fraxinopsis as an advanced gnetophyte with totally reduced strobili is less likely than its cycadoid origin.
The order Dinophytonales consists of the single genus Dinophyton (Fig. 10.1). Although its ovule is medullosalean, the leaves are coniferous, and cupulate structures around the ovule suggest a gnetophyte affiliation (Ash 1970; Krassilov, Ash 1988; Anderson et al. 2007). It is certain that the Dinophyton cupulate zygomorphic flower is a strobilus. The possibility that microsporophyll with one adaxial sporangium is voltzialean bract with one reduced sporangium cannot be excluded, but this is unlikely. It is then even less likely to consider this structure as a compound strobilus homologous to extant gnetophytes. Since the male and female Dinophyton flowers should be homologous, it is best to describe them as three cycles which are homologous to three fused whorls in the cycadeoid manner. The outer whorl of bracts is basally fused in the female flowers and free in the male strobili. The middle whorl is formed by a cupule in the case of the female strobilus and by free bracts in the male strobilus. These male bracts subtend the ‘microsporophylls’ with one abaxial sporangium (Krassilov, Ash 1988). The parallel orientation of the ‘microsporophyll’ with the subtending bract contrasting to the decussate orientation of alternating whorls allows to interpret this bract-leaf complex as one leaf. Indeed, this complex seems to be a true microsporophyll, not only a leaflet

62
with a sporangium. The female cupule can be equivalent to the whorl of these true microsporophylls lacking fertile leaflets. The most distant cycle is missing in the male flower, while this cycle is represented by a single ovule in the female one. It is important to understand the reduction of compound cycadoid leaf in this order. Since the sterile leaves are simplified to microphylls or reduced appendages, the sporophylls are also reduced to microphylls or appendages. So, the microsporophyll could be interpreted as consisting of a sterile part (bract) and a fertile part (leaflet with one sporangium) and the megasporophyll is reduces to a single ovule. So, the reduction of the most distal whorl of four ovules to one ovule is very simple (and therefore likely) process in the plant morphogenesis.
Seemingly unrelated order Axelrodiales (Cornet 1986) is very close to the order Dinophytonales if the morphological interpretation above is accepted. The single so-called floral subunit of Axelrodia (Anderson et al. 2007) consists of a few cycles of various bracts (Fig. 10.2). The two proximal cycles are protecting a cupule, which represents a fused whorl of bracts. However, the content of the female cupules is not clear based on the fossils. There are probably two megasporophylls with adaxial ovules (Cornet 1996) or only two ovules (Cornet 1986) inside of the cupule. Similar floral units were found in Zamiostrobus (Fig. 10.3). These partial flowers (floral subunits) subsequently form a compound strobilus with flower-like appearance. The cupule of Archaestrobilus (Fig. 10.4a) had several sterile scales and a central seed inside. These scales could be sterile megasporophylls or the whole structure could be a single metamorphosed bennettitalean megasporophyll with interseminal scales and one ovule. The female cupule is homologic with a metamorphosed whorl of fused microsporophylls
Fig. 10. The leaf evolution in the subclass Cycadidae – Dinophytonales (1) and Axelrodiales (2–6). 1. Dinophyton spinosus, a female ‘flower’ (a), a leaf (b), a male ‘flower’ (c) and a leaflet wit sporangium (d); 2. Axelrodia burgeri, a flower or floral subunit homologous to Dinophyton flower and two extracted megasporophylls with seeds (below); 3. Zamiostrobus, a floral subunit of Primaraucaria a type; 4. Archaestrobilus cupulanthus, a female flower (a), a male flower (b) and single abaxial bivalved leaflet with sporangia; 5. Synangispadixis tidwellii, a male flower with one extracted abaxial bivalved leaflet with two synangia; 6. Sanmiguelia lewisii, a leaf equivalent to a fused whorl of leaves. Modified after Ash (1970) (1a–c), Anderson et al. (2007) (1d), Cornet (1986) (2–3,6) and Cornet (1996) (4–5). Author: M. Hrabovský.

63
as it is seen in the male Archaeostrobilus cupules (Fig. 10.4b). These microsporophylls are megaphylls with ultimate bivalved leaflets. The ultimate leaflets with sporangia were abaxial in a position to the microsporophyll. There are several sterile scales inside the male cupules of Archaeostrobilus, while cupules of Synangispadixis (Fig. 10.5) were probably filled with parenchyma. The sterile leaf of Sanmiguelia (Fig. 10.6) resembles the leaf of equisetalean genus Schizoneura. This is not surprising, since all these group have leaves formed by fused whorl of reduced megaphylls. The sterile leaf should be homologous to the sporophyll, so if the cupule is defined as a fused whorl of microsporophylls, fused whorls of sterile leaves are expected on the stem.
leaf evolution in conifers (Coniferidae Juss.4, Fig. 11.)
Coniferae are plants characteristic by bearing strobili. The strobili are formed by megasporophylls or microsporophylls. Megasporophylls are reduced to a single ovule except of Cordaitanthus (Procordaiconus) pseudofluitans (Fig. 11.1) with several ovules (Ignatiev, Meyen 1989). This exception suggests a megaphyllous origin of the coniferous leaf. Based on Fig.1, this can be interpreted as the reduction of an appendage either to one microphyll or to a cladodified axis. The second option is an independent evolution of coniferous leaf from an original archaeophyll bearing several ovules, on the contrary to other gymnosperms with only one ovule per archaeophyll. Ovules can be terminal or sessile on a scale. The scale can be homologous to the sterile segment of archaeophyll or, if the megasporophyll is interpreted as a reduced axis of an appendage, the scale could be the remnant of the appendage. Since there were no scales with more ovules found (not to be confused with a seed scale, which is fused from several ovuliferous scales), the hypothesis of origin from appendage axis is less credible. The microsporophylls consist of one to several sporangia and can similarly be interpreted as microphyll. The evolutionary trend in conifers is the creation of compound strobili by simplification and cladodification of single strobili.
The leaves of the representatives of the order Cordaitales (Fig. 11.2–3) could be morphologically interpreted as archaeophylls. The female strobili bear several sterile bracts and a few pedunculate ovules (both homologous to archaeophylls). The male strobili of Cordaitanthus, Mesoxylon (Fig. 11.4) or Pechostrobus are equivalent to female strobili (Anderson et al. 2007). But the strobili of Kuznetskia seem to be cladodified and fused with subtending bracts as well as the strobili of Cladostrobus, which are arranged in a compound strobilus. In the same way, the female strobili are fused with subtending bracts in the order Palissyales. In the order Ferugliocladales the female strobili are simplified to one axillar ovule.
4 The original name Coniferae published by de Jussieu (1774). It was changed to the form Coniferidae by Tahktajan (1956, nomen nudum, p. 257) and then published as valid name by Krassilov (1989). Christenhusz et al. (2011) rejected the name Coniferidae and used the name Pinidae for a paraphyletic clade of conifers excluding the subclass Gnetidae. I include the clade Gnetidae into conifers. The argument that the name Coniferae is not based on the existing name of genus is not sufficient, because the names as gymnosperms, angiosperms, eudicots or monocots are a similar case and are just as common as the name conifers. The author of the name Pininae is Conard (1919). It was changed to Pinidae by Novák (1961, p. 132). Cronquist et al. (1966) appropriated Pinidae and a few other existing names without proper credit.

64
Fig. 11. The leaf evolution in the subclass Coniferidae – Calamitales (1–4), Voltziales (5–9), Pinales (10–12), Cupressales (13,18–23), Araucariales (14–17), Erdtmannithecales (24), Gnetales (25–28) and Dechellyiales (29). 1. Cordaitanthus (Procordaiconus) pseudofluitans, a female strobilus with proximal sterile scales and distal megasporophylls subtended by bract; 2. Cordaites gerelovae, a leaf of archaeophyll type; 3. Vojnovskya paradoxa, a leaf of archaeophyll type; 4. Mesoxylon priapi, a male strobilus with proximal sterile scales and distal microsporophylls subtended by bract; 5. Aetophyllum, a cladodified female strobilus subtended by bract; 6. Darneya mougeoti, a cladodified male strobilus fused with the subtending bract; 7. Lutanthus robustus, a cladodified male strobilus fused with the subtending bract; 8. Lutanthus ornatus, a cladodified male strobilus fused with the subtending bract; 9. Barthelia furcate, a leaf; 10. Picea abies, a pollen scale homologous to a cladodified male strobilus fused with a subtending bract; 11. Abies alba, a needle; 12. Pseudotsuga menziesii, a bract; 13. Cunninghamia lanceolata, a seed scale homologous to a cladodified female strobilus fused with a subtending bract; 14. Agathis dammara, a leaf of archaeophyll type; 15. Rissikistrobus plenus, a cladodified compound cone; 16. Podocarpus, a fleshy compound cone; 17. Phyllocladus, a phylloclade; 18. Cupressus, a compound cone; 19. Thuja, a scale leaf; 20. Cephalotaxus wilsoniana, a compound cone; 21. Taxus baccata, a simplified compound cone; 22. Cephalotaxus, a compound male cone; 23. Taxus, a modified compound male cone; 24. Bayeritheca, a modified compound male cone; 25. Welwitschia mirabilis, a leaf and a modified compound male cone; 26. Ephedra, a modified compound male cone; 27. Gnetum, an euphyll and a modified compound male cone; 28. Drewria potomacensis, a leaf; 29. Dechellyia gormanii, a leaf. Modified after Anderson et al. (2007) (1–2,4–5,7–10,13,15,25–29); Mamay (1976) (3); Grauvogel-Stamm (1978) (6); Lo, Wang (2001) (20); Dörken, Nimsch (2016) (22–23); Kvaček, Pacltová (2001) (24). Author: M. Hrabovský.

65
In the order Voltziales and their relatives there is a tendention to simplify female strobili to a few scales, partially fused with the subtending bracts (Fig. 11.5). These simplified strobili formed the compound cone. Male compound cones in conifers are more advanced than the female. The male strobili are reduced to a few microsporophylls fused with subtending bracts (Fig. 11.6–8). The homology of seed scales subtended by a bract and pollen scales is supported by anomalous hermaphroditic cones of recent conifers (Rudall et al. 2010). Therefore, this bract-strobilus complex (pollen scale) is not a single microsporophyll. Morphologically, the leaves of Voltziales are microphylls with several veins or single midvein (Fig. 11.9).
The recent order Pinales differs from the previous order by the seed scales formed by cladodified strobili composed from two to three scales (Rudall et al. 2010). The subtending bract (Fig. 11.12) is free from the seed scale. The cladodified male strobili are reduced to two sporangia fused with the subtending bract (Fig. 11.10). The leaves are one-veined microphylls, which could be called lycophylls, but it is better to use the popular name needles for them (Fig. 11.11).
The orders Araucariales and Cupressales are sometimes included within the order Pinales, but in this case the monophyly is controversial due to the relatedness of Pinales with gnetophytes (Christenhusz et al. 2011) and it is better to recognize more orders than to include the order Gnetales within the order Pinales. The female strobilus in Araucariales and Cupressales is formed by two to several scales fused to each other and with subtending bract (Fig. 11.13). In both orders there is a tendention to simplify or metamorphose compound cones. The family Araucariaceae (Araucariales) has archaeophylls (Fig. 11.14) or single veined needles. The compound cones of the family Podocarpaceae from the same order are reduced to a few seed scales fused with bracts (Tomlinson 1992; Mill et al. 2004). The fossil genus Rissikistrobus (Fig. 11.15) had these simplified cones arranged in a bicompound cone whereas recent genera have these simplified cones fleshy (Fig. 11.16). a special case is recent genus Phyllocladus which has cladodified shoots (Fig. 11.17) and a less simplified type of cone. The female strobili are simplified to one terminal ovule with a rudimentary scale, subtended by a bract. The rudimentary scale may become an aril in the late ontogenesis. The simplified female strobili are arranged in a compound cone (Tomlinson et al. 1989).
The cone of the family Cupressaceae (Cupressales, Fig. 11.18) is equivalent to a compound cone of Pinales and Araucariales. The scale-like or needle-like microphylls (Fig. 11.19) are arranged either spirally or in decussate pairs. In the family Taxaceae (Cupressales) compound cones tends to be simplified similarly to Podocarpaceae. The female compound cone of Cephalotaxus consists of bracts, which are bearing strobili represented by terminal apex and two ovules (Fig. 11.20). But usually only one ovule from the whole compound cone becomes a seed (Lo, Wang 2001). The ovules lack scales and therefore the fleshy seed coat could be derived from the integument (a sarcotesta) (Conteras et al. 2017). But Dörken et al. (2019) believe that the scale is fused with the ovule integument and then it is not the integument, which is fleshy, but the scale that can be interpreted as an aril. To determine the origin of aril, we can look at the related genus Torreya, which has an aril fused with integument (Stützel, Röwekamp 1999). In generally, taxaceous genera have compound female cone reduced to bracts and to single terminal strobilus subtended by two fused bracts. The terminal strobilus consists of one terminal ovule without scale. Therefore, arillate structures in Torreya or Pseudotaxus are

66
formed just by the bracts of the compound cones, not by scales (Stützel, Röwekamp 1999; Dörken et al. (2019). Therefore, the aril of Torreya, Taxus (Fig. 11.21) or Pseudotaxus cannot be homologous to the aril/sarcotesta of Cephalotaxus. And if we consider the loss of scales from terminal strobili in Phyllocladus, Taxus or Torreya, the loss of these scales in Cephalotaxus is more likely than their fusion with the integument. The sarcotesta was common in early conifers and it is present even in recent Podocarpaceae, where the sarcotesta with fleshy seed scales forms the epimatium (Tomlinson 1992). And in that case, the aril is clearly differentiated into an outer fleshy layer and the inner fleshy sarcotesta (Mill et al. 2004). The compound male strobili (Fig. 11.22) are arranged into bicompound strobilus, subtended by bracts. But the compound strobilus tends to reduce to two terminal voltzialean strobili (bracts with several sporangia) which are fused into one peltate sporangiophore (Fig. 11.23). So, this peltate sporangiophore is homologous to the compound male strobilus of Cupressales. The bracts under these compound strobili are present in Cephalotaxus or Pseudotaxus, but absent in Taxus (Dörken et al. 2011). Male cones of Taxus are thus a simplified bicompound strobilus consisting of the cupressalean compound strobili.
The peltate sporangiophores of Cretaceous Erdtmannithecales could be interpreted in the same way (Fig. 11.24). This order could also be compared with bennettitaleans, but these had their evolutionary radiation node dated to the Triassic. The Cretaceous period is the time of advanced conifers as Taxaceae or gnetophytes. Putative gnetophytes Angarolepis and Aegianthus (Krassilov, Bugdaeva 1988) may be homologous to Erdtmannithecales or Taxales.
The incomprehensible systematic reassignment of the order Gnetales indicates a misunderstanding of their morphology. Understanding of trends among recent conifers is necessary for the placement of this order. These trends lead to reduction of scales under ovules, and even to the reduction of the whole strobilus to one ovule (e.g. Phyllocladus, Taxaceae). Compound strobili are simplified to various lateral structures, which are subsequently arranged in a bicompound strobilus. Even the two terminal strobili in the male compound strobilus are fused together in Taxus. Now, if we look at extant gnetophytes, we can find the same evolutionary trends. Since Taxaceae are advanced forms of Cupressales and Podocarpaceae of Araucariales, the question stands: advanced forms of which group are the gnetophytes? Because they are quite different from Pinales, I think that they evolved from Voltziales and morphological parallels should be considered between these orders. Mundry, Stützel (2004b) suggested the possible morphology of gnetophyte ancestor. I can’t disagree with it, even if it’s not complete. In the plant morphology, the principle of the simplest possible solution applies. The Welwitschia strobilus has a simpler explanation than the one suggested. The different positions of ovules in the male fructifications of Ephedra, Gnetum and Welwitschia may be confusing, but there is a simple morphological formula for the homology in Gnetales. The basic structure that is metamorphosing in extant conifers is the compound cone – strobilus of cordaitalean strobili. And these metamorphosed compound cones form bicompound strobilus (e.g. Rissikistrobus, Taxaceae). In the order Gnetales, the smallest degree of metamorphosis of the compound cone (called sometimes ‘flower’) is in Welwitschia (Fig. 11.25), not in Ephedra or Drewriaceae. The compound cone of Welwitschia has, in fact, all three decussate pair of bracts which could in the past subtend six lateral strobili and a terminal one. In the male ‘flowers’, the lower transverse bracts are free, the median

67
and upper bracts are fused. The upper pair of bracts is bearing six synangia, three per one bract. Mundry, Stützel (2004b) did not recognize this pair as bracts bearing ultimate strobili. But look at Voltziales, Pinales, Araucariales or Cupressales. The terminal strobilus in these groups is reduced to a few sporangia, always fused with the subtending bract, and this is also the case in the Gnetales. The interpretation of female cone by Mundry, Stützel (2004b) based on Martens (1961) is again too complicated. Lignier, Tison (1912) or Church (1914) reported only one pair of bracts in the female ‘flower’. So, the lower transverse bracts in the female flower form the whole envelope of seed, the other pairs of bracts are absent. And in both male and female flower, the terminal strobilus is reduced to one ovule, although it is only fertile in female flowers. These flowers (compound strobili) are a part of a bicompound cone. This bicompound cone is homologous with the cones of Ephedra or Drewriaceae. The male flower of Ephedra lacks transversal bracts (Fig. 11.26). The median bracts are fused. The upper pair is also fused and forms the anterophore. The anterophore is homologous with two strobili of Cordaianthus or with a peltate taxaceous sporangiophore. The drewriaceous Eamesia also has an anterophore homologous to the Ephedra anterophore, but the lower two pairs of bracts are not fused (Yang et al. 2018). The sterile ovules are absent in the male flowers. But, since the bicompound strobilus was originally hermaphroditic, the sterile female flowers can be sometimes preserved in the male bicompound strobilus, what led to many previous misinterpretations. The female flower of Ephedra has one lower pair of fused transversal bracts, other pairs of bracts are absent as in Welwitschia. The bracts in the female bicompound strobilus subtending the female flowers become fleshy in ontogenesis. There are no male flowers in the female bicompound strobilus, while in the male bicompound strobilus the female flowers can be in the distal position. When the structure of Welwitschia and Ephedra flowers is understood, the ‘inflorescence’ of Gnetum is not difficult to interpret. Based on anatomy and floral development (Endress 1996; Becker et al. 2003) the “whorls” can be homologised with two fused bicompound strobili subtended by two fused bracts. The basipetal development of flowers (compound strobili) indicates the proximal position of female flowers and distal position of male flowers in original unmetamorphosed fructification, what is the exact opposite of the Ephedra complex cone. But since all flowers were originally hermaphroditic as in Welwitschia, such a change of flower sexuality is possible. The female flowers consist of the fused lower transverse bracts (outer envelope) and the fused median bracts (inner envelope). The third pair of the upper transverse bracts is absent. In the male flowers (Fig. 11.27) the lower pair of transverse bracts is absent as in Ephedra. Like Ephedra, the fused median bracts envelope the upper pair of transverse bracts, which are fused and form the anterophore. Each upper bract has only one sporangium, therefore the anterophore bears two sporangia. The Gnetum flowers differ from the Ephedra only by the number of sporangia in the male flowers, by the presence of the median bracts in the female flowers and by the absence of bracts under flowers. The sterile leaves of Gnetales are archaeophylls in their nature. They are arranged either in decussate pairs or in whorls of three (Yang 2004). The leaves of Drewriaceae are comparable with voltzialeans archaeophylls apart from anastomosed lateral veins (Fig. 11.28). The Welwitschia leaves are then the largest and the longest living microphylls in the world. Ephedraceous leaves are reduced to scales with two veins. Gnetum lost microphylls with the exception of a few microphyllous bracts in the inflorescence and flowers. The leaves of Gnetum are

68
euphylls with anastomosed veins. They arose from foliation (leaf formation process) of a complex of axes bearing archaeophylls.
The order Eoanthales could be related to Gnetales according to Krassilov (1986). It only differs by four bracts in whorls and naked ovules subtended by bracts. The axillar position of the ovule may indicate that the ovule is a single strobilus and whole flower-like fructification is a compound strobilus. If the affinity to Gnetales is real, the lower whorl of bracts in Eoantha should be homologous to the lower transverse bracts of Gnetales, the ‘gynoecial’ bracts are then equivalent to the median bracts in gnetaceous flower and the upper rosette of bracts is comparable to the upper transverse gnetaceous bracts. The terminal ovule is absent in Eoantha. If this is true, the common ancestor of Gnetales and Eoanthales had a hermaphroditic compound strobilus with alternating whorls of male and female cordaitalean strobili simplified to one ovule and few synangia.
Other putative gnetopsid order Dechellyiales from Triassic (S.R. Ash in Anderson et al. 2007) is clearly different from previous orders. Its male compound voltzialean strobilus and archaeophylls (Fig. 11.29) classify it rather among early conifers. But the female structures are really similar to gnetophytes. The bract, called erroneously megasporophyll, subtended a single ovule – the simplified strobilus.
conclusion
The reformed systematics based on statistical analyses offers many phylogenetic trees as well as systematic categories. I tried to revive the traditional paleobotanical concepts, which could be distorted due to lack of data, but of course, they were certainly closer to the reality then recent systematics. Current view of the evolution of higher plants is more reminiscent of a story where all fossil findings are transitional structures to recent plants, than of science. If we look at current vascular plants, we can clearly recognize horsetails from true ferns and wisk ferns or magnoliids from monocots and eudicots. There are no transitional plant groups. The evolutionary tree does not consist of smooth lines where each point of the line is a transitionary species. a branch between two nodes can be represented by a single morphological change, with many species radiating from a single node in a similar direction. Even if some species are protruding from normal or are of uncertain phylogenetic position, they are a part of the known large plant groups. a knowledge of morphology is sufficient to place these groups, molecular systematics only fine-tunes this placement and corrects the errors caused by the lack of this knowledge. If we were in Mesozoic, the same criterion would apply.
I recognize three large groups of gymnosperm plants. Because of parallelism, they can be sometimes very similar. But the cupulate gymnosperms always had microsporophyll morphologically equivalent to psilophytes even if they lack the cupule. The microsporophylls of acupulate soriferous gymnosperms, even if they form cupule from fused whorls of leaves, are equivalent to microsporophylls of true ferns. The conifers may have megasporophylls reduced to microphylls and these are arranged in strobili. All fossils gymnosperms could be classified into these three groups. I did not find any species that would represent a fourth group. The evolution should be perceived in the terms of founder effect, where an individual is changed by mutation or hybridisation and establishes a new plant lineage. I have described up to now nine morphological

69
groups. Some of them may have arisen from a mutual hybridisation, but they are clearly separated morphologically, and no transient forms are known. All vascular plants tend to form strobili, all seed plants tend to form flowers.
references
Anderson, J. M., Anderson, H. M., 2003: Heyday of the gymnosperms: systematics and biodiversity of the Late Triassic Molteno fructifications. National Botanical Institute: Pretoria.
Anderson, J. M., Anderson, H. M., Cleal, C. J., 2007: Brief history of the gymnosperms: classification, biodiversity, phytogeography and ecology. National Botanical Institute: Pretoria.
Anderson, H. M., Barbacka, M., Bamford, M. K., Holmes, W. B. K., Anderson, J. M., 2019: Dicroidium (foliage) and affiliated wood Part 3 of a reassessment of Gondwana Triassic plant genera and a reclassification of some previously attributed. Alcheringa. An Australasian Journal of Palaeontology, 44 (1): 64-92.
Andrews, H. N., 1941: Dichophyllum moorei and certain associated seeds. Annals of the Missouri Botanical Garden, 28: 375-384.
Archangelsky, S., Cúneo, R., 1990: Polyspermophyllum, a new Permian gymnosperm from Argentina, with considerations about the Dicranophyllales. Review of Palaeobotan and Palynology, 63: 117-135.
Arnold, C. A., 1939: Lagenospermum imparirameum sp. nov., a seedbearing fructification from the Mississippian of Pennsylvania and Virginia. Bulletin of the Torrey Botanical Club, 66 (5): 297-303.
Ash, S. R., 1970: Dinophyton, a problematical new plant genus from the Upper Triassic of the south-western United States. Palaeontology, 13: 646-663.
Beck, C. B., 1960: The Identity of Archaeopteris and Callixylon. Brittonia, 12 (4): 351-368.Becker, A., Saedler, H., Theissen, G., 2003: Distinct MADS-box gene expression patterns in the reproductive
cones of the gymnosperm Gnetum gnemon. Development Genes and Evolution, 213 (11): 567-572.Beltrán, M., Bodnar, J., Melchor, R., 2021: The genera Heidiphyllum and Dordrechtites, and associated fossil
wood from the Triassic Ischichuca Formation, northwestern Argentina. Journal of South American Earth Sciences, 107: 103142.
Benson, M., 1921: The grouping of Vascular plants. The New Phytologist, 20 (2): 82-89.Bergene, J. A., Taylor, E. L., Taylor, T. N., 2017: Dordrechtites arcanus sp. nov., an anatomically preserved
gymnospermous reproductive structure from the Middle Triassic of Antarctica. International Journal of Plant Sciences, 174 (2): 250-265.
Blomenkemper, P., Kerp, H., Hamad, A. A., Bomfleur, B., 2020: Contributions towards whole-plant reconstructions of Dicroidium plants (Umkomasiaceae) from the Permian of Jordan. Review of Palaeobotany and Palynology, 278: 104210.
Bose, M. N., Pal, P. K., Harris, T. M., 1985: The Pentoxylon plant. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 310 (1142): 77-108.
Brousmiche, C., 1982: Sur la synonymie de Crossotheca boulayi Zeiller, 1886–88 et Crossotheca bourozii Danzé, 1956 avec l’espéce-type du genre: Crossotheca crepinii Zeiller, 1883. Geobios, 15 (5): 679-703.
Burnett, G. T., 1835: Outlines of botany. Henry Renshaw: London.Cariglino, B., 2015. New glossopterid polysperms from the Permian La Golondrina Formation (Santa
Cruz Province, Argentina): Potential affinities and biostratigraphic implications. Revista Brasileira de Paleontologia, 18 (3): 379-390.
Carrizo, M., Archangelsky, S., 2013: Kladistamuos golondrinensis nov. gen. et comb., a fertile foliage from the mid to late Permian of Patagonia, Argentina. Review of Palaeobotany and Palynology, 196: 1-8.
Cesari, S. N., 2014: Eonotosperma arrondoi Césari gen. et sp. nov., a pteridosperm from the Early Carboniferous of Argentina. Ameghiniana, 34 (2): 169-178.
Chase, M. W., Reveal, J. L., 2009: a phylogenetic classification of the land plants to accompany APG III. Botanical Journal of the Linnean Society, 161 (2): 122-127.
Christenhusz, M. J. M., Reveal, J. L., Farjon, A., Gardner, M. F., Mill, R. R., Chase, M. W., 2011: a new classification and linear sequence of extant gymnosperms. In Phytotaxa, 19: 55-70.
Church, A. H., 1914: On the Floral Mechanism of Welwitschia mirabilis (Hooker). Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character, 205: 115-151.
Cleal, C. J., Shute, C. H., Hilton, J., Carter, J., 2009: a revision of the Pennsylvanian‐aged Eremopteris ‐bearing seed plant. International Journal of Plant Sciences, 170 (5): 666-698.

70
Conard, H. S., 1919: The classification of vascular plants: a review. In Shreve, F. The Plant World. Plant World Association: Baltimore.
Contreras, D. L., Duijnstee, I. A. P., Ranks, S., Marshall, C. R., Looy, C. V., 2017: Evolution of dispersal strategies in conifers: Functional divergence and convergence in the morphology of diaspores. Perspectives in Plant Ecology, Evolution and Systematics, 24: 93-117.
Cornet, B., 1986: The leaf venation and reproductive structures of a Late Triassic angiosperm, Sanmiguelia lewisii. Evolutionary Theory, 7: 231-309.
Cornet, B., 1996: a New Gnetophyte from the Late Carnian (Late Triassic) of Texas and its bearing on the origin of the angiosperm carpel and stamen. In: Taylor D. W., Hickey L. J. (eds.) Flowering Plant Origin, Evolution & Phylogeny. Springer: Boston.
Coturel, E. P., Césari,S. N., 2017: Revision of Fedekurtzia (pteridosperm) and allied fronds from the Carboniferous of Gondwana. Acta Palaeobotanica, 57 (2): 135-151.
Crane, P. R., 1985: Phylogenetic analysis of seed plants and the origin of angiosperms. Annals of the Missouri Botanical Garden, 72: 716-793.
Crane, P. R., Herendeen, P., 2009: Bennettitales from the Grisethorpe Bed (Middle Jurassic) at Cayton Bay, Yorkshire, UK. American Journal of Botany, 96 (1): 284-295.
Cronquist, A., 1960: The divisions and classes of plants. The Botanical Review, 26 (4): 425-482. Cronquist, A., Takhtajan, A., Zimmermann, W., 1966: On the higher taxa of Embryobionta. Taxon, 15 (4):
129-134. De Haan, H. R. M., 1920: Contribution to the knowledge of the morphological value and the phylogeny of the
ovule and its integ-uments. Recueil des Travaux des botaniques Néerlandais 17: 219-324.De Jussieu, A. L., 1774: Genera plantarum. Herissant: Paris.Delevoryas, T., Gould, R. E., 1971: An unusual fossil fructification from the Jurassic of Oaxaca, Mexico.
American Journal of Botany, 58 (7): 616-620.Donoghue, M. J., Doyle, J. A., 2000: Seed plant phylogeny: Demise of the anthophyte hypothesis? Current
Biology, 10: 106-109.Dörken, V. M., Nimsch, H., 2016: Some new aspects about the evolution of pollen cones and perisporangiate
microsporangiophores in Taxaceae. Bulletin of the Cupressus Conservation Project, 5 (1): 3-21.Dörken, V. M., Zhang, Z. X., Mundry, I. B., Stützel, T., 2011: Morphology and anatomy of male cones of
Pseudotaxus chienii (W.C. Cheng) W.C. Cheng (Taxaceae). Flora, 206: 444-450.Dörken, V. M., Nimsch, H., Rudall, P. J., 2019: Origin of the Taxaceae aril: evolutionary implications of
seed-cone teratologies in Pseudotaxus chienii, Annals of Botany, 123 (1): 133-143.Douglas, A. W., Stevenson, D. W., Little, D. P., 2007: Ovule development in Ginkgo biloba L., with emphasis
on the collar and nucellus. International Journal of Plant Sciences, 168 (9): 1207-1236.Doweld, A. B., 1998: On the morphological nature of the cupules of some Upper Devonian/Lower Carboniferous
seed plants. Geophytology, 26 (2): 1-12. Doweld, A. B., 2001: Prosyllabus Tracheophytorum. Geos: Moskva.Doyle, J. A., Donoghue, M. J., 1986: Seed plant phylogeny and the origin of angiosperms: An experimental
cladistic approach. Botanical Review 52: 321-431.Elgorriaga, A., Escapa, I. H., Cunéo, R., 2019a: Relictual Lepidopteris (Peltaspermales) from the Early
Jurassic Cañadón Asfalto Formation, Patagonia, Argentina. nternational Journal of Plant Sciences 180 (6): 578-596.
Elgorriaga, A., Escapa, I. H., Cunéo, R., 2019b: Southern Hemisphere Caytoniales: vegetative and reproductive remains from the Lonco Trapial Formation (Lower Jurassic), Patagonia. Journal of Systematic Palaeontology, 17 (17): 1-19.
Endress, P. K., 1996: Structure and function of female and bisexual organ complexes in Gnetales. International Journal of Plant Sciences, 157 (6), Supplement: Biology and Evolution of the Gnetales: 113-125.
Escapa, I. H., Taylor, E. L., Cuneo, R., Bomfleur, B., Bergene, J., Serbet, R., Taylor, T. N., 2011: Triassic floras of Antarctica: Plant diversity and distribution in high paleolatitude communities. Palaios, 26 (9): 522-544.
Fischer, T. C., Meller, B., Kustatscher, E., Butzmann, R., 2010. Permian ginkgophyte fossils from the Dolomites resemble extant O-ha-tsuki aberrant leaf-like fructifications of Ginkgo biloba L. BMC Evolutionary Biology 10, 337
Florin, R. 1949: The Morphology of Trichopitys heteromorpha Saporta, a seed plant of Palaeozoic age, and the evolution of the female flowers in the Ginkgoinae. Acta Horti Bergiani, 15 (5): 79-109.
Frenguelli, J., 1946: El origen de las angiospermas. Boletin de la Sociedad Argentina de Botánica, 1 (3): 169-208.

71
Friis, E. M., Crane, P. R., Pedersen, K. R., Bengston, S., Donoghue, P. C. J., Grimm, G. W., Stampanoni, M., 2007: Phase contrast X‐ray microtomography links Cretaceous seeds with Gnetales and Bennettitales. Nature, 450: 549-552.
Fujii, K., 1896: On the different views hitherto proposed regarding the morphology of the flowers of Ginkgo biloba L. Shokubutsugaku Zasshi, 10 (118): 104-110.
Galtier, J., Béthoux, O., 2002: Morphology and growth habit of Dicksonites pluckenetii from the Upper Carboniferous of Graissessac (France). Geobios, 35 (5): 525-535.
Gordenko, N. V., Broushkin, A. V., 2013: Ginkgoales: Some problems of systematics and phylogeny. Paleontological Journal, 49 (5): 546-551.
Gordenko, N. V., 2010: Vladimariales ordo nov. (Gymnospermae) from the Middle Jurassic deposits of the Mikhailovskii Rudnik Locality (Kursk Region, European Russia). Paleontological Journal, 44 (10): 1281-1307.
Grauvogel-Stamm, L., 1978: La flore du Grès à Voltzia (Buntsandstein supérieur) des Vosges du nord (France). Morphologie, anatomie, interprétations phylogénique et paléogéographique. Institut de Géologie – Université Louis-Pasteur: Strasbourg.
Harris, T. M., 1951: The fructification of Czekanowskia and its allies. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 235 (628): 483-508.
Harris, T. M., 1962: The occurrence of the fructification Carnoconites in New Zealand. Transactions of the Royal Society of New Zealand, Geology, 1: 17-27.
Herendeen, P.S., Friis, E.M., Pedersen, K.R., Crane, P.R., 2017: Palaeobotanical Redux: Revisiting the Age of the Angiosperms. Nature Plants, 3: 17015.
Herman, A., Spicer, R. A., 2010: Mid-Cretaceous floras and climate of the Russian high Arctic (Novosibirsk Islands, Northern Yakutiya). Palaeogeography Palaeoclimatology Palaeoecology, 295 (3): 409-122.
Herr, J. M., 1995: The origin of the ovule. American Journal of Botany, 82(4): 547-564.Herrera, F., Shi, G., Ichinnorov, N., Takahashi, M., Bugdaeva, E. V., Herendeen, P. S., Crane, P. R., 2017:
The presumed ginkgophyte Umaltolepis has seed-bearing structures resembling those of Peltaspermales and Umkomasiales. PNAS, 114 (12): E2385-E2391.
Hetherington, A. J., 2020: New views on old seeds: a new description of Genomosperma sheds light on early seed evolution. New Phytologist, 229 (3): 1189-1191.
Hrabovský, M., 2020: Leaf evolution and classification. 1. Lycopodiopsida. Acta Botanica Universitatis Comenianae, 55: 19-37.
Ignatiev, I. A., Meyen, S. V., 1989: Suchoviella – Gen. nov. from the permian of angaraland and a review of the systematics of Cordaitanthales. Review of Palaeobotany and Palynology, 57 (3-4): 313-339.
Ignatiev, I. A., Moseychik, Y. B., 2015. K nadrodovoy sistematike pteridospermov Karbona Angaridy. Paleobotanicheskiy vremennik, supplement of Lethaea rossica, 2: 191–201.
Karstens, W. K. H., 1945: Variability of the female reproductive organs in Ginkgo biloba L. Blumea, 5: 532-553
Kimura, T., Ohana, T., 1978: Czekanowskia nipponica sp. nov. from the Upper Cretaceous Omichidani Formation, Ishikawa Prefecture in the Inner Zone of Central Japan. Proceedings of the Japan Academy. Ser. B: Physical and Biological Sciences, 54 (10): 595-600.
Kirchner, M., van Konijnenburg-van Cittert, J. H. A., 1994: Schmeissneria microstachys (Prel, 1833) Kirchner et Van Konijnenburg-Van Cittert, comb. nov. and Karkenia haupymannii Kirchner et Van Konijnenburg-Van Cittert, sp. nov., plants with ginkgoalean affinities from the Liassic of Germany. Review of Palaeobotany and Palynology, 83 (1-3): 199-215.
Klotz, G. 1985: Zur Typologie des Blattes. Flora, 176: 189-196.Krassilov, V. A., Ash, S. R., 1988: On Dinophyton – protognetalean Mesozoic plant. Palaeontographica
Abteilung B, 208 (1-3): 33-38.Krassilov, V. A., Bugdaeva, E. V., 1988: Gnetalean plants from the jurassic of Ust-Balej, East Siberia. Review
of Palaeobotany and Palynology, 53 (3-4): 359-376.Krassilov, V. A., 1975: Dirhopalostachyaceae - a new family of proangiosperms and its bearing on the problem
of angiosperm ancestry. Palaeontographica Abteilung В, 153 (1-3): 100-110.Krassilov, V. A., 1986: New floral structures from the Lower Cretaceous of Lake Baikal area. Review of
palaeobotany and palynology, 47: 9-16.Krassilov, V. A., 1989: Obshchie printsipy evoljutsionnoj sistematiki i sistema semennykh rastenij. In Kruchinina
N. V., Modzalevskaja T. L. Filogeneticheskie aspekty paleontologii. Vsesoyuznoe Paleontologicheskoe Obshchestvo, Akademiya Nauk SSSR: Leningrad.

72
Kustatscher, E., van Konijnenburg - van Cittert, J. H. A., Bauer, K.; Krings, M., 2016: Strobilus organization in the enigmatic gymnosperm Bernettia inopinata from the Jurassic of Germany. Review of Palaeobotany and Palynology, 232: 151-161.
Kvaček, J., Pacltová, B., 2001: Bayeritheca hughesii gen. et sp. nov., a new Eucommiidites-bearing pollen organ from the Cenomanian of Bohemia. Cretaceous Research, 22 (6): 695-704.
Lignier, O., Tison, A., 1912: Les Gnétales leurs fleurs et leur position systématique. Annales des Sciences Naturelles. Botanique, 9 (16): 55-185.
Link, H. F., 1824: Elementa philosophiae botanicae. Haude & Spener: Berlin.Liu, L., Wang, D.-M., Meng, M., Xue, J.-Z., 2017: Further study of Late Devonian seed plant Cosmosperma
polyloba: its reconstruction and evolutionary significance. BMC Evolutionary Biology, 17: 149.Lo, H.-C., Wang, Y.-F., 2001: Development of ovulate cones from initiation of reproductive buds to
fertilization in Cephalotaxus wilsoniana Hay. Proceedings of the National Science Council, Republic of China. Part B, Life sciences, 25 (2): 97-118.
Long, A. G., 1944: On the prothallus of Lagenstoma ovoides Will. Annals of Botany, 8: 105-120.Long, A. G., 1965: VI. – On the Cupule Structure of Eurystoma angulare. Transactions of the Royal Society
of Edinburgh, 66 (6): 111-128. Mamay, S. H., 1976: Vojnovskyales in the Lower Permian of North America. The Paleobotanist, 25: 290-297.Martens, P., 1961: Structure et ontogenèse du cône et de la fleur mâles de Welwitschia mirabilis Hooker.
Cellule, 62: 5-91.Mcloughlin, S., Prevec, R., 2019: The architecture of Permian glossopterid ovuliferous reproductive organs,
Alcheringa: An Australasian Journal of Palaeontology, 43 (4): 480-510.McLoughlin, S., Pott, C., Sobbe, I. H., 2018: The diversity of Australian Mesozoic bennettitopsid reproductive
organs. Palaeobiodiversity and Palaeoenvironments, 98: 71-95.Meyen, S. V., 1969: Pteridospermy verkhnego paleozoya i mezozoya. Nauka: Moskva.Meyen, S. V., 1984: Basic features of gymnosperm systematics and phylogeny as evidenced by the fossil
record. The Botanical Review, 50 (1): 1-111.Meyen, S. V., 1987: Osnovy paleobotaniki (Fundamentals of paleobotany). Nedra:Moskva.Mill, R. R., Möller, M., Glidewell, S. M., Masson, D., Williamson, B., 2004: Comparative anatomy and
morphology of fertile complexes of Prumnopitys and Afrocarpus species (Podocarpaceae) as revealed by histology and NMR imaging, and their relevance to systematics. Botanical Journal of the Linnean Society, 145 (3): 295-316.
Miller, C. N., 1985: a critical review of S. V. Meyen’s “Basic features of gymnosperm systematics and phylogeny as evidenced by the fossil record” The Botanical Review, 51: 295-318
Moisan, P., Voigt, S., Pott, C., Buchwitz, M., Schneider, J. W., & Kerp, H., 2011: Cycadalean and bennettitalean foliage from the Triassic Madygen Lagerstätte (SW Kyrgyzstan, Central Asia). Review of Palaeobotany and Palynology, 164 (1-2): 93-108.
Mundry, M., Stützel, T., 2004a: Morphogenesis of leaves and cones of male short-shoots of Ginkgo biloba L. Flora - Morphology, Distribution, Functional Ecology of Plants, 199 (5): 437-452.
Mundry, M., Stützel, T., 2004b: Morphogenesis of the reproductive shoots of Welwitschia mirabilis and Ephedra distachya (Gnetales), and its evolutionary implications. Organisms Diversity & Evolution, 4 (1-2): 91-108.
Mundry, M., Stützel, T., 2008: Morphogenesis of male sporangiophores of Zamia amblyphyllidia D. W. Stev. Plant Biology, 5 (3): 297-310.
Naugolnykh, S. V., 2007: Foliar seed-bearing organs of Paleozoic ginkgophytes and the early evolution of the Ginkgoales. Paleontological Journal, 41 (8): 815-859.
Naugolnykh, S. V., 2012a: a new Carboniferous pteridosperm of Angaraland: Angaranthus victorii Naugolnykh, gen. et spec. nov. (Angaranthaceae, fam. nov., Callistophytales). Wulfenia, 19: 39-55.
Naugolnykh, S. V., 2012b: a new species of Psygmophyllum Schimper from the Lower Permian of the Urals. Paleontological Journal, 46 (2): 208-218.
Němejc, F., 1950: The natural systematic of plants in the light of the present palaeontological documents. Sborník Národního Muzea v Praze, 6 (3): 1-84.
Němejc, F., 1963: Paleobotanika 2. ČSAV, Praha.Němejc, F., 1968: Paleobotanika 3. ČSAV, Praha.Novák, F. A., 1954: Taxonomie telomofyt. Státní pedagogické nakladatelství: Praha.Novák, F. A., 1961: Vyšší rostliny. Nakladatelství Československé Akademie Věd: Praha.Osborn, J. M., Taylor, T. N., Crane, P. R., 1991: The Ultrastructure of Sahnia Pollen (Pentoxylales).
American Journal of Botany, 78 (11): 1560-1569.

73
Pacyna, G., Zdebska, D., 2010: Upper Carboniferous seed fern (Pteridospermophyta) pollen organs from Silesia (Poland) and related evolution considerations. Polish Botanical Society, 100: 1-81.
Patterson, C., 1982: Morphological characters and homology. In Joysey, K. A., Friday, A. E. (eds.) Problems of phylogenetic reconstruction. Academic Press, London, pp. 21-74.
Pax, F., 1894: Prantl‘s Lehrbuch Der Botanik. Engelmann: Leipzig.Popa, M. E., 2019: Review of the bennettitalean genus Weltrichia. Journal of Palaeogeography, 8: 12.Pott, C., 2016: Westersheimia pramelreuthensis from the Carnian (Upper Triassic) of Lunz, Austria: More
evidence for a unitegmic seed coat in early Bennettitales. International Journal of Plant Sciences, 177: 771-791.
Pott, C., Fischer, T., Aschauer, B., 2017: Lunzia austriaca – a bennettitalean microsporangiate structure with Cycadopites -like in situ pollen from the Carnian (Upper Triassic) of Lunz, Austria. Grana, 56 (5): 321-338.
Prevec, R., 2011: a structural re-interpretation and revision of the type material of the glossopterid ovuliferous fruitification Scutum from South Africa. Palaeontologica Africana, 46: 1-19.
Raymond, A., McCarty, R., 2009: Pachytesta crenulata, a new medullosan ovule from the Pennsylvanian of Iowa. Review of Palaeobotany and Palynology, 156 (3-4): 283-306.
Read, C. B., 1933: a new Trichopitys from the Carboniferous of Colorado. Journal of the Washington Academy of Sciences, 23 (10): 461-463.
Rothwell, G. W., Stockey, R. A., 2002: Anatomically preserved Cycadeoidea (Cycadeoidaceae), with a reevaluation of systematic characters for the seed cones of Bennettitales. American Journal of Botany, 89 (9): 1447-1458.
Rothwell, G. W., 1982: Cordaianthus duquesnensis sp. nov., anatomically preserved ovulate cones from the Upper Pennsylvanian of Ohio. American Journal of Botany, 69: 239–247.
Rothwell, G. W., Scheckler, S. E., Gillespie, W. H., 1989: Elkinsia gen. nov., a Late Devonian gymnosperm with cupulate ovules. Botanical Gazette, 150 (2): 170-189.
Rothwell, G. W., Crepet, W. L., Stockey, R. A., 2009: Is the anthophyte hypothesis alive and well? New evidence from the reproductive structures of Bennettitales. American Journal of Botany, 96 (1): 296-322.
Rudall, P. J., Hilton, J., Vergara-Silva, F., Bateman, R. M., 2011: Recurrent abnormalities in conifer cones and the evolutionary origins of flower-like structures. Trends in Plant Science, 16 (3): 151-159.
Sakisaka M., 1927: On the morphological significance of seed-bearing leaves of Ginkgo (with Plate X). Shokubutsugaku Zasshi, 41 (483): 273-278.
Schwarz, E. H. L., 1912: South African Geology. Blackie & Son: Glasgow.Serbet, R., Hageman, S., Hoffman, B. L, Taylor, T. N, Taylor, E. L., 2016: a new Pennsylvanian pollen organ
from northwestern Missouri with affinities in the Lyginopteridales. Review of palaeobotany and palynology, 234: 136-146.
Seward, A. C., 1917: Fossil plants: for students of botany and geology 3. University Press: Cambridge.Sharma, B. D., 1974: Ovule ontogeny in Williamnsonia Carr. Palaeontographica B, 148: 137-143. Shi, G., Leslie, A. B., Herendeen, P. S., Herrera, F., Ichinnorov, N., Takahashi, M., Knopf, P., Crane,
P. R., 2016: Early Cretaceous Umkomasia from Mongolia: implications for homology of corystosperm cupules. New Phytologist, 210: 1418-1429.
Skog, J. E., Gensel, P. G., 1980: a Fertile Species of Triphyllopteris from the Early Carboniferous (Mississippian) of Southwestern Virginia. American Journal of Botany, 67 (4): 440-451.
Srivastava, S. C., Banerji, J., 2001: Pentoxylon plant: a reconstruction and interpretation. Plant cell biology and development, 13: 11-18.
Srivastava, R. K., 1952: Abnormal Leaves of Cycas revoluta Thunb. Nature, 170: 253. Stewart, W. N., Rothwell, G. W., 1993: Paleobotany and the evolution of plants. Cambridge University Press,
Cambridge.Stidd, B. M., 1981: The current status of medullosan seed ferns. Review of Palaeobotany and Palynology, 32
(1): 63-101. Stockey, R. A., Rothwell, G. W., 2003: Anatomically preserved Williamsonia (Williamsoniaceae): Evidence
for bennettitalean reproduction in the Late Cretaceous of western North America. International Journal of Plant Sciences, 164: 251-262.
Stützel, T., Röwekamp, I., 1999: Female reproductive structures in Taxales. Flora, 194 (2): 145-157.Suthar, O. P., Sharma, B. D., 1989: a new interpretation on the structure of Sahnia nipaniensis Mttre from the
Rajmahal Hills. Palaeobotanist, 37 (1): 90-93.Tahktajan, A. L., 1956: Vysshie rasteniya 1. Izd-vo Akademii nauk SSSR: Leningrad.Taylor, T. N., Millay, M. A., 1979: Pollination biology and reproduction in early seed plants. Review of
Palaeobotany and Palynology, 27: 329-355.

74
Taylor, E., Taylor, T., Krings, M., 2009: Paleobotany. Academic Press, Amsterdam.Tomlinson, P. B., 1992: Aspects of Cone Morphology and Development in Podocarpaceae (Coniferales).
International Journal of Plant Sciences, 153 (4): 572-588.Tomlinson, P. B., Takaso, T., Rattenbury, J. A., 1989: Cone and ovule ontogeny in Phyllocladus
(Podocarpaceae). Botanical Journal of the Linnean Society, 99 (3): 209-221.Van Konijnenburg-van Cittert, J. H. A., 1992: An enigmatic Liassic microsporophyll, yielding Ephedripites
pollen. Review of Palaeobotany and Palynology, 71 (1-4): 239-254.Van Konijnenburg-van Cittert, J. H. A., 2010: The Early Jurassic male ginkgoalean inflorescence
Stachyopitys preslii Schenk and its in situ pollen. Scripta Geologica Special Issue, 7: 141-149.Vega, J. C., Archangelsky, S., 1996: Austrocalyx jejenensis Vega and Archangelsky, gen. et sp. nov., a cupulate
rhacopteroid pteridosperm from the Carboniferous of Argentina. Review of Palaeobotany and Palynology, 91: 107-119.
Wang, J., Pfefferkorn, H. W., 2009: Nystroemiaceae, a new family of Permian gymnosperms from China with an unusual combination of features. Proceedings of the Royal Society B: Biological Sciences, 277 (1679): 301-309.
Wang, X., 2010: Schmeissneria: An angiosperm from the Early Jurassic. Journal of Systematics and Evolution, 48 (5): 326-335.
Wang, X., 2017: a biased, misleading review on early angiosperms. Natural Science, 9 (12): 399-405.Wang, X., Duan, S., Geng, B., Cui, J., & Yang, Y., 2007: Schmeissneria: a missing link to angiosperms?
BMC Evolutionary Biology, 7(1): 14.Wang, D., Liu, L., Meng, M., Xue, J., Liu, T., Guo, Y., 2014: Cosmosperma polyloba gen. et sp. nov., a seed
plant from the Upper Devonian of South China. Naturwissenschaften, 101 (8): 615-622.Wang, D. M., Meng, M. C., Guo, Y., 2016: Pollen organ Telangiopsis sp. of Late Devonian seed plant and
associated vegetative frond. PLoS One, 11 (1): e0147984.Wettstein, R., 1899: Die weibliche Blüte von Ginkgo. Österreichische Botanische Zeitschrift, 49 (12): 417-
425.Wettstein, R., 1903: Handbuch der Systematischen Botanik 2. Franz Deuticke: Leipzig.Wiggers, A., 1840: Grundriss der Pharmacognosie. Bandenhoeck und Ruprecht: Göttingen.Worsdell, W. C., 1904: The structure and morphology of the ovule: an historical sketch. Annals of Botany,
18: 57-86.Yang, Y., 2004: Ontogeny of triovulate cones of Ephedra intermedia and origin of the outer envelope of ovules
of Ephedraceae. American Journal of Botany, 91 (3): 361-368.Yang, Y., Lin, L., Ferguson, D. K., Wang, Y., 2018: Macrofossil evidence unveiling evolution of male cones
in Ephedraceae (Gnetidae). BMC Evolutionary Biology, 18(1): 125.Zabinkova, N., 1964: Names of taxa above the rank of order. Taxon, 13 (5): 157-160.Zhou, Z., 1997: Mesozoic ginkgoalean megafossils: a Systematic Review. In Hori, T., Ridge, R. W., Tulecke,
W., Del Tredici, P., Trémouillaux-Guiller, J., Tobe, H. Ginkgo biloba - a global treasure. Springer-Verlag: Tokyo.
Zimmermann, W., 1930: Die Phylogenie der Pflanzen. Fischer: Jena.
Abstrakt
Predchádzajúca teória o pôvode mikrofylových a megafylových listov sa môže aplikovať aj na semenné rastliny. Existencia mikrofylu archeofylového typu prináša novú perspektívu na pôvod vajíčka. Nahosemenné rastliny sú klasifikované do rôznych fylogenetických systémov. Tento príspevok prináša nové morfologické pohľady na tieto klasifikácie a rozoznáva tri skupiny nahosemenných rastlín – primárne čiaškovú líniu, primárne bezčiaškovú líniu a strobilovú mirkofylovú líniu. Každá z týchto troch skupín sa mohla vyvíjať rozličnými smermi, preto mnohé morfologické podobnosti medzi nimi viedli k toľkým fylogenetickým interpretáciám. Teória kvetorastov (Antophyta) sa na základe nových morfologických interpretácií javí byť polyfyletická, lebo všetky typy strobilov nahosemenných rastlín mohli viesť k tvorbe kvetov.
Kľúčové slová: pôvod vajíčka, ihličnany, semenné paprade

75
Acta Botanica Universitatis Comenianae Vol. 57, 2021
ruderal vegetation of tHe class ArtemisieteA vulgAris in Bratislava city 2. alliance Convolvulo Arvensis-
Agropyrion repentis ruderálna vegetácia triedy ArtemisieteA vulgAris v Bratislave
2. zväz Convolvulo Arvensis-Agropyrion repentis
Alena Rendeková*, Eva Zahradníková, Ján Miškovic, Martina Michalová, Karol Mičieta
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 81102 Bratislava, Slovakia
Received 7 April 2021; Received in revised form 10 May 2021; Accepted 21 May 2021
Abstract
This paper describes the ruderal vegetation from the alliance Convolvulo arvensis-Agropyrion repentis Görs 1966 (class Artemisietea vulgaris Lohmeyer et al. ex von Rochow 1951) in Bratislava. This paper represents a continuation of the previous one, which was focused on the vegetation of the alliance Dauco-Melilotion Görs 1966. We present our own phytosociological relevés of the recorded associations and communities and refer about their species composition and distribution in Bratislava. We also describe the comparison with the data published from other parts of Slovakia. We recorded 35 relevés from four associations and one community from the alliance, including one relatively rare association Asparago-Chondrilletum juncei Passarge 1978.
Key words: man-affected habitats, phytosociology, southwestern Slovakia, (sub)xerophilous synanthropic plant communities
introduction
Ruderal vegetation is one of the most important elements of inhabited areas. Vegetation in cities and other urban areas provides many beneficial functions such as improvement of the microclimate, uptake of contaminants or noise reduction (Sukopp 2002). It can serve as refuge area for endangered species of plants and invertebrates (Forman 2014, Májeková et al. 2014). However, there is also a negative aspect of ruderal plant communities: they often contain high number of invasive species, which can present a risk for the maintenance of the biodiversity (Mack et al. 2000, Pyšek et al. 2002, Pyšek, Richardson 2010, Medvecká et al. 2012, Pyšek et al. 2012). Therefore, for environmental and many other reasons, ruderal vegetation has been researched by many scientists (e.g. Krippelová, Mucina 1988, Mucina et al. 1993, Jarolímek et al. 1997, Chytrý 2009).
The ruderal plant communities of Bratislava were studied by Jarolímek, Valachovič
* Corresponding author: [email protected]

76
(2016), and Valachovič (2016). A recent comprehensive research was carried out by Rendeková and some parts of its results have already been published (Rendeková 2016, Rendeková, Mičieta 2017, Rendeková et al. 2014, 2015, 2017, 2018, 2019 a, b, 2020), but some parts are still being prepared for publishing.
This paper represents a continuation of the previous one published by Rendeková et al. (2020), which brought the results of the study of the alliance Dauco-Melilotion (class Artemisietea vulgaris). This paper is focused on the alliance Convolvulo arvensis-Agropyrion repentis – another alliance from the class Artemisietea vulgaris, which is relatively abundant in the ruderal areas of Slovakia (Jarolímek et al. 1997).
We aimed to record the ruderal plant communities of the alliance Convolvulo arvensis-Agropyrion repentis in the area of Bratislava, describe their species composition and compare it to other areas of Slovakia. The alliance Convolvulo arvensis-Agropyrion repentis includes ruderal and semiruderal grassland and herbland vegetation distributed in the nemoral and subboreal zones of Europe (Jarolímek et al. 1997, Mucina et al. 2016).
material and methods
The study area lies within the borders of the Bratislava city, the capital of Slovakia, located in southwestern Slovakia, Central Europe. The city covers an area of approximately 367.9 km2 and the population is approximately 425,500 people. The climate has moderate to warm continental character. Bratislava is situated in one of the warmest and driest parts of Slovakia. The natural soils in ruderal areas of the city are covered by anthropogenic ones (Feráková, Jarolímek 2011).
Phytosociological research was done in accordance with the methods of Zürich-Montpelliere school (Braun-Blanquet 1964) during the vegetation seasons of the years 2011–2013. The modified Braun-Blanquet cover-abundance scale, extended by 2a, 2b and 2m values was used in phytosociological relevés (Barkman et al. 1964). The locations of recorded relevés are presented in the Fig. 1. The relevés were imported into TURBOWIN database (Hennekens, Schaminée 2001), and edited in the JUICE programme (Tichý 2002).
Relevés were assigned to associations by numerical classification (cluster analysis), which was performed in the SYN-TAX 2000 programme (Podani 2001) and according to the presence of diagnostic species in relevés. The detailed desciption of the analysis and parameters used is presented in the previous paper of Rendeková et al. (2020).
The relevés are presented in a phytosociological table (Tab. 1), which was created in the JUICE programme (Tichý 2002). Relevés in the table are arranged according to the results of numerical classification. Constancy classes are listed in the last column of each association or community recorded by more than two relevés. Taxa are ordered into groups according to their syntaxonomical relations. Diagnostic, characteristic and constant taxa were determined according to the publications of Jarolímek et al. (1997) and Jarolímek, Šibík (2008). Within the groups, taxa are ordered according to the decreasing frequency. Cover-abundance values 2a and 2b are presented in abbreviated forms a and b.
The nomenclature of the taxa follows Marhold, Hindák (1998), and the nomenclature of the syntaxa follows Jarolímek, Šibík (2008). Taxonomically problematic taxa were included in aggregates, namely Achillea millefolium agg. and Rosa canina agg.
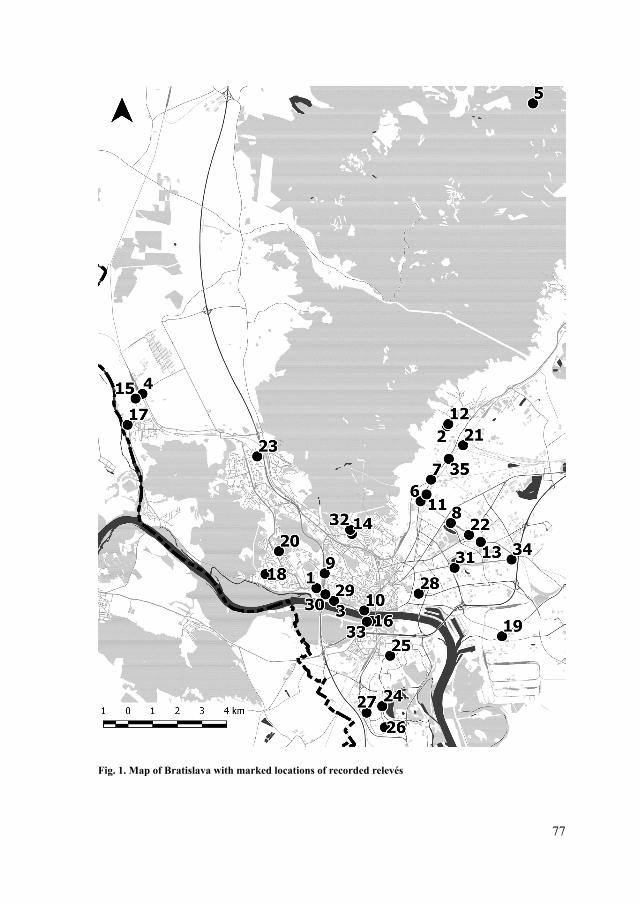
77
Fig. 1. Map of Bratislava with marked locations of recorded relevés

78
results and discussion
Since the results of the numerical classification were already published in the previous paper of Rendeková et al. (2020), here we focuse only on the description of the recorded communities from the alliance Convolvulo arvensis-Agropyrion repentis, which was missing.
The list of recorded communities and their description:
Class Artemisietea vulgaris Lohmeyer et al. ex von Rochow 1951Ordo Agropyretalia repentis Görs 1966
Alliance Convolvulo arvensis-Agropyrion repentis Görs 1966Association Asparago-Chondrilletum juncei Passarge 1978Association Convolvulo-Brometum inermis Eliáš 1979 Association Falcario vulgaris-Agropyretum repentis Th. Müller et Görs 1969 Association Lepidio drabae-Agropyretum repentis Th. Müller et Görs 1969 Elytrigia repens community [Convolvulo-Agropyrion]
We recorded four ruderal associations and one ruderal community from the alliance Convolvulo arvensis-Agropyrion repentis in Bratislava. Associations Convolvulo-Brometum inermis, Falcario vulgaris-Agropyretum repentis (Fig. 2), Lepidio drabae-Agropyretum repentis, and Elytrigia repens community [Convolvulo arvensis-Agropyrion repentis] were found in many ruderal areas. They occupied predominantly the edges of the pathways, roads, and railway tracks, as well as various abandoned areas. The association Asparago-Chondrilletum juncei was only found in one locality – an abandoned sunny ruderal area near the reailway track in the Rača district. This association is relatively rare in Slovakia (Jarolímek et al. 1997), but it may be possible that some stands in Bratislava could be overlooked during our research. Another relatively interesting recorded association is Convolvulo-Brometum inermis, which is documented only from a few other areas in Slovakia by relevés (Jarolímek et al. 1997).
The species Bromus inermis, Cardaria draba, Chondrilla juncea, Elytrigia repens, and Falcaria vulgaris (Fig. 3) dominated in the recorded communities from the alliance Convolvulo arvensis-Agropyrion repentis (Tab. 1). Most of the dominant taxa belong to the diagnostic, characteristic and constant species of the alliance. Another diagnostic, characteristic and dominant taxa of the alliance Convolvulo arvensis-Agropyrion repentis (Achillea millefolium agg., Artemisia vulgaris, Convolvulus arvensis) and many of the class Artemisietea vulgaris and its other lower syntaxa (e.g. Ballota nigra, Medicago sativa, Silene latifolia subsp. alba, Stenactis annua) also occured in the recorded stands (Tab. 1). Molinio-Arrhenatheretea was another class with abundant diagnostic and characteristic taxa. It was represented by taxa such as Arrhenatherum elatius, Dactylis glomerata, Plantago lanceolata, Poa pratensis, and Taraxacum sect. Ruderalia (Tab. 1).
Many grass species (e.g. Arrhenatherum elatius, Bromus inermis, Dactylis glomerata, Elytrigia repens, and Poa pratensis) typically occured in the recorded stands. In many relevés, grasses Arrhenatherum elatius, Bromus inermis and Elytrigia repens reached not only high frequency but also a relatively high cover-abundance values. In the association

79
Lepidio drabae-Agropyretum repentis also grass Poa bulbosa was frequent. The species Convolvulus arvensis also occured frequently and with a relatively high cover-abundance values in almost all recorded associations and in the recorded community (Tab. 1). This species is constant species of the alliance Convolvulo arvensis-Agropyrion repentis.
Ruderal communities from the alliance Convolvulo arvensis-Agropyrion repentis were also recorded in Bratislava forty years ago by Jarolímek (1983). The species composition of the communities which we recorded more recently is similar to those of the stands recorded in the past, but in the more recent relevés, the neophytes Medicago sativa and Stenactis annua occur more frequently. The invasive species Stenactis annua is also slightly more frequent in our relevés than in the relevés from other parts of Slovakia (Jarolímek et al. 1997). The thermophilic species Ballota nigra and Clematis vitalba are slightly more frequent; the rest of the floristic composition is similar.
In comparison to other parts of Slovakia, we only found more differences in the species composition of the stands dominated by Cardaria draba. Slightly similar stands were recorded in Malé Karpaty Mts. by Jarolímek (1986), but the author classified these relevés as the association Lepidietum drabae Timár 1950. These relevés were dominated by Cardaria draba, but the rest of the species composition differed in comparison to our relevés from Bratislava; the relevés recorded by Jarolímek contained higher number of diagnostic species of the classes Polygono arenatri-Poetea annuae and Stellarietea mediae. Stands with Cardaria draba as dominat species were classified as Lepidietum drabae Timár 1950 also by Zaliberová (1982) and Kopecký (1982). Stands recorded by them are also different from ones which we recorded in Bratislava by a higher representation of diagnostic taxa of the classes Polygono arenatri-Poetea annuae and Stellarietea mediae. On the contrary, our relevés contain some taxa from the class Artemisietea vulgaris (e.g. Artemisia vulgaris, Elytrigia repens), which are not present in the stands recorded by Zaliberová. In the relevés recorded by Kopecký in Praha, diagnostic species of the class Stellarietea mediae also pravail over the species from the class Artemisietea vulgaris. In our relevés from Bratislava, both classes are represented, but species from Artemisietea vulgaris reach higher cover-abundance values (Tab. 1), which confirms the classification of these relevés as the relevés from the association Lepidio drabae-Agropyretum repentis Th. Müller et Görs 1969.
Our results deepen the knowledge of ruderal plant communities of Bratislava. Since the invasive neophyte Stenactis annua occurs more frequently in Bratislava than in other parts of Slovakia, the results also contribute to the knowledge of its distribution in Slovakian ruderal vegetation.

80
Fig. 2. Stand of the association Falcario vulgaris-Agropyretum repentis in the ruderal area in Bratislava (Vrakuňa district). Author of photo: A.Rendeková
Fig. 3. Falcaria vulgaris – the dominant species in some ruderal stands from the ordo Convolvulo arvensis-Agropyrion repentis recorded in Bratislava. Author of photo: A.Rendeková
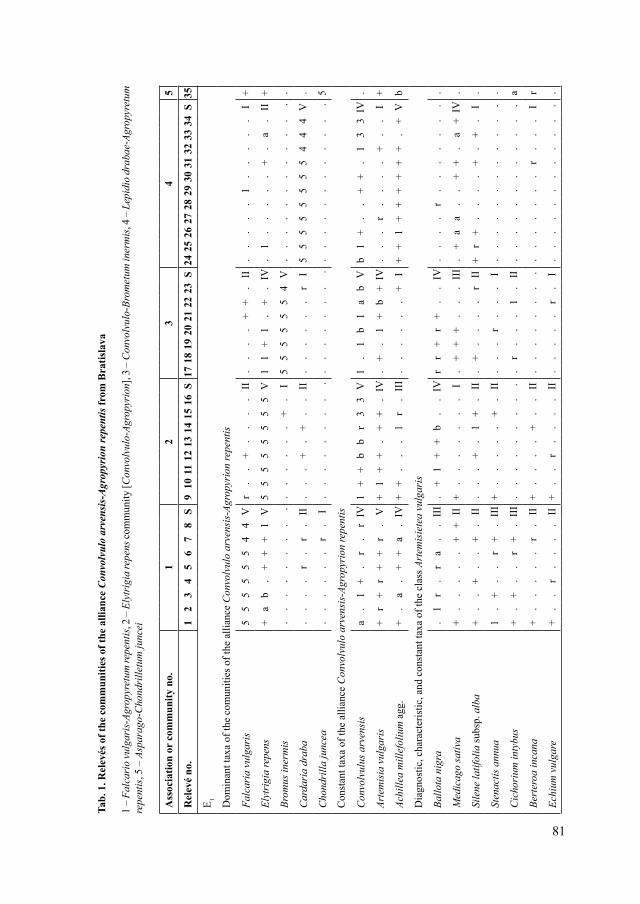
81
Tab.
1. R
elev
és o
f the
com
mun
ities
of t
he a
llian
ce C
onvo
lvul
o ar
vens
is-A
grop
yrio
n re
pent
is fr
om B
ratis
lava
1 –
Falc
ario
vulg
aris
-Agr
opyr
etum
repe
ntis
, 2 –
Ely
trig
ia re
pens
com
mun
ity [C
onvo
lvul
o-Ag
ropy
rion
], 3
– C
onvo
lvul
o-Br
omet
um in
erm
is, 4
– L
epid
io d
raba
e-Ag
ropy
retu
m
repe
ntis
, 5 –
Asp
arag
o-C
hond
rille
tum
junc
ei
Ass
ocia
tion
or c
omm
unity
no.
12
34
5
Rel
evé
no.
12
34
56
78
S9
1011
1213
1415
16S
1718
1920
2122
23S
2425
2627
2829
3031
3233
34S
35
E 1 Dom
inan
t tax
a of
the
com
uniti
es o
f the
alli
ance
Con
volv
ulo
arve
nsis
-Agr
opyr
ion
repe
ntis
Falc
aria
vul
gari
s 5
55
55
54
4V
r.
.+
..
..
II.
..
.+
+.
II.
..
..
1.
..
..
I+
Elyt
rigi
a re
pens
+
ab
.+
++
1V
55
55
55
55
V1
1+
1.
+.
IV.
1.
..
..
+.
a.
II+
Brom
us in
erm
is
.
..
..
..
. .
..
..
..
+.
I5
55
55
54
V.
..
..
..
..
..
..
Car
dari
a dr
aba
..
..
r.
r.
II.
..
+.
+.
.II
..
..
..
rI
55
55
55
55
44
4V
.
Cho
ndri
lla ju
ncea
.
..
..
.r
.I
..
..
..
..
..
..
..
..
..
..
..
..
..
..
.5
Con
stan
t tax
a of
the
allia
nce
Con
volv
ulo
arve
nsis
-Agr
opyr
ion
repe
ntis
Con
volv
ulus
arv
ensi
sa
.1
+.
r.
rIV
1+
+b
br
33
V1
.1
b1
ab
Vb
1+
..
++
.1
33
IV.
Arte
mis
ia v
ulga
ris
+
r+
r+
+r
.V
+1
++
.+
+.
IV.
+.
1+
b+
IV.
..
r.
..
.+
..
I+
Achi
llea
mill
efol
ium
agg
.
+
.a
.+
+a
.IV
++
..
.1
r.
III
..
..
..
+I
++
1+
++
++
+.
+V
b
Dia
gnos
tic, c
hara
cter
istic
, and
con
stan
t tax
a of
the
clas
s Art
emis
iete
a vu
lgar
is
Ballo
ta n
igra
.
1r
.r
a.
.II
I.
+1
++
b.
.IV
rr
+r
+.
.IV
..
..
r.
..
..
. .
.
Med
icag
o sa
tiva
+.
..
..
++
II+
..
..
..
.I
.+
++
..
.II
I.
+a
a.
.+
+.
a+
IV.
Sile
ne la
tifol
ia su
bsp.
alb
a
+.
.+
..
+.
II.
..
+.
1+
.II
.+
..
..
rII
+r
+.
..
.+
.+
.I
.
Sten
actis
ann
ua
1.
+.
.r
+.
III
+.
..
..
+.
II.
..
r.
..
I.
..
..
..
..
..
..
Cic
hori
um in
tybu
s
+.
+.
.r
+.
III
..
..
..
..
..
r.
..
1.
II.
..
..
..
..
..
.a
Bert
eroa
inca
na
+.
..
..
r.
II+
..
..
+.
.II
..
..
..
. .
..
..
..
.r
..
.I
r
Echi
um v
ulga
re
+.
.r
..
..
II+
..
r.
..
.II
..
..
.r
.I
..
..
..
..
..
. .
.
82
Ass
ocia
tion
or c
omm
unity
no.
12
34
5
Rel
evé
no.
12
34
56
78
S9
1011
1213
1415
16S
1718
1920
2122
23S
2425
2627
2829
3031
3233
34S
35
Dau
cus c
arot
a
+
..
..
..
.I
r.
..
..
..
I.
..
..
r.
I.
..
..
..
.r
..
I+
Car
duus
aca
ntho
ides
..
..
..
r.
I+
..
..
r.
.II
..
..
.r
.I
.r
..
..
..
..
.I
.
Cir
sium
arv
ense
..
..
..
..
..
+.
..
..
.I
1.
..
+.
.II
..
r.
..
..
..
. .
.
Anch
usa
offici
nalis
.
..
.r
..
.I
..
..
..
..
..
..
..
+.
I.
..
..
..
+.
..
I.
Rese
da lu
tea
.
..
..
..
. .
..
..
..
..
..
..
..
r.
I.
r.
..
+.
..
..
I.
Cal
amag
rost
is e
pige
jos
.
..
.b
..
.I
..
..
..
..
..
..
..
..
..
..
..
..
..
..
.a
Tana
cetu
m v
ulga
re
.
..
..
r.
.I
..
..
..
..
..
..
..
..
..
..
..
..
..
..
.r
Picr
is h
iera
cioi
des
.
..
..
..
. .
..
..
..
..
..
..
..
r.
I.
..
..
..
..
..
.r
Med
icag
o lu
pulin
a
..
..
..
..
..
..
..
..
. .
..
..
..
. .
.r
r.
..
..
..
.II
I.
Dia
gnos
tic, c
hara
cter
istic
, and
con
stan
t tax
a of
the
clas
s Mol
inio
-Arr
hena
ther
etea
Arrh
enat
heru
m e
latiu
s
1
.+
.b
1+
1IV
..
+.
++
++
IV1
1a
a.
++
V.
..
..
..
.a
+.
I1
Dac
tylis
glo
mer
ata
.
.r
+r
.1
.II
I+
.+
..
++
+IV
++
..
r+
+IV
+.
1.
..
.+
.+
rII
I+
Tara
xacu
m se
ct. R
uder
alia
r
r.
..
..
.II
..
r.
..
+.
IIr
..
+.
.r
III
+r
++
.+
r+
+.
.IV
.
Poa
prat
ensi
s
.
..
..
..
. .
..
..
.+
++
II+
..
+.
..
IIb
1a
+.
.+
11
+1
V.
Plan
tago
lanc
eola
ta
+.
r.
..
.r
II.
..
..
..
+I
..
.+
..
.I
++
+.
.+
.+
.r
.II
I.
Loliu
m p
eren
ne
..
+.
..
+.
II.
..
..
..
. .
..
.+
.+
.II
.r
+r
..
..
..
.II
.
Poly
gonu
m a
rena
stru
m
.+
..
..
..
I.
r.
..
..
.I
..
..
..
. .
..
.+
1.
.+
..
.II
.
Trifo
lium
pra
tens
e
..
..
..
..
..
..
..
.r
.I
..
..
..
. .
++
+.
..
..
..
.II
.
Dia
gnos
tic, c
hara
cter
istic
, and
con
stan
t tax
a of
the
clas
s Ste
llari
etea
med
iae
Hor
deum
mur
inum
.
.r
..
..
.I
..
+.
+.
++
III
.r
+r
..
+II
I.
+.
++
..
..
..
II.
Cap
sella
bur
sa-p
asto
ris
..
..
..
..
..
..
..
..
. .
r.
..
..
.I
.+
++
++
.+
rr
+V
.
Brom
us st
erili
s
..
+.
..
..
I.
..
.+
+.
.II
..
..
..
. .
..
.+
+.
..
..
.I
.
83
Ass
ocia
tion
or c
omm
unity
no.
12
34
5
Rel
evé
no.
12
34
56
78
S9
1011
1213
1415
16S
1718
1920
2122
23S
2425
2627
2829
3031
3233
34S
35
Stel
lari
a m
edia
.
..
..
..
. .
..
..
..
..
..
..
..
..
..
.+
..
+1
..
.+
II.
Lact
uca
serr
iola
.
+.
..
..
.I
r.
..
..
r.
II.
..
..
..
..
..
..
..
..
..
..
Vici
a an
gust
ifolia
..
.r
..
..
I.
..
..
..
. .
..
..
..
. .
..
..
..
..
.r
rI
.
Lam
ium
pur
pure
um
..
..
..
..
..
..
..
..
. .
..
..
..
. .
.+
+.
..
+.
..
.II
.
Papa
ver r
hoea
s
..
..
..
.+
I.
..
..
.+
.I
..
..
..
. .
..
..
..
..
..
. .
.
Vero
nica
per
sica
.
..
..
..
. .
..
..
..
..
..
..
..
..
..
++
..
..
..
..
I.
Vero
nica
arv
ensi
s
.
..
..
..
. .
..
..
..
..
..
..
..
..
..
..
..
..
+.
.r
I.
Dia
gnos
tic, c
hara
cter
istic
, and
con
stan
t tax
a of
the
clas
s Gal
io-U
rtic
etea
Cle
mat
is v
italb
a
.
.a
.3
3.
.II
rr
..
r.
..
II.
+.
..
+.
II.
..
..
..
..
..
..
Gal
ium
apa
rine
.
..
..
..
. .
..
+r
+.
..
IIr
+.
..
.+
III
..
..
.r
+.
..
.I
.
Urt
ica
dioi
ca
..
..
..
..
..
..
a.
.+
+II
1+
..
+.
+II
I.
..
..
..
..
..
..
Vero
nica
subl
obat
a
..
..
..
..
..
..
..
..
. .
..
..
..
. .
..
..
a+
+.
1.
1II
I.
Hum
ulus
lupu
lus
..
..
.r
..
I.
..
..
..
. .
1.
..
+.
.II
..
..
..
..
..
. .
.
Rubu
s cae
sius
.
..
..
..
. .
..
..
..
..
..
r.
..
+.
II.
..
..
..
..
..
..
Oth
er ta
xa
Erod
ium
cic
utar
ium
..
..
..
..
..
..
..
+.
.I
..
..
..
. .
.1
.1
.1
..
.+
.II
.
Poa
bulb
osa
..
..
..
..
..
..
..
..
. .
..
..
..
. .
.+
..
..
1a
.a
rII
I.
Aila
nthu
s alti
ssim
a (ju
v.)
.
..
..
..
aI
..
..
..
..
..
..
.r
r.
II.
..
.+
..
..
..
I.
Rum
ex c
risp
us
.
..
..
..
. .
..
.r
..
..
I.
..
.r
..
I+
..
..
..
.+
..
I.
Col
ymba
da sc
abio
sa
.
..
..
.+
.I
r.
..
..
..
I.
..
..
+.
I.
..
..
..
..
..
..
Brom
us h
orde
aceu
s
.
..
..
..
. .
..
..
..
..
..
..
..
..
..
..
+.
..
+.
+.
II.
Ono
bryc
his v
iciif
olia
r
..
..
..
.I
..
..
..
..
..
..
..
..
..
..
..
..
..
r.
I

84
Ass
ocia
tion
or c
omm
unity
no.
12
34
5
Rel
evé
no.
12
34
56
78
S9
1011
1213
1415
16S
1718
1920
2122
23S
2425
2627
2829
3031
3233
34S
35
Med
icag
o fa
lcat
a
.
..
b.
..
+II
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
Secu
rige
ra v
aria
.
..
..
.+
.I
..
..
..
..
..
..
.+
..
I.
..
..
..
..
..
..
Rum
ex p
atie
ntia
.
..
..
..
. .
..
..
..
r.
I.
.r
..
..
I.
..
..
..
..
..
..
Hyp
eric
um p
erfo
ratu
m
..
..
..
..
..
..
..
..
. .
..
..
r.
.I
..
..
..
..
+.
.I
.
Erop
hila
ver
na
..
..
..
..
..
..
..
..
. .
..
..
..
. .
..
..
.1
..
..
+I
Orn
ithog
alum
um
bella
tum
..
..
..
..
..
..
..
..
. .
..
..
..
. .
..
..
.+
..
..
+I
Lath
yrus
tube
rosu
s
..
..
..
..
..
..
..
..
. .
..
..
..
. .
..
..
..
..
rr
.I
.
Taxa
reco
rded
in o
nly
one
rele
vé:
E 1: Al
liari
a pe
tiola
ta (
18):
+, A
rist
oloc
hia
clem
atiti
s (1
5):
+, A
rtem
isia
abs
inth
ium
(2)
: +,
Ast
raga
lus
cice
r (2
2):
+, A
trip
lex
patu
la (
13):
r, At
ripl
ex t
atar
ica
(10)
: r,
Che
nopo
dium
str
ictu
m (3
1): r
, Cir
sium
vul
gare
(35)
: r, C
onyz
a ca
nade
nsis
(12)
: +, D
escu
rain
ia s
ophi
a (1
1): r
, Ery
ngiu
m c
ampe
stre
(7):
r, Fr
axin
us e
xcel
sior
(juv
.) (1
): r,
Ger
aniu
m p
usill
um (2
6): r
, Geu
m u
rban
um (1
8): r
, Gle
chom
a he
dera
cea
(16)
: 1, H
eder
a he
lix (1
3): +
, Hol
cus l
anat
us (2
1): b
, Hol
oste
um u
mbe
llatu
m (2
9): r
, Jac
ea p
rate
nsis
(7
): 1,
Jug
lans
regi
a (ju
v.) (
22):
r, K
naut
ia a
rven
sis
(22)
: +, M
edic
ago
×var
ia (4
): +,
Mel
ilotu
s offi
cina
lis (2
1): +
, Ono
pord
um a
cant
hium
(12)
: r, P
oa a
nnua
(10)
: r, P
oa
com
pres
sa (
17):
r, Ro
sa c
anin
a ag
g. (j
uv.)
(8):
r, Ru
bus s
ubge
n. R
ubus
(21)
: +, R
umex
obt
usifo
lius (
15):
+, S
ambu
cus e
bulu
s (17
): +,
Sap
onar
ia o
ffici
nalis
(21)
: +, S
enec
io
vulg
aris
(30)
: +, S
isym
briu
m lo
esel
ii (8
): r,
Sonc
hus
arve
nsis
(14)
: r, S
orgh
um h
alep
ense
(22)
: r, S
tella
ria
holo
stea
(31)
: r, T
rifo
lium
arv
ense
(4):
r, Tr
ifoliu
m c
ampe
stre
(3
3): +
, Tri
foliu
m re
supi
natu
m (3
1): r
, Val
eria
nella
car
inat
a (2
6): +
, Val
eria
nella
locu
sta
(33)
: +, V
eron
ica
hede
rifo
lia (3
2): 1
, Vic
ia la
thyr
oide
s (34
): +,
Vic
ia v
illos
a (1
8): r
E 2: Ai
lant
hus a
ltiss
ima
(9):
+

85
Localities and other information of relevés from Tab. 1:
Data are arranged in the following order: Locality, coordinates, altitude, slope, aspect, relevé area, total cover, cover of E1, cover of E2, cover of E3, cover of E0, height of herb layer, height of shrub layer, height of tree layer, soil type, date, author of relevé
Association Falcario vulgaris-Agropyretum repentis:
Relevé no. 1, Podunajská rovina Flatland, Bratislava, Karlova Ves district, Botanická street, abandoned area 20 m from the tram stop Botanická záhrada, N 48°08′53.70″, E 17°04′20.10″, ± 7 m, altitude: 165 m, slope: 15°, aspect: 145° (SE), relevé area: 18.00 m², total cover: 95%, cover of E1: 95%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 10–50 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand and gravel, 15.7.2011, A.Rendeková
Relevé no. 2, Malé Karpaty Mts., Bratislava, Rača district, Alstrova street, abandoned area along the road, N 48°12′25.60″, E 17°08′36.50″, ± 6 m, altitude: 180 m, slope: 0°, aspect: -, relevé area: 10.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 65–120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 26.6.2013, A.Rendeková
Relevé no. 3, Podunajská rovina Flatland, Bratislava, Staré Mesto district, Žižkova street, abandoned area near the pathway, N 48°08′37.20″, E 17°04′56.20″, ± 6 m, altitude: 147 m, slope: 4°, aspect: 198° (SSW), relevé area: 24.00 m², total cover: 99%, cover of E1: 99%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 60–160–170 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 16.7.2012, A.Rendeková
Relevé no. 4, Borská nížina Lowland, Bratislava, Devínska Nová Ves district, abandoned area 24 m from the Devínska Nová Ves Railway station, N 48°13′07.90″, 16°58′38.90″, ± 8 m, altitude: 149 m, slope: 0°, aspect: -, relevé area: 10.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 26.6.2013, A.Rendeková
Relevé no. 5, Podunajská rovina Flatland, Bratislava, Vrakuňa district, Hradská street, abandoned area near the railway track, N 48°19′27.10″, E 17°11′24.80″, ± 5 m, altitude: 136 m, slope: 0°, aspect: -, relevé area: 12.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 80–160 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 16.7.2012, A.Rendeková
Relevé no. 6, Podunajská rovina Flatland, Bratislava, Nové Mesto district, Račianska street, 200 m from Mladá garda, abandoned area near the railway track, N 48°10′47.30″, E 17°07′44.40″, ± 7 m, altitude: 143 m, slope: 7°, aspect: 235° (SW), relevé area: 16.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 85–150 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand and gravel, 26.7.2012, A.Rendeková
Relevé no. 7, Podunajská rovina Flatland, Bratislava, Račianska street, abandoned area 100 m from the Bratislava-Vinohrady Railway station, N 48°11′15.70″, E 17°08′05.00″, ± 6 m, altitude: 139 m, slope: 0°, aspect: -, relevé area: 12.00 m², total cover: 90%, cover of E1: 90%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 60–90 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 26.7.2012, A.Rendeková
Relevé no. 8, Podunajská rovina Flatland, Bratislava, Nové Mesto district, 160 m from the Bratislava-Nové Mesto Railway station, N 48°10′19.00″, E 17°08′43.80″, ± 6 m, altitude: 148 m, slope: 40°, aspect: 200° (SSW), relevé area: 24.00 m², total cover: 85%, cover of E1: 85%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 15–60–100 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand and gravel, 26.6.2012, A.Rendeková

86
Elytrigia repens community [Convolvulo arvensis-Agropyrion repentis]:
Relevé no. 9, Malé Karpaty Mts., Bratislava, Mlynská dolina, 26 m from bus stop Zoologická záhrada, N 48°09′13.10″, E 17°04′36.40″, ± 6 m, altitude: 176 m, slope: 10°, aspect: 324° (NW), relevé area: 10.00 m², total cover: 100%, cover of E1: 100%, cover of E2: 1%, cover of E3: -, cover of E0: -, height of herb layer: 80–90 cm, height of shrub layer: 1.6 m, height of tree layer: -, soil type: loamy + admixture of sand, 20.8.2012, A.Rendeková
Relevé no. 10, Podunajská rovina Flatland, Bratislava, Staré Mesto district, Nábrežie arm. gen. Ludvíka Svobodu street, edge of the road, N 48°08′24.60″, E 17°05′54.10″, ± 5 m, altitude: 140 m, slope: 0°, aspect: -, relevé area: 12.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 80–165 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 6.8.2012, A.Rendeková
Relevé no. 11, Podunajská rovina Flatland, Bratislava, 100 m from Račianska street, abandoned area near the railway bridge, N 48°10′56.40″, E 17°07′56.10″, ± 12 m, altitude: 140 m, slope: 0°, aspect: -, relevé area: 12.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 65–100 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 24.5.2013, A.Rendeková
Relevé no. 12, Malé Karpaty Mts., Bratislava, Rača district, edge of the road at the crossroad of Alstrova street and Úžiny street, N 48°12′28.30″, E 17°08′38.80″, ± 6 m, altitude: 176 m, slope: 0°, aspect: -, relevé area: 8.00 m², total cover: 98%, cover of E1: 98%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 90–120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 26.6.2013, A.Rendeková
Relevé no. 13, Podunajská rovina Flatland, Bratislava, Ružinov district, the crossroad of Krasinského street and Trnavská cesta street, edge of the road 10 m from the railway underpass, N 48°09′54.20″, E 17°09′42.50″, ± 6 m, altitude: 146 m, slope: 0°, aspect: -, relevé area: 6.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 24.5.2013, A.Rendeková
Relevé no. 14, Malé Karpaty Mts., Bratislava, Kramáre district, Vlárska street, edge of the pathway, N 48°10′04.60″, E 17°05′29.80″, ± 6 m, altitude: 238 m, slope: 4°, aspect: 180° (S), relevé area: 12.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 20.5.2013, A.Rendeková
Relevé no. 15, Borská nížina Lowland, Bratislava, Devinska Nová Ves district, Mlynská street, edge of the road, N 48°13′01.70″, 16°58′25.40″, ± 6 m, altitude: 142 m, slope: 0°, aspect: -, relevé area: 6.00 m², total cover: 98%, cover of E1: 98%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 100 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 26.6.2013, A.Rendeková
Relevé no. 16, Podunajská rovina Flatland, Bratislava, Petržalka district, Viedenská cesta street, edge of the road 260 m from the bridge, N 48°08′10.80″, E 17°06′09.40″, ± 6 m, altitude: 133 m, slope: 6°, aspect: 160° (SSE), relevé area: 16.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 110 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 18.5.2013, A.Rendeková
Association Convolvulo-Brometum inermis:
Relevé no. 17, Borská nížina Lowland, Bratislava, Devínska Nová Ves district, edge of the pathway 200 m from the bus stop Na hriadkach, N 48°12′27.40″, 16°58′09.80″, ± 6 m, altitude: 144 m, slope: 0°, aspect: -, relevé area: 14.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 100–120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 26.5.2013, A.Rendeková
Relevé no. 18, Malé Karpaty Mts., Bratislava, Dlhé diely district, Iskerníková street, abandoned area near the pathway, N 48°09′12.10″, E 17°02′40.30″, ± 6 m, altitude: 220 m, slope: 16°, aspect: 154° (SSE), relevé area: 24.00 m², total cover: 98%, cover of E1: 98%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 10.6.2013, A.Rendeková

87
Relevé no. 19, Podunajská rovina Flatland, Bratislava, Vlčie hrdlo, Slovnaftská street, edge of the road near Malý Dunaj River, N 48°07′50.80″, E 17°10′24.20″, ± 6 m, altitude: 142 m, slope: 20°, aspect: 258° (SWW), relevé area: 13.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 125–165 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 6.6.2013, A.Rendeková
Relevé no. 20, Malé Karpaty Mts., Bratislava, Karlova Ves district, the crossroad of Borská street and Pernecká street, N 48°09′42.10″, E 17°03′06.60″, ± 9 m, altitude: 200 m, slope: 0°, aspect: -, relevé area: 12.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 100–140 cm, height of shrub layer: -, -, soil type: loamy, 28.5.2013, A.Rendeková
Relevé no. 21, Podunajská rovina Flatland, Bratislava, Rača district, Žabí majer, abandoned area near the railway track, N 48°12′00.70″, E 17°09′07.80″, ± 8 m, altitude: 140 m, slope: 0°, aspect: -, relevé area: 22.00 m², total cover: 98%, cover of E1: 98%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 100 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand and gravel, 6.7.2013, A.Rendeková
Relevé no. 22, Podunajská rovina Flatland, Bratislava, Nové Mesto district, Rožňavská street, 40 m from the railway track, N 48°10′03.40″, E 17°09′19.50″, ± 8 m, altitude: 141 m, slope: 0°, aspect: -, relevé area: 24.00 m², total cover: 96%, cover of E1: 96%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 120 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 16.7.2013, A.Rendeková
Relevé no. 23, Malé Karpaty Mts., Bratislava, Dúbravka district, Agátová street, edge of the pathway, N 48°11′46.10″, E 17°02′24.00″, ± 14 m, altitude: 216 m, slope: 0°, aspect: -, relevé area: 10.00 m², total cover: 90%, cover of E1: 90%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 80 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 6.6.2013, A.Rendeková
Association Lepidio drabae-Agropyretum repentis:
Relevé no. 24, Podunajská rovina Flatland, Bratislava, Petržalka district, Jantárová cesta street, abandoned area near the road, N 48°06′19.20″, E 17°06′29.20″, ± 6 m, altitude: 146 m, slope: 0°, aspect: -, relevé area: 14.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 45 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 8.5.2013, A.Rendeková
Relevé no. 25, Podunajská rovina Flatland, Bratislava, Petržalka district, area near the road at the crossroad of Rusovská cesta street and Jantárová cesta street, N 48°07′25.30″, E 17°06′44.80″, ± 8 m, altitude: 142 m, slope: 4°, aspect: 160° (SSE), relevé area: 12.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 60 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 8.5.2013, A.Rendeková
Relevé no. 26, Podunajská rovina Flatland, Bratislava, Petržalka district, abandoned area among the buildings at the crossroad of the Jasovská street and Jantárová cesta street, N 48°05′51.50″, E 17°06′34.50″, ± 10 m, altitude: 146 m, slope: 0°, aspect: -, relevé area: 20.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 45 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 8.5.2013, A.Rendeková
Relevé no. 27, Podunajská rovina Flatland, Bratislava, Petržalka district, Budatínska street, edge of the road near the bus stop ZŠ Budatínska, N 48°06′10.80″, E 17°05′58.40″, ± 12 m, altitude: 136 m, slope: 0°, aspect: -, relevé area: 10.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 56 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 12.5.2013, A.Rendeková
Relevé no. 28, Podunajská rovina Flatland, Bratislava, Mlynské nivy street, edge of the road, N 48°08′46.60″, E 17°07′40.80″, ± 8 m, altitude: 136 m, slope: 0°, aspect: -, relevé area: 8.00 m², total cover: 98%, cover of E1: 98%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 45 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 2.5.2013, A.Rendeková
Relevé no. 29, Podunajská rovina Flatland, Bratislava, Staré Mesto district, the crossroad of Žižkova street and Nábrežie arm. gen. Ľudvíka Svobodu street, edge of the pathway, N 48°08′37.20″, E 17°04′54.70″, ± 6 m,

88
altitude: 150 m, slope: 40°, aspect: 188° (S), relevé area: 12.00 m², total cover: 85%, cover of E1: 85%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 30 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 26.4.2013, A.Rendeková
Relevé no. 30, Podunajská rovina Flatland, Bratislava, Staré Mesto district, the crossroad of Nad lomom street and Nábrežie arm. gen. Ludvíka Svobodu street, edge of the road, N 48°08′45.60″, E 17°04′37.80″, ± 9 m, altitude: 150 m, slope: 30°, aspect: 186° (S), relevé area: 8.00 m², total cover: 95%, cover of E1: 95%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 40 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 28.4.2013, A.Rendeková
Relevé no. 31, Podunajská rovina Flatland, Bratislava, Ružinov district, abandoned area at the crossroad of Bajkalská street and Ružinovská street, N 48°09′20.40″, E 17°08′51.10″, ± 8 m, altitude: 142 m, slope: 0°, aspect: -, relevé area: 24.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 45 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 16.5.2013, A.Rendeková
Relevé no. 32, Malé Karpaty Mts., Bratislava, Kramáre district, Pod Krásnou hôrkou street, edge of the road, N 48°10′10.00″, E 17°05′26.30″, ± 7 m, altitude: 240 m, slope: 25°, aspect: 180°(S), relevé area: 8.00 m², total cover: 95%, cover of E1: 95%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 50 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 5.6.2013, A.Rendeková
Relevé no. 33, Podunajská rovina Flatland, Bratislava, Petržalka district, 89 m from Viedenská cesta street, edge of the road, N 48°08′09.80″, E 17°05′59.10″, ± 5 m, altitude: 130 m, slope: 20°, aspect: 167° (SSE), relevé area: 8.00 m², total cover: 100%, cover of E1: 100%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 60 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy + admixture of sand, 18.5.2013, A.Rendeková
Relevé no. 34, Podunajská rovina Flatland, Bratislava, Ružinov district, abandoned area 20 m from the tram stop Astronomická, N 48°09′31.20″, E 17°10′42.90″, ± 7 m, altitude: 136 m, slope: 0°, aspect: -, relevé area: 8.00 m², total cover: 85%, cover of E1: 85%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 40 cm, height of shrub layer: -, height of tree layer: -, soil type: loamy, 30.4.2013, A.Rendeková
Association Asparago-Chondrilletum juncei:
Relevé no. 35, Podunajská rovina Flatland, Bratislava, Rača district, 70 m from the crossroad of Pekná cesta street and Račianska street, 20 m from the railway track, abandoned area, (Note: sunny site), N 48°11′42.80″, E 17°08′39.90″, ± 8 m, altitude: 139 m, slope: 0°, aspect: -, relevé area: 9.00 m², total cover: 90%, cover of E1: 90%, cover of E2: -, cover of E3: -, cover of E0: -, height of herb layer: 110 cm, height of shrub layer: -, height of tree layer: -, soil type: sandy + admixture of gravel, 8.8.2013, A.Rendeková
acknowledgements
This study was supported by the Operation Program of Research and Innovation for the project: Advancing University Capacity and Competence in Research, Development and Innovation, ITMS2014+: 313021X329, co-financed by the European Regional Development Fund.

89
references
Barkman, J. J., Doing, H., Segal, S., 1964: Kritische Bemerkungen und Vorschläge zur quantitativen Vegetationsanalyse. Acta Bot. Neerl., 13: 394-419.
Braun-Blanquet, J., 1964: Pflanzensoziologie. Grundzüge der Vegetationskunde. Ed. 3. Springer Verlag, Wien, New York.
Chytrý, M. (ed.), 2009: Vegetace České republiky 2. Ruderální, plevelová, skalní a suťová vegetace. Academia, Praha.
Feráková, V., Jarolímek, I., 2011: Bratislava. In: Kelcey, J.-K., Müller, N., (eds.), Plants and habitats of European cities, p. 79-129, Springer, New York.
Forman, R.T.T., 2014: Urban Ecology, Science of cities. Cambridge University Press, Camridge UK. Hennekens, S. M., Schaminée, J. H. J., 2001: TURBOVEG, a comprehensive data base management system
for vegetation data. J. Veg. Sci., 12: 589-591.Jarolímek, I., 1983: Ruderálne spoločenstvá Bratislavy. PhD. thesis, depon. in Institute of Botany SAV,
Bratislava.Jarolímek, I., 1986: Ruderálne spoločenstvá juhozápadnej časti Malých Karpát. Acta Bot. Slov. Bratislava,
Ser. A 9: 9-10.Jarolímek, I., Šibík, J. (eds.), 2008: Diagnostic, constant and dominant species of the higher vegetation units
of Slovakia. Veda, Bratislava.Jarolímek, I., Valachovič, M., 2016: Dve zriedkavé ruderálne spoločenstvá z Bratislavy. Bull. Slov. Bot.
Spoločn., 38(1): 79-84.Jarolímek, I., Zaliberová, M., Mucina, L., Mochnacký, S., 1997: Rastlinné spoločenstvá Slovenska. 2.
Synantropná vegetácia. Veda, Bratislava.Kopecký, K., 1982: Die Ruderalpflanzengesellschaften im südwestlichen Teil von Praha (3). Preslia, 54: 67-
89.Krippelová, T., Mucina, L., 1988: Charakteristika vyšších syntaxónov triedy Stellarietea mediae na
Slovensku. Preslia, 60: 41-58.Mack, R. N., Simberloff, D., Lonsdale, W. M., Evans, H., Clout, M., Bazzaz, F. A., 2000: Biotic invasions:
causes, epidemiology, global consequences, and control. Ecol. Appl. 10: 689-710.Májeková, J., Letz, D. R., Slezák, M., Zaliberová, M., Hrivnák, R., 2014: Rare and threatened vascular
plants of the railways in Slovakia. Biodiv. Res. Conserv., 35: 75-85.Marhold, K., Hindák, F. (eds.), 1998: Zoznam nižších a vyšších rastlín Slovenska. Veda, Bratislava.Medvecká, J., Kliment, J., Májeková, J., Halada, Ľ., Zaliberová, M., Gojdičová, E., Feráková, V., Jarolímek,
I., 2012: Inventory of alien species of Slovakia. Preslia, 84: 257-309. Mucina, L., Grabherr, G., Ellmauer, T. (eds.), 1993: Die Pfanzengesellschaften Österreichs. Teil I.
Anthropogene Vegetation. Gustav Fischer Verlag, Jena.Mucina, L., Bültmann, H., Dierssen, K., Theurillat, J.-P., Raus, T., Čarni, A., Šumberová, K., Willner,
W., Dengler, J., Gavilán García, R., Chytrý, M., Hájek, M., Di Pietro, R., Iakushenko, D., Pallas, J., Daniëls, F. J. A., Bergmeier, E., Santos Guerra, A., Ermakov, N., Valachovič, M., Schaminée, J. H. J., Lysenko, T., Didukh, Y. P., Pignatti, S., Rodwell, J. S., Capelo, J., Weber, H. E., Solomeshch, A., Dimopoulos, P., Aguiar, C., Hennekens, S. M., Tichý, L., 2016: Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci.19(Suppl. 1): 3-264.
Podani, J., 2001: SYN-TAX 2000. Computer Program for Data Analysis in Ecology and Systematics for Windows 95, 98 & NT. User’s manual. Scientia Publ., Budapest.
Pyšek, P., Richardson, D. M., 2010: Invasive species, environmental change and management, and health. Annu. Rev. Env. Res., 35: 25-55.
Pyšek, P., Sádlo, J., Mandák, B., 2002: Catalogue of alien plants of the Czech Republic. Preslia, 74: 97-186Pyšek, P., Jarošík, V., Hulme, P. E., Pergl, J., Hejda, M., Schaffner, U., Vilá, M., 2012: A global assessment
of alien invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Glob. Change Biol., 18: 1725-1737.
Rendeková, A., 2016: Little-known ruderal plant communities recorded in Bratislava. Acta Botanica Universitatis Comenianae, 51: 23-29.
Rendeková, A., Mičieta, K., 2017: The trampled communities of the class Polygono arenastri-Poetea annuae Rivas-Martínez 1975 corr. Rivas-Martínez et al. 1991 in the ruderal vegetation of Bratislava and their biodiversity. Acta Botanica Universitatis Comenianae, 52: 57-69.

90
Rendeková, A., Kerekeš, E., Miškovic, J., 2014: Rare and interesting ruderal plant communities of Bratislava. Acta Botanica Universitatis Comenianae, 49: 13-18.
Rendeková, A., Mičieta, K., Miškovic, J., 2019a: The stands of invasive neophyte Stenactis annua in the ruderal vegetation of Bratislava. Acta Botanica Universitatis Comenianae, 54: 21-29.
Rendeková, A., Mičieta, K., Miškovic, J., 2020: Ruderal vegetation of the class Artemisietea vulgaris in Bratislava city 1. Alliance Dauco-Melilotion. Acta Botanica Universitatis Comenianae, 56: 27-45.
Rendeková, A., Miškovic, J., Kerekeš, E., 2015: Asociácia Echio-Melilotetum R. Tx. 1947. na území Bratislavy a zmeny v jej druhovom zložení. Zborník recenzovaných príspevkov zo študentskej vedeckej konferencie PríF UK, 2015: 637-642.
Rendeková, A., Miškovic, J. Mičieta, K., 2017: Spoločenstvá inváznych neofytov zväzu Senecionion fluviatilis R. Tx. 1950 v ruderálnej vegetácii Bratislavy a ich biodiverzita. Acta Universitatis Matthiae Belii series Environmental management, 19(2): 39-54.
Rendeková, A., Miškovic, J., Mičieta, K., 2019b: Ruderal plant communities from the ordo Lamio albi-Chenopodietalia boni-henrici Kopecký 1969 in Bratislava city. Acta Botanica Universitatis Comenianae, 54: 7-19.
Rendeková, A., Hrabovský, M., Miškovic, J., Mičieta, K., 2018: Expanzívny druh Sambucus ebulus v asociácii Sambucetum ebuli v Bratislave. Acta Universitatis Matthiae Belii series Environmental management, 20(1): 56-64.
Sukopp, H., 2002: On the early history of urban ecology in Europe. Preslia, 74: 373-393.Tichý, L., 2002: JUICE, software for vegetation classification. J. Veg. Sci., 13: 451-453.Valachovič, M., 2016: Zaujímavejšie fytocenologické zápisy – zápis č. 11-15. Bull. Slov. Bot. Spoločn., 38(1): 97-98.Zaliberová, M., 1982: Poznámky k ruderálnym spoločenstvám niektorých obcí južnej časti Východoslovenskej
nížiny. Acta Bot. Slov. Bratislava Ser. A6: 183-200.
Abstrakt
Príspevok opisuje ruderálnu vegetáciu zväzu Convolvulo arvensis-Agropyrion repentis Görs 1966 (trieda Artemisietea vulgaris Lohmeyer et al. ex von Rochow 1951) v Bratislave. Príspevok je pokračovaním predchádzajúceho článku, ktorý bol zameraný na vegetáciu zväzu Dauco-Melilotion Görs 1966. Uvádzame vlastné fytocenologické zápisy zaznamenaných asociácií a spoločenstiev a referujeme o ich druhovom zložení a rozšírení na území Bratislavy. Opisujeme aj porovnanie s údajmi z iných čatsí Slovenska. Zaznamenali sme 35 zápisov zo štyroch asociácií a jedného spoločenstva, vrátane pomerne vzácnej asociácie Asparago-Chondrilletum juncei Passarge 1978.
Kľúčové slová: človekom ovplyvnené habitaty, fytocenológia, juhozápadné Slovensko, (sub)xerofilné synantropné rastlinné rastlinné spoločenstvá

91
Acta Botanica Universitatis Comenianae Vol. 57, 2021
sPomienka na kolegyňu, Pracovníčku katedry Botaniky Prírodovedeckej fakulty univerzity komenskéHo
v Bratislavecommemoration of a colleague, a staff memBer of tHe dePartment
of Botany, faculty of natural sciences, comenius university in Bratislava
rndr. anna uHríková 18. 2. 1939 – 14. 1. 2021
Karol Mičieta*, Duňa Javorčíková, Terézia Schwarzová, Katarína Mišíková, Jozef Dušička, Ľubomír Kováčik, Soňa Jančovičová
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany, Révová 39, 811 02 Bratislava, Slovakia
Received 3 April 2021, Received in revised form 3 April 2021, Accepted 5 April 2021
* Corresponding author: [email protected]
RNDr. Anna Uhríková na oslave 70. výročia založenia Katedry botaniky konanej v novembri 2010 (foto: Ľubomír Kováčik)

92
Smutná správa nám rozochvela pamäť. Kľúč svetla otvára svit spomienok v stĺporadí stretnutí, príbehov, činov, nádejí, radostí zo svitov hviezd či zistených počtov chromozómov, pohnútok a správ pri voňavej kávičke, o neúnavnom a neprestajnom pohybe v zornom uhle bez obmedzenia času, vo vianočných starostiach o blízkych a krehkých oblátkach, vo vonných spomienkach na fašiangové šišky, či sústredenosti na každodenné upresňovanie a usmerňovanie okamihov začiatkov a večných pokusov rozlúštenia striedania blízkostí, počiatkov, prílivov i tichého uvädania kvetov.
Hanka Uhríková bola prítomná srdcom doma, v rodine, na pracovisku, všade. Rodáčka z Vrbového najviac milovala krajinu okolia Prašníka, v časti chotára Boorovci, kde bolo hospodárstvo jej rodičov, na ktorom vyrastala. Neskôr tu pri práci aj oddychu, uprostred lúk, políčok, ovocných sadov čerpala životnú energiu, pre srdečný vzťah k ľudom, k prírode, zanietenosti k práci, pre silu jej osobnosti.
Po maturite študovala na Prírodovedeckej fakulte Univerzity Komenského v Bratislave, odbor biológia, zameranie mikrobiológia, ktorý ukončila v roku 1963. Rýchlo sa stala platnou členkou, výkonnou hybnou silou rozvíjajúceho sa pracovného kolektívu na Katedre botaniky pod vedením doc. J. Májovského, ktorý sa s veľkým zanietením a intenzitou začal zaoberať karyotaxonomickým štúdiom taxónov flóry Slovenska. I keď Hanka Uhríková nie je autorkou metodiky, jej historický význam spočíva v stovkách analýz počtu chromozómov rôznych taxónov, ktoré jej z terénu priniesli spolupracovníci a spriaznení botanici, či z exemplárov, o ktoré sa starala na experimentálnom políčku. Významne prispela k zavedeniu cytologických metodík do laboratórnej praxe, k ich zjednodušeniu a bežnému používaniu nielen pre školených cytológov, ale i pre široké spektrum záujemcov, študentov, diplomantov. Významne sa podieľala a prispela k výsledkom karyotaxonomického výskumu taxónov flóry Slovenska, ktorý bol zároveň aj ich enumeráciou a patril vo výskume ku špičke nielen v Československu, ale aj v Európe, čo umožnilo publikovať výsledky nielen v domácich časopisoch, prevažne v Acta Botanica Universitatis Comenianae (v tom čose pod iným názvom), ale aj v prestížnom európskom časopise Taxon. Výsledky výskumu prezentuje súborné dielo Karyotaxonomický prehľad flóry Slovenska (1987), neskoršie doplnený o Prvý doplnok karyotaxonomického prehľadu (Supplementum 1, 2000). Doktorka Uhríková v spolupráci hlbšie študovala problematiku mnohých taxónov napr. rodov Astragalus, Mercurialis, Muscari, Sedum telephium agg., Sorbus a Stellaria.
Za svoju záslužnú prácu bola ocenená viacerými vyznamenaniami. Bola dlhoročnou členkou Slovenskej botanickej spoločnosti a Československej biologickej spoločnosti. Svojou srdečnosťou, zanietenosťou, starostlivosťou o svoje okolie a ľudí si získala široký okruh svojich spolupracovníkov, študentov, najmä priateľov.
V roku 2003 odišla na dôchodok. Nikdy nevynechala možnosť stretnutia s kolegami i s novými pracovníkmi katedry. Jej príchod na naše vianočné katedry bol vždy radostnou slávnosťou. Tešila sa, že sa konečne môže naplno venovať veľkej záhrade na Prašníku. Túto pohodu si však, žiaľ, nemohla užívať natrvalo. V roku 2018 ostala pripútaná na vozík. Ani táto ťažká rana však Hanku nezlomila. Vďaka svojej húževnatosti, podpore manžela a rodiny, nestrácala svoju dobrosrdečnosť a optimizmus.
Milá Hanka, nemohli sme Ťa odprevadiť na Tvojej poslednej ceste, ale ostávaš trvalo zapísaná v našich srdciach. Spomíname na spoločné krásne chvíle. Lúčime sa, ale nezabúdame, ďakujeme. Česť Tvojej pamiatke.
Karol Mičieta

93
Kolegovia ju oslovovali Hana alebo Hanka, študenti pani doktorka. Na Katedre botaniky pôsobila veľa rokov, bola jej súčasťou. Ako jedna z hodinových súčiastok, bez ktorej by inak zastal čas. Raz sa všetkým zastaví, ale dokým budeme spomínať, dovtedy budú potíšku, ale predsa tĺcť. Prijmi od nás, Hanka, Tvojich kolegov, študentov a priateľov ešte zopár živých spomienok.
Je mi smutno, že Hana tak skoro odišla. V tento fašiangový čas by určite pokračovala v „šiškovej“ tradícii. Ale, žiaľ, už viac nebudú jej fantastické šišky s malinovou penou.
Ja si ju pamätám ešte zo študentských čias. Od roku 1963 už z Herbára. Končila na mikrobiológii, ale cesty osudu ju zaviali na Katedru botaniky. Eva Blašková-Kmeťová a ja sme boli v piatom ročníku a Hana nás zaúčala do tajomstva prípravy preparátov a iných prepotrebných činností v laboratóriu. V tom čase existoval na Katedre tzv. apartmán pre prípadných hostí katedry. Pán doc. Májovský usúdil, že bude dobrým riešením, keď ho dá Hane k dočasnému užívaniu, nakoľko v tých časoch nebolo jednoduché získať v Bratislave bývanie. A tak sa Hana stala súčasťou Herbára a my dve sme ju mali stále pri sebe, keď sme si nevedeli s niečim rady. Bol to fantastický rok. Plný práce ale aj potrebného uvoľnenia. Niekedy sme tam boli naozaj 24 hodín. A ako začala s nami, tak pokračovala s ďalšími generáciami študentov až do jej odchodu na dôchodok.
Bola aj vynikajúcou hubárkou. Huby bola jej vášeň a v teréne popri zbere materiálu na karyológiu sa aj „hubárčilo“. Pri hubách nepoznala sestru – brata. Ja som mala šťastie, že som vôbec netúžila po pôvabnici fialovej a tak som mohla v lese byť v jej blízkosti. Pri pečiarkach to však bolo iné.
Hana bola plná energie, elánu, smiechu a keď sa pre niečo nadchla alebo zistila, že sa deje neprávosť, vložila sa do problému s neopísateľnou vervou. A bolo jej jedno, kto stál za problémom – profesor, docent... či herbarista. Napokon, aj po Novembri mala veľký podiel na tom, že opäť bola na svete samostatná Katerda botaniky.
Duňa Javorčíková
Hanku som poznala ešte z internátu počas štúdia, znovu som sa s ňou stretla v roku 1962 v pôrodnici. Hanka priviedla na svet syna, ja dcéru. Aj tu prejavila svoju prirodzenú vlastnosť pomáhať a preto mliečko, ktoré jej syn nevládal spapať darovala detičkám, ktoré také šťastie nemali, medzi inými to bola aj moja dcéra. Naše deti sa potom spolu s deťmi iných kolegýň a kolegov stretávali a boli súčasťou výskumných ciest a terénnych prác na Katedre botaniky. Benevolentný doc. Májovský zakročil len vtedy, keď sa deti príliš rozšantili a hrozilo, že by mohli popadať zo skál. Hanka bola dušou kolektívu, v ktorom sa všetky problémy prerokúvali, niekedy aj veľmi búrlivo, ale nakoniec sa vždy obnovila rodinná atmosféra. Mala veľa dôležitých úloh. Bola nadšenou vedkyňou, erudovanou laborantkou, priateľskou a chápavou učiteľkou, a vždy sa bez váhania pustila do každej roboty, ktorú bolo treba urobiť.
Terézia Schwarzová

94
Hanka bola človekom, ktorého si zapamätáte na celý život. Dodnes počuť na katedre jej smiech, zvonivý hlas, cítiť vôňu medovníkov, oblátok a jabĺk, ktoré nám na Vianoce nosila. Ja to budem počuť a cítiť na každom vianočnom posedení... Chýbaš mi, Hanka.
Katarína Mišíková
Hanku som prvýkrát zaregistroval ako začínajúci študent štvrtého ročníka magisterského štúdia, keď ma ako záujemcu o diplomovku na Katedre botaniky so širokým úsmevom odkázala na vedúceho katedry. Potom už sprevádzala môj život na katedre intenzívne až do jej odchodu do dôchodku. Vždy bola ochotná poradiť pri koncipovaní cvičení z morfológie, podeliť sa so životnými skúsenosťami v oblasti vzťahov, pomôcť v knižničných službách či v rámci nejakých osláv so šibalským úsmevom pripomenúť „dať si za jeden na zdravie“. Pre mňa bola príjemným človekom s veľkým srdcom, ktorý vždy vo mne vedel vyvolať pocit vnútornej radosti. Všetky krásne spomienky tu žijú naďalej.
Jozef Dušička
Hana mala pracovňu hneď prvé dvere vľavo od vchodu. Strategická pozícia. Ani hore, ani v podzemí, tak akurát v strede diania. Bola to magická miestnosť, niečo ako stred vesmíru či magnet, ktorý pritiahol každého okoloidúceho. Vtiahlo Vás to dovnútra – vypočulo, pochválilo aj pokarhalo. Niekedy sa za dverami šuškalo, inokedy sa ozýval hurónsky smiech, tiež sa štrngalo – štamperlíkmi, petriho miskami aj podložnými sklíčkami. A potom si Hana sadla za čiernu Meoptu a mikroskopovala. A keď nemikroskopovala, učila. Taký koreň, onaká stonka, listy sediace alebo stopkaté. Mali sme ju radi. Mala som ju rada aj pre jej medovníky a ríbezľové víno, z ktorého sa mi dodnes krúti hlava. Na to sa predsa nedá zabudnúť.
Soňa Jančovičová
Poznámka redakcie: Bibliografia RNDr. Anny Uhríkovej, ktorú zostavila Terézia Schwarzová, bola publikovaná v časopise Bulletin slovenskej botanickej spoločnosti, ročník 43, číslo 1, str. 148–150 (2021).

ACTA BOTANICA UNIVERSITATIS COMENIANAE
Volume 57
Vydala Univerzita Komenského v Bratislave vo Vydavateľstve UKVyšlo v júni 2021
Technická redaktorka: Eva Zahradníková
ISSN: 2729-7330

instruction for authors
Manuscripts are accepted in English or Slovak. Each manuscript must include: title, name and address of author (authors), e-mail of corresponding author, abstract and maximum of 5 key words which should not repeat in the title of the manuscript (these parts must be submitted in both languages).
Avoid any special type of text formatting except for italic and bold. Start every paragraph on a new line, please do not use spaces or tabulator to create indents. The maximum range of the manuscript should be 15 norm pages, or per agreement with editorial staff. Send the menuscript to the executive editor’s email address. Accepted file formats are: *.doc, *.docx, *.rtf for the manuscript, *.tiff, *.jpg, *.png for pictures. Graphs should be also saved in a picture format.
Indicate the placement of all tables, black and white photos, drawings and maps combined with the legends directly in the text (adding also English legend is recommended for manuscripts written in Slovak) and attach the picture files separately.
Tables should contain cells, not data divided by spaces or tabulators, if possible. For figure labels, use the Arial font. The size of the font after scaling the picture into proper dimensions should be 8 or higher. The dimensions of any figure should not exceed 124 mm in width and 193 mm in height.
For references to illustrative matter in the text use the abbreviations Tab. 1, Fig. 1, Obr. 1, Figs. 1−4, Obr. 1 − 4 (range of values should be given with spaces in Slovak, without in English). The names of genera, species, infraspecific taxa and syntaxa must be written in italics. Acronyms of herbaria should be given according to Index Herbariorum: http://sweetgum.nybg.org/science/ih/. Do not abbreviate the journal titles in the references or unify them according to http://journalseek.net/.
Pokyny pre autorov
Redakcia prijíma príspevky napísané v anglickom aj slovenskom jazyku. Každý príspevok musí obsahovať názov, meno a adresu autora (autorov), e-mail korešpondujúceho autora, abstrakt a maximálne päť kľúčových slov, ktoré by sa nemali opakovať so slovami v názve (tieto časti článku musia byť v oboch jazykoch).
Nepoužívajte iné formátovanie textu okrem kurzívy a tučného písma. Maximálny rozsah príspevku je 15 normovaných strán, resp. po dohode s redakčnou radou. Príspevok pošlite v elektronickej forme na adresu výkonného redaktora. Akceptované formáty: *.doc, *docx, *.rtf pre textovú časť, *.tiff, *.jpg, *.png pre obrázky. Grafy by mali byť tiež uložené vo formáte obrázka.
Vyznačte umiestnenie všetkých tabuliek, čierno-bielych fotografií, kresieb a máp s príslušnou legendou (pri slovensky písanom texte sa odporúčajú legendy aj v angličtine) a pripojte všetky obrázky ako osobitné prílohy.
Tabuľky by, pokiaľ je to možné, mali obsahovať bunky, nie text zarovnaný pomocou medzier alebo tabulátorov. Pre popisky obrázkov používajte písmo typu Arial. Veľkosť písma by po upravení na správnu veľkosť obrázku mala byť 8 alebo viac bodov. Obrázky nesmú presiahnuť šírku 124 mm a výšku 193 mm. Pri odkazovaní sa na obrázky a tabuľky v texte používajte skratky Tab. 1, Fig. 1, Obr. 1, Figs. 1−4, Obr. 1 − 4 (rozsah hodnôt uvádzajte v slovenskom texte s medzerami, v anglickom bez medzier). Mená rodov, druhov, vnútrodruhových taxónov a syntaxónov píšte kurzívou. Skratky herbárov uvádzajte podľa Index Herbariorum: http://sweetgum.nybg.org/science/ih/. Názvy časopisov v zozname literatúry neskracujte, resp. ich zjednoťte podľa: http://journalseek.net/.
References / Literatúra Pilát, A., Dermek, A., 1974: Hríbovité huby. Veda, Bratislava.Oliveira, M., Ribeiro, H., Delgado, J. L., Abreu, I., 2009: Seasonal and intradiurnal variation of allergenic
fungal spores in urban and rural areas of the North of Portugal. Aerobiologia, 25: 85−98.Holec, J., 2008: Gymnopilus P. Karst. In: Knudsen, H., Vesterholt, J. (eds.), Funga Nordica, p. 857−861,
Nordsvamp, Copenhagen.Thiers, B., 2020, continuously updated: Index Herbariorum: a global directory of public herbaria and
associated staff. New York Botanical Garden’s Virtual Herbarium. Available from: http://sweetgum.nybg.org/ih (accessed: 2 Jan 2020).
Mikuš, M., 1999: Rastlinstvo Dostálovej doliny. MSc thesis depon. in Comenius University in Bratislava.
References in the text / Citácie v texteMájovský (1965), Mičieta, Murín (1998), Harman et al. (1992), (Knasmüller 1984), (Pišút, Lackovičová 1998), (Ma et al. 1996), Anonymus (2005), Ryvarden (on line).

contents
Peľová sezóna 2017 na území BratislavyPollen season 2017 in the Bratislava areaDušička, J., Ščevková, J. ...................................................................................................... 3
Let’s go to the field. Botanical excursion 4Poďme spolu do terénu. Botanická exkurzia 4Mišíková, K., Dušička, J., Miškovic, J., Michalová, M., Jančovičová, S. ........................ 9
Bryophyte flora of the Stupava town and its surroundings (Western Slovakia) 2. Bryophytes on meadows in agricultural landscapeBryoflóra Stupavy a jej okolia (Západné Slovensko) 2. Machorasty lúk v agrikultúrnej krajineMišíková, K., Mišík, M. . ...................................................................................................... 25
Leaf evolution and classification 3. GymnospermopsidaMorfogenéza a klasifikácia listov 3. Nahosemenné rastlinyHrabovský, M. ..................................................................................................................... 37
Ruderal vegetation of the class Artemisietea vulgaris in Bratislava city 1. Alliance Convolvulo arvensis-Agropyrion repentisRuderálna vegetácia triedy Artemisietea vulgaris v Bratislave 1. Zväz Convolvulo arvensis-Agropyrion repentisRendeková, A., Zahradníková, E., Miškovic, J., Michalová, M., Mičieta, K. .............. 73
Spomienka na kolegyňu, pracovníčku Katedry botaniky Prírodovedeckej fakulty Univerzity Komenského v BratislaveCommemoration of a colleague, a staff member of the Department of Botany, Faculty of Natural Sciences, Comenius University in BratislavaRNDr. Anna Uhríková 18. 2. 1939 – 14. 1. 2021Mičieta, K., Javorčíková, D., Schwarzová, T., Mišíková, K., Dušička, J., Kováčik, Ľ.,Jančovičová, S. .................................................................................................................... 89
ISSN 2729-7330
Related Documents











