Two Sinorhizobium meliloti glutaredoxins regulate iron metabolism and symbiotic bacteroid differentiationSofiane M. Benyamina, 1 Fabien Baldacci-Cresp, 1 Jérémy Couturier, 2 Kamel Chibani, 2 Julie Hopkins, 1 Abdelkader Bekki, 3 Philippe de Lajudie, 4 Nicolas Rouhier, 2 Jean-Pierre Jacquot, 2 Geneviève Alloing, 1 Alain Puppo 1 * and Pierre Frendo 1 1 UMR ‘Institut Sophia Agrobiotech’ INRA 1355-CNRS 7254 – Université de Nice-Sophia Antipolis, 400 routes des Chappes, 06903 Sophia-Antipolis cedex, France. 2 UMR INRA UHP 1136, Interactions Arbres Microorganismes, IFR 110 EFABA, Université de Lorraine, Faculté des Sciences, BP 70239, 54506 Vandoeuvre cedex, France. 3 Laboratoire de Biotechnologie des Rhizobiums et Amélioration des Plantes, Département de Biotechnologie, Université d’Oran Es Senia, Oran, Algeria. 4 UMR ‘Laboratoire des Symbioses Tropicales et Méditerranéennes’ IRD – Université de Montpellier, 34398 Montpellier cedex 5, France. Summary Legumes interact symbiotically with bacteria of the Rhizobiaceae to form nitrogen-fixing root nodules. We investigated the contribution of the three glutare- doxin (Grx)-encoding genes present in the Sinorhizo- bium meliloti genome to this symbiosis. SmGRX1 (CGYC active site) and SmGRX3 (CPYG) recombinant proteins displayed deglutathionylation activity in the 2-hydroethyldisulfide assay, whereas SmGRX2 (CGFS) did not. Mutation of SmGRX3 did not affect S. meliloti growth or symbiotic capacities. In contrast, SmGRX1 and SmGRX2 mutations decreased the growth of free-living bacteria and the nitrogen fixation capacity of bacteroids. Mutation of SmGRX1 led to nodule abortion and an absence of bacteroid differ- entiation, whereas SmGRX2 mutation decreased nodule development without modifying bacteroid development. The higher sensitivity of the Smgrx1 mutant strain as compared with wild-type strain to oxidative stress was associated with larger amounts of glutathionylated proteins. The Smgrx2 mutant strain displayed significantly lower levels of activity than the wild type for two iron-sulfur-containing enzymes, aconitase and succinate dehydrogenase. This lower level of activity could be associated with deregulation of the transcriptional activity of the RirA iron regulator and higher intracellular iron content. Thus, two S. meliloti Grx proteins are essential for symbiotic nitrogen fixation, playing independent roles in bacterial differentiation and the regulation of iron metabolism. Introduction Rhizobia are aerobic Gram-negative bacteria from the rhizosphere that interact with leguminous plants to develop nitrogen-fixing symbiotic nodules under condi- tions of nitrogen limitation. During this process, an exchange of recognition signals between the plant and the bacterial partners allows the bacteria to penetrate into the host plant root and triggers the development of a new nitrogen-fixing organ, the root nodule. The rhizobia then develop into intracellular symbionts, bacteroids, sur- rounded by a plant-derived membrane, the peribacteroid membrane. The resulting intracellular symbiotic structure is known as the symbiosome. Within the symbiosome, the bacteria modify their metabolism so that they can fix atmospheric nitrogen. Rhizobia involved in symbiosis with legumes of the galegoid group, such as Medicago species, differentiate irreversibly into a nitrogen-fixing form, the bacteroid, which does not survive nodule senes- cence (Mergaert et al., 2006). This bacterial differentiation process involves structural changes, endoreduplication and changes of membrane permeability (Mergaert et al., 2006). Differentiation occurs in stages, corresponding to five different types of bacteroid, from type 1 (dividing bacteria recently released from infection threads) to type 5 (senescent bacteroid no longer able to fix nitrogen) (Vasse et al., 1990). The bacterial differentiation process within the plant involves major changes in bacterial gene expression within the plant (Barnett et al., 2004; Becker et al., 2004; Capela et al., 2006). Bacterial antioxidant systems play a role in the control of symbiotic nitrogen fixation efficiency (Pauly et al., 2006; Chang et al., 2009). In the nitrogen-fixing symbiosis between Sinorhizobium meliloti and Medicago plants, Received 1 March, 2012; revised 29 June, 2012; accepted 2 July, 2012. *For correspondence. E-mail [email protected]; Tel. (+33) 492 386631; Fax (+33) 492 386587. Environmental Microbiology (2013) 15(3), 795–810 doi:10.1111/j.1462-2920.2012.02835.x © 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Two Sinorhizobium meliloti glutaredoxins regulate ironmetabolism and symbiotic bacteroid differentiationemi_2835 795..810
Sofiane M. Benyamina,1 Fabien Baldacci-Cresp,1
Jérémy Couturier,2 Kamel Chibani,2 Julie Hopkins,1
Abdelkader Bekki,3 Philippe de Lajudie,4
Nicolas Rouhier,2 Jean-Pierre Jacquot,2
Geneviève Alloing,1 Alain Puppo1* andPierre Frendo1
1UMR ‘Institut Sophia Agrobiotech’ INRA 1355-CNRS7254 – Université de Nice-Sophia Antipolis, 400 routesdes Chappes, 06903 Sophia-Antipolis cedex, France.2UMR INRA UHP 1136, Interactions ArbresMicroorganismes, IFR 110 EFABA, Université deLorraine, Faculté des Sciences, BP 70239, 54506Vandoeuvre cedex, France.3Laboratoire de Biotechnologie des Rhizobiums etAmélioration des Plantes, Département deBiotechnologie, Université d’Oran Es Senia, Oran,Algeria.4UMR ‘Laboratoire des Symbioses Tropicales etMéditerranéennes’ IRD – Université de Montpellier,34398 Montpellier cedex 5, France.
Summary
Legumes interact symbiotically with bacteria of theRhizobiaceae to form nitrogen-fixing root nodules.We investigated the contribution of the three glutare-doxin (Grx)-encoding genes present in the Sinorhizo-bium meliloti genome to this symbiosis. SmGRX1(CGYC active site) and SmGRX3 (CPYG) recombinantproteins displayed deglutathionylation activity inthe 2-hydroethyldisulfide assay, whereas SmGRX2(CGFS) did not. Mutation of SmGRX3 did not affectS. meliloti growth or symbiotic capacities. In contrast,SmGRX1 and SmGRX2 mutations decreased thegrowth of free-living bacteria and the nitrogen fixationcapacity of bacteroids. Mutation of SmGRX1 led tonodule abortion and an absence of bacteroid differ-entiation, whereas SmGRX2 mutation decreasednodule development without modifying bacteroiddevelopment. The higher sensitivity of the Smgrx1mutant strain as compared with wild-type strain tooxidative stress was associated with larger amounts
of glutathionylated proteins. The Smgrx2 mutantstrain displayed significantly lower levels of activitythan the wild type for two iron-sulfur-containingenzymes, aconitase and succinate dehydrogenase.This lower level of activity could be associated withderegulation of the transcriptional activity of the RirAiron regulator and higher intracellular iron content.Thus, two S. meliloti Grx proteins are essential forsymbiotic nitrogen fixation, playing independentroles in bacterial differentiation and the regulation ofiron metabolism.
Introduction
Rhizobia are aerobic Gram-negative bacteria from therhizosphere that interact with leguminous plants todevelop nitrogen-fixing symbiotic nodules under condi-tions of nitrogen limitation. During this process, anexchange of recognition signals between the plant andthe bacterial partners allows the bacteria to penetrate intothe host plant root and triggers the development of a newnitrogen-fixing organ, the root nodule. The rhizobia thendevelop into intracellular symbionts, bacteroids, sur-rounded by a plant-derived membrane, the peribacteroidmembrane. The resulting intracellular symbiotic structureis known as the symbiosome. Within the symbiosome, thebacteria modify their metabolism so that they can fixatmospheric nitrogen. Rhizobia involved in symbiosis withlegumes of the galegoid group, such as Medicagospecies, differentiate irreversibly into a nitrogen-fixingform, the bacteroid, which does not survive nodule senes-cence (Mergaert et al., 2006). This bacterial differentiationprocess involves structural changes, endoreduplicationand changes of membrane permeability (Mergaert et al.,2006). Differentiation occurs in stages, corresponding tofive different types of bacteroid, from type 1 (dividingbacteria recently released from infection threads) to type5 (senescent bacteroid no longer able to fix nitrogen)(Vasse et al., 1990). The bacterial differentiation processwithin the plant involves major changes in bacterial geneexpression within the plant (Barnett et al., 2004; Beckeret al., 2004; Capela et al., 2006).
Bacterial antioxidant systems play a role in the controlof symbiotic nitrogen fixation efficiency (Pauly et al., 2006;Chang et al., 2009). In the nitrogen-fixing symbiosisbetween Sinorhizobium meliloti and Medicago plants,
Received 1 March, 2012; revised 29 June, 2012; accepted 2 July,2012. *For correspondence. E-mail [email protected]; Tel. (+33) 492386631; Fax (+33) 492 386587.
bs_bs_banner
Environmental Microbiology (2013) 15(3), 795–810 doi:10.1111/j.1462-2920.2012.02835.x
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

deficiencies in catalase or glutathione (GSH) impair bac-terial differentiation and/or nitrogen fixation efficiency(Sigaud et al., 1999; Jamet et al., 2003; Hanyu et al.,2009). GSH usually represents the most abundant low-molecular-weight thiol in cells and has several well-knownfunctions. Among its known functions, it acts as a redoxbuffer and as a signalling molecule, and it is also involvedin the detoxification of radicals and xenobiotics (Rouhieret al., 2008; Foyer and Noctor, 2011). Given the variousroles attributed to GSH, the bacterial symbiotic phenotypeobserved in conditions of GSH deficiency may result fromdefects in several processes.
Glutaredoxins (Grxs) are small ubiquitous oxidoreduc-tases of the thioredoxin (Trx) superfamily that preferen-tially reduce the disulfide bonds formed between cysteineresidues of proteins and glutathione (Rouhier et al., 2008;Dalle-Donne et al., 2009; Meyer et al., 2009). On theother hand, it has been suggested that Grxs act either asscaffold proteins for the de novo synthesis of iron-sulfurclusters (ISCs) or as carrier proteins or chaperones forthe transfer and delivery of ISCs from scaffold to accep-tor apoproteins (Rodriguez-Manzaneque et al., 2002;Mühlenhoff et al., 2003; Bandyopadhyay et al., 2008).Thus, although Grxs have three-dimensional structuressimilar to those of Trxs, their biochemical characterizationindicates that they have different physiological functions.
We investigated the roles of S. meliloti GRXs in free-living bacteria and during nitrogen-fixing symbiosis, byanalysing the phenotypes of mutant strains, each lackingone of the three putative GRX genes identified in thegenome. The Smgrx3 mutant had no discernable pheno-type, but the deletion of SmGRX1 and SmGRX2 showedthat these proteins encoded by these two genes playedimportant but distinct roles in the growth of free-livingbacteria and in their symbiotic capacity.
Results
Biochemical characterization of S. meliloti glutaredoxins
A search for putative grx genes in the S. meliloti genomeled to the identification of three genes encoding proteinsdisplaying significant sequence similarity to glutaredoxins:SmGRX1 (NP_386724, SMc02443), SmGRX2 (NP_385887, SMc00538) and SmGRX3 (NP_435396,SMa0280). A phylogenetic analysis clearly revealed thatthese three proteins belong to different classes (Fig. 1).SmGRX1 (85 amino acids, 9.4 kDa) is a class I Grx with adithiol CGYC active site. SmGRX2 (111 amino acids,12 kDa) is a class II Grx, with a monothiol CGFS activesite. SmGRX3 (262 amino acids, 28.8 kDa) is an elon-gated GRX with two domains, an N-terminal Grx domainwith a CPYG active site and a C-terminal domain with amethylamine utilization protein (MauE) motif, which con-
tains five predicted transmembrane domains (Fig. 1).Class I and II Grxs are present in most organisms, butSmGRX3 orthologues, which constitute class V, are foundonly in some proteobacteria and cyanobacteria, as previ-ously described (Couturier et al., 2009). A careful inspec-tion of these Grx sequences revealed the presence of aconserved N-terminal CxxC/S/G active site and of theresidues characteristic of GSH binding sites (Fig. 1).
We assessed the functionality of these Grxs, by produc-ing the recombinant proteins in Escherichia coli. Two con-structs were generated for the production of SmGRX3.The first, encoding the putative full-length protein, did notallow the production of the recombinant protein in anystrains or culture conditions tested. The second, encodingthe Grx domain and lacking the last 165 amino acids atthe C-terminus (97 amino acids, 10.6 kDa), was success-fully expressed in E. coli. As reported for other monothioland dithiol Grxs, lysed bacterial cells producing SmGRX1and SmGRX2 were visibly brownish in colour (Rouhieret al., 2007). Moreover, the proteins separated into twopeaks in gel filtration experiments: an apoform with anapparent size corresponding to a monomer, and a holo-form, with a molecular weight corresponding to a dimerand containing an ISC. At the end of an aerobic purifica-tion, only SmGRX1 retained a UV-visible spectrumconsistent with the presence of a [2Fe-2S] cluster, high-lighting differences in cluster lability between the two pro-teins (Fig. S1). Analysis of the purified apoproteins bySDS-PAGE under reducing and non-reducing conditionsrevealed that SmGRX2 and SmGRX3 formed a smallproportion of covalent dimers, whereas the dithiol Grx,SmGrx1, did not, most likely due to the capacity of the twocysteines of the active site to form an intramolecular dis-ulfide bond (Fig. S2). While it gives us some indicationson the catalytic and redox properties of these Grxs, thephysiological significance of this finding is still unclear. Weassessed the reductase activity of each SmGrx isoform,using 2-hydroethyldisulfide (HED) as a substrate in thepresence of a GSH-reducing system (Fig. 2). UnlikeSmGRX2, SmGRX1 and SmGRX3 reduced the b-ME-SGmixed disulfide formed between HED and GSH during theinitial pre-incubation period of the reaction. This reductionwas more efficient with SmGRX1 than SmGRX3 (kcat of11.90 s-1 and 0.94 s-1 respectively).
Grx deficiencies affect the growth offree-living S. meliloti
We investigated the contribution of the various Grxs to thegrowth of free-living S. meliloti, by generating mutantstrains lacking SmGRX1, SmGRX2 and SmGRX3,named Smgrx1, Smgrx2 and Smgrx3, respectively, fromthe wild-type (wt) S. meliloti Rm2011 strain. We checkedthat the corresponding proteins were indeed absent, by
796 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

Western blot analysis of protein extracts of the mutantstrains (Fig. S3). SmGRX1 and SmGRX2 were detectedin the soluble fraction of protein extracts of the wt Rm2011strain, whereas SmGRX3 was detected in the cellularpellet, suggesting that SmGRX3 is a periplasmic ormembrane-bound protein.
The growth curves of the wt S. meliloti Rm2011 strainand the Smgrx1, Smgrx2 and Smgrx3 mutant strains areshown in Fig. 3A. Smgrx3 grew at a rate not significantlydifferent from that of the wt strain. In contrast, both theSmgrx1 and Smgrx2 mutant strains grew more slowly,Smgrx1 being the most severely affected. Thus, SmGRX1and SmGRX2 are important for the growth of free-livingS. meliloti.
We investigated the importance of the three Grxs forprotection against oxidative stress, by carrying out diskdiffusion assays to determine the resistance of themutants to hydrogen peroxide (H2O2) (Fig. 3B). Thegrowth of Smgrx2 and Smgrx3 was not significantlyaffected by H2O2 treatment, as compared with the wtRm2011 strain, whereas the growth of Smgrx1 was sig-
nificantly inhibited by this treatment. Thus, SmGRX1may play a role in protecting against oxidative stress inS. meliloti.
SmGRX1 is involved in deglutathionylation reactions
Grxs are involved in the regulation of the cellular redoxstate, in particular through the control of protein glutathio-nylation state. We investigated the role of the three S. me-liloti Grxs in these mechanisms, by quantifying free andprotein-associated GSH in the three mutant strainsgrowing in free-living conditions. The protein-associatedGSH fraction was rendered accessible by reducing theprotein pellet with borohydride after the removal of freeGSH (Greetham et al., 2010). No significant difference intotal and reduced free GSH content was observedbetween the mutant strains and the wt Rm2011 strain(Fig. 4A). Protein-bound GSH was significantly higher inthe Smgrx1 strain than in the Smgrx2, Smgrx3 and wtRm2011 strains (Fig. 4B). We also analysed the expres-sion of two stress-responsive genes (Sigaud et al., 1999),
Fig. 1. Unrooted, neighbour-joining (NJ) tree of the Grx family in a-proteobacteria. The analysis was performed with MEGA 4 software, asdescribed in Experimental procedures. Branch lengths are proportional to phylogenetic distances. For clarity, the protein names have beenremoved. The typical domain organization has been indicated for each Grx class. The sequence logos of active site and glutathione bindingsite motifs were generated with Weblogo (http://weblogo.berkeley.edu/) (Crooks et al., 2004).
Roles of glutaredoxins in S. meliloti 797
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

encoding the KatA and KatC catalases, in the Smgrxmutant strains (Fig. 4C). Some variability was observed,but only the expression of katC was significantly upregu-lated in the Smgrx2 mutant strain and the levels of varia-tion were lower than those observed in S. meliloti mutantstrains lacking GSH (Harrison et al., 2005). These resultssuggest that, in our standard growth conditions, theSmgrx mutant strains were not under significant oxidativestress that might significantly modify GSH content or
redox state and catalase gene expression. However,these results also indicate that the dithiol enzymeSmGRX1 is one of the proteins catalysing protein deglu-tathionylation in S. meliloti.
SmGRX2 deficiency affects iron homeostasis
Grxs are known to be involved in the regulation of ironmetabolism in multiple organisms, either through a role inISC assembly machineries or through the regulation ofiron-responsive transcription factors (Haas et al., 2008;Kaplan and Kaplan, 2009; Mercier and Labbe, 2009;Mühlenhoff et al., 2010; Rouhier et al., 2010). Their role inISC assembly machineries was investigated by determin-
Fig. 2. GSH-dependent reductase activity of SmGrxs.A and B. The dependence of HED reduction on SmGrx1 (A) andSmGrx3 (B) concentration was determined in the presence of 2 mMGSH and 0.7 mM HED.C. Comparison of the reductase activity of the three SmGRXs. Theassay was carried out with 100 nM SmGRX1, and 200 nMSmGRX2 or SmGRX3, in the presence of 2 mM GSH and 0.7 mMHED.The data reported are the means � SD of three independentexperiments.
Fig. 3. Growth of S. meliloti Smgrx mutant strains.A. Growth curves of S. meliloti strains. At time zero, rhizobial cellswere diluted with minimal medium (M9) to an OD600 of 0.1. Wemonitored the growth of the wt Rm2011 strain and of the Smgrx1,Smgrx2 and Smgrx3 mutant strains, by measuring optical densityat 600 nm. The data shown are the means of three experiments.B. Inhibition of the growth of the Rm2011 (2011), Smgrx1 (Grx1),Smgrx2 (Grx2) and Smgrx3 (Grx3) strains under oxidative stress.The data shown are the means � SEM of six independentexperiments. The asterisk (*) indicates a significant difference(P � 0.05).
798 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

ing, in the free-living grx mutant strains, the activity of twoISC-containing enzymes, succinate dehydrogenase andaconitase, and, as a control, two enzymes with no ISC,malate dehydrogenase (MDH) and isocitrate dehydroge-nase (ICDH) (Fig. 5A). No significant difference in MDH
and ICDH activity levels was observed between the grxmutant strains and the wt Rm2011 strain. Succinate dehy-drogenase and aconitase activity levels were significantlylower than in the wild type only in the Smgrx2 mutantstrain, although a non-significant decrease was alsoobserved in the Smgrx1 mutant strain.
The role of Grxs in iron homeostasis was investigatedby assessing the expression of iron-regulated genes. Theregulation of iron homeostasis involves two major tran-scription factors in rhizobia: Iron Responsive Regulator (irrgene) and Rhizobium Iron Responsive A (rirA) (Johnstonet al., 2007). It has been suggested that RirA contains anISC cofactor governing its capacity to downregulate genetranscription (Johnston et al., 2007). The expression ofrhtA, hmuS and rhbA, three genes regulated by RirA andencoding proteins involved in iron transport (Chao et al.,2005), was analysed by quantitative reverse transcription-polymerase chain reaction (RT-qPCR) in the wt Rm2011and Smgrx1, Smgrx2 and Smgrx3 mutant strains(Fig. 5B). No significant difference in expression wasobserved between Smgrx1 or Smgrx3 and Rm2011. Incontrast, rhtA, hmuS and rhbA were significantly overex-pressed in Smgrx2 compared with wt Rm2011.
The disruption of genes encoding class II Grxs in Sac-charomyces cerevisiae (Grx3 to 5) leads to the accumu-lation of iron in the cells (Mühlenhoff et al., 2003; 2010).We therefore determined total iron content in the Smgrxmutant strains (Fig. 5C). No significant difference in ironcontent was observed between the Smgrx1 and Smgrx3mutant strains and the wt Rm2011 strain. In contrast, theiron content was significantly higher in the Smgrx2 strainthan in wt Rm2011.
Thus, SmGRX2 is involved in the regulation of ironhomeostasis, either through the direct or indirect modula-tion of RirA or through a role as a scaffold or carrierprotein in ISC assembly machineries.
Deficiencies of SmGRX1 and 2 affect the nodulationprocess and nitrogen fixation efficiency
We investigated the role of these Grxs in the symbioticprocess, by assessing the capacity of the Smgrx1,Smgrx2 and Smgrx3 mutant strains to produce nitrogen-
Fig. 4. Quantification of GSH in Smgrx mutant strains.A. Total and reduced GSH contents were determined in thewild-type Rm2011 strain and the Smgrx1, Smgrx2 and Smgrx3mutant strains.B. The protein-bound GSH content was determined for the wtRm2011 strain and the Smgrx1, Smgrx2 and Smgrx3 mutantstrains.C. Levels of katA and katC expression in the S. meliloti wild-typeRm2011 (2011) strain and in the Smgrx1 (Grx1), Smgrx2 (Grx2)and Smgrx3 (Grx3) strains.The data shown are the means of three experiments � SEM. Theasterisk (*) indicates a significant difference (P � 0.05).
Roles of glutaredoxins in S. meliloti 799
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

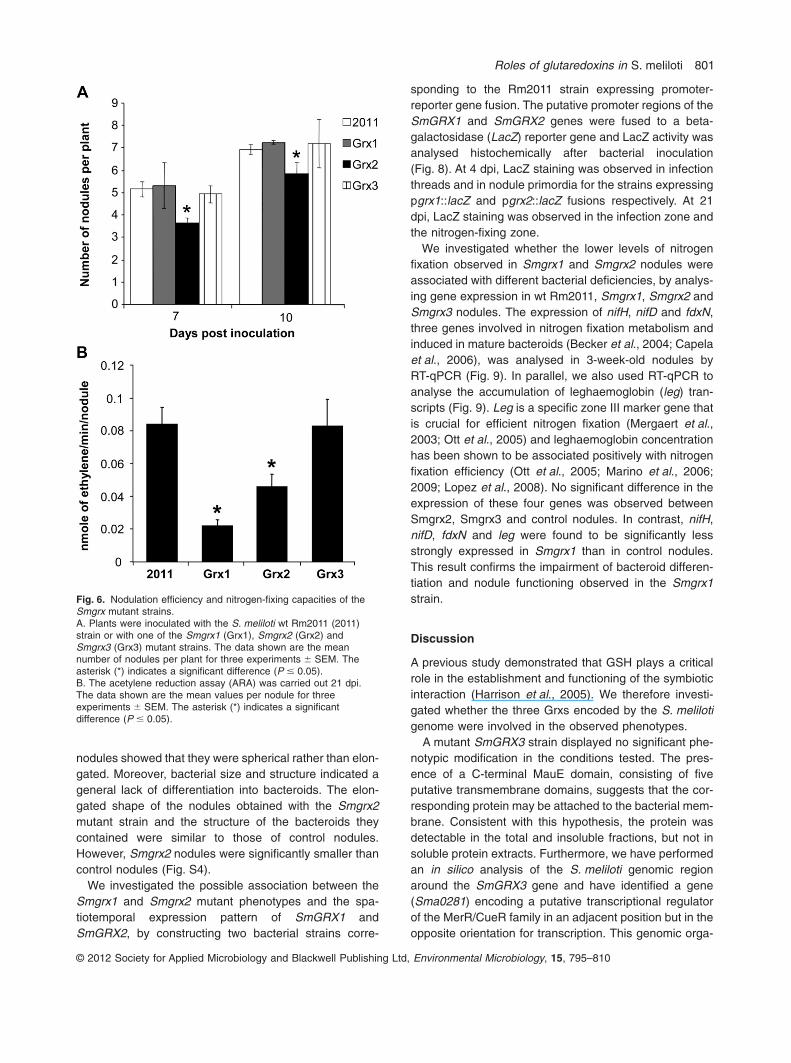
fixing nodules after inoculation with Medicago truncatula(Fig. 6). The nodulation efficiency of the mutant strainswas measured 7 and 10 days post inoculation (dpi)(Fig. 6A). No significant difference in the number ofnodules was observed between plants inoculated with theSmgrx1 and Smgrx3 strains and plants inoculated withthe wt Rm2011 strain. In contrast, significantly fewernodules were observed on plants inoculated with theS. meliloti Smgrx2 strain than on those inoculated withRm2011. We assessed nodule functionality, by determin-ing nitrogen-fixing capacity at 21 dpi, using the acetylenereduction assay (Fig. 6B). We observed no significant dif-ference in nitrogen fixation capacity between plants inocu-lated with the Smgrx3 mutant strain and plants inoculatedwith the Rm2011 strain. In contrast, plants inoculated with
Smgrx1 or Smgrx2 had a significantly lower nitrogen fixa-tion capacity than control plants. Moreover, a significantdifference was also observed between plants inoculatedwith Smgrx1 and plants inoculated with Smgrx2. Thus,both SmGRX1 and SmGRX2 are important for nitrogenfixation efficiency and SmGRX2 deletion affects the effi-ciency of nodule formation.
We investigated the effect of GRX deletion on nodulestructure by microscopic analyses of nodules at 21 dpi(Fig. 7). The wt Rm2011 nodule contained all the usualnodule zones: meristem (zone I), infection zone (zone II)and nitrogen-fixing zone (zone III). Infection with theSmgrx3 mutant strain resulted in a similar nodule struc-ture. Moreover, the bacteroids in zone III were similar tothose of the control. Microscopic analysis of Smgrx1
Fig. 5. Analysis of iron metabolism in S. meliloti Smgrx mutant strains.A. Relative enzyme activities of Fe-S cluster-containing and Fe-S cluster-free proteins. The activities of two Fe-S proteins (succinatedehydrogenase and aconitase) and two non-Fe-S proteins (malate dehydrogenase and isocitrate dehydrogenase) were determined in lysatesof cells of the wt Rm2011 (2011), Smgrx1 (Grx1), Smgrx2 (Grx2) and Smgrx3 (Grx3) strains.B. Expression level of rthA, hmuS and rhbA in the S. meliloti wt Rm2011 strain and in the Smgrx1 (Grx1), Smgrx2 (Grx2) and Smgrx3 (Grx3)mutant strains. The expression levels recorded for the Rm2011 strain were taken as 1.C. Iron content was determined in cells of the S. meliloti wt Rm2011 (2011) strain and the Smgrx1 (Grx1), Smgrx2 (Grx2) and Smgrx3 (Grx3)mutant strains.The data shown are the means of three experiments � SEM. The asterisk (*) indicates a significant difference (P � 0.05).
800 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810
nodules showed that they were spherical rather than elon-gated. Moreover, bacterial size and structure indicated ageneral lack of differentiation into bacteroids. The elon-gated shape of the nodules obtained with the Smgrx2mutant strain and the structure of the bacteroids theycontained were similar to those of control nodules.However, Smgrx2 nodules were significantly smaller thancontrol nodules (Fig. S4).
We investigated the possible association between theSmgrx1 and Smgrx2 mutant phenotypes and the spa-tiotemporal expression pattern of SmGRX1 andSmGRX2, by constructing two bacterial strains corre-
sponding to the Rm2011 strain expressing promoter-reporter gene fusion. The putative promoter regions of theSmGRX1 and SmGRX2 genes were fused to a beta-galactosidase (LacZ) reporter gene and LacZ activity wasanalysed histochemically after bacterial inoculation(Fig. 8). At 4 dpi, LacZ staining was observed in infectionthreads and in nodule primordia for the strains expressingpgrx1::lacZ and pgrx2::lacZ fusions respectively. At 21dpi, LacZ staining was observed in the infection zone andthe nitrogen-fixing zone.
We investigated whether the lower levels of nitrogenfixation observed in Smgrx1 and Smgrx2 nodules wereassociated with different bacterial deficiencies, by analys-ing gene expression in wt Rm2011, Smgrx1, Smgrx2 andSmgrx3 nodules. The expression of nifH, nifD and fdxN,three genes involved in nitrogen fixation metabolism andinduced in mature bacteroids (Becker et al., 2004; Capelaet al., 2006), was analysed in 3-week-old nodules byRT-qPCR (Fig. 9). In parallel, we also used RT-qPCR toanalyse the accumulation of leghaemoglobin (leg) tran-scripts (Fig. 9). Leg is a specific zone III marker gene thatis crucial for efficient nitrogen fixation (Mergaert et al.,2003; Ott et al., 2005) and leghaemoglobin concentrationhas been shown to be associated positively with nitrogenfixation efficiency (Ott et al., 2005; Marino et al., 2006;2009; Lopez et al., 2008). No significant difference in theexpression of these four genes was observed betweenSmgrx2, Smgrx3 and control nodules. In contrast, nifH,nifD, fdxN and leg were found to be significantly lessstrongly expressed in Smgrx1 than in control nodules.This result confirms the impairment of bacteroid differen-tiation and nodule functioning observed in the Smgrx1strain.
Discussion
A previous study demonstrated that GSH plays a criticalrole in the establishment and functioning of the symbioticinteraction (Harrison et al., 2005). We therefore investi-gated whether the three Grxs encoded by the S. melilotigenome were involved in the observed phenotypes.
A mutant SmGRX3 strain displayed no significant phe-notypic modification in the conditions tested. The pres-ence of a C-terminal MauE domain, consisting of fiveputative transmembrane domains, suggests that the cor-responding protein may be attached to the bacterial mem-brane. Consistent with this hypothesis, the protein wasdetectable in the total and insoluble fractions, but not insoluble protein extracts. Furthermore, we have performedan in silico analysis of the S. meliloti genomic regionaround the SmGRX3 gene and have identified a gene(Sma0281) encoding a putative transcriptional regulatorof the MerR/CueR family in an adjacent position but in theopposite orientation for transcription. This genomic orga-
Fig. 6. Nodulation efficiency and nitrogen-fixing capacities of theSmgrx mutant strains.A. Plants were inoculated with the S. meliloti wt Rm2011 (2011)strain or with one of the Smgrx1 (Grx1), Smgrx2 (Grx2) andSmgrx3 (Grx3) mutant strains. The data shown are the meannumber of nodules per plant for three experiments � SEM. Theasterisk (*) indicates a significant difference (P � 0.05).B. The acetylene reduction assay (ARA) was carried out 21 dpi.The data shown are the mean values per nodule for threeexperiments � SEM. The asterisk (*) indicates a significantdifference (P � 0.05).
Roles of glutaredoxins in S. meliloti 801
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

nization is conserved in several cyanobacteria andproteobacteria, suggesting that SmGRX3 may protectbacteria against heavy metals. Nevertheless, in theabsence of phenotype for the mutant strain, the possibleroles of SmGRX3 will not be discussed further in thecontext of this study.
Unlike the SmGRX3 mutation, mutations of SmGRX1and SmGRX2 affected both the growth of free-living bac-teria and the capacity to promote nitrogen-fixing symbio-sis with M. truncatula. The observation of differentdefective phenotypes for the two single mutants indicatesthat the functions of these two Grxs are not identical in the
processes studied. Moreover, the detection of defectivephenotypes also indicates that these functions are notcompensated by the Trx-reducing system which is com-posed of a putative thioredoxin reductase SmTRXB andtwo putative thioredoxins SmTRXA and SmTRXC. TheTrx and Grx systems are therefore not redundant inthis particular case, probably because the phenotypesobserved are linked to specific Grx functions, i.e. thecapacity to perform deglutathionylation reactions and/orto bind Fe-S clusters. On the contrary, the lack of pheno-types for most Grx mutants and the fact that E. colimutants combining mutations affecting the three dithiol
Fig. 7. Structure of 3-week-old M. truncatula nodules and bacteroids. Nodules were induced by S. meliloti Rm2011 (A, E, I), Smgrx1 (B, F, J),Smgrx2 (C, G, K) or Smgrx3 (D, H, L). Spatial development zones (I, II and III) are indicated in nodule cross-sections (A). Scalebars = 100 mM (A, D, G, J), 10 mM (B, E, H, K) and 5 mM (C, F, I, L).
802 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

Grxs (GRX1, 2 and 3) and the two Trxs (TRX1 and 2)were viable and very often aphenotypic led to the propo-sition that these two pathways are largely redundant inthis bacterium (Lillig et al., 2008; Meyer et al., 2009). Onlya double mutant lacking thioredoxin reductase (trxB) andglutathione reductase (gor) is lethal under standard con-ditions, and this phenotype can be rescued by dithiothrei-tol, an artificial reductant (Faulkner et al., 2008). Thesituation is less clear-cut for the monothiol enzyme GRX4,the orthologue of SmGRX2, because mutation of the geneencoding this enzyme, which was initially thought to belethal, is actually not lethal unless other mutations arealso present (Butland et al., 2008). Thus, SmGRX1 andSmGRX2 seem to have more specific roles in S. melilotithan the corresponding enzymes in E. coli.
SmGRX1 is involved in protein deglutathionylation andis important for bacterial differentiation into bacteroids
Dithiol Grxs are known to be involved in the regulation ofredox homeostasis, through their oxidoreductase activity,but they may also act as redox sensors by binding an ISC,as suggested for human Grx2 (Lillig et al., 2005; Gallogly
Fig. 8. Histochemical localization of bacterialgrx1 and grx2 gene expression duringS. meliloti–M. truncatula nodule development.b-Galactosidase activity was assessed, withX-gal used as the substrate for (A and D)S. meliloti wt Rm2011; (B and E) grx1::lacZ;(C and F) grx2::lacZ in nodule primordia andinfection threads, 4 dpi (A through C) and in21-day-old nodules (D through F). Theasterisk (*) indicates nodule primordia. Spatialdevelopment zones (I, II and III) are indicatedon nodule cross-sections. Scale bars = 50 mM(4 dpi) and 100 mM (21 dpi).
Fig. 9. RT-qPCR analysis of the expression of genes involved innitrogen fixation. The levels of expression of nifH, nifD and fdxNwere measured in 3-week-old nodules induced by the S. melilotiRm2011, Smgrx1 (Grx1), Smgrx2 (Grx2) and Smgrx3 (Grx3)strains. Expression levels in control nodules induced with theS. meliloti wt strain Rm2011 were set at 1. Expression analysis wasperformed in triplicate for each experiment, and the mean relativeexpression levels for three independent experiments are presented.Error bars indicate the SEM. The asterisk (*) indicates a significantdifference (P � 0.05).
Roles of glutaredoxins in S. meliloti 803
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

et al., 2009). The phenotypes observed for the Smgrx1mutant strain demonstrate that SmGrx1, the only dithiolGrx in S. meliloti, plays a very important role in bacterialgrowth, bacteroid differentiation and oxidative stressresponse. However, it remains unclear which of the twofunctions described above is important for which pheno-type. Indeed, the observed capacity of SmGRX1 to bind anISC when produced as a recombinant protein in E. coliindicates that it may modulate protein activity or conferadditional functions to the protein. However, SmGRX1 hasa highly efficient thioltransferase activity, with a kcat of11.9 s-1 in the HED assay. This value is within the rangereported for the other prokaryote and eukaryote dithiolGrxs characterized to date (Lillig et al., 2005; Couturieret al., 2011). The deletion of SmGRX1 seems to have nosignificant effect on the redox balance in free-living bacte-ria, as no change was observed in GSH content and redoxstate or in the expression of katA and katC in the Smgrx1mutant strain. However, the higher protein-bound GSHcontent of this mutant suggests that this protein plays akey role in protein deglutathionylation. This may accountfor the higher sensitivity of this mutant strain to an oxida-tive stress generated by exogenous H2O2. Several lines ofevidence suggest that the establishment of symbiosis, andparticularly bacterial differentiation, is a stressful process.Bacterial differentiation is induced by nodule cysteine-rich(NCR) peptides, which resemble defensins and have anti-microbial activity (Van de Velde et al., 2010; Haag et al.,2011). Given the lack of bacterial differentiation in theSmgrx1 mutant, it is possible that SmGRX1 reacts directlywith these peptides to regulate their activity, as demon-strated for human defensins with Trx (Schroeder et al.,2011). Alternatively, the differentiation deficiency may belinked to the lower fitness of the Smgrx1 mutant.
SmGRX2 has iron-related functions
In eukaryotes, some organellar class II Grxs with a strictlyconserved monothiol CGFS active site have been shownto be involved in ISC biogenesis (Rodriguez-Manzanequeet al., 2002; Mühlenhoff et al., 2003; Ye and Rouault,2010). The results of yeast complementation experimentssuggest that this role is probably conserved in all king-doms (Molina-Navarro et al., 2006; Bandyopadhyay et al.,2008). However, several fungal cytosolic class II Grxshave been shown to regulate iron metabolism by modu-lating the activity of transcription factors, such as Aft1,Php4 and Fep1, through a physical interaction (Pujol-Carrion et al., 2006; Kumanovics et al., 2008; Mercier andLabbe, 2010). In the specific case of S. cerevisiae, Aft1has been shown to form a complex with Grx3/4 and atleast two other proteins: FRA1, an aminopeptidase P-likeprotein, and FRA2, a BolA-like protein (Kumanovics et al.,2008). In many bacteria, such as E. coli, iron homeostasis
is controlled by the Fur regulator (Lee and Helmann,2007). However, other bacteria as Rhizobiaceae haveother transcriptional regulators, such as Irr or RirA(Johnston et al., 2007). Sinorhizobium meliloti has a Furhomologue, but this protein is involved in the regulation ofthe Mn2+ transporter operon sitABCD (Chao et al., 2004).Our results demonstrate that, in this bacterium, a unicel-lular organism with a single major subcellular compart-ment, the cytosol, a single monothiol Grx may fulfil bothfunctions initially described in fungi. Indeed, the low levelsof activity of two iron-sulfur proteins, aconitase and suc-cinate dehydrogenase, in the Smgrx2 mutant strain andthe incorporation of a labile ISC by the Smgrx2 proteinstrongly suggest that this protein may act as a scaffoldprotein or as carrier protein, as suggested for yeast Grx5(Mühlenhoff et al., 2003). The defect in the ISC assemblymachinery may be general, because nitrogenase activityis significantly weaker in this mutant than in the wild type.One of the specificities of the biological nitrogen fixation isthe presence of nitrogenase. Nitrogenase is composed ofa Fe protein containing an Fe-S cluster and a molybde-num (Mo)-Fe protein which contains the P cluster (8Fe-7S) and the FeMo cofactor (7 Fe-1Mo-9S-X-homocitrate) (Seefeldt et al., 2009), with X being mostprobably a carbon (Spatzal et al., 2011). Thus, ironmetabolism and Fe-S cluster biogenesis are particularlyimportant during the nitrogen fixing symbiosis.
The significant upregulation of rhtA, hmuS and rhbA,three genes negatively regulated by RirA, which is pre-sumably itself an iron-sulfur protein (Chao et al., 2005), inthe Smgrx2 mutant suggests that SmGRX2 is involved inregulating RirA. In addition, many prokaryotes, includingmost of the Rhizobiaceae, have a gene encoding a puta-tive BolA protein (SMc00487 in the S. meliloti genome)upstream from the SmGRX2 gene. Considering the exist-ence of a Grx-BolA-dependent regulation of Aft1 in S. cer-evisiae, it could be hypothesized that BolA participates tothe SmGRX2 regulation of activity. Incidentally, a doublegrx4-bolA mutant has been shown to be lethal in E. coli(Butland et al., 2008). Finally, the iron content of theSmgrx2 mutant strain is significantly higher than that ofthe wild type (about double). However, it is not possible toclearly attribute the origin of this increase to one or otherof the functions (iron sensing or Fe-S cluster biogenesis),because observations in yeast suggest that both types ofderegulation may lead to such an accumulation (Mühlen-hoff et al., 2003; 2010).
Grx4 transcript levels in E. coli were previously shownto be regulated by iron depletion in a previous study(Fernandes et al., 2005), but this study provides the firstdemonstration that Grx participates in the assembly ofISC and most likely to the regulation of iron homeostasisin prokaryotes, most previous observations having beencarried out in fungi.
804 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

Grx mutations do not produce phenotypes similar tothose of gsh mutants
The SmgshA mutant strain, which contains no detectableg-glutamylcysteine (gEC) or GSH, is unable to producenodules (Harrison et al., 2005). The SmgshB mutant,which produces the intermediate compound g-glutamylcysteine, displays both delayed infection andearly senescence of the bacteroids (Harrison et al., 2005).This phenotype, which is associated with the deregulationof plant carbon metabolism and the formation of hypertro-phied amyloplasts in the nitrogen-fixing zone, was notobserved with the Smgrx1 and Smgrx2 mutant strains.The Smgrx2 mutant strain had a less pronounced pheno-type than the SmgshB mutant strain. Indeed, nitrogenfixation was more efficient in the Smgrx2 mutant strainthan in the SmgshB mutant strain, and no early senes-cence phenotype was detected for the Smgrx2 mutantstrain. In contrast, the Smgrx1 mutant strain had a moresevere phenotype than the SmgshB mutant strain, as itwas unable to differentiate into bacteroids. SmGrx1 dis-plays a GSH-dependent activity. Thus, in the SmgshBmutant strain, SmGrx1 may perform some of its functionswith g-glutamylcysteine (gEC) as an alternative reductant,because this compound accumulated in large amounts inthis mutant. The use of gEC-reducing power has alreadybeen reported in some E. coli mutant strains (Faulkneret al., 2008). However, we cannot exclude the possibilitythat the phenotype observed in the SmgshB mutant strainwas not linked to Grxs.
In conclusion, SmGRX1 and SmGRX2 are criticalproteins for optimal development of the nitrogen-fixingsymbiosis, but they seem to play different roles, in deglu-tathionylation reactions and in the regulation of ironmetabolism respectively. An analysis of Fe-S andFe-molybdenum cluster metabolism during nitrogen-fixingsymbiosis might shed light on the role of SmGrx2 in nitro-genase complex assembly. In parallel, an analysis ofSmGrx1 protein targets should help to identify the pro-teins involved in bacterial differentiation with functionsregulated by SmGrx1.
Experimental procedures
Bacterial strains and growth conditions
The strains and plasmids used in this work are indicated inTable S1. Escherichia coli strains were grown at 37°C inLuria–Bertani (LB) medium, and S. meliloti strains weregrown at 30°C in LB medium supplemented with 2.5 mMMgSO4 and 2.5 mM CaCl2 (LBMC) or in M9 minimal medium.Antibiotics for the selection of E. coli strains were used at theindicated concentrations: ampicillin, 100 mg ml-1; chloram-phenicol, 10 mg ml-1; kanamycin, 25 mg ml-1; streptomycin,100 mg ml-1; tetracycline, 10 mg ml-1; and gentamicin, 10 mgml-1. For the selection of S. meliloti strains, antibiotics were
used at the indicated concentrations: streptomycin, 200 mgml-1; neomycin, 200 mg ml-1; tetracycline, 20 mg ml-1.
The H2O2 resistance assay was performed with grxmutants by the disk diffusion method, with LBMC agar andpaper disks imbibed with 50 mM H2O2. We spotted 100 ml ofeach bacterial suspension (OD = 2.5) onto LBMC agar. TheH2O2 disks were laid onto the agar so that they came intocontact with the bacteria. The plates were incubated at 28°Cuntil a circular clear zone, in which bacterial growth wasinhibited, surrounded the paper disk. The diameter of thezone of inhibition was measured for each mutant and com-pared with that for the wt Rm2011 strain, to provide an esti-mate of susceptibility to oxidants.
Identification of glutaredoxins from S. meliloti andphylogenetic analyses
The amino acid sequences of the SmGRX1 (NP_386724,SMc02443), SmGRX2 (NP_385887, SMc00538) andSmGRX3 (NP_435396, SMa0280) proteins were retrievedfrom the full S. meliloti proteome available from GenBank(February 2009, 6218 sequences), with the HMMER programand the Pfam hidden Markov model (HMM) profile corre-sponding to Grx (PF00462.16) (Eddy, 1998; Finn et al.,2009). These sequences were used as queries for the iden-tification of homologues from other a-proteobacteria withBLASTP (http://blast.ncbi.nlm.nih.gov/). CLUSTALW was used toalign the amino acid sequences, which were then importedinto the Molecular Evolutionary Genetics Analysis (MEGA)package version 4.1 (Tamura et al., 2007). Phylogeneticanalyses were conducted by the neighbour-joining (NJ)method, implemented in MEGA, with the pair-wise deletionoption for handling alignment gaps and the Poisson correc-tion model for distance calculation. Bootstrap tests were con-ducted with 1000 replicates. Branch lengths are proportionalto phylogenetic distances.
Cloning of grx genes for recombinant protein production
The sequences encoding SmGRX1, SmGRX2 and SmGRX3were amplified from genomic regions and inserted betweenthe NcoI and BamHI restriction sites of pET3d (Novagen,Lyons), with the primers listed in Table S2. For SmGRX3, afull-length and a shortened DNA sequence were amplifiedand inserted in pET3d. The shortened sequence encoded aprotein lacking the last 165 amino acids corresponding tothe C-terminal domain with five putative transmembranedomains, to which an alanine residue was added in thesecond position by cloning. This protein thus began with theN-terminal sequence 1MALDRTIP8 and ended with a 89PVAD-PKATT97 sequence at the C-terminus.
Production and purification of the recombinant proteins
For protein production, E. coli BL21(DE3), containing thepSBET plasmid, was co-transformed with the various recom-binant plasmids (Schenk et al., 1995). Cultures were succes-sively amplified up to 2.4 l in LB medium supplemented withampicillin and kanamycin (50 mg ml-1), at 37°C. Protein
Roles of glutaredoxins in S. meliloti 805
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

production was induced in the exponential growth phasefor 4 h at 37°C by adding 100 mM isopropyl b-D-thiogalactopyranoside. The cultures were then centrifuged for15 min at 4400 g. In the case of SmGRX3, the cells wereresuspended in 30 ml of buffer A (30 mM Tris-HCl pH 8.0,1 mM EDTA, 200 mM NaCl). For SmGRX1 and SmGRX2,the cells were resuspended in buffer B (30 mM Tris-HClpH 8.0, 200 mM NaCl). All the cells were finally stored at-20°C.
Cells were lysed by sonication (3 ¥ 1 min pulses, separatedby 1 min) and the soluble and insoluble fractions were sepa-rated by centrifugation for 30 min at 27 000 g. The solublefraction was then further fractionated with ammonium sulfate,in two steps. The fraction precipitating between 40% and 80%of the saturation contained the recombinant protein, asshown by SDS-PAGE. SmGRXs were purified by size exclu-sion chromatography after loading on an ACA44 (5 ¥ 75 cm)column equilibrated in buffer A for SmGRX3, or buffer B forSmGRX1 and SmGRX2. The fractions containing the proteinwere pooled, dialysed by ultrafiltration to remove NaCl andloaded onto a DEAE (diethylaminoethyl) cellulose column(Sigma) equilibrated in buffer C (30 mM Tris-HCl pH 8.0,1 mM EDTA) for SmGRX3 or buffer D (30 mM Tris-HClpH 8.0) for SmGRX1 and SmGRX2. SmGRX3 was passedthrough the DEAE column, concentrated by ultrafiltrationunder nitrogen pressure (YM10 membrane, Amicon) andstored in the same buffers at -20°C. SmGRX1 and SmGRX2were eluted with a gradient of 0 to 0.4 M NaCl, dialysedagain, concentrated and stored at -20°C.
Protein purity was checked by SDS-PAGE and proteinconcentrations were determined spectrophotometri-cally, using molar extinction coefficients at 280 nm of6085 M-1 cm-1 for SmGRX1, 11 585 M-1 cm-1 for SmGRX2and 5960 M-1 cm-1 for the shortened SmGRX3.
Thioltransferase activity (HED assay)
We assessed 2-hydroethyldisulfide (HED) reduction at 25°Cin steady-state conditions, by following NADPH oxidation at340 nm in the presence of a Grx reducing system, as previ-ously described (Couturier et al., 2011). After subtracting theactivity observed in the absence of Grxs, activity is expressedin nmol of NADPH oxidized per nmol GRX per second, usinga molar extinction coefficient of 6220 M-1 cm-1 at 340 nm forNADPH.
Antibody production and Western blot analysis
Polyclonal antibodies were raised against the recombinantproteins in rabbit (genecust). The analysis of the specificity ofthe different sera indicated that there was no cross-reactionsince they did not recognize 50 ng of the other recombinantGRXs. Proteins extracted from the different bacterial strains(30 mg) were separated using SDS-PAGE (14%, w/v gel) andelectrotransferred onto a nitrocellulose membrane. The seraraised against SmGRX1, SmGRX2 and SmGRX3 were usedat a dilution of 1:1000. Bound antibodies were revealed usingan anti-rabbit immunoglobulin G alkaline phosphatase conju-gate diluted 1:20 000 (Sigma).
Construction of the S. meliloti glutaredoxin mutants
For Smgrx1 and Smgrx2 mutants, the entire SmGRX1 andSmGRX2 coding sequences (SMc02443 and SMc00538respectively) were deleted after amplifying the chromosomalregions flanking SMc00538 and SMc02443 by PCR with theprimers listed in Table S2. PCR fragments corresponding tothe upstream and downstream flanking regions of SMc02443were generated with the primer pairs SMc02443-F1/SMc02443-R1 and SMc02443-F2/SMc02443-R2 respec-tively. PCR fragments corresponding to the upstream anddownstream flanking regions of SMc00538 were generatedwith the primer pairs SMc00538-F1/SMc00538-R1 andSMc00538-F2/SMc00538-R2 respectively.
For each gene, the PCR products corresponding to theupstream and downstream flanking regions were in pGEM-T.Then, upstream and downstream flanking regions of the openreading frame were ligated together in the pGEM-T allowingthe creation of a BamHI restriction site between the twoflanking region. The lacZ-kanamycin resistance cartridgefrom pKOK5 (Kokotek and Lotz, 1989) was then inserted intothe BamHI site, replacing the missing GRX open readingframe. Fragments carrying the lacZ-kanamycin resistancecartridge flanked by the SMc02443- or the SMc00538-flanking regions were then inserted into the NcoI site of thesuicide plasmid pSUP202 (Simon et al., 1983).
For the S. meliloti Smgrx3 mutant, the SmGRX3(Sma0280) gene was disrupted, by amplifying a 528 bp inter-nal fragment of the Sma 280 coding region by PCR with theSma0280-F and Sma0280-R primers. The PCR product wasinserted into pGEM-T and then subcloned between the NcoIand EcoRI sites of pSUP202 to generate pSUP202grx3.
The recombinant suicide plasmids were transferred intothe Rm2011 recipient strain by triparental mating with E. coliMT616 as the helper strain, as previously described (Harri-son et al., 2005). The double-recombinant Smgrx1 andSmgrx2 clone resulting from an exchange of the lacZ-kanamycin resistance cartridge for the wild-type grx1 andgrx2 genes was selected as Smr, Nmr and Tcs. A single-recombinant Smgrx3 clone resulting from the integration ofpSUP202grx3, was selected as Smr and Tcr.
Plant nodulation and nitrogen fixation assays
Medicago truncatula was used as the host plant for assess-ments of nodulation and nitrogen fixation by the S. melilotistrains. Surface-sterilized germinating seedlings were grownon modified Fahraeus medium supplemented with 1.5%agarose (Pucciariello et al., 2009) in plates. Two days later,their roots were inoculated with 200 ml of a suspension of theappropriate bacterial strain at an OD600 of 0.01. N2 fixationactivity was determined by assessing C2H2 reduction by gaschromatography, as previously described (Hérouart et al.,1996).
Extraction of proteins from S. meliloti andenzyme assays
Cells were grown in LBMC medium, and 3 ¥ 109 cells werecentrifuged at 3300 g for 5 min. The resulting pellet was
806 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

resuspended in 500 ml of buffer (50 mM Tris-HCl, pH 7.4,100 mM NaCl), and the cells were lysed by sonication.Insoluble components were removed by centrifugation(18 000 g, 5 min, 4°C). Protein concentration was determinedwith the Bio-Rad (Marnes-la-Coquette, France) protein assaykit, with bovine serum albumin as the standard. The activitiesobtained in all assays were corrected by subtracting the slope(DAbs min-1) of the baseline curve.
Malate dehydrogenase activity was assessed by followingthe consumption of NADH (e = 6.22 mM-1 cm-1) spectropho-tometrically at 340 nm. We added 30 mg of crude proteinextract to 0.2 mM NADH and 0.4 mM oxaloacetate in buffer(50 mM Tris-HCl, pH 7.4, 100 mM NaCl), for a total reactionvolume of 1 ml, and measured the decrease in absorption at340 nm (Kispal et al., 1997; Albrecht et al., 2010).
Isocitrate dehydrogenase activity was assessed bymeasuring the absorption of converted NADPH(e = 6.22 mM-1 cm-1) at 340 nm (Vives-Rego et al., 1981). Weadded 30 mg of crude protein extract to a solution of 2 mMisocitrate trisodium and NADP (1 mM) in buffer [50 mM Tris-HCl, pH 7.4, 1.5 mM MgCl2 and 0.1% (v/v) Triton X-100], in atotal reaction volume of 1 ml.
Succinate dehydrogenase activity was measured with theartificial electron acceptor dichlorophenol-indophenol(DCPIP) (e = 19.1 mM-1 cm-1). We added 30 mg of crudeprotein extract to solution of 10 mM succinate in buffer[50 mM Tris-HCl, pH 7.4, 0.1 mM EDTA, 1 mM KCN, 25 mMDCPIP and 0.1% (v/v) Triton X-100], in a total reactionvolume of 1 ml, and measured the change in absorption at600 nm (Hatefi and Stiggall, 1978).
Aconitase activity was assessed by measuring the absorp-tion of converted NADPH (e = 6.22 mM-1 cm-1) at 340 nm(Drapier and Hibbs, 1996). We added 30 mg of crude proteinextract to a solution of 20 mM trisodium citrate, isocitratedehydrogenase (0.4 U ml-1), and NADP (1 mM) in buffer[50 mM Tris-HCl, pH 7.4, 1.5 mM MgCl2 and 0.1% (v/v) TritonX-100], in a total reaction volume of 1 ml.
Determination of protein-bound and freeglutathione concentrations
Thiols were extracted with perchloric acid, derivatized withmonobromobimane and quantified after separation byreverse-phase HPLC, as previously described (Harrisonet al., 2005). Protein-bound glutathione (S-glutathionylation)levels were determined as previously described (Greethamet al., 2010). After extraction, GSH was derivatized withmonobromobimane and quantified after separation byreverse-phase HPLC, as previously described (Fahey andNewton, 1987). Commercial GSH (Sigma) and g-ECS (LGCStandards, Molsheim, France) were used as standards.
Quantification of iron in the cells
Cells were grown in LBMC medium. We added 0.5 ml offreshly prepared reagent A (0.6 N HCl, 0.142 M KMnO4) to 3¥ 109 cells resuspended in 1 ml of H2O. This digestion mixturewas incubated in a capped tube for 2 h at 60°C. We thenadded 0.1 ml of reagent B (6.5 mM ferrozin, 13.1 mM neocu-proin, 2 M ascorbic acid, 5 M ammonium acetate). The result-
ing solution was incubated for 30 min at room temperature,centrifuged and the absorbance of the supernatant was mea-sured at 562 nm, as previously described (Fish, 1988).Absorbance values were converted to nmoles of Fe with astandard curve generated with ferrous sulfate.
Microscopy
Nodules collected 21 days post infection were fixed by incu-bation overnight with 3.6% glutaraldehyde and 4% formalde-hyde in 0.1 M phosphate buffer (pH 7.2), at 4°C, with gentleshaking. Samples were washed, dehydrated and embeddedin Technovit 7100 (Kulzer Histo-Technik, Heraeus France,Villebon, France), according to the manufacturer’s instruc-tions. Sections (5 mm) were cut and stained with toluidineblue. Sections were examined under an Imager Z.1 (Zeiss,Le Pecq, France) microscope coupled to an Axiocam MRmcamera (Zeiss, Le Pecq, France).
LacZ expression analysis
b-Galactosidase activity was detected histochemically oncross-sections, as previously described (Boivin et al., 1990)This activity was assayed throughout the entire root, at 4 dpi,or on nodules, at 21 dpi, for plants inoculated with strainsRm2011 and transconjugant strains carrying pgrx1::lacZ orpgrx2::lacZ by recombination. The entire root or nodule wasfixed by incubation with 2% glutaraldehyde for 30 min undervacuum and then for 1 h at atmospheric pressure. It was thenrinsed three times, for 10 min each, with buffer Z (0.1 Mphosphate buffer pH 7.4, 10 mM KCl, 1 mM MgSO4). Theentire root or 80 mm longitudinal slices of nodules were thentransferred to the staining solution: 860 ml of buffer Z, 50 ml of100 mM K3[Fe(CN)6], 50 ml of 100 mM K4[Fe(CN)6], 40 mlof 2% 5-bromo-4-chloro-3-indolyl-b-D-galactoside in N,N-dimethylformamide (X-Gal) (Boivin et al., 1990). Transversesections of nodules (80 mm) were obtained with a vibratome.The stained sections were examined under an Imager Z.1(Zeiss, Le Pecq, France) microscope coupled to an AxiocamMRm camera (Zeiss, Le Pecq, France).
Quantitative RT-PCR analyses
Reverse transcription was performed with the Omniscript RTkit (Qiagen, Courtaboeuf, France) and random primers (Invit-rogen, Eragny Sur Oise, France), on total RNA extracted fromexponentially growing bacterial cultures or from nodules. ThecDNAs were synthesized according to the manufacturer’sprotocol, with 2 mg of total RNA. RT-qPCR was performedwith a DNA Engine Opticon 2 Continuous FluorescenceDetection system (MJ Research, Bio-Rad, Marnes-la-Coquette, France) and qPCR MasterMix Plus for SYBER®green I (Eurogentec, Angers, France). In each reaction, weused 5 ml of 100-fold diluted cDNA and 0.3 mM primers (thesequences used are described in Table S2). The PCR con-ditions were 50°C for 5 min, then 95°C for 10 min, followed by40 cycles of 95°C for 30 s and 60°C for 1 min. Each reactionwas performed in triplicate and the results shown are themeans of three independent biological experiments. Datawere quantified with Opticon Monitor 2 (MJ Research). Two
Roles of glutaredoxins in S. meliloti 807
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

constitutively expressed genes, the 16S rRNA gene (a bac-terial gene) and Mtc 27 (a plant gene), were used as refer-ences for normalization.
Statistical analyses
The significance of differences was assessed with Student’st-test.
Acknowledgements
We thank Dr Renaud Brouquisse and Claude Castella forfruitful discussions and help on enzymatic and Western blotanalyses. Sofiane Benyamina was recipient of fellowshipsfrom the European programme Averroes and the French pro-gramme Théthys-Arcus-CERES. This work was supported byCentre National de la Recherche Scientifique, Nice-SophiaAntipolis University, Institut National de la RechercheAgronomique and by Institut Universitaire de France grants toJean Pierre Jacquot and Nicolas Rouhier.
References
Albrecht, A.G., Netz, D.J., Miethke, M., Pierik, A.J.,Burghaus, O., Peuckert, F., et al. (2010) SufU is an essen-tial iron-sulfur cluster scaffold protein in Bacillus subtilis.J Bacteriol 192: 1643–1651.
Bandyopadhyay, S., Gama, F., Molina-Navarro, M.M., Gual-berto, J.M., Claxton, R., Naik, S.G., et al. (2008) Chloro-plast monothiol glutaredoxins as scaffold proteins for theassembly and delivery of [2Fe-2S] clusters. EMBO J 27:1122–1133.
Barnett, M.J., Toman, C.J., Fisher, R.F., and Long, S.R.(2004) A dual-genome Symbiosis Chip for coordinate studyof signal exchange and development in a prokaryote–hostinteraction. Proc Natl Acad Sci USA 101: 16636–16641.
Becker, A., Berges, H., Krol, E., Bruand, C., Ruberg, S.,Capela, D., et al. (2004) Global changes in gene expres-sion in Sinorhizobium meliloti 1021 under microoxic andsymbiotic conditions. Mol Plant Microbe Interact 17: 292–303.
Boivin, C., Camut, S., Malpica, C.A., Truchet, G., and Rosen-berg, C. (1990) Rhizobium meliloti genes encoding catabo-lism of trigonelline are induced under symbiotic conditions.Plant Cell 2: 1157–1170.
Butland, G., Babu, M., Diaz-Mejia, J.J., Bohdana, F., Phanse,S., Gold, B., et al. (2008) eSGA: E. coli synthetic geneticarray analysis. Nat Methods 5: 789–795.
Capela, D., Filipe, C., Bobik, C., Batut, J., and Bruand, C.(2006) Sinorhizobium meliloti differentiation during symbio-sis with alfalfa: a transcriptomic dissection. Mol PlantMicrobe Interact 19: 363–372.
Chang, C., Damiani, I., Puppo, A., and Frendo, P. (2009)Redox changes during the legume-rhizobium symbiosis.Mol Plant 2: 370–377.
Chao, T.C., Becker, A., Buhrmester, J., Puhler, A., andWeidner, S. (2004) The Sinorhizobium meliloti fur generegulates, with dependence on Mn(II), transcription of thesitABCD operon, encoding a metal-type transporter.J Bacteriol 186: 3609–3620.
Chao, T.C., Buhrmester, J., Hansmeier, N., Puhler, A., andWeidner, S. (2005) Role of the regulatory gene rirA in thetranscriptional response of Sinorhizobium meliloti to ironlimitation. Appl Environ Microbiol 71: 5969–5982.
Couturier, J., Jacquot, J.P., and Rouhier, N. (2009) Evolutionand diversity of glutaredoxins in photosynthetic organisms.Cell Mol Life Sci 66: 2539–2557.
Couturier, J., Stroher, E., Albetel, A.N., Roret, T., Muthura-malingam, M., Tarrago, L., et al. (2011) Arabidopsis chlo-roplastic glutaredoxin C5 as a model to explore moleculardeterminants for iron-sulfur cluster binding into glutaredox-ins. J Biol Chem 286: 27515–27527.
Crooks, G.E., Hon, G., Chandonia, J.M., and Brenner, S.E.(2004) WebLogo: a sequence logo generator. GenomeRes 14: 1188–1190.
Dalle-Donne, I., Rossi, R., Colombo, G., Giustarini, D., andMilzani, A. (2009) Protein S-glutathionylation: a regulatorydevice from bacteria to humans. Trends Biochem Sci 34:85–96.
Drapier, J.C., and Hibbs, J.B., Jr (1996) Aconitases: a classof metalloproteins highly sensitive to nitric oxide synthesis.Methods Enzymol 269: 26–36.
Eddy, S.R. (1998) Profile hidden Markov models. Bioinfor-matics 14: 755–763.
Fahey, R.C., and Newton, G.L. (1987) Determination of low-molecular-weight thiols using monobromobimane fluores-cent labeling and high-performance liquid chromatography.In Methods in Enzymology. Jakoby, W.B., and Griffith, O.W.(eds). San Diego, CA, USA: Academic Press, pp. 85–96.
Faulkner, M.J., Veeravalli, K., Gon, S., Georgiou, G., andBeckwith, J. (2008) Functional plasticity of a peroxidaseallows evolution of diverse disulfide-reducing pathways.Proc Natl Acad Sci USA 105: 6735–6740.
Fernandes, A.P., Fladvad, M., Berndt, C., Andresen, C., Lillig,C.H., Neubauer, P., et al. (2005) A novel monothiol glutare-doxin (Grx4) from Escherichia coli can serve as a substratefor thioredoxin reductase. J Biol Chem 280: 24544–24552.
Finn, R.D., Mistry, J., Tate, J., Coggill, P., Heger, A., Polling-ton, J.E., et al. (2009) The Pfam protein families database.Nucleic Acids Res 38: D211–D222.
Fish, W.W. (1988) Rapid colorimetric micromethod for thequantitation of complexed iron in biological samples.Methods Enzymol 158: 357–364.
Foyer, C.H., and Noctor, G. (2011) Ascorbate and glu-tathione: the heart of the redox hub. Plant Physiol 155:2–18.
Gallogly, M.M., Starke, D.W., and Mieyal, J.J. (2009) Mecha-nistic and kinetic details of catalysis of thiol-disulfideexchange by glutaredoxins and potential mechanisms ofregulation. Antioxid Redox Signal 11: 1059–1081.
Greetham, D., Vickerstaff, J., Shenton, D., Perrone, G.G.,Dawes, I.W., and Grant, C.M. (2010) Thioredoxins functionas deglutathionylase enzymes in the yeast Saccharomy-ces cerevisiae. BMC Biochem 11: 3.
Haag, A.F., Baloban, M., Sani, M., Kerscher, B., Pierre, O.,Farkas, A., et al. (2011) Protection of Sinorhizobiumagainst host cysteine-rich antimicrobial peptides is criticalfor symbiosis. PLoS Biol 9: e1001169.
Haas, H., Eisendle, M., and Turgeon, B.G. (2008) Sidero-phores in fungal physiology and virulence. Annu Rev Phy-topathol 46: 149–187.
808 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

Hanyu, M., Fujimoto, H., Tejima, K., and Saeki, K. (2009)Functional differences of two distinct catalases inMesorhizobium loti MAFF303099 under free-living andsymbiotic conditions. J Bacteriol 191: 1463–1471.
Harrison, J., Jamet, A., Muglia, C.I., Van de Sype, G., Aguilar,O.M., Puppo, A., and Frendo, P. (2005) Glutathione plays afundamental role in growth and symbiotic capacity ofSinorhizobium meliloti. J Bacteriol 187: 168–174.
Hatefi, Y., and Stiggall, D.L. (1978) Preparation and proper-ties of succinate: ubiquinone oxidoreductase (complex II).Methods Enzymol 53: 21–27.
Hérouart, D., Sigaud, S., Moreau, S., Frendo, P., Touati, D.,and Puppo, A. (1996) Cloning and characterization of thekatA gene of Rhizobium meliloti encoding a hydrogenperoxide-inducible catalase. J Bacteriol 178: 6802–6809.
Jamet, A., Sigaud, S., Van de Sype, G., Puppo, A., andHerouart, D. (2003) Expression of the bacterial catalasegenes during Sinorhizobium meliloti–Medicago sativa sym-biosis and their crucial role during the infection process.Mol Plant Microbe Interact 16: 217–225.
Johnston, A.W., Todd, J.D., Curson, A.R., Lei, S., Nikolaidou-Katsaridou, N., Gelfand, M.S., and Rodionov, D.A. (2007)Living without Fur: the subtlety and complexity of iron-responsive gene regulation in the symbiotic bacteriumRhizobium and other alpha-proteobacteria. Biometals 20:501–511.
Kaplan, C.D., and Kaplan, J. (2009) Iron acquisition andtranscriptional regulation. Chem Rev 109: 4536–4552.
Kispal, G., Csere, P., Guiard, B., and Lill, R. (1997) The ABCtransporter Atm1p is required for mitochondrial iron homeo-stasis. FEBS Lett 418: 346–350.
Kokotek, W., and Lotz, W. (1989) Construction of a lacZ-kanamycin-resistance cassette, useful for site-directedmutagenesis and as a promoter probe. Gene 84: 467–471.
Kumanovics, A., Chen, O.S., Li, L., Bagley, D., Adkins, E.M.,Lin, H., et al. (2008) Identification of FRA1 and FRA2 asgenes involved in regulating the yeast iron regulon inresponse to decreased mitochondrial iron-sulfur clustersynthesis. J Biol Chem 283: 10276–10286.
Lee, J.W., and Helmann, J.D. (2007) Functional specializa-tion within the Fur family of metalloregulators. Biometals20: 485–499.
Lillig, C.H., Berndt, C., Vergnolle, O., Lonn, M.E., Hudemann,C., Bill, E., and Holmgren, A. (2005) Characterization ofhuman glutaredoxin 2 as iron-sulfur protein: a possiblerole as redox sensor. Proc Natl Acad Sci USA 102: 8168–8173.
Lillig, C.H., Berndt, C., and Holmgren, A. (2008) Glutaredoxinsystems. Biochim Biophys Acta 1780: 1304–1317.
Lopez, M., Herrera-Cervera, J.A., Iribarne, C., Tejera, N.A.,and Lluch, C. (2008) Growth and nitrogen fixation in Lotusjaponicus and Medicago truncatula under NaCl stress:nodule carbon metabolism. J Plant Physiol 165: 641–650.
Marino, D., Gonzalez, E.M., and Arrese-Igor, C. (2006)Drought effects on carbon and nitrogen metabolism of peanodules can be mimicked by paraquat: evidence for theoccurrence of two regulation pathways under oxidativestresses. J Exp Bot 57: 665–673.
Marino, D., Pucciariello, C., Puppo, A., and Frendo, P. (2009)The redox state, a referee of the legume-rhizobia symbioticgame. In Advances in Botanical Research: Oxidative Stress
and Redox Regulation in Plants, Vol. 52. Jacquot, J.-P. (ed.).Burlington, MA, USA: Academic Press, pp. 115–151.
Mercier, A., and Labbe, S. (2009) Both Php4 function andsubcellular localization are regulated by iron via a multistepmechanism involving the glutaredoxin Grx4 and the expor-tin Crm1. J Biol Chem 284: 20249–20262.
Mercier, A., and Labbe, S. (2010) Iron-dependent remodelingof fungal metabolic pathways associated with ferrichromebiosynthesis. Appl Environ Microbiol 76: 3806–3817.
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N.,Vaubert, D., Kondorosi, A., and Kondorosi, E. (2003) Anovel family in Medicago truncatula consisting of more than300 nodule-specific genes coding for small, secretedpolypeptides with conserved cysteine motifs. Plant Physiol132: 161–173.
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A.,Catrice, O., et al. (2006) Eukaryotic control on bacterial cellcycle and differentiation in the Rhizobium-legume symbio-sis. Proc Natl Acad Sci USA 103: 5230–5235.
Meyer, Y., Buchanan, B.B., Vignols, F., and Reichheld, J.P.(2009) Thioredoxins and glutaredoxins: unifying elementsin redox biology. Annu Rev Genet 43: 335–367.
Molina-Navarro, M.M., Casas, C., Piedrafita, L., Belli, G., andHerrero, E. (2006) Prokaryotic and eukaryotic monothiolglutaredoxins are able to perform the functions of Grx5 inthe biogenesis of Fe/S clusters in yeast mitochondria.FEBS Lett 580: 2273–2280.
Mühlenhoff, U., Gerber, J., Richhardt, N., and Lill, R. (2003)Components involved in assembly and dislocation of iron-sulfur clusters on the scaffold protein Isu1p. EMBO J 22:4815–4825.
Mühlenhoff, U., Molik, S., Godoy, J.R., Uzarska, M.A.,Richter, N., Seubert, A., et al. (2010) Cytosolic monothiolglutaredoxins function in intracellular iron sensing and traf-ficking via their bound iron-sulfur cluster. Cell Metab 12:373–385.
Ott, T., van Dongen, J.T., Gunther, C., Krusell, L., Des-brosses, G., Vigeolas, H., et al. (2005) Symbiotic leghemo-globins are crucial for nitrogen fixation in legume rootnodules but not for general plant growth and development.Curr Biol 15: 531–535.
Pauly, N., Pucciariello, C., Mandon, K., Innocenti, G., Jamet,A., Baudouin, E., et al. (2006) Reactive oxygen and nitro-gen species and glutathione: key players in the legume-Rhizobium symbiosis. J Exp Bot 57: 1769–1776.
Pucciariello, C., Innocenti, G., Van de Velde, W., Lambert, A.,Hopkins, J., Clement, M., et al. (2009) Homo)glutathionedepletion modulates host gene expression during the sym-biotic interaction between Medicago truncatula andSinorhizobium meliloti. Plant Physiol 151: 1186–1196.
Pujol-Carrion, N., Belli, G., Herrero, E., Nogues, A., andTorre-Ruiz, M.A. (2006) Glutaredoxins Grx3 and Grx4regulate nuclear localisation of Aft1 and the oxidativestress response in Saccharomyces cerevisiae. J Cell Sci119: 4554–4564.
Rodriguez-Manzaneque, M.T., Tamarit, J., Belli, G., Ros, J.,and Herrero, E. (2002) Grx5 is a mitochondrial glutaredoxinrequired for the activity of iron/sulfur enzymes. Mol Biol Cell13: 1109–1121.
Rouhier, N., Unno, H., Bandyopadhyay, S., Masip, L., Kim,S.K., Hirasawa, M., et al. (2007) Functional, structural, and
Roles of glutaredoxins in S. meliloti 809
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810

spectroscopic characterization of a glutathione-ligated[2Fe-2S] cluster in poplar glutaredoxin C1. Proc Natl AcadSci USA 104: 7379–7384.
Rouhier, N., Lemaire, S.D., and Jacquot, J.P. (2008) The roleof glutathione in photosynthetic organisms: emerging func-tions for glutaredoxins and glutathionylation. Annu RevPlant Biol 59: 143–166.
Rouhier, N., Couturier, J., Johnson, M.K., and Jacquot, J.P.(2010) Glutaredoxins: roles in iron homeostasis. TrendsBiochem Sci 35: 43–52.
Schenk, P.M., Baumann, S., Mattes, R., and Steinbiss, H.H.(1995) Improved high-level expression system for eukary-otic genes in Escherichia coli using T7 RNA polymeraseand rare ArgtRNAs. Biotechniques 19: 196–198, 200.
Schroeder, B.O., Wu, Z., Nuding, S., Groscurth, S., Marcino-wski, M., Beisner, J., et al. (2011) Reduction of disulphidebonds unmasks potent antimicrobial activity of humanbeta-defensin 1. Nature 469: 419–423.
Seefeldt, L.C., Hoffman, B.M., and Dean, D.R. (2009) Mecha-nism of Mo-dependent nitrogenase. Annu Rev Biochem78: 701–722.
Sigaud, S., Becquet, V., Frendo, P., Puppo, A., and Hérouart,D. (1999) Differential regulation of two divergent Sinorhizo-bium meliloti genes for HPII-like catalases during free-living growth and protective role of both catalases duringsymbiosis. J Bacteriol 181: 2634–2639.
Simon, R., Priefer, U., and Pühler, A. (1983) ) A broad hostrange mobilization system for in vivo genetic engineering:transposon mutagenesis in gram negative bacteria. Bio-technology (N Y) 1: 784–791.
Spatzal, T., Aksoyoglu, M., Zhang, L., Andrade, S.L., Schle-icher, E., Weber, S., et al. (2011) Evidence for interstitialcarbon in nitrogenase FeMo cofactor. Science 334: 940.
Tamura, K., Dudley, J., Nei, M., and Kumar, S. (2007) MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Mol Biol Evol 24: 1596–1599.
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M.,Ishihara, H., Kevei, Z., et al. (2010) Plant peptides governterminal differentiation of bacteria in symbiosis. Science327: 1122–1126.
Vasse, J., de Billy, F., Camut, S., and Truchet, G. (1990)Correlation between ultrastructural differentiation of
bacteroids and nitrogen fixation in alfalfa nodules.J Bacteriol 172: 4295–4306.
Vives-Rego, J., Juarez, A., Imperial, J., and Pares, R. (1981)Correlation between isocitrate dehydrogenase activity andglutamate excretion by Citrobacter intermedius C3. J GenMicrobiol 122: 167–170.
Ye, H., and Rouault, T.A. (2010) Human iron-sulfur clusterassembly, cellular iron homeostasis, and disease. Bio-chemistry 49: 4945–4956.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Fig. S1. UV-visible absorption spectrum of the SmGrx1 holo-form. The spectrum, recorded after an aerobic purification in30 mM Tris-HCl pH 8.0, is very similar to those recorded forother Fe-S cluster-containing Grxs, with two absorptionbands in the 300–500 nm region characteristic of a [2Fe-2S]centre (Rouhier et al., 2007).Fig. S2. Analysis of the capacity of SmGRXs to form cova-lent oligomers. Five micrograms of the SmGRX1 (lanes 2–5),SmGRX2 (lanes 1–4) and SmGRX3 (lanes 3–6) recombinantproteins were separated by SDS-PAGE (17% polyacrylamidegel) in reducing (lanes 1–3, presence of 140 mM b-mercaptoethanol in the Laemmli buffer) and non-reducing(lanes 4–6, absence of b-mercaptoethanol) conditions.Fig. S3. Detection of GRXs in S. meliloti. The presence ofGrxs was evaluated, by Western blot analysis, in total proteinextracts from the wild-type and mutant strains. The recombi-nant (r) Grx were used as controls. Note that for SmGRX3, atruncated version corresponding to the Grx domain (10 kDa)was compared with the full-length protein of 28 kDa.Fig. S4. Analysis of the size of Smgrx mutant nodules. Thesize of 3-week-old nodules was analysed by microscopy. Thedata shown are the means for 15 nodules from threeexperiments � SEM.Table S1. Bacterial strains and plasmids used in this study.Table S2. Primers used for RT-qPCR analysis, mutant con-struction and cloning of the recombinant proteins.
810 S. M. Benyamina et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 15, 795–810
Related Documents





![apresentados.,.] 1996 00326 Workshopainfo.cnptia.embrapa.br/digital/bitstream/item/203514/1/Coutinho... · Bactérias dos gêneros Rhizobium, Bradyrhizobium, Sinorhizobium, e Azorhizobium,](https://static.cupdf.com/doc/110x72/5e56a3be2c56cc78020e5738/apresentados-1996-00326-bactrias-dos-gneros-rhizobium-bradyrhizobium-sinorhizobium.jpg)





