Ulva diversity in the Yellow Sea during the large-scale green algal blooms in 2008–2009Feng Liu, 1,2 Shao Jun Pang, 1 * Na Xu, 1,2 Ti Feng Shan, 1 Song Sun, 3 Xiang Hu 4 and Jian Qiang Yang 4 1 Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, 2 Graduate University of Chinese Academy of Sciences, Beijing 100049, 3 Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, and 4 North China Sea Branch of the State Oceanic Administration, Qingdao 266033, China SUMMARY In the Yellow Sea of China, large-scale green tides have broken out for three consecutive years from 2007 to 2009. As part of the efforts to localize the algal source, two cruises were conducted in the early stage and the outbreak stage of the bloom in 2009. We analyzed the morphological and genetic diversity of drifting Ulva specimens and culture-derived isolates from seawater sampled in different localities. For phylogenetic analy- ses, the nuclear encoded ribosomal DNA internal tran- scribed spacer region (ITS nrDNA) and the plastid encoded large subunit of ribulose-1, 5-bisphosphate carboxylase/oxgenase gene (rbcL) were used. Our molecular and morphological data indicate that the dominant free-floating Ulva species in 2008 and 2009 possibly belonged to a single strain of the U. linza- procera-prolifera (LPP) clade. The ITS sequences from bloom-forming algal samples with dense branches were identical to those from U. linza-like specimens without branches derived from the Yellow Sea. Microscopic individuals of the dominant Ulva strain were detected in eight stations, revealing that spore dispersal in the water helped to enlarge biomass in the water during the outbreak stage of green tide in the Yellow Sea. Key words: gametes, green tides, phylogeny, the Yellow Sea, Ulva prolifera. INTRODUCTION In the Yellow Sea of China, large-scale green algal blooms have occurred for three consecutive years from 2007 to 2009 (Jiang et al. 2008; Sun et al. 2008). The dominant blooming algal species in 2007 and 2008 was identified to belong to the Ulva linza-procera- prolifera (LPP) clade based on molecular data (Hayden et al. 2003; Leliaert et al. 2008, 2009; Wang et al. 2010; Liu et al. 2010a). According to satellite images from 2008, the green tide originally accumulated near the coasts of Jiangsu province and was sequentially transported north into the Yellow Sea and then onshore northwest to Qingdao coasts by seasonal winds and surface currents (Sun et al. 2008; Liu et al. 2009). It was reported that the green patches had grown rapidly and covered from 80 km 2 impacting 1000 km 2 on 15 May to 1200 km 2 impacting 40 000 km 2 on 31 May in 2008 (Liu et al. 2009). The surprisingly efficient expansion of the drifting bloom remained unidentified although models by various investigators have been proposed, including vegetative and reproductive propa- gation while drifting (Sun et al. 2008; Ye et al. 2008; Zhang et al. 2009). It is widely accepted that localiza- tion of the bloom algal source and the persistence of the bloom are instrumental in understanding the blooming mechanism and are a prerequisite for man- agement decisions (Blomster et al. 2002). Many Ulva species are highly tolerant of variable salinity, temperature, irradiance and nutrient concen- trations (Tan et al. 1999; Dan et al. 2002; Cohen & Fong 2006; Conley et al. 2009). Species in the genus show great intraspecific morphological and cytological variations at different growth phases, in different seasons and environmental conditions (Blomster et al. 1998, 1999). This plasticity often leads to difficulties in species identification of individual thalli based on morphological and cytological characters (Blomster et al. 2002; Leskinen et al. 2004). Therefore, molecu- lar methods are essential for reliable identification of Ulva species. The nuclear-encoded ribosomal DNA internal transcribed spacer region (ITS nrDNA) and the plastid-encoded large subunit of ribulose-1, 5-bisphosphate carboxylase/ oxgenase (rbcL) gene have been commonly used for phylogenetic reconstruction in Ulva and delimitation of its species (Blomster et al. 1998; Tan et al. 1999; Shimada et al. 2003, 2008; Leskinen et al. 2004). *To whom correspondence should be addressed. Email: [email protected] Communicating editor: I. Mine. Received 10 September 2009; accepted 12 May 2010. doi: 10.1111/j.1440-1835.2010.00586.x Phycological Research 2010; 58: 270–279 © 2010 Japanese Society of Phycology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ulva diversity in the Yellow Sea during the large-scale greenalgal blooms in 2008–2009pre_586 270..279
Feng Liu,1,2 Shao Jun Pang,1* Na Xu,1,2 Ti Feng Shan,1 Song Sun,3 Xiang Hu4 and Jian Qiang Yang4
1Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao266071, 2Graduate University of Chinese Academy of Sciences, Beijing 100049, 3Key Laboratory of Marine Ecologyand Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, and 4NorthChina Sea Branch of the State Oceanic Administration, Qingdao 266033, China
SUMMARYIn the Yellow Sea of China, large-scale green tides havebroken out for three consecutive years from 2007 to2009. As part of the efforts to localize the algal source,two cruises were conducted in the early stage and theoutbreak stage of the bloom in 2009. We analyzed themorphological and genetic diversity of drifting Ulvaspecimens and culture-derived isolates from seawatersampled in different localities. For phylogenetic analy-ses, the nuclear encoded ribosomal DNA internal tran-scribed spacer region (ITS nrDNA) and the plastidencoded large subunit of ribulose-1, 5-bisphosphatecarboxylase/oxgenase gene (rbcL) were used. Ourmolecular and morphological data indicate that thedominant free-floating Ulva species in 2008 and 2009possibly belonged to a single strain of the U. linza-procera-prolifera (LPP) clade. The ITS sequences frombloom-forming algal samples with dense branches wereidentical to those from U. linza-like specimens withoutbranches derived from the Yellow Sea. Microscopicindividuals of the dominant Ulva strain were detected ineight stations, revealing that spore dispersal in thewater helped to enlarge biomass in the water during theoutbreak stage of green tide in the Yellow Sea.
Key words: gametes, green tides, phylogeny, the YellowSea, Ulva prolifera.
INTRODUCTION
In the Yellow Sea of China, large-scale green algalblooms have occurred for three consecutive years from2007 to 2009 (Jiang et al. 2008; Sun et al. 2008).The dominant blooming algal species in 2007 and2008 was identified to belong to the Ulva linza-procera-prolifera (LPP) clade based on molecular data (Haydenet al. 2003; Leliaert et al. 2008, 2009; Wang et al.2010; Liu et al. 2010a). According to satellite imagesfrom 2008, the green tide originally accumulated nearthe coasts of Jiangsu province and was sequentially
transported north into the Yellow Sea and then onshorenorthwest to Qingdao coasts by seasonal winds andsurface currents (Sun et al. 2008; Liu et al. 2009). Itwas reported that the green patches had grown rapidlyand covered from 80 km2 impacting 1000 km2 on 15May to 1200 km2 impacting 40 000 km2 on 31 May in2008 (Liu et al. 2009). The surprisingly efficientexpansion of the drifting bloom remained unidentifiedalthough models by various investigators have beenproposed, including vegetative and reproductive propa-gation while drifting (Sun et al. 2008; Ye et al. 2008;Zhang et al. 2009). It is widely accepted that localiza-tion of the bloom algal source and the persistence ofthe bloom are instrumental in understanding theblooming mechanism and are a prerequisite for man-agement decisions (Blomster et al. 2002).
Many Ulva species are highly tolerant of variablesalinity, temperature, irradiance and nutrient concen-trations (Tan et al. 1999; Dan et al. 2002; Cohen &Fong 2006; Conley et al. 2009). Species in the genusshow great intraspecific morphological and cytologicalvariations at different growth phases, in differentseasons and environmental conditions (Blomster et al.1998, 1999). This plasticity often leads to difficultiesin species identification of individual thalli based onmorphological and cytological characters (Blomsteret al. 2002; Leskinen et al. 2004). Therefore, molecu-lar methods are essential for reliable identification ofUlva species. The nuclear-encoded ribosomal DNAinternal transcribed spacer region (ITS nrDNA) andthe plastid-encoded large subunit of ribulose-1,5-bisphosphate carboxylase/ oxgenase (rbcL) gene havebeen commonly used for phylogenetic reconstruction inUlva and delimitation of its species (Blomster et al.1998; Tan et al. 1999; Shimada et al. 2003, 2008;Leskinen et al. 2004).
*To whom correspondence should be addressed.Email: [email protected] editor: I. Mine.Received 10 September 2009; accepted 12 May 2010.doi: 10.1111/j.1440-1835.2010.00586.x
Phycological Research 2010; 58: 270–279
© 2010 Japanese Society of Phycology

In 2009, when the large-scale green tide reoccurredin the Yellow Sea, two cruises were conducted in theearly stage and the outbreak stage. We present resultson the morphological and molecular diversity of Ulvasamples taken from latitude 32–36°, as well as culture-derived Ulva samples from surface water during CruiseB. In addition, vegetative growth and spore dispersal ofthe dominant algal strain were tested under laboratoryconditions in order to study growth rate and mode ofreproduction.
MATERIALS AND METHODS
Grid station settings of the two cruises
Cruise A was conducted on the R/V Beidou of the YellowSea Fisheries Research Institute (YSFRI) from 15 Mayto 20 May in 2009 (Fig. 1a). Cruise B on the R/V XiangYang Hong no. 8 of North China Sea Branch of the StateOceanic Administration was conducted from 21 June to30 June in 2009 (Fig. 1b). These cruises surveyed thesouth-west coast (32–36 °N, 120–123 °E) in grid sta-
tions. Stations were named as A1 to A25 in Cruise Aand B1 to B18 in Cruise B.
Sampling and quantification of the Ulvamicroscopic stages in seawater
Free-floating Ulva samples were collected on board thevessels at different stations in the Yellow Sea. Aftercollection, the algae were cleaned, photographed andfurther cultured in the Provasoli Enriched Seawatermedium (PES) (Berges et al. 2001) in an artificialclimate incubator (Jiangnan, China) at 15–18°C under80–100 mmol photons m-2 s-1 in a 12:12 h light : dark(LD) regime. In total, 20 free-floating Ulva sampleswere selected in the two cruises for further analysis,including seven samples (A11, A12, A14, A15, A16,A17 and A19) from Cruise A, and 13 samples (B4, B5,B7, B8, B9, B10, B11, B12, B13, B14, B15, B16 andB18) from Cruise B, representing the drifting algaesampled in 2009 during the cruises. The samples werenamed after the corresponding stations (Table 1). Eightunialgal Ulva strains (2008 a, b, c, d, e, f, g and h),
Fig. 1. Maps of collection sites in Yellow Sea: ‘a’ and ‘b’ indicate Cruise A on the R/V Beidou from 15 to 20 May 2009 and Cruise B on
the R/V Xiang Yang Hong no. 08 from 21 to 30 June 2009, respectively. The numbers stand for the stations. Filled and empty circles
represented the stations with and without free-floating Ulva biomass, respectively.
© 2010 Japanese Society of Phycology
271Ulva green tide in the Yellow Sea

originally harvested from Tuandao bay and Qingdao No.1 bathing beach in June 2008, representing the algalspecies of the 2008 green tide, were together analyzedin the following phylogenetic experiments.
One-liter seawater samples were collected at differ-ent stations in Cruise B and kept in the incubator underthe above conditions on the vessels. After the cruise,the samples were then transferred to the laboratory, andaerated after adding NO3
- (823 mmol/L) and PO43-
(73 mmol/L) to reach to the PES levels under the sameculture conditions (Pang et al. 2010). Ulva microscopicstages in seawater were quantified using the methoddescribed by Liu et al. (2010b). After 6–7 weeks, allUlva strains derived from the water samples were exam-ined and analyzed microscopically. Three distinctgroups were determined according to their basic mor-phological characters including thallus morphology andthe degree of branching (Table 1).
DNA extraction, ITS nrDNA and rbcL geneamplification and sequencing
The 45 Ulva samples were washed three times withsterilized seawater and dried using filter paper. Then100 mg of material for each sample was ground to a finepowder in liquid nitrogen and transferred to the 2 mLtube. DNA was extracted using the cetyltrimethylammo-nium bromide (CTAB) method (Wang et al. 2006). Theconcentration and the quality of isolated DNA wereassessed by electrophoresis on 1.0% agarose gel. DNAconcentration of each sample was adjusted to 50 ng/mL.ITS and rbcL gene sequences were amplified andsequenced using primers published in Leskinen andPamilo (1997) and Hayden et al. (2003). Total genomicDNA (30–40 ng) was added to 50 mL polymerase chainreaction (PCR) reactions containing final concentrationsof 1 ¥ PCR buffer (Takara, Dalian, China), 2 mM MgCl2
(Takara), 0.8 mM dNTPs (Takara), 25 mM of eachprimer, 1.6 U Taq Polymerase (Takara). Amplificationproducts were separated by 1.0% agarose gel electro-phoresis and fragments of an expected length were cutfrom the gel and purified using a DNA Gel Extraction Kit(Bio Basic Inc., Markham, Canada) according to themanufacturer’s instructions. Sequencing reactions wereperformed using ABI 3730 XL automated sequencers byShanghai Biosune Biotechnology Co., Ltd. and eachsequencing reaction was repeated twice.
Phylogenetic analysis
Internal transcribed spacer and rbcL sequences wereobtained from 45 samples of free-floating thalli andculture derived isolates from seawater. Sequences werealigned using Clustal W (Thompson et al. 1994).Neighbor-joining (NJ) and maximum parsimony (MP)analyses were performed with Mega 4.0 (Tamura et al.2007). Ulvaria fusca and Umbraulva amamiensis wereselected as outgroup taxa. The reliability of brancheswas evaluated with non-parametric bootstrapping(1000 replicates) (Felsenstein 1985). The evolutionarydistances of the NJ tree were computed using theKimura 2-parameter method (Kimura 1980). TheMP tree was obtained using the Close-Neighbor-Interchange algorithm (Nei & Kumar 2000) with searchlevel 3 in which the initial trees were obtained with therandom addition of sequences. All positions containinggaps and missing data were eliminated from the dataset(complete deletion option).
Determination of vegetative growth andspore dispersal under laboratory conditions
A tube culture system was designed to test thegrowth rates and spore dispersal of the dominant Ulva
Table 1. Description of different Ulva groups which were free-floating or derived from the seawater
Group Samples Collectiondate
Source and collection sites Gross morphologicalcharacteristics
1 2008a, b, c, d, e, f, g and h June 2008 Free-floating, Tuandao Bay andQingdao First Beach
Filiform and tubular, monostromatic,dense branches
2 A11, A12, A14, A15, A16, A17and A19
May 2009 Free-floating, Yellow Sea (Fig. 1a) Filiform and tubular, monostromatic,dense branches (Fig. 3a)
3 B4, B5, B7, B8, B9, B10, B11, B12,B13, B14, B15, B16 and B18
June 2009 Free-floating, Yellow Sea (Fig. 1b) Filiform and tubular, monostromatic,dense branches (Fig. 3b–d)
4 B4a, B7a, B8a, B11a, B14a andB16a
July 2009 Culture-derived from the YellowSea seawater (Fig. 1b)
Filiform and tubular, monostromatic,dense branches (Fig. 3e)
5 B4b, B6b, B8b, B10b, B11b, B13b,B14b, B15b and B16b
Filiform and tubular, monostromatic,branches (Fig. 3f,g)
6 B1c and B8c Ribbon, monostromatic, no branch(Fig. 3h)
The names of Ulva samples were corresponding to the station names of the two cruises. In the ‘Samples’ column, ‘a’ and ‘b’ indicateCruise A and Cruise B, respectively, and the number stands for the station of the cruises (Fig. 1).
© 2010 Japanese Society of Phycology
272 F. Liu et al.

strain (Fig. 2). A fine algal filament (approximately0.1–0.2 g) was fixed on a 4 mm glass tube inside aglass cylinder (A) (2.5 cm in diameter), which wasexposed to fluorescent white light to guarantee eachpart of the algal filaments was exposed to identicalirradiance level. A pump transported sterilized,enriched seawater through the tube to the alga andthe flow-rate of the water pumped through the glasstube was about 6–8 L h-1. The entire culture water(PES) was 4 L and renewed weekly. The culture wasrun in ambient temperature (18–21°C). For eachtesting irradiance (230 and 490 mmol photonsm-2 s-1), four identical culture systems were per-formed for each treatment. Light was measured usinga spherical photometer (Biospherical, San Diego, CA,USA). Fresh weight of initial and ending biomass ofthe filaments were evaluated. Relative growth rate(RGR) was calculated as RGR = (ln W2 – ln W1)/t,where W1 are the initial fresh weights and W2 thefresh weight after t days. Total numbers of dischargedspores from the fixed filaments during the cultureperiod were based on counting the young germlingsattached on the tank (B) wall after 11 days of cultureand values were averaged.
RESULTS
Field observations and morphology offree-floating Ulva
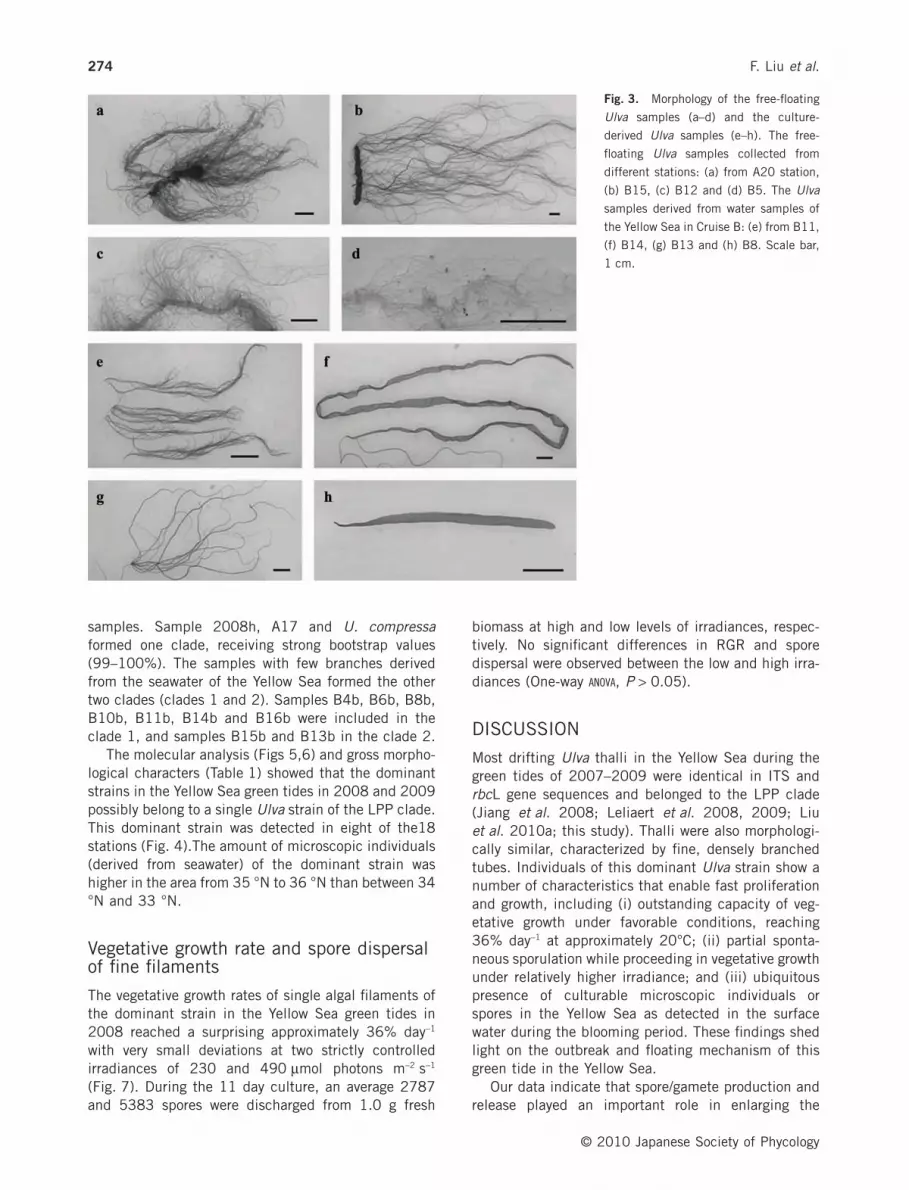
In Cruise A, free-floating Ulva thalli were found in eightout of the 25 stations. Small patches of free-floatingUlva were spotted in the area of 33.0–34.5 °N and121.0–122.5 °E (Fig. 1a). In Cruise B, large patches(approximately 2–3 km2) of free-floating Ulva wereobserved near station B8. Large-scale free-floating Ulvabiomass reached 36°N (Fig. 1b). All sampled free-floating Ulva thalli were characterized by denselybranched, narrow tubes. The main tube was filled withoxygen as a result of photosynthesis and thus kept thebiomass floating on water surface (Fig. 3a–d).Sequences were obtained from 20 free-floating Ulvasamples (Table 1).
Culture-derived Ulva
The number of culture-derived Ulva isolates from 1 Lsurface seawater differed markedly among the grid sta-tions (Fig. 4a). Ulva individuals were not detected intwo stations only (B9 and B18). In eight stations (B4,B7, B8, B10, B11, B12, B16 and B17), Ulva thalliwith densely branched, narrow tubes were detected(Fig. 4b). Ulva strains derived from the seawater werecategorized into three groups according to the degree ofbranching and thallus gross morphology (Fig. 3e–h).Seventeen representatives from the three morphologicalgroups were selected for molecular analysis, includingsamples B4a, B7a, B8a, B11a, B14a and B16a ingroup 4, samples B4b, B6b, B8b, B10b, B11b, B13b,B14b, B15b and B16b) in group 5 and samples B1cand B8c in group 6 (Table 1).
Phylogenetic analysis
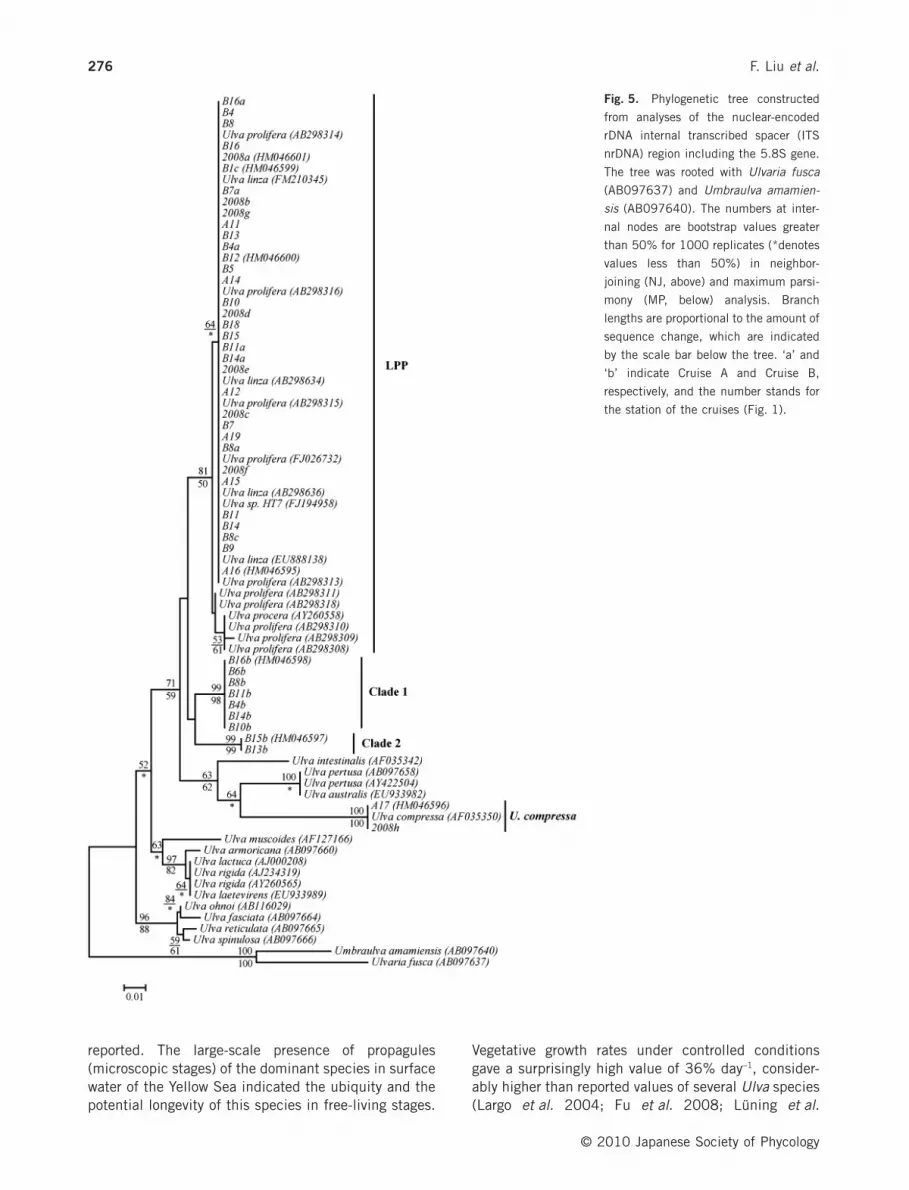
The ITS and rbcL datasets included 45 Ulva samplescombined with additional taxa from GenBank, and were560 and 1298 characters in length, respectively. TheITS1 ranged between 188 and 201 bp, and the ITS2from 177 to 187 bp. The phylogenetic analyses basedon the two datasets showed that all 45 samples fromthis study fell into four clades with high bootstrapsupport: the U. linza-procera-prolifera (LPP) clade (34samples), clade 1 (seven samples), clade 2 (twosamples) and the U. compressa clade (two samples)(Figs 5,6). The LPP clade contained all the free-floatingsamples (26 samples) except samples 2008h and A17,the Ulva samples with dense branches (six samples)and unbranched samples (two samples) derived fromthe water samples. The ITS sequences of the twounbranched U. linza-like samples, B1c and B8c, wereidentical to those of the dominant free-floating
Fig. 2. Schematic drawing to show the tube culture system to
determine the relative growth rate (RGR) and spore dispersal of
unialgal filaments. In such a system, ‘a’ refers to the glass cylin-
der in which a fine filament of Ulva sp. was attached on a glass
tube. An underwater pump transported the water from the tank ‘b’
to ‘a’. The water was recirculated. Fluorescent white light was
shining through the tube evenly on each ramification of the alga.
Culture-derived sporelings were determined after 11 day culture
from the tank wall in ‘b’. The system was run at ambient tem-
peratures (18–21°C). Water was renewed weekly.
© 2010 Japanese Society of Phycology
273Ulva green tide in the Yellow Sea
samples. Sample 2008h, A17 and U. compressaformed one clade, receiving strong bootstrap values(99–100%). The samples with few branches derivedfrom the seawater of the Yellow Sea formed the othertwo clades (clades 1 and 2). Samples B4b, B6b, B8b,B10b, B11b, B14b and B16b were included in theclade 1, and samples B15b and B13b in the clade 2.
The molecular analysis (Figs 5,6) and gross morpho-logical characters (Table 1) showed that the dominantstrains in the Yellow Sea green tides in 2008 and 2009possibly belong to a single Ulva strain of the LPP clade.This dominant strain was detected in eight of the18stations (Fig. 4).The amount of microscopic individuals(derived from seawater) of the dominant strain washigher in the area from 35 °N to 36 °N than between 34°N and 33 °N.
Vegetative growth rate and spore dispersalof fine filaments
The vegetative growth rates of single algal filaments ofthe dominant strain in the Yellow Sea green tides in2008 reached a surprising approximately 36% day-1
with very small deviations at two strictly controlledirradiances of 230 and 490 mmol photons m-2 s-1
(Fig. 7). During the 11 day culture, an average 2787and 5383 spores were discharged from 1.0 g fresh
biomass at high and low levels of irradiances, respec-tively. No significant differences in RGR and sporedispersal were observed between the low and high irra-diances (One-way ANOVA, P > 0.05).
DISCUSSION
Most drifting Ulva thalli in the Yellow Sea during thegreen tides of 2007–2009 were identical in ITS andrbcL gene sequences and belonged to the LPP clade(Jiang et al. 2008; Leliaert et al. 2008, 2009; Liuet al. 2010a; this study). Thalli were also morphologi-cally similar, characterized by fine, densely branchedtubes. Individuals of this dominant Ulva strain show anumber of characteristics that enable fast proliferationand growth, including (i) outstanding capacity of veg-etative growth under favorable conditions, reaching36% day-1 at approximately 20°C; (ii) partial sponta-neous sporulation while proceeding in vegetative growthunder relatively higher irradiance; and (iii) ubiquitouspresence of culturable microscopic individuals orspores in the Yellow Sea as detected in the surfacewater during the blooming period. These findings shedlight on the outbreak and floating mechanism of thisgreen tide in the Yellow Sea.
Our data indicate that spore/gamete production andrelease played an important role in enlarging the
Fig. 3. Morphology of the free-floating
Ulva samples (a–d) and the culture-
derived Ulva samples (e–h). The free-
floating Ulva samples collected from
different stations: (a) from A20 station,
(b) B15, (c) B12 and (d) B5. The Ulva
samples derived from water samples of
the Yellow Sea in Cruise B: (e) from B11,
(f) B14, (g) B13 and (h) B8. Scale bar,
1 cm.
© 2010 Japanese Society of Phycology
274 F. Liu et al.

biomass of the dominant strain during the outbreakstage of the green tide in the Yellow Sea (Fig. 4). Aremarkable feature of the dominant bloom-forming Ulvastrain was its spontaneous, partial sporulation that wasobserved in laboratory cultures. Partial sporulationrefers to a small portion of the vegetative thalli in thefilaments forming spores or gametes that were dis-charged spontaneously while the vegetative growth ofthe thalli still maintains. Most of the species in thegenus of Ulva are characterized by apical sporulation(Smith 1947; Lüning et al. 2008). In our cultures,free-swimming gametes could be detected nearly at anytime while the thalli remained intact, in contrast withthe disappearance of the thalli after sporulations in theapical sporulated species. This feature gives the domi-nant Ulva strain the advantage of longevity of thalli thatcontinuously discharge reproduction cells while driftingand in the meantime maintaining fast vegetativegrowth.
Previous studies have already pointed out that thedominant blooming alga in the Yellow Sea likelybelongs to a single Ulva species. Jiang et al. (2008)reported that the drifting alga in the green tide along
Qingdao coast in the summer of 2007 was composed ofa single strain based on ITS sequence data. Leliaertet al. (2008, 2009) and Liu et al. (2010a) found thatthe dominant Ulva in the Yellow Sea during the 2008bloom belonged to the Ulva linza-procera-prolifera(LPP) clade as circumscribed by Shimada et al.(2008). It is surprising that such a large-scale greentide was composed of a single taxon, and possibly asingle strain for three consecutive years, in each eventover the past two years covering more than thousands ofsquare kilometers in the Yellow Sea. Liu et al. (2010a)showed evidence that this dominant strain was notdetected in the coastal waters of Qingdao in the winterfollowing the 2008 bloom, indicating that this domi-nant strain was not native to the coasts of Qingdao.These observations bring up an apparent and pertinentquestion: have these drifting green-tide algae originatedfrom one location/region/niche in three consecutiveyears?
Species in Ulva are normal components of benthicseaweed populations (Tseng et al. 1962). Detection ofmicroscopic stages derived from spores of seaweeds infree surface water in the open oceans is not regularly
Fig. 4. Number range of culture-derived Ulva strains from 1-L of surface seawater samples from the Yellow Sea collected in Cruise B (a),
and number range of culture-derived dominant strains of the Yellow Sea green tide from 1 L surface seawater samples in Cruise B (b), as
determined by both morphology and two DNA sequences (ITS and rbcL).
© 2010 Japanese Society of Phycology
275Ulva green tide in the Yellow Sea
reported. The large-scale presence of propagules(microscopic stages) of the dominant species in surfacewater of the Yellow Sea indicated the ubiquity and thepotential longevity of this species in free-living stages.
Vegetative growth rates under controlled conditionsgave a surprisingly high value of 36% day-1, consider-ably higher than reported values of several Ulva species(Largo et al. 2004; Fu et al. 2008; Lüning et al.
Fig. 5. Phylogenetic tree constructed
from analyses of the nuclear-encoded
rDNA internal transcribed spacer (ITS
nrDNA) region including the 5.8S gene.
The tree was rooted with Ulvaria fusca
(AB097637) and Umbraulva amamien-
sis (AB097640). The numbers at inter-
nal nodes are bootstrap values greater
than 50% for 1000 replicates (*denotes
values less than 50%) in neighbor-
joining (NJ, above) and maximum parsi-
mony (MP, below) analysis. Branch
lengths are proportional to the amount of
sequence change, which are indicated
by the scale bar below the tree. ‘a’ and
‘b’ indicate Cruise A and Cruise B,
respectively, and the number stands for
the station of the cruises (Fig. 1).
© 2010 Japanese Society of Phycology
276 F. Liu et al.

Fig. 6. Phylogenetic tree constructed
from an analysis of rbcL gene sequences.
The tree was rooted with Ulvaria fusca
(AB097611) and Umbraulva amamien-
sis (AB097614). The numbers at inter-
nal nodes were bootstrap values greater
than 50% for 1000 replicates (*denotes
values less than 50%) in neighbor-
joining (NJ, above) and maximum parsi-
mony (MP, below) analysis. Branch
lengths are proportional to the amount of
sequence change, which are indicated
by the scale bar below the tree. ‘a’ and
‘b’ indicate Cruise A and Cruise B,
respectively, and the number stands for
the station of the cruises (Fig. 1).
© 2010 Japanese Society of Phycology
277Ulva green tide in the Yellow Sea

2008). This high growth rate and reproductive potentialmay be the key to the success of this particular Ulvastrain in occupying the nutrient-rich niches in theYellow Sea.
Worldwide, the occurrence of green tides has beenattributed to high nutrient levels in the water (Charlieret al. 2007; Leliaert et al. 2009). The coasts where theinitial floating biomass of Ulva was spotted are charac-terized by extensive mariculture of animals (crabs,shrimps and clams) which results in elevated levels ofN and P in the nearshore coastal waters of Jiangsuprovince (Pang et al. 2010). Rich nutrients and wideabundance of Ulva microscopic stages in the coastalwaters, in combination with elevated temperature andirradiance in early spring is attributed to the occurrenceof green tide in 2007–2009, and potentially in thefuture to come, in that region in the Yellow Sea.
ACKNOWLEDGMENTS
The authors thank the North China Sea Branch of theState Oceanic Administration and Yellow Sea FisheriesResearch Institute (YSFRI) for organizing the cruises,and Zong Ling Wang, Tie Zhu Mi, Zong Ying Liang andPing Dai for their assistance in the collection of waterand algal samples on the vessel, and Shanghai BiosuneBiotechnology Co., Ltd for its technical assistancein sequencing. Financial support came from (i) theMinistry of Science and Technology of China(2008BAC49B01); and (ii) a project on green tideissued by the State Oceanic Administration (SOA).
REFERENCES
Berges, J. A., Franklin, D. J. and Harrison, P. J. 2001. Evo-lution of an artificial seawater medium: improvements in
enriched seawater, artificial water over the last twodecades. J. Phycol. 37: 1138–45.
Blomster, J., Maggs, C. A. and Stanhope, M. J. 1998. Molecu-lar and morphological analysis of Enteromorpha intestina-lis and E. compressa (Chlorophyta) in the British Isles.J. Phycol. 34: 319–40.
Blomster, J., Maggs, C. A. and Stanhope, M. J. 1999. Exten-sive intraspecific morphological variation in Enteromorphamuscoides (Chlorophyta) revealed by molecular analysis.J. Phycol. 35: 575–86.
Blomster, J., Bäck, S., Fewer, D. P. et al. 2002. Novel mor-phology in Enteromorpha (Ulvophyceae) forming greentides. Am. J. Bot. 89: 1756–63.
Charlier, R. H., Morand, P., Finkl, C. W. and Thys, A. 2007.Green tides on the Brittany coasts. Environ. Res. Eng.Manag. 3: 52–9.
Cohen, R. A. and Fong, P. 2006. Using opportunistic greenmacroalgae as indicators of nitrogen supply and sources toestuaries. Ecol. Appl. 16: 1405–20.
Conley, D. J., Paerl, H. W., Howarth, R. W. et al. 2009.Controlling eutrophication: nitrogen and phosphorus.Science 323: 1014–5.
Dan, A., Hiraoka, M., Ohno, M. and Critchley, A. T. 2002.Observations on the effect of salinity and photon fluencerate on the induction of sporulation and rhizoid formationin the green alga Enteromorpha prolifera (Müller) J.Agardh(Chlorophyta, Ulvales). Fish. Sci. 68: 1182–8.
Felsenstein, J. 1985. Confidence limits on phylogenies:An approach using the bootstrap. Evolution 39: 783–91.
Fu, G., Yao, J. T., Liu, F. L. et al. 2008. Effect of temperatureand irradiance on the growth and reproduction of Entero-morpha prolifera J. Ag. (Chlorophycophyta, Chloro-phyceae). Chin. J. Oceanol. Limnol. 26: 357–62.
Hayden, H. S., Blomster, J., Maggs, C. A., Silva, P. C., Stan-hope, M. J. and Waaland, J. R. 2003. Linnaeus was rightall along: Ulva and Enteromorpha are not distinct genera.Eur. J. Phycol. 38: 277–94.
Jiang, P., Wang, J. F., Cui, Y. L., Li, Y. X., Lin, H. Z. and Qin,S. 2008. Molecular phylogenetic analysis of attachedUlvaceae species and free-floating Enteromorpha fromQingdao coasts in 2007. Chin. J. Oceanol. Limnol. 26:276–9.
Kimura, M. 1980. A simple method for estimatingevolutionary rate of base substitutions through comparativestudies of nucleotide sequences. J. Mol. Evol. 16: 111–20.
Largo, D. B., Sembrano, J., Hiraoka, M. and Ohno, M. 2004.Taxonomic and ecological profile of ‘green tides’ species ofUlva (Ulvales, Chlorophyta) in central Philippines. Hydro-biologia 512: 247–53.
Leliaert, F., Malta, E. J., Engelen, A. H., Mineur, F. and DeClerck, O. 2008. Quindao algal bloom culprit identified.Mar. Pollut. Bull. 56: 1516.
Leliaert, F., Zhang, X., Ye, N. et al. 2009. Identity of theQingdao algal bloom. Phycol. Res. 57: 147–51.
Fig. 7. Relative growth rate (RGR, in percentage day-1, left) and
spore dispersal during the 11 day culture period (in spores g-1,
right) of the unialgal filaments in the tube test system under two
irradiance levels (230 and 490 mmol photons m-2 s-1).
© 2010 Japanese Society of Phycology
278 F. Liu et al.

Leskinen, E. and Pamilo, P. 1997. Evolution of the ITSsequences of ribosomal DNA in Enteromorpha (Chloro-phyceae). Hereditas 126: 17–23.
Leskinen, E., Alstrom-Rapaport, C. and Pamilo, P. 2004.Phylogeographical structure, distribution and geneticvariation of the green algae Ulva intestinalis and U. com-pressa (Chlorophyta) in the Baltic Sea area. Mol. Ecol. 13:2257–65.
Liu, D. Y., Keesing, J. K., Xing, Q. G. and Shi, P. 2009. World’slargest macroalgal bloom caused by expansion of seaweedaquaculture in China. Mar. Pollut. Bull. 58: 888–95.
Liu, F., Pang, S. J., Chopin, T. et al. 2010a. The dominantUlva strain of the 2008 green algal bloom in the Yellow Seawas not detected in the coastal waters of Qingdao in thefollowing winter. J. Appl. Phycol. DOI:10.1007/s10811-009-9489-7.
Liu, F., Pang, S. J., Shan, T. F., Xu, N., Gao, S. Q. and Sun,S. 2010b. A novel method to quantify the microscopicstages of Ulva species in seawater and its applications ingreen tide of the Yellow Sea. Chin. Sci. Bull. 55: 466–73(in Chinese with English abstract).
Lüning, K., Kadel, P. and Pang, S. J. 2008. Control of repro-duction rhythmicity by environmental and endogenoussignals in Ulva pseudocurvata (Chlorophyta). J. Phycol. 44:866–73.
Nei, M. and Kumar, S. 2000. Molecular Evolution and Phy-logenetics. Oxford University Press, New York.
Pang, S. J., Liu, F., Shan, T. F. et al. 2010. Tracking the algalorigin of the Ulva bloom in the Yellow Sea by a combinationof molecular, morphological and physiological analyses.Mar. Environ. Res. 69: 207–15.
Shimada, S., Hiraoka, M., Nabata, S., Lima, M. and Masuda,M. 2003. Molecular phylogenetic analyses of the JapaneseUlva and Enteromorpha (Ulvales, Ulvophyceae), withspecial reference to the free-floating Ulva. Phycol. Res. 51:99–108.
Shimada, S., Yokoyama, N., Arai, S. and Hiraoka, M. 2008.Phylogeography of the genus Ulva (Ulvophyceae, Chloro-
phyta), with special reference to the Japanese freshwaterand brackish taxa. J. Appl. Phycol. 20: 979–89.
Smith, G. M. 1947. On the reproduction of some Pacific coastspecies of Ulva. Am. J. Bot. 34: 80–7.
Sun, S., Wang, F., Li, C. L. et al. 2008. Emerging challenges:massive green algae blooms in the Yellow Sea. NaturePrecedings hdl:10101/npre.2008.2266.1.
Tamura, K., Dudley, J., Nei, M. and Kumar, S. 2007. MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Mol. Biol. Evol. 24: 1596–9.
Tan, I. H., Blomster, J., Hansen, G. et al. 1999. Molecularphylogenetic evidence for a reversible morphogeneticswitch controlling the gross morphology of two commongenera of green seaweeds, Ulva and Enteromorpha. Mol.Biol. Evol. 16: 1011–8.
Thompson, J. D., Higgins, D. G. and Gibson, T. J. 1994.CLUSTAL W: improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice.Nucleic Acids Res. 22: 4673–80.
Tseng, C. K., Zhang, D. R., Zhang, J. F. et al. 1962. EconomicSeaweeds of China. Science Press, Beijing (in Chinese),pp. 41–50.
Wang, D., Wang, X. L., Li, D. P., Wang, F. J. and Duan, D. L.2006. The genetic analysis and germplasm identificationof the gametophytes of Undaria pinnatifida (Phaeo-phyceae) with RAPD method. J. Appl. Phycol. 18:801–9.
Wang, J. F., Jiang, P., Cui, Y. L. et al. 2010. Molecular analy-sis of green-tide-forming macroalgae in the Yellow Sea.Aquat. Bot. 93: 25–31.
Ye, N. H., Zhuang, Z. M., Jin, X. S. et al. 2008. China is onthe track tackling Enteromorpha spp forming green tide.Nature Precedings hdl:10101/npre.2008.2352.1.
Zhang, X., Wang, H., Mao, Y. et al. 2009. Somatic cells serveas a potential propagule bank of Enteromorpha proliferaforming a green tide in the Yellow Sea, China. J. Appl.Phycol. 22: 173–80.
© 2010 Japanese Society of Phycology
279Ulva green tide in the Yellow Sea
Related Documents











