Understanding Germination and Pathogenicity in Zygomycota Species through Genomic and Transcriptomic Approaches by Poppy Sephton Clark A thesis submitted to the University of Birmingham for the degree of DOCTOR OF PHILOSOPHY School of Biosciences College of Life and Environmental Sciences University of Birmingham August 2019

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Understanding Germination and Pathogenicity in Zygomycota Species through Genomic and Transcriptomic
Approaches by Poppy Sephton Clark
A thesis submitted to the University of Birmingham for the degree of DOCTOR OF PHILOSOPHY
School of Biosciences College of Life and Environmental Sciences
University of Birmingham August 2019

University of Birmingham Research Archive
e-theses repository This unpublished thesis/dissertation is copyright of the author and/or third parties. The intellectual property rights of the author or third parties in respect of this work are as defined by The Copyright Designs and Patents Act 1988 or as modified by any successor legislation. Any use made of information contained in this thesis/dissertation must be in accordance with that legislation and must be properly acknowledged. Further distribution or reproduction in any format is prohibited without the permission of the copyright holder.

1
Abstract
Mucorales spores are the causative agents of the emerging disease mucormycosis.
Mucorales species are also responsible for high quantities of food spoilage annually. The
mechanism by which Mucorales spores cause disease and rot relies upon spore
germination, however the mechanism underlying germination in these species remains
poorly understood. Presented here are results which characterise Mucorales spore
germination, through phenotypic and transcriptional studies (RNA-Seq), which followed
the defined germination phenotype throughout. Hallmark pathways are identified
through analysis of differentially expressed genes and co-transcriptional networks,
providing targets for germination inhibition. With the resulting transcriptional data, the
genome of Rhizopus delemar was enriched and analysed, thus providing better information
on the Mucoralean genome. Comparative genomics was also employed to better
understand genotypic variation between Mucorales species. To examine the differences
in pathogenicity between species, and assess the impact of germination stage on
pathogenicity, the transcriptional profile (RNA-Seq) of selected Mucorales species was
examined upon phagocytosis by innate immune cells. To better understand the
corresponding host response, the transcriptional response (single cell RNA-Seq) of innate
immune cells to Mucorales infection was also examined. Finally, germination targets
identified through the described analyses were targeted with suspected inhibitors to
confirm function in germination regulation. This work has furthered our basic understanding
of germination in these ancient fungi, indicated pathways essential to the germination
programme of Mucorales species, and demonstrated a crucial role played by many of these
pathways in host-fungal interactions of the Mucorales.
Sephton- Clark, Poppy

Acknowledgements
I would like to express my gratitude to Dr Elizabeth Ballou and Dr Kerstin Voelz, both advisors
have offered incredible support, encouragement and guidance. A special thanks to Dr Voelz
for taking me on and supporting me through the initial stages of my study, and to Dr Ballou
for encouraging me to continue along my research path and offering amazing support and
mentorship along the way.
I would like to express my appreciation to Dr Christina Cuomo, with whom I spent invaluable
time training with, and Professor Robin May, whose encouragement during my
undergraduate studies led me to continue my research in the fungal field. And of course, a
huge thanks to the amazing HAPI Lab members (past and present!). It has been a joy to
work in such a positive environment, with enthusiastic scientists so supportive of one
another!
I would like to say a special thank you to Daniel, for all of his support, curiosity
and encouragement. For celebrating triumphs with me, sharing frustrations with me and
always encouraging me to go after my goals. It is truly appreciated!
Last but not least, I would like to say thank you to my parents, for encouraging me for as
long as I can remember! For encouraging curiosity and always showing an interest in
whatever I was most fascinated with, thank you! Finally, I would like to say thank you to my
Gran, for all of her encouragement along the way!

Table of Contents
Chapter 1: Introduction .................................................................................. 5
Introduction: Mucorales .................................................................................................... 5
Food Spoilage ............................................................................................................................ 8
Mucormycosis ............................................................................................................................ 9
Mucorales spores and germination regulation ......................................................................... 10
Germination as a mechanism of pathogenicity ......................................................................... 12
Project Aims ................................................................................................. 15
Literature Review ............................................................................................................ 16
Introduction to fungal morphotypes: Spores and Hyphae ........................................................ 16 Importance of spores and hyphae in pathogenicity and food spoilage ..................................................... 20
Spore Composition .................................................................................................................. 22 The spore cell wall ...................................................................................................................................... 22 Spore compartmentalization and dormancy factors .................................................................................. 24 Water availability and metabolic activity ................................................................................................... 25
The Spore Germination Program .............................................................................................. 26 Spore polarization ...................................................................................................................................... 26 Hyphal outgrowth and extension ............................................................................................................... 28
Regulation of Germination ....................................................................................................... 29 The nutritional environment and germination .......................................................................................... 30 Germination Regulation via Ph, Temperature, Light and Environmental Gases ......................................... 35 Signalling molecules ................................................................................................................................... 40
Materials and Methods…………..………………………………………………………………………………………..43
Microbial Culture ............................................................................................................. 44
Fungal Culture ......................................................................................................................... 44 Spore Isolation ............................................................................................................................................ 44 Spore Growth ............................................................................................................................................. 44 Endosymbiont Curing ................................................................................................................................. 44
Bacterial Culture ...................................................................................................................... 45
Germination Phenotyping ................................................................................................ 46
Spore Germination Assay ......................................................................................................... 46 Live cell Imaging ......................................................................................................................................... 46 Flow Cytometry .......................................................................................................................................... 46 XTT Assay .................................................................................................................................................... 46 Exosome Release ........................................................................................................................................ 47
Genomic DNA Extraction, Sequencing and Analysis ......................................................... 48
Genomic DNA Extraction .......................................................................................................... 48 Fungal DNA Extraction ................................................................................................................................ 48 Bacterial DNA Extraction ............................................................................................................................ 48
Genomic DNA Sequencing ........................................................................................................ 48 Fungal Sequencing ...................................................................................................................................... 48
1

Bacterial Sequencing .................................................................................................................................. 49
Genome Sequence Analysis ..................................................................................................... 49 Rhizopus microsporus Genome Assembly .................................................................................................. 49 Rhizopus microsporus Variant Identification .............................................................................................. 49 Comparative Genomics .............................................................................................................................. 50
Rhizopus delemar genome annotation update ......................................................................... 50
Transcription and Inhibition of Rhizopus delemar Germination ........................................ 51
Germination RNA-Seq .............................................................................................................. 51 RNA Isolation .............................................................................................................................................. 51 Data Analysis .............................................................................................................................................. 53
Germination Inhibition ............................................................................................................ 53 Inhibition Assessment of Targets Determined via RNA-Seq ....................................................................... 53 Strathclyde Natural Compound Library ...................................................................................................... 54
Rhizopus-macrophage interactions .................................................................................. 55
Macrophage ............................................................................................................................ 55 Macrophage Culture ................................................................................................................................... 55 In vitro Phagocytosis assay ......................................................................................................................... 55 Phagocytosis Live Cell Imaging ................................................................................................................... 55
RNA-Seq .................................................................................................................................. 56 Bulk Rhizopus RNA-Seq .............................................................................................................................. 56 Single Cell Macrophage RNA-Seq ............................................................................................................... 56 RNA-Seq Data Analysis ............................................................................................................................... 57
Other ............................................................................................................................... 58
Rhizopus delemar Protoplast Formation .................................................................................. 58
Zebrafish .................................................................................................................................. 58 Macrophage isolation and RNA Extraction ................................................................................................. 58
Chapter 2: Mucorales Spore Germination Characterisation .......................... 60
Germination phenotype diversity .................................................................................... 61
Results ............................................................................................................................. 65
Cell Size and Germination Rate ................................................................................................ 65
Morphology and Cell Wall Dynamics ........................................................................................ 67
Extracellular Vesicle Release .................................................................................................... 72
Discussion ................................................................................................................................ 74
Chapter 3: Hallmarks of the Mucorales genome ........................................... 77
Mucoralean Genomics ..................................................................................................... 78 Genome Statistics ....................................................................................................................................... 81
Results ............................................................................................................................. 81
Resulting Characteristics of the Reannotated R. delemar Genome ........................................... 81
Biochemical Pathways Present in R. delemar ........................................................................... 82 R. delemar WGD Enrichment ................................................................................................... 85 R. microsporus Genome Assembly and Statistics ...................................................................... 86
2

Comparative Genomics ............................................................................................................ 88
Comparison with Aspergillus genomes ..................................................................................... 90 Discussion ........................................................................................................................ 90
Chapter 4: Transcriptional States of Germination ......................................... 92
Germination Regulation .................................................................................................. 94
Aspergillus species ................................................................................................................... 94 Conidial Transcripts .................................................................................................................................... 95Germination Metabolism ........................................................................................................................... 98
Neurospora crassa ................................................................................................................... 99
Fusarium species ...................................................................................................................... 100
Mucorales ...............................................................................................................................101 Results ........................................................................................................................... 103
Transcriptional Trends over Germination ................................................................................ 103
Differential Expression Throughout Germination .................................................................... 107
Metabolic Activity over Germination ...................................................................................... 114
Pathways Upregulated at Alternate Germination Phases ........................................................ 118
Co-transcriptional Networks ................................................................................................... 125 Duplicated Gene Pair Expression .............................................................................................................. 127
Potential Roles of Plant and Fungal Hormones as Germination Regulators ............................. 129
Potential Regulators With Known Functions In The Fungal Kingdom ....................................... 132
Comparisons of Transcription Throughout Germination .......................................................... 137 Discussion ...................................................................................................................... 140
Chapter 5: Transcriptional Regulation of Rhizopus-Macrophage Interactions .................................................................................................................. 143
Host-Pathogen Interactions in Mucormycosis ................................................................ 145
Results ........................................................................................................................... 148
The Fungal Response .............................................................................................................. 149
The Macrophage Response ..................................................................................................... 161
Modulating the Infection Outcome ......................................................................................... 166 Discussion ...................................................................................................................... 169
Chapter 6: Germination Inhibition .............................................................. 171
Manipulation and Inhibition of Germination .................................................................. 172
Results ........................................................................................................................... 175
Germination Inhibitors Identified Through Transcriptional Studies ......................................... 175
Germination Inhibitors Identified Through Natural Compound Screening ............................... 178 Discussion ...................................................................................................................... 180
3

References ................................................................................................. 181
Appendix ................................................................................................... 211
4

Chapter 1: Introduction
Introduction: Mucorales
The following work has been adapted from the book chapter “Spore Germination of
Pathogenic Filamentous Fungi” (Sephton-Clark and Voelz 2017), for which I performed the
literature search, wrote the manuscript, completed revisions, and prepared the figures.
Mucorales species (Figure 1), belonging to the Mucorales order of the Zygomycota division
(Mucoromycotina subdivision), are ancient diverging pathogenic fungi, capable of causing
mucormycosis. These species are also known as food spoiling agents, predominantly spoiling
soft fruits, vegetables and baked goods. Mucorales species are disseminated in their spore
form, which swell and produce aseptate hyphae upon germination (Hoffmann et al. 2013).
They reproduce sexually, via the combination of two hyphae (of opposite mating types)
producing zygospores, or asexually (Mendoza et al. 2014). Asexual reproduction is quicker
and leads to the formation of sporangiospores. These structures contain many spores which
are dispersed via water, air, or animal disruption (Moore-Landecker 2011). Sexual
reproduction introduces genetic variation into the population, allowing for the adaptation to
changing environments (Mendoza et al. 2014), whereas asexual reproduction and sporulation
provides an advantage in terms of dissemination, dispersal and colonisation of new
territories. Propagated spores are found ubiquitously throughout the environment and
remain dormant until favourable conditions prompt germination. Upon germination spores
swell and grow to produce aseptate hyphae (Hoffmann et al. 2013). Once hyphal growth is
5

initiated, Mucorales species are characterised by rapid growth which allows them to cause
infection and food spoilage.
6

Figure 1. Phylogeny of the Mucorales A) Figure adapted from “Phylogenetic and Phylogenomic
Definition of Rhizopus Species”, Gryganskyi et al, G3: GENES, GENOMES, GENETICS, 2018. This
figure shows the phylogeny of 5 Rhizopus species, including multiple R. delemar and R.
microsporus species, as well as Mucor circinelloides. This analysis has been performed based
on 192 orthologues genes, and compares this phylogeny to parsimony phylogeny. Genome
sizes have also been included in bold. B) Figure adapted from “An integrated genomic and
transcriptomic survey of mucormycosis-causing fungi”. Chibucos et al, Nature
Communications, 2016. This analysis shows the broader phylogenetic relationship of 38
Mucorales species, including multiple R. oryzae species, based on the relationship between
76 orthologous proteins. C) Phylogenetic tree based on NCBI taxonomy, generated with
phylot. The asterisk denotes species worked with throughout chapter 2-6.
C
Rhi
zopu
s de
lem
ar R
A 99
-880
Lichtheimia
Can
dida
alb
ican
s
Cunninghamella
Aspergillus nidulans
Rhizopus microsporus
Batrachochytriu
m dendrobatidis
Cryptococcus neoformans
Mucor circinelloides
*
*
*
*
A B
7

Food Spoilage
Worldwide one in eight people are malnourished, whilst it is predicted that a third of
all food produced annually is wasted. Reducing this waste is crucial to improving food
security. Food spoilage is a significant contributor toward food waste; it has
been estimated that 20% of harvested fruit and vegetables are spoiled by microbes (Jay
1992; Barth et al. 2009), whilst in east Asian countries, rice losses due to bruising,
moulds and pest spoilage can be as high as 80% (Fox and Fimeche 2013) . It has also
been estimated that up to 5% of baked goods are spoiled by fungi every year, with
fungal spoilage estimated to cost $10,000,000 a year in Australia alone (Dao and
Dantigny 2011). Food spoilage due to fungal contamination occurs at pre-harvest,
storage, processing and packaging stages of food production (Bond et al. 2013) .
The ability to grow in acidic conditions, as well as at temperature extremes, has led
to the spoilage of fruit juices, pasteurized and refrigerated foods, predominantly by fungi
(Dao and Dantigny 2011).
Fungi of the Mucorales order are capable of invading plant tissue due to their rapid growth,
with fruits and vegetables providing an optimal pH, high water content and nutrient source
for growth (Turgeman et al. 2016). Sweet potatoes, cherries, peaches and tomatoes
in particular are commonly affected by Rhizopus spp. spoilage (J W Eckert and Sommer
1967). Rhizopus may even spoil unbroken fruits, as it is able to penetrate the skin by
secreting esterase enzymes (Baggio et al. 2016). Control of spoilage agents can be
achieved through storage of produce below 5°C, however it is not viable to store all fruit
and vegetables at this temperature (Joseph W Eckert and Ogawa 1989). Biological
agents such as Candida guilliermondii and Acremonium cephalosporium have been
used effectively to decrease Rhizopus spoilage of grapes (Zahavi et al. 2000), whilst
Pichia membranefaciens effectively
8

inhibits spoilage within peach wounds through a proposed mechanism of competitive wound
colonisation (Qing and Shiping 2000; Bonaterra et al. 2003). Although storage at low
temperatures works well as a preventative measure, this is not feasible in all countries. With
few alternative options, it is necessary to develop new measures to reduce food spoilage and
increase food security.
Mucormycosis
Human mucormycosis, an emerging fungal infection caused by members of the Mucorales
order, has become a growing concern due to difficulty treating, resulting in mortality rates of
up to 90% (Trzaska et al. 2015; Brown et al. 2012; Kontoyiannis et al. 2012). Rhizopus, Mucor
and Lichthemia species are thought to account for 70-80% of all mucormycosis
infections (Gomes, Lewis, and Kontoyiannis 2011), whilst Cunninghamella has been
reported as one of the most aggressive and pathogenic species (Petraitis et al. 2013a).
Mucormycosis mainly affects immunocompromised patients, such as those having
undergone transplants, or in many cases individuals suffering from ketoacidic phases
due to diabetes (Lanternier and Lortholary 2009). Mucormycosis is especially prevalent
in countries with high counts of uncontrolled diabetes, as is the case in India
(Chakrabarti and Singh 2014). Mucormycosis diagnosis due to traumatic injuries is
common, whilst nosocomial acquisition in susceptible patients is also on the rise (Skiada et
al. 2012). Mucorales species manage to cause invasive infection due to their ability to
germinate, grow and proliferate within the host. They avoid killing by the host in
immunocompromised individuals, causing angioinvasion and tissue necrosis (Ibrahim et
al. 2012). Current treatment consists of lipid forms of Amphotericin B and surgical
debridement (Spellberg and Ibrahim 2010). Statins also effectively decrease the growth
of Rhizopus delemar, by attenuating germination and increasing the pathogens
9

susceptibility to oxidative stress (Bellanger et al. 2016). Although alternative treatment
options are being explored, mortality rates remain high, available treatment options for
mucormycosis are severely limited and the outcome often leads to patients having affected
areas amputated.
Mucorales spores and germination regulation
Mucorales spores have been detected in a range of environments, from the sands of Saudi
Arabia to the forests of China (Murgia et al. 2019; Walther et al. 2013). They appear dark due
to the melanin within the cell wall, a feature common to many fungi which is thought to
protect against UV damage (Moore-Landecker 2011). These hardy spores can survive
temperatures of 60-70°C (maintaining viability), however once germination is initiated, these
spores become increasingly prone to damage (Turgeman et al. 2013). The composition of the
Mucorales cell wall changes over germination; under aerobic conditions chitin increases over
germination, with chitosomes acting as a reservoir for chitin synthase (Kamada, Bracker, and
Bartnicki-Garcia 1991). At rest, Mucorales spore cell walls contain large quantities of lipids,
accounting for 10-40% of the cell wall (Feofilova et al. 2015). The cell wall also contains
considerable chitin/chitinosan (11.6%), sugars (49.3%), protein (16.1%), phosphate (2.6%)
and melanin (10.3%) (Bartnicki-Garcia 1968).
Germination may be initiated upon cell wall breakage, removal of unfavourable conditions or
the introduction of specific cues and nutrients - such as water, carbon and nitrogen (Feofilova
et al. 2012; Mendoza et al. 2014). Essential nutrients for triggering germination of Rhizopus
oligosporus includes glucose, phosphates and a mixture of amino acids, with leucine
displaying a strong inductive effect on germination (Thanh, Rombouts, and Nout 2005). Blue
10

and green light have also been proposed as germination regulators for the light-sensitive
protein possessing Mucorales species Mucor circinelloides (Herrera-Estrella and Horwitz
2007). Germination of Rhizopus delemar can be achieved with acidified glucose alone, as pH
regulates germination via the recruitment of aquaporins (RdAQP1 and RdAQP2) that enable
swelling (Turgeman et al. 2016). In Mucorales spores, the amount of RNA and protein within
the spore appears to increase exponentially as soon as germination is induced, however DNA
synthesis has not been reported to occur until 30 minutes before the production of germ
tubes (Cano and Ruiz-Herrera 1988).
11

Spores of Cryptococcus neoformans and yeast cells of Candida albicans have been shown
to be key to dissemination throughout the host (Walsh et al. 2019; Seman et al. 2018).
C. neoformans spores are the infectious propagules which lead to greater dissemination
and mortality, when compared to infection with the yeast form (Walsh et al. 2019).
Similarly, Aspergillus species are disseminated throughout the host via their
conidial (spore) forms, however hyphae are often required for tissue damage and
invasion (Bertuzzi, Schrettl, Alcazar-Fuoli, Cairns, Munoz, et al. 2014; Seman et al. 2018;
Ben-ami et al. 2009; Ben-Ami et al. 2009). Spores are the infectious particles of Aspergillus
species and germination is central to pathogenicity (Zhao et al. 2006; Fortwendel et
al. 2005). Aspergillus has long been used as a model for understanding the lesser studied
Mucorales species, and though research in this field provides a framework, a full
understanding of Mucormycete pathogenicity requires comprehensive investigation into
mucorales species. The transition from Mucorales spore to hyphae appears to be a key
pathogenicity factor (Inglesfield et al. 2018; Mendoza et al. 2014), however the
underlying mechanisms which control this event in Mucorales species is poorly understood.
As filamentous growth leads to tissue damage, the rate of germination can also be
a contributing factor to virulence. In Rhizopus species, iron availability is known to
regulate virulence (Andrianaki et al. 2018) as iron limitation leads to inhibition of
germination (Kousser et al. 2019), and excess iron induces expression of the invasion
mediating CotH (Gebremariam et al. 2016). Cunninghamella spp. are also known as
one of the more aggressively invasive Mucorales species sets, the rate of germination
of Cunninghamella spp. is increased, when compared to that of other Mucorales species.
Mucor circinelloides shows increased virulence in its hyphal form, when compared to the
yeast form
12
Germination as a mechanism of pathogenicity

(Herrera-Estrella and Horwitz 2007; Walsh et al. 2019; Seman et al. 2018; Ben-ami et al.
2009; Zhao et al. 2006; Fortwendel et al. 2005; Inglesfield et al. 2018; Andrianaki et al. 2018;
Lee et al. 2013).
In immunocompetent individuals phagocytes inhibit spore germination (Inglesfield et
al. 2018), a mechanism key to infection control in immunocompetent patients. Phagocytes
are rapidly recruited to the site of infection, and form innate granuloma-like
structures around spores, leading to a latent infection. Phagocytes of
immunocompromised patients fail to inhibit germination and subsequently life
threatening infections develop. Despite the vital role played by the innate
immune system in controlling mucormycosis, the interaction between Mucor
species and innate immune cells is poorly understood.
There are several challenges when working with species of the Mucorales order.
These include: a complete absence of chromosomal level genome assemblies (the
highest resolution Rhizopus assembly consists of 83 contigs) (Ma et al. 2009; Horn et
al. 2015; Mondo et al. 2017); the repetitive nature of Mucorales genomes which
makes for difficult assembly; unclear species phylogeny (Gryganskyi et al. 2018;
Hoffmann et al. 2013); minimal or absent genome annotation; limited genetic tools
for manipulation (until recently) (Garcia, Vellanki, and Lee 2018); and large
phenotypic variation and genotypic variation between species within the order.
Further to this, understanding of the pathogenicity mechanisms employed by
Mucorales species is limited (Gebremariam et al. 2014), compared to better
studied species such as Cryptococcus, Candida and Aspergillus spp. Aside from the
work presented here, there have been few comparative genomic and phenotypic studies
13

of Mucorales species. To date, there have been no transcriptional (high-resolution)
studies of Mucorales germination, few studies which explore the transcriptional
basis of Mucorales-host interactions and only one which explores germination
inhibition as a means to inhibit pathogenicity (Trzaska et al. 2015).
14

Project Aims
This project aims to further understanding of the mechanisms of germination in the
Mucorales species, and determine how this programme of morphological change and rapid
growth contributes to pathogenicity. Once a basic phenotypic and transcriptional
understanding of germination has been established, this project aims to detect mechanisms
key to pathogenicity and identify pathways targetable to inhibit germination and reduce
pathogenicity.
15

Literature Review
The following literature review will give an overview of knowledge about fungal morphology
and germination regulation in multiple fungal species. Subsequent chapters will include
literature reviews on the current knowledge of: phenotypes of fungal germination (Chapter
2), hallmarks of the fungal genome (Chapter 3), transcriptional regulation of fungal
germination (Chapter 4), fungal-immune cell interactions (Chapter 5) and inhibition and
modulation of fungal germination (Chapter 6), relevant to the work presented in these
sections.
Introduction to fungal morphotypes: Spores and Hyphae
Spores may be formed either through sexual or asexual reproduction. Asexual reproduction
is thought to provide an advantage due to the speed at which the spores can be produced
and disseminated. Sexual reproduction, though often a longer process, presents an advantage
through introduction of genetic variation into the population (Moore-Landecker 2011;
Mendoza et al. 2014). The method of reproduction may be determined by the environment
encountered by the fungi. For example, It has been suggested that the decision to reproduce
sexually is regulated by trehalose homeostasis in Cryptococcus neoformans (Botts et al. 2014),
whilst Aspergillus species conidiate when grown in nutritionally sparse conditions (Adams,
Wieser, and Yu 1998). Although the nutritional triggers of sexual reproduction in the
Mucorales order have not been well studied, trisporic acid is capable of triggering this process
(Schimek and Wostemeyer 2012). Mucorales species produce this pheromone prior to sexual
reproduction: both mating types must co-operate to complete production, as they rely on
16

one another for the interchange of intermediates required for this biosynthetic pathway (Lee
and Heitman 2014).
Asexual spores are genetically identical to their parent cells and may be formed through the
specific process of sporulation, or through the transformation of an existing cell. The asexual
spores of basidiomycetes, ascomycetes, mucormycetes and chytridiomycetes are known as
conidia, arthrospores or conidia, sporangiospores and zoospores (Table 1). Arthrospores are
produced through the conversion of an existing cell, whilst conidia, zoospores and
sporangiospores are formed through a specific process that produces new spores, known as
sporulation or conidiation. Blastospores form in a budding manner, budding away from
hyphae, swollen cells or vesicles. Specialized spore-producing cells known as phialides are
also capable of producing blastospores.
The sexual spores of basidiomycetes, ascomycetes and mucormycetes are known as
basidiospores, ascospores and zygospores, respectively. Sexual spores are usually formed via
the fusion of hyphae, zoospores or gametangia of opposite mating types. This process may
be initiated by the release of fungal hormones, as described for the mucormycetes (Austin,
Bu’lock, and Gooday 1969). The genomic structure of the mating locus has been described for
Rhizopus and Mucorales species and although sequence comparisons revealed locus
conservation, the results enable increased phylogeny resolution (Gryganskyi et al. 2010).
17
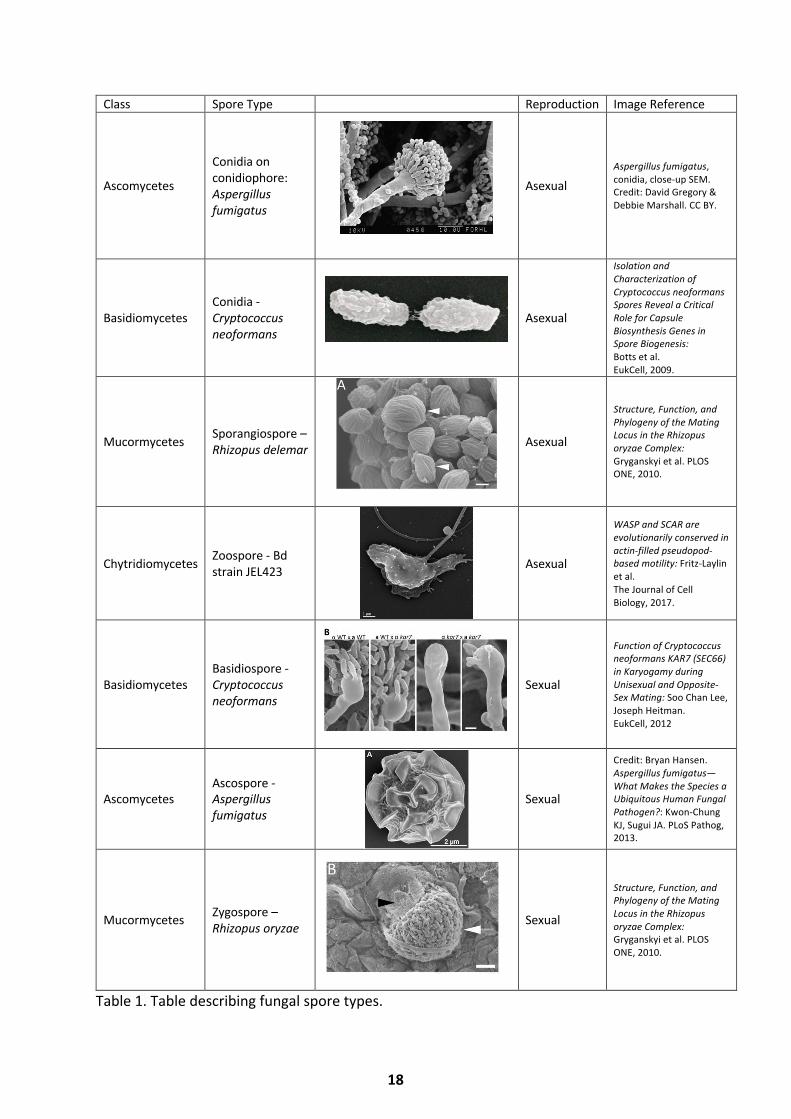
Class Spore Type Reproduction Image Reference
Ascomycetes
Conidia on conidiophore: Aspergillus fumigatus
Asexual Aspergillus fumigatus, conidia, close-up SEM. Credit: David Gregory & Debbie Marshall. CC BY.
Basidiomycetes Conidia - Cryptococcus neoformans
Asexual
Isolation and Characterization of Cryptococcus neoformans Spores Reveal a Critical Role for Capsule Biosynthesis Genes in Spore Biogenesis: Botts et al. EukCell, 2009.
Mucormycetes Sporangiospore – Rhizopus delemar Asexual
Structure, Function, and Phylogeny of the Mating Locus in the Rhizopus oryzae Complex: Gryganskyi et al. PLOS ONE, 2010.
Chytridiomycetes Zoospore - Bd strain JEL423 Asexual
WASP and SCAR are evolutionarily conserved in actin-filled pseudopod-based motility: Fritz-Laylin et al. The Journal of Cell Biology, 2017.
Basidiomycetes Basidiospore - Cryptococcus neoformans
Sexual
Function of Cryptococcus neoformans KAR7 (SEC66) in Karyogamy during Unisexual and Opposite-Sex Mating: Soo Chan Lee, Joseph Heitman. EukCell, 2012
Ascomycetes Ascospore - Aspergillus fumigatus
Sexual
Credit: Bryan Hansen. Aspergillus fumigatus—What Makes the Species a Ubiquitous Human Fungal Pathogen?: Kwon-Chung KJ, Sugui JA. PLoS Pathog, 2013.
Mucormycetes Zygospore – Rhizopus oryzae Sexual
Structure, Function, and Phylogeny of the Mating Locus in the Rhizopus oryzae Complex: Gryganskyi et al. PLOS ONE, 2010.
Table 1. Table describing fungal spore types.
18

Once spores are formed, they are usually kept suspended within sacs or fruiting bodies. These
structures may contain thousands of spores and differentiate from fungal filaments, to which
they remain associated. The number of spores contained in fruiting bodies can be described
as a balancing act: maximizing the number increases spread of the species, increasing chances
of survival, whilst too many may result in stalk collapse (Santorelli et al. 2008). Dispersion of
spores from sacs is achieved through wind, water or animal disturbance. Release of some
spores is also determined seasonally to coincide with the humidity and temperatures which
provide an optimal climate for the germination of different species.
Germination is induced through favourable conditions and generally occurs in the following
stages: isotropic growth or swelling, cell polarization, hyphal emergence and
hyphal elongation (Figure 2) (Bonazzi et al. 2014). The process of germination
culminates in the production of septate or aseptate hyphae depending on fungal species.
Throughout isotropic growth, the spores can visibly be seen to swell. During this phase
spores will take up water, whilst expanding and reorganizing their cell walls. Spores also
increase their metabolic activity and may use external or internal energy sources in
order to initiate transcription and translation (Griffin 1996; Novodvorska et al. 2016).
Polarization of the cell wall following swelling determines where the hyphal tube will
emerge and utilizes cell machinery such as the polarisome to support this process. This
stage involves an extensive remodelling of the cell wall to enable hyphal extension. When
hyphae emerge and extend, they grow to form what can be seen as a hyphal mat or
matrix, in which the hyphae may overlap and even grow towards nutritional or light sources
(Lucas, Kendrick, and Givan 1975; Dussutour et al. 2010).
19

Figure 2. Schematic of germination. Adapted from Sephton-Clark & Voelz (2018).
Importance of spores and hyphae in pathogenicity and food spoilage
Although we are exposed to thousands of fungal spores every day (Moore-Landecker 2011),
not every spore we encounter is pathogenic. Of those which are pathogenic, the
developmental stage of the spore can affect the outcome of the encounter; whilst dormant
spores may not be pathogenic, the germinated counter parts are often capable of causing
disease in humans, animals or plants. To maintain dormancy, dispersed spores may employ
nutrient and enzymatic compartmentalization, alongside the release of germination
autoinhibitors, molecules which help regulate the maintenance of dormancy (Feofilova et al.
2012). Dormancy enables spores to avoid germination under conditions that may not be
optimal, or could even be harmful to the germlings. Under suboptimal conditions the
germling may perish due to environmental stress or lack of nutrients. Conversely, if spores
stay dormant too long, they may miss a golden opportunity to exploit their environment for
fungal growth and dissemination. As a result, the maintenance of dormancy is carefully
regulated.
Cues which initiate germination may include the host environment, signalling spores to
initiate pathogenic growth within or on a host. Once metabolically active, germinating spores
Light
Organic Nutrients
Inorganic Nutrients
Signalling Molecules
Isotropic Growth Hyphal Emergence Hyphal Growth
Temperature
pH
20

have the potential to express their repertoire of virulence factors. Damage to the host may
be caused by the release of toxins, pathogenicity factors, or immunostimulatory
components such as cell wall constituents. Plant pathogenic Fusarium spp. are known to
produce and release phytotoxins which aid pathogenicity and invasion, through the
induction of host cell death (Nishiuchi et al. 2006). The human pathogen Aspergillus flavus,
is known to cause respiratory diseases, like many other Aspergillus sp. This may be through
pulmonary infection in an immunocompromised individual, or simply through an allergic
reaction induced by the inhalation of the conidia. A. flavus is also known to produce the
mycotoxin, aflatoxin, which can lead to liver damage and even cancers, if the toxin is
consumed (Hedayati et al. 2007). Similarly, Candida albicans produces candidalysin, a
cytotoxic peptide which is toxic towards tissue, and causes damage during
infection. Specifically, the hyphal form of C. albicans releases candidalysin, which aids
dissemination and host invasion (Moyes et al. 2016).
In many cases, filamentous growth initiated during spore germination may be the
underlying cause of disease. Filamentous growth can lead to tissue invasion and
disruption which may prove fatal to the host. The lesser known but often more
invasive Aspergillus terreus has been shown to produce accessory conidia, capable of
increased germination rates when compared to the primary conidia produced by
Aspergillus species, with accessory conidia forming hyphae within two hours of
germination initiation. This increased germination rate may account directly for the
aggressive pathogenicity of this species (Deak et al. 2009).
The emergence and extension of hyphae produced by plant pathogenic fungi, which is
frequently accompanied by the release of extracellular degradative enzymes, is often key to
infection. Infection caused by the wheat and barley pathogen Fusarium graminearum relies
21

on hyphal extension and the release of these enzymes in order to invade the host tissue
(Zheng et al. 2012). Hyphal growth is also key to the infection caused by Batrachochytrium
dendrobatidis and Batrachochytrium salamandrivorans. Once the motile chytrid zoospores
of the amphibian fungal pathogens Batrachochytrium dendrobatidis and Batrachochytrium
salamandrivorans have made contact with the mucus membranes of their hosts, they are
also known to utilise swelling and hyphal growth as a mechanism to invade host tissue (van
Rooij et al. 2012).
Spore Composition
The spore cell wall
The cell wall of spores offers increased protection against environmental factors allowing
survival of resting spores. The general structure and components within a spore cell wall are
relatively conserved between fungal species. While fungal cell walls mainly consist of
polysaccharides, with lipids and proteins usually accounting for a smaller percentage, the
ratio of these components largely depends on species.
The composition of the Mucor rouxii spore wall has been well documented and is known to
contain around 42.6% glucose, 16.1% protein, 10.3% melanin, 9.8% lipid, 9.5% chitosan,
2.1% chitin, 2.6% phosphate, 4.8% mannose and 1.9% glucuronic acid (Reyes and Bartnicki-
garcia 1964). Similarly, the S. cerevisiae strain AM3 ascospore wall consists of large
quantities of glucose (55%), mannan (17%), chitin/chitosan (9%), and protein (11%), along
with smaller quantities of phosphates and organic acids (Briza et al. 1988). The cell walls
of Aspergillus oryzae conidia show a much lower lipid content, at around 2%. However the
carbohydrate content is similar at about 30% in Aspergillus niger (Sumi 1928; Feofilova
et al. 1988), demonstrating the variability in ratio between species. The spore cell wall
22

of Aspergillus fumigatus has even been recorded to contain several lectins, capable of
binding sialic acid and fucose, along with other sugars (Houser et al. 2013). Interestingly, the
presence of some lectins has been shown to inhibit spore germination of penicillia
(Barkai-Golan, Mirelman, and Sharon 1978). The conidial cell wall of Aspergillus
fumigatus has also been shown to contain hydrophobic rodlets, known as RodA&B,
thought to give structural support to the spore wall, as well as enhancing the
adhesion of conidia to surfaces, and masking conidia from host immune recognition
(Paris et al. 2003; Aimanianda et al. 2009).
The cell walls of many spore forming pathogenic fungi are also known to contain melanin.
This pigment colours the spores brown, with an increased melanin content causing the
spores to appear darker and most melanin mutants displaying an ‘albino’ phenotype.
Fungal melanin consists of a highly complex structure, which exists in a stacked planar
sheet structure and is likely formed from DOPA oligomers or polymers (Nosanchuck et al.
2015). The inclusion of melanin into fungal cells varies markedly between
species. Melanin accompanies the hydrophobic rodlets in the outer layer of Aspergillus
fumigatus, where it is thought to confer pathogenicity (Akoumianaki et al. 2016), and
provide structural support (Pihet et al. 2009). The cell wall of an Aspergillus fumigatus
mutant, which does not possess melanin, showed decreased electronegativity,
hydrophobicity and a significant change in the structure of the conidial wall itself. This
includes a loss of the outer hydrophobic rodlet layer which likely accounts for the loss of
hydrophobicity (Bayry et al. 2014; Pihet et al. 2009). The accessory conidia of the
highly pathogenic Aspergillus terreus have a wall which is compositionally very
different to those of other Aspergillus species. The wall of A. terreus lacks the
rodlet coating on the outermost layer, as well as the melanised underlayer, and
also contains less ergosterol, demonstrating that variability even within a genus can
be vast (Deak et al. 2009).
23

In addition, the cell wall of spores from the dimorphic human pathogen Blastomyces
dermatitidis is known to contain melanin, which has been shown to offer protection
against UV (Ultra Violet) (Nosanchuk et al. 2004), whilst antioxidant carotenes may also
be present to provide protection against reactive oxygen species (ROS). In the
opportunistic human pathogen Cryptococcus neoformans, melanin-
containing vesicles known as fungal melanosomes are thought to deliver melanin to
the cell wall, and once trapped form the layers seen in C. neoformans (Eisenman et al.
2005, 2009). The presence of melanin in C. neoformans is associated with an increase
in virulence. It is thought that it provides protection against reactive oxygen
species (ROS) which phagocytic cells use against pathogens (Moore-Landecker 2011;
Schnitzler et al. 1999).
Spore compartmentalization and dormancy factors
The structure of spores is, in part, responsible for their ability to survive in extreme
environments. It should be noted that in general spore walls, structurally, are
very sturdy, although prone to morphing when under severely dehydrating
conditions (Sarmiento et al. 2006). By storing compounds which will be used in the initial
stages of germination separately from the enzymes which will catabolize them to release
energy for the initiation of germination, spores maintain dormancy and energy stores
(Dijksterhuis et al. 2007). For example, the storage molecule trehalose is highly
abundant in the cytoplasm of dormant Aspergillus nidulans conidia. However, it is not
metabolized in dormant conidia, indicating a lack of free trehalase enzymes. Trehalose is
rapidly mobilized and used as an energy source to fuel germination (Elbein 1974;
Svanström et al. 2014; Novodvorska et al. 2016).
In addition, dormancy factors can contribute to the maintenance of spores in a resting state.
In Rhizopus oligosporus the autoinhibitor, nonanoic acid, maintains dormancy through a pH
24

regulated germination inhibition mechanism (Breeuwer et al. 1997). A transcriptomic
approach which focused on dormant spores of A. niger, A. fumigatus and A. oryzae revealed
that the autoinhibitor bZIP-type transcription factor AftA plays a significant role in
maintaining dormancy. aftA mutants begin the process of germination much earlier
than wild type conidia, and before the correct nutrients are available. Regulators like
AftA are thought to be a common mechanism used by spores to maintain dormancy, with
AftA involved in the regulation of dormancy and germination through interaction with
conidia-associated genes such as the calA-family genes which encode a thaumatin-like
protein (Hagiwara et al. 2016).
Water availability and metabolic activity
The maintenance of little or no metabolic activity allows spores to preserve dormancy.
Metabolic activity of ‘dormant’ Aspergillus niger conidia measured in an
aqueous environment was seen to be severely reduced when compared to
germinated conidia. However, these readings may not present the true metabolic activity
of dormant spores, as the presence of water alone has been recorded to trigger
germination in A. niger (Teertstra et al. 2017; Novodvorska et al. 2016). Upon
germination, spores rapidly take in water (Turgeman et al. 2016), and maintaining a low
water content is thought to aid spore survival of desiccation. The water content of
Aspergillus oryzae conidia has been recorded to be as low as 17.8%, a third of the reported
percentage for S. cerevisiae yeast cells (Illmer, Erlebach, and Schinner 1999). It is thought
that the lower the water content, the better equipped the spores are to survive extreme
heat, although the thermostability of other spore components such as proteins and lipids
will also play a role in this context (Sumi 1928; Nicholson et al. 2000).
25

The Spore Germination Program
Spore polarization
Prior to hyphal emergence, polarization of the spores occurs. In many fungi this
process is reliant upon polarizomes, a protein complex which regulates polarity through
its effects on actin, chitosomes and microvesicles which support the synthesis of chitin
(Ruiz-Herrera and San-Blas 2003). This reliance appears conserved across fungi; work
done by Kamada et al. (1991) in Mucor rouxii spores demonstrates that
chitosomes act as a reservoir for chitin synthase throughout growth, as well as during
dormancy (Kamada, Bracker, and Bartnicki-Garcia 1991). The recruitment of polarizome
components and polarization-essential proteins has been well studied; during
germination the actin cytoskeleton of S. cerevisiae spores is regulated by polarizome
components Spa2p, Pea2p, Bud6p, Bni1p, Msb3 and Msb4. Over the course of
germination, these components, along with Ras2p, regulate the formation and
polarization of actin patches prior to polarized growth (Kono et al. 2005; Park and Bi
2007). The GTPase Cdc42p (cell division cycle 42) has been shown to be involved in
regulation and recruitment of the polarizome, as well as the GTP binding proteins known as
septins (Loeb et al. 1999; Bassilana, Blyth, and Arkowitz 2003; Momany and Talbot
2017). This conserved process of polarizome recruitment may also be
accompanied by coiled coil cytoskeletal proteins locating to the cell wall. These proteins
are thought to act as stress bearing structures to limit damage done by weakening the cell
wall through polarized growth (Fuchino et al. 2013). In P. discolor, this process is followed
by the formation of an ergosterol cap at the point of polarized growth (Van Leeuwen et
al. 2008). Small GTPases, such as Rho1p, are also involved in the regulation of
germination of S. cerevisiae (Kono et al. 2005). These hydrolytic enzymes have been
shown to be involved in cytoskeletal arrangement, exocytosis and cell wall composition
of the yeast. Cytoskeletal rearrangement also appears crucial to the
Spore polarization
26

polarization of Magnaporthe oryzae. The polarization of the plant pathogen Magnaporthe
oryzae has been shown to be regulated by the transcription factor Tpc1 (Transcription factor
for Polarity Control 1) which interacts with the pathogenicity factor Mst12, that regulates
infectious growth downstream of Pmk1 (mitogen-activated protein kinase 1) as well as
NADPH oxidase (G. Park et al. 2002). Tpc1 is dependent upon MAPK signalling for normal
regulation of cytoskeletal components, specifically F-actin components required for plant
invasion (Galhano et al. 2017). Kinase signalling is also implicated in polarization; the
pathogenic yeast Penicillium marneffei relies upon the p21-activated kinase pakA, which
localizes to the site of germination and plays a major role in the cAMP-PKA pathway, to
regulate polarization (Boyce and Andrianopoulos 2007). Kinases are also important for
polarization in S. cerevisiae; Cdk1 (Cyclin dependent kinase 1) and the cyclins Clb1-4
promote isotropic growth of the cells. The accumulation of Clb2 specifically within the
cytoplasm is thought to stabilize the switch between isotropic growth and
polarization (Machu et al. 2014). RAS signalling also appears important for germination
regulation; the G-protein alpha-subunits GasA and C are implicated in the regulation of
P. marneffei germination, both dependently and independently of RasA (Zuber, Hynes,
and Andrianopoulos 2003). GasA and C mutants also show defects in asexual
development, as well as conidial yield (Boyce and Andrianopoulos 2007; Boyce, Hynes,
and Andrianopoulos 2005; Pérez-Sánchez et al. 2010; Zuber, Hynes, and
Andrianopoulos 2003). Many pathways involved in polarization regulation appear
broadly conserved across filamentous fungal species, however it is likely there are also
many species specific germination processes (Huang and Hull 2017).
The polarization process in Mucor rouxii is impaired by the presence
of cAMP phosphodiesterase inhibitors, revealing that protein kinase A plays a
role 27

in the differentiation of M. rouxii spores. This is further evidenced by germ tube
emergence correlating directly with the amount of active PKA present, as threshold PKA
levels result in germ tube emergence (Pereyra, Mizyrycki, and Moreno 2000). PKA is also
important for the proper polarization of Neurospora crassa, and appears crucial to
germination of Aspergillus fumigatus conidia (Zhao et al. 2006; Bruno et al. 1996).
The mechanism of polarization and initial germination appears to be broadly conserved
across filamentous fungal pathogens (Boyce and Andrianopoulos 2015; D’Souza and
Heitman 2001; Fortwendel 2016), however with this conservation there appears to
be cross functionality: genes required for normal germination are frequently found to
have various other functions, often in asexual development and the production of conidia.
For example, the histidine kinase DRK1/drkA found in Blastomyces dermatidis,
Histoplasma capsulatum and P. marneffei is known to be required for sporulation,
virulence, cell wall integrity and production of infectious spores (Nemecek, Wuthrich, and
Klein 2006; Boyce et al. 2011).
Hyphal outgrowth and extension
Germ tube emergence follows swelling and is a result of polarized growth, with the hyphal
tip emerging from the site of polarization. Upon hyphal production, hyphae of most
Fusarium species will immediately penetrate agar under laboratory conditions, although a
few species produce hyphae that first grow along the top before penetration. As
pathogenesis is often initiated by conidial germination and germ tube production, the more
penetrating the species, the more pathogenic (Ruiz-Roldan et al. 2010; Petraitis et al.
2013b). For example, it is thought that the pathogenicity of Fusarium verticillioides
depends on the FPH1 (frustrated philade, conidiogenesis regulator) linked SIG1 (surface
vs. invasive germination), which regulates the invasive germination phenotype,
28

responsible for pathogenicity (Glenn, 2006).
To exit dormancy and establish growth, appropriate internal or external signals are required.
Germination can be initiated by the removal of dormancy factors, or by the introduction of
environmental cues. Similarly to the process of seed germination, the removal of dormancy
factors such as the dehydrin-like proteins DprA and DprB in Aspergillus fumigatus may induce
germination (Hoi et al. 2011), whilst the removal of the autoinhibitor methyl 3,4-
dimethoxycinnamate is essential for the germination of the plant pathogen Uromyces
phaseoli (Macko et al. 1970; Hogan 2006). Although this induction method may trigger
germination, it may not necessarily support full outgrowth of spores.
External cues capable of initiating germination and regulating growth include temperature,
light, nutrient availability, water availability, pH, quorum sensing molecules and osmolarity
(Alavi et al. 2013; Turgeman et al. 2016; Nguyen Van Long et al. 2017). Whilst species such
as Aspergillus niger can start germination when in contact with water alone, the initiation
of germination in Rhizopus delemar under starvation conditions induces atypical
morphology, fragmented DNA and increased susceptibility to cell death
(Turgeman et al. 2013; Novodvorska et al. 2016). Light responses have been well
studied in plants, although they do not all possess germinating spores. Dependent on the
wavelength, light is also capable of both inducing and inhibiting germination in fungi
(Franklin and Quail 2010; Röhrig, Kastner, and Fischer 2013; Possart, Fleck, and
Hiltbrunner 2014; Aron Maftei et al. 2014; Idnurm and Heitman 2005; Brunk et al. 2015).
29
Regulation of Germination

The nutritional environment and germination
An appropriate nutritional environment to support fungal growth is essential for initiation of
spore germination. The following sections will discuss the organic and inorganic nutrient
signals that can induce spore germination and sustain growth.
Organic nutrients
Organic cues are one of the most commonly used germination initiation factors for fungi.
Whilst carbon and nitrogen sources are generally indispensable for growth, macronutrients
known to trigger germination in various fungi are manifold and species-specific. Essential
nutrients for germination and activation of a higher metabolic rate in Rhizopus oligosporus
include glucose, phosphates and a mixture of amino acids, with leucine capable of triggering
germination (Thanh, Rombouts, and Nout 2005). Although Aspergillus niger may start to
germinate in water alone, D-tagatose, D-lyxose, and 2-deoxy-D-glucose will all trigger
germination, though they will not support outgrowth. For outgrowth D-glucose, D-mannose,
or D-xylose is required (Hayer, Stratford, and Archer 2013). However, not all carbon sources
are capable of triggering germination. For example D-galactose, L-glucose, and D-arabinose
have all been shown to be incapable of triggering germination in Aspergillus niger, whilst they
may support outgrowth if a complementary triggering sugar is present (Hayer, Stratford, and
Archer 2013). This supports the notion that different stages of germination may be carried
out regardless of whether conditions will support full germination (Hayer, Stratford, and
Archer 2013). Similarly, spores of Cryptococcus neoformans are able to germinate on minimal
water agarose medium, suggesting nutritional cues are not needed to trigger germination for
this species (Velagapudi et al. 2009). Blastomyces dermatitidis spores will germinate on a low
30

glucose substrate, but can also utilize complex carbon sources from decomposing matter, a
useful trait for exploiting carbon sources available in the environment. Additionally, B.
dermatitidis requires a nitrogen source such as allantoin, creatinine, quanidoacetic acid,
guanidine or cysteine. Interestingly, B. dermatitidis is also capable of growth at extremely
high ammonia concentrations, of up to 42-62mmol/l. This is particularly significant as the
growth of most soil inhabiting fungi will be inhibited at 2.1-4.2mmol/l. The ability to grow in
such extreme conditions provides B. dermatidis with an environmental niche which it may
exploit to increase its ubiquity (Baumgardner and Laundre 2001; Baumgardner 2009). The
composition of the carbon source can even lead to opposing effects on germination, whilst
L/D-leucine and/or unsaturated long chain fatty acids will induce germination in Microsporum
gypseum, saturated short chain fatty acids are inhibitory towards germination (Barash,
Conway, and Howard 1967).
Sulphur, a constituent key to life and a component of a few key amino acids, is required for
the growth of many fungal species (Marzluf 1993). It enables strengthening and proper
folding of protein structures through disulphide bridges, and is key to the functionality of
many essential enzymes such as permeases and proteases. A source of sulphur such as biotin,
thiamine or thioctic acid is required for the formation of Histoplasma capsulatum yeast cells.
Whilst it is not required for the formation of hyphae, both mycelial and yeast forms utilise
sulphur metabolism. It has been suggested that the sulphur requirement of the dimorphic
fungus Histoplasma capsulatum, specifically cysteine, is temperature dependent, indicating
the sulphur metabolism may be important for morphology and pathogenicity (Howard et al.
1980; Maresca and Kobayashi 1989).
31

Inorganic nutrients
Alongside macronutrients, micronutrients are also required by many fungi to initiate
germination and sustain growth. For example, studies investigating calcium metabolism in
Sporothrix schenckii showed that the presence of exogenous calcium stimulated mycelial
production, with hyphal emergence corresponding to calcium uptake (Rivera-Rodriguez and
Rodriguez-del Valle 1992).
Iron is essential for the proper growth of many fungi (Tamayo et al. 2014; Philpott 2006; C.
Zhang 2014). Although too little iron may result in growth defects, too much can also be
extremely toxic. Tight regulation of iron homeostasis is therefore essential. External
siderophores are often used to scavenge iron from the surrounding environment, whilst
internal siderophores store iron and limit toxicity. Aspergillus species employ the two
transcription factors SreA and HapX to regulate iron homeostasis via siderophores, such as
the intracellular siderophore SidC. Aspergillus nidulans conidia lacking SidC are known to be
more susceptible to oxidative stress and exhibit delayed germination when grown in
reduced iron environments (Eisendle et al. 2006). Interestingly, genes coding for
the biosynthesis of triacetylfusarinine C, a common fungal siderophore, are absent in
Aspergillus niger, indicating that there are many homeostatic mechanisms which
are implicated in maintaining the correct metal homeostasis (Franken et al.
2014). The siderophore rhizoferrin is used by Mucorales species to scavenge iron
from the surrounding environment. When Mucorales spores are germinated in iron
limited conditions, they can be seen to produce atypical hyphae (Lewis et al. 2011;
Kousser et al. 2019) and iron availability is known to regulate virulence in various
Mucorales species (Ibrahim 2011). Notably, Mucorales species such as Rhizopus
delemar may also bind iron provided by exogenous siderophores, such as the iron
overload treatment deferoxamine, increasing spore germination.
32

In turn, this leads to higher susceptibility to mucormycosis in individuals who have elevated
iron levels within their blood e.g. those with uncontrolled diabetes, patients receiving
blood transfusions or those suffering from hemochromatosis. The combination of
increased iron levels and treatment with the siderophore deferoxamine creates an
optimal growth environment for the spores of Mucorales species (Boelaert et al. 1993;
Gebremariam et al. 2016; Spellberg et al. 2016; Spellberg, Edwards, and Ibrahim 2016;
Ibrahim, Spellberg, and Edwards 2016).
Micronutrients are often required by spores to overcome environmental and host stresses.
Spores utilize enzymes such as catalase and superoxide dismutase (SOD) to avoid damage by
reactive oxygen species (ROS). These enzymes are dependent on the presence of transition
metals, namely Zn and Cu, to be able to perform their redox functions. Levels of SOD and
catalase were shown to increase in Aspergillus niger grown at extreme temperatures,
demonstrating their roles in survival in harsh conditions (Abrashev et al. 2005). SOD1, 2 and
3 are all required for the correct germination and outgrowth of Aspergillus fumigatus, whilst
deletions of individual SOD proteins alone leads to growth perturbations and increased
sensitivity to ROS (Lambou et al. 2010; Plante et al. 2017). Similar work done in Neurospora
crassa showed that catalase is important for the survival and viability of conidia. Catalase-1
mutants showed an increased susceptibility to hydrogen peroxide and a decreased ability to
germinate after exposure to light, when compared to the wild type control (Wang, Yoshida,
and Hasunuma 2007).
The acquisition of micronutrients from low availability environments allows many fungal
pathogens to survive and proliferate within hosts (Ballou and Wilson 2016). Nutritional
33

34
immunity is recognized as a way in which a host may limit the growth of a pathogen.
However, pathogens have developed high specificity scavenging systems to combat this.
Using high affinity chelators regulated by transcription factors such as ZafA (Aspergillus
fumigatus), Csr1 (Candida albicans) and other Zap1 orthologues to regulate zinc
homeostasis, pathogenic fungi may scavenge zinc from the host environment
(Böttcher et al. 2015; Moreno et al. 2007). ZafA is required for the correct germination of
Aspergillus fumigatus (Moreno et al. 2007), whilst Zap1 orthologues are thought
to contribute towards virulence of fungi. Similarly, copper homeostasis has been
shown to be important for the pathogenicity of fungi. Copper is also
essential to the germination of Schizosaccharomyces pombe. If lacking copper
transporters Ctr4 and 5, germination is halted at hyphal production, whilst the copper
transporter Ctr6 has also been suggested to play a role in germination. Ctr6 is localised
to the spore membrane at the end of the sporulation process, and spores lacking Ctr6
show decreased viability when grown in a low copper environments, than those with a
functioning Ctr6 (Plante et al. 2014).
Micronutrients are not only required for the initial growth of pathogenic fungi, but often
they are also implicated in ability of fungi to switch between dimorphic growth forms, as
is the case for Blastomyces dernmatidis. B. dermatidis is known to produce a
siderophore which aids in pathogenicity. Although it is capable of
growth without supplementary ferric iron, Blastomyces requires exogenous
magnesium and calcium for hyphal growth (Giles and Czuprynski 2004). The GATA
transcription factor SREB, found frequently amongst fungal species, has been shown
to regulate siderophore production in Blastomyces dermatidis, as well as lipid
droplet formation, triacylglycerol and ergosterol synthesis. As lipid droplets are
thought to be used as energy sources for Blastomyces dermatidis when transitioning
from yeast to mycelial growth, SREB is also essential for hyphal growth (Gauthier et al.
34

2010; Marty et al. 2015). Histoplasma capsulatum, another dimorphic pathogen, utilizes a
siderophore for pathogenicity and iron scavenging within host cells. Histoplasma
capsulatum has been shown to scavenge iron from host ferritin/fe-transferrin with the
SID1 encoded ferric reductase. Without the iron scavenger, Histoplasma capsulatum
is rendered incapable of replicating within host macrophages (Newman and
Smulian 2013). Histoplasma capsulatum also utilises the transition metal requiring
extracellular SOD and catalase enzymes to survive host ROS. As a result, decreasing the
levels of iron and zinc available to Histoplasma capsulatum leads to higher
susceptibility to macrophage killing (Garfoot and Rappleye 2016).
Germination Regulation via pH, Temperature, Light and Environmental Gases
Several exogenous factors, other than nutrient availability, are known to regulate
germination of fungi. The abiotic factors discussed below may be important signals that aid
pathogenic fungi to exploit their host environments at choice moments, or they may
be signals that it is not yet a suitable environment in which to germinate.
pH
pH has been well established as a factor which influences germination in many fungal
species and many pathogenic fungi grow best at the pH of their host
environment. The host environment may also be liable to pH change. Once
phagocytosed by host phagocytes, fungi are often subject to drastic pH changes within
the phagosome, as the phagocytes attempt to disarm the phagocytosed fungi through
fusion and fission events with both endosomes and lysosomes. Often, a matured
phagosome will degrade the captured microbe, therefore maturation arrest (or
phagosome escape) is a tactic often employed by fungi such as C. neoformans, to
avoid degradation (Smith, Dixon, and May 2014; Yates, Hermetter, and Russell 2005;
Gresnigt et al. 2018).35

The germination of the Mucorales species Rhizopus delemar is induced at lower pHs; this is
true in both artificial growth media, along with in vivo replicating conditions (Turgeman et al.
2016). The pH drop of the blood of diabetics undergoing a ketoacidic phase has even been
linked to an increased susceptibility to mucormycosis, as the decreased pH in turn leads to
an increased availability of iron and beta-hydroxybutyric acid for the fungus (Artis et al.
1982; Gebremariam et al. 2016). The pH dependent induction of germination in Rhizopus
delemar is known to be regulated by two aquaporins (RdAQP1 and RdAQP2). The optimal
pH for germination initiation was found to be between pH 4-5 (Turgeman et al. 2016).
Other fungi are capable of growing at a vast range of pH’s. The human fungal pathogen
Sporothrix schenckii is found ubiquitously in soils and can grow between pH 3.5-9.4 (Tapia
Noriega et al. 1993), whilst Candida albicans is capable of colonizing host environments of
extreme pH’s and growing in pH’s from 2-10 (Odds 1988). Many fungi rely on the zinc finger
transcription factor PacC, and the signalling pathway in which it is implicated, to sense the
pH of their surroundings. The PacC signalling pathway has also been shown to effect
germination in Aspergillus nidulans. Conidia lacking PacC components showed decreased
growth, when compared to the wildtype, whilst the acquisition of iron is also thought to be
dependent on PacC in A. nidulans (Bertuzzi, Schrettl, Alcazar-Fuoli, Cairns, Muñoz, et al. 2014).
A. nidulans further relies on the signal transduction components encoded by the PalA-I genes
to respond to environmental pH (Denison 2000). Given a suboptimal pH, fungi may modulate
the pH of their surrounding environment through the release of pH altering molecules.
Through the secretion of organic acids and ammonia, fungi are able to either increase or
decrease the pH of their surroundings (Cornet and Gaillardin 2014; Vylkova 2017).
36

Temperature
In some fungal species, temperature has been shown to effect germination more than other
factors. The germination of Cryptococcus neoformans was shown to be more reliant on
temperature than it is on nutrient availability (Forsythe, Vogan, and Xu 2016). The small
GTPase RAS1 has been shown to regulate the filamentous growth of Cryptococcus
neoformans through the mitogen-activated protein kinase (MAPK) and RAS specific signalling
pathways in response to shifting to higher temperatures (Alspaugh et al. 2000). Neurospora
crassa mcb strains which lack the cAMP dependent protein kinase are incapable of polarized
growth at higher temperatures, demonstrating that the complex triggers for germination are
interlinked (Bruno et al. 1996). Interestingly, Aspergillus nidulans RasA, which is also involved
in the polarization of conidia, regulates the germination of conidia in response to nutrients
through possible interactions with heat sensitive factors encoded by spore germination-
deficient (sgd) genes. These temperature dependent factors are also known to be important
for germination and polarized growth (Osherov and May 2000). Studies investigating the
impact of temperature on A. fumigatus germination revealed that over the course of
germination higher temperatures (37°C) upregulated carbohydrate, lipid and secondary
metabolism pathways, whereas lower temperatures (24°C) upregulated RNA metabolism and
processing (Sueiro-Olivares et al. 2015).
Whilst spore production is often induced in suboptimal conditions as a survival mechanism,
some fungi, such as the blight- causing fungus Exserohilum monoceras, require
specific temperatures in order to produce spores. The optimum temperature for
Exserohilum monoceras growth and spore production is between 27°C and 28°C (Zhang and
Watson 1997).
37

Light
Environmental factors such as light play a role in the regulation of spore germination and
growth in some species. Light induces germination and hyphae formation in arbuscular
mycorrhizal species such as Gigaspora gigantea, but will inhibit germination and growth of
many other fungal species, such as Puccinia graminis and Aspergillus fumigatus, dependent
on the wavelength (Nagahashi, Douds, and Buee 2000). The crop pathogen Puccinia
graminis showed inhibited germination when exposed to both white and far red
light, whilst germination occurred normally in dark conditions (Lucas, Kendrick, and
Givan 1975). Aspergillus fumigatus conidia showed decreased germination rates when
exposed to blue light, red light and far red light (Fuller et al. 2013; Röhrig, Kastner, and
Fischer 2013).
Light is sensed by fungi through photoreceptors which interact with and bind to fungal
chromophores, ultimately transmitting and eliciting a cellular response to light. Light
responses through photoreceptors have been studied extensively in the fungal plant
pathogen Neurospora crassa. It has been shown that both conidia production and release can
be regulated by the presence of blue light, through the con-6 and con-10 light responsive
genes (Linden, Rodriguez-Franco, and Macino 1997). A whole transcriptome response to light
exposure revealed that transcripts con-6 and con-10 were not only upregulated upon light
exposure, but also play a role in the germination of N. crassa conidia (Wu et al. 2014).
Similarly, con-6 and 10 homologues conF and J were found to induce conidia formation in
Aspergillus nidulans. A double mutant also harboured germination defects, demonstrating
the role of blue and red light to be involved in both conidia formation and germination,
which appears to be conserved across filamentous fungi (Suzuki et al. 2013). It has been
38

suggested that the light signalling pathways, such as the cellular response to blue light, in
Neurospora crassa may be dependent on internal ROS, as SOD1 mutations showed
increased light dependent accumulation of metabolites, suggesting ROS factors resulting
from light exposure may act as alternative signals to transmit the signalling responses
to light (Yoshida and Hasunuma 2004; Belozerskaya et al. 2012). In order to overcome the
oxidative stress which light induces, Neurospora crassa upregulates the production of
antioxidants and protective photopigments in response to light exposure (Wu et al. 2014).
Light exposure also increased the resistance of A. fumigatus conidia to UV and ROS,
demonstrating the ability to respond in a protective manner to potentially harmful stimuli
(Fuller et al. 2013; Röhrig, Kastner, and Fischer 2013). Alternate mechanisms such as
proton pumps are also activated by light; the light activated proton pump, CarO, is
involved in the formation of Fusarium fujikuroi hyphae, with hyphal growth rates
decreased under light conditions (Brunk et al. 2015; Garcia-Martinez et al. 2015).
The connection between light and germination has been exploited in both medicine and
agriculture (Lucas, Kendrick, and Givan 1975; Schmidt-Heydt et al. 2011; Baltazar et al. 2015;
Trzaska et al. 2017). Pulsed light containing UV spectra decreases the ability of spores to
germinate on food sources such as wheat grain, providing a useful food spoilage preventative
measure (Aron Maftei et al. 2014).
Gases
Atmospheric gases are most commonly used within food packaging to inhibit fungal
germination, to prevent food spoilage. Through [O2] reduction, or [CO2] increases (15-20%),
spoilage by species such as Rhizopus stolonifer is successfully controlled (Barkai-Golan 2001).
39

Although higher CO2 concentrations often result in germination inhibition, germination is
supported in Fusarium roseum at up to 32% CO2 (Wells and Uota 1970), and Rhizopus
oligosporus germination is supported by CO2 when O2 becomes limited (de Reu et al. 1995).
The signalling molecules known as quorum sensing molecules are capable
of regulating growth in both plant and human pathogenic fungi. One of the best-studied
examples is the quorum sensing molecule farnesol, a secondary metabolite produced
by Candida species when in stationary phase, capable of modulating the innate
immune response (Leonhardt et al. 2015) and inhibiting Candida albicans hyphal
growth at high concentrations (Piispanen et al. 2011). Farnesol is capable of regulating the
growth phase of Candida albicans populations, as well as biofilm formation (Ramage et al.
2002). Even at low cell densities, a condition which usually induces hyphal growth in
Candida albicans, it will not form hyphae if the quorum sensing molecule farnesol is
present (Kruppa et al. 2004). Furthermore, farnesol, produced in higher quantities
by Candida albicans grown between 37 and 40 °C, is known to inhibit the
production of hyphae by disrupting the degradation of the hyphal growth repressor
Nrg1. Nrg1 must be both down regulated via the cAMP-PKA pathway, as well as
degraded for Candida albicans to produce hyphae, a complex regulation which controls
the morphogenesis of Candida albicans and the resulting pathogenicity (Lu et al. 2014).
Farnesol has also been shown to inhibit the germination of the common plant
pathogen Fusarium graminearum, with higher concentrations of purified farnesol inducing
apoptosis (Semighini, Murray, and Harris 2008). Similarly, the presence of farnesol was
40
Signalling Molecules

shown to impair growth of Aspergillus nidulans and induce apoptosis like characteristics in
the hyphal form of Aspergillus niger (Semighini et al. 2006).
Whilst bacteria have been shown to modulate germination through nutrient competition
(Kousser et al. 2019), the presence of the bacterial and fungal cell wall component GlcNac is
also capable of inducing the switch to filamentous growth in Blastomyces dermatidis,
Histoplasma capsulatum and Candida albicans. This response in Histoplasma capsulatum
is thought to be mediated by the GlcNac transporters NGT1/2, required for the
filamentous response (Gilmore et al. 2013), whilst the DNA-binding containing RON1 is
involved in regulating this switch in C. albicans (Naseem et al. 2017).
Hormones produced by both plants and fungi are also capable of regulating the
germination of select fungal species. Gibberellic acids have been shown to stimulate the
germination of the plant pathogen N. crassa, whilst auxins show a dose dependent
modulation of germination. At low concentrations, auxins promote germination of the
plant pathogen Fusarium delphinoides, whilst at high concentrations it is inhibitory towards
germination (Kulkarni et al. 2013; Tomita, Murayama, and Nakamura 1984). The antagonist
to gibberellic acids, abscisic acid, has been shown to promote germination of the rice
blast fungus M. oryzae (Spence et al. 2015). Jasmonic acid was shown to decrease
germination of Fusarium oxysporum spores, whilst the germination of corn pathogen
Harpophora maydis can be subdued by the presence of sialic acid (Krol et al. 2015; Degani,
Drori, and Goldblat 2015). The gaseous hydrocarbon and plant hormone, ethylene, is
released by plants and known to initiate the ripening of fruits. Ethylene is also capable of
triggering germination of the plant pathogen Colletotrichum musae, so that germination
occurs simultaneously with fruit ripening. Upon germination, the fungus releases
extracellular cutinase; this enzyme hydrolyses the waxy cutin layer coating the
41

plant, enabling pathogen access to the cuticle of the plant. Other plant pathogens,
such as Fusarium solani pisi, even store the cutinase enzyme in the spores, enhancing
access to the plant upon spore contact and germination (Kolattukudy et al. 1995).
Plant hormones have also been shown to modulate the growth of some human fungal
pathogens. For example, sialic acid is capable of decreasing hyphal growth of Aspergillus
flavus. However, it is striking that plant hormones have mainly been seen to regulate the
germination of classical plant pathogens which rarely cause human infections. This suggests
fungal plant pathogens may have evolved or diverged to produce hormones which are almost
identical to those mentioned above, allowing fungi to regulate the growth of the plants which
they rely on for nutrients and leading to a dynamic co-regulation of growth between fungi
and their hosts.
42

Materials and Methods
This work has been adapted from the following articles: “Pathways of Pathogenicity:
Transcriptional Stages of Germination in the Fatal Fungal Pathogen Rhizopus
delemar”(Sephton-Clark et al. 2018) and “Host-pathogen transcriptomics of macrophages,
Mucorales and their endosymbionts: a polymicrobial pas de trois” (Sephton-clark et al.
2019). For both I conceived, designed and performed the experiments, collected the data,
performed the analysis and interpretation, wrote the manuscript, completed revisions, and
prepared the figures.
43

Microbial Culture Fungal Culture Spore Isolation
Spores were harvested from 10 day old cultures maintained on sabouraud
dextrose agar plates by washing with 10ml of phosphate buffered saline (PBS) (Sigma-Aldrich).
Spores in PBS solution were centrifuged for 3 minutes at 3000 rpm and washed with 5ml
of PBS. The pellet was resuspended in 3-5ml of PBS and the concentration of spores in the
suspension calculated by counting with a haemocytometer.
Spore Growth
Sabouraud Dextrose Broth
Spores harvested as above were grown in Sabouraud (SAB) broth (10 g/liter mycological
peptone, 20 g/liter dextrose), sourced from Sigma-Aldrich, at a range of temperatures (RT
to 37°C, protocol dependent), at 100-250 rpm.
Sabouraud Dextrose Agar Plate
100-200 spores, harvested as above, were spread evenly onto SAB agar plates.
Plates were incubated lid side up away from light at RT for 2-3 days, before being
turned (lid side down) and incubated in the same conditions for 7-8 days.
Endosymbiont Curing
To cure spores of their respective bacterial endosymbionts, spores were cultured with
ciprofloxacin, as described in Itabangi et al. 2019 (Itabangi et al. 2019). Briefly, spores were
passaged for a month in 60 μg/mL ciprofloxacin and cultured on ciprofloxacin plates
prior to use. Once cured, spores were sub-cultured at least twice in ciprofloxacin-free
media before use.
44

Bacterial Culture
Bacterial endosymbiont species, isolated by Herbert Itabangi, were grown in lysogeny broth
(also referred to as Luria-Bertani media) (LB), sourced from Sigma-Aldrich, at 37°C, for
12 hours, with 250rpm shaking prior to processing for genomic DNA extraction.
45

Germination Phenotyping
Spore Germination Assay
Live cell Imaging
Spores were harvested as above and used at 1x105 Spores/ml in 200μl of Sabouraud dextrose
Broth. Images of the spores were taken every 10 minutes over the course of 24 hours with
brightfield microscopy. Images were taken at 20X objective on a Zeiss Axio Observer. Images
were analysed with ImageJ V1.
Flow Cytometry
Spores were harvested as above and used at a concentration of 2x107 Spores/ml in 200μl of
SAB. Spores were incubated for between 0-6 hours, then fixed in 4% paraformaldehyde (PFA),
sourced from Sigma-Aldrich and prepared according to manufacturer’s instructions, before
staining with either calcofluor white (CFW) or Fluorescein isothiocyanate isomer I (FITC).
Spores stained with 250-1000 μg/ml of CFW were kept at RT, for 20 minutes, prior to washing
with PBS. Spores stained with 100 μg/ml FITC were kept at RT, for 30 minutes, prior to washing
with PBS.
XTT Assay
The XTT assay was used to measure metabolic activity of spores and germlings
(Antachopoulos et al. 2006; Moss et al. 2008). To germinating spores, XTT sodium salt and
menadione were added as described by Antachopoulos et al. After 80 minutes, optical density
readings at 450nm were taken, these reading correspond to metabolic activity. Readings were
46

also taken at 600nm to determine fungal biomass. Readings were corrected by the
appropriate controls during analysis.
Exosome Release
Spores were harvested as above and grown in SAB broth for 0-6 hours at 25°C. The
extracellular vesicles were isolated according to manufacturer’s instructions Exo-spin™.
Extracellular vesicle (EV) size was determined with dynamic light scattering (dls), a method
which determines the size of particles dispersed in a liquid, based on the detection of laser
beam scattering.
47

Genomic DNA Extraction, Sequencing and Analysis
Genomic DNA Extraction
Fungal DNA Extraction
Spores germinated in SAB broth at 37°C, 100-200 rpm shaking, for 12 hours were pelleted at
3000rpm, and washed in PBS. To washed spores 1ml of Qiagen DNEasy PowerLyzer Microbial
Kit lysis buffer (Solution SL) was added. The mixture was bead beaten at 6500 rpm for 2
rounds (30 seconds per round). Samples were immediately placed on ice post beating and
DNA was extracted from the resulting solution as per Qiagen DNEasy PowerLyzer Microbial
Kit instructions (column based extraction). Nanodrop measurements were taken to
determine DNA quality, and agarose gel electrophoresis (1%) indicated DNA integrity.
Bacterial DNA Extraction
Bacterial cells grown in LB broth at 37°C, 100-200 rpm shaking, for 12 hours were processed
according to the of Qiagen DNEasy PowerLyzer Microbial Kit (column based extraction).
Nanodrop measurements were taken to determine DNA quality, and agarose gel
electrophoresis indicated DNA integrity.
Genomic DNA Sequencing
Fungal Sequencing
Library preparation and sequencing of genomic DNA was performed by MicrobesNG. Briefly,
Nextera XT Library Prep Kit was used for library preparation and pooled libraries were
quantified using the Kapa Biosystems Library Quantification Kit for Illumina. Libraries were
48

sequenced on the Illumina HiSeq using a 250bp paired end protocol. 30X coverage was
obtained for all fungal species sequenced.
Bacterial Sequencing
Library preparation and sequencing of genomic DNA was performed by MicrobesNG. Briefly,
Nextera XT Library Prep Kit was used for library preparation and pooled libraries were
quantified using the Kapa Biosystems Library Quantification Kit for Illumina. Libraries were
sequenced on the Illumina HiSeq using a 250bp paired end protocol. 5X coverage was
obtained for all bacterial species sequenced. Species identification was carried out with
Kraken(Wood and Salzberg 2014).
Genome Sequence Analysis
Rhizopus microsporus Genome Assembly
Raw reads obtained from the MicrobesNG protocol above were quality checked with FastQC
(Andrews). Reads were assembled with SPAdes (Bankevich et al. 2012), using the R.
microsporus assembly (Mondo et al. 2017) as a reference (-untrusted contigs).
Rhizopus microsporus Variant Identification
Raw reads obtained from the MicrobesNG protocol above were aligned to the R. microsporus
assembly (Mondo et al. 2017) with bwa (Li and Durbin 2010). Samtools (Li et al. 2009)
and Picard (Broadinstitute) were used to extract alignment metrics. Haplotypecaller
(McKenna et al. 2010) was used to identify single nucleotide polymorphisms (SNPs) in the
aligned reads. SNPs identified in the aligned reads were used to replace their alternatives in
49

the reference genome with GATK (FastaAlternateReferenceMaker) (McKenna et al. 2010).
Subsequent reads generated via RNA-Seq were aligned to this hybrid.
Comparative Genomics
Fisher's exact test was used to detect enrichment of Pfam terms between Rhizopus
delemar and Rhizopus microsporus (R. delemar & R. microsporus), terms with a corrected
(FDR, false discovery rate) P value of < 0.01 were considered significant. Orthologue genes of
R. delemar and R. microsporus were identified using blast+. R (version 3.3.3) was used to carry
out hypergeometric testing of KEGG and GO terms to determine enrichment.
Orthologue genes of R. delemar and Aspergillus niger were identified using blast+.
The genome of R. delemar was reannotated by incorporating the RNA-Seq data via
BRAKER (version 2.1.0) (Hoff et al. 2016), this was fed into the Broad Institute
annotation pipeline, which removed sequences that overlapped with repetitive elements,
numbered, and named genes as previously described (Haas et al. 2011). Completeness of
annotation was analyzed with BUSCO (version 3) (Simão et al. 2015). Duplicated gene pairs
were identified based on parameters outlined by Ma et al (Ma et al. 2009) with
Python (version 3.5.5). GO term enrichment of duplicated gene pairs was determined
in R (version 3.3.3). Pathway Tools (version 21.0) was used to obtain information on
specific pathways (Karp et al. 2016).
Comparative Genomics
Rhizopus delemar Genome Annotation Update
50

Transcription and Inhibition of Rhizopus delemar Germination
Germination RNA-Seq
RNA Isolation
Total RNA was extracted from R. delemar spores (harvested as above) which germinated in
SAB broth for 0,1,2,3,4,5,6,12,16 and 24 hours. To extract Total RNA, the washed samples
were immediately immersed in 1ml of Trizol and lysed via bead beating at 6500 rpm for 2
rounds, 30 seconds each. Samples were then either immediately frozen at -20°C and stored
for RNA extraction, or immediately placed on ice for RNA extraction.
All further work was done on ice to inhibit RNases. After lysis via bead beating, samples were
moved to clean Eppendorf tubes and 0.2ml of chloroform was added for every 1ml of Trizol
used in the sample preparation. Samples were incubated for 3 minutes, then spun at 12,000
G at 4°C for 15 minutes to separate out the aqueous and organic phases.
The aqueous phase, containing RNA, was placed into a fresh tube and the interphase and
organic phase containing proteins and DNA were stored at -20°C for any further work. To the
aqueous phase, an equal volume of 100% EtOH was added, before the samples were loaded
onto RNeasy RNA extraction columns. The columns were spun at 8000g for 30 seconds at 4°C
and the flow through discarded as RNA should now be bound to the silica column. 700µl of
buffer RW1 was added to remove and carbohydrates, lipids or proteins bound to the
column. It should be noted that at this point, only RNA longer than 200bp will remain
bound to the membrane, therefore if regulatory, small non-coding RNA etc... is also
being isolated, this method will not be suitable. The column was spun at 8000g for 30
seconds at 4°C and the
51

flow through discarded. The washing buffer RPE was then added to the column to remove
any contaminating salts which may affect results seen with the NanoDrop, or any other
equipment which relies on measuring absorbance to determine quantity and quality of RNA.
500µl of the buffer was added to the column, followed by a spin at 8000g for 30 seconds at
4°C, the flow through discarded. Another 500µl of the buffer was added to the column,
followed by a spin at 8000g for 2 minutes at 4°C, the flow through discarded. The RNA was
then eluted in two rounds, each using 30µl of RNase free dH2O to elute RNA by spinning the
columns at 8000g for 1 minute at 4°C. 3µl aliquots of each sample was then taken for testing
with the Nano drop, whilst the remainder of the sample was immediately frozen and stored
at -20°C.
Three biological repeats were performed and the quality of the resulting RNA was
checked by Agilent to ensure all RIN scores were above 8, indicating that there was
minimal RNA degradation within the sample and a good ribosomal RNA ratio (Schroeder et
al. 2006). 1ug of Total RNA was used for library preparation.
The following Library preparation was performed in accordance with the NEBNext pipeline.
mRNA was isolated with NEB’s Poly(A) magnetic isolation module. The isolated mRNA
was then fragmented to around 200bp with Mg2+ ions. From this, ds cDNA was
produced and underwent PCR amplification and indexing. The quality of the library was
checked by Agilent BioAnalyzer and the concentration was determined by qPCR prior to
sequencing.
52

Samples were sequenced using the Illumina HiSeq platform. 15 samples were loaded per lane,
with 140 million reads expected per lane. 100bp paired end sequencing was employed
(2X100bp).
FastQC (version 0.11.5)(Andrews) was employed to ensure the quality of all samples, a Phred
value of over 36 was found for every sample. Hisat2 (version 2.0.5) (Kim, Langmead, and
Salzberg 2015) was used to align reads to the indexed genome of R. delemar (JGI:
PRJNA13066) (Ma et al. 2009). HTSeq (version 0.8.0) was used to quantify the output
(Anders, Pyl, and Huber 2015). Trinity and edgeR (version 3.16.5) were then used to analyze
differential expression (Grabherr et al. 2009). Pathway Tools (version 21.0) was used to
obtain information on specific pathways (Karp et al. 2016). Cytoscape (version 3.5.1) was
used to analyse co-transcriptional networks.
Germination Inhibition
Inhibition Assessment of Targets Determined via RNA-Seq
Images of 1X105 spores/ml in SAB were taken every 10 minutes to determine germination
characteristics. Images were taken at 20X objective on a Zeiss Axio Observer. Calcofluor white
(CFW), fluorescein isothiocyanate (FITC) (Sigma-Aldrich), and the ROS stain carboxy-H2DCFDA
(6-carboxy-2=,7=-dichlorodihydrofluorescein diacetate [C400]; Invitrogen) were incubated
with live spores, according to the manufacturer’s instructions, prior to imaging. To assess
inhibition, spores were incubated with either: 1 to 5 mM hydrogen peroxide, 1.5 to 10 nM
antimycin A (Sigma-Aldrich) or Nikkomycin Z 120ug/ml prior to imaging. Bright-field and
fluorescent images were then analyzed using ImageJ V1.
Data Analysis
53

Strathclyde Natural Compound Library
Spores were harvested as above and used at a concentration of 1x105 Spores/ml in 200ul of
SAB. To measure metabolic activity the XTT assay was used (Antachopoulos et al. 2006; Moss
et al. 2008). The compound A10 was added to germinating spores at concentrations of 1000,
100, 75, 50, 25, 10, 1, and 0 ug/ml. To control for effects introduced by the extract solvent
(DMSO), DMSO was added to samples at an equal volume to the highest volume of A10
extract used. 3% Acetic Acid was used as a positive control. To measure metabolic activity and
inhibition the XTT assay was used (Antachopoulos et al. 2006; Moss et al. 2008). Optical
density readings at 450nm were taken at 80 minutes after the addition of the XTT mix.
Readings were also taken at 600nm to determine biomass.
54

Rhizopus-macrophage interactions
Macrophage
Macrophage Culture
Macrophages from the J774.A1 cell line were cultured in Dulbecco’s Modified Eagle Medium
(DMEM), (complemented with 10% foetal bovine serum, 1% penicillin, 1% streptomycin and
1% L-glutamine), sourced from Sigma-Aldrich. Macrophages were grown at 37°C, in 5% CO2.
In vitro Phagocytosis assay
Macrophages were incubated for one hour in serum-free DMEM prior to infection. Spores
were pre-swollen in SAB (2 hr for R. delemar, 4 hr for R. microsporus). Washed spores were
incubated at 5:1 MOI with 1 x 105 macrophages as described in Itabangi et al (Itabangi et al.
2019), to ensure that >95% of macrophages contained one spore or more. After a 1 hour of
incubation, excess spores were washed off the surface and the macrophages were incubated
for a further 2 hours, before processing for RNA-Seq experiments. For live cell imaging
experiments, images were taken starting immediately after the excess spores were removed.
Phagocytosis Live Cell Imaging
Time course images were taken to determine how LPS pre-treatment (100ng/ml) (Myers,
Tsang, and Swanson 2010) of macrophages, and Nikkomycin Z pre-treatment (120ug/ml over
the course of swelling in SAB) of spores effected phagocytic outcome. Images were taken at
20x on a Zeiss Axio Observer, with images taken every 5 minutes. Bright-field and fluorescent
images were then analysed using ImageJ V1.
55

RNA-Seq
Bulk Rhizopus RNA-Seq
RNA was extracted from spores which had either been incubated with the macrophages, or
incubated in DMEM for the equivalent time period. To remove the macrophages, triton at 1%
was used to lyse the macrophages, the resulting solution was then centrifuged for 3 min at
3,000 rpm, and washed, leaving only spores. The DMEM control also received the same
treatment. To extract total RNA, the washed samples were immediately immersed in TRIzol
and lysed via bead beating at 6,500rpm for 60s. Samples were then either immediately frozen
at −20°C and stored for RNA extraction or placed on ice for RNA extraction. After lysis, 0.2 ml
of chloroform was added for every 1 ml of TRIzol used in the sample preparation. Samples
were incubated for 3 min and then spun at 12,000 g at 4°C for 15 min. To the aqueous phase,
an equal volume of 100% ethanol (EtOH) was added, before the samples were loaded onto
RNeasy RNA extraction columns (Qiagen). The manufacturer’s instructions were followed
from this point onwards. RNA quality was checked by Agilent, with all RNA integrity number
(RIN) scores above 7 (Schroeder et al. 2006). One microgram of total RNA was used for cDNA
library preparation. Library preparation was done in accordance with the NEBNext pipeline,
with library quality checked by Agilent. Samples were sequenced using the IIlumina HiSeq
platform; 100-bp paired-end sequencing was employed (2 × 100 bp) (>10 million reads per
sample).
Single Cell Macrophage RNA-Seq
For single cell sequencing experiments, macrophages were infected with fungal spores, as
outlined above. Uninfected macrophages, used as a negative control, were treated in the
56

same manner and underwent mock washes and media changes at identical time points to
infected macrophages. Macrophages were isolated and released from the bottom of their
wells with accutase, as per manufacturer’s instructions (Technologies). Once in solution, the
macrophages were loaded onto the 10X genomics single cell RNA sequencing pipeline for
single cell isolation and library preparation. In total, 1082 single cells were sequenced. The
libraries were sequenced on the IIlumina Sequencing Platform.
RNA-Seq Data Analysis
For the bulk RNA-Seq data, FastQC (version 0.11.5)(Andrews) was employed to ensure the
quality of all samples. Hisat2 (version 2.0.5)(Kim, Langmead, and Salzberg 2015) was used to
align reads to the indexed genome of R. delemar RA 88-880 (PRJNA13066)(Ma et al. 2009)
and the indexed genome of R. microsporus (Mondo et al. 2017). HTSeq (version 0.8.0)(Anders,
Pyl, and Huber 2015) was used to quantify the output. Trinity and edgeR (version 3.16.5) were
then used to analyse differential expression (Grabherr et al. 2009). For the single cell RNA-
Seq data, the 10X genomics analysis pipeline (Loupe Cell Browser V 2.0.0, Cell Ranger Version
V 2.0.0) was used to align reads to the mus musculus genome (version MM10), and quantify
the output. For single cell analysis, the samples were then aggregated using this pipeline, to
allow comparisons between samples.
57

Other
Rhizopus delemar Protoplast Formation
R. delemar spores were harvested as above and incubated in 10ml of SAB broth at 37°C until
germ tubes roughly equal in length to the diameter of the spores had formed. The germlings
were then spun at 3000 RPM for 3 minutes and the supernatant removed. The germlings were
washed in PBS and resuspended on 10ml of 0.6M KCl, with the addition of 50mg of glucanex.
The germlings were incubated at 30°C, 100RPM and checked periodically for protoplast
formation using brightfield microscopy. The bodies that formed were imaged with brightfield
microscopy. To check for protoplast formation, the resulting bodies were immersed in
deionised water to check for lysis. Protoplasts formed were incapable of germinating.
Zebrafish
Macrophage isolation and RNA Extraction
Zebrafish husbandry and infections were performed by Aleksandra Jasiulewicz. Zebrafish
larvae expressing either mCherry macrophages (mCHerry+), GFP neutrophils (GFP+) or
wildtype (AB) were used in the following work. The Zebrafish Larvae, infected and uninfected,
were suspended in fixation solution (10ml 4% PFA solution + 5ml 3% Sucrose), on ice for 10
minutes. 1ml of the fixation solution was used for every 10 larvae. The fixation solution was
then removed and the larvae were dissociated in 1ml of 0.25% Trypsin (preheated to 37°C)
per 10 larvae, the mixture was incubated at 37°C for 10 minutes, mixing via pipetting
periodically. To terminate dissociation, 10% FBS and 1ul of 1M CaCl2 were added per 1ml of
trypsin to the mixture. The solution was then spun at 800G for 3 minutes to pellet the
dissociated cells and remove the dissociation media. The cells were resuspended in an
58

appropriate amount of PBS for sorting (1ml per 10 larvae) and kept on ice until sorting. Sorting
was carried out using a BD FACSaria sorter, cells were sorted based on fluorescent signal when
compared to the signal obtained from wild-type (AB) zebrafish cells. The sorted cells were
then kept on ice (for no longer than 2 hours) until RNA extraction. Total RNA was extracted
following the Recoverall guidelines, modifying the protease digestion step to 3 hours at 50°
(Hrvatin et al. 2014).
59

Chapter 2: Mucorales Spore Germination
Characterisation
This work has been adapted from the book chapter “Spore Germination of Pathogenic
Filamentous Fungi” (Sephton-Clark and Voelz 2017), for which I performed the literature
search, wrote the manuscript, completed revisions, and prepared the figures. Selected results
have been previously published in and “Pathways of Pathogenicity: Transcriptional Stages of
Germination in the Fatal Fungal Pathogen Rhizopus delemar” (Sephton-Clark et al. 2018) for
which I conceived, designed and performed the experiments, collected the data, performed
the analysis and interpretation, wrote the manuscript, completed revisions, and prepared the
figures.
This chapter will introduce the varying germination phenotypes observed across Mucorales
species. Results obtained through live cell imaging, FACS and extracellular vesicle isolation
demonstrate that species within the Mucorales order exhibit a range of germination rates
and physiologies. Phenotypic heterogeneity to this extent has implications for diagnostic
testing, pathogenicity and phylogenetic assignment.
60

Germination phenotype diversity
Large variation is seen in spore physiology between fungal species; ungerminated spores may
range in diameter from 2.8 to 8.85um (Reponen et al. 2001; Li et al. 2011), and present in a
variety of shapes from spherical to ellipsoidal (Reponen et al. 2001). Some spores require an
initial swelling/isotropic growth phase prior to hyphal germ tube emergence, the timing of
which also varies between fungal species.
Filamentous fungi rely upon germination to transform from spore form to filamentous form.
Germination may occur at a variety of rates, and optimal germination conditions vary
between species. Aspergillus species will germinate fully using a sugar cane bagasse
substrate, by 13 hours. In comparison, Rhizopus microsporus will germinate under the same
conditions in 11 hours, whilst Myceliophthora thermophila fails to germinate completely
(Hassouni et al. 2007). For a single species such as Aspergillus niger, environmental
requirements will also govern germination rate. Within an optimal temperature range (19-
45°C) A. niger will germinate in 12 hours, however at a suboptimal temperature range (50-
60°C), A niger fails to germinate (Hassouni et al. 2007). It is unclear how Mucorales spores
germinate in relation to one another, and whether this impacts pathogenicity.
Over the course of germination, the fungal spore wall changes in a dynamic nature. Spore cell
wall components are produced and organized by several cell wall associated enzymes
including chitin synthase and deacetylase, b(1,3)-glucan synthase, a(1,3)-glucan synthase
and Mannosyltransferase for polysaccharide production, with the corresponding transferase
enzymes responsible for remodelling polysaccharide portions of the cell wall (Karkowska-
61

kuleta and Kozik 2015). These enzymes enable constant reorganisation over the course of
germination. The production of melanin in Aspergillus fumigatus is known to require the
ALB1, ARP1/2, ABR1/2, and AYG1 enzymes which make up the dihydroxynaphthalene melanin
pathway (Tsai et al. 1999; Bernard and Latge 2001; Pihet et al. 2009). These enzymes are also
important for restructuring and reorganizing the cell wall throughout germination. They also
play large roles in the production of filaments, as well as the production of new spores
themselves. The Aspergillus fumigatus genes chsC,G&E, all encoding for chitin synthases, are
required for proper conidiation, as well as hyphae formation, whilst the functionality of chitin
synthases also appears to be required for the acquisition of melanin to the cell wall of C.
neoformans (Mellado et al. 1995, 1996; Aufauvre-Brown et al. 1997; Tsai et al. 1999; Beauvais
et al. 2001; Anand, Yadav, and Yadav 2016; Tsirilakis et al. 2012). Although the genomes of
Mucorales species show the presence of predicted cell wall remodelling enzymes, the roles
which they play throughout germination is unknown.
The structural changes of germinating Aspergillus fumigatus conidia have been tracked to
show both compositional and spatial changes over this time, a characteristic which is also
observed in Mucorales spores (Figure 2,3,4) (Dague et al. 2008). Over the course of
germination, a loss of hydrophobicity can be seen in Aspergillus, which is due to a
rearrangement of the hydrophobic rodlet outer layer as the Aspergillus conidia
swell. After 2 hours of germination, the hydrophobic rods form
clusters, surrounded by hydrophilic polysaccharides, resulting in a heterogeneous
Aspergillus coat (Dague et al. 2008). Work done in the plant pathogen Phanerochaete
chrysosporium shows a similar trend, as the outer rodlet coat is disbanded over the
course of germination (Dufrêne et al. 1999). The conidial cell wall of the pathogenic
mold Scedosporium boydii also shows a decrease in hydrophobicity over the course of
62

germination, along with a decreasing electrostatic charge, an increased accessibility of
polysaccharides and lectins within the cell wall and a remodelling of cell wall proteins.
Conidial GPI anchored proteins are swapped for hyphae specific GPI anchored proteins over
germination (Ghamrawi et al. 2015). Taken together, the highly dynamic restructuring of the
cell wall of spores as they germinate is dependent on many enzymes and components, which
appear to act cooperatively in order to provide structure and environmental protection. How
these enzymes behave in Mucorales species throughout germination is unknown.
This cell wall remodelling may aid the release of extracellular vesicles (EV), which fungal
species such as Magnaporthe oryzae release over the course of germination (Rybak and
Robatzek 2019). Extracellular vesicles have been implicated in fungal pathogenesis
(Panepinto et al. 2009; Yoneda and Doering 2006)(Alspaugh 2015), and have been reported
to carry RNA, lipids, fungal cell wall components, complex sugars and enzymes (Albuquerque
et al. 2008; Vallejo et al. 2011; Da Silva et al. 2015; Eisenman et al. 2009; Muxing Liu et al.
2018b). Recently Liu et al detected the presence of small RNAs in EVs isolated from R.
delemar (Liu et al. 2018a). sRNAs are thought to play roles in modulating plant-
pathogen interactions (Zhao, Liang, and Zhou 2018) and have the potential to modulate host
immunity in response to infection, proving a potent potential virulence factor. Whilst fungal
EVs often appear to carry cargo implicated in pathogenesis, EVs from Candida albicans have
been shown to offer ‘protection’ in a Galleria mellonella infection model (Vargas et al.
2015). Despite their role, it remains unclear where EVs originate, and exactly how they
traverse the fungal cell wall. Whilst Rodrigues et al propose the cytosol as a potential EV
reservoir, Wolf et al. provide evidence for EV formation via membrane budding
(Rodrigues et al. 2013; Wolf et al. 2014). Recent work has shown that the cell wall
of C. albicans, and C. neoformans, is permeable to vesicles 60-80nm in size. The
viscoelastic properties of the cell wall may therefore allow for EV traversal (Walker et al.
2018). 63

The mechanism by which EVs traverse the cell wall remains unclear, however several
studies have revealed the presence of cell wall degrading enzymes within fungal EVs,
offering a potential mechanism for this understudied process. The release of extracellular
vesicles over the course of germination has been poorly studied to date, and remains
unknown in Rhizopus sp.
The definition of germination varies widely between studies and fungal species, an
approach which standardises the definition and measurement of germination would benefit
the field. For the work presented here, germination is said to have been reached upon the
formation of a hypha, the length of which is greater than or equal to the diameter of the
spore body from which it emerges. The following results address gaps in the literature
regarding comparative germination phenotypes, germination dynamics of the
Mucorales cell wall and extracellular vesicle release.
64
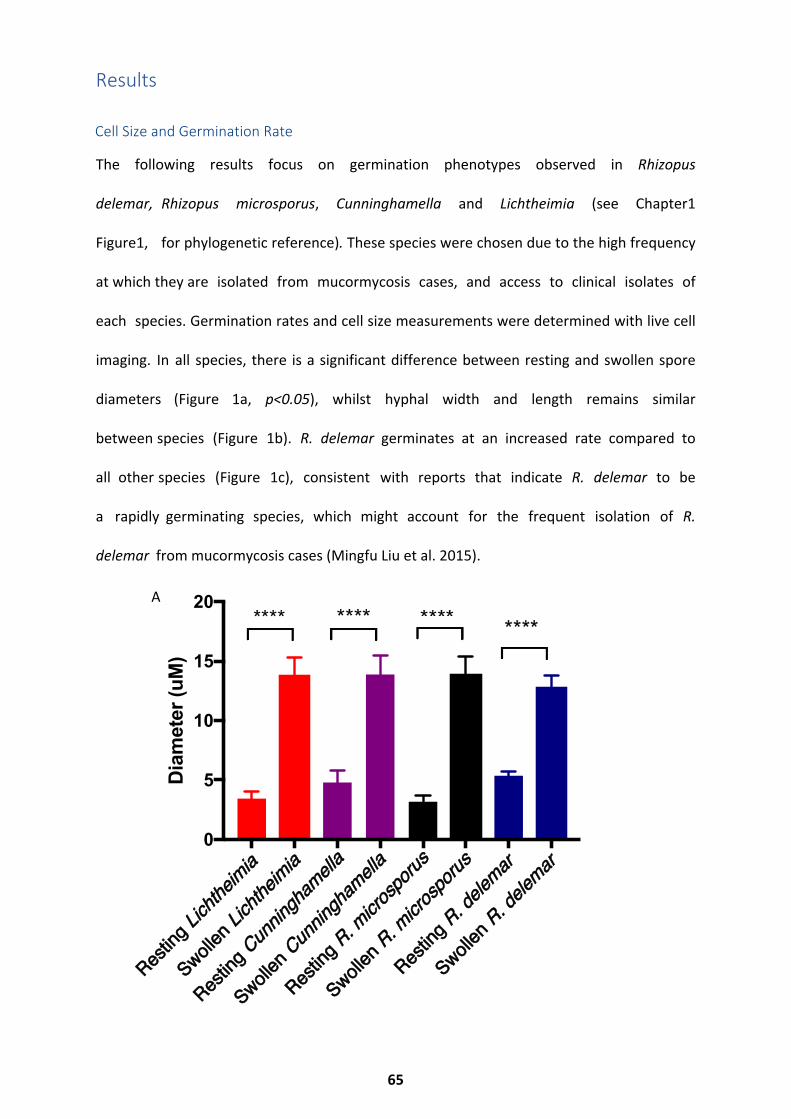
Results
The following results focus on germination phenotypes observed in Rhizopus
delemar, Rhizopus microsporus, Cunninghamella and Lichtheimia (see Chapter1
Figure1, for phylogenetic reference). These species were chosen due to the high frequency
at which they are isolated from mucormycosis cases, and access to clinical isolates of
each species. Germination rates and cell size measurements were determined with live cell
imaging. In all species, there is a significant difference between resting and swollen spore
diameters (Figure 1a, p<0.05), whilst hyphal width and length remains similar
between species (Figure 1b). R. delemar germinates at an increased rate compared to
all other species (Figure 1c), consistent with reports that indicate R. delemar to be
a rapidly germinating species, which might account for the frequent isolation of R.
delemar from mucormycosis cases (Mingfu Liu et al. 2015).
0
5
10
15
20
Dia
met
er (u
M)
A
Restin
g Lich
theim
ia
Restin
g Cun
ningh
amella
Restin
g R. m
icros
porus
Restin
g R. d
elemar
Swollen C
unnin
gham
ella
Swollen R
. micr
ospo
rus
Swollen R
. dele
mar
Swollen L
ichthe
imia
Cell Size and Germination Rate
65
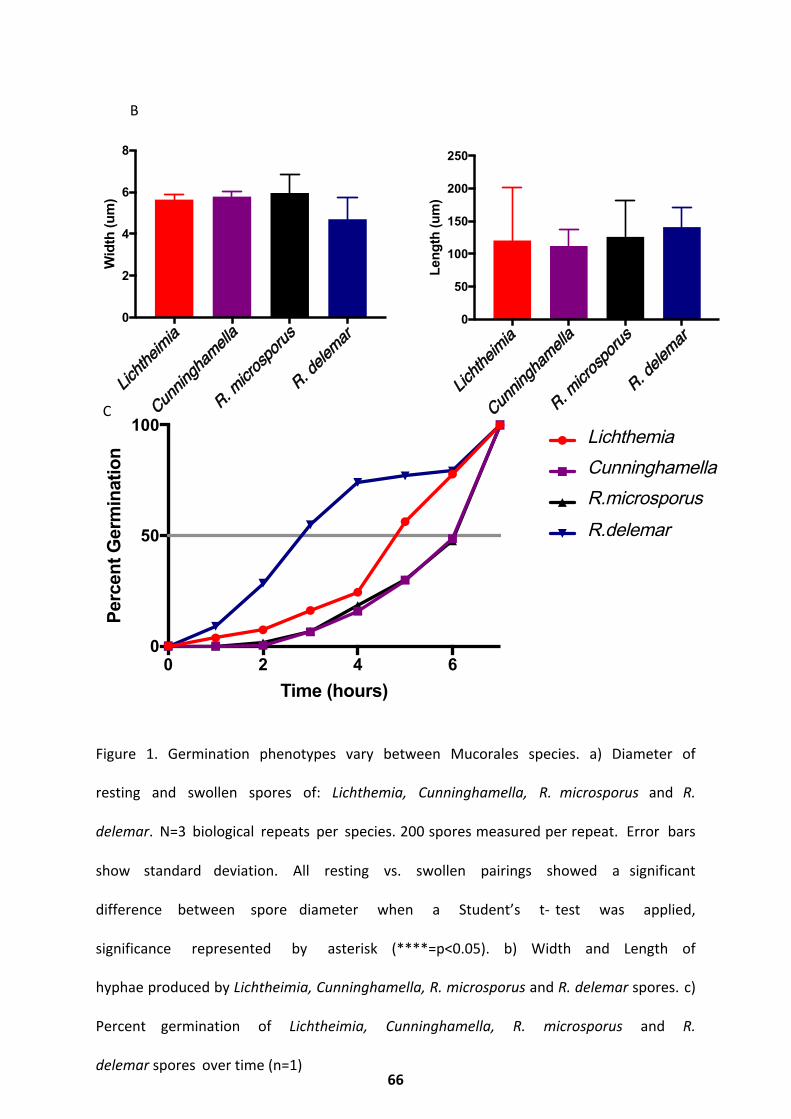
Figure 1. Germination phenotypes vary between Mucorales species. a) Diameter of
resting and swollen spores of: Lichthemia, Cunninghamella, R. microsporus and R.
delemar. N=3 biological repeats per species. 200 spores measured per repeat. Error bars
show standard deviation. All resting vs. swollen pairings showed a significant
difference between spore diameter when a Student’s t- test was applied,
significance represented by asterisk (****=p<0.05). b) Width and Length of
hyphae produced by Lichtheimia, Cunninghamella, R. microsporus and R. delemar spores. c)
Percent germination of Lichtheimia, Cunninghamella, R. microsporus and R.
delemar spores over time (n=1)
0 2 4 60
50
100
Time (hours)
Per
cent
Ger
min
atio
n
Lichthemia
Cunninghamella
R.microsporus
R.delemar
0
2
4
6
8
Wid
th (u
m)
0
50
100
150
200
250
Leng
th (u
m)
B
C Lichth
eimia
Cunnin
gham
ella
R. micr
ospo
rus
R. dele
mar
Lichth
eimia
Cunnin
gham
ella
R. micr
ospo
rus
R. dele
mar
66

The cell wall alters over the course of germination in multiple fungal species (Dague
et al. 2008; Dufrene et al. 1999). Multiple studies have focused on rodlet dynamics in
Aspergillus species as a measure for cell wall restructuring, however the genome
of R. delemar lacks genes with predicted functions in rodlet formation, and SEM
images of Rhizopus arrhizus confirm a lack of rodlets (Wurster et al. 2017). Like other
Mucorales species, the genome of R. delemar appears enriched for chitin synthases (Ma et
al. 2009) and deacetylases (Chibucos et al. 2016), twice as many seen for many dikaryotic
fungi (Ma et al. 2009). For this reason, cell wall chitin/chitinosan content was chosen as
an initial measure of cell wall plasticity over germination. Flow cytometry revealed
dynamic processes which take place within the cell wall over the course of swelling
(Figure 2). Over the 6-hour course of isotropic swelling and germ tube emergence,
CFW staining becomes more intense, indicating that more chitin/chitinosan is available to
the stain. By 6 hours all species show an intense stain for CFW, and both Mucor cbs and
Mucor nrrl (Figure 2. C,D) show slightly increased staining at 0 hours, indicating
that Mucor species display more chitin/chitinosan on the resting spore surface.
Morphology and Cell Wall Dynamics
67
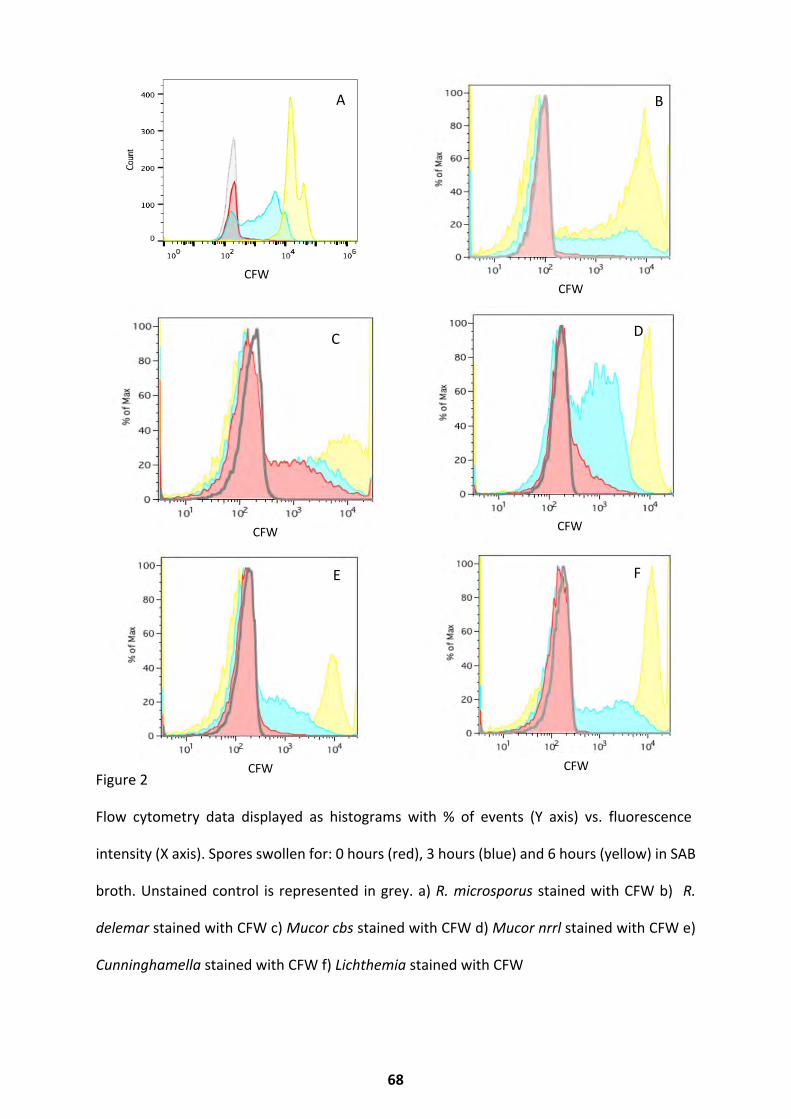
Figure 2
Flow cytometry data displayed as histograms with % of events (Y axis) vs. fluorescence
intensity (X axis). Spores swollen for: 0 hours (red), 3 hours (blue) and 6 hours (yellow) in SAB
broth. Unstained control is represented in grey. a) R. microsporus stained with CFW b) R.
delemar stained with CFW c) Mucor cbs stained with CFW d) Mucor nrrl stained with CFW e)
Cunninghamella stained with CFW f) Lichthemia stained with CFW
A
CFW
B
CFW
C
CFW
D
CFW
E
CFW
F
CFW
68

To better characterise the distinct phases of germination, R. delemar and R. microsporus were
chosen for higher resolution phenotyping. Germination is characterized by three distinct
transitions: dormancy to swelling, swelling to germ tube emergence, and the switch to
sustained filamentous growth. For both species the switch from dormancy to swelling was
triggered by exposure to rich media. Swelling, characterized by an isotropic increase in size,
continued for 4 to 6 h (Figure 3). Once fully swollen, germ tubes emerged from the spore
bodies. Most R.delemar spore bodies (75.5%, Fig. 1C) produced hyphae that exceeded the
diameter of the spore body in length by 4-5 hours, whilst R. microsporus requires 6-7 hours
to reach this point. At this time point, the spores were considered fully germinated. Hyphal
growth was monitored up to the 24 hours post germination initiation for R. delemar to
inform subsequent RNA-Seq studies.
69
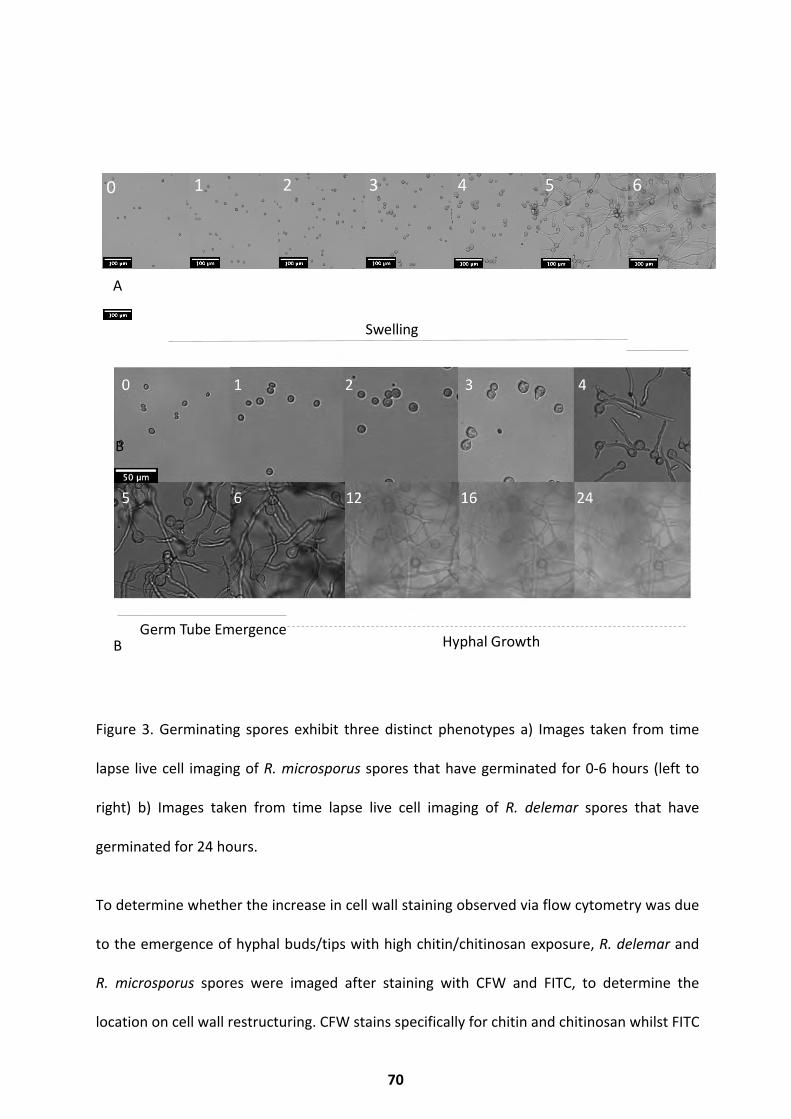
Figure 3. Germinating spores exhibit three distinct phenotypes a) Images taken from time
lapse live cell imaging of R. microsporus spores that have germinated for 0-6 hours (left to
right) b) Images taken from time lapse live cell imaging of R. delemar spores that have
germinated for 24 hours.
To determine whether the increase in cell wall staining observed via flow cytometry was due
to the emergence of hyphal buds/tips with high chitin/chitinosan exposure, R. delemar and
R. microsporus spores were imaged after staining with CFW and FITC, to determine the
location on cell wall restructuring. CFW stains specifically for chitin and chitinosan whilst FITC
Swelling
Germ Tube Emergence Hyphal Growth
A
B
B
0 1 2 3 4 5 6
70
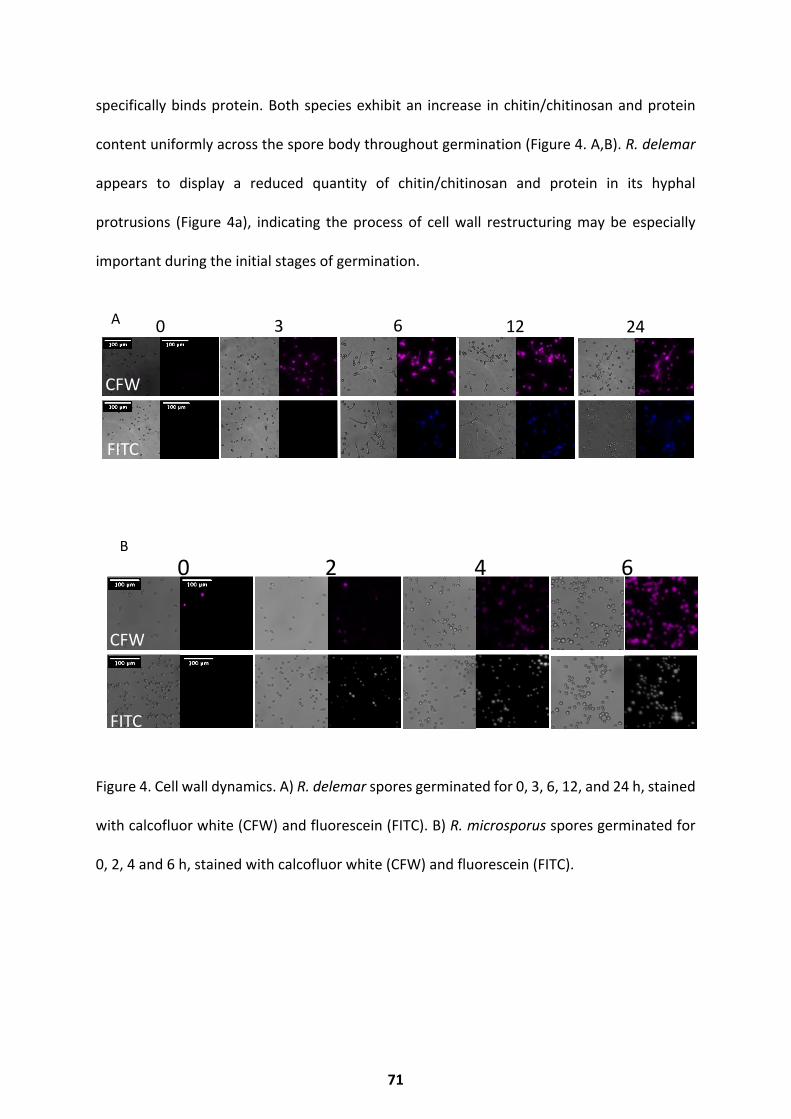
specifically binds protein. Both species exhibit an increase in chitin/chitinosan and protein
content uniformly across the spore body throughout germination (Figure 4. A,B). R. delemar
appears to display a reduced quantity of chitin/chitinosan and protein in its hyphal
protrusions (Figure 4a), indicating the process of cell wall restructuring may be especially
important during the initial stages of germination.
Figure 4. Cell wall dynamics. A) R. delemar spores germinated for 0, 3, 6, 12, and 24 h, stained
with calcofluor white (CFW) and fluorescein (FITC). B) R. microsporus spores germinated for
0, 2, 4 and 6 h, stained with calcofluor white (CFW) and fluorescein (FITC).
0 3 6 12 24
CFW
FITC
ROS
0 2 4 6
CFW
FITC
A
B
71

To determine whether Mucorales species also release extracellular vesicles (EVs) over
germination, as has been reported in other species (Rodrigues et al. 2008), an EV isolation
protocol was applied to the supernatant of germinating R. delemar spores. EVs were
successfully isolated and the size of these particles determined. The concentration of
EVs isolated increases post germination initiation (Table 1) and after 4 hours, the size of
the particles released increases significantly, with the average size for 0-2 hours being
2.74nm and the average size for 4-6 hours being 269nm (Figure 5), however the charge
remains neutral (Table 2). pH has been shown to influence charges across EV membranes,
therefore influencing EV ability to fuse with external cells via micropinocytosis (Yáñez-Mó et
al. 2015). As germinating R. delemar spores are able to germinate in a range of pHs, and may
influence the pH of their microenvironment as they do so, effective EV cargo delivery may
be altered depending on germination stage or environment.
Extracellular Vesicle Release
72
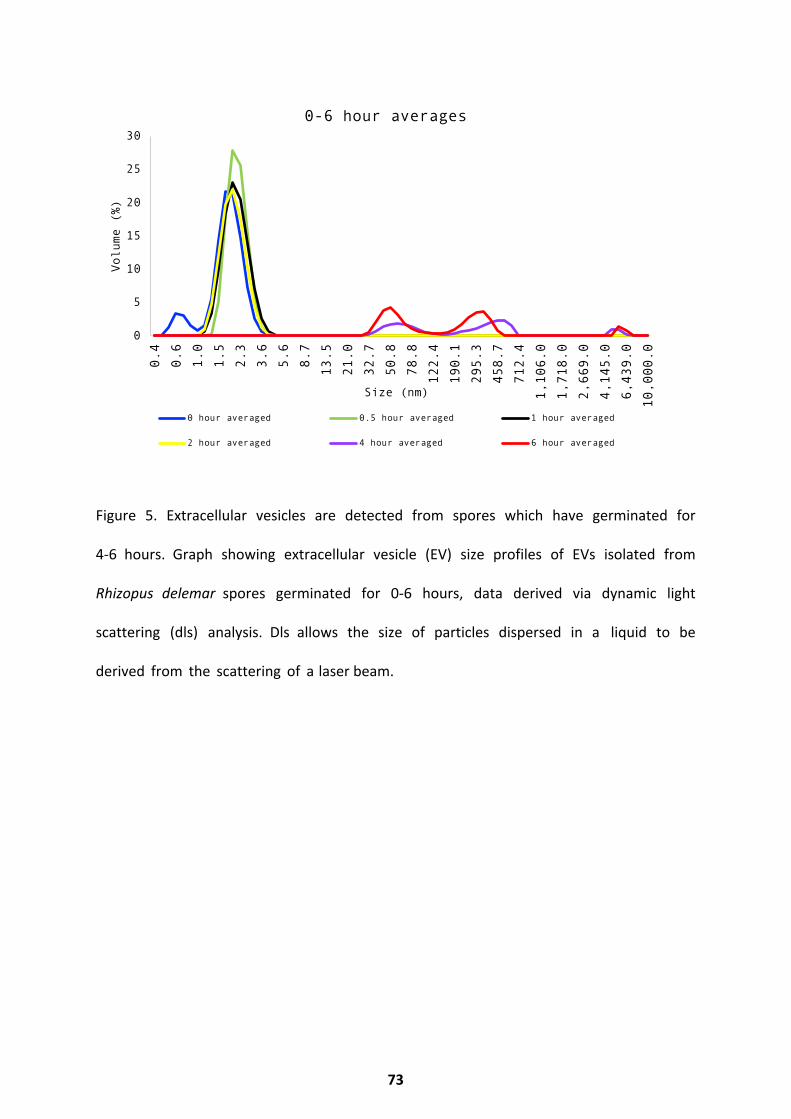
Figure 5. Extracellular vesicles are detected from spores which have germinated for
4-6 hours. Graph showing extracellular vesicle (EV) size profiles of EVs isolated from
Rhizopus delemar spores germinated for 0-6 hours, data derived via dynamic light
scattering (dls) analysis. Dls allows the size of particles dispersed in a liquid to be
derived from the scattering of a laser beam.
0
5
10
15
20
25
30
0.4
0.6
1.0
1.5
2.3
3.6
5.6
8.7
13.5
21.0
32.7
50.8
78.8
122.4
190.1
295.3
458.7
712.4
1,106.0
1,718.0
2,669.0
4,145.0
6,439.0
10,000.0
Volume (%)
Size (nm)
0-6 hour averages
0 hour averaged 0.5 hour averaged 1 hour averaged
2 hour averaged 4 hour averaged 6 hour averaged
73
Condition Average Concentration (particles/spore) 0 hour 3.78 1 hour 10.40 2 hour 7.87 4 hour 8.96 6 hour 16.99
Table 1: showing the concentrations of EVs from spores germinated in SAB, recorded by nano sight.
Condition Average charge (mV) 0 hour -19.750.5 hour -12.831 hour -11.432 hour -11.484 hour -11.496 hour -12.06SAB control (no spores present) -14.82
Table 2: showing the charge of the isolated EVs from spores germinated in SAB, recorded by
zetasizer. By applying an electric field to particles analyzed by dynamic light scattering, the zeta
potential (potential difference across phase boundaries, measured in mV) was determined.
Conclusions
The species studied here display similar spore and hyphae sizes throughout
germination. Conversely, germination timing is varied between Mucorales species
(Figure 1c). Despite timing differences, the germination phenotype of swelling, germ
tube emergence and finally hyphal growth appears conserved across the species
studied. Trends in cell wall component (chitin/chitinosan and protein) exposure
varies between species. Closely-related species exhibit similar exposure patterns to one
another over germination (Figure 2: Mucor cbs and Mucor nrrl), however these patterns
are distinct from other species. Finally, germinating Rhizopus delemar spores were found
to release extracellular vesicles after 4-6 hours (Figure 5). Small particle peaks (1-4nm)
were also detected from spores that germinated for 0.5-2 hours, however given the
size, these peaks likely represent artifacts or cell wall components. These results and the
potential for further work will be discussed here.
74

Discussion
The phenotypic variation over germination highlights the broad phylogeny of the Mucorales
order, which has been better described taxonomically through DNA based reclassification
(Hoffmann et al. 2013; Gryganskyi et al. 2010; Abe et al. 2007). The species studied here
display similar resting spore, swollen spore and hyphal sizes (Figure 1a,b), thus making
diagnosis difficult through single time point imaging alone (Yang et al. 2016). However the
differential germination dynamics of these species (Figure 1c, 2) might provide a cost
effective alternative for diagnosis through multiple time point imaging and cell staining. All
species appear to undergo cell wall remodelling during germination (Figure 2, 4), however
the extent to which each species remodels is variable. Cell wall remodelling and unmasking
in response to pH has been linked to pathogenicity in Candida albicans (Sherrington et
al. 2017), and it may be possible that the variation in remodelling of these species is
linked to variation in pathogenicity. The known Mucorales pathogenicity factor CotH
resides within the cell wall of pathogenic Mucorales species, and unmasking of
this factor over germination may result in increased pathogenicity. This would support the
observation that swollen Mucorales spores are more pathogenic than ungerminated
ones (Chibucos et al. 2016). Mucorales cell wall remodelling may occur in response to pH
changes, as we see for the pH dependent recruitment of aquaporins over germination
(Turgeman et al. 2016). The EV data shown here adds to the sparse literature on fungal EV
release over germination. It is likely that these EVs contain sRNAs (Liu et al. 2018b),
however they may include other components such as enzymes or pathogenicity
factors which might aid environment dependent colonisation. Both seedlings and
neutrophils have been shown to release EVs capable of interacting with their respective
fungal pathogens (Sclerotinia sclerotiorum and Aspergillus fumigatus) to inhibit growth
(Regente et al. 2017; Shopova et al. 2019).
75

It is possible that Rhizopus delemar might release EVs in a similar manner to ensure spore
survival and propagation, despite environmental responses to the spores.
Further Work
This field would benefit from further work focusing on the impact of environment
on germination. Most fungal germination studies use rich media, however this does not
reflect the ‘real world’ environments which many fungi have evolved to inhabit.
Studies which address germination phenotypes in a range of ‘realistic’ media (soil,
blood, guano etc.) would greatly increase our understanding of how these pathogens
interact with their surroundings, and advance our understanding of their pathogenesis.
This work would also benefit from an in-depth analysis of the precise alterations which
occur in the Mucorales cell wall over germination, and any molecules they may secrete (eg
EVs). High resolution mass spectrometry and scanning electron microscopy would allow
these areas to be explored further.
76

Chapter 3: Hallmarks of the Mucorales genome
Results have been previously published or adapted from the following articles: “Pathways of
Pathogenicity: Transcriptional Stages of Germination in the Fatal Fungal Pathogen Rhizopus
delemar”(Sephton-Clark et al. 2018) and “Host-pathogen transcriptomics of macrophages,
Mucorales and their endosymbionts: a polymicrobial pas de trois” (Sephton-Clark et al.
2019). For both I conceived, designed and performed the experiments, collected the data,
performed the analysis and interpretation, wrote the manuscript, completed revisions, and
prepared the figures.
This chapter will introduce characteristics of the Mucoralean genome. Results describing
genome assembly, annotation and comparisons of two Mucorales species, Rhizopus delemar
and Rhizopus microsporus, show the conservation of a core set of fungal features, but
divergence between the two genomes in enrichment of substrate binding, transporter
activity, endosome activity and DNA repair.
77
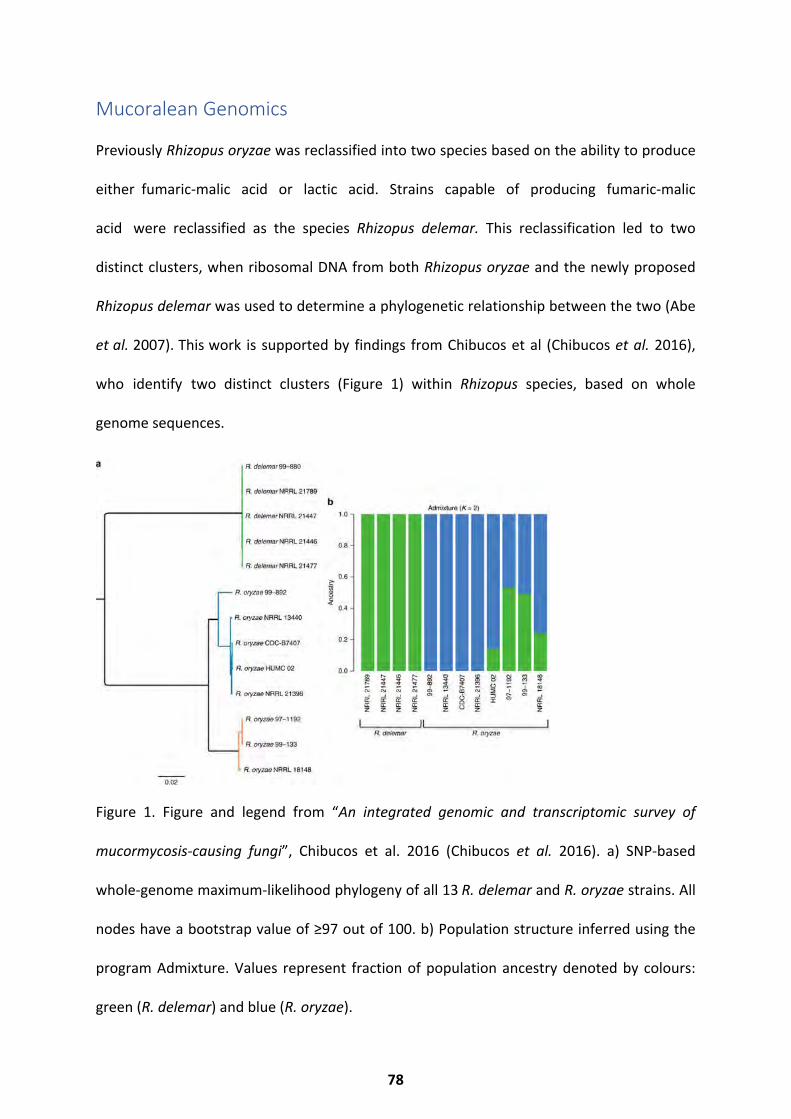
Mucoralean Genomics
Previously Rhizopus oryzae was reclassified into two species based on the ability to produce
either fumaric-malic acid or lactic acid. Strains capable of producing fumaric-malic
acid were reclassified as the species Rhizopus delemar. This reclassification led to two
distinct clusters, when ribosomal DNA from both Rhizopus oryzae and the newly proposed
Rhizopus delemar was used to determine a phylogenetic relationship between the two (Abe
et al. 2007). This work is supported by findings from Chibucos et al (Chibucos et al. 2016),
who identify two distinct clusters (Figure 1) within Rhizopus species, based on whole
genome sequences.
Figure 1. Figure and legend from “An integrated genomic and transcriptomic survey of
mucormycosis-causing fungi”, Chibucos et al. 2016 (Chibucos et al. 2016). a) SNP-based
whole-genome maximum-likelihood phylogeny of all 13 R. delemar and R. oryzae strains. All
nodes have a bootstrap value of ≥97 out of 100. b) Population structure inferred using the
program Admixture. Values represent fraction of population ancestry denoted by colours:
green (R. delemar) and blue (R. oryzae).
78

Prior to reclassification, the genome of Rhizopus oryzae 99-880 (now Rhizopus delemar 99-
880) was sequenced and assembled from whole genome shotgun library sequencing data by
Ma et al. (Ma et al. 2009). Although transcriptomic data was not available for assembly and
annotation, the resulting assembly identified the 45.26Mb genome to be repetitive (20%
transposable elements), and consisting of 13,895 protein coding genes. A whole genome
duplication event, followed by subsequent gene duplication events, indicated by the
segments of duplicate gene pairs scattered throughout the genome, resulted in multiple gene
family expansions. These gene families have roles in ergosterol biosynthesis, cell growth,
signal transduction, protease secretion and subtilase secretion (Ma et al. 2009). Notably,
analysis revealed a lack of the common siderophore producing non-ribosomal peptide
synthases, highlighting R. delemar’s reliance on Rhizoferrin for iron scavenging. This is
especially relevant in the context of this species’ requirement for iron in order to germinate,
and its implications in pathogenesis (Ibrahim et al. 2007; Andrianaki et al. 2018; Kousser et
al. 2019).
The chitin deactelyase family is also expanded in R. delemar, compared to non-early divergent
fungi, as it is for other mucoralean species. There appear to be 2,658 core clusters present in
the 19 Mucorales species included in the analysis carried out by Chibucos et al. We see that
there is a range of genome sizes in this study, with the smallest genome consisting of 22.3Mb,
whilst the largest is 96.7Mb. Highly pathogenic Mucorales species from this study were found
to contain a unique set of 174 genes, conserved in Rhizopus species but lacking in non-
Rhizopus species and thus labelled as putative virulence genes. Amongst the virulence genes
conserved across the Mucoralean family, CotH, which codes for a surface protein uniquely
expressed by Mucoralean species, is present in varying copy numbers between Mucorales
79

79
species. The genomes of Rhizopus delemar, Rhizopus oryzae and Rhizopus microsporus and
Lichtheimia corymbifera all contain over 6 CotH genes, whilst other Mucorales species contain
fewer than 6. The number of CotH genes present has been linked directly to pathogenicity,
however species containing fewer than 6 copies, such as Mucor circinelloides, may still cause
mucormycosis (Chibucos et al. 2016). It is clear that CotH plays an important role in
virulence, however it does not account for all interactions between Mucorales species and
host.
Aside from CotH, few key pathogenicity genes/characteristics have been identified within all
Mucorales species. Around 54% of clinical Mucorales isolates are were found to harbour
bacterial endosymbionts from the Burkholderia family in a study carried out by Ibrahim et
al. (Ibrahim et al. 2008), and though previous work demonstrated that the presence of
these endosymbionts did not impact infection of endothelial cells (Ibrahim et al. 2008),
recent work by Itabangi et al (Itabangi et al. 2019) has shown that the bacterial
endosymbiont within Rhizopus microsporus impacts the fungus’ interaction and
regulation of the host immune system. A bacterial endosymbiont also impacts sexual
reproduction in R. microsporus (Mondo et al. 2017), demonstrating the unique relationship
crucial to development and pathogenicity between Rhizopus microsporus and its bacterial
endosymbiont.
Although several genome sequences exist for Rhizopus microsporus (Horn et al. 2015;
Mondo et al. 2017) a comprehensive analysis of the genome is not available. It is
unknown whether Rhizopus microsporus underwent a whole genome duplication, similarly
to Rhizopus delemar. The following results will provide a thorough analysis of both the
Rhizopus microsporus and Rhizopus delemar genome, and include results based on
comparative genomics analysis carried out to compare the two species.
80
Genome Statistics
Size (Mb) GC% Protein rRNA tRNA Gene Rhizopus delemar
45.32 35.6 17,459 5 239 17,703
Rhizopus microsporus
25.53 37.7 10,891 Not predicted
70 10,959
Results
We utilized our RNA-Seq data to revise the current annotation of the available R.
delemar genome (revised GFF file available at: https://github.com/psephtonclark/
RhizDeleAnno), using BRAKER 2.1.0 (Hoff et al. 2016) to improve gene structures and
incorporate these into an updated annotation. Compared to the previous annotation (Ma et
al. 2009), this updated set included 475 new predicted genes, 370 new protein family
domains (Pfam terms), 103 new pathway predictions (KEGG-EC), and 96 new
transmembrane domains (TMHMM terms) (Figure 2). The updated annotation was assessed
for completeness with BUSCO v3 (Simão et al. 2015) and was shown to include a good
representation of expected core eukaryotic genes, with minimal missing BUSCOs
(benchmarked universal single-copy orthologues) (2%)
Resulting Characteristics of the Reannotated R. delemar Genome
81

Figure 2. Table displaying genome annotation statistics, comparing the original R. delemar
annotation (column 1, ‘Vesper Annotation’) and the annotation updated with our RNA-Seq
data (column 2, ‘‘Vesper Annotation with RNA Seq data incorporated’’).
Biochemical Pathways Present in R. delemar
A custom genome database for R. delemar was created with PathoLogic to allow for
PathwayTools analysis of the reannotated R. delemar genome (Database available at
https://github.com/psephtonclark/rhior3cyc). From this, metabolic, signalling and
transporter pathways can be visualised with the PathwayTools GUI (Figure 3). This annotation
classified genes with predicted or known functions into the following categories: Enzymatic
reactions (2061), Transport reactions (13), Activation/inactivation pathways (4), Biosynthesis
pathways (166), Degradation pathway (93), Detoxification pathway (8), Precursor Metabolism
(20), Macromolecule Modification pathway (4), Metabolic cluster (6), Superpathways (31).
This annotation also identified 4377 enzymes, 93 transporters and 1517 compounds. These
-
82

pathways and molecules have been visualized in Figure 3. All pathway predictions are based
on genomic information. Annotations of model fungal species inform these predictions,
therefore the predictions will be more accurate, the more closely related the species are.
Key metabolic pathways identified include:
• Carbohydrate Biosynthesis and degradation(sugar, polysaccharide, oligosaccharide)
• Amino Acid Biosynthesis and degradation (polyamine, proteinogenic, modification, b alanine
biosynthesis, L-citrulline biosynthesis, L-Ornithine)
• Fermentation and alcohol degradation
• Respiration
• Glycolysis
• TCA Cycle
• Lipoate biosynthesis
• NAD/NADP metabolism
• Polyprenol biosynthesis
• Porphyrin compound biosynthesis
• Tetrahydrobiopterin biosynthesis
• Tetrapyrrole biosynthesis
• Vitamin biosynthesis
• Secondary Metabolite biosynthesis and degradation (Phenylpropanoid, Sugar derivative, Terpenoid)
• Lipid biosynthesis and degradation (choline, FA, phospholipid, sphingolipid, sterol)
• Aminoacyl-tRNA charging
• Nucleoside/tide biosynthesis and degradation
• Aldehyde degradation
83

Figure 3. Pathway visualization based on the R. delemar genome. The PathoLogic database required to generate this can be found in the
online appendix (https://github.com/psephtonclark/rhior3cyc). This annotation was developed via the following
method: predicted gene functions were incorporated into the PathoLogic annotation. Pathways were predicted as described by Karp et al
(Karp et al. 2002), and can be visualized with the PathwayTools GUI. For each prediction there is information available detailing
compound structure, enzymatic reactions and other metabolic functions. Metabolomic and transcriptomic data can be overlaid onto the
annotation, thus pathways up/down regulated (condition dependent) will be highlighted in various colors (this example shows pink, green,
orange and red representing pathways down regulated in hyphal, swelling, germination initiation - 1hr, and resting spores - 0hr, respectively).
= amino acids, = carbohydrates, ◇ = proteins, 0 = purines, = pyrimidines, ▽ = cofactors, T = tRNA's, ⃝ = other
Detoxification
AldehydeDegradation
CarboxylatesDegradation
Nucleosides and Nucleotides Degradation
TCA cycle
Amines and PolyaminesDegradation
Secondary MetabolitesDegradation
Fermentation
Cofactors, Prosthetic Groups, Electron Carriers Biosynthesis
Carbohydrates Degradation
Amino Acids DegradationAminoacyl-tRNA Charging
Nucleosides and Nucleotides Biosynthesis
Metabolic RegulatorsBiosynthesis
Amino Acids Biosynthesis
Other
Degradation/Utilization/Assimilation - Other
Alcohols Degradation
Respiration
Carbohydrates Biosynthesis
Fatty Acid and Lipid Biosynthesis
Secondary Metabolites Biosynthesis
Fatty Acid and Lipids Degradation
Polymeric CompoundsDegradation
Glycolysis
Inorganic Nutrients Metabolism
Amino AcidsCarbohydratesProteinsPurinesPyrimidinesCofactorstRNAsOther
(Filled) Phosphorylated
Rhior_1224DownRhior_26DownRhior_1DownRhior_0Down
Rhizopus delemar RA 99-880 Cellular Overview
This diagram was automatically generated by SRI International Pathway Tools version 21.0, authors S. Paley and P.D. Karp.
84
□△ 0
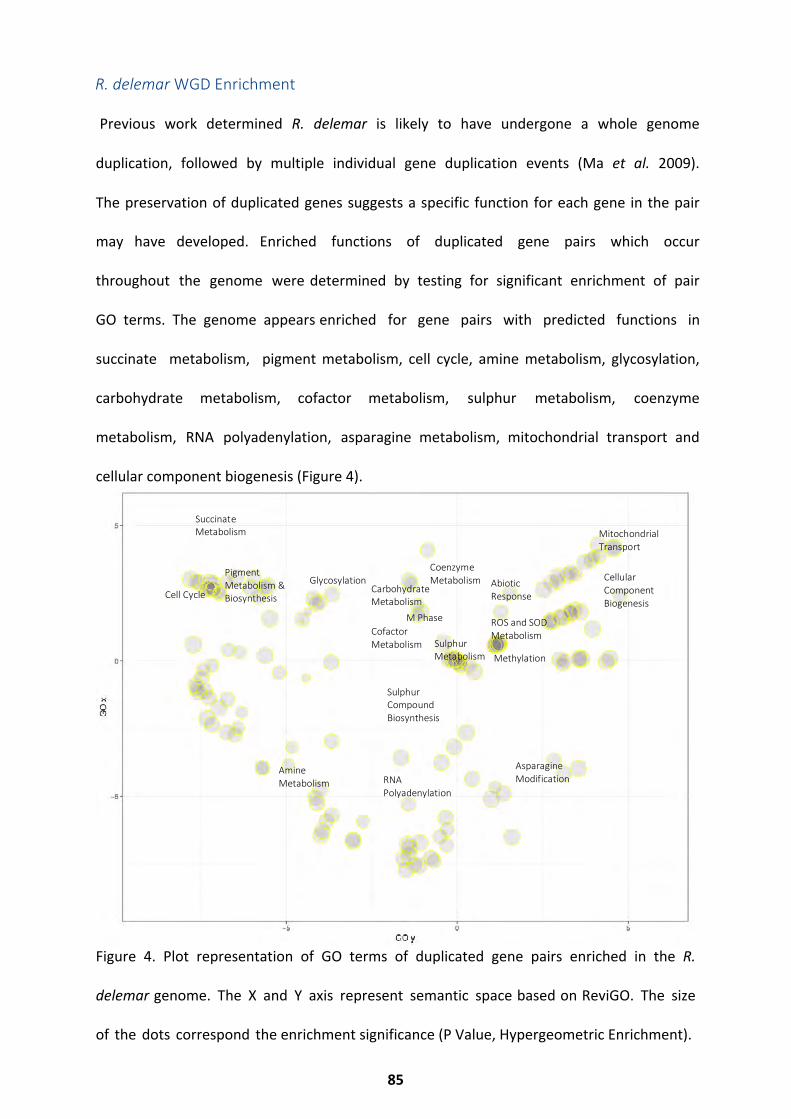
Previous work determined R. delemar is likely to have undergone a whole genome
duplication, followed by multiple individual gene duplication events (Ma et al. 2009).
The preservation of duplicated genes suggests a specific function for each gene in the pair
may have developed. Enriched functions of duplicated gene pairs which occur
throughout the genome were determined by testing for significant enrichment of pair
GO terms. The genome appears enriched for gene pairs with predicted functions in
succinate metabolism, pigment metabolism, cell cycle, amine metabolism, glycosylation,
carbohydrate metabolism, cofactor metabolism, sulphur metabolism, coenzyme
metabolism, RNA polyadenylation, asparagine metabolism, mitochondrial transport and
cellular component biogenesis (Figure 4).
Figure 4. Plot representation of GO terms of duplicated gene pairs enriched in the R.
delemar genome. The X and Y axis represent semantic space based on ReviGO. The size
of the dots correspond the enrichment significance (P Value, Hypergeometric Enrichment).
Glycosylation Pigment Metabolism & Biosynthesis
Succinate Metabolism
Cell Cycle Carbohydrate Metabolism
Mitochondrial Transport
Cellular Component Biogenesis
Abiotic Response
Coenzyme Metabolism
ROS and SOD Metabolism
Methylation Sulphur Metabolism
Cofactor Metabolism
M Phase
Sulphur Compound Biosynthesis
Amine Metabolism RNA
Polyadenylation
Asparagine Modification
R. delemar WGD Enrichment
85

R. microsporus Genome Assembly and Statistics
A genome assembly of R. microsporus was produced with SPAdes, using short read data as
input. For this, Mondo et al’s assembly was taken as a reference (Mondo et al., 2017),
producing an assembly with 4778 contigs. With a GC content of 37.7% (Figure 5), our assembly
appeared identical to that produced by Mondo et al. The low resolution of the assembly
produced (4778 contigs) is likely due to the lack of long read information and likelihood of a
highly repetitive genome, which increases the difficulty of contig joining when relying on short
read data alone. As Mondo et al’s assembly achieved a higher resolution (131 scaffolds) by
making use of long read data, I used this assembly, but replaced nucleotides identified as
SNPs (Figure 6) in our R. microsporus strain, to produce a reference genome specific to the
R. microsporus strain used throughout the project (University of Birmingham R. microsporus
strain). Briefly, I did this by aligning the raw reads obtained through short read sequencing to
the assembly produced by Mondo et al. I then called variants present when comparing the
sequence of the assembly to the aligned reads. I then inserted the variant nucleotides from
our strain into the assembly, in place of the previous nucleotides. In total, 6132 SNPs were
identified and replaced (Figure 6).
86
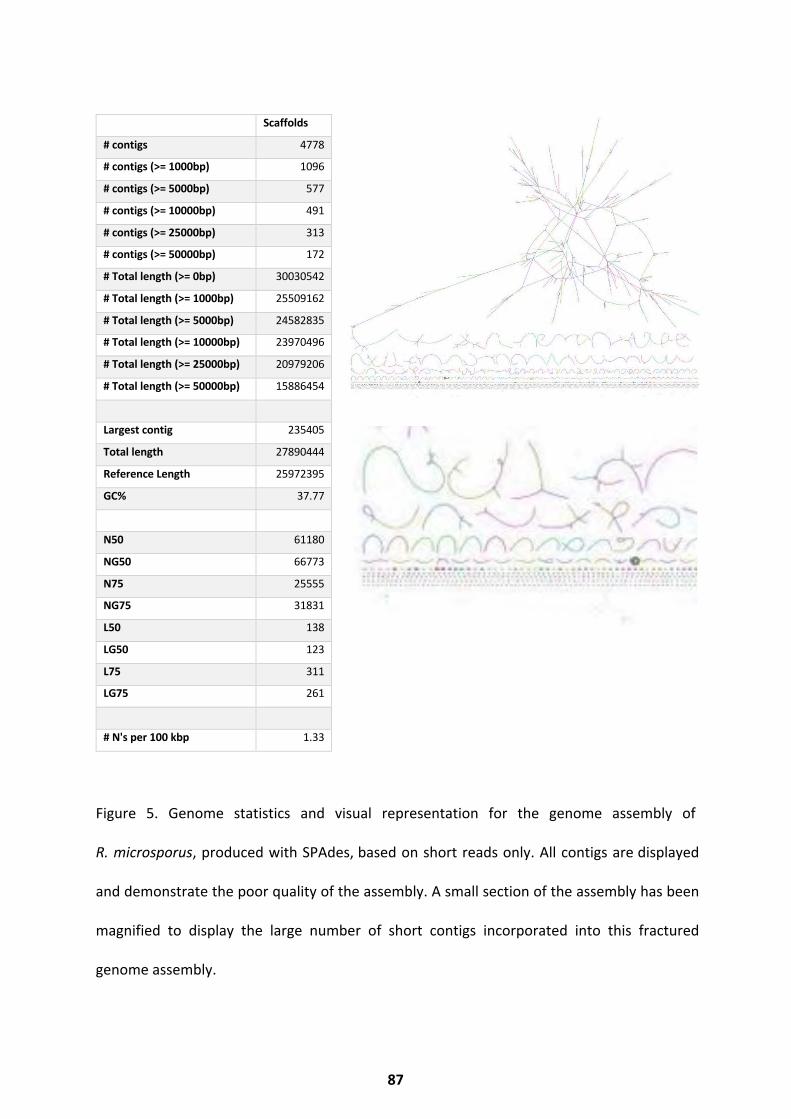
Scaffolds
# contigs 4778
# contigs (>= 1000bp) 1096
# contigs (>= 5000bp) 577
# contigs (>= 10000bp) 491
# contigs (>= 25000bp) 313
# contigs (>= 50000bp) 172
# Total length (>= 0bp) 30030542
# Total length (>= 1000bp) 25509162
# Total length (>= 5000bp) 24582835
# Total length (>= 10000bp) 23970496
# Total length (>= 25000bp) 20979206
# Total length (>= 50000bp) 15886454
Largest contig 235405
Total length 27890444
Reference Length 25972395
GC% 37.77
N50 61180
NG50 66773
N75 25555
NG75 31831
L50 138
LG50 123
L75 311
LG75 261
# N's per 100 kbp 1.33
Figure 5. Genome statistics and visual representation for the genome assembly of
R. microsporus, produced with SPAdes, based on short reads only. All contigs are displayed
and demonstrate the poor quality of the assembly. A small section of the assembly has been
magnified to display the large number of short contigs incorporated into this fractured
genome assembly.
87

Figure 6. Single nucleotide polymorphisms and indels identified when comparing Mondo et al’s assembly to
aligned reads representing the UoBirmingham R. microsporus strain. The IGV graph (top) demonstrates that
Variants (blue columns corresponding to genome location of variants) were identified across the entire genome.
To generate this, the UoB R. microsporus VCF file (variant call data) was aligned against the Mondo et al R.
microsporus genome. Variants highlighted in blue.
Comparative Genomics
R. delemar genes with homology to genes in R. microsporus were identified via blast+. A
comprehensive list of these genes can be found in the online appendix
(https://github.com/psephtonclark/RhizoOrthologues). 8,356 genes were identified with a
bit score > 90 (E Value < 1 x 10-20) and a percentage identity match of over 60% (Pearson,
2013). 2,121 highly conserved genes were found (identity % > 80%), and those with an identity
percentage of over 90% (234 genes) had roles in ribosome structure, translation, protein
metabolism, phosphorus metabolism, organelle structure, transcription, respiration, ATP
metabolism, sugar metabolism, catalytic activity, ion transport and binding.
Fisher's exact test identified enriched Pfam terms when the genome contents of R.
delemar and R. microsporus are compared. Compared to R. microsporus, the genome of R.
delemar is
88
Scaffold 1 Scaffold 131
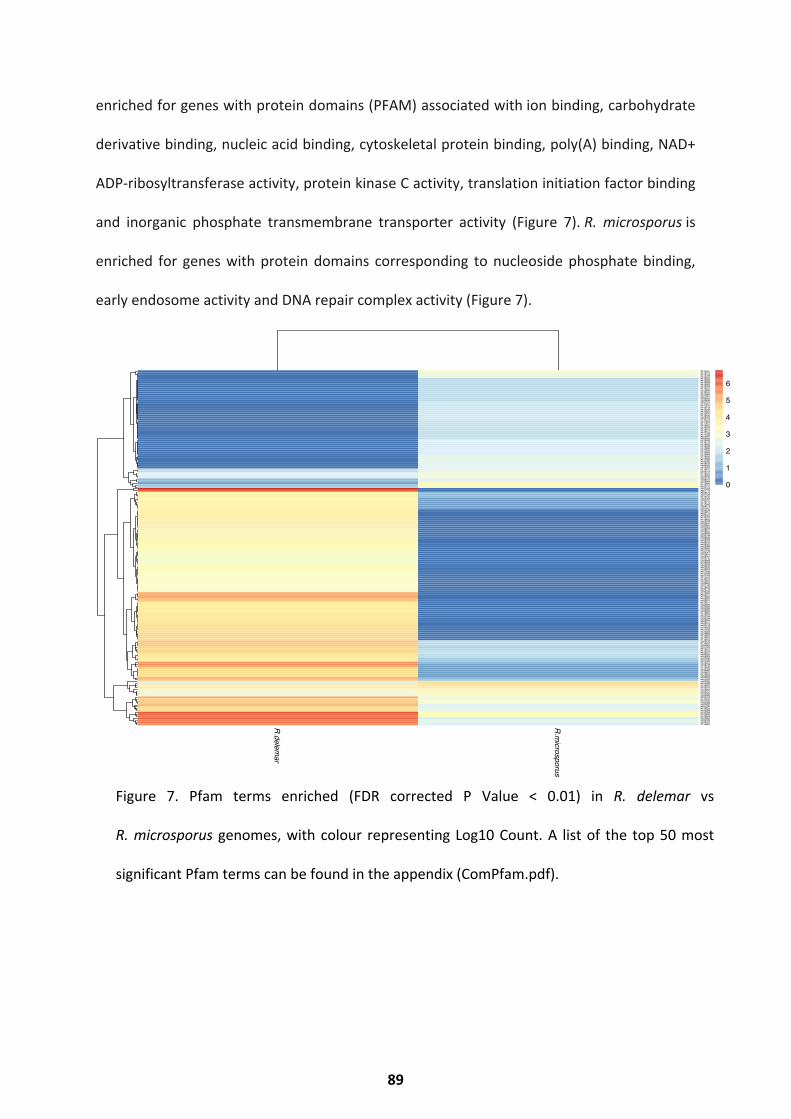
enriched for genes with protein domains (PFAM) associated with ion binding, carbohydrate
derivative binding, nucleic acid binding, cytoskeletal protein binding, poly(A) binding, NAD+
ADP-ribosyltransferase activity, protein kinase C activity, translation initiation factor binding
and inorganic phosphate transmembrane transporter activity (Figure 7). R. microsporus is
enriched for genes with protein domains corresponding to nucleoside phosphate binding,
early endosome activity and DNA repair complex activity (Figure 7).
Figure 7. Pfam terms enriched (FDR corrected P Value < 0.01) in R. delemar vs
R. microsporus genomes, with colour representing Log10 Count. A list of the top 50 most
significant Pfam terms can be found in the appendix (ComPfam.pdf).
R.delemar
R.microsporus
PF13041PF12773PF12796PF16619PF18439PF16983PF16589PF16363PF16159PF14575PF13855PF13555PF10805PF01061PF00454PF00488PF02724PF17921PF17675PF16118PF14662PF13567PF13479PF13428PF09972PF10779PF17862PF17817PF16367PF13533PF13424PF13417PF12799PF11932PF00091PF02786PF12760PF13173PF16842PF16588PF13843PF12330PF10500PF10662PF13304PF13426PF14362PF00036PF18345PF09726PF13561PF13472PF00013PF13401PF13920PF03142PF05347PF00505PF12894PF13191PF00176PF01498PF07714PF05175PF07728PF14510PF03009PF00107PF13176PF12550PF00173PF07647PF00433PF04851PF03144PF00564PF13014PF12854PF02222PF03810PF00891PF02353PF08477PF08581PF05496PF00158PF05406PF08028PF08726PF00610PF06293PF02689PF02390PF01749PF06367PF02874PF05001PF02770PF08448PF06609PF13232PF09439PF00626PF05673PF01636PF14598PF01163PF13912PF10591PF03602PF08443PF01740PF13429PF02719PF05190PF12895PF00077PF13606PF08284PF00271PF13371PF00690PF00806PF00658PF08282PF13881PF13374PF02796PF03184PF08659PF09011PF00512PF05148PF13504PF01209PF14259PF09668PF13857PF13833PF13812PF03732PF13637PF00025PF01926PF13181PF13518PF00651PF00083PF00005PF07646PF00788PF13551PF08238PF13516PF12762PF08662PF04670PF13650PF00665PF07653PF02992PF07282PF00096PF12937PF00076PF14604PF00071PF00069PF04082PF00153PF14531PF01535PF00098PF07690PF00270PF14529PF00009PF13358PF00400PF13894PF00385PF00078PF03372PF13893
0
1
2
3
4
5
6
89

Comparison with Aspergillus genomes
R. delemar genes with homology to genes in Aspergillus niger were identified via blast+.
A comprehensive list of these genes can be found in the
online appendix (https://github.com/psephtonclark/Orthologues). 13,613 genes were
identified with a bit score > 80 (E Value < 1 x 10-20), of these 2,494 genes had a percentage
identity match of over 50% (Pearson, 2013). Highly conserved genes (155 with
identity % > 80%) had roles in ribosomal processes, translation, transcription, protein
metabolism, enzyme activity (kinase, transferase, hydrolase), ion transport and amine
processing.
Conclusion and Discussion
Through incorporation of RNA-Seq data and short read data, an improved R. delemar genome
and updated R. microsporus genome (specific to the UoB R. microsporus strain) were
produced. The extra R. delemar annotation and duplicated genome information will inform
further RNA-Seq analysis, whilst RNA-Seq analysis of the UoB specific R. microsporus is now
possible. Pathway analysis of the R. delemar genome revealed predicted pathways, which
show minimal missing eukaryotic components (Figure 2). As expected, there appears to be
greater conservation between the R. delemar and R. microsporus genomes, than between the
R. delemar and A. niger genomes. R. delemar does however appear enriched for many more
functions when compared to R. microsporus, a trait which may be explained by the larger
genome size and likely whole genome duplication event. Unsurprisingly, the conserved fungal
gene core shared by R. microsporus, R. delemar and A. niger is highly populated with genes
predicted (GO) to play roles in protein synthesis, a general process highly conserved across
filamentous fungi (amongst other kingdoms). Species-specific genes likely offer an advantage
90

to that species, which may be environment dependent. For example, the enrichment of
endosome activity in the rice seedling blight fungus R. microsporus (Lackner and Hertweck,
2011) may aid plant pathogenesis, as it does in U. maydis (Bielska et al. 2014). The R. delemar
genome appears enriched for phosphate transportation, which may support growth in low
micronutrient conditions. This is consistent with C. albicans reliance upon phosphate transport
in stress and starvation conditions (Ikeh et al. 2017).
Further work
To better our understanding of the Mucoralean genome, DNA sequencing of ‘ultra-long’ reads,
via Oxford Nanopore or PacBio technologies, would allow resolution of highly repetitive
regions, resulting in chromosomal level genome detail. This would inform better gene
predictions and, combined with a whole genome CRISPR-Cas9 knockout library, allow for high
quality functional gene annotations. Knockout library analysis may also reveal novel drug
targets, potentially increasing our ability to treat mucormycosis.
91

Chapter 4: Transcriptional States of Germination
This work has been adapted from the book chapter “Spore Germination of Pathogenic
Filamentous Fungi” (Sephton-Clark and Voelz 2017), for which I performed the literature
search, wrote the manuscript, completed revisions, and prepared the figures. Results have
been previously published in “Pathways of Pathogenicity: Transcriptional Stages of
Germination in the Fatal Fungal Pathogen Rhizopus delemar” (Sephton-Clark et al. 2018) for
which I conceived, designed and performed the experiments, collected the data, performed
the analysis and interpretation, wrote the manuscript, completed revisions, and prepared the
figures.
This chapter will introduce transcriptional regulation of germination in a range of fungal
species. Results will describe the transcriptional landscape of Rhizopus delemar over
germination. This fine scale RNA-Seq study sheds light on pathways highly regulated over the
course of germination, identifying pathways which could be targeted to inhibit germination,
providing new treatment options for mucormycosis. Dormant spores are shown to be
transcriptionally unique, containing a subset of transcripts absent in later developmental
stages. A large shift in the expression profile is prompted by the initiation of germination,
with genes involved in respiration, chitin, cytoskeleton, and actin regulation appearing
to be important for this transition. A period of transcriptional consistency can be seen
throughout isotropic swelling, before the transcriptional landscape shifts again at the
onset of hyphal growth. In addition, I compare germinating R. delemar spores
(transcriptionally) to germinating Aspergillus niger spores. The results identify a core set
of orthologous genes which show similar expression patterns over germination; however
germinating R. delemar
92

also exhibits a uniquely expressed set of genes. These results provide us with a greater
understanding of the regulation of germination and highlight processes involved in
transforming R. delemar from a resting spore state to a hyphal mass.
93

Germination Regulation
During spore germination, previously dormant spores adapt a metabolically active state that
is characterized by rapid changes in the transcriptional landscape, metabolism, cell wall
composition and cell physiology. Together these changes lead to isotropic growth which is
defined by swelling of the spore and a marked increase in spore size, eventually leading to
hyphal growth. The transcriptional, biochemical and physiological changes that take place
over this time course will be addressed species by species in the following section.
Aspergillus species
Transcriptional profiling of germinating Aspergillus niger shows the largest differences in gene
expression between resting conidia and germlings (Novodvorska et al. 2013; van Leeuwen et
al. 2013). Conidia having germinated for 2 hours (T2), though barely reaching isotropic
growth, show the greatest change in abundances of transcripts of all major classes, when
compared to resting conidia. Dormant conidia were enriched in transcripts involved in protein
synthesis, whilst T2 conidia contained transcripts involved in protein synthesis, energy
production, cell cycle, transcription and translation. Most strikingly, many transcripts present
in resting conidia were completely depleted in all other stages studied, demonstrating that
the largest transcriptional shift appears at the beginning of germination. T4 conidia showed
little alteration in the transcriptional landscape, compared to T2, however metabolic
transcripts, such as the NADPH-dependent carbonyl reductase homologue, appear to be the
most upregulated when compared to T2 (van Leeuwen et al. 2013).
94

It has been suggested that resting Aspergillus spp. conidia contain vast quantities of
pre-packaged messenger RNA molecules, ready to be translated into proteins
and cell components essential for growth and metabolism, once dormancy is broken
(Lamarre et al. 2008; Novodvorska et al. 2016). This could explain the vast decrease in
transcripts seen between resting and germinated conidia, accounting for the
rapid conidial germination response. Others have argued that the large quantity of
transcripts present in resting conidia are instead an artifact of conidiation and are
degraded upon germination (van Leeuwen et al. 2013).
Transcripts present in large amounts in dormant conidia, which were swiftly depleted,
play roles in the degradation of internal trehalose stores, as well as mannitol
metabolism, signalling and hydrophobin production. This combination of stored
transcripts are likely useful for both the initiation of germination, as well as conidiation,
which would support a combination of the models proposed above (van Leeuwen et al.
2013; Novodvorska et al. 2013, 2016). Transcripts for heat shock proteins and ROS
protection were also found to be high in dormant conidia and depleted upon
germination. Eight proteins involved in ROS protection were found to be specifically
enriched. These include catalase A, thioredoxin reductase, and mitochondrial
peroxiredoxin (Oh et al. 2010). A triple superoxide dismutase (SOD1,2&3) deletion in
Aspergillus fumigatus resulted in delayed germination, whilst individual deletions
resulted growth inhibition at high temperatures and sensitivity to ROS (Morales
Hernandez et al. 2010; Lambou et al. 2010). This indicates stress response and ROS
signalling is likely crucial to germination initiation.
Conidial Transcripts
95

A similar study analysing the transcriptional landscape of germinating Aspergillus niger
elucidated further information on the metabolic changes during the early stages of dormancy
disruption (Novodvorska et al. 2013). Dormant conidia were shown to possess many
transcripts involved in carbon starvation, trehalose and mannitol biosynthesis. Trehalose
mobilisation and utilisation occurs rapidly at the onset of germination in A. nidulans, as does
mannitol utilisation (Thammahong et al. 2017; Novodvorska et al. 2013), making it likely that
these transcripts were pre-packaged to aid germination initiation. Transcripts involved in
glycerol synthesis and the metabolism of conidial sugar solutes were also abundant in the
dormant conidia and are thought to be important in the immediate onset of germination to
provide energy, as their presence may allow for the immediate access to sugars required for
growth.
The presence of abundant metabolic transcripts and ROS response transcripts within dormant
conidia supports the argument that conidia come with pre-packaged mRNA ready to mobilise
carbohydrates and protect from ROS damage at germination onset. However, these
transcripts may also be required for maintaining dormancy. Internal ROS has been shown to
act as a germination signalling molecule in Colletotrichum gloeosporioides and therefor ROS
responders (SOD, catalase etc.) might aid in dormancy maintenance (Li et al. 2018; Nesher
et al. 2011)(van Leeuwen et al. 2013). Similarly, low concentrations of ATP have been shown
to inhibit trehalase, thus inhibiting trehalose mobilisation and germination initiation
(Thevelein, den Hollander, and Shulman 1982), therefore low levels of metabolic activity
may be required for ATP production to maintain dormancy. For the above transcripts
to play a role in maintaining Aspergillus sp. dormancy, translation would be
96

required; spores of Saccharomyces cerevisiae have been found to contain transcripts
which are constantly translated throughout dormancy, and it has been suggested that
this ongoing expression plays a role in long term spore survival (Brengues, Pintard, and
Lapeyre 2002). If the purpose of conidial transcripts are to aid germination initiation, the
mRNA may be capped and packaged to increase stability. Highly stable mRNA has
been detected in dormant S. cerevisiae spores, these mRNAs were stable for several
months, but rapidly degraded upon germination onset (Brengues, Pintard, and
Lapeyre 2002). If transcripts within the conidia are remnants of conidiation, to
reduce degradation there may be either a lack of degradative enzymes such as
ribonucleases existing freely within the conidia, or these enzymes are kept
compartmentalized, as many fungal species compartmentalise enzymes necessary for
secondary metabolism (Kistler and Broz 2015).
I suspect that there is more than one explanation for the vast transcriptional shift upon
germination initiation. Given that ‘resting’ A. niger conidia show metabolic activity
(Novodvorska et al. 2016), it is likely that selected transcripts are translated during
dormancy (Brengues, Pintard, and Lapeyre 2002). Selected transcripts might play a role in
dormancy maintenance as autoinhibitors do, or by controlling ROS and ATP as previously
discussed. As transcripts are rapidly degraded upon germination and play roles in trehalose
mobilisation, it is likely that some transcripts are also pre-packaged (polyadenylated and
capped) ready for germination initiation, as is seen for S. cerevisiae and M. tuberculosis
(Brengues, Pintard, and Lapeyre 2002; Ignatov et al. 2015). There is less support for
that argument that the transcripts found in dormant spores are artifacts of
conidiation, as the longest lived mRNA in transcriptionally inactive S. cerevisiae has a
half-life of under 100 minutes (Wang et al. 2002).
97

Germination Metabolism
Follow up work on the metabolic changes in germinating Aspergillus niger showed that
dormant conidia suspended in water alone exhibited low levels of respiration (Novodvorska
et al. 2016). This either suggests that a nutrient source is less essential for initiating the
process of germination, as can be seen in C. neoformans, or that dormant conidia in fact
maintain a low level of metabolism at all times, as is supported by Brengues et al (Brengues,
Pintard, and Lapeyre 2002). Transcripts involved in oxidative phosphorylation and therefore
respiration, were also shown to be present in dormant A. niger conidia (Novodvorska et al.
2016). Proteomics showed that at early time points, proteins containing the Has domain with
potential aldehyde-lyase activity, proteins with potential enoyl-[acyl-carrier-protein]
reductase activity and proteins with translation elongation factor activity were abundant.
These proteins are suspected to play roles in carbohydrate metabolism, oxidative
phosphorylation and translation, respectively (Novodvorska et al. 2016).
Initial germination relies on fermentation to derive energy, probably from internal carbon
sources such as trehalose. Fermentation rapidly switches to respiration after 25 minutes of
germination. After 60 minutes of germination, glucose uptake can be detected, suggesting
conidia may use their internal carbon sources to support the initial stages of germination,
accounting for the metabolic activity measured in water alone (Novodvorska et al. 2016). An
array of glucose and hexose transporters (An02g03540, An15g03940, An05g01290,
An16g08940, An02g01270) were upregulated during the early stages of germination, with the
low affinity and high affinity transporters An02g03540 (an A. nidulans homologue) and a mstA
homologue, respectively, appearing most upregulated at the early and later time points
(Novodvorska et al. 2016). Several G protein coupled receptors were also highly transcribed
98

at the early stages of germination (Novodvorska et al. 2016). As G protein coupled receptors
are known to play roles in signalling and nutrient sensing (Choi et al. 2015), it is unsurprising
that several serine and threonine phosphatases, linked with glucose uptake, were also found
to be upregulated at these time points.
The presence of sorbic acid prolonged the fermentation phase of germination in A. niger,
altering the metabolism of several substrates including trehalose and ergosterol
(Novodvorska et al. 2016). The germination of Aspergillus species is also reliant upon
RAS/MAPKinase signalling along with cAMP/PKA signalling in response to nutrients. An
upregulation of amino acid metabolism also appears necessary for germination, as proteins
within germinating Aspergillus fumigatus show a specific requirement for lysine (Osherov et
al. 2002; Lamarre et al. 2008).
Neurospora crassa
A transcriptional study of germination in the model organism and plant pathogen Neurospora
crassa showed similar changes during isotropic growth and hyphal emergence to those
described in Aspergillus spp. (Kasuga et al. 2005). Neurospora crassa will begin to germinate
upon immersion in water with the correct salts and carbon sources. Vogel’s medium
is commonly used to initiate germination in laboratory conditions. This is a medium
that contains: sucrose, water, Na3 citrate, KH2PO4, NH4NO3, MgSO4, CaCl2, trace elements
and biotin. Germination results in swelling of the irregular spores, followed by changes in the
cell wall composition and hyphal formation, with most conidia possessing hyphae
at around 4 hours (Kasuga et al. 2005). Biochemical analysis has shown that in the time
immediately after germination induction, internal stores of carbon such as trehalose and
glutamic acid are
99

released and degraded (Kasuga et al. 2005). Transcriptionally, major differences were seen
between resting and germinated spores (Kasuga et al. 2005). Conidia-specific genes such as
the light responsive con-6 and 10, as well as 8, are enriched in resting conidia, but swiftly
depleted after germination (Kasuga et al. 2005), whilst internal amino acid stores are also
rapidly degraded following the initiation of germination. After 30 minutes of germination,
processes such as glucan and chitin synthesis, DNA processing and heat shock protein
production peaked, whilst after 1-4 hours of germination, germlings were enriched in
transcripts involved in RNA synthesis, DNA processing, ribosome biogenesis, protein
biosynthesis, amino acid biosynthesis and enzymes & proteins involved in cellular
transportation (Kasuga et al. 2005). Interestingly, the peak in amino acid production
correlates with the restoration of amino acid stores at around 3 hours post germination
induction. Respiratory transcripts peaked after 8-16 hours of germination (Kasuga et al. 2005),
correlating with the observation that oxygen consumption commences after 2-4 hours of
germination (Kasuga et al. 2005; Schmit and Brody 1976).
Fusarium species
The conservation of cellular reprogramming during germination is further evidenced by the
protein content present in germinating conidia of Fusarium oxysporum. At early time points
(3 hours), germlings were enriched for proteins involved in metabolic processes, redox
processes, RNA processing, transcription and translation. Later time points (7-11 hours)
showed an increase in proteins associated with ergosterol biosynthesis (Deng et al. 2015).
Several conidiation and germination essential proteins and factors are known to exist for the
plant pathogen Fusarium graminearum. GEA1 (Germinated Ascospores 1) is essential
for normal conidia production, as well as development of the ascus wall, in which the conidia
100

are contained (Son, Lee, et al. 2013). The gene FgATG15 also appears to be essential for
normal conidia formation, as mutants produce abnormally shaped conidia. In addition,
FgATG15 is required for the turnover of stored lipids, a process often essential for
germination (Nguyen et al. 2011). Similarly to other filamentous fungi, the production
of conidia in Fusarium graminearum relies upon the presence of an AbaA (Aspergillus)
orthologue, known to be involved in the regulation of cell cycle pathways, regulating
these processes through the conserved AbaA-Weta signalling pathway in Aspergillus spp.
and Fusarium sp. (Son, Kim, et al. 2013).
In summary, the timings of general mechanisms such as translation, metabolism and DNA
synthesis correspond well between N. crassa and Aspergillus spp. Interestingly, the timing of
more specific processes such as heat shock protein production also appears conserved
between species. It has been suggested that the presence of heat shock proteins at these
early stages in Neurospora crassa aids the correct folding of newly synthesized proteins
(Osherov et al. 2002; Kasuga et al. 2005). Similarities between transcriptional changes over
germination have also been found when comparing data from this study to changes seen in
germinating D. discoideum and U. maydis (Zahiri, Babu, and Saville 2005). Together with the
similarities seen in Aspergillus spp., this suggests the broad transcriptional changes seen
over the course of germination are highly conserved between fungi and other eukaryotes,
indicating that a 'core germination program' which evolved prior to recent eukaryotic
species divergence, may exist (Kasuga et al. 2005).
Mucorales
It has been shown that the fatty acid composition of Mucor rouxii is altered over
the course of germination, with quantities of gamma linoleic acid increasing over
101

germination, whilst other fatty acids fluctuate over germination (Khunyoshyeng et al. 2002).
The content of chitin synthase also increases over the course, predominantly under aerobic
conditions, thought to provide chitin to the cell wall as the spores swell and the cell wall
expands (Kamada, Bracker, and Bartnicki-Garcia 1991). It is known that Mucor
circinelloides relies in part in the Atf1/2 (putative transcription factors) pathway to
germinate within phagocytes (Pérez-Arques et al. 2019), and aquaporin recruitment is
required for germination of Rhizopus delemar (Turgeman et al. 2016). Aside from the
transcriptional profiles described above, the transcriptional landscape of germinating
fungi has been largely unexplored, and remains understudied for Mucorales species.
102

Results
In order to address this gap, I performed a transcriptional analysis of Rhizopus delemar spore
germination. My phenotypic analysis of spore germination (Chapter 2, Figure 3b) established
the temporal pattern for the development of spores from dormancy to filamentous growth.
These dramatic morphological changes require vast cellular reprogramming. I performed
transcriptional analysis of each stage outlined in this process (Chapter 2, Figure 3b). For high-
resolution capture of the transcriptional regulation of spore germination, mRNA was isolated
and sequenced from resting spores (0 h), swelling spores (1, 2, 3, 4, and 5 h) and during
filamentous growth (6, 12, 16, and 24 h). Three biological replicates were produced for each
time point, and mRNA from each sample was sequenced with Illumina HiSeq technology, with
100-bp paired end reads. Reads were aligned to the R. delemar genome (Ma et al. 2009),
giving an average alignment rate of over 95% per sample, with an average of 68% (12,170
genes) of all genes expressed over all time points. Genes which were never expressed in this
time course likely have roles in other processes, such as sexual reproduction, or may be
expressed under specific conditions, such as nutrient starvation.
Transcriptional Trends over Germination
Principal-component analysis (PCA) of TMM (trimmed mean of M-values) normalized read
counts per gene (Figure 1) showed that the biological replicates grouped closely together,
with time points grouping into 3 clusters separated by time (principal component 1 [PC1]) and
stage (PC2), as determined by k-means clustering).
103
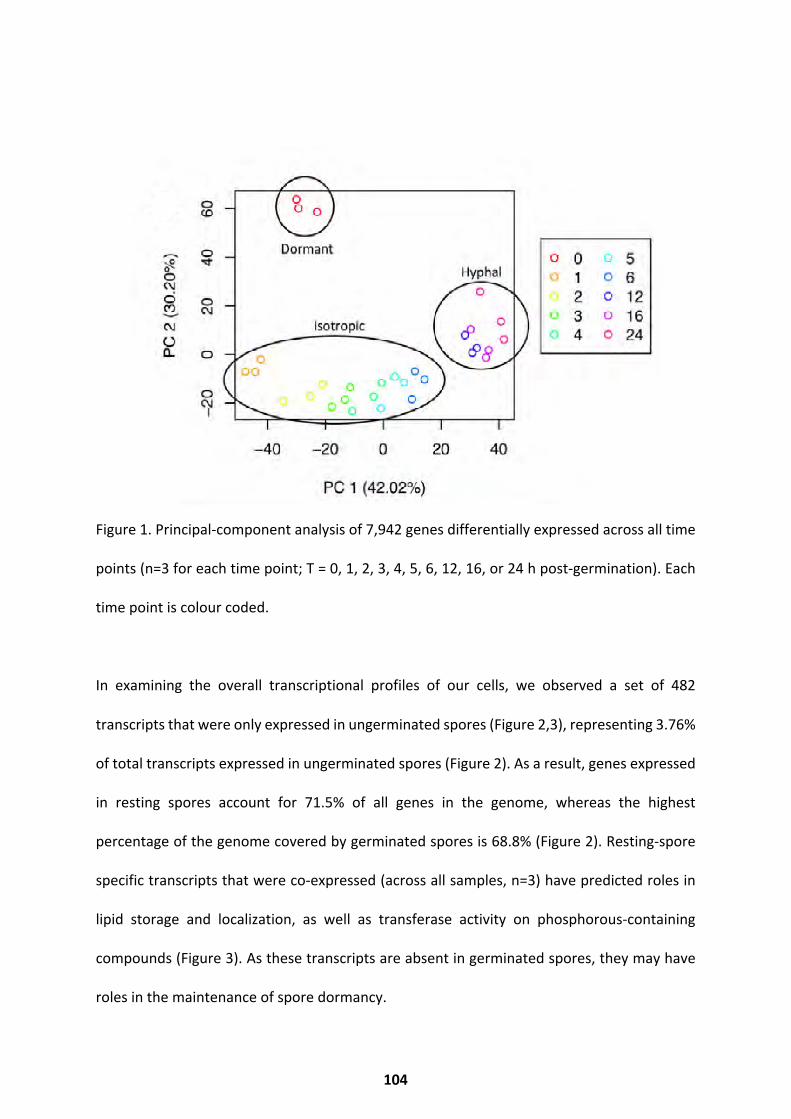
Figure 1. Principal-component analysis of 7,942 genes differentially expressed across all time
points (n=3 for each time point; T = 0, 1, 2, 3, 4, 5, 6, 12, 16, or 24 h post-germination). Each
time point is colour coded.
In examining the overall transcriptional profiles of our cells, we observed a set of 482
transcripts that were only expressed in ungerminated spores (Figure 2,3), representing 3.76%
of total transcripts expressed in ungerminated spores (Figure 2). As a result, genes expressed
in resting spores account for 71.5% of all genes in the genome, whereas the highest
percentage of the genome covered by germinated spores is 68.8% (Figure 2). Resting-spore
specific transcripts that were co-expressed (across all samples, n=3) have predicted roles in
lipid storage and localization, as well as transferase activity on phosphorous-containing
compounds (Figure 3). As these transcripts are absent in germinated spores, they may have
roles in the maintenance of spore dormancy.
104
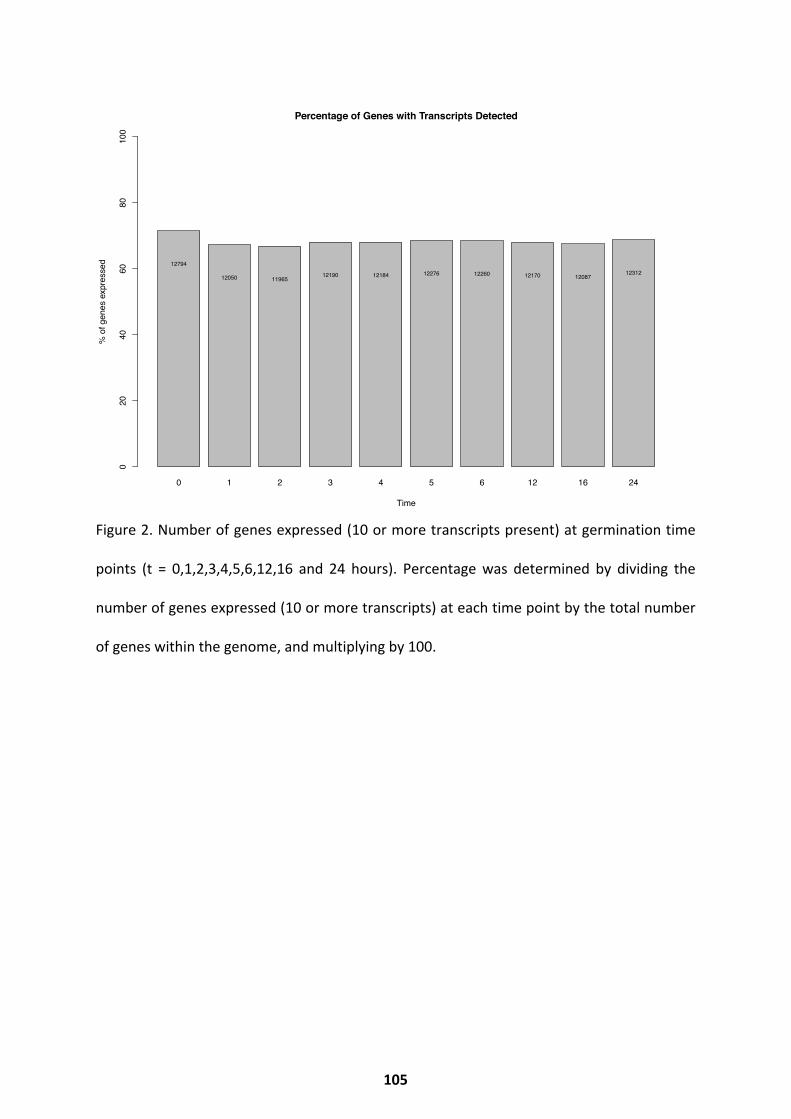
Figure 2. Number of genes expressed (10 or more transcripts present) at germination time
points (t = 0,1,2,3,4,5,6,12,16 and 24 hours). Percentage was determined by dividing the
number of genes expressed (10 or more transcripts) at each time point by the total number
of genes within the genome, and multiplying by 100.
0 1 2 3 4 5 6 12 16 24
Percentage of Genes with Transcripts Detected
Time
% o
f gen
es e
xpre
ssed
020
4060
8010
0
12794
12050 1196512190 12184 12276 12260 12170 12087
12312
105
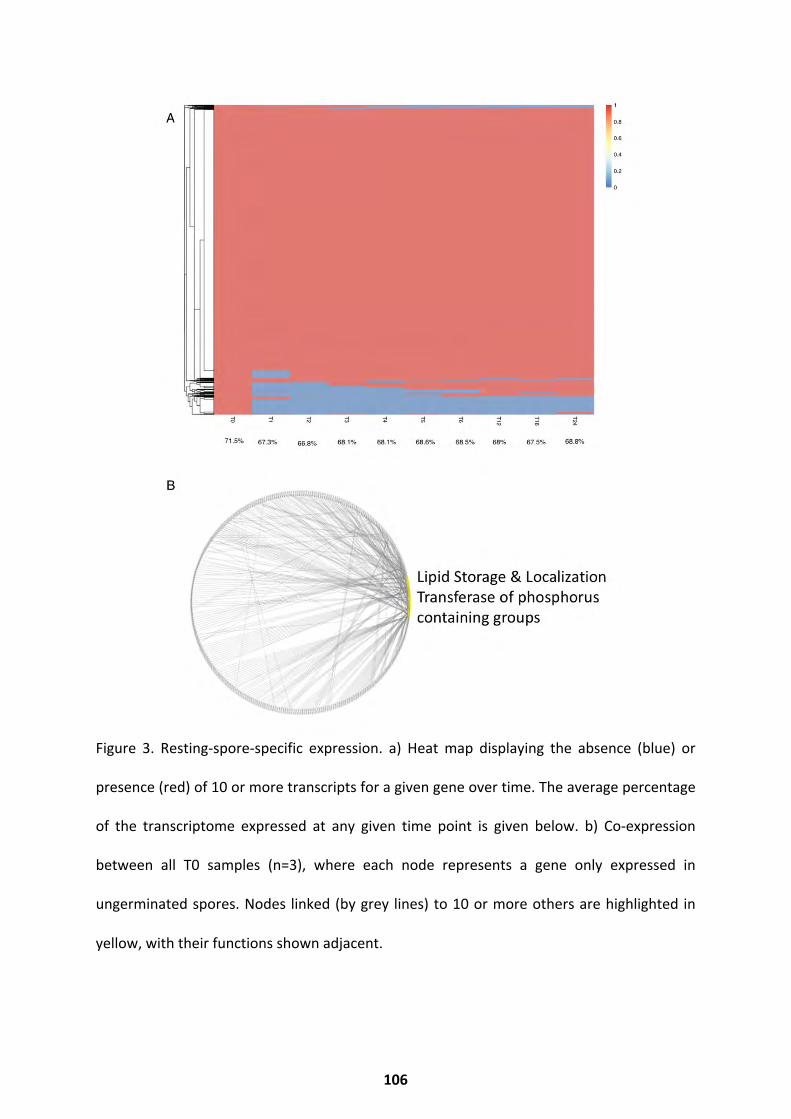
Figure 3. Resting-spore-specific expression. a) Heat map displaying the absence (blue) or
presence (red) of 10 or more transcripts for a given gene over time. The average percentage
of the transcriptome expressed at any given time point is given below. b) Co-expression
between all T0 samples (n=3), where each node represents a gene only expressed in
ungerminated spores. Nodes linked (by grey lines) to 10 or more others are highlighted in
yellow, with their functions shown adjacent.
106

A series of analyses were performed to identify the transcriptional changes occurring during
spore germination. Principle component analyses highlighted that the fungal transcriptome
displayed a time-dependent shift across 3 major clusters corresponding to the phenotypic
developmental stages swelling, germ tube emergence, and hyphal growth, indicating that
spore germination is underpinned by progressive shifts in transcriptional regulation (Figure
4,5). The transcriptome of resting spores was distinct from that of all other developmental
stages, changing dramatically between 0 and 1 h. Thereafter, the transcriptional profiles of
swelling spores and of those developing germ tubes were distinct but clustered together (2
to 6 h). Furthermore, fully established filamentous growth was characterized by a specific
transcriptional signature (12, 16, and 24 h) (Figure 4,5). Consistent with stage-specific
transcriptional changes, progressive change in differential gene expression was observed
during examination of the transcriptional profiles of each time point.
Differential Expression Throughout Germination
A total of 7,924 genes were differentially expressed across the entire time course (Figure 5a).
Analysis of differentially expressed genes by k-means clustering identified seven major
clusters of expression variation over time (Figure 5a,b). Genes in clusters 1 and 3 are
expressed at low levels in resting spores, with abundance increasing upon germination (1 h)
(Figure 4b). Both clusters are enriched (hypergeometric test, corrected P value of < 0.05) for
transcripts with predicted roles in regulation of the cytoskeleton, protein metabolism, the
electron transport chain, translation, and sugar metabolism (Figure 5c), suggesting these
processes are important for germination initiation. Clusters 4 and 6 show gene expression
levels moving from low to high over time, peaking during hyphal growth (Figure 5b). These
clusters are enriched (hypergeometric test, corrected P value of < 0.05) for transcripts with
107

predicted functions related to kinase, transferase, transposase, and oxidoreductase activities,
along with pyrimidine and phosphorous metabolism, stress response, transport, and
signalling (Figure 5c). This is consistent with the established roles for these processes in
starting and maintaining vegetative growth (van Leeuwen et al. 2013; Balmant, Sugai-guérios,
and Coradin 2015; Yao et al. 2016). Cluster 5 contains genes that have high expression levels
in both ungerminated spores and the hyphal form, but low levels during initial swelling (Figure
5b). Cluster 5 is enriched (hypergeometric test, corrected P value of < 0.05) for transcripts
with predicted functions in regulation of the cytoskeleton, transferase and hydrolase
activities, and phosphorous metabolism (Figure 5c). This suggests that these functions may
be repressed during isotropic growth to maintain swelling. Clusters 7 and 2 contain genes
with expression levels peaking in ungerminated spores (Figure 5b). These clusters are
enriched (hypergeometric test, corrected P value of < 0.05) for transcripts with predicted
functions relating to glycerone kinase, pyrophosphatase, transferase, hydrolase, and
oxidoreductase activities, as well as cofactor and coenzyme metabolism, pyrimidine, sulfur,
nitrogen, sugar, and aromatic compound metabolism. These clusters are also enriched for
reduction-oxidation (redox) processes, respiration, and stress responses (Figure 5c). Notably,
every cluster is enriched for transcripts involved in ion transport regulation, specifically
potassium, sodium, and hydrogen ions. This suggests tight regulation of transmembrane
transport of these particular ions is important for the survival of R. delemar.
108

Figure 4. Heatmap displaying sample correlation. R. delemar samples (T0-T24, n=3) are
hierarchically clustered based on gene expression patterns (Grabherr, Brian J. Haas,
Moran Yassour Joshua Z. Levin, Dawn A. Thompson, Ido Amit, Xian Adiconis, Lin Fan,
Raktima Raychowdhury, Qiandong Zeng, Zehua Chen, Evan Mauceli, Nir Hacohen, Andreas
Gnirke, Nicholas Rhind, Federica di Palma, Bruce W., and Friedman 2013). Colours
represent sample correlation, red = similar, green = dissimilar.
cond_0
cond_1
cond_2
cond_3
cond_4
cond_5
cond_6
cond_12
cond_16
cond_24
cond_0
cond_1
cond_2
cond_3
cond_4
cond_5
cond_6
cond_12
cond_16
cond_24
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_3_rep3
cond_4_rep3
cond_3_rep2
cond_4_rep2
cond_5_rep3
cond_6_rep3
cond_5_rep2
cond_6_rep2
cond_5_rep1
cond_6_rep1
cond_3_rep1
cond_4_rep1
cond_24_rep1
cond_12_rep2
cond_24_rep2
cond_24_rep3
cond_12_rep1
cond_16_rep1
cond_12_rep3
cond_16_rep2
cond_16_rep3
cond_1_rep1cond_1_rep2cond_1_rep3cond_2_rep1cond_2_rep2cond_2_rep3cond_0_rep1cond_0_rep2cond_0_rep3cond_3_rep3cond_4_rep3cond_3_rep2cond_4_rep2cond_5_rep3cond_6_rep3cond_5_rep2cond_6_rep2cond_5_rep1cond_6_rep1cond_3_rep1cond_4_rep1cond_24_rep1cond_12_rep2cond_24_rep2cond_24_rep3cond_12_rep1cond_16_rep1cond_12_rep3cond_16_rep2cond_16_rep3
sample correlation matrix diffExpr.P0.001_C2.matrix.log2
0.5 0.7 0.9Value
Color Key
2 4 6 8 10 12
010
000
2000
030
000
4000
0
Number of Clusters
With
in C
lust
er S
um o
f Squ
ares
110
109
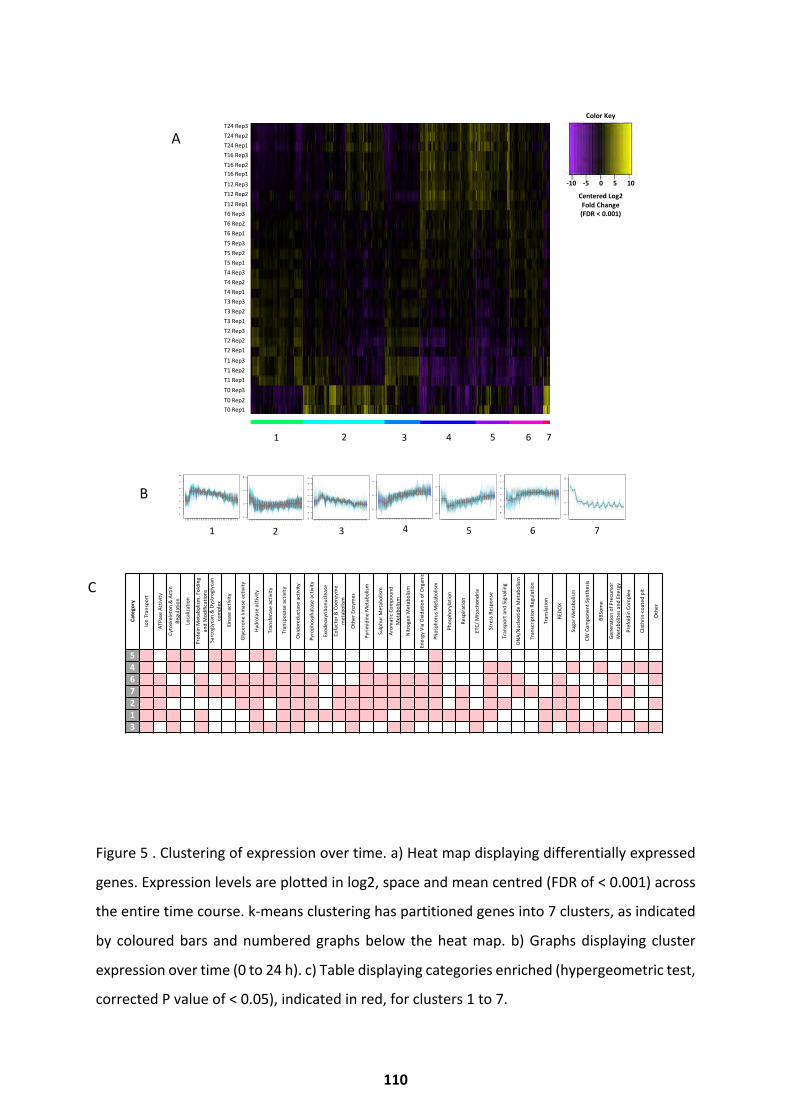
Figure 5 . Clustering of expression over time. a) Heat map displaying differentially expressed
genes. Expression levels are plotted in log2, space and mean centred (FDR of < 0.001) across
the entire time course. k-means clustering has partitioned genes into 7 clusters, as indicated
by coloured bars and numbered graphs below the heat map. b) Graphs displaying cluster
expression over time (0 to 24 h). c) Table displaying categories enriched (hypergeometric test,
corrected P value of < 0.05), indicated in red, for clusters 1 to 7.
T24Rep3T24Rep2T24Rep1T16Rep3T16Rep2T16Rep1
T12Rep3T12Rep2
T12Rep1T6Rep3T6Rep2T6Rep1T5Rep3T5Rep2T5Rep1T4Rep3T4Rep2T4Rep1T3Rep3T3Rep2T3Rep1T2Rep3T2Rep2T2Rep1
T1Rep3T1Rep2T1Rep1
T0Rep3
T0Rep2T0Rep1
Catego
ry
Ion
Tran
spor
t
ATPa
seA
ctiv
ity
Cyto
skel
eton
&A
ctin
Regu
latio
n
Loca
lizat
ion
Prot
ein
Met
abol
ism,F
oldi
ng
and
Mod
ifica
tions
Sarc
ogly
can&D
ystr
ogly
can
com
plex
Kina
sea
ctiv
ity
Gly
cero
neki
nasea
ctiv
ity
Hydr
olas
eac
tivity
Tran
sfer
ase
activ
ity
Tran
spos
ase
activ
ity
Oxi
dore
duct
ase
activ
ity
Pyro
phos
phat
ase
activ
ity
Exod
eoxy
ribon
ucle
ase
Cofa
ctor&C
oenz
yme
met
abol
ism
Oth
erE
nzym
es
Pyrim
idin
eM
etab
olism
Sulp
hurM
etab
olism
Arom
aticC
ompo
und
Met
abol
ism
Nitr
ogen
Met
abol
ism
Ener
gyV
iaO
xida
tion
ofO
rgan
ic
Phos
phor
usM
etab
olism
Phos
phor
ylat
ion
Resp
iratio
n
ETC/M
itoch
ondr
ia
Stre
ssR
espo
nse
Tran
spor
tand
Sig
nalin
g
DNA/
Nuc
leot
ide
Met
abol
ism
Tran
scrip
tion
Regu
latio
n
Tran
slatio
n
REDO
X
Suga
rMet
abol
ism
CWC
ompo
nentS
ynth
esis
BBSo
me
Gen
erat
ion
ofP
recu
rsor
Met
abol
itesa
ndE
nerg
y
Pref
oldi
nCo
mpl
ex
Clat
hrin
-coa
ted
pit
Oth
er5467213
−50
5
subcluster_5_log2_medianCentered_fpkm.txt, 889 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
● ● ●
● ●● ● ●
●●
●
● ●●
● ● ●
●●
●●
●●
● ●● ●
●● ●
5
−50
5
subcluster_4_log2_medianCentered_fpkm.txt, 1474 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●
● ●●
●●
●
●
●
●
●
●
●
●
●
●●
●● ● ●
●●
●●
● ●
●
●●
4
−6−4
−20
24
6
subcluster_6_log2_medianCentered_fpkm.txt, 904 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●
● ●
●●
●
●●
●
● ●●
● ● ● ● ● ● ●●
●●
● ●●
● ●
●
●●
6
−50
510
subcluster_7_log2_medianCentered_fpkm.txt, 158 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●
● ●
●
● ● ●
● ●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
● ●
●
● ●
7
−50
510
subcluster_2_log2_medianCentered_fpkm.txt, 2185 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●● ●
●● ●
● ●● ●
●● ●
●
● ●
●
● ●●
●
●
●
● ●● ●
●
●●
2
−6−4
−20
24
6
subcluster_1_log2_medianCentered_fpkm.txt, 1378 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●
●●
● ● ●●
●●
●
●●
●
●
●●
●
● ●●
●
●●
●●
●●
●
●
●
1
−6−4
−20
24
68
subcluster_3_log2_medianCentered_fpkm.txt, 954 trans
cent
ered
log2
(fpkm
+1)
cond
_0_r
ep1
cond
_0_r
ep2
cond
_0_r
ep3
cond
_1_r
ep1
cond
_1_r
ep2
cond
_1_r
ep3
cond
_2_r
ep1
cond
_2_r
ep2
cond
_2_r
ep3
cond
_3_r
ep1
cond
_3_r
ep2
cond
_3_r
ep3
cond
_4_r
ep1
cond
_4_r
ep2
cond
_4_r
ep3
cond
_5_r
ep1
cond
_5_r
ep2
cond
_5_r
ep3
cond
_6_r
ep1
cond
_6_r
ep2
cond
_6_r
ep3
cond
_12_
rep1
cond
_12_
rep2
cond
_12_
rep3
cond
_16_
rep1
cond
_16_
rep2
cond
_16_
rep3
cond
_24_
rep1
cond
_24_
rep2
cond
_24_
rep3
●● ●
●
●●
●
●
●
●
●●
●
●●
●●
●
●●
●●
●
● ● ● ●● ● ●
3
54 6 721 3
ColorKey
-10-50510
CenteredLog2FoldChange(FDR<0.001)
A
B
C
110

Ungerminated spores have a radically different expression profile from germinated spores
(6,456 significantly differentially expressed genes; false-discovery rate [FDR] of < 0.001): this
is reflected by the functions of transcripts enriched in ungerminated spores. By pairwise
comparisons of differentially expressed genes between time points, the largest
transcriptional changes were seen during the first hour of germination (3,476 genes
upregulated and 2,573 genes downregulated [Figure 6a]). The large quantity of apparently
down regulated transcripts may in fact represent transcripts which have been degraded, or
immediately utilised upon germination initiation. Following this, there was a period of
transcriptional consistency over the course of isotropic swelling, where few or no genes
were found differentially expressed (Figure 6a). A noticeable shift in differential
expression then bridges the beginning and later stages of hyphal growth (6 to 12 h
[Figure 6a]). At the beginning of germination, an increase is observed in expression of
transcripts with predicted roles in stress response, mitochondrial ribonucleases (MRP),
the prefoldin complexes, organophosphate and sulfur metabolism, and
transposase, ATPase, nucleoside triphosphatase, and glycerone kinase activities (Figure
6b). A decrease in expression of genes with predicted functions in the organization of
the actin cytoskeleton, carbohydrate metabolism, translation initiation factors,
hexon binding, and phosphodiesterase, arylformamidase, galactosylceramidase, and
precorrin-2 dehydrogenase activities is also seen (Figure 6b). Notably, some categories are
both positively and negatively regulated at the beginning of germination: transcripts
predicted to have roles in ion channel activity and hydrolase and pyrophosphatase
activities do not always trend together (Figure 6b). It is likely these processes may involve
several regulatory mechanisms implicated in initializing germination.
After initiation (1 to 2 h), there is an overall trend of downregulation. The majority of
transcripts that were upregulated at 1 h are downregulated at 2 h (Figure 6b), suggesting a
111

reorganization of the transcriptome upon germination initiation. Notably, metabolism of
sulfur, organophosphate, and thiamine diphosphate remains downregulated at both 2 and 3
h. After the transcriptional stability during isotropic growth and hyphal emergence,
transcripts with predicted roles in stress response, respiration, ATPase and nucleoside
triphosphatase activities, and redox increase during early hyphal growth (Figure 6b). Between
6 and 12 h, the proportion of downregulated transcripts decreases, with hydrolase and
pyrophosphatase activities appearing both up and downregulated.
112
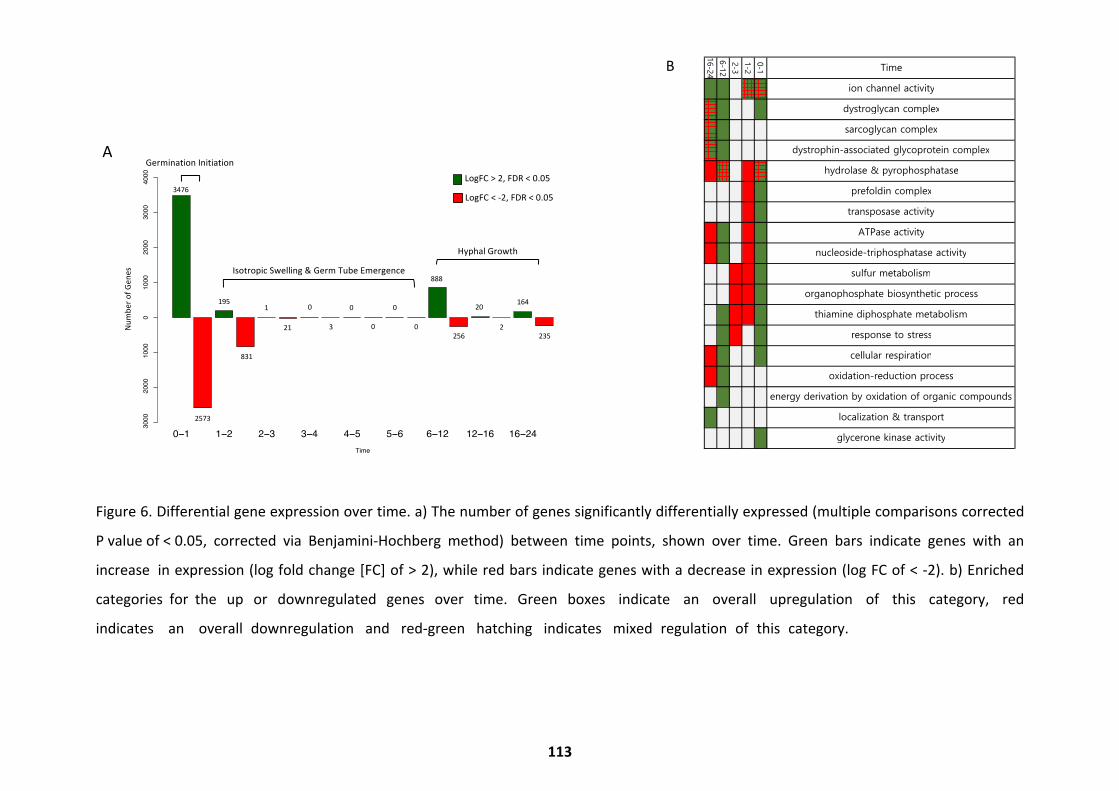
Figure 6. Differential gene expression over time. a) The number of genes significantly differentially expressed (multiple comparisons corrected
P value of < 0.05, corrected via Benjamini-Hochberg method) between time points, shown over time. Green bars indicate genes with an
increase in expression (log fold change [FC] of > 2), while red bars indicate genes with a decrease in expression (log FC of < -2). b) Enriched
categories for the up or downregulated genes over time. Green boxes indicate an overall upregulation of this category, red
indicates an overall downregulation and red-green hatching indicates mixed regulation of this category.
0−11−2
2−33−4
4−55−6
6−1212−16
16−24Tim
e
Number of Genes
−3000 −2000 −1000 0 1000 2000 3000 4000
3476
1951
00
0
868
20164
−2573
−831
−21−3
00
−256
−2
−235
A
B
Germ
inationInitiationIsotropicSw
elling&
Germ
TubeEm
ergence
HyphalG
rowth
NumberofGenes
3476
10
00
888
20164
195
00
2573
831
213
2562
235
LogFC<-2,FDR
<0.05
LogFC>2,FDR
<0.05
Time
ion channel activity
dystroglycan complex
sarcoglycan complex
dystrophin-associated glycoprotein complex
hydrolase & pyrophosphatase
prefoldin complex
transposase activity
ATPase activity
nucleoside-triphosphatase activity
sulfur metabolism
organophosphate biosynthetic process
thiamine diphosphate metabolism
response to stress
cellular respiration
oxidation-reduction process
energy derivation by oxidation of organic compounds
localization & transport
glycerone kinase activity
0-1
1-2
2-3
6-12
16-24
0−1 1−2 2−3 3−4 4−5 5−6 6−12 12−16 16−24Time
Num
ber o
f Gen
es
−300
0−2
000
−100
00
1000
2000
3000
4000
3476
1951 0 0 0
868
20164
−2573
−831
−21 −3 0 0
−256
−2
−235
A
B
GerminationInitiation
IsotropicSwelling&GermTubeEmergence
HyphalGrowth
Num
bero
fGen
es
3476
1 0 0 0
888
20164195
0 0
2573
831
21 3256
2235
LogFC <-2,FDR<0.05
LogFC >2,FDR<0.05
Tim
e
ion
chan
nelac
tivi
ty
dys
trogly
can
com
ple
x
sarc
ogly
can
com
ple
x
dys
trophi
n-as
soci
ated
gly
copro
tein
com
ple
x
hydro
lase
&pyr
opho
spha
tase
pre
fold
in c
om
ple
x
tran
sposa
seac
tivi
ty
ATP
ase
activi
ty
nucl
eoside-
trip
hosp
hata
seac
tivi
ty
sulfu
rm
etab
olis
m
org
anopho
spha
tebio
synt
hetic
pro
cess
thia
min
edip
hosp
hate
met
abolis
m
resp
ons
e to
stres
s
cellu
lar
resp
irat
ion
oxi
dat
ion-
reduc
tion
pro
cess
ener
gy
der
ivat
ion
by
oxi
dat
ion
oforg
anic
com
poun
ds
loca
lizat
ion
&tran
sport
gly
cero
neki
nase
activi
ty
0-1
1-2
2-3
6-12
16-24
B
113

Similar trends are observed when analysis of is carried out to determine the expression
of genes with predicted functions (Pfam) in carbohydrate metabolism (Figure 7a), inorganic
ion metabolism (Figure 7b), lipid metabolism (Figure 7c), amino acid metabolism (Figure 7d)
and signalling (Figure 7e). Resting spore expression appears most similar to post-
germination initiation expression. When carbohydrate metabolism is examined (Figure
7a) two subsets appear to gradually increase in expression over time,
demonstrating the sustained requirement for respiration as biomass increases. Resting
spores appear to express a subset of genes involved in inorganic ion metabolism (Figure 7b),
which appear lowly expressed post germination-initiation, whilst other subsets are stably
expressed throughout. Resting spores also express a subset of genes involved in lipid
metabolism (Figure 7c), whilst other subsets appear highly expressed only after
germination is initiated. This demonstrates the potential reliance of resting spore on
stored lipids to initiate and sustain germination. Expression of genes with predicted
roles in amino acid metabolism appears relatively stable over germination (Figure
7d), whilst other small subsets appear highly expressed within resting spores (0h), and
again in filamentous form (12-24h). A slight increase in expression can also be noted at the
onset of germination (1h), indicating spores rely on protein synthesis to kick-start
germination and biomass production, as expected. Expression of genes with predicted
functions in signalling show 3 distinct trends (Figure 7e): One subset appears highly
expressed in resting spores (0h), a second subset is highly expressed at the onset of
germination and throughout swelling(1-6h) and a third is highly expressed within
filamentous forms (12-24h). These subsets correspond with the phenotypic stages
outlined for germination of these spores, and demonstrate the reliance on unique
signalling pathways to initiate and maintain these phenotypic stages.
Metabolic Activity over Germination
114
A
B
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_008323RO3G_012185RO3G_001059RO3G_004115RO3G_000632RO3G_011127RO3G_004528RO3G_008260RO3G_014039RO3G_002362RO3G_008239RO3G_006092RO3G_004685RO3G_008251RO3G_001638RO3G_014633RO3G_004999RO3G_009004RO3G_016877RO3G_010968RO3G_016768RO3G_013162RO3G_012427RO3G_013624RO3G_014140RO3G_007730RO3G_005361RO3G_007853RO3G_013976RO3G_005729RO3G_008951RO3G_004751RO3G_016738RO3G_004921RO3G_005877RO3G_017335RO3G_007078RO3G_009715RO3G_008034RO3G_001942RO3G_002646RO3G_005964RO3G_015358RO3G_017108RO3G_008584RO3G_010045RO3G_002082RO3G_012639RO3G_003861RO3G_004078RO3G_001101RO3G_004220RO3G_010972RO3G_006347RO3G_013397RO3G_002878RO3G_017698RO3G_007012RO3G_001621RO3G_010931RO3G_008164RO3G_004044RO3G_007331RO3G_014894RO3G_006801RO3G_011425RO3G_012420RO3G_012419RO3G_000184RO3G_001635RO3G_013857RO3G_004050RO3G_006184RO3G_004425RO3G_008235RO3G_013623RO3G_010776RO3G_012402RO3G_012058RO3G_006644RO3G_006723RO3G_015767RO3G_004983RO3G_013713RO3G_005612RO3G_007534RO3G_013999RO3G_010040RO3G_002079RO3G_004077RO3G_011844RO3G_008302RO3G_009331RO3G_016651RO3G_017232RO3G_002460RO3G_013782RO3G_014473RO3G_009341RO3G_006226RO3G_004228RO3G_002098RO3G_005446RO3G_007868RO3G_002765RO3G_013845RO3G_015933RO3G_007811RO3G_001497RO3G_012210RO3G_015589RO3G_005668RO3G_013904RO3G_013412RO3G_008732RO3G_006954RO3G_005593RO3G_003325RO3G_016577RO3G_017362RO3G_008703RO3G_003470RO3G_008907RO3G_014733RO3G_013450RO3G_005022RO3G_002745RO3G_013990RO3G_002184RO3G_005854RO3G_002305RO3G_014443RO3G_009535RO3G_015186RO3G_003642RO3G_000355RO3G_003296RO3G_014472RO3G_014451RO3G_001373RO3G_015025RO3G_008847RO3G_014223RO3G_016455RO3G_015407RO3G_007401RO3G_003412RO3G_012581RO3G_010873RO3G_012763
0
5
10
15
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_009004RO3G_012073RO3G_014633RO3G_001166RO3G_009943RO3G_002989RO3G_012036RO3G_012618RO3G_001942RO3G_008323RO3G_009608RO3G_006082RO3G_012413RO3G_005845RO3G_007727RO3G_002370RO3G_005797RO3G_005023RO3G_010987RO3G_009644RO3G_002905RO3G_017452RO3G_005055RO3G_005319RO3G_004915RO3G_010605RO3G_009715RO3G_007605RO3G_011964RO3G_001303RO3G_015117RO3G_000243RO3G_006639RO3G_010455RO3G_006640RO3G_006147RO3G_007732RO3G_014055RO3G_014118RO3G_011762RO3G_008718RO3G_007953RO3G_005090RO3G_012321RO3G_017837RO3G_005152RO3G_011152RO3G_001114RO3G_016619RO3G_006312RO3G_017038RO3G_016064RO3G_003227RO3G_001339RO3G_007143RO3G_009336RO3G_007591RO3G_014221RO3G_007959RO3G_012220RO3G_003176RO3G_014024RO3G_015783RO3G_005988RO3G_001553RO3G_012410RO3G_012281RO3G_016289RO3G_011710RO3G_002312RO3G_014368RO3G_011347RO3G_017800RO3G_009663RO3G_003379RO3G_006723RO3G_009245RO3G_010477RO3G_013921RO3G_016843RO3G_006368RO3G_009246RO3G_015256RO3G_007775RO3G_008588RO3G_010688RO3G_005612
0
5
10
15
116
115
C
D
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_003904RO3G_007535RO3G_007998RO3G_015853RO3G_016448RO3G_003874RO3G_014067RO3G_008958RO3G_015754RO3G_015627RO3G_001549RO3G_015348RO3G_016315RO3G_008791RO3G_012132RO3G_004127RO3G_010246RO3G_015164RO3G_007979RO3G_010030RO3G_004402RO3G_012829RO3G_003224RO3G_009527RO3G_008510RO3G_008509RO3G_010431RO3G_010223RO3G_008234RO3G_008099RO3G_005919RO3G_017062RO3G_003905RO3G_014387RO3G_012652RO3G_008488RO3G_014326RO3G_011635RO3G_008622RO3G_003469RO3G_013001RO3G_015575RO3G_016681RO3G_011822RO3G_015320RO3G_008710RO3G_009620RO3G_009086RO3G_015330RO3G_000546RO3G_017340RO3G_001223RO3G_001705RO3G_012062RO3G_011722RO3G_010276RO3G_010483RO3G_007119RO3G_000706RO3G_013088RO3G_009533RO3G_006161RO3G_001493RO3G_015564RO3G_011743RO3G_003457RO3G_008635RO3G_015669RO3G_004225RO3G_014735
0
2
4
6
8
10
12
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_003025RO3G_016154RO3G_012545RO3G_008259RO3G_005612RO3G_006189RO3G_008584RO3G_007010RO3G_012989RO3G_007502RO3G_005368RO3G_002723RO3G_006723RO3G_003542RO3G_010906RO3G_009146RO3G_015687RO3G_017286RO3G_006890RO3G_005112RO3G_010844RO3G_004802RO3G_011418RO3G_016366RO3G_012700RO3G_009159RO3G_013994RO3G_011714RO3G_008151RO3G_013710RO3G_003036RO3G_001695RO3G_009237RO3G_001280RO3G_013434RO3G_006615RO3G_007665RO3G_005383RO3G_002471RO3G_011847RO3G_005901RO3G_005301RO3G_003642RO3G_015339RO3G_007591RO3G_007244RO3G_006081RO3G_011196RO3G_000697RO3G_013969RO3G_012926RO3G_016118RO3G_007651RO3G_007708RO3G_014305RO3G_010418RO3G_000945RO3G_017795RO3G_013468RO3G_007218RO3G_012520RO3G_009730RO3G_002537RO3G_002355RO3G_006653RO3G_000148RO3G_006147RO3G_008727RO3G_006006RO3G_012978RO3G_002307RO3G_008848RO3G_015491RO3G_011172RO3G_004575RO3G_011660RO3G_009223RO3G_002715RO3G_010704RO3G_017438RO3G_017437RO3G_013738RO3G_003714RO3G_013750RO3G_001678RO3G_011793RO3G_016642RO3G_011303RO3G_017134RO3G_003231RO3G_006094RO3G_008323RO3G_016233RO3G_016753RO3G_014633RO3G_009004RO3G_010244RO3G_014646RO3G_015786RO3G_010673RO3G_009202RO3G_005927RO3G_015749RO3G_005770RO3G_006314RO3G_014414RO3G_015076RO3G_017764RO3G_004502RO3G_006897RO3G_009855RO3G_001166RO3G_014468RO3G_017770RO3G_002446RO3G_012145RO3G_005601RO3G_016127RO3G_009512RO3G_008578RO3G_006400RO3G_013976RO3G_004233RO3G_003929RO3G_007052RO3G_008747RO3G_009125RO3G_012611RO3G_007256RO3G_001942RO3G_008413RO3G_014770RO3G_005214RO3G_009715RO3G_000961RO3G_001157RO3G_010059RO3G_012647RO3G_011773RO3G_004872RO3G_007427
0
5
10
15
116
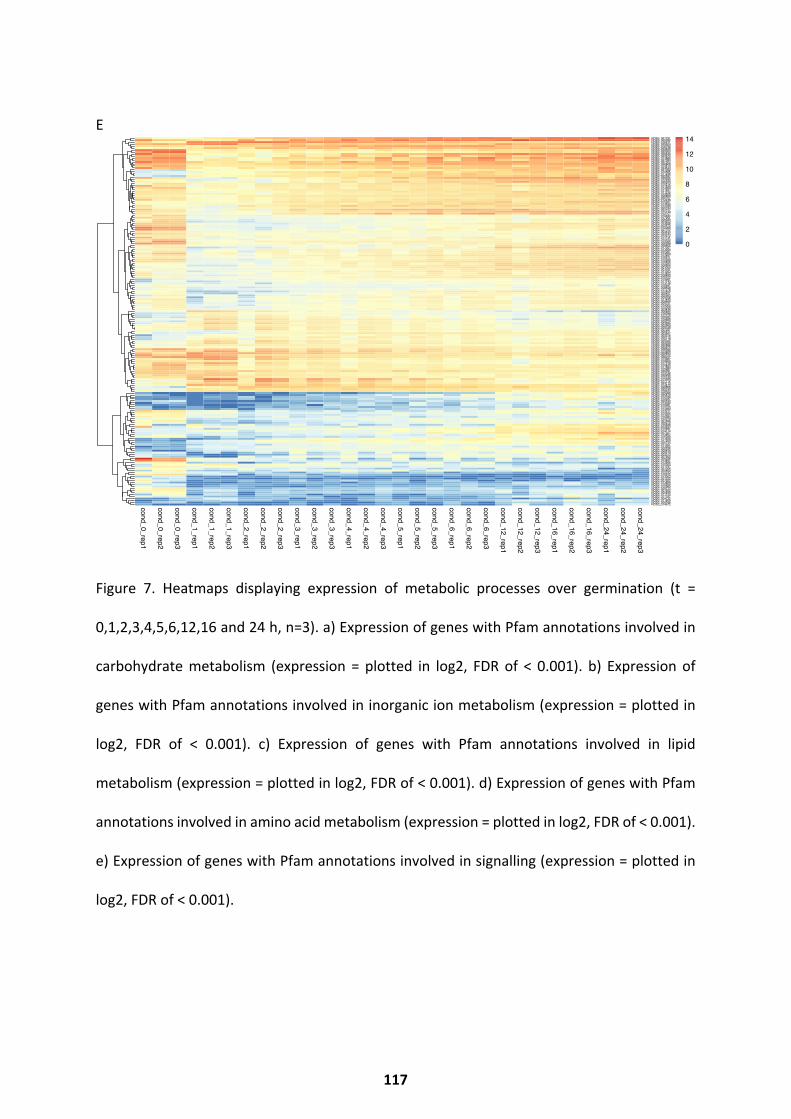
E
Figure 7. Heatmaps displaying expression of metabolic processes over germination (t =
0,1,2,3,4,5,6,12,16 and 24 h, n=3). a) Expression of genes with Pfam annotations involved in
carbohydrate metabolism (expression = plotted in log2, FDR of < 0.001). b) Expression of
genes with Pfam annotations involved in inorganic ion metabolism (expression = plotted in
log2, FDR of < 0.001). c) Expression of genes with Pfam annotations involved in lipid
metabolism (expression = plotted in log2, FDR of < 0.001). d) Expression of genes with Pfam
annotations involved in amino acid metabolism (expression = plotted in log2, FDR of < 0.001).
e) Expression of genes with Pfam annotations involved in signalling (expression = plotted in
log2, FDR of < 0.001).
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_001791RO3G_005082RO3G_016911RO3G_005924RO3G_016794RO3G_005438RO3G_008363RO3G_002202RO3G_009540RO3G_002670RO3G_014880RO3G_007083RO3G_007504RO3G_006808RO3G_013513RO3G_004543RO3G_001092RO3G_015565RO3G_011246RO3G_005397RO3G_006305RO3G_012030RO3G_005323RO3G_010512RO3G_017849RO3G_014693RO3G_011467RO3G_011154RO3G_013506RO3G_006869RO3G_000707RO3G_012238RO3G_015027RO3G_013534RO3G_013289RO3G_000183RO3G_004107RO3G_015149RO3G_012757RO3G_010484RO3G_017261RO3G_008039RO3G_004553RO3G_013928RO3G_002570RO3G_012088RO3G_007702RO3G_003444RO3G_003134RO3G_012572RO3G_011731RO3G_015102RO3G_010064RO3G_006588RO3G_009655RO3G_017057RO3G_013264RO3G_005387RO3G_014669RO3G_012487RO3G_010611RO3G_015596RO3G_016808RO3G_015293RO3G_008604RO3G_001009RO3G_003181RO3G_011953RO3G_013852RO3G_008490RO3G_016702RO3G_015881RO3G_011143RO3G_013148RO3G_016691RO3G_008406RO3G_017278RO3G_012450RO3G_005401RO3G_000963RO3G_008358RO3G_011490RO3G_008466RO3G_009751RO3G_014005RO3G_010306RO3G_003436RO3G_006894RO3G_016098RO3G_012120RO3G_000952RO3G_016699RO3G_009229RO3G_003564RO3G_002669RO3G_009158RO3G_007287RO3G_000671RO3G_003647RO3G_003316RO3G_000710RO3G_016178RO3G_005062RO3G_004039RO3G_000886RO3G_005972RO3G_008594RO3G_015286RO3G_009893RO3G_000722RO3G_008227RO3G_013067RO3G_016909RO3G_017159RO3G_010840RO3G_011683RO3G_015987RO3G_009091RO3G_013742RO3G_000266RO3G_015740RO3G_015060RO3G_017712RO3G_003713RO3G_014470RO3G_006886RO3G_005190RO3G_016644RO3G_005096RO3G_005879RO3G_012180RO3G_012037RO3G_002533RO3G_014030RO3G_010281RO3G_015753RO3G_014380RO3G_013277RO3G_012322RO3G_014354RO3G_016284RO3G_012079RO3G_001466RO3G_009279RO3G_016488RO3G_004933RO3G_016891RO3G_014170RO3G_007327RO3G_013802RO3G_016029RO3G_011926RO3G_017309RO3G_005744RO3G_011301RO3G_013262RO3G_000526RO3G_009434RO3G_009519RO3G_003313RO3G_007655RO3G_009296RO3G_013481RO3G_013906RO3G_015300RO3G_011797RO3G_001004RO3G_005469RO3G_012233RO3G_010915RO3G_013003RO3G_001332RO3G_007652RO3G_013894RO3G_010659RO3G_010980RO3G_005212RO3G_001952RO3G_000839RO3G_011155RO3G_017167RO3G_004732RO3G_011800RO3G_006166RO3G_014672
0
2
4
6
8
10
12
14
117

Pathways Upregulated at Alternate Germination Phases
PathwayTools annotation of the R. delemar genome allows analyses of individual pathway
regulation over the course of germination. There are currently only 4 fungal genomes (S.
cerevisiae, C. albicans, E. dermatitidis and P. rubens, available at: https://biocyc.org/organism-
summary?object=CALBI) publicly available on biocyc, which have annotated PGDB’s
(PathoLogic Databases: format compatible with PathwayTools). This annotation is the first
available (https://github.com/psephtonclark/rhior3cyc) for a Mucorales species.
Pathways which appear highly regulated during discrete germination phases (Figure 8) will be
described here. The following pathways were found to be upregulated at resting (0h) and
during hyphal growth (12-24h): protein glycosylation, chitin biosynthesis, trehalose
degradation, ribonucleotide biosynthesis, glycolysis and galactose degradation. This is
consistent with other findings which show chitin biosynthesis to be important for hyphal
emergence and extension in A. nidulans (Yamazaki et al. 2008), protein turnover to be a
hallmark of filamentous vegetative growth (Sun et al. 2017; Anderson et al. 2016) and
respiration to be important for maintaining fungal biomass (Prasad, Kurup, and Maheshwari
1979). It should be noted that glycolysis also appears down regulated at 0-1 hours,
indicating complex regulation of this process may be required to maintain dormancy
and initiate germination. Galactose degradation appears downregulated throughout
swelling (1-6h), highlighting its importance for initiation and hyphal growth.
Within resting spores (0h), hypusine and glyoxylate pathways are upregulated and NADP
mitochondrial/cytosolic conversion appears to increase, whilst spermidine pathways also
appear upregulated in both resting and initial spores (0-1h). Similarly, cysteine and glutamine
118

pathways are also upregulated at this time. Inositol degradation appears downregulated in
resting and initial spores (0-1h), indicating inositol may be important at the onset of
germination, whilst phyate degradation appears upregulated within resting spores (0h),
indicating a role in dormancy maintenance. Xyoglucan and chitin degradation appear
downregulated in resting spores (0h), along with peroxisome processes. This is consistent
with conidial A. niger, which upregulates transcripts involved in amino acid metabolism and
glyoxylate pathways, and downregulates peroxisome processes (Novodvorska et al. 2013).
At germination initiation (1h), Fe-S cluster biosynthesis appears downregulated, whilst
phyate degradation is upregulated. Within swelling spores (3-6h), cysteine and glutamine
catabolic and metabolic pathways appear upregulated, whilst NADP conversion, Cytochrome C,
ATPase and NADPH oxidoreductase activities appear downregulated. This is consistent
with A. niger germlings, which show upregulation of metabolic processes including
amino acid processing (van Leeuwen et al. 2013). Within hyphal forms (12-24h), Fe-S
cluster biosynthesis, tetrahydrofolate biosynthesis, oxoglutarate and isovalerate
decarboxylation, Acetyl CoA metabolism, glycolysis, ATPase and peroxisome activity appear
upregulated, indicating a reliance on aerobic respiration for hyphal growth. This is
consistent with findings which show aerobic respiration to be important for the
hyphal growth of many fungal species (Seto and Tazaki 1975; Lew and Levina 2004;
Solaiman and Saito 1997; Watanabe et al. 2006).
119
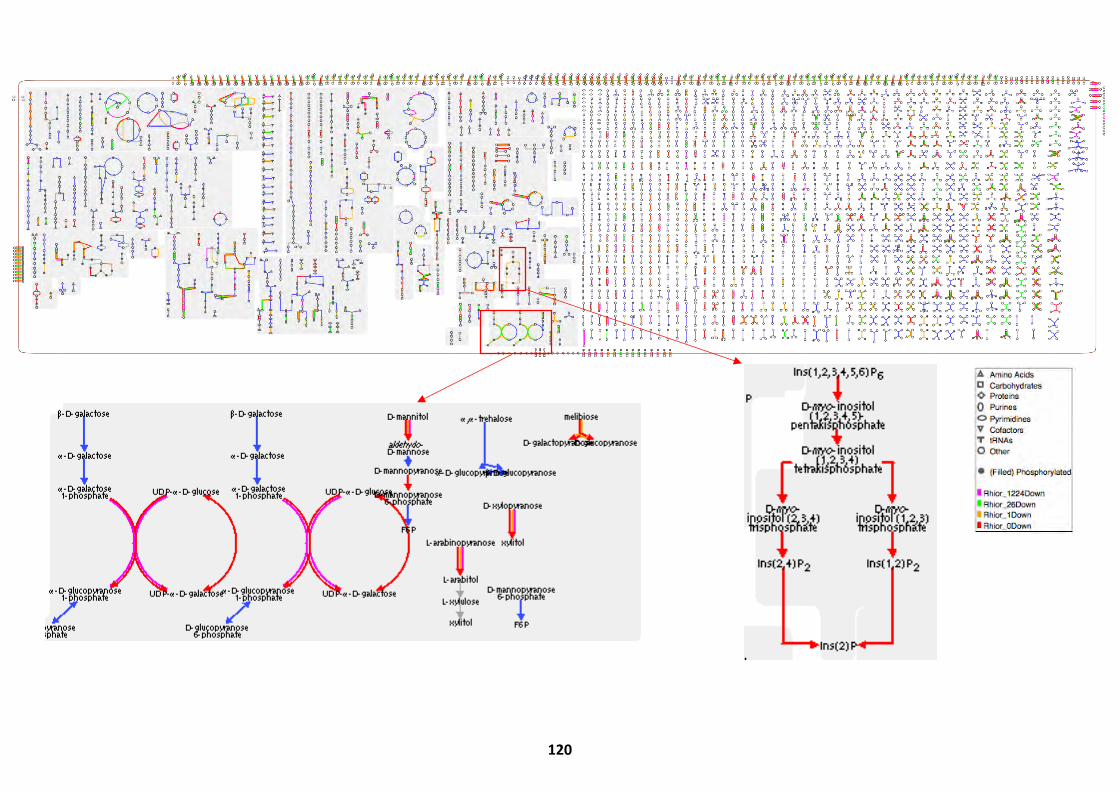
120

119
121
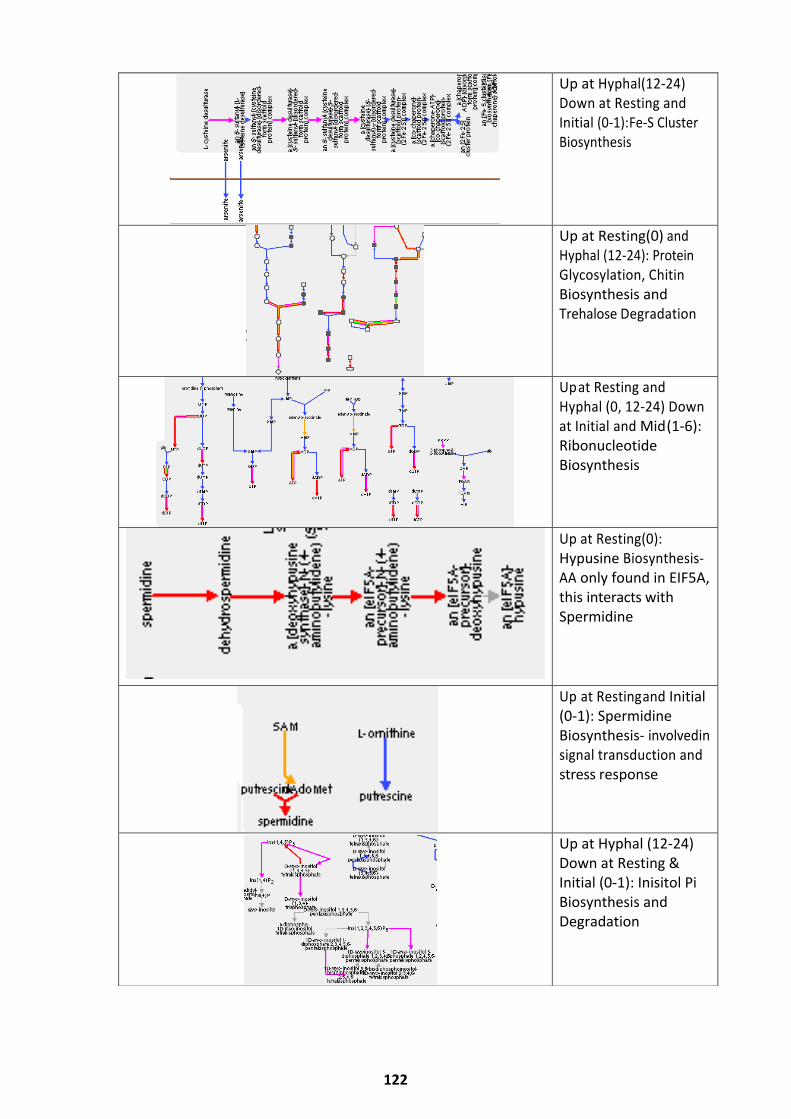
Up at Hyphal (12-24) Down at Resting and Initial (0-1): Fe-S Cluster Biosynthesis
Up at Resting(0) and Hyphal (12-24): Protein Glycosylation, Chitin Biosynthesis and Trehalose Degradation
Up at Resting and Hyphal (0, 12-24) Down at Initial and Mid (1-6): Ribonucleotide Biosynthesis
Up at Resting (0): Hypusine Biosynthesis- AA only found in EIF5A, this interacts with Spermidine
Up at Resting and Initial (0-1): Spermidine Biosynthesis- involved in signal transduction and stress response
Up at Hyphal (12-24) Down at Resting & Initial (0-1): Inisitol Pi Biosynthesis and Degradation
122
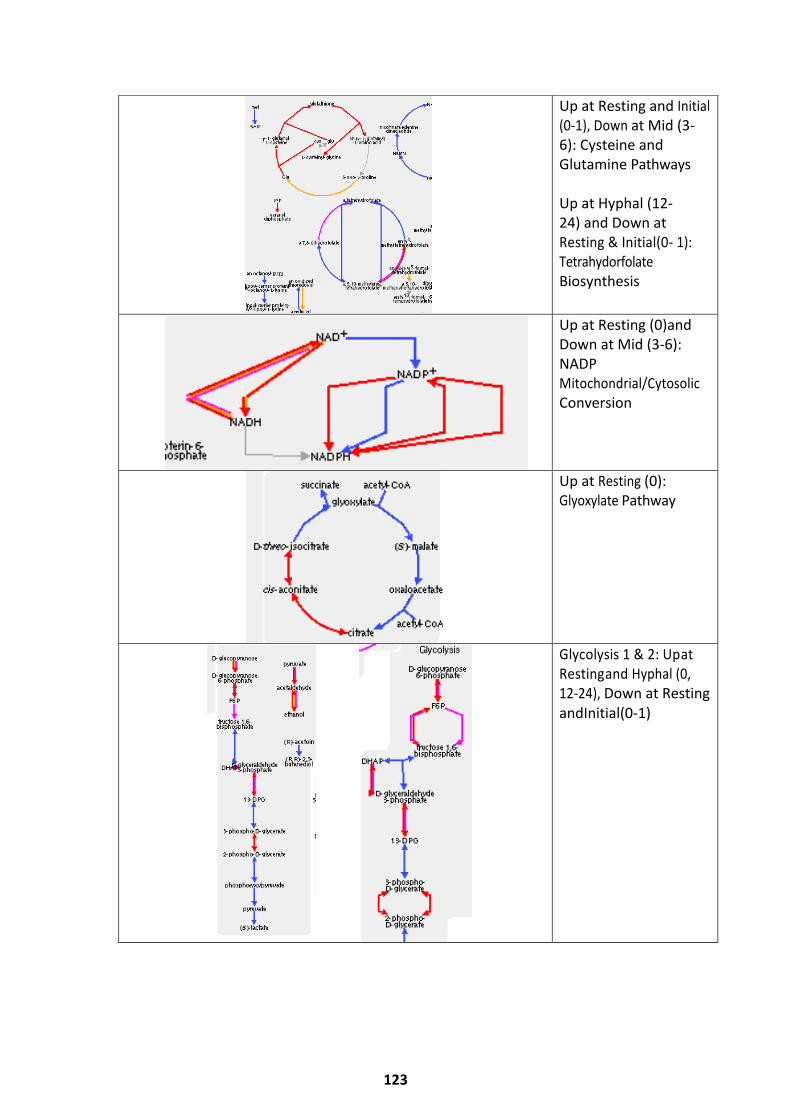
Up at Resting and Initial (0-1), Down at Mid (3-6): Cysteine and Glutamine Pathways
Up at Hyphal (12- 24) and Down atResting & Initial (0- 1):TetrahydorfolateBiosynthesis
Up at Resting (0) and Down at Mid (3-6): NADP Mitochondrial/Cytosolic Conversion
Up at Resting (0): Glyoxylate Pathway
Glycolysis 1 & 2: Up at Resting and Hyphal (0, 12-24), Down at Restingand Initial (0-1)
123

• Down at Mid (3-6): Cytochrome C, ATPase and NADPH Oxidoreductase• Up at Hyphal (12-24): CoA Ligase• Up at Hyphal (12-24): ATPase• Up at Resting (0) and Down Mid (3-6): Myrosinase Production• Down at Resting (0): Monoxygenase• Down at Initial (1) & Mid (3-6): Glucoronosyl transferase : UDP generation• Up at Hyphal (12-24), Down at Mid (3-6): Phosphatidylcholine transferase
Figure 8. Pathways differentially regulated over the course of germination. Differential
pathway expression was determined by highlighting pathways which included genes
significantly differentially expressed (multiple comparisons corrected P value of < 0.05)
between time points. Genes with increased expression (LogFC > 2) and decreased
expression (LogFC < -2) were selected for this analysis. Up and down regulated pathways
were highlighted using the custom R. delemar pathologic annotation and PathwayTools.
Up at Hyphal (12-24): Oxoglutarate and Isovalerate decarboxylation (CoA Regeneration)
Up at Hyphal (12-24): Acetyl CoA Ligase and Synthase
Down at Resting (0): Xyoglucan and Chitin Degradation
Up at Hyphal (12-24), Down at Resting and Mid (0, 3-6): Peroxisome
125
124

Co-transcriptional Networks
To determine whether phenotypes expressed a core set of genes, co-transcriptional network
analysis was performed. In total, resting spores and those germinated for up to two hours
shared expression of 4,738 transcripts. These transcripts were enriched for processes which
include cellular and primary metabolism, ribosome biogenesis, ncRNA processes, RNA
metabolism and protein metabolism (Figure 9). This core set may be key to initiating cellular
metabolism and protein processing which support the rapid morphological changes which
occur at germination initiation. Swelling spores (3-6h) co-expressed only 109 transcripts
(Figure 10), with roles in metabolism, RNA, tRNA and nucleotide processing and protein
processing. Though much smaller, this core set appears similar in function to the one
identified within 0-2h resting and swelling spores.
R. delemar in its hyphal form (12-24h) shares 1677 co-expressed transcripts. These had roles
in metabolism, tRNA, RNA and nucleotide processing, protein processing and macromolecule
processing (Figure 11). Again, the core set identified here appears highly similar in
functionality to the core sets for resting and swelling spores. This analysis suggests there is a
core set of transcripts expressed to maintain function of R. delemar. As all transcripts included
in this analysis were identified to be significantly differentially expressed across the entire
course of germination (and therefore not constituently expressed for the entirety of
germination), it may be possible that alternate genes with similar functions are expressed
throughout the germination of R. delemar. This mechanism would allow for regulation of
these processes alternately at different morphological stages.
125

Figure 9. Co-transcriptional network of genes (grey) and network enrichment visualisation
(yellow) expressed between 0 and 2 hours post germination. Yellow nodes represent
enriched predicted functions (GO term, P < 0.05), reduced to specific terms with ReviGO,
listed on the left.
Figure 10. Co-transcriptional network of genes (grey) and network enrichment visualisation
(yellow) expressed between 3 and 6 hours post germination. Yellow nodes represent
enriched predicted functions (GO term, P < 0.05), reduced to specific terms with ReviGO,
listed on the left.
109 Nodes
126

Figure 11. Co-transcriptional network of genes (grey) and network enrichment visualisation (yellow)
expressed between 12 and 24 hours post germination. Yellow nodes represent enriched
predicted functions (GO term, P < 0.05), reduced to specific terms with ReviGO, listed on the left.
As the genome of R. delemar is likely to have undergone a whole genome duplication event
(Ma et al. 2009), duplicate gene pairs were identified (Ma et al. 2009) and the expression
profiles were analysed to identify pairs which displayed alternate expression profiles over
the course of germination. As these duplicates have been maintained within the
genome, it was hypothesised that some duplicates may have diverged in function.
Evidence of divergence may be visible via alternate expression profiles of the
duplicates. In total 70 pairs, which were differentially expressed over the course of
germination to a significant level (FDR < 0.001), were identified to have alternate
expression profiles (LogFC between each gene within the pair > 2) (Figure 12). Alternate
expression profiles support the hypothesis that these duplicated genes may have diverged in
function or process regulation. Of these genes, pairs with the most highly divergent
expression patterns over the course of germination had the following predicted
functions: Glycosyltransferase, Nucleoporin, SMR domain, Ribosomal protein S2,
Protein Kinase, Metallopeptidase, Phosphatidyl inositol 4-phosphate-5-kinase and a
conserved fungal protein of unknown function (Figure 13).
127
Duplicated Gene Pair Expression
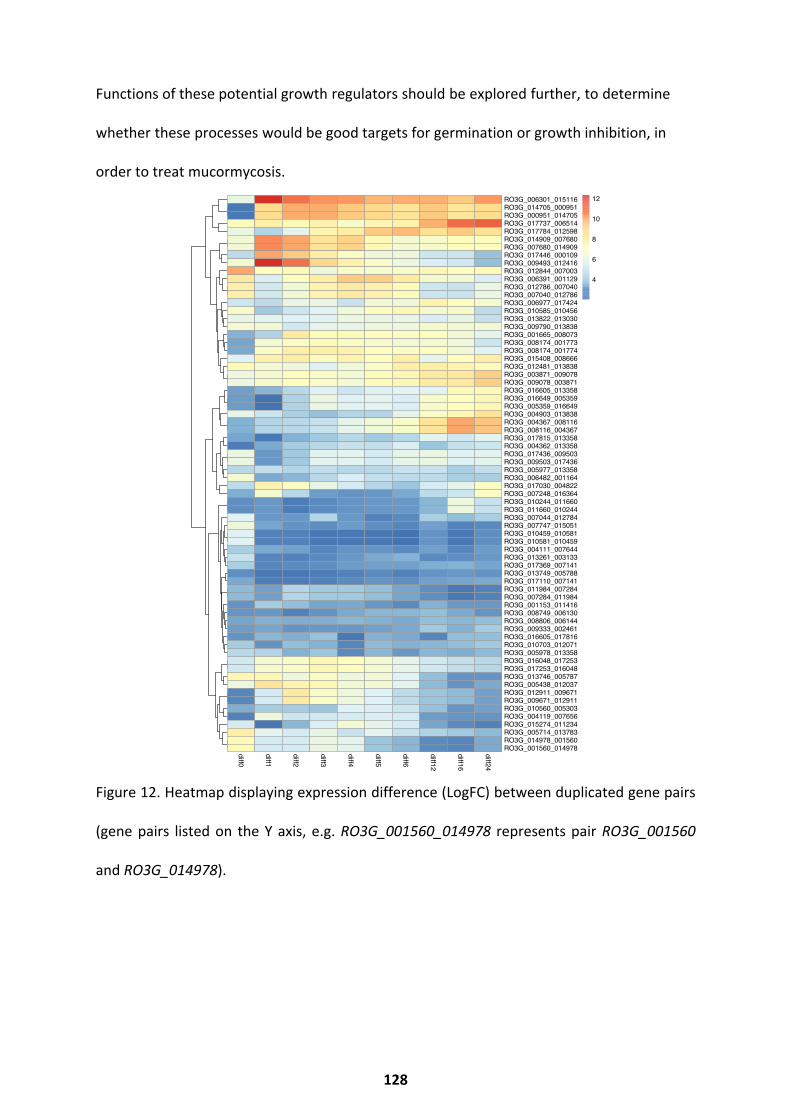
Functions of these potential growth regulators should be explored further, to determine
whether these processes would be good targets for germination or growth inhibition, in
order to treat mucormycosis.
Figure 12. Heatmap displaying expression difference (LogFC) between duplicated gene pairs
(gene pairs listed on the Y axis, e.g. RO3G_001560_014978 represents pair RO3G_001560
and RO3G_014978).
diff0
diff1
diff2
diff3
diff4
diff5
diff6
diff12
diff16
diff24
RO3G_006301_015116RO3G_014705_000951RO3G_000951_014705RO3G_017737_006514RO3G_017784_012598RO3G_014909_007680RO3G_007680_014909RO3G_017446_000109RO3G_009493_012416RO3G_012844_007003RO3G_006391_001129RO3G_012786_007040RO3G_007040_012786RO3G_006977_017424RO3G_010585_010456RO3G_013822_013030RO3G_009790_013838RO3G_001665_008073RO3G_008174_001773RO3G_008174_001774RO3G_015408_008666RO3G_012481_013838RO3G_003871_009078RO3G_009078_003871RO3G_016605_013358RO3G_016649_005359RO3G_005359_016649RO3G_004903_013838RO3G_004367_008116RO3G_008116_004367RO3G_017815_013358RO3G_004362_013358RO3G_017436_009503RO3G_009503_017436RO3G_005977_013358RO3G_006482_001164RO3G_017030_004822RO3G_007248_016364RO3G_010244_011660RO3G_011660_010244RO3G_007044_012784RO3G_007747_015051RO3G_010459_010581RO3G_010581_010459RO3G_004111_007644RO3G_013261_003133RO3G_017369_007141RO3G_013749_005788RO3G_017110_007141RO3G_011984_007284RO3G_007284_011984RO3G_001153_011416RO3G_008749_006130RO3G_008806_006144RO3G_009333_002461RO3G_016605_017816RO3G_010703_012071RO3G_005978_013358RO3G_016048_017253RO3G_017253_016048RO3G_013746_005787RO3G_005438_012037RO3G_012911_009671RO3G_009671_012911RO3G_010560_005303RO3G_004119_007656RO3G_015274_011234RO3G_005714_013783RO3G_014978_001560RO3G_001560_014978
4
6
8
10
12
128
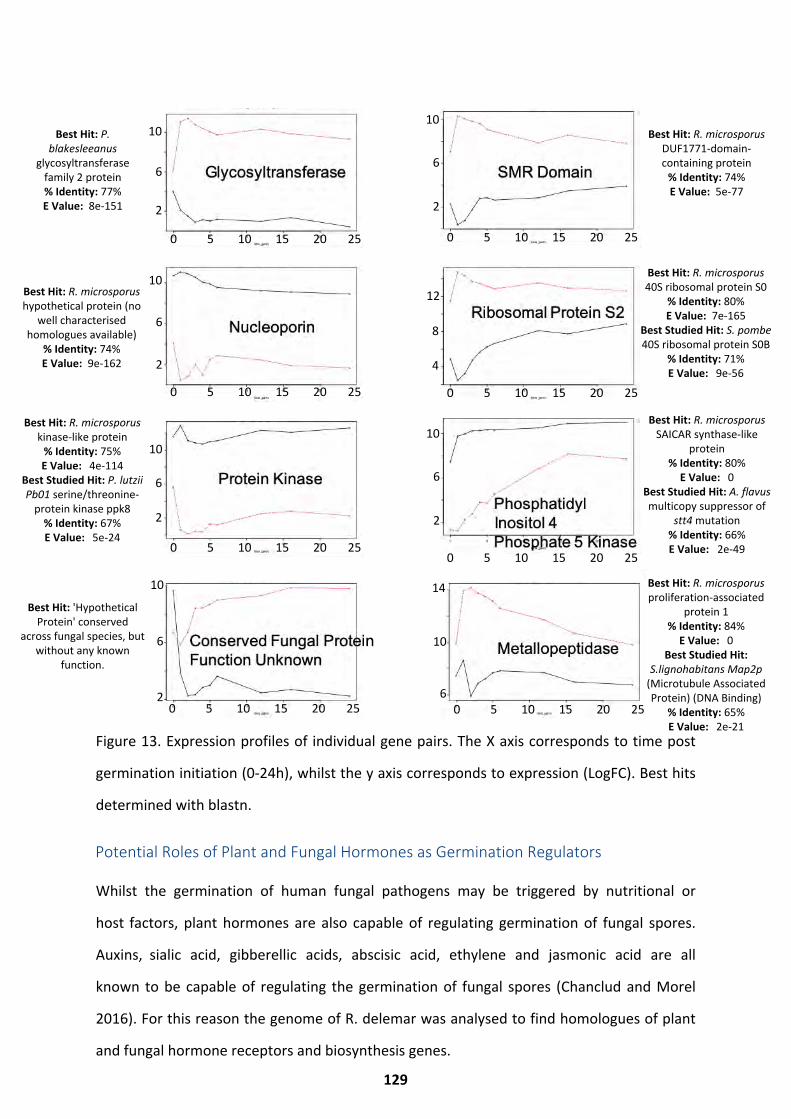
Best Hit: P. blakesleeanus
glycosyltransferase family 2 protein% Identity: 77%E Value: 8e-151
Best Hit: R. microsporus DUF1771-domain-containing protein
% Identity: 74%E Value: 5e-77
Best Hit: R. microsporus hypothetical protein (no
well characterised homologues available)
% Identity: 74%E Value: 9e-162
Best Hit: R. microsporus 40S ribosomal protein S0
% Identity: 80%E Value: 7e-165
Best Studied Hit: S. pombe 40S ribosomal protein S0B
% Identity: 71%E Value: 9e-56
Best Hit: R. microsporus kinase-like protein
% Identity: 75%E Value: 4e-114
Best Studied Hit: P. lutzii Pb01 serine/threonine-
protein kinase ppk8% Identity: 67%E Value: 5e-24
Best Hit: R. microsporus SAICAR synthase-like
protein% Identity: 80%
E Value: 0Best Studied Hit: A. flavus multicopy suppressor of
stt4 mutation% Identity: 66%E Value: 2e-49
Best Hit: 'Hypothetical Protein' conserved
across fungal species, but without any known
function.
Best Hit: R. microsporus proliferation-associated
protein 1% Identity: 84%
E Value: 0Best Studied Hit:
S.lignohabitans Map2p(Microtubule AssociatedProtein) (DNA Binding)
% Identity: 65%E Value: 2e-21
Figure 13. Expression profiles of individual gene pairs. The X axis corresponds to time post
germination initiation (0-24h), whilst the y axis corresponds to expression (LogFC). Best hits
determined with blastn.
Potential Roles of Plant and Fungal Hormones as Germination Regulators
Whilst the germination of human fungal pathogens may be triggered by nutritional or
host factors, plant hormones are also capable of regulating germination of fungal spores.
Auxins, sialic acid, gibberellic acids, abscisic acid, ethylene and jasmonic acid are all
known to be capable of regulating the germination of fungal spores (Chanclud and Morel
2016). For this reason the genome of R. delemar was analysed to find homologues of plant
and fungal hormone receptors and biosynthesis genes.
129
0 5 10 15 20 25 0 5 10 15 20 25
0 5 10 15 20 25 0 5 10 15 20 25
0 5 10 15 20 25 0 5 10 15 20 25
0 5 10 15 20 25 0 5 10 15 20 25
2
10
6
2
10
6
2
10
6
4
12
8
2
10
6
2
10
6
2
10
6
6
14
10
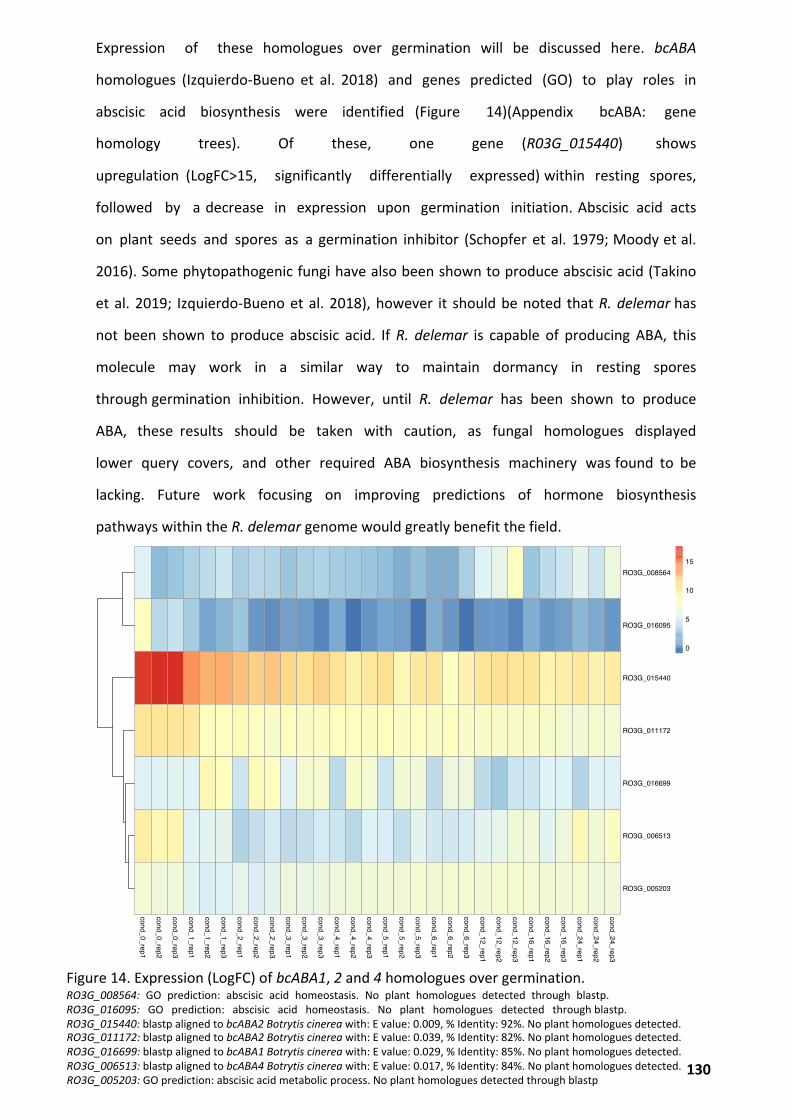
Expression of these homologues over germination will be discussed here. bcABA
homologues (Izquierdo-Bueno et al. 2018) and genes predicted (GO) to play roles in
abscisic acid biosynthesis were identified (Figure 14)(Appendix bcABA: gene
homology trees). Of these, one gene (R03G_015440) shows
upregulation (LogFC>15, significantly differentially expressed) within resting spores,
followed by a decrease in expression upon germination initiation. Abscisic acid acts
on plant seeds and spores as a germination inhibitor (Schopfer et al. 1979; Moody et al.
2016). Some phytopathogenic fungi have also been shown to produce abscisic acid (Takino
et al. 2019; Izquierdo-Bueno et al. 2018), however it should be noted that R. delemar has
not been shown to produce abscisic acid. If R. delemar is capable of producing ABA, this
molecule may work in a similar way to maintain dormancy in resting spores
through germination inhibition. However, until R. delemar has been shown to produce
ABA, these results should be taken with caution, as fungal homologues displayed
lower query covers, and other required ABA biosynthesis machinery was found to be
lacking. Future work focusing on improving predictions of hormone biosynthesis
pathways within the R. delemar genome would greatly benefit the field.
Figure 14. Expression (LogFC) of bcABA1, 2 and 4 homologues over germination. RO3G_008564: GO prediction: abscisic acid homeostasis. No plant homologues detected through blastp. RO3G_016095: GO prediction: abscisic acid homeostasis. No plant homologues detected through blastp. RO3G_015440: blastp aligned to bcABA2 Botrytis cinerea with: E value: 0.009, % Identity: 92%. No plant homologues detected.RO3G_011172: blastp aligned to bcABA2 Botrytis cinerea with: E value: 0.039, % Identity: 82%. No plant homologues detected.RO3G_016699: blastp aligned to bcABA1 Botrytis cinerea with: E value: 0.029, % Identity: 85%. No plant homologues detected.RO3G_006513: blastp aligned to bcABA4 Botrytis cinerea with: E value: 0.017, % Identity: 84%. No plant homologues detected.RO3G_005203: GO prediction: abscisic acid metabolic process. No plant homologues detected through blastp
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_008564
RO3G_016095
RO3G_015440
RO3G_011172
RO3G_016699
RO3G_006513
RO3G_005203
0
5
10
15
130
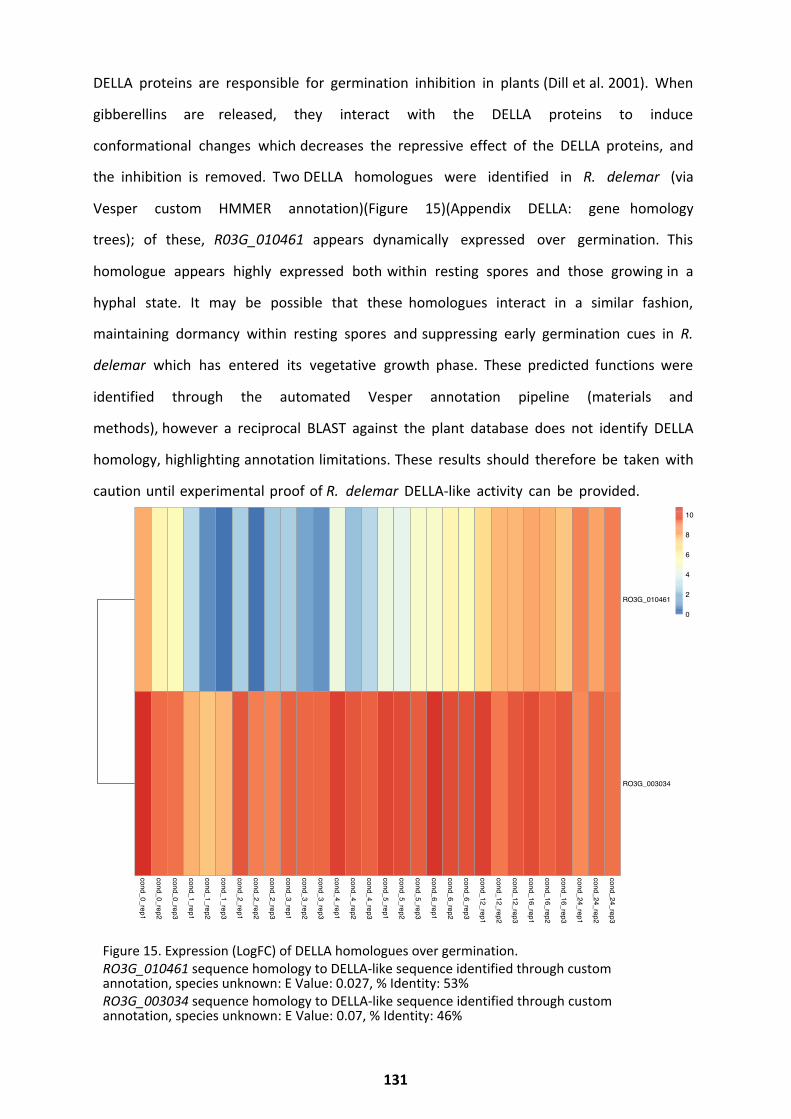
DELLA proteins are responsible for germination inhibition in plants (Dill et al. 2001). When
gibberellins are released, they interact with the DELLA proteins to induce
conformational changes which decreases the repressive effect of the DELLA proteins, and
the inhibition is removed. Two DELLA homologues were identified in R. delemar (via
Vesper custom HMMER annotation)(Figure 15)(Appendix DELLA: gene homology
trees); of these, R03G_010461 appears dynamically expressed over germination. This
homologue appears highly expressed both within resting spores and those growing in a
hyphal state. It may be possible that these homologues interact in a similar fashion,
maintaining dormancy within resting spores and suppressing early germination cues in R.
delemar which has entered its vegetative growth phase. These predicted functions were
identified through the automated Vesper annotation pipeline (materials and
methods), however a reciprocal BLAST against the plant database does not identify DELLA
homology, highlighting annotation limitations. These results should therefore be taken with
caution until experimental proof of R. delemar DELLA-like activity can be provided.
Figure 15. Expression (LogFC) of DELLA homologues over germination. RO3G_010461 sequence homology to DELLA-like sequence identified through custom annotation, species unknown: E Value: 0.027, % Identity: 53%RO3G_003034 sequence homology to DELLA-like sequence identified through custom annotation, species unknown: E Value: 0.07, % Identity: 46%
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_010461
RO3G_003034
0
2
4
6
8
10
131
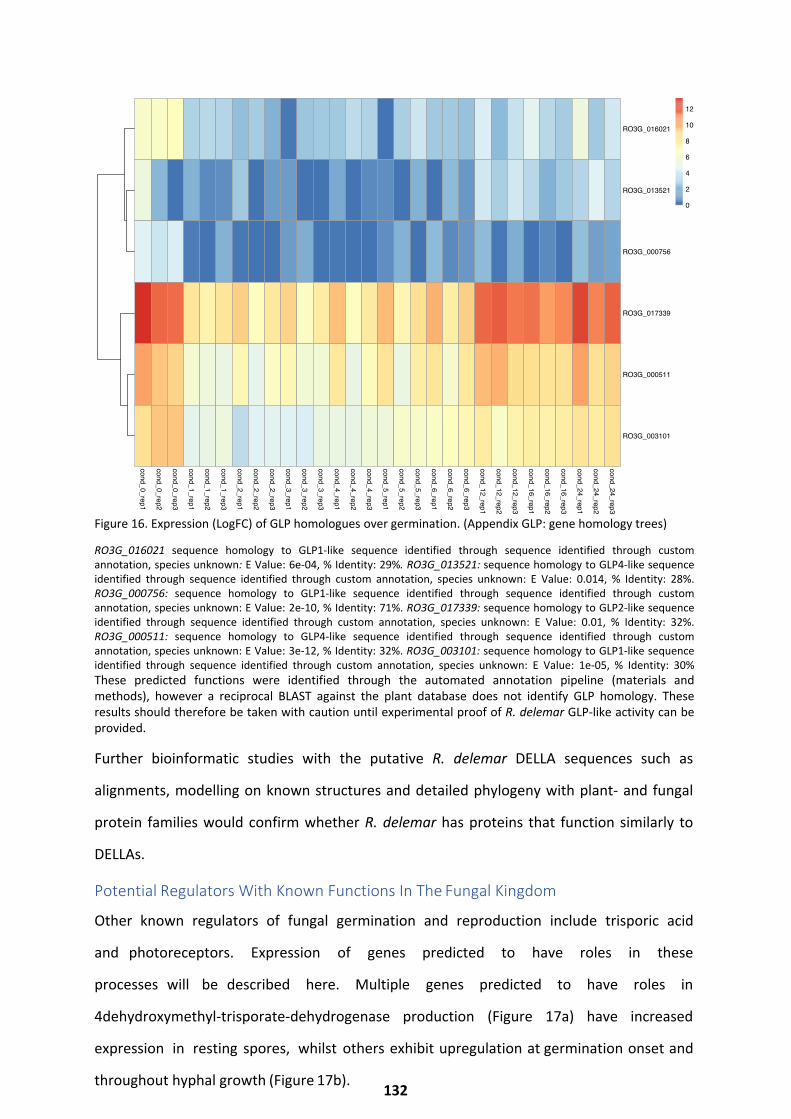
Figure 16. Expression (LogFC) of GLP homologues over germination. (Appendix GLP: gene homology trees)
RO3G_016021 sequence homology to GLP1-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 6e-04, % Identity: 29%. RO3G_013521: sequence homology to GLP4-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 0.014, % Identity: 28%. RO3G_000756: sequence homology to GLP1-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 2e-10, % Identity: 71%. RO3G_017339: sequence homology to GLP2-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 0.01, % Identity: 32%. RO3G_000511: sequence homology to GLP4-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 3e-12, % Identity: 32%. RO3G_003101: sequence homology to GLP1-like sequence identified through sequence identified through custom annotation, species unknown: E Value: 1e-05, % Identity: 30% These predicted functions were identified through the automated annotation pipeline (materials and methods), however a reciprocal BLAST against the plant database does not identify GLP homology. These results should therefore be taken with caution until experimental proof of R. delemar GLP-like activity can be provided.
Further bioinformatic studies with the putative R. delemar DELLA sequences such as
alignments, modelling on known structures and detailed phylogeny with plant- and fungal
protein families would confirm whether R. delemar has proteins that function similarly to
DELLAs.
Potential Regulators With Known Functions In The Fungal Kingdom
Other known regulators of fungal germination and reproduction include trisporic acid
and photoreceptors. Expression of genes predicted to have roles in these
processes will be described here. Multiple genes predicted to have roles in
4dehydroxymethyl-trisporate-dehydrogenase production (Figure 17a) have increased
expression in resting spores, whilst others exhibit upregulation at germination onset and
throughout hyphal growth (Figure 17b).
cond_0_rep1
cond_0_rep2
cond_0_rep3
cond_1_rep1
cond_1_rep2
cond_1_rep3
cond_2_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_4_rep1
cond_4_rep2
cond_4_rep3
cond_5_rep1
cond_5_rep2
cond_5_rep3
cond_6_rep1
cond_6_rep2
cond_6_rep3
cond_12_rep1
cond_12_rep2
cond_12_rep3
cond_16_rep1
cond_16_rep2
cond_16_rep3
cond_24_rep1
cond_24_rep2
cond_24_rep3
RO3G_016021
RO3G_013521
RO3G_000756
RO3G_017339
RO3G_000511
RO3G_003101
0
2
4
6
8
10
12
132
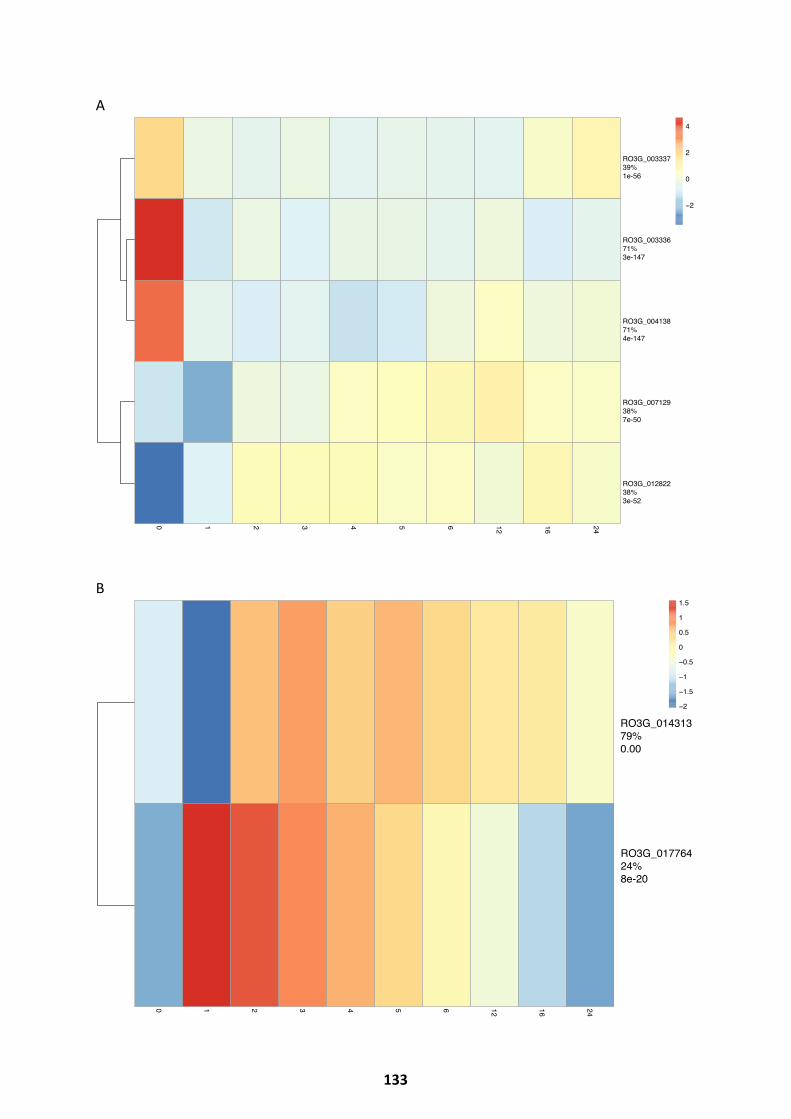
A
B
0 1 2 3 4 5 6 12 16 24
RO3G_00333739%1e-56
RO3G_00333671%3e-147
RO3G_00413871%4e-147
RO3G_00712938%7e-50
RO3G_01282238%3e-52
−2
0
2
4
0 1 2 3 4 5 6 12 16 24
RO3G_01431379%0.00
RO3G_01776424%8e-20
−2
−1.5
−1
−0.5
0
0.5
1
1.5
133
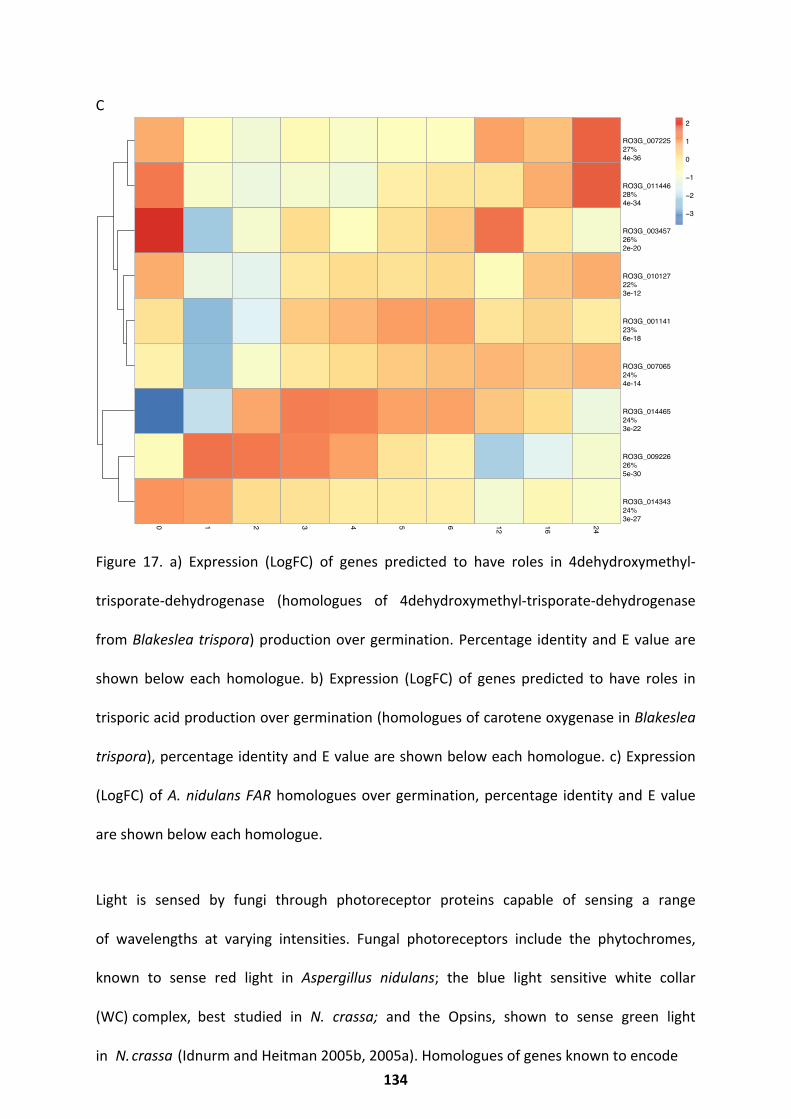
C
Figure 17. a) Expression (LogFC) of genes predicted to have roles in 4dehydroxymethyl-
trisporate-dehydrogenase (homologues of 4dehydroxymethyl-trisporate-dehydrogenase
from Blakeslea trispora) production over germination. Percentage identity and E value are
shown below each homologue. b) Expression (LogFC) of genes predicted to have roles in
trisporic acid production over germination (homologues of carotene oxygenase in Blakeslea
trispora), percentage identity and E value are shown below each homologue. c) Expression
(LogFC) of A. nidulans FAR homologues over germination, percentage identity and E value
are shown below each homologue.
Light is sensed by fungi through photoreceptor proteins capable of sensing a range
of wavelengths at varying intensities. Fungal photoreceptors include the phytochromes,
known to sense red light in Aspergillus nidulans; the blue light sensitive white collar
(WC) complex, best studied in N. crassa; and the Opsins, shown to sense green light
in N. crassa (Idnurm and Heitman 2005b, 2005a). Homologues of genes known to encode
0 1 2 3 4 5 6 12 16 24
RO3G_00722527%4e-36
RO3G_01144628%4e-34
RO3G_00345726%2e-20
RO3G_01012722%3e-12
RO3G_00114123%6e-18
RO3G_00706524%4e-14
RO3G_01446524%3e-22
RO3G_00922626%5e-30
RO3G_01434324%3e-27
−3
−2
−1
0
1
2
134
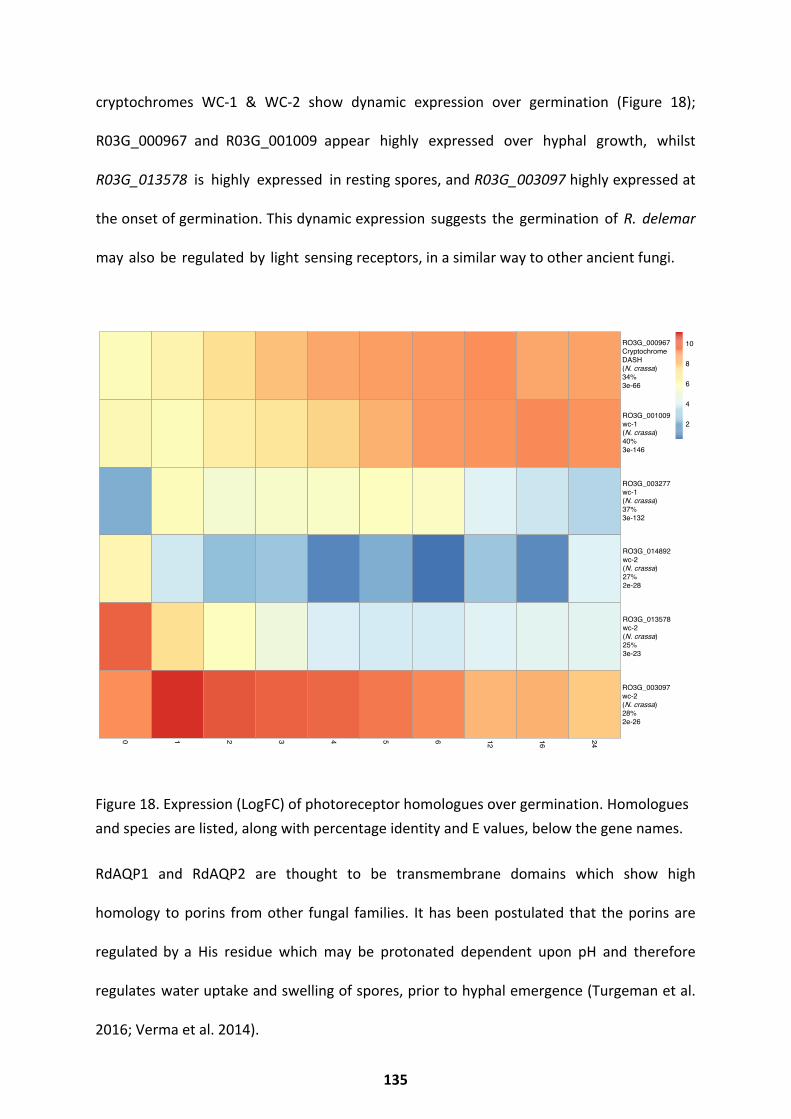
cryptochromes WC-1 & WC-2 show dynamic expression over germination (Figure 18);
R03G_000967 and R03G_001009 appear highly expressed over hyphal growth, whilst
R03G_013578 is highly expressed in resting spores, and R03G_003097 highly expressed at
the onset of germination. This dynamic expression suggests the germination of R. delemar
may also be regulated by light sensing receptors, in a similar way to other ancient fungi.
Figure 18. Expression (LogFC) of photoreceptor homologues over germination. Homologues and species are listed, along with percentage identity and E values, below the gene names.
RdAQP1 and RdAQP2 are thought to be transmembrane domains which show high
homology to porins from other fungal families. It has been postulated that the porins are
regulated by a His residue which may be protonated dependent upon pH and therefore
regulates water uptake and swelling of spores, prior to hyphal emergence (Turgeman et al.
2016; Verma et al. 2014).
0 1 2 3 4 5 6 12 16 24
RO3G_000967Cryptochrome DASH (N. crassa) 34%3e-66
RO3G_001009wc-1(N. crassa) 40%3e-146
RO3G_003277wc-1(N. crassa)37%3e-132
RO3G_014892wc-2(N. crassa)27%2e-28
RO3G_013578wc-2(N. crassa)25%3e-23
RO3G_003097wc-2(N. crassa)28%2e-26
2
4
6
8
10
135
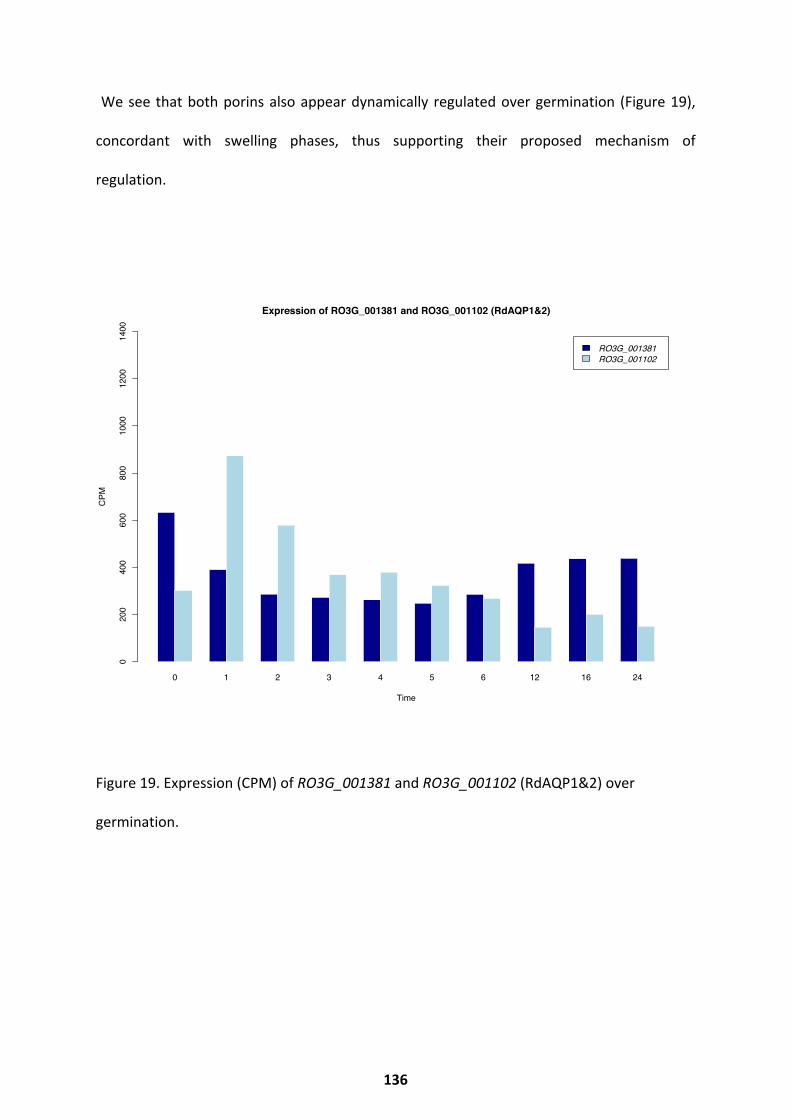
We see that both porins also appear dynamically regulated over germination (Figure 19),
concordant with swelling phases, thus supporting their proposed mechanism of
regulation.
Figure 19. Expression (CPM) of RO3G_001381 and RO3G_001102 (RdAQP1&2) over
germination.
0 1 2 3 4 5 6 12 16 24
RO3G_001381RO3G_001102
Expression of RO3G_001381 and RO3G_001102 (RdAQP1&2)
Time
CPM
0200
400
600
800
1000
1200
1400
136

Comparisons of Transcription Throughout Germination
It is unclear whether the mechanisms that underpin germination are conserved throughout
the diverse fungal kingdom. To explore the extent of conservation, I compared my
transcriptional data set to other available transcriptional data sets for Aspergillus niger,
generated by Van Leeuwen et al and Novodvorska et al (van Leeuwen et al. 2013;
Novodvorska et al. 2013). When expression profiles of homologous genes from A. niger and
R. delemar are compared over the course of germination, genes with common or unique
functions specific to that time point can be identified. The largest shift in the transcriptional
landscape of A. niger can be seen at the initial stage of germination; we also observed this
shift in R. delemar. Transcripts with predicted functions involved in transport and localization,
proteolysis, and glucose, hexose, and carbohydrate metabolism increase at the initial stages
of germination in both A. niger and R. delemar, while transcripts with predicted functions in
translation, tRNA and rRNA processing, and amine carboxylic acid and organic acid
metabolism decrease (Figure 20). We also observe differences between the two data sets:
over isotropic and hyphal growth, homologous genes with predicted functions in valine and
branched-chain amino acid metabolism were upregulated only in R. delemar, while
homologous genes with predicted roles in noncoding RNA (ncRNA) metabolism, translation,
amino acid activation, and ribosome biogenesis were downregulated exclusively in R.
delemar. A 5% increase in genes that are uniquely up- or downregulated in R. delemar is found
in high-synteny regions of the genome, compared to genes that are up or downregulated in
both R. delemar and A. niger. The duplicated nature of the R. delemar genome may allow for
specific and tight regulation of the germination process, a feature unique to R. delemar.
137

It should be noted that A. niger and R. delemar were cultivated under conditions with
different media. Aspergillus complete medium (ACM), used to cultivate A. niger, and
sabouraud dextrose broth (SAB), used to cultivate R. delemar, both contain a complex mix of
salts, inorganic nutrients, and organic components. Peptides are provided in SAB by
mycological peptone, whereas peptides are provided by bacto peptone in ACM. The main
carbon source is the same for both ACM and SAB. Both media have a relatively low pH (ACM,
pH 6.5; SAB, pH 5.6), and it is known that pH is important for regulating germination in both
R. delemar and A. nidulans. There are currently limited studies that address differences in
gene expression, when germination is initiated in filamentous fungi, under different growth
media. Growth characteristics of A. nidulans have been shown to vary when contents of
media differ, while various growth cultivation methods also alter gene expression in
Aspergillus oryzae. The effect of adding or removing specific organic and inorganic nutrients
from media on the growth of filamentous fungi is also better understood. When comparing
data sets or designing experiments to address these issues, the effects of using distinctly
different media should be considered. This is an area that would benefit from further work
aimed at exploring these effects.
138
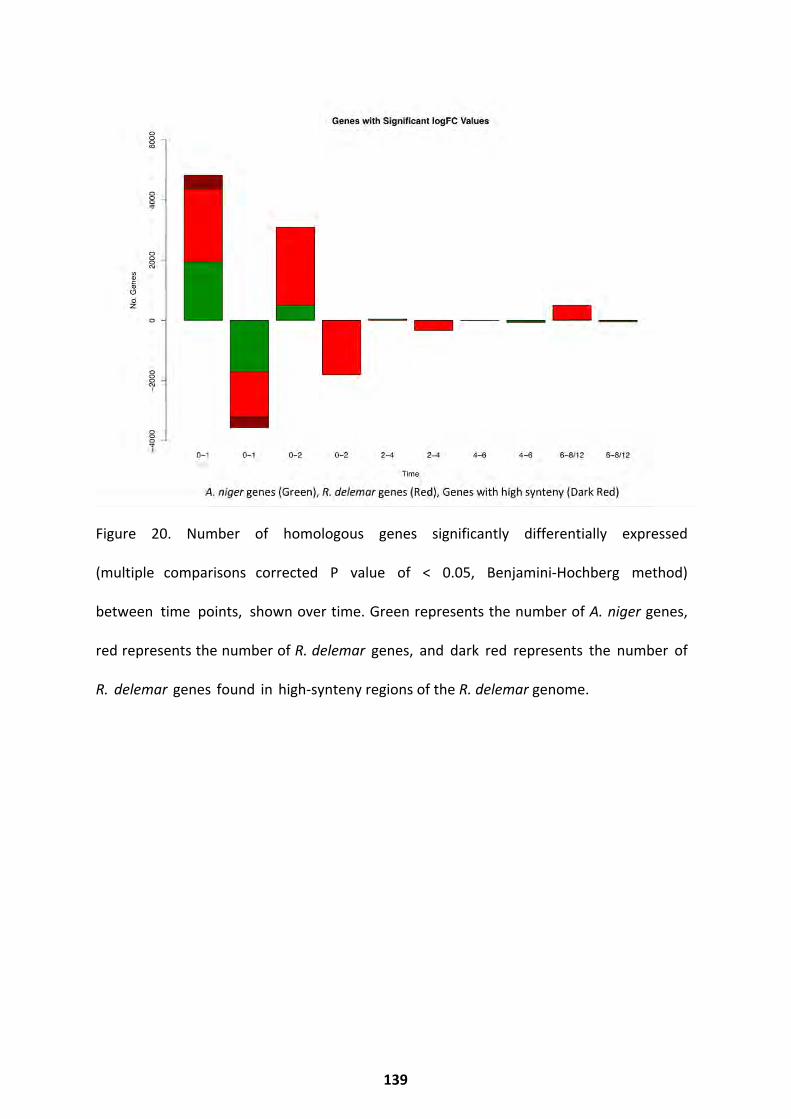
Figure 20. Number of homologous genes significantly differentially expressed
(multiple comparisons corrected P value of < 0.05, Benjamini-Hochberg method)
between time points, shown over time. Green represents the number of A. niger genes,
red represents the number of R. delemar genes, and dark red represents the number of
R. delemar genes found in high-synteny regions of the R. delemar genome.
139

Conclusions
This transcriptional analysis reveals that a temporal transcriptional pattern governs
germination of R. delemar spores (Figure 1). Phenotype-specific (resting spore, swelling
spore, hyphal fungi) transcript clusters (Figure 2-5) are identified, and along with pairwise
differential expression analysis (Figure 6), it is shown that resting spores appear
transcriptionally unique. In comparison, swelling spores appear transcriptionally consistent,
whilst the switch to hyphal growth is underpinned by a significant change in the
transcriptional landscape. Paired with this, analysis of metabolic transcripts (Figure 7),
predicted pathways (Figure 8) and co-transcriptional networks (Figure 9-11) reveal a
reliance of germinating spores on ROS resistance, cell wall remodelling components, protein
synthesis apparatus, and iron acquisition and transport components. Analysis of duplicated
gene pairs highlight individual genes which may be important for germination control.
Homology searching reveals the dynamic regulation of germination and growth
regulators, with homologues in well-studied fungi and plants. Finally, a comparison of
germinating A. niger and R. delemar transcription demonstrates the presence of a
‘core set’ of genes conserved throughout germination, however genes unique to
each species are also abundant.
Discussion
The above results increase our understanding of the molecular mechanisms
controlling germination in R. delemar. They show that the initiation of germination
entails a huge transcriptional shift; ROS resistance and respiration are required for
germination to occur, while actin, chitin, and cytoskeletal components appear to play key
140

roles in initiating isotropic swelling and hyphal growth. Iron acquisition, Fe-S cluster
regulation and sulphur metabolism were also key features of the germination transcriptome,
confirming that R. delemar relies upon iron not only for pathogenesis (Ibrahim et al. 2010),
but also for germination (Kousser et al. 2019). Transcripts co-expressed at the defined
stages of germination (and unique to these stages) share functions in protein synthesis.
This may offer regulation of protein processes, whilst maintaining specificity to
morphological stage. R. delemar shares many transcriptional traits with A. niger at
germination initiation; however, transcriptional features unique to R. delemar indicate
that the duplicated nature of the genome may allow for alternative regulation of this
process. Transcriptional results demonstrate the potential of light, fungal hormones and
plant hormones to regulate germination. Specifically, resting spores show increased
expression of bcABA2 homologues (Figure 14), which may inhibit fungal
germination, similarly to P. patens spore germination inhibition via ABA (Moody et al. 2016).
It should be noted that the custom annotation which predicted DELLA-like and GLP-like
activity did not provide specific species homologues. As plant homologues were not detected
through a traditional blastP approach, these results should be taken with caution. The
hyphal form shows increased expression of both cryptochrome and white
collar homologues, indicating that light sensing may play a role in germination
regulation and hyphal development, as it does in A. fumigatus and the phototrophic P.
blakesleeanus (Fuller et al. 2013; Idnurm et al. 2005). It should be noted that other
homologues of the genes discussed here may exist within the R. delemar genome
(Appendix Homology Trees), however only those which were differentially expressed
(FDR < 0.001) over all time points have been investigated here. As a result, constituent
expression is not discussed here, but should be investigated in further studies. These
results have provided a significant overview of the transcriptome of germinating spores
and expanded current knowledge in the Mucorales field.
141

Further Work
Further work focusing on the elucidation of genes essential to germination would benefit the
field. Whilst co-expression network analysis aimed to reveal these, the networks (Figure
9-11) were ultimately too large to provide a practical list of candidate genes. To determine
germination essential genes, transcriptional studies focusing on the times of phenotypic
change (0-2hr, 4-6hr, 6-12hr), sampling at smaller intervals (every 10 minutes), would
likely provide better information about germination specific genes. Further analysis of
duplicated gene pairs might also aid in the identification of germination specific
genes. Producing knockout mutants of the genes proposed here (Figure 13), which
could be screened for germination defects, would confirm whether these candidate
genes have diverged in function, and whether they play specific roles in germination. A full
transcription factor annotation of the R. delemar genome, combined with an integrated ChIP-
Seq RNA-Seq experimental approach, would also aid in determining germination regulation.
This approach would generate germination-essential gene candidates via analysis of
differentially regulated gene networks. Generation of a CRISPR-Cas9 knock out library in
R. delemar might also identify germination specific genes. Germination specific genes
would be attractive drug targets, as mucormycete germination within the host contributes
to pathogenesis. Use of a metabolomics approach would aid in the validation of
the PathwayTools pathway predictions, and may result in identification of natural
compounds with therapeutic activities, as is common for secondary metabolites of fungi.
142

Chapter 5: Transcriptional Regulation of Rhizopus-Macrophage Interactions
The following text and figures have been previously published, or are adapted from the article
“Host-pathogen transcriptomics of macrophages, Mucorales and their endosymbionts: a
polymicrobial pas de trois” (Sephton-Clark et al. 2019). For this article, I conceived, designed
and performed experiments, collected the data, performed the analysis and interpretation,
wrote the manuscript, completed revisions and prepared the figures.
This chapter will introduce the host-pathogen interactions which govern mucormycosis and
immunity. An introduction to how bacterial endosymbionts impact pathogenicity of
mucorales species will also be given. Results will describe the differential interactions of two
Mucorales species (Rhizopus delemar RA 99-880 and the UoB strain of Rhizopus microsporus)
with the innate immune system, and their interactions with environmental bacterial
endosymbionts. The transcriptional responses of R. delemar and R. microsporus to innate
immune cells, and the corresponding immune cell response will be examined. The variation
in these responses will be assessed, given the presence or absence of bacterial
endosymbionts within the fungi. It can be seen that the fungal response is driven by
interaction with innate immune cells. The effect of the bacterial endosymbiont on the fungus
is species specific, with a minimal effect in the absence of stress, but strongly influencing
fungal expression during spore interactions with innate immune cells. In contrast, the
macrophage response varies depending on the infecting fungal species, but also depending
on endosymbiont status. The most successful macrophages elicit a pro-inflammatory
response, and through germination inhibition macrophage survival is enhanced. This reveals
143

species-specific host responses to related Mucorales species and shows that bacterial
endosymbionts impact the innate immune cell response.
144

Host-Pathogen Interactions in Mucormycosis
Mucormycosis has been studied in the context of mucorales-host interactions using model
organisms and in vitro tissue culture methods. Mouse models have shown that spore
interactions with endothelial cells are vital to causing diseases in immunocompromised
individuals, as pathogenicity is linked to the variable copy number of the gene encoding CotH,
a family of cell surface proteins important for the spores’ adherence to the GRP (glucose
regulated protein) receptors on endothelial cells (Liu et al. 2010; Gebremariam et al. 2014;
Watkins et al. 2018). Mouse studies have also revealed the reliance of mucorales spores on
free iron to cause invasive mucormycosis. The DKA ( Diabetic Ketoacidic) mouse model
is more susceptible to developing mucormycosis (Gebremariam et al. 2016). DKA and other
acidosis disorders result in pH-mediated dissociation of iron from protein
transporters, resulting in elevated concentrations of free iron within individuals. This is
clearly seen in patient data, as individuals with elevated iron concentrations (within
serum) are highly susceptible to mucormycosis (Ibrahim, 2014).
This reliance on iron is also seen in vitro. Iron limitation has been reported as a mechanism
by which macrophages control mucorales spore germination, whilst iron-scavenging
pathways have been reported to confer pathogenicity to Rhizopus spp. (Andrianaki et al.
2018; Chibucos et al. 2016). The reliance on innate immune cells such as macrophages to
control spores has been demonstrated in the established zebra fish model (Voelz et al.
2015). Within the zebra fish model, innate immune cells form granuloma- like structures
around infecting mucorales spores. In doing so they inhibit spore germination,
dissemination and delay invasive infection (Inglesfield et al. 2018). In an
immunocompromised zebra fish model,
145

these innate granulomas are less likely to form, and infecting spores rapidly disseminate,
leading to uncontrolled infection and increased mortality rates (Inglesfield et al. 2018).
The interaction between host and mucorales spore can be further regulated by the presence
of an endosymbiont (Itabangi et al. 2019; Partida-Martinez & Hertweck 2005). Many
Mucorales species have been shown to harbour bacterial endosymbionts whose species can
vary between Mucorales isolates (Ibrahim et al. 2008; Kobayashi & Crouch 2009; Mondo et
al. 2017; Itabangi et al. 2019). Itabangi et al. recently explored the effect of an endosymbiont
on the interaction between R. microsporus and innate immune cells. This study has shown
that a bacterial endosymbiont influences the outcome of Rhizopus microsporus infections in
both zebrafish and murine models, through modulation of both fungal and phagocyte
phenotypes (Itabangi et al. 2019).
It is clear that understanding the interaction between Mucorales spores, their endosymbionts
and innate immune cells is key to understanding mucormycosis. Studies frequently focus
upon the interaction of a single species of the Mucorales order with innate immune cells
(Warris et al. 2005; Chamilos et al. 2008; Schmidt et al. 2013; Kraibooj et al. 2014; Inglesfield
et al. 2018), despite numerous phenotypic and genomic differences existing between
Mucorales species and infecting isolates (Hoffmann et al. 2013; Mendoza et al. 2014;
Schwartze et al. 2014). Several works comparing Aspergillus spp. to Rhizopus spp. have
revealed similar immunostimulatory capacities, but differences in their responses to host
stress (Warris et al. 2005; Chamilos et al. 2008; Schmidt et al. 2013; Kraibooj et al. 2014).
Exploring and understanding fungal responses to the host is essential to improving our
understanding of mucormycosis, yet it remains unclear how Mucorales species respond to,
146

and interact with, the innate immune system, and to what extent this varies by species and
endosymbiont presence.
This work explores the interplay between phagocytes, R. delemar, and R. microsporus using
fungal isolates found to harbour bacterial endosymbionts (Itabangi et al. 2019). This work
investigates the differences between these two fungal species, how they respond
transcriptionally to innate immune cells, and how their respective bacterial endosymbionts
affect this interaction. This work also explores the transcriptional response of innate immune
cells to these infectious spores, and determines how their response is influenced by the
presence of an endosymbiont.
147

Results
To better understand the interactions between Mucorales spores and innate immune cells, I
performed a paired transcriptional study to identify responses of macrophages and fungal
spores, whilst exploring the influence of the endosymbiont on this interaction. Rhizopus
delemar and Rhizopus microsporus spores were either cured via ciprofloxacin treatment to
remove the bacterial endosymbiont (cured) or maintained in media permissive to bacterial
endosymbiosis (wt). Cured spores were passaged twice in the absence of ciprofloxacin to limit
the impact of the drug on transcriptional responses. The cured and wt spores of both R.
delemar and R. microsporus were allowed to swell in sabouraud broth (SAB) until 95% of the
population had reached mid isotropic phase (Sephton-Clark et al. 2018). Due to the
differences in germination rates between the species (Chapter 2), this occurred at 2 hours for
R. delemar and 4 hours for R. microsporus. Swollen spores were then used to infect the J774.1
murine macrophage-like cell line. Swollen fungal spores were co-cultured with macrophages
for one hour, after which unengulfed spores were removed, and phagocytosed spores were
incubated within the macrophages for a further two hours. In all cases 95% phagocytosis
was achieved. Macrophages and spores from the resulting infection were processed to
explore their transcriptional response to this infection scenario. Macrophages that had
phagocytosed swollen fungal spores (cured and wt) were isolated and sequenced via the
10X Genomics Chromium Single Cell Sequencing platform. Macrophages left unexposed
to the fungi were used as a negative control. RNA was also isolated from fungal spores
(cured and wt) which had been engulfed by macrophages, and this was sequenced with a
bulk RNA-Seq approach. Unexposed fungi (cured and wt) were incubated in macrophage
media (serum-free Dulbeccos Modified Eagle Medium) for a matched time and used as
a negative control.
148

The Fungal Response
The overall response to phagocytosis from R. delemar and R. microsporus, obtained through
the bulk RNA-Seq approach, was examined with principle component analysis (PCA) (Figure
1). Large differences between the transcriptomes of both fungal species can be observed,
when exposed or unexposed to macrophages, while the presence or absence of their
respective endosymbionts has a weak but differential effect on PCA (Figure 1). The presence
or absence of the endosymbiont appears to have very little bearing on the transcriptional
patterns displayed by R. delemar, as samples fell into two distinct clusters, most strongly
influenced by macrophage status (Figure 1a). R. microsporus exhibits a similar trend upon
exposure to macrophages, however the presence of the endosymbiont influenced clustering
more so (Figure 1b). Itabangi et al., show that the presence of the endosymbiont Ralstonia
pickettii impacts fungal cell wall organization, resistance to host-relevant stress,
spore germination efficiency, and pathogenesis of R. microsporus (Itabangi et al. 2019).
It should be noted that both sample groups contain an outlier. Given the smaller range of the
R. microsporus PCA axis (Figure 1b), the outlier for this group appears to effect grouping less,
compared to the R. delemar outlier. The outliers were both processed in separate biological
experiments, and both samples obtained a good read level and alignment level when
sequenced and processed with the bioinformatics pipeline. As the cause could not be
determined in either instance, and it is not recommended to calculate significant differential
expression with two biological repeats, the outliers were kept in for analysis. It should
therefore be noted that the results may be skewed due to the outliers presence. This will
be extenuated due to the strict statistical thresholds imposed. To improve the results, a
fourth biological repeat should be performed in the future, time and cost permitting.
149
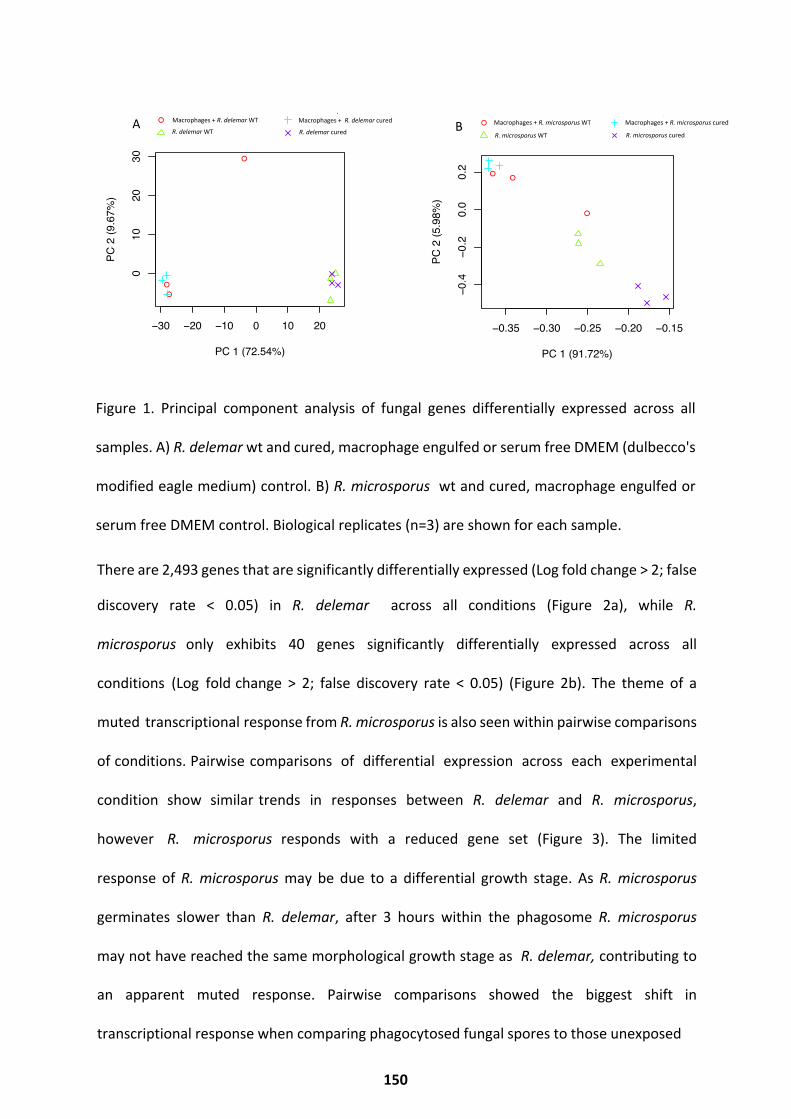
Figure 1. Principal component analysis of fungal genes differentially expressed across all
samples. A) R. delemar wt and cured, macrophage engulfed or serum free DMEM (dulbecco's
modified eagle medium) control. B) R. microsporus wt and cured, macrophage engulfed or
serum free DMEM control. Biological replicates (n=3) are shown for each sample.
There are 2,493 genes that are significantly differentially expressed (Log fold change > 2; false
discovery rate < 0.05) in R. delemar across all conditions (Figure 2a), while R.
microsporus only exhibits 40 genes significantly differentially expressed across all
conditions (Log fold change > 2; false discovery rate < 0.05) (Figure 2b). The theme of a
muted transcriptional response from R. microsporus is also seen within pairwise comparisons
of conditions. Pairwise comparisons of differential expression across each experimental
condition show similar trends in responses between R. delemar and R. microsporus,
however R. microsporus responds with a reduced gene set (Figure 3). The limited
response of R. microsporus may be due to a differential growth stage. As R. microsporus
germinates slower than R. delemar, after 3 hours within the phagosome R. microsporus
may not have reached the same morphological growth stage as R. delemar, contributing to
an apparent muted response. Pairwise comparisons showed the biggest shift in
transcriptional response when comparing phagocytosed fungal spores to those unexposed
●
●
●
−30 −20 −10 0 10 20
010
2030
PC 1 (72.54%)
PC 2
(9.6
7%)
● cond_0cond_1
cond_2cond_3
●
●●
0 10 20 30
−10
−50
5
PC 2 (9.67%)
PC 3
(4.0
0%)
● cond_0cond_1
cond_2cond_3
●
●
●
−30 −20 −10 0 10 20
010
2030
PC 1 (72.54%)
PC 2
(9.6
7%)
● cond_0cond_1
cond_2cond_3
●
●●
0 10 20 30
−10
−50
5
PC 2 (9.67%)
PC 3
(4.0
0%)
● cond_0cond_1
cond_2cond_3
Macrophages + R. delemar WT
R. delemar WT
●
●
●
−30 −20 −10 0 10 20
010
2030
PC 1 (72.54%)
PC 2
(9.6
7%)
● cond_0cond_1
cond_2cond_3
●
●●
0 10 20 30
−10
−50
5
PC 2 (9.67%)
PC 3
(4.0
0%)
● cond_0cond_1
cond_2cond_3
Macrophages + R. delemar cured
R. delemar curedA
●●
●
−0.35 −0.30 −0.25 −0.20 −0.15
−0.4
−0.2
0.0
0.2
PC 1 (91.72%)
PC 2
(5.9
8%)
● cond_0cond_1
cond_2cond_3
●●
●
−0.4 −0.2 0.0 0.2
−0.8
−0.4
0.0
0.4
PC 2 (5.98%)
PC 3
(0.7
8%)
● cond_0cond_1
cond_2cond_3
●
●
●
−30 −20 −10 0 10 20
010
2030
PC 1 (72.54%)
PC 2
(9.6
7%)
● cond_0cond_1
cond_2cond_3
●
●●
0 10 20 30
−10
−50
5
PC 2 (9.67%)
PC 3
(4.0
0%)
● cond_0cond_1
cond_2cond_3
Macrophages + R. microsporus WT
R. microsporus WT
●
●
●
−30 −20 −10 0 10 200
1020
30
PC 1 (72.54%)
PC 2
(9.6
7%)
● cond_0cond_1
cond_2cond_3
●
●●
0 10 20 30
−10
−50
5
PC 2 (9.67%)
PC 3
(4.0
0%)
● cond_0cond_1
cond_2cond_3
Macrophages + R. microsporus cured
R. microsporus curedB
150

to macrophages, regardless of endosymbiont status. When phagocytosed (Figure 4), R.
microsporus upregulates genes enriched in GO categories corresponding to
thiamine metabolism, sulfur metabolism, glycerol metabolism, alcohol
dehydrogenase activity and transmembrane transporter activity (hypergeometric test,
corrected P value < 0.05). This is consistent with the fungal response seen to
macrophage stress (Parente-Rocha et al. 2015), and the micronutrient scavenging
response to nutritional immunity (Ballou and Wilson 2016; Shen et al. 2018;
Andrianaki et al. 2018). Phagocytosed R. microsporus downregulated genes
enriched in GO categories corresponding to rRNA processing, ribosome
biogenesis and ribosome localization (hypergeometric test, corrected P value < 0.05)
(Figure 4), consistent with growth arrest within the phagolysosome (Inglesfield et al.
2018; Andrianaki et al. 2018)
151

152
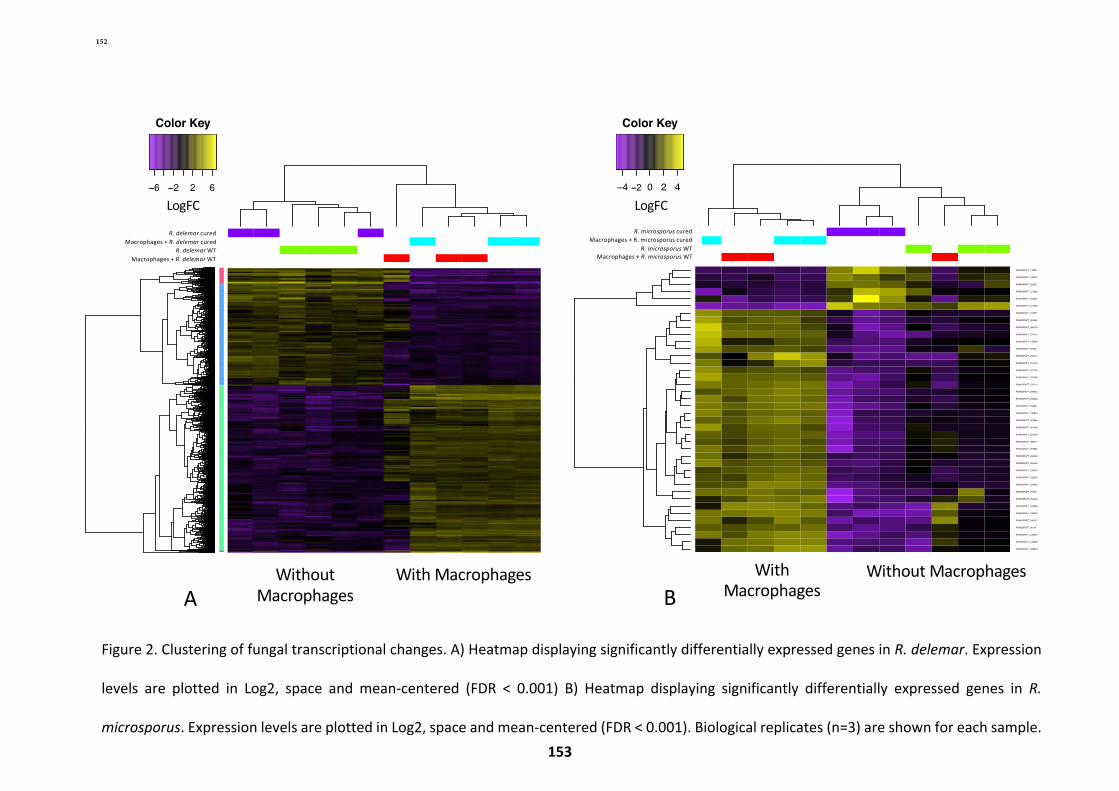
149
cond_0
cond_1
cond_2
cond_3
cond_3_rep2
cond_3_rep1
cond_1_rep3
cond_1_rep1
cond_1_rep2
cond_3_rep3
cond_0_rep2
cond_2_rep1
cond_0_rep3
cond_0_rep1
cond_2_rep3
cond_2_rep2
RO3G_005875RO3G_002036RO3G_009513RO3G_007365RO3G_006644RO3G_016651RO3G_001550RO3G_015694RO3G_009424RO3G_012739RO3G_016465RO3G_015312RO3G_009281RO3G_000465RO3G_003906RO3G_016415RO3G_001012RO3G_016742RO3G_017422RO3G_009519RO3G_013940RO3G_014198RO3G_012571RO3G_013538RO3G_005645RO3G_000866RO3G_007382RO3G_014380RO3G_006165RO3G_006326RO3G_011802RO3G_001062RO3G_008322RO3G_010934RO3G_007683RO3G_012567RO3G_009678RO3G_005635RO3G_017389RO3G_005396RO3G_015241RO3G_006584RO3G_016752RO3G_014734RO3G_010189RO3G_014933RO3G_006623RO3G_014451RO3G_001190RO3G_011698RO3G_007377RO3G_014467RO3G_007769RO3G_014471RO3G_011512RO3G_010037RO3G_016857RO3G_008711RO3G_000900RO3G_000949RO3G_014501RO3G_003598RO3G_004193RO3G_014305RO3G_016120RO3G_010417RO3G_005315RO3G_007433RO3G_016212RO3G_008411RO3G_014396RO3G_015861RO3G_011864RO3G_001309RO3G_009644RO3G_013422RO3G_004367RO3G_009836RO3G_007558RO3G_009004RO3G_001510RO3G_006479RO3G_008489RO3G_001310RO3G_015620RO3G_002493RO3G_013882RO3G_012320RO3G_017274RO3G_006358RO3G_012780RO3G_016571RO3G_003922RO3G_009949RO3G_011212RO3G_010607RO3G_009742RO3G_011162RO3G_002323RO3G_010829RO3G_000954RO3G_002500RO3G_004683RO3G_008249RO3G_006968RO3G_017547RO3G_003102RO3G_000748RO3G_001824RO3G_016254RO3G_003106RO3G_013722RO3G_017160RO3G_009881RO3G_013380RO3G_010796RO3G_002214RO3G_009506RO3G_000908RO3G_008048RO3G_008370RO3G_000468RO3G_012203RO3G_015628RO3G_004544RO3G_003695RO3G_001423RO3G_010990RO3G_006780RO3G_003602RO3G_015213RO3G_004678RO3G_005437RO3G_014646RO3G_005277RO3G_017052RO3G_005059RO3G_014327RO3G_013872RO3G_014340RO3G_011832RO3G_001923RO3G_003091RO3G_014401RO3G_011172RO3G_010903RO3G_005470RO3G_002901RO3G_011817RO3G_003862RO3G_016107RO3G_004721RO3G_006891RO3G_008545RO3G_006894RO3G_015812RO3G_016843RO3G_014481RO3G_001557RO3G_002536RO3G_009159RO3G_008028RO3G_008578RO3G_011184RO3G_004001RO3G_006206RO3G_008727RO3G_002010RO3G_004555RO3G_010673RO3G_013999RO3G_001769RO3G_014851RO3G_005588RO3G_005148RO3G_016300RO3G_012201RO3G_017376RO3G_014162RO3G_001432RO3G_001476RO3G_005274RO3G_006645RO3G_011183RO3G_005137RO3G_013060RO3G_016089RO3G_008500RO3G_007795RO3G_014057RO3G_001097RO3G_014140RO3G_008818RO3G_001707RO3G_016207RO3G_015442RO3G_016268RO3G_006615RO3G_015449RO3G_012452RO3G_009658RO3G_007835RO3G_015881RO3G_003009RO3G_017703RO3G_009187RO3G_016195RO3G_007400RO3G_005904RO3G_006119RO3G_011533RO3G_008558RO3G_016939RO3G_016704RO3G_015490RO3G_013446RO3G_001677RO3G_014971RO3G_007723RO3G_000974RO3G_015579RO3G_013542RO3G_014650RO3G_010433RO3G_005008RO3G_006318RO3G_005899RO3G_000431RO3G_001255RO3G_013281RO3G_011928RO3G_011987RO3G_016325RO3G_013995RO3G_010187RO3G_011399RO3G_011754RO3G_005708RO3G_010002RO3G_013746RO3G_015170RO3G_006145RO3G_007825RO3G_000386RO3G_014298RO3G_012662RO3G_011989RO3G_008929RO3G_005089RO3G_011966RO3G_008351RO3G_012076RO3G_000988RO3G_013888RO3G_005037RO3G_011806RO3G_003166RO3G_004174RO3G_003605RO3G_010132RO3G_002307RO3G_001063RO3G_003293RO3G_003618RO3G_008995RO3G_002456RO3G_012572RO3G_003867RO3G_001808RO3G_005447RO3G_014874RO3G_015798RO3G_009128RO3G_006189RO3G_008227RO3G_015889RO3G_006100RO3G_000266RO3G_000017RO3G_004981RO3G_000249RO3G_013469RO3G_004160RO3G_013571RO3G_016889RO3G_012981RO3G_006622RO3G_003859RO3G_000984RO3G_013624RO3G_004086RO3G_009343RO3G_003912RO3G_013623RO3G_002325RO3G_002533RO3G_011448RO3G_009889RO3G_004813RO3G_009414RO3G_017006RO3G_004010RO3G_007225RO3G_009541RO3G_017073RO3G_000704RO3G_004144RO3G_011905RO3G_017443RO3G_007718RO3G_002767RO3G_004748RO3G_001481RO3G_004138RO3G_005359RO3G_014419RO3G_000874RO3G_001929RO3G_006646RO3G_008103RO3G_010057RO3G_009932RO3G_015347RO3G_011507RO3G_007366RO3G_015504RO3G_009920RO3G_016769RO3G_006128RO3G_015028RO3G_003075RO3G_014724RO3G_002169RO3G_002326RO3G_015779RO3G_005912RO3G_004858RO3G_001951RO3G_008696RO3G_010122RO3G_002747RO3G_007856RO3G_014412RO3G_013534RO3G_009509RO3G_004009RO3G_010098RO3G_001675RO3G_003232RO3G_001224RO3G_004400RO3G_012565RO3G_001525RO3G_005384RO3G_011454RO3G_014441RO3G_008478RO3G_008389RO3G_010257RO3G_015439RO3G_011530RO3G_002806RO3G_009277RO3G_006429RO3G_001179RO3G_007722RO3G_005278RO3G_011400RO3G_007989RO3G_000696RO3G_007013RO3G_000223RO3G_011670RO3G_009007RO3G_004047RO3G_008047RO3G_013858RO3G_017846RO3G_008470RO3G_009453RO3G_001380RO3G_006163RO3G_002990RO3G_007772RO3G_014309RO3G_013251RO3G_017741RO3G_008045RO3G_017756RO3G_003030RO3G_012535RO3G_003256RO3G_002353RO3G_011859RO3G_005015RO3G_004934RO3G_001371RO3G_002497RO3G_002551RO3G_008625RO3G_012790RO3G_003575RO3G_006271RO3G_010000RO3G_017357RO3G_013444RO3G_012124RO3G_011914RO3G_003683RO3G_004982RO3G_006439RO3G_011972RO3G_007258RO3G_014363RO3G_014525RO3G_006507RO3G_000556RO3G_014119RO3G_016237RO3G_016217RO3G_004957RO3G_008902RO3G_017162RO3G_013541RO3G_002095RO3G_008376RO3G_014347RO3G_016149RO3G_011814RO3G_008492RO3G_001091RO3G_005980RO3G_012645RO3G_001314RO3G_013717RO3G_010669RO3G_006731RO3G_007227RO3G_016162RO3G_010964RO3G_003596RO3G_016488RO3G_006225RO3G_013385RO3G_013353RO3G_012385RO3G_004015RO3G_016299RO3G_008972RO3G_007545RO3G_007954RO3G_017167RO3G_005914RO3G_010693RO3G_004597RO3G_016193RO3G_006883RO3G_014166RO3G_011548RO3G_011427RO3G_010988RO3G_016321RO3G_004457RO3G_003077RO3G_001223RO3G_012700RO3G_000187RO3G_014713RO3G_013910RO3G_000464RO3G_011685RO3G_007738RO3G_016298RO3G_008394RO3G_001933RO3G_014920RO3G_010667RO3G_001755RO3G_006363RO3G_012755RO3G_002327RO3G_013896RO3G_006023RO3G_012388RO3G_010991RO3G_008836RO3G_009649RO3G_000644RO3G_007864RO3G_000452RO3G_010159RO3G_011809RO3G_011841RO3G_007086RO3G_006980RO3G_007845RO3G_014553RO3G_004842RO3G_005698RO3G_008809RO3G_011204RO3G_014335RO3G_007727RO3G_005024RO3G_005620RO3G_004033RO3G_005583RO3G_008764RO3G_002632RO3G_007945RO3G_004168RO3G_001758RO3G_011273RO3G_011942RO3G_007531RO3G_009294RO3G_009193RO3G_006474RO3G_005407RO3G_002879RO3G_007556RO3G_006800RO3G_000854RO3G_005724RO3G_006463RO3G_014002RO3G_005670RO3G_000922RO3G_002907RO3G_012218RO3G_007191RO3G_011340RO3G_003411RO3G_010664RO3G_009140RO3G_016206RO3G_004715RO3G_003289RO3G_007124RO3G_017079RO3G_003877RO3G_014136RO3G_013777RO3G_017712RO3G_011406RO3G_016873RO3G_007480RO3G_012573RO3G_014403RO3G_010685RO3G_016572RO3G_015340RO3G_015649RO3G_004456RO3G_001085RO3G_010660RO3G_008793RO3G_011937RO3G_017408RO3G_004333RO3G_012764RO3G_017062RO3G_014678RO3G_010405RO3G_014206RO3G_009647RO3G_016187RO3G_015555RO3G_005669RO3G_006285RO3G_010114RO3G_000687RO3G_013157RO3G_016741RO3G_009222RO3G_006742RO3G_014319RO3G_002009RO3G_007661RO3G_010407RO3G_006743RO3G_001812RO3G_014048RO3G_013232RO3G_007287RO3G_001970RO3G_010548RO3G_000222RO3G_007364RO3G_003664RO3G_005193RO3G_000537RO3G_013087RO3G_017131RO3G_005147RO3G_013309RO3G_013495RO3G_006113RO3G_009253RO3G_013889RO3G_004963RO3G_017103RO3G_008250RO3G_017242RO3G_013773RO3G_005041RO3G_007449RO3G_015884RO3G_009272RO3G_008537RO3G_000819RO3G_015359RO3G_009549RO3G_001753RO3G_001723RO3G_013068RO3G_009664RO3G_007546RO3G_009275RO3G_016194RO3G_011558RO3G_017766RO3G_008932RO3G_002312RO3G_012545RO3G_005935RO3G_014210RO3G_000388RO3G_005702RO3G_003025RO3G_006604RO3G_017698RO3G_016176RO3G_002790RO3G_001488RO3G_000910RO3G_015687RO3G_016617RO3G_011138RO3G_001142RO3G_003285RO3G_010311RO3G_012269RO3G_011700RO3G_013748RO3G_008974RO3G_004995RO3G_000505RO3G_005420RO3G_011417RO3G_016524RO3G_001961RO3G_017352RO3G_005890RO3G_014186RO3G_001847RO3G_007277RO3G_002479RO3G_011470RO3G_015097RO3G_006016RO3G_014906RO3G_010102RO3G_010840RO3G_014517RO3G_002047RO3G_006505RO3G_017234RO3G_015259RO3G_010310RO3G_008416RO3G_005320RO3G_009931RO3G_015426RO3G_015156RO3G_001350RO3G_017371RO3G_001232RO3G_006500RO3G_010124RO3G_012729RO3G_008157RO3G_014054RO3G_008252RO3G_015758RO3G_012211RO3G_004249RO3G_017259RO3G_001635RO3G_004742RO3G_010792RO3G_012942RO3G_002720RO3G_016788RO3G_001553RO3G_008577RO3G_003398RO3G_000871RO3G_001754RO3G_007495RO3G_000490RO3G_006970RO3G_007765RO3G_011762RO3G_006913RO3G_002909RO3G_008963RO3G_002635RO3G_004800RO3G_000686RO3G_011240RO3G_016740RO3G_011178RO3G_011125RO3G_015567RO3G_008114RO3G_007544RO3G_012445RO3G_003610RO3G_002502RO3G_010986RO3G_004225RO3G_015425RO3G_012893RO3G_008121RO3G_004079RO3G_015074RO3G_004979RO3G_008635RO3G_016771RO3G_008151RO3G_002942RO3G_014892RO3G_006403RO3G_007775RO3G_003043RO3G_010608RO3G_011152RO3G_004167RO3G_012852RO3G_012705RO3G_012008RO3G_012685RO3G_012444RO3G_003569RO3G_000860RO3G_001544RO3G_003288RO3G_008486RO3G_010604RO3G_017177RO3G_001154RO3G_017833RO3G_001571RO3G_000394RO3G_011438RO3G_010827RO3G_006159RO3G_004082RO3G_000257RO3G_013314RO3G_003504RO3G_000868RO3G_010708RO3G_011324RO3G_016620RO3G_003168RO3G_001116RO3G_017087RO3G_007333RO3G_015024RO3G_005124RO3G_009005RO3G_010144RO3G_003337RO3G_004821RO3G_001656RO3G_009999RO3G_001509RO3G_011993RO3G_004027RO3G_006575RO3G_016927RO3G_010895RO3G_005573RO3G_014764RO3G_009878RO3G_008089RO3G_013338RO3G_006349RO3G_010426RO3G_001759RO3G_002499RO3G_012894RO3G_003055RO3G_004198RO3G_012551RO3G_009952RO3G_008536RO3G_011858RO3G_009157RO3G_014189RO3G_012511RO3G_010452RO3G_010182RO3G_003017RO3G_003172RO3G_011445RO3G_016159RO3G_013390RO3G_007966RO3G_013408RO3G_009011RO3G_010286RO3G_014968RO3G_000882RO3G_002444RO3G_008922RO3G_005219RO3G_010931RO3G_007508RO3G_001562RO3G_002910RO3G_011658RO3G_009095RO3G_008651RO3G_003499RO3G_005011RO3G_000518RO3G_005751RO3G_007489RO3G_008585RO3G_016320RO3G_007060RO3G_011437RO3G_002014RO3G_012819RO3G_006495RO3G_016464RO3G_013447RO3G_011396RO3G_011948RO3G_000839RO3G_008076RO3G_004186RO3G_010127RO3G_001181RO3G_013729RO3G_015740RO3G_013243RO3G_017320RO3G_001508RO3G_013557RO3G_002055RO3G_008794RO3G_006878RO3G_008232RO3G_004825RO3G_011970RO3G_001138RO3G_000171RO3G_017717RO3G_006105RO3G_001092RO3G_000146RO3G_002213RO3G_005296RO3G_016658RO3G_007376RO3G_006198RO3G_007645RO3G_002553RO3G_003174RO3G_010170RO3G_011503RO3G_011695RO3G_011025RO3G_016345RO3G_005023RO3G_014074RO3G_001822RO3G_006468RO3G_010615RO3G_014753RO3G_008308RO3G_011664RO3G_014051RO3G_008646RO3G_010984RO3G_007288RO3G_000966RO3G_017824RO3G_016921RO3G_012831RO3G_003564RO3G_001746RO3G_005063RO3G_015940RO3G_012096RO3G_003653RO3G_009223RO3G_011260RO3G_005319RO3G_012904RO3G_007091RO3G_007725RO3G_013202RO3G_004219RO3G_009504RO3G_013710RO3G_008235RO3G_010418RO3G_014418RO3G_013770RO3G_000667RO3G_012653RO3G_014397RO3G_015871RO3G_005102RO3G_008712RO3G_011201RO3G_005285RO3G_000231RO3G_010948RO3G_015080RO3G_017166RO3G_004814RO3G_001005RO3G_013241RO3G_000391RO3G_008728RO3G_007866RO3G_005185RO3G_016416RO3G_004159RO3G_003119RO3G_005839RO3G_005364RO3G_010054RO3G_016851RO3G_013744RO3G_013630RO3G_009609RO3G_014292RO3G_006453RO3G_001552RO3G_012161RO3G_006661RO3G_013067RO3G_017294RO3G_003394RO3G_013787RO3G_005640RO3G_001928RO3G_011153RO3G_003228RO3G_005639RO3G_001228RO3G_014769RO3G_011156RO3G_017777RO3G_015650RO3G_003147RO3G_011136RO3G_007103RO3G_008241RO3G_007211RO3G_005379RO3G_015014RO3G_007327RO3G_014430RO3G_014429RO3G_005721RO3G_000834RO3G_003066RO3G_001777RO3G_004430RO3G_009505RO3G_001501RO3G_015145RO3G_001215RO3G_012462RO3G_014255RO3G_001559RO3G_008094RO3G_007600RO3G_005131RO3G_015094RO3G_014771RO3G_014630RO3G_009655RO3G_000853RO3G_011930RO3G_009613RO3G_002752RO3G_011397RO3G_001844RO3G_002562RO3G_013326RO3G_008628RO3G_005425RO3G_010691RO3G_015100RO3G_004109RO3G_009837RO3G_010012RO3G_010682RO3G_000790RO3G_014410RO3G_009190RO3G_013369RO3G_012314RO3G_000901RO3G_001186RO3G_005861RO3G_009240RO3G_010920RO3G_011746RO3G_000224RO3G_006496RO3G_007841RO3G_008044RO3G_001927RO3G_006313RO3G_008172RO3G_008526RO3G_008849RO3G_011251RO3G_016938RO3G_011506RO3G_010035RO3G_014072RO3G_005438RO3G_005098RO3G_009324RO3G_013078RO3G_008532RO3G_000173RO3G_009282RO3G_000963RO3G_006193RO3G_002045RO3G_004189RO3G_017831RO3G_014073RO3G_016000RO3G_008083RO3G_017161RO3G_004245RO3G_004952RO3G_010921RO3G_001102RO3G_003448RO3G_016023RO3G_006306RO3G_011630RO3G_009738RO3G_006110RO3G_004366RO3G_009524RO3G_010238RO3G_006352RO3G_005777RO3G_006106RO3G_013188RO3G_016208RO3G_003038RO3G_001894RO3G_006975RO3G_010915RO3G_011741RO3G_006307RO3G_007215RO3G_016286RO3G_008544RO3G_008944RO3G_004515RO3G_013041RO3G_010477RO3G_012848RO3G_008474RO3G_009245RO3G_016647RO3G_010139RO3G_015794RO3G_002309RO3G_002416RO3G_016312RO3G_012626RO3G_012226RO3G_011733RO3G_011484RO3G_012549RO3G_009743RO3G_012765RO3G_017765RO3G_005924RO3G_007375RO3G_001486RO3G_004520RO3G_008528RO3G_001827RO3G_017286RO3G_016072RO3G_002200RO3G_010221RO3G_005838RO3G_004584RO3G_011359RO3G_007161RO3G_007005RO3G_010613RO3G_017776RO3G_016819RO3G_003472RO3G_006435RO3G_013330RO3G_017737RO3G_002474RO3G_009725RO3G_016598RO3G_002431RO3G_001493RO3G_011451RO3G_008032RO3G_006256RO3G_006718RO3G_015440RO3G_010018RO3G_002191RO3G_017832RO3G_006890RO3G_004874RO3G_002887RO3G_015019RO3G_006195RO3G_001569RO3G_017769RO3G_011373RO3G_006269RO3G_007684RO3G_003040RO3G_005931RO3G_011258RO3G_013849RO3G_004740RO3G_008409RO3G_002175RO3G_002233RO3G_006156RO3G_012068RO3G_003380RO3G_007757RO3G_006281RO3G_006038RO3G_008885RO3G_005123RO3G_007633RO3G_005740RO3G_006386RO3G_010500RO3G_008343RO3G_003844RO3G_015444RO3G_006998RO3G_010454RO3G_014214RO3G_011965RO3G_012493RO3G_010851RO3G_004729RO3G_016593RO3G_013001RO3G_006902RO3G_017009RO3G_013237RO3G_007010RO3G_012394RO3G_007990RO3G_001144RO3G_004605RO3G_008358RO3G_014967RO3G_009429RO3G_014907RO3G_014116RO3G_016587RO3G_012988RO3G_015262RO3G_011953RO3G_016164RO3G_008879RO3G_017301RO3G_014008RO3G_007687RO3G_009322RO3G_012906RO3G_011992RO3G_012897RO3G_014433RO3G_015285RO3G_007394RO3G_016691RO3G_001305RO3G_015026RO3G_004687RO3G_007250RO3G_002824RO3G_014228RO3G_001308RO3G_017074RO3G_008731RO3G_010038RO3G_000971RO3G_017462RO3G_010467RO3G_013878RO3G_008057RO3G_016709RO3G_013415RO3G_010123RO3G_014522RO3G_007787RO3G_012993RO3G_004232RO3G_012131RO3G_010095RO3G_011031RO3G_003652RO3G_006459RO3G_017159RO3G_004606RO3G_008647RO3G_001431RO3G_015168RO3G_008754RO3G_008109RO3G_005211RO3G_003866RO3G_012383RO3G_015452RO3G_017382RO3G_010844RO3G_002322RO3G_009616RO3G_011268RO3G_009958RO3G_005919RO3G_004935RO3G_009528RO3G_015781RO3G_005478RO3G_010596RO3G_013570RO3G_010294RO3G_015380RO3G_013578RO3G_005832RO3G_003042RO3G_004165RO3G_000844RO3G_001339RO3G_013329RO3G_013636RO3G_016573RO3G_014739RO3G_006182RO3G_016621RO3G_010224RO3G_011170RO3G_000228RO3G_004488RO3G_011155RO3G_008899RO3G_010305RO3G_006512RO3G_010431RO3G_004552RO3G_003343RO3G_013040RO3G_001344RO3G_001574RO3G_009078RO3G_012722RO3G_015811RO3G_013481RO3G_012840RO3G_014652RO3G_013336RO3G_010710RO3G_013360RO3G_003505RO3G_008004RO3G_015691RO3G_005152RO3G_002904RO3G_011459RO3G_007780RO3G_014176RO3G_012090RO3G_008717RO3G_012004RO3G_001122RO3G_006670RO3G_014006RO3G_013264RO3G_007270RO3G_009507RO3G_000504RO3G_015883RO3G_001067RO3G_003393RO3G_009598RO3G_001280RO3G_005755RO3G_016240RO3G_013074RO3G_010173RO3G_012023RO3G_004040RO3G_001252RO3G_000622RO3G_003383RO3G_003342RO3G_012001RO3G_017469RO3G_006028RO3G_002426RO3G_006151RO3G_011724RO3G_010556RO3G_007771RO3G_005836RO3G_005268RO3G_010126RO3G_011946RO3G_001926RO3G_008828RO3G_008734RO3G_002428RO3G_002983RO3G_014890RO3G_010315RO3G_003132RO3G_014205RO3G_003095RO3G_011629RO3G_005727RO3G_010788RO3G_013398RO3G_009150RO3G_002501RO3G_006360RO3G_007354RO3G_011455RO3G_013903RO3G_005668RO3G_002992RO3G_009241RO3G_014888RO3G_009954RO3G_017793RO3G_013494RO3G_003426RO3G_009953RO3G_008584RO3G_003392RO3G_010950RO3G_011379RO3G_015865RO3G_000466RO3G_004598RO3G_009246RO3G_003869RO3G_015559RO3G_004530RO3G_015568RO3G_007740RO3G_016738RO3G_001624RO3G_001219RO3G_007952RO3G_010307RO3G_011247RO3G_003407RO3G_007118RO3G_007069RO3G_012009RO3G_009595RO3G_006146RO3G_013257RO3G_007331RO3G_001560RO3G_014492RO3G_011213RO3G_009946RO3G_015258RO3G_011131RO3G_004095RO3G_010223RO3G_002057RO3G_007534RO3G_010235RO3G_017841RO3G_009593RO3G_000923RO3G_001216RO3G_003227RO3G_004829RO3G_010181RO3G_007319RO3G_004433RO3G_012664RO3G_008914RO3G_014171RO3G_008234RO3G_005929RO3G_004839RO3G_015298RO3G_007981RO3G_011449RO3G_005783RO3G_011319RO3G_009342RO3G_016113RO3G_006168RO3G_003103RO3G_012766RO3G_003415RO3G_015578RO3G_012016RO3G_005275RO3G_002212RO3G_009851RO3G_016626RO3G_001704RO3G_009997RO3G_007628RO3G_008412RO3G_013998RO3G_010292RO3G_016874RO3G_002755RO3G_002883RO3G_008313RO3G_005445RO3G_016791RO3G_014359RO3G_003028RO3G_014453RO3G_007152RO3G_011252RO3G_015634RO3G_017796RO3G_002937RO3G_015060RO3G_008598RO3G_012136RO3G_012195RO3G_013154RO3G_014004RO3G_017031RO3G_008371RO3G_001830RO3G_005672RO3G_010085RO3G_012449RO3G_014321RO3G_006616RO3G_008469RO3G_005067RO3G_002260RO3G_017706RO3G_012683RO3G_009736RO3G_010968RO3G_017150RO3G_017791RO3G_006041RO3G_002542RO3G_012282RO3G_012283RO3G_008248RO3G_015189RO3G_005614RO3G_003728RO3G_011019RO3G_011023RO3G_002008RO3G_004581RO3G_010011RO3G_000960RO3G_003300RO3G_014026RO3G_017351RO3G_004685RO3G_001315RO3G_017709RO3G_003853RO3G_003142RO3G_000840RO3G_010975RO3G_001828RO3G_004917RO3G_005189RO3G_012891RO3G_014716RO3G_015186RO3G_002905RO3G_013968RO3G_004556RO3G_000951RO3G_001364RO3G_017411RO3G_016172RO3G_012842RO3G_004856RO3G_015257RO3G_005307RO3G_012513RO3G_000711RO3G_009205RO3G_005075RO3G_013846RO3G_017453RO3G_000909RO3G_017114RO3G_008824RO3G_010972RO3G_012169RO3G_003155RO3G_000123RO3G_008515RO3G_002886RO3G_016061RO3G_017640RO3G_012876RO3G_008945RO3G_002152RO3G_012715RO3G_000393RO3G_014168RO3G_008237RO3G_003027RO3G_011434RO3G_001632RO3G_001862RO3G_000631RO3G_014204RO3G_009252RO3G_012999RO3G_006993RO3G_005306RO3G_004196RO3G_001221RO3G_006287RO3G_014465RO3G_014324RO3G_007984RO3G_016257RO3G_015762RO3G_009568RO3G_013859RO3G_006292RO3G_002707RO3G_009897RO3G_001924RO3G_011488RO3G_005256RO3G_002215RO3G_013327RO3G_004460RO3G_008397RO3G_017461RO3G_015243RO3G_003169RO3G_013546RO3G_006629RO3G_012913RO3G_010574RO3G_001876RO3G_010435RO3G_009437RO3G_005144RO3G_014045RO3G_010415RO3G_005058RO3G_004459RO3G_004722RO3G_004689RO3G_000757RO3G_011414RO3G_012528RO3G_013911RO3G_008806RO3G_016698RO3G_011022RO3G_011412RO3G_005631RO3G_009349RO3G_003108RO3G_007485RO3G_011132RO3G_011981RO3G_013387RO3G_016355RO3G_007812RO3G_005634RO3G_006081RO3G_012566RO3G_000359RO3G_012716RO3G_001940RO3G_012290RO3G_013071RO3G_008951RO3G_004464RO3G_014682RO3G_009544RO3G_007105RO3G_000201RO3G_013559RO3G_013548RO3G_010149RO3G_004438RO3G_012687RO3G_001003RO3G_015108RO3G_011843RO3G_016902RO3G_007302RO3G_008749RO3G_015849RO3G_005643RO3G_015051RO3G_017253RO3G_006503RO3G_000852RO3G_010160RO3G_010800RO3G_006004RO3G_010510RO3G_002467RO3G_016689RO3G_002659RO3G_005512RO3G_007879RO3G_015879RO3G_006448RO3G_017812RO3G_012235RO3G_003404RO3G_017069RO3G_011277RO3G_010949RO3G_009992RO3G_005959RO3G_009554RO3G_004432RO3G_014061RO3G_006196RO3G_005213RO3G_010919RO3G_013275RO3G_009628RO3G_007272RO3G_014039RO3G_016469RO3G_007876RO3G_003760RO3G_016736RO3G_010589RO3G_005336RO3G_008391RO3G_003579RO3G_000108RO3G_001225RO3G_003868RO3G_014867RO3G_014019RO3G_017282RO3G_009423RO3G_015301RO3G_001688RO3G_005954RO3G_012381RO3G_005292RO3G_007129RO3G_017315RO3G_011344RO3G_004134RO3G_008767RO3G_007991RO3G_006498RO3G_001222RO3G_014667RO3G_016075RO3G_016585RO3G_011916RO3G_008766RO3G_016903RO3G_016733RO3G_016451RO3G_011134RO3G_003876RO3G_002800RO3G_010203RO3G_002846RO3G_011300RO3G_014355RO3G_012935RO3G_012123RO3G_012810RO3G_008656RO3G_000893RO3G_017265RO3G_007648RO3G_005504RO3G_003969RO3G_001185RO3G_013175RO3G_008715RO3G_016124RO3G_010300RO3G_012193RO3G_017060RO3G_000196RO3G_009739RO3G_003533RO3G_014449RO3G_005604RO3G_016528RO3G_003874RO3G_015932RO3G_011462RO3G_003430RO3G_000248RO3G_011001RO3G_006048RO3G_007427RO3G_016083RO3G_005497RO3G_002131RO3G_008602RO3G_003578RO3G_005716RO3G_005385RO3G_004490RO3G_002181RO3G_000243RO3G_017090RO3G_012520RO3G_008239RO3G_004495RO3G_003083RO3G_014745RO3G_017721RO3G_014861RO3G_017434RO3G_009167RO3G_010443RO3G_013091RO3G_001196RO3G_017762RO3G_015093RO3G_004497RO3G_007796RO3G_005028RO3G_016290RO3G_002531RO3G_014755RO3G_005957RO3G_011034RO3G_005993RO3G_002046RO3G_002874RO3G_003233RO3G_001842RO3G_005402RO3G_011330RO3G_015790RO3G_006961RO3G_002993RO3G_000244RO3G_004695RO3G_012073RO3G_008398RO3G_012936RO3G_017891RO3G_016074RO3G_012402RO3G_016919RO3G_011127RO3G_007730RO3G_008681RO3G_003550RO3G_016011RO3G_016221RO3G_002661RO3G_001066RO3G_012407RO3G_010304RO3G_014081RO3G_003901RO3G_013847RO3G_009334RO3G_005876RO3G_016913RO3G_008541RO3G_002674RO3G_014929RO3G_002156RO3G_012837RO3G_015348RO3G_001874RO3G_012856RO3G_013266RO3G_006486RO3G_016500RO3G_014317RO3G_002450RO3G_002306RO3G_005048RO3G_005388RO3G_001644RO3G_000524RO3G_016367RO3G_006092RO3G_010985RO3G_014337RO3G_007421RO3G_004738RO3G_009196RO3G_017010RO3G_015336RO3G_008418RO3G_001001RO3G_004915RO3G_004734RO3G_014742RO3G_014837RO3G_008881RO3G_014385RO3G_010479RO3G_015417RO3G_017183RO3G_007484RO3G_006737RO3G_002186RO3G_015192RO3G_013575RO3G_013855RO3G_006738RO3G_003046RO3G_014349RO3G_015825RO3G_003348RO3G_002583RO3G_009623RO3G_016216RO3G_008066RO3G_002625RO3G_000930RO3G_011419RO3G_008657RO3G_002480RO3G_007709RO3G_009618RO3G_000890RO3G_015648RO3G_007817RO3G_004528RO3G_001214RO3G_009987RO3G_004595RO3G_015120RO3G_003599RO3G_016297RO3G_013179RO3G_009918RO3G_010270RO3G_009034RO3G_008791RO3G_011757RO3G_016283RO3G_017145RO3G_003930RO3G_013289RO3G_001935RO3G_009632RO3G_009493RO3G_008984RO3G_006981RO3G_016460RO3G_010707RO3G_005465RO3G_005982RO3G_004253RO3G_003332RO3G_005865RO3G_017272RO3G_001153RO3G_016614RO3G_013715RO3G_015320RO3G_013588RO3G_004968RO3G_002035RO3G_009332RO3G_009824RO3G_009823RO3G_017405RO3G_009363RO3G_012400RO3G_005831RO3G_010779RO3G_000539RO3G_008064RO3G_008040RO3G_006681RO3G_004532RO3G_000118RO3G_005623RO3G_009521RO3G_004534RO3G_004329RO3G_002140RO3G_009130RO3G_001558RO3G_008132RO3G_017644RO3G_004516RO3G_014017RO3G_009601RO3G_005880RO3G_009173RO3G_012278RO3G_004355RO3G_008327RO3G_009631RO3G_000327RO3G_002091RO3G_017816RO3G_017837RO3G_010736RO3G_003330RO3G_003682RO3G_007755RO3G_014344RO3G_004314RO3G_001117RO3G_001490RO3G_014969RO3G_015175RO3G_000976RO3G_016153RO3G_012457RO3G_015117RO3G_007186RO3G_004091RO3G_012239RO3G_013933RO3G_004526RO3G_011742RO3G_011560RO3G_013789RO3G_009295RO3G_008162RO3G_013983RO3G_005797RO3G_011792RO3G_011476RO3G_016282RO3G_012910RO3G_001869RO3G_017093RO3G_009484RO3G_007745RO3G_001115RO3G_006143RO3G_007523RO3G_006658RO3G_014696RO3G_016116RO3G_007284RO3G_017022RO3G_003134RO3G_003677RO3G_007504RO3G_003657RO3G_005587RO3G_008732RO3G_002816RO3G_011823RO3G_010483RO3G_000494RO3G_008665RO3G_011407RO3G_016122RO3G_015791RO3G_015322RO3G_003594RO3G_010883RO3G_012185RO3G_016040RO3G_015775RO3G_007702RO3G_008225RO3G_002823RO3G_015297RO3G_015317RO3G_003163RO3G_015793RO3G_015346RO3G_012066RO3G_015307RO3G_007882RO3G_009081RO3G_016067RO3G_007561RO3G_004921RO3G_013990RO3G_005222RO3G_015875RO3G_016666RO3G_008392RO3G_007058RO3G_014733RO3G_011413RO3G_004211RO3G_017012RO3G_010184RO3G_001515RO3G_002513RO3G_010562RO3G_009430RO3G_015806RO3G_015112RO3G_012458RO3G_006013RO3G_003052RO3G_005608RO3G_005618RO3G_009713RO3G_016898RO3G_005796RO3G_011006RO3G_003151RO3G_005857RO3G_014550RO3G_014912RO3G_013328RO3G_010129RO3G_015212RO3G_001529RO3G_014133RO3G_009293RO3G_004543RO3G_011643RO3G_010006RO3G_012081RO3G_016809RO3G_004804RO3G_002672RO3G_014503RO3G_012289RO3G_003700RO3G_003708RO3G_006668RO3G_009067RO3G_009064RO3G_011478RO3G_001220RO3G_013719RO3G_003647RO3G_016070RO3G_016610RO3G_009033RO3G_012802RO3G_003666RO3G_005939RO3G_000143RO3G_000446RO3G_017335RO3G_007158RO3G_001320RO3G_015591RO3G_012210RO3G_015551RO3G_014237RO3G_000191RO3G_016688RO3G_015161RO3G_008751RO3G_002436RO3G_017412RO3G_004990RO3G_017378RO3G_011644RO3G_004462RO3G_015563RO3G_013547RO3G_011716RO3G_012839RO3G_008001RO3G_008710RO3G_014992RO3G_013056RO3G_005981RO3G_011839RO3G_011144RO3G_008707RO3G_002896RO3G_002072RO3G_010231RO3G_002093RO3G_006341RO3G_005453RO3G_007706RO3G_009529RO3G_011494RO3G_015848RO3G_002903RO3G_015598RO3G_013035RO3G_006074RO3G_008390RO3G_009485RO3G_005305RO3G_016052RO3G_003679RO3G_011355RO3G_007318RO3G_005853RO3G_012071RO3G_012699RO3G_015390RO3G_005162RO3G_001178RO3G_007820RO3G_012430RO3G_005571RO3G_013431RO3G_004451RO3G_010229RO3G_004055RO3G_005518RO3G_009607RO3G_017418RO3G_003540RO3G_009138RO3G_009063RO3G_004202RO3G_004115RO3G_005077RO3G_017034RO3G_002463RO3G_011523RO3G_011165RO3G_002538RO3G_013288RO3G_000254RO3G_000883RO3G_017314RO3G_005748RO3G_014934RO3G_005597RO3G_005996RO3G_009556RO3G_008364RO3G_003439RO3G_012098RO3G_014049RO3G_012267RO3G_006953RO3G_004097RO3G_004178RO3G_004101RO3G_004251RO3G_009218RO3G_005047RO3G_004494RO3G_001160RO3G_010579RO3G_011038RO3G_002534RO3G_006409RO3G_001323RO3G_004491RO3G_003563RO3G_007770RO3G_006291RO3G_003705RO3G_011782RO3G_008357RO3G_000242RO3G_007744RO3G_013591RO3G_007822RO3G_012625RO3G_007356RO3G_010473RO3G_006096RO3G_010983RO3G_001261RO3G_001462RO3G_015991RO3G_008987RO3G_004162RO3G_009604RO3G_001187RO3G_006402RO3G_005488RO3G_011535RO3G_010185RO3G_003701RO3G_003702RO3G_001489RO3G_005594RO3G_007143RO3G_015743RO3G_000221RO3G_016220RO3G_016123RO3G_009537RO3G_008243RO3G_000475RO3G_016270RO3G_000477RO3G_000476RO3G_002217RO3G_005516RO3G_008024RO3G_002918RO3G_004356RO3G_005483RO3G_003617RO3G_003763RO3G_017100RO3G_003019RO3G_009495RO3G_002933RO3G_013852RO3G_002412RO3G_012013RO3G_009436RO3G_006427RO3G_003154RO3G_017122RO3G_005999RO3G_014996RO3G_002224RO3G_010981RO3G_015299RO3G_006027RO3G_008758RO3G_009669RO3G_016059RO3G_000434RO3G_012220RO3G_010175RO3G_000456RO3G_004092RO3G_007147RO3G_001161RO3G_006725RO3G_005197RO3G_005485RO3G_009715RO3G_004458RO3G_001377RO3G_005565RO3G_006803RO3G_012413RO3G_005582RO3G_003581RO3G_012287RO3G_009089RO3G_004087RO3G_013982RO3G_017423RO3G_009106RO3G_003872RO3G_002148RO3G_007159RO3G_009646RO3G_006406RO3G_009922RO3G_015989RO3G_014868RO3G_017048RO3G_001376RO3G_009249RO3G_000463RO3G_009091RO3G_005612RO3G_006723RO3G_009085RO3G_010711RO3G_016532RO3G_016152RO3G_010246RO3G_006032RO3G_006034RO3G_015655RO3G_001698RO3G_010785RO3G_001549RO3G_013156RO3G_015099RO3G_016094RO3G_001805RO3G_012212RO3G_016224RO3G_013149RO3G_013907RO3G_002919RO3G_006665RO3G_008152RO3G_017708RO3G_011423RO3G_000510RO3G_008857RO3G_010179RO3G_009912RO3G_011163RO3G_014293RO3G_002144RO3G_012392RO3G_013186RO3G_015177RO3G_007078RO3G_004745RO3G_002225RO3G_010776RO3G_011311RO3G_012146RO3G_007059RO3G_001638RO3G_015402RO3G_012141RO3G_002645RO3G_001495RO3G_004335RO3G_014240RO3G_003409RO3G_005383RO3G_005025RO3G_012099RO3G_002114RO3G_010030RO3G_013155RO3G_008154RO3G_016768RO3G_010268RO3G_016502RO3G_002646RO3G_001885RO3G_007043RO3G_007044RO3G_012437RO3G_003486RO3G_017722RO3G_008697RO3G_015351RO3G_006982RO3G_017430RO3G_006425RO3G_009516RO3G_007777RO3G_001240RO3G_016150RO3G_006183RO3G_015040RO3G_008908RO3G_009422RO3G_014548RO3G_008886RO3G_004975RO3G_007034RO3G_016043RO3G_012756RO3G_016148RO3G_008379RO3G_005509RO3G_016850RO3G_013299RO3G_013178RO3G_010860RO3G_001799RO3G_012475RO3G_011384RO3G_012686RO3G_002316RO3G_001238RO3G_003054RO3G_015958RO3G_016366RO3G_012410RO3G_007829RO3G_013247RO3G_014707RO3G_011128RO3G_016278RO3G_002745RO3G_016154RO3G_001942RO3G_013634RO3G_012012RO3G_001341RO3G_001819RO3G_007524RO3G_012938RO3G_005625RO3G_016115RO3G_004822RO3G_008353RO3G_006080RO3G_009473RO3G_004098RO3G_014708RO3G_002969RO3G_016466RO3G_013614RO3G_000982RO3G_005795RO3G_001893RO3G_009710RO3G_008346RO3G_013099RO3G_009296RO3G_016813RO3G_009355RO3G_016537RO3G_017027RO3G_002154RO3G_011984RO3G_005446RO3G_008122RO3G_015697RO3G_003645RO3G_002146RO3G_016539RO3G_016538RO3G_009331RO3G_011844RO3G_001745RO3G_007362RO3G_016680RO3G_016462RO3G_001256RO3G_014719RO3G_003035RO3G_003141RO3G_008817RO3G_015801RO3G_006133RO3G_009645RO3G_012784RO3G_015550RO3G_001342RO3G_005633RO3G_009086RO3G_011731RO3G_005073RO3G_009472RO3G_009622RO3G_015882RO3G_012943RO3G_010719RO3G_012442RO3G_013838RO3G_013566RO3G_003676RO3G_012326RO3G_016681RO3G_016575RO3G_006424RO3G_005283RO3G_007525RO3G_011564RO3G_012844RO3G_006384RO3G_003036RO3G_016781RO3G_004107RO3G_002960RO3G_011388RO3G_012785RO3G_007081RO3G_002240RO3G_013176RO3G_010720
samples vs. features diffExpr.P0.001_C2.matrix.log2.centered
−6 −2 2 6Value
Color Key
LogFC
With MacrophagesWithout Macrophages
R. delemar curedMacrophages + R. delemar cured
Macrophages + R. delemar WTR. delemar WTcond_3_rep2
cond_3_rep1
cond_1_rep3
cond_1_rep1
cond_1_rep2
cond_3_rep3
cond_0_rep2
cond_2_rep1
cond_0_rep3
cond_0_rep1
cond_2_rep3
cond_2_rep2
RO3G_005875RO3G_002036RO3G_009513RO3G_007365RO3G_006644RO3G_016651RO3G_001550RO3G_015694RO3G_009424RO3G_012739RO3G_016465RO3G_015312RO3G_009281RO3G_000465RO3G_003906RO3G_016415RO3G_001012RO3G_016742RO3G_017422RO3G_009519RO3G_013940RO3G_014198RO3G_012571RO3G_013538RO3G_005645RO3G_000866RO3G_007382RO3G_014380RO3G_006165RO3G_006326RO3G_011802RO3G_001062RO3G_008322RO3G_010934RO3G_007683RO3G_012567RO3G_009678RO3G_005635RO3G_017389RO3G_005396RO3G_015241RO3G_006584RO3G_016752RO3G_014734RO3G_010189RO3G_014933RO3G_006623RO3G_014451RO3G_001190RO3G_011698RO3G_007377RO3G_014467RO3G_007769RO3G_014471RO3G_011512RO3G_010037RO3G_016857RO3G_008711RO3G_000900RO3G_000949RO3G_014501RO3G_003598RO3G_004193RO3G_014305RO3G_016120RO3G_010417RO3G_005315RO3G_007433RO3G_016212RO3G_008411RO3G_014396RO3G_015861RO3G_011864RO3G_001309RO3G_009644RO3G_013422RO3G_004367RO3G_009836RO3G_007558RO3G_009004RO3G_001510RO3G_006479RO3G_008489RO3G_001310RO3G_015620RO3G_002493RO3G_013882RO3G_012320RO3G_017274RO3G_006358RO3G_012780RO3G_016571RO3G_003922RO3G_009949RO3G_011212RO3G_010607RO3G_009742RO3G_011162RO3G_002323RO3G_010829RO3G_000954RO3G_002500RO3G_004683RO3G_008249RO3G_006968RO3G_017547RO3G_003102RO3G_000748RO3G_001824RO3G_016254RO3G_003106RO3G_013722RO3G_017160RO3G_009881RO3G_013380RO3G_010796RO3G_002214RO3G_009506RO3G_000908RO3G_008048RO3G_008370RO3G_000468RO3G_012203RO3G_015628RO3G_004544RO3G_003695RO3G_001423RO3G_010990RO3G_006780RO3G_003602RO3G_015213RO3G_004678RO3G_005437RO3G_014646RO3G_005277RO3G_017052RO3G_005059RO3G_014327RO3G_013872RO3G_014340RO3G_011832RO3G_001923RO3G_003091RO3G_014401RO3G_011172RO3G_010903RO3G_005470RO3G_002901RO3G_011817RO3G_003862RO3G_016107RO3G_004721RO3G_006891RO3G_008545RO3G_006894RO3G_015812RO3G_016843RO3G_014481RO3G_001557RO3G_002536RO3G_009159RO3G_008028RO3G_008578RO3G_011184RO3G_004001RO3G_006206RO3G_008727RO3G_002010RO3G_004555RO3G_010673RO3G_013999RO3G_001769RO3G_014851RO3G_005588RO3G_005148RO3G_016300RO3G_012201RO3G_017376RO3G_014162RO3G_001432RO3G_001476RO3G_005274RO3G_006645RO3G_011183RO3G_005137RO3G_013060RO3G_016089RO3G_008500RO3G_007795RO3G_014057RO3G_001097RO3G_014140RO3G_008818RO3G_001707RO3G_016207RO3G_015442RO3G_016268RO3G_006615RO3G_015449RO3G_012452RO3G_009658RO3G_007835RO3G_015881RO3G_003009RO3G_017703RO3G_009187RO3G_016195RO3G_007400RO3G_005904RO3G_006119RO3G_011533RO3G_008558RO3G_016939RO3G_016704RO3G_015490RO3G_013446RO3G_001677RO3G_014971RO3G_007723RO3G_000974RO3G_015579RO3G_013542RO3G_014650RO3G_010433RO3G_005008RO3G_006318RO3G_005899RO3G_000431RO3G_001255RO3G_013281RO3G_011928RO3G_011987RO3G_016325RO3G_013995RO3G_010187RO3G_011399RO3G_011754RO3G_005708RO3G_010002RO3G_013746RO3G_015170RO3G_006145RO3G_007825RO3G_000386RO3G_014298RO3G_012662RO3G_011989RO3G_008929RO3G_005089RO3G_011966RO3G_008351RO3G_012076RO3G_000988RO3G_013888RO3G_005037RO3G_011806RO3G_003166RO3G_004174RO3G_003605RO3G_010132RO3G_002307RO3G_001063RO3G_003293RO3G_003618RO3G_008995RO3G_002456RO3G_012572RO3G_003867RO3G_001808RO3G_005447RO3G_014874RO3G_015798RO3G_009128RO3G_006189RO3G_008227RO3G_015889RO3G_006100RO3G_000266RO3G_000017RO3G_004981RO3G_000249RO3G_013469RO3G_004160RO3G_013571RO3G_016889RO3G_012981RO3G_006622RO3G_003859RO3G_000984RO3G_013624RO3G_004086RO3G_009343RO3G_003912RO3G_013623RO3G_002325RO3G_002533RO3G_011448RO3G_009889RO3G_004813RO3G_009414RO3G_017006RO3G_004010RO3G_007225RO3G_009541RO3G_017073RO3G_000704RO3G_004144RO3G_011905RO3G_017443RO3G_007718RO3G_002767RO3G_004748RO3G_001481RO3G_004138RO3G_005359RO3G_014419RO3G_000874RO3G_001929RO3G_006646RO3G_008103RO3G_010057RO3G_009932RO3G_015347RO3G_011507RO3G_007366RO3G_015504RO3G_009920RO3G_016769RO3G_006128RO3G_015028RO3G_003075RO3G_014724RO3G_002169RO3G_002326RO3G_015779RO3G_005912RO3G_004858RO3G_001951RO3G_008696RO3G_010122RO3G_002747RO3G_007856RO3G_014412RO3G_013534RO3G_009509RO3G_004009RO3G_010098RO3G_001675RO3G_003232RO3G_001224RO3G_004400RO3G_012565RO3G_001525RO3G_005384RO3G_011454RO3G_014441RO3G_008478RO3G_008389RO3G_010257RO3G_015439RO3G_011530RO3G_002806RO3G_009277RO3G_006429RO3G_001179RO3G_007722RO3G_005278RO3G_011400RO3G_007989RO3G_000696RO3G_007013RO3G_000223RO3G_011670RO3G_009007RO3G_004047RO3G_008047RO3G_013858RO3G_017846RO3G_008470RO3G_009453RO3G_001380RO3G_006163RO3G_002990RO3G_007772RO3G_014309RO3G_013251RO3G_017741RO3G_008045RO3G_017756RO3G_003030RO3G_012535RO3G_003256RO3G_002353RO3G_011859RO3G_005015RO3G_004934RO3G_001371RO3G_002497RO3G_002551RO3G_008625RO3G_012790RO3G_003575RO3G_006271RO3G_010000RO3G_017357RO3G_013444RO3G_012124RO3G_011914RO3G_003683RO3G_004982RO3G_006439RO3G_011972RO3G_007258RO3G_014363RO3G_014525RO3G_006507RO3G_000556RO3G_014119RO3G_016237RO3G_016217RO3G_004957RO3G_008902RO3G_017162RO3G_013541RO3G_002095RO3G_008376RO3G_014347RO3G_016149RO3G_011814RO3G_008492RO3G_001091RO3G_005980RO3G_012645RO3G_001314RO3G_013717RO3G_010669RO3G_006731RO3G_007227RO3G_016162RO3G_010964RO3G_003596RO3G_016488RO3G_006225RO3G_013385RO3G_013353RO3G_012385RO3G_004015RO3G_016299RO3G_008972RO3G_007545RO3G_007954RO3G_017167RO3G_005914RO3G_010693RO3G_004597RO3G_016193RO3G_006883RO3G_014166RO3G_011548RO3G_011427RO3G_010988RO3G_016321RO3G_004457RO3G_003077RO3G_001223RO3G_012700RO3G_000187RO3G_014713RO3G_013910RO3G_000464RO3G_011685RO3G_007738RO3G_016298RO3G_008394RO3G_001933RO3G_014920RO3G_010667RO3G_001755RO3G_006363RO3G_012755RO3G_002327RO3G_013896RO3G_006023RO3G_012388RO3G_010991RO3G_008836RO3G_009649RO3G_000644RO3G_007864RO3G_000452RO3G_010159RO3G_011809RO3G_011841RO3G_007086RO3G_006980RO3G_007845RO3G_014553RO3G_004842RO3G_005698RO3G_008809RO3G_011204RO3G_014335RO3G_007727RO3G_005024RO3G_005620RO3G_004033RO3G_005583RO3G_008764RO3G_002632RO3G_007945RO3G_004168RO3G_001758RO3G_011273RO3G_011942RO3G_007531RO3G_009294RO3G_009193RO3G_006474RO3G_005407RO3G_002879RO3G_007556RO3G_006800RO3G_000854RO3G_005724RO3G_006463RO3G_014002RO3G_005670RO3G_000922RO3G_002907RO3G_012218RO3G_007191RO3G_011340RO3G_003411RO3G_010664RO3G_009140RO3G_016206RO3G_004715RO3G_003289RO3G_007124RO3G_017079RO3G_003877RO3G_014136RO3G_013777RO3G_017712RO3G_011406RO3G_016873RO3G_007480RO3G_012573RO3G_014403RO3G_010685RO3G_016572RO3G_015340RO3G_015649RO3G_004456RO3G_001085RO3G_010660RO3G_008793RO3G_011937RO3G_017408RO3G_004333RO3G_012764RO3G_017062RO3G_014678RO3G_010405RO3G_014206RO3G_009647RO3G_016187RO3G_015555RO3G_005669RO3G_006285RO3G_010114RO3G_000687RO3G_013157RO3G_016741RO3G_009222RO3G_006742RO3G_014319RO3G_002009RO3G_007661RO3G_010407RO3G_006743RO3G_001812RO3G_014048RO3G_013232RO3G_007287RO3G_001970RO3G_010548RO3G_000222RO3G_007364RO3G_003664RO3G_005193RO3G_000537RO3G_013087RO3G_017131RO3G_005147RO3G_013309RO3G_013495RO3G_006113RO3G_009253RO3G_013889RO3G_004963RO3G_017103RO3G_008250RO3G_017242RO3G_013773RO3G_005041RO3G_007449RO3G_015884RO3G_009272RO3G_008537RO3G_000819RO3G_015359RO3G_009549RO3G_001753RO3G_001723RO3G_013068RO3G_009664RO3G_007546RO3G_009275RO3G_016194RO3G_011558RO3G_017766RO3G_008932RO3G_002312RO3G_012545RO3G_005935RO3G_014210RO3G_000388RO3G_005702RO3G_003025RO3G_006604RO3G_017698RO3G_016176RO3G_002790RO3G_001488RO3G_000910RO3G_015687RO3G_016617RO3G_011138RO3G_001142RO3G_003285RO3G_010311RO3G_012269RO3G_011700RO3G_013748RO3G_008974RO3G_004995RO3G_000505RO3G_005420RO3G_011417RO3G_016524RO3G_001961RO3G_017352RO3G_005890RO3G_014186RO3G_001847RO3G_007277RO3G_002479RO3G_011470RO3G_015097RO3G_006016RO3G_014906RO3G_010102RO3G_010840RO3G_014517RO3G_002047RO3G_006505RO3G_017234RO3G_015259RO3G_010310RO3G_008416RO3G_005320RO3G_009931RO3G_015426RO3G_015156RO3G_001350RO3G_017371RO3G_001232RO3G_006500RO3G_010124RO3G_012729RO3G_008157RO3G_014054RO3G_008252RO3G_015758RO3G_012211RO3G_004249RO3G_017259RO3G_001635RO3G_004742RO3G_010792RO3G_012942RO3G_002720RO3G_016788RO3G_001553RO3G_008577RO3G_003398RO3G_000871RO3G_001754RO3G_007495RO3G_000490RO3G_006970RO3G_007765RO3G_011762RO3G_006913RO3G_002909RO3G_008963RO3G_002635RO3G_004800RO3G_000686RO3G_011240RO3G_016740RO3G_011178RO3G_011125RO3G_015567RO3G_008114RO3G_007544RO3G_012445RO3G_003610RO3G_002502RO3G_010986RO3G_004225RO3G_015425RO3G_012893RO3G_008121RO3G_004079RO3G_015074RO3G_004979RO3G_008635RO3G_016771RO3G_008151RO3G_002942RO3G_014892RO3G_006403RO3G_007775RO3G_003043RO3G_010608RO3G_011152RO3G_004167RO3G_012852RO3G_012705RO3G_012008RO3G_012685RO3G_012444RO3G_003569RO3G_000860RO3G_001544RO3G_003288RO3G_008486RO3G_010604RO3G_017177RO3G_001154RO3G_017833RO3G_001571RO3G_000394RO3G_011438RO3G_010827RO3G_006159RO3G_004082RO3G_000257RO3G_013314RO3G_003504RO3G_000868RO3G_010708RO3G_011324RO3G_016620RO3G_003168RO3G_001116RO3G_017087RO3G_007333RO3G_015024RO3G_005124RO3G_009005RO3G_010144RO3G_003337RO3G_004821RO3G_001656RO3G_009999RO3G_001509RO3G_011993RO3G_004027RO3G_006575RO3G_016927RO3G_010895RO3G_005573RO3G_014764RO3G_009878RO3G_008089RO3G_013338RO3G_006349RO3G_010426RO3G_001759RO3G_002499RO3G_012894RO3G_003055RO3G_004198RO3G_012551RO3G_009952RO3G_008536RO3G_011858RO3G_009157RO3G_014189RO3G_012511RO3G_010452RO3G_010182RO3G_003017RO3G_003172RO3G_011445RO3G_016159RO3G_013390RO3G_007966RO3G_013408RO3G_009011RO3G_010286RO3G_014968RO3G_000882RO3G_002444RO3G_008922RO3G_005219RO3G_010931RO3G_007508RO3G_001562RO3G_002910RO3G_011658RO3G_009095RO3G_008651RO3G_003499RO3G_005011RO3G_000518RO3G_005751RO3G_007489RO3G_008585RO3G_016320RO3G_007060RO3G_011437RO3G_002014RO3G_012819RO3G_006495RO3G_016464RO3G_013447RO3G_011396RO3G_011948RO3G_000839RO3G_008076RO3G_004186RO3G_010127RO3G_001181RO3G_013729RO3G_015740RO3G_013243RO3G_017320RO3G_001508RO3G_013557RO3G_002055RO3G_008794RO3G_006878RO3G_008232RO3G_004825RO3G_011970RO3G_001138RO3G_000171RO3G_017717RO3G_006105RO3G_001092RO3G_000146RO3G_002213RO3G_005296RO3G_016658RO3G_007376RO3G_006198RO3G_007645RO3G_002553RO3G_003174RO3G_010170RO3G_011503RO3G_011695RO3G_011025RO3G_016345RO3G_005023RO3G_014074RO3G_001822RO3G_006468RO3G_010615RO3G_014753RO3G_008308RO3G_011664RO3G_014051RO3G_008646RO3G_010984RO3G_007288RO3G_000966RO3G_017824RO3G_016921RO3G_012831RO3G_003564RO3G_001746RO3G_005063RO3G_015940RO3G_012096RO3G_003653RO3G_009223RO3G_011260RO3G_005319RO3G_012904RO3G_007091RO3G_007725RO3G_013202RO3G_004219RO3G_009504RO3G_013710RO3G_008235RO3G_010418RO3G_014418RO3G_013770RO3G_000667RO3G_012653RO3G_014397RO3G_015871RO3G_005102RO3G_008712RO3G_011201RO3G_005285RO3G_000231RO3G_010948RO3G_015080RO3G_017166RO3G_004814RO3G_001005RO3G_013241RO3G_000391RO3G_008728RO3G_007866RO3G_005185RO3G_016416RO3G_004159RO3G_003119RO3G_005839RO3G_005364RO3G_010054RO3G_016851RO3G_013744RO3G_013630RO3G_009609RO3G_014292RO3G_006453RO3G_001552RO3G_012161RO3G_006661RO3G_013067RO3G_017294RO3G_003394RO3G_013787RO3G_005640RO3G_001928RO3G_011153RO3G_003228RO3G_005639RO3G_001228RO3G_014769RO3G_011156RO3G_017777RO3G_015650RO3G_003147RO3G_011136RO3G_007103RO3G_008241RO3G_007211RO3G_005379RO3G_015014RO3G_007327RO3G_014430RO3G_014429RO3G_005721RO3G_000834RO3G_003066RO3G_001777RO3G_004430RO3G_009505RO3G_001501RO3G_015145RO3G_001215RO3G_012462RO3G_014255RO3G_001559RO3G_008094RO3G_007600RO3G_005131RO3G_015094RO3G_014771RO3G_014630RO3G_009655RO3G_000853RO3G_011930RO3G_009613RO3G_002752RO3G_011397RO3G_001844RO3G_002562RO3G_013326RO3G_008628RO3G_005425RO3G_010691RO3G_015100RO3G_004109RO3G_009837RO3G_010012RO3G_010682RO3G_000790RO3G_014410RO3G_009190RO3G_013369RO3G_012314RO3G_000901RO3G_001186RO3G_005861RO3G_009240RO3G_010920RO3G_011746RO3G_000224RO3G_006496RO3G_007841RO3G_008044RO3G_001927RO3G_006313RO3G_008172RO3G_008526RO3G_008849RO3G_011251RO3G_016938RO3G_011506RO3G_010035RO3G_014072RO3G_005438RO3G_005098RO3G_009324RO3G_013078RO3G_008532RO3G_000173RO3G_009282RO3G_000963RO3G_006193RO3G_002045RO3G_004189RO3G_017831RO3G_014073RO3G_016000RO3G_008083RO3G_017161RO3G_004245RO3G_004952RO3G_010921RO3G_001102RO3G_003448RO3G_016023RO3G_006306RO3G_011630RO3G_009738RO3G_006110RO3G_004366RO3G_009524RO3G_010238RO3G_006352RO3G_005777RO3G_006106RO3G_013188RO3G_016208RO3G_003038RO3G_001894RO3G_006975RO3G_010915RO3G_011741RO3G_006307RO3G_007215RO3G_016286RO3G_008544RO3G_008944RO3G_004515RO3G_013041RO3G_010477RO3G_012848RO3G_008474RO3G_009245RO3G_016647RO3G_010139RO3G_015794RO3G_002309RO3G_002416RO3G_016312RO3G_012626RO3G_012226RO3G_011733RO3G_011484RO3G_012549RO3G_009743RO3G_012765RO3G_017765RO3G_005924RO3G_007375RO3G_001486RO3G_004520RO3G_008528RO3G_001827RO3G_017286RO3G_016072RO3G_002200RO3G_010221RO3G_005838RO3G_004584RO3G_011359RO3G_007161RO3G_007005RO3G_010613RO3G_017776RO3G_016819RO3G_003472RO3G_006435RO3G_013330RO3G_017737RO3G_002474RO3G_009725RO3G_016598RO3G_002431RO3G_001493RO3G_011451RO3G_008032RO3G_006256RO3G_006718RO3G_015440RO3G_010018RO3G_002191RO3G_017832RO3G_006890RO3G_004874RO3G_002887RO3G_015019RO3G_006195RO3G_001569RO3G_017769RO3G_011373RO3G_006269RO3G_007684RO3G_003040RO3G_005931RO3G_011258RO3G_013849RO3G_004740RO3G_008409RO3G_002175RO3G_002233RO3G_006156RO3G_012068RO3G_003380RO3G_007757RO3G_006281RO3G_006038RO3G_008885RO3G_005123RO3G_007633RO3G_005740RO3G_006386RO3G_010500RO3G_008343RO3G_003844RO3G_015444RO3G_006998RO3G_010454RO3G_014214RO3G_011965RO3G_012493RO3G_010851RO3G_004729RO3G_016593RO3G_013001RO3G_006902RO3G_017009RO3G_013237RO3G_007010RO3G_012394RO3G_007990RO3G_001144RO3G_004605RO3G_008358RO3G_014967RO3G_009429RO3G_014907RO3G_014116RO3G_016587RO3G_012988RO3G_015262RO3G_011953RO3G_016164RO3G_008879RO3G_017301RO3G_014008RO3G_007687RO3G_009322RO3G_012906RO3G_011992RO3G_012897RO3G_014433RO3G_015285RO3G_007394RO3G_016691RO3G_001305RO3G_015026RO3G_004687RO3G_007250RO3G_002824RO3G_014228RO3G_001308RO3G_017074RO3G_008731RO3G_010038RO3G_000971RO3G_017462RO3G_010467RO3G_013878RO3G_008057RO3G_016709RO3G_013415RO3G_010123RO3G_014522RO3G_007787RO3G_012993RO3G_004232RO3G_012131RO3G_010095RO3G_011031RO3G_003652RO3G_006459RO3G_017159RO3G_004606RO3G_008647RO3G_001431RO3G_015168RO3G_008754RO3G_008109RO3G_005211RO3G_003866RO3G_012383RO3G_015452RO3G_017382RO3G_010844RO3G_002322RO3G_009616RO3G_011268RO3G_009958RO3G_005919RO3G_004935RO3G_009528RO3G_015781RO3G_005478RO3G_010596RO3G_013570RO3G_010294RO3G_015380RO3G_013578RO3G_005832RO3G_003042RO3G_004165RO3G_000844RO3G_001339RO3G_013329RO3G_013636RO3G_016573RO3G_014739RO3G_006182RO3G_016621RO3G_010224RO3G_011170RO3G_000228RO3G_004488RO3G_011155RO3G_008899RO3G_010305RO3G_006512RO3G_010431RO3G_004552RO3G_003343RO3G_013040RO3G_001344RO3G_001574RO3G_009078RO3G_012722RO3G_015811RO3G_013481RO3G_012840RO3G_014652RO3G_013336RO3G_010710RO3G_013360RO3G_003505RO3G_008004RO3G_015691RO3G_005152RO3G_002904RO3G_011459RO3G_007780RO3G_014176RO3G_012090RO3G_008717RO3G_012004RO3G_001122RO3G_006670RO3G_014006RO3G_013264RO3G_007270RO3G_009507RO3G_000504RO3G_015883RO3G_001067RO3G_003393RO3G_009598RO3G_001280RO3G_005755RO3G_016240RO3G_013074RO3G_010173RO3G_012023RO3G_004040RO3G_001252RO3G_000622RO3G_003383RO3G_003342RO3G_012001RO3G_017469RO3G_006028RO3G_002426RO3G_006151RO3G_011724RO3G_010556RO3G_007771RO3G_005836RO3G_005268RO3G_010126RO3G_011946RO3G_001926RO3G_008828RO3G_008734RO3G_002428RO3G_002983RO3G_014890RO3G_010315RO3G_003132RO3G_014205RO3G_003095RO3G_011629RO3G_005727RO3G_010788RO3G_013398RO3G_009150RO3G_002501RO3G_006360RO3G_007354RO3G_011455RO3G_013903RO3G_005668RO3G_002992RO3G_009241RO3G_014888RO3G_009954RO3G_017793RO3G_013494RO3G_003426RO3G_009953RO3G_008584RO3G_003392RO3G_010950RO3G_011379RO3G_015865RO3G_000466RO3G_004598RO3G_009246RO3G_003869RO3G_015559RO3G_004530RO3G_015568RO3G_007740RO3G_016738RO3G_001624RO3G_001219RO3G_007952RO3G_010307RO3G_011247RO3G_003407RO3G_007118RO3G_007069RO3G_012009RO3G_009595RO3G_006146RO3G_013257RO3G_007331RO3G_001560RO3G_014492RO3G_011213RO3G_009946RO3G_015258RO3G_011131RO3G_004095RO3G_010223RO3G_002057RO3G_007534RO3G_010235RO3G_017841RO3G_009593RO3G_000923RO3G_001216RO3G_003227RO3G_004829RO3G_010181RO3G_007319RO3G_004433RO3G_012664RO3G_008914RO3G_014171RO3G_008234RO3G_005929RO3G_004839RO3G_015298RO3G_007981RO3G_011449RO3G_005783RO3G_011319RO3G_009342RO3G_016113RO3G_006168RO3G_003103RO3G_012766RO3G_003415RO3G_015578RO3G_012016RO3G_005275RO3G_002212RO3G_009851RO3G_016626RO3G_001704RO3G_009997RO3G_007628RO3G_008412RO3G_013998RO3G_010292RO3G_016874RO3G_002755RO3G_002883RO3G_008313RO3G_005445RO3G_016791RO3G_014359RO3G_003028RO3G_014453RO3G_007152RO3G_011252RO3G_015634RO3G_017796RO3G_002937RO3G_015060RO3G_008598RO3G_012136RO3G_012195RO3G_013154RO3G_014004RO3G_017031RO3G_008371RO3G_001830RO3G_005672RO3G_010085RO3G_012449RO3G_014321RO3G_006616RO3G_008469RO3G_005067RO3G_002260RO3G_017706RO3G_012683RO3G_009736RO3G_010968RO3G_017150RO3G_017791RO3G_006041RO3G_002542RO3G_012282RO3G_012283RO3G_008248RO3G_015189RO3G_005614RO3G_003728RO3G_011019RO3G_011023RO3G_002008RO3G_004581RO3G_010011RO3G_000960RO3G_003300RO3G_014026RO3G_017351RO3G_004685RO3G_001315RO3G_017709RO3G_003853RO3G_003142RO3G_000840RO3G_010975RO3G_001828RO3G_004917RO3G_005189RO3G_012891RO3G_014716RO3G_015186RO3G_002905RO3G_013968RO3G_004556RO3G_000951RO3G_001364RO3G_017411RO3G_016172RO3G_012842RO3G_004856RO3G_015257RO3G_005307RO3G_012513RO3G_000711RO3G_009205RO3G_005075RO3G_013846RO3G_017453RO3G_000909RO3G_017114RO3G_008824RO3G_010972RO3G_012169RO3G_003155RO3G_000123RO3G_008515RO3G_002886RO3G_016061RO3G_017640RO3G_012876RO3G_008945RO3G_002152RO3G_012715RO3G_000393RO3G_014168RO3G_008237RO3G_003027RO3G_011434RO3G_001632RO3G_001862RO3G_000631RO3G_014204RO3G_009252RO3G_012999RO3G_006993RO3G_005306RO3G_004196RO3G_001221RO3G_006287RO3G_014465RO3G_014324RO3G_007984RO3G_016257RO3G_015762RO3G_009568RO3G_013859RO3G_006292RO3G_002707RO3G_009897RO3G_001924RO3G_011488RO3G_005256RO3G_002215RO3G_013327RO3G_004460RO3G_008397RO3G_017461RO3G_015243RO3G_003169RO3G_013546RO3G_006629RO3G_012913RO3G_010574RO3G_001876RO3G_010435RO3G_009437RO3G_005144RO3G_014045RO3G_010415RO3G_005058RO3G_004459RO3G_004722RO3G_004689RO3G_000757RO3G_011414RO3G_012528RO3G_013911RO3G_008806RO3G_016698RO3G_011022RO3G_011412RO3G_005631RO3G_009349RO3G_003108RO3G_007485RO3G_011132RO3G_011981RO3G_013387RO3G_016355RO3G_007812RO3G_005634RO3G_006081RO3G_012566RO3G_000359RO3G_012716RO3G_001940RO3G_012290RO3G_013071RO3G_008951RO3G_004464RO3G_014682RO3G_009544RO3G_007105RO3G_000201RO3G_013559RO3G_013548RO3G_010149RO3G_004438RO3G_012687RO3G_001003RO3G_015108RO3G_011843RO3G_016902RO3G_007302RO3G_008749RO3G_015849RO3G_005643RO3G_015051RO3G_017253RO3G_006503RO3G_000852RO3G_010160RO3G_010800RO3G_006004RO3G_010510RO3G_002467RO3G_016689RO3G_002659RO3G_005512RO3G_007879RO3G_015879RO3G_006448RO3G_017812RO3G_012235RO3G_003404RO3G_017069RO3G_011277RO3G_010949RO3G_009992RO3G_005959RO3G_009554RO3G_004432RO3G_014061RO3G_006196RO3G_005213RO3G_010919RO3G_013275RO3G_009628RO3G_007272RO3G_014039RO3G_016469RO3G_007876RO3G_003760RO3G_016736RO3G_010589RO3G_005336RO3G_008391RO3G_003579RO3G_000108RO3G_001225RO3G_003868RO3G_014867RO3G_014019RO3G_017282RO3G_009423RO3G_015301RO3G_001688RO3G_005954RO3G_012381RO3G_005292RO3G_007129RO3G_017315RO3G_011344RO3G_004134RO3G_008767RO3G_007991RO3G_006498RO3G_001222RO3G_014667RO3G_016075RO3G_016585RO3G_011916RO3G_008766RO3G_016903RO3G_016733RO3G_016451RO3G_011134RO3G_003876RO3G_002800RO3G_010203RO3G_002846RO3G_011300RO3G_014355RO3G_012935RO3G_012123RO3G_012810RO3G_008656RO3G_000893RO3G_017265RO3G_007648RO3G_005504RO3G_003969RO3G_001185RO3G_013175RO3G_008715RO3G_016124RO3G_010300RO3G_012193RO3G_017060RO3G_000196RO3G_009739RO3G_003533RO3G_014449RO3G_005604RO3G_016528RO3G_003874RO3G_015932RO3G_011462RO3G_003430RO3G_000248RO3G_011001RO3G_006048RO3G_007427RO3G_016083RO3G_005497RO3G_002131RO3G_008602RO3G_003578RO3G_005716RO3G_005385RO3G_004490RO3G_002181RO3G_000243RO3G_017090RO3G_012520RO3G_008239RO3G_004495RO3G_003083RO3G_014745RO3G_017721RO3G_014861RO3G_017434RO3G_009167RO3G_010443RO3G_013091RO3G_001196RO3G_017762RO3G_015093RO3G_004497RO3G_007796RO3G_005028RO3G_016290RO3G_002531RO3G_014755RO3G_005957RO3G_011034RO3G_005993RO3G_002046RO3G_002874RO3G_003233RO3G_001842RO3G_005402RO3G_011330RO3G_015790RO3G_006961RO3G_002993RO3G_000244RO3G_004695RO3G_012073RO3G_008398RO3G_012936RO3G_017891RO3G_016074RO3G_012402RO3G_016919RO3G_011127RO3G_007730RO3G_008681RO3G_003550RO3G_016011RO3G_016221RO3G_002661RO3G_001066RO3G_012407RO3G_010304RO3G_014081RO3G_003901RO3G_013847RO3G_009334RO3G_005876RO3G_016913RO3G_008541RO3G_002674RO3G_014929RO3G_002156RO3G_012837RO3G_015348RO3G_001874RO3G_012856RO3G_013266RO3G_006486RO3G_016500RO3G_014317RO3G_002450RO3G_002306RO3G_005048RO3G_005388RO3G_001644RO3G_000524RO3G_016367RO3G_006092RO3G_010985RO3G_014337RO3G_007421RO3G_004738RO3G_009196RO3G_017010RO3G_015336RO3G_008418RO3G_001001RO3G_004915RO3G_004734RO3G_014742RO3G_014837RO3G_008881RO3G_014385RO3G_010479RO3G_015417RO3G_017183RO3G_007484RO3G_006737RO3G_002186RO3G_015192RO3G_013575RO3G_013855RO3G_006738RO3G_003046RO3G_014349RO3G_015825RO3G_003348RO3G_002583RO3G_009623RO3G_016216RO3G_008066RO3G_002625RO3G_000930RO3G_011419RO3G_008657RO3G_002480RO3G_007709RO3G_009618RO3G_000890RO3G_015648RO3G_007817RO3G_004528RO3G_001214RO3G_009987RO3G_004595RO3G_015120RO3G_003599RO3G_016297RO3G_013179RO3G_009918RO3G_010270RO3G_009034RO3G_008791RO3G_011757RO3G_016283RO3G_017145RO3G_003930RO3G_013289RO3G_001935RO3G_009632RO3G_009493RO3G_008984RO3G_006981RO3G_016460RO3G_010707RO3G_005465RO3G_005982RO3G_004253RO3G_003332RO3G_005865RO3G_017272RO3G_001153RO3G_016614RO3G_013715RO3G_015320RO3G_013588RO3G_004968RO3G_002035RO3G_009332RO3G_009824RO3G_009823RO3G_017405RO3G_009363RO3G_012400RO3G_005831RO3G_010779RO3G_000539RO3G_008064RO3G_008040RO3G_006681RO3G_004532RO3G_000118RO3G_005623RO3G_009521RO3G_004534RO3G_004329RO3G_002140RO3G_009130RO3G_001558RO3G_008132RO3G_017644RO3G_004516RO3G_014017RO3G_009601RO3G_005880RO3G_009173RO3G_012278RO3G_004355RO3G_008327RO3G_009631RO3G_000327RO3G_002091RO3G_017816RO3G_017837RO3G_010736RO3G_003330RO3G_003682RO3G_007755RO3G_014344RO3G_004314RO3G_001117RO3G_001490RO3G_014969RO3G_015175RO3G_000976RO3G_016153RO3G_012457RO3G_015117RO3G_007186RO3G_004091RO3G_012239RO3G_013933RO3G_004526RO3G_011742RO3G_011560RO3G_013789RO3G_009295RO3G_008162RO3G_013983RO3G_005797RO3G_011792RO3G_011476RO3G_016282RO3G_012910RO3G_001869RO3G_017093RO3G_009484RO3G_007745RO3G_001115RO3G_006143RO3G_007523RO3G_006658RO3G_014696RO3G_016116RO3G_007284RO3G_017022RO3G_003134RO3G_003677RO3G_007504RO3G_003657RO3G_005587RO3G_008732RO3G_002816RO3G_011823RO3G_010483RO3G_000494RO3G_008665RO3G_011407RO3G_016122RO3G_015791RO3G_015322RO3G_003594RO3G_010883RO3G_012185RO3G_016040RO3G_015775RO3G_007702RO3G_008225RO3G_002823RO3G_015297RO3G_015317RO3G_003163RO3G_015793RO3G_015346RO3G_012066RO3G_015307RO3G_007882RO3G_009081RO3G_016067RO3G_007561RO3G_004921RO3G_013990RO3G_005222RO3G_015875RO3G_016666RO3G_008392RO3G_007058RO3G_014733RO3G_011413RO3G_004211RO3G_017012RO3G_010184RO3G_001515RO3G_002513RO3G_010562RO3G_009430RO3G_015806RO3G_015112RO3G_012458RO3G_006013RO3G_003052RO3G_005608RO3G_005618RO3G_009713RO3G_016898RO3G_005796RO3G_011006RO3G_003151RO3G_005857RO3G_014550RO3G_014912RO3G_013328RO3G_010129RO3G_015212RO3G_001529RO3G_014133RO3G_009293RO3G_004543RO3G_011643RO3G_010006RO3G_012081RO3G_016809RO3G_004804RO3G_002672RO3G_014503RO3G_012289RO3G_003700RO3G_003708RO3G_006668RO3G_009067RO3G_009064RO3G_011478RO3G_001220RO3G_013719RO3G_003647RO3G_016070RO3G_016610RO3G_009033RO3G_012802RO3G_003666RO3G_005939RO3G_000143RO3G_000446RO3G_017335RO3G_007158RO3G_001320RO3G_015591RO3G_012210RO3G_015551RO3G_014237RO3G_000191RO3G_016688RO3G_015161RO3G_008751RO3G_002436RO3G_017412RO3G_004990RO3G_017378RO3G_011644RO3G_004462RO3G_015563RO3G_013547RO3G_011716RO3G_012839RO3G_008001RO3G_008710RO3G_014992RO3G_013056RO3G_005981RO3G_011839RO3G_011144RO3G_008707RO3G_002896RO3G_002072RO3G_010231RO3G_002093RO3G_006341RO3G_005453RO3G_007706RO3G_009529RO3G_011494RO3G_015848RO3G_002903RO3G_015598RO3G_013035RO3G_006074RO3G_008390RO3G_009485RO3G_005305RO3G_016052RO3G_003679RO3G_011355RO3G_007318RO3G_005853RO3G_012071RO3G_012699RO3G_015390RO3G_005162RO3G_001178RO3G_007820RO3G_012430RO3G_005571RO3G_013431RO3G_004451RO3G_010229RO3G_004055RO3G_005518RO3G_009607RO3G_017418RO3G_003540RO3G_009138RO3G_009063RO3G_004202RO3G_004115RO3G_005077RO3G_017034RO3G_002463RO3G_011523RO3G_011165RO3G_002538RO3G_013288RO3G_000254RO3G_000883RO3G_017314RO3G_005748RO3G_014934RO3G_005597RO3G_005996RO3G_009556RO3G_008364RO3G_003439RO3G_012098RO3G_014049RO3G_012267RO3G_006953RO3G_004097RO3G_004178RO3G_004101RO3G_004251RO3G_009218RO3G_005047RO3G_004494RO3G_001160RO3G_010579RO3G_011038RO3G_002534RO3G_006409RO3G_001323RO3G_004491RO3G_003563RO3G_007770RO3G_006291RO3G_003705RO3G_011782RO3G_008357RO3G_000242RO3G_007744RO3G_013591RO3G_007822RO3G_012625RO3G_007356RO3G_010473RO3G_006096RO3G_010983RO3G_001261RO3G_001462RO3G_015991RO3G_008987RO3G_004162RO3G_009604RO3G_001187RO3G_006402RO3G_005488RO3G_011535RO3G_010185RO3G_003701RO3G_003702RO3G_001489RO3G_005594RO3G_007143RO3G_015743RO3G_000221RO3G_016220RO3G_016123RO3G_009537RO3G_008243RO3G_000475RO3G_016270RO3G_000477RO3G_000476RO3G_002217RO3G_005516RO3G_008024RO3G_002918RO3G_004356RO3G_005483RO3G_003617RO3G_003763RO3G_017100RO3G_003019RO3G_009495RO3G_002933RO3G_013852RO3G_002412RO3G_012013RO3G_009436RO3G_006427RO3G_003154RO3G_017122RO3G_005999RO3G_014996RO3G_002224RO3G_010981RO3G_015299RO3G_006027RO3G_008758RO3G_009669RO3G_016059RO3G_000434RO3G_012220RO3G_010175RO3G_000456RO3G_004092RO3G_007147RO3G_001161RO3G_006725RO3G_005197RO3G_005485RO3G_009715RO3G_004458RO3G_001377RO3G_005565RO3G_006803RO3G_012413RO3G_005582RO3G_003581RO3G_012287RO3G_009089RO3G_004087RO3G_013982RO3G_017423RO3G_009106RO3G_003872RO3G_002148RO3G_007159RO3G_009646RO3G_006406RO3G_009922RO3G_015989RO3G_014868RO3G_017048RO3G_001376RO3G_009249RO3G_000463RO3G_009091RO3G_005612RO3G_006723RO3G_009085RO3G_010711RO3G_016532RO3G_016152RO3G_010246RO3G_006032RO3G_006034RO3G_015655RO3G_001698RO3G_010785RO3G_001549RO3G_013156RO3G_015099RO3G_016094RO3G_001805RO3G_012212RO3G_016224RO3G_013149RO3G_013907RO3G_002919RO3G_006665RO3G_008152RO3G_017708RO3G_011423RO3G_000510RO3G_008857RO3G_010179RO3G_009912RO3G_011163RO3G_014293RO3G_002144RO3G_012392RO3G_013186RO3G_015177RO3G_007078RO3G_004745RO3G_002225RO3G_010776RO3G_011311RO3G_012146RO3G_007059RO3G_001638RO3G_015402RO3G_012141RO3G_002645RO3G_001495RO3G_004335RO3G_014240RO3G_003409RO3G_005383RO3G_005025RO3G_012099RO3G_002114RO3G_010030RO3G_013155RO3G_008154RO3G_016768RO3G_010268RO3G_016502RO3G_002646RO3G_001885RO3G_007043RO3G_007044RO3G_012437RO3G_003486RO3G_017722RO3G_008697RO3G_015351RO3G_006982RO3G_017430RO3G_006425RO3G_009516RO3G_007777RO3G_001240RO3G_016150RO3G_006183RO3G_015040RO3G_008908RO3G_009422RO3G_014548RO3G_008886RO3G_004975RO3G_007034RO3G_016043RO3G_012756RO3G_016148RO3G_008379RO3G_005509RO3G_016850RO3G_013299RO3G_013178RO3G_010860RO3G_001799RO3G_012475RO3G_011384RO3G_012686RO3G_002316RO3G_001238RO3G_003054RO3G_015958RO3G_016366RO3G_012410RO3G_007829RO3G_013247RO3G_014707RO3G_011128RO3G_016278RO3G_002745RO3G_016154RO3G_001942RO3G_013634RO3G_012012RO3G_001341RO3G_001819RO3G_007524RO3G_012938RO3G_005625RO3G_016115RO3G_004822RO3G_008353RO3G_006080RO3G_009473RO3G_004098RO3G_014708RO3G_002969RO3G_016466RO3G_013614RO3G_000982RO3G_005795RO3G_001893RO3G_009710RO3G_008346RO3G_013099RO3G_009296RO3G_016813RO3G_009355RO3G_016537RO3G_017027RO3G_002154RO3G_011984RO3G_005446RO3G_008122RO3G_015697RO3G_003645RO3G_002146RO3G_016539RO3G_016538RO3G_009331RO3G_011844RO3G_001745RO3G_007362RO3G_016680RO3G_016462RO3G_001256RO3G_014719RO3G_003035RO3G_003141RO3G_008817RO3G_015801RO3G_006133RO3G_009645RO3G_012784RO3G_015550RO3G_001342RO3G_005633RO3G_009086RO3G_011731RO3G_005073RO3G_009472RO3G_009622RO3G_015882RO3G_012943RO3G_010719RO3G_012442RO3G_013838RO3G_013566RO3G_003676RO3G_012326RO3G_016681RO3G_016575RO3G_006424RO3G_005283RO3G_007525RO3G_011564RO3G_012844RO3G_006384RO3G_003036RO3G_016781RO3G_004107RO3G_002960RO3G_011388RO3G_012785RO3G_007081RO3G_002240RO3G_013176RO3G_010720
−6 −2 2 6Value
Color Key
cond_0
cond_1
cond_2
cond_3
cond_2_rep1
cond_0_rep2
cond_0_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_1_rep1
cond_0_rep3
cond_1_rep2
cond_1_rep3
RHIMIDRAFT_283813
RHIMIDRAFT_245669
RHIMIDRAFT_242387
RHIMIDRAFT_96114
RHIMIDRAFT_248707
RHIMIDRAFT_196349
RHIMIDRAFT_197836
RHIMIDRAFT_234322
RHIMIDRAFT_99785
RHIMIDRAFT_314936
RHIMIDRAFT_232533
RHIMIDRAFT_245053
RHIMIDRAFT_290452
RHIMIDRAFT_294525
RHIMIDRAFT_307836
RHIMIDRAFT_288174
RHIMIDRAFT_251305
RHIMIDRAFT_307408
RHIMIDRAFT_267844
RHIMIDRAFT_198874
RHIMIDRAFT_79038
RHIMIDRAFT_240622
RHIMIDRAFT_289002
RHIMIDRAFT_270714
RHIMIDRAFT_253108
RHIMIDRAFT_281133
RHIMIDRAFT_257184
RHIMIDRAFT_239731
RHIMIDRAFT_87630
RHIMIDRAFT_138553
RHIMIDRAFT_272110
RHIMIDRAFT_288130
RHIMIDRAFT_203580
RHIMIDRAFT_313047
RHIMIDRAFT_221939
RHIMIDRAFT_252367
RHIMIDRAFT_271682
RHIMIDRAFT_252937
RHIMIDRAFT_195307
RHIMIDRAFT_138931
samples vs. features diffExpr.P0.001_C2.matrix.log2.centered
−4 0 2 4Value
Color Key
With Macrophages
Without Macrophages
R. microsporus curedMacrophages + R. microsporus cured
Macrophages + R. microsporus WTR. microsporus WT
A B
LogFC
Figure 2. Clustering of fungal transcriptional changes. A) Heatmap displaying significantly differentially expressed genes in R. delemar. Expression
levels are plotted in Log2, space and mean-centered (FDR < 0.001) B) Heatmap displaying significantly differentially expressed genes in R.
microsporus. Expression levels are plotted in Log2, space and mean-centered (FDR < 0.001). Biological replicates (n=3) are shown for each sample.
152
153
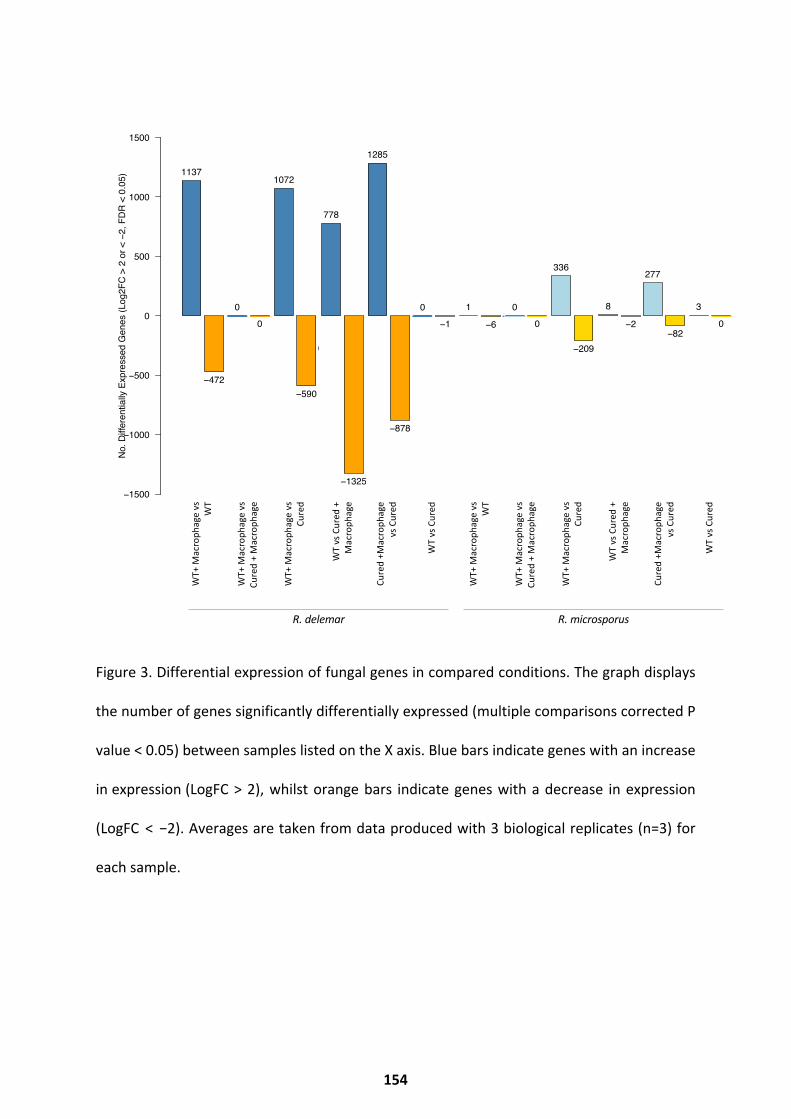
Figure 3. Differential expression of fungal genes in compared conditions. The graph displays
the number of genes significantly differentially expressed (multiple comparisons corrected P
value < 0.05) between samples listed on the X axis. Blue bars indicate genes with an increase
in expression (LogFC > 2), whilst orange bars indicate genes with a decrease in expression
(LogFC < −2). Averages are taken from data produced with 3 biological replicates (n=3) for
each sample.
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82D
EM v
DE
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar SamplesN
o. D
iffer
entia
lly E
xpre
ssed
Gen
es (L
og2F
C >
2 o
r < −
2, F
DR
< 0
.05)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
DEM
v D
E
DEM
v D
E
MEM
v M
E
MEM
v M
E
DEM
v D
M
DEM
v D
M
MEM
v M
M
MEM
v M
M
DEM
v D
DEM
v D
MEM
v M
MEM
v M
DE
v D
M
DE
v D
M
ME
v M
M
ME
v M
M
DE
v D
DE
v D
ME
v M
ME
v M
DM
v D
DM
v D
MM
v M
MM
v M
Differentially Expressed Genes from Pairwise Comparisons of R. microsporus and R. delemar Samples
No.
Diff
eren
tially
Exp
ress
ed G
enes
(Log
2FC
> 2
or <
−2,
FD
R <
0.0
5)
−1500
−1000
−500
0
500
1000
1500
1137
−472
1
−6
0
0
0
0
1072
−590
336
−209
778
−1325
8
−2
0
−1
3
0
1285
−878
277
−82
WT+
Mac
roph
age
vs
WT
WT+
Mac
roph
age
vs
Cure
d +
Mac
roph
age
WT+
Mac
roph
age
vs
Cure
d
Cure
d +M
acro
phag
e vs
Cur
ed
WT
vs C
ured
WT
vs C
ured
+
Mac
roph
age
WT+
Mac
roph
age
vs
WT
WT+
Mac
roph
age
vs
Cure
d +
Mac
roph
age
WT+
Mac
roph
age
vs
Cure
d
Cure
d +M
acro
phag
e vs
Cur
ed
WT
vs C
ured
WT
vs C
ured
+
Mac
roph
age
R. delemar R. microsporus
154
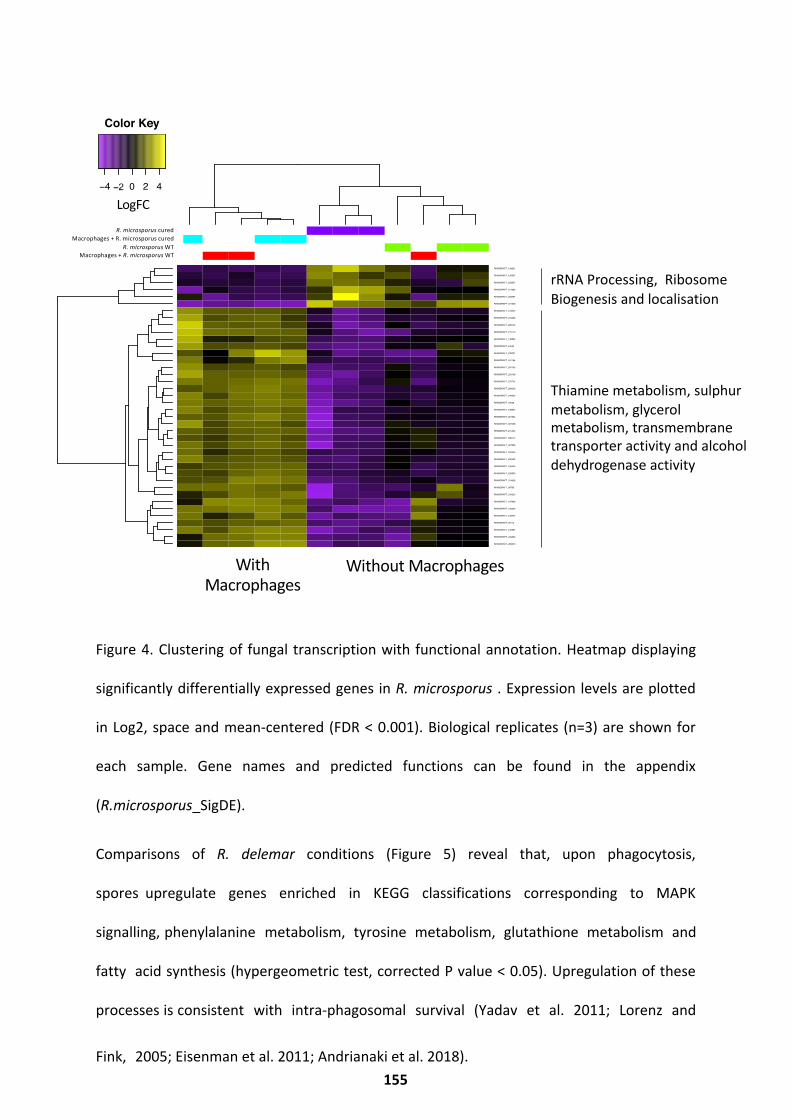
Figure 4. Clustering of fungal transcription with functional annotation. Heatmap displaying
significantly differentially expressed genes in R. microsporus . Expression levels are plotted
in Log2, space and mean-centered (FDR < 0.001). Biological replicates (n=3) are shown for
each sample. Gene names and predicted functions can be found in the appendix
(R.microsporus_SigDE).
Comparisons of R. delemar conditions (Figure 5) reveal that, upon phagocytosis,
spores upregulate genes enriched in KEGG classifications corresponding to MAPK
signalling, phenylalanine metabolism, tyrosine metabolism, glutathione metabolism and
fatty acid synthesis (hypergeometric test, corrected P value < 0.05). Upregulation of these
processes is consistent with intra-phagosomal survival (Yadav et al. 2011; Lorenz and
Fink, 2005; Eisenman et al. 2011; Andrianaki et al. 2018).
cond_0
cond_1
cond_2
cond_3
cond_2_rep1
cond_0_rep2
cond_0_rep1
cond_2_rep2
cond_2_rep3
cond_3_rep1
cond_3_rep2
cond_3_rep3
cond_1_rep1
cond_0_rep3
cond_1_rep2
cond_1_rep3
RHIMIDRAFT_283813
RHIMIDRAFT_245669
RHIMIDRAFT_242387
RHIMIDRAFT_96114
RHIMIDRAFT_248707
RHIMIDRAFT_196349
RHIMIDRAFT_197836
RHIMIDRAFT_234322
RHIMIDRAFT_99785
RHIMIDRAFT_314936
RHIMIDRAFT_232533
RHIMIDRAFT_245053
RHIMIDRAFT_290452
RHIMIDRAFT_294525
RHIMIDRAFT_307836
RHIMIDRAFT_288174
RHIMIDRAFT_251305
RHIMIDRAFT_307408
RHIMIDRAFT_267844
RHIMIDRAFT_198874
RHIMIDRAFT_79038
RHIMIDRAFT_240622
RHIMIDRAFT_289002
RHIMIDRAFT_270714
RHIMIDRAFT_253108
RHIMIDRAFT_281133
RHIMIDRAFT_257184
RHIMIDRAFT_239731
RHIMIDRAFT_87630
RHIMIDRAFT_138553
RHIMIDRAFT_272110
RHIMIDRAFT_288130
RHIMIDRAFT_203580
RHIMIDRAFT_313047
RHIMIDRAFT_221939
RHIMIDRAFT_252367
RHIMIDRAFT_271682
RHIMIDRAFT_252937
RHIMIDRAFT_195307
RHIMIDRAFT_138931
samples vs. features diffExpr.P0.001_C2.matrix.log2.centered
−4 0 2 4Value
Color Key
With Macrophages
Without Macrophages
R. microsporus curedMacrophages + R. microsporus cured
Macrophages + R. microsporus WTR. microsporus WT
LogFC
rRNA Processing, Ribosome Biogenesis and localisation
Thiamine metabolism, sulphurmetabolism, glycerol metabolism, transmembrane transporter activity and alcohol dehydrogenase activity
155

Unexposed R. delemar spores upregulate genes enriched in KEGG classifications
corresponding to ketone body synthesis, protein processing via the endoplasmic
reticulum, amino sugar and nucleotide sugar metabolism (hypergeometric test,
corrected P value < 0.05). This is consistent with metabolic activation and cell wall
biogenesis (Figure 5).
Figure 5. Functions of genes differentially expressed in R. delemar. Enriched KEGG categories
for the up/down regulated genes over sample comparisons (listed on the X axis). The
enrichment of the category is indicated by the colour bar. White corresponds to no
enrichment, and yellow to red corresponds to the given P value of the enrichment. Data
produced with 3 biological replicates (n=3) for each sample.
WT + M
acrophage vs WT (up)
WT + M
acrophage vs WT (down)
WT + M
acrophage vs Cured (up)
WT + M
acrophage vs Cured (down)
WT vs Cured + M
acrophage (up)
WT vs Cured + M
acrophage (down)
Cured + Macrophage vs Cured (up)
Cured + Macrophage vs Cured (up)
Fatty acid degradationStarch and sucrose metabolismButirosin and neomycin biosynthesisFolate biosynthesisGalactose metabolismTyrosine metabolismArachidonic acid metabolismCarbohydrate digestion and absorptionEndocytosisTerpenoid backbone biosynthesisDNA replicationPyruvate metabolismGlycosphingolipid biosynthesis − ganglio seriesC5−Branched dibasic acid metabolismGlycosaminoglycan degradationTryptophan metabolismChloroalkane and chloroalkene degradationHistidine metabolismPentose and glucuronate interconversionsAmino sugar and nucleotide sugar metabolismCytosolic DNA−sensing pathwayGlycerophospholipid metabolismPlant−pathogen interactionSynthesis and degradation of ketone bodiesProtein processing in endoplasmic reticulumStreptomycin biosynthesisPeroxisomeProximal tubule bicarbonate reclamationMethane metabolismFructose and mannose metabolismArginine and proline metabolismPentose phosphate pathwayalpha−Linolenic acid metabolismMAPK signalling pathwayNOD−like receptor signalling pathwayEther lipid metabolismBiosynthesis of unsaturated fatty acidsLysine degradationAscorbate and aldarate metabolismLimonene and pinene degradationIsoquinoline alkaloid biosynthesisPhenylalanine metabolismTropane, piperidine and pyridine alkaloid biosynthesisButanoate metabolismCyanoamino acid metabolismbeta−Alanine metabolismGlutathione metabolismPPAR signalling pathway
0
0.02
0.04
0.06
0.08
(down)
156
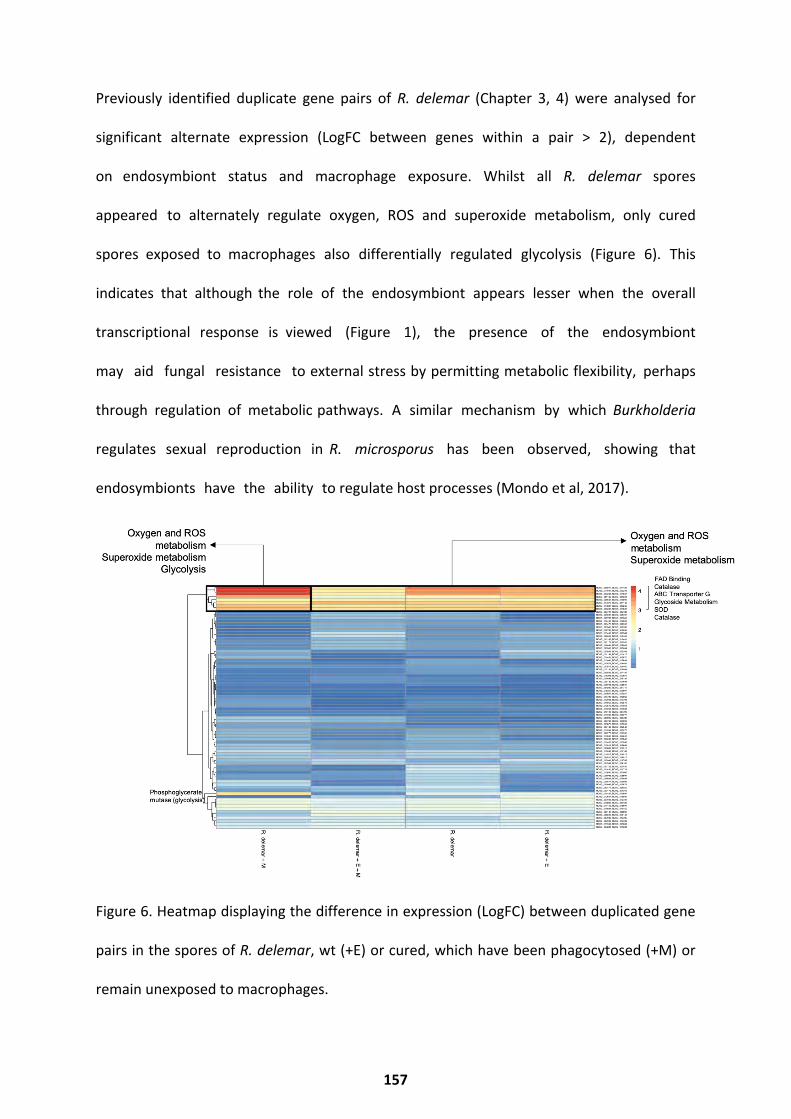
Previously identified duplicate gene pairs of R. delemar (Chapter 3, 4) were analysed for
significant alternate expression (LogFC between genes within a pair > 2), dependent
on endosymbiont status and macrophage exposure. Whilst all R. delemar spores
appeared to alternately regulate oxygen, ROS and superoxide metabolism, only cured
spores exposed to macrophages also differentially regulated glycolysis (Figure 6). This
indicates that although the role of the endosymbiont appears lesser when the overall
transcriptional response is viewed (Figure 1), the presence of the endosymbiont
may aid fungal resistance to external stress by permitting metabolic flexibility, perhaps
through regulation of metabolic pathways. A similar mechanism by which Burkholderia
regulates sexual reproduction in R. microsporus has been observed, showing that
endosymbionts have the ability to regulate host processes (Mondo et al, 2017).
Figure 6. Heatmap displaying the difference in expression (LogFC) between duplicated gene
pairs in the spores of R. delemar, wt (+E) or cured, which have been phagocytosed (+M) or
remain unexposed to macrophages.
157

When comparing transcriptional profiles of wt and cured spores incubated in serum-free
DMEM (sfDMEM) for 3 hours (time matched to phagocytosis assay), very few transcriptional
changes in either species in response to curing were observed. In R. delemar, a single gene,
predicted to be a putative protein phosphatase, was repressed. In R. microsporus , three
genes were induced: an autophagy-related protein, a C2H2 zinc finger transcription factor,
and ribosomal protein L2.
Despite these small changes, loss of the endosymbiont appears to impact the size of
transcriptional responses of both fungal species to macrophages (Figure 3), and Itabangi et al
have shown the importance of the endosymbiont for virulence (Itabangi et al, 2019).
Specifically, an overall increase in the number of fungal genes differentially regulated upon
phagocytosis for both species can be observed. Exposure of wt R. microsporus to
macrophages induced the expression of one gene with no known function and repressed 6
genes. The repressed genes include an autophagy-related protein and a zinc finger
transcription factor, as well as 4 unannotated genes (Figure 3). In contrast, when cured R.
microsporus spores were exposed to macrophages, 277 genes were significantly induced and
82 repressed, compared to unexposed cured spores (Figure 3). Induced genes were enriched
(hypergeometric test, corrected P value < 0.05) for the following GO categories: organelle
organisation, pre-ribosome and ribosome activity, ATPase activity, hydrolase activity,
pyrophosphatase activity, helicase activity, nucleic acid binding, RNA metabolism, nitrogen
metabolism, chromatin silencing. Repressed genes were enriched (hypergeometric test,
corrected P value < 0.05) for the following GO categories: oxidoreductase activity, hydrogen
sulphide metabolism, glycolysis, sulphur metabolism, hexose catabolism, siderophore
158

activity, iron assimilation, nitrogen metabolism, carboxylic acid metabolism. This suggests an
overall failure to properly respond to host stresses such as iron starvation in the absence of
the bacterial endosymbiont.
A similar impact of the endosymbiont was observed for R. delemar. While the overall fungal
response to phagocytosis is characterized by a robust transcriptional response, induced genes
in wt samples (1137 genes, Figure 3) were enriched for KEGG classifications corresponding to:
alanine metabolism, PPAR signalling, aromatic compound biosynthesis and degradation,
lysine metabolism, lipid metabolism, MAPK signalling, sugar metabolism, tyrosine
metabolism, secondary metabolite biosynthesis (Figure 5). Repressed genes in wt samples
(472 genes, Figure 3) were enriched for KEGG classifications corresponding to: carbohydrate
metabolism, secondary metabolite biosynthesis, ketone body processing, protein processes,
MAPK signalling (Figure 5). In contrast, induced genes in cured samples (1285 genes, Figure
3) were enriched for KEGG classifications corresponding to: fatty acid metabolism, DNA
replication, amino acid metabolism, glycan metabolism, pyruvate metabolism and secondary
metabolite processing (Figure 5). Repressed genes in cured samples (878 genes, Figure 3)
were enriched for KEGG classifications corresponding to: sugar metabolism, amino acid
metabolism, lipid metabolism, MAPK signalling, NOD-like receptor signalling. Again, this
suggests that the endosymbiont has an overall suppressive impact on fungal transcription in
response to macrophage challenge.
When comparing the transcriptional responses of ortholog genes shared by R. delemar and
R. microsporus, we see only a small proportion behave similarly (213 genes, detailed
in the online appendix, Ortho_Genes). When phagocytosed, wt spores from both species
upregulate ortholog genes involved in fatty acid
159

catabolism, transcription, regulation via polymerase II, and organelle organization.
Phagocytosed cured spores from both species upregulate ortholog genes involved in RNA
processing, chromosome organization and condensed chromosome pathways. When
unexposed, we see wt spores upregulate ortholog genes involved in translocation, protein
binding, siderophore activity, cobalmin processing, and post-translational protein targeting.
Cured unexposed spores upregulate ortholog genes with roles in siderophore activity
and transferase activity. A list of the 213 orthologs and their predicted functions can
be found in online the appendix (Ortho_Genes). Overall, R. delemar and R. microsporus
both respond transcriptionally to the presence of macrophages, however the size
and composition of this response differs between species.
Due to work which has linked iron scavenging to survival within the phagolysosome
(Andrianaki et al. 2018), expression of genes predicted to be involved in ferrous iron
transport was analysed. There are 12 genes in the R. delemar genome with predicted
ferrous iron roles. While 8 showed no significant change over the tested conditions, 3
(R0G3_006623, R0G3_007727, R0G3_011864) appeared highly expressed in wt and
cured phagocytosed spores, compared to unexposed spores. The last gene, ROG3_009943,
is highly expressed in wt spores unexposed to macrophages. Together, this suggests
there may be condition dependent specialization in the expression of ferrous iron
transport in R. delemar.
160
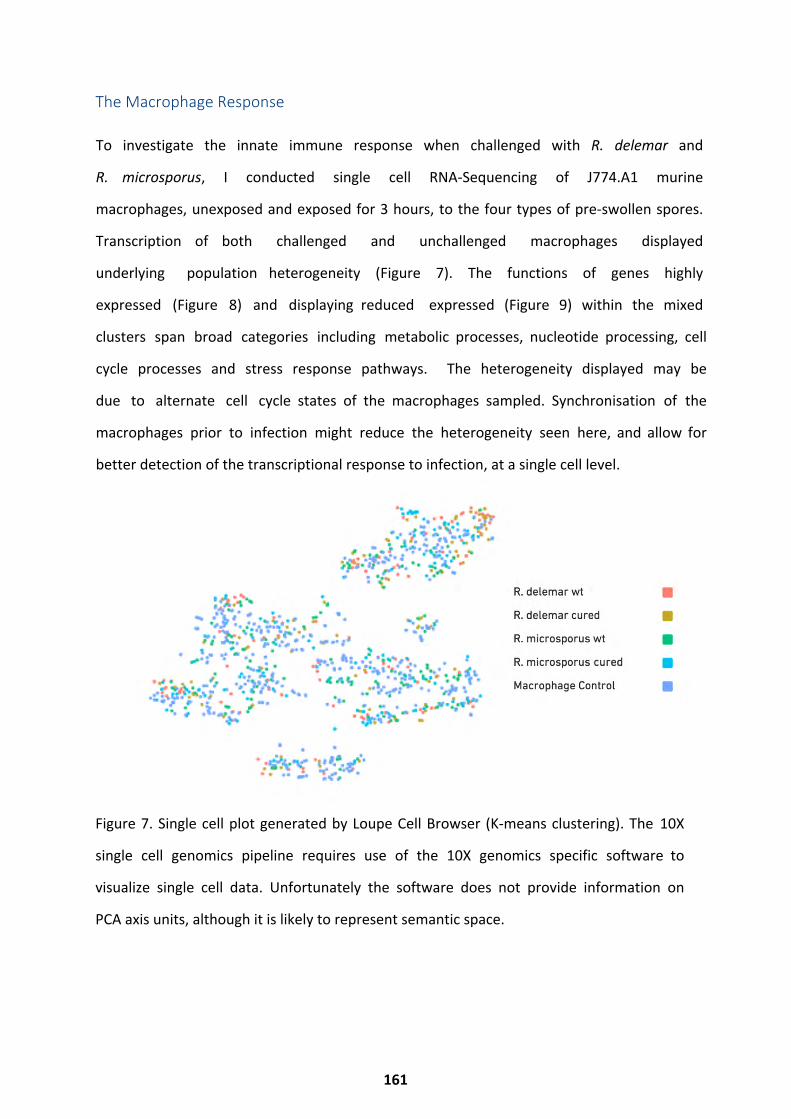
Figure 7. Single cell plot generated by Loupe Cell Browser (K-means clustering). The 10X
single cell genomics pipeline requires use of the 10X genomics specific software to
visualize single cell data. Unfortunately the software does not provide information on
PCA axis units, although it is likely to represent semantic space.
The Macrophage Response
To investigate the innate immune response when challenged with R. delemar and
R. microsporus, I conducted single cell RNA-Sequencing of J774.A1 murine
macrophages, unexposed and exposed for 3 hours, to the four types of pre-swollen spores.
Transcription of both challenged and unchallenged macrophages displayed
underlying population heterogeneity (Figure 7). The functions of genes highly
expressed (Figure 8) and displaying reduced expressed (Figure 9) within the mixed
clusters span broad categories including metabolic processes, nucleotide processing, cell
cycle processes and stress response pathways. The heterogeneity displayed may be
due to alternate cell cycle states of the macrophages sampled. Synchronisation of the
macrophages prior to infection might reduce the heterogeneity seen here, and allow for
better detection of the transcriptional response to infection, at a single cell level.
161
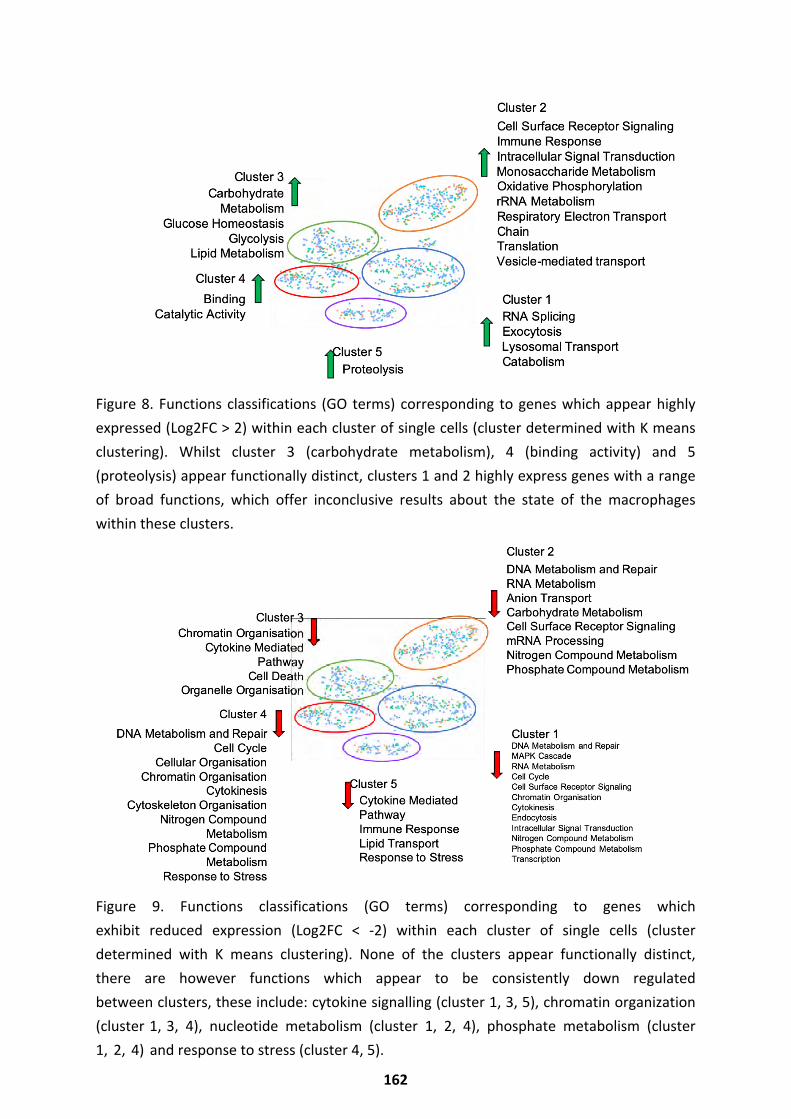
Figure 8. Functions classifications (GO terms) corresponding to genes which appear highly expressed (Log2FC > 2) within each cluster of single cells (cluster determined with K means clustering). Whilst cluster 3 (carbohydrate metabolism), 4 (binding activity) and 5 (proteolysis) appear functionally distinct, clusters 1 and 2 highly express genes with a range of broad functions, which offer inconclusive results about the state of the macrophages within these clusters.
Figure 9. Functions classifications (GO terms) corresponding to genes which exhibit reduced expression (Log2FC < -2) within each cluster of single cells (cluster determined with K means clustering). None of the clusters appear functionally distinct, there are however functions which appear to be consistently down regulated between clusters, these include: cytokine signalling (cluster 1, 3, 5), chromatin organization (cluster 1, 3, 4), nucleotide metabolism (cluster 1, 2, 4), phosphate metabolism (cluster 1, 2, 4) and response to stress (cluster 4, 5).
162
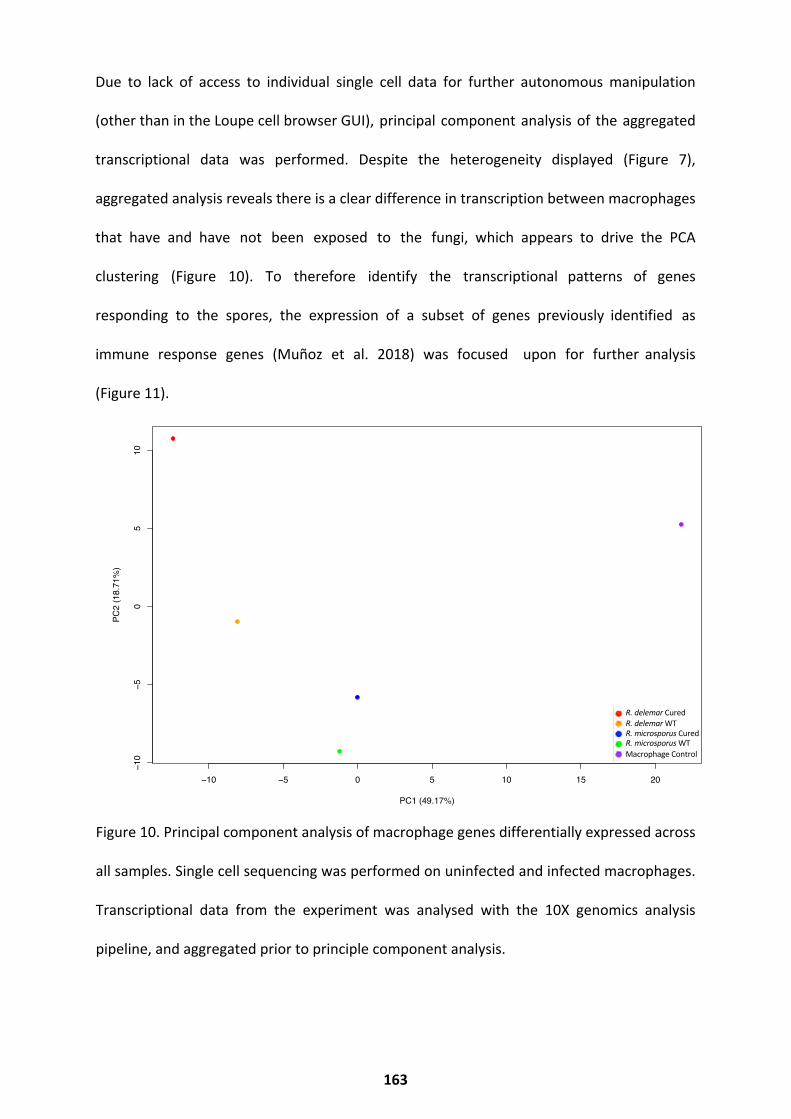
Due to lack of access to individual single cell data for further autonomous manipulation
(other than in the Loupe cell browser GUI), principal component analysis of the aggregated
transcriptional data was performed. Despite the heterogeneity displayed (Figure 7),
aggregated analysis reveals there is a clear difference in transcription between macrophages
that have and have not been exposed to the fungi, which appears to drive the PCA
clustering (Figure 10). To therefore identify the transcriptional patterns of genes
responding to the spores, the expression of a subset of genes previously identified as
immune response genes (Muñoz et al. 2018) was focused upon for further analysis
(Figure 11).
Figure 10. Principal component analysis of macrophage genes differentially expressed across
all samples. Single cell sequencing was performed on uninfected and infected macrophages.
Transcriptional data from the experiment was analysed with the 10X genomics analysis
pipeline, and aggregated prior to principle component analysis.
−10 −5 0 5 10 15 20
−10
−50
510
PC1 (49.17%)
PC2
(18.
71%
)
S1 S2 S3 S4 S5
R. delemar CuredR. delemar WTR. microsporus CuredR. microsporus WTMacrophage Control
163

Across all exposed conditions, relative to unexposed macrophages, there was a profile
consistent with cytokine activation, response to stimulus, and activation of the NF-Kb
pathway. This was accompanied by repression of CCL5, which is involved in T-cell recruitment
(Figure 11). However, different macrophage profiles can be seen in response to the two fungal
species, and these are further influenced by the presence of the endosymbiont. While the
response to wt R. delemar shows the most deviation from the macrophage-only control,
exposure to cured R. delemar also elicited a strong and distinct macrophage response (Figure
11). Exposure to wt R. delemar elicits increased expression of general markers of activation,
including GTPase activity and MHC class II protein binding (LAG3 repressor of T-cell activation,
H2-M2, IFN-gamma induced IIGP1, MX1, KCTD14, PNP2), growth factor binding, IL1 receptor
agonist activity and endocytosis (SERPINE1, ENG, FGFBP3, GM8898, GCNT2, IL1F6).
Specifically, we observe modest increases in the expression of IFN-γ responsive CXCL10 (3.1
fold) and IRG1/IRG11 (5.9 fold), pro-inflammatory SAA3 (3.5 fold), and ENPP4 (2.8 fold), but
also induction of the M2 polarizing PSTPIP2/21 (6.7 fold), the IL-4 responsive signalling
modulator CISH (5.4 fold), and the vascular damage responsive F3/F31 (5.9 fold) (Martinez et
al. 2013). These latter genes are not as strongly induced during exposure to R. microsporus,
which may reflect the aggressive nature of infection by R. delemar relative to R. microsporus.
In contrast, infection with cured R. delemar showed a decrease in the induction of these M2-
polarisation markers (PSTPIP2, 3.5 fold relative to uninduced). The transcriptional profile is
instead shifted to include increased transcription of genes involved in G protein signalling and
phosphoinositide binding (PDE7B, CCL1, SCARF1, RGS16, PLEKHA4) (Figure 11).
164

A similar change in macrophage polarization was observed during exposure to wt and cured
R. microsporus. For both wt and cured R. microsporus, expression of IFN-γ responsive CXCL10,
SAA3, and ENPP4 was comparable to unexposed macrophages. Compared to R. delemar,
exposure to wt R. microsporus induced a more limited expression of genes with roles in
cytokine activation, ERK1 and ERK2 regulation, and regulation of NF-kB cascade (Figure 11).
There was also a weaker induction of the vascular damage responsive F3/F31 genes, and a
relatively stronger upregulation of the PLA2G16 phospholipase, TRIM30D, SLC1A2 and the
M2 polarizing IL-6. However, other key polarizing genes, particularly PSTPIP2/21, SAA3, and
ENPP4 were only weakly induced. Overall, this profile suggests a weak M1-like activation
consistent with poor phagocytosis and reduced overall antifungal activity that can be
observed in macrophages interacting with endosymbiont-harbouring R.microsporus spores
(Itabangi et al. 2019).
Finally, cured R. microsporus induced a strong pro-inflammatory response, which included
upregulation of CXCL3, the neutrophil chemoattractant, consistent with observations of
differences in phagocyte recruitment in zebrafish upon infection with wt vs. cured spores
(Itabangi et al. 2019). Markers of NF-kB activation were also strongly induced in this
population. Cured R. microsporus also strongly induced the expression of TNFRSF8 (CD30), a
marker of lymphocyte activation occasionally associated with subcutaneous fungal infections.
Overall, this is suggestive of a shift to a more pro-inflammatory profile. Itabangi et al. observe
that cured R. microsporus is more sensitive to phagocyte-mediated killing and phagocyte
recruitment compared to wt (Itabangi et al. 2019), to test whether a more successful response
to the spores could be mounted via the induction of a pro-inflammatory response, further in
vitro experiments were carried out (Figure 12).
165
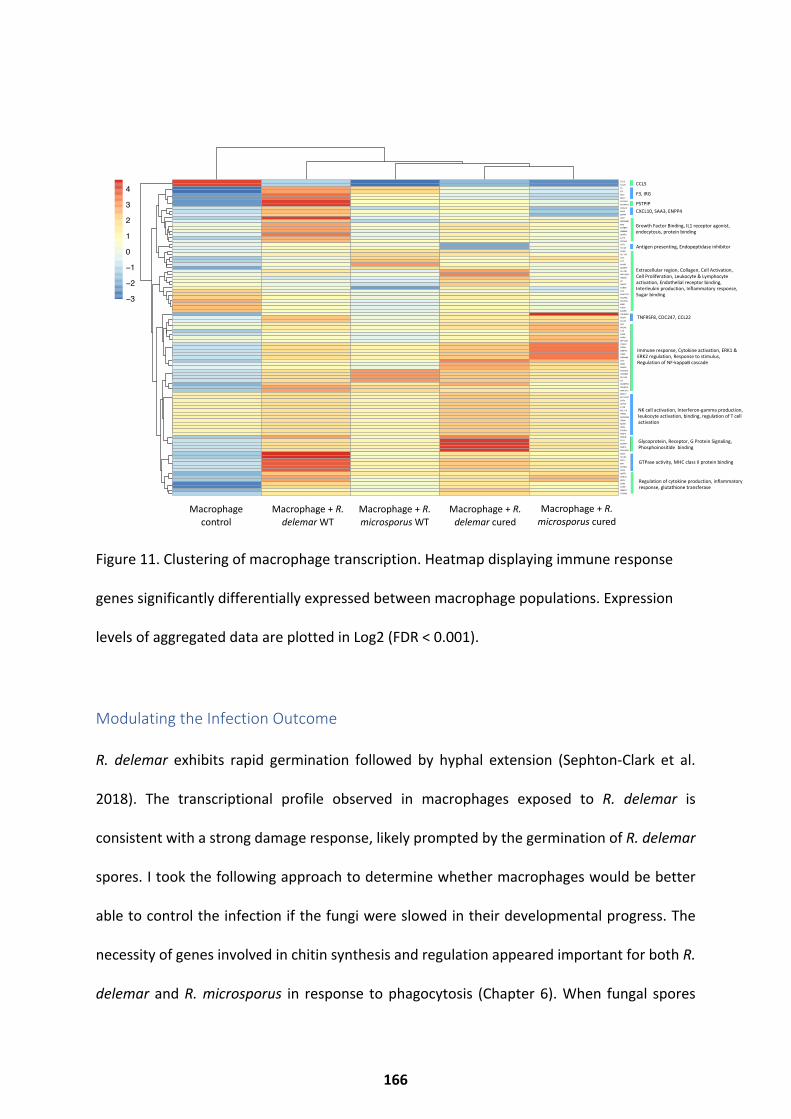
Figure 11. Clustering of macrophage transcription. Heatmap displaying immune response
genes significantly differentially expressed between macrophage populations. Expression
levels of aggregated data are plotted in Log2 (FDR < 0.001).
Modulating the Infection Outcome
R. delemar exhibits rapid germination followed by hyphal extension (Sephton-Clark et al.
2018). The transcriptional profile observed in macrophages exposed to R. delemar is
consistent with a strong damage response, likely prompted by the germination of R. delemar
spores. I took the following approach to determine whether macrophages would be better
able to control the infection if the fungi were slowed in their developmental progress. The
necessity of genes involved in chitin synthesis and regulation appeared important for both R.
delemar and R. microsporus in response to phagocytosis (Chapter 6). When fungal spores
S5 S1 S3 S2 S4
CCL5
CCL51F3
F31IRG1
IRG11PSTPIP2
PSTPIP21CXCL10
SAA3ENPP4
CISHSERPINE1
ENGFGFBP3
GM8898GCNT2
IL1F6ZFP558
CST7H2−Q6
VCANH2−T24
LY75IL27
AKAP2SLAMF7
H2−AB1AW112010GM5431
CPFNDC7
CCND2IL4I1
AA467197COL4A2
COL27A1EDN1
IFI205IL20RB
TNFRSF8CD247
CCL22AK4
HVCN1IL1B
IL23AAGRN
ATP13A4CX3CL1
HTRA1GM8773
TGM2AI504432
FLT4NOD1
RSAD2PLA2G16
TRIM30DSLC1A2IL6
ADAMTS4TNFSF10
AKR1C12MACC1
BC147527CSF3
SLFN5IL12B
BCL11AHTRA4
ADORA2ATRIM5
KLRK1GBP4
PYHIN1FABP3
PDE7BCCL1
SCARF1RGS16
PLEKHA4LAG3
H2−M2IIGP1
MX1KCTD14
PNP2GSTT1
GPR141MEFV
CD83CD831GM6377
TICAM2
−3
−2
−1
0
1
2
3
4CCL5
F3, IRG
PSTPIP
Glycoprotein, Receptor, G Protein Signaling, Phosphoinositide binding
CXCL10, SAA3, ENPP4
Growth Factor Binding, IL1 receptor agonist, endocytosis, protein binding
Antigen presenting, Endopeptidase inhibitor
Extracellular region, Collagen, Cell Activation, Cell Proliferation, Leukocyte & Lymphocyte activation, Endothelial receptor binding, Interleukin production, Inflammatory response, Sugar binding
TNFRSF8, CDC247, CCL22
Immune response, Cytokine activation, ERK1 & ERK2 regulation, Response to stimulus, Regulation of NF-kappaB cascade
NK cell activation, Interferon-gamma production, leukocyte activation, binding, regulation of T cell activation
GTPase activity, MHC class II protein binding
Regulation of cytokine production, inflammatory response, glutathione transferase
Macrophage control
Macrophage + R. delemar WT
Macrophage + R. microsporus WT
Macrophage + R. delemar cured
Macrophage + R. microsporus cured
S5 S1 S3 S2 S4
CCL5
CCL51F3
F31IRG1
IRG11PSTPIP2
PSTPIP21CXCL10
SAA3ENPP4
CISHSERPINE1
ENGFGFBP3
GM8898GCNT2
IL1F6ZFP558
CST7H2−Q6
VCANH2−T24
LY75IL27
AKAP2SLAMF7
H2−AB1AW112010GM5431
CPFNDC7
CCND2IL4I1
AA467197COL4A2
COL27A1EDN1
IFI205IL20RB
TNFRSF8CD247
CCL22AK4
HVCN1IL1B
IL23AAGRN
ATP13A4CX3CL1
HTRA1GM8773
TGM2AI504432
FLT4NOD1
RSAD2PLA2G16
TRIM30DSLC1A2IL6
ADAMTS4TNFSF10
AKR1C12MACC1
BC147527CSF3
SLFN5IL12B
BCL11AHTRA4
ADORA2ATRIM5
KLRK1GBP4
PYHIN1FABP3
PDE7BCCL1
SCARF1RGS16
PLEKHA4LAG3
H2−M2IIGP1
MX1KCTD14
PNP2GSTT1
GPR141MEFV
CD83CD831GM6377
TICAM2
−3
−2
−1
0
1
2
3
4
166

were pre-treated with the chitin synthesis inhibitor Nikkomycin Z (24 μg/ml) they remained
viable but exhibited reduced swelling. By challenging macrophages with spores pre-treated
with Nikkomycin Z (24 μg/ml), macrophage survival was increased at 7.5 hours post infection
(Figure 12). As the macrophages are better able to control these spores, this suggests that
spores undergoing the initial stages of germination may offer less of a challenge for the
macrophages. This is consistent by data shown by Itabangi et al., who examined macrophage
response to both resting and swelling R. microsporus spores (Itabangi et al. 2019).
The transcriptional data show a strong M2 alternative activation signal during R. delemar-
macrophage interaction, but a weaker M2 polarisation during R. microsporus-macrophage
interaction that was further shifted towards NF-kB-mediated M1 upon endosymbiont cure.
To shift the macrophage polarization towards M1 classical activation, macrophages were pre-
treated with NF-kB activating lipopolysaccharide (LPS). This offered a protective effect upon
Mucorales infection, significantly improved the ability of macrophages to control R.
microsporus. At 7.5 hours post infection, 59.7% of macrophages survived when pre-treated
with LPS, compared to 24.6% without (Figure 12).
167
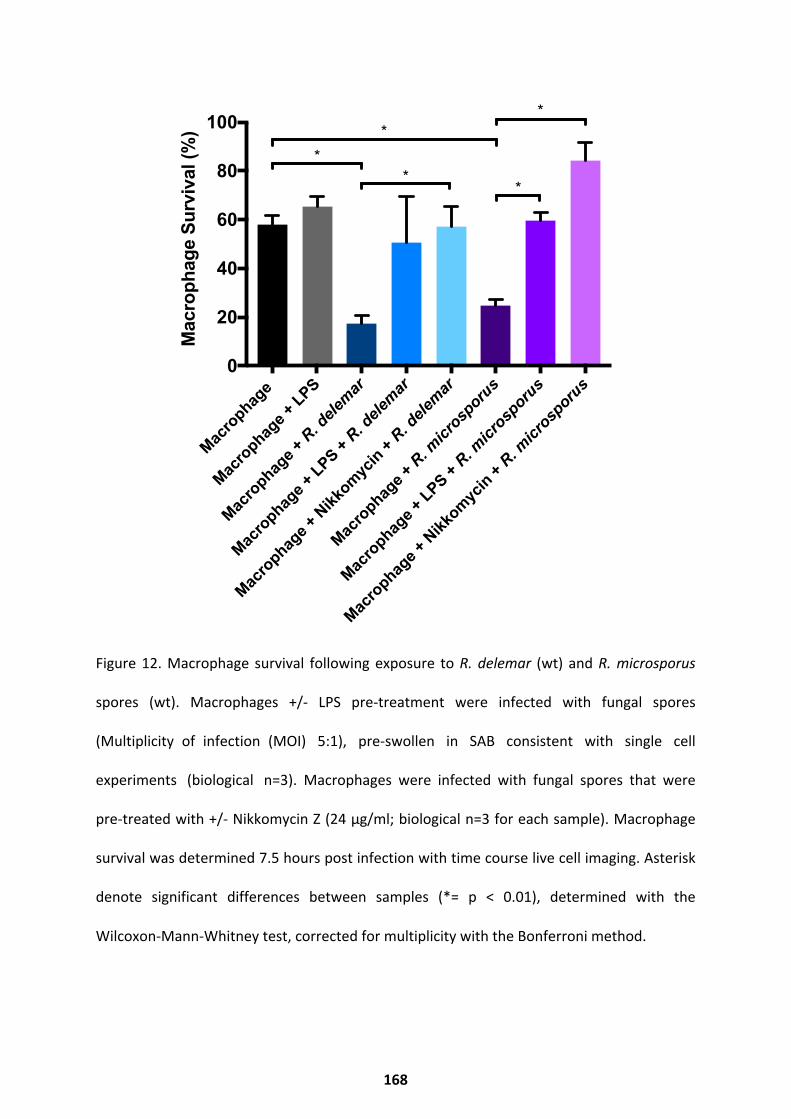
Figure 12. Macrophage survival following exposure to R. delemar (wt) and R. microsporus
spores (wt). Macrophages +/- LPS pre-treatment were infected with fungal spores
(Multiplicity of infection (MOI) 5:1), pre-swollen in SAB consistent with single cell
experiments (biological n=3). Macrophages were infected with fungal spores that were
pre-treated with +/- Nikkomycin Z (24 μg/ml; biological n=3 for each sample). Macrophage
survival was determined 7.5 hours post infection with time course live cell imaging. Asterisk
denote significant differences between samples (*= p < 0.01), determined with the
Wilcoxon-Mann-Whitney test, corrected for multiplicity with the Bonferroni method.
Macro
phage
Macro
phage +
LPS
Macro
phage +
R. d
elem
ar
Macro
phage +
LPS +
R. dele
mar
Macro
phage +
Nikk
omyc
in +
R. dele
mar
Macro
phage +
R. m
icrosp
orus
Macro
phage +
LPS +
R. micr
osporu
s
Macro
phage +
Nikk
omyc
in +
R. micr
osporu
s0
20
40
60
80
100
Mac
roph
age
Sur
viva
l (%
)
**
*
*
*
168

Conclusions
These results show that the fungal response to innate immune cells differs by species in the
Rhizopodaceae family. Although R. delemar and R. microsporus share a small conserved
response to macrophage exposure, the majority of their response differs. Results show that
the fungal transcriptional response appears largely unperturbed by the presence or lack of
an endosymbiont, in the absence of stress. However, the presence of an endosymbiont
greatly effects the response of the host. Activation of the host and inhibition of spore
germination successfully modulates the infection outcome.
Discussion
Surprisingly, the scale of the fungal response varied appreciably between species. As
previously discussed, this may be an artifact of the differential growth rates these species
display. This variation is also observed in other relatively closely related species; C. albicans
and C. glabrata differ substantially in their responses to the host (Brunke and Hube, 2012).
Unsurprisingly, the macrophage response (Figure 11) reflected in vitro observations; the
more ‘M1-like’ the transcriptional response, the better the macrophages control the spores
in vitro. Unfortunately, the macrophage single cell sequencing approach taken here (10X
genomics single cell) resulted in limited meaningful single cell data. This was due to the
unavailability of single cell data to the user, outside of their specified user pipeline (which
provided inflexible and limited analysis options). To improve on the single cell analysis,
further experiments utilising an alternative system such as Drop-Seq should be carried out.
169

As anticipated, activation of pro-inflammatory pathways increased macrophage survival in
response to the spores. This consolidates several studies which demonstrate improved innate
immune cell response to fungal pathogens when primed (Rogers et al. 2013; Municio et al.
2013; Blasi et al. 1995). In addition, this analysis reveals profound differences in the host
response to two related Rhizopus species. In particular, results show a M2/damage-
associated response during infection with wt R. delemar spores that is shifted towards an M1
protective response upon infection with cured R. microsporus spores. The ability to
germinate prior to phagocyte control also appears key to virulence, as blocking spore
germination with the chitin synthase inhibitor Nikkomycin Z improves macrophage survival,
and highlights the requirement for chitin synthase for spore development in these species.
Further Work
Future work focusing on modulation of the innate immune system in both in vitro and in vivo
models would allow a better understanding of the innate immune response to Mucorales
species. Modulation of pathways which include the genes identified here (Figure 11) with
exogenous cytokines, signalling molecules and inhibitors would allow for further elucidation
of the pathways involved in the macrophage response. Clinical modulation or stimulation of
the immune system could provide protection against infection in high-risk patients. Topical or
systemic germination inhibitors might also be explored as options for a prophylactic
treatment in high risk patients. Antimycin A isomers and derivatives would be good
candidates for this, however the cytotoxicity of Antimycin A itself makes it unsuitable for use
in humans.
170

Chapter 6: Germination Inhibition
This chapter will introduce approaches which are capable of inhibiting and modulating fungal
germination. My results demonstrate several compounds capable of inhibiting germination
of Rhizopus spores. These compounds successfully inhibit germination through chitin
synthase inhibition, electron transport chain inhibition and reactive oxygen species pathway
regulation. Further to this, my results demonstrate inhibition of Rhizopus spore germination
through treatment with natural compounds extracted from plants and lichen. Both
compound screening and target based approaches have yielded compounds capable of
germination inhibition, shedding light on the mechanisms which underpin germination.
171

Manipulation and Inhibition of Germination
This work has been adapted from the book chapter “Spore Germination of Pathogenic
Filamentous Fungi” (Sephton-Clark and Voelz 2017), for which I performed the
literature search, wrote the manuscript, completed revisions, and prepared the figures.
Germination is the developmental process underlying initiation of many
fungal diseases. Thus, strategies that inhibit germination have been of much
interest as a means to treat both human and plant fungal infections. Inhibition of
Mucorales spore germination by the host is likely key to preventing infection in
healthy individuals, however germination inhibition often fails in
immunocompromised infection models (Voelz et al. 2015; Rosowski et al. 2018). The
process of germination is therefore an attractive therapeutic target, which has been
explored for Aspergillus species previously. Germination inhibition may be reversible, as
seen for multiple Aspergillus species when inhibited (Nogueira et al. 2019). There are
multiple fungistatic and fungicidal drugs currently available to treat fungal
infections, however most remain innefective against Rhizopus species. Given
the impact of Rhizopus germination on the host, and limited treatments
for mucormycosis, I attempted to identify germination inhibitors.
Manipulation of physical parameters is a common strategy employed to ensure food
safety. Decreasing water potential can aid with fungicidal action, as the germination of
Aspergillus niger species is inhibited by reduced water availability (Long, 2017; Ni, 2005).
Lowering temperature is also a common method for inhibiting germination of food spoiling
fungi (Eckert, 1967; Barth, 2009). The common food spoilage agent Rhizopus delemar will
not germinate below 5 °C (Eckert, 1988), whilst the pathogen M. gypseum will
only tolerate temperatures over 35 °C (Leighton and Stock 1969) demonstrating
temperature is clearly a strong mechanism for regulating germination.
Non-steroidal anti-inflammatory drugs (NSAIDs) such as Ibuprofen, have been shown to
inhibit the germination of several fungal species capable of causing respiratory diseases,
including Aspergillus niger (Dalmont, 2017). The use of aerosolized NSAIDs as a
preventative measure has been suggested as a way to tackle these common afflictions,
however one should be careful to note the appropriate controls for this study are absent,
so conclusions which can be drawn remain limited. If effective, they could be especially
useful in damp 172 housing where pathogens such as Aspergillus niger thrive (Dalmont,
2017).
172

Furthermore, statins have been shown to decrease germination of the human
pathogen Rhizopus oryzae, as well as increasing susceptibility to oxidative stress (Bellanger
et al. 2016).
The topical administration of cheap and accessible substances such as acetic acid to open
wounds has been suggested as another treatment which is effective due to its inhibition of
germination (Trzaska et al. 2015). Acetic acid has been shown to inhibit germination of
mucormycosis causing species. The administration of acetic acid might provide effective
means for preventing this hard-to-treat invasive disease in patients with deep tissue wounds
which may have been exposed to contaminated soils (e.g. blast wounds) (Trzaska et al.
2015). Furthermore, the germination of several pathogenic fungi can be inhibited by
substances such as ethanol (Plumridge et al. 2004; Trzaska et al. 2015; Dao & Dantigny
2011).
Nanoparticles, an excellent alternative to traditional chemical treatments, are capable of
inhibiting germination in the tobacco leaf pathogen P. tabacina. Administration of such Zinc
nanoparticles to tobacco leaves has been suggested as a cheap and efficient way to reduce
pathogenesis and crop losses, as the nanoparticles show effectivity at very low doses
(Wagner et al. 2016).
In addition, a range of biological strategies has been investigated. Several biomolecules can
be used effectively to inhibit the germination of fungi. Chitosan derivatives which
incorporate a pyridine were found to exhibit inhibition of germination on the plant
pathogen B. cinerea, posing yet another method for the treatment of food or plants with a
safe molecule, which would decrease plant disease by fungal infection (Jia et al. 2016).
Similarly, the inhibition of germination of plant pathogens through biocontrol is a currently
expanding field. Colonising crops with plant ‘safe’ bacteria or fungi, which produce
molecules damaging to plant pathogens, can effectively control plant disease.
173

For example, organic compounds released by Streptomyces albulus have been shown to be
capable of inhibiting germination of the plant pathogen Fusarium oxysporum (Wu, 2015).
Pseudomonas antimicrobica produces a molecule capable of decreasing germination rates
of the prolific fungal plant pathogen B. cinerea (Wu, 2015; Walker, 2001).
Whilst many of these recently suggested approaches have great potential to offer new
methods in combating fungal growth and disease, our current understanding of the
underlying mechanisms of inhibition is limited and thus requires further study.
174

Results
Results have been previously published or adapted from the following articles: “Pathways of
Pathogenicity: Transcriptional Stages of Germination in the Fatal Fungal Pathogen Rhizopus
delemar”(Sephton-Clark et al. 2018) and “Host-pathogen transcriptomics of macrophages,
Mucorales and their endosymbionts: a polymicrobial pas de trois” (Sephton-clark et al. 2019).
For both I conceived, designed and performed the experiments, collected the data, performed
the analysis and interpretation, wrote the manuscript, completed revisions, and prepared the
figures.
Germination Inhibitors Identified Through Transcriptional Studies
Transcriptional and phenotypic results identified cell wall remodelling, respiration, REDOX
and stress response to be key pathways differentially regulated during the transition from
resting spore to swelling spore. To determine whether modulation of these pathways would
lead to inhibition of germination, spores were treated with a chitin synthase inhibitor
(Nikkomycin Z), a cytochrome c reductase inhibitor (Antimycin A) and exogenous reactive
oxygen species (ROS) (Hydrogen peroxide).
An increase in intracellular ROS can be observed in R. delemar over the course of germination
(Figure 1a). I investigated the significance of ROS detoxification during germination by testing
for resistance to exogenous (H2O2) and endogenous (mitochondrial-derived) ROS (Figure 1b).
Treatment with 5 mM but not 1 mM H2O2 was sufficient to inhibit spore germination. In
contrast, spores were highly sensitive to treatment with 1.5 or 10nM antimycin A. Inhibition
of the mitochondrial cytochrome c reductase leads to an accumulation of superoxide radicals
175

within the cell. Furthermore, antimycin A may exert a dual effect, as the expression of storage
molecule transcripts appears high in both ungerminated spores and the hyphal form. High
sensitivity to inhibition of oxidative phosphorylation with antimycin A is consistent with
reports that utilization of these storage molecules as energy reserves is important for the
initiation and maintenance of growth (Elbein 1974; Novodvorska et al. 2016; Svanström et al.
2014).
When spores of R. delemar and R. microsporus were treated with the chitin synthesis inhibitor
Nikkomycin Z (24-120 μg/ml), they failed to germinate (Figure 1c) and displayed less
chitin/chitosan in their outer cell wall (Figure 1c: R. delemar 120 μg/ml, R. microsporus 24-
120 μg/ml). At Nikkomycin Z concentrations lower than 24 μg/ml, we see the spores are able
to swell, however development appears halted after swelling.
176
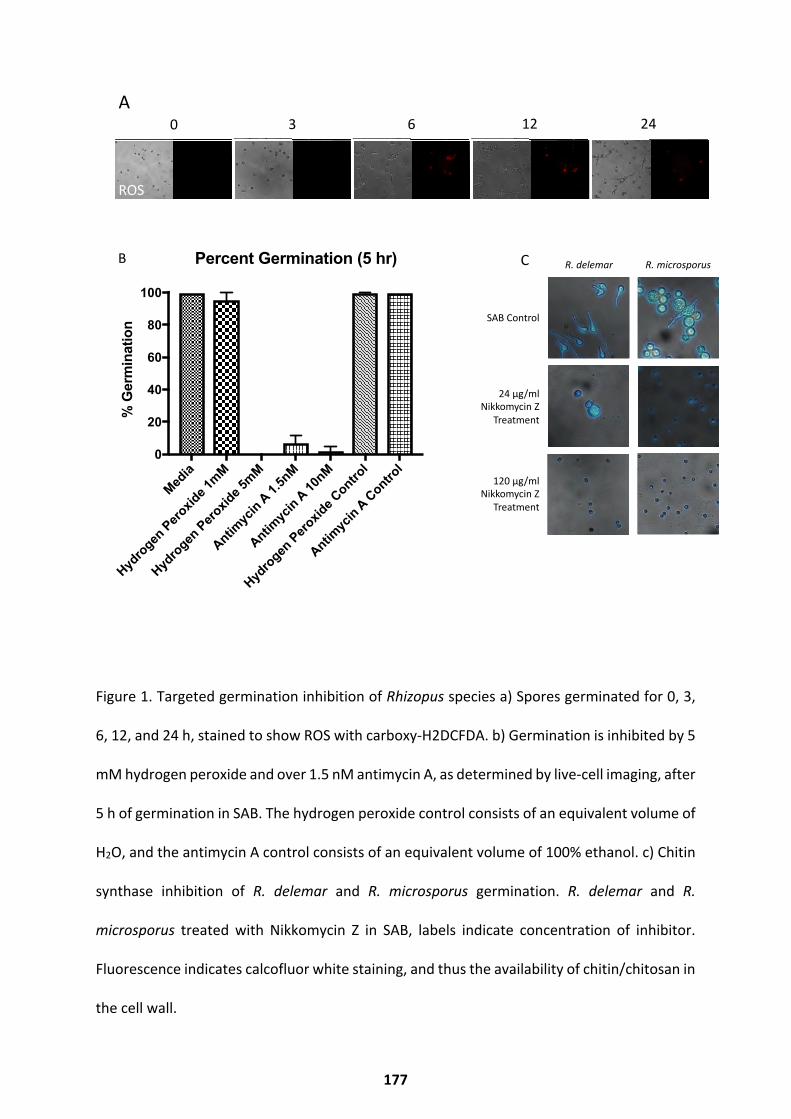
Figure 1. Targeted germination inhibition of Rhizopus species a) Spores germinated for 0, 3,
6, 12, and 24 h, stained to show ROS with carboxy-H2DCFDA. b) Germination is inhibited by 5
mM hydrogen peroxide and over 1.5 nM antimycin A, as determined by live-cell imaging, after
5 h of germination in SAB. The hydrogen peroxide control consists of an equivalent volume of
H2O, and the antimycin A control consists of an equivalent volume of 100% ethanol. c) Chitin
synthase inhibition of R. delemar and R. microsporus germination. R. delemar and R.
microsporus treated with Nikkomycin Z in SAB, labels indicate concentration of inhibitor.
Fluorescence indicates calcofluor white staining, and thus the availability of chitin/chitosan in
the cell wall.
0 3 6 12 24A
B
Media
Hydro
genPero
xide 1m
M
Hydro
genPero
xide 5m
M
Antimyc
inA
1.5nM
Antimyc
inA
10nM
Hydro
genPero
xide Contro
l
Antimyc
inA
Control
0
20
40
60
80
100
% G
erm
inat
ion
Percent Germination (5 hr)
CFW
FITC
ROS
0 3 6 12 24A
B
Media
Hydro
genPero
xide 1m
M
Hydro
genPero
xide 5m
M
Antimyc
inA
1.5nM
Antimyc
inA
10nM
Hydro
genPero
xide Contro
l
Antimyc
inA
Control
0
20
40
60
80
100
% G
erm
inat
ion
Percent Germination (5 hr)
CFW
FITC
ROS
R. delemar R. microsporus
SAB Control
24 µg/ml Nikkomycin Z
Treatment
120 µg/ml Nikkomycin Z
Treatment
0 3 6 12 24A
B
Media
Hydro
gen Pero
xide 1
mM
Hydro
gen Pero
xide 5
mM
Antimyc
in A 1.5n
M
Antimyc
in A 10nM
Hydro
gen Pero
xide C
ontrol
Antimyc
in A Contro
l0
20
40
60
80
100
% G
erm
inat
ion
Percent Germination (5 hr)
CFW
FITC
ROS
C
177

A screen identified compounds from a natural compound library, provided by
Strathclyde University, capable of inhibiting germination of Mucorales species. The
candidate identified to inhibit germination, A10, is an extract from either plant, lichen or
fungi. A10 inhibits the metabolic activity of R. microsporus in a dose dependent manner
(Figure 2), with high doses inhibiting as potently as the known inhibitor acetic acid (Trzaska
et al. 2015). The flavonoid chrysin, a known antifungal (Shimura et al. 2007), was
identified from a list of compounds (provided by Strathclyde University) as a potential
active molecule within A10.
Germination Inhibitors Identified Through Natural Compound Screening
178
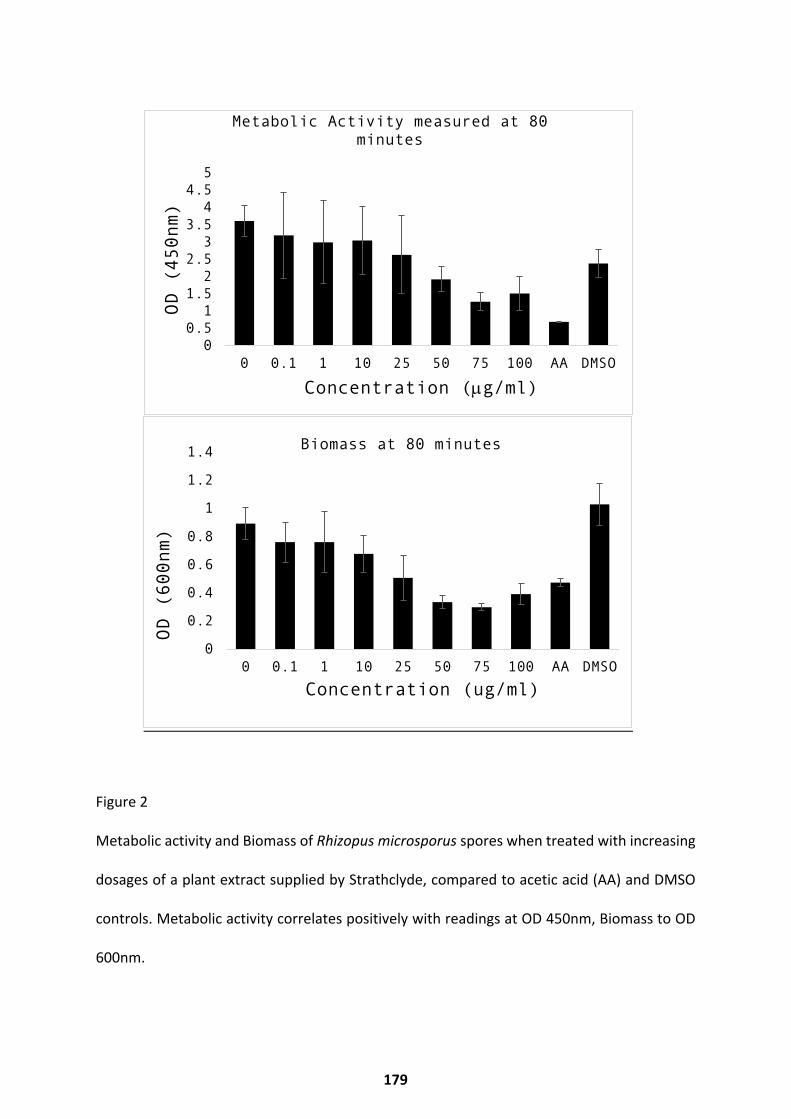
Figure 2
Metabolic activity and Biomass of Rhizopus microsporus spores when treated with increasing
dosages of a plant extract supplied by Strathclyde, compared to acetic acid (AA) and DMSO
controls. Metabolic activity correlates positively with readings at OD 450nm, Biomass to OD
600nm.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 0.1 1 10 25 50 75 100 AA DMSO
OD (
600n
m)
Concentration (ug/ml)
Biomass at 80 minutes
00.51
1.52
2.53
3.54
4.55
0 0.1 1 10 25 50 75 100 AA DMSO
OD (
450n
m)
Concentration (µg/ml)
Metabolic Activity measured at 80 minutes
179

Discussion
The inhibitors identified in this work highlight the roles of chitin synthase and ROS signalling
in regulating germination in Rhizopus species. Hydrogen peroxide has the potential to be
used as a topical prophylactic treatment for high-risk patients, as solutions of 3-6% H2O2
(3% = 0.88M) are well tolerated by the skin (Mahran et al. 2019). H2O2 appears fungicidal to
Rhizopus species at 5mM, a concentration which is unlikely to cause skin irritation. Multiple
pharmaceutical companies have attempted to develop Nikkomycin Z for clinical use as an
antifungal (Galgiani, 2016), however it has not yet made it to market as an approved
antifungal treatment. Nikkomycin Z serves as a good candidate for the treatment of
mucormycosis, and should be explored further as a treatment option with in vivo models.
The potential for germination inhibition with natural compounds should be further explored
with high resolution mass spectrometry of the fungicidal A10, to determine the structures of
compounds within A10. A germination screen with purified forms of these compounds
would offer candidates for testing with mammalian cell lines, to determine toxicity and drug
development suitability.
180

References
A Ayumi, Y Oda, K Asano, And T Sone. 2007. “Rhizopus Delemar Is The Proper Name For Rhizopus Oryzae Fumaric-Malic Acid Producers.” Mycologia
99 99 (5) : 714–22. Https://Doi.Org/10.1080/15572536.2007.11832535.R Abrashev, P Dolashka, R Christova, L Stefanova, And M Angelova. 2005. “Role Of
Antioxidant Enzymes In Survival Of Conidiospores Of Aspergillus Niger Under Conditions Of Temperature Stress.” Journal Of Applied Microbiology 99 (4) : 902–9. Https://Doi.Org/10.1111/J.1365-2672.2005.02669.X.
T H Adams, J K Wieser, And J H Yu. 1998. “Asexual Sporulation In Aspergillus Nidulans.” Microbiology And Molecular Biology Reviews : MMBR 62 (1): 35–54. Http://Www.Ncbi.Nlm.Nih.Gov/Pubmed/9529886%5Cnhttp://Www.Pubmedcentral.Nih.Gov/Articlerender.Fcgi?Artid=PMC98905.
A Vishukumar, J Bayry, S Bozza, O KniemeyerK Perruccio, S Ramulu Elluru, C Clavaud, Et Al. 2009. “Surface Hydrophobin Prevents Immune Recognition Of Airborne Fungal Spores.” Nature 460 (7259) : 1117–21. Https://Doi.Org/10.1038/Nature08264.
Akoumianaki, Tonia, I Kyrmizi, I Valsecchi, M Gresnigt, G Samonis, E Drakos, D Boumpas, Et Al. 2016. “Aspergillus Cell Wall Melanin Blocks LC3-Associated Phagocytosis To Promote Pathogenicity.” Cell Host &
Microbe 19 (1) : 79–90. Https://Doi.Org/10.1016/J.Chom.2015.12.002. Alan, E Celis-Aguilar, Burgos-Páez, Villanueva-Ramos, Alma De La, Mora-
Manjarrez-Velázquez, Escobar-Aispuro, Becerril, Valdez-Flores Francisco, And Beatriz Caballero-Rodríguez. 2019. “An Emergent Entity : Indolent Mucormycosis Of The Paranasal Sinuses . A Multicenter Study,” No. 2193: 92–100.
Alavi, Peyman, H Müller, M Cardinale, C Zachow, M Sánchez, J Martínez, And G Berg. 2013. “The DSF Quorum Sensing System Controls The Positive Influence Of Stenotrophomonas Maltophilia On Plants.” Plos ONE 8 (7) : 1–9. Https://Doi.Org/10.1371/Journal.Pone.0067103.
Albuquerque, P Costa, E Nakayasu, M Rodrigues, S Frases, A Casadevall, R Zancope-Oliveira, I Almeida, And J Nosanchuk. 2008. “Vesicular Transport In Histoplasma Capsulatum: An Effective Mechanism For Trans-Cell Wall Transfer Of Proteins And Lipids In Ascomycetes.” Cellular Microbiology 10 (8) : 1695–1710.
Alspaugh, Cavallo, Perfect, And Heitman. 2000. “RAS1 Regulates Filamentation, Mating And Growth At High Temperature Of Cryptococcus Neoformans.” Molecular Microbiology 36 (2) : 352–65.
Alspaugh, Andrew. 2015. “Virulence Mechanisms And Cryptococcus Neoformans Pathogenesis.” Fungal Genetics And Biology 78: 55–58. Https://Doi.Org/10.1016/J.Fgb.2014.09.004.
Anand, Gautam, Yadav, And Yadav. 2016. “Purification And Characterization Of Polygalacturonase From Aspergillus Fumigatus MTCC 2584
181

And Elucidating Its Application In Retting Of Crotalaria Juncea Fiber.” 3 Biotech 6 6 (2) : 1–7. Https://Doi.Org/10.1007/S13205-016-0517-4.Anders, PT Pyl, And W Huber. 2015. “Htseq-A Python
Framework To Work With High-Throughput Sequencing Data.” Bioinformatics 31 (2): 166–69. Https://Doi.Org/10.1093/Bioinformatics/Btu638.
Andrews S. Fastqc: A Quality Control Tool For High Throughput Sequence Data. Available Online At:
Http://Www.Bioinformatics.Babraham.Ac.Uk/Projects/Fastqc. Anderson, Hane, Stoll, Pain, Hastie, P Kaur, C Hoogland, J Gorman, And KB Singh.
2016. “Mass-Spectrometry Data For Rhizoctonia Solani Proteins Produced During Infection Of Wheat And Vegetative Growth.” Data In Brief 8: 267–71. Https://Doi.Org/Https://Doi.Org/10.1016/J.Dib.2016.05.042.
Andrianaki, Angeliki, Kyrmizi, K Thanopoulou, C Baldin, E Drakos, S.M. Soliman, AC. Shetty, Et Al. 2018. “Iron Restriction Inside MacrophagesRegulates Pulmonary Host Defense Against Rhizopus Species.” Nature
Communications 9 (1). Https://Doi.Org/10.1038/S41467-018-05820-2.Antachopoulos, Meletiadis, Roilides, Sein, Walsh. Rapid Susceptibility Testing
Of Medically Important Zygomycetes By XTT Assay. 2006;44(2):553–60. A Maftei, Nicoleta, A Ramos-Villarroel, A Nicolau, O Martín-Belloso,
And R Soliva-Fortuny. 2014. “Pulsed Light Inactivation Of Naturally Occurring Moulds On Wheat Grain.” Journal Of The Science Of Food And
Agriculture 94 (4): 721–26. Https://Doi.Org/10.1002/Jsfa.6324. Artis, William, J Fountain, H Delcher, And H Jones. 1982. “A
Mechanism Of Susceptibility To Mucormycosis In Diabetic Ketoacidosis: Transferrin And Iron Availability” 31 (December): 1109–14.
Aufauvre-Brown, E Mellado, N Gow, And D Holden. 1997. “Aspergillus Fumigatus Chse: A Gene Related To CHS3 Of Saccharomyces Cerevisiae And Important For Hyphal Growth And Conidiophore Development But Not Pathogenicity.” Fungal Genetics And Biology : FG & B 21 (1): 141–52.
Austin, Bu'Lock, And Gooday. 1969. “Trisporic Acids: Sexual Hormones From Mucor Mucedo And Blakeslea Trispora.” Nature 223 (5211): 1178–79. Https://Doi.Org/10.1038/2231178a0.
Baggio, Gonçalves, Lourenço, Tanaka, Pascholati, And L. Amorim. 2016. “Direct Penetration Of Rhizopus Stolonifer Into Stone Fruits Causing Rhizopus Rot.” Plant Pathology 65 (4): 633–42.
Https://Doi.Org/10.1111/Ppa.12434. Baldin, And Ibrahim. 2017. “Molecular Mechanisms Of
Mucormycosis—The Bitter And The Sweet.” PLOS Pathogens 13 (8): E 1 E1006408. Https://Doi.Org/10.1371/Journal.Ppat.1006408. Ballou And Wilson. 2016. “The Roles Of Zinc And Copper Sensing
In Fungal Pathogenesis.” Current Opinion In Microbiology 32: 128–34. Https://Doi.Org/10.1016/J.Mib.2016.05.013.
182

Balmant, Wellington, Harumi Sugai-Guérios, And Hey Coradin. 2015. “A Model For Growth Of A Single Fungal Hypha Based On Well-Mixed Tanks In Series : Simulation Of Nutrient And Vesicle Transport In Aerial Reproductive Hyphae,” 1–22. Https://Doi.Org/10.1371/Journal.Pone.0120307.
Baltazar, Ludmila, Ray, Santos, Cisalpino, Friedman, And Nosanchuk2015. “Antimicrobial Photodynamic Therapy: An Effective Alternative Approach To Control Fungal Infections.” Frontiers In Microbiology 6 (MAR): 1–11. Https://Doi.Org/10.3389/Fmicb.2015.00202.
Bankevich, Nurk, Antipov, Gurevich, Dvorkin, Kulikov, Et Al. Spades: A New Genome Assembly Algorithm And Its Applications To Single-Cell Sequencing. J Comput Biol [Internet]. 2012;19(5):455–77. Available From: Http://Www.Ncbi.Nlm.Nih.Gov/Pubmed/22506599%0Ahttp://Www.Pubmedcentral.Nih.Gov/Articlerender.Fcgi?Artid=PMC3342519
Barash, Conway, And Howard. 1967. “Carbon Catabolism And Synthesis Of Macromolecules During Spore Germination Of Microsporum Gypseum.”
Journal Of Bacteriology 93 (2): 656–62. Barkai-Golan, Mirelman, And Sharon. 1978. “Studies On Growth Inhibition By L e Lectins Of Penicillia And Aspergilli.” Archives Of Microbiology 116 (2): 119–21. Barkia-Golan and Rivka. 2001. “Postharvest Diseease Initiation: Postharvest
Disease of Fruits and Vegetables.” In POSTHARVEST DISEASES OF FRUITS AND
VEGETABLES, Edited By RIVKA BARKAI-GOLAN. Elsevier. Https://Doi.Org/Https://Doi.Org/10.1016/B978-0-444-50584-2.50020-4.
Barth, Hankinson, Zhuang, And Breidt. 2009. Compendium Of The Microbiological Spoilage Of Foods And Beverages.
Microbiological Spoilage Of Fruits And Vegetables. Springer Science+Business Media. Https://Doi.Org/10.1007/978-1-4419-0826-1.
Bartnicki-Garcia. 1968. “Cell Wall Chemistry, Morphogenesis and Taxonomy of F u n Fungi.” Annual Review Of Microbiology. Bassilana, Martine, Blyth, And Arkowitz. 2003. “Cdc24, The GDP-GTP
Exchange Factor For Cdc42, Is Required For Invasive Hyphal Growth Of Candida A Albicans.” Eukaryotic Cell 2 (1): 9–18. Https://Doi.Org/10.1128/
EC.2.1.9-18.2003.
Hypothesis.”Medical Mycology : Official Publication Of The International
Society For Human And Animal Mycology, No. 906390942: 1–8. Https://Doi.Org/10.1080/13693780802572547.
Baumgardner And Laundre. 2001. “Studies On The Molecular Ecology Of Blastomyces Dermatitidis.” Mycopathologia 152 (2): 51–58.
Bayry, Jagadeesh, Beaussart, Dufrêne, Sharma, Bansal, Kniemeyer, Aimanianda, Et Al. 2014.“Surface Structure Characterization Of Aspergillus Fumigatus Conidia Mutated In The Melanin Synthesis Pathway And Their Human Cellular Immune Response.” Infection And Immunity 82 (8): 3141–53. Https://Doi.Org/10.1128/IAI.01726-14.
Baumgardner. 2009. “Microecology Of Blastomyces Dermatitidis: The Ammonia
183

Beauvais, Bruneau, Mol, Buitrago, Latgé, And Legrand. 2001. “Glucan Synthase Complex Of Aspergillus Fumigatus Glucan Synthase Complex Of Aspergillus Fumigatus.” Journal Of Bacteriology 183 (7): 2273–79. Https://Doi.Org/10.1128/JB.183.7.2273.
Bellanger, Pauline, Tatara, Shirazi, Gebremariam, Albert, Lewis, Ibrahim, And Kontoyiannis. 2016. “Statin Concentrations Below The Minimum Inhibitory Concentration Attenuate The Virulence Of Rhizopus Oryzae.” Journal Of Infectious Diseases 214 (1): 114–21. Https://Doi.Org/10.1093/Infdis/Jiw090.
Belozerskaya, Gessler, Isakova, And Deryabina.
“Aspergillus Fumigatus Inhibits Angiogenesis Through The Production Of Gliotoxin And Other Secondary Metabolites.” Blood 114 (26): 5393–99. Https://Doi.Org/10.1182/Blood-2009-07-231209.
Ben-Ami, Luna, Lewis, Walsh, And Kontoyiannis. 2009. “A Clinicopathological Study Of Pulmonary Mucormycosis In Cancer Patients : Extensive Angioinvasion But Limited Inflammatory Response.” Journal Of
Infection 59 (2): 134–38. Https://Doi.Org/10.1016/J.Jinf.2009.06.002. Bernard, And Latge. 2001. “Aspergillus Fumigatus Cell Wall: Composition And
Biosynthesis.” Med.Mycol. 39 Suppl 1 (September): 9–17. Ht HTtps://Doi.Org/10.1080/Mmy.39.1.9.17. Bertuzzi, Schrettl, Alcazar-Fuoli, Cairns, Muñoz, Walker, Herbst, Et Al. 2014.
“The PH-Responsive Pacc Transcription Factor Of Aspergillus Fumigatus Governs Epithelial Entry And Tissue Invasion During Pulmonary Aspergillosis.” Plos
Pathogens 10 (10). Https://Doi.Org/10.1371/Journal.Ppat.1004413.
Blasi, Barluzzi, Mazzolla, Tancini, Saleppico, Puliti, Pitzurra, And Bistoni. 1995. “Role Of Nitric Oxide And Melanogenesis In The Accomplishment Of Anticryptococcal Activity By The BV-2 Microglial Cell Line.” Journal Of
Neuroimmunology 58 (1): 111–16. Https://Doi.Org/10.1016/0165-5728(95)00016-U.
Boelaert, Johan, De Locht, Van Cutsem, Kerrels, Cantinieaux, Verdonck, Van Landuyt, And Schneidert. 1993.“Mucormycosis During Deferoxamine Therapy Is A Siderophore-Mediated Infection In Vitro And In Vivo Animal Studies” 91 (December 1992): 1979–86.
Ben-Ami, Lewis, Leventakos, And Kontoyiannis. 2009.
Bielska, Higuchi, Schuster, Steinberg, Kilaru, Talbot and Steinberg. 2014"Long-distance endosome trafficking drives fungal effector production during plant infection." Nature Communications. doi: 10.1038/ncomms6097 (2014)
2012. “Neurospora Crassa Light Signal Transduction Is Affected By ROS.” Journal Of Signal Transduction 2012: 1–13. Https://Doi.Org/10.1155/2012/791963.
184

Bonaterra, Mari, Casalini, And Montesinos. 2003. “Biological Control Of Monilinia Laxa And Rhizopus Stolonifer In Postharvest Of Stone Fruit By Pantoea Agglomerans EPS125 And Putative Mechanisms Of Antagonism.” International Journal Of Food Microbiology 84 (1): 93–104.
Bonazzi, Daria, Julien,Romao, Seddiki, Piel, Boudaoud, And Minc. 2014. “Symmetry Breaking In Spore Germination Relies On An Interplay Between Polar Cap Stability And Spore Wall Mechanics.” Developmental Cell 28 (5): 534–46. Https://Doi.Org/10.1016/J.Devcel.2014.01.023.
Bond, Meacham, Bhunnoo, And Benton. 2013. “Food Waste Within Global Food Systems.” Www.Foodsecurity.Ac.Uk.
Böttcher, Bettina, Palige, Jacobsen, Hube, And Brunke. 2015. “Csr1/Zap1 Maintains Zinc Homeostasis And Influences Virulence In Candida Dubliniensis But Is Not Coupled To Morphogenesis.” Eukaryotic Cell 14 (7): 661–70. Https://Doi.Org/10.1128/EC.00078-15.
Botts, Huang, Borchardt, And Hull. 2014.“Developmental Cell Fate And Virulence Are Linked To Trehalose Homeostasis In Cryptococcus Neoformans.” Eukaryotic Cell 13 (9): 1158–68. Https://Doi.Org/10.1128/EC.00152-14.
Boyce And Andrianopoulos. 2007. “A P21-Activated Kinase Is Required For Conidial Germination In Penicillium Marneffei.” Plos Pathogens 3 (11): 1556–69. Https://Doi.Org/10.1371/Journal.Ppat.0030162.
Boyce And Andrianopoulos. 2015. “Fungal Dimorphism: The Switch From Hyphae To Yeast Is A Specialized Morphogenetic Adaptation Allowing Colonization Of A Host.” FEMS Microbiology Reviews 39 (6): 797–811. Https://Doi.Org/10.1093/Femsre/Fuv035.
Boyce, Hynes, And Andrianopoulos. 2005. “The Ras And Rho Gtpases Genetically Interact To Co-Ordinately Regulate Cell Polarity During Development In Penicillium Marneffei.” Molecular Microbiology 55 (5): 1487–1501. Https://Doi.Org/10.1111/J.1365-2958.2005.04485.X.
Boyce, Schreider, Kirszenblat, And Andrianopoulos. 2011. “The Two-Component Histidine Kinases Drka And Slna Are Required For In Vivo Growth In The Human Pathogen Penicillium Marneffei.” Molecular Microbiology 82 (5): 1164–84. Https://Doi.Org/10.1111/J.1365-2958.2011.07878.X.
Breeuwer, De Reu, Drocourt, Rombouts, And Abee. 1997. “Nonanoic Acid, A Fungal Self-Inhibitor, Prevents Germination Of Rhizopus Oligosporus Sporangiospores By Dissipation Of The PH Gradient.” Applied And Environmental
Microbiology 63 (1): 178–85. Https://Www.Ncbi.Nlm.Nih.Gov/Pubmed/16535482.
Brengues, Pintard, And Lapeyre. 2002. “MRNA Decay Is Rapidly Induced After Spore Germination Of Saccharomyces Cerevisiae.” The Journal Of
Biological Chemistry 277 (43): 40505–12. Https://Doi.Org/10.1074/Jbc.M206700200.
185

Briza, Ellinger, Winkler, And Breitenbach. 1988. “Chemical Composition Of The Yeast Ascospore Wall. The Second Outer Layer Consists Of Chitosan.” Journal
Of Biological Chemistry 263 (23): 11569–74. Broadinstitute. Picard Tools [Internet]. Http://Broadinstitute.Github.Io/Picard/. [Cited
Brown, Denning, Gow, Levitz, Netea, And White. 2012.
Fungus Fusarium Fujikuroi Is A Light-Driven Proton Pump That Retards Spore Germination.” Scientific Reports, 1–11. Https://Doi.Org/10.1038/Srep07798.
Bruno, Aramayo, Minke, Metzenberg, And Plamann. 1996. “Loss Of Growth Polarity And Mislocalization Of Septa In A Neurospora Mutant Altered In The Regulatory Subunit Of CAMP-Dependent Protein Kinase.” The EMBO Journal 15 (21): 5772–82.
Cano And Ruiz-Herrera. 1988. “Developmental Stages During The Germination Of Mucor Sporangiospores.” Experimental Mycology 12 (1): 47–59.
Chakrabarti, Arunaloke, And Singh. 2014. “Mucormycosis In India: Unique Features.”
Chamilos, Lewis, G Lamaris, Walsh, And Kontoyiannis. 2008. “Zygomycetes Hyphae Trigger An Early , Robust Proinflammatory Response In Human Polymorphonuclear Neutrophils Through Toll-Like Receptor 2 Induction But Display Relative Resistance To Oxidative Damage �” 52 (2): 722–24. Https://Doi.Org/10.1128/AAC.01136-07.
Chanclud And Benoit Morel. 2016. “Plant Hormones: A Fungal Point Of View.” Molecular Plant Pathology 17 (8): 1289–97.
Chibucos, Soliman, Gebremariam, Lee, Daugherty, Shetty, Crabtree, Et Al. 2016.“An Integrated Genomic And Transcriptomic Survey Of Mucormycosis-Causing Fungi.” Nature Publishing Group 7: 1–11. Https://Doi.Org/10.1038/Ncomms12218.
Choi, Jaehyuk, Hee Jung, Kronstad, Culture Collection, Incheon National, Michael Smith Laboratories, And Food Systems. 2015. “The CAMP/Protein Kinase A Signaling Pathway In Pathogenic Basidiomycete Fungi: Connections With Iron Homeostasis.” Journal Microbiology 53 (9): 579–87. Https://Doi.Org/10.1007/S12275-015-5247-5.The.
Cornet And Gaillardin. 2014. “PH Signaling In Human Fungal Pathogens: A New Target For Antifungal Strategies.” Eukaryotic Cell 13 (3): 342–52. Https://Doi.Org/10.1128/EC.00313-13.
D’Souza, Cletus, And Heitman. 2001. “Conserved CAMP Signaling Cascades Regulate Fungal Development And Virulence.” FEMS Microbiology Reviews 25 (3):
2019 Jul 4]. Available From: Http://Broadinstitute.Github.Io/Picard/
“Hidden Killers : Human Fungal Infections” 13. Https://Doi.Org/10.1126/Scitranslmed.3004404.
Brunk, Avalos, Terpitz, And Garcıa-Martinez. 2015. “The Caro Rhodopsin Of The
Brunke And Hube. 2012. "Two unlike cousins: Candida albicans and C. glabrata infection strategies" Cellular Microbiology. https://doi.org/10.1111/cmi.12091
Mycoses 57 (S3): 85–90. Https://Doi.Org/10.1111/Myc.12243.
Https://Doi.Org/10.1016/0147-5975(88)90015-1.
Https://Doi.Org/10.1111/Mpp.12393.
186

349–64. Https://Doi.Org/10.1016/S0168-6445(01)00058-4. Dague, Etienne, Alsteens, Latgé, And Dufrêne. 2008. “High-
Resolution Cell Surface Dynamics Of Germinating Aspergillus Fumigatus Conidia.” Biophysical Journal 94 (2): 656–60.
“Aspergillus Terreus Accessory Conidia Are Unique In Surface Architecture, Cell Wall Composition And Germination Kinetics.” Plos ONE 4 (10): 1–7. Https://Doi.Org/10.1371/Journal.Pone.0007673.
Degani, Ofir, Drori, And Goldblat. 2015. “Plant Growth Hormones Suppress The Development Of Harpophora Maydis, The Cause Of Late Wilt In Maize.” Physiology And Molecular Biology Of Plants 21 (1): 137–49. Https://Doi.Org/10.1007/S12298-014-0265-Z.
Deng, Ming, Yang, Wei Di He, Yu Li, Yang, Wu Zuo, Gao, Et Al. 2015.“Proteomic Analysis Of Conidia Germination In Fusarium Oxysporum F. Sp. Cubense Tropical Race 4 Reveals New Targets In Ergosterol Biosynthesis Pathway For Controlling Fusarium Wilt Of Banana.” Applied Microbiology
And Biotechnology 99 (17): 7189–7207. Https://Doi.Org/10.1007/S00253-015-6768-X.
Denison 2000. “PH Regulation Of Gene Expression In Fungi.” Fungal Genetics And
Biology : FG & B 29 (2): 61–71. Https://Doi.Org/10.1006/Fgbi.2000.1188. Dijksterhuis, Nijsse, Hoekstra, And Golovina. 2007. “High Viscosity And
Dufrêne, Boonaert, Gerin, Asther, And Rouxhet. 1999.“Direct Probing Of The Surface Ultrastructure And Molecular Interactions Of Dormant And Germinating Spores Of Phanerochaete Chrysosporium.” Journal
Of Bacteriology 181 (17): 5350–54. Dussutour, Latty, Beekman, And Simpson. 2010. “Amoeboid Organism
Solves Complex Nutritional Challenges.” Proceedings Of The National Academy Of
Sciences 107 (10): 4607–11. Https://Doi.Org/10.1073/Pnas.0912198107. Eckert And Sommer. 1967. “Control Of Diseases Of Fruits And Vegetables By Post-
Harvest Treatment.” Annual Review Of Phytopathology 5 (1): 391–428. Https://Doi.Org/10.1146/Annurev.Py.05.090167.002135.
Eckert And Ogawa. 1989. “Chemical Control Of POSTHARVEST DISEASES : Deciduous Fruits , Berries , Vegetables And Root / Tuber Crops.” Annual Review Of Phytopathology 26 (64): 433–69.
Eisendle, Schrettl, Kragl, Müller, Illmer, And Haas. 2006.“The Intracellular Siderophore Ferricrocin Is Involved In Iron Storage, Oxidative-Stress Resistance, Germination, And Sexual Development
Https://Doi.Org/10.1529/Biophysj.107.116491. Dalmont, Biles, Konsure, Dahal, Rowsey, Broge, Poudyal, Gurung, Shrestha, Cluck, Howard.
Dao And Dantigny. 2011. “Control Of Food Spoilage Fungi By Ethanol.”
Deak, Eszter, Wilson, White, Carr, And Balajee. 2009.
2017."Nonsteroidal Anti-inflammatory Drugs (NSAIDS) Inhibit the Growth and Reproduction of Chaetomium globosum and Other Fungi Associated with Water-Damaged Buildings" Mycopathologia. doi: 10.1007/s11046-017-0188-7
Food Control 22 (3–4): 360–68. Https://Doi.Org/10.1016/J.Foodcont.2010.09.019.
Anisotropy Characterize The Cytoplasm Of Fungal Dormant Stress-Resistant Spores.” Eukaryotic Cell 6 (2): 157–70. Https://Doi.Org/10.1128/EC.00247-06.
187
Dill, Jung And Sun. 2001. "The DELLA motif is essential for gibberellin-induced degradation of RGA" PNAS. https://doi.org/10.1073/pnas.251534098

In Aspergillus Nidulans.” Eukaryotic Cell 5 (10): 1596–1603.
Eisenman, Frases, André , Rodrigues, And Casadevall. 2009.“Vesicle-Associated Melanization In Cryptococcus Neoformans.” Microbiology 155 (12): 3860–67. Https://Doi.Org/10.1099/Mic.0.032854-0.
Eisenman, Siu-Kei Chow, Tsé, Mcclelland, And Casadevall. 2011. “The Effect Of L-DOPA On Cryptococcus Neoformans Growth And Gene Expression,” No. August: 329–36. Https://Doi.Org/10.4161/Viru.2.4.16136.
Eisenman, Nosanchuk, Webber, Emerson, Camesano, And Casadevall. 2005. “Microstructure Of Cell Wall-Associated Melanin In The Human Pathogenic Fungus Cryptococcus Neoformans.” Biochemistry 44 (10): 3683–93. Https://Doi.Org/10.1021/Bi047731m.
Elbein 1974. “The Metabolism Of Α,Α-Trehalose.” In , 227–56. Https://Doi.Org/10.1016/S0065-2318(08)60266-8.
Feofilova, Ivashechkin, Alekhin, And Sergeeva. 2012. “Fungal Spores: Dormancy, Germination, Chemical Composition, And Role In Biotechnology (Review).” Applied Biochemistry And Microbiology 48 (1): 1–11. Https://Doi.Org/10.1134/S0003683812010048.
Feofilova, Daragan-Sushchova, Volokhova, Velichko, Shirokova, And Sinitsin. 1988. “Changes In The Chemical Composition Of Aspergillus
Feofilova, Sergeeva, Mysyakina, And Bokareva. 2015. “Lipid Composition In Cell Walls And In Mycelial And Spore Cells Of Mycelial Fungi” 84
Forsythe, Vogan, And Xu. 2016. “Genetic And Environmental Influences On The Germination Of Basidiospores In The Cryptococcus Neoformans Species Complex.” Nature Publishing Group, No. August: 1–12. Https://Doi.Org/10.1038/Srep33828.
Fortwendel, Jarrod 2016. “Orchestration Of Morphogenesis In Filamentous Fungi: Conserved Roles For Ras Signaling Networks.” Fungal Biology Reveiws 29 (2): 54–62. Https://Doi.Org/10.1016/J.Fbr.2015.04.003.Orchestration.
Fortwendel, Jarrod, Wei Zhao, Bhabhra, Park, Perlin, Askew, And Rhodes. 2005. “A Fungus-Specific Ras Homolog Contributes To The Hyphal Growth And Virulence Of Aspergillus Fumigatus.” Eukaryotic Cell 4 (12): 1982 LP – 1989. Https://Doi.Org/10.1128/EC.4.12.1982-1989.2005.
Fox And Fimeche. 2013. “Global Food: Waste Not, Want Not.” Institute Of
Mechanical Engineers, London, Jan, 1–14. Franken, Angelique, Lechner, Werner, Haas, Lokman, Ram, Van Den Hondel,
De Weert, And Punt. 2014. “Genome Mining And Functional Genomics For Siderophore Production In Aspergillus Niger.” Briefings In Functional Genomics 13
Https://Doi.Org/10.1128/EC.00057-06.
Japonicus Cells In The Course Of Their Growth.” Microbiologiya 57: 778–84.
(2): 170–76. Https://Doi.Org/10.1134/S0026261715020046.
188

(6): 482–92. Https://Doi.Org/10.1093/Bfgp/Elu026. Franklin, Keara, And Quail. 2010. “Phytochrome Functions In Arabidopsis
Development.” Journal Of Experimental Botany 61 (1): 11–24. Https://Doi.Org/10.1093/Jxb/Erp304.
Fuchino, K., S. Bagchi, S. Cantlay, L. Sandblad, D. Wu, J. Bergman, M. Kamali-Moghaddam, Flardh, And Ausmees. 2013. “Dynamic Gradients Of An Intermediate Filament-Like Cytoskeleton Are Recruited By A Polarity Landmark During Apical Growth.” Proceedings Of The National Academy Of Sciences 110 (21): E1889–97. Https://Doi.Org/10.1073/Pnas.1305358110.
Fuller, Ringelberg, Loros, And Dunlap. 2013. “The Fungal Pathogen Aspergillus Fumigatus Regulates Growth , Metabolism , And Stress Resistance In Response To Light.” Mbio 4 (2): 11–13. Https://Doi.Org/10.1128/Mbio.00142-13.Editor.
Galhano, Illana, Ryder, Rodríguez-Romero, Demuez, Badaruddin, Martinez-Rocha, Et Al. 2017."Tpc1 Is An Important Zn(II)2Cys6transcriptional Regulator Required For
Polarized Growth And Virulence In The Rice Blast Fungus." Plos Pathogens. Vol. 13. Https://Doi.Org/10.1371/Journal.Ppat.1006516.
Garcia-Martinez, Brunk, Avalos, And Terpitz. 2015. “The Caro Rhodopsin Of The Fungus Fusarium Fujikuroi Is A Light-Driven Proton Pump That Retards Spore Germination.” Scientific Reports 5 (January): 7798. Https://Doi.Org/10.1038/Srep07798.
Garcia, Alexis, Vellanki, And Lee. 2018. “Genetic Tools For Investigating Mucorales Fungal Pathogenesis.” Current Clinical Microbiology
Garfoot And Rappleye. 2016. “Histoplasma Capsulatum Surmounts Obstacles To Intracellular Pathogenesis.” The FEBS Journal 283: 619–33.
Https://Doi.Org/10.1111/Febs.13389. Gauthier, Sullivan, Gallardo, Brandhorst, Vanden Wymelenberg, Cuomo, Suen, Currie,
And Klein. 2010. “SREB, A GATA Transcription Factor That Directs Disparate Fates In Blastomyces Dermatitidis Including Morphogenesis And Siderophore Biosynthesis.” Plos Pathogens 6 (4): E1000846. Https://Doi.Org/10.1371/Journal.Ppat.1000846.
Gebremariam, Lin, Liu, Kontoyiannis, French, Edwards Jr, Filler, And Ibrahim. 2016. “Bicarbonate Correction Of Ketoacidosis Alters Host- Pathogen Interactions And Alleviates Mucormycosis” 126 (6). Https://Doi.Org/10.1172/JCI82744.Mice.
Gebremariam, Liu, Luo, Bruno, Phan, Waring, Edwards Jr, Filler, Yeaman,And Ibrahim. 2014. “Coth3 Mediates Fungal Invasion Of Host Cells During Mucormycosis” 124 (1). Https://Doi.Org/10.1172/JCI71349.In.
Ghamrawi, Gastebois, Zykwinska, Vandeputte, Marot, Mabilleau, Cuenot, And Bouchara. 2015.
Galgiani. 2016. "The Search for the Cure for Valley Fever Nikkomycin Z Development at the University of Arizona", https://vfce.arizona.edu/sites/vfce/files/bio5_summary_nikz_development_plan_1.pdf
Reports 5 (3): 173–80. Https://Doi.Org/10.1007/S40588-018-0097-7.
189

“A Multifaceted Study Of Scedosporium Boydii Cell Wall Changes During Germination And Identification Of GPI-Anchored Proteins.” Plos ONE 10 (6): 1–24. Https://Doi.Org/10.1371/Journal.Pone.0128680.
Ghuman, Voelz. 2017. “Innate And Adaptive Immunity To Mucorales.” Journal Of Fungi 3 (4): 48. Https://Doi.Org/10.3390/Jof3030048.
Giles, Steven, And Czuprynski. 2004. “Extracellular Calcium And Magnesium, But Not Iron, Are Needed For Optimal Growth Of Blastomyces Dermatitidis Yeast Form Cells In Vitro.” Clinical And Diagnostic Laboratory
Immunology 11 (2): 426–29. Https://Doi.Org/10.1128/CDLI.11.2.426-429.2004. Gilmore, Naseem, Konopka, And Sil. 2013. “N-Acetylglucosamine (Glcnac)
Triggers A Rapid, Temperature-Responsive Morphogenetic Program In Thermally Dimorphic Fungi.” Plos Genetics 9 (9). Https://Doi.Org/10.1371/Journal.Pgen.1003799.
Glenn. 2006. “Natural Variation Of Ascospore And Conidial Germination By Fusarium Verticillioides And Other Fusarium Species.” Mycological Research 110: 211–19. Https://Doi.Org/10.1016/J.Mycres.2005.09.004.
Gomes, Marisa, Lewis, And Kontoyiannis. 2011. “Mucormycosis Caused By Unusual Mucormycetes, Non-Rhizopus, -Mucor, And -Lichtheimia Species.” Clinical Microbiology Reviews 24 (2): 411–45. Https://Doi.Org/10.1128/CMR.00056-10.
Grabherr, Nir, Haas, Yassour, Levin, Thompson, Amit, Adiconis, Fan, Raychowdhury,Zeng, Chen, Evan Mauceli, Nir Hacohen, Andreas Gnirke, Nicholas Rhind, Federica Di Palma, Bruce W., And And Regev Friedman. 2013. “Trinity: Reconstructing A Full-Length Transcriptome Without A Genome From RNA-Seq Data.” Nature Biotechnology 29 (7): 644–52. Https://Doi.Org/10.1038/Nbt.1883.Trinity.
Gresnigt, Becker, Leenders, Alonso, Wang, Meis, Bain, Erwig, And Van De Veerdonk. 2018. “Differential Kinetics Of Aspergillus Nidulans And Aspergillus Fumigatus Phagocytosis.” Journal Of Innate Immunity 10 (2): 145–60. Https://Doi.Org/10.1159/000484562.
Griffin 1996. Fungal Physiology. John Wiley & Sons. Gryganskyi, Andrii , Golan, Dolatabadi, Mondo, Robb, Idnurm, Muszewska, Et Al. 201 2018."Phylogenetic And Phylogenomic Definition Of Rhizopus Species.” G3
(Bethesda, Md.) 8 (6): 2007–18. Https://Doi.Org/10.1534/G3.118.200235.
Gryganskyi, Andrii, Lee, Litvintseva, Smith, Bonito, Porter, Anishchenko, Heitman, And Vilgalys. 2010. “Structure, Function, And Phylogeny Of The Mating Locus In The Rhizopus Oryzae Complex.” PLOS ONE 5 (12): E15273. Https://Doi.Org/10.1371/Journal.Pone.0015273.
190

Hagiwara, Daisuke, Takahashi, Kusuya, Kawamoto, Kamei, And Gonoi. 2016. “Comparative Transcriptome Analysis Revealing Dormant Conidia And Germination Associated Genes In Aspergillus Species: An Essential Role For Atfa In Conidial Dormancy.” BMC Genomics 17 (1): 358. Https://Doi.Org/10.1186/S12864-016-2689-Z.
Haas, Zeng, Pearson, Cuomo, Wortman. Approaches To Fungal Genome Annotation. Mycology. 2011;2(3):118–41.
Hassouni, Hicham, Ismaili-Alaoui, Lamrani, Gaime-Perraud, Augur, And Roussos. 2 2007.“Comparative Spore Germination Of Filamentous Fungi On Solid State
Fermentation Under Different Culture Conditions.” Micologia Aplicada
International 19 (1): 7–14. Http://Www.Documentation.Ird.Fr/Intranet/Publi/Depot/2010-06-03/010048975.Pdf%5Cn%3cgo To ISI%3E://BIOABS:BACD200700176791.
Hayer, Kimran, Stratford, And Archer. 2013. “Structural Features Of Sugars That Trigger Or Support Conidial Germination In The Filamentous Fungus Aspergillus Niger.” Applied And Environmental Microbiology 79 (22): 6924–31. Https://Doi.Org/10.1128/AEM.02061-13.
Hedayati, Pasqualotto,Warn, Bowyer, And Denning. 2007. “Aspergillus Flavus: Human Pathogen, Allergen And Mycotoxin Producer.” Microbiology (Reading, England) 153 (Pt 6): 1677–92. Https://Doi.Org/10.1099/Mic.0.2007/007641-0.
Herrera-Estrella, Alfredo, And Horwitz. 2007. “Looking Through The Eyes Of Fungi: Molecular Genetics Of Photoreception.” Molecular Microbiology 64
Hoff, Lange, Lomsadze, Borodovsky, And Stanke. 2016. “BRAKER1: Unsupervised RNA-Seq-Based Genome Annotation With Genemark-ET And AUGUSTUS.” Bioinformatics 32 (5): 767–69. Https://Doi.Org/10.1093/Bioinformatics/Btv661.
Hoffmann, Benny, Kirk, And Voigt. 2013. “The Family Structure Of The Mucorales : A Synoptic Revision Based On Comprehensive Multigene-Genealogies,” 57–76.
Hogan 2006. “Talking To Themselves : Autoregulation And Quorum Sensing In Fungi MINIREVIEW Talking To Themselves : Autoregulation And Quorum Sensing In Fungi.” Eukaryotic Cell 5 (4): 613–19. Https://Doi.Org/10.1128/EC.5.4.613.
Hoi, Lamarre, Beau, Meneau, Berepiki, Barre, Mellado, Read, And Latge. 2011. “A Novel Family Of Dehydrin-Like Proteins Is Involved In Stress Response In The Human Fungal Pathogen Aspergillus Fumigatus.” Molecular Biology Of The Cell 22 (11): 1896–1906. Https://Doi.Org/10.1091/Mbc.E10-11-0914.
(1): 5–15. Https://Doi.Org/10.1111/J.1365-2958.2007.05632.X.
191

Horn, Üzüm, Möbius, Guthke, Linde, And Hertweck. 2015. “Draft Genome Sequences Of Symbiotic And Nonsymbiotic Rhizopus Microsporus Strains CBS 344.29 And ATCC 62417.” Genome
Announcements 3 (1): E01370-14. Https://Doi.Org/10.1128/Genomea.01370-14.
Houser, Komarek, Kostlanova, Cioci, Varrot, Kerr, Lahmann, Et Al. 2013.“A Soluble Fucose-Specific Lectin From Aspergillus Fumigatus Conidia - Structure, Specificity And Possible Role In Fungal Pathogenicity.” Plos ONE 8 (12): 1–15. Https://Doi.Org/10.1371/Journal.Pone.0083077.
Howard, Dabrowa, Otto, And Rhodes. 1980. “Cysteine Transport And Sulfite Reductase Activity In A Germination-Defective Mutant Of Histoplasma
Capsulatum.” Journal Of Bacteriology 141 (1): 417–21.
Following Intracellular Sorting. Plos One. 2014;9(3):1–6. Huang And Hull. 2017. “Sporulation: How To Survive On Planet
Earth (And Beyond).” Current Genetics, 1–8. Https://Doi.Org/10.1007/S00294-017-0694-7.
Ibrahim. 2011. “Host Cell Invasion In Mucormycosis : Role Of Iron.” Current
Opinion In Microbiology 14 (4): 406–11.
Ibrahim, Gebermariam, Fu, Lin, Husseiny, French, Schwartz, Skory, Edwards Jr, And Spellberg. 2007. “The Iron Chelator Deferasirox Protects Mice From Mucormycosis Through Iron Starvation” 117 (9): 2649–57. Https://Doi.Org/10.1172/JCI32338.The.
Ibrahim, Gebremariam, Lin, Luo, Husseiny, Skory, Fu, French,Edwards, And Spellberg. 2010. “The High Affinity Iron Permease Is A Key Virulence Factor Required For Rhizopus Oryzae Pathogenesis.” Molecular Microbiology 77 (June 2010): 587–604. Https://Doi.Org/10.1111/J.1365-2958.2010.07234.X.
Ibrahim, Gebremariam, Liu, Chamilos, Kontoyiannis, Mink, Kwon-Chung, Et Al.
2008. “Bacterial Endosymbiosis Is Widely Present Among Zygomycetes But Does Not Contribute To The Pathogenesis Of Mucormycosis.” The Journal Of
Infectious Diseases 198 (7): 1083–90. Https://Doi.Org/10.1086/591461.
Ibrahim, Spellberg, And Edwards. 2016. “Iron Acquisition : A Novel Perspective On Mucormycosis Pathogenesis And Treatment.”
Ibrahim, Spellberg, Walsh, And P Kontoyiannis. 2012. “Pathogenesis Of Mucormycosis.” Clinical Infectious Diseases 54 (Suppl 1): 1–7. Https://Doi.Org/10.1093/Cid/Cir865.
Idnurm And Heitman. 2005a. “Light Controls Growth And
Hrvatin S, Deng F,O’Donnell, Gifford, Melton. MARIS: Method For Analyzing RNA
Https://Doi.Org/10.1016/J.Mib.2011.07.004.
Https://Doi.Org/10.1097/QCO.0b013e3283165fd1.
192

Development Via A Conserved Pathway In The Fungal Kingdom.” Plos Biology 3
Idnurm And Heitman. 2005b. “Photosensing Fungi: Phytochrome In The Spotlight.” Current Biology 15 (20): 829–32.
Ignatov, Salina, Fursov, Skvortsov, Azhikina, And Kaprelyants. 2015. “Dormant Non-Culturable Mycobacterium Tuberculosis Retains Stable Low-Abundant MRNA.” BMC Genomics 16 (November): 954. Https://Doi.Org/10.1186/S12864-015-2197-6.
Illmer, Erlebach, And Schinner. 1999. “A Practicable And Accurate Method To Differentiate Between Intra- And Extracellular Water Of Microbial Cells.” FEMS
Microbiology Letters 178 (1): 135–39. Https://Doi.Org/10.1016/S0378-1097(99)00351-1.
Inglesfield, Jasiulewicz, Hopwood, Tyrrell, Youlden, Mazon-Moya, Millington, Mostowy, Voelz And, Jabbari 2018. “Robust Phagocyte Recruitment Controls The Opportunistic Fungal Pathogen Mucor Circinelloides In Innate Granulomas In Vivo” 9 (2): 1–20.
Itabangi, Sephton-Clark, Zhou, Insua, Probert, Correia, Moynihan, Et Al. 2019.
Jay 1992. “Spoilage Of Fruits And Vegetables.” In Modern Food
Microbiology, 187–98. Dordrecht: Springer Netherlands.
Kamada, Bracker, And Bartnicki-Garcia. 1991. “Chitosomes And Chitin Synthetase In The Asexual Life Cycle Of Mucor Rouxii: Spores, Mycelium And Yeast Cells.” Journal Of General Microbiology 137 (6): 1241–52. Https://Doi.Org/10.1099/00221287-137-6-1241.
Karkowska-Kuleta, Justyna, And Kozik. 2015. “Cell Wall Proteome Of Pathogenic Fungi.” ABP 62 (3).
Karp, Latendresse, Paley, Krummenacker, Ong, Billington, Et Al. Pathway Tools Version 19.0 Update: Software For Pathway/Genome Informatics
Kasuga, Takao, Townsend, Tian, Gilbert, Mannhaupt, Taylor, And Glass. 2005. “Long-Oligomer Microarray Profiling In Neurospora Crassa Reveals The Transcriptional Program Underlying Biochemical And Physiological Events Of Conidial Germination.” Nucleic Acids Research 33 (20): 6469–85. Https://Doi.Org/10.1093/Nar/Gki953.
Khunyoshyeng, Sauvarat, Cheevadhanarak, Rachdawong, And
(4): 0615–26. Https://Doi.Org/10.1371/Journal.Pbio.0030095.
Https://Doi.Org/10.1016/J.Cub.2005.09.045.
Ikeh, Ahmed and Quinn. 2017. "Phosphate Acquisition and Virulence in Human Fungal Pathogens." Microorganisms. doi: 10.3390/microorganisms5030048
Https://Doi.Org/10.1007/978-94-011-6480-1_8.
And Systems Biology. Brief Bioinform. 2016;17(5):877–90
193
“A Bacterial Endosymbiont Enables Fungal Immune Evasion During Fatal Mucormycete Infection.” Biorxiv.
Mercier, Et Al. 2018. “Biosynthesis Of Abscisic Acid In Fungi: Identification Of A Sesquiterpene Cyclase As The Key Enzyme In Botrytis Cinerea.” Environmental Microbiology 20 (7): 2469–82. Https://Doi.Org/10.1111/1462-2920.14258.
Izquierdo-Bueno, Inmaculada, Gonzalez-Rodriguez, Simon, Dalmais, Pradier, Pecheur,

Tanticharoen. 2002. “Differential Expression Of Desaturases And Changes In Fatty Acid Composition During Sporangiospore Germination And Development In Mucor Rouxii.” Fungal Genetics And Biology 37 (1): 13–21. Https://Doi.Org/10.1016/S1087-1845(02)00028-2.
Kistler, Corby, And Broz. 2015. “Cellular Compartmentalization Of Secondary Metabolism .” Frontiers In Microbiology .
Kim, Langmead, Salzberg. HISAT: A Fast Spliced Aligner With Low Memory Requirements. Nat Methods. 2015;12(4):357–60.
Kobayashi And Crouch. 2009. “Bacterial/Fungal Interactions: From Pathogens To Mutualistic Endosymbionts.” Annual Review Of Phytopathology 47:
Kolattukudy, Pappachan, Rogers, Li, Hwang, And Flaishman. 1995.“Surface Signaling In Pathogenesis.” PNAS 92 (May): 4080–87.
Kono, Keiko, Matsunaga, Hirata, Suzuki, Abe, And Ohya. 2005. “Involvement Of Actin And Polarisome In Morphological Change During Spore Germination Of Saccharomyces Cerevisiae.” Yeast 22 (2): 129–39. Https://Doi.Org/10.1002/Yea.1205.
Kontoyiannis, Lewis, Lotholary, Spellberg, Petrikkos, Roillides, Ibrahim, And Walsh. 2012. “Future Directions In Mucormycosis Research.” Clinical Infectious Diseases 54 (Suppl 1): 79–85. Https://Doi.Org/10.1093/Cid/Cir886.
Kousser, Clark, Sherrington, Voelz, And Hall. 2019. “Pseudomonas Aeruginosa Inhibits Rhizopus Microsporus Germination Through Sequestration Of Free Environmental Iron.” Scientific Reports 9 (1): 5714. Https://Doi.Org/10.1038/S41598-019-42175-0.
Kraibooj, Kaswara, Reung Park, Dahse, Skerka, Voigt, And Figge. 2014. “Virulent Strain Of Lichtheimia Corymbifera Shows Increased Phagocytosis By Macrophages As Revealed By Automated Microscopy Image Analysis.” Mycoses 57 (S3): 56–66. Https://Doi.Org/10.1111/Myc.12237.
Krol, Igielski, Pollmann, And Kepczynska. 2015. “Priming Of Seeds With Methyl Jasmonate Induced Resistance To Hemi-Biotroph Fusarium Oxysporum F.Sp. Lycopersici In Tomato Via 12-Oxo-Phytodienoic Acid, Salicylic Acid, And Flavonol Accumulation.” Journal Of Plant Physiology 179 (May): 122–32. Https://Doi.Org/10.1016/J.Jplph.2015.01.018.
Kruppa, Krom, Chauhan, Bambach, Cihlar, Richard, And Calderone. 2004. “The Two-Component Signal Transduction Protein Chk1p Regulates Quorum Sensing In Candida Albicans The Two-Component Signal Transduction Protein Chk1p Regulates Quorum Sensing In Candida Albicans” 3 (4): 1062–65. Https://Doi.Org/10.1128/EC.3.4.1062.
Kulkarni, Guruprasad, Sanjeevkumar, Kirankumar, Santoshkumar, And
Https://Www.Frontiersin.Org/Article/10.3389/Fmicb.2015.00068.
63–82. Https://Doi.Org/10.1146/Annurev-Phyto-080508-081729.
194

Karegoudar. 2013. “Indole-3-Acetic Acid Biosynthesis In Fusarium Delphinoides Strain GPK, A Causal Agent Of Wilt In Chickpea.” Applied Biochemistry And
Biotechnology 169 (4): 1292–1305. Https://Doi.Org/10.1007/S12010-012-0037-6.
Lamarre, okol, Debeaupuis, Henry, Lacroix, Glaser, Coppée, François, And Latgé. 2008. “Transcriptomic Analysis Of The Exit From Dormancy Of Aspergillus Fumigatus Conidia.” BMC Genomics 15: 1–15. Https://Doi.Org/10.1186/1471-2164-9-417.
Lambou, Lamarre, Beau, Dufour, And Latge. 2010.“Functional Analysis Of The Superoxide Dismutase Family In Aspergillus Fumigatus.” Molecular Microbiology 75 (4): 910–23. Https://Doi.Org/10.1111/J.1365-2958.2009.07024.X.
Lanternier And Lortholary. 2009. “Zygomycosis And Diabetes Mellitus.” Clinical
Microbiology And Infection. Lee And Heitman. 2014. “Sex In The Mucoralean Fungi.” Mycoses
57 Suppl 3 (0 3): 18–24. Https://Doi.Org/10.1111/Myc.12244. Lee, Li, Calo, And Heitman. 2013. “Calcineurin Plays Key
Roles In The Dimorphic Transition And Virulence Of The Human Pathogenic Zygomycete Mucor Circinelloides” 9 (9). Https://Doi.Org/10.1371/Journal.Ppat.1003625.
Leeuwen, Krijgsheld, Bleichrodt, Menke, Stam, Stark, Wösten, And Dijksterhuis. 2013. “Germination Of Conidia Of Aspergillus Niger Is Accompanied By Major Changes In RNA Profiles.” Studies In Mycology 74: 59–70. Https://Doi.Org/10.3114/Sim0009.
Leeuwen, Smant, Boer, And Dijksterhuis. 2008. “Filipin Is A Reliable In Situ Marker Of Ergosterol In The Plasma Membrane Of Germinating Conidia (Spores) Of Penicillium Discolor And Stains Intensively At The Site Of Germ Tube Formation.” Journal Of Microbiological Methods 74 (2–3): 64–73. Https://Doi.Org/10.1016/J.Mimet.2008.04.001.
Leighton And Stock. 1969. “Heat-Induced Macroconidia Germination In Microsporum Gypseum.” Appl Microbiol 17 (3): 473–75. Http://Www.Ncbi.Nlm.Nih.Gov/Entrez/Query.Fcgi?Cmd=Retrieve&Db=Pubmed&Dopt=Citation&List_Uids=5780403.
Leonhardt, Spielberg, Weber, Albrecht-Eckardt, Bläss, Claus, Dagmar Barz, Et Al. 2015. “The Fungal Quorum-Sensing Molecule Farnesol Activates Innate Immune Cells But Suppresses Cellular Adaptive Immunity.” Edited By Mihai Zychlinsky Netea Arturo. Mbio 6 (2): E00143-15. Https://Doi.Org/10.1128/Mbio.00143-15.
Lew, Roger And Levina. 2004. “Oxygen Flux Magnitude And Location Along Growing Hyphae Of Neurospora Crassa.” FEMS Microbiology Letters 233
Lewis, Russell, Pongas, Albert, Ben-Ami, Walsh, And Kontoyiannis. 2011. “Activity Of Deferasirox In Mucorales: Influences Of Species And Exogenous Iron.”
Lackner And Hertweck. 2011. "Impact of Endofungal Bacteria on Infection Biology, Food Safety, and Drug Development."PlosPathogens. doi: 10.1371/journal.ppat.1002096
(1): 125–30. Https://Doi.Org/10.1016/J.Femsle.2004.01.050.
195

Antimicrobial Agents And Chemotherapy 55 (1): 411–13. Https://
2018. “Reactive Oxygen Species And Gibberellin Acid Mutual Induction To Regulate Tobacco Seed Germination.” Frontiers In Plant Science 9 (October): 1279. Https://Doi.Org/10.3389/Fpls.2018.01279.
“Fob1 And Fob2 Proteins Are Virulence Determinants Of Rhizopus Oryzae Via Facilitating Iron Uptake From Ferrioxamine,” 1–33. Https://Doi.Org/10.1371/Journal.Ppat.1004842.
Liu, Muxing, Bruni, Taylor, Zhang, And Wang 2018a.“Comparative Genome-Wide Analysis Of Extracellular Small Rnas From The Mucormycosis Pathogen Rhizopus Delemar.” Scientific Reports 8 (1): 5243. Https://Doi.Org/10.1038/S41598-018-23611-Z.
Loeb, Kerentseva, Pan, Sepulveda-Becerra, And Liu. 1999.
“Saccharomyces Cerevisiae G1 Cyclins Are Differentially Involved In Invasive And Pseudohyphal Growth Independent Of The Filamentation Mitogen-Activated Protein Kinase Pathway.” Genetics 153 (4): 1535–46.
Lu, Yang, Su, Unoje, And Liu. 2014. “Quorum Sensing Controls Hyphal Initiation In
Candida Albicans Through Ubr1-Mediated Protein Degradation.” Proceedings
Of The National Academy Of Sciences 111 (5): 1975–80. Https://Doi.Org/10.1073/Pnas.1318690111.
Lucas, Kendrick, And Givan. 1975. “Photocontrol Of Fungal Spore Germination.” Plant Physiology, 847–49.
Ma, Ibrahim, Skory, Grabherr, Burger, Butler, Elias, Et Al. 2009. “Genomic Analysis Of The Basal Lineage Fungus Rhizopus Oryzae Reveals A Whole-Genome Duplication.” Plos Genetics 5 (7). Https://Doi.Org/10.1371/Journal.Pgen.1000549.
Doi.Org/10.1128/AAC.00792-10. Li And Durbin. 2010. Fast And Accurate Long-Read Alignment With Burrows-Wheeler
Transform. Bioinformatics;26(5):589–95. Li, Handsaker, Wysoker, Fennell, Ruan, Homer, Et Al. The Sequence
Li, Charles, Cervantes, Springer, Boekhout, Ruiz-Vazquez, Torres-Martinez, Heitman, And Lee. 2011.“Sporangiospore Size Dimorphism Is Linked To Virulence Of Mucor Circinelloides” 7 (6). Https://Doi.Org/10.1371/Journal.Ppat.1002086.
Li, Zhan, Gao, Zhang, Lin, Gong, Guan, And Hu. 2018.
Linden, Rodriguez-Franco, And Macino. 1997. “Mutants Of Neurospora Crassa
Liu, Lin, Gebremariam, Luo, Skory, French, Chou, And Edwards. 2015.
196
Alignment/Map Format And Samtools. Bioinformatics. 2009;25(16):2078–9.
Defective In Regulation Of Blue Light Perception.” Molecular & General Genetics: MGG 254 (2): 111–18.

Machu, Eluère, Signon, Noëlle Simon, De La Roche Saint-André, And Bailly. 2014. “Spatially Distinct Functions Of Clb2 In The DNA Damage Response.” Cell
Cycle 13 (3): 383–98. Https://Doi.Org/10.4161/Cc.27354.
Macko, Staples, Gershon, And Renwick. 1970. “Self-Inhibitor Of Bean Rust Uredospores: Methyl 3,4-Dimethoxycinnamate.” Science (New York, N.Y.) 170 (3957): 539–40.
Mahran, Twisy, Elghazally And Badran. 2019. "Evaluation of different concentrations
Marcio, Rodrigues, Nakayasu, Oliveira, Nimrichter, Nosanchuck, Almeida, Casadevall.
Maresca, Bruno, And Kobayashi. 1989. “Dimorphism In Histoplasma Capsulatum : A Model For The Study Of Cell Differentiation In Pathogenic Fungi” 53 (2): 186–209.
Martinez, Fernando, Helming, Milde, Varin, Melgert, Draijer, Thomas, Et Al. 2013. “Genetic Programs Expressed In Resting And IL-4 Alternatively Activated Mouse And Human Macrophages: Similarities And Differences.” Blood 121 (9): E57-69. Https://Doi.Org/10.1182/Blood-2012-06-436212.
Marty, Broman, Zarnowski, Dwyer, Bond, Lounes-Hadj Sahraoui, Fontaine, Et Al. 2015. “Fungal Morphology, Iron Homeostasis, And Lipid Metabolism Regulated By A GATA Transcription Factor In Blastomyces Dermatitidis.” Plos Pathogens 11 (6): 1–40. Https://Doi.Org/10.1371/Journal.Ppat.1004959.
Marzluf. 1993. “Reggulation Of Sulfur And Nitrogen Metabolism In Filamentous Fungi.” Annual Review Of Microbiology, 31–55.
Mckenna, Hanna, Banks, Sivachenko, Cibulskis, Kernytsky, Et Al. The Genome Analysis Toolkit: A Mapreduce Framework For Analyzing Next-Generation DNA Sequencing Data. Genome Res [Internet]. 2010 Sep;20(9):1297–303. Available From: Https://Www.Ncbi.Nlm.Nih.Gov/Pubmed/20644199
Mellado, Aufauvre-Brown, Gow, And Holden. 1996. “The Aspergillus Fumigatus Chsc And Chsg Genes Encode Class III Chitin Synthases With Different
Functions.” Molecular Microbiology 20 (3): 667–79. Mellado, Aufauvre-Brown, Specht, Robbins, And Holden. 1995. “A
Multigene Family Related To Chitin Synthase Genes Of Yeast In The Opportunistic Pathogen Aspergillus Fumigatus.” Molecular & General Genetics : MGG 246 (3): 353–59.
Mendoza, Vilela, Voelz, Ibrahim, Voigt, Lee, Gigliotti, Et Al. 2014.“Human Fungal Pathogens Of Mucorales And Entomophthorales.” Cold Spring
Harbor Perspectives In Medicine 10 (1101). Http://Perspectivesinmedicine.Cshlp.Org/.
Momany And Talbot. 2017. “Septins Focus Cellular Growth For Host Infection By Pathogenic Fungi.” Frontiers In Cell And Developmental Biology
Mondo, Stephen, Lastovetsky, Gaspar, Schwardt, Barber, Riley, Sun, Grigoriev, And
2008."Extracellular Vesicles Produced by Cryptococcus neoformans Contain Protein Components Associated with Virulence" Euk Cell. doi: 10.1128/
EC.00370-07
of hydrogen peroxide solution (3% and 6%) as a potential new therapeutic option" JCocDerm. doi: 10.1111/jocd.13021
5 (April): 1–7. Https://Doi.Org/10.3389/Fcell.2017.00033.
Pawlowska. 2017.
197

“Bacterial Endosymbionts Influence Host Sexuality And Reveal Reproductive Genes Of Early Divergent Fungi.” Nature Communications 8 (1): 1843. Https://Doi.Org/10.1038/S41467-017-02052-8.
Salazar Solis, And Torres Guzman. 2010. “Catalase Overexpression Reduces The Germination Time And Increases The Pathogenicity Of The Fungus Metarhizium Anisopliae.” Applied Microbiology And Biotechnology 87 (3): 1033–44. Https://Doi.Org/10.1007/S00253-010-2517-3.
Moreno, Ibrahim-Granet, Vicentefranqueira, Amich, Ave, Leal, Latgé,And Antonio Calera. 2007. “The Regulation Of Zinc Homeostasis By The Zafa Transcriptional Activator Is Essential For Aspergillus Fumigatus Virulence.” Molecular Microbiology 64 (5): 1182–97. Https://Doi.Org/10.1111/J.1365-2958.2007.05726.X.
Moss, Kim, Nandakumar, Marten. Quantifying Metabolic Activity Of
Moyes, Wilson, Richardson, Mogavero, X. Tang, Wernecke, Höfs, Et Al. 2016. “Candidalysin Is A Fungal Peptide Toxin Critical For Mucosal Infection.” Nature 532 (7597): 64–68. Https://Doi.Org/10.1038/Nature17625.
Municio, Alvarez, Montero, Hugo, Rodríguez, Domingo, Alonso, Fernández, And Sánchez Crespo. 2013. “The Response Of Human Macrophages To Β-Glucans Depends On The Inflammatory Milieu.” Plos ONE 8 (4). Https://Doi.Org/10.1371/Journal.Pone.0062016.
Munoz, Delorey, Ford, Yu Li, Thompson, Rao, And Cuomo. 2018.“Coordinated Host-Pathogen Transcriptional Dynamics Revealed Using Sorted Subpopulations And Single, Candida Albicans Infected Macrophages.” Biorxiv, January, 350322. Https://Doi.Org/10.1101/350322.
Murgia, Manuela, Fiamma, Barac, Deligios, Mazzarello, Paglietti, Cappuccinelli, Et Al. 2019. “Biodiversity Of Fungi In Hot Desert Sands.” Microbiologyopen 8 (1): E00595. Https://Doi.Org/10.1002/Mbo3.595.
Myers, Tsang, And Swanson. 2010. “Activation Of Murine Macrophages.” Current Protocols Immunology 171 (10): 5447–53. Https://Doi.Org/10.1002/0471142735.Im1402s83.Activation.
Fungal Spores.” Plant And Soil 219 (1/2): 71–79. Http://Www.Jstor.Org/Stable/42950671.
Nemecek, Wuthrich, And Klein. 2006. “Global Control Of Dimorphism And Virulence In Fungi.”
Moody, Saidi, Gibbs, Choudhary, Holloway, Vesty, Bansal, Bradshaw And Coates.2016. "An ancient and conserved function for Armadillo-related proteins in the control of spore and seed germination by abscisic acid." New Phytol. doi: 10.1111/nph.13938
Moore-Landecker. 2011. “Fungal Spores.” ELS.
Moralez-Hernandez, Erika, Enrique Padilla Guerrero, Angelica, Gonzalez Hernandez,
Nagahashi, Douds, And Buee. 2000. “Light-Induced Hyphal Branching Of Germinated AM
198
Science (New York, N.Y.) 312 (5773): 583–88. Https://Doi.Org/10.1126/
Https://Doi.Org/10.1002/9780470015902.A0000378.Pub2.
Filamentous Fungi Using A Colorimetric XTT Assay. 2008;780–3.

Nesher, Minz, Kokkelink, Tudzynski, And Sharon. 2011. “Regulation Of Pathogenic Spore Germination By Cgrac1 In The Fungal Plant Pathogen Colletotrichum Gloeosporioides.” Eukaryotic Cell 10 (8): 1122–30. Https://Doi.Org/10.1128/EC.00321-10.
Newman, And Smulian. 2013. “Iron Uptake And Virulence In Histoplasma Capsulatum.” Current Opinion In Microbiology 16 (6): 700–707. Https://Doi.Org/10.1016/J.Mib.2013.09.001.
Nguyen, Long Nam, Bormann, Thi Thu Le, Starkel, Olsson, Nosanchuk, Giese, And chafer. 2011. “Autophagy-Related Lipase Fgatg15 Of Fusarium Graminearum Is Important For Lipid Turnover And Plant Infection.” Fungal Genetics And Biology :
FG & B 48 (3): 217–24. Https://Doi.Org/10.1016/J.Fgb.2010.11.004.
Nguyen, Vasseur, Coroller, Dantigny, Le Panse, Weill, Mounier, And Rigalma. 2017. “Temperature, Water Activity And PH During Conidia Production Affect The Physiological State And Germination Time Of Penicillium Species.” International Journal Of Food Microbiology 241: 151–60. Https://Doi.Org/10.1016/J.Ijfoodmicro.2016.10.022.
Nicholson, Munakata, G Horneck, H Melosh, And P Setlow. 2000. “Resistance Of Bacillus Endospores To Extreme Terrestrial And Extraterrestrial Environments.” Microbiology And Molecular Biology Reviews : MMBR 64 (3): 548–72. Https://Doi.Org/10.1128/MMBR.64.3.548-572.2000.
Nishiuchi, Takumi, D Masuda, H Nakashita, K Ichimura, K Shinozaki, S Yoshida, M Kimura, I Yamaguchi, And K Yamaguchi. 2006. “Fusarium Phytotoxin Trichothecenes Have An Elicitor-Like Activity In Arabidopsis Thaliana, But The Activity Differed Significantly Among Their Molecular Species.” Molecular
Plant-Microbe Interactions : MPMI 19 (5): 512–20. Https://Doi.Org/10.1094/MPMI-19-0512.
Nosanchuk, Van Duin, Mandal, Aisen, Legendre And Casadevall. 2004. “Blastomyces Dermatitidis Produces Melanin In Vitro And During Infection.” FEMS Microbiology Letters 239 (1): 187–93. Https://Doi.Org/10.1016/J.Femsle.2004.08.040.
Novodvorska, Hayer, Pullan, Wilson, Blythe, Stam, Stratford, And Archer. 2013. “Transcriptional Landscape Of Aspergillus Niger At Breaking Of Conidial Dormancy ”
Activity In Dormant Conidia Of Aspergillus Niger And Developmental Changes During Conidial Outgrowth.” Fungal Genetics And Biology 94: 23–31. Https://Doi.Org/10.1016/J.Fgb.2016.07.002.
Odds. 1988. Candida And Candidosis. 2nd Edn. London: Bailliere Tindall.
199
Naseem, Min, Spitzer, Gardin And Konopka. 2017."Regulation of Hyphal Growth and N-Acetylglucosamine Catabolism by Two Transcription Factors in Candida albicans" Genetics. https://doi.org/10.1534/genetics.117.201491
Nogueira, Pereira, Jenull, Kuchler and Lion. 2019. "Klebsiella pneumoniae prevents
spore germination and hyphal development of Aspergillus species" SciRep. https://doi.org/10.1038/s41598-018-36524-8
Nosanchuk, Stark And Casadevall. 2015. "Fungal Melanin: What do We Know About Structure?" FrontMicrob. doi: 10.3389/fmicb.2015.01463
Novodvorska, Stratford, Blythe, Wilson, Beniston, And Archer. 2016. “Metabolic

Oh, Taek, Chun-Seob Ahn, Kim, Ro, Lee, And Kim. 2010.“Proteomic Analysis Of Early Phase Of Conidia Germination In Aspergillus Nidulans.” Fungal Genetics And Biology 47 (3): 246–53. Https://Doi.Org/10.1016/J.Fgb.2009.11.002.
Osherov, Mathew, Romans, And May. 2002. “Identification Of Conidial-Enriched Transcripts In Aspergillus Nidulans Using Suppression Subtractive Hybridization.” Fungal Genetics And Biology 37: 197–204.
Requires RAS Signaling And Protein Synthesis.” Genetics 155 (2): 647–56. Https://Doi.Org/10.1007/Bf00444092.
Panepinto, Komperda, Frases, Park, Djordjevic, Casadevall, And Williamson. 2009. “Sec6-Dependent Sorting Of Fungal Extracellular Exosomes And Laccase Of Cryptococcus Neoformans.” Molecular Microbiology 71 (5): 1165–76. Https://Doi.Org/10.1111/J.1365-2958.2008.06588.X.
Parente-Rocha, Alves, Flávia, Baeza, Bail, Pelleschi Taborda, Luiz Borges, And Maria De. 2015.“Macrophage Interaction With Paracoccidioides Brasiliensis Yeast Cells Modulates Fungal Metabolism And Generates A Response To Oxidative Stress,” 1–18. Https://Doi.Org/10.1371/Journal.Pone.0137619.
Paris, Debeaupuis, Crameri, Charlès, Prévost, Philippe, Latgé, Et Al. 2003. “Conidial Hydrophobins Of Aspergillus Fumigatus.” Applied And Environmental
Microbiology 69 (3): 1581–88. Https://Doi.Org/10.1128/AEM.69.3.1581. Park, Xue, L Zheng, S Lam, And J Xu. 2002. “MST12 Regulates Infectious Growth
But Not Appressorium Formation In The Rice Blast Fungus Magnaporthe Grisea.” Mol Plant Microbe Interact 15 (3): 183–92. Https://Doi.Org/10.1094/MPMI.2002.15.3.183.
Park And Bi. 2007. “Central Roles Of Small Gtpases In The Development Of Cell Polarity In Yeast And Beyond.” Microbiology And Molecular Biology Reviews
Partida-Martinez And Hertweck. 2005. “Pathogenic Fungus Harbours Endosymbiotic Bacteria For Toxin Production” 437 (October).
Pearson 2013. “An Introduction To Sequence Similarity (‘Homology’)
Pereyra, Mizyrycki, And Moreno. 2000. “Threshold Level Of Protein Kinase A Activity And Polarized Growth In Mucor Rouxii.” Microbiology (Reading, England)
( Pt 8 (August): 1949–58. Https://Doi.Org/10.1099/00221287-146-8-1949. Pérez-
And Garre. 2019. “Mucor Circinelloides Thrives Inside The Phagosome
Osherov And May. 2000. “Conidial Germination In Aspergillus Nidulans
200
71 (1): 48–96. Https://Doi.Org/10.1128/MMBR.00028-06.
Https://Doi.Org/10.1038/Nature03997.
Searching.” Current Protocols In Bioinformatics Chapter 3 (June): Unit3.1-Unit3.1. Https://Doi.Org/10.1002/0471250953.Bi0301s42.
Perez-Arques, Navarro-Mendoza, Murcia, Lax, Martínez-García, Heitman, Nicolás,

Through An Atf-Mediated Germination Pathway.” Edited By Anuradha Idnurm Chowdhary Alexander Kronstad, James. Mbio 10 (1): E02765-18. Https://Doi.Org/10.1128/Mbio.02765-18.
Pérez-Sánchez, González, Colón-Lorenzo, González-Velázquez, González-Méndez, And Rodríguez-Del Valle. 2010. “Interaction Of The Heterotrimeric G Protein Alpha Subunit SSG-1 Of Sporothrix Schenckii With Proteins Related To Stress Response And Fungal Pathogenicity Using A Yeast Two-Hybrid Assay.” BMC
Microbiology. Https://Doi.Org/10.1186/1471-2180-10-317.
Petraitis, Vidmantas, Petraitiene, Antachopoulos, Hughes, Cotton, Kasai, Harrington,
“Increased Virulence Of Cunninghamella Bertholletiae In Experimental Pulmonary Mucormycosis : Correlation With Circulating Molecular Biomarkers , Sporangiospore Germination And Hyphal Metabolism,” No. January: 72–82. Https://Doi.Org/10.3109/13693786.2012.690107.
Philpott, Caroline C. 2006. “Iron Uptake In Fungi: A System For Every Source.” Biochimica Et Biophysica Acta - Molecular Cell Research 1763 (7): 636–45. Https://Doi.Org/10.1016/J.Bbamcr.2006.05.008.
Pihet, Vandeputte, Tronchin, Renier, Saulnier, Georgeault, Mallet, Chabasse, Symoens And Bouchara. 2009. “Melanin Is An Essential Component For The Integrity Of The Cell Wall Of Aspergillus Fumigatus Conidia.” BMC Microbiology 9 (1): 177. Https://Doi.Org/10.1186/1471-2180-9-177.
Piispanen, Bonnefoi, Carden, Deveau, Bassilana, And Hogan. 2011.“Roles Of Ras1 Membrane Localization During Candida Albicans Hyphal Growth And Farnesol Response.” Eukaryotic Cell 10 (11): 1473 LP – 1484. Https://Doi.Org/10.1128/EC.05153-11.
Plante, Ioannoni, Beaudoin, And Labbã. 2014.
“Characterization Of Schizosaccharomyces Pombe Copper Transporter Proteins In Meiotic And Sporulating Cells.” Journal Of Biological Chemistry 289 (14): 10168–81. Https://Doi.Org/10.1074/Jbc.M113.543678.
Plante, Normant, Ramos-Torres, And Labbé. 2017. “Cell-Surface Copper Transporters And Superoxide Dismutase 1 Are Essential For Outgrowth During Fungal Spore Germination.” Journal Of Biological Chemistry 292 (28): 11896–914. Https://Doi.Org/10.1074/Jbc.M117.794677.
Possart, Fleck, And Hiltbrunner. 2014. “Shedding (Far-Red) Light On Phytochrome Mechanisms And Responses In Land Plants.” Plant Science
217–218 (316): 36–46. Https://Doi.Org/10.1016/J.Plantsci.2013.11.013.
Et Al. 2013.
201

Prasad, Kurup, And Maheshwari. 1979. “Effect Of Temperature On Respiration Of AMesophilic And A Thermophilic Fungus.” Plant Physiology 64 (2): 347–48. Https://Doi.Org/10.1104/Pp.64.2.347.
Qing And Shiping. 2000. “Postharvest Biological Control Of Rhizopus Rot Of Nectarine Fruits By Pichia Membranefaciens.” Plant Disease 84 (11): 1212–16. Https://Doi.Org/10.1094/PDIS.2000.84.11.1212.
Ramage, Saville, Wickes, López-Ribot, And Lo. 2002“Inhibition Of Candida Albicans Biofilm Formation By Farnesol , A Quorum-Sensing Molecule Inhibition Of Candida Albicans Biofilm Formation By Farnesol , A Quorum-Sensing Molecule Downloaded From Http://Aem.Asm.Org/ On September 16 , 2013 By Danish Veterina.” Applied And Environmental
Microbiology 68 (11): 5459–63. Https://Doi.Org/10.1128/AEM.68.11.5459. Regente, Mariana, Marcela Pinedo, Hélène San Clemente, Thierry Balliau, Elisabeth
Jamet, And Laura De La Canal. 2017. “Plant Extracellular Vesicles Are Incorporated By A Fungal Pathogen And Inhibit Its Growth.” Journal Of
Experimental Botany 68 (20): 5485–95. Https://Doi.Org/10.1093/Jxb/Erx355. Reponen, Grinshpun, Conwell, Wiest, And Anderson. 2001.
“Aerodynamic Versus Physical Size Of Spores: Measurement And Implication For Respiratory Deposition.” Grana 40 (3): 119–25. Https://Doi.Org/10.1080/00173130152625851.
Reu, Johan De, Griffiths, Rombouts, And Nout. 1995. “Effect Of Oxygen And Carbon Dioxide On Germination And Growth Ofrhizopus Oligosporus On Model Media And Soya Beans.” Applied Microbiology And Biotechnology 43 (October): 908–13. Https://Doi.Org/10.1007/BF02431927.
Reyes And Bartnicki-Garcia. 1964. “Chemistry Of Spore Wall In Mucor Rouxii.” Archives of Biochemistry and Physics 133: 125–33.
Rivera-Rodriguez And Rodriguez-Del Valle. 1992. “Effects Of Calcium Ions On The Germination Of Sporothrix Schenckii Conidia.” Journal Of Medical And Veterinary
Mycology : Bi-Monthly Publication Of The International Society For Human And
Animal Mycology 30 (3): 185–95. Robinson, Mccarthy, And Smyth. 2009. “Edger: A Bioconductor Package
For Differential Expression Analysis Of Digital Gene Expression Data.” Bioinformatics 26 (1): 139–40. Https://Doi.Org/10.1093/Bioinformatics/Btp616.
Rodrigues, Franzen, Nimrichter, And Miranda. 2013.“Vesicular Mechanisms Of Traffic Of Fungal Molecules To The Extracellular Space.” Current Opinion In Microbiology 16 (4): 414–20. Https://Doi.Org/Https://Doi.Org/10.1016/J.Mib.2013.04.002.
Rogers, Williams, Jie Feng, Lewis, And Qing Wei. 2013. “Role Of Bacterial Lipopolysaccharide In Enhancing Host Immune Response To Candida Albicans.” Clinical And Developmental Immunology 2013. Https://Doi.Org/10.1155/2013/320168.
202

Röhrig, Kastner, And Fischer. 2013. “Light Inhibits Spore Germination Through Phytochrome In Aspergillus Nidulans.” Current Genetics, 1–8. Https://Doi.Org/10.1007/S00294-013-0387-9.
Rooij, Pascale Van, Martel, D’Herde, Brutyn, Croubels, Ducatelle, Haesebrouck, And Pasmans.2012. “Germ Tube Mediated Invasion Of Batrachochytrium Dendrobatidis In Amphibian Skin Is Host Dependent.” Plos ONE 7 (7): 1–8. Https://Doi.Org/10.1371/Journal.Pone.0041481.
Ruiz-Herrera And San-Blas. 2003. “Chitin Synthesis As Target For Antifungal Drugs.” Current Drug Targets. Infectious Disorders 3 (1): 77–91.
Ruiz-Roldan, Kohli, Roncero, Philippsen, Di Pietro, And Espeso. 2010. “Nuclear Dynamics During Germination, Conidiation, And Hyphal Fusion Of Fusarium Oxysporum.” Eukaryotic Cell 9 (8): 1216–24. Https://Doi.Org/10.1128/EC.00040-10.
Rybak And Robatzek. 2019. “Functions Of Extracellular Vesicles In Immunity And Virulence.” Plant Physiology 179 (4): 1236–47.
Santorelli, Thompson, Villegas, Svetz, Dinh, Parikh, Sucgang, Et Al. 2008. “Facultative Cheater Mutants Reveal The Genetic Complexity Of Cooperation In Social Amoebae.” Nature 451 (7182): 1107–10. Https://Doi.Org/10.1038/Nature06558.
Sarmiento, Ciarmela, Sánchez Thevenet, Minvielle, And Basualdo. 2006. “Comparison Of Preparation Techniques Of Mixed Samples (Fungi-Helminth Eggs) For Scanning Electron Microscopy By Critical Point Drying.” Parasitology Research 99 (4): 455–58. Https://Doi.Org/10.1007/S00436-006-0187-Y.
Schimek And Wostemeyer. 2012. “Biosynthesis, Extraction, Purification, And Analysis Of Trisporoid Sexual Communication Compounds From Mated Cultures Of Blakeslea Trispora.” Methods In Molecular Biology (Clifton, N.J.) 898: 61–74. Https://Doi.Org/10.1007/978-1-61779-918-1_3.
Schmidt-Heydt, Rüfer, Raupp, Bruchmann, Perrone, And Geisen. 2011. “Influence Of Light On Food Relevant Fungi With Emphasis On Ochratoxin Producing Species.” International Journal Of Food Microbiology 145 (1): 229–37. Https://Doi.Org/10.1016/J.Ijfoodmicro.2010.12.022.
Schmidt, Tramsen, Perkhofer, Lass-Flörl, Hanisch, Röger, Klingebiel, Koehl, And Lehrnbecher. 2013. “Immunobiology Rhizopus Oryzae Hyphae Are Damaged By Human Natural Killer ( NK ) Cells , But Suppress NK Cell Mediated Immunity.” Immunobiology 218 (7): 939–44. Https://Doi.Org/10.1016/J.Imbio.2012.10.013.
Schmit And Brody. 1976. “Biochemical Genetics Of Neurospora
Schnitzler, Peltroche-Llacsahuanga, Bestier, Zündorf, Lütticken, And Haase. 1999.
Https://Doi.Org/10.1104/Pp.18.01557.
203
“Effect Of Melanin And Carotenoids Of Exophiala (Wangiella) Dermatitidis On Phagocytosis, Oxidative Burst, And Killing By Human Neutrophils.” Infection And
Immunity 67 (1): 94–101.
Rosowski, Raffa, Knox, Golenberg, Keller And Huttenlocher. 2018. " Macrophages inhibit Aspergillus fumigatus germination and neutrophil-mediated fungal killing" PlosPathog. doi: 10.1371/journal.ppat.1007229
Crassa Conidial Germination.” Bacteriology Reviews 40 (1): 1–41.

Schroeder, Mueller, Stocker, Salowsky, Leiber, Gassmann, Lightfoot, Menzel, Granzow, And Ragg. 2006. “The RIN: An RNA Integrity Number For Assigning Integrity Values To RNA Measurements.” BMC Molecular Biology 7 (1): 3. Https://Doi.Org/10.1186/1471-2199-7-3.
Schwartze, Volker, Winter, Shelest, Marcet-Houben, Horn, Wehner, Linde, Et Al. 2014. “Gene Expansion Shapes Genome Architecture In The Human Pathogen Lichtheimia Corymbifera: An Evolutionary Genomics Analysis In The Ancient Terrestrial Mucorales (Mucoromycotina).” Plos Genetics 10 (8). Https://Doi.Org/10.1371/Journal.Pgen.1004496.
Seman, Moore, Scherer, Blair, Manandhar, Jones, And Wheeler. 2018. “Yeast And Filaments Have Specialized, Independent Activities In A Zebrafish Model Of Candida Albicans Infection.” Edited By George S Deepe. Infection And
Immunity 86 (10): E00415-18. Https://Doi.Org/10.1128/IAI.00415-18.
Semighini, Murray, And Harris. 2008. “Inhibition Of Fusarium Graminearum Growth And Development By Farnesol.” FEMS Microbiology Letters 279 (2): 259–64. Https://Doi.Org/10.1111/J.1574-6968.2007.01042.X.
Semighini, Hornby, Dumitru, Nickerson, And Harris. 2006. “Farnesol-Induced Apoptosis In Aspergillus Nidulans Reveals A Possible Mechanism For Antagonistic Interactions Between Fungi.” Molecular Microbiology 59 (3): 753–64. Https://Doi.Org/10.1111/J.1365-2958.2005.04976.X.
Sephton-Clark, Munoz, Ballou, Cuomo, And Voelz. 2018. “Pathways Of Pathogenicity : Transcriptional Stages Of Germination In The Fatal Fungal Pathogen Rhizopus Delemar.” Msphere 3 (5): 1–16.
Sephton-Clark And Voelz: Advances In Applied Microbiology 2017. “Spore Germination Of Pathogenic Filamentous Fungi.” In . Academic
Sephton-Clark, Munoz, Itabangi, Voelz, Cuomo, And Ballou. 2019. “Host-
Seto And Tazaki. 1975. “Growth And Respiratory Activity Of Mold Fungus
(Trichoderma Lignorum).” The Botanical Magazine Tokyo 88 (4): 255–66. Https://Doi.Org/10.1007/BF02488368.
Press. Https://Doi.Org/Https://Doi.Org/10.1016/Bs.Aambs.2017.10.002.
Pathogen Transcriptomics Of Macrophages, Mucorales And Their Endosymbionts: A Polymicrobial Pas De Trois.” Biorxiv.
204
Schopfer, Bajracharya And Plachy. 1979. "Control of Seed Germination by Abscisic Acid." Plant Physiol.

Sherrington, Sorsby, Mahtey, Kumwenda, Lenardon, Brown, Ballou, Maccallum, And Hall. 2017. “Adaptation Of Candida Albicans To Environmental PH Induces Cell Wall Remodelling And Enhances Innate Immune Recognition.” Plos
Pathogens 13 (5): 1–28. Https://Doi.Org/10.1371/Journal.Ppat.1006403.
Shimura, Matsuura, Takada, And Koda. 2007. “An Antifungal Compound Involved In Symbiotic Germination Of Cypripedium Macranthos Var. Rebunense (Orchidaceae).” Phytochemistry 68 (10): 1442–47. Https://Doi.Org/10.1016/J.Phytochem.2007.03.006.
Shopova, Iordana, Belyaev, Dasari, Jahreis, Stroe, Cseresnyés, Medyukhina, Et Al. 2019.“Human Neutrophils Produce Antifungal Extracellular Vesicles Against Aspergillus Fumigatus.” Biorxiv, January, 620294. Https://Doi.Org/10.1101/620294.
Silva, Roberta Peres Da, Puccia, Rodrigues, L Oliveira, Joffe, César, Nimrichter, Goldenberg, And Alves. 2015. “Extracellular Vesicle-Mediated Export Of Fungal RNA.” Scientific Reports 5: 7763.
Simão, Waterhouse, Ioannidis, Kriventseva, And Zdobnov. 2015. “BUSCO: Assessing Genome Assembly And Annotation Completeness With Single-Copy Orthologs.” Bioinformatics 31 (19): 3210–12. Https://Doi.Org/10.1093/Bioinformatics/Btv351.
Skiada, Rigopoulos, Larios, Petrikkos, And Katsambas. 2012. “Global Epidemiology Of Cutaneous Zygomycosis.” Clinics In Dermatology 30 (6): 628–32. Https://Doi.Org/10.1016/J.Clindermatol.2012.01.010.
Smith, Dixon, And May. 2014. “The Fungal Pathogen Cryptococcus Neoformans Manipulates Macrophage Phagosome Maturation.” Cellular
Microbiology, November. Https://Doi.Org/10.1111/Cmi.12394 [Doi]. Solaiman And Saito. 1997. “Use Of Sugars By Intraradical Hyphae Of Arbuscular
Mycorrhizal Fungi Revealed By Radiorespirometry.” The New Phytologist 136 (3): 533–38. Http://Www.Jstor.Org/Stable/2558957.
Son, Hokyoung, Myung-Gu Kim, K Min, Young-Su Seo, JY Lim, GJ Choi, C Kim, Suhn-Kee Chae And Yin-Won Lee. 2013. “Abaa Regulates Conidiogenesis In The Ascomycete Fungus Fusarium Graminearum.” Plos ONE 8 (9). Https://Doi.Org/10.1371/Journal.Pone.0072915.
Son, Hokyoung, Lee And Yin-Won Lee. 2013. “A Novel Gene , GEA1 , Is Required For Ascus Cell-Wall Development In The Ascomycete Fungus Fusarium Graminearum.” Microbiology 159 (2013): 1077–85. Https://Doi.Org/10.1099/Mic.0.064287-0.
Spellberg, Edwards, And Ibrahim. 2016. “Novel Perspectives On Mucormycosis : Pathophysiology , Presentation , And Management” 18 (3): 556–69. Https://Doi.Org/10.1128/CMR.18.3.556.
Spellberg, Yue Fu, Edwards, And Ibrahim. 2016. “Combination Therapy With
205

Amphotericin B Lipid Complex And Caspofungin Acetate Of Disseminated Zygomycosis In Diabetic Ketoacidotic Mice” 49 (2): 830–32. Https://Doi.Org/10.1128/AAC.49.2.830.
Spellberg And Ibrahim. 2010. “Recent Advances In The Treatment Of Mucormycosis,” No. August: 423–29. Https://Doi.Org/10.1007/S11908-010-0129-9.
Spence, Lakshmanan, Donofrio, And Bais. 2015. “Crucial Roles Of Abscisic Acid Biogenesis In Virulence Of Rice Blast Fungus Magnaporthe Oryzae.” Frontiers In Plant Science 6 (1082): 1–13. Https://Doi.Org/10.3389/Fpls.2015.01082.
Sueiro-Olivares, Fernandez-Molina, Abad-Diaz-De-Cerio, Gorospe, Pascual, Guruceaga, Ramirez-Garcia, Et Al. 2015. “Aspergillus Fumigatus Transcriptome Response To A Higher Temperature During The Earliest Steps Of Germination Monitored Using A New Customized Expression Microarray.” Microbiology (Reading,
England) 161 (2015): 490–502. Https://Doi.Org/10.1099/Mic.0.000021. Sumi. 1928. “Über Die Chemischen Bestandteile Der Sporen Von Aspergillus
Oryzae.” Biochem. Z, No. 195: 161–74. Sun, Li, Liu, Yang, Liang, You-Liang Peng, And Huang. 2017.
“Large-Scale Identification Of Lysine Acetylated Proteins In Vegetative Hyphae Of The Rice Blast Fungus.” Scientific Reports 7 (1): 15316. Https://Doi.Org/10.1038/S41598-017-15655-4.
Suzuki, Sarikaya Bayram, Ozgur Bayram, And Braus. 2013. “Conf And Conj Contribute To Conidia Germination And Stress Response In The Filamentous Fungus Aspergillus Nidulans.” Fungal Genetics And Biology : FG & B 56 (July): 42–53. Https://Doi.Org/10.1016/J.Fgb.2013.04.008.
Svanström, Van Leeuwen, Dijksterhuis, And Melin. 2014. “Trehalose Synthesis In Aspergillus Niger: Characterization Of Six Homologous Genes, All With Conserved Orthologs In Related Species.” BMC Microbiology 14 (1): 90. Https://Doi.Org/10.1186/1471-2180-14-90.
Tamayo, Gamez-Gallego, Azcan-Aguilar, And Ferrol. 2014. “Genome-Wide Analysis Of Copper, Iron And Zinc Transporters In The Arbuscular Mycorrhizal Fungus Rhizophagus Irregularis.” Frontiers In Plant
Science 5 (October): 1–13. Https://Doi.Org/10.3389/Fpls.2014.00547. Tapia, Rivera Garay, Sabanero, Trejo Basurto, And Sabanero Lopez.
1993. “Sporothrix Schenckii: Cultures In Different Soils.” Revista
Latinoamericana De Microbiologia 35 (2): 191–94.Technologies, Stemcell “Accutase Directions For Use.”
Https://Cdn.Stemcell.Com/Media/Files/Pis/29637-PIS_1_0_2.Pdf?_Ga=2.10286005.1436903603.1548350382-38118485.1548350382.
Teertstra, Tegelaar, Dijksterhuis, Golovina, Ohm, And W Sten. 2017. “Maturation Of Conidia On Conidiophores Of Aspergillus Niger.”
206
Takino, Kozaki, Ozaki, Liu, Minami And Oikawa. 2019."Elucidation of biosynthetic pathway of a plant hormone abscisic acid in phytopathogenic fungi." Biosci Biotechnol Biochem. doi: 10.1080/09168451.2019.1618700

Fungal Genetics And Biology 98: 61–70. Https://Doi.Org/10.1016/J.Fgb.2016.12.005.
Thammahong, Puttikamonkul, Perfect, Brennan, And Cramer. 2017. “Central Role Of The Trehalose Biosynthesis Pathway In The Pathogenesis Of Human Fungal Infections: Opportunities And Challenges For Therapeutic Development.” Microbiology And Molecular Biology Reviews 81 (2): E00053-16. Https://Doi.Org/10.1128/MMBR.00053-16.
Thanh, Rombouts, And Nout. 2005. “Effect Of Individual Amino Acids And Glucose On Activation And Germination Of Rhizopus Oligosporus Sporangiospores In Tempe Starter.” Journal Of Applied Microbiology 99 (5): 1204–14. Https://Doi.Org/10.1111/J.1365-2672.2005.02692.X.
Thevelein, Den Hollander, And Shulman. 1982. “Changes In The Activity And Properties Of Trehalase During Early Germination Of Yeast Ascospores: Correlation With Trehalose Breakdown As Studied By In Vivo 13C NMR.” Proceedings Of The National Academy Of Sciences Of The United States Of
America 79 (11): 3503–7. Https://Doi.Org/10.1073/Pnas.79.11.3503. Tomita, Murayama, And Nakamura. 1984. “Effects Of Auxin And
Gibberellin On Elongation Of Young Hyphae In Neurospora Crassa.” Plant And
Physiology 25 (2): 355–58. Https://Doi.Org/10.1093/Oxfordjournals.Pcp.A076722. Trzaska, Wrigley, Thwaite, And May. 2017.
“Species-Specific Antifungal Activity Of Blue Light.” Scientific Reports 7 (1): 4605. Https://Doi.Org/10.1038/S41598-017-05000-0.
Trzaska, Correia, Villegas, May, And Voelz. 2015. “PH Manipulation As A Novel Strategy For Treating Mucormycosis” 59 (11):
6968–74. Https://Doi.Org/10.1128/AAC.01366-15.Address. Tsai, Wheeler, Chang, And Kwon-Chung. 1999. “A Developmentally
Regulated Gene Cluster Involved In Conidial Pigment Biosynthesis In Aspergillus Fumigatus.” Journal Of Bacteriology 181 (20): 6469–77.
Tsirilakis, Kim, Vicencio, Andrade, Casadevall, And Goldman. 2012. “Methylxanthine Inhibit Fungal Chitinases And Exhibit Antifungal Activity.” Mycopathologia. Https://Doi.Org/10.1007/S11046-011-9483-X.
Turgeman, Kakongi, Schneider, Vinokur, Teper-Bamnolker, Carmeli, Levy, Skory,Lichter, And Eshel. 2013. “Induction Of Rhizopus Oryzae Germination Under Starvation Using Host Metabolites Increases Spore Susceptibility To Heat Stress.” Biochemistry And Cell Biology, No. 4.
Turgeman, Shatil-Cohen, Moshelion, And Teper-Bamnolker. 2016.“The Role Of Aquaporins In PH-Dependent Germination Of Rhizopus Delemar Spores,” 1–18. Https://Doi.Org/10.1371/Journal.Pone.0150543.
Sinigaglia-Coimbra, Almeida, And Puccia. 2011. “The Pathogenic Fungus Paracoccidioides Brasiliensis Exports Extracellular Vesicles Containing
Cell Physiology 25 (2): 355–58. Https://Doi.Org/10.1093/Oxfordjournals.Pcp.A076722.
6968–74. Https://Doi.Org/10.1128/AAC.01366-15.
Vallejo, Matsuo, Ganiko, Soares Medeiros, Miranda, Silva, Freymüller-Haapalainen,
207

Highly Immunogenic Α-Galactosyl Epitopes.” Eukaryotic Cell 10 (3): 343–51.
Vargas, Rocha, Leite Oliveira, Costa Albuquerque, Frases, Santos, Nosanchuk, Oliveira Gomes, Medeiros, And Miranda. 2015. “Compositional And Immunobiological Analyses Of Extracellular Vesicles Released By C Andida Albicans.” Cellular
Microbiology 17 (3): 389–407. Velagapudi, Ping Hsueh, Geunes-Boyer, Wright, And Heitman. 2009.
“Spores As Infectious Propagules Of Cryptococcus Neoformans.” Infection And Immunity 77 (10): 4345–55. Https://Doi.Org/10.1128/IAI.00542-09.
Verma, Kumar, Duti Prabh, Sankararamakrishnan, Gomes, Agasse, Thiebaud, Delrot, Et Al. 2014.“New Subfamilies Of Major Intrinsic Proteins In Fungi Suggest Novel Transport Properties In Fungal Channels: Implications For The Host-Fungal Interactions.” BMC Evolutionary Biology 14 (1): 173. Https://Doi.Org/10.1186/S12862-014-0173-4.
Vylkova. 2017. “Environmental PH Modulation By Pathogenic Fungi As A Strategy To Conquer The Host.” Plos Pathogens 13 (2): 1–6.
Walsh, Botts, Mcdermott, Ortiz, Wüthrich, Klein, And Hull. 2019. “Infectious Particle Identity Determines Dissemination And Disease Outcome For The Inhaled Human Fungal Pathogen Cryptococcus.” Plos Pathogens, 1–30.
Walther, Pawłowska, Alastruey-Izquierdo, Wrzosek, Rodriguez-Tudela, Dolatabadi, Chakrabarti, And De Hoog. 2013. “DNA Barcoding In Mucorales: An Inventory Of Biodiversity.” Persoonia 30 (June): 11–47. Https://Doi.Org/10.3767/003158513X665070.
Wang, Yoshida, And Hasunuma. 2007. “Catalase-1 (CAT-1) And Nucleoside Diphosphate Kinase-1 (NDK-1) Play An Important Role In Protecting Conidial Viability Under Light Stress In Neurospora Crassa.” Molecular Genetics And
Genomics : MGG 278 (3): 235–42. Https://Doi.Org/10.1007/S00438-007-0244-Y.
Wang, Long Liu, Storey, Tibshirani, Herschlag, And O'Brown. 2002. “Precision And Functional Specificity In MRNA Decay.” Proceedings Of The
National Academy Of Sciences Of The United States Of America 99 (9): 5860–65. Https://Doi.Org/10.1073/Pnas.092538799.
Warris, Netea, Verweij, Gaustad, And Kullberg. 2005.“Cytokine Responses And Regulation Of Interferon-Gamma Release By Human Mononuclear Cells To Aspergillus Fumigatus And Other Filamentous Fungi,” No. November: 613–21. Https://Doi.Org/10.1080/13693780500088333.
Watanabe, Ogasawara, Mikami, And Matsumoto. 2006“Hyphal Formation Of Candida Albicans Is Controlled By Electron Transfer System.” Biochemical And Biophysical Research Communications 348 (1): 206–11.
208
Https://Doi.Org/10.1371/Journal.Ppat.1006149. Walker, Sood, Lenardon, Milne, Olson, Jensen, Wolf, Casadevall, Adler-Moore And Gow.
2018. "The Viscoelastic Properties of the Fungal Cell Wall Allow Traffic of AmBisome as Intact Liposome Vesicles" mBio. DOI: 10.1128/mBio.02383-17

Https://Doi.Org/10.1016/J.Bbrc.2006.07.066. Wells And Uota. 1970. “Germination And Growth Of Five Fungi In Low-Oxygen
And High-Carbon Dioxide Atmospheres.” Phytopathology 60 (1): 50–53.
Wolf, Espadas-Moreno, Luque-Garcia, And Casadevall. 2014.“Interaction Of Cryptococcus Neoformans Extracellular Vesicles With The Cell Wall.” Eukaryotic Cell 13 (12): 1484–93. Https://Doi.Org/10.1128/Ec.00111-14.
Wood, Salzberg. 2014 Kraken: Ultrafast Metagenomic Sequence Classification Using
Wu, Yang, Smith, Peterson, Dekhang, Zhang, Zucker, Et Al. 2014. “Genome-Wide Characterization Of Light-Regulated Genes In Neurospora Crassa.” G3; Genes|Genomes|Genetics 4 (9): 1731–45. Https://Doi.Org/10.1534/G3.114.012617.
Wurster, Thielen, Weis, Walther, Elias, Waaga-Gasser, Dragan, Et Al. 2017. “Mucorales Spores Induce A Proinflammatory Cytokine Response In Human Mononuclear Phagocytes And Harbor No Rodlet Hydrophobins.” Virulence 8 (8): 1708–18. Https://Doi.Org/10.1080/21505594.2017.1342920.
Yamazaki, Tanaka, Kaneko, Ohta, And Horiuchi. 2008. “Aspergillus Nidulans Chia Is A Glycosylphosphatidylinositol (GPI)-Anchored Chitinase Specifically Localized At Polarized Growth Sites.” Fungal Genetics And
Biology 45 (6): 963–72. Https://Doi.Org/Https://Doi.Org/10.1016/J.Fgb.2008.02.008.
Yáñez-Mó, Minder, Andreu, Bedina Zavec, Borràs, Buzas, Et Al. 2015. “Biological Properties Of Extracellular Vesicles And Their Physiological Functions.” Journal Of Extracellular Vesicles 4 (May): 27066. Https://Doi.Org/10.3402/Jev.V4.27066.
Yang, Ho Lee, Kim, Seok Ki, Jae Huh, And Yong Lee. 2016. “Identification Of Mucorales From Clinical Specimens: A 4-Year Experience In A Single Institution.” Annals Of Laboratory Medicine 36 (1): 60–63. Https://Doi.Org/10.3343/Alm.2016.36.1.60.
Yao, Guo, Wang, Zhang, Xu, And Tang. 2016. “A Cytoplasmic Cu-Zn Superoxide Dismutase SOD1 Contributes To Hyphal Growth And Virulence Of Fusarium Graminearum.” Fungal Genetics And Biology 91: 32–42. Https://Doi.Org/10.1016/J.Fgb.2016.03.006.
Yates, Hermetter, And Russell. 2005. “The Kinetics Of Phagosome Maturation As A Function Of Phagosome/Lysosome Fusion And Acquisition Of Hydrolytic Activity.” Traffic (Copenhagen, Denmark) 6 (5): 413–20. Https://Doi.Org/10.1111/J.1600-0854.2005.00284.X.
Yoneda And Doering. 2006. “A Eukaryotic Capsular Polysaccharide Is Synthesized Intracellularly And Secreted Via Exocytosis.” Molecular Biology Of
The Cell 17 (12): 5131–40.
Https://Doi.Org/10.1094/Phyto-60-50.
Exact Alignments. Genome Biol.
209

Yoshida And Hasunuma. 2004. “Reactive Oxygen Species Affect Photomorphogenesis In Neurospora Crassa.” Journal Of Biological Chemistry 279
Zahavi, Cohen, Weiss, Schena, Daus, Kaplunov, Zutkhi, Ben-Arie, And Droby. 2000. “Biological Control Of Botrytis, Aspergillus And Rhizopus Rots On Table And Wine Grapes In Israel.” Postharvest Biology And Technology 20 (2): 115–24. Https://Doi.Org/10.1016/S0925-5214(00)00118-6.
Zahiri, Babu, And Saville. 2005. “Differential Gene Expression During Teliospore
Zhang And Watson. 1997. “Effect Of Dew Period And Temperature On The Ability Of Exserohilum Monoceras To Cause Seedling Mortality Of Echinochloa Species.” Plant Disease 81 (6): 629–34. Https://Doi.Org/10.1094/PDIS.1997.81.6.629.
Zhao, Panepinto, Fortwendel, Fox, Oliver, Askew, And Rhodes. 2006. “Deletion Of The Regulatory Subunit Of Protein Kinase A In Aspergillus Fumigatus Alters Morphology, Sensitivity To Oxidative Damage, And Virulence.” Infection And Immunity 74 (8): 4865 LP – 4874. Https://Doi.Org/10.1128/IAI.00565-06.
Zhao, Liang, And Zhou. 2018. “Small RNA Trafficking At The Forefront Of Plant- Pathogen Interactions.” F1000Research 7 (October): F1000 Faculty Rev-1633. Https://Doi.Org/10.12688/F1000research.15761.1.
Regulates Hyphal Growth, Stress Responses, And Plant Infection In Fusarium Graminearum.” Plos ONE 7 (11): 1–12. Https://Doi.Org/10.1371/
Journal.Pone.0049495.Zuber, Hynes, And Andrianopoulos. 2003. “The G-Protein Alpha-Subunit Gasc
Plays A Major Role In Germination In The Dimorphic Fungus Penicillium Marneffei.” Genetics 164 (2): 487–99.
(8): 6986–93. Https://Doi.Org/10.1074/Jbc.M310060200.
Zhang, Caiguo. 2014. “Essential Functions Of Iron-Requiring Proteins In DNA
Zheng, Zhang, Zhou, Wang, Xiang, Zheng, And Xu. 2012. “The Fghog1 Pathway
Germination In Ustilago Maydis.” Molecular Genetics And Genomics : MGG 273 (5): 394–403. Https://Doi.Org/10.1007/S00438-005-1142-9.
Replication, Repair And Cell Cycle Control.” Protein & Cell 5 (10): 750–60. Https://Doi.Org/10.1007/S13238-014-0083-7.
210
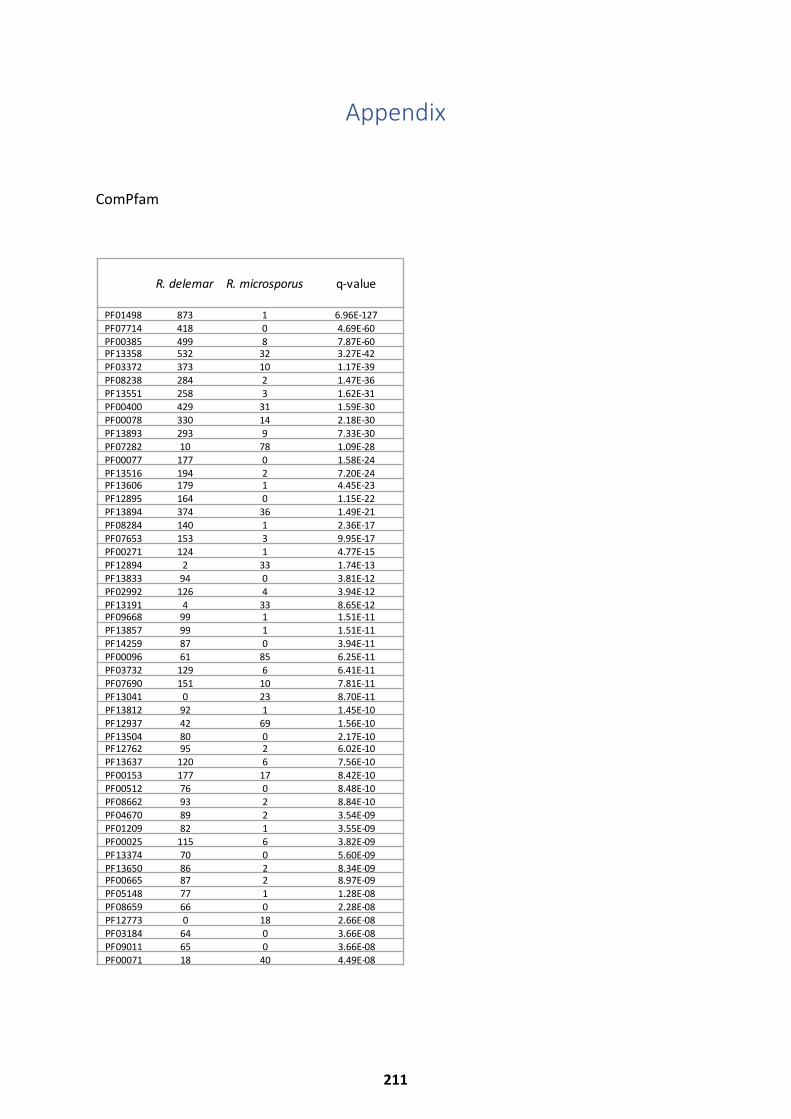
Appendix
ComPfam
R. delemar R. microsporus q-value
PF01498 873 1 6.96E-127PF07714 418 0 4.69E-60PF00385 499 8 7.87E-60PF13358 532 32 3.27E-42PF03372 373 10 1.17E-39PF08238 284 2 1.47E-36PF13551 258 3 1.62E-31PF00400 429 31 1.59E-30PF00078 330 14 2.18E-30PF13893 293 9 7.33E-30PF07282 10 78 1.09E-28PF00077 177 0 1.58E-24PF13516 194 2 7.20E-24PF13606 179 1 4.45E-23PF12895 164 0 1.15E-22PF13894 374 36 1.49E-21PF08284 140 1 2.36E-17PF07653 153 3 9.95E-17PF00271 124 1 4.77E-15PF12894 2 33 1.74E-13PF13833 94 0 3.81E-12PF02992 126 4 3.94E-12PF13191 4 33 8.65E-12PF09668 99 1 1.51E-11PF13857 99 1 1.51E-11PF14259 87 0 3.94E-11PF00096 61 85 6.25E-11PF03732 129 6 6.41E-11PF07690 151 10 7.81E-11PF13041 0 23 8.70E-11PF13812 92 1 1.45E-10PF12937 42 69 1.56E-10PF13504 80 0 2.17E-10PF12762 95 2 6.02E-10PF13637 120 6 7.56E-10PF00153 177 17 8.42E-10PF00512 76 0 8.48E-10PF08662 93 2 8.84E-10PF04670 89 2 3.54E-09PF01209 82 1 3.55E-09PF00025 115 6 3.82E-09PF13374 70 0 5.60E-09PF13650 86 2 8.34E-09PF00665 87 2 8.97E-09PF05148 77 1 1.28E-08PF08659 66 0 2.28E-08PF12773 0 18 2.66E-08PF03184 64 0 3.66E-08PF09011 65 0 3.66E-08PF00071 18 40 4.49E-08
211

RHIMIDRAFT_288130RHIMIDRAFT_288174RHIMIDRAFT_307408RHIMIDRAFT_79038RHIMIDRAFT_267844RHIMIDRAFT_221939RHIMIDRAFT_248707RHIMIDRAFT_203580RHIMIDRAFT_270714RHIMIDRAFT_283813RHIMIDRAFT_195307RHIMIDRAFT_307836RHIMIDRAFT_290452RHIMIDRAFT_232533RHIMIDRAFT_253108RHIMIDRAFT_257184RHIMIDRAFT_245053RHIMIDRAFT_245669RHIMIDRAFT_87630RHIMIDRAFT_314936RHIMIDRAFT_240622RHIMIDRAFT_234322RHIMIDRAFT_138553RHIMIDRAFT_242387RHIMIDRAFT_196349RHIMIDRAFT_99785RHIMIDRAFT_252937RHIMIDRAFT_198874RHIMIDRAFT_138931RHIMIDRAFT_271682RHIMIDRAFT_313047RHIMIDRAFT_294525RHIMIDRAFT_272110RHIMIDRAFT_252367RHIMIDRAFT_96114RHIMIDRAFT_251305RHIMIDRAFT_239731RHIMIDRAFT_281133RHIMIDRAFT_289002RHIMIDRAFT_197836
R. microsporus genes significantly (FDR<0.001) differentially expressed (40 genes). Thepredicted functions for each gene can be found in the GO file, within the online appendix(R_micro_GO).
212

213
hypothetical protein HMPREF1544_02453 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein A0J61_00230 [Choanephora cucurbitarum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA00373 [Lichtheimia ramosa]
hypothetical protein LRAMOSA09367 [Lichtheimia ramosa]
hypothetical protein HMPREF1544_05937 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome P450 CYP5313 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein A0J61_00670 [Choanephora cucurbitarum]
hypothetical protein HMPREF1544_05316 [Mucor circinelloides f. circinelloides 1006PhL]
P450 monooxygenase [Mucor ambiguus]
cytochrome P450 CYP5313 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein HMPREF1544_10867 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome p450 monooxygenase [Mucor ambiguus]
cytochrome P450 CYP5313 [Mucor circinelloides f. lusitanicus CBS 277.49]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein LRAMOSA00735 [Lichtheimia ramosa]
p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein BCR43DRAFT_510841 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein [Absidia glauca]
cytochrome P450 [Absidia repens]
cytochrome P450 [Absidia repens]
cytochrome P450 [Hesseltinella vesiculosa]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
cytochrome P450 [Absidia repens]
lanosterol 14-alpha demethylase [Hesseltinella vesiculosa]
RecName: Full=Lanosterol 14-alpha demethylase; AltName: Full=CYPLI; AltName: Full=Cytochrome P450 51; AltName: Full=Cytochrome P450-14DM; AltName: Full=Cytochrome P450-LIA1; AltName: Full...
hypothetical protein [Absidia glauca]
hypothetical protein [Parasitella parasitica]
hypothetical protein MUCCIDRAFT_110910 [Mucor circinelloides f. lusitanicus CBS 277.49]
lanosterol 14-alpha demethylase [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome P450 [Mucor ambiguus]
cytochrome P450 [Mucor ambiguus]
hypothetical protein [Parasitella parasitica]
hypothetical protein HMPREF1544_11256 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome p450 superfamily protein [Mucor ambiguus]
hypothetical protein MUCCIDRAFT_162569 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein HMPREF1544_02140 [Mucor circinelloides f. circinelloides 1006PhL]
CYP5312 protein [Mucor circinelloides f. lusitanicus CBS 277.49]
cytochrome p450 4v2 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA01771 [Lichtheimia ramosa]
cytochrome p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA00755 [Lichtheimia ramosa]
cytochrome P450 [Absidia repens]
cytochrome p450 704c1-like [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA08735 [Lichtheimia ramosa]
hypothetical protein [Absidia glauca]
cytochrome P450 CYP5203 [Phycomyces blakesleeanus NRRL 1555(-)]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
hypothetical protein A0J61_01881 [Choanephora cucurbitarum]
hypothetical protein RMATCC62417_06674 [Rhizopus microsporus]
hypothetical protein RMATCC62417_12643 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_04064 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus]
BcABA1, cytochrome P450 monooxygenase [Botrytis cinerea T4]
cytochrome P450 [Hesseltinella vesiculosa]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein LRAMOSA03645 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome P450 [Rhizopus microsporus var. microsporus]
hypothetical protein RMATCC62417_02362 [Rhizopus microsporus]
hypothetical protein RMCBS344292_14484 [Rhizopus microsporus]
hypothetical protein RMATCC62417_17980 [Rhizopus microsporus]
hypothetical protein RMCBS344292_18706 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus]
CYP5208 protein [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein [Parasitella parasitica]
cytochrome P450 [Mucor ambiguus]
BactB [Mucor circinelloides f. circinelloides]
hypothetical protein HMPREF1544_10037 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome P450 [Absidia repens]
CYP5206 protein [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein RO3G_16665 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_00866 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_08351 [Rhizopus delemar RA 99-880]
cytochrome P450 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_17241 [Rhizopus microsporus]
hypothetical protein RMATCC62417_08225 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus]
hypothetical protein RMCBS344292_03990 [Rhizopus microsporus]
0.3
BcABA1 Homolgy Tree
214
hypothetical protein RO3G_00211 [Rhizopus delemar RA 99-880]
cytochrome P450 [Rhizopus microsporus]
hypothetical protein RMATCC62417_01088 [Rhizopus microsporus]
hypothetical protein RMATCC62417_15808 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus var. microsporus]
cytochrome P450 [Rhizopus microsporus]
hypothetical protein RMCBS344292_00313 [Rhizopus microsporus]
hypothetical protein RO3G_12495 [Rhizopus delemar RA 99-880]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein LRAMOSA10222 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein [Parasitella parasitica]
cytochrome P450 [Mucor ambiguus]
hypothetical protein A0J61_01881 [Choanephora cucurbitarum]
cytochrome P450 CYP5203 [Mucor circinelloides f. lusitanicus CBS 277.49]
cytochrome P450 704C1-like [Mucor ambiguus]
hypothetical protein HMPREF1544_04052 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein [Parasitella parasitica]
hypothetical protein RMATCC62417_00219 [Rhizopus microsporus]
hypothetical protein RMATCC62417_15337 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus var. microsporus]
cytochrome P450 [Rhizopus microsporus]
hypothetical protein RMATCC62417_06674 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_04064 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus]
cytochrome P450 [Hesseltinella vesiculosa]
hypothetical protein LRAMOSA10151 [Lichtheimia ramosa]
cytochrome p450 704c1-like [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA08735 [Lichtheimia ramosa]
hypothetical protein [Absidia glauca]
hypothetical protein [Parasitella parasitica]
hypothetical protein RO3G_06009 [Rhizopus delemar RA 99-880]
cytochrome P450 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_12454 [Rhizopus microsporus]
cytochrome P450 [Rhizopus microsporus]
hypothetical protein RMATCC62417_14135 [Rhizopus microsporus]
hypothetical protein RMCBS344292_14501 [Rhizopus microsporus]
hypothetical protein RMATCC62417_06177 [Rhizopus microsporus]
hypothetical protein LRAMOSA10595 [Lichtheimia ramosa]
hypothetical protein LRAMOSA01665 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein [Parasitella parasitica]
cytochrome P450 [Mucor ambiguus]
cytochrome P450 CYP5203 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein HMPREF1544_01794 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein LRAMOSA04912 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome P450 CYP5203 [Mucor circinelloides f. lusitanicus CBS 277.49]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein LRAMOSA01155 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
cytochrome P450 [Hesseltinella vesiculosa]
cytochrome P450 [Absidia repens]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA07518 [Lichtheimia ramosa]
cytochrome P450 [Absidia repens]
cytochrome P450 [Hesseltinella vesiculosa]
cytochrome P450 [Hesseltinella vesiculosa]
CYP5205 protein [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein LRAMOSA09445 [Lichtheimia ramosa]
hypothetical protein RO3G_10498 [Rhizopus delemar RA 99-880]
CYP5205 protein [Mucor circinelloides f. lusitanicus CBS 277.49]
BcABA2, cytochrome P450 monooxygenase [Botrytis cinerea T4]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 CYP5313 [Phycomyces blakesleeanus NRRL 1555(-)]
cytochrome P450 [Hesseltinella vesiculosa]
hypothetical protein [Absidia glauca]
cytochrome P450 [Absidia repens]
hypothetical protein [Parasitella parasitica]
hypothetical protein BCR43DRAFT_510841 [Syncephalastrum racemosum]
hypothetical protein A0J61_00670 [Choanephora cucurbitarum]
cytochrome p450 monooxygenase [Mucor ambiguus]
hypothetical protein HMPREF1544_10867 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome P450 CYP5313 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein [Parasitella parasitica]
P450 monooxygenase [Mucor ambiguus]
hypothetical protein HMPREF1544_05316 [Mucor circinelloides f. circinelloides 1006PhL]
cytochrome P450 CYP5313 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein LRAMOSA00735 [Lichtheimia ramosa]
p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
p450 monooxygenase [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA01501 [Lichtheimia ramosa]
cytochrome p450 [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA00734 [Lichtheimia ramosa]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
cytochrome P450 [Syncephalastrum racemosum]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
cytochrome P450 [Absidia repens]
cytochrome P450 [Absidia repens]
0.3BcABA2 Homolgy Tree 215
NAD-binding protein [Absidia repens]
hypothetical protein [Absidia glauca]
Peroxisomal 2,4-dienoyl-CoA reductase SPS19 [Choanephora cucurbitarum]
Putative 2,4-dienoyl-CoA reductase (NADPH2) [Rhizopus microsporus]
Putative 2,4-dienoyl-CoA reductase (NADPH2) [Rhizopus microsporus]
Putative 2,4-dienoyl-CoA reductase (NADPH2) [Rhizopus microsporus]
peroxisomal 2,4-dienoyl-CoA reductase [Hesseltinella vesiculosa]
2,4-dienoyl-CoA reductase [Absidia repens]
hypothetical protein [Absidia glauca]
hypothetical protein LRAMOSA10708 [Lichtheimia ramosa]
-dienoyl-reductase [Lichtheimia corymbifera JMRC:FSU:9682]
NAD(P)-binding protein [Hesseltinella vesiculosa]
hypothetical protein BCR43DRAFT_459156 [Syncephalastrum racemosum]
hypothetical protein [Absidia glauca]
carbonyl reductase family member 4-like protein [Syncephalastrum racemosum]
hypothetical protein [Parasitella parasitica]
hypothetical protein HMPREF1544_10064 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein MUCCIDRAFT_134408 [Mucor circinelloides f. lusitanicus CBS 277.49]
phospholipid-transporting ATPase [Mucor ambiguus]
NAD(P)-binding protein [Hesseltinella vesiculosa]
hypothetical protein [Absidia glauca]
3-oxoacyl-reductase FabG-like protein [Absidia repens]
3-oxoacyl-reductase [Rhizopus microsporus]
hypothetical protein RMATCC62417_16579 [Rhizopus microsporus]
hypothetical protein RMCBS344292_07747 [Rhizopus microsporus]
hypothetical protein LRAMOSA10250 [Lichtheimia ramosa]
3-oxoacyl-(acyl-carrier-protein) reductase [Lichtheimia corymbifera JMRC:FSU:9682]
3-ketoacyl-(acyl-carrier-protein) reductase [Lichtheimia corymbifera JMRC:FSU:9682]
BcABA4 [Botrytis cinerea T4]
hypothetical protein [Absidia glauca]
hypothetical protein BCR42DRAFT_446447 [Absidia repens]
hypothetical protein [Absidia glauca]
Putative Glucose 1-dehydrogenase [Rhizopus microsporus]
hypothetical protein PHYBLDRAFT_128144 [Phycomyces blakesleeanus NRRL 1555(-)]
3-oxoacyl-[acyl-carrier-protein] reductase FabG [Choanephora cucurbitarum]
3-hydroxyacyl-CoA-dehydrogenase, mitochondrial [Mucor ambiguus]
glucose 1-dehydrogenase [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein MUCCIDRAFT_156993 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein [Parasitella parasitica]
hypothetical protein BCR43DRAFT_514859 [Syncephalastrum racemosum]
Putative Glucose 1-dehydrogenase [Lichtheimia ramosa]
short-chain dehydrogenase [Lichtheimia corymbifera JMRC:FSU:9682]
NAD(P)-binding protein [Hesseltinella vesiculosa]
hypothetical protein [Absidia glauca]
hypothetical protein BCR43DRAFT_453530 [Syncephalastrum racemosum]
3-oxoacyl-[acyl-carrier-protein] reductase FabG [Choanephora cucurbitarum]
hypothetical protein [Parasitella parasitica]
hypothetical protein MUCCIDRAFT_46792 [Mucor circinelloides f. lusitanicus CBS 277.49]
NAD(P)-binding protein [Mucor ambiguus]
3-oxoacyl-[acyl-carrier protein] reductase [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein PHYBLDRAFT_26103 [Phycomyces blakesleeanus NRRL 1555(-)]
hypothetical protein RO3G_16300 [Rhizopus delemar RA 99-880]
Putative 3-oxoacyl-[acyl-carrier protein] reductase [Rhizopus microsporus]
NAD(P)-binding protein [Rhizopus microsporus]
NAD(P)-binding protein [Rhizopus microsporus var. microsporus]
Putative 3-oxoacyl-[acyl-carrier protein] reductase [Rhizopus microsporus]
Putative 3-oxoacyl-[acyl-carrier protein] reductase [Rhizopus microsporus]
Putative 3-oxoacyl-[acyl-carrier protein] reductase [Rhizopus microsporus]
hypothetical protein LRAMOSA06177 [Lichtheimia ramosa]
sex determination protein tasselseed-2 [Lichtheimia corymbifera JMRC:FSU:9682]
3-oxoacyl-(acyl-carrier-protein) reductase [Mucor ambiguus]
hypothetical protein HMPREF1544_11236 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein BCR42DRAFT_402595 [Absidia repens]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
NAD(P)-binding protein [Hesseltinella vesiculosa]
hypothetical protein [Absidia glauca]
hypothetical protein [Absidia glauca]
hypothetical protein BCR42DRAFT_468603 [Absidia repens]
hypothetical protein [Absidia glauca]
hypothetical protein BCR42DRAFT_417976 [Absidia repens]
hypothetical protein BCR42DRAFT_338541 [Absidia repens]
hypothetical protein BCR42DRAFT_421293 [Absidia repens]
hypothetical protein BCR42DRAFT_395194 [Absidia repens]
hypothetical protein BCR42DRAFT_421288 [Absidia repens]
Sorbose reductase SOU1 [Choanephora cucurbitarum]
hypothetical protein [Parasitella parasitica]
L-xylulose reductase [Rhizomucor pusillus]
hypothetical protein HMPREF1544_00552 [Mucor circinelloides f. circinelloides 1006PhL]
hypothetical protein MUCCIDRAFT_148022 [Mucor circinelloides f. lusitanicus CBS 277.49]
3-oxoacyl-ACP reductase [Mucor ambiguus]
hypothetical protein RO3G_01452 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_01846 [Rhizopus delemar RA 99-880]
NAD(P)-binding protein [Rhizopus microsporus]
hypothetical protein RMCBS344292_12835 [Rhizopus microsporus]
NAD(P)-binding protein [Rhizopus microsporus var. microsporus]
hypothetical protein RMATCC62417_05492 [Rhizopus microsporus]
hypothetical protein RMATCC62417_10579 [Rhizopus microsporus]
hypothetical protein BCR42DRAFT_339880 [Absidia repens]
hypothetical protein PHYBLDRAFT_125203 [Phycomyces blakesleeanus NRRL 1555(-)]
hypothetical protein BCR43DRAFT_521475 [Syncephalastrum racemosum]
short-chain dehydrogenase reductase sdr [Lichtheimia corymbifera JMRC:FSU:9682]
hypothetical protein LRAMOSA11265 [Lichtheimia ramosa]
hypothetical protein BCR43DRAFT_455482 [Syncephalastrum racemosum]
hypothetical protein [Absidia glauca]
hypothetical protein BCR42DRAFT_365058 [Absidia repens]
hypothetical protein [Absidia glauca]
hypothetical protein BCR42DRAFT_492670 [Absidia repens]
hypothetical protein MUCCIDRAFT_151684 [Mucor circinelloides f. lusitanicus CBS 277.49]
hypothetical protein HMPREF1544_08721 [Mucor circinelloides f. circinelloides 1006PhL]
TerpG [Mucor circinelloides f. circinelloides]0.2BcABA4 Homology Tree 216
hypothetical protein RO3G_07800 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_12412 [Rhizopus delemar RA 99-880]
hypothetical protein BCV71DRAFT_267961 [Rhizopus microsporus]
hypothetical protein RMATCC62417_05318 [Rhizopus microsporus]
hypothetical protein RMATCC62417_05318 [Rhizopus microsporus]
Putative STE/STE20/PAKA protein kinase [Rhizopus microsporus]
Putative STE/STE20/PAKA protein kinase [Rhizopus microsporus]
Putative STE/STE20/PAKA protein kinase [Rhizopus microsporus]
Pkinase-domain-containing protein [Rhizopus microsporus]
Pkinase-domain-containing protein [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_11067 [Rhizopus microsporus]
hypothetical protein RMATCC62417_17169 [Rhizopus microsporus]
hypothetical protein BCV72DRAFT_71979 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_04428 [Rhizopus microsporus]
hypothetical protein RMATCC62417_09711 [Rhizopus microsporus]
hypothetical protein RMCBS344292_07609 [Rhizopus microsporus]
hypothetical protein RMATCC62417_07950 [Rhizopus microsporus]
TPR-like protein [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_06885 [Rhizopus microsporus]
hypothetical protein RMATCC62417_07950 [Rhizopus microsporus]
hypothetical protein RO3G_03312 [Rhizopus delemar RA 99-880]
hypothetical protein RMATCC62417_14752 [Rhizopus microsporus]
hypothetical protein RMATCC62417_14752 [Rhizopus microsporus]
UPF0014-domain-containing protein [Rhizopus microsporus var. microsporus]
hypothetical protein RMATCC62417_07219 [Rhizopus microsporus]
hypothetical protein RMATCC62417_11126 [Rhizopus microsporus]
hypothetical protein BCV72DRAFT_231313 [Rhizopus microsporus var. microsporus]
hypothetical protein RMATCC62417_05403 [Rhizopus microsporus]
hypothetical protein BCV71DRAFT_279323 [Rhizopus microsporus]
hypothetical protein RMCBS344292_00256 [Rhizopus microsporus]
hypothetical protein RO3G_11943 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_14028 [Rhizopus delemar RA 99-880]
0.3
DELLA
Dell Homology Tree 217
hypothetical protein RMATCC62417_13746 [Rhizopus microsporus]
hypothetical protein RMATCC62417_13746 [Rhizopus microsporus]
hypothetical protein RMCBS344292_16129 [Rhizopus microsporus]
hypothetical protein RMATCC62417_15451 [Rhizopus microsporus]
hypothetical protein RMATCC62417_15451 [Rhizopus microsporus]
hypothetical protein RO3G_14096 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_03851 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_01805 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16636 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16675 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16666 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16672 [Rhizopus delemar RA 99-880]
GLP1 GID1-like protein [Physcomitrella patens]
hypothetical protein RO3G_16524 [Rhizopus delemar RA 99-880]
hypothetical protein RMATCC62417_02350 [Rhizopus microsporus]
hypothetical protein RMCBS344292_07419 [Rhizopus microsporus]
hypothetical protein BCV71DRAFT_178021 [Rhizopus microsporus]
hypothetical protein BCV72DRAFT_212589 [Rhizopus microsporus var. microsporus]
alpha/beta-hydrolase [Rhizopus microsporus var. microsporus]
hypothetical protein RO3G_08298 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_00099 [Rhizopus delemar RA 99-880]
alpha/beta-hydrolase [Rhizopus microsporus]
hypothetical protein RMCBS344292_02919 [Rhizopus microsporus]
hypothetical protein RO3G_12741 [Rhizopus delemar RA 99-880]
alpha/beta-hydrolase [Rhizopus microsporus]
alpha/beta-hydrolase [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_07649 [Rhizopus microsporus]0.6
GLP1 Homolgy Tree
218
hypothetical protein RO3G_03851 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_01805 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16636 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16672 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16666 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16675 [Rhizopus delemar RA 99-880]
hypothetical protein BCV71DRAFT_178021 [Rhizopus microsporus]
hypothetical protein BCV72DRAFT_212589 [Rhizopus microsporus var. microsporus]
hypothetical protein RMCBS344292_07419 [Rhizopus microsporus]
hypothetical protein RMATCC62417_02350 [Rhizopus microsporus]
hypothetical protein RO3G_02078 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_12741 [Rhizopus delemar RA 99-880]
alpha/beta-hydrolase [Rhizopus microsporus]
hypothetical protein RMCBS344292_07649 [Rhizopus microsporus]
alpha/beta-hydrolase [Rhizopus microsporus var. microsporus]
GLP2 GID1-like protein [Physcomitrella patens]
hypothetical protein RO3G_00099 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_08298 [Rhizopus delemar RA 99-880]
hypothetical protein RMCBS344292_02919 [Rhizopus microsporus]
hypothetical protein RMATCC62417_02738 [Rhizopus microsporus]
alpha/beta-hydrolase [Rhizopus microsporus]
alpha/beta-hydrolase [Rhizopus microsporus var. microsporus]
0.5
GLP2 Homology Tree
219
hypothetical protein RO3G_00099 [Rhizopus delemar RA 99-880]
GLP4 GID1-like protein [Physcomitrella patens]
hypothetical protein RO3G_02534 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_01805 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16636 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16672 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16666 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_16675 [Rhizopus delemar RA 99-880]
hypothetical protein RO3G_03851 [Rhizopus delemar RA 99-880]
hypothetical protein BCV72DRAFT_227955 [Rhizopus microsporus var. microspo...
hypothetical protein RMATCC62417_13084 [Rhizopus microsporus]
hypothetical protein RMCBS344292_04013 [Rhizopus microsporus]
hypothetical protein BCV71DRAFT_82860 [Rhizopus microsporus]0.5
Glp4 Homology Tree
220
Related Documents











