Unique amnesic shellfish toxin composition found in the South East Asian diatom Nitzschia navis-varingica Marc Lawrence J. Romero a , Yuichi Kotaki b, *, Nina Lundholm c , Hikmah Thoha d , Hisao Ogawa b , Juan R. Relox a , Ryuta Terada e , Shigenobu Takeda f , Yoshinobu Takata b , Koichi Haraguchi g , Tetsuya Endo h , Po-Teen Lim i , Masaaki Kodama b , Yasuwo Fukuyo j a Bureau of Fisheries and Aquatic Resources, 860 Arcadia Building, Quezon Avenue, Quezon City 1100, Philippines b School of Marine Biosciences, Kitasato University, Sanriku, Ofunato, Iwate 022-0101, Japan c The Natural History Museum of Denmark, Soelvgade 83 Opg S, DK-1307 Kbh K, Denmark d Center for Oceanographic Research-LIPI, Jl. Pasir Putih No. 1, Ancol Timur, Jakarta 11048, Indonesia e Faculty of Fisheries, Kagoshima University, Shimoarata, Kagoshima 890-0056, Japan f Faculty of Fisheries, Nagasaki University, Bunkyo, Nagasaki 852-8521, Japan g Daiichi College of Pharmaceutical Sciences, Tamagawa, Minami, Fukuoka 815-8511, Japan h Faculty of Pharmaceutical Sciences, Health Sciences University of Hokkaido, Tobetsu, Ishikari, Hokkaido 061-0293, Japan i Faculty of Resource Science and Technology, University of Malaysia Sarawak, Kota Samarahan, Sarawak 94300, Malaysia j Asian Natural Environmental Science Center, The University of Tokyo, Yayoi, Bunkyo, Tokyo 113-8657, Japan 1. Introduction Domoic acid (DA) and its derivatives (e.g., isodomoic acids A, B, C, G, and H) (Maeda et al., 1986; Zaman et al., 1997) are insecticidal agents first isolated from the macro-alga Chondria armata (Takemoto & Daigo, 1958). Later, DA was identified as the toxin responsible for amnesic shellfish poisoning (ASP) (Wright et al., 1989) followed by identification of the isomers isodomoic acids D, E, and F as minor components (Wright et al., 1990), and the causative organism was traced and identified as the diatom Pseudo-nitzschia multiseries (Bates et al., 1989). A number of DA- producing Pseudo-nitzschia spp. have been found in subsequent searches primarily in temperate areas, although the toxin level varied widely (Garrison et al., 1992; Martin et al., 1990; Lundholm et al., 1994; Rhodes et al., 1996; Rhodes, 1998; Orsini et al., 2002). Among them, three Pseudo-nitzschia species (P. multiseries, P. australis, and P. seriata) are known to produce high levels of DA (Bates et al., 1989; Garrison et al., 1992; Lundholm et al., 1994; Rhodes et al., 1996). During a more recent search for DA-producing diatoms in tropical waters, a major DA-producing benthic diatom was isolated from Vietnam (Kotaki et al., 2000). This organism was identified as a new species, Nitzschia navis-varingica (Lundholm and Moestrup, 2000; Bates, 2000). This species has also been isolated from brackish water areas in the Philippines and Japan (Kotaki et al., 2004). Recently, some of the Philippine strains of N. navis-varingica Harmful Algae 10 (2011) 456–462 ARTICLE INFO Article history: Received 23 September 2010 Received in revised form 22 February 2011 Accepted 23 February 2011 Available online 1 March 2011 Keywords: Amnesic shellfish poisoning Domoic acid Estuary Isodomoic acid Nitzschia navis-varingica Pennate diatom ABSTRACT Nitzschia navis-varingica is a diatom that is known to produce significant levels of amnesic shellfish poisoning (ASP) toxins. A total of 33 N. navis-varingica strains were isolated from four brackish water localities in the Philippines and Indonesia, and cultured to characterize the toxins produced. The isolates were analyzed for domoic acid (DA) and isodomoic acids A (IA) and B (IB) by HPLC with fluorescence detection. Two toxin composition types were detected that have not been previously described: strains producing only IB and strains producing DA–IA–IB. These two types were isolated from two different localities. Eighteen strains were isolated from the Philippines (northern Luzon Island). Among them, 10 isolates from Alaminos produced only IB with an average toxin content of 3.05 pg cell 1 , seven isolates from Bulacan produced DA and IB with average toxin contents of 0.68 pg cell 1 and 1.18 pg cell 1 , respectively. One isolate from Cavite produced DA, IA, and IB with a toxin content of 0.58, 0.20, and 0.92 pg cell 1 , respectively. Fifteen isolates from Indonesia (Bone, South Sulawesi) produced only DA (four isolates) or DA with trace amounts of IB (eleven isolates), with an average toxin content of 2.38 pg cell 1 and 0.06 pg cell 1 , respectively. Sub-strains were established from strains producing either of the three toxin types: IB, DA–IA–IB, and DA-trace IB. Results showed that the toxin composition type was the same for parent and sub-strains, indicating that the toxin composition is a stable character for a strain. ß 2011 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +81 192 44 2121; fax: +81 192 44 2125. E-mail address: [email protected] (Y. Kotaki). Contents lists available at ScienceDirect Harmful Algae journal homepage: www.elsevier.com/locate/hal 1568-9883/$ – see front matter ß 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.hal.2011.02.006

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Harmful Algae 10 (2011) 456–462
Unique amnesic shellfish toxin composition found in the South East Asiandiatom Nitzschia navis-varingica
Marc Lawrence J. Romero a, Yuichi Kotaki b,*, Nina Lundholm c, Hikmah Thoha d, Hisao Ogawa b,Juan R. Relox a, Ryuta Terada e, Shigenobu Takeda f, Yoshinobu Takata b, Koichi Haraguchi g,Tetsuya Endo h, Po-Teen Lim i, Masaaki Kodama b, Yasuwo Fukuyo j
a Bureau of Fisheries and Aquatic Resources, 860 Arcadia Building, Quezon Avenue, Quezon City 1100, Philippinesb School of Marine Biosciences, Kitasato University, Sanriku, Ofunato, Iwate 022-0101, Japanc The Natural History Museum of Denmark, Soelvgade 83 Opg S, DK-1307 Kbh K, Denmarkd Center for Oceanographic Research-LIPI, Jl. Pasir Putih No. 1, Ancol Timur, Jakarta 11048, Indonesiae Faculty of Fisheries, Kagoshima University, Shimoarata, Kagoshima 890-0056, Japanf Faculty of Fisheries, Nagasaki University, Bunkyo, Nagasaki 852-8521, Japang Daiichi College of Pharmaceutical Sciences, Tamagawa, Minami, Fukuoka 815-8511, Japanh Faculty of Pharmaceutical Sciences, Health Sciences University of Hokkaido, Tobetsu, Ishikari, Hokkaido 061-0293, Japani Faculty of Resource Science and Technology, University of Malaysia Sarawak, Kota Samarahan, Sarawak 94300, Malaysiaj Asian Natural Environmental Science Center, The University of Tokyo, Yayoi, Bunkyo, Tokyo 113-8657, Japan
A R T I C L E I N F O
Article history:
Received 23 September 2010
Received in revised form 22 February 2011
Accepted 23 February 2011
Available online 1 March 2011
Keywords:
Amnesic shellfish poisoning
Domoic acid
Estuary
Isodomoic acid
Nitzschia navis-varingica
Pennate diatom
A B S T R A C T
Nitzschia navis-varingica is a diatom that is known to produce significant levels of amnesic shellfish
poisoning (ASP) toxins. A total of 33 N. navis-varingica strains were isolated from four brackish water
localities in the Philippines and Indonesia, and cultured to characterize the toxins produced. The isolates
were analyzed for domoic acid (DA) and isodomoic acids A (IA) and B (IB) by HPLC with fluorescence
detection. Two toxin composition types were detected that have not been previously described: strains
producing only IB and strains producing DA–IA–IB. These two types were isolated from two different
localities. Eighteen strains were isolated from the Philippines (northern Luzon Island). Among them, 10
isolates from Alaminos produced only IB with an average toxin content of 3.05 pg cell�1, seven isolates
from Bulacan produced DA and IB with average toxin contents of 0.68 pg cell�1 and 1.18 pg cell�1,
respectively. One isolate from Cavite produced DA, IA, and IB with a toxin content of 0.58, 0.20, and
0.92 pg cell�1, respectively. Fifteen isolates from Indonesia (Bone, South Sulawesi) produced only DA
(four isolates) or DA with trace amounts of IB (eleven isolates), with an average toxin content of
2.38 pg cell�1 and 0.06 pg cell�1, respectively. Sub-strains were established from strains producing
either of the three toxin types: IB, DA–IA–IB, and DA-trace IB. Results showed that the toxin composition
type was the same for parent and sub-strains, indicating that the toxin composition is a stable character
for a strain.
� 2011 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Harmful Algae
journal homepage: www.e lsev ier .com/ locate /ha l
1. Introduction
Domoic acid (DA) and its derivatives (e.g., isodomoic acids A, B,C, G, and H) (Maeda et al., 1986; Zaman et al., 1997) are insecticidalagents first isolated from the macro-alga Chondria armata
(Takemoto & Daigo, 1958). Later, DA was identified as the toxinresponsible for amnesic shellfish poisoning (ASP) (Wright et al.,1989) followed by identification of the isomers isodomoic acids D,E, and F as minor components (Wright et al., 1990), and thecausative organism was traced and identified as the diatomPseudo-nitzschia multiseries (Bates et al., 1989). A number of DA-
* Corresponding author. Tel.: +81 192 44 2121; fax: +81 192 44 2125.
E-mail address: [email protected] (Y. Kotaki).
1568-9883/$ – see front matter � 2011 Elsevier B.V. All rights reserved.
doi:10.1016/j.hal.2011.02.006
producing Pseudo-nitzschia spp. have been found in subsequentsearches primarily in temperate areas, although the toxin levelvaried widely (Garrison et al., 1992; Martin et al., 1990; Lundholmet al., 1994; Rhodes et al., 1996; Rhodes, 1998; Orsini et al., 2002).Among them, three Pseudo-nitzschia species (P. multiseries, P.
australis, and P. seriata) are known to produce high levels of DA(Bates et al., 1989; Garrison et al., 1992; Lundholm et al., 1994;Rhodes et al., 1996).
During a more recent search for DA-producing diatoms intropical waters, a major DA-producing benthic diatom was isolatedfrom Vietnam (Kotaki et al., 2000). This organism was identified asa new species, Nitzschia navis-varingica (Lundholm and Moestrup,2000; Bates, 2000). This species has also been isolated frombrackish water areas in the Philippines and Japan (Kotaki et al.,2004). Recently, some of the Philippine strains of N. navis-varingica

Fig. 1. Structure of domoic acid and isodomoic acids A and B.
Fig. 2. Sampling areas in the Philippines, Indonesia, and Malaysia.
M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462 457
were found to produce not only DA but also isodomoic acids A (IA)and B (IB) as major toxin components (Kotaki et al., 2005) (Fig. 1).ASP toxin production was then re-investigated using the strains ofN. navis-varingica from the Philippines and a few other Asiancountries (Japan, Vietnam, and Thailand) and three types of ASPtoxin composition (DA, DA–IB, IA–IB) were detected. The mostabundant toxin composition type was a combination of DA and IB(DA–IB), with the ratio of IB ranging from trace to ca. 80%. This typewas found in strains from Japan, Thailand, Vietnam, and thePhilippines (Kotaki, 2008; Kotaki et al., 2008; Romero et al., 2008).Isolates from restricted areas in the northern part of the Philippineshad another toxin composition (IA and IB) (Bajarias et al., 2006;Kotaki, 2008; Kotaki et al., 2008). However, the composition andgeographical distribution of toxins from southernmost Asia, suchas Indonesia and Malaysia, and most of the northern part of thePhilippines have not been studied.
We report here the results of a toxin screening of 33 N. navis-
varingica strains from South Sulawesi in Indonesia and threelocalities in the Philippines. We document two new toxincomposition types (IB and DA–IA–IB) from two different localitiesin the Philippines, although IA and IB might not pose a major healthrisk because of their lower toxicity than DA (Munday et al., 2008).
2. Materials and methods
2.1. Isolation and identification of N. navis-varingica
Samples were collected from the surface of mangrove roots orpalm tree leaves hanging down into the water in estuarine areasusing a handy scoop net (20 mm). Sampling was conducted in threeareas of Luzon Island, Philippines, including several brackish sites,in December 2007 and December 2008: Alaminos in Pangasinan,and Bulacan and Cavite in Manila Bay (Fig. 2). Sampling was alsoconducted in several other brackish sites: Panyula, Bone in SouthSulawesi (July 2008), Jakarta Bay (March 2009), Lampung Bay(March 2009), and Sangihe Island (May 2009), all in Indonesia, andfinally in Kota Kinabalu, Malaysia (May 2008) (Fig. 2).
Crude cultures were prepared by mixing aliquots (0.2 mL or1.0 mL) of net sample with 30 mL of f/2 medium in 50 mL tissueculture flasks (Greiner bio-one, Tokyo, Japan), and incubating themat 25 8C under an irradiance level of 60 mmol photons m�2 s�1 witha 16:8 h light:dark cycle. f/2 medium was prepared (Guillard, 1983)using sea water diluted with distilled water to a salinity of ca. 27 psu.Uni-algal cultures of N. navis-varingica were established from crudecultures by capillary washing of single cells under a light microscope(LM) according to the morphological characteristics of N. navis-
varingica (Lundholm and Moestrup, 2000; Kotaki et al., 2005). The
cells possess two yellow-brown chloroplasts at each end of the celland are lanceolate in valve view. Cells are 38–110 mm long and 9–11 mm wide. In girdle view, the cells are rectangular and slightlyindented at the middle. Most cells form ribbon-shaped colonieswhile growing (Kotaki et al., 2004). Experience have shown, thatusing these light microscopical characters allow us to isolate strainsthat have a high likelihood of being N. navis-varingica. Onerepresentative strain from each collection site was furtherinvestigated morphologically to confirm the identification usingtransmission electron microscopy (TEM) according to the method ofLundholm and Moestrup (2000). Subsamples were fixed in formalin(final concentration 3.7%). Representative strains identified usingTEM were PBULD 07-4 (Bulacan, Manila Bay), PCAVA 07-2 (Cavite,Manila Bay), PALMC 07-1 (Alaminos, Pangasinan) (all from thePhilippines), and IBNA 08-3, IBNB 08-4, and IBNC 08-3 (Panyula,Bone in South Sulawesi, all from Indonesia).
2.2. Analysis of ASP toxins in N. navis-varingica cultures
Isolates were cultured in 30 mL f/2 medium in 50 mL tissueculture tubes under the same culture conditions described inSection 2.1. The cultures were harvested after a 3-week cultureperiod (mid-stationary growth phase). A 3 mL aliquot of wholeculture was retrieved, boiled for 5 min and then centrifuged afterbeing cooled to room temperature (Bajarias et al., 2006). A 2 mLaliquot was also retrieved for determining the cell density (Kotakiet al., 2000). Toxin profile analysis was done for DA, IA, and IB usingHPLC-fluorescence analysis with pre-column derivatization using

M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462458
9-fluorenylmethylchloroformate (FMOC-Cl) according to theslightly modified method of Pocklington et al. (1990) in which aDevelosil ODS-5 column (4.6 � 250 mm, Nomura, Seto, Aichi,Japan) and a mobile phase of 40% acetonitrile in 20 mM phosphatebuffer (pH 2.5) was used. The cellular toxin content was expressedon a per cell basis (pg cell�1), which is calculated by dividing thetotal amount of each toxin (ng mL�1) in the entire culture by thecorresponding density of cells in the culture. The number of cellswas determined using 5 mL of culture under LM in a Sedgewick-Rafter counting chamber. Samples were counted in triplicate andthe average cell number was used for the cellular toxin contentcalculation. Standard toxins were purified from a mass culture of N.
navis-varingica that produces both IA and IB (for IA and IBstandards) and an extract of the red alga C. armata that produceshigh amount of DA and small/trace amount of IA, IB, 50-epi-DA, etc.(for DA standard) collected in Kagoshima Prefecture, Japan. Thesewere calibrated using commercial Canadian standard toxin (DACS-1D, DA) or purified toxins (IA and IB) (Kotaki et al., 2005).
Confirmation of toxin production of the representative strains wasperformed by LC–MS/MS using multiple reaction monitoring (MRM)(Takata et al., 2009). A 1 L mass culture of each strain was filtered(mixed cellulose ester 3 mm pore size filters, Advantec, Tokyo, Japan)at the end of the three-week culture period and the cells wereextracted by boiling followed by ultrafiltration (10,000 Da cut-off).LC–MS was performed on an HP1100 LC system (Agilent Technolo-gies, Santa Clara, CA) coupled to an Applied Biosystems/MDS Sciex API2000 triple quadruple mass spectrometer (Applied Biosystems, FosterCity, CA). LC separation was performed using a slight modification ofthe method described in the Canadian standard DACS-1D manual. AWakosil Navi C18-5 column (2 mm� 250 mm; Wako, Tokyo, Japan)was employed, and analytes were eluted over 20 min using a linear
Fig. 3. HPLC-fluorescence analyses of ASP toxins from representative Nitzschia navis-varin
containing IA, IB and DA (8.7, 9.2, and 17.5 ng/mL) analyzed at the same time as the resp
PALMC 08-1 strain isolated from Alaminos, Pangasinan, (B) PBULD 07-5 isolated from Bu
Island, the Philippines and (D) IBNB 08-3 isolated from Panyula, Bone, South Sulawesi, In
derived from the FMOC-Cl reagent. The amount of each derivative often varies due to con
and/or due to reaction conditions (Pocklington et al., 1990), *a: Side-products and/or rea
the culture (e.g., neutral amino acids) that react with the FMOC-Cl reagent. Analytical co
acetonitrile in 20 mM phosphate buffer (pH 2.5); column temperature, 55 8C; flow rate
gradient from 10% acetonitrile in 0.1% formic acid to 20% acetonitrilein 0.1% formic acid with a flow rate of 0.2 mL min�1. The MS analysiswas achieved by MRM with a turboion-spray interface in positive ionmode. Three MRM transitions from the protonated DA ion weremonitored (m/z 312–266, m/z 312–248, and m/z 312–161). Standardtoxin was prepared from an extract of C. armata and calibrated usingcommercial Canadian standard toxin (DACS-1D) or purified toxins (IAand IB) (Kotaki et al., 2005).
2.3. Toxin composition of N. navis-varingica sub-strains
Sub-strains were prepared by capillary washing of single cells oftwo representative parental Philippine strains: PALMC 07-1(Alaminos, IB type), PCAVA 07-2 (Cavite, DA–IA–IB type), andone Indonesian parental strain: IBNB 08-2 (Panyula, Bone, DA-trace IB type), after having been pre-cultured for one week underthe same conditions as the batch cultures in Section 2.1. Sub-strains were cultured for three weeks as described in Section 2.1,harvested and analyzed for toxin production as described inSection 2.2. The toxin composition was then compared with that ofthe parental strain. Parental strains were maintained for more thanhalf year by inoculating aliquots of old culture into new f/2medium every month followed by culturing for one week underthe same conditions in Section 2.1.
3. Results
3.1. Isolation and identification of N. navis-varingica
Ten isolates of N. navis-varingica were obtained from Alaminos(PALM 07/08 strains, one in 2007 and nine in 2008), seven were
gica strains. Upper chromatogram of (A), (B), (C) and (D); standard toxins DAKS 04-1
ective sample. Lower chromatogram of (A), (B), (C) and (D); sample extracts of (A)
lacan, Manila Bay, (C) PCAVA 07-2 isolated from Cavite, Manila Bay, all in the Luzon
donesia. DA, domoic acid; IA, isodomoic acid A; IB, isodomoic acid B. *Side-products
tamination with compounds (e.g., other amino acids) that compete with ASP toxins
ction products derived from contaminants in the medium and other compounds in
nditions: Develosil ODS-5 column (4.6 mm � 250 mm, Nomura); mobile phase, 40%
, 1 mL min�1; fluorescence detection, Ex. = 264 nm, Em. = 313 nm.
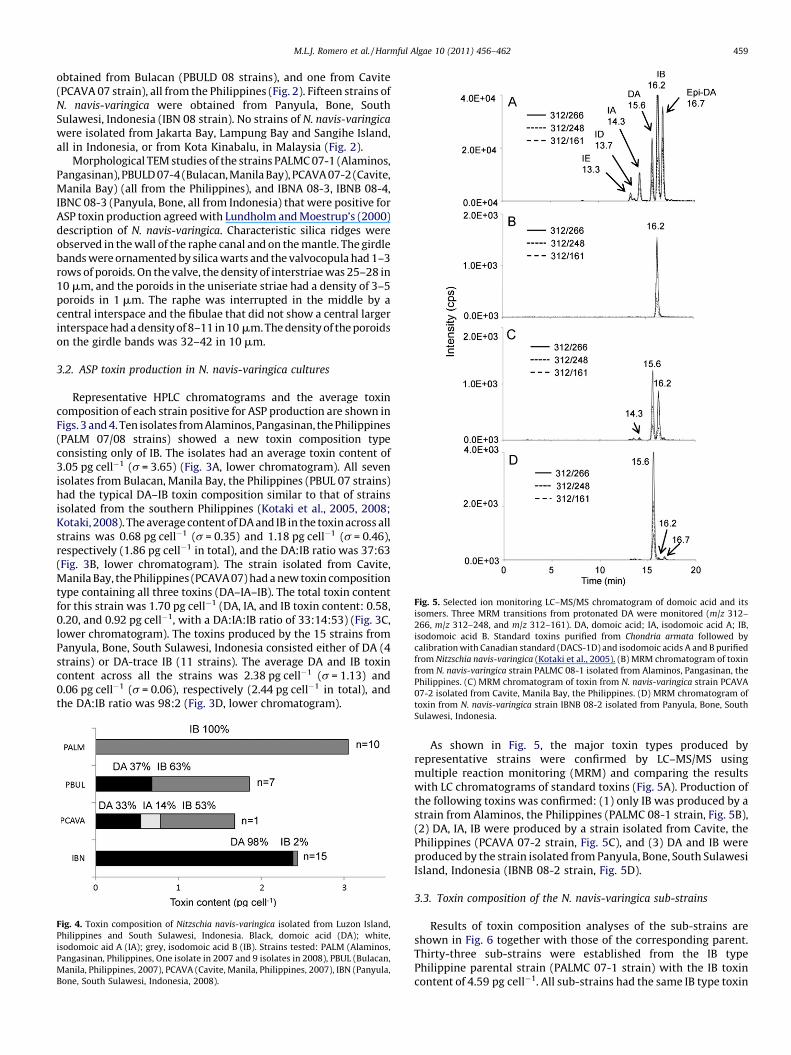
Fig. 5. Selected ion monitoring LC–MS/MS chromatogram of domoic acid and its
isomers. Three MRM transitions from protonated DA were monitored (m/z 312–
266, m/z 312–248, and m/z 312–161). DA, domoic acid; IA, isodomoic acid A; IB,
isodomoic acid B. Standard toxins purified from Chondria armata followed by
calibration with Canadian standard (DACS-1D) and isodomoic acids A and B purified
from Nitzschia navis-varingica (Kotaki et al., 2005). (B) MRM chromatogram of toxin
from N. navis-varingica strain PALMC 08-1 isolated from Alaminos, Pangasinan, the
Philippines. (C) MRM chromatogram of toxin from N. navis-varingica strain PCAVA
07-2 isolated from Cavite, Manila Bay, the Philippines. (D) MRM chromatogram of
toxin from N. navis-varingica strain IBNB 08-2 isolated from Panyula, Bone, South
M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462 459
obtained from Bulacan (PBULD 08 strains), and one from Cavite(PCAVA 07 strain), all from the Philippines (Fig. 2). Fifteen strains ofN. navis-varingica were obtained from Panyula, Bone, SouthSulawesi, Indonesia (IBN 08 strain). No strains of N. navis-varingica
were isolated from Jakarta Bay, Lampung Bay and Sangihe Island,all in Indonesia, or from Kota Kinabalu, in Malaysia (Fig. 2).
Morphological TEM studies of the strains PALMC 07-1 (Alaminos,Pangasinan), PBULD 07-4 (Bulacan, Manila Bay), PCAVA 07-2 (Cavite,Manila Bay) (all from the Philippines), and IBNA 08-3, IBNB 08-4,IBNC 08-3 (Panyula, Bone, all from Indonesia) that were positive forASP toxin production agreed with Lundholm and Moestrup’s (2000)description of N. navis-varingica. Characteristic silica ridges wereobserved in the wall of the raphe canal and on the mantle. The girdlebands were ornamented by silica warts and the valvocopula had 1–3rows of poroids. On the valve, the density of interstriae was 25–28 in10 mm, and the poroids in the uniseriate striae had a density of 3–5poroids in 1 mm. The raphe was interrupted in the middle by acentral interspace and the fibulae that did not show a central largerinterspace had a density of 8–11 in 10 mm. The density of the poroidson the girdle bands was 32–42 in 10 mm.
3.2. ASP toxin production in N. navis-varingica cultures
Representative HPLC chromatograms and the average toxincomposition of each strain positive for ASP production are shown inFigs. 3 and 4. Ten isolates from Alaminos, Pangasinan, the Philippines(PALM 07/08 strains) showed a new toxin composition typeconsisting only of IB. The isolates had an average toxin content of3.05 pg cell�1 (s = 3.65) (Fig. 3A, lower chromatogram). All sevenisolates from Bulacan, Manila Bay, the Philippines (PBUL 07 strains)had the typical DA–IB toxin composition similar to that of strainsisolated from the southern Philippines (Kotaki et al., 2005, 2008;Kotaki, 2008). The average content of DA and IB in the toxin across allstrains was 0.68 pg cell�1 (s = 0.35) and 1.18 pg cell�1 (s = 0.46),respectively (1.86 pg cell�1 in total), and the DA:IB ratio was 37:63(Fig. 3B, lower chromatogram). The strain isolated from Cavite,Manila Bay, the Philippines (PCAVA 07) had a new toxin compositiontype containing all three toxins (DA–IA–IB). The total toxin contentfor this strain was 1.70 pg cell�1 (DA, IA, and IB toxin content: 0.58,0.20, and 0.92 pg cell�1, with a DA:IA:IB ratio of 33:14:53) (Fig. 3C,lower chromatogram). The toxins produced by the 15 strains fromPanyula, Bone, South Sulawesi, Indonesia consisted either of DA (4strains) or DA-trace IB (11 strains). The average DA and IB toxincontent across all the strains was 2.38 pg cell�1 (s = 1.13) and0.06 pg cell�1 (s = 0.06), respectively (2.44 pg cell�1 in total), andthe DA:IB ratio was 98:2 (Fig. 3D, lower chromatogram).
Fig. 4. Toxin composition of Nitzschia navis-varingica isolated from Luzon Island,
Philippines and South Sulawesi, Indonesia. Black, domoic acid (DA); white,
isodomoic aid A (IA); grey, isodomoic acid B (IB). Strains tested: PALM (Alaminos,
Pangasinan, Philippines, One isolate in 2007 and 9 isolates in 2008), PBUL (Bulacan,
Manila, Philippines, 2007), PCAVA (Cavite, Manila, Philippines, 2007), IBN (Panyula,
Bone, South Sulawesi, Indonesia, 2008).
Sulawesi, Indonesia.
As shown in Fig. 5, the major toxin types produced byrepresentative strains were confirmed by LC–MS/MS usingmultiple reaction monitoring (MRM) and comparing the resultswith LC chromatograms of standard toxins (Fig. 5A). Production ofthe following toxins was confirmed: (1) only IB was produced by astrain from Alaminos, the Philippines (PALMC 08-1 strain, Fig. 5B),(2) DA, IA, IB were produced by a strain isolated from Cavite, thePhilippines (PCAVA 07-2 strain, Fig. 5C), and (3) DA and IB wereproduced by the strain isolated from Panyula, Bone, South SulawesiIsland, Indonesia (IBNB 08-2 strain, Fig. 5D).
3.3. Toxin composition of the N. navis-varingica sub-strains
Results of toxin composition analyses of the sub-strains areshown in Fig. 6 together with those of the corresponding parent.Thirty-three sub-strains were established from the IB typePhilippine parental strain (PALMC 07-1 strain) with the IB toxincontent of 4.59 pg cell�1. All sub-strains had the same IB type toxin

Fig. 6. Toxin composition of the sub-strains of Nitzschia navis-varingica compared
with the respective parental strain. Black, domoic acid (DA); white, isodomoic aid
(IA); grey, isodomoic acid B (IB). Parental strains used: PALM 07-1 (Alaminos,
Pangasinan, Philippines, 2007), PCAV 07-2 (Cavite, Manila, Philippines, 2007), IBN
08-2 (Bone, South Sulawesi, Indonesia, 2008).
M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462460
composition. The toxin composition was 100% IB with an averagetoxin content of 4.63 pg cell�1 (s = 1.28), entirely lacking IA and DA(Fig. 6). Ten sub-strains were established from the DA–IA–IB typePhilippine parental strain (PCAVA 07-2 strain) which had a totaltoxin content of 1.70 pg cell�1 with a DA:IA:IB toxin ratio of33:14:53. The toxin in all the sub-strains was composed of DA–IA–IB, similar to the parental strain, with an average toxin content of0.33 pg cell�1 (s = 0.30), 0.19 pg cell�1 (s = 0.15), and 0.60 pg cell�1
(s = 0.45), respectively, and with a DA:IA:IB toxin ratio of 29:17:54(Fig. 6). Eleven sub-strains were established from the Indonesianparental strain having a DA–IB type toxin composition (IBNB 08-2strain) showing the total toxin content of 1.55 pg cell�1 with a DA:IBtoxin ratio of 96:4. All sub-strains had the same toxin composition asthe parental strain (DA–IB). The average DA and IB toxin content was1.09 pg cell�1 (s = 0.64) and 0.07 pg cell�1 (s = 0.02), respectively,with an average DA:IB ratio of 94:6 (Fig. 6).
4. Discussion
We confirmed that N. navis-varingica can be found in estuarineareas of the northern part of Luzon Island, the Philippines and inSouth Sulawesi, Indonesia (Fig. 2). Interestingly, all 10 strains
Fig. 7. Distribution and toxin composition of Nitzschia navis-varingica in Asian waters. S
Romero et al. (2008) and Kotaki (2008). Black, domoic acid (DA); white, isodomoic aci
isolated from Alaminos, Luzon Island had a toxin composition onlyof IB without even trace amounts of any other isomers. The singleisolate from Cavite, Luzon Island had the toxin content composedof DA–IA–IB. Neither of these two toxin composition types hasbeen detected before in any diatom as major toxin components,and is described for the first time in the present study. Otherisolates from Bulacan, Luzon Island, the Philippines and Bone,South Sulawesi, Indonesia showed the major toxin composition ofDA–IB (including only DA) similar to N. navis-varingica from areasother than Luzon Island (Kotaki et al., 2005, 2008; Bajarias et al.,2006; Kotaki, 2008). Though total toxin content of each isolate hadrather high variability, toxin composition was the same withineach toxin composition type. The variability might come from adifference of culture conditions (e.g. difference of co-existingbacteria, difference of light intensity due to the location of tissueculture tube, etc.).
In this study, N. navis-varingica samples were collected mainlyin dry season. It is thus unknown whether there is a seasonaldifference in the distribution and toxin compositions of N. navis-
varingica. Furthermore, we have not explored the possibility ofother benthic diatoms producing ASP toxins. However, in SakariEstuary, near our college, in Japan, N. navis-varingica could beisolated throughout the year (even in winter season, data notshown), all strains showing the same toxin composition (DA–IB).Our data indicate that N. navis-varingica might show the sametoxin composition during all seasons in tropical areas. Detailedstudies in different seasons at the same locality will be needed tofurther explore the seasonality of the distribution and toxincomposition of N. navis-varingica in tropical areas.
Toxin compositions of N. navis-varingica isolates are summa-rized in Fig. 7 together with previously published results (Kotakiet al., 2005, 2008; Bajarias et al., 2006; Romero et al., 2008; Kotaki,2008). The strains isolated from Panyula, Bone, South Sulawesi,Indonesia showed a DA-trace IB toxin composition type that wassimilar to those of strains isolated from Northern Japan (Kotakiet al., 2008; Kotaki, 2008) and Thailand (Romero et al., 2008), incontrast to the typical DA–IB type strains from Okinawa,southernmost Japan, strains from Vietnam, and some strains fromthe southern Philippines which had a higher proportion of IB (morethan ca. 20%) (Kotaki et al., 2005, 2008; Kotaki, 2008). Our resultsshow that toxin composition is not related to latitude, in contraryto what has previously been suggested, namely that the proportion
ummary of the present studies and Kotaki et al. (2005, 2008), Bajarias et al. (2006),
d A (IA); grey, isodomoic acid B (IB). *The present study.

M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462 461
of IB in the DA–IB type toxin should be higher in lower latitudes(Kotaki et al., 2005, 2008; Kotaki, 2008).
Sub-strains were established from strains representative ofthree different toxin composition types (IB, DA–IA–IB, and DA-trace IB), followed by toxin analysis. All of the sub-strains had thesame toxin composition as the parental strains. As the parentalstrains were maintained for more than half year until use, aboveresults demonstrate that toxin composition is most likely a stablecharacter, although the exact ratio of each toxin component varieswithin each toxin composition type (Fig. 6). The stability of thecharacter is also supported by supplemental batch cultureexperiments that were done using PALMC 07-1 (IB type) andPCAVA 07-2 (DA–IA–IB type) strains for up to 36 days resulting inconfirmation that they maintained the same toxin profile typeduring the entire experimental period (PALMC strain; toxincontent of IB increased up to 4.3 pg cell�1 at 36 day and PCAVAstrain; total toxin content of DA–IA–IB showed maximum of5.0 pg cell�1 at 28 day with the DA:IA:IB ratio of 37:14:49 anddecreased to 4.2 pg cell�1 at 36 day with the DA:IA:IB ratio of34:13:53).
As shown in Fig. 6, toxin content per cell between parent andsub-strain of PALMC 07-1 was almost the same (ca. 4.6 pg cell�1)but in case of PCAVA 07-2 (DA–IA–IB type) and IBNB 08-3 (DA–IBtype), average toxin content per cell of the sub-strains was a littlelower than those of the parents. However, these differences areincluded in the variability of sub-strains (PCAVA; s = 0.87, IBN;s = 0.66).
Trace amounts of other isomers (e.g., isodomoic acids D and Etogether with DA, IA, and IB in the PCAVA 07 strain (Fig. 5C) and 50-epi-DA together with DA and IB in the IBNB 08-2 strain (Fig. 5D)were detected in LC–MS/MS chromatograms of toxins fromrepresentative strains. However, the presence of these additionalminor isomers was not confirmed in HPLC chromatograms withfluorescence detection because of low concentrations using ananalysis method to extract samples without concentration(Pocklington et al., 1990; Kotaki et al., 2004). It is uncertainwhether these minor isomers were artifacts produced by UV orheating (Wright et al., 1990; Quilliam, 2003) or whether they werebio-synthesized. Nevertheless, we did confirm that DA, IA, and IBare the major components of N. navis-varingica toxin, and that thetoxin composition is more complex than previously reported(Kotaki et al., 2005, 2006; Bajarias et al., 2006; Kotaki, 2008). Thepresence of isodomoic acid C (IC) simultaneously with DA wasrecently reported in toxin of P. australis and shellfish that have fedon P. australis (Holland et al., 2003, 2005; Rhodes et al., 2003). Wedid not determine whether IC was present in this study due to alack of standard toxin for comparison. However, it is unlikely thatIC constitutes a major component of the toxin profile because noother apparent peak was observed in any of the HPLC and LC–MS/MS chromatograms.
Interestingly, only N. navis-varingica isolates from northernLuzon Island had a toxin composition different from the major DA–IB type. In total, three different toxin profiles have been confirmedin isolates from Luzon Island: IA–IB (Kotaki et al., 2005, 2008;Bajarias et al., 2006), IB, and DA–IA–IB (present study) (Fig. 7). Thestrains are morphologically similar to those found in other/closelocalities (Haiphong in Vietnam, Okinawa in Japan, the southernPhilippines and Bulacan Estuary, Manila Bay in the Philippines).
One of the strains showing DA–IB toxin composition has beenshown to change its toxin composition to the IA–IB type whengrown in axenic culture and it returned to the original DA–IB typewhen the culture medium was replaced with the cell-free but non-axenic medium of the parental strain (Kotaki et al., 2008). Thissuggests that bacteria might play a role in controlling the toxincomposition, but the mechanism is presently unknown. Onepossibility is that some kinds of bacteria might enhance DA
production and/or block IA production and/or consume IA. Suchbacteria might be absent at sites from which strains having a toxincomposition that includes IA were isolated. Five types of ASP toxincompositions have been identified to date, namely DA, DA–IB, IA–IB, IB, and DA–IA–IB, although the difference between DA and DA–IB is uncertain because the ratio of IB varies (Fig. 7).
For the primary screening of ASP toxin-producing N. navis-
varingica, the culture conditions described in Section 2.1 wereused. The conditions are considered to be sufficiently similar tothose of the seawater from which these N. navis-varingica strainswere isolated. Temperature and salinity appear to not affect thetoxin composition based on preliminary experiments (data notshown), but systematic examination of the factors affecting thetoxin composition of this diatom, including co-existing bacteria, isneeded. The detailed processes of the production of each toxincomponent are being investigated for determining the toxinproduction mechanism.
The toxicity of IA, IB, and IC is reported to be significantly lowerthan DA alone (Munday et al., 2008), suggesting that these toxinspose a lower risk to humans. However, the structural difference ofDA and IB is based on the position of only one double bond: 30–40
(DA) and 40–50 (IA), and in IA and IB, the difference is due to only theconfiguration of Z (IA) and E (IB) at the 10–20 double bond (Fig. 1).Because of the structural similarities, there is a possibility thatmarine animals such as shellfish, fish and mammals may act asvectors of the toxin isomers, which might then be converted fromIB to IA and finally to DA in the animal tissue, although there is noreport showing these bioconversions in the animal tissue. Studiesof such conversions are currently ongoing.
In conclusion, we isolated ASP toxin-producing diatom N. navis-
varingica from selected areas of Luzon Island, the Philippines andSouth Sulawesi, Indonesia. Two new toxin composition types: (1)only IB and (2) DA, IA and IB were confirmed in the N. navis-
varingica isolates from Luzon Island, the Philippines. The stabilityof these toxin compositions was confirmed by comparing the toxincomposition of a parental strain and respective sub-strains. Wesummarized, in total, five ASP toxin composition types in N. navis-
varingica. Investigation of factors affecting the toxin compositionof N. navis-varingica might help in determining the toxinproduction mechanism of diatoms.
Acknowledgements
This work was supported in part by a grant-in-aid for research(C) from the Japan Society for the Promotion of Sciences (JSPS) andby a grant-in-aid for a multilateral cooperative research project(coastal oceanography), from the Ministry of Education, Science,Sports and Culture of Japan. We also thank Ms. Logen F. Enriquez ofBFAR Region I Lucap, Alaminos, Pangasinan, for help in samplecollection.
References
Bajarias, t.l., Kotaki Jr., F.F.A.Y., Romero, J.R.R., Furio, M.L.J., Lundholm, E.F., Koike, N.,Fukuyo, K., Kodama, Y.M., 2006. Screening of diatoms producing domoic acidand its derivatives in the Philippines. Coast. Mar. Sci. 30, 121–129.
Bates, S.S., 2000. Domoic acid-producing diatoms: another genus added. J. Phycol.36, 978–985.
Bates, S.S., Bird, C.J., deFreitas, A.S.W., Foxall, R., Gilgan, M., Hanic, L.A., Johnson, G.R.,McCulloch, A.W., Odense, P., Pocklington, R., Quilliam, M.A., Sim, P.G., Smith, J.C.,SubbaRao, D.V., Todd, E.C.D., Walter, J.A., Wright, J.L.C., 1989. Pennate diatomNitzschia pungens as the primary source of domoic acid, a toxin in shellfish fromeastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 46, 1203–1215.
Garrison, D.L., Conrad, S.M., Eilers, P.P., Waldron, E.M., 1992. Confirmation of domoicacid production by Pseudonitzschia australis (Bacillariophyceae) cultures. J.Phycol. 28, 604–607.
Guillard, R.R.L., 1983. Culture of phytoplankton for feeding marine invertebrates. In:Berg, Jr., C.J. (Ed.), Culture of Marine Invertebrates. Hutchinson Ross PublishingCo., Stroudsberg, PA, pp. 108–132.

M.L.J. Romero et al. / Harmful Algae 10 (2011) 456–462462
Holland, P.T., McNabb, P., Rhodes, L.L., Selwood, A.I., Neil, T., 2003. Amnesic shellfishpoisoning toxins in New Zealand shellfish: detection of an unusual domoic acidisomer using a newly validated LC–MS/MS method. In: Villalba, A., RomaldeBeiras, B.R.J. (Eds.), Molluscan Shellfish Safety. Xanta de Galicia & IOC ofUNESCO, Santiago de Compostela, pp. 29–42.
Holland, P.T., Selwood, A.I., Mountfort, D.O., Wilkins, A.L., McNabb, P., Rhodes, L.L.,Doucette, G.J., Mikulski, C.M., King, K.L., 2005. Isodomoic acid C, an unusualamnesic shellfish poisoning toxin from Pseudo-nitzschia australis. Chem. Res.Toxicol. 18, 814–816.
Kotaki, Y., 2008. Ecobiology of amnesic shellfish toxin producing diatoms. In:Botana, L.M. (Ed.), Seafood and Freshwater Toxins – Pharmacology, Physiology,and Detection. second ed. CRC Press, Taylor & Francis Group, New York, pp.383–396.
Kotaki, Y., Lundholm, N., Katayama, T., Furio, E.F., Romero, M.L., Relox, J.R., Yasu-moto, T., Naoki, H., Hirose, M.Y., Thann, T.D., Thuoc, C.V., Huyen, N.T.M., Thu,P.T., Takata, Y., Kodama, M., Fukuyo, Y., 2008. ASP toxins of pennate diatoms andbacterial effects on the variation in toxin composition. In: Moestrup, Ø., et al.(Eds.), Harmful Algae, The Proceeding of 12th International Conference onHarmful Algae. Intergovermental Oceanographic Commission, UNESCO, Copen-hagen, Denmark, pp. 300–302.
Kotaki, Y., Furio, E.F., Satake, M., Lundholm, N., Katayama, T., Koike, K., Fulgueras,V.P., Bajarias, F.A., Takata, Y., Kobayashi, K., Sato, S., Fukuyo, Y., Kodama, M.,2005. Production of isodomoic acids A and B as major toxin components of apennate diatom Nitzschia navis-varingica. Toxicon 46, 946–953.
Kotaki, Y., Koike, K., Yoshida, M., Thuoc, C.V., Huyen, N.T.M., Hoi, N.C., Fukuyo, Y.,Kodama, M., 2000. Domoic acid production in Nitzschia sp. (Bacillariophyceae)isolated from a shrimp-culture pond in Do Son, Vietnam. J. Phycol. 36, 1057–1060.
Kotaki, Y., Lundholm, N., Onodera, H., Kobayashi, K., Bajarias, F.A., Furio, E.F., Iwataki,M., Fukuyo, Y., Kodama, M., 2004. Wide distribution of Nitzschia navis-varingica,a new domoic acid-producing benthic diatom found in Vietnam. Fish. Sci. 70,28–32.
Lundholm, N., Moestrup, Ø., 2000. Morphology of the marine diatom Nitzschia navis-varingica, sp. nov, (Bacillariophyceae), another producer of the neurotoxindomoic acid. J. Phycol. 36, 1162–1174.
Lundholm, N., Skov, J., Pocklington, R., Moestrup, Ø., 1994. Domoic acid, the toxicamino acid responsible for amnesic shellfish poisoning, now in Pseuodonitzschiaseriata (Bacillariophyceae) in Europe. Phycologia 33, 475–478.
Maeda, M., Kodama, T., Tanaka, T., Yoshizumi, H., Takemoto, T., Nomoto, K., Fujita,T., 1986. Structures of isodomoic acids A B, and C, novel insecticidal aminoacids from the red alga Chondria armata. Chem. Pharm. Bull. 34, 4892–4895.
Martin, J.L., Haya, K., Burridge, L.E., Wildish, D.J., 1990. Nitzschia pseudodelicatissima– a source of domoic acid in the Bay of Fundy, eastern Canada. Mar. Ecol. Prog.Ser. 67, 177–182.
Munday, R., Holland, P.T., McNabb, P., Selwood, A.I., Rhodes, L.L., 2008. Comparativetoxicity to mice of domoic acid and isodomoic acids A, B and C. Toxicon 52, 954–956.
Orsini, L., Sarno, D., Procaccini, G., Poletti, R., Dahlmann, J., Montresor, M., 2002.Toxic Pseudo-nitzschia multistriata (Bacillariophyceae) from the Gulf of Naples:morphology, toxin analysis and phylogenic relationship with other Pseudo-nitzschia species. Eur. J. Phycol. 37, 247–257.
Pocklington, R., Milley, J.E., Bates, S.S., Bird, C.J., deFreitas, A.S.W., Quilliam, M.A.,1990. Trace determination of domoic acid in seawater and phytoplankton byhigh-performance liquid chromatography of the fluorenylmethoxycarbonyl(FMOC) derivative. Int. J. Environ. Anal. Chem. 38, 351–368.
Quilliam, M.A., 2003. Chemical methods for domoic acid, the amnesic shellfishpoisoning (ASP) toxin. In: Hallegraeff, G.M., Anderson, D.M, Cembella, A.D.(Eds.), Manual on Harmful Marine Microalgae. UNESCO, Paris, pp. 247–265.
Rhodes, L., White, D., Syhre, M., Atkinson, M., 1996. Pseudo-nitzschia species isolatedfrom New Zealand coastal waters: domoic acid production in vitro and linkswith shellfish toxicity. In: Yasumoto, T., Oshima, Y., Fukuyo, Y. (Eds.), Harmfuland Toxic Algal Blooms. Intergovermental Oceanographic Commission ofUNESCO, Paris, pp. 155–158.
Rhodes, L.L., 1998. Identification of potentially toxic Pseudo-nitzschia (Bacillario-phyceae) in New Zealand coastal waters, using lectins. New Zeal. J. Mar.Freshwater Res. 32, 537–544.
Rhodes, L.L., Holland, P.T., Adamson, J.E., McNabb, P., Selwood, A.I., 2003. Production of anew isomer of domoic acid by New Zealand isolates of the diatom Pseudo-nitzschiaausutralis. In: Villalba, A.,Romalde, B.R.J.,Beiras, R. (Eds.),Molluscan Shellfish Safety.Xanta de Galicia & IOC of UNESCO, Santiago de Compostela, pp. 43–48.
Romero, M.L.J., Lirdwitayaprasit, T., Kotaki, Y., Lundholm, N., Relox, J.R., Furio, E.F.,Terada, R., Yokoyama, T., Kodama, M., Fukuyo, Y., 2008. Isolation of ASP toxin-producing Nitzschia from Thailand. Mar. Res. Indones. 33, 225–228.
Takata, Y., Sato, S., Ha, D.V., Montojo, U.M., Lirdwitayaprasit, T., Kamolsiripichai-porn, S., Kotaki, Y., Fukuyo, Y., Kodama, M., 2009. Occurrence of domoic acid intropical bivalves. Fish. Sci. 75, 473–480.
Takemoto, T., Daigo, K., 1958. Constituents of Chondria armata. Chem. Pharm. Bull. 6,578–580.
Wright, J.L.C., Boyd, R.K., de Freitas, A.S.W., Falk, M., Foxall, R.A., Jamieson, W.D.,Laycock, M.V., McCulloch, A.W., McInnes, A.G., Odense, P., Pathak, V.P., Quilliam,M.A., Ragan, M.A., Sim, P.G., Thibault, P., Walter, J.A., Gilgan, M., Richard, D.J.A.,Dewar, D., 1989. Identification of domoic acid, a neuroexcitatory amino acid, intoxic mussels from eastern Prince Edward Island. Can. J. Chem. 67, 481–490.
Wright, J.L.C., Falk, M., McInnes, A.G., Walter, J.A., 1990. Identification of isodomoicacid D and two new geometrical isomers of domoic acid in toxic mussels. Can. J.Chem. 68, 22–25.
Zaman, L., Arakawa, O., Shimosu, A., Onoue, Y., Nishio, S., Shida, Y., Noguchi, T., 1997.Two new isomers of domoic acid from a red alga, Chondria armata. Toxicon 35,205–212.
Related Documents











