Uncoupling of Acetylation from Phosphorylation Regulates FoxO1 Function Independent of Its Subcellular Localization * Received for publication, May 1, 2010 Published, JBC Papers in Press, June 2, 2010, DOI 10.1074/jbc.M110.140228 Li Qiang ‡ , Alexander S. Banks § , and Domenico Accili ‡1 From the ‡ Department of Medicine, Columbia University, New York, New York 10032 and § Dana Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts 02115 The activity of transcription factor FoxO1 is regulated by phosphorylation-dependent nuclear exclusion and deacetyla- tion-dependent nuclear retention. It is unclear whether and how these two post-translational modifications affect each other. To answer this question, we expressed FoxO1 cDNAs with com- bined mutations of phosphorylation and acetylation sites in HEK-293 cells and analyzed their subcellular localization pat- terns. We show that mutations mimicking the acetylated state (KQ series) render FoxO1 more sensitive to Akt-mediated phos- phorylation and nuclear exclusion and can reverse the constitu- tively nuclear localization of phosphorylation-defective FoxO1. Conversely, mutations mimicking the deacetylated state (KR series) promote FoxO1 nuclear retention. Oxidative stress and the Sirt1 activator resveratrol are thought to promote FoxO1 deacetylation and nuclear retention, thus increasing its activity. Accordingly, FoxO1 deacetylation was required for the effect of oxidative stress (induced by H 2 O 2 ) to retain FoxO1 in the nucleus. H 2 O 2 also inhibited FoxO1 phosphorylation on Ser- 253 and Thr-24, the key insulin-regulated sites, irrespective of its acetylation. In contrast, the effect of resveratrol was inde- pendent of FoxO1 acetylation and its phosphorylation on Ser- 253 and Thr-24, suggesting that resveratrol acts on FoxO1 in a Sirt1- and Akt-independent manner. The dissociation of deacetylation from dephosphorylation in H 2 O 2 -treated cells indicates that the two modifications can occur independently of each other. It can be envisaged that FoxO1 exists in multiple nuclear forms with distinct activities depending on the balance of acetylation and phosphorylation. FoxO1 and its closely related isoforms FoxO3A and FoxO4 are transcription factors characterized by a conserved winged helix (“forkhead”) DNA binding domain. Genetic epistasis experiments in Caenorhabditis elegans demonstrated a role for these proteins in insulin receptor signaling, spawning studies of their contribution to mammalian metabolism, cellular differen- tiation, and transformation (1). It is now recognized that FoxOs are critical regulators of hepatic gluconeogenesis (2, 3) and pan- creatic -cell function (4 – 8), in addition to differentiation of myotubes (9 –11) and adipocytes (12). Moreover, the C. elegans FoxO ortholog DAF-16 is required for life extension caused by DAF-2 (insulin receptor) mutations, suggesting that FoxO has a role in longevity (13, 14). FoxO activity is regulated by post-translational modifica- tions that affect primarily its subcellular localization (15). Insu- lin and growth factor signaling inhibit FoxO via Akt-dependent phosphorylation and nuclear exclusion (16 –18). Several addi- tional serine/threonine kinases, such as Mst1 (19), Jnk (20), and Sgk promote or inhibit FoxO via nuclear translocation (20 –22) or nuclear exclusion, respectively (23–25). A second regulatory layer is FoxO acetylation by p300, CBP (cAMP-response ele- ment-binding protein-binding protein), and PCAF (p300/CBP- associated factors) in response to oxidative stress or DNA bind- ing (26 –28), followed by deacetylation by class I and II histone deacetylases (26, 28 –30), including Sirt1, the NAD -depen- dent deacetylase encoded by the ortholog of yeast longevity gene Sir2 (31). The effects of phosphorylation and acetylation on FoxO function have been studied extensively but separately. How- ever, these two modifications are likely to occur concurrently in vivo and to reciprocally affect each other. In this study, we gen- erated an allelic series of FoxO1 mutants containing changes to both acetylation and phosphorylation sites and analyzed their regulation in response to physiologic (insulin) and pathophys- iologic cues (oxidative stress, resveratrol) to explore the recip- rocal regulation of acetylation and phosphorylation and their combined effects on FoxO1 cellular localization and biological functions. EXPERIMENTAL PROCEDURES Materials—Dulbecco’s modified Eagle’s medium with 4.5 g/liter glucose, fetal bovine serum, calf serum, trypsin/EDTA, and phosphate-buffered saline were purchased from Mediatech (Manassas, VA). Insulin, H 2 O 2 , resveratrol, nicotinamide, and cycloheximide were purchased from Sigma, Akti-1/2 from EMD, microcystin-LR from Cayman, and leptomycin B from LC Laboratories. Anti-FLAG (M2) affinity gel was purchased from Sigma. Anti-phospho-S253, phospho-T24 FoxO1, and phospho-Akt antibodies (T308) were purchased from Cell Sig- naling. Anti-GFP, 2 anti-tubulin, and anti-FLAG antibodies were purchased from Santa Cruz Biotechnology. Plasmids, Adenoviruses, and Cell Culture—cDNAs encoding murine FoxO1 and carrying the following mutations, T24A, T24A-KQ, T24A-KR, and S253A, were subcloned into pEGFP-N1 to generate FoxO1-GFP fusion proteins. Plasmids * This work was supported by National Institutes of Health Grants DK57539 and DK63608 from the Columbia University Diabetes and Endocrinology Research Center. 1 To whom correspondence should be addressed: 1150 St. Nicholas Ave., Rm. 238, New York, NY 10032. Fax: 212-851-5335; E-mail: da230@ columbia.edu. 2 The abbreviations used are: GFP, green fluorescent protein; ADA, T24A/ S253D/S316A; WT, wild type. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 35, pp. 27396 –27401, August 27, 2010 © 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 27396 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 35 • AUGUST 27, 2010 by guest on April 18, 2016 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Uncoupling of Acetylation from Phosphorylation RegulatesFoxO1 Function Independent of Its Subcellular Localization*
Received for publication, May 1, 2010 Published, JBC Papers in Press, June 2, 2010, DOI 10.1074/jbc.M110.140228
Li Qiang‡, Alexander S. Banks§, and Domenico Accili‡1
From the ‡Department of Medicine, Columbia University, New York, New York 10032 and §Dana Farber Cancer Institute, HarvardMedical School, Boston, Massachusetts 02115
The activity of transcription factor FoxO1 is regulated byphosphorylation-dependent nuclear exclusion and deacetyla-tion-dependentnuclear retention. It is unclearwhether andhowthese two post-translational modifications affect each other. Toanswer this question, we expressed FoxO1 cDNAs with com-bined mutations of phosphorylation and acetylation sites inHEK-293 cells and analyzed their subcellular localization pat-terns. We show that mutations mimicking the acetylated state(KQ series) render FoxO1more sensitive toAkt-mediated phos-phorylation and nuclear exclusion and can reverse the constitu-tively nuclear localization of phosphorylation-defective FoxO1.Conversely, mutations mimicking the deacetylated state (KRseries) promote FoxO1 nuclear retention. Oxidative stress andthe Sirt1 activator resveratrol are thought to promote FoxO1deacetylation and nuclear retention, thus increasing its activity.Accordingly, FoxO1 deacetylation was required for the effect ofoxidative stress (induced by H2O2) to retain FoxO1 in thenucleus. H2O2 also inhibited FoxO1 phosphorylation on Ser-253 and Thr-24, the key insulin-regulated sites, irrespective ofits acetylation. In contrast, the effect of resveratrol was inde-pendent of FoxO1 acetylation and its phosphorylation on Ser-253 and Thr-24, suggesting that resveratrol acts on FoxO1 ina Sirt1- and Akt-independent manner. The dissociation ofdeacetylation from dephosphorylation in H2O2-treated cellsindicates that the twomodifications can occur independently ofeach other. It can be envisaged that FoxO1 exists in multiplenuclear forms with distinct activities depending on the balanceof acetylation and phosphorylation.
FoxO1 and its closely related isoforms FoxO3A and FoxO4are transcription factors characterized by a conserved wingedhelix (“forkhead”) DNA binding domain. Genetic epistasisexperiments in Caenorhabditis elegans demonstrated a role forthese proteins in insulin receptor signaling, spawning studies oftheir contribution tomammalianmetabolism, cellular differen-tiation, and transformation (1). It is now recognized that FoxOsare critical regulators of hepatic gluconeogenesis (2, 3) and pan-creatic �-cell function (4–8), in addition to differentiation ofmyotubes (9–11) and adipocytes (12). Moreover, theC. elegansFoxO ortholog DAF-16 is required for life extension caused by
DAF-2 (insulin receptor)mutations, suggesting that FoxOhas arole in longevity (13, 14).FoxO activity is regulated by post-translational modifica-
tions that affect primarily its subcellular localization (15). Insu-lin and growth factor signaling inhibit FoxO via Akt-dependentphosphorylation and nuclear exclusion (16–18). Several addi-tional serine/threonine kinases, such asMst1 (19), Jnk (20), andSgk promote or inhibit FoxO via nuclear translocation (20–22)or nuclear exclusion, respectively (23–25). A second regulatorylayer is FoxO acetylation by p300, CBP (cAMP-response ele-ment-binding protein-binding protein), and PCAF (p300/CBP-associated factors) in response to oxidative stress orDNAbind-ing (26–28), followed by deacetylation by class I and II histonedeacetylases (26, 28–30), including Sirt1, the NAD�-depen-dent deacetylase encoded by the ortholog of yeast longevitygene Sir2 (31).
The effects of phosphorylation and acetylation on FoxOfunction have been studied extensively but separately. How-ever, these twomodifications are likely to occur concurrently invivo and to reciprocally affect each other. In this study, we gen-erated an allelic series of FoxO1mutants containing changes toboth acetylation and phosphorylation sites and analyzed theirregulation in response to physiologic (insulin) and pathophys-iologic cues (oxidative stress, resveratrol) to explore the recip-rocal regulation of acetylation and phosphorylation and theircombined effects on FoxO1 cellular localization and biologicalfunctions.
EXPERIMENTAL PROCEDURES
Materials—Dulbecco’s modified Eagle’s medium with 4.5g/liter glucose, fetal bovine serum, calf serum, trypsin/EDTA,andphosphate-buffered salinewere purchased fromMediatech(Manassas, VA). Insulin, H2O2, resveratrol, nicotinamide, andcycloheximide were purchased from Sigma, Akti-1/2 fromEMD, microcystin-LR from Cayman, and leptomycin B fromLC Laboratories. Anti-FLAG (M2) affinity gel was purchasedfrom Sigma. Anti-phospho-S253, phospho-T24 FoxO1, andphospho-Akt antibodies (T308) were purchased from Cell Sig-naling. Anti-GFP,2 anti-tubulin, and anti-FLAG antibodieswere purchased from Santa Cruz Biotechnology.Plasmids, Adenoviruses, and Cell Culture—cDNAs encoding
murine FoxO1 and carrying the following mutations, T24A,T24A-KQ, T24A-KR, and S253A, were subcloned intopEGFP-N1 to generate FoxO1-GFP fusion proteins. Plasmids
* This work was supported by National Institutes of Health Grants DK57539and DK63608 from the Columbia University Diabetes and EndocrinologyResearch Center.
1 To whom correspondence should be addressed: 1150 St. Nicholas Ave.,Rm. 238, New York, NY 10032. Fax: 212-851-5335; E-mail: [email protected].
2 The abbreviations used are: GFP, green fluorescent protein; ADA, T24A/S253D/S316A; WT, wild type.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 35, pp. 27396 –27401, August 27, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
27396 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 35 • AUGUST 27, 2010
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from

pEGFP-N1 encoding WT, KQ, KR, ADA, ADA-KR, andADA-KQ FoxO1 have been described previously (8). Weused TransIT transfection reagent from Mirus (Madison,WI) for cell transfection. HEK-293 cells were cultured asdescribed (10). In some experiments, we used adenovirusesencoding FoxO1-KR and -KQ mutants as described previ-ously (8).Protein Analyses—We harvested cells and prepared protein
extracts in buffer containing 20mMTris, pH 7.4, 150mMNaCl,10% glycerol, 2%Nonidet P-40, 1mMEDTA, pH 8.0, 0.2% semi-dehydroascorbate, 0.5% sodium deoxycholate supplementedwith protease and phosphatase inhibitors (Boston Bioprod-ucts). We fractionated 40 �g of protein by gel electrophoresis,followed by Western blot. Immunoprecipitation was carriedout by standardmethods. FLAG immunoprecipitation was car-ried out according to the manufacturer’s instructions, andbound proteins were eluted using a FLAG peptide.
RESULTS AND DISCUSSION
Generation of FoxO1Mutants—In addition to the previouslydescribed constitutively acetylated (FoxO1-KQ, in which lysineat amino acid residues 219, 242, 245, 259, 262, 271, and 291 isreplaced with glutamine) and constitutively deacetylatedmutants (FoxO1-KR, in which the same lysine residues arereplaced with arginine) (Fig. 1A) (8), we generated an allelicseries in which the KR and KQ mutations were introducedalong with the following mutations of the three main phospho-rylation sites: T24A, S253A, or combined T24A/S253D/S316A(ADA mutant) (12). Ser-253 is the main Akt site, whereasThr-24 is phosphorylated by an insulin-activated kinase(s) dis-
tinct from Akt (23, 25). We generated the mutants KR, KQ,S253A, T24A-KQ, T24A-KR, ADA-KQ, and ADA-KR as GFPfusion proteins to facilitate their detection by fluorescencemicroscopy (30). For simplicity, we omit “FoxO1-GFP” fromthe nomenclature.Effects of Acetylation Site Mutations on FoxO1 Subcellular
Localization—We studied the effect of mutating acetylationsites on insulin-induced FoxO1 subcellular translocation. Tothis end, we transfectedwild type (WT), KQ, or KRFoxO1-GFPfusion proteins intoHEK-293 cells.WT localized to the nucleusin serum-free medium and translocated to the cytoplasm uponinsulin stimulation. The Akt inhibitor Akti-1/2 inhibited thisprocess (33) (Fig. 1B). Conversely, the KQ mutant was pre-dominantly cytoplasmic, regardless of whether cells wereincubated in serum-free medium or in the presence of insu-lin and Akt inhibitor. The KR mutant translocated to thecytoplasm after insulin stimulation in a WT-like fashion andwas retained in the nucleus after Akti-1/2 treatment,whereas the S253A mutant was constitutively nuclear underall conditions tested (Fig. 1B) (24).We next examined whether acetylation trumps phosphory-
lation as a signal for FoxO1 retention in the nucleus. To thisend, we compared the phosphorylation-defective mutant ADA(12) with combined phosphorylation/acetylation site mutants,ADA-KQ or ADA-KR. ADAwas constitutively nuclear regard-less of the culture conditions (Fig. 2A). Surprisingly, ADA-KQhad predominantly cytoplasmic localization, whereas ADA-KRwas retained in the nucleus (Fig. 2A). These data indicate thatacetylation trumps phosphorylation as a signal regulatingFoxO1 cellular localization. Inhibition of nuclear export by lep-tomycin B resulted in nuclear accumulation of both KQ andADA-KQ, indicating that acetylation doesn’t prevent nucleartargeting of FoxO1 but likely accelerates its export to the cyto-plasm or retards its nuclear import (Fig. 2B).To rule out that mutation of the phosphorylation sites
affected FoxO1 acetylation, we measured acetyl-FoxO1 levelsin the phosphorylation site mutants, S253A and T24A. As theformer is unaffected by insulin treatment, we measured onlybasal acetylation in the absence of insulin; in the latter, we com-pared acetylation levels in the absence and presence of insulin.However in neither case did we observe changes to FoxO1acetylation (Fig. 2C).Effects of Acetylation Site Mutations on Insulin-induced
FoxO1 Phosphorylation—To understand why acetylation pro-motes FoxO1nuclear exclusion,we examinedwhether it affectsphosphorylation of Ser-253, the site required for insulin-de-pendent nuclear translocation (24). To avoid the potential con-founding effects of Thr-24 phosphorylation on subcellularlocalization (24), we measured Ser-253 phosphorylation inT24A-KQ andT24A-KRmutants following exposure of cells todifferent doses of insulin. Insulin-induced Ser-253 phosphory-lation of the T24A mutant paralleled Akt phosphorylation in adose-dependent manner, with an ED50 � 0.3 nM (Fig. 3). Incontrast, the ED50 for Ser-253 phosphorylation decreased to�0.15 nM in the T24A-KQ mutant and rose to �1.5 nM in theT24A-KRmutant, resulting in a 10-fold difference between thetwo mutants. Interestingly, levels of the T24A-KQ mutantdecreased in insulin-treated cells. Based on prior studies, this is
FIGURE 1. Effect of insulin stimulation and Akt inhibition on FoxO1 acety-lation site mutants. A, diagram summarizing the location of phosphoryla-tion (purple circles) and acetylation sites (green triangles) in FoxO1. B, HEK-293cells transfected with WT FoxO1-GFP or KQ, KR, and S253A mutants. Cellswere treated with insulin and Akti for 30 min, and FoxO1 localization wasvisualized using a fluorescent microscope.
FoxO1 Acetylation and Phosphorylation
AUGUST 27, 2010 • VOLUME 285 • NUMBER 35 JOURNAL OF BIOLOGICAL CHEMISTRY 27397
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from

likely to reflect increased protein degradation through the pro-teasome (8). This process was reversed byAkt inhibition, as wasSer-253 phosphorylation (Fig. 3). These data indicate thatacetylation increases FoxO1 sensitivity to Akt phosphorylationand degradation, suggesting that FoxO1 nuclear exclusion andprotein turnover are integrated through acetylation-basedmechanisms.Uncoupling of Acetylation from Phosphorylation following
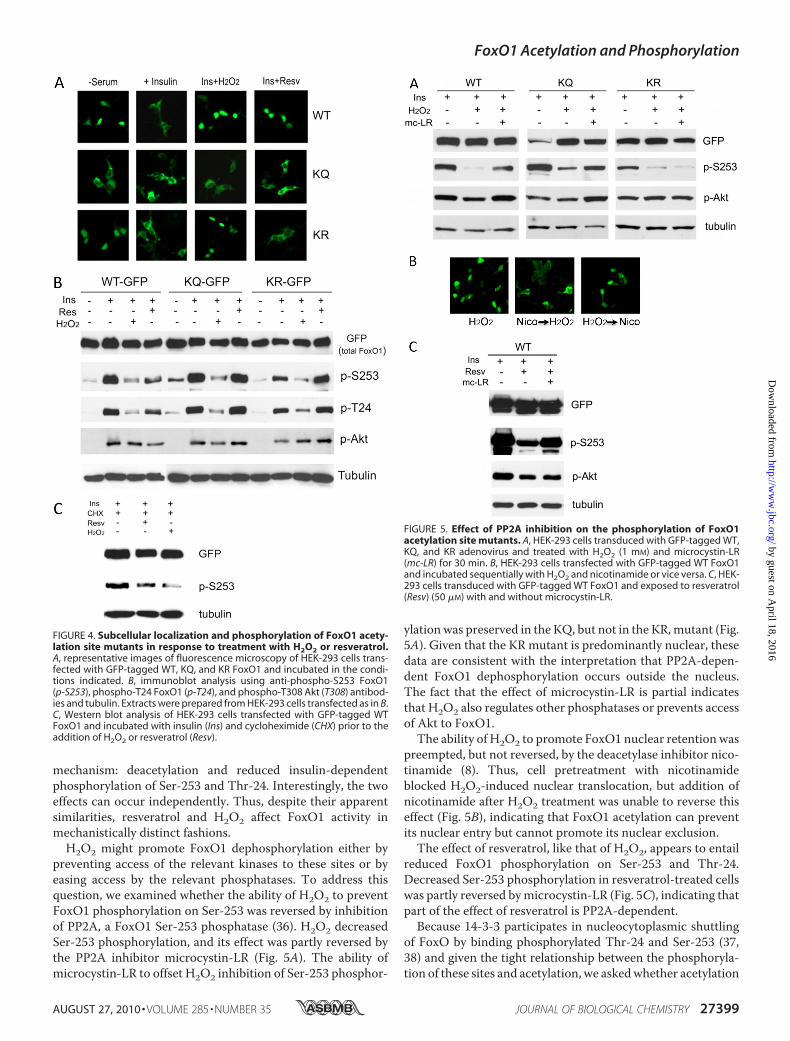
H2O2-induced Oxidative Stress—Oxidative stress and the poly-phenol resveratrol promote FoxO1 nuclear retention (30).Their effects have been ascribed to FoxO1deacetylation (27, 28,34). However, the data in Fig. 3 raise the possibility that theyalso inhibit FoxO1 phosphorylation or promote its dephos-phorylation. To answer this question, we used acetylation siteFoxO1 mutants to examine FoxO1 localization and phosphor-
ylation following incubation of cells with insulin and H2O2, achemical agent used to mimic oxidative stress (35), or insulinand resveratrol, a Sirt1 and AMP-activated protein kinase ago-nist (29, 30). Addition of H2O2 to insulin-treated cells pre-vented FoxO1 nuclear export. This effect was reversed by theconstitutively acetylated KQ mutant but not by the deacety-lated KR mutant, indicating that H2O2 promotes FoxO1deacetylation or requires that FoxO1 be deacetylated to keep itin the nucleus (Fig. 4A). Resveratrol also prevented FoxO1nuclear exclusion in response to insulin but, unlikeH2O2, failedto prevent nuclear exclusion of either KQ or KR mutants (Fig.4A), indicating that its effects are independent of FoxO1acetylation.Next we compared the effects of H2O2 and resveratrol on
insulin-dependent phosphorylation of Ser-253 and Thr-24.Insulin promoted FoxO1 phosphorylation on both sites. Addi-tion ofH2O2 to insulin decreased phosphorylation of both sites.The effect of H2O2 was preserved in the KQ and KR mutants,indicating that it is independent of FoxO1 acetylation (Fig. 4B).Addition of resveratrol to insulin also decreased insulin-depen-dent Ser-253 and Thr-24 phosphorylation in WT FoxO1, butnot in the KQ and KR mutants (Fig. 4B). Neither H2O2 norresveratrol affected insulin-induced Akt phosphorylation to asignificant extent (Fig. 4B), and their effect on Ser-253 phos-phorylation was independent of changes in FoxO1 protein lev-els, as indicated by the fact that they retained their ability todecrease Ser(P)-253 in the presence of the protein synthesisinhibitor, cycloheximide (Fig. 4C).From these experiments, we concluded that the effects of
resveratrol are mediated neither by changes in FoxO1 acetyla-tion nor by dephosphorylation of Ser-253 and Thr-24. In con-trast, H2O2 promotes FoxO1 nuclear retention through a dual
FIGURE 2. Subcellular localization of FoxO1 acetylation site mutants inresponse to insulin. A, HEK-293 cells transfected with GFP-tagged WT orADA-KQ and ADA-KR FoxO1 and incubated in the absence and presence ofinsulin prior to visualizing FoxO1 localization by fluorescence microscopy.B, HEK-293 cells transfected with GFP-tagged KQ and ADA-KQ and incubatedin the absence and presence of serum or leptomycin B (LMB) prior to visual-izing FoxO1 localization by fluorescence microscopy. C, FoxO1 acetylationwas determined by immunoblotting with anti-acetyllysine antiserum follow-ing immunoprecipitation with anti-FLAG (M2) antibody of protein extractsfrom HEK-293 cells transfected with FLAG-tagged FoxO1 mutants S253A (SA)or T24A (TA), the latter treated with and without insulin.
FIGURE 3. Phosphorylation of FoxO1 acetylation site mutants. HEK-293cells were transfected with FLAG-tagged T24A, T24A-KQ, or T24A-KR FoxO1and treated with different concentrations of insulin. Phosphorylation ofFoxO1 and Akt was determined by Western blotting with anti-phospho-S253FoxO1 (p-S253), phospho-T24 FoxO1 (p-T24), and phospho-Akt (T308) anti-bodies. Tubulin was used as the gel loading control.
FoxO1 Acetylation and Phosphorylation
27398 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 35 • AUGUST 27, 2010
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from
mechanism: deacetylation and reduced insulin-dependentphosphorylation of Ser-253 and Thr-24. Interestingly, the twoeffects can occur independently. Thus, despite their apparentsimilarities, resveratrol and H2O2 affect FoxO1 activity inmechanistically distinct fashions.H2O2 might promote FoxO1 dephosphorylation either by
preventing access of the relevant kinases to these sites or byeasing access by the relevant phosphatases. To address thisquestion, we examined whether the ability of H2O2 to preventFoxO1 phosphorylation on Ser-253 was reversed by inhibitionof PP2A, a FoxO1 Ser-253 phosphatase (36). H2O2 decreasedSer-253 phosphorylation, and its effect was partly reversed bythe PP2A inhibitor microcystin-LR (Fig. 5A). The ability ofmicrocystin-LR to offset H2O2 inhibition of Ser-253 phosphor-
ylationwas preserved in the KQ, but not in the KR,mutant (Fig.5A). Given that the KRmutant is predominantly nuclear, thesedata are consistent with the interpretation that PP2A-depen-dent FoxO1 dephosphorylation occurs outside the nucleus.The fact that the effect of microcystin-LR is partial indicatesthat H2O2 also regulates other phosphatases or prevents accessof Akt to FoxO1.The ability of H2O2 to promote FoxO1 nuclear retention was
preempted, but not reversed, by the deacetylase inhibitor nico-tinamide (8). Thus, cell pretreatment with nicotinamideblocked H2O2-induced nuclear translocation, but addition ofnicotinamide after H2O2 treatment was unable to reverse thiseffect (Fig. 5B), indicating that FoxO1 acetylation can preventits nuclear entry but cannot promote its nuclear exclusion.The effect of resveratrol, like that of H2O2, appears to entail
reduced FoxO1 phosphorylation on Ser-253 and Thr-24.Decreased Ser-253 phosphorylation in resveratrol-treated cellswas partly reversed bymicrocystin-LR (Fig. 5C), indicating thatpart of the effect of resveratrol is PP2A-dependent.Because 14-3-3 participates in nucleocytoplasmic shuttling
of FoxO by binding phosphorylated Thr-24 and Ser-253 (37,38) and given the tight relationship between the phosphoryla-tion of these sites and acetylation, we askedwhether acetylation
FIGURE 4. Subcellular localization and phosphorylation of FoxO1 acety-lation site mutants in response to treatment with H2O2 or resveratrol.A, representative images of fluorescence microscopy of HEK-293 cells trans-fected with GFP-tagged WT, KQ, and KR FoxO1 and incubated in the condi-tions indicated. B, immunoblot analysis using anti-phospho-S253 FoxO1(p-S253), phospho-T24 FoxO1 (p-T24), and phospho-T308 Akt (T308) antibod-ies and tubulin. Extracts were prepared from HEK-293 cells transfected as in B.C, Western blot analysis of HEK-293 cells transfected with GFP-tagged WTFoxO1 and incubated with insulin (Ins) and cycloheximide (CHX) prior to theaddition of H2O2 or resveratrol (Resv).
FIGURE 5. Effect of PP2A inhibition on the phosphorylation of FoxO1acetylation site mutants. A, HEK-293 cells transduced with GFP-tagged WT,KQ, and KR adenovirus and treated with H2O2 (1 mM) and microcystin-LR(mc-LR) for 30 min. B, HEK-293 cells transfected with GFP-tagged WT FoxO1and incubated sequentially with H2O2 and nicotinamide or vice versa. C, HEK-293 cells transduced with GFP-tagged WT FoxO1 and exposed to resveratrol(Resv) (50 �M) with and without microcystin-LR.
FoxO1 Acetylation and Phosphorylation
AUGUST 27, 2010 • VOLUME 285 • NUMBER 35 JOURNAL OF BIOLOGICAL CHEMISTRY 27399
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from

affected binding of 14-3-3 to the Ser-253 site. Using immuno-precipitation of FLAG-tagged T24A, T24A-KQ, and T24A-KRmutants, followed by immunoblotting with anti-14-3-3 anti-body, we observed that insulin-induced phosphorylation ofSer-253 was associated with increased binding of T24A to14-3-3 (Fig. 6). Constitutively deacetylated T24A-KR bound14-3-3 more efficiently than T24A (Fig. 6, lanes WT and KRunder �Insulin), even as its phosphorylation on Ser-253 wasreduced. The constitutively acetylated T24A-KQ mutant alsoshowed increased 14-3-3 binding, but, unlike the KRmutant, itwas associated with increased Ser-253 phosphorylation (Fig. 6,compare lanesWT andKQ under�Insulinwith the same lanesunder �Insulin). These data indicate that FoxO1 binding to14-3-3 is alsomodulated by its acetylation state, lending furthersupport to the idea that acetylation affects FoxO1 nucleocyto-plasmic shuttling.Conclusions—The goal of this study was to examine the
reciprocal regulation of two primary posttranslational modifi-cations of FoxO1, acetylation and phosphorylation, and theircombined effects on FoxO1 function. Using constitutivelydeacetylated (KR) and acetylated (KQ) mutants, we show thatacetylation causes a leftward shift in the dose-response curvefor insulin-induced FoxO1 phosphorylation, whereas deacety-lation causes a rightward shift. As a result, the two mutantsdiffer by �10-fold in their insulin sensitivity, suggesting thatacetylation is a major determinant of FoxO1 activity in vivo.A new finding of the present study is that the two modifica-
tions, acetylation and phosphorylation, can be uncoupled fromeach other. Using H2O2 to mimic oxidative stress and induceFoxO1 deacetylation, we show that H2O2 can antagonize insu-lin signaling by promoting either FoxO1 deacetylation ordephosphorylation of Ser-253 and Thr-24, the former in partthrough the serine/threonine phosphatase PP2A. In either case,the expectation is that FoxO1 will be retained in the nucleus.In contrast, and somewhat surprisingly, resveratrol pro-
motes FoxO1 nuclear localization independent of acetylation,as well as of Ser-253 and Thr-24 phosphorylation. These datasupport the recent observation that resveratrol acts by deacety-lation-independent mechanisms (e.g. AMP-activated proteinkinase activation) (39). We cannot exclude the possibility thatresveratrol is unable to induce dephosphorylation of the KQandKRmutants, because of structural alterations caused by thereplacement of acetyllysine with arginine or glutamine.The uncoupling of FoxO1 phosphorylation from acetylation
in H2O2-treated cells has important ramifications for FoxO1
nuclear function. In fact, it has been shown that acetylationaffects FoxO1 affinity to bind its DNA targets (8, 26) and maythus favor DNA binding-independent modalities of FoxO1function, e.g. cell differentiation versus replication and metab-olism (10). Likewise, it has previously been demonstrated thatphosphorylation affects transactivation properties of FoxO1(32), making it theoretically possible that FoxO1 be nuclear andinactive.In conclusion, the findings that phosphorylated FoxO1 can
be retained in the nucleus by decreasing its acetylation and,conversely, that dephosphorylated FoxO1 can be targeted tothe cytoplasm through increased acetylation underscore thatphosphorylation and acetylation are regulated through partlyoverlapping, and partly independent, mechanisms and suggestnew research directions to develop agents that modulate pleio-tropic functions of FoxOs.
Acknowledgment—We thank members of the Accili laboratory forhelpful discussions and advice.
REFERENCES1. Accili, D., and Arden, K. C. (2004) Cell 117, 421–4262. Nakae, J., Biggs, W. H., 3rd, Kitamura, T., Cavenee, W. K., Wright, C. V.,
Arden, K. C., and Accili, D. (2002) Nat. Genet. 32, 245–2533. Nakae, J., Kitamura, T., Silver, D. L., and Accili, D. (2001) J. Clin. Invest.
108, 1359–13674. Buteau, J., and Accili, D. (2007) Diabetes. Obes. Metab. 9, Suppl. 2,
140–1465. Buteau, J., Shlien, A., Foisy, S., and Accili, D. (2007) J. Biol. Chem. 282,
287–2936. Kitamura, T., Kitamura, Y. I., Kobayashi, M., Kikuchi, O., Sasaki, T., De-
pinho, R. A., and Accili, D. (2009)Mol. Cell. Biol. 29, 4417–44307. Kitamura, T., Nakae, J., Kitamura, Y., Kido, Y., Biggs, W. H., 3rd, Wright,
C. V., White, M. F., Arden, K. C., and Accili, D. (2002) J. Clin. Invest. 110,1839–1847
8. Kitamura, Y. I., Kitamura, T., Kruse, J. P., Raum, J. C., Stein, R., Gu,W., andAccili, D. (2005) Cell Metab. 2, 153–163
9. Hribal, M. L., Nakae, J., Kitamura, T., Shutter, J. R., and Accili, D. (2003)J. Cell Biol. 162, 535–541
10. Kitamura, T., Kitamura, Y. I., Funahashi, Y., Shawber, C. J., Castrillon,D. H., Kollipara, R., DePinho, R. A., Kitajewski, J., and Accili, D. (2007)J. Clin. Invest. 117, 2477–2485
11. Sandri,M., Sandri, C., Gilbert, A., Skurk, C., Calabria, E., Picard, A.,Walsh,K., Schiaffino, S., Lecker, S. H., and Goldberg, A. L. (2004) Cell 117,399–412
12. Nakae, J., Kitamura, T., Kitamura, Y., Biggs, W. H., 3rd, Arden, K. C., andAccili, D. (2003) Dev. Cell 4, 119–129
13. Lin, K., Dorman, J. B., Rodan, A., and Kenyon, C. (1997) Science 278,1319–1322
14. Ogg, S., Paradis, S., Gottlieb, S., Patterson, G. I., Lee, L., Tissenbaum,H. A.,and Ruvkun, G. (1997) Nature 389, 994–999
15. Vogt, P. K., Jiang, H., and Aoki, M. (2005) Cell Cycle 4, 908–91316. Biggs,W.H., 3rd,Meisenhelder, J., Hunter, T., Cavenee,W.K., andArden,
K. C. (1999) Proc. Natl. Acad. Sci. U. S. A. 96, 7421–742617. Brunet, A., Bonni, A., Zigmond, M. J., Lin, M. Z., Juo, P., Hu, L. S., Ander-
son, M. J., Arden, K. C., Blenis, J., and Greenberg, M. E. (1999) Cell 96,857–868
18. Nakae, J., Park, B. C., andAccili, D. (1999) J. Biol. Chem. 274, 15982–1598519. Lehtinen, M. K., Yuan, Z., Boag, P. R., Yang, Y., Villen, J., Becker, E. B.,
DiBacco, S., de la Iglesia, N., Gygi, S., Blackwell, T. K., and Bonni, A. (2006)Cell 125, 987–1001
20. Essers, M. A., Weijzen, S., de Vries-Smits, A. M., Saarloos, I., de Ruiter,N. D., Bos, J. L., and Burgering, B. M. (2004) EMBO J. 23, 4802–4812
FIGURE 6. Binding of FoxO1 acetylation site mutants to 14-3-3. HEK-293cells were transfected with FLAG-tagged T24A, T24A-KQ, or T24A-KR FoxO1,treated with insulin, and immunoprecipitated with anti-FLAG antibody (M2)prior to Western blot with the indicated antibodies.
FoxO1 Acetylation and Phosphorylation
27400 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 35 • AUGUST 27, 2010
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from

21. Asada, S., Daitoku, H., Matsuzaki, H., Saito, T., Sudo, T., Mukai, H.,Iwashita, S., Kako, K., Kishi, T., Kasuya, Y., and Fukamizu, A. (2007) CellSignal. 19, 519–527
22. Kawamori, D., Kaneto, H., Nakatani, Y., Matsuoka, T. A., Matsuhisa, M.,Hori, M., and Yamasaki, Y. (2006) J. Biol. Chem. 281, 1091–1098
23. Brunet, A., Park, J., Tran, H., Hu, L. S., Hemmings, B. A., and Greenberg,M. E. (2001)Mol. Cell. Biol. 21, 952–965
24. Nakae, J., Barr, V., and Accili, D. (2000) EMBO J. 19, 989–99625. Nakae, J., Kitamura, T., Ogawa, W., Kasuga, M., and Accili, D. (2001)
Biochemistry 40, 11768–1177626. Daitoku, H., Hatta,M.,Matsuzaki, H., Aratani, S., Ohshima, T.,Miyagishi,
M., Nakajima, T., and Fukamizu, A. (2004) Proc. Natl. Acad. Sci. U. S. A.101, 10042–10047
27. Fukuoka, M., Daitoku, H., Hatta, M., Matsuzaki, H., Umemura, S., andFukamizu, A. (2003) Int. J. Mol. Med. 12, 503–508
28. van der Horst, A., Tertoolen, L. G., de Vries-Smits, L. M., Frye, R. A.,Medema, R. H., and Burgering, B. M. (2004) J. Biol. Chem. 279,28873–28879
29. Brunet, A., Sweeney, L. B., Sturgill, J. F., Chua, K. F., Greer, P. L., Lin, Y.,Tran,H., Ross, S. E.,Mostoslavsky, R., Cohen,H. Y., Hu, L. S., Cheng,H. L.,Jedrychowski, M. P., Gygi, S. P., Sinclair, D. A., Alt, F. W., and Greenberg,M. E. (2004) Science 303, 2011–2015
30. Frescas, D., Valenti, L., and Accili, D. (2005) J. Biol. Chem. 280,20589–20595
31. Imai, S., Armstrong, C.M., Kaeberlein,M., andGuarente, L. (2000)Nature403, 795–800
32. Tomizawa, M., Kumar, A., Perrot, V., Nakae, J., Accili, D., Rechler, M. M.,and Kumaro, A. (2000) J. Biol. Chem. 275, 7289–7295
33. Logie, L., Ruiz-Alcaraz, A. J., Keane,M.,Woods, Y. L., Bain, J.,Marquez, R.,Alessi, D. R., and Sutherland, C. (2007) Diabetes 56, 2218–2227
34. Lagouge, M., Argmann, C., Gerhart-Hines, Z., Meziane, H., Lerin, C.,Daussin, F., Messadeq, N., Milne, J., Lambert, P., Elliott, P., Geny, B.,Laakso, M., Puigserver, P., and Auwerx, J. (2006) Cell 127, 1109–1122
35. Nemoto, S., and Finkel, T. (2002) Science 295, 2450–245236. Yan, L., Lavin, V. A., Moser, L. R., Cui, Q., Kanies, C., and Yang, E. (2008)
J. Biol. Chem. 283, 7411–742037. Obsil, T., Ghirlando, R., Anderson, D. E., Hickman, A. B., and Dyda, F.
(2003) Biochemistry 42, 15264–1527238. Rena, G., Prescott, A. R., Guo, S., Cohen, P., and Unterman, T. G. (2001)
Biochem. J. 354, 605–61239. Canto, C., Gerhart-Hines, Z., Feige, J. N., Lagouge, M., Noriega, L., Milne,
J. C., Elliott, P. J., Puigserver, P., and Auwerx, J. (2009) Nature 458,1056–1060
FoxO1 Acetylation and Phosphorylation
AUGUST 27, 2010 • VOLUME 285 • NUMBER 35 JOURNAL OF BIOLOGICAL CHEMISTRY 27401
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Li Qiang, Alexander S. Banks and Domenico AcciliIndependent of Its Subcellular Localization
Uncoupling of Acetylation from Phosphorylation Regulates FoxO1 Function
doi: 10.1074/jbc.M110.140228 originally published online June 2, 20102010, 285:27396-27401.J. Biol. Chem.
10.1074/jbc.M110.140228Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/285/35/27396.full.html#ref-list-1
This article cites 39 references, 18 of which can be accessed free at
by guest on April 18, 2016
http://ww
w.jbc.org/
Dow
nloaded from
Related Documents











