Protist, Vol. 163, 560–573, July 2012 http://www.elsevier.de/protis Published online date 16 December 2011 ORIGINAL PAPER Ultrastructure and Molecular Phylogeny of Thaumatomonads (Cercozoa) with Emphasis on Thaumatomastix salina from Oslofjorden, Norway Shuhei Ota a,1 , Wenche Eikrem a,b , and Bente Edvardsen a a Marine Biology, Department of Biology, University of Oslo, P.O. Box 1066 Blindern, NO-0316 Oslo, Norway b NIVA, Norwegian Institute for Water Research, Gaustadalléen 21, NO-0349 Oslo, Norway Submitted May 24, 2011; Accepted October 24, 2011 Monitoring Editor: Michael Melkonian A culture of Thaumatomastix was isolated from a sediment sample collected in Oslofjorden and established as a monospecific strain (UIO286). Based on this culture, light and transmission electron microscopy and phylogenetic analyses were carried out. Thaumatomastix species are confined within the order Thaumatomonadida of the class Imbricatea and phylum Cercozoa. They are heterotrophic and their cell bodies are covered with silica scales. Observations of thin sections as well as whole mounts indicate that the morphology and ultrastructure of UIO286 is identical to T. salina, which was initially described from salt pools in Denmark. Detailed examination revealed some new features such as the presence of pseudopodia and silica deposition vesicles producing spine scales. The phylogeny presented here includes ribosomal DNA sequences from both imbricatean cultures and environmen- tal samples. The 18S rDNA phylogenetic tree suggests that (i) Thaumatomastix is paraphyletic within the Thaumatomonadida clade, (ii) there is no close affinity between T. salina and other cultured and sequenced strains, but it is closely related to a sequence obtained from environmental DNA; we pro- pose the present strain to serve as a reference culture of Thaumatomastix species and T. salina. Further, we discuss the distribution, habitats, and evolution of scale formation among euglyphids and thaumatomonads. © 2011 Elsevier GmbH. All rights reserved. Key words: Cercozoan diversity; heterotrophic flagellate; Imbricatea; Oslofjorden; Thaumatomastix; ultrastruc- ture. Introduction The class Imbricatea (Cercozoa) is characterized by solitary cells carrying silica scales and currently comprises four orders: Euglyphida, Spongomona- dida, Thaumatomonadida, and Marimonadida 1 Corresponding author. Present address: Department of Inte- grated Biosciences, Graduate School of Frontier Sciences, University of Tokyo, FSB-601, Kashiwanoha, Kashiwa, Chiba 277-8562, Japan e-mail [email protected], ota [email protected] (S. Ota). (Cavalier-Smith and Chao 2003; Howe et al. 2011). Although Euglyphida and Thaumatomonadida are scaly cercozoans, they differ morphologically; the euglyphids are testate filose amoebae (Meisterfeld 2000), whereas the thaumatomonads are scaly flagellates or amoeboflagellates (Patterson and Zölffel 1991). Well-known thaumatomonad gen- era are Allas Sandon (1927), Thaumatomonas de Saedeleer (1931) and Thaumatomastix Lauterborn (1899), which are biflagellate het- erotrophic flagellates with siliceous body scales. The genus Thaumatomonas is distinguished from © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.protis.2011.10.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Protist, Vol. 163, 560–573, July 2012http://www.elsevier.de/protisPublished online date 16 December 2011
ORIGINAL PAPER
Ultrastructure and Molecular Phylogeny ofThaumatomonads (Cercozoa) with Emphasis onThaumatomastix salina from Oslofjorden, Norway
Shuhei Otaa,1, Wenche Eikrema,b, and Bente Edvardsena
aMarine Biology, Department of Biology, University of Oslo, P.O. Box 1066 Blindern, NO-0316 Oslo, NorwaybNIVA, Norwegian Institute for Water Research, Gaustadalléen 21, NO-0349 Oslo, Norway
Submitted May 24, 2011; Accepted October 24, 2011Monitoring Editor: Michael Melkonian
A culture of Thaumatomastix was isolated from a sediment sample collected in Oslofjorden andestablished as a monospecific strain (UIO286). Based on this culture, light and transmission electronmicroscopy and phylogenetic analyses were carried out. Thaumatomastix species are confined withinthe order Thaumatomonadida of the class Imbricatea and phylum Cercozoa. They are heterotrophicand their cell bodies are covered with silica scales. Observations of thin sections as well as wholemounts indicate that the morphology and ultrastructure of UIO286 is identical to T. salina, which wasinitially described from salt pools in Denmark. Detailed examination revealed some new features suchas the presence of pseudopodia and silica deposition vesicles producing spine scales. The phylogenypresented here includes ribosomal DNA sequences from both imbricatean cultures and environmen-tal samples. The 18S rDNA phylogenetic tree suggests that (i) Thaumatomastix is paraphyletic withinthe Thaumatomonadida clade, (ii) there is no close affinity between T. salina and other cultured andsequenced strains, but it is closely related to a sequence obtained from environmental DNA; we pro-
pose the present strain to serve as a reference culture of Thaumatomastix species and T. salina.Further, we discuss the distribution, habitats, and evolution of scale formation among euglyphids andthaumatomonads.© 2011 Elsevier GmbH. All rights reserved.Key words: Cercozoan diversity; heterotrophic flagellate; Imbricatea; Oslofjorden; Thaumatomastix; ultrastruc-ture.
Introduction
The class Imbricatea (Cercozoa) is characterizedby solitary cells carrying silica scales and currentlycomprises four orders: Euglyphida, Spongomona-dida, Thaumatomonadida, and Marimonadida
Corresponding author. Present address: Department of Inte-grated Biosciences, Graduate School of Frontier Sciences,University of Tokyo, FSB-601, Kashiwanoha, Kashiwa, Chiba277-8562, Japane-mail [email protected], ota [email protected](S. Ota).
(Cavalier-Smith and Chao 2003; Howe et al. 2011).Although Euglyphida and Thaumatomonadida arescaly cercozoans, they differ morphologically; theeuglyphids are testate filose amoebae (Meisterfeld2000), whereas the thaumatomonads are scalyflagellates or amoeboflagellates (Patterson andZölffel 1991). Well-known thaumatomonad gen-era are Allas Sandon (1927), Thaumatomonasde Saedeleer (1931) and ThaumatomastixLauterborn (1899), which are biflagellate het-erotrophic flagellates with siliceous body scales.The genus Thaumatomonas is distinguished from
© 2011 Elsevier GmbH. All rights reserved.doi:10.1016/j.protis.2011.10.007

Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 561
Thaumatomastix by the absence of flagellarscales and by possession of a very short anteriorflagellum (e.g., Wylezich et al. 2007). However,some Thaumatomastix species lack flagellarscales and have a short anterior flagellum (e.g.,T. tripus = Chrysosphaerella tripus; Takahashi andHara 1984), indicating that genus delimitation maybe imprecise and further examination is neededusing both morphological and molecular data.More recently, Howe et al. (2011) transferrednine Thaumatomastix species (basionyms:T. dybsoeana, T. formosa, T. fragilis,T. fusiformis, T. gloenlandica, T. igloolika,T. nigeriensis, T. spinosa, and T. splendida)to the genus Reckertia (Conrad 1920). Moreover,Howe et al. (2011) pointed out that the transfer ofChrysosphaerella triangulata (Balonov 1980) toThaumatomastix (Beech and Moestrup 1986) wasincorrect. Consequently, six species are currentlyrecognized in the genus Thaumatomastix. MarineThaumatomastix species have been recordedin pelagic temperate to polar environments atsalinities ranging from 2 to 35 PSU (Beech andMoestrup 1986; Ikävalko 1998; Moestrup 1979;Takahashi and Hara 1984; Thomsen et al. 1995;Thomsen and Ikävalko 1997; Vørs 1992a,b, 1993).There have been some reports on Thaumatomas-tix morphology using whole-mount transmissionelectron microscopy of collected field material (e.g.,Beech and Moestrup 1986; Thomsen et al. 1995),whereas only a few previous studies have providedphylogenetic information of Thaumatomastix:For instance, Wylezich et al. (2007) proposeda phylogenetic position of Thaumatomonadidawhen describing Thaumatomonas coloniensis,and Chantangsi et al. (2010) and Cavalier-Smithand Chao (2003) proposed a global cercozoanphylogeny. However, there has been no previousstudy integrating morphology, ultrastructure andmolecular phylogeny in Thaumatomastix species.
During a sampling cruise in outer Oslofjordenin June 2010, we collected water and sedimentsamples, from which we established over 60cultured strains. One of the strains (UIO286) iso-lated from the sediment samples was identifiedas Thaumatomastix salina based on morphology.Thaumatomastix salina was originally describedfrom marine salt pools in Denmark as the chrys-ophyte Chrysosphaerella salina (Birch-Andersen1973). Later Beech and Moestrup (1986) reex-amined C. salina and its allies in detail bywhole-mount electron microscopy as well as lightmicroscopy, and found that C. salina was het-erotrophic and showed considerable resemblancein spine morphology to Thaumatomastix species.
Consequently, C. salina was transferred to thegenus Thaumatomastix and new combinationswere provided (Beech and Moestrup 1986). Herewe examined the morphology, ultrastructure, andthe 18S rDNA sequence of cultured T. salina(UIO286) and linked morphology-based identifi-cation to molecular phylogeny. In addition, weobserved silicon deposition vesicles (SDVs) closelyassociated with mitochondria, which has beendescribed previously in thaumatomonads (Karpov1990, 1993, 2000; Karpov and Zhukov 1987;Moestrup 1982; Swale and Belcher 1974, 1975).
Results
Nomenclatural Note
Thaumatomastix salina (Birch-Andersen 1973:142) Beech and Moestrup 1986: 866 emend. S.Ota, Eikrem et Edvardsen
Basionym: Chrysosphaerella salina Birch-Andersen 1973: 140
Emended diagnosis: Cells solitary, heterotrophic,spherical to elliptical, 8-13 �m (mean = 10.7 �m,n = 10) long, 5.3-9.1 �m (mean = 7.5 �m, n = 10)wide. Two flagella of unequal length, posterior flag-ellum naked, longer than body (c. 15 �m); anteriorflagellum short (c. 2.5 �m) with flagellar scales. Cellmovement gliding with rapid flicking motion. Flag-ellate cells may transform to amoeboid cells withbranching pseudopodia. Cell cytoplasm with oneto three (or sometimes more) orange bodies. Cellsurface covered with one layer of inter-connectedspine scales and elliptical scales. Spines 3.5-9.3 �m (mean = 5.5 �m, n = 15) long and 0.1 �mwide. Basal part with two discs, proximal disc0.5 �m; distal disc 0.3 �m in diameter. Tip of spinescale with three ridges. Spineless scales ellipticaland two-tiered, 1.0-1.6 �m (mean = 1.3 �m, n = 15)long, 0.6-1.1 �m (mean = 0.9 �m, n = 15) wide withrims. Elliptical plates with perforations (sometimesnon-perforated). Scales produced within silicadeposition vesicles closely associated with mito-chondria. Ejectosome-like organelles elongatedwith fibrous and amorphous content. Microbodylocated in proximity of nucleus.
Lectotype (ICZN): Birch-Andersen 1973, Bot.Tidsskr. 68: p. 142, fig. 3; Bramsnæsvig, Sjælland,Denmark (for nomenclatural stability under theICZN, we designated a lectotype here).
Habitat: marine or brackish
Distribution (salinity): Denmark (Birch-Andersen
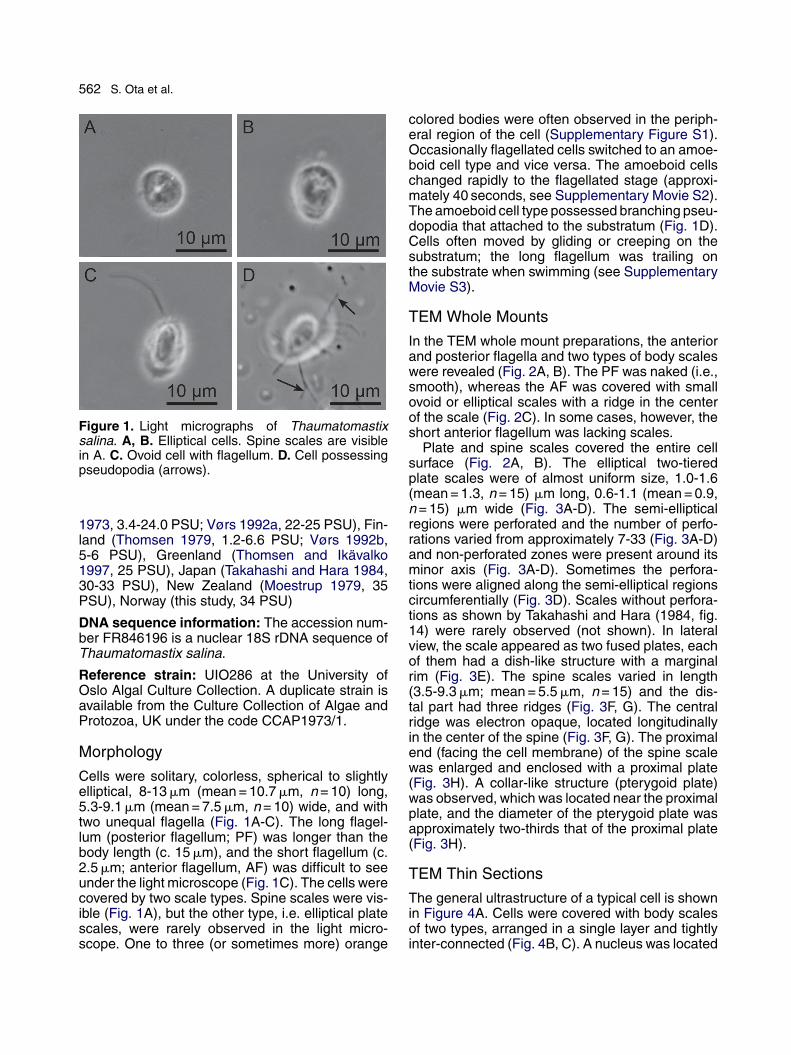
562 S. Ota et al.
Figure 1. Light micrographs of Thaumatomastixsalina. A, B. Elliptical cells. Spine scales are visiblein A. C. Ovoid cell with flagellum. D. Cell possessingpseudopodia (arrows).
1973, 3.4-24.0 PSU; Vørs 1992a, 22-25 PSU), Fin-land (Thomsen 1979, 1.2-6.6 PSU; Vørs 1992b,5-6 PSU), Greenland (Thomsen and Ikävalko1997, 25 PSU), Japan (Takahashi and Hara 1984,30-33 PSU), New Zealand (Moestrup 1979, 35PSU), Norway (this study, 34 PSU)
DNA sequence information: The accession num-ber FR846196 is a nuclear 18S rDNA sequence ofThaumatomastix salina.
Reference strain: UIO286 at the University ofOslo Algal Culture Collection. A duplicate strain isavailable from the Culture Collection of Algae andProtozoa, UK under the code CCAP1973/1.
Morphology
Cells were solitary, colorless, spherical to slightlyelliptical, 8-13 �m (mean = 10.7 �m, n = 10) long,5.3-9.1 �m (mean = 7.5 �m, n = 10) wide, and withtwo unequal flagella (Fig. 1A-C). The long flagel-lum (posterior flagellum; PF) was longer than thebody length (c. 15 �m), and the short flagellum (c.2.5 �m; anterior flagellum, AF) was difficult to seeunder the light microscope (Fig. 1C). The cells werecovered by two scale types. Spine scales were vis-ible (Fig. 1A), but the other type, i.e. elliptical platescales, were rarely observed in the light micro-scope. One to three (or sometimes more) orange
colored bodies were often observed in the periph-eral region of the cell (Supplementary Figure S1).Occasionally flagellated cells switched to an amoe-boid cell type and vice versa. The amoeboid cellschanged rapidly to the flagellated stage (approxi-mately 40 seconds, see Supplementary Movie S2).The amoeboid cell type possessed branching pseu-dopodia that attached to the substratum (Fig. 1D).Cells often moved by gliding or creeping on thesubstratum; the long flagellum was trailing onthe substrate when swimming (see SupplementaryMovie S3).
TEM Whole Mounts
In the TEM whole mount preparations, the anteriorand posterior flagella and two types of body scaleswere revealed (Fig. 2A, B). The PF was naked (i.e.,smooth), whereas the AF was covered with smallovoid or elliptical scales with a ridge in the centerof the scale (Fig. 2C). In some cases, however, theshort anterior flagellum was lacking scales.
Plate and spine scales covered the entire cellsurface (Fig. 2A, B). The elliptical two-tieredplate scales were of almost uniform size, 1.0-1.6(mean = 1.3, n = 15) �m long, 0.6-1.1 (mean = 0.9,n = 15) �m wide (Fig. 3A-D). The semi-ellipticalregions were perforated and the number of perfo-rations varied from approximately 7-33 (Fig. 3A-D)and non-perforated zones were present around itsminor axis (Fig. 3A-D). Sometimes the perfora-tions were aligned along the semi-elliptical regionscircumferentially (Fig. 3D). Scales without perfora-tions as shown by Takahashi and Hara (1984, fig.14) were rarely observed (not shown). In lateralview, the scale appeared as two fused plates, eachof them had a dish-like structure with a marginalrim (Fig. 3E). The spine scales varied in length(3.5-9.3 �m; mean = 5.5 �m, n = 15) and the dis-tal part had three ridges (Fig. 3F, G). The centralridge was electron opaque, located longitudinallyin the center of the spine (Fig. 3F, G). The proximalend (facing the cell membrane) of the spine scalewas enlarged and enclosed with a proximal plate(Fig. 3H). A collar-like structure (pterygoid plate)was observed, which was located near the proximalplate, and the diameter of the pterygoid plate wasapproximately two-thirds that of the proximal plate(Fig. 3H).
TEM Thin Sections
The general ultrastructure of a typical cell is shownin Figure 4A. Cells were covered with body scalesof two types, arranged in a single layer and tightlyinter-connected (Fig. 4B, C). A nucleus was located
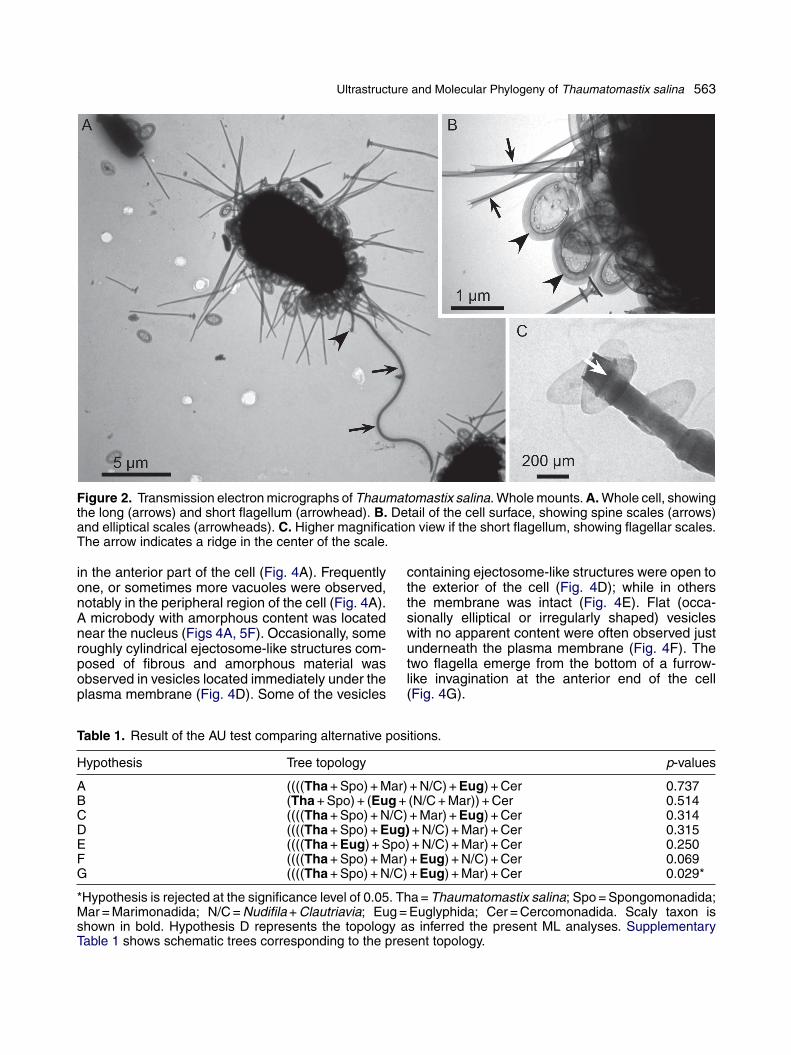
Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 563
Figure 2. Transmission electron micrographs of Thaumatomastix salina. Whole mounts. A. Whole cell, showingthe long (arrows) and short flagellum (arrowhead). B. Detail of the cell surface, showing spine scales (arrows)and elliptical scales (arrowheads). C. Higher magnification view if the short flagellum, showing flagellar scales.The arrow indicates a ridge in the center of the scale.
in the anterior part of the cell (Fig. 4A). Frequentlyone, or sometimes more vacuoles were observed,notably in the peripheral region of the cell (Fig. 4A).A microbody with amorphous content was locatednear the nucleus (Figs 4A, 5F). Occasionally, someroughly cylindrical ejectosome-like structures com-posed of fibrous and amorphous material wasobserved in vesicles located immediately under theplasma membrane (Fig. 4D). Some of the vesicles
containing ejectosome-like structures were open tothe exterior of the cell (Fig. 4D); while in othersthe membrane was intact (Fig. 4E). Flat (occa-sionally elliptical or irregularly shaped) vesicleswith no apparent content were often observed justunderneath the plasma membrane (Fig. 4F). Thetwo flagella emerge from the bottom of a furrow-like invagination at the anterior end of the cell(Fig. 4G).
Table 1. Result of the AU test comparing alternative positions.
Hypothesis Tree topology p-values
A ((((Tha + Spo) + Mar) + N/C) + Eug) + Cer 0.737B (Tha + Spo) + (Eug + (N/C + Mar)) + Cer 0.514C ((((Tha + Spo) + N/C) + Mar) + Eug) + Cer 0.314D ((((Tha + Spo) + Eug) + N/C) + Mar) + Cer 0.315E ((((Tha + Eug) + Spo) + N/C) + Mar) + Cer 0.250F ((((Tha + Spo) + Mar) + Eug) + N/C) + Cer 0.069G ((((Tha + Spo) + N/C) + Eug) + Mar) + Cer 0.029*
*Hypothesis is rejected at the significance level of 0.05. Tha = Thaumatomastix salina; Spo = Spongomonadida;Mar = Marimonadida; N/C = Nudifila + Clautriavia; Eug = Euglyphida; Cer = Cercomonadida. Scaly taxon isshown in bold. Hypothesis D represents the topology as inferred the present ML analyses. SupplementaryTable 1 shows schematic trees corresponding to the present topology.
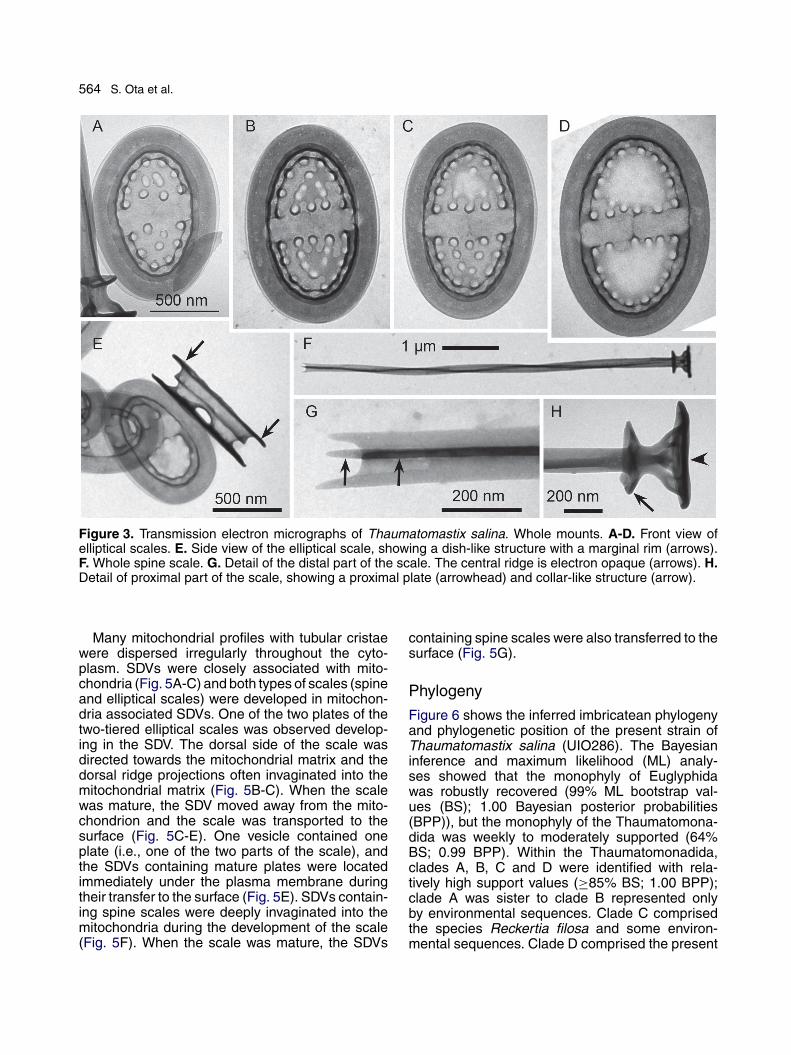
564 S. Ota et al.
Figure 3. Transmission electron micrographs of Thaumatomastix salina. Whole mounts. A-D. Front view ofelliptical scales. E. Side view of the elliptical scale, showing a dish-like structure with a marginal rim (arrows).F. Whole spine scale. G. Detail of the distal part of the scale. The central ridge is electron opaque (arrows). H.Detail of proximal part of the scale, showing a proximal plate (arrowhead) and collar-like structure (arrow).
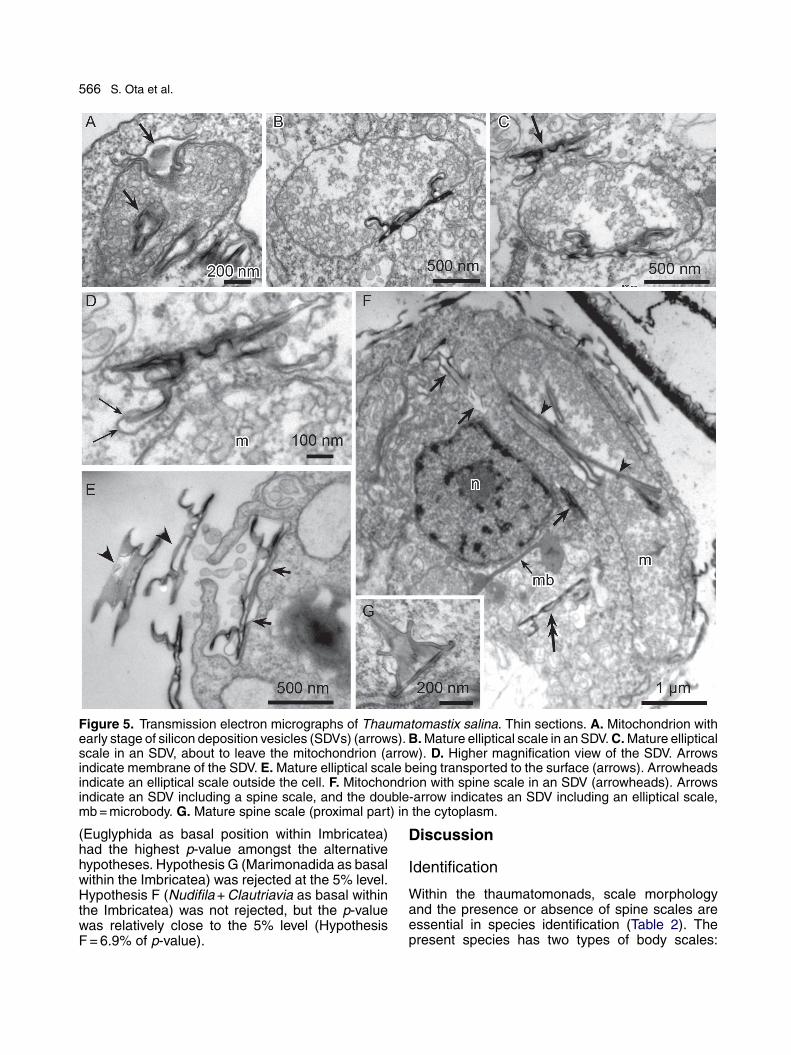
Many mitochondrial profiles with tubular cristaewere dispersed irregularly throughout the cyto-plasm. SDVs were closely associated with mito-chondria (Fig. 5A-C) and both types of scales (spineand elliptical scales) were developed in mitochon-dria associated SDVs. One of the two plates of thetwo-tiered elliptical scales was observed develop-ing in the SDV. The dorsal side of the scale wasdirected towards the mitochondrial matrix and thedorsal ridge projections often invaginated into themitochondrial matrix (Fig. 5B-C). When the scalewas mature, the SDV moved away from the mito-chondrion and the scale was transported to thesurface (Fig. 5C-E). One vesicle contained oneplate (i.e., one of the two parts of the scale), andthe SDVs containing mature plates were locatedimmediately under the plasma membrane duringtheir transfer to the surface (Fig. 5E). SDVs contain-ing spine scales were deeply invaginated into themitochondria during the development of the scale(Fig. 5F). When the scale was mature, the SDVs
containing spine scales were also transferred to thesurface (Fig. 5G).
Phylogeny
Figure 6 shows the inferred imbricatean phylogenyand phylogenetic position of the present strain ofThaumatomastix salina (UIO286). The Bayesianinference and maximum likelihood (ML) analy-ses showed that the monophyly of Euglyphidawas robustly recovered (99% ML bootstrap val-ues (BS); 1.00 Bayesian posterior probabilities(BPP)), but the monophyly of the Thaumatomona-dida was weekly to moderately supported (64%BS; 0.99 BPP). Within the Thaumatomonadida,clades A, B, C and D were identified with rela-tively high support values (≥85% BS; 1.00 BPP);clade A was sister to clade B represented onlyby environmental sequences. Clade C comprisedthe species Reckertia filosa and some environ-mental sequences. Clade D comprised the present
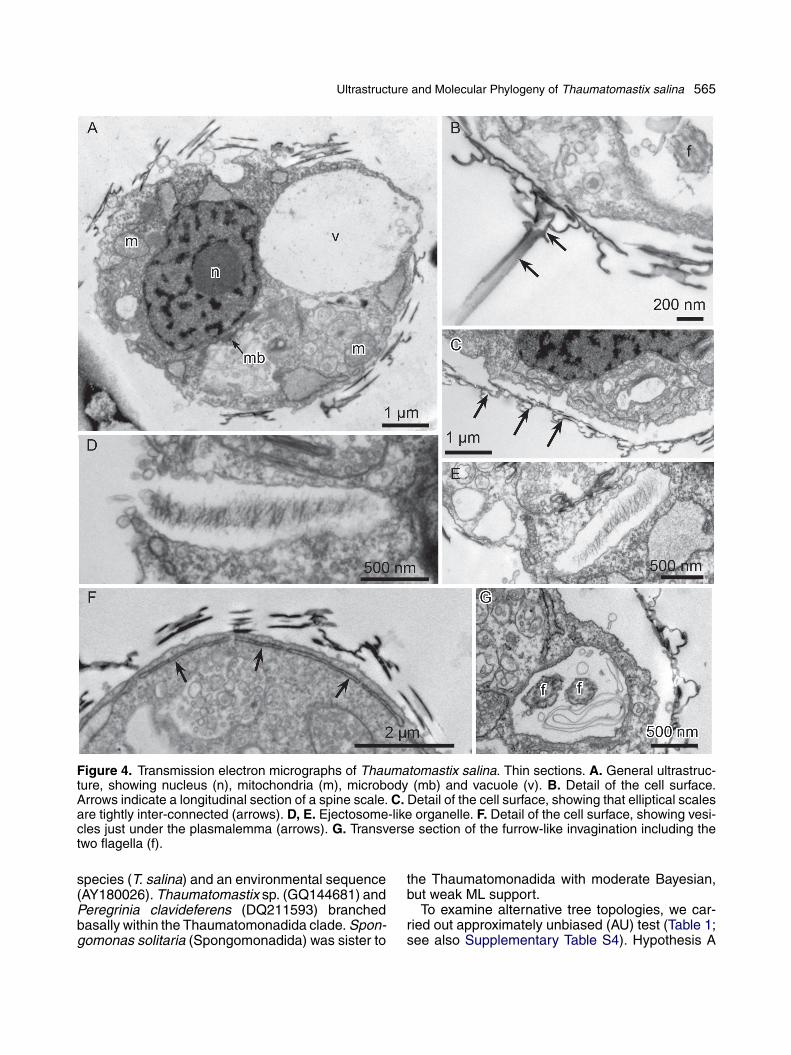
Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 565
Figure 4. Transmission electron micrographs of Thaumatomastix salina. Thin sections. A. General ultrastruc-ture, showing nucleus (n), mitochondria (m), microbody (mb) and vacuole (v). B. Detail of the cell surface.Arrows indicate a longitudinal section of a spine scale. C. Detail of the cell surface, showing that elliptical scalesare tightly inter-connected (arrows). D, E. Ejectosome-like organelle. F. Detail of the cell surface, showing vesi-cles just under the plasmalemma (arrows). G. Transverse section of the furrow-like invagination including thetwo flagella (f).
species (T. salina) and an environmental sequence(AY180026). Thaumatomastix sp. (GQ144681) andPeregrinia clavideferens (DQ211593) branchedbasally within the Thaumatomonadida clade. Spon-gomonas solitaria (Spongomonadida) was sister to
the Thaumatomonadida with moderate Bayesian,but weak ML support.
To examine alternative tree topologies, we car-ried out approximately unbiased (AU) test (Table 1;see also Supplementary Table S4). Hypothesis A
566 S. Ota et al.
Figure 5. Transmission electron micrographs of Thaumatomastix salina. Thin sections. A. Mitochondrion withearly stage of silicon deposition vesicles (SDVs) (arrows). B. Mature elliptical scale in an SDV. C. Mature ellipticalscale in an SDV, about to leave the mitochondrion (arrow). D. Higher magnification view of the SDV. Arrowsindicate membrane of the SDV. E. Mature elliptical scale being transported to the surface (arrows). Arrowheadsindicate an elliptical scale outside the cell. F. Mitochondrion with spine scale in an SDV (arrowheads). Arrowsindicate an SDV including a spine scale, and the double-arrow indicates an SDV including an elliptical scale,mb = microbody. G. Mature spine scale (proximal part) in the cytoplasm.
(Euglyphida as basal position within Imbricatea)had the highest p-value amongst the alternativehypotheses. Hypothesis G (Marimonadida as basalwithin the Imbricatea) was rejected at the 5% level.Hypothesis F (Nudifila + Clautriavia as basal withinthe Imbricatea) was not rejected, but the p-valuewas relatively close to the 5% level (HypothesisF = 6.9% of p-value).
Discussion
Identification
Within the thaumatomonads, scale morphologyand the presence or absence of spine scales areessential in species identification (Table 2). Thepresent species has two types of body scales:

Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 567
0.1
Thaumatomonas sp. SCCAP T1 AY496046
Thaumatomonas seravini AY496044
23/-
Thaumatomonas sp. U4244641/0.85
Thaumatomonas coloniensis DQ21159153/0.75
Thaumatomonas sp. ATCC50250 AF411261
76/0.98
Thaumatomonas sp. AF411260
Thaumatomonas sp. TMT002 DQ98048625/-
Thaumatomonas seravini AF411259
71/-
Allas diplophysa AF411262
79/0.99
Uncultured Amb_18S_1434 EF023966
Uncultured Elev_18S_7011 EF025032
69/1.00
Uncultured Amb_18S_1319 EF023867
100/1.00
Uncultured 3b-D9 FN690393
Uncultured 5-B11 FN69039444/-
Uncultured 8-32 FN690392
Reckertia filosa AY26804092/1.00
55/0.83
Thaumatomastix salina UIO286 FR846196
Uncultured CCW52 AY18002685/1.00
50/0.72
Thaumatomastix sp. GQ144681
53/0.88
Peregrinia clavideferens DQ211593
64/0.99
Spongomonas solitaria CCAP 1971/1 HQ121435
39/0.70
Euglypha rotunda CCAP1520/1 X77692
Euglypha rotunda AJ418782
93/1.00
Euglypha rotunda AJ418784
Trinema enchelys AJ418792
Corythion dubium EF456751
43/0.56
Euglypha acanthophora AJ41878
44/0.78
Assulina muscorum AJ418791
Ovulinata parva strain CCAP1554/1 HQ121432
Paulinella chromatophora X81811
Cyphoderia ampulla GU228896
80/0.94
99/1.00
9/-
Nudifila producta CCAP1911/1 HQ121434
Clautriavia biflagellata FJ919772 38/0.74
25/0.50
Pseudopirsonia mucosa AJ5611
Auranticordis quadriverberis EU48439485/1.00
Cercomonas plasmodialis AF411268
Cercomonas LargeSA AF41126698/1.00
Cavernomonas mira FJ790718
54/-
Eocercomonas sp. AF411269
Uncultured eukaryote clone I_4_50 AB534514
97/1.00
x2
x2
Marimonadida
Sarcomonadea (outgroup)
ThaumatomonadidaB
C
D
Spongomonadida
Euglyphida
A
N/C clade
Figure 6. RAxML phylogenetic tree of imbricateans and sarcomonads (out group) based on 18SrDNA sequences (40 OTUs and 1,419 nucleotide positions). Accession numbers are given after thespecies/environmental clone names. Bootstrap values/Bayesian posterior probabilities are shown near the inter-nal nodes. Black dots on nodes indicate support values of 100%/1.00. The present species (Thaumatomastixsalina) is shown in bold.
spine scales and elliptical scales, and so doThaumatomastix salina, T. bipartita and T. setiferaas well. Thaumatomastix bipartita is distinguishedfrom T. salina by the possession of a bipartitestructure in the spine scales (Beech and Moestrup1986). The AF of T. salina is very short and difficult
to see in light microscopy, whereas in T. setifera theAF is longer and both flagella are clearly visible.Consequently, the present isolate (UIO286) isassigned to T. salina. Our isolate had the ability totransform into an amoeboid stage and it formedpseudopodia. In addition, an ejectosome-like
568 S. Ota et al.
organelle was observed. All together these novelphenotypes called for an emended diagnosis of T.salina, which was provided above. The morphologyof the body scales may vary and Takahashi andHara (1984) showed that the elliptical body scalesmay even lack perforations. Our observationsrevealed that the number of perforations wasvariable, suggesting that non-perforated bodyscales are within the intra-species variation.
The ejectosome-like organelle observed in T.salina was located just beneath the plasma mem-brane. It may also resemble a stage of endo-or exocytosis. Since we are not convinced thatthis organelle is an ejectosome, we use theterm “ejectosome-like organelle”. The longitudinaloutline was slightly cylindrical and contained astructure that had both fibrous and amorphouscomponents. In contrast, the cylindrical ejecto-somes previously known from cercozoan genera(e.g., Gyromitus, Thaumatomonas and Bigelowiella[chlorarachniophytes]) have a homogeneous andsomewhat electron-opaque structure (Cavalier-Smith et al. 2008; Howe et al. 2011; Moestrupand Sengco 2001; Swale and Belcher 1975). Thisorganelle might be a unique ultrastructural featurein Thaumatomastix.
Both elliptical and spine scales were formedin SDVs associated with mitochondria. In thepresent study we did not observe a mitochondrion-associated SDV forming both types of scales
simultaneously, suggesting that one mitochon-drion-associated SDV can form one type of scalesonly. However, Moestrup (1982) noted that in Reck-ertia sagittifera both body scales and flagellarscales were formed in the same mitochondrion-associated SDV simultaneously, and thus furtherobservations are required to confirm this accord-ingly. Recently, mitochondria-associated SDVswere also reported by Zolotarev et al. (2011) inThaumatomastix sp. from the White Sea. Althoughwe do not know the physiological advantages ofmitochondrion-associated SDVs, we infer that theSDVs need some functions of the mitochondria todevelop scales. This feature is only observed inthe thaumatomonad lineage and appears uniqueamong the unicellular eukaryotes. Evolutionaryaspects are discussed below.
Phylogenetic Relationship
The imbricatean (Euglyphida, Thaumatomonadida,Spongomonadida, and Marimonadida) phylogenybased on 18S rDNA sequences presented hereis roughly consistent with previously published MLtrees (Cavalier-Smith and Chao 2003; Ekelundet al. 2004; Howe et al. 2011; Lara et al. 2007;Wylezich et al. 2007) and provides new insightinto the phylogenetic position of T. salina. Thau-matomastix salina was found to be sister to anuncultured environmental DNA sequence CCW52
Table 2. Comparison of morphology in Thaumatomastix species.
Thaumatomastixspecies
References Numberof bodyscaletypes
Type ofbodyscale
Spines Habitattype
Other remarks
T. bipartita Beech and Moestrup,1986
2 elliptical Yes Marine Bifurcated spines
T. patelliformis Takahashi and Hara,1984; Beech andMoestrup, 1986
2 triangular Yes Marine Flagella present;spinelength = 3.4-6.6 �m
T. salina (Birch-Andersen,1973) Beech andMoestrup, 1986
2 elliptical Yes Marine -
T. setifera Lauterborn, 1899 Unknown ? Yes Marine Both flagellaobserved
T. tripus Takahashi and Hara,1984; Beech andMoestrup, 1986
2 triangular Yes Marine Flagella present;spinelength = 7.0-19 �m
T. tauryanini Mikrjukov, 2002 2 triangular Yes Marine no flagella observed
Note: Thaumatomastix species are listed alphabetically. Some Thaumatomastix species are unlisted here: T.punctata is nomen nudum, T. thomseni was transferred to the Reckertia as R. thomseni (Karpov 2011), and T.triangulata was dealt with Chrysophaerella species according to Howe et al. (2011).

Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 569
(AY180026) originating from a marine anoxic envi-ronment off Cape Cod, NE USA (Stoeck andEpstein 2003) with robust statistical support. Thepresent analyses also demonstrate that clade Bis composed only of environmental sequencesderived from soil samples (Lesaulnier et al. 2008);and there are few isolated cultures of taxa closelyrelated to T. salina, probably due to the relativedifficulty of isolating and culturing Thaumatomas-tix species. In the present analyses Peregriniaclavideferens is the deepest branch in the thau-matomonads; this is also consistent with the tree ofWylezich et al. (2007) (Gyromitus sp. HFCC94 wasre-described and renamed as Peregrinia clavidef-erens by Howe et al. (2011)).
The well-supported clade A mainly comprisesThaumatomonas species, but also contains strainATCC50250 (originally labeled as “Thaumatomas-tix sp.”) and Allas diplophysa. As noted previouslyby Cavalier-Smith and Chao (2003) and Wylezichet al. (2007), the present analyses also show thatthere are very small differences in 18S rDNA withinclade A (e.g., more than 99% similarity of 18SrDNA sequences between “Thaumatomastix” sp.strain ATCC50250 and Thaumatomonas colonien-sis). More recently, Howe et al. (2011) provided acomprehensive taxonomy of filoseans (Cercozoa),where they noted the misidentification of strainATCC50250 “Thaumatomastix sp.”, which is actu-ally a Thaumatomonas species. Accordingly, wealso relabeled strain ATCC50250 of the presentOTU as Thaumatomonas in the phylogenetic analy-ses. Howe et al. (2011) also dealt with the taxonom-ical and nomenclatural confusion that surroundsthe morphologically similar genera Allas and Thau-matomonas. They suggested that there are no mor-phological grounds for retaining Thaumatomonasas a genus distinct from Allas, which concurs witha high degree of similarity in 18S rDNA sequencesbetween the genera. However, the authors retainboth genera because of nomenclatural stability; i.e.,they did not regard the more recent name Thau-matomonas (widely used name) as junior synonym,and considered ATCC50365 as a reference cultureof the genus Allas based on the novel scale typediscovered in strain ATCC50365 (Howe et al. 2011).We basically agree with this idea; however furtherculture-based studies are needed prior to a revi-sion of the taxonomy of thaumatomonads, notablythe Allas-Thaumatomonas complex.
Because the basal branching was poorlyresolved in the present phylogenetic analyses,we carried out an approximately unbiased (AU)test to compare seven alternative tree topolo-gies. Although hypothesis D, corresponding to
the present RAxML tree (the best tree) was notrejected, hypothesis A had the highest p-values.Hypothesis G (Marimonadida as basal position)was rejected at the 5% level. Although hypothesis F(Nudifila + Clautriavia as basal position within Imbri-catea) was not rejected at the significance level of5%, the p-value was relatively low (p-value = 6.9%).Among hypotheses A, B, C, D, and E, hypothesisE is the only topology where Thaumatomona-dida and Euglyphida are monophyletic; the othersare topologies that represent a paraphyletic rela-tionship in Thaumatomonadida and Euglyphida.Moreover, the topology of hypothesis B is consis-tent with recently published trees (Chantangsi et al.2010; Howe et al. 2011). The previous phylogeneticanalyses as well as the present AU test suggestthat scale-less imbricateans are likely to be posi-tioned between the thaumatomonad and euglyphidlineages (see below).
Evolution of Scale Formation
As suggested by Wylezich et al. (2007), the pro-cess of scale formation is completely different ineuglyphids and thaumatomonads; euglyphid bodyscales are formed in vesicles associated withthe Golgi body (Meisterfeld 2000), whereas thau-matomonad body scales are formed in vesiclesassociated with mitochondria (Karpov 1990, 1993,2000; Karpov and Zhukov 1987; Moestrup 1982;Swale and Belcher 1974, 1975; this study). In addi-tion, the morphology of both groups is very distinct;the euglyphids are testate amoebae (Lara et al.2007), whereas the thaumatomonads are scale-bearing flagellates (Swale and Belcher 1974, 1975;Thomsen et al. 1995; Wylezich et al. 2007; thisstudy). In the present phylogenetic analyses, theThaumatomonadida and Euglyphida clades wereseparately recovered with high Bayesian statisticalsupports (but low to high ML support), suggestingthat they are distinct lineages. Moreover, the AUtest suggested that several scale-less lineages areplaced between these clades. Based on the AUtest, the scale-less genera Spongomonas, Pseu-dopirsonia (Kühn et al. 2004) and Auranticordis(Chantangsi et al. 2008) are likely to be positionedbetween the thaumatomonad and euglyphid lin-eages (Table 1; Hypotheses A, B, C, and F). Takingthese facts into account, it is deduced that scaleformation within the Imbricatea has distinct originsrather than a single origin, and scale formationmight have evolved at least twice among the imbri-cateans. Our results comply with Patterson’s viewthat the formation of scales in association with

570 S. Ota et al.
mitochondria may be a synapomorphy for thau-matomonads (Patterson 1999).
Distribution and Habitat ofThaumatomonads
Our study from Oslofjorden suggested that thau-matomonads here are restricted to sediments asbenthos, as no thaumatomonad cells or sequenceswere detected from water samples of any depthsor size fractions (Ota et al. unpublished data).This may be consistent with previous reportsby Thomsen et al. (1995) and Thomsen andIkävalko (1997), in which Thaumatomastix specieswere found from sea ice cores as benthos.However, some previous studies reported thatThaumatomastix species occurred in water sam-ples as plankton (Beech and Moestrup 1986;Birch-Andersen 1973; Takahashi and Hara 1984).Wylezich et al. (2007) showed that the contributionof thaumatomonads to the pelagic heterotrophicprotist community was on average only about 4%in brackish water and 0.1% to 4% in fresh watersamples, whereas maximum abundances of thau-matomonads were recorded in the detritus layer orsediments. Based on their observations, they indi-cated that re-suspension of detrital particles couldsignificantly contribute to the distribution of thau-matomonads.
As mentioned above, the uncultured cloneCCW52 (the closest relative of T. salina) originatedfrom a marine anoxic environment. In addition, theuncultured marine eukaryote clones GoC4_B04(FJ153695) and GoC4_A11 (FJ153693), having91% and 90% similarity in partial 18S rDNAsequences with Thaumatomastix sp. (GQ144681),respectively (determined by BLAST search), orig-inated from suboxic and anoxic waters of theGotland Deep (Baltic Sea) (Stock et al. 2009),suggesting that Thaumatomastix and its relativeshave a wide distribution including suboxic andanoxic environments. Further studies includingenvironmental DNA libraries (e.g., cloning and454-pyrosequencing) would elucidate habitat pref-erences and community structures of cercozoansand thaumatomonads (Thaumatomonas, Thau-matomastix, and Allas).
Concluding Remarks
The environmental sequencing approach is widelyused in marine ecology and environmental sci-ences to explore protist biodiversity, and geneticdata are rapidly accumulating in databases from
environmental DNA clone libraries (e.g., Lepèreet al. 2009; Massana et al. 2011; Sauvadet et al.2010) and recently also from pyrosequencing (e.g.,Bråte et al. 2010; Shalchian-Tabrizi et al. 2010).These are powerful approaches that uncoverthe biodiversity of marine environments, but stillmany environmental DNA sequences are of uncer-tain affinities, and it is important that moleculardata is linked to morphological identification. Thepresent strain UIO286 isolated from Oslofjordenwas identified as Thaumatomastix salina based oncomprehensive morphological studies. The phylo-genetic analyses demonstrated that T. salina strainUIO286 is sister to one environmental sequencethat is as-yet uncultured. Thus, we propose UIO286as a reference culture of T. salina for further envi-ronmental and phylogenetic studies. Isolating andculturing efforts are still required to understand pro-tist diversity in marine environments.
Methods
Sampling and culture: T. salina, strain UIO286 (isolator’s ref-erence number: OF258) originated from a sediment samplecollected by B. Edvardsen 23 June 2010 during a BioMarKscruise with R/V Trygve Braarud in the outer Oslofjorden at astation (59.254604N, 10.711379E, 103 m depth) north east ofRauøy in Østfold County, Southern Norway. The sediment wascollected using a Gemini corer, from which the upper 2 cm wastransferred to a flask added bottom seawater (34 PSU, 6 ◦C)and kept cold (∼6 ◦C) and dark until isolation.
T. salina was isolated by S. Ota using the micropipettemethod (Andersen and Kawachi 2005), and the culture wasmaintained at 17 ◦C in sterile filtered seawater (30 PSU salinity)added yeast extract (final conc. 25 mg/L; Becton, Dickinson andCompany) and pepton (final conc. 50 mg/L; Becton, Dickinsonand Company) or half-strength ES medium (Kasai et al. 2009)added yeast extract (final conc. 25 mg/L) and pepton (final conc.50 mg/L).
Light microscopy (LM): Living cells were observed undera Zeiss Axio Scope A1 microscope (Carl Zeiss, Oberkochen,Germany) equipped with Nomarski differential interference con-trast and phase contract optics. Light micrographs were takenwith a Nikon D5000 digital camera (Nikon, Tokyo, Japan).
Transmission electron microscopy (TEM): For wholemounts preparation, droplets of cells fixed with 5% glutaralde-hyde (Electron Microscopy Sciences, Hatfield, PA, USA; finalconc.) were placed on grids covered by a formvar film. After1-2 h, the droplets were removed and the grids were washedtwice in distilled water. Then the cells were stained in saturatedaqueous uranyl acetate (Merck, Darmstadt, Germany) for about90 s, and washed with distilled water. For thin section, 50% glu-taraldehyde was added directly into the culture and fixed 1 h atroom temperature (glutaraldehyde final conc. 2.5%). The cellswere washed four times (5 min each) with 0.1 M sodium cacody-late buffer (pH 7.8) (Agar Scientific Ltd., Essex, UK). Aftercentrifugation and removal of the supernatant, the pellet waspost-fixed in 1% (final conc.) osmium tetroxide overnight at 4 ◦C.The cells were rinsed three times with the same buffer, dehy-drated through a graded ethanol series (30%, 50%, 70%, 90%,

Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 571
and 100% × 4; 15 min each) on ice, and transferred to propy-lene oxide twice (15 min each). Samples were left overnight ina 1:1 mixture of propyleneoxide and Epon’s resin (EM Bed-812based on EPON-812, Sigma), followed by incubation in 100%Epon’s resin overnight at room temperature, and polymerized at60 ◦C overnight (12 h or more). Ultrathin sections were cut ona Reichert Ultracut S ultramicrotome (Leica, Vienna, Austria)using a diamond knife. The sections were mounted on coppergrids coated with polyvinyl formvar films, and stained in satu-rated aqueous uranyl acetate (30 min) and lead citrate (3 min)(Reynolds 1963). Sections and whole mount preparations wereobserved using a FEI/PHILIPSCM-100 TEM (Hillsboro, Ore-gon, USA) at 80 kV at the Electron Microscopy Unit, Dept. ofMolecular Sciences, University of Oslo.
DNA extraction, polymerase chain reaction (PCR), andsequencing: Cells were collected from a two-weeks-old rawculture growing in a 50 ml culture flasks (Nunc, Roskilde, Den-mark) containing 10-20 ml medium and genomic DNA wasextracted using the E.Z.N.A.TM SP Plant DNA kit (Omega-Bio-Tek, Norcross, GA, USA) according to the manufacturer’sprotocol. The SSU (18S) nuclear ribosomal encoding regionwas amplified using primers 1F and 1528R described inEdvardsen et al. (2003), but with the 5 PRIME Taq DNApolymerase (5 PRIME, Hamburg, Germany). The PCR con-dition was as follows: initial denaturation at 94 ◦C for 3 min,34 cycles (denaturation at 94 ◦C for 30 s, annealing at 54 ◦Cfor 30 s, extension at 68 ◦C for 1 min), and final extension at68 ◦C for 10 min. PCR products were run on a 0.8% agarosegel and checked for purity and correct fragment length. ThePCR products were purified using ExoSAP-IT® (USB Corp.,Cleveland, OH, USA) and bidirectionally sequenced using anApplied Biosystems 3730 analyzer (Applied Biosystems, CA,USA) sequencing device at the Department of Biology, Univer-sity of Oslo.
Phylogenetic analyses: Sequences of the 18S rDNA werealigned using ClustalX v. 2.0 (Larkin et al. 2007) and manuallyedited using BioEdit v. 7.0.5.3 (Hall 1999). Maximum likelihood(ML) analysis was carried out using RAxML v. 7.0.3 (Stamatakis2006). The general time reversible model with parametersaccounting for invariable sites (I) and gamma-distributed (G)rate variation across sites with four discrete rate categorieswas used for the 18S rDNA dataset. The bootstrap analyseswere done in 100 replicates for the ML analysis. Bayesianinference under the same evolutional model was performedwith MrBayes v. 3.1.2 (Ronquist and Huelsenbeck 2003). TwoMarkov Chain Monte Carlo (MCMC) runs each with four chainswere performed for 1,000,000 generations, where the aver-age standard deviation of split frequencies was 0.009024.Trees were sampled every 100 generations. Bayesian posteriorprobabilities were calculated from the majority rule consen-sus of the tree sampled after the initial burn-in phase. Inthe present Bayesian and ML analyses, taxon sampling wasmainly based on the previous trees of Lara et al. (2007)and Wylezich et al. (2007) in a Monadofilosan context, andcercomonad sequences were used as outgroup. Some envi-ronmental (uncultured) sequences were also included in theanalyses. The Bayesian posterior probabilities were annotatedonto the ML tree. Bayesian phylogenetic analyses were carriedout on the University of Oslo Bioportal (www.bioportal.uio.no).The SSU rDNA alignment is available from TreeBASE(http://purl.org/phylo/treebase/phylows/study/TB2:S11905).
For examination of alternative trees, site-wise log-likelihoodsof seven alternative trees were calculated using TREE-PUZZLE5.2 (Schmidt et al. 2002) with the same evolutionary model,and approximately unbiased (AU) tests were performed usingCONSEL v.0.1i (Shimodaira and Hasegawa 2001).
Acknowledgements
We are most grateful to Tove Bakar at the Elec-tron Microscopy Unit, University of Oslo, for her kindhelp with the transmission electron microscope.We would like to thank Simon Dittami and threeanonymous reviewers for critical reading of themanuscript. We also thank Elianne Sirnæs Eggeand Sissel Brubak for their kind supports and tech-nical assistance. This work was supported by theEU BiodivERsA project BioMarKs (2008-29765-62739-34).
Appendix A. Supplementary data
Supplementary data associated with this arti-cle can be found, in the online version, atdoi:10.1016/j.protis.2011.10.007.
References
Andersen RA, Kawachi M (2005) Traditional MicroalgaeIsolation Techniques. In Andersen RA (ed) Algal Culturing Tech-niques. Elsevier, Academic Press, Amsterdam, pp 83–100
Balonov IM (1980) On a new species of the genusChrysosphaerella (Chrysophyta). Bot Zh 65:1190–1191
Beech PL, Moestrup Ø (1986) Light and electron microscop-ical observations on the heterotrophic protist Thaumatomastixsalina comb. nov. (syn. Chrysosphaerella salina) and its allies.Nord J Bot 6:865–877
Birch-Andersen P (1973) Chrysosphaerella salina. A newspecies of the Chrysophyceae from salt marsh pools. BotTidsskr 68:140–144
Bråte J, Logares R, Berney C, Ree DK, Klaveness D, Jakob-sen KS, Shalchian-Tabrizi K (2010) Freshwater Perkinsea andmarine-freshwater colonizations revealed by pyrosequencingand phylogeny of environmental rDNA. ISME J 4:1144–1153
Cavalier-Smith T, Chao EE (2003) Phylogeny and classifica-tion of phylum Cercozoa (Protozoa). Protist 154:341–358
Cavalier-Smith T, Lewis R, Chao EE, Oates B, Bass D (2008)Morphology and phylogeny of Sainouron acronematica sp. n.and the ultrastructural unity of Cercozoa. Protist 159:591–620
Chantangsi C, Esson HJ, Leander BS (2008) Morphology andmolecular phylogeny of a marine interstitial tetraflagellate withputative endosymbionts: Auranticordis quadriverberis n. gen. etsp. (Cercozoa). BMC Microbiol 8:e123
Chantangsi C, Hoppenrath M, Leander BS (2010) Evolution-ary relationships among marine cercozoans as inferred fromcombined SSU and LSU rDNA sequences and polyubiquitininsertions. Mol Phylogenet Evol 57:518–527
Conrad W (1920) Sur un flagella nouveau à trichocystes, Reck-ertia sagittifera, n. g., n. sp. Bull Acad Roy Sci Belg 6:541–555

572 S. Ota et al.
De Saedeleer H (1931) Notes de Protistologie IV. Thau-matomonas lauterbornii, n.g., n. sp., flagella nouveau. RecueilL’Instit Zool Torley Rousseau 3:89–103
Edvardsen B, Shalchian-Tabrizi K, Jakobsen KS, MedlinLK, Dahl E, Brubak S, Paasche E (2003) Genetic variabilityand molecular phylogeny of Dinophysis species (Dinophyceae)from Norwegian waters inferred from single cell analyses ofrDNA. J Phycol 39:395–408
Ekelund F, Daugbjerg N, Fredslunda L (2004) Phylogeny ofHeteromita, Cercomonas and Thaumatomonas based on SSUrDNA sequences, including the description of Neocercomonasjutlandica sp. nov., gen. nov. Eur J Protistol 40:119–135
Hall TA (1999) BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symp Ser 41:95–98
Howe AT, Bass D, Scoble JM, Lewis R, Vickerman K,Arndt H, Cavalier-Smith T (2011) Novel cultured protists iden-tify deep-branching environmental DNA clades of Cercozoa:new genera Tremula, Micrometopion, Minimassisteria, Nudifila,Peregrinia. Protist 162:332–372
Ikävalko J (1998) Further observations on flagellates withinsea ice in northern Bothnian Bay, the Baltic Sea. Polar Biol19:323–329
Karpov SA (1990) Analysis of the orders Phalansteriida, Spon-gomonadida и Thaumatomonadida. Zool Zhurn 69:5–12 (inRussian)
Karpov SA (1993) The ultrathin structure of colourless flagel-late Thaumatomonas seravini. Tsitologia 35:8–11 (in Russian)
Karpov SA (2000) Flagellate Phylogeny: UltrastructuralApproach. In Leadbeater BSC, Green JC (eds) The Flagellates.Systematics Association Spec. Publ. Taylor & Francis, London,pp 336–360
Karpov SA (2011) Order Thaumatomonadida. In Protista III.Handbook on Zoology. KMK Scientific Press. Saint Petersburg,Moscow (in Russian) in press
Karpov SA, Zhukov BF (1987) Cytological peculiarities ofcolourless flagellate Thaumatomonas lauterborni. Tsitologia29:1168–1171 (in Russian)
Kasai F, Kawachi M, Erata M, Mori F, Yumoto K, Sato M,Ishimoto M (2009) NIES-collection list of strains, 8th edition.Jap J Phycol (Sôrui) 57(Suppl.):1–350
Kühn S, Medlin L, Eller G (2004) Phylogenetic posi-tion of the parasitoid nanoflagellate Pirsonia inferred fromnuclear-encoded small subunit ribosomal DNA and a descrip-tion of Pseudopirsonia n. gen. and Pseudopirsonia mucosa(Drebes) comb. nov. Protist 155:143–156
Lara E, Heger TJ, Mitchell EA, Meisterfeld R, EkelundF (2007) SSU rRNA reveals a sequential increase in shellcomplexity among the euglyphid testate amoebae (Rhizaria:Euglyphida). Protist 158:229–237
Larkin MA, Blackshields G, Brown NP, Chenna R, McGetti-gan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, LopezR, Thompson JD, Gibson TJ, Higgins DG (2007) Clustal Wand Clustal X version 2.0. Bioinformatics 23:2947–2948
Lauterborn R (1899) Protozoen-Studien IV. Flagellaten ausdem Gebiete des Oberrheins. Zeitschr wiss Zool 65:369–391
Lepère C, Vaulot D, Scanlan DJ (2009) Photosyntheticpicoeukaryote community structure in the South East PacificOcean encompassing the most oligotrophic waters on Earth.Environ Microbiol 11:3105–3117
Lesaulnier C, Papamichail D, McCorkle S, Ollivier B, SkienaS, Taghavi S, Zak D, van der Lelie D (2008) Elevated atmo-spheric CO2 affects soil microbial diversity associated withtrembling aspen. Environ Microbiol 10:926–941
Massana R, Pernice M, Bunge JA, del Campo J (2011)Sequence diversity and novelty of natural assemblagesof picoeukaryotes from the Indian Ocean. ISME J 5:184–195
Meisterfeld R (2000) Testate Amoebae with Filopodia. In LeeJJ, Leedale GF, Bradbury P (eds) An Illustrated Guide to theProtozoa. Society of Protozoologists, Lawrence, KS, USA, pp1054–1084
Mikrjukov R (2002) Thaumatomastix tauryanini sp. n. (Pro-tista, Sarcomonadea, Thaumatomonadida) is a new species ofphagotrophic amoebo-flagellate from the White Sea sediments.Zool Zhur 81:261–265
Moestrup Ø (1979) Identification by electron microscopy ofmarine nanoplankton from New Zealand, including the descrip-tion of four new species. N Z J Bot 17:61–95
Moestrup Ø (1982) Flagellar structure in algae: a review,with new observations particularly on the Chrysophyceae,Phaeophyceae (Fucophyceae), Euglenophyceae and Recker-tia. Phycologia 21:427–528
Moestrup Ø, Sengco M (2001) Ultrastructural studies onBigelowiella natans, gen. et sp. nov., a chlorarachniophyte flag-ellate. J Phycol 37:624–646
Patterson DJ (1999) The Diversity of Eukaryotes. Am Nat154:96–124
Patterson DJ, Zölffel M (1991) Heterotrophic Flagellates ofUncertain Taxonomic Position. In Patterson DJ, Larsen J (eds)The Biology of Free-Living Heterotrophic Flagellates. Claren-don Press, Oxford, pp 427–476
Reynolds ES (1963) The use of lead citrate at high pH asan electron opaque stain in electron microscopy. J Cell Biol17:208–212
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19:1572–1574
Sandon H (1927) The Composition and Distribution of the Pro-tozoan Fauna of the Soil. Oliver and Boyd, Edinburgh
Sauvadet A-L, Gobet A, Guillou L (2010) Comparative anal-ysis between protist communities from the deep-sea pelagicecosystem and specific deep hydrothermal habitats. EnvironMicrobiol 12:2946–2964
Schmidt HA, Strimmer K, Vingron M, von Haeseler A(2002) TREE-PUZZLE: maximum likelihood phylogenetic anal-ysis using quartets and parallel computing. Bioinformatics18:502–504
Shalchian-Tabrizi K, Reier-Røberg K, Ree DK, KlavenessD, Bråte J (2010) Marine-freshwater colonizations of hapto-phytes inferred from phylogeny of environmental 18S rDNASequences. J Eukaryot Microbiol 4:1144–1153

Ultrastructure and Molecular Phylogeny of Thaumatomastix salina 573
Shimodaira H, Hasegawa M (2001) CONSEL: for assessingthe confidence of phylogenetic tree selection. Bioinformatics17:1246–1247
Stamatakis A (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixedmodels. Bioinformatics 22:2688–2690
Stoeck T, Epstein S (2003) Novel eukaryotic lineages inferredfrom small-subunit rRNA analyses of oxygen-depleted marineenvironments. Appl Environ Microbiol 69:2657–2663
Stock A, Jürgens K, Bunge J, Stoeck T (2009) Protistan diver-sity in suboxic and anoxic waters of the Gotland Deep (BalticSea) as revealed by 18S rRNA clone libraries. Aquat MicrobEcol 55:267–284
Swale EMF, Belcher JH (1974) Gyromitus disomatus Skuja– a free living colourless flagellate. Arch Protistenkd 116:211–220
Swale EMF, Belcher JH (1975) Gyromitus limax nov. sp.– free living colourless amoebo-flagellate. Arch Protistenkd117:20–26
Takahashi E, Hara S (1984) Two new marine species ofChrysosphaerella (Chrysophyceae) with a reinvestigation of C.salina. Phycologia 23:103–109
Thomsen HA (1979) Electron microscopical observations onbrackish-water nannoplankton from the Tvärminne area, SWcoast of Finland. Acta Bot Fenn 110:11–37
Thomsen HA, Ikävalko J (1997) Species of Thaumatomastix(Thaumatomastigidae, Protista incertae sedis) from the Arcticsea ice biota (North-East Water Polynya, NE Greenland). J MarSys 10:263–277
Thomsen HA, Kosman C, Ikävalko J (1995) Three newspecies of Thaumatomastix (Thaumatomastigidae, Protistaincertae sedis), a ubiquitous genus from the Antarctic ice biota.Eur J Protistol 31:174–181
Vørs N (1992a) Heterotrofe protister (ekskl. dinoflagellater,loricabærende choanoflagellater og ciliater). In Thomsen HA(ed) Plankton i de indre danske farvande. Havforskning fraMiljøstyrelsen Nr. 11, Copenhagen, pp 195–250
Vørs N (1992b) Heterotrophic amoebae, flagellates and helio-zoa from the Tvärminne area Gulf of Finland, in 1988-1990.Ophelia 36:1–109
Vørs N (1993) Heterotrophic amoebae, flagellates and heliozoafrom Arctic marine waters (North West Territories, Canada andWest Greenland). Polar Biol 13:113–126
Wylezich C, Mylnikov AP, Weitere M, Arndt H (2007)Distribution and phylogenetic relationships of freshwaterthaumatomonads with a description of the new species Thau-matomonas coloniensis n. sp. J Eukaryot Microbiol 54:347–357
Zolotarev VA, Myl’nikova ZM, Myl’nikov AP (2011) Theultrafine structure of amoeboid flagellate Thaumatomastix sp.(Thaumatomonadida (Shirkina) Karpov, 1990). Inland WaterBiol 4:287–292
Available online at www.sciencedirect.com
Related Documents











