_____________ Mun. Ent. Zool. Vol. 6, No. 1, January 2011___________ I This volume is dedicated to the memory of the chief-editor Hüseyin Özdikmen’s father-in-law TURGUT AKÇAY who lived an honorable life MUNIS ENTOMOLOGY & ZOOLOGY Ankara / Turkey

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

_____________ Mun. Ent. Zool. Vol. 6, No. 1, January 2011___________ I
This volume is dedicated to the memory of the chief-editor Hüseyin Özdikmen’s father-in-law
TURGUT AKÇAY
who lived an honorable life
MUNIS
ENTOMOLOGY & ZOOLOGY
Ankara / Turkey

_____________ Mun. Ent. Zool. Vol. 6, No. 1, January 2011___________ II
Scope: Munis Entomology & Zoology publishes a wide variety of papers on all aspects of Entomology and Zoology from all of the world, including mainly studies on systematics, taxonomy, nomenclature, fauna, biogeography, biodiversity, ecology, morphology, behavior, conservation, paleobiology and other aspects are appropriate topics for papers submitted to Munis Entomology & Zoology. Submission of Manuscripts: Works published or under consideration elsewhere (including on the internet) will not be accepted. At first submission, one double spaced hard copy (text and tables) with figures (may not be original) must be sent to the Editors, Dr. Hüseyin Özdikmen for publication in MEZ. All manuscripts should be submitted as Word file or PDF file in an e-mail attachment. If electronic submission is not possible due to limitations of electronic space at the sending or receiving ends, unavailability of e-mail, etc., we will accept ―hard‖ versions, in triplicate, accompanied by an electronic version stored in a floppy disk, a CD-ROM. Review Process: When submitting manuscripts, all authors provides the name, of at least three qualified experts (they also provide their address, subject fields and e-mails). Then, the editors send to experts to review the papers. The review process should normally be completed within 45-60 days. After reviewing papers by reviwers: Rejected papers are discarded. For accepted papers, authors are asked to modify their papers according to suggestions of the reviewers and editors. Final versions of manuscripts and figures are needed in a digital format.
Preparation of Manuscripts All manuscripts must be typed in English, using Microsoft Word. Entire manuscript must be double-spaced, with margins of at least 2-3 cm on all sides of the page (A4). Pages should be numbered consecutively. Authors whose native language is not English are encouraged to have their manuscripts read by a native English-speaking colleague before submission. Nomenclature must be in agreement with the International Code of Zoological Nomenclature (4th edition 1999). Author(s) of species name must be provided when the scientific name of any animal species is first mentioned (the year of publication needs not be given; if you give it, then provide a full reference of this in the reference list). Authors of plant species name need not be given. Metric systems should be used. If possible, use the common font Times New Roman (12 pt) and use as little formatting as possible (use only bold and italics). Special symbols (e.g. male or female sign) should be avoided.

_____________ Mun. Ent. Zool. Vol. 6, No. 1, January 2011___________ III
Title and Name(s) of Author(s): The title should be informative and as possible as brief, in boldface capital letters, not exceed twenty words. The higher taxa containing the taxa dealt with in the paper should be indicated in parentheses. Full name(s) of author(s) should come underneath the title with full address, each on a separate line. The author(s) name (s) should be given in boldface lower case. Abstract: The abstract should be concise and should draw attention to the significant contents of the paper and the author's main conclusions. It should normally not exceed 200 words and should contain no uncommon abbreviations or references. Any new names or new combinations proposed in the paper should be mentioned. The abstract should be followed by a list of key words. Up to seven keywords should be suggested by the author. Text: Regular papers include as the main sections (except in Book Reviews and Scientific Notes etc.); Introduction, Material & Methods, Results, Discussion, Acknowledgments and Literature Cited. The section introduction should be written without a title. However, the main sections may be varies with different types of papers. According to types of papers, main section can be changed. All scientific names (only genus and species group names) should be italicized throughout the paper, including literature cited. References should be cited in the text as Turgut (2003), Turgut & Turgut (2000) or Turgut et al. (2001) (3 or more authors), or alternatively in a parenthesis (Turgut, 2003; Turgut & Turgut, 2000 or Turgut et al., 2001). All literatures in the text must be listed alphabetically in the literature cited in the following format. Journal paper: Turgut, S. 2003. Title of the paper. Title of the journal in full, volume number: page range. Book chapter: Turgut, S. & Turgut, A. 2000. Title of the Chapter. In: Turgut, A., Turgut, B. & Turgut, C. (Eds.), Title of Book. Publisher name and location, page range. Book: Turgut, A., Turgut, B. & Turgut, C. 2001. Title of Book, Publisher name and location, number of pages (e.g. 123 pp). Internet resources: Turgut, S. 2002. Title of website, database or other resources, Publisher name and location (if indicated), number of pages (if known). Available from: http://xxx.xxx.xxx/ (Date of access).

_____________ Mun. Ent. Zool. Vol. 6, No. 1, January 2011___________ IV
Tables, Illustrations and Photographs: Tables, illustrations and photographs should be submitted in a separate file, not embedded in the text. They should be given at the end of the manuscript. Please use the table function in your word processor to build tables so that the cells, rows and columns can remain aligned when font size and width of the table are changed. Illustrations should be clean, sharp, with good contrast. Small illustrations should be grouped into plates. For species illustration, line drawings are preferred, although good quality B&W photographs are also acceptable. Maximum size of printed illustration, including all legends, is 12 x 16 cm. Images must be submitted either in .tif, .jpg, or .pdf (PC compatible format strongly preferred). Digital versions of illustrations should be prepared as follows: photographs should be saved as .pdf or .tif format at 300 dpi. Line figures should be saved in .tif or .jpg at 300 dpi. All illustrations must be numbered consecutively using Arabic numerals. They should be cited ―Fig. 1‖ or ―Figs. 1–4‖ in sequential order. Photographs must be of exceptional quality, good contrast. Scientific Notes and Book Reviews. These are usually short contributions, typically not exceeding one (Book Review) or two (Scientific Notes) printed pages. Scientific notes and book reviews lack an abstract and most of the main headings, except for the acknowledgements and the literature cited sections. Page Charge: There is no page charge for publishing with MEZ. MEZ is indexed in Zoological Record, Biological Abstract, Biosis Preview, Agricola, ……

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 1
NEW SUBSPECIES OF STENURELLA BIFASCIATA (MÜLLER, 1776) (COLEOPTERA, CERAMBYCIDAE)
FROM SOUTH WEST TURKEY
Mikhail L. Danilevsky* * A. N. Severtzov Institute of Ecology and Evolution Russian Academy of Sciences Leninsky prospect 33, Moscow, 119071, RUSSIA. E-mail: [email protected], [email protected], Web: www.cerambycidae.net [Danilevsky, M. 2011. New subspecies of Stenurella bifasciata (Müller, 1776) (Coleoptera, Cerambycidae) from South West Turkey. Munis Entomology & Zoology, 6 (1): 1-5] ABSTRACT: Stenurella bifasciata safronovi ssp. n. is described from South-West Turkey (Antalya, Beldibi env. – type locality and Isparta). S. bifasciata ssp. intermedia Holzschuh, 2006, new rank originally described as a species from Magnisia, is accepted as a subspecies distributed in Greece from south Peloponnesus to Bulgaria and in SW Bulgaria; several forms of previously unknown females are figured. Stenurella bifasciata ssp. ferruginipes (Pic, 1895), new rank described as a variation from «Bitlis» is accepted as a subspecies distributed in Mardin and Bitlis. S. bifasciata ssp. lanceolata (Mulsant & Rey, 1863), new rank, regarded previosly as a synonym of the nominative subspecies, is accepted as a subspecies distributed in Iberian Peninsula and South-East France. The species area of S. bifasciata (Müller, 1776) is extended eastwards to Pacific Ocean. KEY WORDS: Coleoptera, Cerambycidae, Lepturinae, Stenurella, taxonomy, new rank, new subspecies, Palaearcrtic Region, Europe, Spain, France, Greece, Bulgaria, Turkey.
Stenurella bifasciata (Müller, 1776) was described in the genus Leptura Linnaeus, 1758 from «Dania». The species is widely distributed in the north of Palaearctic Region: all over Europe from Portugal to Urals (absent in Great Britain and Scandinavia); in Siberia it was traditionally known (Plavilstshikov, 1936) to about Transbaikalia (absent in Mongolia) and Yakutia (Tsherepanov, 1996), but in my collections several specimens from the North-East part of Khabatovsk Region (Nelkan environs, 57°39‘N, 136°09‘E) are preserved, so the area of the species must reach Pacific Ocean; all over Caucasus with Transcaucasia; in Near East - to North Iran. Records from Syria (Rejzek et al., 2003), Lebanon (Sama et al., 2010) and Iraq (Derwesh, 1965) are known. In Turkey the species is distributed all over the country (Özdikmen, 2007).
Now the species is delimited in three subspecies (Löbl & Smetana, 2010): S. b. limbiventris (Reitter, 1898) distributed along the East bank of Black Sea in Georgia and Turkey; S. b. nigrosuturalis (Reitter, 1895) known from Syria, Lebanon and South-Eastern Turkey; the nominative subspecies is generally accepted to be distributed in the rest of the species area.
In fact the taxonomy of S. bifasciata is much more complicated. Populations, which contain specimens with pale-orange legs, represent a well
delimited subspecies described as Strangalia bifasciata var. ferruginipes Pic, 1895 from «Bitlis». Stenurella bifasciata ssp. ferruginipes (Pic, 1895), new rank is represented in my materials from Mardin (Hop Geçidi) and Bitlis (Tatvan environs).
The taxon described as Stenurella intermedia Holzschuh, 2006 from Greece (Magnisia, Othrys, 1100m, 39˚05‘N, 22˚40‘E) on the base of two males is in fact a local subspecies of S. bifasciata. The true nature of the taxon is clear now after study of 54 specimens (34 males and 20 females) collected in south Greece (many

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 2
localities from Fthiotis to South Peloponnesus) by A. Napolov & I. Roma in May 2010. The relatively big pronotal punctation of S. bifasciata ssp. intermedia Holzschuh, 2006, new rank (the main distinguishing character according to the original description) is really a little larger than in specimens from Central Europe, but just same as in many southern populations from Italy to Caucasus. Two females of S. b. intermedia were also collected by A. Napolov & I. Roma in SW Bulgaria near Kresna (2.6.2010). Several series S. b. intermedia collected by V. Sakalyan in 1986-87 and by T. Ljubomirov in 2009 along south parts of the valleys of Struma and Mesta rivers in SW Bulgaria are preserved in my collection. Females of S. b. intermedia Holz. (Figs 1-3) are very similar to S. b. bifasciata, but black elytral design is considerably reduced to about totally absent (Fig. 3). The nominative subspecies S. b. bifasciata with well developed black elytral design is also distributed in Bulgaria. It is known already in Sophia environs.
The taxon described as Strangalia lanceolata Mulsant & Rey, 1863 from «L‘Espagne» on the base of females with elytra widely darkened along suture is a well formed Iberian subspecies S. bifasciata ssp. lanceolata (Mulsant & Rey, 1863), new rank. The peculiarity of Iberian S. bifasciata was already mentioned by Villiers (1978): ―It est curieux de noter la présence de deux formes prescue identiques, l‘une lanceolata Mulsant, en Méditerranée occidentale et l‘autre, nigrosuturalis Reitter, en Méditerranée orientale.‖ S. b. lanceolata penetrates in South France. Two females with the label: «France, Pyrénées Orientales, Prades, 24-30.6.1986, Schimmel leg.» are preserved in my collection.
The taxon described as Strangalia bifasciata var. nigriventris Pic, 1891 without exact locality (South France?) on the base of females with black abdomen is most probably also a good subspecies. According to Villiers (1978) in France: ―La var. nigriventris semble plus méridionale que la forme typique, mais paraît manquer en Corse.‖
One more subspecies is described bellow from south-west Turkey.
Description
Stenurella bifasciata safronovi ssp. n.
(Figs 4-6) Holotype: male (Fig. 4) with the label: ―TR, Taurus, Antalya, Kemer distr., Beldibi env., 36°44‘N, 30°33‘E 13-21.5. 2008, A. A. Safronov leg.‖ – author‘s collection. 91 paratypes – all in author‘s collection: 10 males and 13 females with same label as holotype; 31 males and 36 females from same locality: 15-26.5.2010, D. A. Safronov leg.; 1 female (Fig , ―Turkey, Isparta: Isparta-Sidre sub., 37°44‘N, 30°33‘E, 1320m, 13.VII.2008, T. Ljubomirov leg.‖ Body length in males: 8.5-11.5mm, in females: 8.4-11.6mm; body width in males: 2.1-2.9mm, in females: 2.2-3.0mm. Body black with partly dark-red abdomen, 2nd - 4th visible sternites totally red, 1st and 5th totally or partly black; legs and antennae totally black; general elytral color dark-brown: elytra in males usually dark-red with black apices, but sometimes (as in holotype - Fig. 4) with large rhomboid black area, which could be more or less reduced; elytra in females usually with central black area widely connected with apical black area (Fig. 5), but not touching laterally epipleurae; black elytral areas in females can be strongly reduced (Fig. 6); pronotal

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 3
punctation is relatively rough, a little rougher than in S. b. intermedia and much rougher then in specimens of S. b. bifasciata from Central Europe. Differential diagnosis. The new taxon differs from all other subspecies by peculiar elytral design in males and females. The close Eastern subspecies S. b. nigrosuturalis (Reitter, 1895) has similarly dark-red elytral and abdomen color, similarly rough pronotal punctation, but elytral black area is much more developed. European subspecies are poorly investigated, but Spanish S. b. lanceolata seems usually have also very rough pronotal punctation, but another character of black elytral design, bisides specimens of S. b. lanceolata can have black abdomen. S. b. intermedia distributed in the south-east of the West Europe have usually strongly reduced black elytral area in females, which sometimes nearly totally absent (Fig. 3). The nomianative subspecies is characterized by relatively fine pronotal punctation and light-red general elytral color, males without central elytral black area, females with more or less wide contrast postmedial black stripe reaching laterally epipleurae. Distribution. S. b. safronovi ssp. n. is distributed in south-west Turkey, it is now known from Antalya and Isparta, but its real area must be much wider. Etymology. The new subspecies is dedicated to Alexandr Safronov (Tula, Russia), who discovered the taxon.
ACKNOWLEDGEMENTS
I am very grateful to Toshko Ljubomirov (Bulgaria, Sofia), Alexander Napolov (Riga, Latvia), Alexandr Safronov (Tula, Russia), Dmitriy Safronov (Tula, Russia) and Vladimir Sakalyan (Bulgaria, Sofia) for providing me with the specimens for study.
LITERATURE CITED Derwesh, A. I. 1965. A preliminary list of identified Insects and some Arachnids of Iraq. Bulletin No. 112. Directorate General of Agricultural Research and Projects. Ministry of Agriculture: 1-123. Holzschuh, C. 2006. Beschreibung von 51 neuen Bockkäfern aus der palaearktischen und orientalischen Region, vorwiegend aus Borneo und China (Coleoptera, Cerambycidae). Entomologica Basiliensia et Collectionis Frey, 28: 205-276. Linnaeus, C. 1758. Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymies, locis. Tomus I. Editio decima, reformata. Holmiae. Impensis Direct. Laurentii Salvii, iv + 824 + [1] pp. Löbl, I. & Smetana A. (ed.) 2010. Catalogue of Palaearctic Coleoptera, Vol. 6. Chrysomeloidea. Stenstrup. Apollo Books, 924 pp. Müller, O. F. 1776. Zoologiae Danicae prodromus, seu animalium Daniae et Norvegiae indigenarum characteres, nomina et synonyma imprimis popularium. Hafniae. Hallageriis, xxxii + 282 pp. Mulsant, E. & Rey, C. 1863: Longicornes nouveaux ou peu connus. Annales de la Société Linnéenne de Lyon (2) 10: 144-184. Özdikmen, H. 2007. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae). Part I - Black Sea Region. Munis Entomology & Zoology, 2 (2): 179-422. Pic, M. 1891: Faune franco-algérienne (Variétés). Pp. 1-50. Matériaux pour servir à l‘étude des longicornes. Premier cahier. Lyon: L. Jacquet, 67 + [1] pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 4
Pic, M. 1895. Longicornes de la collection H. Tournier. L‘Échange, Revue Linnéenne, 11 (127): 75-78. Plavilstshikov, N. N. 1936. Fauna SSSR. Nasekomye zhestokrylye. T. XXI. Zhuki-drovoseki (ch. 1). Moskva - Leningrad: Izdatel‘stvo Akademii Nauk SSSR, 612 + [1] pp. Rejzek, M., Kadlec, S. & Sama, G. 2003. Contribution to the knowledge of Syrian Cerambycidae fauna (Coleoptera). Biocosme Mésogéen, Nice, 20 (1): 7-50. Sama, G., Rapuzzi, P. & Kairouz, A. 2010. Catalogue commenté des Cerambycidae du Liban. An annotated catalogue of the Cerambycidae of Lebanon (Insecta Coleoptera Cerambycidae). Quaderno di Studi e Notizie di Storia Naturale della Romagna, 30: 131-201. Tsherepanov, A. I. 1996. [104. Fam. Cerambycidae – Longicorn or Timber beetles. Pp. 56-140. In: Ler P.A. (red.) Key to the insects of Russian Far East. Vol. III. Coleoptera. Pt. 3.] Vladivostok: Dal‘nauka: 556p. [in Russian; the text was arranged by G. O. Krivolutzkaya and A. L. Lobanov on the base of a manuscript by Tsherepanov] [in Russian]
1 2 3 Figures 1-3. Stenurella bifasciata intermedia Holzschuh, 2006, new rank, females (author‘s collection): 1. Peloponnesus, Karyoupoli, 100m, 22.5.2010, A. Napolov & I. Roma leg.; 2. SW Bulgaria, Pirin, Struma River, 7.1986, V. Sakalyan leg.; 3. SW Bulgaria, Struma - Mesta, 7.1986, V. Sakalyan leg.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 5
4 5 Figures 4-5. Stenurella bifasciata safronovi ssp. n. (author‘s collection): 4. Male, holotype, TR, Taurus, Antalya, Kemer distr., Beldibi env., 36°44‘N, 30°33‘E 13-21.5. 2008, A. A. Safronov leg.; 5. Female, paratype, TR, Taurus, Antalya, Kemer distr., Beldibi env., 36°44‘N, 30°33‘E 15-26.5. 2010, D. A. Safronov leg.‖.
6
Figure 6. Stenurella bifasciata safronovi ssp. n. (author‘s collection): 6. Female, paratype, Turkey, Isparta: Isparta-Sidre sub., 37°44‘N, 30°33‘E, 1320m, 13.VII.2008, T. Ljubomirov leg.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 6
THE LONGICORN BEETLES OF TURKEY (COLEOPTERA: CERAMBYCIDAE)
PART IV – MEDITERRANEAN REGION
Hüseyin Özdikmen* * Gazi Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü, 06500 Ankara / TÜRKĠYE. E-mail: [email protected] [Özdikmen, H. 2008. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part IV – Mediterranean Region. Munis Entomology & Zoology, 6 (1): 6-145] ABSTRACT: The paper gives faunistical, nomenclatural, taxonomical and zoogeographical review of the longicorn beetles of Mediterranean Region in Turkey. KEY WORDS: Cerambycidae, Fauna, Nomenclature, Zoogeography, Taxonomy, Mediterranean Region, Turkey.
TABLE OF CONTENTS
INTRODUCTION 8 COVERED GEOLOGICAL AREA OF THE PRESENT WORK 9 ARRANGEMENT OF INFORMATION 9 CLASSIFICATION 10 PRIONINAE 11 ERGATINI 11 Ergates Audinet-Serville, 1832 11 Callergates Lameere, 1904 11 MACROTOMINI 11 Prinobius Mulsant, 1842 11 REMPHANINI 12 Rhaesus Motschulsky, 1875 12 AEGOSOMATINI 12 Aegosoma Audinet-Serville, 1832 12 PRIONINI 12 Prionus Geoffroy, 1762 12 Mesoprionus Jakovlev, 1887 13 LEPTURINAE 13 RHAGIINI 13 Rhagium Fabricius, 1775 13 Stenocorus Geoffroy, 1762 15 Anisorus Mulsant, 1862 15 Dinoptera Mulsant, 1863 15 Cortodera Mulsant, 1863 16 LEPTURINI 18 Grammoptera Audinet-Serville, 1835 18 Alosterna Mulsant, 1863 19 Vadonia Mulsant, 1863 20 Pseudovadonia Lobanov, Danilevsky et Murzin, 1981 21 Anoplodera Mulsant, 1839 22 Stictoleptura Casey, 1924 22 Anastrangalia Casey, 1924 26 Pedostrangalia Sokolov, 1897 26 Etorofus Matsushita, 1933 27 Carlandrea Sama & Rapuzzi, 1999 27 Pachytodes Pic, 1891 28 Rutpela Nakane et Ohbayashi, 1957 28 Stenurella Villiers, 1974 28 NECYDALINAE 30 NECYDALINI 30 Necydalis Linnaeus, 1758 30

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 7
ASEMINAE 30 SAPHANINI 30 Oxypleurus Mulsant, 1839 30 ANISARTHRONINI 30 Alocerus Mulsant, 1862 30 ASEMINI 31 Arhopalus Serville, 1834 31 SPONDYLIDINAE 31 SPONDYLIDINI 31 Spondylis Fabricius, 1775 31 DORCASOMINAE 32 DORCASOMINI 32 Apatophysis Chevrolat, 1860 32 CERAMBYCINAE 32 ACHRYSONINI 32 Icosium Lucas, 1854 32 HESPEROPHANINI 32 Hesperophanes Dejean, 1835 32 Trichoferus Wollaston, 1854 33 Stromatium Audinet-Serville, 1834 34 PHORACANTHINI 34 Phoracantha Newman, 1840 34 CERAMBYCINI 35 Cerambyx Linnaeus, 1758 35 TRACHYDERINI 36 Purpuricenus Dejean, 1821 36 CALLICHROMATINI 39 Aromia Audinet-Serville, 1833 39 GRACILIINI 39 Penichroa Stephens, 1839 39 Axinopalpis Dejean, 1835 39 OBRIINI 40 Obrium Dejean, 1821 40 Anatolobrium Adlbauer, 2004 40 CERTALLINI 40 Certallum Dejean, 1821 40 DEILINI 41 Delagrangeus Pic, 1892 41 Deilus Audinet-Serville, 1834 42 STENOPTERINI 42 Stenopterus Illiger, 1804 42 HYBODERINI 43 Callimus Mulsant, 1846 43 Lampropterus Mulsant, 1862 43 Procallimus Pic, 1907 44 PSEBIINI 44 Nathrius Brethes, 1916 44 MOLORCHINI 45 Molorchus Fabricius, 1792 45 Glaphyra Newman, 1840 45 Brachypteroma Heyden, 1863 46 STENHOMALINI 47 Stenhomalus White, 1855 47 CALLIDIINI 47 Hylotrupes Serville, 1834 47 Ropalopus Mulsant, 1839 47 Leioderes Redtenbacher, 1849 48 Semanotus Mulsant, 1839 48 Callidium Fabricius, 1775 48 Phymatodes Mulsant, 1839 49 Poecilium Fairmaire, 1864 49 CLYTINI 50 Plagionotus Mulsant, 1842 50 Isotomus Mulsant, 1862 51

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 8
Chlorophorus Chevrolat, 1863 51 Xylotrechus Chevrolat, 1860 54 Pseudosphegesthes Reitter, 1913 54 Rhaphuma Pascoe, 1858 55 Clytus Laicharting, 1784 55 LAMIINAE 56 PARMENINI 56 Parmena Dejean, 1821 56 MESOSINI 57 Mesosa Latreille, 1829 57 BATOCERINI 57 Batocera Dejean, 1835 57 MONOCHAMINI 58 Monochamus Dejean, 1821 58 LAMIINI 58 Morimus Brullé, 1832 58 DORCADIINI 59 Dorcadion Dalman, 1817 59 APODASYINI 68 Anaesthetis Dejean, 1835 68 PTEROPLINI 68 Niphona Mulsant, 1839 68 POGONOCHERINI 69 Pogonocherus Dejean, 1821 69 ACANTHODERINI 69 Aegomorphus Haldeman, 1847 69 ACANTHOCININI 70 Acanthocinus Dejean, 1821 70 Leiopus Audinet-Serville, 1835 70 Exocentrus Dejean, 1835 71 TETROPINI 71 Tetrops Stephens, 1829 71 SAPERDINI 72 Saperda Fabricius, 1775 72 PHYTOECIINI 72 Oberea Dejean, 1835 72 Oxylia Mulsant, 1862 73 Mallosia Mulsant, 1862 74 Semiangusta Pic, 1893 75 Coptosia Fairmaire, 1864 75 Pygoptosia Reitter, 1895 76 Phytoecia Dejean, 1835 76 AGAPANTHIINI 86 Calamobius Guérin-Méneville, 1847 86 Agapanthia Audinet-Serville, 1835 87 ZOOGEOGRAPHICAL REMARKS 92 CONCLUSIONS 96 LITERATURE CITED 109 APPENDIX 115
Clearly, there is no work, except Parts I, II and III of the series, on Turkish Cerambycidae related to the whole territories of Turkey or regions of Turkey at present. Previous works were either short notes on short-lived expeditions or about at most a province and its environment. Also, works including description of new taxons are sometimes encountered. As opposed to this, European fauna has almost been investigated entirely as mentioned in Sama (2002) and Russian fauna has also been given mainly in Danilevsky (2010a). As known, a series work was planned with the aim to expose Turkish Cerambycidae fauna with as much detail as possible and entirely by beginning from the Black Sea Region (see Part I). The present study is an attempt at the forth step of this aim. The Mediterranean Region will be evaluated into two divisions as Western Mediterranean (=Antalya Part) and Eastern Mediterranean (=Adana Part) with regard to some interesting properties.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 9
Finally, with the desire to be useful for entomologists studying on Turkish fauna, longicorn beetles of Mediterranean Region are evaluated and interpreted with respect to their nomenclatural, taxonomical, faunistical and zoogeographical features.
COVERED GEOLOGICAL AREA OF THE PRESENT WORK The research area of the present work covers the Mediterranean Region. As see below, the Mediterranean Region has two parts. 1. Antalya Part (=Western Mediterranean) including three provinces, namely Antalya, Burdur and Isparta bordered by Aegean Region in the West, Adana Part of Mediterranean Region of Turkey (=Eastern Mediterranean) in the East, Aegean and Central Anatolian Regions in the North and Mediterranean Sea in the South. 2. Adana Part (=Eastern Mediterranean) including six provinces as Adana, Hatay, Ġçel, KahramanmaraĢ, Kilis and Osmaniye bordered by Antalya Part of Mediterranean Region of Turkey (=Western Mediterranean) in the West, South-Eastern Anatolian Region in the East, Central Anatolian and Eastern Anatolian Regions in the North and Mediterranean Sea and Syria in the South.
As given in Part I, Turkey is divided into seven regional parts commonly. The Mediterranean Region includes two parts as follows: Mediterranean Region (South and South-West Anatolia)
1. Antalya Part [=Western Mediterranean] (including Burdur, Isparta and
Antalya provinces)
2. Adana Part [=Eastern Mediterranean] (including Ġçel, Adana, Osmaniye, Hatay, KahramanmaraĢ and Kilis provinces)
ARRANGEMENT OF INFORMATION
Information in the present text is given in the following order: The subfamily and the tribe names are given simply. For the genus and subgenus names, the type species are provided under the taxon
names. For each species, the reported Mediterranean Region as connected with the geological
area covered by the present work using the symbols (*WM), (*EM) are given just under the taxon names.
For each species, except examined species in Parts I-III, the whole subspecies are provided under the taxon names.
The data, except examined species in Parts I-III, Other names, Material examined, Records in Turkey, Distribution, Remarks and Chorotype under the title for each taxon is given. The data of examined species occur in both Parts I-III and Part IV: Mediterranean Region, Records in Turkey, Records in Mediterranean Region, Remarks and Chorotype under the title for each taxon is given. Other names. In these parts, as possible as the whole other names including all infraspecific names (synonym, variety, morpha, form, aberration etc.) are provided. Records in Turkey. Except the examined species in Parts I-III, on the first line are given in paranthesis abbreviations of the provinces and lands in Turkey. These parts include previous records that have been given by various authors in different literatures. The whole records are evaluated as only concerning province and locality in the related reference. Each record is accompanied by the author‘s name and publication date of the related reference. For the examined species in Parts I-III, on the first line are given (in paranthesis) only abbreviations of the provinces and lands in Turkey. Records in Mediterranean Region. Under this title, for the examined species in Parts I-III, the distributions data only related to the Mediterranean Region among the records in the whole of Turkey which presented in Parts I-III. Distribution. In these parts, the whole distribution areas in the world are provided for each taxon except the examined species in Parts I-III.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 10
Remarks. In these parts, taxonomical and nomenclatural problems are discussed for some taxons and are given regional and general distribution range in Turkey chiefly, except the examined species in Parts I-III. Chorotype. The present zoogeographical characterization is based on the chorotype classification of Anatolian fauna, recently proposed by Vigna Taglianti et al. (1999). In the text, as far as possible one chorotype description can be identificated for each taxon. But this kind of description can not be possible for some taxons, so two or more chorotypes are used for them. Maps. The maps are given in the Appendix. For each taxon, a map showing distribution patterns in Turkey is used. On the maps, the black colored parts are represented the European Turkey records in Althoff & Danilevsky (1997). So Edirne province is black colored representatively. The other distribution patterns in Turkey are shown with simple black colored parts as seen on maps.
CLASSIFICATION
In this paper, classification and nomenclature of the longhorned beetles suggested by Althoff & Danilevsky (1997), Sama (2002), Danilevsky (2010) and Löbl & Smetana (2010) are followed chiefly. Within the subfamilies all genera are listed in the same order in Danilevsky (2010). Within the genera the species are listed alphabetically. Each name of a species or subspecies is accompanied by the author‘s name and description date.
ABREVIATIONS OF THE PROVINCES AND LANDS IN TURKEY
ADANA (AD) ADIYAMAN (ADY) AFYON (AF) AĞRI (AG) AKSARAY (AK) AMASYA (AM) ANKARA (AN) ANTALYA (ANT) ARDAHAN (AR) ARTVĠN (ART) AYDIN (AY) BALIKESĠR (BL) BARTIN (BR) BATMAN (BA) BAYBURT (BY) BĠLECĠK (BI) BĠNGÖL (BN) BĠTLĠS (BT) BOLU (BO) BURDUR (BU) BURSA (BS) ÇANAKKALE (CA) ÇANKIRI (CN) ÇORUM (CO) DENĠZLĠ (DE) DĠYARBAKIR (DI) DÜZCE (DU) EDĠRNE (ED) ELAZIĞ (EL) ERZĠNCAN (ER) ERZURUM (EZ) ESKĠġEHĠR (ES) GAZĠANTEP (GA) GĠRESUN (GI) GÜMÜġHANE (GU) HAKKARĠ (HA) HATAY (HT) IĞDIR (IG) ISPARTA (IP) ĠÇEL (IC) ĠSTANBUL (IS) ĠZMĠR (IZ)
KAHRAMANMARAġ (KA) KARABÜK (KR) KARAMAN (KM) KARS (KAR) KASTAMONU (KS) KAYSERĠ (KY) KIRIKKALE (KI) KIRKLARELĠ (KK) KIRġEHĠR (KIR) KĠLĠS (KL) KOCAELĠ (KO) KONYA (KN) KÜTAHYA (KU) MALATYA (MA) MANĠSA (MN) MARDĠN (MR) MUĞLA (MG) MUġ (MU) NEVġEHĠR (NE) NĠĞDE (NI) ORDU (OR) OSMANĠYE (OS) RĠZE (RI) SAKARYA (SA) SAMSUN (SM) SĠĠRT (SI) SĠNOP (SN) SĠVAS (SV) ġANLIURFA (SU) ġIRNAK (SK) TEKĠRDAĞ (TE) TOKAT (TO) TRABZON (TB) TUNCELĠ (TU) UġAK (US) VAN (VA) YALOVA (YA) YOZGAT (YO) ZONGULDAK (ZO) THRACE (EUROPEAN TUR.) (TRA) TURKEY (TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 11
Family CERAMBYCIDAE
Subfamily PRIONINAE
Tribe ERGATINI
Ergates Audinet-Serville, 1832 [Type sp.: Prionus serrarius Panzer, 1793
= Cerambyx faber Linnaeus, 1760]
Ergates faber (Linnaeus, 1760) (*WM, EM)
Records in Turkey: (AN-ANT-ART-BO-BS-DU-KA-KS-KO-SN-TB-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Sarısu Forest / KaĢ (Sütleğen) / Central (Tosun, 1975); Mediterranean Region (Çanakçıoğlu, 1983, 1993); Mediterranean Region, KahramanmaraĢ prov.: BaĢkonuĢ (Kanat, 1998); Antalya prov.: Central / KaĢ (Alkan, 2000). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Callergates Lameere, 1904 [Type sp.: Ergates gaillardoti Chevrolat, 1854]
Callergates gaillardoti (Chevrolat, 1854)
(*WM, EM) Other names: akbesianus Pic. Records in Turkey: (AD-ANT-AY-HT-IC-KN-MG-OS-TUR) Records in Turkey: Hatay prov.: Akbez as the type loc. of Ergates akbesianus Pic, 1900 (Pic, 1900); Antalya prov.: Bey Dağları, Kumköy, Adana prov.: KarataĢ (Demelt, 1963); Ġçel prov.: Namrun (Svacha & Danilevsky, 1986); Antalya prov., Adana prov. (Öymen, 1987); Antalya prov.: Alanya (Güzelbağ), Adana prov.: Kozan (Feke) (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Rapuzzi, 2000); Antalya prov.: Alanya (Cırlasun bridge, Çayarası plateau-Sarımut bridge) (Özdikmen & Çağlar, 2004); Aydın prov.: Yenihisar, Adana prov.: (Çukurova Üniv. Campus of Balcalı), Muğla prov.: Köyceğiz (Özdikmen, 2006); Antalya prov.: Manavgat (Demirciler village) (Özdikmen & Demir, 2006); Konya prov.: Beyreli, Antalya prov.: Çayarası-Alanya Sarımut bridge env. as C. akbesianus Pic (Özdikmen & Turgut, 2009); Osmaniye prov.: Zorkun (Özdikmen & Turgut, 2009); Antalya prov.: Çayarası-Alanya Sarımut bridge env., Konya prov.: Beyreli (Özdikmen & Turgut, 2010). Distribution: Europe (Greece: Rodos and Samos Islands), Turkey, Cyprus, Syria, Lebanon, N-Africa (Egypt). Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S and SW Turkey. C. akbesianus (Pic, 1900) was regarded as a synonym of the species by Drumont & Komiya in Löbl & Smetana (2010). Chorotype: E-Mediterranean.
Tribe MACROTOMINI
Prinobius Mulsant, 1842 [Type sp.: Prinobius myardi Mulsant, 1842]
Prinobius myardi Mulsant, 1842
(*WM, EM) Records in Turkey: (AD-ANT-ART-AY-BU-CA-IC-IS-IZ-KS-MG-TO-TB) Records in Mediterranean Region: Adana prov.: Dörtyol as Macrotoma scutellaris (Bodenheimer, 1958); Adana prov. (Gül-Zümreoğlu, 1975); Adana prov. as Macrotoma

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 12
scutellaris (Öymen, 1987); Adana prov.: KarataĢ as Prinobius scutellaris (Adlbauer, 1992); Adana prov. (Lodos, 1998); Burdur prov.: Bucak, Antalya prov.: Kemer / KaĢ-Gürsu / Çakırlar - Ekici, 1971; Öymen, 1987; Tosun, 1975; Yüksel, 1996 (Ex. Alkan, 2000); Antalya prov.: Beldibi / Manavgat / Serik, Ġçel prov.: Erdemli (Tozlu et al., 2002); Antalya prov.: Alanya (Çayarası plateau-Sarımut bridge) (Özdikmen & Çağlar, 2004). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. According to Drumont & Komiya in Löbl & Smetana (2010), atropos Chevrolat, 1854 is a subspecies of the species. It distributes in Cyprus and Middle East. Chorotype: Turano-Europeo-Mediterranean.
Tribe REMPHANINI
Rhaesus Motschulsky, 1875 [Type sp.: Rhaesus persicus Motschulsky, 1875
= Prionus serricollis Motschulsky, 1838]
Rhaesus serricollis (Motschulsky, 1838) (*WM, EM)
Records in Turkey: (AD-ANT-BI-BU-DE-DU-HT-IC-IS-IZ-KA-KN-MG-OS-TRA-TUR) Records in Mediterranean Region: Antalya prov. : Alanya (Demelt, 1963); Antalya prov.: Toros Mountains (Elmalı) (Villiers, 1967); Antalya prov.: Alanya (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983; Çanakçıoğlu & Mol, 1998); Antalya prov.: Alanya (Svacha & Danilevsky, 1986); Antalya prov.: Alanya / Bambus Camp (Adlbauer, 1988); KahramanmaraĢ prov. (Kanat, 1998); Antalya prov.: Alanya (Lodos, 1998); Adana prov.: Balcalı / KarataĢ, Antalya prov.: Central / Çaltıcak / Finike (Turunçova) / Serik, Burdur prov.: Central, Hatay prov.: Central / Ġskenderun (Cırtıman), Ġçel prov.: Erdemli / Tarsus, Osmaniye prov.: Kadirli (Kabayar) (Tozlu et al., 2002); Ġçel prov.: Erdemli (Karahasanlı village) (Özdikmen, 2006); Osmaniye prov.: Bahçe road (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Irano-Mediterranean + Balkano-Anatolian).
Tribe AEGOSOMATINI
Aegosoma Audinet-Serville, 1832
[Type sp.: Cerambyx scabricornis Scopoli, 1763]
Aegosoma scabricorne (Scopoli, 1763) (*WM, EM)
Records in Turkey: (ANT-BL-BR-GU-IP-IS-KA-KN-KR-OS-SM-VA-TRA-TUR) Records in Mediterranean Region: KahramanmaraĢ prov.: Andırın as Megopis scabricornis (Adlbauer, 1992); Antalya prov.: Central, Isparta prov.: Eğirdir (Ağıl) (Tozlu et al., 2002); Antalya prov.: Alanya (Çayarası plateau-Sarımut bridge) (Özdikmen & Çağlar, 2004); Antalya prov.: TaĢkent-Alanya road: Exit of Karapınar, Konya prov.: TaĢkent-Alanya road and TaĢkent: AfĢar town (Özdikmen & Turgut, 2010); Osmaniye prov.: Bahçe road and Zorkun road (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Tribe PRIONINI
Prionus Geoffroy, 1762 [Type sp.: Cerambyx coriarius Linnaeus, 1758]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 13
Prionus coriarius (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AN-ANT-ART-AY-BL-BO-BU-HT-KA-KK-KN-KO-KS-OS-RI-SN-TB-TRA-TUR) Records in Mediterranean Region: Burdur prov.: Bucak (Ekici, 1971); Antalya prov.: Kemer (Beldibi) / KaĢ (Gürsu) / Çakırlar forest (Tosun, 1975); Antalya prov. (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); KahramanmaraĢ prov. (Kanat, 1998); Antalya prov. (Çanakçıoğlu & Mol, 1998); Antalya prov.: KaĢ (Sinekçi village, Sinekçi Beli) (Özdikmen & Çağlar, 2004); Hatay prov.: Hassa (Söğütler) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır Mahallesi) (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki, Konya prov.: Çayarası-Alanya (Özdikmen & Turgut, 2010); Osmaniye prov.: Çiftmazı, Zorkun road and Mitisin plateau (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European + Turano-Europeo-Mediterranean.
Prionus komiyai Lorenc, 1999 (*EM)
Other names: Absent. Records in Turkey: (?HT-TUR) Turkey: ?Hatay prov. (Lorenc, 2006; Drumont & Komiya in Löbl & Smetana, 2010). Distribution: Turkey, Syria, Lebanon. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S Turkey and Middle East. Chorotype: E-Mediterranean.
Mesoprionus Jakovlev, 1887 [Type sp.: Prionus asiaticus Faldermann, 1837]
Mesoprionus besikanus (Fairmaire, 1855)
(*WM, EM) Records in Turkey: (AD-AN-ANT-BI-BS-BU-CA-DE-ER-EZ-IC-IS-IZ-KI-KL-KN-KU-KY-MG-NE-TRA-TUR-US) Records in Mediterranean Region: Antalya prov.: KaĢ (Demelt, 1963); Adana prov., Antalya prov., Burdur prov., Ġçel prov., Kilis prov. (Tozlu et al., 2002); Burdur prov.: Bucak (Özdikmen & ġahin, 2006); Antalya prov.: Manavgat (Demirciler village) (Özdikmen & Demir, 2006); Antalya prov.: Alanya (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Subfamily LEPTURINAE
Tribe RHAGIINI
Rhagium Fabricius, 1775 [Type sp.: Cerambyx inquisitor Linnaeus, 1758]
Subgenus Hagrium Villiers, 1978
[Type sp.: Rhagium bifasciatum Fabricius, 1775]
Rhagium bifasciatum Fabricius, 1775 (*WM)
Records in Turkey: (AM-ANT-ART-BO-BU-DU-EZ-GI-GU-IP-IS-KR-KAR-KS-KK-KO-NI-OR-RI-SA-SN-TB-ZO-TRA-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 14
Records in Mediterranean Region: Burdur prov.: Bucak (Sobya, Kızılgöl place) / Bucak (Kavacık forest), Antalya prov.: Akseki (Zomana place) / Akseki (Emirhasan forest) (Tosun, 1975); Isparta prov.: Eğirdir (Çanakçıoğlu, 1983); Antalya prov.: Akseki, Burdur prov.: Bucak, Isparta prov.: Eğirdir (Çanakçıoğlu, 1993; Çanakçıoğlu & Mol, 1998; Lodos, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European or Turano-Europeo-Mediterranean.
Subgenus Megarhagium Reitter, 1912 [Type sp.: Cerambyx sycophanta Schrank, 1781]
Rhagium elmaliense Schmid, 1999
(*WM) Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov. (Schmid, 1999); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in Antalya prov. (S Turkey). Chorotype: Anatolian.
Rhagium phrygium Daniel, 1906
(*EM) Other names: Absent. Records in Turkey: (IC-KN) Konya prov.: AkĢehir as the type loc. (Daniel, 1906; Plavilstshikov, 1915); Turkey (Winkler, 1924-1932; Sama, 2002); Ġçel prov.: Silifke and Erdemli (Adlbauer, 1988). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Rhagium syriacum Pic, 1892 (*EM)
Other names: Absent. Records in Turkey: (IC-KN) Hatay prov.: Akbez as the type loc. (Pic, 1892); Turkey (Lodos, 1998; Sama, 2002; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S Turkey (Amanos Mts.). Chorotype: SW-Asiatic (Syro-Anatolian).
Subgenus Rhagium Fabricius, 1775 [Type sp.: Cerambyx inquisitor Linnaeus, 1758]
Rhagium inquisitor (Linnaeus, 1758)
(*WM) Records in Turkey: (AM-AN-ANT-ART-BO-BS-BU-DU-EZ-GI-GU-IS-KR-KAR-KS-OR-RI-SA-SN-TB-TRA-TUR) Records in Mediterranean Region: Burdur prov.: Bucak (Sobya and Karlık forest), Antalya prov.: Akseki (Emirhasanbeli) / Elmalı (Avlanbeli forests) (Tosun, 1975); Antalya

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 15
prov.: Akseki / Elmalı, Burdur prov.: Bucak (Çanakçıoğlu, 1983); Antalya prov.: North of Akseki (Central and Irmasan pass) (Adlbauer, 1992); Antalya prov. (Lodos, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Holarctic.
Stenocorus Geoffroy, 1762 [Type sp.: Leptura meridiana Linnaeus, 1758]
Subgenus Stenocorus Geoffroy, 1762
[Type sp.: Leptura meridiana Linnaeus, 1758]
Stenocorus auricomus (Reitter, 1890) (*EM)
Other names: Absent. Records in Turkey: (IC-OS-TUR) Cilicia (Winkler, 1924-1932); Ġçel prov.: Namrun (Demelt, 1967); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); South-Eastern Turkey (Sama, 2002); Osmaniye prov.: Boğaz plateau (Özdikmen et al., 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Anisorus Mulsant, 1862
[Type sp.: Cerambyx quercus Götz, 1783]
Anisorus heterocerus (Ganglbauer, 1882) (*WM, EM)
Other names: nigripennis Pic. Records in Turkey: (ANT-BT-IC-OS-TUR) Cilicia (Winkler, 1924-1932); Antalya prov.: Antitoros Mountains (Demelt & Alkan, 1962); Antalya prov.: Bey Mts. (Demelt, 1963); Ġçel prov.: Silifke, Gülnar, Erdemli, Kuzucubelen, Osmaniye prov.: Nurdağı pass, Antalya prov.: Alanya (Adlbauer, 1988); Ġçel prov.: Erdemli as f. nigripennis Pic, 1896 (Adlbauer, 1988); Turkey (Lodos, 1998); Bitlis prov.: Sarıkonak, Ġçel prov.: Erdemli (Tauzin, 2000); Southern Turkey (Taurus Mountains) (Sama, 2002); (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes in S and SE Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Dinoptera Mulsant, 1863 [Type sp.: Leptura collaris Linnaeus, 1758]
Dinoptera collaris (Linnaeus, 1758)
(*WM, EM) Records in Turkey: (AM-AN-ANT-ART-BO-BS-CN-EZ-IC-IP-IS-KR-KS-KO-OS-RI-SM-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir as Acmeops collaris (Demelt & Alkan, 1962); Isparta prov.: Eğirdir, Ġçel prov.: Namrun (Demelt, 1963); Ġçel prov.: Çamlıyayla (Tozlu et al., 2002); Ġçel prov.: Bolkar Mountain (Namrun) (Özdikmen et al., 2005); Antalya prov.: Alanya, Akseki (Özdikmen & Turgut, 2010); Osmaniye prov.: Zorkun road and Boğaz plateau (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 16
Chorotype: Sibero-European.
Cortodera Mulsant, 1863 [Type sp.: Grammoptera spinosula Mulsant, 1839
= Leptura humeralis Schaller, 1783]
Cortodera alpina (Ménétriés, 1832) (*WM, EM)
Records in Turkey: (AN-ANT-ART-EZ-IC-KAR-KN-MU-NI-VA-TUR) Records in Mediterranean Region: Ġçel prov.: Burna, Antalya prov.: Toros Mountains as C. xanthoptera (Bodemeyer, 1900); Ġçel prov.: Namrun as Cortodera umbripennis (Demelt, 1963). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Cortodera cirsii Holzschuh, 1975 (*WM, EM)
Other names: Absent. Records in Turkey: (AD-KN-NI-OS-TUR) Adana prov. (=Osmaniye prov.): Nurdağı pass as the type loc. (Holzschuh, 1975a); Osmaniye prov.: Nurdağı pass, Niğde prov.: Çiftehan (Adlbauer, 1988); Osmaniye prov.: Nurdağı pass (Tauzin, 2000); Osmaniye prov.: Hasanbeyli env., Nurdağı pass (Rejzek et al., 2001; Özdikmen, 2003); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Konya prov.: TaĢkent (Belpınarı), TaĢkent (Beyreli village, Gevne valey) (Özdikmen & Çağlar, 2004); Konya prov.: Gencek- Derebucak (Özdikmen & Turgut, 2010). Distribution: Turkey. Remarks: It has been reported only from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Cortodera colchica Reitter, 1890 (*WM, EM)
Records in Turkey: (AD-ADY-AK-AN-ANT-ART-BN-BU-BY-EZ-HA-KAR-KY-KN-SV-TUR) Records in Mediterranean Region: Adana prov.: Misis as C. discolor - Fuchs et Breuning, 1971 (Ex. Holzschuh, 1980); Burdur prov.: Sagalassos, Antalya prov.: Akseki (Bademli) (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Cortodera differens Pic, 1898 (*WM)
Other names: steineri Sama. Records in Turkey: (AN-ANT-KN) Ankara prov.: Kızılcahamam (Güvem village), Antalya prov.: Gevne valley (Karapınar), Konya prov.: Hadim-Beyreli road 3 rd km (Gevne valley env.) (Özdikmen & Turgut, 2008a); Antalya prov.: Gevne valley (Karapınar), Konya prov.: Hadim (Özdikmen & Turgut, 2010). Distribution: Greece, Turkey. Remarks: It has been reported only from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species probably distributes only in Western half of Turkey. Chorotype: Turano-Mediterranean (Balkano-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 17
Cortodera discolor Fairmaire, 1866 (*WM, EM)
Records in Turkey: (AK-AN-ANT-IC-IZ-KN-MA-NI-TUR) Records in Mediterranean Region: Ġçel prov.: Kırobası, Central (Özdikmen, 2003); Antalya prov.: Teke pass, Alanya-TaĢkent, Akseki (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Cortodera flavimana (Waltl, 1838) (*WM, EM)
Records in Turkey: (AD-AF-AK-AN-ANT-ART-BO-BS-BY-CN-EZ-GU-IC-IP-IS-IZ-KA-KAR-KN-KO-KR-KS-KY-NI-RI-SM-SN-SV-TO-YO-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Burna, Antalya prov.: Toros Mountains (Bodemeyer, 1900); Isparta prov.: Eğirdir (Demelt & Alkan, 1962); Isparta: prov.: Eğirdir (Demelt, 1963); Antalya prov.: Kalkan, Adana prov.: Tufanbeyli (Güzelim village env.) (Özdikmen & Çağlar, 2004); KahramanmaraĢ prov.: Nurhak (Alçiçek / Tatlar), Ekinözü, AfĢin (Tanır, YeĢiloba, Farta place and Karasalgan place), AfĢin (Emirli, Gerger), Göksun (Göksun-Çardak road / Fındık village / Küçüksu village (Göcük yaylası), Saraycık village, Göksun (Mehmetbey / SE Acıelma village / Kireçköy / Pürin pass / Değirmendere / Saçkoca place / Küçüksu plateau / Korkmaz, Central (NE Çağlayan village / Kavaklı) Pazarcık (Büyüknacar), KahramanmaraĢ-Andırın road (Körsülü bridge env.) (Özdikmen & Okutaner, 2006); Antalya prov.: Bademli pass, Teke pass, Alanya-TaĢkent, Bademli-Bakaran, Alanya (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Cortodera humeralis (Schaller, 1783) (*WM, EM)
Records in Turkey: (ANT-OS) Records in Mediterranean Region: Osmaniye prov.: Nurdağı pass as the locality of holotype of C. humeralis orientalis Adlbauer, 1988 (Adlbauer, 1988); Antalya prov.: Akseki (Cevizli) as C. humeralis orientalis Adlbauer, 1988 (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: S-European.
Cortodera imrasanica Sama & Rapuzzi, 1999 (*WM)
Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: Çakıllı pass (Imrasan pass) as the type loc. (Sama & Rapuzzi, 1999); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Cortodera omophloides Holzschuh, 1975 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Ġçel prov.: Cilician Taurus (Namrun, Tekir) as the type loc. (Holzschuh, 1975a); Ġçel prov.: Erdemli, Fındıkpınarı (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 18
Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Ġçel province in S Turkey. Chorotype: Anatolian.
Cortodera pseudomophlus Reitter, 1889 (*EM)
Other names: Absent. Records in Turkey: (EZ-KA) KahramanmaraĢ prov.: Yenicekale, Erzurum prov.: AĢkale (Adlbauer, 1992). Distribution: Turkey, Caucasus (Armenia, Azerbaijan), Iran, Turkmenistan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species probably distributes only in Eastern half of Turkey. Chorotype: Turano-Mediterranean (Turano-Anatolian).
Cortodera rubripennis Pic, 1891 (*EM)
Other names: obscura Pic. Records in Turkey: (AD-HT-KN-TUR) Hatay prov.: Akbez as the type loc. (Pic, 1891); Adana prov.: Toros Mountains, Pozantı, Bolkar Mountains (Villiers, 1959); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Konya prov.: Gencek-Derebucak (Özdikmen & Turgut, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species probably distributes only in Amanos MTS and near in S Turkey. Chorotype: Anatolian.
Cortodera syriaca Pic, 1901 (*EM)
Other names: aureopubens Pic; korbi Pic. Records in Turkey: (ADY-AK-IC-KA-MU-TUR) Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); KahramanmaraĢ prov.: Göksun, Ġçel prov.: Silifke (Gülnar) (Adlbauer, 1988); Adıyaman prov.: the peak region of Nemrut Mountain (Rejzek & Hoskovec, 1999); MuĢ prov.: Buğlan pass, KahramanmaraĢ prov.: Göksun env. (Rejzek et al., 2001); Aksaray prov.: Central (Özdikmen, 2003). Distribution: Turkey, Syria, Lebanon, Caucasus (Armenia, Azerbaijan), Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species probably distributes mostly in Eastern half of Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Tribe LEPTURINI
Grammoptera Audinet-Serville, 1835 [Type sp.: Leptura praeusta Fabricius, 1787
= Leptura ustulata Schaller, 1783]
Subgenus Grammoptera Audinet-Serville, 1835 [Type sp.: Leptura praeusta Fabricius, 1787
= Leptura ustulata Schaller, 1783]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 19
Grammoptera baudii (Stephens, 1831) (*EM)
ssp. baudii Stephens, 1831 ssp. pistacivora Sama, 1996 Other names: Absent. Records in Turkey: (HT-TUR) Hatay prov.: Yayladağı as G. baudii pistacivora (Sama & Rapuzzi, 1999); Turkey as G. baudii pistacivora (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Cyprus, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S Turkey. It is represented by the subspecies G. baudii pistacivora Sama, 1996 in Turkey. The nominotypical subspecies occurs only in Cyprus. Chorotype: E-Mediterranean.
Grammoptera merkli Frivaldszky, 1884 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-IC-MN-TUR) Anatolia (Winkler, 1924-1932); Antalya prov.: Korkuteli (Demelt & Alkan, 1962); Antalya prov.: Korkuteli, Bey Mountain (Demelt, 1963); Ġçel prov.: Erdemli, Tarsus (Çamlıyayla) (Adlbauer, 1988); Ġçel prov.: Namrun (Svacha & Danilevsky, 1988); Turkey (Lodos, 1998); Manisa prov. (Sama & Rapuzzi, 1999); Antalya prov.: Akseki (Özdikmen & Turgut, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Alosterna Mulsant, 1863 [Type sp.: Leptura tabacicolor De Geer, 1775]
Alosterna anatolica Adlbauer, 1992
(*WM) Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: S elmalı, Akseki (Yarpuz) (Adlbauer, 1992); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Alosterna tabacicolor (De Geer, 1775) (*WM)
Records in Turkey: (ANT-ART-TB-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Akseki (Yarpuz) (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 20
Vadonia Mulsant, 1863 [Type sp.: Leptura unipunctata Fabricius, 1787]
Vadonia bisignata (Brullé, 1832)
(*WM) Records in Turkey: (ANT-ART-IP-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros Mountains (Bey Mountains) (Demelt & Alkan, 1962); Antalya prov.: Bey Mountain / Alanya, Isparta prov. (Demelt, 1963). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Vadonia bitlisiensis Chevrolat, 1882 (*WM)
Records in Turkey: (ANT-BT-ER-EZ-GU-TU-VA-TUR) Records in Mediterranean Region: Antalya prov.: Akseki-Güzelsu (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian).
Vadonia ciliciensis Daniel & Daniel, 1891 (*WM)
Records in Turkey: (ANT-BU-DE-TUR) Records in Mediterranean Region: Burdur prov.: Bucak (Kavacık forest), Antalya prov.: Elmalı (Çığlıkara, Suluçukur place and Bucak forest) (Tosun, 1975); Burdur prov.: Bucak, Antalya prov.: Elmalı (Çanakçıoğlu, 1983; Alkan, 2000). Distribution: Turkey. Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Anatolian.
Vadonia danielorum Holzschuh, 1984 (*WM)
Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: Termessos as the type loc. (Holzschuh, 1984a); Antalya prov.: TaĢağıl and Termessos (Adlbauer, 1992); Turkey (Sma & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Vadonia frater Holzschuh, 1981 (*EM)
Other names: Absent. Records in Turkey: (AD-TUR) Adana prov.: Nurdağı pass the type loc. (Holzschuh, 1981a); Turkey (Sma & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Adana province in S Turkey. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 21
Vadonia moesiaca (Daniel & Daniel, 1891) (*WM)
Records in Turkey: (ANT-CN-KK-TUR) Records in Mediterranean Region: Antalya prov.: TaĢağıl (Adlbauer, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Vadonia monostigma (Ganglbauer, 1882) (*WM)
Records in Turkey: (AM-ANT-BO-KS-SM-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Bey Mountains (Antitoros) (Demelt & Alkan, 1962; Demelt, 1963). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Vadonia soror Holzschuh, 1981 (*WM, EM)
ssp. soror Holzschuh, 1981 ssp. tauricola Holzschuh, 1993 Other names: Absent. Records in Turkey: (ANT-DE-IC-TUR) Denizli prov.: Pamukkale (Holzschuh, 1981); Ġçel prov.: Silifke (Gülnar) and Kuzucubelen (Adlbauer, 1988); Antalya prov. as ssp. tauricola (Hoskovec & Rejzek, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S and SW Turkey. Chorotype: Anatolian.
Vadonia unipunctata (Fabricius, 1787) (*WM, EM)
Records in Turkey: (AF-AK-AM-AN-ANT-ART-BI-BN-BO-BT-BU-BY-CN-EL-EZ-IP-IZ-KA-KAR-KIR-KO-KR-KS-KY-KU-MA-NE-NI-OS-RI-SV-TO-US-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mountains (Bodemeyer, 1900); Isparta prov.: Eğirdir (Demelt & Alkan, 1962; Demelt, 1963); Isparta prov. (Tuatay et al., 1972); Burdur prov.: Bucak (Adlbauer, 1988); Isparta prov.: Yalvaç (Eleği village) (Özdikmen & Çağlar, 2004); Isparta prov. (Özdikmen et al., 2005); Osmaniye prov.: Yarpuz road (KarataĢ place) / YeĢil village (Hasanbeyli) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: AfĢin (Kabaağaç / Emirli (Gergel) / Göksun (Göksun-Çardak road, Gücük plateau / Mehmetbey (Özdikmen & Okutaner, 2006); Osmaniye prov.: Entry of Yarpuz (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European or Turano-Europeo-Mediterranean.
Pseudovadonia Lobanov, Danilevsky et Murzin, 1981 [Type sp.: Leptura livida Fabricius, 1777]
Pseudovadonia livida (Fabricius, 1777)
(*WM, EM) Records in Turkey: (ADY-AM-AN-ANT-ART-BI-BO-BR-BS-BT-BY-CN-ER-EZ-GA-GI-GU-HT-IC-IP-IS-IZ-KAR-KK-KO-KR-KS-MN-NI-OS-RI-SM-US-TRA-TUR) Records in Mediterranean Region: Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains / Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near as Leptura livida m. pecta (Demelt & Alkan, 1962); Antalya prov.:

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 22
Kemer / Kumluca (Yeniceköy) / Termessos / Manavgat-Sorgun, Ġçel prov.: Erdemli (Aslanlı), Osmaniye prov.: Nurdağı pass as Pseudovadonia livida pecta (Adlbauer, 1988); Antalya prov.: Arapsuyu (Tozlu et al., 2002); Isparta prov.: Yalvaç (Bağkonak, Sultan mountains) (Özdikmen & Çağlar, 2004); Isparta prov.: Gölcük (Çakıören) (Özdikmen et al., 2005); Osmaniye prov.: Zorkun plateau road (OlukbaĢı place) / Yarpuz road (KarataĢ place) / Bahçe (Yaylalar village), Gaziantep prov.: Nurdağı (plateau of Kazdere village) / KuĢçubeli pass, Hatay prov.: Hassa (Zeytinoba village, Aktepe) (Özdikmen & Demirel, 2005); Antalya prov.: Irmasan pass, Hatay prov.: Yayladağı, Ġçel prov.: Erdemli-Güzeloluk / Güzeloluk / Silifke (Ortagören to Mut) (Malmusi & Saltini, 2005); Antalya prov.: GündoğmuĢ, SeydiĢehir-Antalya road, Akseki, Akseki-Güzelsu, Ġbradı, Exit of Ġbradı 5th km, Irmasan, Karapınar, Alanya (KeĢbelen plateau, Gökbel plateau), Akseki (Mahmutlu village env., Güçlüköy env., Murtiçi-Güzelsu, Güzelsu village) (Özdikmen & Turgut, 2010); Gaziantep prov.: Akbez, Hatay prov.: Entry of Belen, Osmaniye prov.: Zorkun road (Çiftmazı), Hasanbeyli (Kalecikli village), Karaçay, Akyar village, Zorkun road (Fenk plateau), Yarpuz road (Yukarı Haraz plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Anoplodera Mulsant, 1839 [Type sp.: Leptura sexguttata Fabricius, 1775]
Subgenus Anoplodera Mulsant, 1839
[Type sp.: Leptura sexguttata Fabricius, 1775]
Anoplodera rufipes (Schaller, 1783) (*EM)
Records in Turkey: (AN-BN-BO-BS-EZ-GU-IC-KK-KS-OR-RI-TB-TUR) Records in Mediterranean Region: Ġçel prov.: Erdemli (Adlbauer, 1988); Southern Turkey as A. rufipes lucidipes Sama, 1999 (Sama, 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Anoplodera sexguttata (Fabricius, 1775) (*EM)
Records in Turkey: (GU-HT-KO-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as Anoplodera 6-guttata (Fairmaire, 1884). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European + Europeo-Mediterranean.
Stictoleptura Casey, 1924 [Type sp.: Leptura cribripennis LeConte, 1859]
Subgenus Aredolpona Nakane & Ohbayashi, 1957
[Type sp.: Leptura rubra Linnaeus, 1758]
Stictoleptura rubra (Linnaeus, 1758) (*WM)
Records in Turkey: (ANT-ART-BO-BU-DU-IS-KO-SA-SN-TUR) Records in Mediterranean Region: Antalya prov.: Serik (Belek forest), Burdur prov.: Bucak (Eğrice place / Karlık forest) (Tosun, 1975); Antalya prov. (Erdem & Çanakçıoğlu, 1977); Burdur prov.: Bucak (Çanakçıoğlu, 1983); Antalya prov. (Lodos, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 23
Subgenus Stictoleptura Casey, 1924 [Type sp.: Leptura cribripennis LeConte, 1859]
Stictoleptura cordigera (Füsslins, 1775)
(*WM, EM) Records in Turkey: (ADY-AK-AN-ANT-ART-BL-BN-BO-BT-BU-CA-DE-ED-EZ-GA-GU-HT-IC-IS-IZ-KA-KK-KN-KO-MG-MN-MU-NE-NI-OS-TE-TU-YA-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as L. hastata (Fairmaite, 1881); Antalya prov.: Alanya (Demelt & Alkan, 1962; Demelt, 1963); Hatay prov.: Amanos Mountains (Nurdağı pass), Ġçel prov.: Alata (Villiers, 1967); Osmaniye prov.: Central (Öymen, 1987); Antalya prov.: Alanya, Ġçel prov.: Anamur / Erdemli (Adlbauer, 1988); Ġçel prov., Antalya prov. (Lodos, 1998); Antalya prov.: Elmalı (Tauzin, 2000); Antalya prov.: Kemer (Olimpos mountain) / KaĢ (Gömbe, Sinekçi village, Sinekçibeli) (Özdikmen & Çağlar, 2004); Osmaniye prov.: Zorkun plateau road (OlukbaĢı place) / Yarpuz / Yarpuz road (KarataĢ place), KahramanmaraĢ prov.: Türkoğlu (Kaledibi village), Gaziantep prov.: KuĢçubeli pass, Hatay prov.: Hassa (Akbez, GeneluĢağı village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Nurhak (Nurhak-Malatya road, Tatlar) / Pazarcık (Aksu bridge / Sakarkaya village (Kısık env. / Göynük env.) / Sakarkaya-Çağlayancerit road) / Göksun (Mehmetbey) / Çağlayancerit (Ç.Cerit-Bozlar road, Aksu) (Özdikmen & Okutaner, 2006); Antalya prov.: Ġbradı, Exit of Ġbradı 5th km (Gevenli pass env.), Akseki (Ceceler village), Alanya (Karapınar village, DikmetaĢ plateau) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Stictoleptura excisipes (Daniel & Daniel, 1891) (*WM, EM)
Records in Turkey: (AD-ANT-BL-IC-NI-OS-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros and Bey Mts. as Leptura excisipes (Demelt & Alkan, 1962; Demelt, 1963); Adana prov.: Tekir, Kozan, Himmetli, Antalya prov.: Kemer, Manavgat, TaĢağıl, Alanya, Güzelbağ, Akseki, Ġçel prov.: Silifke (Gülnar), Kuzucubelen, Erdemli (Adlbauer, 1988); Ġçel prov.: Kuzucubelen (Tauzin, 2000); Osmaniye prov.: Hasanbeyli, Ġçel prov.: Çamlıyayla env., Aydınlar env. (Kurzawa pers. comm.); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km), GündoğmuĢ, Akseki-Manavgat road (GündoğmuĢ return), GündoğmuĢ (Senir town env.), Murtiçi-Güzelsu, Akseki-Güzelsu, Ibradı, Akseki (Irmasan), Alanya (Karapınar village), Akseki (Mahmutlu village env., Güçlüköy env., Murtiçi-Güzelsu, Güzelsu village) (Özdikmen & Turgut, 2010). Distribution: Turkey, Syria. Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. The species distributes mostly in S and W Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Stictoleptura fulva (DeGeer, 1775) (*WM, EM)
Records in Turkey: (AD-ANT-ART-BO-BR-BS-BU-HT-IC-IP-IS-IZ-KA-KK-KN-KO-KR-KS-MN-OS-ZO-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun (Demelt & Alkan, 1962; Demelt, 1963); Hatay prov.: around Dörtyol (Öymen, 1987); Isparta prov.: Eğirdir (AĢağı Gökdere), Antalya prov.: Alanya (Gökbel plateau) (Özdikmen & Çağlar, 2004); Osmaniye prov.: Yarpuz road (KarataĢ place) / entry of Yarpuz (Cebel, turn of Oruçgazi road) / Çulhalı village, Hatay prov.: Hassa (Akbez, GeneluĢağı village / Zeytinoba village (Aktepe) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı (KarataĢ) (Özdikmen & Demir, 2006); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (BaĢkonuĢ forest) (Özdikmen & Okutaner, 2006); Antalya prov.: Alanya / Mahmutlar (Gödre plateau), Alanya (Entry of Gökbel plateau), Akseki (Yarpuz env., Çukurköy-Mahmutlu, Murtiçi-Güzelsu), Ġbradı-Derebucak road (Özdikmen & Turgut, 2010); Hatay prov.: Akbez (GüzeluĢağı village),

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 24
Osmaniye prov.: Haraz plateau, Yarpuz road (Yukarı Haraz plateau), Yarpuz road (8th km), Zorkun road (Fenk plateau, Çiftmazı Gölyeri, Ürün plateau), Hasanbeyli, Cebel road (Çürükarmut plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Stictoleptura gevneensis Özdikmen & Turgut, 2008 (*WM)
Other names: Absent. Records in Turkey: (ANT) Antalya prov.: Alanya, Gevne valley (Sarımut- Çayarası) as the type loc.(Özdikmen & Turgut, 2008b). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. The species was regarded by Sama & Löbl in Löbl & Smetana (2010) as a synonym of S. rufa rufa. Chorotype: Anatolian.
Stictoleptura heydeni (Ganglbauer, 1889) (*EM)
Other names: incisipennis Reitter; maceki Holzschuh. Records in Turkey: (AD-IC-TUR) Turkey (Lobanov et al., 1981; Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama & Rapuzzi, 2000; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: Erdemli (Adlbauer, 1988); Adana prov.: Pozantı (Tekir plateau) (Özdikmen, 2006). Distribution: Turkey, Syria, Israel, Lebanon, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S Turkey. According to Sama (1991), Leptura ustulata Ménétriés, 1832 must be replaced with Leptura heydeni Ganglbauer, 1889. However, Leptura ustulata Ménétriés, 1832 was regarded by Sama & Löbl in Löbl & Smetana (2010) as a synonym of S. tonsa. According to Sama & Löbl in Löbl & Smetana (2010), Stictoleptura heydeni (Ganglbauer, 1889) is absent in Transcaucasia as the species (distributed in Palestine and neihbour regions of Turkey) absent in East Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian) or SW-Asiatic (Irano-Anatolian).
Stictoleptura pallens (Brullé, 1832) (*EM)
Records in Turkey: (AD-KA-KK-KO-NI-TRA-TUR) Records in Mediterranean Region: Adana prov.: Pozantı (Bodemeyer, 1900); KahramanmaraĢ prov.: Central (Karbasan village, Körsülü bridge env.) (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Stictoleptura pallidipennis (Tournier, 1872) (*?WM)
Records in Turkey: (ANT-ART-BT-EZ-GI-IS-RI-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros Mountains (Demelt & Alkan, 1962; Demelt, 1963). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Probably the species distributes in North-Eastern Turkey. So, the records of Demelt & Alkan, 1962 and Demelt, 1963 should be very likely a different taxon.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 25
Chorotype: SW-Asiatic (Anatolo-Caucasian).
Stictoleptura rufa (Brullé, 1832) (*WM, EM)
Records in Turkey: (ANT-BI-BY-ER-EZ-GU-IC-TU-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros (Demelt & Alkan, 1962); Antalya prov.: Alanya (Demelt, 1963); Ġçel prov.: Silifke (Gülnar) (Adlbauer, 1988); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km) as S. rufa rufa (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Apenninian).
Stictoleptura sambucicola (Holzschuh, 1982) (*EM)
Other names: Absent. Records in Turkey: (IC-KA-TUR) Ġçel prov.: Tarsus (Çamlıyayla) (Adlbauer, 1988); KahramanmaraĢ prov.: Central (Yenicekale road) and Andırın (Parmaksız plateau) (Özdikmen & Okutaner, 2006); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species distributes only in S Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Stictoleptura tesserula (Charpentier, 1825) (*EM)
Records in Turkey: (EZ-KN-KR-KS-OS-RI-TUR) Records in Mediterranean Region: Osmaniye prov.: Yarpuz road (Yukarı Haraz plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Stictoleptura tonsa (Daniel & Daniel, 1891) (*EM)
Records in Turkey: (AM-ART-BR-ER-EZ-GU-KAR-KO-OS-TB-YA-TUR) Records in Mediterranean Region: Osmaniye prov.: Central (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Stictoleptura tripartita (Heyden, 1889) (*EM)
Records in Turkey: (ADY-ER-EZ-HA-KA-KAR-KY-MA-RI-TU-YO) Records in Mediterranean Region: KahramanmaraĢ prov.: Göksun (Mehmetbey) (Özdikmen & Okutaner, 2006). Distribution: Turkey, Syria, Iraq, Iran. Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Irano-Anatolian + Irano-Palaestinian + Mesopotamian + Syro-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 26
Anastrangalia Casey, 1924 [Type sp.: Leptura sanguinea LeConte, 1859]
Anastrangalia dubia (Scopoli, 1763)
(*EM) Records in Turkey: (AF-ART-BI-BO-BR-CN-CO-GI-GU-KAR-KIR-KN-KR-KS-MG-NE-OS-RI-SN-TB-TO-ZO-YO-TUR) Records in Mediterranean Region: Osmaniye prov.: Yarpuz road (KarataĢ place) (Özdikmen & Demirel, 2005) Osmaniye prov.: Hasanbeyli (Kalecikli village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Anastrangalia montana (Mulsant & Rey, 1863) (*WM, EM)
Records in Turkey: (AD-ANT-CA-HT-IC-IZ-KN-OS-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros (Demelt & Alkan, 1962); Antalya prov.: Bey Mountain, Ġçel prov.: Namrun (Demelt, 1963); Antalya prov.: TaĢağıl, Adana prov.: Kozan (Adlbauer, 1988); Ġçel prov.: Gülek (Toros Mts.) – Ganglbauer, 1885, Hatay prov.: Amanos Mts. (Akbez) – Pic, 1892, Ġçel prov.: Çamlıyayla, Antalya prov.: Çakıllı pass and Adana prov. - Adlbauer, 1988 (Sama, 1994b); Antalya prov.: Perge (Gülek) (Tauzin, 2000); Antalya prov.: Finike (Alakır dam) (Özdikmen & Çağlar, 2004); Osmaniye prov.: Yarpuz road (KarataĢ place), Çulhalı village (Özdikmen & Demirel, 2005); Ġçel prov.: Mut-Silifke road (exit of Karabağ) (Özdikmen, 2006); Antalya prov.: GündoğmuĢ, SeydiĢehir-Antalya road, Akseki (Bademli beli-Tekebeli) (Özdikmen & Turgut, 2010); Osmaniye prov.: Hınzırlı plateau (Kalaycıbatıran), Yarpuz road (Yukarı Haraz plateau, Mitisin plateau), Zorkun road (Fenk plateau), Zorkun-Karıncalı-Hassa road (Küllü plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: E-Mediterranean (Palaestino-Cyprioto-Taurian + Aegean).
Pedostrangalia Sokolov, 1897 [Type sp.: Pedostrangalia kassjanovi Sokolov, 1897
= Leptura imberbis Menetries, 1832]
Subgenus Pedostrangalia Sokolov, 1897 [Type sp.: Pedostrangalia kassjanovi Sokolov, 1897
= Leptura imberbis Menetries, 1832]
Pedostrangalia revestita (Linnaeus, 1767) (*WM)
Records in Turkey: (ANT-GU) Records in Mediterranean Region: Antalya prov.: Manavgat / Alanya (Demelt & Alkan, 1962); Antalya prov.: Alanya (Kargıcak) Özdikmen & Demir, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Subgenus Neosphenalia Löbl, 2010 [Type sp.: Leptura verticalis Germar, 1822]
Pedostrangalia emmipoda (Mulsant, 1863)
(*WM, EM) Records in Turkey: (AD-ADY-ANT-BN-BT-BU-CA-EL-GA-HA-HT-IC-IP-IZ-KA-KN-MA-MN-MU-NI-OS-TU-TUR-US)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 27
Records in Mediterranean Region: Adana prov.: Pozantı (Bodemeyer, 1900); Isparta prov.: Eğirdir, Antalya prov.: Antitoros (Bey Mountains)( Demelt & Alkan, 1962); Antalya prov.: Manavgat, Alanya, Bey Mts., Isparta prov.: Eğirdir, Ġçel prov.: Namrun (Demelt, 1963); Adana prov.: Misis (Fuchs et Breuning, 1971); Adana prov.: Bolkar Mts. (Tuatay et al., 1972); Antalya prov.: Yeni Karaman, Termessos, Alanya, Ġçel prov.: Tarsus (Çamlıyayla), Silifke (Gülnar), Kuzucubelen, Erdemli, Osmaniye prov.: Nurdağı pass, KahramanmaraĢ prov., Adana prov.: Kozan (Adlbauer, 1988); Ġçel prov.: Çamlıyayla (Sama, 1996); Antalya prov.: Finike, Alanya, Alakır dam, Beymelek city, Gömbe, KaĢ, Burdur prov.: Bucak (Özdikmen & Çağlar, 2004); Ġçel prov.: Bulgar Mt. (Namrun) (Özdikmen et al., 2005); Osmaniye prov.: Zorkun plateau road, Yarpuz, entry of Yarpuz, YeĢil village (Hasanbeyli), Gaziantep prov.: KuĢçubeli pass, Hatay prov.: Hassa (Akbez), Ġskenderun (Kozaklı village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road, Göksun (Andırın-Göksun road) (Özdikmen & Okutaner, 2006); Antalya prov.: Ġbradı, Exit of Ġbradı 5th km (Gevenli pass env.), Alanya (Karapınar village, DikmetaĢ plateau, Karapınar-Sarımut), Akseki (Mahmutlu village env., Murtiçi-Güzelsu, Güzelsu village, Bademli beli-Tekebeli) (Özdikmen & Turgut, 2010); Hatay prov.: Akbez (Gülpınarı plateau, GüzeluĢağı village), Osmaniye prov.: Zorkun Road (Çiftmazı Gölyeri), Yarpuz road (Yukarı Haraz plateau, 8th km), Cebel road (Çürükarmut plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Syro-Anatolian).
Pedostrangalia verticalis (Germar, 1822) (*WM)
Records in Turkey: (ANT-EZ-GU-IZ-TUR) Records in Mediterranean Region: Antalya prov.: Alanya (Karapınar village) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Apenninian).
Etorofus Matsushita, 1933 [Type sp.: Etorofus variicornis Matsushita, 1933 (nec Dalman, 1817)
= Etorofus nemurensis Matsushita, 1933]
Subgenus Etorofus Matsushita, 1933 [Type sp.: Etorofus variicornis Matsushita, 1933 (nec Dalman, 1817)
= Etorofus nemurensis Matsushita, 1933]
Etorufus pubescens (Fabricius, 1787) (*WM)
Records in Turkey: (BU-KU-US-YO-ZO-TUR) Records in Mediterranean Region: Burdur prov.: YeĢilova (Çaltepe) (Özdikmen & Çağlar, 2004). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Carlandrea Sama & Rapuzzi, 1999 [Type sp.: Strangalia syriaca Pic, 1891]
Carlandrea syriaca (Pic, 1891)
(*WM, EM) Other names: Absent. Records in Turkey: (ANT-IC-TU-TUR) Antalya prov.: Antitoros (Bey Mountains) (Demelt & Alkan, 1962; Demelt, 1963); Tunceli prov.: Ovacık (Fuchs & Breuning, 1971); Ġçel prov.: Erdemli (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 28
Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in S Turkey. Chorotype: Anatolian.
Pachytodes Pic, 1891 [Type sp.: Leptura cerambyciformes Schrank, 1781]
Pachytodes erraticus (Dalman, 1817)
(*WM, EM) Records in Turkey: (AF-AM-AN-ANT-ART-BI-BO-BR-BS-BT-CN-CO-EL-ER-EZ-GA-GU-HT-IP-IS-IZ-KAR-KK-KO-KN-KR-KS-MN-MU-RI-SM-SN-SV-TB-TO-TU-YO-ZO-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as Judolia erratica (Fairmaire, 1884); Isparta prov. (Tuatay et al., 1972); Hatay prov.: Dörtyol env. (Öymen, 1987); Antalya prov.: Akseki (Adlbauer, 1992); Isparta prov. (Lodos, 1998); Antalya prov.: Finike (Alakır dam) (Özdikmen & Çağlar, 2004); Isparta prov. (Özdikmen et al., 2005); Antalya prov.: Alanya (Karapınar village) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Rutpela Nakane et Ohbayashi, 1957 [Type sp.: Leptura maculata Poda von Neuhaus, 1761]
Rutpela maculata (Poda von Neuhaus, 1761)
(*WM, EM) Records in Turkey: (AD-AM-ANT-ART-BL-BN-BO-BS-CA-CN-HT-IC-IS-KK-KN-KO-KR-KS-MA-MU-OS-RI-SM-SN-SV-TB-TO-TU-VA-TUR) Records in Mediterranean Region: Antalya prov.: Irmasan pass as type locality of R. maculata irmasanica Sama, 1996 and Akseki (Sama, 1996a); Hatay prov.: Akbez, Ġçel prov.: Namrun (Bolkar Mountains) (Sama, 1996a); Osmaniye prov.: Zorkun plateau road (OlukbaĢı place) / Yarpuz road (KarataĢ place), Hatay prov.: Dörtyol (TopaktaĢ plateau) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı (KarataĢ) (Özdikmen & Demir, 2006); Osmaniye prov.: Küllü-Islahiye road (Hınzırlı plateau), Zorkun road (Çiftmazı Gölyeri), Yarpuz road (Yukarı Haraz plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Stenurella Villiers, 1974 [Type sp.: Leptura melanura Linnaeus, 1758]
Stenurella bifasciata (Müller, 1776)
(*WM, EM)
Records in Turkey: (AD-AF-AK-AM-AN-ANT-ART-BI-BN-BO-BR-BS-BT-BU-CA-CN-CO-ER-EZ-GA-GU-HT-IC-IZ-KA-KK-KN-KO-KR-KS-KY-MG-MN-NE-OS-RI-SM-TB-US-YA-YO-ZO-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as Strangalia cruciata (Fairmaire, 1884); Antalya prov.: near Central (Demelt & Alkan, 1962); Antalya prov.: Central / Alanya (Demelt, 1963); Antalya prov.: Kemer / Manavgat, Ġçel prov.: Anamur (Adlbauer, 1988); Antalya prov.: Alanya (Mahmutlar) / Finike (Alakır dam) / KaĢ (Gömbe, Sinekçi village, Sinekçibeli) (Özdikmen & Çağlar, 2004); Burdur prov.: Ağlasun (Özdikmen et al., 2005); Osmaniye prov.: Zorkun plateau / Zorkun plateau road (Ürün plateau / OlukbaĢı place) / Yarpuz / Yarpuz road (KarataĢ place) / Bahçe (Buğdaycık village) / Ömerli village (Asar plateau) / turn of Düziçi road / Düziçi (Karasi village / Berka dam) / exit of Çotlu village, KahramanmaraĢ prov.: Sumaklı village / Türkoğlu (Doluca village / Kaledibi

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 29
village), Gaziantep prov.: Nurdağı (plateau of Kazdere village) / KuĢçubeli pass, Hatay prov.: Hassa (Akbez, GeneluĢağı village / Zeytinoba village) / Kırıkhan (plateau of Ceylanlı village) / Belen (Güzelyayla) / Ġskenderun (Kozaklı village) / Antakya (Seldiren village) / Dörtyol (Yahyalı plateau) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env./ BaĢkonuĢ forest) / KahramanmaraĢ-Göksun road (entry of Tekir) / Andırın-Çokak road (Çınar district / Parmaksız plateau / Akifiye) / Andırın (Çokak-Geben road, Ġtoğlu place / Andırın-Geben road) / Göksun (Mehmetbey) / Pazarcık (Osmandede village) (Özdikmen & Okutaner, 2006); Adana prov.: Pozantı (entry of Fındıklı / Pozantı, Tekir plateau) (Özdikmen, 2006); Antalya prov.: Alanya / Mahmutlar (Gödre plateau), Akseki-Manavgat road (GündoğmuĢ return), GündoğmuĢ road, Ġbradı, Alanya (DikmetaĢ plateau, Sarımut-Çayarası, Sarımut-Karapınar, Entry of Gökbel plateau, Gökbel plateau), Ġbradı-BeyĢehir road (Gevenli pass env.) as S. bifasciata nigrusuturalis (Özdikmen & Turgut, 2010); Osmaniye prov.: Zorkun road (Çiftmazı Gölyeri, Mitisin plateau, Ürün plateau, Fenk plateau, Karacalar village, Çiftmazı), Hasanbeyli, Karaçay, Zorkun-Karıncalı-Hassa road (Küllü plateau), ArslantaĢ-Osmaniye road (Kazmaca village), Cebel road (Çürükarmut plateau), Yarpuz road (8th km, Yukarı Haraz plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European + SW-Asiatic chorotypes.
Stenurella jaegeri (Hummel, 1825) (*EM)
Records in Turkey: (GU-HT-KA-RI-TB-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); KahramanmaraĢ prov.: Göksun (Tauzin, 2000). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic + Turanian (Ponto-Caspian).
Stenurella melanura (Linnaeus, 1758) (*WM)
Records in Turkey: (ANT-ART-BO-BS-CA-IS-IZ-KO-KS-OR-TUR) Records in Mediterranean Region: Antalya prov.: Alanya (Demelt, 1963); Antalya prov.: Akseki (Cevizli) (Adlbauer, 1992); Antalya prov. (Lodos, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Moreover, these old records probably can belong to S. pamphyliae. Chorotype: Sibero-European.
Stenurella pamphyliae Rapuzzi & Sama, 2009 (*WM)
Other names: Absent. Records in Turkey: (ANT) Antalya prov.: W GündoğmuĢ as the locality of holotype, Antalya prov.: W GündoğmuĢ, Alanya, Lyk. Taurus, 6,5 km N Mahmuttar, Alanya (Obaalacami), Alanya (Dim Çayı) as the localities of paratypes (Rapuzzi & Sama, 2009); Antalya prov.: Alanya (DikmetaĢ plateau) as S. melanura (Özdikmen & Turgut, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species distributes only in Antalya province in S Turkey. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 30
Subfamily NECYDALINAE
Tribe NECYDALINI
Necydalis Linnaeus, 1758 [Type sp.: Necydalis major Linnaeus, 1758]
Subgenus Necydalis Linnaeus, 1758
[Type sp.: Necydalis major Linnaeus, 1758]
Necydalis hadullai Szallies, 1994 (*WM)
Other names: Absent. Records in Turkey: (IP) Isparta prov.: Kovada, S of Eğridir as the type loc. (Szallies, 1994). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Isparta province in SW Turkey. Chorotype: Anatolian.
Subfamily ASEMINAE
Tribe SAPHANINI
Oxypleurus Mulsant, 1839 [Type sp.: Oxypleurus nodieri Mulsant, 1839]
Oxypleurus nodieri Mulsant, 1839
(*WM) Other names: scutellaris A. Costa; bewickii Wollaston; pinicola Wollaston. Records in Turkey: (ANT-TUR) Antalya prov.: Bey Mt. (Demelt, 1963); S Turkey (Sama, 2002); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Europe (Portugal, Spain, France, Italy, Croatia, Bosnia-Herzegovina, Greece, Ukraine), Caucasus (Georgia), Turkey, North Africa (Algeria, Morocco, Canary Islands, Madeira, Tunusia). Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from Antalya province in S Turkey. Chorotype: Mediterranean.
Tribe ANISARTHRONINI
Alocerus Mulsant, 1862 [Type sp.: Alocerus fulvus Mulsant, 1862 = Callidium moesiacum Frivaldsky, 1837]
Alocerus moesiacus (Frivaldsky, 1837)
(*WM) Records in Turkey: (ANT-IP-KK-TUR) Records in Mediterranean Region: Isparta prov.: Eğridir (Kovada) (Adlbauer, 1992); Antalya prov. (Tozlu et al., 2002). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Mediterranean.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 31
Tribe ASEMINI
Arhopalus Serville, 1834 [Type sp.: Cerambyx rusticus Linnaeus, 1758]
Arhopalus ferus (Mulsant, 1839)
(*WM, EM) Records in Turkey: (AD-AM-AN-ANT-AY-BI-BO-CA-ES-HT-IZ-KK-KU-MG-TO-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Antalya prov.: Gebiz (Bozburan Mountain) (Villiers, 1967); Adana prov.: Balcalı / Kozan / KarataĢ, Antalya prov.: Central / Akseki / Serik / Alanya / Topçam (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region as A. tristis. Chorotype: Palearctic.
Arhopalus rusticus (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AN-ANT-ART-BL-BO-BU-BY-DE-GU-IS-KAR-KR-KS-KU-MG-OR-OS-RI-SM-SN-TB-TO-TUR) Records in Mediterranean Region: Antalya prov.: Manavgat (Karavca place) / Korkuteli (Yazır forests) / Serik (Belek forest) (Tosun, 1975); Antalya prov.: Manavgat / Korkuteli / Serik (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Burdur prov.: Burdur lake (Özdikmen, 2007); Osmaniye prov.: Mitisin plateau (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Holarctic.
Arhopalus syriacus (Reitter, 1895) (*WM, EM)
Records in Turkey: (AD-ANT-ART-GA-IC-IZ-MG-NE-OS-SU) Records in Mediterranean Region: Antalya prov.: Bey Mountain (Demelt, 1963); Antalya prov.: Kepez forests (Tosun, 1975); Antalya prov.: Kepez (Ex. Alkan, 2000); Adana prov.: Balcalı / Kozan / Bağtepe, Antalya prov.: Central (Tozlu et al., 2002); Ġçel prov.: Mut (Alahandüzü) (Özdikmen & ġahin, 2006); Osmaniye prov.: Zorkun road (Fenk plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Mediterranean.
Subfamily SPONDYLIDINAE
Tribe SPONDYLIDINI
Spondylis Fabricius, 1775 [Type sp.: Attelabus buprestoides Linnaeus, 1758]
Spondylis buprestoides (Linnaeus, 1758)
(*EM) Records in Turkey: (AN-ART-BS-IS-KAR-KR-OS-SN-TB-TUR) Records in Mediterranean Region: Osmaniye prov.: Mitisin plateau (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palaearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 32
Subfamily DORCASOMINAE
Tribe DORCASOMINI
Apatophysis Chevrolat, 1860 [Type sp.: Apatophysis toxotoides Chevrolat, 1860
= Polyarthron barbarum Lucas, 1858]
Subgenus Apatophysis Chevrolat, 1860 [Type sp.: Apatophysis toxotoides Chevrolat, 1860
= Polyarthron barbarum Lucas, 1858]
Apatophysis kadleci Danilevsky, 2008 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Ġçel prov.: Namrun as the type loc. (Danilevsky, 2008); Turkey (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Ġçel province in S Turkey. Chorotype: Anatolian.
Subfamily CERAMBYCINAE
Tribe ACHRYSONINI
Icosium Lucas, 1854 [Type sp.: Icosium tomentosum Lucas, 1854]
Icosium tomentosum Lucas, 1854
(*WM) ssp. tomentosum Lucas, 1854 ssp. atticum Ganglbauer, 1882 Other names: chevrolati Stål. Records in Turkey: (ANT-IZ-TUR) Ġzmir prov.: Bornova (Demelt & Alkan, 1962); Ġzmir prov.: Bornova, Antalya prov.: on beach (Demelt, 1963); Antalya prov.: Central (Öymen, 1987); Antalya prov.: Manavgat, Side (Adlbauer, 1992); Antalya prov. (Tozlu et al., 2002); Turkey (Özdikmen & Demir, 2006; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Europe (Portugal, Spain, France, Italy, Malta, Croatia, Bosnia-Herzegovina, Greece ), North Africa (Algeria, Morocco, Libya, Tunusia), Caucasus (Azerbaijan), Turkey, Cyprus, Israel, Jordan. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S and SW Turkey. Chorotype: Mediterranean.
Tribe HESPEROPHANINI
Hesperophanes Dejean, 1835 [Type sp.: Callidium sericeum Fabricius, 1787]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 33
Hesperophanes sericeus (Fabricius, 1787) (*WM)
Records in Turkey: (AY-DE-ER-IP-IZ-TUR) Records in Mediterranean Region: Isparta prov. (Tozlu et al., 2002). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Turano-Europeo-Mediterranean.
Trichoferus Wollaston, 1854 [Type sp.: Trichoferus senex Wollaston, 1854
= Trichoferus fasciculatus senex Wollaston, 1854]
Trichoferus fasciculatus (Faldermann, 1837) (*WM)
Records in Turkey: (AN-ANT-BR-BS-IZ-MG-MN-TB-TUR) Records in Mediterranean Region: Mediterranean Region as Hesperophanes fasciculatus (Ġyriboz, 1940); Antalya prov.: Perge (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean.
Trichoferus griseus (Fabricius, 1792) (*WM, EM)
Records in Turkey: (AD-ANT-AY-GA-HT-IC-IZ-KN-MN-OS-TUR) Records in Mediterranean Region: Antalya prov. (Demelt, 1963); Antalya prov. – Demelt, 1963 (Ex. - Gül-Zümreoğlu, 1975); Antalya prov.: KaĢ as Hesperophanes griseus (Öymen, 1987); Antalya prov., Aegean Region, ?Mediterranean Region (Lodos, 1998); Adana prov., Antalya prov., Hatay prov., Ġçel prov., Osmaniye prov. (Tozlu et al., 2002); Ġçel prov.: Mut (Özdikmen & ġahin, 2006); Antalya prov.: Manavgat (Central, Titreyengöl, Bucak) (Özdikmen & Demir, 2006). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Mediterranean.
Trichoferus kotschyi (Ganglbauer, 1883) (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-IC-TUR) Anatolia (Winkler, 1924-1932); Antalya prov.: Bey Mts. (Demelt, 1963); Antalya prov.: Alanya (Adlbauer, 1988); Turkey (Lodos, 1998; Kadlec & Rejzek, 2001); Antalya prov. (Tozlu et al., 2002; Sama, 2002); Içel prov. (Kurzawa pers. comm.). Distribution: Europe (Greece: Samos Island), Turkey, Syria, Lebanon. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S and SW Turkey. Chorotype: E-Mediterranean.
Trichoferus preissi (Heyden, 1894) (*WM, EM)
Other names: Absent. Records in Turkey: (AD-ANT-IC-SU-AER-TUR) Antalya prov.: Perge (Demelt, 1963; Adlbauer, 1992); Turkey (Lodos, 1998; Kadlec & Rejzek, 2001; Sama & Löbl in Löbl & Smetana, 2010); Adana prov., Antalya prov., Ġçel prov. (Tozlu et al., 2002); Ġçel prov. (Sama, 2002); Antalya prov.: Manavgat (Sarılar village) (Özdikmen & Demir, 2006). Distribution: Turkey, Syria, Iran.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 34
Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey. Chorotype: SW-Asiatic (Irano-Anatolian + Syro-Anatolian).
Trichoferus sbordonii Sama, 1982 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: Alanya (Sama, 1982; Kadlec et Rejzek, 2001); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Stromatium Audinet-Serville, 1834 [Type sp.: Callidium barbatum Fabricius, 1775]
Stromatium unicolor (Olivier, 1795)
(*WM, EM) Records in Turkey: (AD-AF-AM-AN-ANT-BL-BS-CA-DE-EL-ER-EZ-GA-GI-GU-HT-IC-IS-IZ-KA-KK-MA-MG-MN-OR-OS-SM-TB-TRA-TUR) Records in Mediterranean Region: Adana prov.: KarataĢ as Stromatium fulvum (Adlbauer, 1992); Adana prov., Ġçel prov., Antalya prov. as Stromatium fulvum (Lodos, 1998); Adana prov.: Central / Ceyhan, Antalya prov.: Central / Lara / Finike / Turunçova / Kumluca / Manavgat / Serik, Hatay prov.: Central / Erzin / Ġskenderun (Denizciler), Ġçel prov.: Central / Kızılbağ / Erdemli / Dağlı / Silifke, Osmaniye prov.: Central (Tozlu et al., 2002); Ġçel prov.: AtaĢ road (Karaduvar) (Özdikmen & Çağlar, 2004); Antalya prov. (Özdikmen et al., 2005); Antalya prov.: Manavgat (Demirciler village / Sülek village) (Özdikmen & Demir, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır) (Özdikmen & Okutaner, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Subcosmopolitan.
Tribe PHORACANTHINI
Phoracantha Newman, 1840 [Type sp.: Stenocorus semipunctatus Fabricius, 1775]
Phoracantha recurva Newman, 1840
(*EM) Other names: papua Gressitt. Records in Turkey: (IC-TUR) Ġçel prov.: AtaĢ road (Karaduvar) (Özdikmen & Çağlar, 2004); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Europe (Portugal, Spain, Italy, Malta, Greece), Turkey, Middle East (Lebanon, Israel), North Africa (Morocco, Tunusia, Libya), Afrotropical Region, Nearctic Region, Neotropical Region, Australian Region. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from Ġçel province in S Turkey. Chorotype: Cosmopolitan.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 35
Phoracantha semipunctata (Fabricius, 1775) (*WM, EM)
Records in Turkey: (AD-ANT-HT-IC-KL-MG-OS-SU-TUR) Records in Mediterranean Region: Ġçel prov.: Tarsus (Karabucak) (Demelt, 1967; Erdem, 1968; Erdem & Çanakçıoğlu, 1977); Ġçel prov.: Tarsus (Karabucak), Erdemli, Silifke, Adana prov., Hatay prov., Antalya prov. (Çanakçıoğlu, 1983); Adana prov. (Adlbauer, 1992); Ġçel prov.: Tarsus (Karabucak), Erdemli, Silifke, Adana prov., Hatay prov.: Arsus, Antalya prov. (Kanat, 1998); Ġçel prov.: Tarsus, other parts of Ġçel, Hatay prov., Antalya prov. (Lodos, 1998); Adana prov., Antalya prov., Kilis prov., Osmaniye prov. (Tozlu et al., 2002). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Cosmopolitan.
Tribe CERAMBYCINI
Cerambyx Linnaeus, 1758
[Type sp.: Cerambyx cerdo Linnaeus, 1758]
Subgenus Cerambyx Linnaeus, 1758 [Type sp.: Cerambyx cerdo Linnaeus, 1758]
Cerambyx cerdo Linnaeus, 1758
(*WM, EM) Records in Turkey: (AD-ADY-AN-ANT-ART-BR-BS-CA-DE-HT-IC-IS-IZ-KA-KK-KN-KO-KS-KY-MG-NI-OS-SA-SK-SM-SN-TU-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as C. cerdo acuminatus (Fairmaire, 1884); Ġçel prov.: Bolkar Mountains (Bodemeyer, 1906); Antalya prov.: near Aspendos (Belkıs) as C. cerdo acuminatus (Demelt & Alkan, 1962); Antalya prov.: Aspendos as C. cerdo acuminatus (Demelt, 1963); KahramanmaraĢ prov. (Kanat, 1998); Adana prov.: Pozantı (Ulusoy et al., 1999); Antalya prov.: Alanya (Çayarası) (Özdikmen & Çağlar, 2004); Osmaniye prov.: Mitis‘s plateau (Özdikmen & Demir, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır) (Özdikmen & Okutaner, 2006); Osmaniye prov.: Mitisin plateau (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Cerambyx dux (Faldermann, 1837) (*WM, EM)
Records in Turkey: (AD-ADY-AN-ANT-BI-BN-BS-BU-DE-EL-ER-EZ-GA-HT-IC-IP-IS-IZ-KA-KAR-KK-KN-KS-KY-MA-MG-NI-OS-TO-TU-VA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Antalya prov.: Toros Mountains (Bodemeyer, 1900); Ġçel prov.: Bolkar Mountains (Bodemeyer, 1906); Hatay prov.: Dörtyol (Bodenheimer, 1958); KahramanmaraĢ prov., Hatay prov.: Dörtyol (Nizamlıoğlu & Gökmen, 1964); Mediterranean Region (Lodos, 1998); Adana prov.: Pozantı (Ulusoy et al., 1999); Hatay prov.: Antakya (Tauzin, 2001); Antalya prov.: Central / GazipaĢa, Burdur prov.: Central, Hatay prov.: Cırtıman / Ġskenderun (Denizciler), Isparta prov.: Eğirdir, Ġçel prov.: Anamur, Osmaniye prov.: Çiftmazı / OlukbaĢı (Tozlu et al., 2002); Adana prov. (Özdikmen et al., 2005); Gaziantep prov.: KuĢçubeli pass / Islahiye (Kabaklar village / Köklü village), Hatay prov.: Ġskenderun (entry of Kurtbağı village, Üçgüllük) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır / Sakarkaya village, Kısık / BotaĢ) / Çağlayancerit (Bozlar) / Nurhak (Nurhak-Malatya road, Tatlar) (Özdikmen & Okutaner, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır), Osmaniye prov.: Central (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Balkan).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 36
Cerambyx miles Bonelli, 1812 (*WM, EM)
Records in Turkey: (AD-ADY-AF-ANT-BT-ED-DE-DI-IC-IS-KA-KN-NI-OS-US-TRA-TUR) Ġçel prov.: Bolkar Mts. (Bodemeyer, 1906); Adana prov.: Pozantı (Ulusoy et al., 1999); KahramanmaraĢ prov.: Pazarcık (Özdikmen & Okutaner, 2006); Antalya prov.: Alanya (Sarımut-Karapınar) (Özdikmen & Turgut, 2010); Osmaniye prov.: Düziçi (between Böcekli-Hıdırlı) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: S-European.
Cerambyx nodulosus Germar, 1817
(*WM, EM) Records in Turkey: (AD-ADY-ANT-ART-BN-BS-BT-EZ-IC-IP-IS-IZ-KA-KN-MG-MN-MR-OS-US-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun (Demelt, 1967); Osmaniye prov.: Nurdağı pass (Adlbauer, 1992); Isparta prov.: Eğirdir (Tauzin, 2000); Adana prov.: Seyhan, Antalya prov.: Manavgat, Ġçel prov.: Tarsus (BağlarbaĢı) (Tozlu et al., 2002); Antalya prov.: Alanya (Çayarası plateau-Sarımut bridge), Ġçel prov.: Gözne (Özdikmen & Çağlar, 2004); KahramanmaraĢ prov.: Pazarcık (BotaĢ) (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Apenninian) + Turano-European (Turano-Sarmato-Pannonian).
Cerambyx welensii (Küster, 1846) (*WM, EM)
Records in Turkey: (ADY-ANT-IC-IS-KA-KM-KN-OS-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Bolkar Mts. as C. velutinus (Bodemeyer, 1906); Antalya prov. (Tozlu et al., 2002); Antalya prov.: Alanya (Çayarası plateau-Sarımut bridge) (Özdikmen & Çağlar, 2004); KahramanmaraĢ prov.: Pazarcık, Ġçel prov.: Ortagören-Mut (Malmusi & Saltini, 2005); Antalya prov.: Alanya (Sarımut env.), Akseki (Yarpuz env.) (Özdikmen & Turgut, 2010); Osmaniye prov.: Düziçi (between Böcekli-Hıdırlı) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: S-European.
Subgenus Microcerambyx Miksic et Georgijevic, 1973 [Type sp.: Cerambyx scopolii Fuessly, 1775]
Cerambyx scopolii Fuessly, 1775
(*WM, EM) Records in Turkey: (ANT-ART-BN-BO-ED-IC-IS-KAR-KK-NI-OS-RI-SA-SM-SN-TB-TO-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Burna, Antalya prov.: Toros Mountains (Bodemeyer, 1900); Osmaniye prov.: Nurdağı pass as C. scopolii nitidus (Pic, 1892) (Adlbauer, 1988); Osmaniye prov.: Zorkun road (Fenk plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Tribe TRACHYDERINI
Purpuricenus Dejean, 1821 [Type sp.: Cerambyx kaehleri Linnaeus, 1758]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 37
Subgenus Purpuricenus Dejean, 1821 [Type sp.: Cerambyx kaehleri Linnaeus, 1758]
Purpuricenus budensis (Götz, 1783)
(*WM, EM) Records in Turkey: (AD-ADY-AF-AM-AN-ANT-ART-AY-BL-BN-BO-BS-BU-CA-CO-DE-ED-EZ-GA-GU-HT-IC-IP-IS-IZ-KA-KI-KN-KO-MG-MN-MU-NI-OS-RI-SI-SM-SN-TO-TU-YO-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Adana prov.: Pozantı (Bodemeyer, 1900); Ġçel prov.: Bolkar Mountains (Villiers, 1959); Antalya prov.: Toros Mountains, Isparta prov.: Eğirdir (Demelt, 1963); Isparta prov.: Eğirdir (Tuatay et al., 1972); Isparta prov.: Eğirdir according to records of Institute Ġzmir/Bornova (Ex. Gül-Zümreoğlu, 1975); Gaziantep prov.: FevzipaĢa, (Sama, 1982); Antalya prov.: Yeni Karaman / Alanya (Akseki / Güzelbağ), Ġçel prov.: Anamur / Erdemli / Silifke / Kuzucubelen, Adana prov.: Kozan (Himmetli) (Adlbauer, 1988); Osmaniye prov.: Nurdağı pass as P. budensis productus Plavilstshikov, 1940 (Adlbauer, 1992); Isparta prov. (Lodos, 1998); Adana prov.: Bahçe (Tauzin, 2000); Adana prov.: Balcalı / Kozan, Antalya prov.: Central, Burdur prov.: Ağlasun / Bucak (Çamlık), Ġçel prov.: Kızılbağ, Osmaniye prov.: Çiftmazı (Tozlu et al., 2002); Isparta prov.: Sütçüler (Kasımlı, Ġncidere), Antalya prov.: Ġbradı-plateau road / Kumluca (Gölcük village) / Akseki (Geylan plateau road) / Alanya (Mahmutlar) / Kemer (Olimpos Mountain), Ġçel prov.: Kocavilayet (Emirler village) (Özdikmen & Çağlar, 2004); Isparta prov.: Eğridir, Burdur prov.: Ağlasun (Özdikmen, et al., 2005); Osmaniye prov.: Yarpuz road (KarataĢ place) / Çulhalı village / YeĢil village (Hasanbeyli) / Bahçe (Buğdaycık village) / turn of Düziçi road / Düziçi (Çamiçi village) / exit of Çotlu village, KahramanmaraĢ prov.: Sumaklı village / Türkoğlu (Doluca village), Hatay prov.: Belen (Çerçikaya village, Madenli) (Özdikmen & Demirel, 2005); Antalya prov.: Kemer / Manavgat (Bucak, ġeyhler village) (Özdikmen & Demir, 2006); Adana prov.: near Pozantı (Özdikmen, 2006); KahramanmaraĢ prov.: Central (Kavaklı) / KahramanmaraĢ-Andırın road (Körsülü bridge env.) / Pazarcık (Sakarkaya-Çağlayancerit road / Sakarkaya village, Kısık) / Çağlayancerit (Bozlar) / Andırın (KahramanmaraĢ-Andırın road, Çuhadırlı village) (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki-Manavgat road, GündoğmuĢ, Akseki-Güzelsu, Ġbradı, Exit of Ġbradı 5th km, Alanya (Karapınar village, Karapınar-Sarımut, 6 km to DikmetaĢ plateau), Akseki (Mahmutlu village env., Murtiçi-Güzelsu, Güzelsu village, Güçlüköy env., Bademli beli-Tekebeli), SeydiĢehir-Manavgat road (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Purpuricenus dalmatinus Sturm, 1843 (*WM, EM)
Records in Turkey: (ADY-ANT-AY-BN-HT-IC-IZ-KA-MG-MN-MR-MU-OS-SI-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Antalya prov.: near Central and Manavgat (Demelt & Alkan, 1962); Antalya prov.: Central and Kumköy (Demelt, 1963); Osmaniye prov.: Nurdağı pass, KahramanmaraĢ prov.: Andırın, Ġçel prov.: Erdemli (Adlbauer, 1988); Antalya prov.: Termessos (Tauzin, 2000); Antalya prov.: Alanya (KuĢkayası place), Kemer (Olimpos mountain), Osmaniye prov.: OlukbaĢı plateau, Ġçel prov.: Kocavilayet (Emirler village) (Özdikmen & Çağlar, 2004); Ġçel prov.: Erdemli-Güzeloluk road (Özdikmen, 2007); Antalya prov.: GündoğmuĢ, Akseki-Güzelsu (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: E-Mediterranean.
Purpuricenus desfontainei (Fabricius, 1792)
(*WM, EM) Records in Turkey: (AD-ANT-ART-CA-DE-HT-IZ-MN-OS-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 38
Records in Mediterranean Region: Antalya prov.: Antitoros / Toros Mountain (Demelt & Alkan, 1962 Demelt, 1963); Antalya prov.: Yeni Karaman / Central, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Adana prov.: Bahçe (Tauzin, 2000); Hatay prov.: Hassa (Zeytinoba village, Aktepe) (Özdikmen & Demirel, 2005); Antalya prov.: Kemer (Özdikmen & Demir, 2006); Antalya prov.: GündoğmuĢ, Akseki-Güzelsu, Akseki (Mahmutlu village env.) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian) + N-Africa.
Purpuricenus interscapillatus Plavilstshikov, 1937 (*WM, EM)
ssp. interscapillatus Plavilstshikov, 1937 ssp. nudicollis Demelt, 1968 ssp. sasanus Kadlec, 2006 Other names: humeralis Pic; longevittatus Pic. Records in Turkey: (AD-ANT-HT-IC-KN-NI-OS-TUR) Antalya prov.: Antitoros Mountains as P. budensis interscapillatus Plavilstshikov, 1937 (Demelt & Alkan, 1962); Konya prov. as P. nudicollis (Sama, 1982); Antalya prov.: Korkuteli, Karaman prov. as P. nudicollis (Adlbauer, 1988); Hatay prov.: Antakya, Dörtyol, Ġçel prov.: Namrun, Nurdağı pass as P. longevittatus (Sabbadini & Pesarini, 1992); Anatolia as P. longevittatus (Sama & Rapuzzi, 2000); Adana prov.: Pozantı (Tekir plateau), Niğde prov.: Niğde-Bor road (Okçu village), Ġçel prov.: Erdemli-Güzeloluk road, Mersin-Gözne road as P. longevittatus (Özdikmen, 2006); Antalya prov.: Alanya (DikmetaĢ plateau), Konya prov.: Hadim-Alanya road as Purpuricenus interscapillatus nudicollis Demelt, 1968 (Özdikmen & Turgut, 2010); Hatay prov.: Yukarı Ekinci village, Sazlık, Osmaniye prov.: Hasanbeyli (Kalecikli village), Zorkun road (Çiftmazı, Karacalar village) as Purpuricenus interscapillatus interscapillatus Plavilstshikov, 1937 (Özdikmen et al., 2010); Osmaniye prov.: Zorkun road (Karacalar village) as Purpuricenus interscapillatus nudicollis Demelt, 1968 (Özdikmen et al., 2010); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Cyprus, Middle East (Syria, Lebanon, Israel), Iran. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species is represented by two subspecies in Turkey. They have been known only from S Turkey. P. interscapillatus sasanus occurs only in Iran. Chorotype: E-Mediterranean (Palaestino-Cyprioto-Taurian) + SW-Asiatic (Irano-Anatolian).
Purpuricenus nigronotatus Pic, 1907 (*EM)
Other names: konradi Bernhauer. Records in Turkey: (IC-OS-TUR) Osmaniye prov.: Nurdağı pass as P. konradi (Adlbauer, 1988); Ġçel prov.: Cilicia (Karsantı), Toros Mountains, Osmaniye prov.: Nurdağı pass (Sabbadini & Pesarini, 1992); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey. Chorotype: Anatolian.
Purpuricenus wachanrui Levrat, 1858 (*EM)
Other names: aleppensis Witte; haussknechti Witte; bilunatus Schaufuss; schoenfeldti Heyden; atricolor Pic; quadrinototus Pic; diversipennis Pic. Records in Turkey: (AD-ADY-BN-BT-DI-MA-TU-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 39
Type loc. Turkey (Levrat, 1858); Malatya prov. (Heyden,1888); Anatolia (Winkler, 1924-1932); Bingöl prov.: Central (Fuchs & Breuning, 1971); E of Adana prov.: Misis, Bingöl prov.: Central, Tunceli prov. as P. wachanrui bilunatus Schauf.(Fuchs & Breuning, 1971); Bingöl prov. (Sama, 1982); Diyarbakır prov., Bingöl prov. (Adlbauer, 1988); Turkey (Lodos, 1998); Adıyaman prov.: Karadut village, Nemrut Mt. (Rejzek & Hoskovec, 1999); Tunceli prov.: Munzur valley (Tauzin, 2000); Adıyaman prov.: Nemrut Mt. (Rejzek et al., 2001); Bitlis prov. (Tozlu et al., 2002). Distribution: Turkey, Iran, Iraq. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from SE Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).
Tribe CALLICHROMATINI
Aromia Audinet-Serville, 1833
[Type sp.: Cerambyx moschatus Linnaeus, 1758]
Aromia moschata (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AD-ADY-AN-ANT-ART-AY-BI-BL-BN-BS-BU-CA-EZ-IC-IP-IS-IZ-KA-KN-KO-MN-OS-SM-TO-TU-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Central as A. moschata var. thoracica (Demelt & Alkan, 1962; Demelt, 1963); Adana prov. as A. moschata ambrosiaca (Villiers, 1967); Adana prov. (Gül-Zümreoğlu, 1972); Antalya prov. (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Antalya prov.: Elmalı as A. moschata ambrosiaca (Adlbauer, 1992); Antalya prov., KahramanmaraĢ prov. (Kanat, 1998); Antalya prov., Mediterranean Region (Lodos, 1998); Isparta prov. as A. moschata thoracica (Tauzin, 2000); Antalya prov.: Central / Arapsuyu / Manavgat / Korkuteli / Serik, Burdur prov.: Bucak (Çamlık), KahramanmaraĢ prov.: Central as A. moschata ambrosiaca (Tozlu et al., 2002); Antalya prov.: Elmalı (near Çalpınar) (Özdikmen & Çağlar, 2004); Ġçel prov.: Mersin as A. moschata thoracica (Malmusi & Saltini, 2005) ; Antalya prov.: Manavgat as A. moschata ambrosiaca (Özdikmen & Demir, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır) / BaĢkonuĢ forest area / Andırın (HaĢtırın village) / Nurhak (Tatlar) as A. moschata ambrosiaca (Özdikmen & Okutaner, 2006); Osmaniye prov.: Düziçi (between Böcekli-Hıdırlı) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palearctic.
Tribe GRACILIINI
Penichroa Stephens, 1839 [Type sp.: Callidium fasciatum Stephens, 1831]
Penichroa fasciata (Stephens, 1831)
(*WM, EM) Records in Turkey: (AM-AN-ANT-IC-SM-TO-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Manavgat (Sorgun forest) (Tosun, 1975); Ġçel prov.: Erdemli, Antalya prov.: Alanya (Adlbauer, 1988); Antalya prov.: Central / Finike (Turunçova) (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Axinopalpis Dejean, 1835 [Type sp.: Obrium gracile Krynicki, 1832]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 40
Axinopalpis gracilis (Krynicki, 1832) (*WM, EM)
ssp. gracilis Krynicki, 1832 ssp. christinae Rapuzzi, 1995
Other names: latior Pic. Records in Turkey: (IC-IP-TUR) Anatolia (Winkler, 1924-1932; Sama, 2002); Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Isparta prov.: ġarkikaraağaç (Sama, 1982); Ġçel prov.: Erdemli (Adlbauer, 1988). Distribution: Europe (Italy, Slovenia, Croatia, Bosnia and Herzegovina, Serbia, Macedonia, Albania, Greece, Bulgaria, Romania, Hungary, Austria, Czechia, Slowakia, Poland, Lithuania, Ukraine, Moldovia, European Russia), Caucasus (Georgia), Iran, Turkey, Syria, Israel. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species is represented only by nominotypical subspecies in Turkey. It has been known only from S Turkey. A. gracilis christinae occurs only in Greece. Chorotype: Turano-Mediterranean (Turano-apenninian).
Tribe OBRIINI
Obrium Dejean, 1821 [Type sp.: Cerambyx cantharinus Linnaeus, 1767]
Obrium cantharinum (Linnaeus, 1767)
(*WM) Records in Turkey: (ANT-BO-IP-TUR) Records in Mediterranean Region: Antalya prov.: Elmalı (Avlanbeli), Isparta prov.: ġarkikaraağaç (Kızıldağ forest) (Tosun, 1975). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Anatolobrium Adlbauer, 2004 [Type sp.: Anatolobrium eggeri Adlbauer, 2004]
Anatolobrium eggeri Adlbauer, 2004
(*WM) Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: Alanya as the type loc. (Adlbauer, 2004); Antalya prov.: Between Alanya-Çayarası plateau (Sarımut bridge env.) (Özdikmen & Okutaner, 2006). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Tribe CERTALLINI
Certallum Dejean, 1821 [Type sp.: Saperda ruficollis Fabricius, 1787
= Cerambyx ebulinus Linnaeus, 1767]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 41
Certallum ebulinum (Linnaeus, 1767) (*WM, EM)
Records in Turkey: (AD-ADY-AK-AM-AN-ANT-AY-BI-BL-BS-CA-CN-DE-DI-ER-GA-HT-IC-IP-IS-IZ-KA-KN-KY-MG-MN-MR-NE-NI-OS-SN-SU-TB-?YO-TRA-TUR) Records in Mediterranean Region: Adana prov.: Toros Mountains (Pozantı, Bolkar Mountains) (Villiers, 1959); Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains, Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near (Demelt & Alkan, 1962); Osmaniye prov.: Central / Toprakkale, Hatay prov.: Arsuz / YeniĢehir / Antakya (Reyhanlı), Osmaniye prov.: Bahçe, Adana prov.: Ceyhan (Yumurtalık, Misis) as Certallum ebulinum ruficolle (Fuchs et Breuning, 1971); Adana prov.: Pozantı, Ġçel prov.: Bolkar Mountains (Namrun, Çamlıyayla) / Tarsus (Tuatay et al., 1972); Ġçel prov.: Bolkar Mountains, Adana prov., Isparta prov. (Ex. - Gül-Zümreoğlu, 1975); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Adana prov., Hatay prov.: Antakya, Antalya prov., Isparta prov. (Lodos, 1998); Adana prov.: Balcalı, Antalya prov.: Side, Hatay prov.: Dörtyol (Ġcadiye) / Kuzuculu / Erzin / Ġskenderun (Sarımazı), Ġçel prov.: Tarsus (Çamlıyayla) (Tozlu et al., 2002); Ġçel prov.: Bulgar Mountain (Namrun), Adana prov.: Taurus (Pozantı), Ġçel prov.: Silifke, Osmaniye prov. (Özdikmen, et al., 2005); Ġçel prov.: Mut-Karaman road (Karabağ place, Hatıra Forest) (Özdikmen, 2006); Hatay prov.: Yayladağı / Kırıkhan / from Harbiye to Yayladağı, KahramanmaraĢ prov.: Pazarcık, Ġçel prov.: Tarsus-Çamlıyayla / Çamlıyayla / Tarsus / from Tarsus to Çamlıyayla (MeĢelik), Isparta prov.: Senirkent, (Malmusi & Saltini, 2005); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env. / Karbasan village env.) / Türkoğlu (ġekeroba env.) / Pazarcık (Osmandede village / Evri-Karahöyük road) (Özdikmen & Okutaner, 2006); Antalya prov.: GündoğmuĢ-Akseki road (Özdikmen & Turgut, 2010); Gaziantep prov.: Nurdağı (Exit of Ġslahiye), Hatay prov.: Ġskenderun-Arsuz, Erzin (Gökdere village), Belen, Akbez (GüzeluĢağı village), Arsuz (Akçalı), Erzin (Erzin Ġçmeler district), Osmaniye prov.: Kesmeburun village (Castabala (Hierapolis)), Karagedik village, between Kumarlı-Kazmaca villages, FakıuĢağı village, FakıuĢağı village, Sarpınağzı village, Çardak village, Bahçe, Osmaniye-Gaziantep road 5th km., Akyar village, Zorkun road (Çiftmazı), KuĢcubeli pass, Entry of Yarpuz, Castabala (Hierapolis) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Tribe DEILINI
Delagrangeus Pic, 1892 [Type sp.: Delagrangeus angustissimus Pic, 1892]
Subgenus Delagrangeus Pic, 1892
[Type sp.: Delagrangeus angustissimus Pic, 1892]
Delagrangeus angustissimus Pic, 1892 (*EM)
ssp. angustissimus Pic, 1892 ssp. troodi Sama, 1994 ssp. liviae Pesarini & Sabbadini, 2004 Other names: carbonarius Reitter; asiaticus Pic. Records in Turkey: (IC-TUR) Ġçel prov.: Namrun (Demelt, 1963; Holzschuh, 1975b); Ġçel prov.: Silifke (Olba, Gülnar) (Adlbauer, 1988); Turkey (Sama & Rapuzzi, 1999, 2000). Distribution: Europe (Greece: Rodos), Turkey, Cyprus, Lebanon. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. It is represented by the nominotypical subspecies in Turkey. The other known subspecies, D. angustissimus trooidi occurs in Cyprus and Lebanon and D. angustissimus liviae occurs only in Greece (Rodos). Chorotype: E-Mediterranean.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 42
Deilus Audinet-Serville, 1834 [Type sp.: Callidium fugax Olivier, 1790]
Deilus fugax (Olivier, 1790)
(*WM, EM) Records in Turkey: (ANT-BO-DE-HT-IZ-MN-TUR) Records in Mediterranean Region: Antalya prov.: Termessos (Adlbauer, 1992); Hatay prov.: Kırıkhan (Malmusi & Saltini, 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Tribe STENOPTERINI
Stenopterus Illiger, 1804 [Type sp.: Necydalis rufa Linnaeus, 1767]
Stenopterus atricornis Pic, 1891
(*WM) Other names: Absent. Records in Turkey: (ANT-KN-TUR) Antalya prov.: Termessos, Ġçel prov.: Erdemli (Adlbauer, 1988); Turkey (Sama & Löbl in Löbl & Smetana); Antalya prov.: Alanya (Karapınar village), Akseki (Çukurköy-Mahmutlu, Güçlüköy env.), Konya prov.: Hadim-Bozkır road (Yazdamı village env.) (Özdikmen & Turgut, 2010). Distribution: Europe (Greece), Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S and CS Turkey. Chorotype: E-Mediterranean.
Stenopterus flavicornis Küster, 1846 (*WM, EM)
Records in Turkey: (ANT-HA-HT-IZ-KN-TRA-TUR) Hatay prov.: Akbez (Fairmaire, 1884); Antalya prov.: Bey Mts., Alanya – Demelt, 1963 (Ex. Alkan, 2000). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: E-Mediterranean.
Stenopterus kraatzi Pic, 1892 (*WM, EM)
Records in Turkey: (AD-ADY-ANT-BL-HT-IC-IZ-KS-TUR) Records in Mediterranean Region: Antalya prov.: Bey Mountain / Alanya as Stenopterus flavicornis morpha kraatzi Pic, 1892 (Demelt, 1963); Ġçel prov.: Tarsus, Adana prov. (Tauzin, 2000); Hatay prov.: Yayladağı (Malmusi & Saltini, 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Anatolian.
Stenopterus rufus (Linnaeus, 1767) (*WM, EM)
Records in Turkey: (ADY-AM-AN-ANT-ART-BI-BO-BR-BS-CA-CN-CO-EZ-GA-GU-HT-IC-IS-IZ-KA-KK-KN-KO-KR-KS-KY-MN-NI-OS-RI-SM-SN-TB-TO-TU-YA-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Antitoros (Demelt & Alkan, 1962); Ġçel prov.: Namrun (Demelt, 1963); Ġçel prov.: Alata as Stenepterus rufus syriacus (Villiers,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 43
1967); Antalya prov.: TaĢağıl / Alanya (Central / Güzelbağ), Ġçel prov.: Anamur / Silifke (Gülnar) / Kuzucubelen / Erdemli, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Antalya prov. as Stenopterus rufus syriacus (Tauzin, 2000); Antalya prov.: Alanya (between Çayarası and Cırlasun / Mahmutlar) / Kalkan / Kemer (Olimpos Mt.) (Özdikmen & Çağlar, 2004); Osmaniye prov.: Zorkun plateau road (Ürün plateau) / Yarpuz road (KarataĢ place) / Düziçi (Karasi village), KahramanmaraĢ prov.: Türkoğlu (Doluca village / Kaledibi village), Hatay prov.: Hassa (Akbez, GeneluĢağı village) / Antakya (Seldiren village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Andırın (Andırın-Geben road) (Özdikmen & Okutaner, 2006); Antalya prov.: GündoğmuĢ (Ümütlü village), Ġbradı, Alanya (Karapınar village, DikmetaĢ plateau, Karapınar-Sarımut), Akseki (Çukurköy-Mahmutlu, Güçlüköy env.) as S. rufus syriacus (Özdikmen & Turgut, 2010); Osmaniye prov.: Zorkun road (Çiftmazı, Karacalar village, Ürün plateau), Yarpuz road forest store env., Yarpuz road (Yukarı Haraz plateau), Hınzırlı plateau (Kalaycıbatıran), Cebel road (Çürükarmut plateau), Hasanbeyli as S. rufus syriacus (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Tribe HYBODERINI
Callimus Mulsant, 1846 [Type sp.: Callimus bourdini Mulsant, 1846
= Saperda angulata Schrank, 1789]
Callimus akbesianus Pic, 1892 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Ġçel prov.: Erdemli (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from in S Turkey. Chorotype: Anatolian.
Callimus angulatus (Schrank, 1789) (*WM, EM)
Records in Turkey: (ADY-ANT-BO-IC-IS-KK-OR-SM-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Termessos, Ġçel prov.: Erdemli (Adlbauer, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Lampropterus Mulsant, 1862 [Type sp.: Necydalis femoratus Germar, 1824]
Subgenus Lampropterus Mulsant, 1862 [Type sp.: Necydalis femoratus Germar, 1824]
Lampropterus femoratus (Germar, 1824)
(*WM, EM) Records in Turkey: (AD-ADY-AM-AN-ANT-ART-BL-BN-BS-BU-CA-DI-ED-EZ-GA-HA-HT-IC-IS-IZ-KA-KI-KK-KN-MA-MG-MN-MU-NI-OS-YO-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Ġçel prov.: Burna, Antalya prov.: Toros Mts. (Bodemeyer, 1900); Ġçel prov.: Toros Mts. (Bolkar Mts.) as Callimus adonis (Villiers, 1959); Antalya prov.: Central as Callimellum adonis (Demelt &

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 44
Alkan, 1962; Demelt, 1963); Adana prov. as Callimellum narcissus (Demelt, 1963); Hatay prov.: Amanos Mts. (Nurdağı pass), Ġçel prov.: Alata (Villiers, 1967); Adana prov.: Nur Mt., Ġçel prov.: Erdemli (Öymen, 1987); Antalya prov.: Kemer / Yeni Karaman / Termessos / Alanya (Güzelbağ), Burdur prov.: Sagalassos, Ġçel prov.: Anamur / Silifke (Gülnar) / Erdemli / Kuzucubelen / Tarsus (Çamlıyayla), Osmaniye prov.: Nurdağı pass, KahramanmaraĢ prov., Adana prov.: Kozan (Adlbauer, 1988); Antalya prov. as Callimellum adonis (Lodos, 1998); Antalya prov.: Arapsuyu (Tozlu et al., 2002); Antalya prov.: Kalkan / Alanya (Çayarası village), Burdur prov.: Bucak (Çamlık village) (Özdikmen & Çağlar, 2004); Adana prov. (Özdikmen, et al., 2005); Hatay prov.: Ġskenderun (Güzelyayla road / entry of Kurtbağı village / Kozaklı village) / Hassa (Akbez / Zeytinoba village), Osmaniye prov.: Zorkun plateau road (Ürün plateau) / Düziçi (Çamiçi village / Kurtlar village), KahramanmaraĢ prov.: Sumaklı village / Türkoğlu (Doluca village / Kaledibi village), Gaziantep prov.: Nurdağı (plateau of Kazdere village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Pazarcık (Büyük Nacar / Kocalar village / Sakarkaya village (Pekmez stream district / Kısık env.) / Central / Bağdınısağır) / KahramanmaraĢ-Andırın road (Karbasan village env. / Yenicekale env.) / Andırın (Çınar pass / Andırın-Geben road) (Özdikmen & Okutaner, 2006); Antalya prov.: Alanya-TaĢkent (Exit of Karapınar village), Akseki-Manavgat road (GündoğmuĢ return 5th km), SeydiĢehir-Antalya road, Akseki-Manavgat road (GündoğmuĢ return), Akseki (Murtiçi-Güzelsu), Güzelsu, Ġbradı, Exit of Ġbradı 5th km (Gevenli pass env.), Alanya (Karapınar village, DikmetaĢ plateau, Karapınar-Sarımut, Sarımut-Çayarası), Akseki (Mahmutlu village env., Çukurköy-Mahmutlu), Ġbradı-Derebucak road (Özdikmen & Turgut, 2010); Hatay prov.: Dörtyol-Yeniyurt, Sazlık, Erzin-kaplıcalar, Hassa-Söğüt road (Exit of Hassa), Akbez (Gülpınarı plateau), Osmaniye prov.: Akyar village, Yarpuz road forest store env., Bahçe-Ġnderesi road, Kalecik-Hasanbeyli road, Bahçe, Haraz plateau, Karaçay, Entry of Yarpuz, Hasanbeyli (Kalecikli village), Yarpuz road, 8th km, Yarpuz road (Yukarı Haraz plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-E-Mediterranean).
Procallimus Pic, 1907 [Type sp.: Callimus egregius Mulsant & Rey, 1863]
Procallimus egregius (Mulsant & Rey, 1863)
(*WM, EM) Other names: obscurithorax Pic. Records in Turkey: (ANT-HT-IC-TUR) Anatolia (Winkler, 1924-1932); Antalya prov., Ġçel prov.: Namrun (Demelt, 1963); Hatay prov.: Antakya (Ġskenderun) (Villiers, 1967); Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: Tarsus (Çamlıyayla) (Adlbauer, 1988). Distribution: Europe (Ukraine, European Russia), Caucasus (Georgia), Turkey, Syria, Lebanon. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: E-Mediterranean.
Tribe PSEBİİNİ
Nathrius Brethes, 1916 [Type sp.: Nathrius porteri Brethes, 1916 = Leptidea brevipennis Mulsant, 1839]
Nathrius brevipennis (Mulsant, 1839)
(*WM, EM) Records in Turkey: (ANT-HT-IC-IS-IZ-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 45
Records in Mediterranean Region: Ġçel prov.: Erdemli, Antalya prov.: Alanya (Adlbauer, 1988); Hatay prov.: Yayladağı, Harbiye-Yayladağı, Ġçel prov.: Kızkalesi, Tarsus-Çamlıyayla (Malmusi & Saltini, 2005). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Subcosmopolitan.
Tribe MOLORCHINI
Molorchus Fabricius, 1792 [Type sp.: Necydalis minor Linnaeus, 1758]
Molorchus juglandis Sama, 1982
(*WM, EM) Other names: Absent. Records in Turkey: (ANT-IC-NI-TUR) Antalya prov.: Alanya as the type loc., Ġçel prov.: Bolkar Mts., Namrun (Sama, 1982); Niğde prov.: Çiftehan (Adlbauer, 1988); S Turkey: Taurus Mts. (Sama & Rapuzzi, 2000); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Caucasus (Georgia), Turkey, Israel, Lebanon. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian + Syro-Anatolian).
Molorchus minor (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (ANT-BI-BO-BS-BU-GI-IC-KS-NI-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mountains (Bodemeyer, 1900); Ġçel prov.: Namrun (Demelt, 1967); Antalya prov.: Elmalı (Avlanbeli forest), Burdur prov.: Bucak (Kavacık / Kızılgöl forest) (Tosun, 1975); Antalya prov.: Akseki (Central / Yarpuz / Irmasan pass) (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Glaphyra Newman, 1840 [Type sp.: Glaphyra semiusta Newman, 1840]
Glaphyra Newman, 1840
[Type sp.: Glaphyra semiusta Newman, 1840]
Glaphyra kiesenwetteri (Mulsant & Rey, 1861) (*WM, EM)
Records in Turkey: (AM-AN-ANT-BI-IC-IZ-KN-KS-TRA-TUR) Records in Mediterranean Region: Ġçel prov: Erdemli, Namrun (Central / Çamlıyayla) as M. kiesenwetteri anatolicus Adlbauer, 1988 (Adlbauer, 1988); Antalya prov.: Akseki (Yarpuz / Cevizli) as M. kiesenwetteri anatolicus Adlbauer, 1988 (Adlbauer, 1992); Ġçel prov.: Namrun (Erdemli) as M. kiesenwetteri hircus (Abeille de Perin, 1881) (Tauzin, 2000). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-E-Mediterranean + Turano-Apenninian) + Turano-European (Turano-Sarmato-Pannonian + Ponto-Pannonian).
Glaphyra marmottani Brisout de Barneville, 1863 (*WM, EM)
ssp. marmottani Brisout de Barneville, 1863 ssp. crovatoi Sama, 1995

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 46
ssp. frischi Sama, 1995 Other names: discicollis Heyden. Records in Turkey: (OS-TUR) Osmaniye prov.: Zorkun plateau (Adlbauer, 1992); Osmaniye prov.: Zorkun plateau as the type locality of G. marmottani frischi (Sama, 1995); Turkey (Sama, 1995; Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Osmaniye prov.: Zorkun plateau as G. marmottani frischi (Tauzin, 2000); South-Eastern Turkey G. marmottani frischi (Sama, 2002). Distribution: Europe (Spain, France, Italy, Bulgaria, Germany, Austria, Czechia, Slowakia, Poland, European Russia), Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. It is represented by two subspecies in Turkey as G. marmottani marmottani and G. marmottani frischi (it occurs only in S Turkey). G. marmottani crovatoi occurs only in Italy. Chorotype: S, C and E-European.
Glaphyra tenuitarsis (Holzschuh, 1981) (*EM)
Other names: sterbai Adlbauer, 1988. Records in Turkey: (AD-IC-KA-TUR) Ġçel prov.: Namrun as the type loc. (Holzschuh, 1981a); KahramanmaraĢ prov.: Göksun as M. sterbai (Adlbauer, 1988); Ġçel prov.: Namrun, KahramanmaraĢ prov.: Göksun, Adana prov.: Pozantı (Sama, 1995); Ġçel prov.: Namrun (Tauzin, 2000); Southern Turkey: Taurus Mountains (Sama, 2002); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from in S Turkey. Chorotype: Anatolian.
Brachypteroma Heyden, 1863 [Type sp.: Brachypteroma ottomanum Heyden, 1863]
Brachypteroma holtzi Pic, 1905
(*EM)
Other names: Absent. Records in Turkey: (IC-OS-TUR) Anatolia (Winkler, 1924-1932); Ġçel prov. (Demelt, 1967); Osmaniye prov. (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria, Lebanon. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).
Brachypteroma ottomanum Heyden, 1863
(*EM)
Records in Turkey: (AY-BS-IC-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun (Demelt, 1963). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: Turano-Mediterranean (Turano-Apenninian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 47
Tribe STENHOMALINI
Stenhomalus White, 1855 [Type sp.: Stenhomalus fenestratus White, 1855]
Subgenus Obriopsis Müller, 1948
[Type sp.: Obrium bicolor Kraatz, 1862]
Stenhomalus bicolor (Kraatz, 1862) (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-IC-NE-NI-OS-TUR) Antalya prov.: Toros Mts., Niğde prov.: Çamardı (Bodemeyer, 1900); Antalya prov.: Korkuteli (Demelt & Alkan, 1962; Demelt, 1963; Ġren & Ahmed, 1973); Antalya prov.: KaĢ, Ġçel prov.: Silifke (Gülnar), Erdemli, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Turkey (Lodos, 1998; Sama, 2002; Sama & Löbl in Löbl & Smetana, 2010); NevĢehir prov.: Uçhisar (Özdikmen, 2006). Distribution: Europe (Italy, Croatia, Bosnia & Herzegovina, Slovenia, Serbia, Macedonia, Bulgaria, Greece, Hungary, Austria, Czechia, Sweden, Slowakia), Turkey, Cyprus, Syria, Israel. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S and C Turkey. Chorotype: SE-European.
Tribe CALLIDIINI
Hylotrupes Serville, 1834
[Type sp.: Cerambyx bajulus Linnaeus, 1758]
Hylotrupes bajulus (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AD-AM-AN-ANT-ART-AY-BI-BO-BR-BS-CA-DE-DU-ER-EZ-GI-GU-HT-IC-IP-IS-IZ-KA-KAR-KN-KR-KS-KU-KY-OS-RI-SN-SV-TB-US-ZO-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Antalya prov.: Antitoros Mountains (Demelt & Alkan, 1962); Antalya prov.: Toros Mountains / on beach, Ġçel prov.: Silifke (Demelt, 1963); Ġçel prov.: Silifke (Tuatay et al., 1972); Antalya prov.: Central / Sarısu place, Isparta prov. (Tosun, 1975); Antalya prov. (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Antalya prov.: Alanya (Adlbauer, 1988); KahramanmaraĢ prov., Mediterranean Region (Kanat, 1998); Adana prov.: Kozan (Bağtepe) / Yumurtalık, Antalya prov.: Kumluca / Serik, Hatay prov.: Central, Isparta prov.: Central / Senirkent / Sütçüler, Ġçel prov.: Erdemli / Tarsus (Tozlu et al., 2002); Ġçel prov.: Silifke / Fındıkpınarı, Adana prov., Isparta prov. (Özdikmen, et al., 2005); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır) (Özdikmen & Okutaner, 2006); Osmaniye prov.: Zorkun road (Fenk plateau) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Subcosmopolitan.
Ropalopus Mulsant, 1839 [Type sp.: Callidium clavipes Fabricius, 1775]
Subgenus Ropalopus Mulsant, 1839
[Type sp.: Callidium clavipes Fabricius, 1775]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 48
Ropalopus clavipes (Fabricius, 1775) (*EM)
Records in Turkey: (AD-ADY-AN-BL-BO-CN-CO-DE-ED-ER-HT-IC-IS-IZ-KO-KU-MN-MU-NI-OS-US-TRA-TUR) Records in Mediterranean Region: Osmaniye prov.: Nurdağı pass, Adana prov.: Kozan (Himmetli) (Adlbauer, 1988); Hatay prov.: Antakya (Lodos, 1998); Adana prov.: Pozantı (Ulusoy et al., 1999); Ġçel prov.: from Tarsus to Çamlıyayla (Malmusi & Saltini, 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European or Sibero-European.
Leioderes Redtenbacher, 1849 [Type sp.: Leioderes kollari Redtenbacher, 1849]
Leioderes tuerki Ganglbauer, 1885
(*EM) Records in Turkey: (AD-AM-HT-IC-IZ-MN-TUR) Records in Mediterranean Region: Ġçel prov.: Burna (?=Çamlıyayla) as Leioderes türki (Bodemeyer, 1900); Taurus as Leioderes türki (Winkler, 1924-1932); Ġçel prov.: Namrun as Leioderes türki (Demelt, 1963); Ġçel prov.: Erdemli as Leioderes türki (Adlbauer, 1988); Hatay prov.: Akbez as the type loc. of Leioderes tuerki v. nigripes; Ġçel prov.: Bolkar Mts. (Cilicischen Taurus) as Leioderes bodemeyeri (Sama, 1992); Adana prov.: Ceyhan (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: E-Mediterranean (Palaestino-Taurian) or SW-Asiatic (Syro-Anatolian).
Semanotus Mulsant, 1839 [Type sp.: Cerambyx undatus Linnaeus, 1758]
Semanotus russicus (Fabricius, 1777)
(*WM) Records in Turkey: (AD-AM-HT-IC-IZ-MN-TUR) Records in Mediterranean Region: Antalya prov.: Elmalı (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European (Turano-Sarmato-Pannonian) + Turano-Mediterranean (Turano-Apenninian + Turano-E-Mediterranean).
Callidium Fabricius, 1775 [Type sp.: Cerambyx violaceus Linnaeus, 1758]
Subgenus Callidium Fabricius, 1775
[Type sp.: Cerambyx violaceus Linnaeus, 1758]
Callidium syriacum (Pic, 1892) (*WM, EM)
Other names: orientale Pic. Records in Turkey: (ANT-HT-TUR) Hatay prov.: Akbez as the type loc. – Pic, 1892 (Ex. Sama, 1992); Antalya prov.: Toros Mts. (Irmasan pass) (Sama, 1992); Hatay prov.: Akbez (Sama & Rapuzzi, 2000); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 49
Phymatodes Mulsant, 1839 [Type sp.: Cerambyx variabilis Linnaeus, 1760
= Cerambyx testaceus Linnaeus, 1758]
Subgenus Phymatodes Mulsant, 1839 [Type sp.: Cerambyx variabilis Linnaeus, 1760
= Cerambyx testaceus Linnaeus, 1758]
Phymatodes testaceus (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (ADY-ANT-ART-BO-CA-GU-HT-IC-IS-NI-OS-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Bolkar Mts. as P. testaceus var. analis (Bodemeyer, 1906); Hatay prov.: Dörtyol (Öymen, 1987); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Antalya prov.: Central (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Holarctic.
Subgenus Phymatodellus Reitter, 1913 [Type sp.: Callidium rufipes Fabricius, 1777]
Phymatodes magnanii (Sama & Rapuzzi, 1999)
(*WM, EM) Other names: Absent. Records in Turkey: (ANT-IC-TUR) Antalya prov.: Akseki (Çakıllı pass), Ġçel prov.: N Erdemli as the type loc. (as Poecilium (Phymatodellus) magnanii) (Sama & Rapuzzi, 1999); Turkey as Phymatodes (Phymatodellus) magnanii (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from in S Turkey. Chorotype: Anatolian.
Phymatodes rufipes (Fabricius, 1777) (*WM, EM)
Records in Turkey: (ANT-BO-HT-IC-NI-OS-SA-TUR) Records in Mediterranean Region: Hatay prov.: Akbez as type loc. of P. rufipes syriacum; Antalya prov.: Toros Mountains (Bodemeyer, 1900); Osmaniye prov.: Nurdağı pass as P. rufipes syriacum (Adlbauer, 1988); Ġçel prov.: Çamlıyayla, Hatay prov.: Yayladağı as P. rufipes syriacum (Sama & Rapuzzi, 1999); South-Eastern Turkey as P. rufipes syriacum (Sama, 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Poecilium Fairmaire, 1864 [Type sp.: Leptura alni Linnaeus, 1767]
Poecilium alni (Linnaeus, 1767)
(*EM) Records in Turkey: (BO-IS-OS-SA-TRA-TUR) Records in Mediterranean Region: Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Osmaniye prov.: KuĢçubeli pass (Gaziantep border) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: W-Palaearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 50
Poecilium lividum (Rossi, 1794) (*EM)
Records in Turkey: (IC-IS-SA-TUR) Records in Mediterranean Region: Ġçel prov.: Erdemli (Adlbauer, 1988). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Subcosmopolitan. Mediterranean for Palearctic Region.
Tribe CLYTINI
Plagionotus Mulsant, 1842 [Type sp.: Leptura detrita Linnaeus, 1758]
Subgenus Plagionotus Mulsant, 1842 [Type sp.: Leptura detrita Linnaeus, 1758]
Plagionotus arcuatus (Linnaeus, 1758)
(*WM, EM) Records in Turkey: (ART-BI-CA-DU-IP-IS-KS-MU-OS-SM-TO-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir (Tuatay et al., 1972); Osmaniye prov.: Central (Tozlu et al., 2002); Isparta prov.: Eğirdir (Özdikmen et al., 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: W-Palaearctic.
Plagionotus detritus (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AD-ANT-EZ-HT-IS-KA-MN-SN-TRA-TUR) Records in Mediterranean Region: Adana prov., Hatay prov., Antalya prov. (Lodos, 1998); KahramanmaraĢ prov.: Central (Tozlu et al., 2002). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Subgenus Neoplagionotus Kasatkin, 2005 [Type sp.: Clytus bobelayei Brullé, 1832]
Plagionotus bobelayei (Brullé, 1832)
(*WM, EM) Records in Turkey: (ADY-AG-ANT-ART-BN-EZ-GU-HT-IC-IP-IZ-KAR-KI-KN-MA-MU-SM-TU-YO-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir as Plagionotus speciosus (Demelt & Alkan, 1962); Isparta prov.: Eğirdir (Demelt, 1963); Ġçel prov.: Erdemli-Güzeloluk, Hatay prov.: Yayladağı (Malmusi & Saltini, 2005); Antalya prov.: Alanya (Sarımut-Çayarası) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European (Turano-Sarmato-Pannonian).
Subgenus Echinocerus Mulsant, 1863 [Type sp.: Cerambyx floralis Pallas, 1773]
Plagionus floralis (Pallas, 1773)
(*WM, EM) Records in Turkey: (AD-ADY-AF-AG-AM-AN-ANT-AR-ART-BI-BO-BS-BU-BY-CA-CN-CO-DE-EL-ER-ES-EZ-GI-GU-IC-IG-IP-IZ-KA-KAR-KIR-KK-KM-KN-KO-KR-KS-KY-MA-MN-MU-NI-OS-SM-SN-SV-TB-TO-TU-US-YO-ZO-TRA-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 51
Records in Mediterranean Region: Antalya prov.: Toros Mountains (Bodemeyer, 1900); Isparta prov.: Eğirdir, Ġçel prov.: Namrun (Demelt, 1963); Isparta prov.: Eğirdir (Tuatay et al., 1972); Isparta prov.: Uluborlu (Gül-Zümreoğlu, 1975); Adana prov.: Kozan (Savruk), Antalya prov.: Central (Karain cave) (Tozlu et al., 2002); Isparta prov.: Keçiborlu (Özbahçe village / Yenitepe) / Eğirdir (between Eğirdir and Gelendost) / Yalvaç (Sultan Mountains), Antalya prov.: Alanya (Gökbel plateau), Burdur prov.: Gölhisar (Çameli road) (Özdikmen & Çağlar, 2004); Isparta prov.: Eğirdir (Özdikmen et al., 2005); Osmaniye prov.: entry of Yarpuz (Cebel, turn of Oruçgazi road) / Bahçe (Yaylalar village) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı (entry of Fındıklı), Ġçel prov.: between Mut-Karaman / Mut-Karaman road (Gökçeören pine grove / DeğirmenbaĢı) (Özdikmen, 2006); KahramanmaraĢ prov.: Göksun (Andırın-Göksun road / Göksun-Çardak / KamıĢcık village / Mehmetbey / Meyremçil plateau) / Ekinözü (Türkeli / Alpınar village) / Pazarcık (Sakarkaya-Çağlayancerit road / Sakarkaya village (Göynük env.) / BaĢkonuĢ forest / Andırın-Çokak road (Akifiye / Parmaksız plateau) / Andırın (Çokak-Geben road / Geben (Ardıççalısı) (Özdikmen & Okutaner, 2006); Antalya prov.: Exit of Ġbradı 5th km (Gevenli pass env.), Alanya (DikmetaĢ plateau, Sarımut-Çayarası) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Isotomus Mulsant, 1862 [Type sp.: Callidium semipunctatum Fabricius, 1798
=Callidium speciosum Schneider,1787]
Isotomus syriacus (Schneider, 1787) (*EM)
Other names: Absent. Records in Turkey: (HT-IC-TUR) Hatay prov.: Amanos Mountains (Akbez) – Pic, 1902 (Ex. Sama, 1996); Ġçel prov.: Çamlıyayla (Sama, 1996); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from in S Turkey. Chorotype: Anatolian.
Chlorophorus Chevrolat, 1863 [Type sp.: Callidium annulare Fabricius, 1787]
Chlorophorus aegyptiacus (Fabricius, 1775)
(*EM) Records in Turkey: (AM-AN-BL-BO-BS-CA-DE-HT-IS-IZ-MG-MN-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Chlorophorus dinae (Schneider, 1787) (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-HT-KA-KN-OS-TUR) Hatay prov.: Antakya (ġenköy) as the type loc. (Rapuzzi & Sama, 1999); Osmaniye prov.: Yarpuz road (KarataĢ place), entry of Yarpuz (Cebel: turn of Oruçgazi road), Bahçe (Yaylalar village), Hatay prov.: Dörtyol (Yahyalı plateau) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Andırın (Çınar pass) (Özdikmen & Okutaner, 2006); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Antalya prov.: Akseki-Güzelsu, Alanya (Karapınar village), Akseki (Murtiçi-Güzelsu) (Özdikmen & Turgut, 2010). Distribution: Turkey, Syria.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 52
Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Chlorophorus figuratus (Scopoli, 1763) (*WM, EM)
Records in Turkey: (ANT-GU-IC-IS-KN-KO-KS-SM-TB-TO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Alanya (KuĢkayası place), Ġçel prov.: Gözne (Özdikmen & Çağlar, 2004). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Chlorophorus gratiosus Marseul, 1868 (*WM, EM)
Records in Turkey: (ANT-HT-IC-IZ-NI-TUR) Records in Mediterranean Region: Ġçel prov.: Silifke as Chlorophorus gratiosus m. muchei Heyr. (Demelt, 1967); Antalya prov.: Ovacık, Manavgat (ġelale), Termessos, Alanya (Central, GazipaĢa, Güzelbağ), Dim stream, Kemer, Ġçel prov.: Anamur, Silifke (Gülnar), Erdemli, Kuzucubelen (Adlbauer, 1988); Hatay prov.: Yayladağı, Ġçel prov.: Güzeloluk, Ortagören-Mut (Malmusi & Saltini, 2005); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km), GündoğmuĢ, Akseki-Manavgat road (GündoğmuĢ return, Murtiçi-Güzelsu), Akseki-Güzelsu, Ġbradı, Exit of Ġbradı 5th km (Gevenli pass env.), Akseki (Mahmutlu village env., Güçlüköy env., Güzelsu village, Murtiçi-Güzelsu) as C. gratiosus sparsus (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. It is represented only by the subspecies C. gratiosus sparsus in Turkey. Chorotype: E-Mediterranean (Palestino-Taurian).
Chlorophorus hungaricus Seidlitz, 1891 (*EM)
Records in Turkey: (AD-AN-BO-BR-GA-IC-KA-KN-KO-KR-KS-NI-OS-SV-TUR) Records in Mediterranean Region: Adana prov.: Nurdağı pass (Sama, 1982); Osmaniye prov.: Nurdağı pass, Ġçel prov.: Erdemli / Silifke (Mut) (Adlbauer, 1988); Adana prov.: Pozantı (entry of Fındıklı), Ġçel prov.: between Gözne-Mersin / entry of Çukurbağ / Mut-Karaman road (DeğirmenbaĢı) (Özdikmen, 2006); KahramanmaraĢ prov.: Central (Tekir) / KahramanmaraĢ-Andırın road (BaĢkonuĢ forest) / Andırın-Geben road (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European (Ponto-Pannonian).
Chlorophorus nivipictus (Kraatz, 1879) (*WM, EM)
Other names: cinctiventris Chevrolat; splichali Fleischer. Records in Turkey: (AD-ANT-IC-KN-NI-OS-VA-TUR) Niğde prov.: Çamardı, Adana prov.: Pozantı (Bodemeyer, 1900); Turkey: Taurus as splichali Fleischer, 1908; Antalya prov.: Antitoros Mts. (Demelt & Alkan, 1962); Antalya prov.: Bey Mt. And Alanya, Ġçel prov.: Namrun (Demelt, 1963); Van prov. (Sama, 1982); Ġçel prov.: Tarsus (Çamlıyayla), Erdemli, Silifke, Kuzucubelen, Antalya prov.: Alanya (Güzelbağ, Akseki, Central), Osmaniye prov.: Nurdağı pass, Adana prov.: Kozan (Adlbauer, 1988); Ġçel prov.: Külek (Gülek, Cilician Taurus) as the type loc. – Kraatz, 1879 (Ex. Sama, 1994, 1996); Turkey (Lodos, 1998; Rapuzzi & Sama, 1999); Osmaniye prov. (Tozlu et al., 2002); Konya prov.: Bozkır: Çağlayan town, 1210 m, N 37 10 E 32 11, 11.VI.2008, 1 specimen (Özdikmen & Turgut, 2010); Osmaniye prov.: Zorkun road (Fenk plateau), Yarpuz road (Yukarı Haraz plateau), Hınzırlı plateau (Kalaycıbatıran) (Özdikmen et al., 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 53
Distribution: Europe (Greece: Samos), Turkey, Syria, Iran. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from in S Turkey. Chorotype: SW-Asiatic (Irano-Anatolian) or E-Mediterranean.
Chlorophorus sartor (Müller, 1766) (*WM, EM)
Records in Turkey: (AD-AM-AN-ANT-ART-AY-BI-BL-BR-BS-BU-CA-CN-DE-EL-ES-EZ-GA-GU-HT-IC-IP-IS-IZ-KA-KK-KN-KR-KS-KY-MG-MN-OS-RI-SM-SN-TE-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov., Isparta prov.: Eğirdir (Demelt & Alkan, 1962; Demelt, 1963); Ġçel prov.: Alata (Villiers, 1967); Ġçel prov.: Silifke (Tuatay et al., 1972); Isparta prov.: Atabey (Ġslamköy) (Gül-Zümreoğlu, 1975); Gaziantep prov.: FevzipaĢa (Sama, 1982); Antalya prov.: Yeni Karaman / Alanya (Güzelbağ), Ġçel prov.: Anamur, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Isparta prov., Hatay prov.: Antakya (Lodos, 1998); Antalya prov.: Arapsuyu (Tozlu et al., 2002); Isparta prov.: YeniĢarbademli-Gedikli (near BeyĢehir lake), Antalya prov.: Alanya (Gökbel plateau) (Özdikmen & Çağlar, 2004); Ġçel prov.: Silifke, Burdur prov.: Ağlasun / Ġnsuyu (Özdikmen et al., 2005); Osmaniye prov.: Zorkun plateau road (Ürün plateau / OlukbaĢı place) / Çulhalı village / Düziçi (Çamiçi village), KahramanmaraĢ prov.: Türkoğlu (Doluca village), Gaziantep prov.: Nurdağı (plateau of Kazdere village), Hatay prov.: Hassa (Akbez, GeneluĢağı village / Zeytinoba village, Aktepe) / Kırıkhan (TaĢoluk village) / Belen (Güzelyayla road) (Özdikmen & Demirel, 2005); Adana prov., Ġçel prov.: Mersin / Güzeloluk (Malmusi & Saltini, 2005); Antalya prov.: Manavgat (Demirciler village) (Özdikmen & Demir, 2006); Ġçel prov.: exit of Fındıkpınarı (Özdikmen, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır / Sakarkaya village (Kısık env.) / KahramanmaraĢ-Göksun road (entry of Tekir) / Andırın (KahramanmaraĢ-Andırın road, Çuhadırlı village) (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km), Akseki-Güzelsu, Ġbradı, Alanya (DikmetaĢ plateau, Sarımut-Karapınar), Akseki-Manavgat road (GündoğmuĢ return: 30 km to GündoğmuĢ), Hatay prov.: Sazlık, Osmaniye prov.: Yarpuz road forest store env., Zorkun road (Karacalar village), ArslantaĢ-Osmaniye road (Kazmaca village), Cebel road (Çürükarmut plateau), Yarpuz road (Yukarı Haraz plateau), Yarpuz road, 8th km, Düziçi (Böcekli village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Chlorophorus trifasciatus (Fabricius, 1781)
(*WM, EM) Records in Turkey: (ANT-BI-IC-IS-KN-KO-KR-KS-KU-OS-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun (Demelt & Alkan, 1962; Demelt, 1963); Antalya prov.: Kemer (Olimpos Mt.) (Özdikmen & Çağlar, 2004); Osmaniye prov.: KarataĢ Dam env., Zorkun road (Karacalar village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Mediterranean.
Chlorophorus varius (Müller, 1766) (*WM, EM)
Records in Turkey: (AD-ADY-AK-AM-AN-ANT-ART-AY-BI-BL-BO-BR-BU-CA-CN-DE-ER-ES-EZ-GU-HA-HT-IC-IG-IP-IS-IZ-KA-KI-KIR-KK-KM-KN-KO-KR-KS-KY-MA-MG-MN-MR-MU-NE-NI-OS-SU-TB-TO-US-ZO-VA-TRA-TUR) Records in Mediterranean Region: Adana prov.: Pozantı as ornatus Herbst (Bodemeyer, 1900); Antalya prov.: Alanya as C. varius damascenus Chevrolat, 1854 (Demelt & Alkan, 1962; Demelt, 1963); Antalya prov.: Alanya (Ex. Gül-Zümreoğlu, 1975); Isparta prov.: Atabey (Ġslamköy) (Gül-Zümreoğlu, 1975); Antalya prov.: Alanya (Adlbauer, 1988); Adana prov., Antalya prov. (Lodos, 1998); Adana prov.: Balcalı, Antalya prov.: Kumluca / Manavgat / Serik, Burdur prov.: Bucak (Çamlık), Hatay prov.: Ġskenderun

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 54
(Denizciler), Isparta prov.: Senirkent, Ġçel prov.: Tarsus, Osmaniye prov.: Central / Kadirli (Kabayar) (Tozlu et al., 2002); Antalya prov.: KaĢ (Gömbe, Sinekçi village, Sinekçibeli) (Özdikmen & Çağlar, 2004); Osmaniye prov.: turn of Düziçi road, Gaziantep prov.: Nurdağı (plateau of Kazdere village) / Islahiye (Esenli village), Hatay prov.: Belen (Güzelyayla road) (Özdikmen & Demirel, 2005); Antalya prov.: Manavgat (Central / Demirciler village) (Özdikmen & Demir, 2006); Adana prov.: Pozantı-Ġçel road, Ġçel prov.: exit of Atakent / Mut-Silifke road / Erdemli-Güzeloluk road / Mersin-Gözne road (Çukurkeklik) / Silifke-Mut road (Göksu bridge) (Özdikmen, 2006); KahramanmaraĢ prov.: Pazarcık (Bağdınısağır / Aksu bridge / Sakarkaya village (Kısık)) / Çağlayancerit (Bozlar) / KahramanmaraĢ-Andırın road (Körsülü bridge env.) / AfĢin (Çardak-AfĢin road) / Nurhak (Nurhak-Malatya road, Tatlar) / Andırın (Andırın-Çokak road, Çınar place / Parmaksız plateau) / Çağlayancerit (Ç.cerit-Düzbay road / Ç.cerit-Bozlar road) as C. varius damascenus (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return: 30 km to GündoğmuĢ) as C. varius damascenus (Özdikmen & Turgut, 2010); Hatay prov.: Yukarı Ekinci village, KahramanmaraĢ prov.: Pazarcık, Bağdınısağır, Osmaniye prov.: Zorkun road (Ürün plateau, Karacalar village), ArslantaĢ-Osmaniye road (Kazmaca village), Bahçe road (Çona village), Düziçi (Böcekli village), Toprakkale (Antakya road 1st km) as C. varius damascenus (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palearctic.
Xylotrechus Chevrolat, 1860 [Type sp.: Clytus sartorii Chevrolat, 1860]
Subgenus Xylotrechus Chevrolat, 1860
[Type sp.: Clytus sartorii Chevrolat, 1860]
Xylotrechus arvicola (Olivier, 1795) (*EM)
Records in Turkey: (AD-GI-HT-KK-TB-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Ġskenderun (Amanos Mountains) (Öymen, 1987); Adana prov.: Kozan (Feke) (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: W-Palaearctic.
Pseudosphegesthes Reitter, 1913 [Type sp.: Clytus cinereus Laporte De Castelnau & Gory, 1836]
Pseudosphegesthes longitarsus Holzschuh, 1974
(*WM) Other names: Absent. Records in Turkey: (ADY-ANT-IP-MU-TUR) MuĢ prov.: Buğlan pass as the type loc. (Holzschuh, 1974); Antalya prov.: Alanya (GündoğmuĢ), Isparta prov.: Eğridir (Kovada) (Adlbauer, 1992); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Antalya prov.: GündoğmuĢ, Termessos (Danilevsky, 1999); Adıyaman prov.: Karadut village (Rejzek & Hoskovec, 1999). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species probably occurs in whole S Turkey. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 55
Rhaphuma Pascoe, 1858 [Type sp.: Rhaphuma placida Pascoe, 1858]
Rhaphuma gracilipes (Faldermann, 1835)
(*EM) Other names: angusticollis Mulsant; tenuicornis Fairmaire; sachalinensis Matsumura; rosinae Pic. Records in Turkey: (HT) Hatay prov.: Akbez (Fairmaire, 1884). Distribution: Belarus, European Russia, Poland, Siberia, Far East Russia, China, Kazakhstan, Mongolia, Korea, Japan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species is distributed in Northern Plaearctic region. So it is not confirmed and impossible for Turkey. Chorotype: Sibero-European.
Clytus Laicharting, 1784 [Type sp.: Leptura arietis Linnaeus, 1758]
Clytus ciliciensis (Chevrolat, 1863) (*EM)
Other names: bifarius Heyden; reitteri Théry; griseofasciatus Pic; delagrangei Pic. Records in Turkey: (HT-IC-KA-OS-TUR) Anatolia (Winkler, 1924-1932); Hatay prov.: Ġskenderun (Demelt, 1963); Ġçel prov.: Gözne (Villiers, 1967); Ġçel prov.: Erdemli, Kuzucubelen (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: Erdemli (Limonlu) (Özdikmen et al., 2005); Osmaniye prov.: Düziçi (Gökçayır village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Ekinözü (Türkeli), Pazarcık (Aksu bridge), Central (Türkoğluaraplar village, Kavaklı), KahramanmaraĢ-Andırın road (Karbasan village env.) (Özdikmen & Okutaner, 2006); Ġçel prov.: Erdemli-Güzeloluk road (Özdikmen, 2006); Hatay prov.: Sazlık, Erzin-kaplıcalar district, Osmaniye prov.: Kalecik-Hasanbeyli road, Hasanbeyli (Kalecikli village), Karaçay, Zorkun road (Çiftmazı) (Özdikmen et al., 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey (Amanos Mts. and near). Chorotype: SW-Asiatic (Syro-Anatolian).
Clytus gulekanus Pic, 1904 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Anatolia (Winkler, 1924-1932); Ġçel prov.: Tarsus, Çamlıyayla (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Ġçel province in S Turkey. Chorotype: Anatolian.
Clytus madoni Pic, 1891 (*EM)
Other names: preapicalis Pic. Records in Turkey: (HT-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 56
Hatay prov.: Antakya (Adlbauer, 1992); Turkey (Lodos, 1998; Sama & Rapuzzi, 2000; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria, Israel, Lebanon. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey (Amanos Mts.). Chorotype: E-Mediterranean (Palaestino-Taurian).
Clytus rhamni Germar, 1817 (*WM, EM)
Records in Turkey: (AD-ADY-AM-AN-ANT-ART-BI-BS-BY-CA-CN-GA-GU-HT-IC-IP-IS-IZ-KA-KK-KN-KO-KR-KS-KY-MA-OS-RI-SM-SN-SV-TO-YA-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains / Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near (Demelt & Alkan, 1962); Ġçel prov.: Silifke (Tuatay et al., 1972); Gaziantep prov.: FevzipaĢa as C. rhamni temesiensis (Sama, 1982); Hatay prov.: Amanos Mountains (near Dörtyol) (Öymen, 1987); Antalya prov.: Central / Kemer / Alanya (Güzelbağ) / Manavgat / Patara / Termessos / Yeni Karaman, Ġçel prov.: Anamur / Silifke (Central / Gülnar) / Kuzucubelen / Tarsus (Çamlıyayla) / Kanlıdivane, Osmaniye prov.: Nurdağı pass, KahramanmaraĢ prov.: Andırın, Adana prov.: Kozan as C. rhamni temesiensis (Adlbauer, 1988); Antalya prov.: Arapsuyu, Hatay prov.: Erzin (Tozlu et al., 2002); Antalya prov.: Alanya (Mahmutlar) / Kemer (Olimpos Mt.) (Özdikmen & Çağlar, 2004); Ġçel prov.: Silifke (Özdikmen et al., 2005); Osmaniye prov.: Zorkun plateau / Zorkun plateau road (Ürün plateau / OlukbaĢı place) / Yarpuz road (KarataĢ place) (Özdikmen & Demirel, 2005); Adana prov., Ġçel prov.: Erdemli- Güzeloluk / Güzeloluk / from Tarsus to Çamlıyayla / from Ortagören to Mut (Malmusi & Saltini, 2005); Ġçel prov.: Uzuncaburç / Mersin-Gözne road (entry of Yeniköy) (Özdikmen, 2006); Antalya prov.: Akseki (Murtiçi-Güzelsu), Ġbradı (Özdikmen & Turgut, 2010); Osmaniye prov.: Zorkun road (Çiftmazı Gölyeri), Cebel road (Çürükarmut plateau), Yarpuz road (Yukarı Haraz plateau, 8th km, Karacalar village, Ürün plateau), Yarpuz road forest store env. (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Clytus taurusiensis (Pic, 1903) (*EM)
Other names: bytinskii Heyrovsky. Records in Turkey: (HT-IC-OS-TUR) Osmaniye prov.: Bahçe (Adlbauer, 1992); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Hatay prov.: Antakya (Teknepınar) (Özdikmen & Demirel, 2005); Ġçel prov.: Uzuncaburç road (Özdikmen,2006). Distribution: Turkey, Israel. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey (Amanos Mts.). Chorotype: E-Mediterranean (Palaestino-Taurian).
Subfamily LAMIINAE
Tribe PARMENINI
Parmena Dejean, 1821 [Type sp.: Lamia unifasciata Rossi, 1790]
Parmena lukati Sama, 1994
(*WM) Other names: Absent.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 57
Records in Turkey: (ANT-TUR) Antalya prov.: Kekova as the type loc. (Sama, 1994a; Tauzin, 2000); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Parmena mutilloides Sabbadini & Pesarini, 1992 (*WM)
Other names: Absent. Records in Turkey: (ANT-IP-KN-TUR) Konya prov.: BeyĢehir, Antalya prov.: Perge (Sama, 1994a); Konya prov.: BeyĢehir (Tauzin, 2000); Isparta prov.: Yalvaç (Bağkonak, Sultan Mts.) (Özdikmen & Hasbenli, 2004); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S and CSW Turkey. Chorotype: Anatolian.
Tribe MESOSINI
Mesosa Latreille, 1829 [Type sp.: Cerambyx curculionoides Linnaeus, 1760]
Subgenus Mesosa Latreille, 1829
[Type sp.: Cerambyx curculionoides Linnaeus, 1760]
Mesosa curculionoides (Linnaeus, 1760) (*WM)
Records in Turkey: (ANT-KK-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Akseki (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Tribe BATOCERINI
Batocera Dejean, 1835 [Type sp.: Cerambyx rubus Linnaeus, 1758]
Batocera rufomaculata (DeGeer, 1775)
(*WM, EM) Other names: cruentata Gmelin; diana Nonfried. Records in Turkey: (AD-ANT-HT-IC-OS-TUR) Hatay prov., Adana prov., Ġçel prov. (Lodos, 1998); Adana prov.: Yumurtalık, Kozan, Osmaniye prov.: Kadirli, Hatay prov.: Ġskenderun (Gedik, Central), Ġçel prov.: Silifke (Tozlu & Özbek, 2000); Turkey (Sama & Rapuzzi, 2000; Özbek, 2002); Hatay prov.: Dörtyol, Osmaniye prov., Adana prov., Ġçel prov., Antalya prov., Mediterranean Region (Özbek & Tozlu, 2000); Adana prov., Antalya prov., Hatay prov., Ġçel prov., Osmaniye prov. (Tozlu et al., 2003); Ġçel prov.: Central (Karaduvar), AtaĢ road (Karaduvar) (Özdikmen & Hasbenli, 2004); Hatay prov.: Ġskenderun (Sarımazı) (Özdikmen & ġahin, 2006); Hatay prov.: Ġskenderun (Karayılan) (Özdikmen, 2006); Hatay prov.: Arsus (Özdikmen & Demir, 2006); Ġçel prov.: Erdemli, Osmaniye prov.: Central (FakıuĢağı village, Rahime Hatun), Kanlıgeçit village (Özdikmen, 2006); Osmaniye prov.: FakıuĢağı village (Özdikmen et al., 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 58
Distribution: Turkey, Syria, Lebanon, Jordan, Israel, Pakistan, India, China, Comores, Oman, Yemen, Africa, Madagascar and surrounding Islands (St. Thomas, Maurice Island, St. Croix, Seychelles, Reunion Island, Mascarene Islands), Andaman & Nicobar Islands, Barbados, British Virgin Islands, Iran. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey. Chorotype: Afrotropico-Indo-Mediterranean + Neotropic + Oriental.
Tribe MONOCHAMINI
Monochamus Dejean, 1821 [Type sp.: Cerambyx sutor Linnaeus, 1758]
Subgenus Monochamus Dejean, 1821 [Type sp.: Cerambyx sutor Linnaeus, 1758]
Monochamus galloprovincialis (Olivier, 1795)
(*WM, EM) Records in Turkey: (AD-ANT-ART-AY-BO-BS-DE-IC-IP-KAR-KN-KR-KS-MG-OR-SN-TB-TUR) Records in Mediterranean Region: Antalya prov.: Central as M. galloprovincialis v. tauricola Pic, 1912 (Demelt & Alkan, 1962); Antalya prov.: Central / Bey Mountain as M. galloprovincialis ssp. pistor v. tauricola Pic, 1912 (Demelt, 1963); Antalya prov.: Akseki-Cevizli (Urlupelit place) / Dağbucağı place, Isparta prov.: Eğridir (Çamyol forest) (Tosun, 1975); Antalya prov.: Akseki, Isparta prov.: Eğirdir, Mediterranean Region (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Ġçel prov.: Anamur, Antalya prov.: Kumluca (Adlbauer, 1992); Isparta prov., Adana prov., Antalya prov. (Lodos, 1998); Antalya prov.: Central (Tozlu et al., 2003); Antalya prov.: Akseki (Yarpuz env.) as M. galloprovincialis tauricola (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Tribe LAMIINI
Morimus Brullé, 1832 [Type sp.: Lamia lugubris Fabricius, 1792
= Cerambyx asper Sulzer, 1776]
Morimus funereus (Mulsant, 1862) (*WM)
Records in Turkey: (AM-AN-ANT-BI-BO-BR-BS-BU-CA-DU-ES-KK-KO-KR-TO-TUR) Records in Mediterranean Region: Antalya prov.: Akseki (Irmasan pass) (Adlbauer, 1992); Burdur prov.: Bucak (Özdikmen & ġahin, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. According to Sama & Löbl in Löbl & Smetana (2010), it does not occur in Turkey. So, these records should be belong to M. orientalis. Chorotype: Turano-European (Ponto-Pannonian).
Morimus ganglbaueri Reitter, 1894 (*EM)
Records in Turkey: (DE-IS-KA-SA-TUR) Records in Mediterranean Region: KahramanmaraĢ prov. (Kanat, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. According to Sama & Löbl in Löbl & Smetana (2010), it is a synonym of M. asper. Chorotype: Turano-Mediterranean (Balkano-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 59
Tribe DORCADIINI
Dorcadion Dalman, 1817 [Type sp.: Cerambyx glycyrrhizae Pallas, 1773]
Subgenus Cribridorcadion Pic, 1901
[Type sp.: Dorcadion mniszechi Kraatz, 1873]
Dorcadion amanense Breuning, 1943 (*EM)
Other names: Absent. Records in Turkey: (TUR) S Turkey: Amanos Mts. (Breuning, 1943, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Amanos Mts.). Chorotype: Anatolian.
Dorcadion anatolicum Pic, 1900 (*WM, EM)
Other names: ininterruptum Pic; mediointerruptum Pic; subuniforme Pic; brevipenne Pic; chehirense Pic; humeraliinterruptum Breuning; postapertum Breuning; pseudomerkli Breuning; flavosignatum Breuning; seydisehirense Breuning; brignolii Breuning. Records in Turkey: (AD-ANT-IP-KA-KN-TUR) Konya prov. (Pic, 1900); Anatolia as D. brevipenne (Aurivillius, 1921); Turkey (Aurivillius, 1921; Winkler, 1924-1932; Lodos, 1998); Turkey as D. brevipenne Pic, 1902 (Winkler, 1924- 1932; Lodos, 1998); Konya prov. (Breuning, 1962); Konya prov.: AkĢehir (Demelt, 1963); Konya prov.: AkĢehir, Sultan Mts., Ilgın and Kızılören, Antalya prov.: Irmasan pass, Adana prov.-KahramanmaraĢ prov.: between Tufanbeyli and Göksun (Braun, 1978); Konya prov.: Dedegöl Mts. (Adlbauer, 1988); Isparta prov.: Eğirdir, Konya prov. (Önalp, 1990); Konya prov.: TaĢkent (Çukuryurt pass) (Özdikmen & Hasbenli, 2004a); KahramanmaraĢ prov.: Göksun (Özdikmen & Okutaner, 2006); Anatolia (Danilevsky in Löbl & Smetana, 2010); Konya prov.: TaĢkent (FaĢikan plateau), Hadim-Beyreli road (Özdikmen & Turgut, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S and CS Turkey. Subspecific status of this species needs to be clarified. Chorotype: Anatolian.
Dorcadion arcivagum Thomson, 1867 (*EM)
Other names: cilicium Pic. Records in Turkey: (AD-IC-TUR) Ġçel prov.: Gülek (Thomson, 1867); Turkey: Cilician Taurus (Ganglbauer, 1884; Aurivillius, 1921); Turkey (Winkler, 1924-1932; Lodos, 1998); Ġçel prov: Gülek, Adana prov. (Missis) (Breuning, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 60
Dorcadion carinipenne Pic, 1900 (*EM)
Other names: quinquevittatum Breuning. Records in Turkey: (GA-HT-TUR) Hatay prov.: Akbez (Aurivillius, 1921); Hatay prov.: Akbez, Gaziantep prov. (Breuning, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Amanos Mts. and near). Chorotype: Anatolian.
Dorcadion cinerarium (Fabricius, 1787) (*EM)
Records in Turkey: (AN-BS-CN-ES-GA-IC-KA-KI-KS-KY-OR-SM-SV-TO-YO-TUR) Records in Mediterranean Region: Ġçel prov. (from map in Braun, 1979); Gaziantep prov.: Islahiye (Özdikmen et al., 2005); KahramanmaraĢ prov.: Göksun (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Moreover, this species can not be represented in Turkey. According to Lazarev (2009), Dorcadion populations from Turkey were wrongly regarded as different forms of D. cinerarium (D. micans Thomson, 1867; D. sericatulum Kraatz, 1873; D. macropus Kraatz, 1873). He also stated Dorcadion populations that were attributed before to D. cinerarium from Turkey (as subspecies, variations, morphs, forms or aberrations) belong to another partly not described species (see Özdikmen, 2010). Chorotype: E-Mediterranean (NE-Mediterranean).
Dorcadion culminicola Thomson, 1868 (*EM)
Other names: Absent. Records in Turkey: (TUR) Turkey: Taurus (Ganglbauer, 1884; Aurivillius, 1921); Anatolia (Winkler, 1924-1932); Cilician Taurus (Breuning, 1962); Turkey (Lodos, 1998); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Cilicia). Chorotype: Anatolian.
Dorcadion delagrangei Pic, 1894 (*EM)
Other names: fuscopubescens Pic. Records in Turkey: (GA-HT-KL-OS-TUR) Hatay prov.: Akbez (Aurivillius, 1921); Gaziantep prov., Bolkar Mts., Amanos Mts. as D. accola (Breuning, 1962); Hatay prov.: Akbez as D. delagrangei (Pic, 1894) (Ex. Braun, 1978); Kilis prov., Osmaniye prov.: Nurdağ (Braun, 1978); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Amanos Mts. and near). Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 61
Dorcadion deyrollei Ganglbauer, 1884 (*EM)
Other names: melas Breuning; subalbipenne Breuning. Records in Turkey: (BT-KA-VA-TUR) Bitlis prov. as the type loc. (Ganglbauer, 1884); NE Turkey (Plavilstshikov, 1958); Bitlis prov., Van prov., KahramanmaraĢ prov.: Ahır Mt. (Breuning, 1962); Van prov.: Kuzgunkıran (Fuchs & Breuning, 1971; Holzschuh, 1980; Önalp, 1990); Turkey (Lodos, 1998); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from SE Turkey. Chorotype: Anatolian.
Dorcadion divisum Germar, 1839 (*WM, EM)
Records in Turkey: (AD-ADY-AN-BL-BS-BU-CA-DI-ES-IC-IP-IZ-KN-MN-MR-NI-OS-SV- TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Burna as D. köchlini Pic (Bodemeyer, 1900); Cilicia, West and South Anatolia eastwards to South-Eastern Anatolian Region (Breuning, 1962); Burdur prov. (Demelt, 1963); Burdur prov. – Demelt, 1963 (Ex. Gül-Zümreoğlu, 1975); Adana prov.: Tufanbeyli as D. divisum m. intercisum (Braun, 1978); Isparta prov.: Eğirdir (Önalp, 1990); Osmaniye prov.: Between Kaypak-Alamanpınarı (Özdikmen, 2006). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Dorcadion drusoides Breuning, 1962 (*EM)
Other names: Absent. Records in Turkey: (TUR) Turkey (not Syria): Amanos Mts. (Breuning, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Amanos Mts.). Chorotype: Anatolian.
Dorcadion halepense (Kraatz, 1873) (*EM)
Other names: internesignatum Pic; latealbum Pic; internenotatum Pic; latebivittatum Pic; tabense Pic; pseudomardinense Breuning; brunneosericans Breuning; akbesianum Breuning; pseudosimile Breuning; dorsoinvittatum Breuning. Records in Turkey: (GA-HT-TUR) Gaziantep prov. (Breuning, 1962); Gaziantep prov.: Sakçagözü, Kilis prov. (Braun, 1978); Gaziantep prov.: Toros Mountains, Hatay prov.: Akbez, Toros Mountains (Önalp, 1990); Turkey (Lodos, 1998); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey (Amanos Mts. and near). Chorotype: SW-Asiatic (Syro-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 62
Dorcadion holtzi (Pic, 1905) (*EM)
Other names: Absent. Records in Turkey: (IC) Ġçel prov.: Bolkar Mts. as D. culminicola var. holtzi (Bodemeyer, 1906); Cilician Taurus: Funduk-Pungar (Breuning, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from S Turkey (Cilicia). Chorotype: Anatolian.
Dorcadion infernale Mulsant & Rey, 1863 (*WM, EM)
Records in Turkey: (AM-AN-ANT-BI-BU-CO-DI-ES-IC-IZ-KA-KN-NI-SV-US-TUR) Records in Mediterranean Region: Burdur prov.: Karakent as Dorcadion infernale; Antalya prov.: Toros Mountains as Dorcadion infernale var. revestitum (Bodemeyer, 1900); KahramanmaraĢ prov.: Ahır Mountain (Önalp, 1990); Ġçel prov.: Tarsus (Çamlıyayla) (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Anatolian.
Dorcadion ispartense Breuning, 1962 (*WM)
Other names: Absent. Records in Turkey: (IP) Isparta prov.: Bozanönü (Breuning, 1962). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Isparta province in SW Turkey. Chorotype: Anatolian.
Dorcadion kraetschmeri Bernhauer, 1988 (*WM)
Other names: Absent. Records in Turkey: (ANT-TUR) Antalya prov.: Akseki (Bernhauer, 1988); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Dorcadion lohsei Braun, 1976 (*EM)
Other names: sertavuli Breuning. Records in Turkey: (IC-KM-KN-TUR) Karaman prov., Ġçel prov.: Mut (Sertavul pass) (Braun, 1978); Ġçel prov.: Sertavul pass (Sama, 1982; Adlbauer, 1992); Turkey (Lodos, 1998); Anatolia (Danilevsky in Löbl & Smetana, 2010); Konya prov.: TaĢkent (FaĢikan plateau) (Özdikmen & Turgut, 2010). Distribution: Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 63
Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey. Chorotype: Anatolian.
Dorcadion menradi Holzschuh, 1989 (*EM)
Other names: Absent. Records in Turkey: (KA-MA-TUR) KahramanmaraĢ prov.: AfĢin, Malatya prov. (Holzschuh, 1989); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey. Chorotype: Anatolian.
Dorcadion micans Thomson, 1867 (*EM)
ssp. micans Thomson, 1867 ssp. susheriense Breuning, 1970 Other names: amasinum Pic; atripes Reitter; corallicornis Pic; subobesum Pic; macropus Kraatz; sericatulum Kraatz; obscurans Pic; paracinerarium Breuning; subreductum Pic; tokatense Pic. Records in Turkey: (AM-AN-BS-CA-CO-IC-IZ-KM-SV-SU-TO-YO-TUR) Amasya prov. as D. amasinum (Aurivillius, 1921); Anatolia as D. caucasicum a. corallicorne Pic, 1904, D. caucasicum a. corallicorne Pic, 1904, D. caucasicum v. macropus Kraatz, 1873 and D. amasinum Pic, 1898 (Winkler, 1924-1932); East Anatolia, southward to Bolkar Mountains as D. c. m. micans / Amasya prov. and Ankara prov. as D. c. m. corallicorne / Ankara prov. and Bursa prov. as D. c. m. sericatulum / Amasya prov. and Tokat prov. as D. c. m. macropus / North Turkey as D. c. m. subobesum and D. c. m. subreductum, D. c. m. perroudi, D. c. m. amasinum (Breuning, 1962); Ankara prov.: Elmadağ as D. c. micans (Demelt, 1963); Ankara prov.: GölbaĢı as D. c. micans (Perissinotto & Luchini, 1966); Çorum prov.: Boğazkale as D. cinerarium m. amasinum (Perissinotto & Luchini, 1966); Amasya as D. c. micans (Breuning et Villiers, 1967); Yozgat prov.: Central as D. cinerarium amasinum (Fuchs et Breuning, 1971); ġanlıurfa prov. as D. sericatum micans (Gül & Zümreoğlu, 1972); Ġzmir prov.: Bornova, Çanakkale prov.: Ezine as D. cinerarium m. corallicorne (Gül-Zümreoğlu, 1975); Ġçel prov.: Sertavul pass, Karaman prov. as D. cinerarium m. paracinerarium (Braun, 1978 and 1979); Turkey as D. c. subobesum, D. c. micans (Braun, 1979); Sivas prov. as D. cinerarium susheriense Breuning, 1970 (Ex. Braun, 1979); Yozgat prov.: Central as D. c. micans (Ex. Holzschuh, 1980); Turkey as D. amasinum (Lodos, 1998); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species is distributed rather widely in Turkey. It evaluated in Part I under D. cinerarium. This species is endemic to Turkey. It has two subspecies. Danilevsky in Löbl & Smetana (2010) gave it as a separate species. Chorotype: Anatolian.
Dorcadion miminfernale Breuning, 1970 (*EM)
Other names: Absent. Records in Turkey: (KA-KY) KahramanmaraĢ prov.: Binboğa Mts. (Breuning, 1970); Kayseri prov.: Bakırdağı, KahramanmaraĢ prov.: Göksun and Yalakköy (Braun, 1978).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 64
Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The species has been known only from S Turkey. This species is endemic to CSE Turkey. Danilevsky in Löbl & Smetana (2010) accepted it as a synonym of D. infernale infernale. Chorotype: Anatolian.
Dorcadion oezdurali Önalp, 1988 (*EM)
Other names: Absent. Records in Turkey: (ADY-KA) KahramanmaraĢ prov.: Ahır Mt. (Önalp, 1990); Adıyaman prov.: Nemrut Mt. (Rejzek & Hoskovec, 1999); KahramanmaraĢ prov.: Göksun (Mehmetbey) (Özdikmen & Okutaner, 2005); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from CSE Turkey. Chorotype: Anatolian.
Dorcadion ortrudae Braun, 1980 (*EM)
Other names: Absent. Records in Turkey: (KA-TUR) KahramanmaraĢ prov.: Göksun (Adlbauer, 1988); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known only from CSE Turkey. Chorotype: Anatolian.
Dorcadion paracinerarium Breuning, 1974 (*EM)
Other names: Absent. Records in Turkey: (IC-KM) Ġçel prov.: Mut (Sertavul pass) (Breuning, 1974); Ġçel prov.: Mut (Sertavul pass), Karaman prov. (Braun, 1978 and 1979); Ġçel prov. (Demelt ex Braun, 1979). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to SCW Turkey (Cilician Taurus). According to Braun (1979), D. paracinerarium Breuning, 1974 is a morpha of D. cinerarium (Fabricius, 1787). Chorotype: Anatolian.
Dorcadion petrovitzi Heyrovsky, 1964 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Ġçel prov. (Heyrovsky, 1964); Ġçel prov.: Namrun (Demelt, 1967); Turkey (Lodos, 1998). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to SCW Turkey (Cilician Taurus). It has been known only from the type locality. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 65
Dorcadion pseudinfernale Breuning, 1943 (*EM)
Other names: pseudimmutatum Breuning. Records in Turkey: (TUR) S Turkey: Amanos Mts. (Breuning, 1943, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to Amanos Mts. in SC Turkey. It has been known only from type locality until now. Chorotype: Anatolian.
Dorcadion saulcyi Thomson, 1865 (*EM)
ssp. saulcyi Thomson, 1865 ssp. javeti Kraatz, 1873 Other names: simile Kraatz; destinoi Fairmaire; fenestratum Pic; adanense Pic; univestitum Pic; subelongatum Pic; tabense Pic; adanense m. mersinense Breuning; haleppense Kraatz. Records in Turkey: (AD-GA-HT-IC-MA-OS-TUR) Hatay prov.: Akbez as D. destinoi (Fairmaire, 1884); Turkey: Ġçel prov. as D. saulcyi v. javeti (Ganglbauer, 1884; Aurivillius, 1921); Malatya prov. as D. simile (Ganglbauer, 1886; Aurivillius, 1921); Hatay prov.: Akbez as D. destinoi (Ganglbauer, 1886; Aurivillius, 1921); Adana prov. as D. destinoi v. adanense (Aurivillius, 1921); Anatolia as D. saulcyi v. javeti (Winkler, 1924-1932); Adana prov. as D. destinoi v. adanense Pic, 1905 (Winkler, 1924-1932); Malatya prov. as D. simile (Kraatz, 1884); Anatolia as D. saulcyi v. simile (Winkler, 1924-1932); Cilicia: Ġçel prov.: Gülek, Malatya prov. (Breuning, 1962); Hatay prov.: Ġskenderun, Osmaniye prov.: Hasanbeyli (Demelt, 1963); Osmaniye prov.: Hasanbeyli, Amanos Mountains D. saulcyi javeti (Fuchs et Breuning, 1971); Osmaniye prov.: Hasanbeyli, Amanos Mountain as D. saulcyi fenestratum (Fuchs et Breuning, 1971); Osmaniye prov.: Toprakkale, Hatay prov.: Antakya (Yayladağı) (Braun, 1978); Anatolia (Önalp, 1990); Hatay: Akbez, Amanos Mountains D. saulcyi javeti (ex Önalp, 1990); Hatay prov.: Akbez, Adana prov. as D. destinoi (Önalp, 1990); Hatay prov.: Antakya (Harbiye) (Adlbauer, 1992); Turkey (Lodos, 1998); Turkey as D. destinoi (Lodos, 1998); Gaziantep prov.: Islahiye (Özdikmen et al., 2005); Anatolia (Danilevsky in Löbl & Smetana, 2010); Hatay prov.: Ġskenderun (Sakıtlı plateau) as D. saulcyi saulcyi (Özdikmen et al., 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species has two subspecies. D. saulcyi javeti Kraatz, 1873 that described from Akbez in Turkey (not Syria), probably occurs only in Amanos Mts. for Turkey and Syria. So all old records from Amanos Mts. in Turkey should be belonging to the subspecies, D. saulcyi javeti Kraatz, 1873. It is represented by both subspecies in central parts of S Turkey. Besides, Breuning (1962) mentioned the type locality, Syria is mistaken information (―irrige Angabe‖). I also think the type locality of this species should be Turkey. So this species is endemic to Turkey. It is distributed in SC, SCW and CSE Turkey. Chorotype: Anatolian.
Dorcadion scabricolle Dalman, 1817 (*WM, EM)
Records in Turkey: (AD-AF-AG-AN-ANT-AR-BI-BL-BS-CO-ER-EZ-GU-IC-IP-KA-KAR-KN-KS-KY-MA-NI-SV-US-VA-YO-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mts. as D. s. v. caramanicum (Bodemeyer, 1900); Cilician Taurus as D. s. caramanicum Daniel, 1903 (Breuning, 1962); Ġçel prov.: Sertavul pass, KahramanmaraĢ prov.: Göksun / Elbistan, Adana prov.: Saimbeyli / Tufanbeyli (Braun, 1978); Isparta prov., Ġçel prov., KahramanmaraĢ prov. (from map in

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 66
Braun, 1978); KahramanmaraĢ prov.: Göksun (Central / Küçüksu plateau / Korkmaz) (Özdikmen & Okutaner, 2006); KahramanmaraĢ prov.: AfĢin (Emirilyas village, Mağaraözü district) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Dorcadion semibrunneum Pic, 1903 (*WM, EM)
ssp. semibrunneum Pic, 1903 ssp. anamasum Pic, 1934 Other names: mediocreimpressum Pic; medioimpressum Pic; boszdaghanum Breuning. Records in Turkey: (AG-EZ-IP-IZ-KA-KAR-SV-TUR) ?Ġzmir prov.: Bozdağ as D. mniszechi var. semibrunneum (Bodemeyer, 1906); Anatolia as D. (C.) mniszechi v. semibrunneum (Aurivillius, 1921); Anatolia as D. mniszechi v. semibrunneum (Winkler, 1924-1932);?Ġzmir prov.: Bozdağ to ÖdemiĢ as D. mniszechi ssp. semibrunneum, Pisidian Taurus: ?Isparta prov.: Anamas Mts. as D. mniszechi ssp. anamasum (Breuning, 1962); KahramanmaraĢ prov.: Çağlayancerit (Bozlar) as D. semibrunneum (Özdikmen & Okutaner, 2006); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*).This species has two subspecies as the nominotypical subspecies and D. semibrunneum anamasum Pic, 1934 which described from Pisidian Taurus: ?Isparta prov.: Anamas Mts. in the original combination as D. mniszechi ssp. anamasum Pic, 1934. Both subspecies are distributed only in Anatolia. In this status, probably D. semibrunneum semibrunneum Pic, 1903 is distributed in W Anatolia and D. semibrunneum anamasum Pic, 1934 is distributed in S Anatolia. So, the old KahramanmaraĢ record of Özdikmen & Okutaner (2005) belongs to the subspecies, D. mniszechi ssp. anamasum Pic, 1934 very likely. Chorotype: Anatolian.
Dorcadion septemlineatum Waltl, 1838 (*WM)
Records in Turkey: (AF-AN-BI-BL-BO-BS-BU-CA-ES-GA-IP-IS-KN-KO-KR-KU-SA-TRA-TUR) Records in Mediterranean Region: Burdur prov.: Karakent as D. septemlineatum novemlineatum (Bodemeyer, 1900); Isparta prov.: Eğirdir (Önalp, 1990). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Dorcadion sinuatevittatum Pic, 1937 (*WM, EM)
Other names: brevemediolineatum Pic. Records in Turkey: (DE-BU-KA-TUR) Denizli prov.: Carian Taurus, Babadağ (Breuning, 1962); Burdur prov. as D. bangi (Özdikmen et al., 2005); KahramanmaraĢ prov.: Göksun (Mehmetbey bridge) as D. bangi heinzorum Braun, 1975 (Özdikmen & Okutaner, 2006); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*). This species has been known from S Turkey. It has been known only from type locality until now. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 67
Dorcadion smyrnense (Linneaus, 1757) (*EM)
Records in Turkey: (BI-BL-BS-ES-IS-IZ-KN-KU-OS-US-TRA-TUR) Records in Mediterranean Region: Osmaniye prov.: Bahçe as D. crux var. integrofasciatum (Fuchs et Breuning, 1971). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Anatolian.
Dorcadion subinterruptum Pic, 1900 (*EM)
Other names: indivisum Pic; interruptevittatum Breuning; exiguum Breuning; posticeconjunctum Breuning. Records in Turkey: (ES-?IC-IZ-KN-TUR) Konya prov. as D. divisum v. indivisum and D. divisum ab. subinterruptum (Aurivillius, 1921); Ġzmir prov.: Bozdağ, EskiĢehir prov., Konya prov., ?Ġçel prov.: Bolkar Mts. (Breuning, 1962); Konya prov.: Ilgın (Braun, 1978); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). The endemic species has been known W, C and S Turkey. Chorotype: Anatolian.
Dorcadion syriense Breuning, 1943 (*EM)
Other names: Absent. Records in Turkey: (TUR) Turkey: Amanos Mts. (Breuning, 1943 and 1962). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to SC Turkey. It has been known only from type locality until now. The type locality was given as Syria: Amanos Mts. by Breuning. Amanos Mts. is in S Turkey, not Syria. Chorotype: Anatolian.
Dorcadion tuerki Ganglbauer, 1884 (*EM)
Other names: internestriatum Breuning; stramentosoides Breuning; posticefibulatum Plavilstshikov. Records in Turkey: (TUR) S Turkey: Amanos Mts. (=Nur Mts.) (Demelt, 1963). Distribution: Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to Iran now. Since Danilevsky (2010b) gave D. türki v. transcaspicum Jakovlev, 1901 which was regarded as a subspecies of D. türki by Breuning (1962), as a synonym of D. komarowi Jakovlev, 1887. However, it has been recorded by Demelt (1963) from Amanos Mts. (S Turkey) for Turkey. This doubtful record probably belongs to another Dorcadion species. It needs to be confirmed. So this species is not confirmed for Turkey now. Chorotype: Iranian or SW-Asiatic (Irano-Anatolian).
Dorcadion variegatum Ganglbauer, 1884 (*EM)
Other names: Absent.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 68
Records in Turkey: (GA-KA-TUR) KahramanmaraĢ prov., Gaziantep prov. (Breuning, 1962); Anatolia (Danilevsky in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).This species is distributed only in central parts of S Anatolia for Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Subgenus Maculatodorcadion Breuning, 1942 [Type sp.: Dorcadion quadrimaculatum Küster, 1848]
Dorcadion triste Frivaldsky, 1845
(*WM) Records in Turkey: (ANT-BL-BS-IZ-TUR) Records in Mediterranean Region: Antalya prov.: Ova SE Korkuteli, Bakacak beli N Saklıkent as the type loc. of D. triste phrygicum Peks, 1889 (Peks, 1993). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*). This species is endemic to W and SW Turkey. It has two subspecies as D. triste triste (western subspecies) and D. triste phrygicum (eastern subspecies). The later was collected by Chris Bruggeman (Belgium) from Antalya prov.: Aspendos in April 2010 (pers. comm.. 2010). Chorotype: Anatolian.
Tribe APODASYINI
Anaesthetis Dejean, 1835 [Type sp.: Saperda testacea Fabricius, 1781]
Anaesthetis anatolica Holzschuh, 1969
(*WM, EM) Other names: Absent. Records in Turkey: (ANT-IC-KA-TUR) Antalya prov.: Ġncekum, Ġçel prov.: Silifke, KahramanmaraĢ prov.: Andırın (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria, Israel, Lebanon. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).This species is distributed only in S Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).
Tribe PTEROPLIINI
Niphona Mulsant, 1839 [Type sp.: Niphona picticornis Mulsant, 1839]
Subgenus Niphona Mulsant, 1839
[Type sp.: Niphona picticornis Mulsant, 1839]
Niphona picticornis Mulsant, 1839 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-HT-IC-KA-MG-TUR) Hatay prov.: Akbez (Fairmaire, 1884); Turkey (Ġyriboz, 1940; Bodenheimer, 1941, 1958; Ġren & Ahmed, 1973; Sama & Löbl in Löbl & Smetana, 2010); Antalya prov.: Manavgat (Demelt &

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 69
Alkan, 1962; Demelt, 1963); Ġçel prov.: Erdemli, KahramanmaraĢ prov.: Andırın, Antalya prov.: Alanya (Adlbauer, 1988); Antalya prov.: Manavgat, from Ġçel prov. to Muğla prov.: Dalaman (Lodos, 1998); Whole Mediterranean Region (Sama, 2002); Antalya prov. (Tozlu et al., 2003); Muğla prov.: Fethiye (Günlüklü place) (Özdikmen & Hasbenli, 2004). Distribution: Turkey, Syria, Israel, Lebanon. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).This species is distributed only in S Turkey. Chorotype: Mediterranean.
Tribe POGONOCHERINI
Pogonocherus Dejean, 1821 [Type sp.: Cerambyx hispidus Linnaeus, 1758]
Subgenus Pogonocherus Dejean, 1821
[Type sp.: Cerambyx hispidus Linnaeus, 1758]
Pogonocerus anatolicus Mulsant, 1839 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-IC-TUR) Anatolia (Winkler, 1924-1932); Antalya prov.: near Central, Kumköy (Schimitschek, 1944, 1953); Antalya prov.: Alanya (Demelt, 1963); Antalya prov.: Serik (Belek forest), Kumköy (Tosun, 1975); Ġçel prov.: Silifke, Antalya prov.: Alanya (Güzelbağ), Kemer (Adlbauer, 1988); Turkey (Lodos, 1998; Özdikmen & ġahin, 2006; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: Silifke (Tauzin, 2000). Distribution: Europe (Rodos), Turkey, Cyprus, Syria. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S Turkey. Chorotype: E-Mediterranean.
Pogonocerus perroudi Mulsant, 1839 (*EM)
Records in Turkey: (CA-HT-IC-IS-IZ-KAR-KR-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Ġçel prov.: Anamur (Adlbauer, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Mediterranean.
Tribe ACANTHODERINI
Aegomorphus Haldeman, 1847 [Type sp.: Aegomorphus decipiens Haldeman, 1847
= Lamia modesta Gyllenhal, 1817]
Aegomorphus clavipes (Schrank, 1781) (*EM)
Records in Turkey: (AM-ED-HT-IS-IZ-KO-MN-TU-TRA-TUR) Records in Mediterranean Region: Hatay prov.: around Dörtyol (Amanos Mts.) (Öymen, 1987); Hatay prov.: Amanos Mountains / Akbez (Sama, 1994c). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 70
Tribe ACANTHOCININI
Acanthocinus Dejean, 1821 [Type sp.: Cerambyx aedilis Linnaeus, 1758]
Acanthocinus aedilis (Linnaeus, 1758)
(*WM) Records in Turkey: (AM-AN-ANT-ART-BI-BL-BO-BS-CA-DE-ES-EZ-GI-GU-IP-IZ-KAR-KR-KS-KU-MG-SN-TO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Akseki-Cevizli (Urlupelit forest) / Nebiler forest / Korkuteli (Hacıbekar forest), Isparta prov.: Eğridir (Sarıidrisler forest) (Tosun, 1975); Antalya prov.: Akseki, Isparta prov.: Eğirdir (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Antalya prov. (Lodos, 1998). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Acanthocinus griseus (Fabricius, 1792) (*WM, EM)
Records in Turkey: (AD-ANT-BS-DE-ED-EZ-HT-IC-IZ-KO-MG-MN-TB-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun as Acanthocinus griseus ssp. novaki Tippm. (Demelt, 1967); Antalya prov.: Manavgat / TaĢağıl / KaĢ / Sütleğen (Sinekçibeli place) / Akseki (Murtiçi place) / Nebiler forest (Tosun, 1975); Adana prov., Antalya prov. (Erdem & Çanakçıoğlu, 1977); Ġçel prov.: Anamur, Hatay prov.: Antakya (Tekkepınar) (Adlbauer, 1992); Adana prov. (Lodos, 1998); Adana prov.: Kozan (Tozlu et al., 2003). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
Leiopus Audinet-Serville, 1835 [Type sp.: Cerambyx nebulosus Linnaeus, 1758]
Leiopus nebulosus (Linnaeus, 1758)
(*WM) Records in Turkey: (ART-IP-IS-KIR-TO-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Gölcük (Özdikmen et al., 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Leiopus syriacus (Ganglbauer, 1884) (*WM, EM)
Other names: major Pic. Records in Turkey: (ANT-BU-IC-KA-NI-TUR) Antalya prov.: Toros Mountains, Niğde prov.: Çamardı (Bodemeyer, 1900); Burdur prov. (Demelt, 1963); Ġçel prov.: Tarsus (Çamlıyayla), Niğde prov.: Çiftehan (Adlbauer, 1988); Turkey (Sama & Rapuzzi, 2000; Sama & Löbl in Löbl & Smetana, 2010); KahramanmaraĢ prov.: AfĢin (Çardak-AfĢin road, Ördek village), Pazarcık (Büyük Nacar) (Özdikmen & Okutaner, 2006). Distribution: Turkey, Cyprus, Syria, Israel, Lebanon. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 71
Exocentrus Dejean, 1835 [Type sp.: Callidium lusitanicum Olivier, 1790
= Cerambyx lusitanus Linnaeus, 1767]
Subgenus Exocentrus Dejean, 1835 [Type sp.: Callidium lusitanicum Olivier, 1790
= Cerambyx lusitanus Linnaeus, 1767]
Exocentrus adspersus Mulsant, 1846 (*EM)
Records in Turkey: (IS-KA-KO-TRA-TUR) Records in Mediterranean Region: KahramanmaraĢ prov.: Andırın (Adlbauer, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Exocentrus punctipennis Mulsant & Guillebeau, 1856 (*WM)
Records in Turkey: (ANT-IS-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Manavgat (Adlbauer, 1992). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Exocentrus ritae Sama, 1985 (*EM)
Other names: Absent. Records in Turkey: (IC-TUR) Ġçel prov.: Çamlıyayla (Namrun) (Sama, 1985); Ġçel prov.: Erdemli (Adlbauer, 1988); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from SC Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Tribe TETROPINI
Tetrops Stephens, 1829 [Type sp.: Leptura praeusta Linnaeus, 1758]
Tetrops praeustus (Linnaeus, 1758)
(*WM) Records in Turkey: (AN-ANT-BI-CO-IS-KN-NI-SA-SM-SN-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mountains (Bodemeyer, 1900); Ġçel prov. (Özdikmen et al., 2005); Antalya prov.: Alanya-TaĢkent (Exit of Karapınar village), Alanya (Karapınar-Sarımut) as T. praeustus anatolicus (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palearctic.
Tetrops warnckei Holzschuh, 1977 (*WM)
Other names: Absent. Records in Turkey: (ANT-TUR)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 72
Antalya prov.: Toros Mts. (Akseki) (Holzschuh, 1977); Anatolia (Holzschuh, 1981b; Sama & Löbl in Löbl & Smetana, 2010); Antalya prov.: Akseki (Yarpuz) (Adlbauer, 1992). Distribution: Turkey. Remarks: It has been reported only from Western Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from Antalya province in S Turkey. Chorotype: Anatolian.
Tribe SAPERDINI
Saperda Fabricius, 1775 [Type sp.: Cerambyx carcharias Linnaeus, 1758]
Subgenus Lopezcolonia Alonso-Zarazaga, 1998
[Type sp.: Saperda tremulae Fabricius, 1775 = Leptura octopunctata Scopoli, 1772]
Saperda punctata (Linnaeus, 1767)
(*WM) Records in Turkey: (ANT-ART-BY-GI-GU-IS-IZ-TB-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Manavgat (Demelt, 1963). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: W-Palaearctic.
Subgenus Compsidia Mulsant, 1839 [Type sp.: Cerambyx populneus Linnaeus, 1758]
Saperda populnea (Linnaeus, 1758)
(*WM, EM) Records in Turkey: (ART-BL-BO-BS-BT-BU-BY-CA-DE-DU-ED-ER-GI-GU-HT-IS-IZ-KA-KN-KO-KY-MN-MU-SA-SM-SN-TB-VA-YO-ZO-TUR) Records in Mediterranean Region: Hatay prov.: around Ġskenderun (Öymen, 1987); Burdur prov.: Gökçebağ (Adlbauer, 1992); KahramanmaraĢ prov. (Kanat, 1998); KahramanmaraĢ prov.: Pazarcık (Aksu bridge) (Özdikmen& Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Holarctic.
Saperda quercus Charpentier, 1825 (*WM, EM)
Records in Turkey: (AD-ADY-ANT-IC-KA-OS-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun as S. quercus ocellata (Demelt, 1967); Adana prov.: Tekir, Osmaniye prov.: Nurdağı pass, KahramanmaraĢ prov.: Andırın as S. quercus ocellata (Adlbauer, 1988); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km) as S. quercus ocellata (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: E-Mediterranean or Turano-Mediterranean (Balkano-Anatolian) + E-Mediterranean (Palaestino-Taurian).
Tribe PHYTOECIINI
Oberea Dejean, 1835 [Type sp.: Cerambyx linearis Linnaeus, 1760]
Subgenus Oberea Dejean, 1835
[Type sp.: Cerambyx linearis Linnaeus, 1760]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 73
Oberea linearis (Linnaeus, 1760) (*EM)
Records in Turkey: (ART-GI-HT-IS-OR-OS-RI-TB-TUR) Records in Mediterranean Region: Hatay prov.: Ġskenderun (Breuning et Villiers, 1967); Osmaniye prov.: Entry of Yarpuz (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Oberea oculata (Linnaeus, 1758) (*WM, EM)
Records in Turkey: (AD-ADY-AN-ANT-DE-EZ-HT-IC-IP-IZ-KA-KN-KO-MG-NI-TU-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Manavgat (Öymen, 1987); Antalya prov.: Çukurbağ (Adlbauer, 1992); Adana prov., Antalya prov., Hatay prov., Ġçel prov.: Erdemli, Silifke and Tarsus (Çanakçıoğlu, 1993; Çanakçıoğlu & Mol, 1998); Çukurova (South Turkey) (Lodos, 1998); KahramanmaraĢ prov.: Yenicekale (kayıĢlı), Isparta prov.: Yalvaç (Sultan Mts.) (Özdikmen & Hasbenli, 2004). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palaearctic.
Subgenus Amaurostoma Müller, 1906 [Type sp.: Cerambyx erythrocephalus Schrank, 1776]
Oberea erythrocephala (Schrank, 1776)
(*WM, EM) Records in Turkey: (AF-AM-AN-ANT-ART-BY-CO-ER-EZ-GU-IS-KA-KAR-KN-KO-KS-NI-OS-SV-VA-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Alanya as O. erythrocephala var. insidiosa Muls. (Demelt & Alkan, 1962); Antalya prov.: Antitoros Mountains as O. erythrocephala schurmanni Heyr. (Demelt & Alkan, 1962); Antalya prov.: Alanya as O. erythrocephala insidiosa, Antalya prov.: Alanya / Thermessos as O. erythrocephala schurmanni – Demelt, 1961 and 1963 (Ex. Öymen, 1987); Antalya prov.: Alanya (Adlbauer, 1988); Antalya prov.: Alanya (Dim stream) as O. erythrocephala schurmanni (Adlbauer, 1988); Osmaniye prov.: Nurdağı pass as O. erythrocephala taygetana (Adlbauer, 1988); KahramanmaraĢ prov.: Göksun (Göksun-Çardak road, exit of Saraycık) as O. erythrocephala schurmanni (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palaearctic.
Oxylia Mulsant, 1862 [Type sp.: Phytoecia languida Ménétriés, 1839]
Oxylia argentata (Ménétriés, 1832)
(*WM, EM) Records in Turkey: (ADY-AG-AN-ANT-ART-BT-BY-CO-DI-EL-ER-EZ-GI-GU-HT-IC-IP-IZ-KAR-KI-KN-KS-NI-SI-SN-YO-TUR) Records in Mediterranean Region: Ġçel prov.: Bolkar Mountains (Villiers, 1959); Isparta prov.: Eğirdir as Oxylia languida Muls. (Demelt & Alkan, 1962); Ġçel prov.: Namrun (Bolkar Mt.) (Demelt, 1967); Antalya prov.: Ġrmasan pass (Rejzek et al., 2001); Isparta prov.: Yalvaç (between Sücüllü-Yalvaç dam) (Özdikmen & Hasbenli, 2004); Hatay prov.: Ġskenderun (Güzelyayla road) (Özdikmen & Demirel, 2005); Isparta prov.: Uluborlu as Oxylia languida (Özdikmen et al., 2005); Antalya prov.: Akseki-Güzelsu (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 74
Oxylia duponcheli (Brullé, 1832) (*WM, EM)
Records in Turkey: (AK-AN-ANT-ART-ES-IC-KA-KM-KN-MA-MN-OS-TUR) Records in Mediterranean Region: Osmaniye prov.: Zorkun plateau road (OlukbaĢı place) (Özdikmen & Demirel, 2005); Ġçel prov.: entry of Fındıkpınarı / Mut-Karaman road (Gökçeören pine grove) (Özdikmen, 2006); KahramanmaraĢ prov.: Göksun (Göksun-Çardak) (Özdikmen & Okutaner, 2006); Antalya prov.: Ġbradı-Akseki road, Akseki (Güzelsu village, EmiraĢıklar village env.), Ġbradı (BaĢlar village env.) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Mallosia Mulsant, 1862 [Type sp.: Saperda graeca Sturm, 1843]
Subgenus Semnosia Daniel, 1904
[Type sp.: Saperda scovitzii Faldermann, 1837]
Mallosia mirabilis (Faldermann, 1837) (*EM)
ssp. mirabilis Faldermann, 1837 ssp. devexula Holzschuh, 1989 Other names: kotschyi Hampe; ganglbaueri Kraatz; bitlisiensis Pic; delagrangei Pic; multimaculata Pic; semirubra Pic. Records in Turkey: (BT-HA-KA-MA-TUR) Malatya prov. (Heyden, 1888); Anatolia as S. mirabilis a. ganglbaueri Kraatz, 1884 (Winkler, 1924-1932); Bitlis prov. as S. mirabilis a. bitlisiensis Pic, 1902 (Winkler, 1924-1932); KahramanmaraĢ prov.: Elbistan (Demelt, 1967); Hakkari prov.: Sat (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Iraq, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from SE Turkey. It is represented by all subspecies in Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).
Subgenus Eumallosia Danilevsky, 1990 [Type sp.: Mallosia herminae Reitter, 1890]
Mallosia imperatrix Abeille de Perrin, 1885
(*EM) Other names: robusta Pic; tauricola Daniel. Records in Turkey: (ADY-BN-BT-EZ-HA-OS-VA-TUR) Nur Mt. (Demelt, 1967); Osmaniye prov.: Nurdağı pass, Bingöl prov. (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Adıyaman prov.: Karadut village, Nemrut Mt. as ssp. tauricola Daniel, 1904 (Rejzek & Hoskovec, 1999); Hakkari prov.: Suvarthulil (Tauzin, 2000); Bingöl prov., Bitlis prov., Erzurum prov. (Tozlu et al., 2003); Turkey as ssp. tauricola Daniel, 1904 (Tozlu et al., 2003); Van prov.: Van-Bahçesaray road (Narlıca) as ssp. tauricola Daniel, 1904 (Özdikmen, 2006). Distribution: Turkey, Syria, Lebanon, Iraq, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from SE Turkey. It is represented by all subspecies in Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 75
Semiangusta Pic, 1893 [Type sp.: Conizonia delagrangei Pic, 1891]
Semiangusta delagrangei (Pic, 1891)
(*EM) Other names: brevior Pic. Records in Turkey: (AD-MA-TUR) Malatya prov. (Demelt, 1967); Adana prov.: Toprakkale (Adlbauer, 1988); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S and SE Turkey. Chorotype: Anatolian.
Coptosia Fairmaire, 1864 [Type sp.: Phytoecia languida Fairmaire, 1864
= Phytoecia albovittigera Heyden, 1863]
Subgenus Coptosia Fairmaire, 1864 [Type sp.: Phytoecia languida Fairmaire, 1864
= Phytoecia albovittigera Heyden, 1863]
Coptosia bithynensis (Ganglbauer, 1884) (*WM, EM)
Records in Turkey: (AD-ADY-ANT-BI-BS-DI-EL-EZ-HT-IC-IP-IZ-MU-OS-TUR) Records in Mediterranean Region: Adana prov.: Kozan, Isparta prov.: Eğirdir, Ġçel prov.: Çamlıyayla as Conizonia bithynensis Ganglbauer (Demelt, 1963); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Antalya prov.: Ġbradı, Ġbradı-Akseki road (Özdikmen & Turgut, 2010); Hatay prov.: Antakya (Saint Pierre church env.) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Balkan).
Coptosia ganglbaueri Pic, 1936 (*EM)
Other names: Absent. Records in Turkey: (ADY-HA-MA-OS-TUR) Malatya prov. (Heyden, 1888); Hakkari prov.: Habur-Deresi valley (BeytüĢĢebap) (Adlbauer, 1992); Adıyaman prov.: Karadut village (Rejzek & Hoskovec, 1999); SE Anatolia (Sama & Rapuzzi, 2000); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Osmaniye prov.: Osmaniye-Gaziantep road 5th km (Özdikmen et al., 2010). Distribution: Turkey, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from SE Turkey. It is represented by all subspecies in Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).
Coptosia compacta (Ménétriés, 1832) (*WM)
Records in Turkey: (ADY-ANT-BT-EZ-KAR-MA-SI-TO-TUR) Records in Mediterranean Region: Malatya prov. (Heyden, 1888); Turkey (Winkler, 1924-1932; Danilevsky & Miroshnikov, 1985; Lodos, 1998); Tokat prov.: Yakacık (Gökdere), Erzurum prov.: Central / AĢkale (Adlbauer, 1992); Adıyaman prov.: Plateaus under the peak region of Nemrut Mountain (Rejzek & Hoskovec, 1999); Kars prov.: SarıkamıĢ (80 km NE Horasan), Bitlis prov.: Tatvan env., Adıyaman prov.: Nemrut Mountain (Karadut env.)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 76
(Rejzek et al., 2001); Antalya prov.: KaĢ (Kemer village) as C. Sancta (Özdikmen & Hasbenli, 2004); Siirt prov. as C. sancta (Özdikmen et al., 2005). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. According to Sama & Löbl in Löbl & Smetana (2010), C. sancta Reiche, 1877 is a subspecies of C. compacta. So, this species is represented by two subspecies in Turkey as C. compacta compacta Ménétriés, 1832 (Caucasus, NE Turkey, Iran) and C. compacta sancta Reiche, 1877 (Middle East and S Turkey). Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Coptosia schuberti Fuchs, 1965 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-IC-TUR) Ġçel prov.: Namrun as the type loc. (Fuchs, 1965); Ġçel prov.: Namrun (Demelt, 1967); Ġçel prov.: Tarsus (Çamlıyayla), Antalya prov.: Akseki (Yarpuz) (Adlbauer, 1992); Antalya prov.: 5 km N Yarpuz, Ġçel prov.: Namrunkale (Çamlıyayla, Sebil village) (Rejzek et al., 2001). Distribution: Turkey. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from S Turkey. Chorotype: Anatolian.
Pygoptosia Reitter, 1895 [Type sp.: Phytoecia speciosa Frivaldszky, 1884]
Pygoptosia speciosa (Frivaldszky, 1884)
(*EM) Other names: lineolata Reitter; pictipennis Pic. Records in Turkey: (ADY-DI-KA-MR-TUR) Diyarbakır prov. (Frivaldszky, 1884); Anatolia as P. speciosa a. pictipennis Pic, 1909 (Winkler, 1924-1932); KahramanmaraĢ prov. (Breuning et Villiers, 1967); Turkey (Lodos, 1998); Adıyaman prov.: Karadut village, Nemrut Mt. (Rejzek & Hoskovec, 1999); Adıyaman prov.: Nemrut Mt. (Rejzek et al., 2001); Adıyaman prov.: Nemrut Mt., Mardin prov.: Hop pass (Çınaraltı village) (Rejzek et al., 2003). Distribution: Turkey, Syria, Lebanon, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from SE Turkey. It is represented by all subspecies in Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).
Phytoecia Dejean, 1835 [Type sp.: Cerambyx cylindricus Linnaeus, 1758]
Subgenus Pilemia Fairmaire, 1864
[Type sp.: Phytoecia tigrina Mulsant, 1851]
Phytoecia annulata Hampe, 1862 (*EM)
Records in Turkey: (ADY-HT-KAR-OS-SV-TUR) Records in Mediterranean Region: S Turkey: Hatay prov.: Akbez (not Syria) as the type locality of Pilemia wawerkana (Reitter, 1905); Osmaniye prov.: Bahçe (Kızlaç village, Aslanlı Beli) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 77
Phytoecia griseomaculata (Pic, 1891) (*EM)
Other names: laterufonotata Pic; maculifera Holzschuh. Records in Turkey: (AD-EZ-HT-IC-OS-TUR) S Turkey: Hatay prov.: Akbez (not Syria) as the type locality of Pilemia tigrina var. griseomaculata (Pic, 1891); Erzurum prov. and near (Özbek, 1978); Ġçel prov.: Namrun and Adana prov.: Tekir and Nurdağı pass as the type loc. Of P. maculifera (Holzschuh, 1984b); Osmaniye prov.: Nurdağı pass as P. maculifera (Adlbauer, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: Arslanköy as P. maculifera (Rejzek et al., 2001); Osmaniye prov.: Bahçe, (Kızlaç village, Aslanlı Beli) as P. maculifera (Özdikmen et al., 2010). Distribution: Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known mostly from SE Turkey. Chorotype: SW-Asiatic (Syro-Anatolian).
Phytoecia hirsutula (Frölich, 1793) (*WM, EM)
Records in Turkey: (ADY-AF-ANT-BI-BY-EZ-HA-IC-IP-IZ-KAR-KN-OS-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun (Demelt, 1963); Antalya prov. – Demelt, 1961 (Ex. Öymen, 1987); Ġçel prov.: Erdemli (Adlbauer, 1988); Isparta prov.: Yalvaç (Bağkonak, Sultan mountains) (Özdikmen & Hasbenli, 2004); Antalya prov.: Alanya (KeĢbelen plateau), Akseki (Mahmutlu village env., Çukurköy-Mahmutlu), Ġbradı-Akseki road (Özdikmen & Turgut, 2010); Osmaniye prov.: Boğaz plateau, Zorkun road (Çiftmazı) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-E-Mediterranean).
Phytoecia tigrina Mulsant, 1851 (*WM)
Records in Turkey: (BI-IP-IZ-MA-MN-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Yalvaç (Sultan Mts.) (Özdikmen & Hasbenli, 2004). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Turano-European (Turano-Sarmato-Pannonian).
Subgenus Helladia Fairmaire, 1864 [Type sp.: Saperda flavescens Brullé, 1832]
Phytoecia adelpha Ganglbauer, 1886
(*EM) Other names: edessensis Reitter; scapipicta Reitter; urfanensis Reitter; damascena Pic; chtaurensis Heyrovsky; spaceki Heyrovsky; strandi Heyrovsky; adanensis Breuning; nigrofemorata Breuning; atrofemorata Breuning; maraschensis Breuning; cyprica Breuning. Records in Turkey: (AD-GA-HT-IC-MR-OS-TUR) Hatay prov.: Akbez, Ġçel prov. (Pic, 1900a); Hatay prov.: Akbez as Phytoecia edessensis Reitter, 1898 (Aurivillius, 1921); Anatolia as Phytoecia (Helladia) adelpha (Winkler, 1924-1932); Turkey (Breuning, 1951 and 1966; Sama & Löbl in Löbl & Smetana, 2010); Mardin prov.: Sultan (Fuchs et Breuning, 1971 ex Holzschuh, 1980); Gaziantep prov. as P. orbicollis Reiche et Saulcy, 1858 (Demelt, 1963 ex Öymen, 1987); Adana prov.: Misis and Feke as P. orbicollis Reiche et Saulcy, 1858 (Adlbauer, 1992); Turkey (Lodos, 1998); Ġçel prov.: Gülek

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 78
and Namrunkale (Çamlıyayla, Sebil village) (Rejzek et al., 2001). Osmaniye prov.: Hasanbeyli, Hatay prov.: Akbez (Özdikmen et al., 2010). Distribution: Caucasus (Armenia), Turkey, Cyprus, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species probably is distributed in Eastern half of Turkey. According to Sama & Löbl in Löbl & Smetana (2010), it is a subspecies of P. orbicollis. Chorotype: SW-Asiatic (Anatolo-Caucasian + Syro-Anatolian).
Phytoecia alziari Sama, 1992 (*WM, EM)
Other names: Absent. Records in Turkey: (ANT-HT-KA-OS-TUR) Hatay prov.: Arsuz (Fuchs et Breuning, 1971); Hatay prov.: Ġskenderun – Demelt, 1963 (Ex. Öymen, 1987); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); South-East Anatolia as H. millefolii alziari (Sama & Rapuzzi, 2000); South-eastern Mediterranean Region as H. millefolii alziari (Tozlu et al., 2003); KahramanmaraĢ prov.: Nurhak (Alçiçek village) / Ekinözü (AĢağıörten / 1th Source env.) / Göksun (Mehmetbey / Kireçköy / Göksun-Çardak yolu) / Pazarcık (Aksu bridge / Kocalar village) / Çağlayancerit / Andırın-Çokak road (Özdikmen & Okutaner, 2006); Antalya prov.: GündoğmuĢ-Akseki road (Özdikmen & Turgut, 2010); Osmaniye prov.: Bahçe (Kızlaç village, Aslanlı Beli), Hatay prov.: Harbiye, Harbiye-Yayladağı road, Akbez (Özdikmen et al., 2010); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey, Cyprus, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S Turkey. Chorotype: E-Mediterranean (Palaestino-Cypriato-Taurian).
Phytoecia armeniaca Frivaldszky, 1878 (*WM, EM)
Records in Turkey: (ADY-ANT-BT-DI-EZ-KA-KAR-KN-MA-MU-NI-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mts. (Bodemeyer, 1900); KahramanmaraĢ prov.: Ekinözü (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Moreover, it has two subspecies now as nominotypical subspecies and Phytoecia (Helladia) armeniaca testaceovittata (Pic, 1934). It is represented only by the nominotypical subspecies in S and E Turkey. - iranica Holzschuh, 1981, - natali Lobanov, 1994 and - holzschuhi Özdikmen, 2008 (unavailable name) are other names for this species. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Phytoecia demelti (Sama, 2003) (*EM)
Other names: obscuripennis Demelt. Records in Turkey: (IC-KA-TUR) Ġçel prov.: Silifke, KahramanmaraĢ prov., Hatay prov.: Akbez (in Turkey, not Syria) (Demelt, 1963b; Sama, 2003); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from S Turkey. Chorotype: Anatolian.
Phytoecia ferrugata Ganglbauer, 1884 (*WM, EM)
ssp. ferrugata Ganglbauer, 1884

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 79
ssp. dilaticollis Pic, 1900 Other names: houskai Heyrovsky. Records in Turkey: (HT-MR-TUR) Mardin prov.: Sultan as - houskai Heyrovsky (Fuchs & Breuning, 1971); Hatay prov.: Ġskenderun (Sama & Rejzek, 2001); Distribution: Turkey, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S and SE Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).
Phytoecia humeralis (Waltl, 1838) (*WM, EM)
Records in Turkey: (AD-ADY-AK-AM-AN-ANT-AY-BU-DE-DI-ED-ES-HA-HT-IC-IP-IZ-KA-KN-MN-NI-OS-US-TUR) Records in Mediterranean Region: Hatay prov.: YeniĢehir / Antakya (Reyhanlı) (Fuchs et Breuning, 1971); Ġçel prov.: Anamur (Ex. Gül-Zümreoğlu, 1975); Isparta prov.: Kovada (Gül-Zümreoğlu, 1975); Adana prov.: Misis, Hatay prov.: Topboğazı, Osmaniye prov.: FevzipaĢa, Burdur prov.: Bucak, Ġçel prov.: Silifke (Mut) (Adlbauer, 1988); Antalya prov.: GündoğmuĢ / Perge env., Ġçel prov.: Namrunkale (Çamlıyayla, Sebil village) (Rejzek et al., 2001); Adana prov.: Central (Tozlu et al., 2003); Hatay prov.: Ġskenderun (Topboğazı) (Özdikmen et al., 2005); KahramanmaraĢ prov.: Çakalçullu road (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: Göksun (Mehmetbey bridge env. / Küçüksu village, Göcük plateau) / Elbistan (Elbistan-TaĢburun road, TaĢoluk / Karaelbistan, Cumhuriyet district) / Ekinözü / AfĢin (Tanır, YeĢiloba, Karasalgan) / Pazarcık (ġahintepe village / Büyük Nacar / Armutlu village / Sakarkaya village / Evri-Karahöyük road / Evri (Küçükcennetpınarı)) / Central (Kavaklı) (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km), Akseki (Güzelsu village, Mahmutlu village env.), Hatay prov.: Kırıkhan–Belen road (Kıcı), Hassa–Kırıkhan road (10 km to Kırıkhan), Serinyol, Alahan castle, Akbez, Gaziantep prov.: FevzipaĢa–Islahiye road (Özdikmen & Turgut, 2010); Antalya prov.: Akseki-Manavgat road (GündoğmuĢ return 5th km), Akseki (Güzelsu village, Mahmutlu village env.) as P. (H.) humeralis caneri Özdikmen & Turgut, 2010 (Özdikmen & Turgut, 2010); Osmaniye prov.: Yaylalık village-Türkoğlu road, Zorkun road (Çiftmazı), Entry of Yarpuz, Hasanbeyli, Toprakkale, Bahçe (Kızlaç village, Aslanlı Beli), Hatay prov.: Kırıkhan–Belen road (Kıcı), Hassa–Kırıkhan road (10 km to Kırıkhan), Serinyol, Alahan castle, Akbez, Samandağı (Hüseyinli village), Samandağı (Üzengili village), Gaziantep prov.: Akbez (Gülpınarı plateau), FevzipaĢa–Islahiye road as P. (H.) humeralis caneri Özdikmen & Turgut, 2010 (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Moreover, it has three subspecies now as nominotypical subspecies and Phytoecia (Helladia) humeralis insignata Chevrolat, 1854 and Phytoecia (Helladia) humeralis caneri Özdikmen & Turgut, 2010. This species is distributed rather widely in Anatolia for Turkey. It is represented by the nominotypical subspecies and Phytoecia (Helladia) humeralis caneri in Turkey. - scapulata Mulsant, 1851; - scapularis Chevrolat, 1882; - frontalis Chevrolat, 1882; - mersinensis Pic, 1900; - bethaniensis Pic, 1900; - bytinskii Heyrovsky, 1948; - flavoreducta Breuning, 1951 are other names for this species. Chorotype: SW-Asiatic or E-Mediterranean.
Phytoecia praetextata (Steven, 1817) (*EM)
Records in Turkey: (AN-BY-DU-EZ-GU-HT-IC-KS-SV-ZO-TUR) Records in Mediterranean Region: Turkey: Hatay prov.: Akbez as the type loc. of H. praetextata nigricollis Pic, 1891; Ġçel prov. as H. praetextata nigricollis – Demelt, 1963 (Ex. Öymen, 1987); Ġçel prov.: Tarsus (Çamlıyayla) (Adlbauer, 1988); Ġçel prov.: Namrunkale (Çamlıyayla, Sebil village) as H. praetextata nigricollis (Rejzek et al., 2001). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 80
Chorotype: E-Mediterranean (NE-Mediterranean).
Subgenus Musaria Thomson, 1864 [Type sp.: Leptura affinis Harrer, 1784]
Phytoecia anatolica Fuchs & Breuning, 1971
(*EM) Other names: Absent. Records in Turkey: (AD-MU-TUR) Adana prov.: Misis as the type loc. (Fuchs & Breuning, 1971); MuĢ prov.: Buğlan pass (Rejzek et al., 2003); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from S and SE Turkey. Chorotype: Anatolian.
Phytoecia astarte Ganglbauer, 1886 (*EM)
ssp. astarte Ganglbauer, 1886 ssp. perrini Pic, 1892 ssp. lederi Pic, 1899 Other names: Absent. Records in Turkey: (AD-ANT-IC-NI-OS-TUR) Anatolia as the type loc. (Ganglbauer, 1886); Anatolia (Winkler, 1924-1932); Niğde prov.: UlukıĢla (Demelt, 1967); Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Osmaniye prov.: Nurdağı pass, Ġçel prov.: Silifke (Mut) (Adlbauer, 1988); Adana prov.: Nurdağı pass (Rejzek et al., 2001); Antalya prov.: Alanya (Karapınar village) (Özdikmen & Turgut, 2010). Distribution: Caucasus (Georgia, Armenia, Azerbaijan), Turkey, Syria, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S Turkey. It is represented by two subspecies in Turkey as the nominotypical subspecies (in S and SE Turkey) and P. astarte lederi (in NE and E Turkey). Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Phytoecia wachanrui Mulsant, 1851 (*WM, EM)
Records in Turkey: (ANT-BI-GA-KA-KN-MR-OS-TRA-TUR-YO) Osmaniye prov.: Hasanbeyli – Demelt, 1963 (Ex. Öymen, 1987); KahramanmaraĢ prov.: Püren pass (Göksun) (Adlbauer, 1992); Antalya prov.: Alanya-TaĢkent (Özdikmen & Turgut, 2007); Antalya prov.: Alanya-TaĢkent (Exit of Karapınar village) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: SW-Asiatic (Irano-Anatolian) or E-Mediterranean.
Subgenus Neomusaria Plavilstshikov, 1928 [Type sp.: Saperda balcanica Frivaldszky, 1835]
Phytoecia merkli Ganglbauer, 1884
(*EM) Records in Turkey: (AD-ADY-AM-AN-ES-IC-KN-NI-OS-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Namrun as Phytoecia merkli m. alepensis Pic (Demelt, 1967); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Ġçel prov.:

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 81
Gülek – Ganglbauer, 1884; Adana prov. as -inapicalis Pic, 1905 as a subspecies of modesta, it is a synonym of merkli (Sama, 1993); Osmaniye prov.: Hasanbeyli env. (Nurdağı pass), Ġçel prov.: Erdemli (Rejzek et al., 2001); Osmaniye prov.: Zorkun road (Çiftmazı) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: SW-Asiatic.
Phytoecia waltli Sama, 1991 (*EM)
Other names: modesta Waltl. Records in Turkey: (AD-IC-OS-TUR) Adana prov., Southeastern coast of Turkey (Sama, 1993); Ġçel prov.: Arslanköy (Rejzek et al., 2001); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Osmaniye prov.: Haraz plateau (Özdikmen et al., 2010). Distribution: Turkey, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).
Subgenus Phytoecia Dejean, 1835 [Type sp.: Cerambyx cylindricus Linnaeus, 1758]
Phytoecia annulipes Mulsant & Rey, 1863
(*EM) Other names: Absent. Records in Turkey: (AK-AN-OS-TUR) Asia Minor and European Turkey as P. annulipes (Winkler, 1924-1932); Turkey as P. annulipes (Lodos, 1998); Osmaniye prov.: Zorkun, Aksaray prov.: Güzelyurt (Selime), Ankara prov.: Kızılcahamam (Yenimahalle village) as P. icterica annulipes (Özdikmen, 2006); Turkey (Sama & Löbl in Löbl & Smetana, 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species was accepted as a separate species by Sama & Löbl in Löbl & Smetana (2010). Chorotype: Anatolian.
Phytoecia asiatica Pic, 1891 (*EM)
ssp. asiatica Pic, 1891 ssp. sublineata Holzschuh, 1984 Other names: achilleae Holzschuh. Records in Turkey: (HT-MU-TUR) Type loc.: East Turkey (Holzschuh, 1971); Turkey (Danilevsky & Miroshnikov, 1985; Sama & Löbl in Löbl & Smetana, 2010); Hatay prov.: Akbez as the type loc. (Sama, Rapuzzi & Rejzek, 2007); MuĢ prov.: Buğlan pass (Hoskovec & Rejzek, 2010). Distribution: Caucasus (Armenia, Azerbaijan), Turkey, Syria. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known only from S (with nominative subspecies) and E (P. achilleae sublineata Holzschuh, 1984) Turkey. According to Sama & Löbl in Löbl & Smetana (2010), there are two distinct subspecies in the World. In Turkey, it is represented by both subspecies. P. achilleae sublineata Holzschuh, 1984 is distributed in Caucasus and NE, E Turkey. Phytoecia achilleae Holzschuh, 1971 that was evaluated in Part I in the section dealing with the Black Sea Region, is a synonym of the nominotypical subspecies.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 82
Chorotype: SW-Asiatic (Anatolo-Caucasian + Syro-Anatolian).
Phytoecia bangi Pic, 1897 (*EM)
Other names: Absent. Records in Turkey: (IC-KY-MR-NI-OS-TUR) Mardin prov. as the type loc. (ex. Holzschuh, 1975b); Anatolia (Winkler, 1924-1932); Osmaniye prov.: Amanos Mts., Ġçel prov.: Karabucak-Tarsus, Kayseri prov. (Holzschuh, 1975b); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Niğde prov.: Altınhisar (Alıçören) (Özdikmen et al., 2005). Distribution: Turkey, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S and SE Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).
Phytoecia caerulea (Scopoli, 1772) (*WM, EM)
Records in Turkey: (AD-AF-AK-AN-ANT-AY-BI-BO-BU-DE-DU-ES-EZ-GA-HT-IC-IP-IS-IZ-KA-KM-KN-KR-KS-KU-KY-MG-MN-NE-NI-OS-SM-SV-YO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains, Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near as P. caerulea baccueti (Demelt & Alkan, 1962); Osmaniye prov.: Toprakkale (Fuchs et Breuning, 1971); Isparta prov.: Central / Kovada, Burdur prov. as Phytoecia caerulea var. baccueti (Gül-Zümreoğlu, 1975); Burdur prov.: Bucak as P. caerulea baccueti (Adlbauer, 1988); Burdur prov.: Bucak (Çamlık village), Adana prov.: Feke (Musalar village) (Özdikmen & Hasbenli, 2004); Isparta prov.: Gölcük / Central (Özdikmen et al., 2005); Adana prov.: Pozantı (Özdikmen & Demir, 2006); Ġçel prov.: Mut-Karaman road (Gökçeören pine grove) (Özdikmen, 2006); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env.) / AfĢin (Tanır-AfĢin road / Tanır) / Pazarcık (Aksu bridge) (Özdikmen & Okutaner, 2006); Antalya prov.: Güzelbağ-Alanya (Exit of Güzelbağ), Akseki-Manavgat road (GündoğmuĢ return 5th km) as P. caerulea baccueti (Özdikmen & Turgut, 2010); Osmaniye prov.: Sarpınağzı village, Akyar village, Bahçe, Kumarlı-Kazmaca, Kesmeburun village (Castabala (Hierapolis)), Kaypak, Hasanbeyli, Issızca village, Kırıklı village, Hatay prov.: Hassa–Kırıkhan road, Alahan castle, Harbiye, Harbiye-Yayladağı road, Yayladağ, Erzin, Akbez, Gaziantep prov.: Nurdağı (Exit of Ġslahiye), Nurdağı (Exit of Ġslahiye 5th km) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Phytoecia croceipes Reiche & Saulcy, 1858 (*EM)
Records in Turkey: (AD-DI-HT-IZ-KN-NI-OS-TUR) Records in Mediterranean Region: Hatay prov.: Akbez (Fairmaire, 1884); Adana prov.: Pozantı (Bodemeyer, 1900); Osmaniye prov.: Bahçe (Adlbauer, 1988); Hatay prov. (Tozlu et al., 2003). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: SW-Asiatic (Irano-Anatolian + Syro-Anatolian).
Phytoecia cylindrica (Linnaeus, 1758) (*EM)
Records in Turkey: (AN-IS-IZ-KA-KO-KS-KY-NI-TRA-TUR) Records in Mediterranean Region: KahramanmaraĢ prov.: Pazarcık (Kocalar village) / Tekir (Ministry of Forest Tekir Nursery) (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 83
Chorotype: Sibero-European.
Phytoecia geniculata Mulsant, 1862 (*WM, EM)
Records in Turkey: (AD-AN-ANT-AY-BI-BS-BU-DE-ED-GA-HT-IC-IS-IZ-KA-KS-MN-OS-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Antakya (Reyhanlı) (Fuchs et Breuning, 1971); Burdur prov.: Bucak, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Antalya prov.: Perge env. (Rejzek et al., 2001); Ġçel prov.: Central (Tozlu et al., 2003); Adana prov. (Özdikmen et al., 2005); Hatay prov.: Ġskenderun (Güzelyayla road) / Samandağı (Büyükkaya stream, Fidanlı, Uzunbağ), Osmaniye prov.: Düziçi (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env.) / Pazarcık (Evri-Karahöyük road / Evri, Küçükcennetpınarı village / Osmandede village) / Central (Karbasan village env.) / BaĢkonuĢ forest (Özdikmen & Okutaner, 2006); Osmaniye prov.: Kumarlı-Kazmaca, Entry of Yarpuz, Gaziantep prov.: Akbez (Gülpınarı plateau), Hatay prov.: Dörtyol (Kuzuculu) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-Balkan).
Phytoecia icterica (Schaller, 1783) (*EM)
Records in Turkey: (AF-AN-BO-BT-BY-CO-EZ-GA-HT-IS-KA-KAR-KL-KN-KS-KU-OS-YO-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Erzin (Tozlu et al., 2003); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env. / Karbasan village env.) / Pazarcık (Aksu bridge / Kocalar village / Kısık village / Osmandede village) / Central (Türkoğluaraplar village / Kavaklı) / AfĢin (Tanır) / Elbistan-Ekinözü road (Ekinözü env.) (Özdikmen & Okutaner, 2006); Antalya prov.: Akseki-GündoğmuĢ (Özdikmen & Turgut, 2010); Osmaniye prov.: Bahçe, FakıuĢağı village, Kumarlı-Kazmaca, Haraz plateau, Hınzırlı plateau (Kalaycıbatıran district), Bahçe-Ġnderesi road, Kalecik-Hasanbeyli road, Zorkun road (Çiftmazı), Castabala (Hieropolis), Karaçay, Issızca village, Toprakkale, Kırıklı village, Hasanbeyli, Akyar village, Bıçakçı village, Kaypak, Hatay prov.: Harbiye, Kuzuculu, Samandağı (Hüseyinli village, Lahit area (Nekropol)), Akbez, Dörtyol (Kuzuculu), Erzin, Erzin-Dörtyol road, Gaziantep prov.: Nurdağı (Exit of Ġslahiye), Islahiye, Kilis prov.: Hassa–Kilis road (Hisar village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Moreover, According to Sama & Löbl in Löbl & Smetana (2010), P. annulipes is a separate species. Chorotype: European or Turano-European.
Phytoecia manicata Reiche & Saulcy, 1858 (*WM, EM)
Records in Turkey: (AD-DI-HT-IP-IZ-KA-KN-OS-SI-TUR) Records in Mediterranean Region: Adana prov.: Kozan – Demelt, 1961 and 1963 (Ex. Öymen, 1987); Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Isparta prov.: Yalvaç (Özdikmen & Hasbenli, 2004); Hatay prov.: Ġskenderun, entry of Kurtbağı village, Dörtyol (Yahyalı plateau), Osmaniye prov.: Düziçi (Gökçayır village), Bahçe (Buğdaycık village) (Özdikmen & Demirel, 2005); KahramanmaraĢ prov.: KahramanmaraĢ-Andırın road (Körsülü bridge env., Karbasan village env.), Nurhak (Alçiçek village), Göksun (Mehmetbey bridge env.), Pazarcık (Aksu bridge, Kısık village, Sakarkaya village, Evri), Campus of Sütçü Ġmam University, Çağlayancerit, Tekir, AfĢin (Soğucak-Nadir road), Andırın (Kabaklar village), Andırın-Çokak road (Çınar district, Parmaksız plateau), Central, Göksun-Çardak road (Özdikmen & Okutaner, 2006); Adana prov.: Pozantı-Mersin road (Özdikmen, 2006); Osmaniye prov.: Kesmeburun village (Castabala (Hierapolis)), Kalecik-Hasanbeyli road, Karaçay, Kalecik-Hasanbeyli road, Yarpuz road (Forest store env.), Bıçakçı village,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 84
Toprakkale, Issızca village, Kaypak, Akyar village, Hatay prov.: Erzin (Gökdere village), Erzin-kaplıcalar district (Özdikmen et al., 2010). Remarks: It was treated in detail in Part III in the section dealing with the Aegean Region. Chorotype: E-Mediterranean (Palaestino-Taurian) + Turano-Mediterranean (Balkano-Anatolian).
Phytoecia nigricornis (Fabricius, 1782) (*EM)
Records in Turkey: (AD-NI-TRA-TUR) Records in Mediterranean Region: Adana prov.: Pozantı as Phytoecia nigricornis v. solidaginis (Bodemeyer, 1900). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Sibero-European.
Phytoecia pubescens Pic, 1895 (*EM)
Records in Turkey: (AD-AM-AN-DI-EZ-HT-IS-KO-TUR) Records in Mediterranean Region: Adana prov.: Namrun, Nurdağı pass (Danilevsky, 1993); Diyarbakır prov., Erzurum prov., Hatay prov. (Tozlu et al., 2003). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Turano-E-Mediterranean).
Phytoecia pustulata (Schrank, 1776) (*EM)
Records in Turkey: (AM-BI-BO-DU-KA-KN-OS-SM-TRA-TUR) Records in Mediterranean Region: Osmaniye prov. : Nurdağı pass (Adlbauer, 1988); KahramanmaraĢ prov.: Nurhak (Nurhak-Malatya road, Tatlar) (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Phytoecia rufipes (Olivier, 1795) (*EM)
ssp. rufipes Olivier, 1795 ssp. latior Pic, 1895 Other names: umbellatarum Gistel; sibirica Gebler; ludovici Pic; coeca Küster; ledereri Mulsant; femoralis Mulsant. Records in Turkey: (AD-ADY-AK-HT-IC-KA-NI-TUR) Hatay prov.: Akbez as the type loc. of P. rufipes latior (Pic, 1895); Adana prov.: KarataĢ (Breuning & Villiers, 1967); Turkey – Danilevsky & Miroshnikov, 1985 Turkey – Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Ġçel prov.: around Erdemli (Öymen, 1987); Adıyaman prov.: Karadut village (Rejzek & Hoskovec, 1999); Malatya prov.: Erguvan (Özdikmen et al., 2005); KahramanmaraĢ prov.: AfĢin (Tanır-AfĢin road) (Özdikmen & Okutaner, 2006); Aksaray prov.: Hasan Mountain (AĢağı Dikmen), Niğde prov.: Altunhisar (Akçaören), Çamlıyayla, Adana prov.: Pozantı-Mersin road as P. rufipes latior (Özdikmen, 2006). Distribution: Europe (Portugal, Spain, France, Italy, Croatia, Bosnia-Hewrzegovina, Bulgaria, Greece (Crete), Slovenia, Switzerland, Ukraine, European Russia), Siberia, Kazakhstan, Kirgizia, Turkmenistan, Tajikistan, Uzbekistan, Caucasus (Georgia, Armenia), Turkey, Syria, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S and CSE Turkey. There are two distinct subspecies in the World. In Turkey, it is represented by both subspecies. P.rufipes latior is distributed only in S Turkey and Syria.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 85
Chorotype: Sibero-European or Turano-European.
Phytoecia virgula (Charpentier, 1825) (*WM, EM)
Records in Turkey: (ADY-AK-AM-AN-BI-BN-BO-BR-BU-DE-ER-ES-EZ-HT-IP-IS-IZ-KA-KAR-KN-KR-KS-MN-NI-OS-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir (Demelt & Alkan, 1962); Hatay prov.: Ġskenderun (Tuatay et al., 1972); Burdur prov.: Bucak (Adlbauer, 1988); Hatay prov.: Dörtyol (Ġcadiye) / Ġskenderun (Sarımazı) (Tozlu et al., 2003); Isparta prov.: Yalvaç (Sultan Mts.) (Özdikmen & Hasbenli, 2004); KahramanmaraĢ prov.: Göksun (Alparslan TürkeĢ picnic area / Göksun-Kayseri road, Mehmetbey bridge env. / Kireçköy) / Pazarcık (Aksu bridge / Central / Evri-Karahöyük road) (Özdikmen & Okutaner, 2006); Osmaniye prov.: FakıuĢağı village, Kesmeburun village (Castabala (Hierapolis)), Karaçay, Issızca village, Bahçe (Kızlaç village, Aslanlı Beli), Akyar village, Hasanbeyli, Hatay prov.: Alahan castle, Akbez (Mozaik areas), Dörtyol (Kuzuculu) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Subgenus Opsilia Mulsant, 1862 [Type sp.: Opsilia flavicans Mulsant, 1862
= Leptura coerulescens Scopoli, 1763 ]
Phytoecia coerulescens (Scopoli, 1763) (*WM, EM)
Records in Turkey: (AD-ADY-AK-AM-AN-ANT-AR-ART-AY-BO-BS-BU-BY-CN-CO-DE-DI-ER-ES-EZ-GA-GU-IC-IP-IS-IZ-KA-KAR-KIR-KK-KM-KN-KS-KY-MA-MG-MN-NE-NI-OS-SM-SN-SV-TB-YO-ZO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mts. as virescens Fabricius (Bodemeyer, 1900); Adana prov.: Toros Mountains (Pozantı, Bolkar Mts.) (Villiers, 1959); Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains, Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near (Demelt & Alkan, 1962); Adana prov.: Çallıdağı as Opsilia coerulescens grisescens (Breuning et Villiers, 1967); Adana prov. (Tuatay et al., 1972); Isparta prov. (Gül-Zümreoğlu, 1975); Isparta prov. as virescens Fabricius (Gül-Zümreoğlu, 1975); Antalya prov.: Alanya, Ġçel prov.: Tarsus (Çamlıyayla) / Güzeloluk, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Ġçel prov.: Arslanköy (Rejzek et al., 2001); Antalya prov.: Arapsuyu (Tozlu et al., 2003); Isparta prov.: Sütçüler (YeĢildere place) / Eğirdir (Kovada National Park / between Eğirdir and Gelendost) / Yalvaç (Sultan Mts.), Antalya prov.: Alanya (between Çayarası and Cırlasun) / KaĢ (Gömbe, Sinekçibeli), Burdur prov.: Bucak (Çamlık village) (Özdikmen & Hasbenli, 2004); Gaziantep prov.: Islahiye (Yağızlar village, Altınüzüm) / Nurdağı (Belpınar village) (Özdikmen & Demirel, 2005); Adana prov., Isparta prov.: Uluborlu (Özdikmen et al., 2005); Adana prov.: Pozantı-Mersin road, Ġçel prov.: Mut-Karaman road (DeğirmenbaĢı / Gökçeören pine grove) (Özdikmen, 2006); KahramanmaraĢ prov.: Ekinözü (Central /Alpınar village) / AfĢin (Tanır-AfĢin road / Tanır / Emirli, Gerger) / Göksun (Küçüksu village, Göcük plateau / Göksun-Çardak road, Kocaahmet village) / Tekir / Andırın-Çokak road 4. km (Özdikmen & Okutaner, 2006); Antalya prov.: Güzelbağ-Alanya (Exit of Güzelbağ), Alanya (Güzelbağ), Ġbradı-Akseki road, Akseki (Güzelsu village), Ġbradı (BaĢlar village env.) (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Palaearctic.
Subgenus Blepisanis Pascoe, 1866 [Type sp.: Saperda bohemani Pascoe, 1858]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 86
Phytoecia samai Özdikmen & Turgut, 2008 (*EM)
Other names: Absent. Records in Turkey: (OS) Osmaniye prov.: Küllü village (Amanos Mts.) as the type loc. (Özdikmen & Turgut, 2008c); Osmaniye prov.: Küllü village (Amanos Mts.), Zorkun-Karıncalı-Hassa road (Küllü plateau, Amanos Mts.) (Özdikmen et al., 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from the type locality in S Turkey (Amanos Mts.). According to Sama & Löbl in Löbl & Smetana (2010), it is a synonym of P. vittipennis leuthneri. Chorotype: Anatolian.
Phytoecia vittipennis Reiche, 1877 (*WM, EM)
Records in Turkey: (AD-ADY-AN-ANT-BU-DE-ER-EZ-IZ-KA-KN-MN-NI-OS-YO-TUR) Records in Mediterranean Region: Antalya prov. – Demelt, 1961 and 1963 (Ex. Öymen, 1987); Adana prov. (Rejzek et al., 2001); Burdur prov.: YeĢilova (EĢeler Mt.) (Özdikmen & Hasbenli, 2004); Osmaniye prov.: Zorkun plateau road (Ürün plateau), Yarpuz road (KarataĢ place), entry of Yarpuz (Cebel), Çulhalı village and YeĢil village (Hasanbeyli), KahramanmaraĢ prov.: Türkoğlu (Kaledibi village) (Özdikmen & Demirel, 2005); Osmaniye prov.: Zorkun-Karıncalı-Hassa road (Küllü plateau), Küllü village (Özdikmen et al, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean.
Tribe AGAPANTHIINI
Calamobius Guérin-Méneville, 1847 [Type sp.: Saperda gracilis Creutzer, 1799
= Saperda filum Rossi, 1790]
Calamobius filum (Rossi, 1790) (*WM, EM)
Records in Turkey: (AD-AN-ANT-BO-BS-BU-CA-GA-HT-IC-IP-IS-IZ-KA-KL-KN-KO-MG-MN-OS-SA-TRA-TUR) Records in Mediterranean Region: Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Isparta prov.: Eğirdir (Akbelenli-Sütçüler / AĢağı Gökdere), Antalya prov.: Kemer (Olimpos Mt.), Burdur prov.: Bucak (Çamlık village) (Özdikmen & Hasbenli, 2004); Hatay prov.: Ġskenderun (Gediksaray / Güzelyayla road) / Samandağı (Çevlik Rock Cemeteries / Büyükkaya stream (Fidanlı, Uzunbağ)) / Ġskenderun-Belen (Atik plateau) / Kırıkhan (Alabeyli village) / Hassa (Sögüt village road), Gaziantep prov.: Nurdağı (entry of Karaburçlu village) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı-Mersin road, Ġçel prov.: Fındıkpınarı / Silifke-Uzuncaburç road (Özdikmen, 2006); KahramanmaraĢ prov.: Göksun (Saraycık village / Kireçköy) / Püren pass / KahramanmaraĢ-Andırın road (Körsülü bridge env. / Karbasan village env.) / Pazarcık (Kocalar village / Armutlu village) / KahramanmaraĢ-Kavaklı road (entry of Kavaklı / Campus of Sütçü Ġmam University / Central (Türkoğluaraplar village) (Özdikmen & Okutaner, 2006); Antalya prov.: Alanya-TaĢkent (Exit of Karapınar village), Alanya (ġıhlar village plateau, Karapınar village), GündoğmuĢ, Akseki-Manavgat road (GündoğmuĢ return), Alanya (Karapınar village, DikmetaĢ plateau), Ġbradı, Ġbradı-Akseki road (Özdikmen & Turgut, 2010); Osmaniye prov.: Kesmeburun village (Castabala (Hierapolis)), Kalecik-Hasanbeyli road, Karaçay, Bahçe, Zorkun road (Çiftmazı), Akyar village, Bahçe-Ġnderesi road, Yarpuz road (Forest store env.), Osmaniye-Gaziantep road, Kesmeburun village (Castabala castle), Toprakkale, Issızca village, Bahçe (Kızlaç village, Aslanlı Beli), Hatay prov.: Samandağı (Lahit area (Nekropol)),

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 87
Kuzuculu, Akbez, Erzin-kaplıcalar district, Dörtyol-Yeniyurt, Belen, Entry of Belen (Çakallı), Akbez (GüzeluĢağı village), Gaziantep prov.: Akbez (Gülpınarı plateau), FevzipaĢa-Islahiye road, Kilis- Gaziantep road, Kilis prov.: Hassa–Kilis road (Hisar village), Hassa–Kilis road (Gözkaya village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Europeo-Mediterranean.
Agapanthia Audinet-Serville, 1835 [Type sp.: Cerambyx cardui Linnaeus, 1767]
Subgenus Synthapsia Pesarini & Sabbadini, 2004
[Type sp.: Saperda kirbyi Gyllenhal, 1817]
Agapanthia kirbyi (Gyllenhal, 1817) (*WM, EM)
Records in Turkey: (AD-AF-AK-AM-AN-ANT-BI-BS-BT-BU-CO-ED-ER-ES-EZ-IC-IP-IZ-KA-KAR-KIR-KN-KO-KY-MN-NI-OS-SI-TO-VA-TRA-TUR) Records in Mediterranean Region: Adana prov.: Toros Mountains (Pozantı, Bolkar Mts.) (Villiers, 1959); Ġçel prov.: Alata (Breuning et Villiers, 1967); Ġçel prov.: Mersin (Kuzucubelen) (Öymen, 1987); Isparta prov.: Eğirdir, Ġçel prov.: Çamalan, Toros Mountains, Adana prov.: Cilicia, KahramanmaraĢ prov. (Önalp, 1988); Burdur prov.: Sagalassos, Ġçel prov.: Erdemli, Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Adana prov.: Balcalı (Tozlu et al., 2003); Antalya prov.: Kemer (Olimpos Mt.) / Gömbe (Sütleğen / Sinekçibeli pass, Sinekçi village), Burdur prov.: Bucak (Çamlık village), Isparta prov.: Yalvaç (Sultan Mts. / Yarıkkaya village) (Özdikmen & Hasbenli, 2004); Isparta prov. (Özdikmen et al., 2005); Adana prov.: Pozantı (Özdikmen & Demir, 2006); Ġçel prov.: Silifke-Kırobası road, Adana prov.: Pozantı-Mersin road (Özdikmen, 2006); Antalya prov.: GündoğmuĢ, Akseki (Güzelsu village), Ġbradı (Özdikmen & Turgut, 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-European.
Subgenus Epoptes Gistel, 1857 [Type sp.: Lamia asphodeli Latreille, 1804]
Agapanthia asphodeli (Latreille, 1804)
(*WM, EM) Records in Turkey: (AD-AN-ANT-AY-BI-CA-HT-IP-IZ-YO-TUR) Records in Mediterranean Region: Hatay prov.: Ġskenderun (Demelt, 1963); Antalya prov. – Demelt, 1961 (Ex. Öymen, 1987); Hatay prov.: Akbez as A. annularis = A. reyi (Önalp, 1988); Antalya prov.: Alanya, Adana prov.: Pozantı, Hatay prov.: Amanos (Akbez) (Önalp, 1989); Turkey as A. annularis = A. reyi (Lodos, 1998); Isparta prov.: Isparta-Burdur road, Antalya prov.: near Manavgat waterfall and Kemer (Olimpos Mt.), Isparta prov.: Yalvaç (Sultan Mts.) (Özdikmen & Hasbenli, 2004); Isparta prov.: Yalvaç (Sultan Mts.) as A. annularis = A. reyi (Özdikmen & Hasbenli, 2004). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: European.
Agapanthia cynarae (Germar, 1817) (*EM)
Records in Turkey: (AM-BI-BS-CO-ED-EZ-IC-IS-KN-KO-TRA-TUR) Records in Mediterranean Region: Ġçel prov.: Erdemli (Adlbauer, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: E-Mediterranean (NE-Mediterranean) + Turano-European (Ponto-Pannonian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 88
Agapanthia dahli (Richter, 1821) (*EM)
Records in Turkey: (AD-AN-BS-EZ-GA-GU-HT-OS-SI-TUR) Records in Mediterranean Region: Hatay prov.: YeniĢehir – Fuchs et Breuning, 1971 (Ex. Holzschuh, 1980); Adana prov.: Toprakkale / Kozan (Feke), Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Osmaniye prov.: Karagedik village, Kesmeburun village (Castabala (Hierapolis)), KuĢcubeli pass, Gaziantep prov.: Nurdağı (Exit of Ġslahiye 5th km) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European or Turano-European.
Agapanthia detrita Kraatz, 1882 (*EM)
Other names: bucharica Pic. Records in Turkey: (AN-EZ-HT) Ankara prov., Erzurum prov. (Önalp, 1989); Hatay prov.: Ġskenderun (Topboğazı) (Özdikmen et al., 2005). Distribution: Kazakhstan, Kirgizia, Tajikistan, Uzbekistan, Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S, C and E Turkey. Chorotype: Centralasiatic-Mediterranean or Centralasiatic + Anatolian.
Agapanthia kindermanni Pic, 1905 (*EM)
Other names: amicula Holzschuh. Records in Turkey: (AD-IC-OS-TUR) Adana prov.: Tekir, Ġçel prov.: Namrun (Çamlıyayla) as the type loc. of A. amicula (Holzschuh, 1989); Adana prov.: Kozan, Feke (Adlbauer, 1992); Turkey (Sama & Löbl in Löbl & Smetana, 2010); Osmaniye prov.: Kalecik-Hasanbeyli road (Özdikmen et al., 2010). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from S Turkey. Chorotype: Anatolian.
Agapanthia lateralis Ganglbauer, 1884 (*WM, EM)
Records in Turkey: (AF-AG-AK-AM-AN-ANT-BI-BO-CA-CN-CO-ES-GA-IC-IP-IS-IZ-KA-KM-KN-KR-KS-MG-MN-NE-NI-OS-TE-TO-ZO-TRA-TUR) Records in Mediterranean Region: Antalya prov.: Toros Mountains (Bodemeyer, 1900); Ġçel prov.: Toros Mountains (Bolkar Mountains) (Villiers, 1959); Antalya prov.: near Central / Belkıs (Aspendos, Cumali) / Antitoros Mountains (Bey Mountains, Korkuteli) / Alanya and near, Isparta prov.: Eğirdir and near (Demelt & Alkan, 1962); Antalya prov.: Kemer / Patara, Ġçel prov.: Güzeloluk / Erdemli / Silifke (Adlbauer, 1988); Isparta prov.: Eğirdir / Taurus / Central, Antalya prov. (Önalp, 1989); Antalya prov.: Alanya (DemirtaĢ / Mahmutlar) / Kalkan, Isparta prov.: BaĢkonak (Yalvaç road) / Isparta-Burdur road / Eğirdir (AĢağı Gökdere) / Yalvaç (Kuyucak village / Çetince small town / Sultan Mts. / Bağkonak) / Keçiborlu (Yeditepe) (Özdikmen & Hasbenli, 2004); Isparta prov.: Central / Eğirdir (Özdikmen et al., 2005); Antalya prov.: Kemer (Özdikmen & Demir, 2006); Ġçel prov.: Silifke-Kırobası road (Özdikmen, 2006); KahramanmaraĢ prov.: AfĢin (Tanır, YeĢiloba) (Özdikmen & Okutaner, 2006); Antalya prov.: Güzelbağ-Alanya (Exit of Güzelbağ) (Özdikmen & Turgut, 2010); Osmaniye prov.: Kesmeburun village (Castabala castle), Gaziantep prov.: Nurdağı-Islahiye (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 89
Chorotype: Anatolian.
Agapanthia verecunda Chevrolat, 1882 (*EM)
Other names: delagrangei Pic. Records in Turkey: (BT-HT-KN-MG-MR-TUR) Muğla prov.: Marmaris, Konya prov.: AkĢehir, Mardin prov.: Taurus, Hatay prov.: Akbez, Bitlis prov. (Önalp, 1989). Distribution: Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The endemic species has been known only from S and CS Turkey. Chorotype: Anatolian.
Agapanthia villosoviridescens (DeGeer, 1775) (*WM, EM)
Records in Turkey: (AF-AN-AY-BS-DE-ED-EZ-HA-HT-IP-KA-SA-TRA-TUR) Hatay prov.: Akbez as – lineatocollis Mulsant (Fairmaire, 1884); Isparta prov.: Keçiborlu (Gül-Zümreoğlu, 1975); KahramanmaraĢ prov.: Pazarcık (Kısık village) (Özdikmen & Okutaner, 2006). Remarks: It was treated in detail in Part II in the section dealing with the Marmara Region. Chorotype: Sibero-European.
Agapanthia walteri Reitter, 1898 (*EM)
Records in Turkey: (AM-BN-EZ-HA-HT-KAR-MR-TU-TUR) Hatay prov.: YeniĢehir (Fuchs et Breuning, 1971). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Subgenus Homohlephara Pesarini et Sabadini, 2004 [Type sp.: Saperda maculicornis Gyllenhal, 1817]
Agapanthia maculicornis (Gyllenhal, 1817)
(*EM) ssp. maculicornis Gyllenhal, 1817 ssp. davidi Slama, 1986 Other names: Absent. Records in Turkey: (HA-KA-TUR) Hakkari prov.: Yüksekova (Fuchs & Breuning, 1971); Turkey (Önalp, 1989; Lodos, 1998); KahramanmaraĢ prov.: Ekinözü (Türkeli) (Özdikmen & Okutaner, 2006). Distribution: Europe (Italy, Bosnia-Herzegovina, Macedonia, Greece, Romania, Hungary, Slovenia, Moldova, Ukraine, European Russia, European Kazakhstan), W Siberia, Turkey. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S and SE Turkey. There are two distinct subspecies in the World. In Turkey, it is represented only by the nominotypical subspecies. A. maculicornis davidi is distributed only in Italy. Chorotype: Turano-Mediterranean (Turano-Apenninian).
Subgenus Agapanthoplia Pesarini et Sabadini, 2004 [Type sp.: Agapanthia coeruleipennis Frivaldszky, 1878]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 90
Agapanthia coeruleipennis Frivaldszky, 1878 (*WM, EM)
Other names: brevis Pic. Records in Turkey: (ADY-ANT-BN-EZ-HA-IC-IP-KA-KY-MA-MR-MU-TU-TUR) Anatolia as the type loc. (Frivaldszky, 1878); Malatya prov. (Heyden, 1888); Anatolia as A. coeruleipennis brevis Pic, 1891 (Winkler, 1924-1932); East Anatolia (Demelt, 1967); Hakkari prov.: Bajirgi, Yüksekova, ġemdinli, MuĢ prov.: Mountainous area, Bingöl prov.: Central (Fuchs & Breuning, 1971); Erzurum prov. and near (Özbek, 1978); Isparta prov., Antalya prov., Ġçel prov. – Demelt, 1963; KahramanmaraĢ prov., MuĢ prov., Bingöl prov., Hakkari prov. – Fuchs & Breuning, 1971 (Ex. Önalp, 1988); KahramanmaraĢ prov. (Önalp, 1988); Isparta prov.: Eğirdir, Antalya prov., Ġçel prov.: Namrun – Demelt, 1963; KahramanmaraĢ prov., MuĢ prov., Bingöl prov., Hakkari prov.: Yüksekova, ġemdinli – Fuchs & Breuning, 1971 (Ex. Önalp, 1988); KahramanmaraĢ prov.: Püren pass (Göksun), Kayseri prov.: PınarbaĢı (Sarız) (Adlbauer, 1992); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Adıyaman prov.: Nemrut Mt. (Rejzek & Hoskovec, 1999); MuĢ prov.: Buğlan pass, KahramanmaraĢ prov.: Göksun env., Adıyaman prov.: Nemrut Mt., Ġçel prov.: Aslanköy (Rejzek et al., 2001); Tunceli prov. (Tozlu et al., 2003); Mardin prov. (Özdikmen et al., 2005). Distribution: Turkey, Syria, Iran. Remarks: It has been reported from Western and Eastern Parts of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S and E Turkey. Chorotype: SW-Asiatic (Irano-Anatolian + Syro-Anatolian).
Subgenus Agapanthia Audinet-Serville, 1835 [Type sp.: Cerambyx cardui Linnaeus, 1767]
Agapanthia suturalis (Fabricius, 1787)
(*WM, EM) Records in Turkey: (AD-AN-ANT-ART-AY-BI-BN-BS-BU-BY-CA-CN-DE-DI-ED-EL-ER-ES-EZ-GA-GU-HT-IC-IS-IZ-KA-KAR-KIR-KK-KL-KN-KO-KS-MG-MN-OS-RI-SI-SV-TRA-TUR) Records in Mediterranean Region: Hatay prov.: Arsuz (YeniĢehir) / Antakya (Reyhanlı), Osmaniye prov.: Toprakkale, Adana prov.: Misis (Fuchs et Breuning, 1971); Osmaniye prov., Hatay prov.: Antakya (Gül-Zümreoğlu, 1975); Adana prov.: Misis, Ġçel prov.: Erdemli (Kızkalesi), Antalya prov.: Manavgat (ġelale), Burdur prov.: Bucak (Adlbauer, 1988); Ġçel prov.: Mut, Adana prov.: Cilicia (Önalp, 1989); Adana prov., Hatay prov.: Antakya (Lodos, 1998); Adana prov.: Balcalı / Ceyhan, Antalya prov.: Kumluca, Artvin prov.: Central (Ormanlı) / Ardanuç (Akarsu) / ġavĢat (Çayağzı), Bayburt prov.: Maden, Bilecik prov.: Central, Bingöl prov.: Solhan (Buğlan pass), Çanakkale prov.: Central, Diyarbakır prov.: Silvan, Erzincan prov.: Central (Bahçe) / Üzümlü / Bayırbağ, Erzurum prov.: AĢkale (Kop Mt.) / Ilıca (Eğerti) / Ġspir (MadenköprübaĢı) / Narman (Beyler) / Oltu / Karakaban / Çamlıbel / Pasinler / Çalıyazı / Pazarroad / Kartal Plateau / ġenkaya (Ġçmesuyu) / Ormanlı, Hatay prov.: Erzin / Ġskenderun (Sarımazı), Rize prov.: ÇamlıhemĢin (Ayder), Sivas prov.: Central / TürkeĢlik / Ümranlı (Kızıldağ) as A. suturalis (Fabricius, 1787) (Tozlu et al., 2003); Antalya prov.: Isparta road (Özdikmen & Hasbenli, 2004); Adana prov., Hatay prov.: Ġskenderun (Central / Esentepe), Osmaniye prov. (Özdikmen et al., 2005); Hatay prov.: Ġskenderun (Güzelyayla road / entry of Kurtbağı village) / Samandağı (Büyükkaya stream, Fidanlı, Uzunbağ) / Ġskenderun-Belen (Atik plateau) / Kırıkhan (Alabeyli village) / Hassa (Akbez / Zeytinoba village) / Belen (Müftüler village), Osmaniye prov.: entry of Nohutköy / Düziçi / Zorkun plateau road (Ürün plateau) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı (Özdikmen & Demir, 2006); Adana prov.: Pozantı-Mersin road (Özdikmen, 2006); KahramanmaraĢ prov.: AfĢin (Çardak-AfĢin road) / Pazarcık (Aksu bridge / ġahintepe village / Armutlu village) / KahramanmaraĢ-Kavaklı road (entry of Kavaklı) / Türkoğlu (Kılılı) / Göksun (KamıĢcık village) / Central (Özdikmen & Okutaner, 2006); Antalya prov.: Ġbradı-Akseki road, Ġbradı (Özdikmen & Turgut, 2010); Osmaniye prov.: Kalecik-Hasanbeyli road, Zorkun road (Çiftmazı), Karaçay, Kesmeburun

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 91
village (Castabala (Hierapolis)), Bahçe, Osmaniye-Gaziantep road, Bıçakçı village, Kesmeburun village (Castabala castle), Hasanbeyli, Hatay prov.: Samandağı (Lahit area (Nekropol)), Kuzuculu, Akbez, Kuzuculu, Belen, Entry of Belen (Çakallı), Erzin-kaplıcalar district, Gaziantep prov.: Kilis-Gaziantep road (Oğuzeli return), FevzipaĢa-Islahiye road, Kilis prov.: Hassa–Kilis road (Deliosmanlı village) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region as A. cardui. Chorotype: Mediterranean.
Subgenus Smaragdula Pesarini & Sabbadini, 2004 [Type sp.: Saperda violacea Fabricius, 1775]
Agapanthia amitina Holzschuh, 1989
(*EM) Other names: Absent. Records in Turkey: (IC-OS) Osmaniye prov.: Zorkun plateau, Ġçel prov.: Tarsus (Çamlıyayla) (Adlbauer, 1992). Distribution: Turkey, Iran. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S Turkey. Chorotype: SW-Asiatic (Irano-Anatolian).
Agapanthia chalybaea Faldermann, 1837 (*WM, EM)
Records in Turkey: (BI-IP-IS-KA-KN-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir, KahramanmaraĢ prov., Taurus (South Turkey) (Önalp, 1988). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Agapanthia frivaldszkyi Ganglbauer, 1884 (*WM, EM)
Records in Turkey: (AM-AN-BI-?DE-IC-IP-IS-MU-NI-SA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir (Demelt & Alkan, 1962); Isparta prov., Ġçel prov.: Namrun – Demelt, 1963 (Ex. Öymen, 1987). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Agapanthia lais Reiche & Saulcy, 1858 (*EM)
Other names: Absent. Records in Turkey: (HT-KA-MR-OS-TUR) Osmaniye prov.: Nurdağı pass (Adlbauer, 1988); Mardin prov., KahramanmaraĢ prov.: Ahır Mt. (Önalp, 1988); Turkey (Lodos, 1998; Sama & Löbl in Löbl & Smetana, 2010); Hatay prov.: Akbez (GüzeluĢağı village) (Özdikmen et al., 2010). Distribution: Turkey, Syria, Israel, Lebanon, Jordan. Remarks: It has been reported only from Eastern Part of Mediterranean Region as connected with the geological area covered by the present work (*).The species has been known from S and SE Turkey. Chorotype: E-Mediterranean (Palaestino-Taurian).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 92
Agapanthia osmanlis Reiche & Saulcy, 1858 (*EM)
Records in Turkey: (ART-BI-BY-ER-EZ-GU-HT-IS-KAR-SM-SV-TUR) Records in Mediterranean Region: Hatay prov.: Erzin (Tozlu et al., 2003). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
Agapanthia violacea (Fabricius, 1775) (*WM, EM)
Records in Turkey: (AD-AF-AK-AN-BI-BO-BS-CO-DE-DU-ED-EZ-GA-HT-IC-IP-IS-IZ-KA-KIR-KK-KN-KO-KR-KS-KY-MG-MN-NE-NI-OS-SA-ZO-TRA-TUR) Records in Mediterranean Region: Isparta prov.: Eğirdir, Adana prov. (Önalp, 1988); Isparta prov.: Yalvaç (Eleği village / Sultan Mts.) (Özdikmen & Hasbenli, 2004); Isparta prov.: Gölcük (Özdikmen et al., 2005); Hatay prov.: Ġskenderun (Belen, Atik plateau) (Özdikmen & Demirel, 2005); Adana prov.: Pozantı (Özdikmen & Demir, 2006); Adana prov.: Pozantı (entry of Fındıklı), Ġçel prov.: Uzuncaburç road / Mut-Karaman road (DeğirmenbaĢı) (Özdikmen, 2006); KahramanmaraĢ prov.: Pazarcık (Armutlu village, Kartalkaya dam env.) (Özdikmen & Okutaner, 2006); Osmaniye prov.: Entry of Yarpuz, Haraz plateau, Hatay prov.: Akbez, Hassa–Kırıkhan road, Gaziantep prov.: Kilis-Gaziantep road (Oğuzeli return) (Özdikmen et al., 2010). Remarks: It was treated in detail in Part I in the section dealing with the Black Sea Region. Chorotype: Sibero-European.
ZOOGEOGRAPHICAL REMARKS
The present zoogeographical characterization is based on the chorotype classification of Anatolian fauna, recently proposed by Vigna Taglianti et al. (1999). In this part, the taxons which have same chorotype(s) are arranged into a single paragraph. Chorotypes are given alphabetically.
The species that has the Afrotropico-Indo-Mediterranean + Neotropic + Oriental chorotype is Batocera rufomaculata (DeGeer, 1775).
The species that has the Centralasiatic-Mediterranean or Centralasiatic + Anatolian chorotype is Agapanthia detrita Kraatz, 1882.
The species that have the Cosmopolitan chorotype are Phoracantha recurva Newman, 1840 and Phoracantha semipunctata (Fabricius, 1775).
The species that have the E-Mediterranean chorotype are Callergates gaillardoti (Chevrolat, 1854); Prionus komiyai Lorenc, 1999; Grammoptera baudii (Stephens, 1831); Stictoleptura heydeni (Ganglbauer, 1889); Anastrangalia montana (Mulsant & Rey, 1863); Trichoferus kotschyi (Ganglbauer, 1883); Purpuricenus dalmatinus Sturm, 1843; Purpuricenus interscapillatus Plavilstshikov, 1937; Delagrangeus angustissimus Pic, 1892; Stenopterus atricornis Pic, 1891; Stenopterus flavicornis Küster, 1846; Procallimus egregius (Mulsant & Rey, 1863); Brachypteroma holtzi Pic, 1905; Leioderes tuerki Ganglbauer, 1885; Chlorophorus gratiosus Marseul, 1868 ; Clytus madoni Pic, 1891; Clytus taurusiensis (Pic, 1903); Dorcadion cinerarium (Fabricius, 1787); Anaesthetis anatolica Holzschuh, 1969; Pogonocerus anatolicus Mulsant, 1839; Leiopus syriacus (Ganglbauer, 1884); Saperda quercus Charpentier, 1825; Coptosia ganglbaueri Pic, 1936; Phytoecia alziari Sama, 1992; Phytoecia ferrugata Ganglbauer, 1884; Phytoecia praetextata (Steven, 1817); Phytoecia waltli Sama, 1991; Phytoecia manicata Reiche & Saulcy, 1858; Agapanthia cynarae (Germar, 1817) and Agapanthia lais Reiche & Saulcy, 1858.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 93
The species that have the European chorotype are Rhagium bifasciatum Fabricius, 1775; Stictoleptura fulva (DeGeer, 1775); Pedostrangalia revestita (Linnaeus, 1767); Etorufus pubescens (Fabricius, 1787); Rutpela maculata (Poda von Neuhaus, 1761); Cerambyx scopolii Fuessly, 1775; Ropalopus clavipes (Fabricius, 1775); Plagionotus detritus (Linnaeus, 1758); Clytus rhamni Germar, 1817; Mesosa curculionoides (Linnaeus, 1760); Leiopus nebulosus (Linnaeus, 1758); Exocentrus adspersus Mulsant, 1846; Exocentrus punctipennis Mulsant & Guillebeau, 1856; Oberea linearis (Linnaeus, 1760); Phytoecia icterica (Schaller, 1783) and Agapanthia asphodeli (Latreille, 1804).
The species that has the European + Europeo-Mediterranean chorotype is Anoplodera sexguttata (Fabricius, 1775).
The species that has the S, C and E-European chorotype is Glaphyra marmottani Brisout de Barneville, 1863.
The species that have the Holarctic chorotype are Rhagium inquisitor (Linnaeus, 1758); Arhopalus rusticus (Linnaeus, 1758); Phymatodes testaceus (Linnaeus, 1758) and Saperda populnea (Linnaeus, 1758).
The species that have the Mediterranean chorotype are Oxypleurus nodieri Mulsant, 1839; Alocerus moesiacus (Frivaldsky, 1837); Arhopalus syriacus (Reitter, 1895); Icosium tomentosum Lucas, 1854; Trichoferus griseus (Fabricius, 1792); Chlorophorus trifasciatus (Fabricius, 1781); Niphona picticornis Mulsant, 1839; Pogonocerus perroudi Mulsant, 1839 and Agapanthia suturalis (Fabricius, 1787).
The species that have the Palaearctic chorotype are Stictoleptura rubra (Linnaeus, 1758); Arhopalus ferus (Mulsant, 1839); Spondylis buprestoides (Linnaeus, 1758); Aromia moschata (Linnaeus, 1758); Chlorophorus varius (Müller, 1766); Aegomorphus clavipes (Schrank, 1781); Tetrops praeustus (Linnaeus, 1758); Oberea oculata (Linnaeus, 1758); Oberea erythrocephala (Schrank, 1776) and Phytoecia coerulescens (Scopoli, 1763).
The species that have the S-European chorotype are Cortodera humeralis (Schaller, 1783); Cerambyx miles Bonelli, 1812 and Cerambyx welensii (Küster, 1846).
The species that has the SE-European chorotype is Stenhomalus bicolor (Kraatz, 1862).
The species that have the Sibero-European chorotype are Dinoptera collaris (Linnaeus, 1758); Alosterna tabacicolor (De Geer, 1775); Pseudovadonia livida (Fabricius, 1777); Anoplodera rufipes (Schaller, 1783); Pachytodes erraticus (Dalman, 1817); Stenurella bifasciata (Müller, 1776); Stenurella melanura (Linnaeus, 1758); Obrium cantharinum (Linnaeus, 1767); Molorchus minor (Linnaeus, 1758); Plagionus floralis (Pallas, 1773); Chlorophorus figuratus (Scopoli, 1763); Monochamus galloprovincialis (Olivier, 1795); Acanthocinus aedilis (Linnaeus, 1758); Acanthocinus griseus (Fabricius, 1792); Phytoecia cylindrica (Linnaeus, 1758); Phytoecia nigricornis (Fabricius, 1782); Phytoecia rufipes (Olivier, 1795); Agapanthia dahli (Richter, 1821); Agapanthia villosoviridescens (DeGeer, 1775) and Agapanthia violacea (Fabricius, 1775).
The species that has the Sibero-European + Turano-Europeo-Mediterranean chorotype is Prionus coriarius (Linnaeus, 1758).
The species that have the Subcosmopolitan chorotype are Stromatium unicolor (Olivier, 1795); Nathrius brevipennis (Mulsant, 1839); Hylotrupes bajulus (Linnaeus, 1758) and Poecilium lividum (Rossi, 1794).
The species that have the SW-Asiatic chorotype are Rhagium syriacum Pic, 1892; Anisorus heterocerus (Ganglbauer, 1882); Cortodera alpina (Ménétriés, 1832); Cortodera colchica Reitter, 1890; Cortodera syriaca Pic, 1901; Vadonia

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 94
bitlisiensis Chevrolat, 1882; Stictoleptura excisipes (Daniel & Daniel, 1891); Stictoleptura pallidipennis (Tournier, 1872); Stictoleptura sambucicola (Holzschuh, 1982); Stictoleptura tonsa (Daniel & Daniel, 1891); Stictoleptura tripartita (Heyden, 1889); Pedostrangalia emmipoda (Mulsant, 1863); Trichoferus preissi (Heyden, 1894); Purpuricenus wachanrui Levrat, 1858; Molorchus juglandis Sama, 1982; Callidium syriacum (Pic, 1892); Chlorophorus dinae (Schneider, 1787); Chlorophorus nivipictus (Kraatz, 1879); Clytus ciliciensis (Chevrolat, 1863); Dorcadion halepense (Kraatz, 1873); Dorcadion scabricolle Dalman, 1817; Dorcadion variegatum Ganglbauer, 1884; Exocentrus ritae Sama, 1985; Oxylia argentata (Ménétriés, 1832); Mallosia mirabilis (Faldermann, 1837); Mallosia imperatrix Abeille de Perrin, 1885 ; Pygoptosia speciosa (Frivaldszky, 1884); Coptosia compacta (Ménétriés, 1832); Phytoecia annulata Hampe, 1862; Phytoecia griseomaculata (Pic, 1891); Phytoecia adelpha Ganglbauer, 1886; Phytoecia armeniaca Frivaldszky, 1878; Phytoecia humeralis (Waltl, 1838); Phytoecia astarte Ganglbauer, 1886; Phytoecia wachanrui Mulsant, 1851; Phytoecia merkli Ganglbauer, 1884; Phytoecia asiatica Pic, 1891; Phytoecia bangi Pic, 1897; Phytoecia croceipes Reiche & Saulcy, 1858; Agapanthia walteri Reitter, 1898; Agapanthia coeruleipennis Frivaldszky, 1878; Agapanthia amitina Holzschuh, 1989 and Agapanthia chalybaea Faldermann, 1837.
The species that has the SW-Asiatic + Turanian (Ponto-Caspian) chorotype is Stenurella jaegeri (Hummel, 1825).
The species that have the Turano-European chorotype are Aegosoma scabricorne (Scopoli, 1763); Vadonia unipunctata (Fabricius, 1787); Stictoleptura cordigera (Füsslins, 1775); Stictoleptura tesserula (Charpentier, 1825); Stenopterus rufus (Linnaeus, 1767); Phymatodes rufipes (Fabricius, 1777); Plagionotus bobelayei (Brullé, 1832); Chlorophorus hungaricus Seidlitz, 1891; Chlorophorus sartor (Müller, 1766); Morimus funereus (Mulsant, 1862); Phytoecia tigrina Mulsant, 1851; Phytoecia caerulea (Scopoli, 1772); Phytoecia pustulata (Schrank, 1776); Phytoecia virgula (Charpentier, 1825) and Agapanthia kirbyi (Gyllenhal, 1817).
The species that has the Turano-European (Turano-Sarmato-Pannonian) + Turano-Mediterranean (Turano-Apenninian + Turano-E-Mediterranean)SW-Asiatic + Turanian (Ponto-Caspian) chorotype is Semanotus russicus (Fabricius, 1777).
The species that have the Turano-Europeo-Mediterranean chorotype are Ergates faber (Linnaeus, 1760); Prinobius myardi Mulsant, 1842; Anastrangalia dubia (Scopoli, 1763); Hesperophanes sericeus (Fabricius, 1787); Cerambyx cerdo Linnaeus, 1758; Purpuricenus budensis (Götz, 1783); Penichroa fasciata (Stephens, 1831); Certallum ebulinum (Linnaeus, 1767); Deilus fugax (Olivier, 1790); Callimus angulatus (Schrank, 1789) and Calamobius filum (Rossi, 1790).
The species that have the Turano-Mediterranean chorotype are Rhaesus serricollis (Motschulsky, 1838); Mesoprionus besikanus (Fairmaire, 1855); Cortodera differens Pic, 1898; Cortodera discolor Fairmaire, 1866; Cortodera flavimana (Waltl, 1838); Cortodera pseudomophlus Reitter, 1889; Vadonia bisignata (Brullé, 1832); Vadonia moesiaca (Daniel & Daniel, 1891); Vadonia monostigma (Ganglbauer, 1882); Stictoleptura pallens (Brullé, 1832); Stictoleptura rufa (Brullé, 1832); Pedostrangalia verticalis (Germar, 1822); Trichoferus fasciculatus (Faldermann, 1837); Cerambyx dux (Faldermann, 1837); Purpuricenus desfontainei (Fabricius, 1792); Axinopalpis gracilis (Krynicki, 1832); Lampropterus femoratus (Germar, 1824); Brachypteroma ottomanum Heyden, 1863; Chlorophorus aegyptiacus (Fabricius, 1775); Morimus

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 95
ganglbaueri Reitter, 1894; Dorcadion divisum Germar, 1839; Dorcadion septemlineatum Waltl, 1838; Oxylia duponcheli (Brullé, 1832); Coptosia bithynensis (Ganglbauer, 1884); Phytoecia hirsutula (Frölich, 1793); Phytoecia geniculata Mulsant, 1862; Phytoecia pubescens Pic, 1895; Phytoecia vittipennis Reiche, 1877; Agapanthia maculicornis (Gyllenhal, 1817); Agapanthia frivaldszkyi Ganglbauer, 1884 and Agapanthia osmanlis Reiche & Saulcy, 1858.
The species that have the Turano-Mediterranean + Turano-European chorotype are Cerambyx nodulosus Germar, 1817 and Glaphyra kiesenwetteri (Mulsant & Rey, 1861).
The species that have the W-Palaearctic chorotype are Poecilium alni (Linnaeus, 1767); Plagionotus arcuatus (Linnaeus, 1758); Xylotrechus arvicola (Olivier, 1795) and Saperda punctata (Linnaeus, 1767).
On the other hand, following 68 taxa are endemic to Turkey: The species that have the Anatolian chorotype are Rhagium elmaliense
Schmid, 1999; Rhagium phrygium Daniel, 1906; Stenocorus auricomus (Reitter, 1890); Cortodera cirsii Holzschuh, 1975; Cortodera imrasanica Sama & Rapuzzi, 1999; Cortodera omophloides Holzschuh, 1975; Cortodera rubripennis Pic, 1891; Grammoptera merkli Frivaldszky, 1884; Alosterna anatolica Adlbauer, 1992; Vadonia ciliciensis Daniel & Daniel, 1891; Vadonia danielorum Holzschuh, 1984; Vadonia frater Holzschuh, 1981; Vadonia soror Holzschuh, 1981; Stictoleptura gevneensis Özdikmen & Turgut, 2008; Carlandrea syriaca (Pic, 1891); Stenurella pamphyliae Rapuzzi & Sama, 2009; Necydalis hadullai Szallies, 1994; Apatophysis kadleci Danilevsky, 2008; Trichoferus sbordonii Sama, 1982; Purpuricenus nigronotatus Pic, 1907; Anatolobrium eggeri Adlbauer, 2004; Stenopterus kraatzi Pic, 1892; Callimus akbesianus Pic, 1892; Glaphyra tenuitarsis (Holzschuh, 1981); Phymatodes magnanii (Sama & Rapuzzi, 1999); Isotomus syriacus (Schneider, 1787); Pseudosphegesthes longitarsus Holzschuh, 1974; Clytus gulekanus Pic, 1904; Parmena lukati Sama, 1994; Parmena mutilloides Sabbadini & Pesarini, 1992; Dorcadion amanense Breuning, 1943; Dorcadion anatolicum Pic, 1900; Dorcadion arcivagum Thomson, 1867; Dorcadion carinipenne Pic, 1900; Dorcadion culminicola Thomson, 1868; Dorcadion delagrangei Pic, 1894; Dorcadion deyrollei Ganglbauer, 1884; Dorcadion drusoides Breuning, 1962; Dorcadion holtzi (Pic, 1905); Dorcadion infernale Mulsant & Rey, 1863; Dorcadion ispartense Breuning, 1962; Dorcadion kraetschmeri Bernhauer, 1988; Dorcadion lohsei Braun, 1976; Dorcadion menradi Holzschuh, 1989; Dorcadion micans Thomson, 1867; Dorcadion miminfernale Breuning, 1970; Dorcadion oezdurali Önalp, 1988; Dorcadion ortrudae Braun, 1980; Dorcadion paracinerarium Breuning, 1974; Dorcadion petrovitzi Heyrovsky, 1964; Dorcadion pseudinfernale Breuning, 1943; Dorcadion saulcyi Thomson, 1865; Dorcadion semibrunneum Pic, 1903; Dorcadion sinuatevittatum Pic, 1937; Dorcadion smyrnense (Linneaus, 1757); Dorcadion subinterruptum Pic, 1900; Dorcadion syriense Breuning, 1943; Dorcadion triste Frivaldsky, 1845; Tetrops warnckei Holzschuh, 1977; Semiangusta delagrangei (Pic, 1891); Coptosia schuberti Fuchs, 1965; Phytoecia demelti (Sama, 2003); Phytoecia anatolica Fuchs & Breuning, 1971; Phytoecia annulipes Mulsant & Rey, 1863; Phytoecia samai Özdikmen & Turgut, 2008; Agapanthia kindermanni Pic, 1905; Agapanthia lateralis Ganglbauer, 1884 and Agapanthia verecunda Chevrolat, 1882.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 96
CONCLUSIONS
Apparently, Turkey has continental properties. Turkey is the origin of many taxons. It is a refigium for effected living creatures from geological and climatical changes that have more biological importance than any land in the world. As seen in the whole world, incredible variations have also been seen among the insects which are the most influenced living creatures from these changes occurring in the past in Turkey. As a result of this, Turkey has a rich biodiversity. Resulting from this biodiversity faunistical richness can be virtually accepted to the same degree as Continental Europe. As a result of this study, totally 280 species of 92 genera of 8 subfamilies are also determined in the whole territories of the Mediterranean Region of Turkey [Antalya Part = Western Mediterranean Ragion = WM (including Burdur, Isparta and Antalya provinces), Adana Part = Eastern Mediterranean Region = EM (including Ġçel, Adana, Osmaniye, Hatay, KahramanmaraĢ and Kilis provinces)]. A simple faunistical list of determined species is given below.
A LIST OF LONGICORN BEETLES FROM MEDITERRANEAN REGION IN TURKEY
Subfamily PRIONINAE
1. Ergates faber (Linnaeus, 1760) (*WM, EM) 2. Callergates gaillardoti (Chevrolat, 1854) (*WM, EM) 3. Prinobius myardi Mulsant, 1842 (*WM, EM) 4. Rhaesus serricollis (Motschulsky, 1838) (*WM, EM) 5. Aegosoma scabricorne (Scopoli, 1763) (*WM, EM) 6. Prionus coriarius (Linnaeus, 1758) (*WM, EM) 7. Prionus komiyai Lorenc, 1999 (*EM) 8. Mesoprionus besikanus (Fairmaire, 1855) (*WM, EM)
Subfamily LEPTURINAE
1. Rhagium bifasciatum Fabricius, 1775 (*WM) 2. Rhagium elmaliense Schmid, 1999 (*WM) 3. Rhagium phrygium Daniel, 1906 (*EM) 4. Rhagium syriacum Pic, 1892 (*EM) 5. Rhagium inquisitor (Linnaeus, 1758) (*WM) 6. Stenocorus auricomus (Reitter, 1890) (*EM) 7. Anisorus heterocerus (Ganglbauer, 1882) (*WM, EM) 8. Dinoptera collaris (Linnaeus, 1758) (*WM, EM) 9. Cortodera alpina (Ménétriés, 1832) (*WM, EM) 10. Cortodera cirsii Holzschuh, 1975 (*WM, EM) 11. Cortodera colchica Reitter, 1890 (*WM, EM) 12. Cortodera differens Pic, 1898 (*WM) 13. Cortodera discolor Fairmaire, 1866 (*WM, EM) 14. Cortodera flavimana (Waltl, 1838) (*WM, EM) 15. Cortodera humeralis (Schaller, 1783) (*WM, EM) 16. Cortodera imrasanica Sama & Rapuzzi, 1999 (*WM) 17. Cortodera omophloides Holzschuh, 1975 (*EM) 18. Cortodera pseudomophlus Reitter, 1889 (*EM) 19. Cortodera rubripennis Pic, 1891 (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 97
20. Cortodera syriaca Pic, 1901 (*EM) 21. Grammoptera baudii (Stephens, 1831) (*EM) 22. Grammoptera merkli Frivaldszky, 1884 (*WM, EM) 23. Alosterna anatolica Adlbauer, 1992 (*WM) 24. Alosterna tabacicolor (De Geer, 1775) (*WM) 25. Vadonia bisignata (Brullé, 1832) (*WM) 26. Vadonia bitlisiensis Chevrolat, 1882 (*WM) 27. Vadonia ciliciensis Daniel & Daniel, 1891 (*WM) 28. Vadonia danielorum Holzschuh, 1984 (*WM) 29. Vadonia frater Holzschuh, 1981 (*EM) 30. Vadonia moesiaca (Daniel & Daniel, 1891) (*WM) 31. Vadonia monostigma (Ganglbauer, 1882) (*WM) 32. Vadonia soror Holzschuh, 1981 (*WM, EM) 33. Vadonia unipunctata (Fabricius, 1787) (*WM, EM) 34. Pseudovadonia livida (Fabricius, 1777) (*WM, EM) 35. Anoplodera rufipes (Schaller, 1783) (*EM) 36. Anoplodera sexguttata (Fabricius, 1775) (*EM) 37. Stictoleptura rubra (Linnaeus, 1758) (*WM) 38. Stictoleptura cordigera (Füsslins, 1775) (*WM, EM) 39. Stictoleptura excisipes (Daniel & Daniel, 1891) (*WM, EM) 40. Stictoleptura fulva (DeGeer, 1775) (*WM, EM) 41. Stictoleptura gevneensis Özdikmen & Turgut, 2008 (*WM) 42. Stictoleptura heydeni (Ganglbauer, 1889) (*EM) 43. Stictoleptura pallens (Brullé, 1832) (*EM) 44. Stictoleptura pallidipennis (Tournier, 1872) (*?WM) 45. Stictoleptura rufa (Brullé, 1832) (*WM, EM) 46. Stictoleptura sambucicola (Holzschuh, 1982) (*EM) 47. Stictoleptura tesserula (Charpentier, 1825) (*EM) 48. Stictoleptura tonsa (Daniel & Daniel, 1891) (*EM) 49. Stictoleptura tripartita (Heyden, 1889) (*EM) 50. Anastrangalia dubia (Scopoli, 1763) (*EM) 51. Anastrangalia montana (Mulsant & Rey, 1863) (*WM, EM) 52. Pedostrangalia revestita (Linnaeus, 1767) (*WM) 53. Pedostrangalia emmipoda (Mulsant, 1863) (*WM, EM) 54. Pedostrangalia verticalis (Germar, 1822) (*WM) 55. Etorufus pubescens (Fabricius, 1787) (*WM) 56. Carlandrea syriaca (Pic, 1891) (*WM, EM) 57. Pachytodes erraticus (Dalman, 1817) (*WM, EM) 58. Rutpela maculata (Poda von Neuhaus, 1761) (*WM, EM) 59. Stenurella bifasciata (Müller, 1776) (*WM, EM) 60. Stenurella jaegeri (Hummel, 1825) (*EM) 61. Stenurella melanura (Linnaeus, 1758) (*WM) 62. Stenurella pamphyliae Rapuzzi & Sama, 2009 (*WM)
Subfamily NECYDALINAE
1. Necydalis hadullai Szallies, 1994 (*WM)
Subfamily ASEMINAE
1. Oxypleurus nodieri Mulsant, 1839 (*WM) 2. Alocerus moesiacus (Frivaldsky, 1837) (*WM) 3. Arhopalus ferus (Mulsant, 1839) (*WM, EM) 4. Arhopalus rusticus (Linnaeus, 1758) (*WM, EM) 5. Arhopalus syriacus (Reitter, 1895) (*WM, EM)
Subfamily SPONDYLIDINAE
1. Spondylis buprestoides (Linnaeus, 1758) (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 98
Subfamily DORCASOMINAE
1. Apatophysis kadleci Danilevsky, 2008 (*EM)
Subfamily CERAMBYCINAE
1. Icosium tomentosum Lucas, 1854 (*WM) 2. Hesperophanes sericeus (Fabricius, 1787) (*WM) 3. Trichoferus fasciculatus (Faldermann, 1837) (*WM) 4. Trichoferus griseus (Fabricius, 1792) (*WM, EM) 5. Trichoferus kotschyi (Ganglbauer, 1883) (*WM, EM) 6. Trichoferus preissi (Heyden, 1894) (*WM, EM) 7. Trichoferus sbordonii Sama, 1982 (*WM, EM) 8. Stromatium unicolor (Olivier, 1795) (*WM, EM) 9. Phoracantha recurva Newman, 1840 (*EM) 10. Phoracantha semipunctata (Fabricius, 1775) (*WM, EM) 11. Cerambyx cerdo Linnaeus, 1758 (*WM, EM) 12. Cerambyx dux (Faldermann, 1837) (*WM, EM) 13. Cerambyx miles Bonelli, 1812 (*WM, EM) 14. Cerambyx nodulosus Germar, 1817 (*WM, EM) 15. Cerambyx welensii (Küster, 1846) (*WM, EM) 16. Cerambyx scopolii Fuessly, 1775 (*WM, EM) 17. Purpuricenus budensis (Götz, 1783) (*WM, EM) 18. Purpuricenus dalmatinus Sturm, 1843 (*WM, EM) 19. Purpuricenus desfontainei (Fabricius, 1792) (*WM, EM) 20. Purpuricenus interscapillatus Plavilstshikov, 1937 (*WM, EM) 21. Purpuricenus nigronotatus Pic, 1907 (*EM) 22. Purpuricenus wachanrui Levrat, 1858 (*EM) 23. Aromia moschata (Linnaeus, 1758) (*WM, EM) 24. Penichroa fasciata (Stephens, 1831) (*WM, EM) 25. Axinopalpis gracilis (Krynicki, 1832) (*WM, EM) 26. Obrium cantharinum (Linnaeus, 1767) (*WM) 27. Anatolobrium eggeri Adlbauer, 2004 (*WM) 28. Certallum ebulinum (Linnaeus, 1767) (*WM, EM) 29. Delagrangeus angustissimus Pic, 1892 (*EM) 30. Deilus fugax (Olivier, 1790) (*WM, EM) 31. Stenopterus atricornis Pic, 1891 (*WM) 32. Stenopterus flavicornis Küster, 1846 (*WM, EM) 33. Stenopterus kraatzi Pic, 1892 (*WM, EM) 34. Stenopterus rufus (Linnaeus, 1767) (*WM, EM) 35. Callimus akbesianus Pic, 1892 (*EM) 36. Callimus angulatus (Schrank, 1789) (*WM, EM) 37. Lampropterus femoratus (Germar, 1824) (*WM, EM) 38. Procallimus egregius (Mulsant & Rey, 1863) (*WM, EM) 39. Nathrius brevipennis (Mulsant, 1839) (*WM, EM) 40. Molorchus juglandis Sama, 1982 (*WM, EM) 41. Molorchus minor (Linnaeus, 1758) (*WM, EM) 42. Glaphyra kiesenwetteri (Mulsant & Rey, 1861) (*WM, EM) 43. Glaphyra marmottani Brisout de Barneville, 1863 (*WM, EM) 44. Glaphyra tenuitarsis (Holzschuh, 1981) (*EM) 45. Brachypteroma holtzi Pic, 1905 (*EM) 46. Brachypteroma ottomanum Heyden, 1863 (*EM) 47. Stenhomalus bicolor (Kraatz, 1862) (*WM, EM) 48. Hylotrupes bajulus (Linnaeus, 1758) (*WM, EM) 49. Ropalopus clavipes (Fabricius, 1775) (*EM) 50. Leioderes tuerki Ganglbauer, 1885 (*EM) 51. Semanotus russicus (Fabricius, 1777) (*WM) 52. Callidium syriacum (Pic, 1892) (*WM, EM) 53. Phymatodes testaceus (Linnaeus, 1758) (*WM, EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 99
54. Phymatodes magnanii (Sama & Rapuzzi, 1999) (*WM, EM) 55. Phymatodes rufipes (Fabricius, 1777) (*WM, EM) 56. Poecilium alni (Linnaeus, 1767) (*EM) 57. Poecilium lividum (Rossi, 1794) (*EM) 58. Plagionotus arcuatus (Linnaeus, 1758) (*WM, EM) 59. Plagionotus detritus (Linnaeus, 1758) (*WM, EM) 60. Plagionotus bobelayei (Brullé, 1832) (*WM, EM) 61. Plagionus floralis (Pallas, 1773) (*WM, EM) 62. Isotomus syriacus (Schneider, 1787) (*EM) 63. Chlorophorus aegyptiacus (Fabricius, 1775) (*EM) 64. Chlorophorus dinae (Schneider, 1787) (*WM, EM) 65. Chlorophorus figuratus (Scopoli, 1763) (*WM, EM) 66. Chlorophorus gratiosus Marseul, 1868 (*WM, EM) 67. Chlorophorus hungaricus Seidlitz, 1891 (*EM) 68. Chlorophorus nivipictus (Kraatz, 1879) (*WM, EM) 69. Chlorophorus sartor (Müller, 1766) (*WM, EM) 70. Chlorophorus trifasciatus (Fabricius, 1781) (*WM, EM) 71. Chlorophorus varius (Müller, 1766) (*WM, EM) 72. Xylotrechus arvicola (Olivier, 1795) (*EM) 73. Pseudosphegesthes longitarsus Holzschuh, 1974 (*WM) 74. Clytus ciliciensis (Chevrolat, 1863) (*EM) 75. Clytus gulekanus Pic, 1904 (*EM) 76. Clytus madoni Pic, 1891 (*EM) 77. Clytus rhamni Germar, 1817 (*WM, EM) 78. Clytus taurusiensis (Pic, 1903) (*EM)
Subfamily LAMIINAE
1. Parmena lukati Sama, 1994 (*WM) 2. Parmena mutilloides Sabbadini & Pesarini, 1992 (*WM) 3. Mesosa curculionoides (Linnaeus, 1760) (*WM) 4. Batocera rufomaculata (DeGeer, 1775) (*WM, EM) 5. Monochamus galloprovincialis (Olivier, 1795) (*WM, EM) 6. Morimus funereus (Mulsant, 1862) (*WM) 7. Morimus ganglbaueri Reitter, 1894 (*EM) 8. Dorcadion amanense Breuning, 1943 (*EM) 9. Dorcadion anatolicum Pic, 1900 (*WM, EM) 10. Dorcadion arcivagum Thomson, 1867 (*EM) 11. Dorcadion carinipenne Pic, 1900 (*EM) 12. Dorcadion cinerarium (Fabricius, 1787) (*EM) 13. Dorcadion culminicola Thomson, 1868 (*EM) 14. Dorcadion delagrangei Pic, 1894 (*EM) 15. Dorcadion deyrollei Ganglbauer, 1884 (*EM) 16. Dorcadion divisum Germar, 1839 (*WM, EM) 17. Dorcadion drusoides Breuning, 1962 (*EM) 18. Dorcadion halepense (Kraatz, 1873) (*EM) 19. Dorcadion holtzi (Pic, 1905) (*EM) 20. Dorcadion infernale Mulsant & Rey, 1863 (*WM, EM) 21. Dorcadion ispartense Breuning, 1962 (*WM) 22. Dorcadion kraetschmeri Bernhauer, 1988 (*WM) 23. Dorcadion lohsei Braun, 1976 (*EM) 24. Dorcadion menradi Holzschuh, 1989 (*EM) 25. Dorcadion micans Thomson, 1867 (*EM) 26. Dorcadion miminfernale Breuning, 1970 (*EM) 27. Dorcadion oezdurali Önalp, 1988 (*EM) 28. Dorcadion ortrudae Braun, 1980 (*EM) 29. Dorcadion paracinerarium Breuning, 1974 (*EM) 30. Dorcadion petrovitzi Heyrovsky, 1964 (*EM) 31. Dorcadion pseudinfernale Breuning, 1943 (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 100
32. Dorcadion saulcyi Thomson, 1865 (*EM) 33. Dorcadion scabricolle Dalman, 1817 (*WM, EM) 34. Dorcadion semibrunneum Pic, 1903 (*WM, EM) 35. Dorcadion septemlineatum Waltl, 1838 (*WM) 36. Dorcadion sinuatevittatum Pic, 1937 (*WM, EM) 37. Dorcadion smyrnense (Linneaus, 1757) (*EM) 38. Dorcadion subinterruptum Pic, 1900 (*EM) 39. Dorcadion syriense Breuning, 1943 (*EM) 40. Dorcadion variegatum Ganglbauer, 1884 (*EM) 41. Dorcadion triste Frivaldsky, 1845 (*WM) 42. Anaesthetis anatolica Holzschuh, 1969 (*WM, EM) 43. Niphona picticornis Mulsant, 1839 (*WM, EM) 44. Pogonocerus anatolicus Mulsant, 1839 (*WM, EM) 45. Pogonocerus perroudi Mulsant, 1839 (*EM) 46. Aegomorphus clavipes (Schrank, 1781) (*EM) 47. Acanthocinus aedilis (Linnaeus, 1758) (*WM) 48. Acanthocinus griseus (Fabricius, 1792) (*WM, EM) 49. Leiopus nebulosus (Linnaeus, 1758) (*WM) 50. Leiopus syriacus (Ganglbauer, 1884) (*WM, EM) 51. Exocentrus adspersus Mulsant, 1846 (*EM) 52. Exocentrus punctipennis Mulsant & Guillebeau, 1856 (*WM) 53. Exocentrus ritae Sama, 1985 (*EM) 54. Tetrops praeustus (Linnaeus, 1758) (*WM) 55. Tetrops warnckei Holzschuh, 1977 (*WM) 56. Saperda punctata (Linnaeus, 1767) (*WM) 57. Saperda populnea (Linnaeus, 1758) (*WM, EM) 58. Saperda quercus Charpentier, 1825 (*WM, EM) 59. Oberea linearis (Linnaeus, 1760) (*EM) 60. Oberea oculata (Linnaeus, 1758) (*WM, EM) 61. Oberea erythrocephala (Schrank, 1776) (*WM, EM) 62. Oxylia argentata (Ménétriés, 1832) (*WM, EM) 63. Oxylia duponcheli (Brullé, 1832) (*WM, EM) 64. Mallosia mirabilis (Faldermann, 1837) (*EM) 65. Mallosia imperatrix Abeille de Perrin, 1885 (*EM) 66. Semiangusta delagrangei (Pic, 1891) (*EM) 67. Coptosia bithynensis (Ganglbauer, 1884) (*WM, EM) 68. Coptosia ganglbaueri Pic, 1936 (*EM) 69. Coptosia compacta (Ménétriés, 1832) (*WM) 70. Coptosia schuberti Fuchs, 1965 (*WM, EM) 71. Pygoptosia speciosa (Frivaldszky, 1884) (*EM) 72. Phytoecia annulata Hampe, 1862 (*EM) 73. Phytoecia griseomaculata (Pic, 1891) (*EM) 74. Phytoecia hirsutula (Frölich, 1793) (*WM, EM) 75. Phytoecia tigrina Mulsant, 1851 (*WM) 76. Phytoecia adelpha Ganglbauer, 1886 (*EM) 77. Phytoecia alziari Sama, 1992 (*WM, EM) 78. Phytoecia armeniaca Frivaldszky, 1878 (*WM, EM) 79. Phytoecia demelti (Sama, 2003) (*EM) 80. Phytoecia ferrugata Ganglbauer, 1884 (*WM, EM) 81. Phytoecia humeralis (Waltl, 1838) (*WM, EM) 82. Phytoecia praetextata (Steven, 1817) (*EM) 83. Phytoecia anatolica Fuchs & Breuning, 1971 (*EM) 84. Phytoecia astarte Ganglbauer, 1886 (*EM) 85. Phytoecia wachanrui Mulsant, 1851 (*WM, EM) 86. Phytoecia merkli Ganglbauer, 1884 (*EM) 87. Phytoecia waltli Sama, 1991 (*EM) 88. Phytoecia annulipes Mulsant & Rey, 1863 (*EM) 89. Phytoecia asiatica Pic, 1891 (*EM) 90. Phytoecia bangi Pic, 1897 (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 101
91. Phytoecia caerulea (Scopoli, 1772) (*WM, EM) 92. Phytoecia croceipes Reiche & Saulcy, 1858 (*EM) 93. Phytoecia cylindrica (Linnaeus, 1758) (*EM) 94. Phytoecia geniculata Mulsant, 1862 (*WM, EM) 95. Phytoecia icterica (Schaller, 1783) (*EM) 96. Phytoecia manicata Reiche & Saulcy, 1858 (*WM, EM) 97. Phytoecia nigricornis (Fabricius, 1782) (*EM) 98. Phytoecia pubescens Pic, 1895 (*EM) 99. Phytoecia pustulata (Schrank, 1776) (*EM) 100. Phytoecia rufipes (Olivier, 1795) (*EM) 101. Phytoecia virgula (Charpentier, 1825) (*WM, EM) 102. Phytoecia coerulescens (Scopoli, 1763) (*WM, EM) 103. Phytoecia samai Özdikmen & Turgut, 2008 (*EM) 104. Phytoecia vittipennis Reiche, 1877 (*WM, EM) 105. Calamobius filum (Rossi, 1790) (*WM, EM) 106. Agapanthia kirbyi (Gyllenhal, 1817) (*WM, EM) 107. Agapanthia asphodeli (Latreille, 1804) (*WM, EM) 108. Agapanthia cynarae (Germar, 1817) (*EM) 109. Agapanthia dahli (Richter, 1821) (*EM) 110. Agapanthia detrita Kraatz, 1882 (*EM) 111. Agapanthia kindermanni Pic, 1905 (*EM) 112. Agapanthia lateralis Ganglbauer, 1884 (*WM, EM) 113. Agapanthia verecunda Chevrolat, 1882 (*EM) 114. Agapanthia villosoviridescens (DeGeer, 1775) (*WM, EM) 115. Agapanthia walteri Reitter, 1898 (*EM) 116. Agapanthia maculicornis (Gyllenhal, 1817) (*EM) 117. Agapanthia coeruleipennis Frivaldszky, 1878 (*WM, EM) 118. Agapanthia suturalis (Fabricius, 1787) (*WM, EM) 119. Agapanthia amitina Holzschuh, 1989 (*EM) 120. Agapanthia chalybaea Faldermann, 1837 (*WM, EM) 121. Agapanthia frivaldszkyi Ganglbauer, 1884 (*WM, EM) 122. Agapanthia lais Reiche & Saulcy, 1858 (*EM) 123. Agapanthia osmanlis Reiche et Saulcy, 1858 (*EM) 124. Agapanthia violacea (Fabricius, 1775) (*WM, EM)
Also Turanoclytus sieversi Ganglbauer, 1890 (syn.= Clytus deyrollei Pic, 1897 and Xylotrechus akbesianus Pic, 1902) that was recorded by Sama & Löbl in Löbl & Smetana (2010) for Anatolia, is present very likely in Mediterranean Region of Turkey. So, 281 longicorn beetles species occur in Mediterranean Region of Turkey. The following species which are 172 species of 77 genera of 6 subfamilies occur only in Antalya Part (= Western Mediterranean Region = WM) of Mediterranean Region (including Burdur, Isparta and Antalya provinces):
A LIST OF LONGICORN BEETLES FROM
WESTERN MEDITERRANEAN REGION (*WM)
Subfamily PRIONINAE
1. Ergates faber (Linnaeus, 1760) (*WM, EM) 2. Callergates gaillardoti (Chevrolat, 1854) (*WM, EM) 3. Prinobius myardi Mulsant, 1842 (*WM, EM) 4. Rhaesus serricollis (Motschulsky, 1838) (*WM, EM) 5. Aegosoma scabricorne (Scopoli, 1763) (*WM, EM) 6. Prionus coriarius (Linnaeus, 1758) (*WM, EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 102
7. Mesoprionus besikanus (Fairmaire, 1855) (*WM, EM)
Subfamily LEPTURINAE
1. Rhagium bifasciatum Fabricius, 1775 (*WM) 2. Rhagium elmaliense Schmid, 1999 (*WM) 3. Rhagium inquisitor (Linnaeus, 1758) (*WM) 4. Anisorus heterocerus (Ganglbauer, 1882) (*WM, EM) 5. Dinoptera collaris (Linnaeus, 1758) (*WM, EM) 6. Cortodera alpina (Ménétriés, 1832) (*WM, EM) 7. Cortodera cirsii Holzschuh, 1975 (*WM, EM) 8. Cortodera colchica Reitter, 1890 (*WM, EM) 9. Cortodera differens Pic, 1898 (*WM) 10. Cortodera discolor Fairmaire, 1866 (*WM, EM) 11. Cortodera flavimana (Waltl, 1838) (*WM, EM) 12. Cortodera humeralis (Schaller, 1783) (*WM, EM) 13. Cortodera imrasanica Sama & Rapuzzi, 1999 (*WM) 14. Grammoptera merkli Frivaldszky, 1884 (*WM, EM) 15. Alosterna anatolica Adlbauer, 1992 (*WM) 16. Alosterna tabacicolor (De Geer, 1775) (*WM) 17. Vadonia bisignata (Brullé, 1832) (*WM) 18. Vadonia bitlisiensis Chevrolat, 1882 (*WM) 19. Vadonia ciliciensis Daniel & Daniel, 1891 (*WM) 20. Vadonia danielorum Holzschuh, 1984 (*WM) 21. Vadonia moesiaca (Daniel & Daniel, 1891) (*WM) 22. Vadonia monostigma (Ganglbauer, 1882) (*WM) 23. Vadonia soror Holzschuh, 1981 (*WM, EM) 24. Vadonia unipunctata (Fabricius, 1787) (*WM, EM) 25. Pseudovadonia livida (Fabricius, 1777) (*WM, EM) 26. Stictoleptura rubra (Linnaeus, 1758) (*WM) 27. Stictoleptura cordigera (Füsslins, 1775) (*WM, EM) 28. Stictoleptura excisipes (Daniel & Daniel, 1891) (*WM, EM) 29. Stictoleptura fulva (DeGeer, 1775) (*WM, EM) 30. Stictoleptura gevneensis Özdikmen & Turgut, 2008 (*WM) 31. Stictoleptura pallidipennis (Tournier, 1872) (*?WM) 32. Stictoleptura rufa (Brullé, 1832) (*WM, EM) 33. Anastrangalia montana (Mulsant & Rey, 1863) (*WM, EM) 34. Pedostrangalia revestita (Linnaeus, 1767) (*WM) 35. Pedostrangalia emmipoda (Mulsant, 1863) (*WM, EM) 36. Pedostrangalia verticalis (Germar, 1822) (*WM) 37. Etorufus pubescens (Fabricius, 1787) (*WM) 38. Carlandrea syriaca (Pic, 1891) (*WM, EM) 39. Pachytodes erraticus (Dalman, 1817) (*WM, EM) 40. Rutpela maculata (Poda von Neuhaus, 1761) (*WM, EM) 41. Stenurella bifasciata (Müller, 1776) (*WM, EM) 42. Stenurella melanura (Linnaeus, 1758) (*WM) 43. Stenurella pamphyliae Rapuzzi & Sama, 2009 (*WM)
Subfamily NECYDALINAE
1. Necydalis hadullai Szallies, 1994 (*WM)
Subfamily ASEMINAE
1. Oxypleurus nodieri Mulsant, 1839 (*WM) 2. Alocerus moesiacus (Frivaldsky, 1837) (*WM) 3. Arhopalus ferus (Mulsant, 1839) (*WM, EM) 4. Arhopalus rusticus (Linnaeus, 1758) (*WM, EM) 5. Arhopalus syriacus (Reitter, 1895) (*WM, EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 103
Subfamily CERAMBYCINAE
1. Icosium tomentosum Lucas, 1854 (*WM) 2. Hesperophanes sericeus (Fabricius, 1787) (*WM) 3. Trichoferus fasciculatus (Faldermann, 1837) (*WM) 4. Trichoferus griseus (Fabricius, 1792) (*WM, EM) 5. Trichoferus kotschyi (Ganglbauer, 1883) (*WM, EM) 6. Trichoferus preissi (Heyden, 1894) (*WM, EM) 7. Trichoferus sbordonii Sama, 1982 (*WM, EM) 8. Stromatium unicolor (Olivier, 1795) (*WM, EM) 9. Phoracantha semipunctata (Fabricius, 1775) (*WM, EM) 10. Cerambyx cerdo Linnaeus, 1758 (*WM, EM) 11. Cerambyx dux (Faldermann, 1837) (*WM, EM) 12. Cerambyx miles Bonelli, 1812 (*WM, EM) 13. Cerambyx nodulosus Germar, 1817 (*WM, EM) 14. Cerambyx welensii (Küster, 1846) (*WM, EM) 15. Cerambyx scopolii Fuessly, 1775 (*WM, EM) 16. Purpuricenus budensis (Götz, 1783) (*WM, EM) 17. Purpuricenus dalmatinus Sturm, 1843 (*WM, EM) 18. Purpuricenus desfontainei (Fabricius, 1792) (*WM, EM) 19. Purpuricenus interscapillatus Plavilstshikov, 1937 (*WM, EM) 20. Aromia moschata (Linnaeus, 1758) (*WM, EM) 21. Penichroa fasciata (Stephens, 1831) (*WM, EM) 22. Axinopalpis gracilis (Krynicki, 1832) (*WM, EM) 23. Obrium cantharinum (Linnaeus, 1767) (*WM) 24. Anatolobrium eggeri Adlbauer, 2004 (*WM) 25. Certallum ebulinum (Linnaeus, 1767) (*WM, EM) 26. Deilus fugax (Olivier, 1790) (*WM, EM) 27. Stenopterus atricornis Pic, 1891 (*WM) 28. Stenopterus flavicornis Küster, 1846 (*WM, EM) 29. Stenopterus kraatzi Pic, 1892 (*WM, EM) 30. Stenopterus rufus (Linnaeus, 1767) (*WM, EM) 31. Callimus angulatus (Schrank, 1789) (*WM, EM) 32. Lampropterus femoratus (Germar, 1824) (*WM, EM) 33. Procallimus egregius (Mulsant & Rey, 1863) (*WM, EM) 34. Nathrius brevipennis (Mulsant, 1839) (*WM, EM) 35. Molorchus juglandis Sama, 1982 (*WM, EM) 36. Molorchus minor (Linnaeus, 1758) (*WM, EM) 37. Glaphyra kiesenwetteri (Mulsant & Rey, 1861) (*WM, EM) 38. Glaphyra marmottani Brisout de Barneville, 1863 (*WM, EM) 39. Stenhomalus bicolor (Kraatz, 1862) (*WM, EM) 40. Hylotrupes bajulus (Linnaeus, 1758) (*WM, EM) 41. Semanotus russicus (Fabricius, 1777) (*WM) 42. Callidium syriacum (Pic, 1892) (*WM, EM) 43. Phymatodes testaceus (Linnaeus, 1758) (*WM, EM) 44. Phymatodes magnanii (Sama & Rapuzzi, 1999) (*WM, EM) 45. Phymatodes rufipes (Fabricius, 1777) (*WM, EM) 46. Plagionotus arcuatus (Linnaeus, 1758) (*WM, EM) 47. Plagionotus detritus (Linnaeus, 1758) (*WM, EM) 48. Plagionotus bobelayei (Brullé, 1832) (*WM, EM) 49. Plagionus floralis (Pallas, 1773) (*WM, EM) 50. Chlorophorus dinae (Schneider, 1787) (*WM, EM) 51. Chlorophorus figuratus (Scopoli, 1763) (*WM, EM) 52. Chlorophorus gratiosus Marseul, 1868 (*WM, EM) 53. Chlorophorus nivipictus (Kraatz, 1879) (*WM, EM) 54. Chlorophorus sartor (Müller, 1766) (*WM, EM) 55. Chlorophorus trifasciatus (Fabricius, 1781) (*WM, EM) 56. Chlorophorus varius (Müller, 1766) (*WM, EM) 57. Pseudosphegesthes longitarsus Holzschuh, 1974 (*WM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 104
58. Clytus rhamni Germar, 1817 (*WM, EM)
Subfamily LAMIINAE
1. Parmena lukati Sama, 1994 (*WM) 2. Parmena mutilloides Sabbadini & Pesarini, 1992 (*WM) 3. Mesosa curculionoides (Linnaeus, 1760) (*WM) 4. Batocera rufomaculata (DeGeer, 1775) (*WM, EM) 5. Monochamus galloprovincialis (Olivier, 1795) (*WM, EM) 6. Morimus funereus (Mulsant, 1862) (*WM) 7. Dorcadion anatolicum Pic, 1900 (*WM, EM) 8. Dorcadion infernale Mulsant & Rey, 1863 (*WM, EM) 9. Dorcadion ispartense Breuning, 1962 (*WM) 10. Dorcadion kraetschmeri Bernhauer, 1988 (*WM) 11. Dorcadion scabricolle Dalman, 1817 (*WM, EM) 12. Dorcadion semibrunneum Pic, 1903 (*WM, EM) 13. Dorcadion septemlineatum Waltl, 1838 (*WM) 14. Dorcadion sinuatevittatum Pic, 1937 (*WM, EM) 15. Dorcadion triste Frivaldsky, 1845 (*WM) 16. Anaesthetis anatolica Holzschuh, 1969 (*WM, EM) 17. Niphona picticornis Mulsant, 1839 (*WM, EM) 18. Pogonocerus anatolicus Mulsant, 1839 (*WM, EM) 19. Acanthocinus aedilis (Linnaeus, 1758) (*WM) 20. Acanthocinus griseus (Fabricius, 1792) (*WM, EM) 21. Leiopus nebulosus (Linnaeus, 1758) (*WM) 22. Leiopus syriacus (Ganglbauer, 1884) (*WM, EM) 23. Exocentrus punctipennis Mulsant & Guillebeau, 1856 (*WM) 24. Tetrops praeustus (Linnaeus, 1758) (*WM) 25. Tetrops warnckei Holzschuh, 1977 (*WM) 26. Saperda punctata (Linnaeus, 1767) (*WM) 27. Saperda populnea (Linnaeus, 1758) (*WM, EM) 28. Saperda quercus Charpentier, 1825 (*WM, EM) 29. Oberea oculata (Linnaeus, 1758) (*WM, EM) 30. Oberea erythrocephala (Schrank, 1776) (*WM, EM) 31. Oxylia argentata (Ménétriés, 1832) (*WM, EM) 32. Oxylia duponcheli (Brullé, 1832) (*WM, EM) 33. Coptosia bithynensis (Ganglbauer, 1884) (*WM, EM) 34. Coptosia compacta (Ménétriés, 1832) (*WM) 35. Coptosia schuberti Fuchs, 1965 (*WM, EM) 36. Phytoecia hirsutula (Frölich, 1793) (*WM, EM) 37. Phytoecia tigrina Mulsant, 1851 (*WM) 38. Phytoecia alziari Sama, 1992 (*WM, EM) 39. Phytoecia armeniaca Frivaldszky, 1878 (*WM, EM) 40. Phytoecia ferrugata Ganglbauer, 1884 (*WM, EM) 41. Phytoecia humeralis (Waltl, 1838) (*WM, EM) 42. Phytoecia wachanrui Mulsant, 1851 (*WM, EM) 43. Phytoecia caerulea (Scopoli, 1772) (*WM, EM) 44. Phytoecia geniculata Mulsant, 1862 (*WM, EM) 45. Phytoecia manicata Reiche & Saulcy, 1858 (*WM, EM) 46. Phytoecia virgula (Charpentier, 1825) (*WM, EM) 47. Phytoecia coerulescens (Scopoli, 1763) (*WM, EM) 48. Phytoecia samai Özdikmen & Turgut, 2008 (*EM) 49. Phytoecia vittipennis Reiche, 1877 (*WM, EM) 50. Calamobius filum (Rossi, 1790) (*WM, EM) 51. Agapanthia kirbyi (Gyllenhal, 1817) (*WM, EM) 52. Agapanthia asphodeli (Latreille, 1804) (*WM, EM) 53. Agapanthia lateralis Ganglbauer, 1884 (*WM, EM) 54. Agapanthia villosoviridescens (DeGeer, 1775) (*WM, EM) 55. Agapanthia coeruleipennis Frivaldszky, 1878 (*WM, EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 105
56. Agapanthia suturalis (Fabricius, 1787) (*WM, EM) 57. Agapanthia chalybaea Faldermann, 1837 (*WM, EM) 58. Agapanthia frivaldszkyi Ganglbauer, 1884 (*WM, EM)
The following species which are 233 species of 78 genera of 7 subfamilies occur only in Adana Part (= Eastern Mediterranean Region = EM) of Mediterranean Region (including Ġçel, Adana, Osmaniye, Hatay, KahramanmaraĢ and Kilis provinces):
A LIST OF LONGICORN BEETLES FROM EASTERN MEDITERRANEAN REGION (*EM)
Subfamily PRIONINAE
1. Ergates faber (Linnaeus, 1760) (*WM, EM) 2. Callergates gaillardoti (Chevrolat, 1854) (*WM, EM) 3. Prinobius myardi Mulsant, 1842 (*WM, EM) 4. Rhaesus serricollis (Motschulsky, 1838) (*WM, EM) 5. Aegosoma scabricorne (Scopoli, 1763) (*WM, EM) 6. Prionus coriarius (Linnaeus, 1758) (*WM, EM) 7. Prionus komiyai Lorenc, 1999 (*EM) 8. Mesoprionus besikanus (Fairmaire, 1855) (*WM, EM)
Subfamily LEPTURINAE
1. Rhagium phrygium Daniel, 1906 (*EM) 2. Rhagium syriacum Pic, 1892 (*EM) 3. Stenocorus auricomus (Reitter, 1890) (*EM) 4. Anisorus heterocerus (Ganglbauer, 1882) (*WM, EM) 5. Dinoptera collaris (Linnaeus, 1758) (*WM, EM) 6. Cortodera alpina (Ménétriés, 1832) (*WM, EM) 7. Cortodera cirsii Holzschuh, 1975 (*WM, EM) 8. Cortodera colchica Reitter, 1890 (*WM, EM) 9. Cortodera discolor Fairmaire, 1866 (*WM, EM) 10. Cortodera flavimana (Waltl, 1838) (*WM, EM) 11. Cortodera humeralis (Schaller, 1783) (*WM, EM) 12. Cortodera omophloides Holzschuh, 1975 (*EM) 13. Cortodera pseudomophlus Reitter, 1889 (*EM) 14. Cortodera rubripennis Pic, 1891 (*EM) 15. Cortodera syriaca Pic, 1901 (*EM) 16. Grammoptera baudii (Stephens, 1831) (*EM) 17. Grammoptera merkli Frivaldszky, 1884 (*WM, EM) 18. Vadonia frater Holzschuh, 1981 (*EM) 19. Vadonia soror Holzschuh, 1981 (*WM, EM) 20. Vadonia unipunctata (Fabricius, 1787) (*WM, EM) 21. Pseudovadonia livida (Fabricius, 1777) (*WM, EM) 22. Anoplodera rufipes (Schaller, 1783) (*EM) 23. Anoplodera sexguttata (Fabricius, 1775) (*EM) 24. Stictoleptura cordigera (Füsslins, 1775) (*WM, EM) 25. Stictoleptura excisipes (Daniel & Daniel, 1891) (*WM, EM) 26. Stictoleptura fulva (DeGeer, 1775) (*WM, EM) 27. Stictoleptura heydeni (Ganglbauer, 1889) (*EM) 28. Stictoleptura pallens (Brullé, 1832) (*EM) 29. Stictoleptura rufa (Brullé, 1832) (*WM, EM) 30. Stictoleptura sambucicola (Holzschuh, 1982) (*EM) 31. Stictoleptura tesserula (Charpentier, 1825) (*EM) 32. Stictoleptura tonsa (Daniel & Daniel, 1891) (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 106
33. Stictoleptura tripartita (Heyden, 1889) (*EM) 34. Anastrangalia dubia (Scopoli, 1763) (*EM) 35. Anastrangalia montana (Mulsant & Rey, 1863) (*WM, EM) 36. Pedostrangalia emmipoda (Mulsant, 1863) (*WM, EM) 37. Carlandrea syriaca (Pic, 1891) (*WM, EM) 38. Pachytodes erraticus (Dalman, 1817) (*WM, EM) 39. Rutpela maculata (Poda von Neuhaus, 1761) (*WM, EM) 40. Stenurella bifasciata (Müller, 1776) (*WM, EM) 41. Stenurella jaegeri (Hummel, 1825) (*EM)
Subfamily ASEMINAE
1. Arhopalus ferus (Mulsant, 1839) (*WM, EM) 2. Arhopalus rusticus (Linnaeus, 1758) (*WM, EM) 3. Arhopalus syriacus (Reitter, 1895) (*WM, EM)
Subfamily SPONDYLIDINAE
1. Spondylis buprestoides (Linnaeus, 1758) (*EM)
Subfamily DORCASOMINAE
1. Apatophysis kadleci Danilevsky, 2008 (*EM)
Subfamily CERAMBYCINAE
1. Trichoferus griseus (Fabricius, 1792) (*WM, EM) 2. Trichoferus kotschyi (Ganglbauer, 1883) (*WM, EM) 3. Trichoferus preissi (Heyden, 1894) (*WM, EM) 4. Trichoferus sbordonii Sama, 1982 (*WM, EM) 5. Stromatium unicolor (Olivier, 1795) (*WM, EM) 6. Phoracantha recurva Newman, 1840 (*EM) 7. Phoracantha semipunctata (Fabricius, 1775) (*WM, EM) 8. Cerambyx cerdo Linnaeus, 1758 (*WM, EM) 9. Cerambyx dux (Faldermann, 1837) (*WM, EM) 10. Cerambyx miles Bonelli, 1812 (*WM, EM) 11. Cerambyx nodulosus Germar, 1817 (*WM, EM) 12. Cerambyx welensii (Küster, 1846) (*WM, EM) 13. Cerambyx scopolii Fuessly, 1775 (*WM, EM) 14. Purpuricenus budensis (Götz, 1783) (*WM, EM) 15. Purpuricenus dalmatinus Sturm, 1843 (*WM, EM) 16. Purpuricenus desfontainei (Fabricius, 1792) (*WM, EM) 17. Purpuricenus interscapillatus Plavilstshikov, 1937 (*WM, EM) 18. Purpuricenus nigronotatus Pic, 1907 (*EM) 19. Purpuricenus wachanrui Levrat, 1858 (*EM) 20. Aromia moschata (Linnaeus, 1758) (*WM, EM) 21. Penichroa fasciata (Stephens, 1831) (*WM, EM) 22. Axinopalpis gracilis (Krynicki, 1832) (*WM, EM) 23. Certallum ebulinum (Linnaeus, 1767) (*WM, EM) 24. Delagrangeus angustissimus Pic, 1892 (*EM) 25. Deilus fugax (Olivier, 1790) (*WM, EM) 26. Stenopterus flavicornis Küster, 1846 (*WM, EM) 27. Stenopterus kraatzi Pic, 1892 (*WM, EM) 28. Stenopterus rufus (Linnaeus, 1767) (*WM, EM) 29. Callimus akbesianus Pic, 1892 (*EM) 30. Callimus angulatus (Schrank, 1789) (*WM, EM) 31. Lampropterus femoratus (Germar, 1824) (*WM, EM) 32. Procallimus egregius (Mulsant & Rey, 1863) (*WM, EM) 33. Nathrius brevipennis (Mulsant, 1839) (*WM, EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 107
34. Molorchus juglandis Sama, 1982 (*WM, EM) 35. Molorchus minor (Linnaeus, 1758) (*WM, EM) 36. Glaphyra kiesenwetteri (Mulsant & Rey, 1861) (*WM, EM) 37. Glaphyra marmottani Brisout de Barneville, 1863 (*WM, EM) 38. Glaphyra tenuitarsis (Holzschuh, 1981) (*EM) 39. Brachypteroma holtzi Pic, 1905 (*EM) 40. Brachypteroma ottomanum Heyden, 1863 (*EM) 41. Stenhomalus bicolor (Kraatz, 1862) (*WM, EM) 42. Hylotrupes bajulus (Linnaeus, 1758) (*WM, EM) 43. Ropalopus clavipes (Fabricius, 1775) (*EM) 44. Leioderes tuerki Ganglbauer, 1885 (*EM) 45. Callidium syriacum (Pic, 1892) (*WM, EM) 46. Phymatodes testaceus (Linnaeus, 1758) (*WM, EM) 47. Phymatodes magnanii (Sama & Rapuzzi, 1999) (*WM, EM) 48. Phymatodes rufipes (Fabricius, 1777) (*WM, EM) 49. Poecilium alni (Linnaeus, 1767) (*EM) 50. Poecilium lividum (Rossi, 1794) (*EM) 51. Plagionotus arcuatus (Linnaeus, 1758) (*WM, EM) 52. Plagionotus detritus (Linnaeus, 1758) (*WM, EM) 53. Plagionotus bobelayei (Brullé, 1832) (*WM, EM) 54. Plagionus floralis (Pallas, 1773) (*WM, EM) 55. Isotomus syriacus (Schneider, 1787) (*EM) 56. Chlorophorus aegyptiacus (Fabricius, 1775) (*EM) 57. Chlorophorus dinae (Schneider, 1787) (*WM, EM) 58. Chlorophorus figuratus (Scopoli, 1763) (*WM, EM) 59. Chlorophorus gratiosus Marseul, 1868 (*WM, EM) 60. Chlorophorus hungaricus Seidlitz, 1891 (*EM) 61. Chlorophorus nivipictus (Kraatz, 1879) (*WM, EM) 62. Chlorophorus sartor (Müller, 1766) (*WM, EM) 63. Chlorophorus trifasciatus (Fabricius, 1781) (*WM, EM) 64. Chlorophorus varius (Müller, 1766) (*WM, EM) 65. Xylotrechus arvicola (Olivier, 1795) (*EM) 66. Clytus ciliciensis (Chevrolat, 1863) (*EM) 67. Clytus gulekanus Pic, 1904 (*EM) 68. Clytus madoni Pic, 1891 (*EM) 69. Clytus rhamni Germar, 1817 (*WM, EM) 70. Clytus taurusiensis (Pic, 1903) (*EM)
Subfamily LAMIINAE
1. Batocera rufomaculata (DeGeer, 1775) (*WM, EM) 2. Monochamus galloprovincialis (Olivier, 1795) (*WM, EM) 3. Morimus ganglbaueri Reitter, 1894 (*EM) 4. Dorcadion amanense Breuning, 1943 (*EM) 5. Dorcadion anatolicum Pic, 1900 (*WM, EM) 6. Dorcadion arcivagum Thomson, 1867 (*EM) 7. Dorcadion carinipenne Pic, 1900 (*EM) 8. Dorcadion cinerarium (Fabricius, 1787) (*EM) 9. Dorcadion culminicola Thomson, 1868 (*EM) 10. Dorcadion delagrangei Pic, 1894 (*EM) 11. Dorcadion deyrollei Ganglbauer, 1884 (*EM) 12. Dorcadion divisum Germar, 1839 (*WM, EM) 13. Dorcadion drusoides Breuning, 1962 (*EM) 14. Dorcadion halepense (Kraatz, 1873) (*EM) 15. Dorcadion holtzi (Pic, 1905) (*EM) 16. Dorcadion infernale Mulsant & Rey, 1863 (*WM, EM) 17. Dorcadion lohsei Braun, 1976 (*EM) 18. Dorcadion menradi Holzschuh, 1989 (*EM) 19. Dorcadion micans Thomson, 1867 (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 108
20. Dorcadion miminfernale Breuning, 1970 (*EM) 21. Dorcadion oezdurali Önalp, 1988 (*EM) 22. Dorcadion ortrudae Braun, 1980 (*EM) 23. Dorcadion paracinerarium Breuning, 1974 (*EM) 24. Dorcadion petrovitzi Heyrovsky, 1964 (*EM) 25. Dorcadion pseudinfernale Breuning, 1943 (*EM) 26. Dorcadion saulcyi Thomson, 1865 (*EM) 27. Dorcadion scabricolle Dalman, 1817 (*WM, EM) 28. Dorcadion semibrunneum Pic, 1903 (*WM, EM) 29. Dorcadion sinuatevittatum Pic, 1937 (*WM, EM) 30. Dorcadion smyrnense (Linneaus, 1757) (*EM) 31. Dorcadion subinterruptum Pic, 1900 (*EM) 32. Dorcadion syriense Breuning, 1943 (*EM) 33. Dorcadion variegatum Ganglbauer, 1884 (*EM) 34. Anaesthetis anatolica Holzschuh, 1969 (*WM, EM) 35. Niphona picticornis Mulsant, 1839 (*WM, EM) 36. Pogonocerus anatolicus Mulsant, 1839 (*WM, EM) 37. Pogonocerus perroudi Mulsant, 1839 (*EM) 38. Aegomorphus clavipes (Schrank, 1781) (*EM) 39. Acanthocinus aedilis (Linnaeus, 1758) (*WM) 40. Acanthocinus griseus (Fabricius, 1792) (*WM, EM) 41. Leiopus syriacus (Ganglbauer, 1884) (*WM, EM) 42. Exocentrus adspersus Mulsant, 1846 (*EM) 43. Exocentrus ritae Sama, 1985 (*EM) 44. Saperda populnea (Linnaeus, 1758) (*WM, EM) 45. Saperda quercus Charpentier, 1825 (*WM, EM) 46. Oberea linearis (Linnaeus, 1760) (*EM) 47. Oberea oculata (Linnaeus, 1758) (*WM, EM) 48. Oberea erythrocephala (Schrank, 1776) (*WM, EM) 49. Oxylia argentata (Ménétriés, 1832) (*WM, EM) 50. Oxylia duponcheli (Brullé, 1832) (*WM, EM) 51. Mallosia mirabilis (Faldermann, 1837) (*EM) 52. Mallosia imperatrix Abeille de Perrin, 1885 (*EM) 53. Semiangusta delagrangei (Pic, 1891) (*EM) 54. Coptosia bithynensis (Ganglbauer, 1884) (*WM, EM) 55. Coptosia ganglbaueri Pic, 1936 (*EM) 56. Coptosia schuberti Fuchs, 1965 (*WM, EM) 57. Pygoptosia speciosa (Frivaldszky, 1884) (*EM) 58. Phytoecia annulata Hampe, 1862 (*EM) 59. Phytoecia griseomaculata (Pic, 1891) (*EM) 60. Phytoecia hirsutula (Frölich, 1793) (*WM, EM) 61. Phytoecia adelpha Ganglbauer, 1886 (*EM) 62. Phytoecia alziari Sama, 1992 (*WM, EM) 63. Phytoecia armeniaca Frivaldszky, 1878 (*WM, EM) 64. Phytoecia demelti (Sama, 2003) (*EM) 65. Phytoecia ferrugata Ganglbauer, 1884 (*WM, EM) 66. Phytoecia humeralis (Waltl, 1838) (*WM, EM) 67. Phytoecia praetextata (Steven, 1817) (*EM) 68. Phytoecia anatolica Fuchs & Breuning, 1971 (*EM) 69. Phytoecia astarte Ganglbauer, 1886 (*EM) 70. Phytoecia wachanrui Mulsant, 1851 (*WM, EM) 71. Phytoecia merkli Ganglbauer, 1884 (*EM) 72. Phytoecia waltli Sama, 1991 (*EM) 73. Phytoecia annulipes Mulsant & Rey, 1863 (*EM) 74. Phytoecia asiatica Pic, 1891 (*EM) 75. Phytoecia bangi Pic, 1897 (*EM) 76. Phytoecia caerulea (Scopoli, 1772) (*WM, EM) 77. Phytoecia croceipes Reiche & Saulcy, 1858 (*EM) 78. Phytoecia cylindrica (Linnaeus, 1758) (*EM)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 109
79. Phytoecia geniculata Mulsant, 1862 (*WM, EM) 80. Phytoecia icterica (Schaller, 1783) (*EM) 81. Phytoecia manicata Reiche & Saulcy, 1858 (*WM, EM) 82. Phytoecia nigricornis (Fabricius, 1782) (*EM) 83. Phytoecia pubescens Pic, 1895 (*EM) 84. Phytoecia pustulata (Schrank, 1776) (*EM) 85. Phytoecia rufipes (Olivier, 1795) (*EM) 86. Phytoecia virgula (Charpentier, 1825) (*WM, EM) 87. Phytoecia coerulescens (Scopoli, 1763) (*WM, EM) 88. Phytoecia samai Özdikmen & Turgut, 2008 (*EM) 89. Phytoecia vittipennis Reiche, 1877 (*WM, EM) 90. Calamobius filum (Rossi, 1790) (*WM, EM) 91. Agapanthia kirbyi (Gyllenhal, 1817) (*WM, EM) 92. Agapanthia asphodeli (Latreille, 1804) (*WM, EM) 93. Agapanthia cynarae (Germar, 1817) (*EM) 94. Agapanthia dahli (Richter, 1821) (*EM) 95. Agapanthia detrita Kraatz, 1882 (*EM) 96. Agapanthia kindermanni Pic, 1905 (*EM) 97. Agapanthia lateralis Ganglbauer, 1884 (*WM, EM) 98. Agapanthia verecunda Chevrolat, 1882 (*EM) 99. Agapanthia villosoviridescens (DeGeer, 1775) (*WM, EM) 100. Agapanthia walteri Reitter, 1898 (*EM) 101. Agapanthia maculicornis (Gyllenhal, 1817) (*EM) 102. Agapanthia coeruleipennis Frivaldszky, 1878 (*WM, EM) 103. Agapanthia suturalis (Fabricius, 1787) (*WM, EM) 104. Agapanthia amitina Holzschuh, 1989 (*EM) 105. Agapanthia chalybaea Faldermann, 1837 (*WM, EM) 106. Agapanthia frivaldszkyi Ganglbauer, 1884 (*WM, EM) 107. Agapanthia lais Reiche & Saulcy, 1858 (*EM) 108. Agapanthia osmanlis Reiche et Saulcy, 1858 (*EM) 109. Agapanthia violacea (Fabricius, 1775) (*WM, EM)
LITERATURE CITED
Adlbauer, K. 1988. Neues zur Taxonomie und Faunistik der Bockkäferfauna der Türkei (Coleoptera, Cerambycidae). Entomofauna, 9 (12): 257-297. Adlbauer, K. 1992. Zur Faunistik und Taxonomie der Bockkäferfauna der Türkei II (Coleoptera, Cerambycidae). Entomofauna, 13 (30): 485-509. Adlbauer, K. 2004. A. eggeri Alkan, H. 2000. Türkiye orman Cerambycidae (Insecta, Coleoptera)‘lerinin tanıtımı ve Doğu Karadeniz Bölgesindeki türlerin araĢtırılması. Yüksek Lisans Tezi. Karadeniz Teknik Üniversitesi Fen Bilimleri Enstitüsü, Trabzon, 227 pp. Althoff, J. & Danilevsky, M. L. 1997. A Check-List of Longicorn Beetles (Coleoptera, Cerambycoidea) of Europe. Slovensko Entomološko Društvo Štefana Michielija. Ljubljana, 64 pp. Aurivillius, C. 1921. Coleopterorum Catalogus, pars 73, Cerambycidae: Lamiinae. Berlin. W. Junk & S. Schenkling. 704 pp. Bernhauer, D. 1988. Türkische Dorcadien (1. Beitrag) (Col.: Cerambycidae: Lamiinae). Entomologische Zeitschrift, 98: 97-104. Bodemeyer, H. E. V. 1900. Quer durch Klein Asien, in den Bulghar Dagh; Eine Naturvissenschafliche studien-Reise. Coleopterologisches, 196 pp. Bodemeyer, H. E. V. 1906. Beitrage zur Käferfauna von Klein Asien - Deutsche Entomologische Zeitschrift, 2: 417-437.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 110
Bodenheimer, F. S. 1958. Türkiye‘de ziraate ve ağaçlara zararlı olan böcekler ve bunlarla savaĢ hakkında bir etüt. Bayur Matbaası, Ankara, 347 pp. Braun, W. 1978. Die Dorcadienausbeute der Forschungsreisen von W. Heinz 1963-1977, Faunistiche Aufstellung, Beschreibung einer neuen Unterart und Bemerkungen zur Systematik wenig bekannter Arten (Coleoptera, Cerambycidae). Nachrichtenblatt der Bayerischen Entomologen, 27 (6): 101-116. Braun, W. 1979. Beitrag zur Kenntnis der Gattung Dorcadion Systematisch neu bewertete Dorcadion-Formen (Col., Cerambycidae). Nachrichtenblatt der Bayerischen Entomologen, 28 (5): 81-86. Breuning, S. 1951. Revision du genre Phytoecia Muls. (Coleoptera Cerambycidae). Entom. Arbeiten aus dem Museum G. Frey 2: 1-103, 353-460. Breuning, S. 1962. Revision der Dorcadionini. Entomologische abhandlungen und berichte aus dem staatliche museum für tierkunde in Dresden, 27: 665. Breuning, S. 1966. Catalogue des Lamiares du Monde (Col.: Céramb.). Im Verlag des Museums G. Frey, Tutzing 9: 659-765. Breuning, S. 1970. Nouveaux Dorcadion des collections du Muséum de Paris. L‘Entomologiste, 24: 97-101. Breuning, S. 1974. Descriptions de dorcadions nouveaux d‘Anatolie (Coleoptera: Lamiinae). Bulletin de la Societe Entomologique de Mulhouse, 1974: 65. Breuning, S. & Villiers, A. 1967. Cérambycides de Turquie (2. note). L‘ Entomologiste, 23 (3): 59-63. Çanakçıoğlu, H. 1983. Orman Entomolojisi: Özel bölüm. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 349, Ġstanbul, 535 pp. Çanakçıoğlu, H. 1993. Orman Entomolojisi: Özel bölüm. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 3623, Ġstanbul, 535 pp. Çanakçıoğlu, H. & Mol, T. 1998. Orman Entomolojisi: Zararlı ve yararlı böcekler. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 451, Ġstanbul, 144-174. Daniel, K. 1906. [new taxa]. In: Daniel, K. & Daniel, J., Münchener Koleopterologische Zeitschrift, 3 (1906-1908): 41-45, 170-187. Danilevsky, M. L. 1993. Some preliminary results of the type-material study of the Cerambycidae (Coleoptera) collection of the National Museum of Natural History, Paris. Russian Ent. Journ., 2 (1): 47-50. Danilevsky, M. L. 1999. New taxa of Cerambycidae from Turkey and Transcaucasia (Coleoptera). Elytron, 13: 39-47. Danilevsky, M. L. 2008. Apatophysis Chevrolat, 1860 (Coleoptera, Cerambycidae) of Russia and adjacent regions. Studies and reports of District Museum Prague-East. Taxonomical Series, 4 (1-2): 7-55. Danilevsky, M. L. 2010. Dorcadionini. In Löbl, I. & Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera, Volume 6, Chrysomeloidea. Apollo Books, Stenstrup, pp. 241-264. Danilevsky, M. L. 2010. Systematic list of Longicorn Beetles (Cerambycoidea) of the territory of the former USSR. Available from: http://www.cerambycidae.net/. Danilevsky, M. L. & Miroshnikov A. I. 1985. Timber-Beetles of Caucasus (Coleoptera, Cerambycidae). The Key. Krasnodar, 419 pp. Demelt, C. V. 1963. Beitrag zur Kenntnis der Cerambycidenfauna Kleinasiens und 13. Beitrag zur Biologie palaearkt. Cerambyciden, sowie Beschreibung einer neuen Oberea-Art. Entomologische Blatter, 59 (3) : 132-151. Demelt, C. V. 1967. Nachtrag zur Kenntnis der Cerambyciden-Fauna Kleinasiens. Entomologische Blatter, 63 (2): 106-109. Demelt, C. V. & Alkan, B. 1962. Short information of Cerambycidae Fauna of Turkey. Bitki Koruma Bülteni, 2 (10): 49-56.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 111
Drumont, A. & Komiya, Z. 2010. Prioninae. In Löbl, I. & Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera, Volume 6, Chrysomeloidea. Apollo Books, Stenstrup, pp. 86-94. Ekici, M. 1971. Sedir (Cedrus libani Barre.) zararlı böceklerinin biyolojisi ve mücadelesi. Ormancılık AraĢtırma Enstitüsü Yay., Teknik Bülten Serisi No : 45, 50-51. Erdem, R. 1968. Ormanın faydalı ve Zararlı böcekleri. Ġstanbul Üniversitesi Orman Fakültesi Yay., Ġstanbul, 43-58. Erdem, R. & Çanakçıoğlu, H. 1977. Türkiye odun zararlıları. Ġstanbul Üniversitesi Orman Fakültesi Yay., Ġstanbul, 113-134. Fairmaire, M. L. 1881. Description de quelques coleopteres de Syrie. Annales de la Societe Entomologique de France. 1880: 27-28. Fairmaire, M. L. 1884. Liste des Coleopteres recueillis par M. l‘abbe David a Akbes (Asie-Mineure) et Descriptions des Especes Nouvelles. Annales de la Societe entomologique de France, Octobre (1884): 165-180. Frivaldszky, J. 1878. Coleoptera nova. Uj tehelyröpüek. Termeszetrajzi Füzetek, 2: 9-14. Frivaldszky, J. 1884. Coleoptera nova ex Asia minore. Uj tehelyröpüek Kis-Azsiabol. Termeszetrajzi Füzetek, 8: 1-8. Fuchs, E. 1965. Zwei neue palaearktische Saperdini (Col.). Entomologische Blatter, 61: 110-112. Fuchs, E. & Breuning, S. 1971. Die Cerambycidenausbeute der Anatolienexpedition 1966-67 des Naturhistorischen Museums, Wien. Annalen Naturhistorischen Museum Wien, 75: 435-439. Ganglbauer, L. 1884. Bestimmungs-Tabellen der europaeischen Coleopteren. VIII. Cerambycidae.-Verhandlungen der kaiserlich-kongiglichen zoologish.-botanischen Gesellschaft in Wien, 33: 437-586. Gül-Zümreoğlu, S. 1972. Catalogue of Insect and common pests (1928-1969). T. C. Publications of Agriculture Ministry, Bornova, Ġzmir, 119 pp. Gül-Zümreoğlu, S. 1975. Investigations on taxonomy, host plants and distribution of the Longhorned Beetles (Cerambycidae-Coleoptera) in Aegean Region. T. C. Ministry of Food, Agriculture and Stockbreeding, No : 28, , Ġstiklal Press, Ġzmir, 208 pp. Heyden, L. 1888. Neue und interessante Coleopteren aus Malatia in Mesopotamien. Deutsche Entomologische Zeitschrift, 32 (1): 72-78. Heyrovsky, L. 1964. Eine neue Dorcadion-Art aus Kleinasien. Reichenbachia, 3: 97-98. Holzschuh, C. 1971. Zwei neue Phytoecia-Arten (Col.: Cerambycidae) aus Anatolien und Libanon. Mitteilungen der Forstlichen Bundes-Versuchsanstalt Wien, 94: 67-69. Holzschuh, C. 1974. Neue Bockkafer aus Pakistan, Iran, Anatolien und Mazedonien (Col.: Cerambycidae). Zeitschrift der Arbeitgemeinschaft Österre,chischer Entomologen, 25: 81-100. Holzschuh, C. 1975a. Neue westpalaearktische Bockkafer aus den Gattungen Cortodera, Vadonia und Agapanthia (Coleoptera: Cerambycidae). Zeitschrift der arbeitsgemeinschaft oesterreich entomologen, 26: 77-90. Holzschuh, C. 1975b. Zur Synonymie Palaearktischer Cerambycidae I. (Col.). Koleopterologische Rundschau, 52 (1975): 101-104. Holzschuh, C. 1977. Neue Bockkäfer aus Anatolien und Iran (Col., Cerambycidae). Koleopterologische Runschau, 53: 127-136. Holzschuh, C. 1980. Revision einer Cerambycidenausbeute des Naturhistorischen Museums Wien (Coleoptera). Annalen des Naturhistorischen Museums in Wien, 83: 573-574. Holzschuh, C. 1981a. Zwanzig neue Bockkafer aus Europa und Asien (Col.: Cerambycidae). Zeitschrift der Arbeitgemeinschaft Österre,chischer Entomologen, 33: 65-76.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 112
Holzschuh, C. 1981b. Beitrage zur Kenntnis der Europeischen Tetrops-Arten (Col.: Cerambycidae). Koleopterologische Rundschau, 55: 77-89. Holzschuh, C. 1984a. Beschreibung von 21 neuen Bockkafern aus Europa und Asien (Col.: Cerambycidae). Koleopterologische Rundschau, 57: 141-165. Holzschuh, C. 1984b. Beschreibung neuer Arten aus der unmittelbaren Verwandtschaft von Phytoecia (Pilemia) tigrina (Col.: Cerambycidae). Koleopterologische Rundschau, 57: 167-175. Holzschuh, C. 1989. Beschreibung neuer Bockkafer aus Europa und Asien (Col.: Cerambycidae). Koleopterologische Rundschau, 59: 153-183. Hoskovec, M. & Rejzek, M. 2010. Longhorn Beetles (Cerambycidae) oft he West Palaearctic Region. Available from: http://www.cerambyx.uochb.cz/ İren, Z. & Ahmed, M. K. 1973. Microlepidoptera and pests of fruit-trees in Turkey. Bitki Koruma Bülteni, Ankara, Turkey, 1: 41-42. İyriboz, N. 1940. Ġncir hastalıkları. T. C. Ziraat Vekaleti NeĢriyatı Umumi sayı: 489, Ġzmir, 44-47. Kadlec, S. & Rejzek, M. 2001. Trichoferus samai, a new species from Turkey (Coleoptera: Cerambycidae). Biocosme Mesogeen, 17: 295-302. Kanat, M. 1998. KahramanmaraĢ ormanlarında önemli zararlı böceklerin araĢtarılması. Yüksek Lisans Tezi, Karadeniz Teknik Üniversitesi, Fen Bilimleri Enstitüsü, Trabzon, 41-127. Kraatz, G. 1884. Neue Kafer-Arten aus Malatia im sudlichen Kleinasien. Deutsche Entomologische Zeitschrift, 28: 231-234. Levrat, J. N. G. B. 1858. Description de deux coleopteres nouveaux. Annales de la Societe Linnenne de Lyon 5 (2): 260-263. Lobanov, A. L., Danilevsky, M. L. & Murzin, S. V. 1981. [Systematic list of Longicorn beetles (Coleoptera, Cerambycidae) of the USSR. 1]. Revue d‘Entomologie, 60(4): 784-803 (in Russian). Löbl, I. & Smetana, A. 2010. Catalogue of Palaearctic Coleoptera, Volume 6, Chrysomeloidea. Apollo Books, Stenstrup, 924 pp. Lodos, N. 1998. Entomology of Turkey VI (General, Aplied and Faunistic). Ege Ü. Ziraat Fak. Yayınları No: 529, E. Ü. Faculty of Agriculture Press, Ġzmir, 300 pp. Lorenc, J. 2006. Systematic checklist of the Cerambycoidea of the NW Palaearctic region, Euroasiatic and Primary Mediterran, Part I. Subfamily Parandranae and Prioninae. Available from: http://cerambycidae.cz/CHECKLIST%20pal.%201..pdf Malmusi, M. & Saltini, L. 2005. Cerambycidae raccolti dai componenti del Gruppo Modenese Scienze Naturali durante escursioni in Turchia tra il 1987-2003 (Contributo alla Fauna dei Cerambycidae di Turchia). Quaderno di studi e notizie di storia naturale della Romagna, n. 21, 28 pp. (unpublished). Nizamlıoğlu, K. & Gökmen, N. 1964. Damaging insects on olive in Turkey. Institute of Agriculture of Göztepe, Ġstanbul, 51-56. Önalp, B. 1988. Taxonomic researches on Agapanthia Serville, 1835 species (Coleoptera, Cerambycidae: Lamiinae) I. H. Ü. Eğitim Fakültesi Dergisi, 3: 257-295. Önalp, B. 1989. Taxonomic researches on Agapanthia Serville, 1835 species (Coleoptera, Cerambycidae: Lamiinae) II. H. Ü. Eğitim Fakültesi Dergisi, 4: 197-234. Önalp, B. 1990. Systematic researches on Dorcadion Dalman, 1817 species in Turkey (Coleoptera, Cerambycidae: Lamiinae) I. H. Ü. Eğitim Fakültesi Dergisi, 5: 57-102. Önalp, B. 1991. Systematic researches on Dorcadion Dalman, 1817 species in Turkey (Coleoptera, Cerambycidae: Lamiinae) II. H. Ü. Eğitim Fakültesi Dergisi, 6: 191-227. Öymen, T. 1987. The Forest Cerambycidae of Turkey. Ġ. Ü. Forest Faculty, Ġstanbul, 146 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 113
Özbek, H. 1978. Hylotrupes bajulus (L.) Serville in Erzurum and the near, and some others longhorn beetles. Atatürk Üniversitesi Ziraat Fakultesi Dergisi, 9 (1): 31-44 (in Turkish). Özdikmen, H. 2003. The Genus Cortodera Mulsant, 1863 (Cerambycidae: Coleoptera) in Turkey. Phytoparasitica, 31 (5): 433-441. Özdikmen, H. 2006. Contribution to the knowledge of Turkish longicorn beetles fauna (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 1 (1): 71-90. Özdikmen, H. 2007. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part I – Black Sea Region. Munis Entomology & Zoology 2 (2): 179-422. Özdikmen, H. & Çağlar, Ü. 2004. Contribution to the knowledge of longhorned bettles (Coleoptera, Cerambycidae) from Turkey, Subfamilies Prioninae, Lepturinae, Spondylidinae and Cerambycinae. J. Ent. Res. Soc., 6 (1): 39-69. Özdikmen, H. & Demir, H. 2006. Notes on longicorn beetles fauna of Turkey (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 1 (1): 157-166. Özdikmen, H. & Demirel, E. 2005. Additional Notes to the Knowledge of Longhorned Beetle Collection from Zoological Museum of Gazi University, Ankara, Turkey (GUZM) for Turkish Fauna (Coleoptera, Cerambycidae). J. Ent. Res. Soc., 7 (3): 13-38. Özdikmen, H., Güven, M. & Gören, C. 2010. Longhorned beetles fauna of Amanos Mountains, Southern Turkey (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 5, suppl.: 1141-1167. Özdikmen, H. & Hasbenli, A. 2004. Contribution to the knowledge of longhorned bettles (Coleoptera, Cerambycidae) from Turkey, Subfamily Lamiinae. J. Ent. Res. Soc., 6 (2): 25-49. Özdikmen, H. & Okutaner, A. Y. 2006. The longhorned beetles fauna (Coleoptera, Cerambycidae) of KahramanmaraĢ province. G. U. Journal of Science 19 (2): 77-89. Özdikmen, H., Özdemir, Y. & Turgut, S. 2005. Longhorned Beetles Collection of the Nazife Tuatay Plant Protection Museum, Ankara, Turkey (Coleoptera, Cerambycidae). J. Ent. Res. Soc., 7 (2): 1-33. Özdikmen, H. & Şahin, Ö. 2006. Ġç Anadolu Ormancılık AraĢtırma Müdürlüğü, Entomoloji Müzesi (Türkiye, Ankara) Teke Böcekleri Koleksiyonu (Coleoptera, Cerambycidae). G. U. Journal of Science, 19 (1): 1-8. Özdikmen, H. & Turgut, S. 2008a. A new Cortodera species for Turkey (Coleoptera: Cerambycidae: Lepturinae). Munis Entomology & Zoology, 3 (2): 596-605. Özdikmen, H. & Turgut, S. 2008b. A new species of the genus Stictoleptura Casey, 1924 from Turkey (Coleoptera: Cerambycidae: Lepturinae). Munis Entomology & Zoology, 3 (2): 548-553. Özdikmen, H. & Turgut, S. 2008c. A new species and a new subspecies of the subgenus Phytoecia (Blepisanis) Pascoe, 1866 from Turkey (Coleoptera: Cerambycidae: Lamiinae). Munis Entomology & Zoology, 3 (2): 568-581. Özdikmen, H. & Turgut, S. 2009. A synopsis of Turkish Vesperinae Mulsant, 1839 and Prioninae Latreille, 1802 (Coleoptera: Cerambycidae). Munis Entomology & Zoology 4 (2): 402-423. Turgut, S. & Özdikmen, H. 2010. New data for Turkish longhorned beetles fauna from Southern Turkey (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 5, suppl.: 859-889. Peks, H. 1993. Drei neue Taxa der Gattung Dorcadion aus Anatolien (Coleoptera: Cerambycidae: Lamiinae). Coleoptera. Schwanfelder Coleopterologische Mitteilungen, 1: 1-11. Plavilstshikov, N. N. 1915. Les especes palearctiques du genre Rhagium F. (Coleoptera, Cerambycidae). Revue Russ. d`Entom., XV, 1: 1-19. Plavilstshikov, N. N. 1958. Fauna SSR. Zhestokrylye. T. XXIII, vyp. 1. Zhuki-drovoseki. Ch. 3. Podsemeistvo Lamiinae, ch. 1. Moskva-Leningrad: Izdatel‘stvo Akademii Nauk SSSR, 591 pp. Perissinotto, A. & Luchini, S. R. 1966. Coleotteri Raccolti Nel Vicino e Medio Oriente Nota I. Dorcadion Dalm. (Coleoptera, Cerambycidae). Bollettino della Societa Entomologica Italiana, 96 (9-10): 147-149.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 114
Rapuzzi, P. & Sama, G. 2009. Description of new Cerambycidae from Greece, Turkey, Northern Syria and China (Insecta: Coleoptera: Cerambycidae). Quaderno di Studi e Notizie di Storia Naturale della Romagna, 29: 181-188. Reitter, E. 1905. Übersicht der Arten der Coleopteren-Gattung Pilemia Fairm. Aus der palaearktischen Fauna. Wiener Entomologische Zeitung, 24: 239-240. Rejzek, M., Kadlec, S. & Sama, G. 2003. Conribution to the knowledge of Syrian Cerambycidae fauna (Coleoptera). Biocosme Mésogéen, Nice, 20 (1): 7-50. Rejzek, M. & Hoskovec, M. 1999. Cerambycidae of Nemrut Dağı National Park (Anatolia, South-East Turkey). Biocosme Mésogéen, Nice, 15 (4): 257-272. Rejzek, M., Sama, G. & Alziar, G. 2001. Host Plants of Several Herb-Feeding Cerambycidae Mainly from East Mediterranean Region (Coleoptera : Cerambycidae). Biocosme Mésogéen, Nice, 17(4): 263-294. Sabbadini, A. & Pesarini, C. 1992. Note su Purpuricenus budensis (Goeze) e specie affini. Bollettino della Società italiana di Entomologia, Genova, 124 (1): 55-64. Sama, G. 1982. Contributo allo studio dei coleotteri Cerambycidae di Grecia e Asia Minore. Fragmenta Entomologica, Roma, 16 (2): 205-227. Sama, G. 1985. Exocentrus ritae n.sp. della Turchia orientale. Lavori Soc. ven. Sc. ital., 10: 65-67. Sama, G. 1992. Révision des types de la Collection M.Pic. II. Callidiini de l'Anatolie - L'Entomologiste, 48 (4): 191-196. Sama, G. 1993. Notes on Neomusaria with description of a new species from Turkey. Koleopterologische Rundschau, 63: 293-297. Sama, G. 1994a. Descrizione di Parmena striatopunctata n.sp. della Turchia nord orientale e note sul genere Delagrangeus Pic, 1892. Lambillionea, 94 (4): 553-558. Sama, G. 1994b. Cerambycidae nuovi o poco noti del Mediterraneo orientale. Lambillionea, 94 (1): 9-13. Sama, G. 1994c. Note sulla nomenclatura dei Cerambycidae della regione mediterranea. II. Revisione di alcuni tipi di Kraatz, V. Heyden e Stierlin. Lambillionea, 94 (3): 321-334. Sama, G. 1995. Note sui Molorchini. II. I generi Glaphyra Newman, 1840 e Nathrioglaphyra nov.. Lambillionea, 95 (3): 363-390. Sama, G. 1996. Contribution à la connaissance des Longicornes de Grèce et d‘Asie Mineure. Biocosme mesogéen, 12 (4): 101-116. Sama, G. & Rapuzzi, P. 1999. Cerambycidae nuovi e poco noti di Turchia e Medio Oriente. Lambillionea, 99 (3): 461-468. Sama, G. & Rapuzzi, P. 2000. Note preliminaire pour une faune des Cerambycidae du Liban. Lambillionea, 100 (1): 7-23. Sama, G. 2002. Atlas of the Cerambycidae of Europe and the Mediterranean Area, Volume I, Kabourek, Zlin, 173 pp. Sama, G. 2003. Descrizione di due nuovi Cerambycidae di Turchia: Solaia antonellae n. gen., n. sp. e Helladia demelti n. sp. Quaderno di studi e notizie di storia naturale della Romagna, 17 (suppl.): 69-78. Sama, G., Rapuzzi, P. & Rejzek, M. 2007. New or interesting Phytoeciini from the Middle East, especially from Iran (Coleoptera: Cerambycidae). Folia Heyrovskyana, 14 (4): 163-179. Sama, G. & Rejzek, M. 2001. Helladia imperialis, espèce nouvelle de l‘Iran. Biocosme Mésogéen, 17 (3): 239-246. Sama, G. & Löbl, I. 2010. Cerambycidae (others). In Löbl, I. & Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera, Volume 6, Chrysomeloidea. Apollo Books, Stenstrup.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 115
Schmid, H. 1999. Ein neuer Bockkafer aus Anatolien (Cerambycidae). Koleopterologische Rundschau, 69: 157-158. Szallies, A. 1994. Drei neue Coleopteren-Arten aus der Turkei (Coleoptera: Prostomidae, Cerambycidae). Entomologische Zeitschrift, 104: 259-263. Schmitschek, E. 1944. Forstinsekten der Türkei und Ihre Umwelt Grundlagen der türkischen Forstentomologie, Volk und Reich Verlag Prag, 125-141 pp. Svacha, P. & Danilevsky, M. L. 1986. Cerambycoid Larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea), Part I. Acta Universitatis Carolinae – Biologica, 30: 1-186. Svacha, P. & Danilevsky, M. L. 1988. Cerambycoid Larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea), Part III. Acta Universitatis Carolinae – Biologica, 32: 1-205. Taglianti, A. V., Audisio, P. A., Biondi, M., Bologna, M. A., Carpaneto, G. M., De Biase, A., Fattorini, S., Piattella, E., Sindaco, R., Venchi, A. & Zapparoli, M. 1999. A proposal for a chorotype classification of the Near East fauna, in the framework of the Western Palaearctic Region. Biogeographia 20: 31-59. Tauzin, P. 2000. Complement a l‘inventaire des Coleopteres Cerambycidae de Turquie. L‘Entomologiste, 56 (4): 151-153. Thomson, J. 1867. Physis. Recueil d‘Histoire Naturelle. [Revisionen und Neubeschreibungen von Kafern.]. Vol. 1. Paris: Societe Entomologique de France, 170 pp. Tosun, İ. 1975. Akdeniz Bölgesi iğne yapraklı ormanlarında zarar yapan böcekler ve önemli türlerin parazit ve yırtıcıları üzerine araĢtırmalar. Ġstanbul 201 pp. Tozlu, G. & Özbek, H. 2000. The Tropical Fig Borer, Batocera rufomaculata (Coleoptera: Cerambycidae), new for Turkey. Zoology in the Middle East, 20: 121-124. Tozlu, G., Rejzek, M. & Özbek, H. 2002. A contribution to the knowledge of Cerambycidae (Coleoptera) fauna of Turkey. Part I: Subfamilies Prioninae to Cerambycinae. Biocosme Mèsogèen, Nice, 19 (1-2): 55-94. Tozlu, G., Rejzek, M. & Özbek, H. 2003. A Contribution to the Knowledge of Cerambycidae (Coleoptera) Fauna of Turkey Part II : Subfamily Lamiinae. Biocosme Mèsogèen, Nice, 19 (3): 95-110. Tuatay, N., Kalkandelen, A. & Aysev, N. 1972. Bitki Koruma Müzesi Böcek Kataloğu (1961-1971). T. C. Tarım Bakanlığı, Ankara, 53-55. Ulusoy, R., Vatansever, G. & Uygun, N. 1999. The cherry pests, their natural enemies and observations on some important species in UlukıĢla (Niğde) and Pozantı (Adana) provinces of Turkey. Türkiye Entomoloji Dergisi, 23 (2): 111-120. Villiers, A. 1959. Cérambycides de Turquie. L‘ Entomologiste, 15 (1-2): 7-11. Villiers, A. 1967. Coléoptéres Cérambycides de Turquie (1. Partie) - L‘ Entomologiste, 23 (1): 18-22. Winkler, A. 1924-1932. Catalogus Coleopterorum regionis palaearcticae. Verlag von Albert Winkler, 1135-1226.
APPENDIX
1. Map of Turkey‘s regions and adjacent areas. 2. Map of Turkey‘s provinces and adjacent areas. 3. Map of the provinces of Turkey. 4. Ergates faber (Linnaeus, 1760): Distribution patterns in Turkey. 5. Callergates gaillardoti (Chevrolat, 1854): Distribution patterns in Turkey. 6. Prinobius myardi Mulsant, 1842: Distribution patterns in Turkey. 7. Rhaesus serricollis (Motschulsky, 1838): Distribution patterns in Turkey. 8. Aegosoma scabricorne (Scopoli, 1763): Distribution patterns in Turkey. 9. Prionus coriarius (Linnaeus, 1758): Distribution patterns in Turkey. 10. Prionus komiyai Lorenc, 1999: Distribution patterns in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 116
11. Mesoprionus besikanus (Fairmaire, 1855): Distribution patterns in Turkey. 12. Rhagium bifasciatum Fabricius, 1775: Distribution patterns in Turkey. 13. Rhagium elmaliense Schmid, 1999: Distribution patterns in Turkey. 14. Rhagium phrygium Daniel, 1906: Distribution patterns in Turkey. 15. Rhagium syriacum Pic, 1892: Distribution patterns in Turkey. 16. Rhagium inquisitor (Linnaeus, 1758): Distribution patterns in Turkey. 17. Stenocorus auricomus (Reitter, 1890): Distribution patterns in Turkey. 18. Anisorus heterocerus (Ganglbauer, 1882): Distribution patterns in Turkey. 19. Dinoptera collaris (Linnaeus, 1758): Distribution patterns in Turkey. 20. Cortodera alpina (Ménétriés, 1832): Distribution patterns in Turkey. 21. Cortodera cirsii Holzschuh, 1975: Distribution patterns in Turkey. 22. Cortodera colchica Reitter, 1890: Distribution patterns in Turkey. 23. Cortodera differens Pic, 1898: Distribution patterns in Turkey. 24. Cortodera discolor Fairmaire, 1866: Distribution patterns in Turkey. 25. Cortodera flavimana (Waltl, 1838): Distribution patterns in Turkey. 26. Cortodera humeralis (Schaller, 1783): Distribution patterns in Turkey. 27. Cortodera imrasanica Sama & Rapuzzi, 1999: Distribution patterns in Turkey. 28. Cortodera omophloides Holzschuh, 1975: Distribution patterns in Turkey. 29. Cortodera pseudomophlus Reitter, 1889: Distribution patterns in Turkey. 30. Cortodera rubripennis Pic, 1891: Distribution patterns in Turkey. 31. Cortodera syriaca Pic, 1901: Distribution patterns in Turkey. 32. Grammoptera baudii (Stephens, 1831): Distribution patterns in Turkey. 33. Grammoptera merkli Frivaldszky, 1884: Distribution patterns in Turkey. 34. Alosterna anatolica Adlbauer, 1992: Distribution patterns in Turkey. 35. Alosterna tabacicolor (De Geer, 1775): Distribution patterns in Turkey. 36. Vadonia bisignata (Brullé, 1832): Distribution patterns in Turkey. 37. Vadonia bitlisiensis Chevrolat, 1882: Distribution patterns in Turkey. 38. Vadonia ciliciensis Daniel & Daniel, 1891: Distribution patterns in Turkey. 39. Vadonia danielorum Holzschuh, 1984: Distribution patterns in Turkey. 40. Vadonia frater Holzschuh, 1981: Distribution patterns in Turkey. 41. Vadonia moesiaca (Daniel & Daniel, 1891): Distribution patterns in Turkey. 42. Vadonia monostigma (Ganglbauer, 1882): Distribution patterns in Turkey. 43. Vadonia soror Holzschuh, 1981: Distribution patterns in Turkey. 44. Vadonia unipunctata (Fabricius, 1787): Distribution patterns in Turkey. 45. Pseudovadonia livida (Fabricius, 1777): Distribution patterns in Turkey. 46. Anoplodera rufipes (Schaller, 1783): Distribution patterns in Turkey. 47. Anoplodera sexguttata (Fabricius, 1775): Distribution patterns in Turkey. 48. Stictoleptura rubra (Linnaeus, 1758): Distribution patterns in Turkey. 49. Stictoleptura cordigera (Füsslins, 1775): Distribution patterns in Turkey. 50. Stictoleptura excisipes (Daniel & Daniel, 1891): Distribution patterns in Turkey. 51. Stictoleptura fulva (DeGeer, 1775): Distribution patterns in Turkey. 52. Stictoleptura gevneensis Özdikmen & Turgut, 2008: Distribution patterns in
Turkey. 53. Stictoleptura heydeni (Ganglbauer, 1889): Distribution patterns in Turkey. 54. Stictoleptura pallens (Brullé, 1832): Distribution patterns in Turkey. 55. Stictoleptura pallidipennis (Tournier, 1872): Distribution patterns in Turkey. 56. Stictoleptura rufa (Brullé, 1832): Distribution patterns in Turkey. 57. Stictoleptura sambucicola (Holzschuh, 1982): Distribution patterns in Turkey. 58. Stictoleptura tesserula (Charpentier, 1825): Distribution patterns in Turkey. 59. Stictoleptura tonsa (Daniel & Daniel, 1891): Distribution patterns in Turkey. 60. Stictoleptura tripartita (Heyden, 1889): Distribution patterns in Turkey. 61. Anastrangalia dubia (Scopoli, 1763): Distribution patterns in Turkey. 62. Anastrangalia montana (Mulsant & Rey, 1863): Distribution patterns in Turkey. 63. Pedostrangalia revestita (Linnaeus, 1767): Distribution patterns in Turkey. 64. Pedostrangalia emmipoda (Mulsant, 1863): Distribution patterns in Turkey. 65. Pedostrangalia verticalis (Germar, 1822): Distribution patterns in Turkey. 66. Etorufus pubescens (Fabricius, 1787): Distribution patterns in Turkey. 67. Carlandrea syriaca (Pic, 1891): Distribution patterns in Turkey. 68. Pachytodes erraticus (Dalman, 1817): Distribution patterns in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 117
69. Rutpela maculata (Poda von Neuhaus, 1761): Distribution patterns in Turkey. 70. Stenurella bifasciata (Müller, 1776): Distribution patterns in Turkey. 71. Stenurella jaegeri (Hummel, 1825): Distribution patterns in Turkey. 72. Stenurella melanura (Linnaeus, 1758): Distribution patterns in Turkey. 73. Stenurella pamphyliae Rapuzzi & Sama, 2009: Distribution patterns in Turkey. 74. Necydalis hadullai Szallies, 1994: Distribution patterns in Turkey. 75. Oxypleurus nodieri Mulsant, 1839: Distribution patterns in Turkey. 76. Alocerus moesiacus (Frivaldsky, 1837): Distribution patterns in Turkey. 77. Arhopalus ferus (Mulsant, 1839): Distribution patterns in Turkey. 78. Arhopalus rusticus (Linnaeus, 1758): Distribution patterns in Turkey. 79. Arhopalus syriacus (Reitter, 1895): Distribution patterns in Turkey. 80. Spondylis buprestoides (Linnaeus, 1758): Distribution patterns in Turkey. 81. Apatophysis kadleci Danilevsky, 2008: Distribution patterns in Turkey. 82. Icosium tomentosum Lucas, 1854: Distribution patterns in Turkey. 83. Hesperophanes sericeus (Fabricius, 1787): Distribution patterns in Turkey. 84. Trichoferus fasciculatus (Faldermann, 1837): Distribution patterns in Turkey. 85. Trichoferus griseus (Fabricius, 1792): Distribution patterns in Turkey. 86. Trichoferus kotschyi (Ganglbauer, 1883): Distribution patterns in Turkey. 87. Trichoferus preissi (Heyden, 1894): Distribution patterns in Turkey. 88. Trichoferus sbordonii Sama, 1982: Distribution patterns in Turkey. 89. Stromatium unicolor (Olivier, 1795): Distribution patterns in Turkey. 90. Phoracantha recurva Newman, 1840: Distribution patterns in Turkey. 91. Phoracantha semipunctata (Fabricius, 1775): Distribution patterns in Turkey. 92. Cerambyx cerdo Linnaeus, 1758: Distribution patterns in Turkey. 93. Cerambyx dux (Faldermann, 1837): Distribution patterns in Turkey. 94. Cerambyx miles Bonelli, 1812: Distribution patterns in Turkey. 95. Cerambyx nodulosus Germar, 1817: Distribution patterns in Turkey. 96. Cerambyx welensii (Küster, 1846): Distribution patterns in Turkey. 97. Cerambyx scopolii Fuessly, 1775: Distribution patterns in Turkey. 98. Purpuricenus budensis (Götz, 1783): Distribution patterns in Turkey. 99. Purpuricenus dalmatinus Sturm, 1843: Distribution patterns in Turkey. 100. Purpuricenus desfontainei (Fabricius, 1792): Distribution patterns in Turkey. 101. Purpuricenus interscapillatus Plavilstshikov, 1937: Distribution patterns in
Turkey. 102. Purpuricenus nigronotatus Pic, 1907: Distribution patterns in Turkey. 103. Purpuricenus wachanrui Levrat, 1858: Distribution patterns in Turkey. 104. Aromia moschata (Linnaeus, 1758): Distribution patterns in Turkey. 105. Penichroa fasciata (Stephens, 1831): Distribution patterns in Turkey. 106. Axinopalpis gracilis (Krynicki, 1832): Distribution patterns in Turkey. 107. Obrium cantharinum (Linnaeus, 1767): Distribution patterns in Turkey. 108. Anatolobrium eggeri Adlbauer, 2004: Distribution patterns in Turkey. 109. Certallum ebulinum (Linnaeus, 1767): Distribution patterns in Turkey. 110. Delagrangeus angustissimus Pic, 1892: Distribution patterns in Turkey. 111. Deilus fugax (Olivier, 1790): Distribution patterns in Turkey. 112. Stenopterus atricornis Pic, 1891: Distribution patterns in Turkey. 113. Stenopterus flavicornis Küster, 1846: Distribution patterns in Turkey. 114. Stenopterus kraatzi Pic, 1892: Distribution patterns in Turkey. 115. Stenopterus rufus (Linnaeus, 1767): Distribution patterns in Turkey. 116. Callimus akbesianus Pic, 1892: Distribution patterns in Turkey. 117. Callimus angulatus (Schrank, 1789): Distribution patterns in Turkey. 118. Lampropterus femoratus (Germar, 1824): Distribution patterns in Turkey. 119. Procallimus egregius (Mulsant & Rey, 1863): Distribution patterns in Turkey. 120. Nathrius brevipennis (Mulsant, 1839): Distribution patterns in Turkey. 121. Molorchus juglandis Sama, 1982: Distribution patterns in Turkey. 122. Molorchus minor (Linnaeus, 1758): Distribution patterns in Turkey. 123. Glaphyra kiesenwetteri (Mulsant & Rey, 1861): Distribution patterns in Turkey. 124. Glaphyra marmottani Brisout de Barneville, 1863: Distribution patterns in
Turkey. 125. Glaphyra tenuitarsis (Holzschuh, 1981): Distribution patterns in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 118
126. Brachypteroma holtzi Pic, 1905: Distribution patterns in Turkey. 127. Brachypteroma ottomanum Heyden, 1863: Distribution patterns in Turkey. 128. Stenhomalus bicolor (Kraatz, 1862): Distribution patterns in Turkey. 129. Hylotrupes bajulus (Linnaeus, 1758): Distribution patterns in Turkey. 130. Ropalopus clavipes (Fabricius, 1775): Distribution patterns in Turkey. 131. Leioderes tuerki Ganglbauer, 1885: Distribution patterns in Turkey. 132. Semanotus russicus (Fabricius, 1777): Distribution patterns in Turkey. 133. Callidium syriacum (Pic, 1892): Distribution patterns in Turkey. 134. Phymatodes testaceus (Linnaeus, 1758): Distribution patterns in Turkey. 135. Phymatodes magnanii (Sama & Rapuzzi, 1999): Distribution patterns in Turkey. 136. Phymatodes rufipes (Fabricius, 1777): Distribution patterns in Turkey. 137. Poecilium alni (Linnaeus, 1767): Distribution patterns in Turkey. 138. Poecilium lividum (Rossi, 1794): Distribution patterns in Turkey. 139. Plagionotus arcuatus (Linnaeus, 1758): Distribution patterns in Turkey. 140. Plagionotus detritus (Linnaeus, 1758): Distribution patterns in Turkey. 141. Plagionotus bobelayei (Brullé, 1832): Distribution patterns in Turkey. 142. Plagionus floralis (Pallas, 1773): Distribution patterns in Turkey. 143. Isotomus syriacus (Schneider, 1787): Distribution patterns in Turkey. 144. Chlorophorus aegyptiacus (Fabricius, 1775): Distribution patterns in Turkey. 145. Chlorophorus dinae (Schneider, 1787): Distribution patterns in Turkey. 146. Chlorophorus figuratus (Scopoli, 1763): Distribution patterns in Turkey. 147. Chlorophorus gratiosus Marseul, 1868: Distribution patterns in Turkey. 148. Chlorophorus hungaricus Seidlitz, 1891: Distribution patterns in Turkey. 149. Chlorophorus nivipictus (Kraatz, 1879): Distribution patterns in Turkey. 150. Chlorophorus sartor (Müller, 1766): Distribution patterns in Turkey. 151. Chlorophorus trifasciatus (Fabricius, 1781): Distribution patterns in Turkey. 152. Chlorophorus varius (Müller, 1766): Distribution patterns in Turkey. 153. Xylotrechus arvicola (Olivier, 1795): Distribution patterns in Turkey. 154. Pseudosphegesthes longitarsus Holzschuh, 1974: Distribution patterns in Turkey. 155. Clytus ciliciensis (Chevrolat, 1863): Distribution patterns in Turkey. 156. Clytus gulekanus Pic, 1904: Distribution patterns in Turkey. 157. Clytus madoni Pic, 1891: Distribution patterns in Turkey. 158. Clytus rhamni Germar, 1817: Distribution patterns in Turkey. 159. Clytus taurusiensis (Pic, 1903): Distribution patterns in Turkey. 160. Parmena lukati Sama, 1994: Distribution patterns in Turkey. 161. Parmena mutilloides Sabbadini & Pesarini, 1992: Distribution patterns in Turkey. 162. Mesosa curculionoides (Linnaeus, 1760): Distribution patterns in Turkey. 163. Batocera rufomaculata (DeGeer, 1775): Distribution patterns in Turkey. 164. Monochamus galloprovincialis (Olivier, 1795): Distribution patterns in Turkey. 165. Morimus funereus (Mulsant, 1862): Distribution patterns in Turkey. 166. Morimus ganglbaueri Reitter, 1894: Distribution patterns in Turkey. 167. Dorcadion amanense Breuning, 1943: Distribution patterns in Turkey. 168. Dorcadion anatolicum Pic, 1900: Distribution patterns in Turkey. 169. Dorcadion arcivagum Thomson, 1867: Distribution patterns in Turkey. 170. Dorcadion carinipenne Pic, 1900: Distribution patterns in Turkey. 171. Dorcadion cinerarium (Fabricius, 1787): Distribution patterns in Turkey. 172. Dorcadion culminicola Thomson, 1868: Distribution patterns in Turkey. 173. Dorcadion delagrangei Pic, 1894: Distribution patterns in Turkey. 174. Dorcadion deyrollei Ganglbauer, 1884: Distribution patterns in Turkey. 175. Dorcadion divisum Germar, 1839: Distribution patterns in Turkey. 176. Dorcadion drusoides Breuning, 1962: Distribution patterns in Turkey. 177. Dorcadion halepense (Kraatz, 1873): Distribution patterns in Turkey. 178. Dorcadion holtzi (Pic, 1905): Distribution patterns in Turkey. 179. Dorcadion infernale Mulsant & Rey, 1863: Distribution patterns in Turkey. 180. Dorcadion ispartense Breuning, 1962: Distribution patterns in Turkey. 181. Dorcadion kraetschmeri Bernhauer, 1988: Distribution patterns in Turkey. 182. Dorcadion lohsei Braun, 1976: Distribution patterns in Turkey. 183. Dorcadion menradi Holzschuh, 1989: Distribution patterns in Turkey. 184. Dorcadion micans Thomson, 1867: Distribution patterns in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 119
185. Dorcadion miminfernale Breuning, 1970: Distribution patterns in Turkey. 186. Dorcadion oezdurali Önalp, 1988: Distribution patterns in Turkey. 187. Dorcadion ortrudae Braun, 1980: Distribution patterns in Turkey. 188. Dorcadion paracinerarium Breuning, 1974: Distribution patterns in Turkey. 189. Dorcadion petrovitzi Heyrovsky, 1964: Distribution patterns in Turkey. 190. Dorcadion pseudinfernale Breuning, 1943: Distribution patterns in Turkey. 191. Dorcadion saulcyi Thomson, 1865: Distribution patterns in Turkey. 192. Dorcadion scabricolle Dalman, 1817: Distribution patterns in Turkey. 193. Dorcadion semibrunneum Pic, 1903: Distribution patterns in Turkey. 194. Dorcadion septemlineatum Waltl, 1838: Distribution patterns in Turkey. 195. Dorcadion sinuatevittatum Pic, 1937: Distribution patterns in Turkey. 196. Dorcadion smyrnense (Linneaus, 1757): Distribution patterns in Turkey. 197. Dorcadion subinterruptum Pic, 1900: Distribution patterns in Turkey. 198. Dorcadion syriense Breuning, 1943: Distribution patterns in Turkey. 199. Dorcadion variegatum Ganglbauer, 1884: Distribution patterns in Turkey. 200. Dorcadion triste Frivaldsky, 1845: Distribution patterns in Turkey. 201. Anaesthetis anatolica Holzschuh, 1969: Distribution patterns in Turkey. 202. Niphona picticornis Mulsant, 1839: Distribution patterns in Turkey. 203. Pogonocerus anatolicus Mulsant, 1839: Distribution patterns in Turkey. 204. Pogonocerus perroudi Mulsant, 1839: Distribution patterns in Turkey. 205. Aegomorphus clavipes (Schrank, 1781): Distribution patterns in Turkey. 206. Acanthocinus aedilis (Linnaeus, 1758): Distribution patterns in Turkey. 207. Acanthocinus griseus (Fabricius, 1792): Distribution patterns in Turkey. 208. Leiopus nebulosus (Linnaeus, 1758): Distribution patterns in Turkey. 209. Leiopus syriacus (Ganglbauer, 1884): Distribution patterns in Turkey. 210. Exocentrus adspersus Mulsant, 1846: Distribution patterns in Turkey. 211. Exocentrus punctipennis Mulsant & Guillebeau, 1856: Distribution patterns in
Turkey. 212. Exocentrus ritae Sama, 1985: Distribution patterns in Turkey. 213. Tetrops praeustus (Linnaeus, 1758): Distribution patterns in Turkey. 214. Tetrops warnckei Holzschuh, 1977: Distribution patterns in Turkey. 215. Saperda punctata (Linnaeus, 1767): Distribution patterns in Turkey. 216. Saperda populnea (Linnaeus, 1758): Distribution patterns in Turkey. 217. Saperda quercus Charpentier, 1825: Distribution patterns in Turkey. 218. Oberea linearis (Linnaeus, 1760): Distribution patterns in Turkey. 219. Oberea oculata (Linnaeus, 1758): Distribution patterns in Turkey. 220. Oberea erythrocephala (Schrank, 1776): Distribution patterns in Turkey. 221. Oxylia argentata (Ménétriés, 1832): Distribution patterns in Turkey. 222. Oxylia duponcheli (Brullé, 1832): Distribution patterns in Turkey. 223. Mallosia mirabilis (Faldermann, 1837): Distribution patterns in Turkey. 224. Mallosia imperatrix Abeille de Perrin, 1885: Distribution patterns in Turkey. 225. Semiangusta delagrangei (Pic, 1891): Distribution patterns in Turkey. 226. Coptosia bithynensis (Ganglbauer, 1884): Distribution patterns in Turkey. 227. Coptosia ganglbaueri Pic, 1936: Distribution patterns in Turkey. 228. Coptosia compacta (Ménétriés, 1832): Distribution patterns in Turkey. 229. Coptosia schuberti Fuchs, 1965: Distribution patterns in Turkey. 230. Pygoptosia speciosa (Frivaldszky, 1884): Distribution patterns in Turkey. 231. Phytoecia annulata Hampe, 1862: Distribution patterns in Turkey. 232. Phytoecia griseomaculata (Pic, 1891): Distribution patterns in Turkey. 233. Phytoecia hirsutula (Frölich, 1793): Distribution patterns in Turkey. 234. Phytoecia tigrina Mulsant, 1851: Distribution patterns in Turkey. 235. Phytoecia adelpha Ganglbauer, 1886: Distribution patterns in Turkey. 236. Phytoecia alziari Sama, 1992: Distribution patterns in Turkey. 237. Phytoecia armeniaca Frivaldszky, 1878: Distribution patterns in Turkey. 238. Phytoecia demelti (Sama, 2003): Distribution patterns in Turkey. 239. Phytoecia ferrugata Ganglbauer, 1884: Distribution patterns in Turkey. 240. Phytoecia humeralis (Waltl, 1838): Distribution patterns in Turkey. 241. Phytoecia praetextata (Steven, 1817): Distribution patterns in Turkey. 242. Phytoecia anatolica Fuchs & Breuning, 1971: Distribution patterns in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 120
243. Phytoecia astarte Ganglbauer, 1886: Distribution patterns in Turkey. 244. Phytoecia wachanrui Mulsant, 1851: Distribution patterns in Turkey. 245. Phytoecia merkli Ganglbauer, 1884: Distribution patterns in Turkey. 246. Phytoecia waltli Sama, 1991: Distribution patterns in Turkey. 247. Phytoecia annulipes Mulsant & Rey, 1863: Distribution patterns in Turkey. 248. Phytoecia asiatica Pic, 1891: Distribution patterns in Turkey. 249. Phytoecia bangi Pic, 1897: Distribution patterns in Turkey. 250. Phytoecia caerulea (Scopoli, 1772): Distribution patterns in Turkey. 251. Phytoecia croceipes Reiche & Saulcy, 1858: Distribution patterns in Turkey. 252. Phytoecia cylindrica (Linnaeus, 1758): Distribution patterns in Turkey. 253. Phytoecia geniculata Mulsant, 1862: Distribution patterns in Turkey. 254. Phytoecia icterica (Schaller, 1783): Distribution patterns in Turkey. 255. Phytoecia manicata Reiche & Saulcy, 1858: Distribution patterns in Turkey. 256. Phytoecia nigricornis (Fabricius, 1782): Distribution patterns in Turkey. 257. Phytoecia pubescens Pic, 1895: Distribution patterns in Turkey. 258. Phytoecia pustulata (Schrank, 1776): Distribution patterns in Turkey. 259. Phytoecia rufipes (Olivier, 1795): Distribution patterns in Turkey. 260. Phytoecia virgula (Charpentier, 1825): Distribution patterns in Turkey. 261. Phytoecia coerulescens (Scopoli, 1763): Distribution patterns in Turkey. 262. Phytoecia samai Özdikmen & Turgut, 2008: Distribution patterns in Turkey. 263. Phytoecia vittipennis Reiche, 1877: Distribution patterns in Turkey. 264. Calamobius filum (Rossi, 1790): Distribution patterns in Turkey. 265. Agapanthia kirbyi (Gyllenhal, 1817): Distribution patterns in Turkey. 266. Agapanthia asphodeli (Latreille, 1804): Distribution patterns in Turkey. 267. Agapanthia cynarae (Germar, 1817): Distribution patterns in Turkey. 268. Agapanthia dahli (Richter, 1821): Distribution patterns in Turkey. 269. Agapanthia detrita Kraatz, 1882: Distribution patterns in Turkey. 270. Agapanthia kindermanni Pic, 1905: Distribution patterns in Turkey. 271. Agapanthia lateralis Ganglbauer, 1884: Distribution patterns in Turkey. 272. Agapanthia verecunda Chevrolat, 1882: Distribution patterns in Turkey. 273. Agapanthia villosoviridescens (DeGeer, 1775): Distribution patterns in Turkey. 274. Agapanthia walteri Reitter, 1898: Distribution patterns in Turkey. 275. Agapanthia maculicornis (Gyllenhal, 1817): Distribution patterns in Turkey. 276. Agapanthia coeruleipennis Frivaldszky, 1878: Distribution patterns in Turkey. 277. Agapanthia suturalis (Fabricius, 1787): Distribution patterns in Turkey. 278. Agapanthia amitina Holzschuh, 1989: Distribution patterns in Turkey. 279. Agapanthia chalybaea Faldermann, 1837: Distribution patterns in Turkey. 280. Agapanthia frivaldszkyi Ganglbauer, 1884: Distribution patterns in Turkey. 281. Agapanthia lais Reiche & Saulcy, 1858: Distribution patterns in Turkey. 282. Agapanthia osmanlis Reiche et Saulcy, 1858: Distribution patterns in Turkey. 283. Agapanthia violacea (Fabricius, 1775): Distribution patterns in Turkey.
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 121
1
2

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 122
3
4 5
6 7
8 9
10 11

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 123
12 13
14 15
16 17
18 19
20 21
22 23

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 124
24 25
26 27
28 29
30 31
32 33
34 35

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 125
36 37
38 39
40 41
42 43
44 45
46 47

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 126
48 49
50 51
52 53
54 55
56 57
58 59

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 127
60 61
62 63
64 65
66 67
68 69
70 71

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 128
72 73
74 75
76 77
78 79
80 81
82 83

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 129
84 85
86 87
88 89
90 91
92 93
94 95

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 130
96 97
98 99
100 101
102 103
104 105
106 107

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 131
108 109
110 111
112 113
114 115
116 117
118 119

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 132
120 121
122 123
124 125
126 127
128 129
130 131

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 133
132 133
134 135
136 137
138 139
140 141
142 143

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 134
144 145
146 147
148 149
150 151
152 153
154 155

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 135
156 157
158 159
160 161
162 163
164 165
166 167
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 136
168 169
170 171
172 173
174 175
176 177
178 179

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 137
180 181
182 183
184 185
186 187
188 189
190 191

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 138
192 193
194 195
196 197
198 199
200 201
202 203

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 139
204 205
206 207
208 209
210 211
212 213
214 215

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 140
216 217
218 219
220 221
222 223
224 225
226 227

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 141
228 229
230 231
232 233
234 235
236 237
238 239

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 142
240 241
242 243
244 245
246 247
248 249
250 251

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 143
252 253
254 255
256 257
258 259
260 261
262 263

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 144
264 265
266 267
268 269
270 271
272 273
274 275

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 145
276 277
278 279
280 281
282 283

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 146
A NEW SUBSPECIES OF CARABUS (MORPHOCARABUS) AERUGINOSUS FISCHER VON WALDHEIM, 1822
(COLEOPTERA: CARABIDAE) FROM EAST SAJANS MOUNTAINS (EAST SIBERIA)
Dmitry Obydov*
* Timiryazev‘s State Museum of Biology, 123242, Malaya Gruzinskaya str. 15, Moscow, RUSSIA. E-mail: [email protected] [Obydov, D. 2011. A new subspecies of Carabus (Morphocarabus) aeruginosus Fischer von Waldheim, 1822 (Coleoptera: Carabidae) from east Sajans Mountains (East Siberia). Munis Entomology & Zoology, 6 (1): 146-149] ABSTRACT: A new subspecies of Carabus (Morphocarabus) aeruginosus: Carabus (Morphocarabus) aeruginosus pitschugini ssp. n. is described from east Sajans Mountains (East Siberia). Diagnostic data are given. KEY WORDS: Coleoptera, Carabidae, Carabus (Morphocarabus) aeruginosus, new subspecies, Sajans Mountains, East Siberia.
The wide area of Carabus (Morphocarabus) aeruginosus Fischer von Waldheim, 1822 is inhabited by many unequally differing populations. Some of the groups of populations, undoubtedly, must be considered as subspecies. At the same time, the majority of populations constitute a continuous sequence of forms slightly different in size, coloration and ratios, but it is hardly possible to consider these forms as subspecies. At present a considerable number of forms of Carabus aeruginosus are described. The majority of infraspecific names must be treated as synonyms.
The name Carabus aereus Dejean, 1826 was introduced for specimens with orange elytra and indicated individual color variations occur in various localities of Carabus aeruginosus area. It was mentioned by G. Fischer von Waldheim (1825-1828) who considered Carabus aereus as ―variété de l‘aeruginosus‖.
Originally Carabus incertus Motschulsky, 1844 has been described from ―environs du Baical‖, but the locality mentioned on the label of the lectotype (O.L. Kryzhanovskij designation, 1968) is ―Krasnojarsk‖. Both type series and the populations from Krasnoiarsk and Baikal environs (represented in my materials) are conspecific with typical Carabus aeruginosus and must be attributed to the nominotypical subspecies.
Carabus subcostatus Motschulsky, 1850 has been described based on single female from "Sibir. orient". Often the name Carabus subcostatus was wrongly attributed by many authors to other species, most frequently to Carabus aeruginosiformis Breuning, 1932. The holotype female of Carabus subcostatus is preserved in the collection of the Zoological Museum of Moscow Lomonosov State University (Moscow, Russia). It has relatively short elytra with about equally developed, uninterrupted elytral interspaces and small size. But in the shape of pronotum and other features this form is very close to the typical Carabus aeruginosus. I'm sure that Carabus subcostatus is teratological specimen of Carabus aeruginosus.
The species is distributed from about Urals Mountain to the east as far as Yakutia and Baikal Lake and as far to the north as north-eastern regions of European Russia and Yamal Peninsula and mouth of Yenisei River. The species

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 147
occur in the forested areas; in the north part of the area inhabits tundra and forest-tundra.
The species includes four subspecies: Carabus (Morphocarabus) aeruginosus aeruginosus Fischer von Waldheim, 1822, Carabus (Morphocarabus) aeruginosus herrmanni Mannerheim, 1827, Carabus (Morphocarabus) aeruginosus salechardensis Obydov, 1999 and Carabus (Morphocarabus) aeruginosus pitschugini ssp. n.
Recently I have received small series (2 males and female) of Carabus (Morphocarabus) aeruginosus collected in the eastern Sajans, Tunkinskie Goltzy (East Siberia). The collected specimens differ from other subspecies of Carabus (Morphocarabus) aeruginosus by many features, distributed on periphery of a specific area of Carabus aeruginosus and represent a new subspecies, most probably limited to Tunkinskie Goltzy Mountains. A new subspecies of Carabus (Morphocarabus) aeruginosus Fischer von Waldheim, 1822 is described in this work. Diagnostic data are given.
Carabus (Morphocarabus) aeruginosus pitschugini ssp. n. (Figs. 1, 2)
Holotype: male with label: ―East Siberia, Tunkinskie Goltzy (East Sajans), Mondy env., 24.VI.1979, E. Ya. Berlov leg. Papatypes: male and female, same data and same locality. The holotype is preserved in the collection of the State Museum of Biology (Moscow, Russia). The paratypes are preserved in the collection of Nikolai Yu. PICHUGIN (Vladimir, Russia). Description: Body length in males is 20.0 - 22.4 mm (including mandibles), width 7.3 - 8.3 mm; body length in female is 21.0 mm, width 7.8 mm.
Head not thickened; ratio width of pronotum/width of head 2.10; eyes strongly convex; mandibles relatively short and broad, slightly incurved and sharply pointed at the apex; terebral tooth of the right and left mandibles bi-dentate, strongly prominent; retinaculum of right and left mandibles about equally developed, triangular, prominent; surface of mandibles smooth. Frontal furrows relatively deep and shallow, inside smooth or with few coarse wrinkles. Head nearly smooth. Frons, vertex and neck with few coarse wrinkles. Labrum wider than clypeus, strongly notched, with two lateral setae. Antennae protruding beyond the base of pronotum by three-four apical segments; palpi slightly dilated; penultimate segment of the maxillary palpi equal to the last segment; penultimate segment of the labial palpi with two setae. Mentum tooth very short, much shorter than lateral lobes; submentum with two or four setae.
Pronotum convex; broadest at about middle; ratio width/length 1.50. Disk of pronotum with dense coarse punctures, posteriorly pronotum with dense coarse wrinkles; pronotal sculpture less rough laterally. Median longitudinal line distinct; basal foveae big and deep, inside coarsely-wrinkled. Sides of pronotum narrowly margined; lobes of hind angles short, evenly rounded, bent downwards. Lateral margin with two setiferous pores: one pore at about middle and one pore near hind angle.
Elytra short-oval, strongly convex; widest behind middle; shoulders prominent; sides of elytra narrowly margined. Ratio length/width 1.50; ratio width of elytra/width of pronotum 1.32. Elytral sculpture: all elytral interspaces

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 148
slightly convex, about equally developed, primary elytral interspaces interrupted into relatively long and short links, secondary and tertiaries uninterrupted. Primary foveoles indistinct; striae finely punctured.
Metepisternum smooth, longer than its width; abdominal sternites smooth, sides of abdomen finely wrinkled; sternal sulci absent.
Legs of normal length; fore male tarsi with four dilated segments bearing hairy pads.
Shape of aedeagus and endophallic structure (Fig. 3) in general is characteristic for species.
Head and pronotum black, elytra brown, etytral margins black. Mandibles, palpi, antennae, legs, claws and ventral body surface black or dark brown. Differential diagnosis: The new subspecies differs from nominotypical subspecies of Carabus aeruginosus by following features: body much smaller (Fig.4) and more robust (body length in the new subspecies is 20.0 – 22.4 mm; in Carabus aeruginosus aeruginosus – 25.0 – 35.0 mm); head nearly smooth (in Carabus aeruginosus aeruginosus with dense coarse wrinkles); mentum tooth broader and shorter; pronotum more convex; elytra broader, more convex and shorter (ratio length/width is 1.52, while in Carabus aeruginosus aeruginosus this coefficient is 1,58); elytral sculpture less rough, elytral interspaces less convex; primary elytral foveoles less distinct. From other subspecies of Carabus aeruginosus the new subspecies differs by less rough sculpture of head; less rough elytral sculpture and much smaller body size. Distribution: East Siberia, eastern Sajans, Tunkinskie Goltzy Mountains. Etymology: The new subspecies is named in honour of Russian entomologist Mr. Nikolai Yu. PICHUGIN (Vladimir).
ACKNOWLEDGEMENTS
I wish to express my hearty gratitude to Mr. Nikolai Yu. PICHUGIN (Vladimir) who kindly loaned material for studies.
LITERATURE CITED Breuning, S. 1932. Monographie der Gattung Carabus. Bestimmungs-Tabellen der europäischen Coleopteren, 105 Heft. Troppau: 291-496. Dejean, P. F. M. A. 1826. Species général des Coléoptères de la collection de M. le Comte Dejean. Vol. 2, Méquignon-Marvis éd., Paris: 501 pp. Fischer von Waldheim, G. 1820-1822. Entomographia imperii Rossici, Vol. I, Typogr. Acad. Caes. medico-chir. excusa, Mosquae: 104 pp. Ryzhanovskij, O. L. 1968. [On the types of tribes Carabini and Cychrini (Coleoptera, Carabidae), described by V. I. Motschulsky]. Proceedings of the Zoological Museum of Moscow University, T.XI:169-188. [in Russian]. Mannerheim, G. 1827. Supplementa quaedam in genus Carabum systemati entomologico. Hummel's Essais entomologiques, T. I, No. 6: 21-26. Motschulsky, V. 1844. Insectes de la Sibérie, rapportés d‘un voyage fait en 1839 et 1840. Mém. Acad. Imp. Sci. Saint-Pétersbourg, 5 (1,2,3): 274 pp. Motschulsky, V. 1850. Die Käfer Russlands. Moscou: 91 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 149
Obydov, D. 1999. A revision of the ―Carabus aeruginosus‖ species group (Coleoptera, Carabidae). Coléoptères, 5 (9): 149-174. Obydov, D. 2009. Faune des Carabus de Sibérie & d'Extrême-Orient russe - I. Collection systématique, Vol. 20, Magellanes, France: 148 pp.
Figures 1, 2. 1. Carabus (Morphocarabus) aeruginosus pitschugini ssp. n. (holotype). 2. Carabus (Morphocarabus) aeruginosus pitschugini ssp. n. (paratype).
Figures 3, 4. Fig. 3. Aedeagus and endophallus of Carabus (Morphocarabus) aeruginosus pitschugini ssp. n. Fig. 4. Differences in average body size of Carabus aeruginosus: 1. Carabus aeruginosus pitschugini ssp. n.; 2. Carabus aeruginosus aeruginosus (from Barnaul environs).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 150
USING SOME PLANT ESSENTIAL OILS AS NATURAL FUMIGANTS AGAINST ADULTS OF CALLOSOBRUCHUS
MACULATUS (F.) (COLEOPTERA: BRUCHIDAE)
Mohammad Mahmoudvand*, Habib Abbasipour*, Mohammad Hossein Hosseinpour*, Fahimeh Rastegar* and Moslem Basij*
* Department of Plant Protection, College of Agricultural Sciences, Shahed University, Tehran, IRAN. E-mail: [email protected]; [email protected]; [email protected]; [email protected]; [email protected] [Mahmoudvand, M., Abbasipour, H., Hosseinpour, M. H., Rastegar, F. & Basij, M. 2011. Using some Plant Essential Oils as natural fumigants against adults of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Munis Entomology & Zoology, 6 (1): 150-154] ABSTRACT: Present study was aimed to investigate fumigant toxicity of essential oils extracted from Lippia citrodora Kunth., Rosmarinus officinalis L., Mentha piperita L. and Juniperus Sabina L. against adults of cowpea weevil, Callosobruchus maculatus (Col.: Bruchidae). Essential oils were extracted from aerial parts of plants via hydrodistillation. Adults (1-7 days old) of insects were exposed to a series of concentrations of essential oils for 24 h and then the numbers of dead and alive insects were counted. Results showed that essential oil of M. piperita (LC50 7.86 µl/L air) was significantly more toxic than L. citrodora (LC50 187.51 µl/L air), J. Sabina (LC50 134.35 µl/L air) and R. officinalis (LC50 46.81 µl/L air). Results of this investigation suggested that essential oils of these plants could be used as safe fumigants for controlling of C. maculatus in small scales. KEY WORDS: Lippia citrodora, Rosmarinus officinalis, Mentha piperita, Juniperus sabina; Callosobruchus maculatus, natural fumigants, essential oils.
The major storage pest of cowpea is Callosobruchus maculates (F.) (Col.: Bruchidae) (Epidi et al., 2008). It is an important insect pest of many grains such as cowpea, chickpea, lentil and soybean (Mahfuz & Khalequzzaman, 2007). Stored products pests control mainly depends on fumigation by methyl bromide or phosphine (Butler & Rodriguez, 1996). Due to ozone-depleting properties, application of methyl bromide is being restricted (MBTOC, 1998). Insect resistance to phosphine has been reported from many countries and using of this fumigant may become limited (Daghlish & Collins, 1999). Low toxicity to mammals, biodegradability and high volatility of essential oils posses these compounds as alternatives to conventional fumigants (Shaaya et al., 1997; Li & Zou, 2001).
This study was designed to evaluate the fumigant toxicity of essential oils of four plants, Lippia citrodora (Verbenaceae), Rosmarinus officinalis (Lamiaceae), Mentha piperita (Lamiaceae) and Juniperus sabina (Pinaceae) against adults of C. maculatus. L. citrodora, is native to South America and contain a strong lemon-scented volatile oil. R. officinalis is native to Mediterranean and cultivated throughout the world. M. piperita is used to flavour non-alcoholic beverages, ice-creams and candies and finally J. sabina is bitter and has a disagreeable smell and volatile oil. All of these plants have medicinal values (Prajapati et al., 2004).
MATERIALS AND METHODS
Aerial parts of Lippia citrodora, Rosmarinus officinalis, Mentha piperita and
Juniperus sabina were collected from the garden of medicinal plants of Shahed

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 151
University in july, 2009 and then shade dried at room temperature. Dried materials were subjected to hydrodistillation using Clevenger type apparatus. Extracted essential oils were dried via anhydrous sodium sulphate and stored at 4°C in darkness. In all tests pure essential oils were employed.
The cowpea weevil, C. maculatus was reared on mung bean grains in the growth chamber under conditions of 27 ± 1°C and 65 ± 5% R.H and darkness. Adults of insects (1-7 days old) were used for experiments.
The fumigant toxicity of essential oils has been tested in glass vials (70 ml) each of them containing 10 adults of insects. Filter paper disks (Whatman No. 1) were cut into 2cm diameters and were attached to undersurface of screw caps of glass vials. Filter papers were impregnated with series of concentrations of each essential oil. Then the caps were tightly screwed on glass vials. Concentrations used for essential oils of L. citrodora were 114.24, 142.8, 171.36, 256.04 and 285.80 µl/L air, and for R. officinalis oil were 21.42, 42.84, 71.40, 100 and 128.52 µl/L air. Likewise, concentrations used for M. piperita were 4.28, 5.71, 7.14, 10, 14.28, 17.14 and 21.42 µl/L air and finally for J. sabina were 128.57, 157.14, 200, 228.57 and 271.43 µl/L air. Four replicates were run for each concentration and control. After 3, 6, 9, 12 and 24h from the beginning of exposure, numbers of dead and alive insects were counted. There was no mortality in control.
The 50% and 90% lethal concentrations (LC50 and LC90) values were assessed by Probit analysis (Finney, 1971).
RESULTS
The results of LC50 values of various essential oils are shown in Table 1. Results of this research showed that percentage of insect mortality was increased with the concentration and the time of exposure. In the other hand, toxicity depends on concentration, type of essential oil and exposure time. Figs. 1-4 show that in all cases mortality was increased with increasing the concentration and exposure time.
Essential oils of L. citrodora at lowest concentration (114.24 µl/L air) caused 20% mortality in insect population after 24 h compared to 85% mortality at the highest concentration (285.8 µL/l air) (Fig. 1). Lowest concentration of essential oil of R. officinalis (21.42 µl/L air) achieved 14% mortality after 24 h exposure and at the highest concentration (128.52 µl/L air) caused 88% mortality after the same time (Fig. 2). Lowest concentration of essential oil of M. piperita (4.28 µl/L air) killed 5% of insects after 3 h exposure and 7.5%, 20% and 37.5% after 9, 12 and 24 h exposure, respectively (Fig. 3). Exposing C. maculatus adults to 128.57 µl/L air of J. sabina essential oil for 3h caused no mortality and after 6, 12 and 24h exposure caused 2%, 6% and 14% mortality in insect population. Likewise, with increasing the concentration to 271.43 µl/L air, also mortality was increased to 28% after 6 h exposure, 84% and 96% mortality after 12 and 24 h exposure to essential oil of J. sabina, respectively (Fig. 4).
Comparison of the estimated LC50 values of essential oils indicated that M. piperita essential oils is significantly more toxic to C. maculatus than other essential oils which were reported in this study. Furthermore, adults of C. maculatus were highly more tolerant to essential oil of L. citrodora than essential oils of M. piperita, R. officinalis and J. sabina (Table 1).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 152
DISCUSSION Employing the synthetic insecticides remains undesirable effects such as
mammalian toxicity, food chain disruption and increases resistance in insect population (Regnault-Roger, 1997). Natural compounds from plant origins could be efficient alternatives for conventional fumigants because of their low toxicity to mammals, fast degradability properties and regional availability (Rajendran & Sriranjini, 2008). Essential oils of M. piperita and R. officinalis, displayed considerable fumigant toxicity on adults of C. maculatus. In previous investigations demonstrated that essential oils of Artemisia unnua L. (LC50 14.1 µl/L air) and Eucalyptus camodulensis Deh. (LC50 17.77 µl/L air) have toxic effect on adults of C. maculatus (Hosseinpour et al., 2009; Abbasipour et al., 2009). Comparing the estimated LC50 values of present investigation with LC50 values of mentioned essential oils indicated that essential oils of M. piperita is more toxic to C. maculatus than essential oils of A. annua and E. camodulensis. Results of this investigation clearly illustrated that insects varied in their susceptibility to various essential oils that probably referring to insecticidal ability of their active constituents. Abbasipour et al (2009) reported that essential oils of Rosemarinus officinalis possess toxic effect in gaseous phase against adults of Tribolium castaneum (LC50 103.28 µl/L air). The noticeable signification between the LC50 values of T. castaneum and C. maculatus can be of their differences in morphology, physiology, sensitivity and other factors.
Results of this study suggested that essential oils of L. citrodora, R. officinalis, M. piperita and J. sabina plants are good choice for controlling this pest. Also, we need some additional studies for formulating and improving methods of application of them.
LITERATURE CITED Abbasipour, H., Mahmoudvand, M., Deylami, A. & Hosseinpour, M. H. 2009. Fumigant toxicity of essential oils of Rosemarinus officinalis L. and Eucalyptus camodulensis Deh. against some stored products pests, Proceeding of the 6th Asia-Pasific Congress of Entomology, Entomology in Health, Agriculture and Environment, Beijing, China. Butler, J. H., & Rodriguea, J. M. 1996. Methyl bromide in the atmosphere. In: Bell, C. H., Price, N & Chakrabarti, B. (Eds), The methyl bromide issue Vol. 1, Wiley, West Sussex, England, pp 27-90. Daglish, G. J. & Collins, P. J. 1999. Improving the relevance of assays for phosphine resistance. In: Jin, X., Liang, Q., Liang, Y.S., Tan, X.C., Guan, L.H. (Eds.), Stored Product Protection. Proceedings of the 7th International Working Conference on Stored-Product Protection, October 1998, Beijing, China. Sichuan Publishing House of Science & Technology, Chengdu, China, pp. 584–593. Epidi, T. T., Nwani C. D. & Udoh S. 2008. Efficacy of some Plant Species for the Control of Cowpea Weevil (Callosobruchus maculatus) and Maize Weevil (Sitophilus zeamais). Inernational Journal of Agricultural Biology, 10: 588-590. Finney, D. J. 1971. Probit Analysis, 3th Edition. Cambridge University, London. 333 pp. Hosseinpour, M. H., Askarianzadeh, A., Moharramipour, S. & Jalali Sendi, J. 2009. Fumigant toxicity of essential oils of Artemisia unnua L. (Asteraceae) against two stored products coleopteran insect pests, Proceeding of the 6th Asia-Pasific Congress of Entomology Entomology in Health, Agriculture and Environment, Beijing, China. Li, Y. S., & Zou, H. Y. 2001. Insecticidal activity of extracts from Eupatorium adenophorum against four stored grain insects. Entomological Knowledge, 38: 214–216. Mahfuz, I. & Khalequzzaman M. 2007. Contact and Fumigant Toxicity of Essential Oils Against Callosobruchus maculatus. Journal of Zoology Rajshahi University, 26: 63-66.
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 153
MBTOC. 2002. Report of the Methyl Bromide Technical Options Committee (MBTOC) 2002 Assessment. UNEP, Nairobi, Kenya. Prajaparti, N. D., Purohit, S. S., Sharma, A. K. & Kumar, T. 2004. A Hand Book of Medicinal Plants, Agrobios, India, 1200 pp. Rajendran, S. & Sriranjini, V. 2008. Plant products as fumigants for stored-product insect control. Journal of Stored Products Research, 44: 126-35. Regnault-Roger, C. 1997. The potential of botanical essential oils for insect pest control. Integrated Pest Management Reviews, 2: 25-34. Shaaya, E., Kostjukovski, M., Eilberg, J. & Sukprakarn, C. 1997. Plant oils as fumigants and contact insecticides for the control of stored-product insects. Journal of Stored Products Research, 33: 7–15. Table 1. Fumigant toxicity of L. citrodora, R. officinalis, M. piperita and J. sabina essential oils against C. maculatus. Essential oils na LC50
b (µl/L air)
LC90b
(µl/L air) Slope±SE Df Chi square
(χ2)
Lippia citrodora 200
187.51 (171.33-206.05)
338.65 (290.74-433.71)
4.99±0.68 3 3.96
Rosmarinus officinalis
200
46.81 (39.27-54.22)
135.76 (110-186.07)
2.77±0.34 3 1.34
Mentha piperita 280
7.86 (5.90-9.66)
37.76 (25.02-90.39)
1.88±0.36 5 1.28
Juniperus sabina 200
134.35 (113.86-149.35)
258.51 (228-320.17)
4.50±0.73 3 3.82
na: number of insects b 95% lower and upper fiducially limits are shown in parenthesis.
Figure 1. Mortality of adults of C. maculatus exposed to L. citrodora essential oil in different concentrations and times.
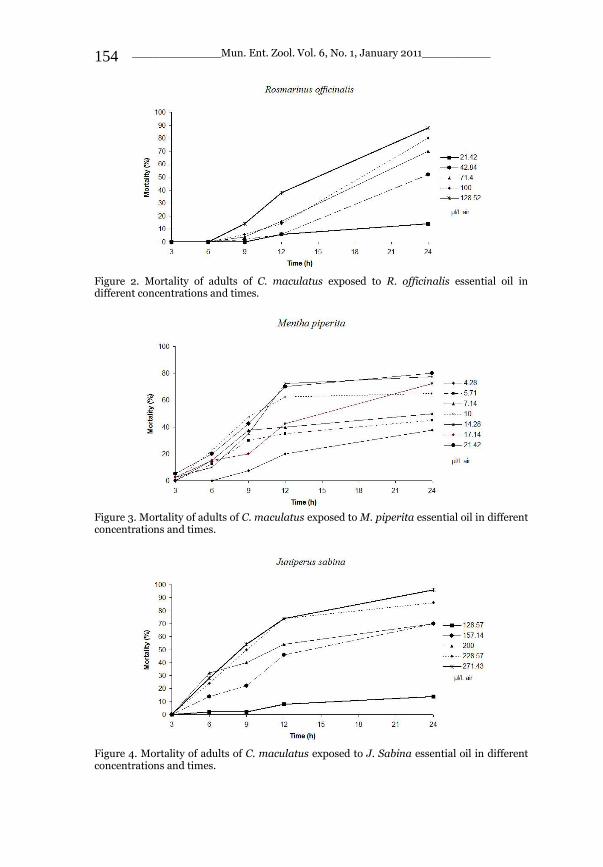
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 154
Figure 2. Mortality of adults of C. maculatus exposed to R. officinalis essential oil in different concentrations and times.
Figure 3. Mortality of adults of C. maculatus exposed to M. piperita essential oil in different concentrations and times.
Figure 4. Mortality of adults of C. maculatus exposed to J. Sabina essential oil in different concentrations and times.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 155
RECENT RECORDS OF RARE AND NEW FOR UKRAINIAN CARPATHIANS SPECIES OF LONGHORN BEETLES
(INSECTA: COLEOPTERA: CERAMBYCIDAE) WITH NOTES ON THEIR DISTRIBUTION
Andrew M. Zamoroka* and Ruslan Yu. Panin**
* Pre-Carpathian national university named after Vasyl Stefanyk, T. Shevchenko str., 57, Ivano-Frankivsk, UKRAINE. E-mail: [email protected] ** Lviv department of Ukrainian Entomological Society at State Museum of Natural History, Teatralna str., 18, Lviv, UKRAINE. E-mail: [email protected]
[Zamoroka, A. M. & Panin, R. Yu. 2011. Recent records of rare and new for Ukrainian Carpathians species of Longhorn beetles (Insecta: Coleoptera: Cerambycidae) with notes on their distribution. Munis Entomology & Zoology, 6 (1): 155-165] ABSTRACT: We listed 11 species of long horn beetles that are rare or new in Carpathians region of Ukraine. These species were recorded during the years 2008-2009 and include new for Ukrainian fauna species Stenopterus flavicornis, Trichoferus campestris, Trichoferus pallidus and Agapanthia intermedia. We also listed species Leptura aurulenta, Batesiata tesserula, Paracorymbia fulva, Dorcadion aethiops and Phytoecia tigrina that have not been recorded for the past 50-110 years. The endangered and listed in Ukrainian Red Data Book species Rosalia alpina and Aromia moschata were also recorded. The areal distribution of all species is described. The Key to Insect taxa of Trichoferus and Stenopterus genera in Ukraine is presented. KEY WORDS: Cerambycidae, Ukrainian Carpathians, Rare species.
The long horned beetles family (Coleoptera: Cerambycidae) is one of the five largest families of Coleoptera that consists of 25000 species found worldwide (Sama and al., 2010). They occupy various habitats and play an important role in natural ecosystems. The long horn beetles are the first level consumers and essential components of food chains of the forest ecosystems. As woodboring insects they are potential pests, and many of them use dead trees of deciduous and coniferous species (Zamoroka, 2009).
To date, over 625 species of Cerambycidae are found in Europe (Althoff and Danilevsky, 1997). This is the most studied Coleoptera family in Europe and Ukraine. According to recent studies, there are 275 (Zahaykevych, 1991) or 280 (Bartenev, 2004) species of long horned beetles found in Ukraine.
The long horn beetles studies in Ukrainian Carpathians were initiated in the middle of the XIX century by Marian Lomnicki, who presented the first detailed faunistic description of Cerambycidae. Based on his studies conducted in various parts of Ukrainian and Polish Carpathians and Halicia, M. Lomnicki compiled and published the first list of 132 long horned beetle species found in the area (Lomnicki, 1884). These studies were interrupted in the first part of the XX century and continued after the Second World War in 1950s. During the second part of the XX century Ivan Zhaykevych continued faunistic studies of Cerambycidae and listed 150 species in Ukrainian Carpathians (Zahaykevych, 1991). This list was further expanded by Zamoroka (2006) and now includes 160 species. Nevertheless, the recent studies indicate that this list is far from being completed (Zamoroka, 2009).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 156
METHODS
The Cerambycids were collected during the years 2008-2009 in Ukrainian Carpathians Mountains including adjoining Pannonia and Pre-Сarpathia plains. Multiple insect collection methods were applied. These included collecting insects on forage plants, on withered, felled and damaged coniferous and broadleaf trees, shaking tree branches, using entomological sweep-net, Berber's soil traps (4% formaldehyde) and light traps (ethyl acetate). Insects were identified using the Key to Insect Orders "Beetles of Central Europe"(Freude H. and al, 1966).
Abbreviations: surr. – surround; loc. – locality; vlg. – village; twn. – town; dstr. – district; reg. – region; coll. – collector; SMNH – State Museum of Natural History (Lviv).
RESULTS AND DISCUSSIONS
According to the recent faunistic publications, there are about 160 species of long horned beetles found in Ukrainian Carpathians (Zahaykevych, 1991, Zamoroka, 2009). Some of these species are very rare and represented by single specimens. Many of these species were recorded last time more than 50 years ago.
In this paper we present the recent record of 11 long horned beetle species that comprise 10 genera, 8 tribes and 3 subfamilies.
The list of recorded rare long horn beetle species found in Ukrainian Carpathians is given below.
Family Cerambycidae Latreille, 1802 Subfamily Lepturinae Latreille, 1802
Tribe Lepturini Latreille, 1804
Leptura aurulenta Fabricius, 1793 Material examined: 2 females 17.VII.2009, Carpathian Biosphere Reserve, "Kuziy" loc., Luh vlg. surr., Rakhiv dstr., Transcarpathian reg., Ukraine, coll. Yuriy Heriak.
The last known data collection for L. aurulenta in Ukrainian Carpathians was performed in Perechyn twn. surr., Transcarpathian reg by I. Zahaykevych in 1957 .To date, there are only few known records of L. aurulenta in Ukrainian Carpathians. Thus, our record is the first publication after 52 years.
L. aurulenta is found in Ukraine in Carpathians (Zahaykevych, 1991), Chernihiv and Kharkiv reg. (Bartenev, 2004).
L. aurulenta is distributed throughout Europe excluding its north part, and it is found in some parts of North Africa including Algeria (Rosa, 2003). In Europe L. aurulenta was recorded in Portugal (Althoff and Danilevsky, 1997), Spain (Rosa, 2003), Britain (Rejzek, 2004), France (Brustel and al, 2002), Luxemburg, Germany (Althoff and Danilevsky, 1997), Austria (Adlbauer, 2001), Czech Republic, Poland (Althoff and Danilevsky, 1997), Ukraine (Zahaykevych, 1991, Bartenev, 2004), Slovakia (Holekova, 1996), Hungary (Csóka and Kovács, 1999), Moldova (Baban, 2006), Romania, Bulgaria, Greece, Albany, Bosnia and Herzegovina, Italy (Althoff and Danilevsky, 1997), Serbia (Pil, 2005), Slovenia (Brelih and al, 2006), and West Russia (Mamontov, 2009).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 157
Batesiata tesserula (Charpentier, 1825) Material examined: 1 female, 17.VII.2009, loc. "Kuziy", Carpathian Biosphere Reserve, Luh vlg. surr., Rakhiv dstr., Transcarpathian reg., Ukraine, coll. Yuriy Heriak.
This is a new record for Carpathian Biosphere Reserve. B. tesserula is found in old beech forests of Reserve. In Ukraine B. tesserula is encountered only in Carpathians, especially in Trancarpathian part.
B. tesserula is distributed in Central and South Europe, Asia Minor, Caucasus and Cental Asia. In Europe it occupies areals in Czech Republic (Szopa, 2002), Ukraine (Zahaykevych, 1991), Romania (Serafim, 2006), Slovakia, Poland, Hungary, Bulgaria, Greece, Albany, and Serbia (Althoff and Danilevsky, 1997). According to Ozdikmen H. (2008) B. tesserula also occurs in Turkey (Asia Minor). Also, it was recorded in Georgia, Azerbaijan and Iran (Bartenev, 2004).
Paracorymbia fulva (De Geer, 1775) Material examined: 1 female, 13.VII.2008, loc. "Chorna Hora", Vynohradiv twn. surr., Trancarpathian reg., Ukraine, coll. Ruslan Panin.
This record of P. fulva in Ukrainian Carpathians is the first record in Ukraine after 80 years. Our record also established a new locality of this species. Prior to our record, P. fulva was known only in two localities: Pasika vlg. surr. (Svalyava dstr., Transcarpathian reg.) and unidentified location named "Osa" (Transcapathian reg.) without existing administrative authorities (Zahaykevych, 1961). Generally, P. fulva is distributed very sporadically in Ukraine. It was recorded in Kharkiv reg. (Bartenev, 2004), Khmelnytsk reg. (Sokolivka vlg. surr., Yarmolynets dstr.) and Transcarpathian reg. (Zahaykevych, 1961).
In Europe P. fulva is widely distributed with the exception of North Europe. It is found in Portugal (Althoff and Danilevsky, 1997), Spain (Pena, 2007 а б), Britain (Rejzek, 2004), France (Brustel and al, 2002), Germany, Czech Republic, Slovakia, Poland, Ukraine (Zahaykevych, 1961, Bartenev, 2004), Hungary (Kovács and al, 2001), Romania (Serafim, 2006), Bulgaria (Serafim, 2006) Greece, Albany, Serbia, Italy (Althoff and Danilevsky, 1997), Slovenia (Brelih and al, 2006), South-Western Russia (Althoff and Danilevsky, 1997, Bartenev, 2004). It was also recorded in Asia Minor, namely the Black Sea region of Turkey (Ozdikmen, 2007).
Subfamily Cerambycinae Latreille, 1802 Tribe Hesperophanini Mulsant, 1839
Trichoferus campestris (Faldermann, 1835)
Material examined: 1 male, 27.VII.2009, Ivano-Frankivsk city (Doroshenka str., 18a), Ukraine, coll. Andrew Zamoroka, collected in the evening using light trap. 1 female, 11.IX.2009, Kalush twn. Ivano-Frankivsk reg., Ukraine, collected in the City Park in evening using light trap, coll. Andrew Zamoroka.
Previously we reported T. campestris as a new species for Ukrainian fauna (Zamoroka 2009). This conclusion was based on the review of Longhorn beetles of Ukraine by A. Bartenev (2004). Unfortunately, the publication of V. Terekhova and A. Bartenev (2007) where authors describe the distribution of T. campestris in Eastern Ukraine was not available for us at the time. According to V. Terekhova

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 158
and A. Bartenev (2007), while T. campestris was first recorded in 1998, it was correctly identified only in 2006.
In Ukrainian Carpathians T. campestris was previously collected in Kryvets vlg. surr. (Bohorodchany dstr., Ivano-Frankivsk reg.) (Zamoroka, 2009). These records of T. campestris are novel for Carpathians. Therefore, it should be considered that T. campestris is more widely distributed in Ukrainian Carpathians and Central Europe than it was previously thought. It was collected from the following localities in Ukraine: Sevastopol (Zamoroka, 2009), Evpatoria (Crimean Autonomous Republic), Odessa, Donetsk, Kharkiv, Dyakove (Luhansk reg.), Natural Reserve "Kam'yani Mohyly" (Volodarske dtr., Donetsk reg.), National park "Homilshanski Lisy" (Zmiiv dst., Kharkiv reg.) (Terkhova, 2007), and Ivano-Frankivsk reg. (Zamoroka, 2009).
Originally, T. campestris was found in Transcaucasia, Central Asia, Mongolia, Korea, North China and Japan (Lobanov, 1982). Nowadays, it is found from Far East to Carpathians, and it is also reported in Canada and USA (Zamoroka, 2009, Grebennikov, and Gill, 2010). T. campestris is spreading to west of Europe.
Trichoferus pallidus (Oliver, 1790) Material examined: 2 males, 20.VII.2009, Berehove twn. surr., Transcarpathian reg., Ukraine, coll. Yuriy Heriak, collected under bark of oak.
Our finding of T. pallidus is the first record in Ukrainian Carpathians and the first certain record in Ukrainian fauna after 110 years. The only known record of this species is from 1900, when T. pallidus was found in surrounding Alushta area (Crimean peninsula) (Bartenev, 2004). In our previous publication (Zamoroka, 2009) we predicted presence of T. pallidus in Transcarpathian plain.
Since T. pallidus is very rare species in Ukraine and it is already included to the Red Lists of neighboring countries such as Poland (Pawłowski and all., 2002) and Slovakia (Baláţ, 2001, Jendek, 2006), we suggest to include T. pallidus to the fourth edition of the Red Book of Ukraine.
In Europe T. pallidus is found in Portugal (Grosso-Silva, 2007), Spain (Pena and all, 2007), France (Brustel and all, 2002), Austria (Adlbauer, 2002), south of Germany, south of Poland (Gutowski, 1986), Czech Republic, Slovakia (Sláma, 1998), Hungary (Kovács and all, 2001), Slovenia (Brelih and all, 2006) Bulgaria, Greece, Romania, and Caucasian coast of Russia (Althoff and Danilevsky, 1997).
Since T. pallidus and T. campestris were not included to the Key to Insect Orders of Trichoferus species in Ukraine, we present the respective Key to Insect Orders below.
The key for identification of species of Trichoferus Wollaston, 1854 in Ukraine
1 (6) Elytra is covered by long upstanding hairs.………………..……………………………………………2 2 (3) The elytra‘s hair coating is not regular, without hairy maculates. The elytra are luster. The long upstanding hairs cover the second half of the elytra. Main coloration of body is fulvous. The body length is 11-20 mm. It is distributed throughout all Ukraine…………………… …………………………………………………………………………. T. campestris (Faldermann, 1835) 3 (2) The hairy coating of elytra is thick and maculated. Long upstanding hairs cover entire surface of elytra …………………………………………………………………………………………………………..4 4 (5) The small flat parts of elytra occupies the area around the shield. Hair on maculae creates irregular pattern. Two distinct hair maculae are present on the pronotum's disk of

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 159
males. General body color is from reddish to fulvous. The body length is 8-16 mm. It is distributed in Crimean peninsula…………………………………………………………………………………… ………………………………….…….T. fasciculatus subsp fasciculatus (Faldermann, 1837) 5 (4) The small flat parts of elytra around the shield are absent. Hair coating is regular without maculated pattern. The main body‘s color is from fulvous to black. The body length is 9-20 mm. It is distributed in Crimea peninsula.……………...T. griseus (Fabricius, 1792) 6 (1) Elytra are not covered by long upstanding hairs. Elytra‘s hair coating is very thick……..7 7 (8) The hairy coating color is yellowish. There are two light and one dark stripes on elytra. Two light stripes are crossed on the second third of elytra and then are hammed by dark stripe. Antenna, legs and body are fulvous to black colored. The body length is 14-21 mm. It is distributed in Crimean peninsula and Transcarpathians lowlands………………………………… ……………………………………………………………………..………………… T. pallidus (Olivier, 1790) 8 (7) The light stripes on elytra are absent. The elytra‘s hair coating is yellowish-fulvous with distinct maculae. The elytra's surface is unequal under hair. The body color is fulvous to black. The body length is 14-22 mm. It is distributed in South and East Ukraine………………… …………………………………………………………………………………….. T. holosericeus Rossi, 1790
Tribe Compsocerini Thomson, 1864
Rosalia alpina (Linnaeus, 1758)
Material examined: 1 male, 11.VІІ.2009, loc. "Ilma", Natural Reserve "Gorgany", Zelena vlg. surr., Nadvirna dstr., Ivano-Frankivsk reg. Ukraine, coll. Volodymyr Tretiak.
R. alpina is endangered species. It is included to the Red List of IUCN as vulnerable (VU) species, to the Red Data Book of Ukraine (VU) (Rizun and al., 2000, Akimov, 2009), Poland (EN) (Pawłowski, 2002), Russia (2-d category = EN) (Dezhkin, 2001), Sweden (RE) (Gärdenfors, 2005) et cet.
Our record of R. alpina show new location of the species in Ukrainian Carpathians, where its distribution is very spotted. While in Transcarpathians it has continuous distribution range in beech forests, in Pre-Carpathians it is only found in few localities. These include samples from areas surrounding the towns of Yaremche (Tymochko and Kyselyuk, 2005), Bolekhiv, Scole and city of Lviv (data of SMNH collection). In general, R. alpina is found in Carpathians and Crimea in Ukraine (Zahaykevych, 1974). Single records are known from Kyiv, Cherkasy (Rizun and al., 2000), Volynia and Kharkiv (Bartenev, 2004).
R. alpina is widely spread in Europe, Caucasus, Asia Minor (Ozdikmen, 2007 black sea), Near East (Bartenev, 2004, Ozdikmen, 2007 black sea) and North Africa. In Europe it is found in Spain (Pena, 2002, 2007), France (Brustel and al, 2002, Dutto, 2005), Switzerland (Althoff and Danilevsky, 1997), Austria (Adlbauer, 2001), Hungary (Csóka and Kovács, 1999), Poland (Pawłowski, 2002), Czech Republic, Slovakia (Sláma M.E.F. 1998, Brelih and al, 2006), Ukraine (Zahaykevych, 1974, Bartenev, 2004), Romania, Bulgaria, Greece, Albany (Althoff and Danilevsky, 1997), Slovenia (Jurc, 2008), Serbia (Pil, 2005), Italy (Vitali, 1999, Dutto, 2005), Moldova (Baban, 2006), and Western Russia (Bartenev, 2004, Althoff and Danilevsky, 1997). According to IUCN R. alpina is distributed in Iran and in several countries of North Africa such as Libia, Algeria, Morocco. R. alpina was excluded from the long horn beetle species list of Israel (Sama, 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 160
Tribe Callichromatini Blanchard, 1845
Aromia moschata (Linnaeus, 1758) Material examined: 1 female, 27.VІІ.2009, Halych National Park, Shevchenkove vlg. surr., Halych dstr., Ivano-Frankivsk reg., Ukraine, coll. Volodymyr Buchko.
A. moschata is included to the Red Data Book of Ukraine (VU) (Rizun and al., 2000, Akimov, 2009).
Our finding shows a new location of A. moschata in Ukrainian Carpathians and represents the first record for Halych National Park. In Ukrainian Carpathians A. moschata was recorded from few localities only. These localities are found in areas surrounding Ivano-Frankivsk region such as the towns of Verkhovyna and Kolomya, villages of Novosilky (Nadvirna dstr.) (Rizun and al., 2000), Zelena (Nadvirna dstr.) and Biloberizka (Verkhovyna dstr) (Zamoroka, 2007), and city of Ivano-Frankivsk (Lomnicki, 1885, Zamoroka, 2007). In Lviv region A. moschata was known from SMNH collections from the following localities: village Shehyni (Mostysky dstr.), towns Truskawec (Drohobych dstr.) and Sambir, city of Lviv. In general, A. moschata is very abundant along rivers in Ukraine (Bartenev, 2004).
A. moschata is distributed in all palearctic regions and occupies the territory from the Atlantic to the Pacific ocean. The two subspecies are found in Europe. These are A. moschata moschata (Linnaeus, 1758) and A. moschata ambrosiaca (Steven, 1809). A. moschata moschata occupies most of Europe including Britain, France, Germany (Althoff and Danilevsky, 1997), Poland, Czech Republic, Austria, Slovakia (Adlbauer, 2001), Ukraine (Zahaykevych, 1991, Bartenev, 2004, Zamoroka, 2007), Hungary (Csóka and Kovács, 1999), Moldova (Baban, 2006), Byelorussia, Russia, Romania, Bulgaria, Albany (Althoff and Danilevsky, 1997), Serbia (Pil, 2005), Slovenia, Northern Italy (Brelih and al, 2006), and Northern Spain (Pena, 2002). It is also found in Caucasus, Kazakhstan and Turkey. A. moschata ambrosiaca occupies countries of Mediterranean basin including Portugal, Spain (Pena, 2002), Italy, Greece (Althoff and Danilevsky, 1997) and Southern France (Brustel and al, 2002).
Tribe Stenopterini Fairmaire, 1868
Stenopterus flavicornis Kuster, 1846 Material examined: 2 females, 2 males, 13.VII.2008, 1♀ 07.VІІ.2009, loc. "Chorna Hora", Vynohradiv twn. surr., Trancarpathian reg. Ukraine, coll. Ruslan Panin.
S. favicornis is new for Ukrainian fauna species. It was not described in early faunistic publications for Ukrainian territory (Zahaykevych, 1991, Althoff and Danilevsky, 1997, Bartenev, 2004).
S. favicornis belongs to small Mediterranean genera Stenopterus Illiger, 1804, which includes 8 species. It is found from North Africa (Sama G. et all, 2005) and Southern Europe (H. Brustel et all, 2002) to Near East (Ozdikmen H., 2007) and Caucasus (Althoff J., Danilevsky M. L., 1997). The seven species that are distributed in Europe are S. ater Linnaeus, 1767, S. rufus (Linnaeus, 1767), S. atricornis Pic, 1891, S. similatus Holzschuh, 1979, S. creticus Sama, 1995, S. mauritanicus Lucas, 1846, and S. favicornis (Althoff J., Danilevsky M. L., 1997, Sama G.and all, 2005). The S. kraatzi (Pic, 1892) was found only in Asia Minor and Near East (Ozdikmen H., 2007).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 161
S. favicornis is widely distributed in Central and South-East Europe. It occupies South-East France (H. Brustel et all, 2002), South Germany, Austria (Adlbauer, 2001), lowlands of Czech Republic, Slovakia, Hungary, Romania, Bulgaria, Balkans and North-East Italy (Althoff J., Danilevsky M. L., 1997). Some locations of S. favicornis in Turkey and Israel were misidentified and belong to S. kraatzi (H. Brustel et all, 2002).
Since S. favicornis is new species for Ukrainian fauna, we present the Key to Insect Orders of Stenopterus species below.
The key for identification of species of Stenopterus Illiger, 1804 in Ukraine
1 (2) The two small naked and luster snags are presented on the pronotum disk. The light hairy stripes on the top and base of pronotum are tore in the middle. The elytra are fulvous with black bases. The legs and antenna are reddish. The body length is 10-15 mm. It is distributed in Trascarpathian plains…………………………………..S. flavicornis Kuster, 1846 2 (1) The three small naked and luster snags are presented on the pronotum disk. Two of them located on the both sides from the medial line, and the third snag is laid on the middle below the two first………………………………………………………………………………………………………..3 3 (4) The deep furrow is found on the external side of the antenna's first segment. The body color differs. Females are black; in rare cases only elytra are particularly reddish. Males are also black. Elytra, antenna and lags are reddish with the black colored tops and bases. Sometimes they are black too. The body length is 6-16 mm. it is distributed in Crimea peninsula…………………………………………..…………………………………… S. ater Linnaeus, 1767 4 (3) The first segment of antenna is without furrow. Males and females are the same color. Elytra, legs and antenna are reddish with the black top and base parts. The body length is 6-16 mm. It is distributed in steppe and forest-steppe biomes of Ukraine……………………………… ………………………………………………………………………………………….S. rufus (Linnaeus, 1767)
Subfamily Lamiinae Latreille, 1825 Tribe Dorcadiini Latreille, 1825
Dorcadion aethiops (Scopoli, 1763)
Material examined: 2 females 15.V.2009 Muzhieve vlg. surr., Trancarpathian reg., Ukraine, coll. Yuriy Heriak.
This is the first after 49 years record in Ukrainian Carpathians. Here D. aethiops is known in few localities near Berehove twn. and Vynohradiv twn. (Zahaykevych, 1961). In Ukraine D. aethiops is found only in Transcarpathia and in low basin of Dnister river (Odessa reg.) (Bartenev, 2004).
D. aethiops is distributed only in Central and South-Eastern Europe including Switzerland (Ozdikmen, 2008), Southern Germany, Austria, Czech Republic, Slovakia (Althoff and Danilevsky, 1997), Ukraine (Zahaykevych, 1961, 1991, Bartenev, 2004), Hungary (Kovács and al, 2001), Moldova (Baban, 2006), Romania (Rozner, 2007), Serbia (Pil, 2005), Slovenia (Brelih and al, 2006), Bulgaria, Croatia, Bosnia and Herzegovina, Macedonia, Albany, Greece (Althoff and Danilevsky, 1997), and European part of Turkey (Ozdikmen, 2008).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 162
Tribe Agapanthiini Mulsant, 1839
Agapanthia intermedia Ganglbauer, 1883 Material examined: 1 male, 18.V.2009, loc. "Kasova Hora", Halych National Park, Burshtyn twn. surr., Halych dstr., Ivano-Frankivsk reg., Ukraine, col. Andriy Zamoroka, by butterfly net; 1 male, 13.VІ.2009, loc. "Kamin'", Halych National Park, Mezhyhirtsi vlg. surr., Halych dstr., Ivano-Frankivsk reg., Ukraine, col. Andrew Zamoroka, collected in soil trap.
This is new species for Carpathian region of Ukraine. Distribution of A. intermedia in Ukraine is unclear. It is known from Ukrainian long horn beetles review of O. Bartenev (2004), unfortunately, these studies do not describe the precise location of the species.
In Europe A. intermedia is widely distributed with exception of the northern part. It is found in Netherlands, Germany, France (Brustel and al, 2002, Brelih and al, 2006), Switzerland, Austria (Adlbauer, 2001, Steiner, 1999, Brelih and al, 2006), Czech republic, Slovakia (Brelih and al, 2006), Poland, Ukraine (Bartenev, 2004), Hungary (Kovács and Hegyessy, 1997), Italy, Slovenia (Brelih and al, 2006), Romania, Moldova, Byelorussia, Lithuania, and Russia (Althoff and Danilevsky, 1997).
Tribe Saperdini Mulsant, 1839
Phytoecia tigrina Mulsant, 1851 Material examined: 3 females, 2 males, 07.V.2009, 1female, 1 male, 18.V.2009, loc. "Kasova Hora", Halych National Park, Burshtyn twn. surr., Halych dstr., Ivano-Frankivsk reg., Ukraine, col. Andrew Zamoroka, collected on stems of Anchusa L.
Our findings represent the first reliable record of Ph. tigrina in Western Podillya and Carpathians regions after 80 and 41 years, respectively. There are two known localities of Ph. tigrina in Ukraine. The first locality "Chorna Hora" is near Vynohradiv town in Transcarpathian (Zahaykevych, 1961). The second locality "Kasova Hora" is situated in Western Podillya near Burshtyn town in Ivano-Frankivsk region. The two specimens collected in 1929 in "Kasova Hora" are now deposited in funds of SMNH collection (Lviv). Their authenticity, however, was doubted and they were never published. Now we confirm distribution Ph. tigrina in Western Podillya and Pre-Carpathians. In our opinion, the locality "Kasova Hora" is the most northern point of the distribution of Ph. tigrina in Europe (49°13 ́northern latitude and 24°41 ́eastern longitude).
Ph. tigrina is found in Europe including Hungary, Slovakia (Holzshuh, 1984), Ukraine (Zahaykevych, 1961), Romania, Bulgaria, Balkans (Holzshuh, 1984); Asia Minor: western Turkey (Ozdikmen, 2004, 2008); Near East (Ozdikmen, 2008) and Caucasus (Bartenev, 2004, Ozdikmen, 2008). The presence of Ph. tigrina in Southern France is not clear (Brustel and al, 2002).
ACKNOWLEDGEMENTS
We wish to thank Yuriy Heriak, Volodymyr Tretiak and Volodymyr Buchko for helping to collect material in the field. Especially thanks to Dr. Alexander Boyko for text correcting.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 163
LITERATURE CITED Adlbauer, K. & Holzer, E. 2002. Vadonia unipunctata (F.) und Trichoferus pallidus (Ol.) – zwei für die Steiermark neue Bockkäfer (Coleoptera, Cerambycidae). Joannea Zool., 4: 83-85. Adlbauer, K. 2001. Nachtrag zur Bockkäferfauna der Steiermark unter dem Aspekt der Artenbedrohung (Coleoptera, Cerambycidae). Joannea Zool., 3: 83–104. Akimov, A. I. (eds.) 2009. Red book of Ukraine. Animals. Kyiv. 624 pp. Althoff, J. & Danilevsky, M. L. 1997. Acheck-list of longicorn beetles (Coleoptera, Cerambycoidae) of Europe. Slovensko entomolosko drustvo Stefana Michielija, Ljubljana, 164. Baban, E. 2006. Diversitatea coleopterelor (Coleoptera: Carabidae, Silphidae, Scarabaeidae, Cerambycidae) din ecosistemele forestiere ale podiĢului Moldovei centrale. Teză de doctor în biologie, 03.00.09, 136 pp. Baláž, D., Marhold, K. & Urban, P. (eds) 2001. Červený zoznam rastlín a ţivočíchov Slovenska. Ochrana prírody, 20, Suplement: 1–160. Bartenev, A. F. 2003. Review of longhorned beetles (Coleoptera, Cerambyciae) of Ukraine. News of Kharkiv Entomological Society, 11, 1-2: 24-43 (in Russian lang.). Bense, U. 1995. Longhorn beetles. Illustrated key to the Cerambycidae and Vesperidae of Europe. Margraf Verlag, Weikersheim, 512 pp. Brelih, S., Drovenik, B. & Pirnat, A. 2006. Gradivo za favno hroščev (Coleoptera) Slovenije, 2. prispevek, Polyphaga: Chrysomeloidea (= Phytophaga): Cerambycidae. Scopolia, 58: 1-351. Brustel, H., Berger, P. & Cocquempot, C. 2002. Catalogue des Vesperidae et des Cerambycidae de la faune de France (Coleoptera). Аnn. Soc. entomol. Fr.(n.s.), 38 (4): 443-461. Csóka, G. & Kovács, T. 1999. Xilofág rovarok – Xylophagous insects. Erdészeti Turományos Intézet, Agroinform Kiadó, 189 pp. Dezhkin, V. V. (ed.) 2001. Red book of Russian Federation: Animals. 862 p. (in Russian lang.). Dutto, M. 2005. Nuove interessanti osservazioni di Rosalia alpina (linnaeus, 1758) nelle Alpi Occidentali (Coleoptera, Cerambycidae). Riv. piem. st. nat., 26: 283-284. Freude, H., Harde, K. W. & Lohse, G. A. 1966. Die Käfer Mitteleuropas. Band 9. Cerambycidae, Chrysomelidae. Krefeld: Goecke & Evers, 296 pp. Gärdenfors, U. (ed.) 2005. Rödlistade arter i Sverige. Uppsala, SE, 496 pp. Grebennikov, V. V. & Gill, B. D. 2010. Trichoferus campestris (Faldermann) (Coleoptera: Cerambycidae), an Asian wood-boring beetle recorded in North America. The Coleopterists Bulletin, 64 (1): 13–20. Grosso-Silva, J. M. 2007. New and interesting beetle (Coleoptera) records from Portugal (5-th note). Boletín Sociedad Entomológica Aragonesa, 1 40: 471−472. Gutowski, J. M. 1986. Trichoferus pallidus (Olivier, 1790) (Col., Cerambycidae) w Polsce. Przegl. zool., 30, 3: 313-317. Holecova, M. & Zach, P. 1996. Prehľad fauny chrobákov (Coleoptera) ţijúcich na duboch na území Slovenska. Folia Faunistica Slovaca, 1: 39-52. Holzshuh, C. 1984. Beschreibung neuer arten aus der unmittelbaren verwandtschaft von Phytoecia (Pilemia) tigrina (Cerambycidae, Col.). Koleopterologische Rundschau, 57: 167-175. Jendek, B. & Jendek, E. 2006. Analýza druhovej ochrany Coleoptera na Slovensku na základe modelovej skupiny fuzáče (Coleoptera, Cerambycidae). Folia faunistica Slovaca, 11 (4): 15-28. Jurc, M., Ogris, N., Pavlin, R. & Borkovic, D. 2008. Forest as a habitat of saproxylic beetles on Natura 2000 sites in Slovenia. Rev. Écol. (Terre Vie), 63: 53-66.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 164
Kovács, T., Hegyessy, G. & Borsos, S. 2001. Checklist of the longhorn beetle fauna of Somogy county (Coleoptera: Cerambycidae). Natura Somogyiensis. Kaposvár, 1: 213-220. Kovács, T. & Hegyessy, G. 1997. A Mátra cincérfaunája (Coleoptera, Cerambycidae). Foliahistorico Naturaliamusei Matraensis, 22: 203–222. Lobanov, A. L., Danilevsky, M. L. & Murzin, S. V. 1982. A systematic list of long horn beetles (Coleoptera, Cerambycidae) of Soviet Union. II. Entomologycal review., LXI, part 2: 252-276. Lomnicki, M. 1875. Chrzaszcze zebrane w okolicy Stanislawowa. Sprawozdanie komisyi fizyjograficznej, T. 20: 154-184. Mamontov, S. N. 2009. Xylophylic coleoptera of forest botanic-geographical district of Tula region. Ph. d. in boil. scien. thesis: 22. Ozdikmen, H. 2007. The longicorn beetles of Turkey (Coleoptera: Cerambycidae) part I – Black Sea region. Mun. Ent. Zool., 2 (2): 179-387. Ozdikmen, H. 2008. The longicorn beetles of Turkey (Coleoptera: Cerambycidae) part II – Marmara region. Mun. Ent. Zool., 3 (1): 7-152. Ozdikmen, H. 2008. The longicorn beetles of Turkey (Coleoptera: Cerambycidae) part III – Aegean region. Mun. Ent. Zool., 3 (1): 355-436. Ozdikmen, H. & Hasbenli, A. 2004. Contribution to the Knowledge of Longhorned Beetles (Coleoptera, Cerambycidae) from Turkey, Subfamily Lamiinae. J. Ent. Res. Soc., 6 (2): 25-49. Pawłowski, J., Kubisz, D. & Mazur, M. 2002. Coleoptera Chrząszcze. In Z. Głowaciński (red.), Czerwona lista zwierząt ginących i zagrożonych w Polsce. Instytut Ochrony Przyrody. PAN: 155. Pena, G. C. F., Vives i Noguera, E. & Zuzarte, A. J. S. 2007. Nuevo catálogo de los Cerambycidae (Coleoptera) de la Península Ibérica, islas Baleares e islas atlanticas: Cana-rias, Açores y Madeira. Monografias S.E.A., 12: 211. Pena, G. C. F., Vives i Noguera E. & Zuzarte, A. J. S. 2007. Addenda et corrigenda al nuevo catálogo de los Cerambycidae (Coleoptera) de la Península Ibérica, islas Baleares e islas atlanticas: Cana-rias, Açores y Madeira (Monografías S.E.A., vol. 12, 2007). Boletín Sociedad Entomológica Aragonesa, n1 40: 595−596. Pena, G. C. F. 2002. Catálogo de los Cerambycidae (Coleoptera) de Aragón. Cat. entomofauna aragon., № 27: 2–44. Pil, N. 2005. Checklist of longhorn beetles (Coleoptera: Cerambycidae) from mt. Fruska Gora. Acta entomologica Serbica, 9/10 (1/2): 105-110. Rejzek, M. 2004. Check-list of Cerambycidae (Col.) of the British Isles. Entomologist's Monthly Magazine, 140: 51-57. Rizun, B. V., Konovalova, I. B. & Yanytsky, T. P. 2000. Rare and vanishing insects in collection of State Museum of Natural History. Lviv. 71 pp. de la Rosa, J. J. 2003. Aportaciones al conocimiento de la fauna de cerambicidos (Coleoptera: Cerambycidae) de la comunidad de Madrid y territorios limitrofes, Bol. S.E.A., № 33: 285-287. Rozner, I. 2007. Data to the Longicorn Beetle Fauna of Transylvania and the Banat (Coleoptera: Cerambycidae). Acta Siculica: 133–141. Sama, G., Ringenbach, J.-C. & Rejzek, M. 2005. A preliminary survey of the Cerambycidae of Libia (Coleoptera). Аnn. Soc. entomol. Fr., 110 (4/5): 439-454. Sama, G., Buse, J., Orbach, E., Friedman, A.-L.-L., Rittner, O. & Chikatunov, V. 2010. A new catalogue of the Cerambycidae (Coleoptera) of Israel with notes on their distribution and host plants. Mun. Ent. Zool., 5 (1): 1-51. Serafim, R. 2006. The catalogue of the palaearctic species of lepturinae (Coleoptera: Cerambycidae) from the patrimony of "Grigore Antipa" National museum of Natural history (Bucharest) (part II). Travaux du Muséum National d‘Histoire Naturelle "Grigore Antipa", Vol. XLIХ.: 203–238.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 165
Sláma, M. E. F. 1998. Tesaříkovití - Cerambycidae České republiky a Slovenské republiky (Brouci – Coleoptera). Vyd. M. Slama, Tisk Tercie, Krhanice: 383 pp. Steiner, S. 1999. Rote Liste der Bockkäfer Kärntens (Coleoptera, Cerambycidae), In Holzinger, W.E., Mildner, P., Rottenburg, T., Wieser, C. (Hrsg.): Rote Listen gefährdeter Tiere Kärntens. Klagenfurt, Naturschutz in Kärnten 15: 269-286. Szopa, R. 2002. Contribution to the knowledge of the longhorned beetles (Coleoptera: Cerambycidae) of the Jablunkovská brázda valley and its close environs. Klapalekiana, 38: 63-83. Terekhova, V. V. & Bartenev, A. F. 2007. New data for distribution and biology of Trichoferus campestris (Faldermann, 1835) (Coleoptera: Cerambycidae) in Ukraine. News of Kharkiv Entomological Society, XIV, 1-2.: 67-68. Tymochko, V. B. & Kyselyuk, A. I. 2005. Recent state of rare species of insects in Carpathian National Park. Conference report. Kyiv: 108-110. Vitali, F. 1999. Nuovi dati corologici per alcune specie di Cerambicidi Italiani. (Coleoptera, Cerambicidae). Annali del Museo civico di storia naturale ―G. Doria‖, vol. VII, № 315: 1-6. Zahaykevych, I. K. 1961. Contribution to knowledge of longhorned beetles (Coleoptera, Cerambyciae) of Ukraine. Proc. of the State Nat. Hist. Museum., 9: 52-59. Zahaykevych, I. K. 1974. The Longhorn beetles. The agricaltural and forest pests, 2: pp. 24-49. Zahaykevych, I. K. 1991. Taxonomy and ecology of longhorned beetles. Kyiv, 420 pp. Zamoroka, A. M. 2005. Influence of urbanization to long horn beetles' fauna (Coleoptera: Cerambycidae) in city of Ivano-Frankivsk. Zoocenosis, 2: 270-271. Zamoroka, A. M. 2007. Long horn beetles (Coleoptera: Cerambycidae) of Ivano-Frankivsk region. Proc. of Precarpathian national university. Biology, 7: 131-132. Zamoroka, A. M. 2009. A new record of longhorned beetle Trichoferus campestris (Faldermann, 1835) (Coleoptera: Cerambycidae) in Ukraine. Proc. of the State Nat. Hist. Museum, 25: 195-206. Zamoroka, A. M. 2009. Ecological features of long horn beetles entomocomplexes (Coleoptera: Cerambycidae) in the forest ecosystems of the north-eastern macroslope of the Ukrainian Carpathians. Ph. d. thesis. Dnipropetrovsk National University, 20 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 166
EFFICIENCY OF INSECT-PROOF SCREENS IN EXCLUDING THREE MAJOR PESTS OF STRAWBEERY
INTO THE GREENHOUSE
Leila Kiani*, Bahram Tafaghodinia**, Mohsen Yazdanian*
and Mohammad Hassan Sarayloo* * Department of Plant Protection, Faculty of Crop Sciences, Gorgan University of Agricultural Sciences and Natural Resources, IRAN. E-mails: [email protected]; [email protected]; [email protected]
** Iranian Research Organization for Science and Technology, Agricultural Institute, Entomology Department, IRAN. E-mail: [email protected]. [Kiani, L., Tafaghodinia, B., Yazdanian, M. & Sarayloo, M. H. 2011. Efficiency of insect-proof screens in excluding three major pests of strawbeery into the greenhouse. Munis Entomology & Zoology, 6 (1): 166-172] ABSTRACT: In recent years, the use of the insect-proof screens in protection of greenhouse crops has been increasingly recognized. In this study, the effectiveness of three types of insect-proof screens for their exclusion of Frankliniella occidentalis, Chaetosiphon fragaefolii and Tetranychus urticae have been evaluated in laboratory conditions (L:D 16:8,
27 2 °C Temp., and 60 10% R.H.). The tested insect-proof screens were the 5×5 (5×5 threads per cm2), 13×23, 30×34 and Agryl p-17. In each cylindrical container, which was sealed by every one of the above mentioned insect-proof screens at one end, 50 individuals were placed. In this experiment, flowers and leaves of strawberry were used as an incentive for the insect to move through the openings of insect-proof screens. The control percent of screens was evaluated by calculating the number of insects in which were existed in the containers after 24 hours. The results of this experiment indicated that the 5×5 insect screen, which is widely used in greenhouses, did not exclude these pests really. Results also indicated that 94% of C. fragaefolii controlled by the 13×23 screen, compared to not efficient of this screen in excluding of F. occidentalis and T. urticae. 100% of C. fragaefolii, 46.66% of F. occidentalis and 13.8% of T. urticae controlled by the 30×34 screen. However, control percent was higher for C. fragaefolii than for F. occidentalis and T. urticae and this was related to the larger size of the strawberry aphid as compared to the western flower thrips and twospotted spider mite. KEY WORDS: Insect-proof screens, Mechanical barrier, Frankliniella occidentalis, Chaetosiphon fragaefolii, Tetranychus urticae, Strawberry, Greenhouse.
Over the past 30 years, western flower thrips, Frankliniella occidentalis (Pergande), has become one of the most prominent agricultural pests throughout the world (Thoeming et al., 2003). F. occidentalis feed by piercing plant cells with their mouthparts and sucking out the contents. The piercing of a flower cell causes the death of single cells, making a bronzed semblance on the strawberry fruit (Obrycki, 2004; Steiner, 2009). Adults and larvae feed in the same way, so both stages contribute to plant damage (Reitz, 2009). The strawberry aphid, C. fragaefolii, can affect yields because it transmits viruses such as the strawberry mild yellow edge virus (SYEV), strawberry crinkle virus (SCV) and strawberry mottle virus (SMV) (Rondon & Cantlliffe, 2004; Cedola & Greco, 2008). Twospotted spider mite, Tetranychus urticae Koch is most economically important pests in strawberries. T. urticae feed on the underside of the leaf, piercing the chloroplast containing palisade and spongy parenchyma cells in the mesophyll layer at a rate of 18-22 cells/min (Rondon et al., 2005; Fraulo et al., 2009).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 167
The use of the insect-proof screens in protection of greenhouse crops has been increasingly recognized in recent years (Bartzanas et al,. 2002; Bailey et al., 2003; Fatnassi et al., 2003; Hanafi et al., 2005; Valera et al., 2006). Although pesticides will remain an important tool for pest management in greenhouse crops, non chemical methods must be introduced to reduce damage to the environment and to delay the development of pest‘s resistance (Fatnassi et al., 2003; Hanafi et al., 2005). The use of insect screens in greenhouses have been found to be an efficient method for reducing insect migration into the greenhouse (Teitel et al., 1999; Bartzanas et al,. 2002; Hanafi et al., 2005), and subsequent for reducing the number of pesticide applications targeting the insect pests and vectors (Soni et al., 2005; Hanafi et al., 2005; Katsoulas et al., 2006).
The efficiency of insect-proof screens as a mechanical barrier (Berlinger et al., 1996; Fatnassi et al., 2003; Shilo et al,. 2004; Hanafi et al., 2005; Valera et al., 2006) depends on the dimensions of the pores. However, the density of the threads alone does not suffice to determine the average dimension of the pores; the diameter of the fibers must be also known. In most instances, the screens are characterized by the term ―mesh‖, which is the number of threads per inch in each direction. For example, a 50-mesh screen has 50 threads per inch of material. If the mesh and thread thickness are known the size of the opening can be obtained by subtracting the thread size from the reciprocal of the mesh; so a 50 mesh screen with a thread thickness of 0.15 mm has openings with a width of 0.35 mm (i.e. 1/50=0.02 inches=0.5 mm; subtracting the thread thickness of 0.15 mm gives 0.35 mm) in each direction. If the openings are rectangular, the screen will have openings of 0.48 mm by 0.27 mm if the thread thickness is 0.15 mm (Hanafi et al., 2005).
There are several commercial screens which have variable efficiencies in excluding insect pests. These screens have various sizes and have been used to exclude a certain number of insect pests of greenhouse crops. Depending on the target pest, the openings size in the screen must clearly be smaller than the size of the insect (Table 1) (Hanafi, 2005).
The maximum sizes of the openings in a insect-proof screen to exclude some important insect pests are given in table 2. In Europe and North Africa, the screens are identified as 6×9, 10×14 or 10×22, which means that these screens have rectangular openings. For example, a 10×20 screen has 10 threads by 20 threads in a centimeter square (Hanafi et al., 2005).
The objective of this work was to experimentally investigate the influence of several types of insect proof screens (13×23 and 30×34 as compared to indexes of positive (Agryl/P17) and negative (5×5) control) used to prevent of three major pests of strawberry of intrusion.
MATERIAL AND METHODS
In this study, we have evaluated in laboratory conditions (L:D 16:8, 27 2 °C
Temp., and 60 10% R.H.) the effectiveness of three types of insect-proof screens for their exclusion of the two spotted spider mite (TSSM), Tetranychus urticae Koch (Acari: Tetranychidae), the western flower thrips, Frankliniella occidentalis(Pergande) (Thysanoptera: Thripidae) and the strawberry aphid, Chaetosiphon fragaefolii(Cockerell) (Homoptera: Aphididae). The insect-proof screens tested were the 13×23 (13×23 threads per cm2) and 30×34, against a negative control (5×5, ability pests to cross) and to a positive control (Agryl/P17, a protective crop cover, not ability pests to cross). The insects were collected from

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 168
an experimental greenhouse of the Iranian Research Organization for Science and Technology (IROST) in Tehran, Iran.
In this experiment we have using one plastic cylindrical for each replicate. In each cylindrical container which was sealed by every one of the above mentioned insect-proof screens at one end (9 cm in diameter and 10 cm in height), 50 individuals were placed. Hence each screen was tested as a barrier between inner and outer space of cylindrical container. In this experiment, flowers and leaves of strawberry were used as an incentive for the insect to move through the openings of insect-proof screens. The control percent of insect screens was evaluated by calculating the number of insects that there were in the containers after 24 hours. We have evaluated two treatments: T1 (13×23), T2 (30×34), as compared to a negative control (5×5) and to a positive control (Agryl/P17). Each treatment was replicated three times. Analyzing data calculate by Design Expert (version 7.0.0) software.
RESULTS AND DISCUSSION
The control percent of insect screens were evaluated by calculating the number of insects that were in the containers after 24 hours. The results of this experiment indicated clearly that the 5×5 insect screen, which is widely used in greenhouses, did not really exclude these pests. The results of control percent of pests evaluated in this experiment through the different screens are indicated in Fig 2. The results of this experiment indicated that, the 13×23 insect screen could control 94% penetration of C. fragaefolii as compared to the 30×34 screen which controlled 100% of this species (Fig. 1.a). There was significant difference between the 13×23 and 30×34 insect screens in the control of C. fragaefolii (F= 27, P< 0.0065, df= 1). However, control percent were higher for C. fragaefolii than for F. occidentalis and T. urticae and this is related to the larger size of the strawberry aphid as compared to the western flower thrips and two spotted spider mite. These results also indicated that the average of control percent and excluding of F. occidentalis was zero for the 13×23 insect screen and 46.66% for the 30×34 one (Fig. 1.b). The comparison of the results also indicated that there was not significant difference between these screens for control of F. occidentalis (F= 7, P<0.0572, df=1). The 13×23 and 30×34 screens excluding of T. urticae, zero and 13.8% respectively. There was a significant difference between these screens for control of this pest (Fig. 1.c). Hence, results shown that these screens are not efficient in control and excluding of T. urticae (F= 36.60, P< 0.0038, df=1).
The results also have been compared with indexes of positive (Agryl/P17) and negative (5×5 screen) control for performance evaluation of these screens in the controlling of tested pests (Fig. 2 and table 3). The comparison of the 13×23 and 30×34 screens with the index of positive control (Agryl/ p-17) indicated the adequate control potency of these screens in exclusion of C. fragaefolii (F=7227.00, P<0.0001, df=3).As a result, these screens can be used as an anti-aphid screen. The comparison between of negative control index (5×5 screen) and the 13×23 screens indicated not efficient of this screen in excluding of F. occidentalis. But there was a significant difference between negative and positive control indexes and the 30×34 screen (F=29.14, P<0.0001, df=3). There was not a significant difference between the 13×23 screen and negative control index(5×5 screen) for control of T. urticae(F=1781.63, P<0.0001, df=3). However, there was a significant difference between the 30×34 screen and index of negative control, but this screen cannot provide a suitable control for excluding of this pest.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 169
Berlinger et al. (1996) indicated that screens also effectively excluded other insects equal or greater in size than whiteflies e.g. aphids and leafminers. The insect-proof screens must be installed before planting and fixed thoroughly to prevent even the smallest opening. Screening greenhouses with a suitable insect-proof screen minimizes pest immigration, significantly decreases pesticide applications, provides the basis for the implementation of an IPM program, and enables the use of bumble bee pollinators. Consequently, all greenhouse strawberry and many other greenhouse crops need protective screens nowadays.
ACKNOWLEDGEMENTS
The authors would like to thank Mr. Rohollah Rajabi for their valuable helps and discussions.
LITERATURE CITED Bailey, B. J., Montero, J. I., Perez Parra, J., Robertson, A. P., Baeza, E. & Kamaruddin, R. 2003. Airflow resistance of greenhouse ventilators with and without insect screens. Biosystems Engineering, 86 (2): 217–229. Bartzanas, T., Boulard, T. & Kittas, C. 2002. Numerical simulation of the airflow and temperature distribution in a tunnel greenhouse equipped with insect-proof screen in the openings. Computers and Electronics in Agriculture, 34: 207–22. Berlinger, M. J., Lebiush-Mordenchi, S. & Rosenfeld, J. 1996. State of the art and the future of IPM in greenhouse vegetables in Israel. IOBC/WPRS Bulletin, 19 (1): 11-14. Cedola, C. & Greco, N. 2008. Presence of the aphid, Chaetosiphon fragaefolii, on strawberry in Argentina. Journal of Insect Science, 10 (9): 1-9. Fatnassi, H., Boulard, T. & Bouirden, L. 2003.Simulation of climatic conditions in full-scale greenhouse fitted with insect-proof screens. Agricultural and Forest Meteorology, 118: 97–111. Fraulo, A. B., Cohen, M. & Liburd, O. E. 2009. Visible/near infrared reflectance (VNIR) spectroscopy for detecting two spotted spider mite (Acari: Tetranychidae) damage in strawberries. Environmental Entomology, 38 (1): 137-142. Hanafi, A., Amouat, S., Miftah, S. & Bouharroud, R. 2005. Efficiency of insect nets in excluding whiteflies and their impact on some natural biological control agents. Available from: http://www.actahot.org/members/ shawpdf?booknrarnr=646-47 Katsoulas, N., Bartzanas, T., Boulard, T., Mermier, M. & Kittas, C. 2006. Effect of vent openings and insect screens on greenhouse ventilation. Biosystems Engineering , 93 (4): 427–436. Obrycki, J. 2004. The effects of thrips on strawberry production in Iowa. Leopold Center Progress Report, 13: 28-30. Reitz, S. R. 2009. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Florida Entomologist, 92 (1): 7-13. Rondon, S. I. & Cantliffe, D. J. 2004. Chaetosiphon fragaefolii (Homoptera: Aphididae): a potential new pest in florida?. Florida Entomologist, 87 (4): 612-615. Rondon, S. I., Cantliffe, D. J. & Price, J. F. 2005. Population dynamics of the cotton aphid, Aphis gossypii(Homoptera: Aphididae), on strawberry grown under protected structure. Florida Entomologist, 88 (2): 152-158. Shilo, E., Teitel, M., Mahrer, Y. & Boulard, T .2004. Air-flow patterns and heat fluxes in roof-ventilated multi-span greenhouse with insect-proof screens. Agricultural and Forest Meteorology, 122: 3–20. Soni, P., Salokhe, V. M. & Tantau, H. J. 2005. Effect of screen mesh size on vertical temperature distribution in naturally ventilated tropical greenhouses. Biosystems Engineering, 92 (4): 469–482.
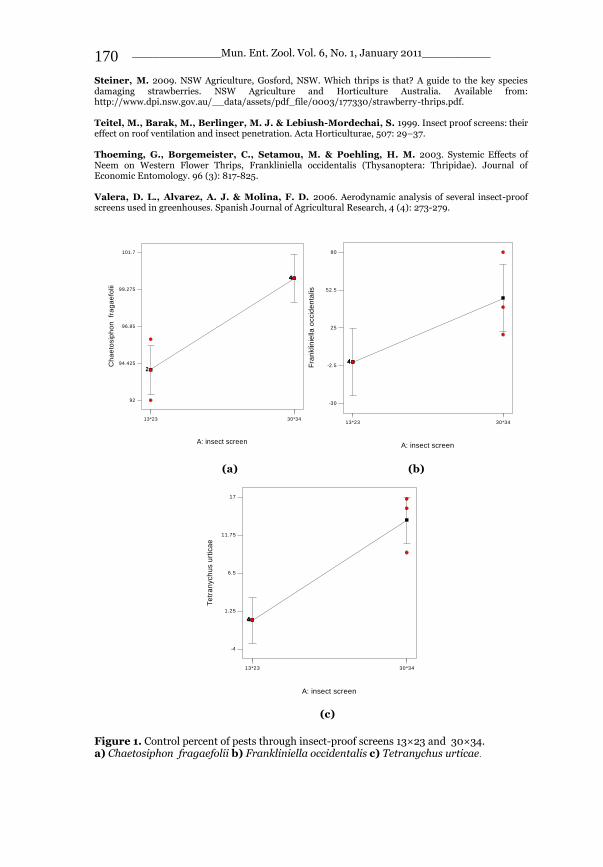
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 170
Steiner, M. 2009. NSW Agriculture, Gosford, NSW. Which thrips is that? A guide to the key species damaging strawberries. NSW Agriculture and Horticulture Australia. Available from: http://www.dpi.nsw.gov.au/__data/assets/pdf_file/0003/177330/strawberry-thrips.pdf. Teitel, M., Barak, M., Berlinger, M. J. & Lebiush-Mordechai, S. 1999. Insect proof screens: their effect on roof ventilation and insect penetration. Acta Horticulturae, 507: 29–37. Thoeming, G., Borgemeister, C., Setamou, M. & Poehling, H. M. 2003. Systemic Effects of Neem on Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Journal of Economic Entomology. 96 (3): 817-825. Valera, D. L., Alvarez, A. J. & Molina, F. D. 2006. Aerodynamic analysis of several insect-proof screens used in greenhouses. Spanish Journal of Agricultural Research, 4 (4): 273-279.
Design-Expert® Software
Chaetosiphon fragaefolii
Design Points
X1 = A: insect screen
A: insect screen
Ch
ae
tosip
ho
n
fra
ga
efo
lii
One Factor
13*23 30*34
92
94.425
96.85
99.275
101.7
2
4
2
444
Design-Expert® Software
Frankliniella occidentalis
Design Points
X1 = A: insect screen
A: insect screen
Fra
nklin
iella
occid
en
talis
One Factor
13*23 30*34
-30
-2.5
25
52.5
80
4444
(a) (b) Design-Expert® Software
Tetranychus urticae
Design Points
X1 = A: insect screen
A: insect screen
Te
tra
nych
us u
rtic
ae
One Factor
13*23 30*34
-4
1.25
6.5
11.75
17
4444
(c)
Figure 1. Control percent of pests through insect-proof screens 13×23 and 30×34. a) Chaetosiphon fragaefolii b) Frankliniella occidentalis c) Tetranychus urticae

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 171
0
20
40
60
80
100
120
23 13 34 30 5 5 Agryl p-17
Contr
ol
per
cent
Type of insect-proof screens
Chaetosiphon
fragaefoliiFrankliniella
occidentalis Tetranychus urticae
Figure 2. Control percent of Chaetosiphon fragaefolii, and Frankliniella occidentalis and Tetranychus urticae through different insect screens (13×23 & 30×34) as compared to a positive control (Agryl/P17) and to a negative control (5×5), in laboratory conditions.
Table 1. Width and length in millimeter of some important insect pests of greenhouse crops (Hanafi et al., 2005).
Insect pest Width(mm) Length(mm)
Thorax Maximum
Serpentine leaf miner
(Liriomyza trifolii) 0.608
0.850 0.177
Sweet potato whitefly
(Bemisia tabaci) 0.615
0.870 0.181
Melon aphid
(Aphis gossypii) 0.355
0.239 0.236
Greenhouse whitefly
(Trialeurodes vaporariorum) 0.288
0.709 0.128
Silverleaf whitefly
(Bemisia argentifolii) 0.239
0.565 0.107
Western flower thrips
(Frankliniella occidentalis) 0.215
0.267 0.126
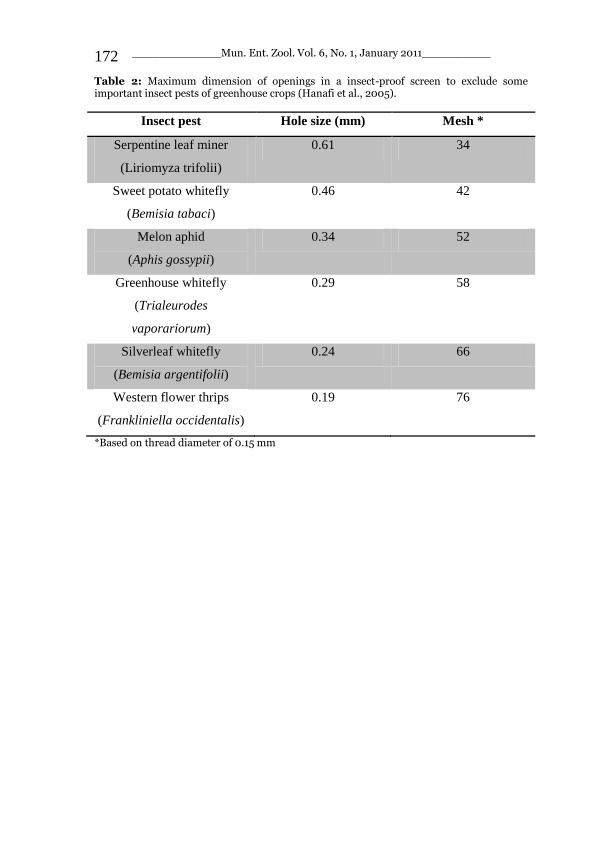
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 172
Table 2: Maximum dimension of openings in a insect-proof screen to exclude some important insect pests of greenhouse crops (Hanafi et al., 2005).
Insect pest Hole size (mm) Mesh *
Serpentine leaf miner
(Liriomyza trifolii)
0.61 34
Sweet potato whitefly
(Bemisia tabaci)
0.46 42
Melon aphid
(Aphis gossypii)
0.34 52
Greenhouse whitefly
(Trialeurodes
vaporariorum)
0.29 58
Silverleaf whitefly
(Bemisia argentifolii)
0.24 66
Western flower thrips
(Frankliniella occidentalis)
0.19 76
*Based on thread diameter of 0.15 mm

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 173
A NEW RECORD OF XYLOTRUPES GIDEON (LINNAEUS) (COLEOPTERA: SCARABAEIDAE) ON PERSIA BOMBYCINA,
KOST. FROM INDIA
Rajesh Kumar*, G. Rajkhowa, N. J. Dhar and R. K. Rajan * Central Muga Eri Research & Training Institute, Central Silk Board, Ministry of Textiles (Govt. of India), Central Silk Board, Lahdoigarh 785700, Assam, INDIA. E-mail: [email protected]
[Kumar, R., Rajkhowa, G., Dhar, N. J. & Rajan, R. K. 2011. A new record of Xylotrupes gideon (Linnaeus) (Coleoptera: Scarabaeidae) on Persia bombycina, Kost. from India. Munis Entomology & Zoology, 6 (1): 173-175] ABSTRACT: Xylotrupes gideon (Linnaeus) is belongs to the family Scarabaeidae in order Coleoptera was noticed feeding on Som plant, Persia bombycina Kost. (Lauraceae) during May to September, 2010 and analysis of literature indicates that this is the first record of X. gideon Linnaeus on Som plantation crop at Farm No. 3 and Farm No. 2, Central Muga Eri Research and Training Institute, Central Silk Board, Ministry of Textiles, Govt. of India, Lahdoigarh, Assam. In the manuscript the host range of X. gideoon (L.) and some morphological characters is discussed. KEY WORDS: Xylotrupes gideon (L.), new record, som, Assam, India.
Som, Persia bombycina Kost (Family: Lauraceae) is primary host plant for Muga Silkworm, Antheraea assamensis, Helfer (Lepidoptera: Saturniidae) in whole Assam and other North Eastern Region of India where the muga silkworm rearing occurs. This tree usually attacked by many pests like, shoot borer, trunk borer, leaf miners, leaf galls, mealy bugs etc. Recently, Xylotrupes gideon (Linnaeus) (Coleoptera: Scarabaeidae: Dynastinae) was noticed feeding on the trunk of the Som plant during the August and September, 2010. North eastern region of India is abode to endemic insect biodiversity. Som plantation is available in North Eastern India and muga silkworm (A. assamensis, Helfer) is endemic to this region in the world, because of climatic conditions. The purpose of this paper is to highlight important observations and conclusions made by previous host plants and new host plant record along with color polymorphism of this beetle.
MATERIALS AND METHODS
All the specimens were collected and preserved in well fumigated wooden boxes. Prior to collection, the beetles were photographed in the field condition from May to September, 2010 at Farm No. 3 and Farm No. 2, Central Muga Eri Research and Training Institute, Central Silk Board, Ministry of Textiles, Govt. of India, Lahdoigarh, Assam. For field observations specimens and damage symptoms were photographed by Sony DSC R1 10.3 mega pixel. The photographs were edited using software ACDSee 9.0 Photo Manager and prepared plate in 600 dpi using software Adobe Photoshop 7.0.
RESULTS AND DISCUSSION
This plantation crop for rearing of Muga Silkworm was reared at Farm No. 3 and Farm No. 2, Central Muga Eri Research and Training Institute (CMER&TI),

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 174
Lahdoigarh, Jorhat, Assam (India). The detail information of observation was collected from two farms of the institute (Farm No. 2 and 3).
The damage symptoms were observed in the field and shows that beetle made hole to the main trunk of the plant (Figs. 1, 2, 3). During rainy season May – September, 2010, It was observed that the beetles come out from the soil and sit on plant trunk in group (Figs. 4, 5). The beetles were found in group of 4-6 full grown adults feeding on the trunk of the plant. The male were larger than the female and infatuated cephalic and prothoracic horn, but females never infatuate with horn. The color polymorphism was also observed and was found that the color of male and female of X. gideon (L.) were varied. X. gideon (L.) was found in three colors i.e. black, blackish brown, reddish brown (Fig. 4-7). The observation for X. gideon (L.) shows that males were dimorphic for horn length, but horn size varies within morphs (Figs. 4-7). Males of X. gideon were generally larger than females, but sometimes observed that few males were smaller then the females (Figs. 4, 5).
X. gideon is well known as occasional pest of coconut (Dangar et al., 1994) and it has also been reported on crops like plum (Kumar et al., 2007), okra (Nair et al., 2001), Oil palms, Sugarcane, Rubber, Banana, Bamboo, Poinciana, Cassia, Litchi and Potato (CABI, 1985). X. gideon is world wide in distribution. Hence, an analysis of literature indicates that this first record of infestation on Som plantation crop by X. gideon (Linnaeus).
ACKNOWLEDGEMENTS
The authors are thankful to Dr. V. V. Ramamurthy, Principal Scientist / In-charge, Identification Service, Division of Entomology, Indian Agricultural Research Institute, New Delhi 110012 for identification of the beetles.
LITERATURE CITED CABI, 1985. Xylotrupes gideon, Distribution Maps of Plant Pests. Map No. 474. Dangar, T. K., Solomon, J. J. & Pillai, G. B. 1994. Infection of the coconut palm beetle, Xylotrupes gideon (Coleoptera: Scarabaeidae), by a nonoccluded baculovirus. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz, 101 (6): 561-566. Nair, S., Prameela, P., Suma, A., Cherian, K. A. & Babu, K. V. 2001. A new record of Xylotrupes gideon (L.) on okra (Abelmoschus esculentus [L.]). Insect Environment, 7 (2): 71. Kumar, J., Sharma, S. D. & Ramesh, Lal. 2007. Host plants of scarabaeid beetles in the lower Kullu valley of Himachal Pradesh. Pest Management and Economic Zoology. 15 (2): 213-216.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 175
Figures 1-7, Xylotrupes gideon (Linnaeus): 1. Som plantation, Persea bombycina Kost shows highlighting beetle feeding, 2. Beetle feeding on trunk, 3. Damage symptoms, 4. Group sitting on trunk during morning time after rain (one male and three female), 5. One male and one female, 6. Male lateral view, 7. Female hiding in litter.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 176
PARASITISM OF PIERIS BRASSICAE (L.) (LEP.: PIERIDAE) ON CABBAGE FARMS IN COMPARISON
WITH WILD HOSTS AND STUDY ON USE OF PTEROMALUS PUPARUM (L.) (HYM. PTEROMALIDAE), AS A BIOLOGICAL CONTROL AGENT VERSUS THIS PEST
Mehdi Razmi*, Yones Karimpour
and Mohammad Hasan Safaralizadeh
* Department of Plant Protection, Faculty of Agriculture, Urmia University, P. O. Box -165, IRAN. E-mail: [email protected] [Razmi, M., Karimpour, Y. & Safaralizadeh, M. H. 201. Parasitism of Pieris brassicae (L.) (Lep.: Pieridae) on cabbage farms in comparison with wild hosts and study on use of Pteromalus puparum (L.) (Hym. Pteromalidae), as a biological control agent versus this pest. Munis Entomology & Zoology, 6 (1): 176-180] ABSTRACT: During the recent decade, the high risks and detrimental consequences associated with the use of chemical and even microbial pesticides have been a recurring theme in pest control, and researchers have been trying to pursue low-risk and environmentally sound tactics. However, biological control is a non-chemical method that utilizes natural enemies for suppression and regulation of insect pest populations. This study was performed in order to assess parasitism rate and parasitoid diversity of Cabbage Large White Butterfly, Pieris brassicae (L.) (Lep.: Pieridae), on cabbage fields in comparison with wild hosts in Urmia. Also In this study some biological and behavioral characteristics of Pteromalus puparum (L.) (Hym.: Pteromalidae , predominant endoparasitoid wasp of P. brassicae, was studied. For this purpose, samples were taken from various cabbage farms and wild hosts of this pest outside the farm. The specimens have been collected from all stages of P. brassicae. In order to parasitoids emerge, samples incubated separately in laboratory conditions at 23 1°C, 40 5 % r.h. and 16:8 h (L:D) photoperiod. The other main purpose of this study was to evaluate the efficacy of P. puparum in the biological control of Cabbage Large White Butterfly. KEY WORDS: Pieris brassicae, Pteromalus puparum, Biological control, Parasitism, Cabbage .
Pieris brassicae and Pieris rapae (Pieridae) are some of the most frequent pests of cabbage crops alongside Mamestra brassicae, and Plutella xylostella (Patriche et al., 2005). The Large White Butterfly (LWB), Pieris brassicae L., is a cosmopolitan insect and is found wherever cruciferous plants are grown (Hill, 1987). Sometimes massive outbreaks of LWB may occur and injury on cabbage cultures may be extensive.
Parasitic wasps which lay their eggs either in or on their hosts are important regulators of insect populations and comprise a large proportion of hymenopteran species (Quicke, 1997).The role of beneficial organisms (such as parasitoids) as natural enemies of insect pests on cabbage is of great consequence (Balevski et al., 2007). Insect parasitoids have been introduced into a variety of ecosystems as bioregulators and there have been many successful cases of their being used to control harmful native arthropods (DeBach Schlinger 1964; DeBach, 1974). Studies on the fauna of parasitic insect of different regions are of great significance for effective pest control (Aliyev, 1999). Moreover, eggs and larvae of P. brassicae are preyed on by generalist predators such as Spiders, Chrysopids, Staphylinids and Carabids (Pfiffner et al., 2009).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 177
Numerous experiments have shown that pest populations often flourish in absence of predators and parasitoids, often killed by the pesticide applications used to control the pest. Chemical pesticides affect more the beneficial insects than the pests themselves, because they are more sensitive (Costea et al., 2002). The goal of our research was to observe how the parasitoids limit P. brassicae populations on the cabbage crops in comparison with wild hosts of this pest outside the farms.
MATERIALS AND METHODS
The experiments were conducted in north-west of Iran (Urmia). In 2008 & 2009, during a survey on this pest LWB, sufficient specimens including egg, larvae (in all stages) and pupae have been collected from different collecting dates. Also, we monitored pupal and larval parasitism as well as parasitism rates in some cabbage fields and regions with wild hosts of this pest like Capparis spinosa (Capparaceae) and Raphanus raphanistrum (Cruciferae). All insects used in the experiments originated from cabbage fields in the Urmia.
In laboratory, egg clusters, larvae and pupae in order to prevent infecting with pathogens (fungi, virus or bacteria), placed in a clean Petri dish separated one by one and each of which with a part of fresh host leaf as an alimentary material. Once a day, Petri dishes were cleaned and larvae were fed fresh leaves of Brassica oleracea or C. spinosa or R. raphanistrum. Food plants were not changed during the experiment. Host larvae were fed until they pupated. Also, egg clusters with up to 150 eggs were kept on moist tissue paper in Petri dishes under 16L : 8D and 23 1°C in an incubator.
Parasitized butterfly pupae were maintained in large Petri dishes (10-cm diameter) and were monitored until adult parasitoid emergence. Then, emerged adult wasps were placed in separate Petri dishes and were provided with water and honey and pollen. The parasitoids are identified and the percentage of parasitism by each parasitoid is estimated too.
To evaluate the efficacy of P. puparum in the biological control of LWB, host stage preference, number of progeny laid by the female, number of parasitoid adults emerged from each pupa, immature period from egg hatch to adult emergence of this parasitoid was estimated. For parasitism, we used females that were 1- to 3-d old after adult emergence.
For experiments, newly pupated hosts (within 24 h after pupation) were exposed to female wasps, which had no pervious contacted with hosts. To avoid super parasitization, a pupa was held together with one mated female wasp in Petri dishe. Furthermore, the parasitioid was removed immediately after the oviposition was observed. Then, the parasitized pupae were cultured at 23 1°C and a photoperiod of 16:8h (L:D).
RESULTS
Laboratory assays made an evidence of the parasitism percentage in
samples collected from farms and wild hosts were 16.93 and 43.25, respectively. The diversity of parasitoids in the samples collected from the wild hosts outside the cabbage fields was higher than the specimens collected from farms. Results showed that 0.56, 1.41, 1.97, 3.66, 44.51 and 47.89 percent of extracted parasitoids belonged to the families: Eulophidae, Ichneumonidae, Chalcididae, Tachinidae, Braconidae and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 178
Pteromalidae respectively. The summary of parasitoids Family recorded during the present investigation is provided in Table and figure 1. The other main purpose of this study was to evaluate the efficacy of P. puparum in the biological control of LWB. Initially, host stage preference of this parasitoid to different instars and developmental stages was determined. Other experiments pointed out that number of progeny laid by the female was 270.6 ± 4.62. Number of parasitoid adults emerged from each pupa was 42.36 ± 2.42, but more than 200 parasitoids can be produced within each host pupae, when more than one female attacked a single host. Total immature period from egg hatch to adult emergence required 17.56 ± 0.33 days. Mating starts soon after eclosion. Longevity was evaluated in two diets including honey mixed with pollen and without feeding that were 16.28 ± 0.38 and 3.04 ± 0.12 days, respectively. The parasitoid wasp, P. puparum, significantly preferred to attack the pupae and pre-pupal stage of P. brassicae.
DISCUSSION
Some P. brassicae larvae in Hokkaido have been parasitized by Cotesia glomerata, but the parasitism rate of P. brassicae larvae tends to be lower than that of P. rapae (Sato & Ohsaki, 2004). During this study we observed that considerable numbers of P. brassicae larvae are efficiently controlled by C. glomerata. This difference in results perhaps because all of the wasps used via Sato and Ohsaki were obtained from P. rapae larvae. However, several of the P. brassicae larvae collected in the field and on wild hosts in our region had been parasitized by C. glomerata.
Also, considerable numbers of P. brassicae pupae are efficiently parasitized by P. puparum and use of this parasitoid in same time with C. glomerata may play an important role in the biological control of P. brassicae on cabbage farms. P. puparum (Linnaeus) is a gregarious pupal parasitoid with a wide host range, including species of the Nympahlidae and Pieridae. This parasitoid contributes to regulation of a population of Papilio xuthus Linnaeus at low density in a suburban area of Fukuoka, Japan (Takag, 1985). This wasp is a predominant pupal endoparasitoid of Pieris rapae (Zhu et al., 2008) and considered to be an important biological control agent of this pest (Harvey et al., 2007).
ACKNOWLEDGEMENTS
We would like to thanks from Dr. Aurel Lozan, Dr. Gerard Delvare, Dr. Hans-Peter Tschorsnig, Dr. Hossein Lotfalizadeh, Dr. Janko Kolarov and Dr. Maximilian Fischer for their valuable helps in the identification of parasitoid specimens. We also thank Dr. Seid Ali Safavi of Urmia University for his help in the statistical examination of the results and Mr. Ebrahim Hasanzade for his help in carrying out the experiment.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 179
LITERATURE CITED Aliyev, A. A. O. 1999. Fauna of the subfamily Ichneumoninae (Hymenoptera, Ichneumonidae) of Azerbaijan with new records. Tr. J. of Zoology, 23: 1-12. Balevski, N., Draganov, H., Velichkova-Kojuharova, M. & Draganova, S. 2007. Beneficial organisms (entomopathogens and entomophagous) on pests in the biocoenoses of cabbage in Bulgaria. Plant Science, 44: 230-235. Costea, G., Mustata, G. H., Lozan, A. 2002. Role of Braconidae (Hymenoptera) in limitation of Lepidoptera cabbage pest's populations in Romania. In: Melika, G. & Thuroczy, C. (Eds.), parasitic wasps: evolution, systematic, biodiversity and biological control. Agroinform, Budapest, pp: 391-395. DeBach, P. 1974. Biological control by natural enemies. Cambridge University Press. DeBach P., Schlinger, E. I. 1964. Biological control of insect pests and weeds. Chapman and Hall, London. Harvey, J. A., Gols, R., Wagenaar, R. & Bezemer, T. M. 2007. Development of an insect herbivore and its pupal parasitoid reflect differences in direct plant defense. J. Chem. Ecol., 33: 1556–1569. Hill, D. S. 1987. Agricultural insect pests of temperate regions and their control. Oxford, 659 pp. Patriche (Costea), G., Andriescu, I. & Mustaţǎ, G. 2005. The hyperparasitoid complex which limits the action of the primary parasitoids of the pieridae species (Insecta: Lepidoptera), defoliators in cabbage crops. Biologie animală, 23-29. Pfiffner, L., Luka, H., Schlatter, C., Juen, A. & Traugott, M. 2009. Impact of wildflower strips on biological control of cabbage lepidopterans. Agriculture, Ecosystems and Environment, 129 : 310–314. Quicke, D. L. J. 1997. Parasitic Wasps. Chapman and Hall, London. Sato, Y. & Ohsaki, N. 2004. Response of the wasp (Cotesiaglomerata) to larvae of the large white butterfly (Pieris brassicae). Ecological Research, 19: 445–449. Takag, M. 1985. The reproductive strategy of the gregarious parasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae). Oecologia (Berlin), 68: 1-6. Zhu, J., Ye, G. & Hu, C. 2008. Morphology and ultrastructure of the venom apparatus in the endoparasitic wasp Pteromalus puparum (Hymenoptera:Pteromalidae). Micron, 39: 926–933.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 180
Table 1: Parasitoids of P. brassicae, recorded during 2008-2009 survey in Urmia (Iran).
Parasitoid Family Order Host pest/Stage Host-crop/s percentage of parasitism
Ichneumonidae Hymenoptera Pupa/P. brassicae
Raphanus/ Capparis
1.41
Brassica oleracea 0.00
Chalcididae Hymenoptera Pupa/P. brassicae
Raphanus/ Capparis
1.97
Brassica oleracea 0.00
Braconidae Hymenoptera Larva/P. brassicae
Raphanus/ Capparis
6.76
Brassica oleracea 37.75
Pteromalidae Hymenoptera Pupa/P. brassicae
Raphanus/ Capparis
47.89
Brassica oleracea 0.00
Eulophidae Hymenoptera Pupa/P. brassicae
Raphanus/ Capparis
0.56
Brassica oleracea 0.00
Tachinidae Diptera Larva/P. brassicae
Raphanus/ Capparis
2.82
Brassica oleracea 0.84
Specimens reared from Raphanus/Capparis and Cabbage in Urmia has been deposited in the University of Urmia at Department of Entomology collection.
percentage of parasitism on cabbage farms in
comparison with wild hosts
0
1020
30
4050
60
Ichn
eum
onid
ae
Chal
cididae
Bra
conida
e
Pte
rom
alidae
Eulop
hidae
Tachinida
e
cabbage fields
wild hosts
Figure 1. Percentage distribution of P. brassicae parasitoids on cabbage farms in comparision with wild hosts.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 181
SOME ADDITIONAL NOTES ABOUT ALEOCHARINAE (COLEOPTERA: STAPHYLINIDAE) FAUNA OF TURKEY
Sinan Anlaş* and Armin Rose **
* Department of Biology, Faculty of Science, University of Ege, 35100 Bornova, Izmir / TURKEY. E-mail: [email protected] ** DZMB, German Centre for Marine Biodiversity Research, Senckenberg Research Institute, Südstrand 44, D-26382 Wilhelmshaven / GERMANY. E-mail: [email protected] [Anlaş, S. & Rose, A. 201. Some additional notes about Aleocharinae (Coleoptera: Staphylinidae) fauna of Turkey. Munis Entomology & Zoology, 6 (1): 181-185] ABSTRACT: In this study, additional records of 27 species of Aleocharinae are given. Additional notes on most of them new to certain Turkish regions and provinces are given. Anaulacaspis laevigata Duvivier, 1883 is the first record for the Turkish fauna. A. pamphylica (Fagel, 1969) represents the first records since the description of the species. KEYWORDS: Staphylinidae, Aleocharinae, new record, fauna, Turkey.
The Aleocharinae are the most diverse subfamily of Staphylinidae, more than 12.000 species worldwide out of 1.740 genera (Herman, 2001). According to AnlaĢ (2009 and an updated version), the subfamily Aleocharinae is represented by 520 species of 81 genera in Turkey, 202 (39,2 %) of which occur as endemics. However, many Turkish provinces are sparsely investigated regarding their Aleocharinae species inventory.
The aim of this study is to enhance scientific knowledge on the distribution of Turkish Aleocharinae.
MATERIAL AND METHOD
The present paper is based primarily on material collected during three field trips to Antalya carried out by Armin Rose in March 2000, 2001, and 2002, as well as recent collections by Sinan AnlaĢ and Ersen Aydın Yağmur.
Classification and nomenclature of the Aleocharinae suggested by Smetana (2004) has been followed in this study.
The material referred to in this study is deposited in the following collections: LEMT ……………………………..Lodos Entomological Museum, Izmir (S. Tezcan) cAnl ………………………………..first author‘s private collection cRos …………………….………….second author‘s private collection cTer …………………….…………..private collection of H. Terlutter (Billerbeck)
RESULTS
In this study, 27 species of 12 genera belonging to 7 tribes of Aleocharinae are reported from Turkey. Anaulacaspis laevigata Duvivier, 1883 is the first record for the Turkish fauna. A. pamphylica (Fagel, 1969) represents the first records since the description of the species. The formerly known distribution of these species for Turkey is given by AnlaĢ (2009).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 182
Subfamily Aleocharinae Fleming, 1821 Tribe Aleocharini Fleming, 1821
Aleochara intricata Mannerheim, 1830 Material: Adıyaman: Gerger, Açma 2 km E, 37°59‘35‘‘N, 38°57‘51‘‘E, 1 ex., 19.IV.2008, leg. Yağmur (cAnl). Antalya: Kemer 5 km NW, rocky riverbank in canyon, on rotting meat, 197 m, 36°36‘03‘‘N, 30°29‘03‘‘E, 1 ex., 30.III.2001, leg. Rose (cRos). Distribution in Turkey: Ankara, Bitlis, Hatay, Istanbul, Konya, Mersin, Osmaniye, ġanlıurfa, Van.
Aleochara lata Gravenhorst, 1802 Material: Antalya: Alanya, TaĢkesiği, east bank of Karpuz river, in dry chicken carrion on sand, 50 m, 36°45‘N, 31°37‘E, 1 ex., 17.III.2000; Central province, near Cakırlar, riverside, on car, 259 m, 36°48‘51‘‘N, 30°28‘15‘‘E, 1 ex., 27.III.2001; Kemer 5 km NW, rocky riverbank in canyon, on rotting meat, 197 m, 36°36‘03‘‘N, 30°29‘03‘‘E, 2 exs., 30.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Aleochara maculipennis Baud idi Selve, 1857 Material: Antalya: Central province, Kumluca 5 km E, dry meadow with stone pines, from cow dung, 550 m, 36°22‘47‘‘N, 30°22‘58‘‘E, 1 ex., 25.III.2001, leg. Rose (cRos). Distribution in Turkey: Hakkari, KahramanmaraĢ, Konya, ġanlıurfa.
Aleochara tristis Gravenhorst, 1806 Material: Antalya: Alanya, mountain road near Ilıca, slope (SE), in old dry cow dung, 340 m, 2 exs., 15.III.2000, leg. Rose (cRos). Izmir: Bayındır, Osmanlar 2 km SE, 420 m, 38°18‘07‘‘N, 27°33‘58‘‘E, 4 exs., 12.XII.2008, leg. AnlaĢ (LEMT, cAnl). Muğla: Fethiye, Akdağlar, 2510 m, 2 exs., 13.VIII.2005, leg. Yağmur (cAnl). Distribution in Turkey: Artvin, Bingöl?, Bitlis, Bursa, Erzurum, Gaziantep, Hatay, KahramanmaraĢ, Konya, Malatya, Manisa, Mardin, Mersin, Osmaniye.
Tribe Athetini Casey, 1910 Acrotona muscorum (Brisout de Barneville, 1860)
Material: Antalya: Alanya, Kuzyaka, hillside meadow with creek, in nearly dry cow dropping, 811 m, 36°47‘24‘‘N, 30°28‘58‘‘E, 2 exs., 27.III.2001, leg. Rose (cRos). Distribution in Turkey: Erzurum, Izmir, KahramanmaraĢ.
Acrotona nigerrima (Aubé, 1850) Material: Antalya: Central province, 273 m, 36°49‘22‘‘N, 30°32‘16‘‘E, 1 ex., 31.III.2001, leg. Rose (cRos). Distribution in Turkey: Izmir.
Acrotona parens (Mulsant & Rey, 1852) Material: Antalya: Kemer 15 km WNW, steep westside slope with stone pines, moist patches, under stones, 1115 m, 36°39‘21‘‘N, 30°24‘48‘‘E, 1 ex., 30.III.2001, leg. Rose (cRos). Distribution in Turkey: Konya, Ordu, Samsun, Tokat (Assing, 2009).
Acrotona parvula (Mannerheim, 1830) Material: Antalya: Kemer 15 km WNW, steep westside slope with stone pines, moist patches, under stones, 1115 m, 36°39‘21‘‘N, 30°24‘48‘‘E, 1 ex., 30.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Atheta aeneicollis (Sharp, 1869) Material: Antalya: Akseki, near Güçlüköy, dry hillside with terraces, 600 m, 36°48‘N, 31°44‘E, 2 exs., 14.III.2000; Alanya, TaĢkesiği, east bank of Karpuz river, 50 m, 36°45‘N, 31°37‘E, 1 ex., 17.III.2000; same data but, in dry chicken carrion on sand, 50 m, 36°45‘N, 31°37‘E, 7 exs., 17.III.2000; Central province, Altınyaka 5 km S, hillside creek, humid

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 183
sycomore tree litter, sifted close to stem, 551 m, 36°28‘42‘‘N, 30°20‘08‘‘E, 6 exs., 30.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Atheta atramentaria (Gyllenhal, 1810) Material: Antalya: Alanya, TaĢkesiği, east bank of Karpuz river, flotsam sifted, on carrion, 50 m, 36°45‘N, 31°37‘E, 1 ex., 17.III.2000; Kuzyaka, hillside meadow with creek, in nearly dry cow dropping, 811 m, 36°47‘24‘‘N, 30°28‘58‘‘E, 1 ex., 27.III.2001; Central province, Kemer 5 km NW, rocky riverbank in canyon, on rotting meat, 197 m, 36°36‘03‘‘N, 30°29‘03‘‘E, 1 ex., 30.III.2001; near Ovacık, semi-moist dip with clay soil, in old cow dropping, 1051 m, 36°38‘37‘‘N, 30°26‘14‘‘E, 1 ex., 31.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Atheta elongatula (Gravenhorst, 1802) Material: Antalya: Alanya, TaĢkesiği, east bank of Karpuz river, in gravel with hollows under fresh flotsam, 50 m, 36°45‘N, 31°37‘E, 2 exs., 17.III.2000, leg. Rose (cRos). Distribution in Turkey: According to Assing (2009), a recent record of this species is known only from Amasya province.
Atheta laevana (Mulsant & Rey, 1852) Material: Antalya: Alanya, mountain road between Sugözü & Ilıca, from mule dung, 400 m, 1 ex., 15.III.2000, leg. Rose (cRos). Distribution in Turkey: Trabzon.
Atheta longicornis (Gravenhorst, 1802) Material: Antalya: Central province 5 km S, from quite fresh mule dung, 273 m, 36°49‘22‘‘N, 30°32‘16‘‘E, 1 ex., 31.III.2001; near Ovacık, semi-moist dip with clay soil, in old cow dropping, 1051 m, 36°38‘37‘‘N, 30°26‘14‘‘E, 1 ex., 31.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Atheta testaceipes (Heer, 1839) Material: Izmir: Karaburun 25 km SE, 10 m, 38°21‘02‘‘N, 26°38‘20‘‘E, 2 exs., 19.XII.2008, leg. AnlaĢ (cAnl). Distribution in Turkey: KahramanmaraĢ, Manisa.
Nehemitropia lividipennis (Mannerheim, 1830) Material: Izmir: Bayındır, Osmanlar 2 km SE, 420 m, 38°18‘07‘‘N, 27°33‘58‘‘E, 2 exs., 12.XII.2008, leg. AnlaĢ (cAnl). Mersin: Mezitli, Çevikli 2 km S, 1 ex., 12.VII.2005, leg. AnlaĢ (cAnl). Distribution in Turkey: Istanbul, Manisa, Tunceli.
Tribe Autaliini Thomson, 1859 Autalia impressa (Olivier, 1795)
Material: Antalya: Central province, near BeĢkonak, Köprülu Kanyon, shore of Köprü Cayi, from fine sand, 126 m, 37°07‘08‘‘N, 31°12‘41‘‘E, 1 ex., 17.III.2002, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Tribe Falagriini Mulsant & Rey, 1873 Anaulacaspis laevigata Duvivier, 1883
Material: Izmir: Bayındır, Yakapınar, 2 exs., 30.VI.2005, leg. AnlaĢ (cAnl). Manisa: Turgutlu, Çıkrıkçı, 5 exs., 23.V.2005, leg. AnlaĢ (LEMT, cAnl). Mersin: Central province, Mezitli, 4 exs., leg. AnlaĢ (cAnl). Remarks: According to Smetana (2004), the species was previously known from Austria, Bosnia Hercegovina, Croatia, Greece, Italy and Uzbekistan. It is here reported from Turkey for the first time.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 184
Anaulacaspis pamphylica (Fagel, 1969) Material: Antalya: Central province, Arif 2 km NW, dripping water under bridge, in gravel, 795 m, 36°31‘23‘‘N, 30°00‘42‘‘E, 23 exs., 26.III.2001; near YarbaĢı, clear river creek, under stones, 1084 m, 36°44‘33‘‘N, 30°20‘24‘‘E, 1 ex., 29.III.2001, leg. Rose (cRos). Distribution in Turkey: A. pamphylica was previously known only one locality from Antalya province, Alanya (Fagel, 1969). The species represents the first records since the description of the species.
Falagrioma thoracica (Stephens, 1832) Material: Kahramanmaraş: Döngel cave, 820 m, 37°51‘35‘‘N, 36°38‘29‘‘E, 1 ex., 30.V.2008, leg. Yağmur (cAnl). Distribution in Turkey: MuĢ, Ordu, Osmaniye, Samsun, Trabzon (Assing, 2009).
Tribe Hypocyphtini Laporte, 1835 Oligota pusillima (Gravenhorst, 1806)
Material: Antalya: Alanya, Side 5 km NW, dry sand dunes, litter of bushes sifted, 30 m, 1 ex., 13.III.2000, leg. Rose (cRos). Distribution in Turkey: Mersin.
Tribe Myllaenini Ganglbauer, 1895 Myllaena intermedia Erichson, 1837
Material: Antalya: Central province, BeĢkonak 1 km N, Köprülü Kanyon, waterfall, from moist mosses and leaves, 178 m, 37°11‘14‘‘N, 31°10‘47‘‘E, 1 ex., 22.III.2002, leg. Rose (cRos). Izmir: ÖdemiĢ, Bozdağlar, road to ski resort, ca. 1600 m, 38°21‘N, 28°06‘E, 2 exs., 21.V.2006, leg. AnlaĢ (cAnl). Distribution in Turkey: Istanbul, Manisa.
Tribe Oxypodini Thomson, 1859 Ocalea ruficollis Eppelsheim, 1888
Material: Antalya: Central province, Kumuca 5 km E, dry meadow with stone pines, shady brookside, sifted, 530 m, 36°22‘49‘‘N, 30°22‘49‘‘E, 1 ex., 25.III.2001; Camyuva 6 km S, stone pine forest, at puddle, 15 m, 36°31‘39‘‘N, 30°31‘39‘‘E, 1 ex., 25.III.2001; near Çatallar, mountain river bank, from moss, under leaves and gravel, 370 m, 36°29‘13‘‘N, 30°04‘56‘‘E, 4 exs., 26.III.2001; Çıralı 3 km W, river shore in deciduous forest, from gravel close to tree root, 71 m, 36°25‘27‘‘N, 30°26‘41‘‘E, 13 exs., 28.III.2001; Söğütcumavi 1 km N, hillside creek, from gravel and leaves, 1463 m, 36°41‘50‘‘N, 30°22‘29‘‘E, 2 exs., 29.III.2001; Kemer 5 km NW, rocky riverbank in canyon, from gravel with leaves and roots, 197 m, 36°36‘03‘‘N, 30°29‘03‘‘E, 2 exs., 30.III.2001, leg. Rose (cRos, cTer). Distribution in Turkey: Istanbul, Manisa.
Oxypoda carbonaria (Heer, 1841) Material: Antalya: Central province, near Ovacık, semi-moist dip with clay soil, in old cow dropping, 1051 m, 36°38‘37‘‘N, 30°26‘14‘‘E, 1 ex., 31.III.2001, leg. Rose (cRos). Distribution in Turkey: Izmir, KahramanmaraĢ.
Oxypoda filiformis L. Redtenbacher, 1849 Material: Antalya: Central province, Göltarya 2 km N, seasoning lake, under stones and sifted from litter near street, 1068 m, 36°34‘20‘‘N, 29°57‘21‘‘E, 1 ex., 26.III.2001, leg. Rose (cRos). Distribution in Turkey: Not locality cited.
Oxypoda flavicornis Kraatz, 1856 Material: Antalya: Akseki, near Güclüköy, single tree on field, litter and formicary sifted, 570 m, 3 exs., 16.III.2000; Central province, Sağırın, Köprülü Kanyon, dry grassland near river, at stones, 30 m, 37°04‘33‘‘N, 31°13‘55‘‘E, 1 ex., 17.III.2002, leg. Rose (cRos). Distribution in Turkey: Istanbul, Izmir, Ordu, Sinop.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 185
Oxypoda longipes Mulsant & Rey, 1861 Material: Antalya: Central province, Saklıkent, slope (SE) with snow fields, nest of small mammal, sifted, 1905 m, 36°50‘24‘‘N, 30°19‘53‘‘E, 1 ex., 18.3III.2002, leg. Rose (cRos). Distribution in Turkey: Hatay, KahramanmaraĢ, Konya, Mersin.
Parocyusa longitarsis (Erichson, 1839) Material: Antalya: Central province, near Çatallar, mountain river bank, from moss, under leaves and gravel, 370 m, 36°29‘13‘‘N, 30°04‘56‘‘E, 3 exs., 26.III.2001; near Çakırlar, riverside, in wet gravel, 259 m, 36°48‘51‘‘N, 30°28‘15‘‘E, 1 ex., 27.III.2001; Çıralı 3 km W, river shore in deciduous forest, from gravel close to tree root, 71 m, 36°25‘27‘‘N, 30°26‘41‘‘E, 1 ex., 28.III.2001; Kemer 5 km NW, rocky riverbank in canyon, from wet mosses, 197 m, 36°36‘03‘‘N, 30°29‘03‘‘E, 15 exs., 30.III.2001, leg. Rose (cRos). Mersin: Central province, Mezitli, Çevikli 2 km S, small stream, 4 exs., 12.VII.2005, leg. AnlaĢ (cAnl). Distribution in Turkey: Izmir, Manisa.
ACKNOWLEDGEMENTS
We are most grateful to our colleagues for their assistance in collecting some of the material and to Prof. Dr. Horst Korge (Berlin, Germany), Dr. Volker Assing (Hannover, Germany), Michael Schülke (Berlin, Germany), and Dr. Christian Maus (Düsseldorf, Germany) for identifications/confirmations of parts of the material.
LITERATURE CITED Anlaş, S. 2009. Distributional checklist of the Staphylinidae (Coleoptera) of Turkey, with new and additional records. Linzer biologische Beiträge, 41 (1): 215-342. Assing, V. 2009. On the Staphylinidae of Turkey. VI. Thirteen new species and additional records (Coleoptera). Koleopterologische Rundschau, 79: 117-172. Fagel, G. 1969. Contribution à la connaissance des Staphylinidae. CIX. Les Melagria Casey de la région paléarctique occidentale. Bulletin Institut Royal des Sciences Naturelles de Belgique, 45 (34): 1-43. Herman, L. H. 2001. Catalog of the Staphylinidae (Insecta: Coleoptera). 1758 to the end of the second millennium. Volumes I-VII. Bulletin of the American Museum of Natural History, 265: 1-4218. Smetana, A. 2004. Subfamily Aleocharinae — In: Löbl I. & Smetana A. (eds), Catalogue of Palaearctic Coleoptera. Volume 2. Hydrophiloidea, Histeroidea, Staphylinoidea. Apollo Books, Stenstrup, 353-494.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 186
GROUND AND TIGER BEETLES (COLEOPTERA: CARABIDAE) FROM KERMAN AND KHORASAN PROVINCES OF IRAN
Hussein Sadeghi*, Somayeh Hasani Saadi*
and Ron Felix** * Department of Plant Protection, Ferdowsi University of Mashhad, IRAN. Email: [email protected]; [email protected] ** Hazelaarlaan 51, 5056 XP Berkel Enschot, NETHERLANDS. Email: [email protected] [Sadeghi, H., Saadi, S. H. & Felix, R. 2011. Ground and tiger beetles (Coleoptera: Carabidae) from Kerman and Khorasan provinces of Iran. Munis Entomology & Zoology, 6 (1): 186-193] ABSTRACT: As a result of a faunistic survey on Ground and tiger beetles which conducted mainly in agroecosystems in different localities of Kerman, Razavi and North Khorasan provinces of Iran, a total of 23 species were identified. Among them, 3 species, namely Nebria xanthacra Chaudoir, Pheropsophus africanus (Dejean), and Poecilus nitens (Chaudoir) are recorded for the first time in Iran. 21 species are new records for Kerman province. Also, 2 species are new for fauna of Khorasan provinces. Data regarding abundance, provincial and geographical distribution of reported species in this study are presented. KEY WORDS: Carabidae, Fauna, New record, Iran.
Carabid beetles, an insect group containing ground and tiger beetles, are an incredibly diverse group of insects with over 40,000 species worldwide (Best & Beegle, 1977; Clark et al., 1994; Lövei & Sunderland, 1996). Members of this family are considered to be mostly opportunistic feeders that consume a variety of foods; however, the majority of species have been observed as primarily predatory, feeding on other insects and related organisms. Most species locate food by random search, although some diurnal species hunt by sight. A few species have also been observed to detect chemical cues from springtails, mollusks, and aphids (Lovei & Sunderland, 1996).
Carabids play a major role in agro ecosystems by contributing to the mortality of a variety of pest organisms including aphids, moth larvae, beetle larvae, mites, and springtails. They have also been used effectively to control slugs in greenhouses (Kromp, 1999). Several ground beetles also are phytophagous. Among the later group, those eat the seeds of troublesome weeds are of particular interest since they help regulate weed populations (Liebman & Gallandt, 1997). At $27 billion per year in management costs, weeds are the most costly pest in North American agriculture (White et al., 2007). For this reason, weed seed consumption or predation by ground beetles is of great interest. Weed seed predation by arthropods such as ground beetles could potentially be used to lower costs associated with weed populations and to increase crop yield. Also, weed seed predators may alter the plant species composition in an area rather than eliminate a weed species. Changing the species composition could give crop species a better chance for success by reducing competition for resources such as light, nutrients, and water, thus increasing crop yields. Understanding weed seed preferences is critical to effectively utilizing weed seed predators in biological control programs.
Although carabid beetles can be found in nearly every available habitat, some species are associated with particular ecosystems, like meadows, woodlands, or crop fields. Because that, the types of carabid beetles found at a location can be a

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 187
valuable biological indicator to assess the impacts of different habitat management and tillage practices.
Compared with the World fauna, the number of recorded carabid species from Iran is less than 1 percent (Löbl & Smetana, 2003; Mohammadzadeh Fard, 2008; Ghahari et al., 2009a,b & Ghahari et al., 2010; Namaghi et al., 2010). In other hand, the carabid fauna in Iran is insufficiently known. Reviewing published papers on carabidae family in Iran (Afshar, 1944; Khajehzadeh, 1998; Alichi & Minaei, 2002a,b; Fallahzadeh et al., 2005; Ghahhari et al., 2009a,b & Ghahari et al., 2010; Hejkal, 2000; Jaeger, 1990; Jaeger, 1992; Lassalle, 2001; Jaskuła, 2007; Magrini & Pavesi, 2003; Mohammadzadeh Fard, 2008; Mohammadzadeh Fard & Hojjat, 2005; Mohammadzadeh Fard & Hodjat, 2008; Namaghi et al., 2010), it is clear that many parts of Iran never been investigated in terms of Carabidae fauna. So, it is expected that further studies will add more information to the knowledge of this family in Iran. The objective of this study was to determine the species composition of ground and tiger beetles inhabiting agro ecosystems and surroundings in Kerman province of Iran. Also, specimens collected from Razavi and North Khorasan provinces were examined.
MATERIALS AND METHODS
This study was conducted between 2009- 2010. The materials from Razavi and North Khorasan provinces were collected by the first author and those from Kerman province were collected by second author, where no detailed studies have been carried out. The majority of the collections were made using pitfall traps. Also, additional specimens collected directly by hands when walking through the agro ecosystems and the surroundings. Data, such as number of specimens, locations and dates were recorded. The majority of identifications were done by the third author.
The voucher specimens are deposited in the department of Plant Protection, College of Agriculture, Ferdowsi University of Mashhad, Iran. Also, a few paratypes are held in personal collection of the third author.
The nomenclature of the Carabidae is given follows the data base www.faunaeur.org and Löbl & Smetana (2003).
RESULTS
In this study, a total of 23 species of 17 genera and 10 subfamilies were recorded. They are as bellow list. Subfamily Pterostichinae Bonelli, 1810 Anthia (Anthia) mannerheimii Chaudoir, 1842 Material: Razavi Khorasan (Mashhad), 59º, 40' E, 36º ,14' N, 2 ♀♀ and 1♂, 10August 2008; Kerman (Bam), 58 º, 20' E, 29 º,7' N, 1♀, 30 Oct. 2009. Previous provincial records for Iran: Khorasan, (Afshar, 1944). General distribution: Iran, Turkmenistan (Löbl & Smetana, 2003). Anthia (Thermophilum) duodecimguttata Bonelli, 1813 Material: Kerman (Bam), 58º, 20' E, 29 º, 7' N, 2 ♀, 30 Oct. 2009. Previous provincial records for Iran: Mazandaran (Ghaemshahr), Semnan (Garmsar), (Ghahari et al., 2009a; Ghahari et al 2010). General distribution: Egypt, Iran, Iraq, Saudi Arabia, Yemen (Löbl & Smetana, 2003).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 188
Amara aenea (DeGeer, 1774) Material: Kerman, 57 º, 1' E, 30 º, 3' N, 1♀, 3 Sept. 2009. Previous provincial records for Iran: Fars, Kohkeyloye and Boyer Ahmad Golestan, Khorasan (Mohammadzadeh Fard & Hodjat, 2008; Ghahari et al., 2009a). General distribution: Palearctic Region, North America, Caucasia (Löbl & Smetana, 2003). Poecilus nitens (Chaudoir, 1850) Material: Kerman, 57 º, 1' E, 30 º, 3' N, 2♀♀, 21 Aug. 2009. Previous provincial records for Iran: new record for Iranian fauna. General distribution: Armenia, Russia south, Uzbekistan, Turkmenistan, Kazakhstan, Tadjikistan (Löbl & Smetana, 2003).
Subfamily Harpalinae Bonelli, 1810 Acinopus picipes (Olivier, 1795) Material: Kerman (Jiroft), 58 º,3' E, 28 º,3' N, 3♀♀, 30 Oct.2009; Kerman (Zarand), 57º, 03' E, 30, 81' N, 1♀, 10 Oct. 2009. Previous provincial records for Iran: Fars, Razavi Khorasan, Mazandaran (Alichi & Minaie, 2002; Namaghi et al., 2010). General distribution: Azarbaijan, Albania, Armenia, Bosnia Herzegovina, Bulgaria, Croatia, France (incl. Corsica, Monaco), Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Malta, Macedonia, Moldavia, Portugal, Romania, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Turkey, Ukraine, Serbia and Montenegro, Cyprus, Iran, Iraq, Israel, Syria, Turkey (Löbl & Smetana, 2003). Pseudoophonus rufipes (DeGeer, 1774) Material: Kerman, 57 º, 1' E, 30 º, 3' N, 1♀, 21 Aug. 2009. Previous provincial records for Iran: Tehran (Varamin), Ardabil (Dasht-e-Moghan), Mazandaran (Ghaemshahr), Fars (Darab), Razavi Khorasan (Kashmar and Mashhad), Semnan (Garmsar) (Ghahari et al., 2009a, 2010). General distribution: Azerbaijan, Albania, Armenia, Austria, Azores, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Croatia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France , Great Britain , Germany, Georgia, Greece, Hungary, Ireland, Italy, Kazakhstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Malta, Macedonia, Moldavia, Netherlands, Norway, Poland, Portugal, Romania, Slovakia, Slovenia, Spain, Russia: South European Territory, Sweden, Switzerland, Turkey, Ukraine, Serbia & Montenegro, Algeria, Morocco, Western Sahara, Tunisia, Afghanistan, Cyprus, Russia: East Siberia, Iran, Iraq, Kyrgyzstan, Kazakhstan, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, West Siberia, Xinjiang (Sinkiang), Nearctic Region (Löbl & Smetana, 2003). Pseudoophonus griseus (Panzer, 1796) Material: Kerman (Baghin, Ghaemabad) (57° E, 30° N), 2♀♀, 11July2009; Jiroft (Dalfard)(57°E, 28°N), 1♀, 12 October 2009. Previous provincial records for Iran: Mazandaran (Ghaemshahr), Golestan(Gonbad), Razavi Khorasan (Kashmar, Bardaksan, Mashhad, Dargaz, Serakhs), South Khorasan (Ghaenat, Ferdows) (Namaghi et al., 2010; Ghahari et al., 2009a, 2010). General distribution: Russia, Moldova, Ukraine, Crimea, Azarbaijan, Daghestan, Armenia, Uzbekistan, Kazakhstan, Turkmenistan, Tajikistan, North and West Africa.
Subfamily Scaritinae Bonelli, 1810 Distichus planus (Bonelli, 1913) Material: Kerman (Zarand), 57 º, 03' E, 30 º, 81' N, 2♀♀, 10 Oct. 2009. Previous provincial records for Iran: Tehran, Razavi Khorasan(Mahvelat) (Ghahhari et al., 2009a; Namaghi et al., 2010). General distribution: Azerbaijan, France, Georgia, Greece, Italy, Malta, Portugal, Spain, Turkey, Egypt, Morocco, Western Sahara, Tunisia, Iran, Iraq, Jordan, Kazakhstan, Pakistan,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 189
Saudi Arabia, Syria, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Yemen (Löbl & Smetana, 2003). Scarites (Parallelomorphus) eurytus (Fischer von Waldheim, 1825) Material: Kerman (Baft), 56 º, 36' E 32 º, 17' N, 4 ♀♀ & 1 ♂, 14 Aug.2009 and 21 Aug. 2009. Previous provincial records for Iran: Fars; Kohkeyloye and Boyer Ahmad (Mohammadzadeh Fard & Hodjat, 2008). General distribution: Russia, Armenia, Kazakhistan, Turkmenistan, Uzbekistan, Mountains of Middle Asia, Syria and Israel (Löbl & Smetana, 2003).
Subfamily Nebriinae Laporte, 1834 Nebria xanthacra Chaudoir, 1850 Material: Kerman, 57 º, 1' E, 30 º, 3' N, 1♀, 3 Sept. 2008. Previous provincial records for Iran: this is a new record for fauna of Iran. General distribution: Asia: India, Turkmenistan (Löbl & Smetana, 2003).
Subfamily Callistinae Laporte, 1834 Chlaenius festivus (Panzer, 1796) Material: Kerman, 57 º, 1' E, 30 º, 3' N, 1♀, 10 Sept. 2009. Previous provincial records for Iran: Kohkeyloye and Boyer Ahmad, Khozestan( Ahvaz), Golestan (Salikandeh), Razavi Khorasan (Kashmar), (Mohammadzadeh Fard& Hodjat, 2008; Shafiei et al.,2004; Ghahari et al., 2009). General distribution: Middle Asia, Central and South Europe, Turkey, Caucasia, Transcaucasia, Iran (Löbl & Smetana, 2003). Pheropsophus africanus (Dejean, 1825) Material: Kerman, 57 º,1' E, 30 º, 3' N, 2♀♀ & 1♂, 13 Sept. 2009. Previous provincial records for Iran: new record for fauna of Iran. General distribution: Africa: Algeria, Libya, Morocco, Tunisia, Asia: Israel, Egypt (Löbl & Smetana, 2003).
Subfamily Broscinae Hope, 1838 Broscus (Cephalotes) laevigatus Dejean, 1828 Material: Kerman, 57 º, 1' E, 30 º, 3' N, 2♀♀, 21 Aug. 2009. Previous provincial records for Iran: Ardabil (Dasht-e-Moghan), Tehran (Varamin), Fars (Darab), East Azarbaijan (Arasbaran), Semnan (Garmsar) (Ghahari et al., 2009a). General distribution: Armenia, Austria, Belgium, Bosnia and Herzegovina, Bulgaria, Russia central, Czech, Denmark, , Finland, France, Great Britain, Germany, Georgia, Hungry, Ireland, Italy, Latvia, Luxemburg, Norway, Poland, Romania, Slovak, Slovenia, Switzerland, West Russia, Turkey, West Siberia, Nearctic's region(Löbl & Smetana, 2003).
Subfamily Carabinae Latreille, 1802 Calosoma (Caminara) olivieri Dejean, 1831 Material: Kerman (Zarand), 57 º,03' E, 30 º,81' N, 1♀, 15Oct.2009; Kerman (Anar), 55 º, 35' E, 30 º, 32' N, 3♀♀, 29 July 2009; Kerman (Jiroft), 58 º,3' E, 28 º,3' N, 4♀♀, 24 Oct. 2008. Previous provincial records for Iran: Razavi Khorasan (Kashmar), Mazandaran (Behshahr), Golestan (Ramian), Khozestan (Ahvaz), (Ghahari et al., 2009a; Shafiei et al., 2004). General distribution: Russia, Tajikistan, Turkey, Turkmenistan and Uzbekistan (Löbl & Smetana, 2003). Calosoma (Callistriga) algiricum Géhin, 1885 Material: Kerman, 57 º, 1' E, 30º, 3' N, 2♀♀, 3 July 2009. Previous provincial records for Iran: East Azarbaijan (Arasbaran) (Ghahari et al., 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 190
General distribution: Italy (Sicilia), North Africa: Algeria, Libya, Morocco, Western Sahara, Tunisia, and Asia: Iran, Israel, Jordan, Saudi Arabia, and Turkmenistan (Löbl & Smetana, 2003).
Subfamily Platyninae Bonelli, 1810 Sphodrus leucophthalmus (Linnaeus, 1758) Material: Kerman, 57º, 1' E, 30º, 3' N, 2♀♀, 3 Sept. 2009. Previous provincial records for Iran: Golestan (Nokandeh), Khozestan (Ahvaz), (Ghahari et al., 2009a; Shafiei et al., 2004). General distribution: North Africa, South and West Asia, Europe, Turkey, Caucasia, India and Canary Island (Löbl & Smetana, 2003). Calathus (Neocalathus) cinctus Motschulsky, 1850 Material: Kerman (Baghin-Ghaemabad) (57° E, 30 °N), 2♀♀ & 2♂♂, 21 July 2009. Previous provincial records for Iran: Fars, Kohkeyloye & Boyer Ahmad (Mohammadzadeh Fard & Hodjat, 2008). General distribution: Albania, Austria, Belgium, Bosnia and Herzegovina, Britain.Incl. Shetlands, Orkneys, Hebrides and Man Is., Bulgaria, Corsica, Crete, Croatia, Cyclades Is., Cyprus, Czech Republic, Danish mainland, Dodecanese Is., Estonia, European Turkey, French mainland, Germany, Greek mainland, Hungary, Ireland, Italian mainland, Latvia, Liechtenstein, Luxembourg, Macedonia, Malta, Moldova, Republic of, Northern Ireland, Norwegian mainland, Poland, Portuguese mainland, Romania, Russia East, Russia North, Russia Northwest, Russia South, Sardinia, Sicily, Slovakia, Slovenia, Spanish mainland, Sweden, Switzerland, The Netherlands, Ukraine, YugoslaviaIncl. Serbia, Kosovo, Voivodina, Montenegro, East Palaearctic, Near East: Asian Turkey, Caucasian Russian republics, Georgia, Armenia, Azarbaidjan, Lebanon, Syria, Israel, Jordan, Sinai Peninsula (Egypt), Arabian peninsula, Iran, Iraq, North Africa Not including Sinai Peninsula (www.faunaeur.org).
Subfamily Lebiinae Bonelli, 1810 Lebia (Lamprias) cyanocephalus Linnaeus, 1758 Material: Kerman (Baghin-Ghaemabad) (57° E, 30° N), 3♀♀ & 1♂, 21 July 2009. Previous provincial records for Iran: Fars, Kohkeyloye & Boyer Ahmad (Mohammadzadeh Fard & Hodjat, 2008). General distribution: Albania, Austria, Balearic Is.Incl. Mallorca I., Menorca I., and Pityuses Is., Belarus, Belgium, Bosnia and Herzegovina, Britain Incl. Shetlands, Orkneys, Hebrides and Man Is., Bulgaria, Corsica, Croatia, Cyprus, Czech Republic, Danish mainland Incl. Borholm I., Dodecanese Is., Estonia, European Turkey, Finland, French mainland, Germany, Greek mainland, Hungary, Italian mainland, Kaliningrad Region, Latvia, Liechtenstein, Lithuania, Luxembourg, Macedonia, Moldova, Republic of, Norwegian mainland, Poland, Portuguese mainland, Russia Central, Russia North, Russia Northwest and south, Sicily, Slovakia, Spanish mainland Incl. Alboran I. , Sweden Incl. Gotland I, Switzerland, The Netherlands, Ukraine, Yugoslavia Incl. Serbia, Kosovo, Voivodina, Montenegro, East Palaearctic, Near East Asian Turkey, Caucasian Russian republics, Georgia, Armenia, Azarbaijan, Lebanon, Syria, Israel, Jordan, Sinai Peninsula (Egypt), Arabian peninsula, Iran, Iraq, North Africa Not including Sinai Peninsula (www.faunaeur.org).
Subfamily Cicindelinae Latreille, 1802 Megacephala euphratica Dejean in Latreille & Dejean, 1822 Material: Kerman (Jiroft), 58 º, 3' E, 28 º, 3' N, 2♀♀, 4 July 2009. Previous provincial records for Iran: it has been reported from Iran, but the locality is unknown. General distribution: Spanish mainland, Crete, Dodecanese Is., Cyprus, East Palaearctic, Near east: Asian Turkey, Caucasian Russian republics, Georgia, Armenia, Azerbaidjan,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 191
Lebanon, Syria, Israel, Jordan, Sinai Peninsula (Egypt), Arabian peninsula, Iran, Iraq and North Africa: Egypt, Tunisia, Libya, Morocco, Algeria, Yemen (www.faunaeur.org). Cicindela asiatica Audouin & Brullé, 1839 Material: North Khorasan (Bojnord), 57º, 20' E , 37º, 28' N, 3♀♀, 4May 2006. Previous provincial records for Iran: Khorasan (Kashmar), (Ghahari et al., 2009a). General distribution: Russia, Caucasia (Löbl & Smetana, 2003). Cicindela clypeata Fischer von Waldheim, 1821 Material: Razavi Khorasan (Mashhad), 59º, 40' E, 36 º, 14' N, 2♀♀ & 1♂, 1 Sep. 2009. Previous provincial records for Iran: this species has been recorded for Iranian fauna, but the locality is unknown. General distribution: Iran, Afghanistan, Kazakhstan, Uzbekistan, Tajikistan, Turkemmania(Löbl & Smetana, 2003) and Russia South (www.faunaeur.org). Cicindela germanica (Linnaeus, 1758) Material: Razavi Khorasan (Chenaran), 59º, 40' E, 36 º,14' N, 2♀♀, 6 May 2009. Previous provincial records for Iran: Northern and Northwestern provinces of Iran (Afshar, 1944). General distribution: Austria, Albania, Andorra, Yugoslavia Incl. Serbia, Kosovo, Voivodina, Montenegro, Ukraine,The Netherlands, Switzerland, Slovenia, Spanish mainland, Slovakia, SicilyIncl. adjacent Italian islands (Lipari Is., Ustica, Egadi Is., Pantelleria I., Pelagie Is.), Russia South, Russia North, Poland, Romania, Moldova, Republic of, Estonia, Macedonia, Luxembourg, Lithuania, Liechtenstein, Latvia, Italian mainland, Ireland, Hungary, Greek main land Incl. Andikithira I., Evvia I., Ionian Is., Samothraki I., Northern Sporades Is., French mainland, Germany, Finland, European Turkey, Czech Republic, Croatia, Bulgaria, Britain Incl. Shetlands, Orkneys, Hebrides and Man Is., Bosnia and Herzegovina, Belgium, East Palaearctic (www.faunaeur.org). Cicindela histrio (Tschitscherine, 1903) Material: Kerman (Zarand), 57 º, 03' E, 30 º, 81' N, 2♀♀ & 1♂, 2 Aug. 2009. Previous provincial records for Iran: according to Lobl & Smetana (2003), this species has been reported from Iran, but the locality is unknown. General distribution: Amman, Saudi Arabia, Pakistan, Afghanistan, Iran, Arabian Emirates (Löbl & Smetana, 2003).
DISCUSSION
Studies on the structure of fauna of carabid beetles and abundance of
particular species are important as this large taxon has an important role in sustainable agriculture. Many species of this family are considered as predators of other arthropods (Kromp, 1999; Clark et al., 1994; Best & Beegle, 1977). Some species consume a variety of weed seeds that can decrease costs associated with weed controls (Liebman & Gallandt, 1997). Carabid beetles can play a significant role in ecologically-based integrated pest management programmes that focus on avoidance or reduction of pest pressure through cultural practices and biological controls. Moreover, being well known both taxonomically and ecologically, carabids are extremely sensitive to several abiotic and biotic factors, respond quickly to habitat alteration and can be easily and cost-effectively collected. For these reasons, ground beetles are increasingly being used in ecological studies in order to evaluate the environmental impacts of man in terrestrial ecosystems (Avgın & Luff, 2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 192
CONCLUSSION
This study showed that the Carabidae fauna of Iran is much more than that has been reported and can be higher after more comprehensive works. Moreover, the economic importance and roles played by the carabid species occurring in different ecosystems of Iran is unclear. So in addition to further faunistic surveys, detailed biological and ecological studies are waiting to be carried out.
ACKNOWLEDGEMENTS
This study was supported partly by Ferdowsi University of Mashhad who is gratefully acknowledged.
LITERATURE CITED Afshar, J. 1944. Les nomes scientifique de quelques coleopteres de l' Iran et leurs importance en agriculture. Min. Agri. Tehran. 22 pp. Alichi, M. & Minaei, K. 2002a. New records on the fauna of the Carabidae in Fars province, 175-176. Proceedings of the 15th Iranian Plant Protection Congress, Razi University, Kermanshah. Alichi, M. & Minaei, K. 2002b. Study on distribution of the beetles belonging the family Carabidae in Shiraz region, 175. Proceedings of the 15th Iranian Plant Protection Congress, Razi University, Kermanshah. Avgın, S. S. & Luff, M. L. 2010. Ground beetles (Coleoptera: Carabidae) as bioindicators of human impact. Munis Entomology & Zoology, 5 (1): 209-215. Best, R. L. & Beegle, C. C. 1977. Food preferences of five species of carabids commonly found in Iowa cornfields. Environmental Entomology, 6 (1): 9-12. Clark, M. S., Luna, J. M., Stone, N. D. & Youngman, R. R. 1994. Generalist predator consumption of armyworm (Lepidoptera: Noctuidae) and effect of predator removal on damage in no-till corn. Environmental Entomology, 23 (3): 617-622. Fallahzadeh, M., Shojace, M. & Ostovan, H. 2005. Report of Broscus punctatus (Col.: Carabidae) from Iran. Journal of Entomological Society of Iran, 24 (2): 140. Ghahari, H., Kesdek, M., Samin, N., Ostovan, H., Havaskary, M. & Imani, S. 2009a. Ground beetles (Coleoptera:Carabidae) of Iranian cotton fields and surrounding grasslands. Munis Entomology & Zoology, 4 (2): 436-450. Ghahari, H., Wojciech, B., Czkowsi, J., Kesdek, M., Otovan, H. & Tabari, M. 2009b. Ground beetles (Coleoptera: Carabidae) from rice fields and surrounding grasslands of Northern Iran. Journal of Biological Control, 23 (2): 105-109. Ghahari, H., Avgin, S. S. & Ostovan, H. 2010. Carabid beetles (Coleoptera: Carabidae) collected from different ecosystems in Iran with new records. Türk. entomol. derg., 34 (2): 179-195. Hejkal, J. 2000. Amara (Amara) elborzensis sp. n. (Coleoptera: Carabidae) from Iran. Klapalekiana. 36 pp. Jaeger, B. 1990. Zur Verbreitung von Bradycellus csikii Laczo, 1912 und B. distinctus Dejean, 1829 sowie Beschreibung von Bradycellus heinzei n.sp. aus demNord-Iran (Col., Carabidae). Entomologische Nachrichten und Berichte, 34: 9-13. Jaeger, B. 1992. Beitrag zur Erforschung der Acupalpus-Fauna des Iran und der Türkei nebst Beschreibung von Acupalpus turcicus n. sp. (Col., Carabidae). Entomologische Nachrichten und Berichte, 36: 223-230. Jaskuła, R. 2007. First record of Lebia trimaculata from Iran (Coleoptera:Carabidae). Entomological Problems, 37 (1-2): 48.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 193
Khajehzadeh, Y. 1998. Investigation on the most important injurious beetles of cereal fields of Khuzestan, the Proceedings of 13th Iranian Plant Protection Congress. Kromp, B. 1999. Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation impacts and enhancement. Agriculture, Ecosystems and Environment, 74: 187-228. Lassalle, B. 2001. Les Sphodristocarabus d‘Iran (Coleoptera, Carabidae). Le Coléoptériste, 43: 169-170. Löbl, I. & Smetana, A. 2003. Catalogue of Palaearctic Coleoptera. Volume I. Archostemata-Myxophaga-Adephaga. ApolloBooks. Stenstrup, Denmark, 819 pp. Lovei, G. L. & Sunderland, K. D. 1996. Ecology and behavior of ground beetles(Coleoptera: Carabidae). Annual Reviews Entomology, 41: 231-256. Magrini, P. & Pavesi, M. 2003. A new Iranian species of Neoblemus (Coleoptera: Carabidae). Bollettino della Societa Entomologica Italiana, 135 (2): 71-78. Mohammadzadeh Fard, S. 2008. Description of female Axonya farsica (Coleoptera: arabidae: Broscinae). Journal of Entomological Society of Iran, 27 (2), Supplement, 3-4. Mohammadzadeh Fard, S. & Hodjat, S. H. 2005. New reocrds on Carabid beetles in Iran. Journal of Entomological Society of Iran, 24 (2). Mohammadzadeh Fard, S., Hodjat, S. H. 2008. A comparison survey on fauna of Family Carabidae ( Insecta: Coleoptera) in natural and agricultural areas. 18th Iranian Plant Protection Congress, Bu-Ali Sina University, Hamadan. p.89. Namaghi, S. H., Avgin, S. S. & Farahi, S. 2010. New Data To The Knowledge Of Ground Beetles (Coleoptera: Carabidae) Fauna of Iran. Journal of Turkish Entomology, 34 (2): 197-210. White, S. S., Renner, K. A., Menalled, F. D. & Landis, D. A. 2007. Feeding preferences of weed seed predators and effect on weed emergence. Weed Science, 55: 606-612.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 194
REDESCRIPTION OF DIAERETIELLA RAPAE (M’INTOSH) (HYMENOPTERA: BRACONIDAE: APHIDIINAE) WITH
MORPHOLOGICAL VARIABILITY OF SEVERAL POPULATIONS FROM INDIA
Mir Samim Akhtar*, Debjani Dey* and Mohd Kamil Usmani**
* Division of Entomology, Indian Agricultural Research Institute, New Delhi, INDIA. E-mail: [email protected] ** Department of Zoology, Aligarh Muslim University, Aligarh, INDIA. [Akthar, M. S., Dey, D. & Usmani, M. K. 2011. Redescription of Diaeretiella rapae (M‘Intosh) (Hymenoptera: Braconidae: Aphidiinae) with morphological variability of several populations from India. Munis Entomology & Zoology, 6 (1): 194-203] ABSTRACT: Diaeretiella rapae (M‘Intosh) is redescribed with additional of recording morphometric ratios, in addition to original measurement of different body parts. Four populations of D. rapae were investigated on the basis of ten character ratios, using numerical methods. The analysis of populations indicated that the specimens reared from different localities showed clear distinction from other populations. KEY WORDS: Aphidiinae, Diaeretiella rapae, morphological variability, systematics.
Aphid parasitoids belonging to the braconid subfamily Aphidiinae (Hymenoptera) are solitary endoparasitoids of aphids (Stary, 1970; 1988). Diaeretiella rapae (M‘Intosh), a polyphagous and cosmopolitan parasitoids of aphids and very important control agents for aphid pest in a variety of agricultural and horticultural crops (Hagvar and Hofsvang, 1991), most important factor for natural control of mustard aphid (Dhiman, 2007; Dogra et al., 2003; Pike et al., 1999). It was exported from China to the USA for biological control of Diuraphis noxia (Mordwilko), a pest of small grains (Bernal and Gonzalez, 1997). Furthermore, D. rapae parasitizing several aphid taxa viz., Aphis ruborum longisetosus, Aphis craccivora, Aphis gossypii, Brachycaudus helichrysi, Brevicoryne brassicae, Hayhurstia atriplicis, Lipaphis erysimi, Macrosiphoniella pseudoartemisae, Macrosiphum (Metopolophium) drihodun, Macrosiphum (Sitobion) rosaeformis, Myzus persicae, Sitobion avenae eleusinae are recorded from India (Dey & Akhtar, 2007). Knowledge of their taxonomy and diversity has great importance for successful biological control programs.
The aim of the present paper is redescription of D. rapae with addition morphmetric ratio which was not present in earlier description along with original measurement of different body parts and taxonomic study using numerical methods of several population of D. rapae from selected pest aphids. In our study we have used taxonomic characters with a high content of diagnostic information.
MATERIAL AND METHODS
The specimens for the present study were collected from various parts of India and deposited in National Pusa Collection (NPC), New Delhi. The essential diagnostic characters of the specimens were examined without any special preparation under Leica MZ12 stereo zoom microscope with suitable illumination. Photos were taken with a Leica MZ16 stereomicroscope with integrated Leica photo camera. The captured images were assembled with CombineZP programme

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 195
and edited in Adobe Photoshop Elements 2.0. The nomenclatural treatment, morphological terminology and taxonomic characters adopted from Sharkey and Wharton, 1997.
Ten specimens in good condition were selected from four populations of Diaeretiella rapae (M‘Intosh) (viz., LMNNT= Lipaphis erysimi on mustard from Nainital, LMDER= Lipaphis erysimi on mustard from Dehra Dun, LMDEL= Lipaphis erysimi on mustard from Delhi and MCDEL= Myzus persicae on cabbage from Delhi). All specimens were individually numbered and kept aside for recording their measurements.
Eighteen taxonomic characters with a high content of diagnostic information relevant to the parasitoid, D. rapae were selected and measurements were taken. All measurements were expressed as ratios as recommended by Tomnic et al. (2005) which were then used for matrix analysis. All the characters and ratios used have been listed in Table 1. The morphometric data was recorded for four populations of D. rapae from three different localities (viz., New Delhi, Nainital and Dehradun); on two different hosts (viz., L. erysimi and M. persicae); on two different host plants (viz., mustard and cabbage). The data was then subjected to cluster analysis using SPSS 10.0.5 software from which a dendrogram was constructed.
RESULTS AND DISCUSSION Genus Diaeretiella Stary, 1960: 242-243.
Type species: Aphidius rapae M‘Intosh, 1855 Diagnosis: Head transverse to subquadrate, wider than mesosoma at tegulae; occiput margined, antenna filiform with 14 segment in female and 18 in male; eyes medium oval; mandibles bidentate. Notauli developed on ascending part of mesoscutum; stigma large triangular; metacarp (R1) longer than stigmal width; radial sector long; marginal cell incomplete; cubital vein effaced beyond basal cell towards the apex of fore wing; hindwing without any complete cell; costal and subcostal vein developed; propodeum areolated with narrow small central areola. Metasoma of female lanceolate and rounded at apex in male; ovipositor sheaths curved upwards, sparsely hairy.
Diaeretiella rapae (M’Intosh, 1855) (Fig. 1-8)
?Aphidius vulgaris Bouche, 1834: 161-162. Aphidius rapae M‘Intosh, 1855: 194. Aphidius (Trionyx) rapae Curtis, 1860: 73-74. Diaeretus chenopodii Foerster, 1867:125. Trioxys piceus Cresson, 1880:260. Lipolexis chenopodiaphidis Ashmead, 1889: 671. Aphidius brassicae Marshall, 1896: 597-598. Diaeretus californicus Baker, 1909: 25. Diaeretus nipponensis Viereck, 1911: 182. Diaeretus obsoletus Kurdjumov, 1913: 25-26. Diaeretus napus Quilis, 1931: 71-72. Diaeretus croaticus Quilis, 1934: 8-9. Diaeretus plesiorapae Blanchard, 1940: 45-48. Diaeretus aphidum Mukerji and Chatterjee, 1950: 4-6.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 196
Female (Fig. 7): Colour: Head frons and occiput brown; antenna brown; scape, pedicel and base of flagellar segment I yellow. Mesosoma yellow; lobes with slightly brown spot; wing venation brown; legs yellow; femur, tibia and tarsi darkened. Tergum I yellow; remaining metasoma brownish. Head (Fig. 3,4) transverse to subquadrate, smooth, shiny, sparsely hairy wider than mesosoma at tegulae, 0.92x wider than width of mesoscutum; occiput margined. Eyes (Fig. 3, 4) medium sized, nearly hemispherical with sparse hairs, prominent, strongly convergent towards clypeus; transverse diameter of eye as wide as width of temple; width of gena as wide as 0.24x longitudinal eye diameter; inter-tentorial line 0.37x of facial line, 2.2x as long as tentori-occular line; clypeus transverse, oval, convex, margined frontally, smooth shiny with six to eight long hairs, separated from face by shallow grooves; mandibles broad basally, uniformly narrowed towards apex. Ocelli medium sized, forming an equilateral triangle. Antenna filiform (Fig. 3, 4), 14 segmented, thickened towards apex, about as long as head, mesosoma and half of metasoma combined; antennal socket ocular line 0.43x of socket diameter; scape and pedicel sub-equal to their maximum width; flagellar segment I and II almost 2.26x as long as its maximum width; flagellar segment II 1.30x as long as length of penultimate segment; length of apical segment sub-equal flagellar segment II.
Mesosoma smooth, shiny, sparsely hairy; mesoscutum falling almost vertically to pronotum, without covering it when viewed from side; length of mesosoma 1.37x its height and 1.54x its width, notauli distinct anteriorly, narrow, crenulate but effaced on disc, fore margin slightly prominent. Forewing (Fig. 1) hyaline, length 2.91x its maximum width; stigma triangular, 4.75x longer than wide; length of metacarp (R1) 0.34x length of stigma; ‗r‘ arising from about basal 0.50 stigma, about 0.25 as long as stigmal width; marginal cell incomplete; r-m and mcu absent; submarginal cells confluent. Hindwing (Fig. 2) more than 4.80x as long as wide, SC+R prominent. Legs with hind femur 5.50x as long as wide; length of hind tibia 1.50x length of hind femur; length of hind tarsal segment I 0.60x length of remaining segments combined; length of hind tarsal segment II to V gradually small.
Propodeum (Fig. 5) with small narrow central areola; disc of areola smooth, shiny; transverse carinae arising from either side of mid lateral part of areola reaching spiracles; median carina originating on top of areola run up to postscutellum; few small carinae present around basal region of areola; propodeal spiracle located on lateral propodeal margin.
Metasoma lanceolate, longer than head mesosoma combined; tergum I slender (Fig. 6), costulate, slightly granulate, dilated towards apex, more than 3.43x longer than wide at spiracles, prominent lateral longitudinal carina, more or less coarsely rugose anteriorly, posterior 0.25 smooth; spiracular tubercles poorly visible, sparsely hairy; combined median length of tergum II and III 2.00x of basal with of tergum II, following tergum smooth shiny, sparsely hairy.
Genitalia: Ovipositor sheath straight or slightly curved upwards, about 2.22x longer than its maximum width, bearing 4 to 6 setae; ovipositor curved upwards, ending in a pointed tip.
Measurements (mm±SD): Body length 1.94±0.07; tenterio-ocular line 0.05±0.01, intertentorial line 0.11±0.01, inter-ocellar line 0.12±0.01, facial line 0.30±0.03, tenterio-facial line 0.22±0.01, width of gena 0.05±0.01, longitudinal eye diameter 0.21±0.03, transverse eye diameter 0.11±0.02, width of temple

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 197
0.13±0.02, length of F I 0.13±0.01, width of F I 0.05±0.01, length of F II 0.13±0.01, width of F II 0.05±0.01, length of penultimate segment of antenna 0.10±0.01, length of apical segment of antenna 0.21±0.01, length of stigma 0.38±0.03, width of stigma 0.08±0.01, length of metacarp (R1) 0.13±0.02, length of radial vein 0.06±0.01, length of tergum I 0.24±0.02, width of tergum I 0.07±0.01, length of ovipositor sheath 0.12±0.01, width of ovipositor sheath 0.05±0.01. Male (Fig. 8): Antenna 16 segmented, colour darker than female; mouthparts and tergum I yellowish; leg brownish black; other colourations as female. Mummy: Dark shining brown. Material examined: 120♀♀ and 30 ♂♂; INDIA: 60 ♀♀and 10 ♂♂: Delhi, IARI, 14. II. 2007, 06. III. 2007, 21. II. 2008, parasitic on Lipaphis erysimi on Mustard, Coll. Mir Samim Akhtar; 20 ♀♀ and 5 ♂♂: Himachal Pradesh, Palampur, 28. III. 2007, parasitic on Lipaphis erysimi on Mustard, Coll. Ajay Kumar; Uttarakhand, 25 ♀♀and 10 ♂♂: Nainital, 17. I. 2007, parasitic on Lipaphis erysimi on Mustard Coll. Poonam Dev; 15 ♀♀and 5 ♂♂: Uttarakhand, Dehradun, 28. II. 2007, parasitic on Lipaphis erysimi on Mustard, Coll. Mir Samim Akhtar. Distributions: Assam, Himachal Pradesh, Delhi, Jammu and Kashmir, Karnataka, Manipur, Meghalaya, Punjab, Sikkim, Uttar Pradesh. Morphological variability:
The extent of variation among the various character ratios as indicated by correlation matrix (Table 3) revealed all interactions between various character ratios to be significantly different from each other except that between TL/TW and MS/LED (r=-0.54). However among all the significantly different ratios maximum interaction was shown between the ratios F1T/F2T and F2L/F2W (r=0.45) followed by F2L/F2W and F1L/F1W (r=0.44); F1L/F2L and F2L/F2W (r=-0.41); M1T/M2T and TL/TW (r=-0.41). Significantly lower interaction was shown by all the others, least being shown in four pairs of ratios, i.e. F1L/F2L and MS/LED(r=0.02); F1L/F2L and TO/IT (r=-0.02); M1T/M2T and F2L/F2W (r=0.02); M1T/M2T and F1L/F1W (r=-0.02).
The ratio of length of malar space: longitudinal eye diameter i.e. MS/LED showed minimum deviation followed by the TO/IT ratio (tentorio-ocular line: inter tentorio line) among all the individuals of all the four populations of D. rapae studied. Although the ratio of F1L/F2L showed less deviation among the individuals of two populations viz., LMDER and LMDEL but showed more deviation among the members of the other two populations viz., LMNNT and MCDEL. Similarly ratio of F1T/F2T showed less deviation in the two populations viz., LMNNT and LMDEL and more deviation among the members of the other two populations viz., LMDER and MCDEL. The remaining character ratios did not show any particular trend as indicated by the differential values of standard deviations. Therefore from the data it can be concluded that among all the character ratios studied only two i.e. MS/LED and TO/IT can be considered as stable characters (Table 2).
All the four populations were significantly different from each other (F= 7.179; p= 0.0001) (Table 4). However, the cluster of the four populations (Fig. 9), showed that two of them viz., LMDEL and MCDEL to be very closely clustered

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 198
(Square Euclidian Distance (SED)= 0.620; both the populations had been collected from the same locality but host plants are different.
Again both the above two populations were closer to LMDER compared to the fourth population LMNNT both from different localities. Among the two populations LMDEL and MCDEL, MCDEL was closer to LMDER (SED= 0.713; localities are different). Comparisons among populations reveal significant differences, viz., LMNNT and LMDER (SED= 1.271), LMDER and LMDEL (SED= 1.534) wherein the localities of collection were different; while among LMNNT and LMDEL (SED= 1.628) and LMNNT and MCDEL (SED= 1.683) both host plants and localities of collection were different.
Analysis of various morphometric ratios by two way multiple factor ANOVA indicated significant variability among D. rapae populations from different places, hosts and host plants (Table 5). A clear cut difference among the reared populations of D. rapae from Delhi, Nainital and Dehradun could be observed. D. rapae females parasitizing Lipaphis erysimi on mustard from Nainital (Uttarakhand in northern hill zone) showed distinct variations among some of the characters studied, viz., in general size was larger, with almost concave temples, a semi-oval head, well developed notauli at least anteriorly, propodeal areola with numerous irregular carinae etc. Cluster analysis also revealed that all the four populations were significantly different from each other (F= 7.179; p= 0.0001) (Table 5). However, the cluster of the four populations (Fig. 12), showed that populations collected from Delhi and Dehradun were closer to each other than to the population from Nainital. Further two of the populations from the same locality Delhi but from two different hosts, L. erysimi and M. persicae and also different host plants, mustard and cabbage were very closely clustered [Square Euclidian Distance (SED) = 0.620]. Again both the above two populations were closer to the population collected from Dehradun but on the host L. erysimi on mustard compared to the Nainital population which was on the same host and host plant. Therefore it can be inferred that the place of occurrence might have an influence on the morphological characters as Nainital is situated on a higher altitude compared to both Delhi and Dehradun.
ACKNOWLEDGEMENTS
We would like to thank Dr. V.V. Ramamurthy, Principal Investigator, Network Project on Insect Biosystematics (NPIB 21-17), Division of Entomology, Indian Agricultural Research Institute, New Delhi for providing necessary facility and Indian Council of Agricultural Research (ICAR), New Delhi for providing financial assistance to NPIB 21-17.
LITERATURE CITED Ashmead, W. H. 1889. Descriptions of new Braconidae in the collection of the U.S. National Museum. Proceedings of the United States National Museum, 11: 611-671. Baker, C. F. 1909. Plant louse parasites I (Fam. Braconidae, subfamily Aphidiinae). Pomona College Journal of Entomology, 1: 22-25. Bernal, J. & Gonzalez, D. 1997. Reproduction of Diaeretiella rapae on Russian wheat aphid hosts at different temperatures. Entomologia Experimentalis et Applicata, 82: 159–166. Blanchard, E. E. 1940. Descripcion de un nuevo Afidiino argentine, util para la agricultura. Revista Chilena de Historia Natural, 44: 45-48. Bouche, P. F. 1834. Naturgeschichte der Insekten, besonders in Hinsicht ihrer ersten Zustande als Larven und Puppen. Berlin, 1: 1-216.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 199
Cresson, E. T. 1880. Report of the commissioner of agriculture. Report of the Entomologist. United States Department of Agriculture Annual Report, 1879: 208-260. Curtis, J. 1860. Farm Insects, being the natural history and economy of the insects injurious to the field crops of Great Britain and Ireland. John van Voorst, London. pp. 528. Dey, D. & Akhtar, M. S. 2007. Diversity of natural enemies of aphids belonging to Aphidiinae (Hymenoptera: Braconidae) in India. Journal of Asia-Pacific Entomology, 10: 281-296. Dhiman, S. C. 2007. Population dynamics of Diaeretiella rapae (M'Intosh): a parasitoid of mustard aphid, Lipaphis erysimi (Kalt.). Journal of Applied Zoological Researches, 18: 117-120. Dogra, I., Devi, N. & Desh-Raj. 2003. Parasitization of mustard aphid, Lipaphis erysimi Kalt. by Diaeretiella rapae M'Intosh in the mid-hill zone of Himachal Pradesh (India). Journal of Entomological Research, 27: 145-149. Foerster, A. 1867. Catalogue of Hymenoptera, Europe, 124-125. Hagvar, E. B. & Hofsvang, T. 1991. Aphid parasitoids (Hymenoptera, Aphidiidae): biology, host selection and use in biological control. Biological News and Information, 12: 13–41. Kurdjumov, N. V. 1913. One new aphid feeding braconid. Revue Russe d'Entom, 13: 25-26. M’Intosh, C. 1855. Book of the Gardens, W. Blackwood and Sons. Edinburgh and London. pp. 194. Marshall, T. A. 1896. Les Braconides. In: Andre E. (ed.) Species des Hymenopteres d'Europe et d'Algerie. Tome 5. pp. 635. Mukerji, S. & Chatterjee, S. N. 1950. Diaeretus aphidum sp. n., a parasite of Pterochlorus persicae Cholodkovsky on Prunus persicae in Baluchistan. Proceedings of the Royal Entomological Society of London, 19: 4-6. Pike, K. S., Stary, P., Miller, T., Allison, D., Graf, G., Boydston, L., Miller, R. & Gillespie, R. 1999. Host range and habitats of the aphid parasitoid Diaeretiella rapae (Hymenoptera: Aphididae) in Washington State. Environmental Entomology, 28: 61–71. Quilis, M. P. 1931. Especies nueves de Aphidiidae espanoles. Revista Espanola de Entomologia, 7: 25-84. Quilis, M. P. 1934. Algunos Aphidiidae de Chekoslavakia. Revista Espanola de Entomologia, 10: 5-19. Sharkey, M. J. & Wharton, R. A. 1997. Morphology and terminology. In: Wharton, RA; PM Marsh & MJ Sharkey (Ed.), Manual of the New World Genera of the Family Braconidae (Hymenoptera). Special Publication of the International Society of Hymenopterists, 1: 19–37. Stary, P. 1970. Biology of aphid parasites (Hymenoptera, Aphidiidae), With respect to integrated control The Hague W. Junk., 575 pp. Stary, P. 1988. Chapter 9. Natural Enemies. 9.1. Parasites 9.1.1 Aphidiidae. In: Minks, AK & P Harrewijn. Aphids, Their biology, Natural Enemies and Control. World crop Pests. 2B, 171-184. Elsevier, Amsterdam. Tomic, M., Tomanovic, Z.E., Kavallieratosb, N.G., Stary, P., Athanassioud, C.G., Tomic, V. & Lucic, L. 2005. Morphological variability of several biotypes of Ephedrus plagiator (Nees, 1811) (Hymenoptera: Braconidae: Aphidiinae) and description of a new species. Zoologischer Anzeiger, 244: 153-162. Viereck, H. T. 1911. Description of six new genera and thirty one new species of Ichneumon flies. Proceedings of the United States National Museum, 40: 170-196.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 200
Figures 1–9. Diaeretiella rapae. 1. Forewing, 2. Hindwing, 3. Head lateral, 4. Head frontal, 5. Propodeum, 6. Tergum I, 7. Adult female, 8. Adult male, 9. Dendogram of generalized distances between four populations of Diaeretiella rapae.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 201
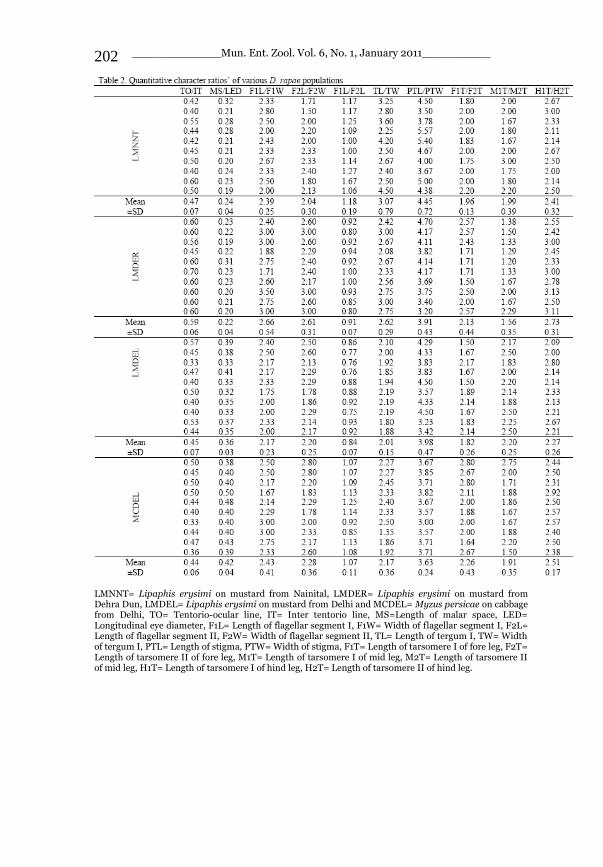
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 202
LMNNT= Lipaphis erysimi on mustard from Nainital, LMDER= Lipaphis erysimi on mustard from Dehra Dun, LMDEL= Lipaphis erysimi on mustard from Delhi and MCDEL= Myzus persicae on cabbage from Delhi, TO= Tentorio-ocular line, IT= Inter tentorio line, MS=Length of malar space, LED= Longitudinal eye diameter, F1L= Length of flagellar segment I, F1W= Width of flagellar segment I, F2L= Length of flagellar segment II, F2W= Width of flagellar segment II, TL= Length of tergum I, TW= Width of tergum I, PTL= Length of stigma, PTW= Width of stigma, F1T= Length of tarsomere I of fore leg, F2T= Length of tarsomere II of fore leg, M1T= Length of tarsomere I of mid leg, M2T= Length of tarsomere II of mid leg, H1T= Length of tarsomere I of hind leg, H2T= Length of tarsomere II of hind leg.
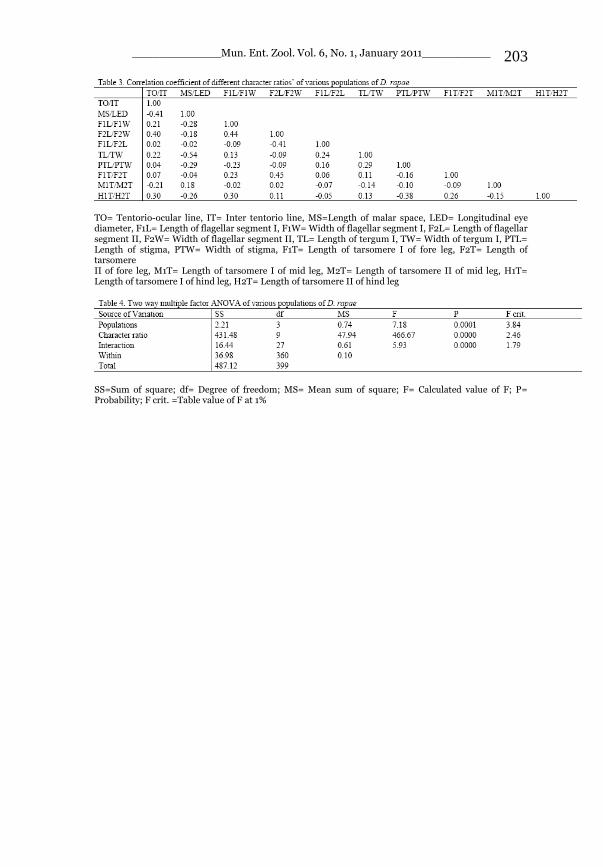
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 203
TO= Tentorio-ocular line, IT= Inter tentorio line, MS=Length of malar space, LED= Longitudinal eye diameter, F1L= Length of flagellar segment I, F1W= Width of flagellar segment I, F2L= Length of flagellar segment II, F2W= Width of flagellar segment II, TL= Length of tergum I, TW= Width of tergum I, PTL= Length of stigma, PTW= Width of stigma, F1T= Length of tarsomere I of fore leg, F2T= Length of tarsomere II of fore leg, M1T= Length of tarsomere I of mid leg, M2T= Length of tarsomere II of mid leg, H1T= Length of tarsomere I of hind leg, H2T= Length of tarsomere II of hind leg
SS=Sum of square; df= Degree of freedom; MS= Mean sum of square; F= Calculated value of F; P= Probability; F crit. =Table value of F at 1%

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 204
INVESTIGATION ON THE INSECTICIDAL EFFICACY OF NOVEL PELLET FORMULATION AGAINST
STORED PRODUCTS BEETLES
Iman Sharifian*, Mohammad-Hasan Safaralizade* and Peyman Najafi-Moghaddam**
* Department of Plant Protection, Agricultural Faculty, Urmia University, P.O. Box 57135-165, Urmia, West Azerbaijan, IRAN. E-mail: [email protected] ** Department of Chemistry, Faculty of Science, Urmia University, Urmia, IRAN, P.O. Box 57135-16. [Sharifian, I., Safaralizade, M. H. & Najafi-Moghaddam, P. 2011. Investigation on the insecticidal efficacy of novel pellet formulation against stored products beetles. Munis Entomology & Zoology, 6 (1): 204-209] ABSTRACT: Applying essential oils fumigant toxicity in stored products pests control has sharpened recently. Since these products have low penetration power and vapor pressure, their use in crop depth is restricted. The aim of this work was using controlled release technology to solve this problem. In this research, 1,8-Cineole based pellets were prepared by Dry Mixing Method and physical load of 1,8-Cineole on PVA, which followed by pressing the mixture, using pellet maker apparatus, to form pellets. Insecticidal efficacy of produced pellets was investigated against adults (1-3 days old) of Tribolium castaneum (Herbst), callosobruchus maculatus (F.) and Rhyzopertha dominica (F.) under 28±2ºC and darkness in laboratory condition. LC50 of 1 gram pellets against T. castaneum in 24 h and 48 h was 1.6 and 0.91 ml active ingredient per L air, respectively. LC50 value for R. dominica and C. maculatus in 12 h was 0.7and 1.48 ml a.i./L air, respectively. Physical load of essential oils and their constituents could be performed on biodegradable polymers and their application in stores will be possible by this manner. KEY WORDS: Controlled release, 1,8-Cineole, PVA, pellet formulation, fumigant toxicity.
Fumigation play the major role in insect pest elimination in stored products. Insect resistance to common fumigants such as phosphine etc. is a global issue now and control failures have been reported in field situations in some countries (Taylor, 1989; Bell & Wilson, 1995; Collins et al., 2002; Benhalima et al., 2004). In addition, there have been some arguments about the genotoxicity of phosphine (Garry et al., 1989).
Studies on fumigant toxicity of essential oils of plants and their constituents have been sharpened (Isman, 2006). Eucalyptus essential oil is one of the most important essential oils which has fumigant toxicity against insects. 1,8-Cineole (1,8-Cineole) is the most important compound of the genus Eucalyptus, and is largely responsible for its pesticidal properties (Batish et al., 2008).
Controlled release technology has emerged as an alternative approach with the promise to solve the problems accompanying the use of some agrochemicals, while avoiding possible side effects with others (Han et al., 2009). The overall aim of controlled release formulation consists of protecting the supply of the reagent and allowing its automatic delivery to the target at controlled rates to maintain its concentration within an optimum concentration over a long period of time. Also controlled release technology could assist in improving protection for stored grains against insect and rodent pests (Kenawy & Sherrington, 1992).
Two different approaches have been reported in the case of the physical combination of biologically active agent with polymeric materials. Firstly the biologically active agent can be encapsulated in a polymeric material and in

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 205
second approach biologically active agent is heterogeneously dispersed or dissolved in a solid polymeric matrix which can be either biodegradable or non-biodegradable. Poly (vinyl alcohol) (PVA) is a water soluble polymer based on petroleum resources with unique properties such as good transparency, lustre, antielectrostatic properties, chemical resistance and toughness (Gohil et al., 2006). It has also good gas barrier properties and good printability. Its water solubility, reactivity, and biodegradability make it a potentially useful material in biomedical, agricultural, and water treatment areas (Chiellini et al., 2003). Because it is a biodegradable polymer and cheap to make Poly(vinyl alcohol) has been used as a polymer matrix for encapsulation of the pharmacological reactive agents (Fundueanu et al., 2007). Without a doubt, its use in crop mass does‘nt has any health problem for human beings. In this research we loaded 1,8-Cineole, botanical constituent, on Poly(vinyl alcohol), biodegradable polymer, and evaluated insecticidal efficacy of produced pellet formulation, against three stored product pests.
MATERIAL AND METHODS
1,8-Cineole with 154.25 g/mol molar mass and Assay (GC, area%) ≥ 98% was purchased from Merck (Darmstadt, Germany). Poly (vinyl alcohol) (Molar Weight: 72000 g/mol; hydrolysis mole: 98 ) was purchased from Merck (Darmstadt, Germany).
All culturing insects were obtained from laboratory cultures from Urmia University, stored product pests incubation room. Rust red flour beetle (Tribolium castaneum (Herbst)), Cowpea beetle (Callosobruchus maculatus (F.)) and lesser grain borer (Rhyzopertha dominica (F.)), were reared on flour mixed with yeast (5% W/W), Cowpea and Wheat at 12-13% r.h., Respectively. All culturing media were purchased from a local market and kept in a freezer at -5ºC for 48 h and then were used as culturing medium. Experimental procedures were carried out at 28±2ºC in a dark room, and 65±5% r.h. was provided only for culturing media.
PVA and 1,8-Cineole mixture with various concentrations were prepared by Dry Mixing Method (DMM) in ice temperature on stirrer with 500 rpm for 4 h. Preparation of mixture was taken place in tight 100 ml balloons and mixing power provided with magnetic stirrer. Then prepared mixtures were transferred to FTIR pellet maker apparatus (Thermo Nicolet part No. 0016-035, USA) and pellets were produced by hydraulic pressure under 100 Kg (Fig. 1). Producing of a pellet from prepared mixture was taken about 1 min of manufacturing time. Average of ten 1 gr pellets diameter and thickness were measured by caliper that were 1.3±0.1 and 0.93±0.01 cm, respectively.
Assessment of fumigant toxicity of pellets without grain was carried out with 20 adults (1-3 days old) exposed to 250 ml glass jars sealed with screwed metal caps fitted with nylon and adhesive tape. Volume of each glass jar was measured by the amount of water it could contain. Preliminary trials in 24 h were undertaken on T. castaneum adults (1-3 days old), to determine optimum weight of pellets. Pellets were placed in containers (5 cm in length and 3 cm in diameter) in each glass jar; hence, controls had no pellet in their containers. Each container had perforated screen (40 meshes) at its end to enable penetration of volatiles emanating from pellet while insects had no contact with the pellet. Four weights (0.25, 0.5, 0.75 and 1 gram pellets) with 5 concentrations (0.5, 1, 1.5, 2 and 2.5 ml active ingredient in 5 gr PVA) were used as factors.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 206
According to preliminary trial results, one gram pellets were used as Experimental weight in original fumigant toxicity tests. Exposure time was 24 and 48 h for T. castaneum (because of its resistance against released 1,8-Cineole, and pellets limited capacity for attraction of active ingredient) and 12 h for R. dominica and C. maculatus. Insects mortality was determined immediately after treatment. Insects showing any movement by closing hot needle to their antenna were considered to be alive.
Preliminary test for determination of optimum pellets weight caused mortality
data, were altered in xarcsin then were analysed (FACTOR) by using the
MSTAT C software. The dose-effect experiments were arranged by randomized complete design and the data were analysed with analysis of variance (ANOVA) by using the MSTAT C software. The LC50 and LC95 values were calculated by probit analysis (SPSS, 16). At the end we added 5 to all SPSS obtained intercepts.
RESULTS AND DISCUSION
The fumigant activity of 1,8-Cineole has been evaluated against similar insects (Prates et al., 1998; Lee et al., 2004; Stamopoulos et al., 2007; Abdolmaleki et al., 2010). Obtained results about susceptibility of experimental insects to 1,8-Cineole, confirm prior researchers findings (Lee et al., 2002; Lee et al., 2004). In present study higher values of LC50 and LC95 were estimated for 1,8-Cineole, because a trace of 1,8-Cineole remained in polymer matrix in 24 h exposure time. We have obtained significant reduction (P<0.01) for LC50 and LC95 values on T. castaneum adults in 48 h in comparison with 24h to confirm this claim.
Preliminary test for determination of optimum pellets weight showed that insecticidal activity varied in all cases, with different pellet doses (P<0.01) (Table 1). Significant increase of mortality was observed in adults with increasing pellet weights (P<0.01). According to preliminary trials, 1 gram pellets had significantly higher effect than other weights thus we used these pellets in our main experiments (Fig. 2).
Fumigant toxicity of 1 gram pellets without grains against each of experimental species in 12 h (against T. castaneum in 24 and 48 h), is shown in table 2. Results indicates that R. dominica and T. castaneum had lowest and highest resistance against 1,8-Cineole and PVA based pellets, respectively. A significant increase of adults mortality was observed in all cases with increasing doses (P<0.01).
Fumigation containers volume were varied (e.g. 250, 500, 710 and 9000 ml) between different works (Keita et al., 2000; Lee et al., 2002; Lee et al., 2004; Stamopoulos et al., 2007; abdolmaleki et al., 2010). In this research we used 250 ml glass vials that have similarities to other fumigation vials in prior works (Lee et al., 2002; Lee et al., 2004).
Application of controlled release formulations in agriculture and study on different polymers for use as pesticide delivery agents were performed by many researchers (Hu et al., 1998; Jana et al., 2001; Riyajan & Sakdapipanich, 2009; Singh et al., 2009 a,b). Petroleum derived polymers with biodegradable, non pollutant residues in environment and non toxic characteristics, could be useful in pesticide delivery systems in agriculture (Riyajan & Sakdapipanich, 2009). However, application of natural polymers is more recommended (Hu et al., 1998; Jana et al., 2001; Singh et al., 2009a,b).
Produced formulations size were varied in different researches. Hu et al. (1998) used 8 cm diameter spheres in their work. Singh et al. (2009b) were

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 207
produced some beads (1.07-1.34 mm in size) and measured them by caliper. In present study 1 gram pellets thickness and diameter were measured by caliper and their average were 0.93±0.01 and 1.3 mm that were recorded as pellets size.
In this study, pellets were produced and expressed fumigant toxicity to T. castaneum, C. maculatus and R. dominica. According to obtained results we propose that 1,8-Cineole could be produced in pellet formulation and these pellets could have sufficient potency for insect pests control. The insecticidal efficacy varied with insect species, dose of pellets, weight of pellets and exposure time.
Physical loading of essential oils and their constituents on biodegradable polymers could be performed and their application in stores will be possible by this manner. Insecticidal efficacy of produced pellets should be evaluated against other stored products pests such as moths and mites in further studies. Also pellets could be produced by other biodegradable polymers such as starch and poly(vinyl acetate) etc for reach to cheaper to make pellets.
ACKNOWLEDGEMENTS
Authors are grateful to Urmia University for the grants that provided them to undertake this research.
LITERATURE CITED Abdolmaleki, A., Saferalizadeh, M. H. & Safavi, S. A. 2010. Mixed effects of 1,8-Cineole, botanical constituent, and reduced atmosphere pressure on stored products beetles in laboratory condition. Munis Entomology & Zoology, 5, suppl.: 1040-1047. Batish, D. B., Singh, H. P., Kohli, R. K. & Kaur, S. 2008. Eucalyptus essential oil as a natural pesticide. Forest Ecology and Management, 256: 2166-2174. Bell, C. H. & Wilson, S. M. 1995. Phosphine tolerance and resistance in Trogoderma granarium Everts (Coleoptera: Dermestidae). Journal of Stored Products Research, 31: 199-205. Benhalima, H., Chaudhry, M. Q., Mills, K. A. & Price, N. R. 2004. Phosphine resistance in stored-products insects collected from various grain storage facilities in morocco. Journal of Stored Products Research, 40: 241-249. Chiellini, E., Corti, A., D'Antone, S. & Solaro, R. 2003. Biodegradation of poly(vinyl alcohol) based materials. Progress in Polymer Science, 28: 963-1014. Collins, P. J., Daglish, G. J., Bengston, M. T., Lambkin, M. & Hervoika, P. 2002. Genetics of resistance to Phosphine in Rhyzopertha dominica (Coleoptera: Bostrichidae). Journal of Economic Entomology, 95: 862-869. Fundueanu, G., Constantin, M., Bortolotti, F., Cortesi, R., Ascenzi, P. & Menegatti, E. 2007. Cellulose acetate butyrate-pH/thermosensitive polymer microcapsules containing aminated poly(vinyl alcohol) microspheres for oral administration of DNA. European. Tournal Pharmacology and Biopharmacology, 66: 11-20. Garry, V. F., Griffith, J., Danzl, T. J., Nelson, R. L., Whorton, E. B., Krueger, L. A. & Cervenka, J. 1989. Human genotoxicity: pesticide applicators and phosphine. Science, 246: 251-255. Gohil, J. M., Bhattacharya, A. & Ray, P. 2006. Studies on the cross-linking of poly(vinyl alcohol). Journal of Polymer Research, 13: 161-169. Han, X., Chen, S. & Hu, X. 2009. Controlled-release fertilizer encapsulated by starch/poly(vinyl alcohol) coating. Desalination, 240: 21-26. Hu, X. P., Shasha, B. S., McGuire, M. R. & Prokopy, R. J. 1998. Controlled release of sugar and toxicant from a novel device for controlling pest insects. Journal of Controlled Release, 50: 257-265.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 208
Isman, B. 2006. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annual Review of Entomology, 51: 45–66. Jana, T., Roy, B. C. & Maiti, S. 2001. Biodegradable film: 6. Modification of the film for control release of insecticides. European Polymer Journal, 37 (4): 861-864. Keita, S. M., Vincent, C., Schmit, J. P., Ramaswamy, S. & Belanger, A. 2000. Effect of various essential oils on Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Journal of Stored Products Research, 36: 355-364. Kenawy, E. R., Sherrington, D. C. & Akelah, A. 1992. Controlled release of agrochemical molecules chemically bound to polymers. Journal of European Polymer, 28: 841-862. Lee, B. H., Lee, S. E., Annis, P. C., Pratt, S. J., Park, B. S. & Tumaalii, F. 2002. Fumigant toxicity of essential oils and monoterpenes against the red flour beetle, Tribolium castaneum Herbst. Journal of Asia-Pacific Entomology, 5 (2): 237-240. Lee, B. H., Annis, P. C., Tumaalii, F. & Choi, W. S. 2004. Fumigant toxicity of essential oils from the Myrtaceae family and 1,8-Cineole against 3 major stored-grain insects. Journal of Stored Products Research, 40: 55-65. Prates, H. T., Santos, J. P., Waquil, J. M., Fabris, J. D., Oliveira, A. B. & Foster, J. E. 1998. Insecticidal activity of monoterpenes against Rhyzopertha dominica (F.) and Tribolium castaneum (Herbst). Journal of Stored Products Research, 34: 243-249. Rajendran, S. & Sriranjini, V. 2008. Plant products as fumigants for stored-product insect control. Journal of Stored Products Research, 44: 126-135. Riyajan, S., & Sakdapipanich, J. T. 2009. Encapsulated neem extract containing Azadirachtin-A within hydrolyzed Poly(vinyl acetate) for controlling its release and photodegradation stability. Chemical Engineering Journal, 159: 591-597. Singh, B., Sharma, D. K., Kumar, R. & Gupta, A. 2009a. Controlled release of the fungicide thiram from starch–alginate–clay based formulation. Applied Clay Science, 45: 76-82. Singh, B., Sharma, D. K., Kumar, R. & Gupta, A. 2009b. Development of a new controlled pesticide delivery system based on neem leaf powder. Journal of Hazardeous Materials, 56: 33-38. Stamopoulos, D. C., Damos, P. & Karagianidou, G. 2007. Bioactivity of five monoterpenoid vapors to Tribolium confusum (Du Val) (Coleoptera: Tenebrionidae). Journal of Stored Products Research, 43: 571-577. Taylor, W. D. 1989. Phosphine a major fumigant at risk. International pest Control, 31: 10-14.
Table 1. The results of ANOVA belonging to dose and weight of pellets.
P F Mean
square
df Source of variations
0.00 460.3 11527.55 4 Pellets dose
Pellets weight
0.00 405.12 7609.31 3
0.00 4.08 306.48 12 Dose× weight
250.43 40 Error
59 Total **: Indicate significant difference at P< 0.01, CV= 7.07 %.
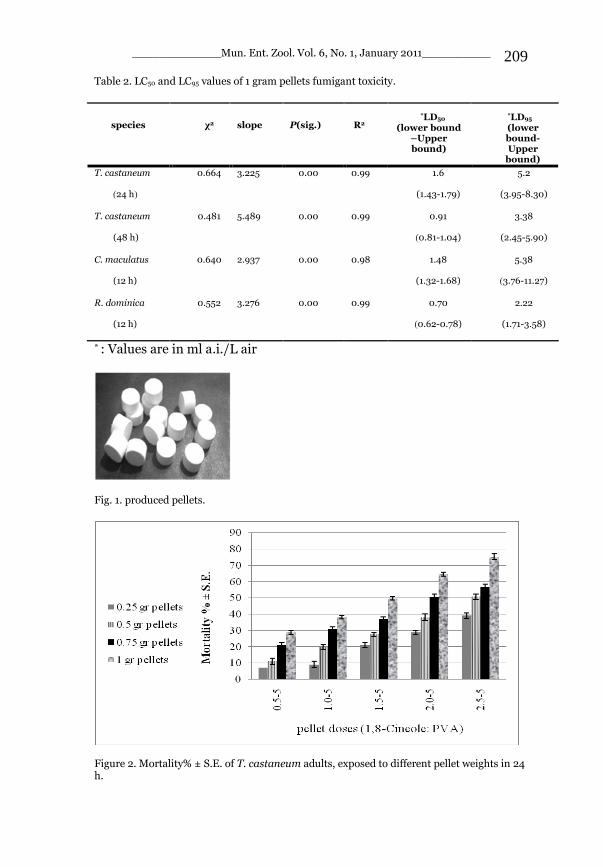
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 209
Table 2. LC50 and LC95 values of 1 gram pellets fumigant toxicity.
*LD95
(lower bound-Upper
bound)
*LD50
(lower bound –Upper bound)
R2 P(sig.) slope χ2 species
5.2
(3.95-8.30)
1.6
(1.43-1.79)
0.99 0.00 3.225 0.664
T. castaneum
24 h
3.38
(2.45-5.90)
0.91
0.81-1.04)
0.99 0.00 5.489 0.481 T. castaneum
(48 h)
5.38
3.76-11.27)
1.48
(1.32-1.68)
0.98 0.00 2.937 0.640 C. maculatus
(12 h)
2.22
(1.71-3.58)
0.70
0.62-0.78)
0.99 0.00 3.276 0.552 R. dominica
(12 h)
* : Values are in ml a.i./L air
Fig. 1. produced pellets.
Figure 2. Mortality% ± S.E. of T. castaneum adults, exposed to different pellet weights in 24 h.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 210
LONGHORNED BEETLES OF BOLU PROVINCE IN TURKEY (COLEOPTERA: CERAMBYCIDAE)
Hüseyin Özdikmen*
* Gazi Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü, 06500 Ankara / TÜRKĠYE. E-mail: [email protected] [Özdikmen, H. 2011. Longhorned beetles of Bolu province in Turkey (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 6 (1): 210-240] ABSTRACT: This work is the first attempt for entire longhorned beetles fauna of Bolu. All known taxa from Bolu province are given with some new faunistical data in the present text. 25 species are new records for Bolu province as Dinoptera collaris (Linnaeus, 1758), Anoplodera rufipes (Schaller, 1783), Stictoleptura rufa (Brullé, 1832) and Rutpela maculata (Poda, 1761) for Lepturinae; Spondylis buprestoides (Linnaeus, 1758) for Spondylidinae; Purpuricenus budensis (Götz, 1783), Obrium brunneum (Fabricius, 1792), Ropalopus clavipes (Fabricius, 1775), Plagionotus floralis (Pallas, 1773), Chlorophorus dominici Sama, 1996, C. sartor (Müller, 1766), C. trifasciatus (Fabricius, 1781) and Clytus rhamni Germar, 1817 for Cerambycinae; Morimus orientalis Reitter, 1894, Dorcadion bodemeyeri Daniel, 1900, D. praetermissum Pesarini & Sabbadini, 1998, Acanthocinus griseus (Fabricius, 1792), Oberea erythrocephala (Schrank, 1776), Oxylia duponcheli (Brullé, 1832), Phytoecia humeralis (Waltl, 1838), P. praetextata (Steven, 1817), P. pubescens Pic, 1895, Agapanthia kirbyi (Gyllenhal, 1817), A. cynarae (Germar, 1817) and A. dahli (Richter, 1821) for Lamiinae. 1 species is new record for Black Sea Region as Morimus orientalis Reitter, 1894. 9 species are new records for Western Black Sea Region as Stictoleptura rufa (Brullé, 1832), Dorcadion bodemeyeri Daniel, 1900, Acanthocinus griseus (Fabricius, 1792), Oxylia duponcheli (Brullé, 1832), Phytoecia humeralis (Waltl, 1838), P. pubescens Pic, 1895, Agapanthia kirbyi (Gyllenhal, 1817), A. cynarae (Germar, 1817) and Agapanthia dahli (Richter, 1820).Moreover, Dorcadion praetermissum mikhaili Özdikmen, 2010 described from Bolu province on the base of the specimens in the present work. KEY WORDS: Cerambycidae, Coleoptera, fauna, new records, Bolu, Turkey.
The research area of the present work covers Bolu province. Bolu is located at 39°57' North, 32°53' East coordinates. It is placed in Western Black Sea Region of NW Anatolia. It is bounded by Zonguldak province in the North, Karabük province in the North-West, Çankırı province in the East, Ankara province in the South, Bilecik and EskiĢehir provinces in the South-West, Düzce and Sakarya provinces in the West (Map 1).
The area of Bolu is 8.294 km² that is 1.06 % of the area of whole Turkey. The main morphological elements of Bolu province are two big Mountain
Range that placed N and S of Bolu plain, and plains between them and platoes which formed from planes at higher parts of Mountains. Centrum of Bolu is in Bolu plain.
The Mountains along the Black Sea divide to branches in Bolu. Valleys and plains extends among them.
Bolu has two different types of vegetation, namely ―Step vegetation‖ and ―Forest vegetation‖. The area of Bolu covers with concentrated forest vegetation. 55 % of whole area covers with forest vegetation. Karadere, Seben and Aladağ forests are rich in terms of flora and vegetation.
The data on this fauna has accumulated in a piecemeal fashion over the twentieth century and this century especially. Various authors have reported some partial data on the fauna in their different works. However, most of works were

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 211
completed in a short time and their works did not focus on fauna of Bolu generally. So the longhorned beetles fauna of Bolu has not been studied completely until now. More detailed information of most evaluated species in the text can obtain in the works of Özdikmen (2007 and 2008a,b).
In this work, some new faunistical data are presented. Besides, according to cited literatures, all known taxa from Bolu province are also given in the text. So I determined that the longhorned beetles fauna of Bolu province consists of 79 species (belong to 7 subfamily, 24 tribe, 43 genera). However, it must be suppose that the fauna is richer from determining fauna now. Since some known taxa in Turkish fauna should be presented in this region. But the taxa which can be supposed in this area are not mentioned in the present text. Consequently it would be expected that a number of additional species and new records are to be expected to occur in Bolu. Finally, this work indicates that Bolu‘s fauna is rather important for Turkey.
Map 1. Bolu province.
ARRANGEMENT OF INFORMATION
Information in the present text is given in the following order: The subfamily and the tribe names are given simply. For the genus and subgenus names, the type species are provided under the
taxon names. For each species, the whole subspecies are provided under the taxon names. The data, Material examined, Old records in Bolu, Records in
Turkey, Remarks and Chorotype under the title for each taxon is given. Material examined. Material examined that is provided for some taxons covers the original records for Bolu province in Turkey. The most materials were collected from various localities in Bolu. They are deposited in Gazi University (Ankara).
The data under the title of Material examined are given according to the following outline as possible as: Bolu(1): Seben(2), Küçükalan(3), 14.VI.2009(4), 2 specimens(5) ((1) Administrative district (Province); (2) Town; (3) Village; (4) Collecting date (day/month/year); (5) Number of specimens). Old records in Bolu. These parts include previous records that have been given by various authors in different literatures from Bolu. The whole records are evaluated with localities in related references. Each record is accompanied by the author‘s name and publication date of the related reference.
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 212
Records in Turkey. The abbreviations of the provinces and lands in Turkey are given in paranthesis. These parts include previous records that have been given by various authors in different literatures. Remarks. In these parts, taxonomical and nomenclatural problems are discussed for some taxons and are given regional and general distribution range in Turkey chiefly. Chorotype. The present zoogeographical characterization is based on the chorotype classification of Anatolian fauna, recently proposed by Vigna Taglianti et al. (1999). In the text, a possible chorotype description can be identified for each taxon. But this kind of description cannot be possible for some taxons, so two or more chorotypes are used for them.
CLASSIFICATION In this paper, classification and nomenclature of the longhorn beetles suggested by Danilevsky (2010a,b) and Löbl & Smetana (2010) are followed chiefly. Within the subfamilies all genera are listed in the same order in Danilevsky (2010b). Within the genera the species are listed alphabetically. Each name of a species or subspecies is accompanied by the author‘s name and description date.
ABREVIATIONS OF THE PROVINCES AND LANDS IN TURKEY
ADANA (AD) ELAZIĞ (EL) MANĠSA (MN) ADIYAMAN (ADY) ERZĠNCAN (ER) MARDĠN (MR) AFYON (AF) ERZURUM (EZ) MUĞLA (MG) AĞRI (AG) ESKĠġEHĠR (ES) MUġ (MU) AKSARAY (AK) GAZĠANTEP (GA) NEVġEHĠR (NE) AMASYA (AM) GĠRESUN (GI) NĠĞDE (NI) ANKARA (AN) GÜMÜġHANE (GU) ORDU (OR) ANTALYA (ANT) HAKKARĠ (HA) OSMANĠYE (OS) ARDAHAN (AR) HATAY (HT) RĠZE (RI) ARTVĠN (ART) IĞDIR (IG) SAKARYA (SA) AYDIN (AY) ISPARTA (IP) SAMSUN (SM) BALIKESĠR (BL) ĠÇEL (IC) SĠĠRT (SI) BARTIN (BR) ĠSTANBUL (IS) SĠNOP (SN) BATMAN (BA) ĠZMĠR (IZ) SĠVAS (SV) BAYBURT (BY) KAHRAMANMARAġ (KA) ġANLIURFA (SU) BĠLECĠK (BI) KARABÜK (KR) ġIRNAK (SK) BĠNGÖL (BN) KARAMAN (KM) TEKĠRDAĞ (TE) BĠTLĠS (BT) KARS (KAR) TOKAT (TO) BOLU (BO) KASTAMONU (KS) TRABZON (TB) BURDUR (BU) KAYSERĠ (KY) TUNCELĠ (TU) BURSA (BS) KIRIKKALE (KI) UġAK (US) ÇANAKKALE (CA) KIRKLARELĠ (KK) VAN (VA) ÇANKIRI (CN) KIRġEHĠR (KIR) YALOVA (YA) ÇORUM (CO) KĠLĠS (KL) YOZGAT (YO) DENĠZLĠ (DE) KOCAELĠ (KO) ZONGULDAK (ZO) DĠYARBAKIR (DI) KONYA (KN) THRACIA (=EUROPEAN
TUR.) (TRA) DÜZCE (DU) KÜTAHYA (KU) TURKEY (TUR) EDĠRNE (ED) MALATYA (MA)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 213
Family CERAMBYCIDAE Subfamily PRIONINAE Tribe ERGATINI Ergates Serville, 1832 [Type sp.: Prionus serrarius Panzer, 1793 = Cerambyx faber Linnaeus, 1767] Ergates faber (Linnaeus, 1761) Ergates faber faber (Linnaeus, 1761) Material examined: Bolu: Çepni village, 25.VII.2010, 1 specimen. Old records in Bolu prov.: Abant (Sakarca plateau) (Defne, 1954); Abant (Alkan, 2000). Records in Turkey: (AN-ANT-ART-BO-BS-DU-KA-KO-KS-SN-TB-TRA-TUR) Remarks: The species distributes mostly in N Turkey. It is represented by the nominative subspecies in Turkey. Chorotype: Turano-Europeo-Mediterranean. Tribe PRIONINI Prionus Geoffroy, 1762 [Type sp.: Cerambyx coriarius Linnaeus, 1758] Prionus coriarius (Linnaeus, 1758) Material examined: Bolu: Gölcük, 27.VII.2010, 1 specimen. Old records in Bolu prov.: Abant (Özdikmen & Demir, 2006). Records in Turkey: (AN-ANT-ART-AY-BL-BO-BU-HT-KA-KI-KK-KN-KO-KS-OS-RI-SN-TB-TRA-TUR) Remarks: According to distribution in Turkey of host plants, probably the species distributes rather widely in Turkey. Chorotype: Sibero-European + Turano-Europeo-Mediterranean. Subfamily LEPTURINAE Tribe XYLOSTEINI Xylosteus Plavilstshikov, 1936 [Type sp.: Xylosteus spinolae Frivaldszky, 1838] Xylosteus kadleci Miroshnikov, 2000 Material examined: Bolu: Abant env., 19.VI.2009, 1 specimen. Old records in Bolu prov.: Abant Lake (Miroshnikov, 2000). Records in Turkey: (BO) Remarks: The endemic species has been known only from the type locality until now. Chorotype: Anatolian. Tribe RHAGIINI Rhagium Fabricius, 1775 [Type sp.: Cerambyx inquisitor Linnaeus, 1758] Subgenus Hagrium Villiers, 1978 [Type sp.: Rhagium bifasciatum Fabricius, 1775]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 214
Rhagium bifasciatum Fabricius, 1775 Old records in Bolu prov.: Abant (Küçükalan and Küçüksu places and Öküzovası forest env.) (Defne, 1954); Aladağ road (Öymen, 1987); Aladağ (Çanakçıoğlu, 1993; Çanakçıoğlu & Mol, 1998). Records in Turkey: (AM-ANT-ART-BO-BU-DU-EZ-GI-GU-IP-IS-KR-KAR-KS-KK-KO-NI-OR-RI-SA-SN-TB-ZO-TRA-TUR) Remarks: The species distributes mostly in North Turkey. Chorotype: European or Turano-Europeo-Mediterranean. According to Sama (2002), the record of Plavilstshikov (1936) from North Africa is erroneous. Subgenus Megarhagium Reitter, 1912 [Type sp.: Cerambyx sycophanta Schrank, 1781] Rhagium fasciculatum Faldermann, 1837 Old records in Bolu prov.: Abant (Küçüksu place) (Defne, 1954). Records in Turkey: (ART-BO-GI-RI-SN-TB-TUR) Remarks: The species distributes only in North Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian). Subgenus Rhagium Fabricius, 1775 [Type sp.: Cerambyx inquisitor Linnaeus, 1758] Rhagium inquisitor (Linnaeus, 1758) Rhagium inquisitor inquisitor (Linnaeus, 1758) Material examined: Bolu: Abant, 19.VI.2009, 2 specimens; Yedigöller- Bolu road, Bolu Mts., 14.VI.2010, Pheromone trap, 2 specimens; Gürdek, Yedigöller, 14.VI.2010, Pheromone trap, 1 specimen; Akçaalan district, Abant road, 12.VII.2010, 2 specimens. Old records in Bolu prov.: Abant (Alkan, 1946); Abant (Karakuz forest, Demircikırığı) / Abant (Küçükalan ve Küçüksu forests), Gerede / Mudurnu (Defne, 1954); Central / Gerede / Mudurnu (Çanakçıoğlu, 1983). Records in Turkey: (AM-AN-ANT-ART-BO-BS-BU-DU-EZ-GI-GU-IS- KAR- KR-KS-OR-RI-SA-SN-TB-TRA-TUR) Remarks: The species has four distinct subspecies in the World. In Turkey, it is represented by two subspecies. R. inquisitor stshukini Semenov, 1898 occurs only in NE Turkey, and the nominative R. inquisitor inquisitor (Linnaeus, 1758) occurs in other parts of Turkey. Chorotype: Holarctic. Dinoptera Mulsant, 1863 [Type sp.: Leptura collaris Linnaeus, 1758] Dinoptera collaris (Linnaeus, 1758) Material examined: Bolu: Abant, 19.VI.2009, 1 specimen; Mudurnu-Göynük road, Karapınar return env., 15 km to Mudurnu, 01.V.2010, 2 specimens; Yeniçağa-Mengen road, 14.VI.2010, 2 specimens; Mengen, Mengen-Yedigöller road, 14.VI.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AM-AN-ANT-ART-BO-BS-CN-EZ-IC-IP-IS-KN-KO-KR-KS-OS-RI-SM-TRA-TUR) Remarks: New to Bolu province. The species distributes rather widely in Turkey. Chorotype: Sibero-European.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 215
Cortodera Mulsant, 1863 [Type sp.: Grammoptera spinosula Mulsant, 1839 = Leptura humeralis Schaller, 1783] Cortodera flavimana (Waltl, 1838) Old records in Bolu prov.: Abant (Sama, 1982; Tauzin, 2000; Özdikmen & Demir, 2006); Yeniçağa Lake – Deliler village, between Yeniçağa–Gerede (exit of Yeniçağa), between Gerede–Kızılcahamam, Mengen (Devrek–Mengen, entry of Mengen)(Özdikmen, 2007). Records in Turkey: (AD-AF-AK-AN-ANT-ART-BO-BS-BY-CN-EZ-GU-IC-IP-IS-IZ- NI-KA-KAR-KN-KO-KR-KS-KY-RI-SM-SN-SV-TO-YO-TRA-TUR) Remarks: The species distributes widely in Turkey due to the host plant, Ranunculus, is a cosmopolite genus of plants. Chorotype: Turano-Mediterranean (Balkano-Anatolian). Cortodera humeralis (Schaller, 1783) Cortodera humeralis humeralis (Schaller, 1783) Old records in Bolu prov.: Ankara-Bolu provinces: Kızılcahamam (Köroğlu Mountains) (Adlbauer, 1992). Records in Turkey: (AN-ANT-ART-BO-OS-TRA) Remarks: The species distributes in N Turkey. Chorotype: S-European. Fallacia Mulsant & Rey, 1863 [Type sp.: Fallacia longicollis Mulsant & Rey, 1863] Fallacia elegans (Faldermann, 1837) Old records in Bolu prov.: Bolu prov.: Abant (Sama, 1996). Records in Turkey: (ART-BO-RI-SN-TB-TUR) Remarks: The species distributes only in North Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian). Tribe LEPTURINI Vadonia Mulsant, 1863 [Type sp.: Leptura unipunctata Fabricius, 1787] Vadonia unipunctata (Fabricius, 1787) Vadonia unipunctata unipunctata (Fabricius, 1787) Material examined: Bolu: between Dereceören-Akçaalan, Abant, 19.VI.2009, 10 specimens; Gerede-Mengen, 06.VII.2009, 2 specimens; between Gerede-Mengen, 06.VII.2009, 2 specimens; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 1 specimen; entry of Göynük-Mudurnu road, Atyaylası pass, exit of Sarılar village, 08.VII.2009, 1 specimen; Yeniçağa-Mengen road, 14.VI.2010, 2 specimens; Mengen, Mengen-Yedigöller road, 14.VI.2010, 1 specimen. Old records in Bolu prov.: Devrek–Mengen road, Mengen (Devrek–Mengen), Yeniçağa (Özdikmen, 2007). Records in Turkey: (AF-AK-AM-AN-ANT-ART-BI-BN-BO-BT-BU-BY-CN-EL-EZ-IP-IZ-KA-KAR-KIR-KN-KO-KR-KS-KY-MA-NE-NI-OS-RI-SV-TO-US-TRA-TUR) Remarks: The species distributes widely in Turkey. It is represented by the nominative subspecies in Turkey. Chorotype: Turano-European or Turano-Europeo-Mediterranean. According to Sama (2002), the records from North Africa are erroneous.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 216
Pseudovadonia Lobanov, Danilevsky & Murzin, 1981 [Type sp.: Leptura livida Fabricius, 1777] Pseudovadonia livida (Fabricius, 1777) Pseudovadonia livida livida (Fabricius, 1777) Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 1 specimen; Karacasu env., 18.VI.2009, Abies normandiana forest, 10 specimens; Abant, Dereceören env., 19.VI.2009, 3 specimens; Gerede, 06.VII.2009, 17 specimens; between Gerede-Mengen, 06.VII.2009, 2 specimens; Mudurnu-Abant road, 3 km to Abant, 40 km to Bolu, 08.VII.2009, 1 specimen; Aynalı village pass, 08.VII.2009, 15 specimens; entry of Göynük-Mudurnu road, Atyaylası pass, exit of Sarılar village, 08.VII.2009, 2 specimens; Abant-Dereceören env., 08.VII.2009, 2 specimens; Bolu-Seben road, 08.VII.2009, 2 specimens; Kıbrıscık-Seben road, 10 km to Seben, 09.VII.2009, 1 specimen; Abant, 30.IV.2010, 1 specimen; Yeniçağa, 14.VI.2010, 1 specimen; Gökçesu-Mengen road, 5 km to Mengen, 14.VI.2010, 1 specimen; Mengen, Mengen-Yedigöller road, 14.VI.2010, 9 specimens; 10 km to Yedigöller, 14.VI.2010, 12 specimens; Atyaylası road, 17.VI.2010, 5 specimens; 40 km to Mudurnu, Abant road, 12.VII.2010, 2 specimens; Gölcük road, 14.VII.2010, 1 specimen; Yeniçağa, 15.VII.2010, 1 specimen; Akçaalan-Abant road, 15.VI.2010, 4 specimens; 8 km to Abant, Abant road, 15.VI.2010, 1 specimen; Abant env., 2010, 2 specimens. Old records in Bolu prov.: Mengen (Devrek–Mengen) (Özdikmen, 2007). Records in Turkey: (ADY-AM-AN-ANT-ART-BI-BO-BR-BS-BT-BY-CN-ER-EZ-GA-GI-GU-HT-IC-IP-IS-IZ-KAR-KK-KN-KO-KR-KS-MN-NI-OS-RI-SM-US-TRA-TUR) Remarks: The species distributes widely in Turkey. It is represented by two subspecies in Turkey. P. livida desbrochersi (Pic, 1891) occurs in E or NE Turkey, and the nominative P. livida livida occurs in other parts of Turkey. Chorotype: Sibero-European + E-Mediterranean (Palaestino-Taurian). Anoplodera Mulsant, 1839 [Type sp.: Leptura sexguttata Fabricius, 1775] Anoplodera rufipes (Schaller, 1783) Anoplodera rufipes rufipes (Schaller, 1783) Material examined: Bolu: 8 km to Abant, Abant road, 15.VI.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AN-BN-BO-BU-EZ-GU-IC-KS-OR-RI-TB-TUR) Remarks: New to Bolu province. The species distributes rather widely in Turkey. The species is represented by two subspecies in Turkey. A. rufipes lucidipes Sama, 1999 occurs only in S Turkey and the nominative A. rufipes rufipes occurs mostly in N Turkey. Chorotype: Sibero-European. Stictoleptura Casey, 1924 [Type sp.: Leptura cribripennis LeConte, 1859] Subgenus Aredolpona Nakane & Ohbayashi, 1957 [Type sp.: Leptura rubra Linnaeus, 1758] Stictoleptura rubra (Linnaeus, 1758) Stictoleptura rubra rubra (Linnaeus, 1758) Old records in Bolu prov.: Abant (Tavak plateau / Öküzovası / Sakarca plateau env. / Abant forests) (Defne, 1954); Bolu prov. (Lodos, 1998). Records in Turkey: (ANT-ART-BO-BU-DU-IS-KO-SA-SN-TUR) Remarks: It distributes rather widely in Turkey. The species is represented by the nominative subspecies in Turkey. Chorotype: Palaearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 217
Stictoleptura fulva (DeGeer, 1775) Material examined: Bolu: Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 1 specimen; Hacıayaz pass, 1080 m, 07.VII.2009, 2 specimens; Bolu-Seben road, 09.VII.2009, 1 specimen; Akçaalan district, Abant road, 12.VII.2010, 3 specimens; Advan village, 18.VII.2010, 21 specimens. Old records in Bolu prov.: Akçakoca (Konuralp) (Sama, 1982). Records in Turkey: (AD-ANT-ART-BO-BR-BU-HT-IC-IP-IS-IZ-KA-KK-KN-KO-KR-KS-MN-OS-ZO-TRA-TUR) Remarks: It distributes rather widely in Turkey. Chorotype: European. Stictoleptura rufa (Brullé, 1832) Stictoleptura rufa dimidiata (Daniel & Daniel, 1891) Material examined: Bolu: Mudurnu-Göynük road, Karapınar return, 15.VI.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (ANT-BI-BO-BY-ER-EZ-GU-IC-TU-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. Probably the species distributes rather widely in Turkey. Danilevsky (2010a) stated “According to J. Vorisek (personal communication, 1992), S. rufa is represented in Caucasus and Turkey by S. r. dimidiata (Daniel, 1891). But according to the original description, "dimidiata" is characterized by black elytral half (or 2/3); such form is unknown in Caucasus. The specimens, similar to Caucasian variations, were identified in Paris Museum as var. attaliensis Dan.”. In this case, the species is probably represented by three subspecies in Turkey. The nominative subspecies S. rufa rufa (Brullé, 1832) occuring in European Turkey and North-West Anatolia near European Turkey, S. rufa attaliensis (Daniel, 1891) occurring only in North-Eastern Turkey and S. rufa dimidiata (Daniel, 1891) occurring in the other parts of Turkey. Chorotype: Turano-Mediterranean (Turano-Apenninian). Anastrangalia Casey, 1924 [Type sp.: Leptura sanguinea LeConte, 1859] Anastrangalia dubia (Scopoli, 1763) Anastrangalia dubia dubia (Scopoli, 1763) Material examined: Bolu: Karacasu env., 18.VI.2009, Abies normandiana forest, 5 specimens; Abant, 19.VI.2009, 6 specimens; Abant, Dereceören env., 19.VI.2009, 1 specimen; between Dereceören-Akçaalan, Abant, 19.VI.2009, 2 specimens; between Gerede-Mengen, 06.VII.2009, 5 specimens; Gökçesu-Mengen road, 5 km to Mengen, 14.VI.2010, 1 specimen; Mudurnu-Göynük road, Karapınar village env., 07.VII.2009, 2 specimens; from Ġkizler plateau road to Abant road, 07.VII.2009, 4 specimens; Meyitler pass, 1130 m, 08.VII.2009, 2 specimens; Abant- Dereceören env., 08.VII.2009, 2 specimens; Mengen, Mengen-Yedigöller road, 14.VI.2010, 1 specimen; Mengen-Yedigöller, 14.VI.2010, 2 specimens; Akçaalan-Abant road, 15.VI.2010, 3 specimens; 8 km to Abant, Abant road, 15.VI.2010, 1 specimen; Atyaylası, 17.VI.2010, 2 specimens; 500 m to Dereceören, 3 specimens; Saba road, 2010, 1 specimen. Old records in Bolu prov.: Abant-Sakarca plateau env. (Sinekli plateau) as Leptura dubia ab. chamomillae F. and Leptura dubia ab. limbata Laich. (Defne, 1954). Records in Turkey: (AF-ART-BI-BO-BR-CN-CO-GI-GU-KAR-KIR-KN-KR-KS-MG-NE-OS-RI-SN-TB-TO-ZO-YO-TUR) Remarks: It distributes mostly in North Turkey but probably distributes rather widely in Turkey. The species is represented by two subspecies in Turkey. A. dubia melanota (Faldermann, 1837) occurs only in NE Turkey and the nominative A. dubia dubia occurs in other parts of Turkey. Chorotype: Turano-Europeo-Mediterranean.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 218
Anastrangalia sanguinolenta (Linnaeus, 1760) Material examined: Bolu: Meyitler pass, 1130 m, 08.VII.2009, 1 specimen; Mengen-Yedigöller, 14.VI.2010, 2 specimens; Akçaalan-Abant road, 15.VI.2010, 1 specimen. Old records in Bolu prov.: Mengen (Çorak, Kadirler) / Bolu Mountain (Özdikmen et al., 2005). Records in Turkey: (AM-AN-ART-BO-BS-EZ-GI-GU-KAR-KN-KR-KS-KY-SM-SN-TB-TO-YO-TUR) Remarks: The species distributes in N Turkey. Chorotype: European or Sibero-European. According to Sama (2002) records from Siberia not confirmed by Cherepanov (1990). Pachytodes Pic, 1891 [Type sp.: Leptura cerambyciformes Schrank, 1781] Pachytodes erraticus (Dalman, 1817) Pachytodes erraticus erraticus (Dalman, 1817) Material examined: Bolu: Karacasu env., 18.VI.2009, Abies normandiana ormanı, 27 specimens; Gerede-Mengen arası, 06.VII.2009, Çam ormanı açıklığı, 3 specimens; Hacıayaz pass, 1080 m, 07.VII.2009, 3 specimens; Aynalı köy pass, 08.VII.2009, 5 specimens; Abant-Dereceören env., 08.VII.2009, 23 specimens; Bolu-Seben road, 08.VII.2009, 5 specimens; Mengen-Yedigöller, 14.VI.2010, 2 specimens; Yeniçağa- Mengen road, 14.VI.2010, 26 specimens; Yedigöller‘e 10 km kala, 14.VI.2010, 1 specimen; Akçaalan-Abant road, 15.VI.2010, 1 specimen; Abant‘a 8 km kala, Abant road, 15.VI.2010, 1 specimen; Mudurnu-Göynük road, Karapınar kavĢağı 6 km sapağı, 15.VI.2010, 3 specimens; Seben env., 16.VI.2010, 1 specimen; Mudurnu 40 km kala, Abant road, 12.VII.2010, 1 specimen; Gölcük road, 14.VII.2010, 3 specimens; Dereceören‘e 500 m kala, 3 specimens. Old records in Bolu prov.: Mengen (Çorak, Kadirler) / Bolu Mountain (Özdikmen et al., 2005). Records in Turkey: (AF-AM-AN-ANT-ART-BI-BO-BR-BS-BT-CN-CO-EL-ER-EZ-GA-GU-HAT-IP-IS-IZ-KAR-KK-KO-KN-KR-KS-MN-MU-RI-SM-SN-SV-TB-TO-TU-YO-ZO-TRA-TUR) Remarks: The species distributes widely in Turkey. The species has two subspecies. It is represented by the nominotypical subspecies in Turkey. Chorotype: Sibero-European. Solaia Sama, 2003 [Type sp.: Solaia antonellae Sama, 2003] Solaia antonellae Sama, 2003 Old records in Bolu prov.: Abant (type loc.) (Sama, 2003). Records in Turkey: (BO) Remarks: The endemic species has been known only from the type locality until now. Chorotype: Anatolian. Rutpela Nakane et Ohbayashi, 1957 [Type sp.: Leptura maculata Poda, 1761] Rutpela maculata (Poda, 1761) Rutpela maculata maculata (Poda, 1761) Material examined: Bolu: Gerede-Mengen, 06.VII.2009, Uzun papatya çiçekleri üzerinden, 4 specimens; Ġkizler yaylası roadndan Abant roadna doğru, 07.VII.2009, 1 specimen; Bolu-Seben road, 08.VII.2009, 1 specimen; Yeniçağa- Mengen road, 14.VI.2010, 2 specimens; Abant, Dereceören env., 19.VI.2009, 1 specimen; Gölcük road, 14.VII.2010, 2 specimens; Dereceören‘e 500 m kala, 2 specimens. Old records in Bolu prov.: Absent.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 219
Records in Turkey: (AD-AM-ANT-ART-BL-BN-BO-BS-CA-CN-HT-IC-IS-KK-KN-KO-KR-KS-MA-MU-OS-RI-SM-SN-SV-TB-TO-TU-VA-TUR) Remarks: New for Bolu province. It distributes widely in Turkey. The species is represented by two subspecies in Turkey. These are; R. maculata irmasanica Sama, 1996 occurs only in South Turkey and the nominative R. maculata maculata occurs in other parts of Turkey. Chorotype: European. Stenurella Villiers, 1974 [Type sp.: Leptura melanura Linnaeus, 1758] Stenurella bifasciata (Müller, 1776) Stenurella bifasciata bifasciata (Müller, 1776) Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 1 specimen; Karacasu env., 18.VI.2009, Abies normandiana forest, 1 specimen; between Gerede-Mengen, 06.VII.2009, 3 specimens; Mudurnu-Göynük road, Karapınar village env., 07.VII.2009, 4 specimens; from Ġkizler plateau road to Abant road, 07.VII.2009, 2 specimens; Göynük road, 30 km to Göynük, SamsaçavuĢ village env., 07.VII.2009, 2 specimens; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 1 specimen; Hacıayaz pass, 1080 m, 07.VII.2009, 1 specimen; Mudurnu-Göynük road, Karapınar village env., 07.VII.2009, 2 specimens; entry of Göynük-Mudurnu road, Atyaylası pass, exit of Sarılar village, 08.VII.2009, 1 specimen; Aynalı village pass, 08.VII.2009, 8 specimens; Meyitler pass, 1130 m, 08.VII.2009, 6 specimens; Mengen, Mengen-Yedigöller road, 14.VI.2010, 1 specimen; Mengen-Yedigöller, 14.VI.2010,2 specimens; 10 km to Yedigöller, 14.VI.2010, 1 specimen; Akçaalan-Abant road, 15.VI.2010, 1 specimen; Atyaylası road, 17.VI.2010 2 specimens; Mudurnu-Göynük road, Karapınar return, 15.VI.2010, 2 specimens; Göynük-Sünnet Lake, 15.VI.2010, 2 specimens; 40 km to Mudurnu, Abant road, 12.VII.2010, 1 specimen. Old records in Bolu prov.: from Konuralp to Akçakoca / Abant (Sama, 1982); Central / Mengen (Özdikmen et al., 2005); between Gerede–Kızılcahamam, Mengen (Devrek–Mengen) (Özdikmen, 2007). Records in Turkey: (AD-AF-AK-AM-AN-ANT-ART-BI-BN-BO-BR-BS-BT-BU-CA-CN-CO-ER-EZ-GA-GU-HT-IC-IZ-KA-KK-KN-KO-KR-KS-KY-MG-MN-NE-OS-RI-SM-TB-US-YA-YO-ZO-TUR) Remarks: The species distributes widely in Turkey. It is represented by three subspecies in Turkey. S. bifasciata nigrosuturalis (Reitter, 1895) occurs in SE Turkey and Lebanon and Syria, S. bifasciata limbiventris (Reitter, 1898) occurs only in NE Turkey and the nominative S. bifasciata bifasciata (Müller, 1776) occurs in other parts of Turkey. Chorotype: Sibero-European + SW-Asiatic. Stenurella melanura (Linnaeus, 1758) Old records in Bolu prov.: from Konuralp to Akçakoca (Sama, 1982). Records in Turkey: (ANT-ART-BO-BS-CA-IS-IZ-KO-KS-OR-TUR) Remarks: It distributes in North, West and South-West Turkey. Chorotype: Sibero-European. Stenurella nigra (Linnaeus, 1758) Material examined: Bolu: Abant, Dereceören env., 19.VI.2009, 1 specimen; Gerede-Mengen, 06.VII.2009, 1 specimen; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 2 specimens; Yeniçağa-Mengen road, 14.VI.2010, 10 specimens; Mengen, Mengen-Yedigöller road, 14.VI.2010, 5 specimens; Mengen-Yedigöller, 14.VI.2010, 5 specimens; 10 km to Yedigöller, 14.VI.2010, 2 specimens; Mudurnu-Göynük road, Karapınar return, 15.VI.2010, 1 specimen; Atyaylası road, 17.VI.2010, 32 specimens. Old records in Bolu prov.: Mengen (Özdikmen, 2007). Records in Turkey: (ART-BO-IS-KO-KR-KS-RI-SM-YA-ZO-TUR) Remarks: It distributes only in North Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 220
Chorotype: European. Stenurella septempunctata (Fabricius, 1792) Stenurella septempunctata suturata (Reiche & Saulcy, 1858) Material examined: Bolu: Abant, Dereceören env., 19.VI.2009, 3 specimens; between Gerede-Mengen, 06.VII.2009, 3 specimens; Gerede-Mengen, 06.VII.2009, 22 specimens; from Ġkizler plateau road to Abant road, 07.VII.2009, 82 specimens; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 4 specimens; Aynalı village pass, 08.VII.2009, 1 specimen; Abant-Dereceören env., 08.VII.2009, 3 specimens; Bolu-Seben road, 08.VII.2009, 6 specimens; Meyitler pass, 1130 m, 08.VII.2009, 2 specimens; Yeniçağa-Mengen road, 14.VI.2010, 22 specimens; Gökçesu-Mengen road, 5 km to Mengen, 14.VI.2010, 1 specimen; Mengen, Mengen-Yedigöller road, 14.VI.2010, 8 specimens; Mengen-Yedigöller, 14.VI.2010, 10 specimens; 10 km to Yedigöller, 14.VI.2010, 2 specimens; Akçaalan-Abant road, 15.VI.2010, 5 specimens; 8 km to Abant, Abant road, 15.VI.2010, 3 specimens; Mudurnu-Göynük road, Karapınar return, 15.VI.2010, 2 specimens; Göynük-Sünnet Lake, 15.VI.2010, 1 specimen; Göynük road, 15.VI.2010, 2 specimens; Çubuk Lake, Göynük, 16.VI.2010, 2 specimens; Atyaylası road, 17.VI.2010, 13 specimens; Akçaalan district, Abant road, 12.VII.2010, 2 specimens; Gölcük road, 14.VII.2010, 16 specimens; Yeniçağa, 15.VII.2010, 2 specimens; 500 m to Dereceören, 2 specimens; Abant env., 2010, 1 specimen. Old records in Bolu prov.: from Konuralp to Akçakoca (Sama, 1982); Devrek–Mengen road, Mengen, Afyon: Kavaklıdere, Karakaya tepesi (Özdikmen, 2007). Records in Turkey: (AF-AM-AN-ART-BI-BO-BS-CA-EZ-GU-IS-IZ-KK-KO-KR-KS-RI-SM-TO-TB-YA-YO-ZO-TUR) Remarks: The species distributes mostly in N Turkey and Northern Central Turkey. There are two distinct subspecies in the World. These are; the nominative subspecies and S. septempunctata suturata occurs in Caucasus and Turkey. Chorotype: Turano-European. Subfamily NECYDALINAE Tribe NECYDALINI Necydalis Linnaeus, 1758 [Type sp.: Necydalis major Linnaeus, 1758] Subgenus Necydalis Linnaeus, 1758 [Type sp.: Necydalis major Linnaeus, 1758]
Necydalis sabatinelli Sama, 1994 Old records in Bolu prov.: Abant (Sama, 1994; Tauzin, 2000). Records in Turkey: (BO) Remarks: The endemic species has been known only from the type locality until now. Chorotype: Anatolian. Subfamily ASEMINAE Tribe SAPHANINI Drymochares Mulsant, 1847 [Type sp.: Drymochares truquii Mulsant, 1847] Drymochares starcki Ganglbauer, 1888 Drymochares starcki ivani Sama & Rapuzzi, 1993 Old records in Bolu prov.: Abant (type locality) as Drymochares starcki ivani Sama & Rapuzzi, 1993; Bolu prov. (Tauzin, 2000).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 221
Records in Turkey: (ART-BO-GI-GU-KS-OR-SM-SN-TO-ZO-TUR) Distribution: Caucasus, Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian). Tribe ASEMINI Arhopalus Serville, 1834 [Type sp.: Cerambyx rusticus Linnaeus, 1758] Arhopalus ferus (Mulsant, 1839) Old records in Bolu prov.: Abant (Alkan, 1946). Records in Turkey: (AD-AM-AN-ANT-AY-BI-BO-CA-ES-HT-IZ-KK-KU-MG-TO-TUR) Remarks: The species probably distributes rather widely in Turkey. Chorotype: Palaearctic. Arhopalus rusticus (Linnaeus, 1758) Arhopalus rusticus rusticus (Linnaeus, 1758) Old records in Bolu prov.: Bolu Mountains (Mengen) (Demelt, 1967); Bolu prov. (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983). Records in Turkey: (AN-ANT-ART-BL-BO-BU-BY-DE-GU-IS-KAR-KR-KS-KU-MG-OR-RI-SM-SN-TB-TO-TUR) Remarks: The species distributes rather widely in Turkey. It is represented by the nominotypical subspecies in Palaearctic Region (incl. Turkey). Known other subspecies are distributed in Nearctic Region. These are; A. rusticus montanus (LeConte, 1873) occurs in United States, Mexico, A. rusticus nubilus (LeConte, 1850) occurs in United States, Mexico, Jamaica, Bahamas, A. rusticus obsoletus (Randall, 1838) occurs in United States, Guatemala, Honduras, Canada, Mexico and A. rusticus hesperus Chemsak & Linsley, 1965 occurs in United States. Chorotype: Holarctic. Tetropium Kirby, 1837 [Type sp.: Tetropium cinnamopterum Kirby, 1837] Tetropium castaneum (Linnaeus, 1758) Old records in Bolu prov.: Seben (Öymen, 1987); Bolu prov. (Lodos, 1998). Records in Turkey: (ART-BO-DU-ZO-TUR) Remarks: The species distributes only in North Turkey. Chorotype: Sibero-European. Subfamily SPONDYLIDINAE Tribe SPONDYLIDINI Spondylis Fabricius, 1775 [Type sp.: Attelabus buprestoides Linnaeus, 1758] Spondylis buprestoides (Linnaeus, 1758) Material examined: Bolu: Göynük road, 7 km to Göynük, 15.VI.2010, on Pinus, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AN-ART-BS-IS-KAR-KR-SN-TB-TUR) Remarks: New to Bolu province. The species distributes mostly in N Turkey. Chorotype: Sibero-European or Sibero-European + N-Africa. Because, according to Sama (2002), records from North Africa (Morocco) need confirmation.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 222
Subfamily CERAMBYCINAE Tribe TRACHYDERINI Purpuricenus Dejean, 1821 [Type sp.: Cerambyx kaehleri Linnaeus, 1758] Subgenus Purpuricenus Dejean, 1821 [Type sp.: Cerambyx kaehleri Linnaeus, 1758] Purpuricenus budensis (Götz, 1783) Material examined: Bolu: Mengen-Yedigöller, 14.VI.2010, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ADY-AF-AM-AN-ANT-ART-AY-BL-BN-BO-BS-BU-CA-CO-DE-ED-EZ-GA-GU-HT-IC-IP-IS-IZ-KA-KI-KN-KO-MG-MN-MU-NI-OS-RI-SI-SM-SN-TO-TU-YO-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. Chorotype: Turano-Europeo-Mediterranean. Tribe OBRIINI Obrium Dejean, 1821 [Type sp.: Cerambyx cantharinus Linnaeus, 1767] Obrium brunneum (Fabricius, 1792) Material examined: Bolu: Atyaylası, 17.VI.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (ART-BI-BO-GI-GU-KS-TB-TUR) Remarks: New to Bolu province. It distributes only in North Turkey. Chorotype: European. Tribe STENOPTERINI Stenopterus Illiger, 1804 [Type sp.: Necydalis rufa Linnaeus, 1767] Stenopterus rufus (Linnaeus, 1767) Stenopterus rufus geniculatus Kraatz, 1863 Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 1 specimen; Göynük road, 30 km to Göynük, SamsaçavuĢ village env., 07.VII.2009, 1 specimen; Bolu-Seben road, 09.VII.2009, 7 specimens; Yeniçağa-Mengen road, 14.VI.2010, 7 specimens; Gökçesu-Mengen road, 5 km to Mengen, 14.VI.2010, 2 specimens; Göynük-Sünnet Lake, 15.VI.2010, 2 specimens; Göynük road, 15.VI.2010, 1 specimen. Old records in Bolu prov.: Mengen (Devrek–Mengen) (Özdikmen, 2007). Records in Turkey: (ADY-AM-AN-ANT-ART-BI-BO-BR-BS-CA-CN-CO-EZ-GA-GU-HT-IC-IS-IZ-KA-KK-KN-KO-KR-KS-KY-MN-NI-OS-RI-SM-SN-TB-TO-TU-YA-YO-TRA-TUR) Remarks: The species distributes widely in Turkey. The species is represented by three subspecies in Turkey. S. rufus geniculatus Kraatz, 1863 occurs mostly in N Turkey, S. rufus syriacus Pic, 1892 occurs in S Turkey (Southern costal region and Amanos Mts.) (Sama, 1995) and the nominative S. rufus rufus (Linnaeus, 1767) occurs in other parts of Turkey. Chorotype: Turano-European. According to Sama (2002), this species is not in North Africa.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 223
Tribe MOLORCHINI Glaphyra Newman, 1840 [Type sp.: Glaphyra semiusta Newman, 1840] Subgenus Glaphyra Newman, 1840 [Type sp.: Glaphyra semiusta Newman, 1840] Glaphyra malmusii Sama, 1995 Material examined: Bolu: Abant, Dereceören env., 19.VI.2009, 1 specimen; Abant, 19.VI.2009, 7 specimens. Old records in Bolu prov.: Abant (Sama, 1995 and 2002; Tauzin, 2000). Records in Turkey: (BO) Remarks: The endemic species has been known only from the type locality until now. Chorotype: Anatolian. Tribe CALLIDIINI Hylotrupes Serville, 1834 [Type sp.: Cerambyx bajulus Linnaeus, 1758] Hylotrupes bajulus (Linnaeus, 1758) Old records in Bolu prov.: Abant env., Samat village, Çepni village (Defne, 1954); Bolu prov. (Erdem & Çanakçıoğlu, 1977; Çanakçıoğlu, 1983); Mudurnu (Öymen, 1987); Abant (Tozlu et al., 2002); Mengen (Elemenler) (Özdikmen, et al., 2005). Records in Turkey: (AD-AM-AN-ANT-ART-AY-BI-BO-BR-BS-CA-DE-DU-ER-EZ-GI-GU-HT-IC-IP-IS-IZ-KA-KAR-KN-KR-KS-KU-KY-RI-SN-SV-TB-US-ZO-TRA-TUR) Remarks: The species distributes widely in Turkey. Chorotype: Subcosmopolitan. Ropalopus Mulsant, 1839 [Type sp.: Callidium clavipes Fabricius, 1775] Subgenus Ropalopus Mulsant, 1839 [Type sp.: Callidium clavipes Fabricius, 1775] Ropalopus clavipes (Fabricius, 1775) Material examined: Bolu: Göynük road, 15.VI.2010, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ADY-AN-BL-BO-CN-CO-DE-ED-ER-HT-IC-IS-IZ-KO-KU-MN-MU-NI-OS-US-TRA-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. Chorotype: European or Sibero-European. Sama (2002) reported that this species distributed in Siberia too. Tribe ANAGLYPTINI Paraclytus Bates, 1884 [Type sp.: Paraclytus excultus Bates, 1884] Paraclytus sexguttatus (Adams, 1817) Material examined: Bolu: Yedigöller-Bolu road, Bolu Mts., 14.VI.2010, Pheromone trap, 3 specimens. Old records in Bolu prov.: Abant Lake (Akçaalan) (Özdikmen & Demir, 2006).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 224
Records in Turkey: (ART-BO-KK-TUR) Remarks: The species distributes only in North Turkey. Chorotype: Turano-Mediterranean (Turano-Balkan). Anaglyptus Mulsant, 1839 [Type sp.: Leptura mystica Linnaeus, 1758]
Subgenus Anaglyptus Mulsant, 1839 [Type sp.: Leptura mystica Linnaeus, 1758]
Anaglyptus mysticoides Reitter, 1894 Old records in Bolu prov.: Bolu prov. (Miroshnikov, 2000b). Records in Turkey: (AF-AM-BO-EZ-SM-TB-TO-TUR) Remarks: The species distributes mostly in North Turkey. Chorotype: SW-Asiatic (Anatolo-Caucasian). Tribe CLYTINI Plagionotus Mulsant, 1842 Type sp.: Leptura detrita Linnaeus, 1758] Subgenus Echinocerus Mulsant, 1862 Type sp.: Cerambyx floralis Pallas, 1773] Plagionotus floralis (Pallas, 1773) Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 6 specimens; Gerede-Mengen, 06.VII.2009, 6 specimens; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 1 specimen; Meyitler pass, 1130 m, 08.VII.2009, 8 specimens; Mudurnu-Abant road, 3 km to Abant, 40 km to Bolu, 08.VII.2009, 1 specimen; Aynalı village pass, 08.VII.2009, 4 specimens; Bolu-Seben road, 09.VII.2009, 4 specimens; Abant-Akçaalın, Ömerler wood store, 08.VII.2009, 2 specimens; Akçaalan district, Abant road, 12.VII.2010, 4 specimens; Yeniçağa, 15.VII.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ADY-AF-AG-AM-AN-ANT-AR-ART-BI-BO-BS-BU-BY-CA-CN-CO-DE-EL-ER-ES-EZ-GI-GU-IC-IG-IP-IZ-KA-KAR-KIR-KK-KM-KN-KO-KR-KS-KY-MA-MN-MU-NI-OS-SM-SN-SV-TB-TO-TU-US-YO-ZO-TRA-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. Chorotype: Sibero-European. Chlorophorus Chevrolat, 1863 [Type sp.: Callidium annulare Fabricius, 1787] Chlorophorus cursor Rapuzzi & Sama, 1999 Old records in Bolu prov.: Abant lake (Rapuzzi & Sama, 1999). Records in Turkey: (AN-BO) Remarks: The endemic species is distributed only in NW and CN Anatolia. Chorotype: Anatolian. Chlorophorus dominici Sama, 1996 Material examined: Bolu: Yeniçağa-Mengen road, 14.VI.2010, 3 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (BO-EZ-GI-GU-KS-SN-TO) Remarks: The species has been known only in North Turkey. Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 225
Chlorophorus hungaricus Seidlitz, 1891 Material examined: Bolu: Mengen-Yedigöller, 14.VI.2010, 3 specimens. Old records in Bolu prov.: Bolu Mt. (Özdikmen et al., 2005). Records in Turkey: (AD-AN-BO-BR-GA-IC-KA-KN-KO-KR-KS-NI-OS-SV-TUR) Remarks: The species distributes rather widely in Turkey. Chorotype: Turano-European (Ponto-Pannonian). Chlorophorus sartor (Müller, 1766) Material examined: Bolu: Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 1 specimen; Bolu-Seben road, 09.VII.2009, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-AM-AN-ANT-ART-AY-BI-BL-BO-BR-BS-BU-CA-CN-DE-EL-ES-EZ-GA-GU-HT-IC-IP-IS-IZ-KA-KK-KN-KR-KS-KY-MG-MN-OS-RI-SM-SN-TE-YO-TRA-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. Chorotype: Turano-European. According to Sama (2002), the records from Siberia not confirmed. Chlorophorus trifasciatus (Fabricius, 1781) Material examined: Bolu: Gökçesu village, Mengen-Gökçesu village road, 06.VII.2009, 1 specimen; Mudurnu-Göynük road, Karapınar village env., 07.VII.2009, 1 specimen; Hacıayaz pass, 1080 m, 07.VII.2009, 1 specimen; from Ġkizler plateau road to Abant road, 07.VII.2009, 1 specimen; Aynalı village pass, 08.VII.2009, 1 specimen; Abant-Akçaalın, Ömerler wood store, 08.VII.2009, 1 specimen; Mengen-Yedigöller, 14.VI.2010, 8 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AN-ANT-BI-BO-IC-IS-KN-KO-KR-KS-KU-OS-TUR) Remarks: New to Bolu province. The species distributes rather widely in western half of Turkey. Chorotype: Mediterranean. Chlorophorus varius (Müller, 1766) Chlorophorus varius varius (Müller, 1766) Material examined: Bolu: Göynük road, 30 km to Göynük, SamsaçavuĢ village env., 07.VII.2009, 1 specimen; Bolu-Seben road, 09.VII.2009, 1 specimen. Old records in Bolu prov.: Yeniçağa (AvĢar village) (Özdikmen & Çağlar, 2004). Records in Turkey: (AD-ADY-AK-AM-AN-ANT-ART-AY-BI-BL-BO-BR-BU-CA-CN-DE-ER-ES-EZ-GU-HA-HT-IC-IG-IP-IS-IZ-KA-KI-KIR-KK-KM-KN-KO-KR-KS-KY-MA-MG-MN-MR-MU-NE-NI-OS-SU-TB-TO-US-ZO-VA-TRA-TUR) Remarks: The species distributes widely in Turkey. The species is represented by two subspecies in Turkey. C. varius damascenus Chevrolat, 1854 occurs in S Turkey and the nominative C. varius varius (Müller, 1766) occurs in other parts of Turkey. Chorotype: Palaearctic. Clytus Laicharting, 1784 [Type sp.: Leptura arietis Linnaeus, 1758] Clytus arietis (Linnaeus, 1758) Clytus arietis arietis (Linnaeus, 1758) Material examined: Bolu: Abant, 19.VI.2009, 2 specimens; Gökçesu-Mengen road, 5 km to Mengen, 14.VI.2010, 3 specimens; entry of Abant, 15.VI.2010, 3 specimens; 8 km to Abant, Abant road, 15.VI.2010, 1 specimen; Abant env., 2010, 1 specimen. Old records in Bolu prov.: Pazarköy env. (Özdikmen, 2007).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 226
Records in Turkey: (AM-AN-ART-BO-CA-CN-DU-EZ-GU-IS-KO-KS-SM-TB-ZO-TRA-TUR) Remarks: The species distributes in N and E Turkey. The species has three subspecies in the World. It is represented by two subspecies in Turkey. C. arietis lederi Ganglbauer, 1881 occurs in Caucasus (Talysh, Kopet-Dag and North Iran), E Turkey and the nominative C. arietis arietis (Linnaeus, 1758) occurs in other parts of N Turkey. Chorotype: European. Clytus rhamni Germar, 1817 Material examined: Bolu: Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 2 specimens; Aynalı village pass, 08.VII.2009, 1 specimen; Göynük-Sünnet Lake, 15.VI.2010, 5 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ADY-AM-AN-ANT-ART-BI-BO-BS-BY-CA-CN-GA-GU-HT-IC-IP-IS-IZ-KA-KK-KN-KO-KR-KS-KY-MA-OS-RI-SM-SN-SV-TO-YA-YO-TRA-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. Chorotype: European. Clytus schurmanni Sama, 1996 Material examined: Bolu: Mudurnu-Göynük road, Karapınar village env., 07.VII.2009, 1 specimen. Old records in Bolu prov.: Köroğlu Beli (Özdikmen et al., 2005). Records in Turkey: (AM-AN-BO-CN-CO-IZ-KIR-KR-KS-TO-YO-TUR) Remarks: The endemic species distributes mostly in central parts of N and NW Anatolia. Chorotype: Anatolian. Subfamily LAMIINAE Tribe MONOCHAMINI Monochamus Dejean, 1821 [Type sp.: Cerambyx sutor Linnaeus, 1758]
Subgenus Monochamus Dejean, 1821 [Type sp.: Cerambyx sutor Linnaeus, 1758] Monochamus galloprovincialis (Olivier, 1795) Material examined: Bolu: Mudurnu-Göynük, 20 km to Göynük, 07.VII.2009, 16 specimens; Aladağ, 01.VIII.2010, 1 specimen. Old records in Bolu prov.: Mengen (Elemenler) (Özdikmen et al., 2005). Records in Turkey: (AD-ANT-ART-AY-BO-BS-DE-IC-IP-KAR-KR-KS-MG-OR-SN-TB-TUR) Remarks: Probably it is distributed rather widely in Turkey. Chorotype: Sibero-European. Tribe LAMIINI Morimus Brullé, 1832 [Type sp.: Lamia lugubris Fabricius, 1832 = Cerambyx asper Sulzer, 1776] Morimus funereus Mulsant, 1862 Old records in Bolu prov.: Mengen (Kadısusuz village, Aslı district) (Özdikmen & Hasbenli, 2004); Yeniçağa-Mengen road (Hızırbaba Tekkesi village) (Özdikmen, 2007). Records in Turkey: (AM-AN-ANT-BI-BO-BR-BS-BU-CA-DU-ES-KK-KO-KR-TO-TUR) Remarks: The species distributes only in Northern West half of Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 227
Chorotype: Turano-European (Ponto-Pannonian). Morimus orientalis Reitter, 1894 Material examined: Bolu: Advan village, 12.VIII.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (BO-EZ-IS-SA-TRA-TUR) Remarks: New to Bolu province and thereby to Black Sea Region. Probably it distributes rather widely in Turkey. Chorotype: SW-Asiatic (Irano-Anatolian). Tribe DORCADIINI Dorcadion Dalman, 1817 [Type sp.: Cerambyx glycyrrhizae Pallas, 1771] Subgenus Cribridorcadion Pic, 1901 [Type sp.: Dorcadion mniszechi Kraatz, 1873] Dorcadion bangi Heyden, 1894 Dorcadion bangi roridum Pesarini & Sabbadini, 1999 Material examined: Bolu: Yeniçağ-Gerede road, exit of Gerede, entry of Aydınlar village, 30.IV.2010, 5 specimens. Old records in Bolu prov.: Gerede (Braun, 1978, 1979). Records in Turkey: (AN-BO-CO-KR-KS) Remarks: This species is endemic to CN and NCW Turkey. According to Braun (1978 and 1979), D. heinzorum Braun, 1975 is a subspecies of D. bangi Heyden, 1894. So it is represented by three subspecies. The nominotypical subspecies D. bangi bangi Heyden, 1894 occurs only in West parts of Western Black Sea Region (Kastamonu and Bolu provinces). D. bangi heinzorum Braun, 1975 occurs in Kastamonu and Çorum provinces. D. bangi roridum was described by Pesarini & Sabbadini (1998) from Bolu province (Gerede). Özdikmen (2007) gave D. heinzorum Braun, 1975 as a subspecies of D. bangi Heyden, 1894. Chorotype: Anatolian. Dorcadion bodemeyeri Daniel, 1900 Material examined: Bolu: Yeniçağ-Gerede road, exit of Gerede, entry of Aydınlar village, 30.IV.2010, 5 specimens. Old records in Bolu prov.: GölbaĢı (Demelt, 1963); Central and GölbaĢı (Önalp, 1990). Records in Turkey: (AF-AM-AN-BO-ES-IZ-KN-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The endemic species distributes mostly in the western half of Anatolia. Chorotype: Anatolian. Dorcadion boluense Breuning, 1962 Dorcadion boluense boluense Breuning, 1962 Old records in Bolu prov.: Akyarma pass (Adlbauer, 1992); Records in Turkey: (AN-BO-TUR) Remarks: This species is endemic to Turkey. It has 3 subspecies. All of them are distributed only in NCW Turkey. The nominotypical subspecies occurs in Bolu province (Bolu Mountains, Gerede, Çifkan Mt., Akyarma pass). D. boluense imitator was described by Pesarini & Sabbadini (1998) from Ankara province (Karagöl) and Çankırı province (IĢık Mountain). Also D. boluense corallinum was described by Pesarini & Sabbadini (1998) from Ankara province (Kızılcahamam, Çamlıdere). Chorotype: Anatolian.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 228
Dorcadion maceki Holzschuh, 1995 Old records in Bolu prov.: Abant Lake (Holzschuh, 1995). Records in Turkey: (BO-TUR) Remarks: This species is endemic to SCW Turkey. It has been known only from type locality until now. Chorotype: Anatolian. Dorcadion pararufipenne Braun, 1976 Old records in Bolu prov.: Abant Mts. (Braun, 1978); Gerede / Central as D. pararufipenne rassei (Braun, 1978); Bolu prov. as D. pararufipenne rassei (Adlbauer, 1992); Abant (Özdikmen & Demir, 2006). Records in Turkey: (AN-BO) Remarks: This species is endemic to NCW Turkey and probably distributes in a local area. It has two subspecies. D. pararufipenne rassei Braun, 1976 occurs probably eastward from the distribution patterns of the nominotypical subspecies. Chorotype: Anatolian. Dorcadion praetermissum Pesarini & Sabbadini, 1998 Dorcadion praetermissum mikhaili Özdikmen, 2010 Material examined: Bolu: Abant, 30.IV.2010, 2 specimens. Old records in Bolu prov.: Abant (Özdikmen, 2010). Records in Turkey: (BO-BS-TUR) Remarks: This species is endemic to NW Anatolia. This species has two subspecies now. D. praetermissum mikhaili was described by Özdikmen (2010) from Bolu province on the base of the present materials. Chorotype: Anatolian. Dorcadion septemlineatum Waltl, 1838 Dorcadion septemlineatum abanti Braun, 1976 Old records in Bolu prov.: Abant Mts. as D. septemlineatum abanti (Braun, 1978); Abant (Sama, 1982); Abant lake as D. septemlineatum abanti (Adlbauer, 1992); Gerede (Yeniçağ) as D. septemlineatum abanti (Özdikmen & Hasbenli, 2004); Dörtdivan as D. septemlineatum abanti (Özdikmen et al., 2005). Records in Turkey: (AF-AN-BI-BL-BO-BS-BU-CA-ES-GA-IP-IS-KN-KO-KR-KU-SA-TRA-TUR) Remarks: The species distributes rather widely in Turkey (especially west half of Turkey). It has five subspecies. All subspecies are represented in Turkey. The nominotypical subspecies occurs also in Greece and Bulgaria. This species is distributed rather widely in Turkey (especially W and N Turkey). The nominotypical subspecies, D. septemlineatum septemlineatum Waltl, 1838, occurs mainly in European Turkey. Chorotype: Turano-Mediterranean (Balkano-Anatolian). Tribe POGONOCHERINI Pogonocherus Dejean, 1821 [Type sp.: Cerambyx hispidus Linnaeus, 1758] Subgenus Pityphilus Mulsant, 1862 [Type sp.: Cerambyx ovatus Goeze, 1777] Pogonocherus decoratus Fairmaire, 1855 Old records in Bolu prov.: Abant Mountains (Demelt, 1967) Records in Turkey: (AN-BO-KS-TUR) Remarks: The species distributes in N Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 229
Chorotype: European or Sibero-European. Tribe ACANTHOCININI Acanthocinus Dejean, 1821 [Type sp.: Cerambyx aedilis Linnaeus, 1758] Acanthocinus aedilis (Linnaeus, 1758) Material examined: Bolu: Mudurnu-Göynük, 20 km to Göynük, 07.VII.2009, 1 specimen. Old records in Bolu prov.: Abant (Alkan, 1946; Tozlu et al., 2003). Records in Turkey: (AM-AN-ANT-ART-BI-BL-BO-BS-CA-DE-ES-EZ-GI-GU-IP-IZ-KAR-KR-KS-KU-MG-SN-TO-TRA-TUR) Remarks: The species distributes widely in Turkey. Chorotype: Sibero-European. Acanthocinus griseus (Fabricius, 1792) Material examined: Bolu: Mengen forest management, 06.VII.2009, Pheromone trap, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ANT-BO-BS-DE-ED-EZ-HT-IC-IZ-KO-MG-MN-TB-TRA-TUR) Remarks: New to Bolu and thereby to Western Black Sea Region. The species distributes rather widely in Turkey. Chorotype: Sibero-European. Tribe SAPERDINI Saperda Fabricius, 1775 [Type sp.: Cerambyx carcharias Linnaeus, 1758] Subgenus Compsidia Mulsant, 1839 [Type sp.: Cerambyx populneus Linnaeus, 1758]
Saperda populnea (Linnaeus, 1758) Saperda populnea populnea (Linnaeus, 1758) Old records in Bolu prov.: Central / Mengen (Sekendiz, 1974). Records in Turkey: (ART-BL-BO-BS-BT-BU-BY-CA-DE-DU-ED-ER-GI-GU-HT-IS-IZ-KA-KN-KO-KY-MN-MU-SA-SM-SN-TB-VA-YO-ZO-TUR) Remarks: The species distributes widely in Turkey. It is represented by the nominotypical subspecies in Turkey. Known other subspecies S. populnea moesta LeConte, 1850 and S. populnea tulari Felt & Joutel, 1904 occurring only in Nearctic Region (United States and Canada). Chorotype: Sibero-European + Nearctic. Sama (2002) stated that records from North Africa are clearly erroneous due to mislabellings. Tribe PHYTOECIINI Oberea Dejean, 1835 [Type sp.: Cerambyx linearis Linnaeus, 1760] Subgenus Amaurostoma Müller, 1906 [Type sp.: Cerambyx erythrocephalus Schrank, 1776]

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 230
Oberea erythrocephala (Schrank, 1776) Oberea erythrocephala amanica Holzschuh, 1993 Material examined: Bolu: between Dereceören-Akçaalan, Abant, 19.VI.2009, on Euphorbia, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AF-AM-AN-ANT-ART-BO-BY-CO-ER-EZ-GU-IS-KA-KAR-KN-KO-KS-NI-OS-SV-VA-TRA-TUR) Remarks: New to Bolu province. The species distributes widely in Turkey. It is represented by three subspecies in Turkey. O. erythrocephala amanica Holzschuh, 1993 occurs in N Turkey, O. erythrocephala schurmanni Heyrovsky, 1962 occurs mainly in C, S and E Turkey and O. erythrocephala erythrocephala (Schrank, 1776) occurs in the other parts of Turkey (especially European Turkey, NW and W Anatolia). The other known subspecies, O. erythrocephala calvescens Müller, 1948 occurs only in Italy. Chorotype: Palaearctic. Oxylia Mulsant, 1862 [Type sp.: Phytoecia languid Ménétriés, 1839] Oxylia duponcheli (Brullé, 1832) Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 1 specimen. Old records in Bolu prov.: Bağlum, Kızılcahamam (Güvem) (Özdikmen et al., 2005); Çal Mountain (Özdikmen & Demir, 2006). Records in Turkey: (AK-AN-ANT-ART-BO-ES-IC-KA-KM-KN-MA-MN-OS-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes rather widely in Turkey. Chorotype: Turano-Mediterranean (Balkano-Anatolian). Phytoecia Dejean, 1835 [Type sp.: Cerambyx cylindricus Linnaeus, 1758] Subgenus Helladia Fairmaire, 1864 [Type sp.: Saperda flavescens Brullé, 1832] Phytoecia humeralis (Waltl, 1838) Phytoecia humeralis humeralis (Waltl, 1838) Material examined: Bolu: Karacasu env., 18.VI.2009, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AD-ADY-AK-AM-AN-ANT-AY-BO-BU-DE-DI-ED-ES-GA-HA-HT-IC-IP-IZ-KA-KN-MN-NI-OS-US-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes widely in Turkey. It is represented by two subspecies in Turkey. P. humeralis caneri Özdikmen & Turgut, 2010 occurs only in S Turkey. Chorotype: E-Mediterranean (Palaestino-Cyprioto-Taurian + NE-Mediterranean). Phytoecia praetextata (Steven, 1817) Phytoecia praetextata praetextata (Steven, 1817) Material examined: Bolu: Yeniçağa-Mengen road, 14.VI.2010, 2 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AN-BO-BY-DU-EZ-GU-HT-IC-KS-SV-ZO-TUR) Remarks: New to Bolu province. The species distributes rather widely in Turkey. It is represented by two subspecies in Turkey. P. praetextata nigricollis occurs in S Turkey and the nominative H. praetextata praetextata occurs mostly in N Turkey. Chorotype: Turano-Mediterranean (Turano-Balkan).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 231
Subgenus Musaria Thomson, 1864 [Type sp.: Leptura affinis Harrer, 1784] Phytoecia affinis (Harrer, 1784) Phytoecia affinis affinis (Harrer, 1784) Material examined: Bolu: Abant, Dereceören env., 19.VI.2009, 10 specimens; 8 km to Abant, Abant road, 15.VI.2010, 2 specimens; Abant-Dereceören env., 08.VII.2009, 4 specimens. Old records in Bolu prov.: Yedigöller National Park (Özdikmen & Hasbenli, 2004). Records in Turkey: (BO-TUR) Remarks: The species distributes in North Turkey. It is represented by nominative subspecies in Turkey. Chorotype: Sibero-European. Subgenus Phytoecia Dejean, 1835 [Type sp.: Cerambyx cylindricus Linnaeus, 1758] Phytoecia icterica (Schaller, 1783) Material examined: Bolu: Abant, 30.IV.2010, 1 örnek. Old records in Bolu prov.: Yeniçağa-Mengen road (Çamlık village) (Özdikmen, 2007). Records in Turkey: (AF-AN-ANT-BO-BT-BY-CO-EZ-HT-IS-KA-KAR-KN-KS-KU-YO-TRA-TUR) Remarks: The species probably distributes rather widely in Turkey. Chorotype: Turano-European. Phytoecia pubescens Pic, 1895 Material examined: Bolu: Kıbrıscık-Seben yolu, Seben‘e 10 km kala, 09.VII.2009, 1 örnek. Old records in Bolu prov.: Absent. Records in Turkey: (AM-AN-GA-KL-KO-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes in N Turkey. Chorotype: Turano-Mediterranean (Turano-E-Mediterranean). Subgenus Opsilia Mulsant, 1862 [Type sp.: Opsilia flavicans Mulsant, 1862 = Leptura coerulescens Scopoli, 1763 ] Opsilia coerulescens (Scopoli, 1763) Opsilia coerulescens coerulescens (Scopoli, 1763) Material examined: Bolu: Kıbrıscık-Seben road, 10 km to Seben, 18.VI.2009, 3 specimens; Abant-Dereceören env., 19.VI.2009, 1 örnek; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 2 specimens; Meyitler pass, 1130 m, 08.VII.2009, 3 specimens; entry of Göynük-Mudurnu road, Atyaylası pass, exit of Sarılar village, 08.VII.2009, 1 specimen; Abant-Dereceören env., 08.VII.2009, 1 specimen; Kıbrıscık-Seben road, 10 km to Seben, 09.VII.2009, 1 specimen; Yeniçağa, 14.VI.2010, 1 specimen; Abant, 30.IV.2010, 2 specimens. Old records in Bolu prov.: Abant (Sama, 1982); Mudurnu (Özdikmen et al., 2005); Bolu-Gerede road (Susuz Kınık village), Entry of Gerede expressway, Gerede–Karabük road (Dikmen and Sungur villages), Mengen (Devrek–Mengen, entry of Mengen) (Özdikmen, 2007). Records in Turkey: (AD-ADY-AK-AM-AN-ANT-AR-ART-AY-BO-BS-BU-BY-CN-CO-DE-DI-ER-ES-EZ-GA-GU-IC-IP-IS-IZ-KA-KAR-KIR-KK-KM-KN-KS-KY-MA-MG-MN-NE-NI-OS-SM-SN-SV-TB-YO-ZO-TRA-TUR) Remarks: The species distributes widely in Turkey. The species is represented by the nominotypical subspecies in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 232
Chorotype: Palaearctic. Tribe AGAPANTHIINI Calamobius Guérin, 1847 [Type sp.: Saperda gracilis Creutzer, 1799. = Saperda filum Rossi, 1790] Calamobius filum (Rossi, 1790) Old records in Bolu prov.: KaynaĢlı, Sakarya prov.: Sapanca (Özdikmen et al., 2005). Records in Turkey: (AD-AN-ANT-BO-BS-BU-CA-GA-HT-IC-IP-IS-IZ-KA-KL-KN-KO-MG-MN-OS-SA-TRA-TUR) Remarks: The species distributes rather widely in Turkey (especially west half of Turkey). Chorotype: Turano-Europeo-Mediterranean. Agapanthia Serville, 1835 [Type sp.: Saperda cardui Fabricius, 1801 = Cerambyx cardui Linnaeus, 1767] Subgenus Synthapsia Pessarini & Sabbadini, 2004 [Type sp.: Saperda kirbyi Gyllenhal, 1817] Agapanthia kirbyi (Gyllenhal, 1817) Material examined: Bolu: between Dereceören-Akçaalan, Abant, 19.VI.2009, 4 specimens; entry of Göynük-Mudurnu road, Atyaylası pass, exit of Sarılar village, 08.VII.2009, 6 specimens; Yeniçağa-Mengen road, 14.VI.2010, 5 specimens; 8 km to Abant, Abant road, 15.VI.2010, 1 specimen; Abant env., 2010, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-AF-AK-AM-AN-ANT-BI-BO-BS-BT-BU-CO-ED-ER-ES-EZ-IC-IP-IZ-KA-KAR-KIR-KN-KO-KY-MN-NI-OS-SI-TO-VA-TRA-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes widely in Turkey. Chorotype: Turano-European. Subgenus Epoptes Gistel, 1857 [Type sp.: Lamia asphodeli Latreille, 1804] Agapanthia cynarae (Germar, 1817) Agapanthia cynarae cynarae (Germar, 1817) Material examined: Bolu: Gerede, 06.VII.2009, 1 specimen; Mudurnu-Göynük road, Sünnet Lake env., 07.VII.2009, 2 specimens. Old records in Bolu prov.: Absent. Records in Turkey: (AD-AN-ANT-AY-BI-BO-CA-HT-IP-IZ-YO-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes mostly in west half of Turkey. It is represented by the nominotypical subspecies in Turkey. Chorotype: European. Agapanthia dahli (Richter, 1820) Material examined: Bolu: Mengen, Mengen-Yedigöller road, 14.VI.2010, 1 specimen. Old records in Bolu prov.: Absent. Records in Turkey: (AD-AN-BO-BS-EZ-GA-GU-HT-OS-SI-TUR) Remarks: New to Bolu province and thereby to Western Black Sea Region. The species distributes rather widely in Turkey. Chorotype: Sibero-European or Turano-European. Since, according to Sama (2002) most records from East Mediterranean and Central Asia of this species probably belong to different species.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 233
Agapanthia lateralis Ganglbauer, 1884 Old records in Bolu prov.: Entry of Gerede expressway, Gerede–Karabük road (entry of Koçumlar village), Yeniçağa (Özdikmen, 2007). Records in Turkey: (AF-AG-AK-AM-AN-ANT-BI-BO-CA-CN-CO-ES-IC-IP-IS-IZ-KA-KIR-KM-KN-KR-KS-MG-MN-NE-NI-TE-TO-ZO-TRA-TUR) Remarks: The species distributes widely in Turkey. Chorotype: E-Mediterranean. Subgenus Smaragdula Pesarini & Sabbadini, 2004 [Type sp.: Saperda violacea Fabricius, 1775] Agapanthia violacea (Fabricius, 1775) Material examined: Bolu: Abant, 30.IV.2010, 1 specimen; Mudurnu-Göynük road, Karapınar return env., 01.V.2010, 1 specimen; Yeniçağa-Mengen road, 14.VI.2010, 1 specimen. Old records in Bolu prov.: Seben (Özdikmen et al., 2005); Gerede–Bolu road, Bolu-Gerede road (Susuz Kınık village), between Gerede–Kızılcahamam, Yeniçağa-Mengen road (Çamlık village env.) (Özdikmen, 2007). Records in Turkey: (AD-AF-AK-AN-BI-BO-BS-CO-DE-DU-ED-EZ-GA-HT-IC-IP-IS-IZ-KA-KIR-KK-KN-KO-KR-KS-KY-MG-MN-NE-NI-OS-SA-ZO-TRA-TUR) Remarks: The species distributes widely in Turkey. Chorotype: Sibero-European.
Consequently, according to old references, the longhorned beetles fauna of Bolu province included 51 species of 33 genera of 6 subfamilies: [Prioninae: 2 species as Ergates faber (Linnaeus, 1761) and Prionus coriarius (Linnaeus, 1758); Lepturinae: 19 species as Xylosteus caucasicola Plavilstshikov, 1936, X. kadleci Miroshnikov, 2000, Rhagium bifasciatum Fabricius, 1775, R. fasciculatum Faldermann, 1837, R. inquisitor (Linnaeus, 1758), Cortodera flavimana (Waltl, 1838), C. humeralis (Schaller, 1783), Fallacia elegans (Faldermann, 1837), Vadonia unipunctata (Fabricius, 1787), Pseudovadonia livida (Fabricius, 1777), Stictoleptura rubra (Linnaeus, 1758), S. fulva (DeGeer, 1775), Anastrangalia dubia (Scopoli, 1763), A. sanguinolenta (Linnaeus, 1761), Pachytodes erraticus (Dalman, 1817), Solaia antonellae Sama, 2003, Stenurella bifasciata (Müller, 1776), S. nigra (Linnaeus, 1758) and S. septempunctata (Fabricius, 1792); Necydalinae: 1 species as Necydalis sabatinellii Sama, 1994; Aseminae: 4 species as Drymochares starcki Ganglbauer, 1888, Arhopalus ferus (Mulsant, 1839), A. rusticus (Linnaeus, 1758) and Tetropium castaneum (Linnaeus, 1758); Cerambycinae: 10 species as Stenopterus rufus (Linnaeus, 1767), Glaphyra malmusii Sama, 1995, Hylotrupes bajulus (Linnaeus, 1758), Paraclytus sexguttatus (Adams, 1817), Anaglyptus mysticoides Reitter, 1894, Chlorophorus cursor Rapuzzi & Sama, 1999, C. hungaricus Seidlitz, 1891, C. varius (Müller, 1776), Clytus arietis (Linnaeus, 1758) and C. schurmanni Sama, 1996; Lamiinae: 15 species as Monochamus galloprovincialis (Olivier, 1795), Morimus funereus Mulsant, 1862, Dorcadion bangi Heyden, 1894, D. boluense Breuning, 1962, D. maceki Holzschuh, 1995; D. pararufipenne Braun, 1976, D. septemlineatum Waltl, 1838, Pogonocherus decoratus Fairmaire, 1855, Acanthocinus aedilis (Linnaeus, 1758), Saperda populnea (Linnaeus, 1758), Phytoecia affinis (Harrer, 1784), P. coerulescens (Scopoli, 1763), Calamobius filum (Rossi, 1790), Agapanthia lateralis Ganglbauer, 1884 and A. violacea (Fabricius, 1775)].
With the present work, 52 species of 31 genera of 5 subfamilies among the
collected specimens from Bolu province were determined:

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 234
[Prioninae: 2 species as Ergates faber (Linnaeus, 1760) and Prionus coriarius (Linnaeus, 1758); Lepturinae: 15 species as Xylosteus kadleci Miroshnikov, 2000, Rhagium inquisitor (Linnaeus, 1758), Dinoptera collaris (Linnaeus, 1758), Vadonia unipunctata (Fabricius, 1787), Pseudovadonia livida (Fabricius, 1777), Anoplodera rufipes (Schaller, 1783), Stictoleptura fulva (DeGeer, 1775), S. rufa (Brullé, 1832), Anastrangalia dubia (Scopoli, 1763), A. sanguinolenta (Linnaeus, 1761), Pachytodes erraticus (Dalman, 1817), Rutpela maculata (Poda, 1761), Stenurella bifasciata (Müller, 1776), S. nigra (Linnaeus, 1758) and S. septempunctata (Fabricius, 1792); Spondylidinae: 1 species as Spondylis buprestoides (Linnaeus, 1758); Cerambycinae: 15 species as Purpuricenus budensis (Götz, 1783), Obrium brunneum (Fabricius, 1792), Stenopterus rufus (Linnaeus, 1767), Glaphyra malmusii Sama, 1995, Ropalopus clavipes (Fabricius, 1775), Paraclytus sexguttatus (Adams, 1817), Plagionotus floralis (Pallas, 1773), Chlorophorus dominici Sama, 1996, C. hungaricus Seidlitz, 1891, C. sartor (Müller, 1766), C. trifasciatus (Fabricius, 1781), C. varius (Müller, 1776), Clytus arietis (Linnaeus, 1758), C. rhamni Germar, 1817 and C. schurmanni Sama, 1996; Lamiinae: 19 species as Monochamus galloprovincialis (Olivier, 1795), Morimus orientalis Reitter, 1894, Dorcadion bangi Heyden, 1894, D. bodemeyeri Daniel, 1900, D. praetermissum Pesarini & Sabbadini, 1998, Acanthocinus aedilis (Linnaeus, 1758), A. griseus (Fabricius, 1792), Oberea erythrocephala (Schrank, 1776), Oxylia duponcheli (Brullé, 1832), Phytoecia humeralis (Waltl, 1838), P. praetextata (Steven, 1817), P. affinis (Harrer, 1784), P. icterica (Schaller, 1783), P. pubescens Pic, 1895, P. coerulescens (Scopoli, 1763), Agapanthia kirbyi (Gyllenhal, 1817), A. cynarae (Germar, 1817), A. dahli (Richter, 1821) and A. violacea (Fabricius, 1775)].
25 of these species are new records to Bolu province:
[Lepturinae: 4 species as Dinoptera collaris (Linnaeus, 1758), Anoplodera rufipes (Schaller, 1783), Stictoleptura rufa (Brullé, 1832) and Rutpela maculata (Poda, 1761); Spondylidinae: 1 species as Spondylis buprestoides (Linnaeus, 1758); Cerambycinae: 8 species as Purpuricenus budensis (Götz, 1783), Obrium brunneum (Fabricius, 1792), Ropalopus clavipes (Fabricius, 1775), Plagionotus floralis (Pallas, 1773), Chlorophorus dominici Sama, 1996, C. sartor (Müller, 1766), C. trifasciatus (Fabricius, 1781) and Clytus rhamni Germar, 1817; Lamiinae: 12 species as Morimus orientalis Reitter, 1894, Dorcadion bodemeyeri Daniel, 1900, D. praetermissum Pesarini & Sabbadini, 1998, Acanthocinus griseus (Fabricius, 1792), Oberea erythrocephala (Schrank, 1776), Oxylia duponcheli (Brullé, 1832), Phytoecia humeralis (Waltl, 1838), P. praetextata (Steven, 1817), P. pubescens Pic, 1895, Agapanthia kirbyi (Gyllenhal, 1817), A. cynarae (Germar, 1817) and A. dahli (Richter, 1821)].
Morimus orientalis Reitter, 1894 is new record for Black Sea Region.
9 species are new records for Western Black Sea Region:
[Stictoleptura rufa (Brullé, 1832), Dorcadion bodemeyeri Daniel, 1900, Acanthocinus griseus (Fabricius, 1792), Oxylia duponcheli (Brullé, 1832), Phytoecia humeralis (Waltl, 1838), P. pubescens Pic, 1895, Agapanthia kirbyi (Gyllenhal, 1817), A. cynarae (Germar, 1817) and Agapanthia dahli (Richter, 1820)]
Moreover, Dorcadion praetermissum mikhaili was described by Özdikmen
(2010) from Bolu province on the base of the specimens in the present work. Finally, Longhorned Beetles Fauna of Bolu province comprises of 79 species of
43 genera of 24 tribes belonging to 7 subfamilies with the newly recorded species in the present work. Number of known species from Bolu province was increased from 51 species to 79 species.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 235
A faunistic list are presented as follows:
In the list, the sign of (*****) is used for recently described subspecies, (****) is used for new record to Black Sea Region, (***) is used for new records to Western Black Sea Region, (**) is used for new records to Bolu province and (*) is used for known species in both old references and the present text. Any sign is not used for the known species only from old references.
LONGHORNED BEETLES FAUNA OF BOLU PROVINCE (COLEOPTERA: CERAMBYCIDAE)
Family CERAMBYCIDAE Subfamily PRIONINAE Latreille, 1802 Tribe ERGATINI Fairmaire, 1864
Genus ERGATES Serville, 1832 Ergates faber (Linnaeus, 1761) (*)
Ergates faber faber (Linnaeus, 1761) (*) Tribe PRIONINI Latreille, 1802
Genus PRIONUS Geoffroy, 1762 Prionus coriarius (Linnaeus, 1758) (*)
Subfamily LEPTURINAE Latreille, 1802 Tribe XYLOSTEINI Fairmaire, 1864
Genus XYLOSTEUS Frivaldszky, 1837 Xylosteus caucasicola Plavilstshikov, 1936 Xylosteus kadleci Miroshnikov, 2000 (*)
Tribe RHAGIINI Mulsant, 1839 Genus RHAGIUM Fabricius, 1775
Subgenus HAGRIUM Villiers, 1978 Rhagium bifasciatum Fabricius, 1775
Subgenus MEGARHAGIUM Reitter, 1913 Rhagium fasciculatum Faldermann, 1837
Subgenus RHAGIUM Fabricius, 1775 Rhagium inquisitor (Linnaeus, 1758) (*)
Rhagium inquisitor inquisitor (Linnaeus, 1758) (*) Genus DINOPTERA Mulsant, 1863
Dinoptera collaris (Linnaeus, 1758) (**) Genus CORTODERA Mulsant, 1863
Cortodera flavimana (Waltl, 1838) Cortodera humeralis (Schaller, 1783)
Cortodera humeralis humeralis (Schaller, 1783) Genus FALLACIA Mulsant & Rey, 1863
Fallacia elegans (Faldermann, 1837) Tribe LEPTURINI Latreille, 1802
Genus VADONIA Mulsant, 1863 Vadonia unipunctata (Fabricius, 1787) (*)
Vadonia unipunctata unipunctata (Fabricius, 1787) (*) Genus PSEUDOVADONIA Lobanov et al., 1981
Pseudovadonia livida (Fabricius, 1777) (*) Pseudovadonia livida livida (Fabricius, 1777) (*)
Genus ANOPLODERA Mulsant, 1839 Anoplodera rufipes (Schaller, 1783) (**)
Anoplodera rufipes rufipes (Schaller, 1783) (**)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 236
Genus STICTOLEPTURA Casey, 1924 Subgenus AREDOLPONA Nakane & Ohbayashi, 1957
Stictoleptura rubra (Linnaeus, 1758) Stictoleptura rubra rubra (Linnaeus, 1758)
Subgenus STICTOLEPTURA Casey, 1924 Stictoleptura fulva (DeGeer, 1775) (*) Stictoleptura rufa (Brullé, 1832 (**) (***)
Stictoleptura rufa dimidiata (Daniel & Daniel, 1891) (**) (***) Genus ANASTRANGALIA Casey, 1924
Anastrangalia dubia (Scopoli, 1763) (*) Anastrangalia dubia dubia (Scopoli, 1763) (*)
Anastrangalia sanguinolenta (Linnaeus, 1761) (*) Genus PACHYTODES Pic, 1891
Pachytodes erraticus (Dalman, 1817) (*) Pachytodes erraticus erraticus (Dalman, 1817) (*)
Genus SOLAIA Sama, 2003 Solaia antonellae Sama, 2003
Genus RUTPELA Nakane & Ohbayashi, 1957 Rutpela maculata (Poda, 1761) (**)
Rutpela maculata maculata (Poda, 1761) (**) Genus STENURELLA Villiers, 1974
Stenurella bifasciata (Müller, 1776) (*) Stenurella bifasciata bifasciata (Müller, 1776) (*)
Stenurella melanura (Linnaeus, 1758) Stenurella nigra (Linnaeus, 1758) (*) Stenurella septempunctata (Fabricius, 1792) (*)
Stenurella septempunctata suturata (Reiche & Saulcy, 1858) (*)
Subfamily NECYDALINAE Latreille, 1825 Tribe NECYDALINI Latreille, 1825
Genus NECYDALIS Linnaeus, 1758 Subgenus NECYDALIS Linnaeus, 1758
Necydalis sabatinellii Sama, 1994 Subfamily ASEMINAE Thomson, 1861 Tribe SAPHANINI Gistel, 1856
Genus DRYMOCHARES Mulsant, 1847 Drymochares starcki Ganglbauer, 1888
Drymochares starcki ivani Sama & Rapuzzi, 1993 Tribe ASEMINI Thomson, 1861
Genus ARHOPALUS Serville, 1832 Arhopalus ferus (Mulsant, 1839) Arhopalus rusticus (Linnaeus, 1758)
Genus TETROPIUM Kirby, 1837 Tetropium castaneum (Linnaeus, 1758)
Subfamily SPONDYLIDINAE Audinet-Serville, 1832 Tribe SPONDYLIDINI Audinet-Serville, 1832
Genus SPONDYLIS Fabricius, 1775 Spondylis buprestoides (Linnaeus, 1758) (**)
Subfamily CERAMBYCINAE Latreille, 1802 Tribe TRACHYDERINI Dupont, 1836
Genus PURPURICENUS Dejean, 1821 Subgenus PURPURICENUS Dejean, 1821
Purpuricenus budensis (Götz, 1783) (**) Tribe OBRIINI Mulsant, 1839
Genus OBRIUM Dejean, 1821 Obrium brunneum (Fabricius, 1792) (**)

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 237
Tribe STENOPTERINI Gistel, 1848 Genus STENOPTERUS Illiger, 1804
Stenopterus rufus (Linnaeus, 1767) (*) Stenopterus rufus geniculatus Kraatz, 1863 (*)
Tribe MOLORCHINI Gistel, 1848 Genus GLAPHYRA Newman, 1840
Subgenus GLAPHYRA Newman, 1840 Glaphyra malmusii Sama, 1995 (*)
Tribe CALLIDIINI Kirby, 1837 Genus HYLOTRUPES Serville, 1834
Hylotrupes bajulus (Linnaeus, 1758) Genus ROPALOPUS Mulsant, 1839
Subgenus ROPALOPUS Mulsant, 1839 Ropalopus clavipes (Fabricius, 1775) (**)
Tribe ANAGLYPTINI Lacordaire, 1868 Genus PARACLYTUS Bates, 1884
Paraclytus sexguttatus (Adams, 1817) (*) Genus ANAGLYPTUS Mulsant, 1839
Subgenus ANAGLYPTUS Mulsant, 1839 Anaglyptus mysticoides Reitter, 1894
Tribe CLYTINI Mulsant, 1839 Genus PLAGIONOTUS Mulsant, 1842
Subgenus ECHINOCERUS Mulsant, 1862 Plagionotus floralis (Pallas, 1773) (**)
Genus CHLOROPHORUS Chevrolat, 1863 Chlorophorus cursor Rapuzzi & Sama, 1999 Chlorophorus dominici Sama, 1996 (**) Chlorophorus hungaricus Seidlitz, 1891 (*) Chlorophorus sartor (Müller, 1766) (**) Chlorophorus trifasciatus (Fabricius, 1781) (**) Chlorophorus varius (Müller, 1776) (*)
Chlorophorus varius varius (Müller, 1776) (*) Genus CLYTUS Laicharting, 1784
Clytus arietis (Linnaeus, 1758) (*) Clytus arietis arietis (Linnaeus, 1758) (*)
Clytus rhamni Germar, 1817 (**) Clytus schurmanni Sama, 1996 (*)
Subfamily LAMIINAE Latreille, 1825 Tribe MONOCHAMINI Gistel, 1848
Genus MONOCHAMUS Dejean, 1821 Subgenus MONOCHAMUS Dejean, 1821
Monochamus galloprovincialis (Olivier, 1795) (*) Tribe LAMIINI Latreille, 1825
Genus MORIMUS Brulle, 1832 Morimus funereus Mulsant, 1862 Morimus orientalis Reitter, 1894 (**) (****)
Tribe DORCADIINI Latreille, 1825 Genus DORCADION Dalman, 1817
Subgenus CRIBRIDORCADION Pic, 1901 Dorcadion bangi Heyden, 1894 (*)
Dorcadion bangi bangi Heyden, 1894 Dorcadion bangi roridum Pesarini & Sabbadini, 1998 (*)
Dorcadion bodemeyeri Daniel, 1900 (**) (***) Dorcadion boluense Breuning, 1962
Dorcadion boluense boluense Breuning, 1962 Dorcadion maceki Holzschuh, 1995 Dorcadion pararufipenne Braun, 1976
Dorcadion pararufipenne pararufipenne Braun, 1976

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 238
Dorcadion pararufipenne rassei Braun, 1976 Dorcadion praetermissum Pesarini & Sabbadini, 1998 (**)
Dorcadion praetermissum mikhaili Özdikmen, 2010 (*****) Dorcadion septemlineatum Waltl, 1838
Dorcadion septemlineatum abanti Braun, 1976 Tribe POGONOCHERINI Mulsant, 1839
Genus POGONOCHERUS Dejean, 1821 Pogonocherus decoratus Fairmaire, 1855
Tribe ACANTHOCININI Blanchard, 1845 Genus ACANTHOCINUS Dejean, 1821
Acanthocinus aedilis (Linnaeus, 1758) (*) Acanthocinus griseus (Fabricius, 1792) (**) (***)
Tribe SAPERDINI Mulsant, 1839 Genus SAPERDA Fabricius, 1775
Subgenus COMPSIDIA Mulsant, 1839 Saperda populnea (Linnaeus, 1758)
Tribe PHYTOECIINI Mulsant, 1839 Genus OBEREA Dejean, 1835
Subgenus AMAUROSTOMA Müller, 1906 Oberea erythrocephala (Schrank, 1776) (**)
Oberea erythrocephala amanica Holzschuh, 1993 (**) Genus OXYLIA Mulsant, 1862
Oxylia duponcheli (Brullé, 1832) (**) (***) Genus PHYTOECIA Dejean, 1835
Subgenus HELLADIA Fairmaire, 1864 Phytoecia humeralis (Waltl, 1838) (**) (***)
Phytoecia humeralis humeralis (Waltl, 1838) (**) (***) Phytoecia praetextata (Steven, 1817) (**)
Phytoecia praetextata praetextata (Steven, 1817) (**) Subgenus MUSARIA Thomson, 1864
Phytoecia affinis (Harrer, 1784) (*) Phytoecia affinis affinis (Harrer, 1784) (*)
Subgenus PHYTOECIA Dejean, 1835 Phytoecia icterica (Schaller, 1783) (*) Phytoecia pubescens Pic, 1895 (**) (***)
Subgenus OPSILIA Mulsant, 1862 Phytoecia coerulescens (Scopoli, 1763) (*)
Phytoecia coerulescens coerulescens (Scopoli, 1763) (*) Tribe AGAPANTHIINI Mulsant, 1839
Genus CALAMOBIUS Guerin-Meneville, 1847 Calamobius filum (Rossi, 1790)
Genus AGAPANTHIA Serville, 1835 Subgenus SYNTHAPSIA Pesarini & Sabbadini, 2004
Agapanthia kirbyi (Gyllenhal, 1817) (**) (***) Subgenus EPOPTES Gistel, 1857
Agapanthia cynarae (Germar, 1817) (**) (***) Agapanthia cynarae cynarae (Germar, 1817) (**) (***)
Agapanthia dahli (Richter, 1821) (**) (***) Agapanthia lateralis Ganglbauer, 1884
Subgenus SMARAGDULA Pesarini & Sabbadini, 2004 Agapanthia violacea (Fabricius, 1775) (*)
* This work supported by the project of GAZI UNIVERSITY (project number BAP-05-
2009-12).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 239
LITERATURE CITED
Adlbauer, K. 1992. Zur Faunistik und Taxonomie der Bockkäferfauna der Türkei II (Coleoptera, Cerambycidae). Entomofauna, 13 (30): 485-509. Alkan, B. 1946. Tarım Entomolojisi. T. C. Tarım Bakanlığı Ankara Yüksek Ziraat Enstitüsü Ders Kitabı 31, Ankara, 232 pp. Alkan, H. 2000. Türkiye orman Cerambycidae (Insecta, Coleoptera)‘lerinin tanıtımı ve Doğu Karadeniz Bölgesindeki türlerin araĢtırılması. Yüksek Lisans Tezi. Karadeniz Teknik Üniversitesi Fen Bilimleri Enstitüsü, Trabzon, 227 pp. Braun, W. 1978. Die Dorcadienausbeute der Forschungsreisen von W. Heinz 1963-1977, Faunistiche Aufstellung, Beschreibung einer neuen Unterart und Bemerkungen zur Systematik wenig bekannter Arten (Coleoptera, Cerambycidae). Nachrichtenblatt der Bayerischen Entomologen, 27 (6): 101-116. Braun, W. 1979. Beitrag zur Kenntnis der Gattung Dorcadion Systematisch neu bewertete Dorcadion-Formen (Col., Cerambycidae). Nachrichtenblatt der Bayerischen Entomologen, 28 (5): 81-86. Çanakçıoğlu, H. 1983. Orman Entomolojisi: Özel bölüm. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 349, Ġstanbul, 535 pp. Çanakçıoğlu, H. 1993. Orman Entomolojisi: Özel bölüm. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 3623, Ġstanbul, 535 pp. Çanakçıoğlu, H. & Mol, T. 1998. Orman Entomolojisi: Zararlı ve yararlı böcekler. Ġstanbul Üniversitesi Orman Fakültesi Yay. No: 451, Ġstanbul, 144-174. Danilevsky, M. L. 2010a. A check-list of Longicorn Beetles (Coleoptera, Cerambycoidea) of Europe. Available from: http://www.cerambycidae.net. Danilevsky, M. L. 2010b. Systematic list of Longicorn Beetles (Cerambycoidea) of the territory of the former USSR. Available from: http://www.cerambycidae.net. Defne, M. Ö. 1954. Batı Karadeniz Bölgesindeki Göknarların Zararlı Böcekleri ve Mücadele Metodları. T. C. Tarım Bakanlığı, Orman Genel Müdürlüğü Yay. No. 105, 60-72. Demelt, C. V. 1963. Beitrag zur Kenntnis der Cerambycidenfauna Kleinasiens und 13. Beitrag zur Biologie palaearkt. Cerambyciden, sowie Beschreibung einer neuen Oberea-Art. Entomologische Blatter, 59 (3) : 132-151. Demelt, C. V. 1967. Nachtrag zur Kenntnis der Cerambyciden-Fauna Kleinasiens. Entomologische Blatter, 63 (2): 106-109. Erdem, R. & Çanakçıoğlu, H. 1977. Türkiye odun zararlıları. Ġstanbul Üniversitesi Orman Fakültesi Yay., Ġstanbul, 113-134. Holzschuh, C. 1995. Beschreibung von 65 neuen Bockkäfer aus Europa und Asien, vorwiegend aus Thailand und China (Coleoptera, Disteniidae und Cerambycidae). FBVA – Berichte; Schriftenreiche der Forstlichen Bundesversuchsanstalt in Wien, 84: 1-63. Lodos, N. 1998. Entomology of Turkey VI (General, Aplied and Faunistic). Ege Ü. Ziraat Fak. Yayınları No: 529, E. Ü. Faculty of Agriculture Press, Ġzmir, 300 pp. Miroshnikov, A. I. 2000a. New longicorn beetles of the tribe Xylosteini from Asia (Coleoptera Cerambycidae). Entomologia Kubanica, 1: 37-54. Miroshnikov, A. I. 2000b. Contribution to the knowledge of the longicorn beetles of the Caucasus. 3. Genus Anaglyptus Mulsant, 1839 (Coleoptera Cerambycidae). Entomologia Kubanica, 1: 61-103. Önalp, B. 1990. Systematic researches on Dorcadion Dalman, 1817 species in Turkey (Coleoptera, Cerambycidae: Lamiinae) I. H. Ü. Eğitim Fakültesi Dergisi, 5: 57-102. Önalp, B. 1991. Systematic researches on Dorcadion Dalman, 1817 species in Turkey (Coleoptera, Cerambycidae: Lamiinae) II. H. Ü. Eğitim Fakültesi Dergisi, 6: 191-227. Öymen, T. 1987. The Forest Cerambycidae of Turkey. Ġ. Ü. Forest Faculty, Ġstanbul, 146 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 240
Özdikmen, H. 2007. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part I - Black Sea Region. Munis Entomology & Zoology 2 (2): 179-422. Özdikmen, H. 2010. The Turkish Dorcadiini with zoogeographical remarks (Coleoptera: Cerambycidae: Lamiinae). Munis Entomology & Zoology, 5 (2): 380-498. Özdikmen, H. & Çağlar, Ü. 2004. Contribution to the knowledge of longhorned bettles (Coleoptera, Cerambycidae) from Turkey, Subfamilies Prioninae, Lepturinae, Spondylidinae and Cerambycinae. J. Ent. Res. Soc., 6 (1): 39-69. Özdikmen, H. & Demir, H. 2006. Notes on longicorn beetles fauna of Turkey (Coleoptera: Cerambycidae). Munis Entomology & Zoology, 1 (1): 157-166. Özdikmen, H. & Hasbenli, A. 2004. Contribution to the knowledge of longhorned bettles (Coleoptera, Cerambycidae) from Turkey, Subfamily Lamiinae. J. Ent. Res. Soc., 6 (2): 25-49. Özdikmen, H., Özdemir, Y. & Turgut, S. 2005. Longhorned Beetles Collection of the Nazife Tuatay Plant Protection Museum, Ankara, Turkey (Coleoptera, Cerambycidae). J. Ent. Res. Soc., 7 (2): 1-33. Özdikmen, H. & Turgut, S. 2010. An overview on the W-Palaearctic subgenus Phytoecia (Helladia) Fairmaire, 1864 with a new subspecies Phytoecia (Helladia) humeralis caneri ssp. n. from Turkey (Coleoptera: Cerambycidae: Lamiinae). Munis Entomology & Zoology, 5 (2): 317-343. Rapuzzi, P. & Sama, G. 1999. Descrizione di due nuovi Chlorophorus di Asia Minore (Coleoptera, Cerambycidae). Lambillionea, 96 (3) : 329-332. Sama, G. 1982. Contributo allo studio dei coleotteri Cerambycidae di Grecia e Asia Minore. Fragmenta Entomologica, Roma, 16 (2): 205-227. Sama, G. 1994. Cerambycidae nuovi o poco noti del Mediterraneo Orientale (Coleoptera, Cerambycidae). Lambillionea 94 (1): 9-13. Sama, G. 1995. Note sui Molorchini. II. I generi Glaphyra Newman, 1840 e Nathrioglaphyra nov.. Lambillionea, 95 (3): 363-390. Sama, G. 1996. Contribution a la connaissance des longicornes de Grece et d'Asie Mineure (Coleoptera, Cerambycidae). Biocosme Mésogéen, Nice, 12 (4): 101-116. Sama, G. 2002. Atlas of the Cerambycidae of Europe and the Mediterranean Area, Volume I, Kabourek, Zlin, 173 pp. Sama, G. 2003. Descrizione di due nuovi Cerambycidae di Turchia: Solaia antonellae n. gen., n. sp. e Helladia demelti n. sp.. Quad. Studi Nat. Romagna, 17 (suppl.): 69-78. Sama, G. & Rapuzzi, P. 1993. Revisione dei generi Saphanus Serville, 1834 e Drymochares Mulsant, 1847. Lambillionea, 93: 278-294. Sekendiz, O. A. 1974. Türkiye hayvansal kavak zararlıları üzerine araĢtırmalar. K. T. Ü. Orman Fakültesi Yayın no: 3, Trabzon, 194 pp. Taglianti, A. V., Audisio, P. A., Biondi, M., Bologna, M. A., Carpaneto, G. M., De Biase, A., Fattorini, S., Piattella, E., Sindaco, R., Venchi, A. & Zapparoli, M. 1999. A proposal for a chorotype classification of the Near East fauna, in the framework of the Western Palaearctic Region. Biogeographia 20: 31-59. Tauzin, P. 2000. Complement a l‘inventaire des Coleopteres Cerambycidae de Turquie. L‘Entomologiste, 56 (4): 151-153. Tozlu, G., Rejzek, M. & Özbek, H. 2002. A contribution to the knowledge of Cerambycidae (Coleoptera) fauna of Turkey. Part I: Subfamilies Prioninae to Cerambycinae. Biocosme Mèsogèen, Nice, 19 (1-2): 55-94. Tozlu, G., Rejzek, M. & Özbek, H. 2003. A Contribution to the Knowledge of Cerambycidae (Coleoptera) Fauna of Turkey Part II : Subfamily Lamiinae. Biocosme Mèsogèen, Nice, 19 (3): 95-110.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 241
STUDIES ON THE CONTACT EFFECT OF SPINOSAD ON ADULT OF TRIBOLIUM CASTANEUM (TENEBRIONIDAE),
SITOPHILUS ORYZAE (CURCULIONIDAE) AND ORYZAEPHILUS SURINAMENSIS (SILVANIDAE)
UNDER LABORATORY CONDITIONS
Golam Reza Sadeghi*, Mojtaba Ghane Jahromi*, Ali Asgar Pourmirza*, Arman Abdolmaleki*
and Asgar Ebadollahi* * Agriculture Faculty, Department of Crop Protection, Urmia University, Urmia, IRAN. E-mail: [email protected] [Sadeghi, G. R., Jahromi, M. G., Pourmirza, A. A., Abdolmaleki, A. & Ebadollahi, A. 2011. Studies on the contact effect of spinosad on adult of Tribolium castaneum (Tenebrionidae), Sitophilus oryzae (Curculionidae) and Oryzaephilus surinamensis (Silvanidae) under laboratory conditions. Munis Entomology & Zoology, 6 (1): 241-245] ABSTRACT: Due to economical importance of stored-products insects and resistance to conventional insecticides, it is necessary to use novel and suitable compounds in control programs. Therefore, we evaluated the toxicity of spinosad to three important storage insects viz, Red flour beetle, Tribolium castaneum (Herbst), Rice weevil, Sitophilus oryzae (L.), and Sawtoothed grain beetle Oryzaephilus surinamensis (L.). One-day old adult insects were exposed to different concentrations of spinosad. The exposure times were 24, 48 and 72 hours in all experiments. Experiments were performed in complete randomized block design with 4 replications. After treatment the samples were held under constant conditions in rearing room at 27±2ºC, 65±5 R.H and 14 D: 10 L photoperiod. In contact tests for T. castaneum the maximum mortality rate was achieved with 500 ppm of spinosad after 72 h exposure. Application of 400 ppm of spinosad after 72 h caused maximum mortality in S. oryzae. For contact treatment the complete control of O. surinamensis was obtained with 500 ppm spinosad at 72 h exposure time. It is possible to recommend 500 ppm of spinosad for complete control of the tested insects through contact application. Estimated LC50 values of spinosad in 72 h after exposure for adults of T. Castaneum, S. oryzae and O. surinamensis were 1287, 279 and 53.17 ppm, respectively. Results showed that O. surinamensis is most susceptible insect to spinosad in these experiments. KEY WORDS: Spinosad, Bioassay, Tribolium Castaneum, Sitophilus oryzae, Oryzaephilus surinamensis.
To protect the agricultural products against insect pests in warehouses, chemical insecticides have been the most effective tool for a long time (Sadeghi & Pourmirza, 2009). To preserve the quantity and quality of stored-product foodstuff particularly cereals it is necessary to decrease the population of the insect pests (Pourmirza & Tajbakhsh, 2001). It is well documented that the number of contact insecticides with lower toxicity to human and environment are very limited (Arthur, 1999; Leesch, 1995). Studies show that the most important and conventionally used contact insecticide to protect wheat against stored-products insects has been malathion (Daglish, 1998), but since 1999 its application in stores is no longer approved (Arthur, 1999). The use of organophosphate was decreased, because of increasing resistance incidence in stored-product insects against these chemicals (Fang et al., 2002). Fumigants have been applied to control stored-products insects for a long time but in recent years the number of usable fumigants has been decreased considerably (Leesch, 1995). Methyl bromide is used as a fumigant to control insects pests on stored

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 242
cereals (United Nations Environmental Programme, 2001). Application of methyl bromide has been largely scaled down because of its carcinogenicity and its effect on depleting ozone layer (Johnson et al., 1998; United Nations Environmental Programme, 2001). Therefore this fumigant will be phase-out in immediate future and could be replaced by new compound such as spinosad (Cisnros et al., 2002). Spinosad is less persistent with formidable insecticidly properties owing low level of toxicity against mammals and birds (Carson & Trumble, 1997; Cisnros et al., 2002; Toews et al., 2003). Moreover, spinosad does not have carcinogenic, mutagenic and tumorigenic effects (Schoonover & Larson, 1995). This compound is also compatible with other integrated pest management program measures (Thompson et al., 1999) and it could be a useful tool in insect control programs (Bessin, 2001).
MATERIALS AND METHODS
Spinosad is a secondary metabolite from aerobic fermentation of Saccharopolyspora spinosa (Mertz & Yao) which is formed on foodstuffs. During this process, spinosad is extracted as a concentrated substance (Anonymous, 1996). By stimulating the nervous system of insects spinosad causes involuntary muscle contraction, insect knock down and severe shakings; the insect is then paralyzed and dies after 4-5 days. These effects are caused by nicotine receptors of acetylcholine. Spinosad also affects GABA (gamma- amino butyric acid) receptors (Salgado, 1998; Salgado et al., 1998; Salgado, 1997). The spinosad used in this study was purchased from American Dow Agro Science Company 48 % SC.
To distribute the insecticides evenly on Petri-dish with the diameter of 5 and height of 1cm surface, cytowet oil was used. Cytowet oil consisted of emulsifier was applied as a 100% pure liquid. Distilled water was used for dilution of spinosad and had the pH value of 6.7. All three insect species were kept in stored-products insects rearing room in Agriculture Faculty of Urmia University at 27±2ºC, 65±5% R.H and 14 D: 10 L photoperiod. The rearing medium for T. castaneum was wheat flour mixed with 5% of yeast (Lale & Yusuf, 2001). Soft Wheat was used for rearing S. oryzae (Padin et al., 2002) and oat used as rearing
medium for O. surinamensus (Tunҫbilek, 1997). Contact bioassay with one-day old insects
After preliminary tests with different concentrations and dose fixing the main tests were performed using 5 concentrations of spinosad and a control group. The concentrations were prepared by diluting of insecticide in distilled water. A drop of cytowet oil which decreased the surface tension was used for even distribution of insecticide on the Petri-dish surface (one drop of cytowet oil was used for a 50 milliliter of solution). All the internal surfaces of Petri-dish were impregnated by 2 milliliter of a given concentration. After all surfaces were impregnated, the surplus liquid was removed and Petri-dishes were left to one hour under ambient conditions. Fifteen one-day old adult insects were introduced to each Petri-dish. The dishes were kept at rearing conditions. Mortality was recorded after 24, 48 and 72 hours and each experiment was replicated four times. After treatment of the Petri-dishes these were dried under ambient conditions. Fifteen one-day old adult insects were introduced to each Petri- dish and Petri- dishes were covered muslin cloth and kept under rearing conditions. Each concentration was replicated four times and for each concentration totally 60 one- day old adult insects of each species were used.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 243
Basic Design and Data Analysis The completely randomized block design through factorial tests was used.
Data transformation were performed as Arcsin x and x whenever needed
and MSTATC program was used for variance analysis. Mortality data from all bioassays were analyzed with SPSS software (SPSS Inc, 1993).
RESULTS AND DISCUSSION
Results of table 1 based on the LC50 values showed that after 24 hours of contact exposure, adults of T. castaneum are more tolerant than two other insects and adults of O. surinamensis are more susceptible than T. castaneum and S. oryzae (T. castaneum> S. oryzae > O. surinamensis). Amounts of RR50 values prove this claim. Results of F values showed that there is no significant difference between the concentrations in the case of T. castaneum, but a significant difference was observed between concentrations that used on S. oryzae in ρ<0.05 and O. surinamensis in ρ< 0.01.
Data from table 2 showed that, after 48 hours since the application of contact exposure, adults of S. oryzae were more tolerant than two other insects and adults of O. surinamensis were more susceptible than T. castaneum and S. oryzae. (S. oryzae > T. castaneum > O. surinamensis). RR50 values prove this claim. The results showed that there is a significant difference between concentrations which were applied against these three species.
Results showed that there is no significant difference between concentrations in the case of T. castaneum, but a significant difference was observed between concentrations which were used against S. oryzae and O. surinamensis. RR50 values in 72h exposure times showed that adults of T. castaneum are more tolerant than two other insects and adults of O. surinamensis are more susceptible than T. castaneum and S. oryzae (T. castaneum> S. oryzae > O. surinamensis) (Table3).
Data from table 4 showed that, F values of time, concentrations and the interaction between concentrations and time periods (Time×Concentration) are significant on the three experimented insects.
With retrospect results of these experiments show that maximum mortality of red flour beetle occurred in the concentration of 500 ppm after 72 h exposure which agree with the results of Toews et al. (2003) and Mcleod et al. (2002) working with some stored-products pest insects. Considering the results from this study the highest mortality of rice weevil adults was obtained in the concentration of 400 ppm after 72 hours which is in agreement with the results of Mcleod et al. (2002) and Fang et al. (2002) which reported a direct relationship between mortality rate of R. dominica and exposure time. The present study showed that 500 ppm concentration of spinosad over 72 hours exposure caused the highest mortality rate of sawtoothed grain beetle. This finding is similiar to the results of Fang et al. (2002) which reported that mortality percentage of R. dominica was increased by increasing the concentration of spinosad. With retrospect it could be concluded that spinosad is merit to be considered as a potential compound in controling the insects in question.
ACKNOWLEDGEMENTS
The authors thank the Faculty of Agriculture, Urmia University, for providing facilities for this research.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 244
LITERATURE CITED Anonymous. 1996. Spinosad technical guide. Dow Elanco (Now Dow AgroSciences LLC), 25. Arthur, F. H. 1999. Evaluation of an encapsulated formulation of cyfluthrin to control Sitophilus oryzae L. on stored wheat. Journal of Stored Products Research, 35: 159-166. Bessin, R. 2001. Colorado potato beetle management. University of Kentucky Entomology. Available in: www .uky.edu/Ag/Entomology/ entfacts /pdfs/ entfa312.pdf. Carson, W. G. & Trumble, J. T. 1997. Effect of insecticides on celery insects. Journal of Arthropod Management Tests, 22: 117. Cisnros, J., Goulson, D., Dewent, C. L., Penagos, I. D., Hernez, O. & Williams, T. 2002. Toxic effects of spinosad on predatory insects. Journal of Biological Control, 23: 156-163. Daglish, G. J. 1998. Efficacy of six grain protectants applied or in combination against three species of Coleoptera. Journal of Stored Products Research, 34: 263-268. Fang, L., Subramayan, B. & Arthur, H. F. 2002. Effectiveness of spinosad on four classes of wheat against five stored insects. Journal of Economic Entomology, 95: 640-650. Johnson, J. A., Vail, P. V., Soderstorm, E. L., Curtis, C. E., Brel, D. G., Tebbets, J. S. & Valero, K. A. 1998. Integration of non chemical post harvest treatments for control of novel orange worm (Lep.: Pyralidae) and Indian meal moth (Lep.: Pyralidae) in walnuts. Journal of Economic Entomology, 91: 1437-1444. Lale, N. E. S. & Yusuf, B. A. 2001. Potential of variety resistance and piper guinease seed oil to control infestation of stored millet seeds and processed product by Tribolium castaneum (Herbst). Journal of Stored Products Research, 37: 63-75. Leesch, J. G. 1995. Fumigant action of acrolein on stored product insects. Journal of Economic Entomology, 88: 326-330. Mcleod, P., Diaz, F. J. & Johnson, D. T. 2002. Toxicity persistence and efficacy of spinosad, chlorfenaptro and thamethoxam on eggplant when applied against the eggplant flea beetle (Coleoptera: Chrysomelidae). Journal of Economic Entomology, 95: 331-335. Padin, S., Dal Bello, G. & Fabrizio, M. 2002. Grain loss closed by Tribolium castaneum, Sitophilus oryzae and Acanthosceloides obtectus in stored durum wheat and beans treated with Beauveria bassiana. Journal of Stored Products Research, 38: 69-74. Pourmirza, A. A. & Tajbakhsh, M. 2001. Studies aceton effect on control stored-product insects. Journal of Agricultural & Techniques Sciences of IRAN, 3: 229-239. Sadeghi, G. R. & Pourmirza, A. A. 2009. Studies on the insecticidal effect of spinosad on the adults of Tribolium castaneum (Herbst), Sitophilus oryzae (L.) and Oryzaephilus surinamensis (L.) under laboratory conditions. Journal of Plant Protection (Agricultural Science and Technology), 22: 13-20. Salgado, V. L. 1997. The mode of action of spinosad and other insect control products. Journal of Down to Earth, 52: 35-44. Salgado, V. L. 1998. Studies on the mode of action of spinosad insect symptoms physiological correlates. Journal of Pesticide Biochemical Physiological, 60: 91-102. Salgado, V. L., Sheets, J. J., Watson, G. B. & Schmidt, A. L. 1998. Studies on the mode of action of spinosad: The internal effective concentration and the concentration dependence of neural excitation. Journal of Pesticide Biochemical and Physiological, 60: 103- 110. Schoonover, J. R. & Larson, L. L. 1995. Laboratory activity of spinosad on non-target beneficial arthropods. Journal of Arthropod Management Tests, 20: 357. SPSS. 1993. SPSS for Windows User‘s Guide Release 6. SPSS Inc. Chicago. Thompson, G. D., Hutchins, S. H. & Sparks, T. C. 1999. Development of spinosad and attributes of a new class of insect control products. Dow AgroScience LLC.9330 Zinosvill Rd., Indianapolis. IN, 46268, USA.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 245
Toews, M. D., Subramayan, B. & Rowan, J. M. 2003. Knockdown and mortality of adults of eight species stored-products beetle exposed to four surfaces treated with spinosad. Journal of Economic Entomology, 96: 1967-1973.
Tunҫbilek, S. A. 1997. Susceptibility of the sawtooth the grain beetle, Oryzaephilus surinamensis (L.), to gamma radiation. Journal of Stored Products Research, 33: 331-334. United Nations Environmental Programme. 2001. Backgrounder basic facts and data on the science and politics of ozone protection. Available in: http: //www.unep.org/ozone or at http: // www.unep.ch/ozone.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 246
GROWTH RESPONSE OF ZONOCERUS VARIEGATUS (LINNAEUS) (ORTHOPTERA: PYRGOMORPHIDAE) NYMPHS
TO FEEDING AT DIFFERENT PERIODS OF THE DAY
Oyebamiji Oyewole Oyegoke*, Akinola John Akintola* and Samuel Adelani Babarinde**
* Department of Pure and Applied Biology, Ladoke Akintola University of Technology, P. M. B. 4000, Ogbomoso, NIGERIA. E-mails: [email protected]; [email protected] ** Department of Agronomy, Ladoke Akintola University of Technology, P. M.B. 4000, Ogbomoso, NIGERIA. E-mail: [email protected] [Oyegoke, O. O., Akintola, A. J. & Babarinde, S. A. 2011. Growth response of Zonocerus variegatus (Linnaeus) (Orthoptera: Pyrgomorphidae) nymphs to feeding at different periods of the day. Munis Entomology & Zoology, 6 (1): 246-250] ABSTRACT: Ninety second instars were used to investigate the growth response of Zonocerus variegatus (L) (Orthoptera: Pyrgomorphidae) nymphs at three feeding periods namely 7.00am-7.00pm, 7.00pm -7.00am and 7.00am-7.00am. Nine cages were constructed and divided into groups of three tagged A, B and C; representing each of the three feeding periods studied. Three replicates per treatment with 10 nymphs per replicate were produced in a completely randomized design. The nymphs were fed with leaves of cassava (Manihot esculenta Cranz). In all the five bodily parameters measured, Z. variegatus nymphs fed better during 7.00am-7.00am period (P< 0.05) than other feeding periods; and this was followed by nymphs that were fed during 7.00am -7.00pm. Consequently, Z. variegatus nymphs recorded higher growth rates in three morphometric and one gravimetric parameter namely mean body length, mean antennal length, mean femur length, and mean body weight during 7.00am-7.00am period. Even, in the mean internotal lenght where significant difference was not shown (P>0.05), higher mean value was still shown by nymphs that were fed during 7.00am-7.00am period. This study has implicated among other factors the role of the length of time of food availability as important in growth response of Z. variegatus nymphs. KEY WORDS: Zonocerus variegatus, nymphs, gravimetric parameters, morphometric parameters.
The variegated grasshopper, Zonocerus variegatus (L.) is a polyphagous and ubiquitous pest of many crops and trees of economic importance in West Africa. Currently, it has become one of the most important grasshopper pests of crops in the humid forests of low altitude and savannas of West and Central Africa (De Grégorio, 1989; Modder, 1984, 1994; Idowu and Modder, 1996). In Nigeria, Z. variegatus has been implicated in the transmission of both okra mosaic virus as well as bacterial burn of cassava (De Visscher, 1990).
Like a typical hemimetabolous insect, the life cycle of Z. variegatus consists of egg, six nymphal instars and the adult stage which is reached by progressive increases in structure and complexity (Chapman et al., 1986; Idowu, 1997; Ademolu, et al., 2006).
Z. variegatus is known to feed as polyphagous insect whose food plants have been found to include a wide range of uncultivated plants, though it exhibits preference (Bernays et al., 1992). Such range of crops on which the insects feed has been confirmed to include coffee and banana as well as a variety of subsistence crops, like cassava, yam and some weed plants (Chapman et al., 1986). Modder (1984) found out that confined nymph of Z. variegatus had the ability to start active feeding in the morning at a temperature as low as 170C with

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 247
extreme massive feeding recorded at 340C. Later in the day (mostly in the evening), they were able to ascend to the top of their food plants to roost. Z. variegatus begins feeding in the morning when the temperature reaches about 260C or 230C with a slowing down at temperature over 350C (Kaufmann, 1965). Some of the primary factors which determine feeding in grasshoppers had been identified; they include time since the previous meal, size and nutritional quality of the meal (Simpson, 1990). This work aims at investigating the effect of feeding periods on some aspects of the growth of Z. variegatus.
MATERIALS AND METHODS
The experiment was carried out between the months of August and September, 2009 in the open field behind the main building of the Department of Pure and Applied Biology, Ladoke Akintola University of Technology (LAUTECH), Ogbomoso, Nigeria.
Cage Construction: Nine wire meshed cages were constructed. Each cage measured 30 x 30 x 45 cm3 in dimension. The topmost part of the cage was not wired but had a replaceable wooden section that served for feed introduction and withdrawal of nymphs during measurements and weighing. Insect Collection: Nymphs of Z. variegatus at the second instar were collected by sweeping net from the premises of the Health Centre of the university. The nymphs were collected into a large paper box measuring 40 x 40 x 50 cm3. The nine wire meshed cages were divided into groups of three with each group representing the three feeding durations under investigation, namely: 7.00am-7.00pm, 7.00pm-7.00am and 7.00am- 7.00am. Nymph introduction and feeding: Ninety second instar nymphs were randomly picked from the large paper box and starved for 24 hours. After starving, the nymphs were divided into three groups of thirty insects with each randomly assigned to three treatments diets in a completely randomized design experiment. Each treatment group was further divided into three replicates of ten nymphs of Z. variegatus per replicate. Thus, 30 nymphs were counted into each of three treatments representing the three feeding periods under investigation viz, 7.00am-7.00pm, 7.00pm-7.00am and 7.00am-7.00am). Nymphs in each treatment were fed with 15g of fleshly plucked cassava, Manihot esculenta (Cranz) leaf, obtained from LAUTECH Teaching and Research Farm, Ogbomoso. The left over feed and accumulated frass after each feeding period were collected before new feed were provided. Data collection The following morphometric and gravimetric parameters were taken at interval of four days for a period of 36 days on the nymphs of each cage/subgroup; after which the data were cumulated: Antenna length, Internodal length, Femur length, Total body length and Body weight. Each of the parameters is defined as follows: Antenna length: The distance of each antenna from the base of scape to the tip of filament. Internodal length: The distance of the thorax lying between the tip of pronotum and base of metanotum. Femur length: The distance between the base of trochanter to the head of tibia.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 248
Total Body length: The distance between the epicranium of the head and tip of the abdomen. Body weight: Weight of an individual nymph expressed in grams.
All length parameters were measured in millimeters. Data collected were subjected to analysis of variance (Steel and Torrie, 1980) and significance between means were separated using Duncan‘s multiple range test at 5 % level of probability.
RESULTS AND DISCUSSION
Tables 1 shows the mean body weight of Z. variegatus nymphs fed with cassava (M. esculenta (Cranz)) leaf during the three feeding periods investigated. It is evident from the table that significant difference (P < 0.05) existed in the body weights of nymphs fed at different periods of the day. In each of the three feeding periods, the mean body weight was in the order 7.00am-7.00am > 7.00am-7.00pm > 7.00pm-7.00am. This implies that Z. variegatus nymphs fed better in the order- A whole day feeding (7.00am-7.00am) > diurnal /daily feeding (7.00am-7.00pm) > Nocturnal feeding (7.00pm-7.00am).
Similarly, significant difference (P < 0.05) was also established in the mean body length of the Z. variegatus nymphs fed at different periods of the day; in this parameter, a growth order similar to the one observed in the mean body weight was obtained, that is, a whole day feeding (7.00am-7.00am) > diurnal /daily feeding (7.00am-7.00pm) > Nocturnal feeding (7.00pm-7.00am).
From Table 1 however, the growth order seen in both the mean body weight and the mean body length was not followed in the mean antennal length of Z. variegatus nymphs fed at different periods of the day. Although, significant difference (P < 0.05) was still evident, the mean antennal length of nymphs measured during 7.00am-7.00am and 7.00am-7.00pm were both higher than those obtained during 7.00pm-7.00am.
Although, no significant difference was observed in the mean femur lengths of Z. variegatus nymphs fed during 7.00am-7.00am and 7.00am-7.00pm, the values of the two periods were however significantly higher than the nymphs fed during 7.00pm-7.00am (nocturnal) feeding. From Tables 1, there were no significant difference in the mean internodal length of Z. variegatus nymphs measured during the three feeding periods (P >0.05). However, the internodal length of nymphs measured during 7.00am-7.00am was still higher than those observed during 7.00am-7.00pm and 7.00pm-7.00am. Consequently, the observation that Z. variegatus nymphs showed better growth response in the order- A whole day feeding (7.00am-7.00am) > diurnal /daily feeding (7.00am-7.00pm) > nocturnal feeding (7.00pm-7.00am) was established in four out of the five parameters determined.
Feeding and foraging in grasshoppers is known to be determined by several demands and constraints. It has been confirmed that grasshoppers‘ demands include such needs like nutrients, minerals, and water (Bernays and Simpson, 1990; Chapman, 1990; Joern and Behmer, 1998). In the like manner, constraints on grasshopper foraging and feeding are known to include food availability, environmental conditions, competition, and predation (Lockwood et al., 1967; Bernays et al., 1997; Rothley et al., 1997; Woods et al., 1997). Although, thermoregulation, along with other factors, has been identified as one of the important determinants of when a grasshopper feeds, the time since the previous meal, size and nutritional quality of the previous meal have also been identified

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 249
(Simpson, 1990). Apart from indicating the usefulness of some parameters as growth indicators in Z. variegatus, this investigation also underlines the period when optimum growth can be obtained in grasshoppers.
CONCLUSION
From the result of this study, it is recommended that when Z. variegatus is to be cultured for laboratory studies, the best feeding period for optimum growth performance is 7.00am-7.00am, which is a whole day feeding. Furthermore, it can be deduced that some bodily parameters are indicative of growth trend in grasshoppers.
LITERATURE CITED Ademolu, K. O., Idowu, A. B. & Dansu, B. M. 2006. Morphometric analysis of Zonocerus variegatus (L.) during post-embryonic development. Proceedings of 2nd International Conference on Science and National Development (Eromosele, I.C. and Popoola, T. O. S. eds.) pp.75-79. College of Natural sciences, University of Agriculture, Abeokuta, Ogun State, Nigeria. Bernays, E. A. & Simpson, S. J. 1990. Nutrition. In Biology of Grasshoppers. Ed. by R. F. Chapman and A. Joern. John Wiley and Sons. New York. Pp. 105-127. Bernays, E. A., Bright, K., Howard, J. J., Raubenheimer, D. & Champagne D. 1992. Variety is the spice of life: frequent switching between foods in the polyphagous grasshopper Taeniopoda eques Burmeister (Orthoptera: Acrididae). Animal Behaviour, 44: 721-731. Bernays, E. A., Angel, J. E. & Augner M. 1997. Foraging by a generalist grasshopper: the distance between food resources influences diet mixing and growth rate (Orthoptera: Acrididae). Journal of Insect Behaviour, 10: 829-840. Chapman, R. F. 1990. Food Selection. In Biology of Grasshoppers. Ed. by R.F. Chapman and A. Joern. John Wiley and Sons. New York. Pp. 39-72. Chapman, R. F., Page, W. W. & MacCaffery, 1986. Bionomics of the variegated grasshopper Zonocerus variegatus in West and Central Africa. Annual Review of Entomology, 31: 479-505. De Grégorio, R. 1989. Liste commentée des travaux consacrés à la morphologie, la biologie, l'éthologie, l'alimentation et le polymorphisme saisonnier du criquet puant, Zonocerus variegatus. II: Alimentation et polymorphisme saisonnier (Orthoptera: Pyrgomorphidae). Bull. Soc. Ent. Fr., 94: 5-6. De Visscher, M. N. 1990. Résultat de l'enquête SAS 1989 sur Zonocerus variegatus (Linne, 1758), Acridien ravageur communément appelé Criquet puant ou Criquet bariole. RIFAS, Montpellier. Idowu, A. B. 1997. The defensive mechanism of Zonocerus variegatus (L). Journal of African. Zoology, 111: 199-203. Idowu, A. B. & Modder, W. W. D. 1996. Possible control of the stinking grasshopper Zonocerus variegatus (L.) (Orthoptera: Pyrgomorphidae) in Ondo state, through human consumption. The Nigerian Field, 61: 7–14. Joern, A. & Behmer, S. T. 1998. Impact of diet quality on demographic attributes in adult grasshoppers and the nitrogen limitation hypothesis. Ecological Entomology, 23: 174-184. Kaufmann, T. 1965. Observation on aggregation, migration and feeding habits of Zonocerus variegatus in Ghana (Orthoptera: Acrididae). Annals of Entomological Society of America, 58: 426-436. Lockwood, J. A., Struttmann, J. M. & Miller, C. J. 1996. Temporal patterns in feeding of grasshoppers (Orthoptera: Acrididae): importance of nocturnal feeding. Environmental Entomology, 25: 570-581. Modder, W. W. D. 1984. The attraction of the weed Chromolaena odorata to grasshopper and associated feeding behaviour. Bulletin of Entomological Research, 74: 239–247.
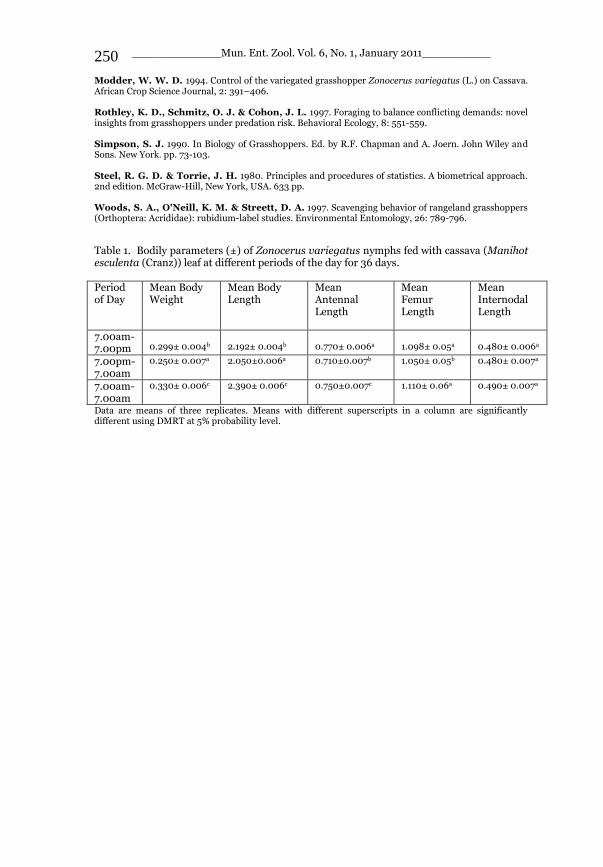
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 250
Modder, W. W. D. 1994. Control of the variegated grasshopper Zonocerus variegatus (L.) on Cassava. African Crop Science Journal, 2: 391–406. Rothley, K. D., Schmitz, O. J. & Cohon, J. L. 1997. Foraging to balance conflicting demands: novel insights from grasshoppers under predation risk. Behavioral Ecology, 8: 551-559. Simpson, S. J. 1990. In Biology of Grasshoppers. Ed. by R.F. Chapman and A. Joern. John Wiley and Sons. New York. pp. 73-103. Steel, R. G. D. & Torrie, J. H. 1980. Principles and procedures of statistics. A biometrical approach. 2nd edition. McGraw-Hill, New York, USA. 633 pp. Woods, S. A., O'Neill, K. M. & Streett, D. A. 1997. Scavenging behavior of rangeland grasshoppers (Orthoptera: Acrididae): rubidium-label studies. Environmental Entomology, 26: 789-796.
Table 1. Bodily parameters (±) of Zonocerus variegatus nymphs fed with cassava (Manihot esculenta (Cranz)) leaf at different periods of the day for 36 days. Period of Day
Mean Body Weight
Mean Body Length
Mean Antennal Length
Mean Femur Length
Mean Internodal Length
7.00am-7.00pm
0.299± 0.004b
2.192± 0.004b
0.770± 0.006a
1.098± 0.05a
0.480± 0.006a
7.00pm-7.00am
0.250± 0.007a 2.050±0.006a 0.710±0.007b 1.050± 0.05b
0.480± 0.007a
7.00am-7.00am
0.330± 0.006c 2.390± 0.006c 0.750±0.007c 1.110± 0.06a 0.490± 0.007a
Data are means of three replicates. Means with different superscripts in a column are significantly different using DMRT at 5% probability level.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 251
FAUNISTIC SURVEY OF THYSANOPTERA IN FARS PROVINCE, IRAN
Majid Fallahzadeh*, Elaheh Azarmi*, Nazila Saghaei**,
Hassan Alemansoor*** and Jalil Alavi****
*Islamic Azad University, Jahrom Branch, Department of Entomology, IRAN. E-mail: [email protected] **Islamic Azad University, Marvdasht Branch, Department of Plant Protection, IRAN. *** Research Center of Agriculture and Natural Resources of Fars province, IRAN. **** Research Center of Agriculture and Natural Resources of North Khorasan province, IRAN. [Fallahzadeh, M., Azarmi, E., Saghei, N., Alemansoor, H. & Alavi, J. 2011. Faunistic survey of Thysanoptera in Fars province, Iran. Munis Entomology & Zoology, 6 (1): 251-261] ABSTRACT: The thrips (Thysanoptera) fauna of Fars province, Iran, was studied during year 2009. A total of 20 species representing 8 genera belonging to 3 families were collected and identified. Aeolothrips balati Pelikán 1958, Aeolothrips citricinctus Bagnall 1933, Aeolothrips modestus zur Strassen 1965 and Thrips trybomi (Karny 1908) are new records for the fauna of Iran. Locality and date of collection, host(s) and distribution data for each species are provided. KEY WORDS: Fars province, Iran, Fauna, New record, Thrips, Thaysanoptera.
The Thrips, or Thysanoptera, is one of the orders of insects distributed through all parts of the world. More than 5500 species of thrips are currently recognised from around the world. The traditional classification of the Order Thysanoptera is adopted, recognising nine Families for living species (plus three fossil families), with two subfamilies in the Phlaeothripidae (the only Family in suborder Tubulifera), and four subfamilies in the Thripidae (one of eight Families comprising suborder Terebrantia) (Mound, 2008).
The thrips in Iran has been a subject of special investigations only in the last few years. The first published records of Iranian Thysanoptera those by Afshar (1938) and several species were subsequently recorded by Davatchi (1949), Salavatian (1959), Farahbakhsh (1961), Shojai (1971), Mortazawiha & Dern (1977), Zahedi (1992), Seyedoleslami & Naderi (1993), Modarres Awal (1994), Cheraghian (1996) and Cheraghian & Hojat (1998). The faunistic and taxonomic papers treated the family in Iran accumulated rapidly through last years (Gilasian, 2000; Javan Moghadam et al., 2000; Mashhadi Jafarlo & Malkeshi, 2000; Moharramipour et al., 2000; Noori et al., 2000; Alavi & zur Strassen, 2002; Etebari & Hesami, 2002; Kalafchi et al., 2002; Minaei et al., 2002; Mojeni, 2002; Moodi, 2002; Saeidi et al., 2002; Teraz & Kheyrandish Koshkoei, 2002; Alavi, 2004; Alemansour & Fallahzadeh, 2004; Jafari & Fallahzadeh, 2004; Jalili Moghadam & Azmayesh Fard, 2004; Khormaly, 2004; Mansouri, et al., 2004; Taghizadeh et al., 2004; Hasani & Fallahzadeh, 2005; Khanjani & Mirab Baluo 2005a,b; Akbarzadeh Shoukat & Shayesteh, 2006; Pirafkan et al., 2006; Alavi et al., 2007; Minaei & Mound, 2008; Minaei & Mound, 2010).
There are 177 nominal species of Thysanoptera reported in Iran, including 132 species of Terebrantia in 47/49 genera, and 45 species of Tubulifera in 15 genera (Bhatti et al., 2009). The objectives of the present study were to provide detailed information on the distribution of Thysanoptera in Fars province, Iran and to

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 252
contribute to the knowledge of the Iranian fauna. It includes general distribution and previously recorded from Iran.
MATERIALS AND METHODS
Thrips specimens were collected at different localities in Fars province, Iran during 2009. The Fars province is located in southern Iran between 27°01′ and 31°51′N and between 50°27′ and 55°45′E, covering an area of 125,000 km2. The climate of the province is arid to semiarid.
The specimens were captured by shaking and beating herbaceous plants, brushes, and branches of trees, shaking leaf litter beneath the trees, and picking flowers of trees and herbaceous plants. The specimens were stored in vials with AGA (60% alcohol 10 parts, glycerol 1 part, acetic acid 1 part) and examined with a microscope after slide preparations with Hoyer‘s medium using the method outlined by Alavi & Kamali (2003). Voucher specimens of all thrips species were deposited in the Insect Collection of the Islamic Azad University, Jahrom Branch.
RESULTS
A total of 20 thrips species representing 8 genera belonging to 3 families were collected from different regions of Fars. The list of species and their distributional data are given below. Terebrantia: Aelothripidae Genus Aeolothrips Haliday, 1836 Aeolothrips balati Pelikán, 1958 Material examined: Shiraz, 1 male, on Bassia scoparia (L.) A. J. Scott (Chenopodiaceae), 22.v.2009. Distribution: Turkey, Bulgaria, Spanish mainland (zur Strassen, 2003), East Palaearctic (Fauna Europaea Web Service, 2004), New record for Iran fauna. Aeolothrips citricinctus Bagnall, 1933 Material examined: Marvdasht, 1 female, 1 male, on Triticum aestivum L. (Poaceae), 31.v.2009. Distribution: Europe (France, Spain, Portugal, Ukraine), North Africa (Tunisia, Algeria, Morocco) on the flower bushes (zur Strassen, 2003; Fauna Europaea Web Service, 2004), New record for Iran fauna. Aeolothrips collaris Priesner, 1919 Material examined: Shiraz, 1 female, 2 male, on Bassia scoparia (L.) A. J. Scott (Chenopodiaceae), 22.v.2009. Distribution: Europe (Albania, Azores, Bulgaria, Canary Islands, Corsica, Croatia, Cyprus, Turkey, Germany, Greek mainland, Italian mainland, Macedonia, Madeira, North Aegean Islands, Portuguese mainland, Russia South, Sardinia, Sicily, Spanish mainland, Ukraine), East Palaearctic, Near East, North Africa, Oriental region on the most of bushes of flower and vegetables (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian Records: Golestan province (Mortazawiha & Dern, 1977; Alavi & zur Strassen, 2002; Alavi, 2004; Alavi et al., 2007), Fars province (Minaei & Alichi, 2000a, b; Minaei et al., 2001), Isfahan province (Ghahhari & Hatami, 2000), Kerman province (Kheyrandish Koshkoei et al., 2000a; Teraz & Kheyrandish Koshkoei, 2002), Khorasan-e-Shomali province (Alavi & Kamali, 2003), Khuzestan province (Cheragian, 1996; Cheraghian & Hojat, 1998; Bagheri & Mosadegh, 2000; Bagheri et al., 2002; Bagheri et al., 2005b).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 253
Aeolothrips intermedius Bagnall, 1934 Material examined: Shiraz, 1 female, on Raphanus sp. (Brassicaceae), 17.iv.2009; Fasa, 1 male, female, on Vicia faba L. (Fabaceae), 15.v.2009. Distribution: Asia, Europe, North Africa, Oriental region on the most of bushes of flower and vegetables (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Tehran, Golestan and Mazandaran provinces (Mortazawiha & Dern, 1977); Khuzestan province (Cheraghian & Hojat, 1998; Bagheri et al., 2005b) Kerman province (Kheyrandish Koshkoei et al., 2000b, Teraz & Kheyrandish Koshkoei, 2002), Fars province (Minaei & Alichi, 2001; Minaei et al., 2001), Tehran and Markazi provinces (Jalili Moghadam & Azmayesh Fard, 2004). Aeolothrips modestus zur Strassen, 1965 Material examined: Shiraz, 1 female, 4 male, on Bassia scoparia (L.) A. J. Scott (Chenopodiaceae), 22.v.2009. Distribution: Canary Islands on flowers (zur Strassen, 2003), New record for Iran fauna. Aeolothrips mongolicus Pelikán, 1985 Material examined: Fasa, 1 male, on Medicago sativa L. (Fabaceae), 15.v.2009; Beyza, 1 male, on Medicago sativa L., 01.vi.2009. Distribution: East Palaearctic (Alavi et al., 2007). Iranian records: Khuzestan province (Cheraghian & Hojat, 1998), Golestan province (Alavi, 2004; Alavi et al., 2007), Khorasan-e-Shomali (Alavi & Kamali, 2003). Aeolothrips tenuicornis Bagnall, 1926 Material examined: Marvdasht, 1 female, 4 male, on Triticum aestivum L. (Poaceae), 31.v.2009. Distribution: Andorra, Britain Island, Canary Islands, French mainland, Germany, Italian mainland, Malta, Portuguese mainland, Selvagens Islands, Sicily, Spanish mainland, Switzerland, The Netherlands, Ukraine, Near East, North Africa on Vitis, Citrus and flower bushes (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Tehran and Qom provinces (Mortazawiha & Dern 1977), Fars province (Minaei & Alichi 2000a, b; Minaei et al., 2001; Minaei et al., 2002), Khuzestan province (Cheraghian & Hojat, 1998; Bagheri et al., 2005b) Golestan province (Alavi et al., 2007). Genus Melanthrips Haliday, 1836 Melanthrips fuscus (Sulzer, 1776) Material examined: Marvdasht, 1 female, on Hirschfeldia incana (L.) Lagr.-Foss. (Brassicaceae), 04.v.2009; 1 male, Shiraz, on Raphanus sp. (Brassicaceae), 17.iv.2009. Distribution: Europe, East Palaearctic, Near East, Nearctic region, North Africa on Cruciferae and the other families of plants (Priesner, 1964b; Fauna Europaea Web Service, 2004). Iranian records: Golestan province (Mortazawiha & Dern, 1977); Khuzestan province (Cheraghian, 1996; Cheraghian & Hojat, 1998), Kerman province (Kheyrandish Koshkoei et al., 2000b, Fars province (Minaei & Alichi 2000a), Khuzestan province (Bagheri et al., 2002) Bagheri et al., 2005b, Tehran and Markazi provinces (Jalili Moghadam & Azmayesh Fard, 2004). Terebrantia: Thripidae Genus Anaphothrips Uzel, 1895 Anaphothrips obscurus (Müller, 1776) Material examined: Fars province: Marvdasht, 1 female, 2 male, on Triticum aestivum L. (Poaceae), 31.v.2009. Distribution: Europe, Australian region, East Palaearctic, Near East, Nearctic region, Neotropical region, North Africa, Oriental region on the Weeds and different kinds of bushes (zur Strassen, 2003; Fauna Europaea Web Service, 2004).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 254
Iranian records: Khorasan-e-Shomali province (Alavi & Kamali, 1995; Alavi & Kamali, 2003), Golestan province (Alavi & zur Strassen, 2002). Anaphothrips sudanensis Trybom, 1911 Material examined: Fars province: Beyza, 1 female, on Allium cepa L. (Liliaceae) 01.vi.2009; 1 male, Fasa, on Triticum aestivum L. (Poaceae), 01.v.2009. Distribution: Europe (Cyprus, Spanish mainland), Afro-tropical region, Australian region, East Palaearctic, Near East, Neotropical region, North Africa, Oriental region on the different species plants as Urochloa maxima (Jacq.) R. Webster (Poaceae), Pennisetum sp. (Poaceae), Sorghum halepense (L.) Pers. (Poaceae), Zea mays L. (Poacrae) (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Khorasan-e-Shomali province (Alavi & Kamali, 1995), Fars province (Minaei & Alichi, 2000a; Minaei et al., 2002), Golestan province (Gilasian et al., 2000; Alavi & zur Strassen, 2002; Alavi et al., 2007), Kerman province (Kheyrandish Koshkoei et al., 2000a; Teraz & Kheyrandish Koshkoei, 2002), Khuzestan province (Behdad, 1996; Cheraghian & Hojat, 1998; Bagheri et al., 2005a). Genus Frankliniella Karny, 1910 Frankliniella tenuicornis (Uzel, 1895) Material examined: Marvdasht, 1 female, on Triticum aestivum L. (Poaceae), 31.v.2009. Distribution: Europe, East Palaearctic, Near East, Nearctic region, North Africa (Palmer et al. 1992; zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Khorasan-e-Shomali province (Alavi & Kamali, 1995; Alavi & Kamali, 2003). Fars province (Minaei & Alichi, 2000a; Minaei et al., 2002), Golestan province (Gilasian et al., 2000; Alavi & zur Strassen, 2002; Alavi, 2004), Isfahan province (Etebari & Hesami, 2002), Marekazi province (Ghotbi et al., 2003), Tehran province (Ghotbi et al., 2003; Jalili Moghadam & Azmayesh Fard, 2004). Genus Limothrips Haliday, 1836 Limothrips angulicornis Jablonowski, 1894 Material examined: Marvdasht, 5 male, on Triticum aestivum L. (Poaceae), 31.v.2009. Distribution: Europe, Australian region, East Palaearctic, Near East, Nearctic region, Neotropical region, North Africa on Hordeum sp. and various species of Poaceae (Priesner, 1964a, zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Khorasan-e-Shomali province (Alavi & Kamali, 1995; Alavi & Kamali, 2003). Golestan province (Gilasian, 2000); Azarbaijan e-Gharbi province (Akbarzadeh Shoukat & Shayesteh, 2006), Kerman province (Kheyrandish Koshkoie et al., 2000a), Khuzestan province (Cheraghian, 1996; Cheraghian & Hojat, 1998). Genus Microcephalothrips Bagnall, 1926 Microcephalothrips abdominalis (D. L. Crawford, 1910) Material examined: Shiraz, 13 female, on Tagetes lucida Cav. (Asteraceae), 16.x.2009. Distribution: Europe (Canary Islands, Italian mainland, Slovenia), Afro-tropical region, Australian region, Near East, Nearctic region, Neotropical region, North Africa, Oriental region on Astereceae and Chenopodiaceae plants (Priesner, 1964a; Mound & Kibby, 1998; Fauna Europaea Web Service, 2004). Iranian records: Khuzestan (Cheraghian & Hojat, 1998; Bagheri et al., 2002), Golestan (Gilasian et al., 2000), Tehran provincs (Jalili Moghadam & Azmayesh Fard, 2004), Markazi province (Ghotbi et al., 2005), Mazandaran province (Minaei et al., 2007). Genus Odontothrips Amyot & Serville, 1843 Odontothrips confusus Priesner, 1926 Material examined: Shiraz, 6 female, 2 male,on Glyssyrhiza glabra L. (Fabaceae), 09.v.2009; Shiraz, 4 female, 2 male, on Sophora alopecuroides L. (Fabaceae), 09.v.2009; Shiraz, 1 male, on Sisymbrium irio L. (Brassicaceae), 28.iii.2009; Maharloo, 1 male, on

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 255
Glyssyrhiza glabra L. (Fabaceae), 15.v.2009; Sepidan, 1 male, on Astragalus sp. (Fabaceae), 25.vii.2009. Distribution: Europe, East Palaearctic, Near East (Fauna Europaea Web Service, 2004). Iranian records: Tehran province (Mortazawiha & Dern, 1977; Pirafkan et al., 2006), Kerman province (Teraz & Kheyrandish Koshkoei, 2002), Lorestan province (Jafari & Fallahzadeh, 2004). Genus Tenothrips Bhatti, 1967 Tenothrips frici (Uzel, 1895) Material examined: Shiraz, 1 female, on Triticum aestivum L. (Poaceae), 22.v.2009. Distribution: Semi-cosmopolitan: Europe, Australian region, East Palaearctic, Near East, Nearctic region, Neotropical region, North Africa, Oriental region (Fauna Europaea Web Service, 2004). Iranian records: Iran (Davatchi, & Taghizadeh, 1954), Golestan province (Gilasian et al., 2000) Fars province (Minaei & Alichi 2000a), Khuzestan province (Bagheri & Alavi, 2006). Genus Thrips Linnaeus, 1758 Thrips meridionalis (Priesner, 1926) Material examined: Shiraz, 3 female, on Raphanus sp. (Brassicaceae), 17.iv.2009; Shiraz, 1 female, on Raphanus sp., 28.iv.2009; Shiraz, 1 female, on Raphanus sp. 26.iii.2009; Shiraz, 1 female, on Lepidium draba L. (Brassicaceae), 01.iv.2009; Shiraz, 1 female, on Ligustrum vulgar L. (Oleaceae), 25.v.2009; Shiraz, 1 female, on Sisymbrium irio L. (Brassicaceae), 28.iii.2009; Shiraz, 1 female, on Sisymbrium irio L., 25.v.2009; Shiraz, 1 female, on Vaccaria hispanica (Mill.) Rauschert (Caryophyllaceae), 17.iv.2009; Shiraz, 1 female, on Malcolmia africana (L.) W. T. Aiton (Brassicaceae), 28.iii.2009; Shiraz, 1 female, on Hirschfeldia incana (L.) Lagr.-Foss. (Brassicaceae), 09.v.2009; Marvdasht, 1 female, on Lepidium draba L. (Brassicaceae), 14.iv.2009; Marvdasht, 1 female, on Brassica napus L. (Brassicaceae), 24.iv.2009; Sepidan, 1 female, on Lepidium draba L. (Brassicaceae), 24.iv.2009; Maharloo, 3 female, on Glyssyrhiza glabra L. (Fabaceae), 15.v.2009. Distribution: Europe, East Palaearctic, Near East on the bushes of flowers as Rosaceae and Fabaceae and truck of trees (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Fars province (Priesner ,1954; Minaei & Alichi, 2000a; Minaei, 2002; Minaei et al., 2002; Alemansour & Fallahzadeh, 2004), Golestan province (Mortazawiha & Dern, 1977; Gilasian et al., 2000, Alavi et al., 2007), Kerman province (Kheyrandish Koshkoei et al., 2000a; Teraz & Kheyrandish Koshkoei, 2002), Khorasan-e-Shomali province (Alavi & Kamali, 2003), Khuzestan province (Cheraghian & Hojat, 1998), Lorestan province (Jafari & Fallahzadeh, 2004), Tehran province (Mortazawiha & Dern, 1977; Jalili Moghadam & Azmayesh Fard, 2004), Yazd province (Mohaghegh & Kheyrandish Koshkoei, 2002). Thrips tabaci Lindeman, 1889 Material examined: Shiraz, 1 female, on Sisymbrium irio L. (Brassicaceae), 28.iii.2009; Shiraz, 4 female, on Raphanus sp. (Brassicaceae), 17.iv.2009; Shiraz, 3 female, on Reseda lutea L. (Resedaceae), 17.iv.2009; Shiraz, 1 female, on Tagetes lucida Cav. (Asteraceae), 10.viii.2009; Marvdasht, 1 female, on Hirschfeldia incana (L.) Lagr.-Foss. (Brassicaceae), 24.iv.2009; Marvdasht, 2 female, on Lepidium draba L. (Brassicaceae), 04.v.2009; Marvdasht, 1 female, on Allium cepa L. (Liliaceae), 31.v.2009. Distribution: Cosmopolitan (zur Strassen, 2003; Fauna Europaea Web Service, 2004). Iranian records: Iran (Afshar, 1938; Salavatian, 1959; Farahbakhsh, 1961; Shojai, 1971; Zahedi, 1992; Modarres Awal, 1994), Azarbaijan-e-Gharbi province (Akbarzadeh Shokat & Rezwani, 1998; Akbarzadeh Shoukat & Shayesteh, 2006), Azarbaijane-Sharghi province (Hassan-Zadeh Salmasi, 1997; Mashhadi Jafarlo & Malkeshi, 2000; Mansouri, et al., 2004; Taghizadeh et al., 2004), Fars province (Javan Moghadam et al., 2000; Noori et al., 2000; Minaei et al., 2002; Alemansour & Fallahzadeh, 2004), Gilan province (Etebari et al., 2000), Golestan province (Gilasian, 2000; Alavi & zur Strassen, 2002; Mojeni, 2002; Alavi, 2004; Khormaly, 2004; Alavi et al., 2007), Hamadan province (Khanjani & Mirab Baluo 2005a, b), Isfahan province (Seyedoleslami & Naderi, 1993; Etebari & Hesami, 2002;

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 256
Kalafchi et al., 2002; Saeidi et al., 2002), Kerman province (Moharramipour et al., 2000; Teraz & Kheyrandish Koshkoei, 2002), Khorasan-e-Janubi province (Moodi, 2002; Shahrokhi, & Rahimi, 2003; Rahimi et al., 2004), Khorasan-e-Razavi province (Noori et al., 2000; Javan Moghadam et al., 2000), Khorasan-e-Shomali province (Alavi & Kamali, 2003), Khuzestan province (Cheraghian, 1996; Cheraghian & Hojat, 1998; Bagheri & Mosadegh, 2000; Bagheri et al., 2002; Bagheri et al., 2005b, c), Lorestan province (Jafari & Fallahzadeh, 2004), Markazi province (Yousefi & Abbasifar, 2004), Mazandaran province (Farahbakhsh, 1961), Tehran province (Davatchi, 1949; Farahbakhsh, 1961; Shojai, 1989; Javan Moghadam et al., 2000; Mirkarimi, 2000; Noori et al., 2000; Hosseininia & Malkeshi, 2004; Jalili Moghadam & Azmayesh Fard, 2004; Khani et al., 2004), Yazd province (Mohaghegh & Kheyrandish Koshkoei, 2002). Thrips trybomi (Karny, 1908) Material examined: Shiraz, 3 male, on Vaccaria hispanica (Mill.) Rauschert (Caryophyllaceae), 17.iv.2009. Distribution: Europe, East Palaearctic on different kind of bushes particularly Tomato's bushes (zur Strassen, 2003; Fauna Europaea Web Service, 2004), New record for Iran fauna. Tubulifera: Phlaeothripidae Genus Haplothrips Amyot & Serville, 1843 Haplothrips flavitibia Williams, 1916 Material examined: Shiraz, 1 female, on Vitis vinifera L. (Vitaceae), 30.v.2009. Distribution: Europe: Britain Island, Germany, Spanish mainland (Mound et al., 1976; Fauna Europaea Web Service, 2004). Iranian records: Kerman province (Kheyrandish Koshkoei et al., 2000a; Teraz & Kheyrandish Koshkoei, 2002), Fars province (Minaei & Alichi 2001, Minaei et al., 2001), Khorasan provine Province (Alavi & Kamali 2003), Khuzestan province (Bagheri & Alavi, 2006), Tehran province (Pirafkan et al., 2006). Haplothrips ganglbaueri Schmutz, 1913 Material examined: Shiraz, 3 female, on Tagetes lucida Cav. (Asteraceae), 15.x.2009. Distribution: South-East Asia on Rice flowers and panicles (Tillekaratn et al., 2007). Iranian records: Iran (Mortazawiha, 1995), Khuzestan province (Cheraghian and Hojat 1998; Bagheri & Alavi, 2006), Fars Province (Minaei and Alichi 2001), Kerman province (Teraz & Kheyrandish Koshkoei, 2002), Khorasan province (Alavi & Kamali 2003), Tehran Province (Pirafkan et al. 2006), Fars and Tehran provinces (Minaei and Mound, 2008).
DISCUSSION
The result of this research and also that of many other researchers, which were conducted on the fauna of Iranian Thysanoptera, indicate that there is a diverse fauna of thrips in Iran. With attention to the prior papers and 4 new records of this paper, the total number of Iranian thrips reaches to 181 species. The number of species probably will increase in the future, because Iran is a large, poorly explored, incorporating various geographical regions, climates, and vegetation zones. We are convinced that additional field work will provide numerous new and interesting records.
ACKNOWLEDGEMENTS
We wish to thank A. Hatami (Fars Research Center for Agriculture and Natural Resources, Iran) for identification of the host plants specimens. We are grateful to Richard zur Strassen (Forschungsinstitut Senckenberg, Frankfurt am Main, Germany), J. S. Bhatti (Department of Zoology, University of Delhi, India),

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 257
L. A. Mound (CSIRO Entomology, Canberra, ACT, Australia) and K. Minaei (Department of Plant Protection, Shiraz University, Iran) for invaluable helps in progress of the project. The research was supported by Islamic Azad University, Jahrom Branch, Fars, Iran.
LITERATURE CITED Afshar, J. 1938. Pests of summer crops, vegetables, industrial plants, and pastures in Iran and their control. General Office of Agriculture, Tehran. 124 pp. [In Persian]. Akbarzadeh Shoukat, G. & Shayesteh, N. 2006. Thrips species found in West Azarbaijan (Orumieh) vineyards, and seasonal abundance of the predominant species (Rubiothrips vitis). Journal of Agricultural Science and Technology, 8: 133-139. Akbarzadeh Shokat, G. & Rezwani, A. 1998. Two new records of thrips from vineyards of West Azarbaidjan. Proceedings of the 13th Iranian Plant Protection Congress, Vol. I, Pests, p. 111. Alavi, J. 2004. Preliminary study on thrips (Thysanoptera) fauna of olive in Golestan province. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 110. Alavi, J. & Kamali, K. 1995. A survey of phytophagous and predaceous Thysanoptera of Bojnourd. Proceedings of the 12th Iranian Plant Protection Congress, p. 340. Alavi, J. & Kamali, K. 2003. The fauna of Thysanoptera in Bojnourd region of Khorasan province, Iran. Thrips, 2: 25-40. Alavi, J. & zur Strassen, R. 2002. Faunistic study of Thysanoptera on soybean in Golestan province. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, 181-182. Alavi, J., zur Strassen, R. & Bagherani, N. 2007. Thrips (Thysanoptera) species associated with wheat and barley in Golestan province, Iran. Journal of Entomological Society of Iran, 27 (1): 1-28. Alemansour, H. & Fallahzadeh, M. 2004. Notes on the Thysanoptera fauna of olive orchards in Fars province. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 103. Bagheri, S. & Alavi, J. 2006. New records of 20 species Thysanoptera on forest and rangeland plants for Iran and Khuzestan province. Proceedings of the 17th Iranian Plant Protection Congress, Vol. I, Pests, p. 78. Bagheri, S., Alavi, J. & Behnamfar, K. 2005a. Collection and identification of 16 species to Thysanoptera, family Thripidae on some medicinal plants in Khuzestan province and introduction of 1 new species for Iran. Abstracts Book of the 2nd Symposium of Medicinal Plants, p. 52. Bagheri, S., Alavi, J. & Mohammadi, R. 2005b. Collection and identification of 8 species predator Thysanoptera on some medicinal plants in Khuzestan province and introduction of 2 new species for Iran. Abstracts Book of the 2nd Symposium of Medicinal Plants, p. 51. Bagheri, S., Alavi, J. & Yousef Naanaie, S. 2005c. Study of Thysanoptera species associated with forest and range plants with report of 2 new species in Khuzestan province. Iranian Journal of Forest and Range Protection Research 2(2), 171-190. [In Persian with English summary]. Bagheri, S. & Mosadegh, M. S. 2000. Natural enemies of Thrips tabaci Lindeman (Thysanoptera: Thripidae) of onion fields in Khuzestan. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 250. Bagheri, S., Mosadegh, M. S. & Kamali, K. 2002. A survey on the biology of Thrips tabaci Lindeman (Thysanoptera: Thripidae) on onions in the north of Khuzestan province, Iran. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 68. Behdad, E. 1996. Iran phytomedicine encyclopaedia, plant Pests and diseases, weeds. Yadbood Publishing. 3153 pp. [In Persian]. Bhatti, J. S., Alavi, J., zur Strassen, R. & Telmadarraiy, Z. 2009.Thysanoptera in Iran 1938–2007. An Overview. Part 1. Thrips, 7: 1–172.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 258
Cheraghian, A. 1996. A faunistic study of Thysanoptera in Ahwaz region. M. Sc. Thesis, College of Agriculture, Shahid-Chamran University, Ahwaz, Iran, 118 pp. [In persian with English summary]. Cheraghian, A. & Hojat, S. H. 1998. A faunistic study of Thysanoptera in Ahwaz region. Proceedings of the 13th Iranian Plant Protection Congress, Vol. I, Pests, p. 211. Davatchi, A. 1949. Important pests of field crops and methods of their control. Bongahe Schimiai, Tehran. 295 pp. [In Persian]. Davatchi, A. 1954. Insect pests in Iran: grasshoppers and other pests of cereals. 1st ed. Tehran University Press. 245 pp. [In Persian]. Davatchi, A. & Taghizadeh, F. 1954. Les insects nuisibles aux aurantiacees en Iran. Entomologie et Phytopathologie Appliquees 14: 1-8 (in Persian), 1-21 (Summary in French). Etebari, K. & Hesami, Sh. 2002. Thysanoptera fauna of grape orchards in Isfahan, Iran. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 182. Etebari, K., Jalali, J. & Tak Shokhan, M. 2000. Thrips (Thysanoptera) fauna of mulberry fields in Gilan province. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 364. Farahbakhsh, Gh. 1961. A checklist of economically important insects and other enemies of plants and agricultural products in Iran. Department of Plant Protection, Tehran. 153 pp. Fauna Europaea Web Service. 2004. Fauna Europaea version 1.1, Available from http://www.faunaeur.org (accessed 20 October 2010). Ghahhari, H. & Hatami, B. 2000. Study on natural enemies of whiteflies (Homoptera: Aleyrodidae) in Isfahan province. Journal of Entomological Society of Iran 20(1), 1- 24. [In Persian with English summary]. Ghotbi, T., Gilasian, E. & Shahraeen, N. 2003. Detection of tospoviruses in individual thrips by ELISA from ornamental plants in Tehran and Markazi provinces. Applied Entomology and Phytopathology, 70: 33-34. Ghotbi, T., Shahraeen, N. & Winter, S. 2005. Occurrence of tospoviruses in ornamental and weed species in Markazi and Tehran provinces in Iran. Plant Disease, 89: 425-429. Gilasian, E. 2000. Faunistic study of Thysanoptera in ornamental and crop plants and the biology of dominant species in Gorgan region. M. Sc. Thesis, College of Agriculture, Tarbiat Modarres University, Tehran, Iran, 148 pp. [In Persian with English summary]. Gilasian, E., Moharramipour, S. & Alavi, J. 2000. One genus and five species of Thysanoptera as new records for Iran fauna. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 341. Hasani, M. R. & Fallahzadeh, M. 2005. Report of Liothrips austriacus (Thysan.: Phlaeothripidae), from Iran. Journal of Entomological Society of Iran, 24: 151-152. Hassan-Zadeh Salmasi, M. 1997. Collecting and identifying insect fauna of onion fields in East Azarbaijan province of Iran. Agricultural Science 6(3-4), 21-42. [In Persian with English summary]. Hosseininia, A. & Malkeshy, S. H. 2004. Comparison of chemical, biological and integrated methods for control of Thrips tabaci Lindeman on carnation under greenhouse conditions. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 300. Jafari, R. & Fallahzadeh, M. 2004. Faunistic study of Thysanoptera on wheat in Lorestan province. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 106. Jalili Moghadam, M. & Azmayesh Fard, P. 2004. Thrips of ornamantal plants in Tehran and Mahallat. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 160. Javan Moghadam, H., Noori, P., Hosseini, S. M. & Amin, Gh. 2000. Control of Thrips tabaci (Lind.) using seed disinfectation method. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 38.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 259
Kalafchi, M., Ebadi, R. & Mobli, M. 2002. A study of population density and damage of onion thrips (Thrips tabaci Lind.) on onion cultivars in Isfahan. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, 68-69. Khani, M., Haghiri, A., Naghdibadi, A. & Yazdani, D. 2004. Notes on the fauna of medicinal plants in Tehran province. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 108. Khanjani, M. & Mirab Baluo, M. 2005a. A study on pests of garlic in Hamadan. Proceedings of National Congress in Sustainable Development of Medicinal Plants, 243-244. Khanjani, M. & Mirab Baluo, M. 2005b. A study on some pests of Bu-Ali Sina medical garden in Hamadan. Proceedings of National Congress in Sustainable Development of Medicinal Plants, 245-246. Kheyrandish Koshkoei, M., Moharramipour, S. & Kamali, K. 2000a. A report on Thysanoptera suborder Terebrantia in Kerman and records of four new species for Iran fauna. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 355. Kheyrandish Koshkoei, M., Moharramipour, S. & Kamali, K. 2000b. A report on Thysanoptera suborder Tubulifera in Kerman and records of three new species for Iran fauna. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 354. Khormaly, S. 2004. Preliminary study on watermelon pests (insects) in Gonbad and Minoodasht. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 297. Mansouri, S. M., Ebadi, R. & Mobli, M. 2004. Comparison of population density and injury of onion thrips (Thrips tabaci Lind.) on selected self and polycross genotypes of onion in Isfahan. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 354. Mashhadi Jafarlo, M. & Malkeshi, S. H. 2000. Survey on efficiency of predator bugs Orius spp. for control of tobacco thrips, Thrips tabaci L. in onion fields in East Azarbaijan province. Proceeding of The Second National Conference on Optimum Utilization of Chemical Fertilizers and Pesticides in Agriculture, Karaj, p. 12. Minaei, K. 2002. The finding of five species of Thysanoptera on oak and hawthorn in forests in Fars province. Proceedings of the 2nd National Plant Protection Conference on Forests and Rangelands, p. 3. Minaei, K. & Alichi, M. 2000a. Faunistic study of the order Thysanoptera in Fars province. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 336. Minaei, K. & Alichi, M. 2000b. The first record of the presence of predatory thrips (Thysanoptera) in Fars province. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 337. Minaei, K. & Alichi, M. 2001. Thrips of the genus Haplothrips (Thys.: Phlaeothripidae) from Shiraz region. Journal of Entomological Society of Iran 20(2), 33-45. [In Persian with English summary]. Minaei, K. & Mound, L. A. 2008. The Thysanoptera Haplothripini (Phlaeothripidae) of Iran. Journal of Natural History, 42: 2617–2658. Minaei, K. & Mound, L. A. 2010. Grass-flower thrips of the genus Chirothrips (Thysanoptera: Thripidae), with a key to species from Iran, Zootaxa 2411: 33–43. Minaei, K., Alichi, M. & Ahmadi, A. A. 2001. The thrips family Aeolothripidae (Thysanoptera: Terebrantia) in the Fars province. Iran Agricultural Research, 20 (1): 53-66. Minaei, K., Alichi, M. & Asadi, Gh. 2002. Introduction to species of Thripidae (Thysanoptera: Thripidae) in Shiraz region. Agricultural Science 12(3), 61-66. [In Persian with English summary]. Minaei, K., Azemayesh Fard, P. & Mound, L. A. 2007. The Thrips genus-group (Thysanoptera: Thripidae) in Iran. Journal of Entomological Society of Iran, 27 (1): 29-36. Mirkarimi, A. 2000. Evaluation of damage of cotton thrips to different cotton varieties in Varamin. Proceeding of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 237. Modarres Awal, M. 1994. List of agricultural pests and their natural enemies in Iran. 1st ed. Ferdowsi University Press. 364 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 260
Mohaghegh, H. & Kheyrandish Koshkoei, M. 2002. The first faunistic study of Thysanoptera in Yazd area. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, 182-183. Moharramipour, S., Kheyrandish Koshkoei, M. & Kamali, K. 2000. Population dynamics of thrips species on rose in Kerman province, Iran. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 146. Mojeni, T. D. 2002. Study on population dynamics of Thrips tabaci L. in the cotton fields of Golestan province. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 45. Moodi, S. 2002. Studies on fauna of Chinese date (Zizyphus jujuba) in Birjand area. Proceeding of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 111. Mortazawiha, A., & Dern, R. 1977. Ein Beitrag zur Thysanopteren fauna des Irans. Entomologie et Phytopathologie Appliquees 45, 8-13. [In German with Persian summary]. Mound, L. A. 2008. Thysanoptera (Thrips) of the World – a checklist. Available from: http://www.ento.csiro.au/thysanoptera/ worldthrips.html (accessed 31 October 2010). Mound, L. A. & Kibby, G. 1998. Thysanoptera: An Identification Guide. CAB International Institute of Entomology and British Museum (Natural History), London. 70 pp. Noori, P., Javan Moghadam, H, Hosseini, S. M. & Amin, G. A. 2000. Population fluctuation of Thrips tabaci Lind. on cotton in cotton growing area of Iran. Proceedings of the 14th Iranian Plant Protection Congress, Vol. I, Pests, p. 36. Pirafkan, S, Kamali, K, Ostovan, H. & Manzari, S. 2006. A faunistic study of Thysanoptera in Pakdasht region. Proceedings of the 17th Iranian Plant Protection Congress, University of Tehran, Karaj; Vol. I. Pests, p. 77. Priesner, H. 1954. On some Thysanoptera from Persia. Annals and Magazine of Natural History, Series 12th, 7 (73): 49-57. Priesner, H. 1964a. Ordnung Thysanoptera (Fransenflügler, Thripse). in Franz H, Bestimmungsbücher zur Bodenfauna Europas, 2: 1-242. Priesner, H. 1964b. A monograph of the Thysanoptera of the Egyptian deserts. Publications de la Institut Desert Egypte, 13: 1-549. Rahimi, H., Moodi, S. & Yazdani, E. 2004. Insect and mite pests and their natural enemies in saffron fields in the south of Khorasan. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 161. Saeidi, Z., Rezvani, A. & Nourbakhsh, S. H. 2002. An investigation on bean thrips.s fauna and evaluation of economic importance of dominant species on native bean variety in Lordegan. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 56. Salavatian, M. 1959. List of pests of cotton, kenaf, flax, sugar beet, sugarcane, tobacco, sesame and castor beans. Entomologie et Phytopathologie Appliques, 18: 62-75. Seyedoleslami, H. & Naderi, A. 1993. Seasonal population fluctuation of important potato sucking insect pests and their natural enemies in the Daran district of Isfahan province. Proceedings of the 11th Plant Protection Congress of Iran, p. 147. Shahrokhi, M. B. & Rahimi, H. 2003. Pests and diseases of Saffron. In: Kafi, M. (Ed.) Saffron production and processing. Center of Excellence for Agronomy, Faculty of Agriculture, Ferdowsi University Press. pp. 137-148 [In Persian]. Shojai, M. 1971. The importance of research on pests of tobacco in Iran. The First Iranian Tobacco and Cigarette Seminar, Rezayeh. Iranian Tobacco Company. pp. 174-182. [In Persian]. Shojai, M. 1989. Entomology (Ethology, Social life and Natural enemies). Vol. 3, 2nd ed. Tehran University Press. 406 pp. [In Persian]. Taghizadeh, M., Hosseini, S. M., Mojeni, T. D. & Amin, G. 2004. Seed disinfectation using some insecticides in the control of onion thrips (Thrips tabaci Lind.) in Moghan area. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 225.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 261
Teraz, A. & Kheyrandish Koshkoei, M. 2002. The first faunistic investigation of Thysanoptera in Jiroft. Proceedings of the 15th Iranian Plant Protection Congress, Vol. I, Pests, p. 184. Yousefi, M. & Abbasifar, A. R. 2004. Evaluation of thrips damage (Thrips tabaci Lindeman) in different onion cultivars in Markazi province. Proceedings of the 16th Iranian Plant Protection Congress, Vol. I, Pests, p. 406. Zahedi, K. 1992. Summer crops and ornamental plants pests and control in Iran. 2nd ed. Iran University Press. 143 pp. [In Persian]. zur Strassen, R. 2003. Die terebranten Thysanopteren Europas und des Mittelmeer-Gebietes. Die Tierwelt Deutschlands 74, 1–277.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 262
EARLY STAGE INDOOR TRAY REARING OF MUGA SILKWORM (ANTHERAEA ASSAMENSIS HELFER)
– A COMPARATIVE STUDY IN RESPECT OF LARVAL CHARACTERS
Himangshu Barman* and Biren Rana
* Central Muga Eri Research and Training Institute, Central Silk Board, Lahdoigarh, Jorhat – 785700, Assam, INDIA. E-mail: [email protected] [Barman, H. & Rana, B. 2011. Early stage indoor tray rearing of Muga Silkworm (Antheraea assamensis Helfer) – a comparative study in respect of larval characters. Munis Entomology & Zoology, 6 (1): 262-267] ABSTRACT: Hatched larvae from 25 dfls of Muga silkworm were simultaneously reared indoor on wooden tray device up to 2nd instar and outdoor providing Som and Soalu leafs as feed throughout one year representing all rearing seasons. Larval weight, larval duration and larval survival were studied in respect of indoor and outdoor rearing on Som-Soalu host plants. Hatched out larvae differ in weight according to seasons highest observed 0.007g per larva during Sept.-Oct. rearing season and lowest being 0.005g per larva during March-April and Nov.-Dec. The larval weight of different instars does not depend upon initial larval weight and size and weight of eggs, depend upon environmental temperature. Larval weight, larval duration and larval survival were found different from each other in Som and Soalu leafs from their indoor and outdoor counterparts. Indoor rearing on detached twigs in wooden tray does not vary mark ably from outdoor rearing on trees. Climatic conditions are the most important factors to be considered in Muga silkworm cultivation regardless of indoor or outdoor rearing. KEY WORDS: Indoor, Outdoor, Rearing, Som, Soalu, Wooden tray, Larvae, Instars, dfls.
Muga silkworm culture is a traditional outdoor rearing practice adopted by people of North Eastern States mainly Assam. Muga silkworm Antheraea assamensis Helfer belongs to Lapidoptera of Saturniidae family and, geographically isolated only to NE region of India. Geographical isolation of this silkworm is indicative of its special requirements for geo-climatic conditions that prevail in this region i.e. high humid temperate climate and forest vegetation of primary and secondary host plants. Thus this species is phylogenetically less adaptive reaching its ecological isolation that is indicative of being on verse of extinction. Although Muga silkworm since time immemorable has been reared for Muga silk still it is purely an outdoor culture in host plant under natural conditions. Only cultural specificity is being managed and took care by Muga rearer. Being exposed to natural environment Muga culture practice encounter lots of problems right from brushing of worms to spinning of cocoons. Outdoor silkworm larvae are invariably expose to nature‘s vagaries such as seasonal climate change, rainfall, strong wind, soaring temperature, besides pests, predators and pathogens inflecting heavy loss particularly in early three instars. Prophylactic measures adopted for pest and disease in outdoor rearing became fruitless due to cross infestation by both pests and pathogens are common in open conditions. In an average in all seasons more than 50% larval loss has been reported by many scientists. Sengupta et al. (1992) reported that during summer more than 50% loss was due to abiotic factors and 80% of the total loss of muga silkworm occurred in second/third instar only. Several workers experimentally practiced indoor rearing of muga silkworm applying different types of rearing devices and, some of them reported effective over outdoor rearing. Singh & Barah

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 263
(1994) conducted partial indoor rearing up to third stages with Som and Soalu twigs in bottle, iron tray and wooden and, reported larval mortality could be reduced marginally as compared to outdoor rearing. Cellular rearing technique developed by Thangavelu & Sahu (1986) for indoor rearing of muga silkworm was found suitable during different seasons for improvement in ERR on Soalu plant, but female cocoon weight and fecundity were found significantly higher on ‗Som‘ plant. Similarly Bhuyan et al. (1991) reported that indoor rearing in iron tray ( 3" x 4" x 4") with water and sand bed covered with slotted cover containing ‗Som‘ twigs showed better ERR (58.8%) as compared to control (51.3%). So, keeping in view of the present constrains faced by muga silkworm cultivation in outdoor conditions, the present comparative studies were undertaken to evaluate wooden tray device in indoor rearing practice of A. assamensis Helfer.
MATERIAL AND METHODS
For the experiments wooden trays of 5'' X 2'' X 3'' with wire mesh (2mm X 2mm size) at either sides with top covering of white cloth fitted with the tray, were taken as indoor rearing device. One Som plant and one Soalu plant of 7-8 years old with plastic net covers were selected from the outdoor garden at a site of complete sunlight for outdoor control rearing. At the same site another host plant stock of same age group were selected to supply leafs for indoor rearing as treatment. Prior to experiments prophylactic measures of disinfestations were given to both outdoor plants and indoor devices.
Twenty five numbers dfls of Antheraea assamensis Helfer were taken from Silkworm Seed Technology Section of the institute. Eggs were kept in perforated brown paper envelop and incubated in B. O. D. incubator at 25°± 1°C. After 6 days of incubation eggs were placed in paper made black box and further incubated for one more day at same temperature. Next the eggs were taken on paper tray and exposed to sunlight. Within two hours the young larvae hatched out prominently making sound. Initial weight of emerged larvae at random was taken by electronic balance. The worms were brushed indoor on detached fresh twigs of Som and Soalu (300 nos. per tray) kept in wooden trays (T). Simultaneously same number of worms was brushed outdoor pre-selected Som and Soalu plants unde3r net cover as control (C). In wooden tray device, perforated polythene sheets and wet foam-pads were used to keep the leaves fresh for longer time. Rearing beds were cleaned once and fresh leafs were given twice a day in the morning and evening hours. During moult, top polythene cover and wet foam pads were removed to keep the bed dry. Just after moult, larval weights were measured and recorded in each instars. Larval duration is counted as number of days required in each instar. After end of each instar, larval survivability was recorded as percentage of living worm. Data of larval weight, larval duration and larval survival of both the treatment and control were recorded in tabulated form and statistically analyzed. The experiments were conducted during five different seasons in one year.
In reaching third instar, all larvae of treatment then transferred and brushed separately on individual pre-selected Som and Soalu trees under net cover in same outdoor garden plot that continued till harvesting.
RESULTS AND DISCUSSION
Results of the experimental data are presented in table-1 and 2. The rearing experiments were carried in different seasons through out the year, e.g. March-April; May-June; July-August; September- October and November-December.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 264
The data in the tables reveals distinct variations in larval weight, larval duration and larval survival in different rearing seasons irrespective of indoor (T) and outdoor (C) rearing. Interestingly, newly hatching out larvae also markedly differs in weight according to season. The heaviest hatching out larvae were found in rearing season of September-October, the weight being recorded at 0.007g and, average lowest weight i.e. 0.005g recorded in rearing seasons March-April and November-December. In the experiment, after first instar highest larval weight in indoor rearing (T) was recorded during March-April in both the cases of Som and Soalu i.e. 0.028 g and 0.033 g respectively. In case of outdoor rearing (C) highest larval weight after first instar was recorded in July-August (0.029g) in Som and, March-April (0.032g) in Soalu. Again, the larval weight after second instar was found highest in July-August in both Som and Soalu leaf feeding tray rearing, being 0.147 g and 0.134 g respectively. Whereas in control (C) after second instar, the same was found 0.173g in Som and 0.170g in Soalu during July-August only. During March-April indoor reared larvae showed enhanced weight over control in both Som and Soalu leafs and also required shorter larval duration than their outdoor counterparts. But their survival in indoor rearing condition is lower than outdoor control. In rearing season May-June, first instar larvae of indoor tray does not exhibit any difference in their weight and duration over outdoor rearing, but lower larval survival was recorded in Soalu. At the end of second instar, larval survival was found slightly lower in both Som and Soalu over control, whereas larval weight was higher in Som and lower in Soalu than their outdoor counterparts. In rearing season July-August larval weight in 1st and 2nd instar were lower in both Som and Soalu than their respective outdoor rearing. In case of larval survival in first instar also except Soalu, lower values were recorded over the control. Larval duration was recorded more or less same in all the cases. No significant difference was observed in larval weight and duration in the 1st instars during rearing period Sept.-Oct. But larval survival was higher in Som over control and lower in Soalu over control. At second instars stage, differences were recorded in all cases except larval duration. Indoor reared larvae exhibited lower value than outdoor except that larval survival was higher i.e. e. 68.0% in Som. During Nov.-Dec, irregular values in respect of larval weight, larval duration in Som and Soalu were recorded. However, higher survival of both first and second instars larvae was recorded over control counterparts of Som and Soalu. During this season, larval survival was all time higher being 97.0% in Som and 92.7% in Soalu at 1st instars and, 89.0% in Som and 84.0% in Soalu at 2nd instars. Data in both the table-1 and table-2 reveals differences in all cases according to food plants (Som & Soalu) of these two rearing conditions.
Thus, it is found from above discussion that indoor rearing of Muga silkworm on detached twigs of Som and Soalu in wooden tray does not differ significantly from outdoor rearing in trees. The former type of rearing prominently exhibited seasonal variations like outdoor rearing. Both the outdoor (C) and indoor (T) rearing were subjected to seasonal climatic changes that mainly included temperature, humidity, leaf moisture, nutritional status of food pant leaves, disease-pest incidence as influencing factors. Das et al. (2004) reported that being multivoltine in nature; Antheraea assamensis Helfer experiences a wide range of temperature (12-37° C) and relative humidity (59-92%) during different climatic seasons of the year. Therefore, it can be inferred that climatic factors are the most influencing factors to be considered in Muga silkworm cultivation regardless of indoor or outdoor rearing. Although there is no significant difference in rearing performances, during unfavorable seasons like very hot climate, very cool and dry climate, indoor rearing may be adopted for Muga silkworm cultivation only by

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 265
developing effectively indoor rearing environment for optimum growth and development. Raja Ram and Sinha (2004) reported that indoor rearing of Muga silkworm on Soalu branches inside perforated polythene bag gave highest ERR during July-August (70.0%) followed by Sept.-Oct. (23.05%) and May-June (15.5%). Talukdar (1999) stated that maintenance of optimum temperature and relative humidity together with disinfestations of rearing room are pre-requisite for good crop harvest.
Further, embryonic tissue growth also differs according to seasonal changes of climate as indicated by variation in weight of newly hatched out larvae in different crop seasons. Hazarika et al. (2004) reported that the size of the egg determines the size and weight of first instar larvae. According to Das et al. (2004) superior egg having 0.0078g weight with 2.8 mm diameter are laid during June and, worst one of 0.0069 weight having 2.0 mm diameter are laid during February. Thus low temperature condition prevailing during November-December and March-April produced hatched larvae of low weight and, high temperature during May to September produced hatched larvae of higher weight as found in our experiment (Table 1).
Since, heaviest larvae were recorded in March-April in first instar and, July-August in second instars in contrast to heaviest newly hatched out larvae during Sept.-Oct., it can be inferred that the larval weight in subsequent instar does not depend upon initial larval weight and hence the size and weight of eggs. Further, as the nutrient compositions in these two host plants differ, it can be very clearly presume the nutrition as the determining factor on larval weight, larval duration and larval survival. Several workers has been worked on indoor rearing of muga silkworm on Som and Soalu host plant and, reported different results, but all found different values in respect of larval weight, larval duration and larval survival in these two host plants. According to Thangavelu et al. (1983) cocoon weight, shell weight, filament length, reel ability and fecundity were higher on Som than Soalu under indoor rearing conditions. Hazarika et al (2004) on the other hand recorded longer larval period and lower cocoon weight but higher shell ratio in indoor wooden box rearing of muga silkworm on Som than outdoor rearing.
ACKNOWLEDGEMENTS
It is great pleasure on part of us to acknowledge ever folding thanks to Dr. R. Chakravorty, then Director, CMERTI, Central Silk Board, M/O Textiles, Govt. of India, Lahdoigarh for his encouragement on indoor rearing of Muga silkworm research and splendid help in successfully completing this work.
LITERATURE CITED Bhuya, N., Borah, B. R., Barah, A. & Sengupta, A. K. 1990-91. Indoor rearing Muga silkworm under specialized conditions for mass rearing. Ann. Rep. RMRS, Boko. pp. 24-26. Das, K., Barah, A., Das. R. & Chakravorty, R. 2004. Oviposition behavior and egg characters of Muga silkworm Antheraea assamensis Helfer (Lepidoptera: Saturniidae) during different seasons. National workshop on Muga silkworm: Biochemistry, Molecular Biology and Biotechnology to improve silk production. RRL, 18-19 Nov. pp. 117-122. Hazarika, L. K., Kataky, A. & Bhuyan, M. 2004. A note on Muga silkworm and indoor rearing of its counterpart. 1st Nat. Semi. On Muga silkworm Biochem., Molecular Biol. & Biotechnology to improve silk production. Org. by RRL, Jorhat, Assam; Nov. 18-19, Abstract, pp 35. Muga Silkworm : Biochemistry, Biotechnology and Molecular Biology. pp. 75-78.
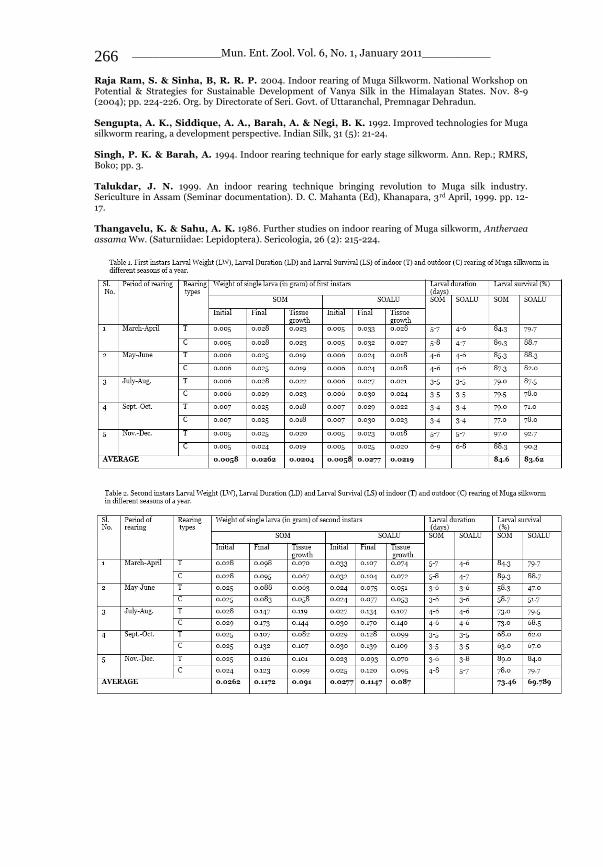
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 266
Raja Ram, S. & Sinha, B, R. R. P. 2004. Indoor rearing of Muga Silkworm. National Workshop on Potential & Strategies for Sustainable Development of Vanya Silk in the Himalayan States. Nov. 8-9 (2004); pp. 224-226. Org. by Directorate of Seri. Govt. of Uttaranchal, Premnagar Dehradun. Sengupta, A. K., Siddique, A. A., Barah, A. & Negi, B. K. 1992. Improved technologies for Muga silkworm rearing, a development perspective. Indian Silk, 31 (5): 21-24. Singh, P. K. & Barah, A. 1994. Indoor rearing technique for early stage silkworm. Ann. Rep.; RMRS, Boko; pp. 3. Talukdar, J. N. 1999. An indoor rearing technique bringing revolution to Muga silk industry. Sericulture in Assam (Seminar documentation). D. C. Mahanta (Ed), Khanapara, 3rd April, 1999. pp. 12-17. Thangavelu, K. & Sahu, A. K. 1986. Further studies on indoor rearing of Muga silkworm, Antheraea assama Ww. (Saturniidae: Lepidoptera). Sericologia, 26 (2): 215-224.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 267

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 268
STUDIES ON SPECIES COMPOSITION OF CARABID BEETLES IN NORTH EASTERN PROVINCES OF IRAN
(COLEOPTERA: CARABIDAE)
Hussein Sadeghi Namaghi*
* Department of Plant Protection, Ferdowsi University of Mashhad, IRAN. E-mail: [email protected] [Namaghi, H. S. 2011. Studies on species composition of carabid beetles in North Eastern provinces of Iran (Coleoptera: Carabidae). Munis Entomology & Zoology, 6 (1): 268-275] ABSTRACT: A preliminary list of 27 carabid species belonging to 6 subfamilies are reported from provinces of Razavi, North and South Khorasan provinces. The list includes 24 records made by the author and those reported in the literature subsequent to 2010. KEY WORDS: Ground beetles, Fauna, Insecta, Khorasan, Iran.
The family Carabidae including ground and tiger beetles, are an incredibly diverse group of insects that can play a major role in agroecosystems. By consuming a variety of insect pests and weed seeds, they can help protect crops from pest damage and associated losses, and decrease costs associated with pest controls. However, in spite of their importance, the knowledge about the fauna of Iranian carabids was limited until recent years. Modarres Awal (1997) reviewing the literature reports only 41 carabid species throughout Iran. Most of them have been reported by foreigner researchers and were collected from a few parts of the country (Hejkal, 2000; Jaeger, 1990; Jaeger, 1992; Lassalle, 2001; Jaskuła, 2007; Magrini & Pavesi, 2003). Recently some faunistic studies (Khajehzadeh, 1998; Alichi & Minaei, 2002a, b; Fallahzadeh et al., 2005; Mohammadzadeh Fard, 2008; Muilwijk& Felix, 2008; Ghahari et al. 2009a, b & Ghahari et al. 2010; Namaghi et al. 2010 and Sadeghi et al. 2011) have added more carabid species to the fauna of Iran. However, many parts of Iran still remained unworked in terms of species composition of this important family of insects. There is no any preliminary checklist of this family in Iran. This study was aimed to determine the species diversity and distribution of ground and tiger beetles in North eastern part of Iran including North, South and Razavi Khorasan provinces, also to present a preliminary list of this insect group for the study area.
MATERIALS AND METHODS
This study was conducted between years 2007 and 2010. Ground beetle specimens were collected from various habitats in Razavi, North and South Khorasan provinces. The collections were made using pitfall traps or collected directly by hand. Addition to these, several other collected specimens by the students have been included. Data, such as the number of specimens, locations and dates were recorded. For identification, part of specimens were sent to Dr. S. Avgin (Osmaniye Korkut Ata University, Turkey) and some were sent to Dr. R. Felix (Berkel Enschot, The Netherlands). The taxonomic status of the species was checked according to the recent update of Fauna Europaea 1.1 (www.faunaeur.org) and Löbl & Smetana (2003).
The voucher specimens are deposited in the department of Plant Protection, College of Agriculture, Ferdowsi University of Mashhad, Iran.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 269
RESULTS
A list of 27carabid species belonging to 6 sub families are reported from provinces of Razavi, North and South Khorasan provinces. The following list includes records made by the author and those reported in the literature subsequent to 2010. I-Subfamily Callistinae Laporte, 1834 Chlaenius festivus (Panzer, 1796) Previous provincial records for Iran: Kohkeyloye and Boyer Ahmad, Khuzestan, Golestan, Razavi Khorasan (Mohammadzadeh Fard& Hodjat, 2008; Shafiei et al., 2004; Ghahari et al. 2009). General distribution: Middle Asia, Central and South Europe, Turkey, Caucasia, Transcaucasia, Iran (Löbl & Smetana, 2003). II-Subfamily Carabinae Latreille, 1802 Tribe Carabini Latreille, 1802 Calosoma (Campalita) maderae tectum Motschulsky, 1844 Materials: 2 specimens, Razavi Khorasan (Mashhad), 27 June 2008, Wheat field; 1 specimen, South Khorasan (Nehbandan), 12 May 2007, Wheat field. Previous provincial records for Iran: This species was previously reported only from Fars, Kohkeyloyeh & Boyer Ahmad, Mazandaran (Mohammad Zadeh Fard & Hodjat, 2008 & Ghahhari et al.2009b). General distribution: Azerbaijan, Armenia, Georgia, Russia: South European Territory, Asia: Iran, Turkey (Löbl & Smetana 2003). Calosoma (Campalita) olivieri Dejean, 1831 Materials: 2 specimens, Razavi Khorasan (Torbat-e-Jam), 23 July 2007, Alfalfa field; 1 specimen, Mashhad, 17 August 2008, Apple orchard. Previous provincial records for Iran: Khorasan, Mazandaran, Golestan (Ghahari et al. 2009a). General distribution: Azores, Malta, North Africa: Algeria, Canary Islands, Egypt, Libya, Morocco (incl. Western Sahara), Tunisia, Asia: Iran, Iraq, Jordan, Pakistan, Saudi Arabia, Syria, Turkmenistan, Uzbekistan (Löbl & Smetana 2003). The range of this species exemplifies a Saharo-Sindian chorotype. Calosoma (Caminara) olivieri Dejean, 1831 Previous provincial records for Iran: Razavi Khorasan, Mazandaran, Golestan , Khuzestan (Ghahari et al. 2009a; Shafiei et al., 2004). General distribution: Russia, Tajikistan, Turkey, Turkmenistan and Uzbekistan (Löbl & Smetana, 2003). III-Subfamily Cicindelinae Latreille, 1802 Tribe CICINDELINI Sloane, 1906 Cicindela asiatica Audouin & Brullé, 1839 Materials: 3 specimens, North Khorasan (Bojnord), 4May 2006, Grasslands. Previous provincial records for Iran: Khorasan (Kashmar), (Ghahari et al., 2009a). General distribution: Russia, Caucasia (Löbl & Smetana, 2003). Cicindela clypeata Fischer von Waldheim, 1821 Materials: 3 specimens, Razavi Khorasan(Mashhad), 1 Sept. 2009. Previous provincial records for Iran: this species has been recorded for Iranian fauna, but the locality is unknown. General distribution: Iran, Afghanistan, Kazakhstan, Uzbekistan, Tajikistan, Turkemmania (Löbl & Smetana, 2003) and Russia South (www.faunaeur.org).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 270
Cicindela germanica (Linnaeus, 1758) Materials: 2 specimens, Razavi Khorasan (Chenaran), 6 May 2009. Previous provincial records for Iran: Northern and Northwestern provinces of Iran (Afshar, 1944). General distribution: Austria, Albania, Andorra, Yugoslavia Incl. Serbia, Kosovo, Voivodina, Montenegro, Ukraine,The Netherlands, Switzerland, Slovenia, Spanish mainland, Slovakia, Sicily Incl. adjacent Italian islands (Lipari Is., Ustica, Egadi Is., Pantelleria I., Pelagie Is.), Russia South, Russia North, Poland, Romania, Moldova, Republic of, Estonia, Macedonia, Luxembourg, Lithuania, Liechtenstein, Latvia, Italian mainland, Ireland, Hungary, Greek main land Incl. Andikithira I., Evvia I., Ionian Is., Samothraki I., Northern Sporades Is., French mainland, Germany, Finland, European Turkey, Czech Republic, Croatia, Bulgaria, Britain Incl. Shetlands, Orkneys, Hebrides and Man Is., Bosnia and Herzegovina, Belgium, East Palaearctic (www.faunaeur.org). Cicindela cf. turkestanicoides Horn, 1938 Materials: 2 specimens, North Khorasn (near Golestan forest), 23June 2007. Previous provincial records for Iran: this species has been recorded from west of Iran, but the locality. According to Dr. Felix (personal communication) record of this species in North eastern parts of Iran is doubtful. General distribution: Asia, Iran (Löbl & Smetana, 2003). IV-Subfamily Harpalinae Bonelli, 1810 Tribe Harpalini Bonelli, 1810 Acinopus (Acinopus) laevigatus laevigatus Ménétriés, 1832 Materials: 2 specimens, Razavi Khorasan(Mashhad), 7 June 2007, Sugar beet field; 2 specimens, South Khorasan (Ferdows), 24 June 2008, Wheat Field surroundings. Previous provincial records for Iran: Fars, Kohkeyloyeh and Boyer Ahmad,Razavi Khorasan, Golestan and Tehran (Mohammad Zadeh Fard & Hodjat, 2008;Ghahhari et al. 2009a: Ghahhari et al. 2009b). General distribution: Azarbaijan, Armenia, Bulgaria, Croatia, Georgia, Greece (incl. Crete), Moldavia, Romania, Russia: South European Territory, Asia: Afghanistan, Gansu (Kansu), Iran, Iraq, Israel, Kyrgyzstan, Kazakhstan, Egypt: Sinai, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Xinjiang (Sinkiang) (Löbl & Smetana 2003). Acinopus (Acinopus) picipes Olivier, 1795 Materials: 3 specimens, Razavi Khorasan(Kashmar), 9 June 2008, Cotton field surroundings. Previous provincial records for Iran: Fars, Razavi Khorasan, Mazandaran (Alichi & Minaie 2002; Namaghi et al. 2010). General distribution: Azarbaijan, Albania, Armenia, Bosnia Herzegovina, Bulgaria, Croatia, France (incl. Corsica, Monaco), Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Malta, Macedonia, Moldavia, Portugal, Romania, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Turkey, Ukraine, Serbia and Montenegro, Cyprus, Iran, Iraq, Israel, Syria, Turkey (Löbl & Smetana, 2003). Dixus clypeatus P. Rossi, 1790 Materials: 3 specimens, Razavi Khorasan (Kashmar), 17 July 2008, grape orchard surroundings. Previous provincial records for Iran: there is no record of this species in Iran. General distribution: Albania, Bosnia Herzegovina, Bulgaria, Croatia, France (incl. Corsica, Monaco), Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Malta, Macedonia, Moldavia, Portugal, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Ukraine, Yugoslavia (Serbia, Montenegro), North Africa: Algeria, Morocco (incl. Western Sahara), Tunisia, Asia: Turkey (Löbl & Smetana 2003). Harpalus (Harpalus) distinguendus distinguendus Duftschmid, 1812 Materials: 2 specimens, Razavi Khorasan (Mashhad), 15 April 2007, Wheat field; Neyshabor , 1 specimen, 25April 2007, Sugar beet field; Torbat-e- Heydariye , 2 specimens,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 271
10 July 2008, Alfalfa field; Torbat-e- Jam, 1 specimen, 3 August 2008, Alfalfa field; Kashmar, 3 specimens, 22 July 2008, Cotton field; 1 specimen, South Khorasan (Ghaenat), 7 June 2007, Sugar beet field. Previous provincial records for Iran: This species has already been reported from Iran , but the locality is unknown (Löbl & Smetana 2003). General distribution: Azarbaijan, Albania, Armenia, Austria, Azores, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Croatia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France (incl. Corsica, Monaco), Germany, Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Lithuania, Luxembourg, Macedonia, Moldavia, The Netherlands, Norway, Russia: North European Territory, Poland, Portugal, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Sweden, Switzerland, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), North Africa: Algeria, Canary Islands, Morocco (incl.Western Sahara), Madeira Archipelago, Asia: Afghanistan, Cyprus, Russia: East Siberia, Iran, Israel, Kyrgyzstan, Kazakhstan, Mongolia, Syria, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Russia: west Siberia, Xinjiang (Sinkiang) (Löbl & Smetana, 2003). Harpalus (Harpalus) honestus honestus Duftschmid, 1812 Materials: 3 specimens, Razavi Khorasan (Chenaran), 18 June 2008, sugarbeet field. Previous provincial records for Iran: Golestan & Razavi Khorasan (Ghahhari et al., 2009a). General distribution: Azarbaijan, Albania, Armenia, Austria, Belgium, Bosnia Herzegovina, Bulgaria, Croatia, Czech Republic, France (incl. Corsica, Monaco), Great Britain (incl. Channel Islands), Germany, Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Liechtenstein, Luxembourg, Macedonia, Netherlands, Poland, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Switzerland, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), Asia: Iran, Syria, Turkey. (Löbl & Smetana, 2003). Harpalus (Harpalus) oblitus oblitus Dejean, 1829 Materials: 2 specimens, Razavi Khorasan(Mashhad), 5 May 2008, Wheat field; 1 specimen, Sabzevar, 5 May 2008, Wheat field; 2 specimens, South Khorasan(Birjand), 6 May 2007, Alfalfa field. Previous provincial records for Iran: This species was already reported from Iran (Löbl & Smetana, 2003), but locality is not given. General distribution: Albania, Bosnia Herzegovina, Bulgaria, Croatia, France (incl. Corsica, Monaco), Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Moldavia, Romania, Slovenia, Russia: South European Territory, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), Asia: Iran, Iraq, Kazakhstan, Syria, Tadzhikistan, Turkey, Russia: west Siberia (Löbl & Smetana, 2003). Harpalus (Harpalus) smaragdinus Duftschmid, 1812 Materials: 3 specimens, South Khorasan (Ferdows), 29 April 2008, Wheat Field. Previous provincial records for Iran: This species was already reported from Iran (Löbl & Smetana 2003), but locality is not given. General distribution: Azarbaijan, Albania, Armenia, Austria, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Croatia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France (incl. Corsica, Monaco), Great Britain (incl. Channel Islands), Germany, Georgia, Hungary, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Lithuania, Luxembourg, Macedonia, Moldavia, The Netherlands, Norway, Russia: North European Territory, Poland, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Sweden, Switzerland, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), Asia: Cyprus, Russia: East Siberia, Iran, Kyrgyzstan, Kazakhstan, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Russia: west Siberia, (Xinjiang) Sinkiang (Löbl & Smetana, 2003). Harpalus (Pseudoophonus) griseus Panzer, 1796 Materials: 2 specimens, Razavi Khorasan (Bardaksan), 19 August 2008, Alfalfa field; 3 specimens, Mashhad, 9 June 2007, Sugar beet field; 2 specimens, Dargaz , 17 July 2007, Wheat Field Surroundings; 2 specimens, Serakhs, 28 April 2008, wheat field; 1 specimen,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 272
South Khorasan (Ghaenat), 9 July 2007, Alfalfa field; 2 specimens, Ferdows, 22 June 2008, Wheat field. Previous provincial records for Iran: Razavi Khorasan & Mazandaran (Ghahhari et al., 2009a; Ghahhari et al., 2009b). General distribution: Azarbaijan, Armenia, Austria, Azores, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Croatia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France (incl. Corsica, Monaco), Great Britain (incl. Channel Islands), Germany, Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Malta, Macedonia, Moldavia, The Netherlands, Norway, Poland, Portugal, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Sweden, Switzerland, Ukraine, Yugoslavia (Serbia, Montenegro), North Africa: Algeria, Egypt, Morocco (incl.Western Sahara), Tunisia, Asia: Afghanistan, Cyprus, Russia: East Siberia, Russia: Far East, Gansu (Kansu), Heilongjiang (Heilungkiang), Iran, Iraq, Israel, Japan, Jiangsu (Kiangsu), Jilin (Kirin), Kyrgyzstan, Kazakhstan, Liaoning, North Korea, South Korea, Shaanxi (Shensi), Shanghai, Shandong (Shantung), Shanxi (Shansi), Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Russia: west Siberia, Xinjiang (Sinkiang), Yunnan, Zhejiang (Chekiang), Oriental Region (Löbl & Smetana, 2003). Harpalus (Pseudoophonus) rufipes DeGeer, 1774 Materials: 1 specimen, Razavi Khorasan (Mashhad), 28 August 2007, Alfalfa field; 2 specimens, Neyshabor, 12 May 2008, Wheat field surroundings. Previous provincial records for Iran: Fars, Kohkeyloyeh and Boyer Ahmad, Tehran, Ardabil, Mazandaran, Semnan & Razavi Khorasan (Mohammad Zadeh Fard & Hodjat, 2008;Ghahhari et al., 2009a). General distribution: Azarbaijan, Albania, Armenia, Austria, Azores, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Croatia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France (incl. Corsica, Monaco), Great Britain (incl. Channel Islands), Germany, Georgia, Greece (incl. Crete), Hungary, Ireland, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Malta, Macedonia, Moldavia, The Netherlands, Norway, Poland, Portugal, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Sweden, Switzerland, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), North Africa: Algeria, Morocco (incl.Western Sahara), Tunisia, Asia: Afghanistan, Cyprus, Russia: East Siberia, Iran, Iraq, Kyrgyzstan, Kazakhstan, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Russia: west Siberia, Xinjiang (Sinkiang), Nearctic Region (Löbl & Smetana, 2003). Tribe Lebiini Bonelli, 1810 Cymindis (Cymindis) andreae Ménétriés, 1832 Materials: 2 specimens, Razavi Khorasan (Mashhad), 12 May 2007, Alfalfa field. Previous provincial records for Iran: Fars, Mazandaran, Kohkihkeyloye and Boyer Ahmad and Khuzestan (Mohammadzadeh Fard & Hodjat, 2008; Shafiei et al., 2004; Ghahhari et al., 2009b). General distribution: Armenia, Azarbaijan, Georgia, Russia: South European Territory, North Africa: Egypt, Asia: Afghanistan, Cyprus, Iran, Iraq, Kyrgyzstan, Kazakhstan, Lebanon, Pakistan, Saudi Arabia, Tadzhikistan, Turkmenistan, Turkey, Yemen (incl. Socotra) (Löbl & Smetana, 2003). Tribe Pterostichini Bonelli, 1810 Pterostichus (Melanius) elongatus Duftschmid, 1812 Materials: 2 specimens, South Khorasan (Ghaenat), 27 June 2007, wheat field. Previous provincial records for Iran: there is no previous record of this species in Iran. Geographical distribution: it is distributed in Europe: Albania, Austria, Bosnia Herzegovina, Bulgaria, Croatia, Czech Republic, France (incl. Corsica, Monaco), Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Moldavia, Romania, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Ukraine, Yugoslavia (Serbia,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 273
Montenegro), North Africa: Morocco (incl.Western Sahara), Asia: Afghanistan, Kazakhstan, Turkey (Löbl & Smetana, 2003). Poecilus (Poecilus) cupreus cupreus Linné, 1758 Materials: 2 specimens, Razavi Khorasan (Chenaran), 28 April 2008, Apple orchard surroundings. Previous provincial records for Iran: Razavi Khorasan, Mazandaran, Fars, Ardabil, Tehran, Semnan (Ghahhari et al., 2009a,b). General distribution: Azarbaijan, Albania, Armenia, Austria, Belgium, Bosnia Herzegovina, Bulgaria, Byelorussia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, Finland, France (incl. Corsica, Monaco), Great Britain (incl. Channel Islands), Germany, Georgia, Greece (incl. Crete), Hungary, Ireland, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Liechtenstein, Lithuania, Moldavia, The Netherlands, Norway, Russia: North European Territory, Poland, Romania, Slovakia, Spain (incl. Gibraltar), Russia: South European Territory, Sweden, Switzerland, Turkey, Ukraine, Yugoslavia (Serbia, Montenegro), Asia: Russia: East Siberia, Kyrgyzstan, Kazakhstan, Syria, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Russia: west Siberia (Löbl & Smetana, 2003). Poecilus (Poecilus) punctulatus Schaller, 1783 Materials: 2 specimens, Razavi Khorasan (Chenaran), 24 July 2008, Wheat field. Previous provincial records for Iran: ther is no previous record of this species in Iran. General distribution: Austria, Belgium, Bulgaria, Byelorussia, Russia: Central European Territory, Czech Republic, Denmark, Estonia, France (incl. Corsica, Monaco), Germany, Hungary, Italy (incl. Sardinia, Sicily, San Marino), Kazakhstan, Latvia, Lithuania, Moldavia, The Netherlands, Russia: North European Territory, Poland, Slovakia, Russia: South European Territory, Sweden, Switzerland, Ukraine, Asia: Kazakhstan, Uzbekistan, Russia: west Siberia (Löbl & Smetana, 2003). Poecilus (Poecilus) subcoeruleus Quensel, 1806 Materials: 3 specimens, Razavi Khorasan (Mashhad), 18 May 2007, Alfalfa field. Previous provincial records for Iran: This species was already reported from Iran (Löbl & Smetana 2003), but locality is not given. General distribution: Bulgaria, Byelorussia, Moldavia, Portugal, Russia: South European Territory, Ukraine, Asia: Iran (Löbl & Smetana, 2003). Tribe Zabrini Bonelli, 1810 Amara (Amara) proxima Putzeys, 1866 Materials: 2 specimens, Razavi Khorasan (Mashhad), 16 August 2007, Sugar beet field. Previous provincial records for Iran: The occurrence of this species in Iran has already been reported (Löbl & Smetana, 2003), but collection sites are not given. General distribution: Azarbaijan, Albania, Armenia, Austria, Bosnia Herzegovina, Bulgaria, Croatia, France (incl. Corsica, Monaco), Germany, Georgia, Greece (incl. Crete), Hungary, Italy (incl. Sardinia, Sicily, San Marino), Macedonia, Romania, Switzerland, Slovakia, Slovenia, Spain (incl. Gibraltar), Russia: South European Territory, Ukraine, Yugoslavia (Serbia, Montenegro), Asia: Iran, Turkmenistan, Turkey (Löbl & Smetana 2003). Amara aenae De Geer, 1774 Previous provincial records for Iran: Fars, Kohkeyloye Boyer Ahmad, Golestan, Khorasan (Mohammadzadeh & Hodjat, 2008; Ghahari et al., 2009a). General distribution: General distribution: Palearctic Region, North America, Caucasia (Löbl & Smetana, 2003). Anthia (Anthia) mannerheimii Chaudoir, 1842 Materials: 3 specimens, Razavi Khorasan (Mashhad), 10 August 2008. Previous provincial records for Iran: Khorasan (Afshar, 1944). General distribution: Iran, Turkmenistan (Löbl & Smetana, 2003).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 274
V-Subfamily Nebriinae Laporte, 1834 Tribe Nebriini Laporte, 1834 Nebria (Nebria) hemprichi Klug, 1832 Materials: 2 specimens, Razavi Khorasan(Fariman), 11 July 2007, Sugarbeet field. Previous provincial records for Iran: Razavi Khorasan, Golestan, Mazandaran (Ghahhari et al., 2009a,b). General distribution: Asia: Israel, Syria, Turkey (Löbl & Smetana, 2003). VI-Subfamily Scaritinae Bonelli, 1810 Tribe Scaritini Bonelli, 1810 Distichus (Distichus) planus Bonelli, 1813 Materials: 2 specimens, Razavi Khorasan(Mah-Velat), 15 May 2007, Alfalfa field. Previous provincial records for Iran T: Tehran and Khorasan (Ghahhari et al., 2009a). General distribution: Azerbaijan, France (incl. Corsica, Monaco), Georgia, Greece (incl. Crete), Italy (incl. Sardinia, Sicily, San Marino), Malta, Portugal, Spain (incl. Gibraltar), Turkey, North Africa: Egypt, Morocco (incl. Western Sahara), Tunisia, Asia: Iran, Iraq, Jordan, Kazakhstan, Pakistan, Saudi Arabia, Syria, Tadzhikistan, Turkmenistan, Turkey, Uzbekistan, Yemen (incl. Socotra) (Löbl & Smetana, 2003).
ACKNOWLEDGEMENTS
The author is grateful to Drs R. Felix (The Netherlands) and S.S. Avgin for their kind help with confirmation or identification of some specimens. The study (project no. p-308) financially supported by Ferdowsi University of Mashhad who is acknowledged.
LITERATURE CITED Anonymous. Data base www.faunaeur.org. Version 1.1, visited on: 15. IV. 2010. Afshar, J. 1944. Les nomes scientifique de quelques coleopteres de l'Iran et leurs importance en agriculture. Min. Agri. Tehran. 22 pp. Alichi, M. & Minaei, K. 2002a. New records on the fauna of the Carabidae in Fars province, 175-176. Proceedings of the 15th Iranian Plant Protection Congress, Razi University, Kermanshah. Alichi, M. & Minaei, K. 2002b. Study on distribution of the beetles belonging the family Carabidae in Shiraz region, 175. Proceedings of the 15th Iranian Plant Protection Congress, Razi University, Kermanshah. Fallahzadeh, M., Shojace, M. & Ostovan, H. 2005. Report of Broscus punctatus (Col.: Carabidae) from Iran. Journal of Entomological Society of Iran, 24 (2): 140. Ghahari, H., Kesdek, M., Samin, N., Ostovan, H., Havaskary, M. & Imani, S. 2009a. Ground beetles (Coleoptera:Carabidae) of Iranian cotton fields and surrounding grasslands. Munis Entomology & Zoology, 4 (2): 436-450. Ghahari, H., Wojciech, B., Czkowsi, J., Kesdek, M., Otovan, H. & Tabari, M. 2009b. Ground beetles (Coleoptera: Carabidae) from rice fields and surrounding grasslands of Northern Iran. Journal of Biological Control, 23 (2): 105-109. Ghahari, H., Avgin, S. S. & Ostovan, H. 2010. Carabid beetles (Coleoptera: Carabidae) collected from different ecosystems in Iran with new records. Türk. entomol. derg., 34 (2): 179-195. Hejkal, J. 2000. Amara (Amara) elborzensis sp. n. (Coleoptera: Carabidae) from Iran. Klapalekiana, 36 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 275
Jaeger, B. 1990. Zur Verbreitung von Bradycellus csikii Laczo, 1912 und B. distinctus Dejean, 1829 sowie Beschreibung von Bradycellus heinzei n.sp. aus demNord-Iran (Col., Carabidae). Entomologische Nachrichten und Berichte, 34: 9-13. Jaeger, B. 1992. Beitrag zur Erforschung der Acupalpus-Fauna des Iran und der Türkei nebst Beschreibung von Acupalpus turcicus n. sp. (Col., Carabidae). Entomologische Nachrichten und Berichte, 36: 223-230. Jaskuła, R. 2007. First record of Lebia trimaculata from Iran (Coleoptera: Carabidae). Entomological Problems, 37 (1-2): 48. Khajehzadeh, Y. 1998. Investigation on the most important injurious beetles of cereal fields of Khuzestan, the Proceedings of 13th Iranian Plant Protection Congress. Lassalle, B. 2001. Les Sphodristocarabus d‘Iran (Coleoptera, Carabidae). Le Coléoptériste, 43: 169-170. Löbl, I. & Smetana, A. 2003. Catalogue of Palaearctic Coleoptera. Volume I. Archostemata-Myxophaga-Adephaga. ApolloBooks. Stenstrup, Denmark, 819 pp. Magrini, P. & Pavesi, M. 2003. A new Iranian species of Neoblemus (Coleoptera: Carabidae). Bollettino della Societa Entomologica Italiana, 135 (2): 71-78. Modarres Awal, M. 1997. List of agricultural pests and their natural enemies in Iran. Revised ed. Ferdowsi University of Mashhad, Iran, no. 429 pp. Mohammadzadeh Fard, S. 2008. Description of female Axonya farsica (Coleoptera: arabidae: Broscinae). Journal of Entomological Society of Iran, 27 (2), Supplement, 3-4. Mohammadzadeh Fard, S. & Hodjat, S. H. 2005. New reocrds of Carabid beetles in Iran. Journal of Entomological Society of Iran, 24 (2). Mohammadzadeh Fard, S., Hodjat, S. H. 2008. A comparison survey on fauna of Family Carabidae ( Insecta: Coleoptera) in natural and agricultural areas. 18th Iranian Plant Protection Congress, Bu-Ali Sina University, Hamadan. p. 89. Muilwijk, J. & Felix, R. 2008. Description of three new species of the tribe Trechini (Col.: Carabidae) from south Iran. J. Entomo. Soc. Iran, 28 (1): 79-85. Namaghi, S. H., Avgin, S. S. & Farahi, S. 2010. New Data ToThe Knowledge Of Ground Beetles (Coleoptera: Carabidae) Fauna of Iran. Journal of Turkish Entomology, 34 (2): 197-210. Sadeghi, H., Hasani Saadi, S. & Felix, R. 2011. Ground and tiger beetles (Coleoptera: Carabidae) from Kerman and Khorasan provinces of Iran. Munis Entomology & Zoology (In Press ).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 276
A SYNOPSIS OF TURKISH XYLOTRECHUS CHEVROLAT, 1860 WITH A NEW RECORD, XYLOTRECHUS STEBBINGI GAHAN,
1906 (COLEOPTERA: CERAMBYCIDAE: CERAMBYCINAE)
Hüseyin Özdikmen* and Serdar Tezcan** * Gazi Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü, 06500 Ankara / Türkiye. E-mails: [email protected] and [email protected] ** Plant Protection Department, Faculty of Agriculture, Ege University, 35100 Bornova, Ġzmir, TURKEY, e-mail: [email protected] [Özdikmen, H. & Tezcan, S. 2011. A synopsis of Turkish Xylotrechus Chevrolat, 1860 with a new record, Xylotrechus stebbingi Gahan, 1906 (Coleoptera: Cerambycidae: Cerambycinae). Munis Entomology & Zoology, 6 (1): 276-281] ABSTRACT: All Turkish taxa of the genus Xylotrechus Chevrolat, 1860 are evaluated and summarized. A new record, Xylotrechus stebbingi Gahan, 1906 from Turkey is also given in the present text. KEY WORDS: Xylotrechus, Clytini, Cerambycinae, Cerambycidae, Coleoptera, Turkey.
The main aim of this work is to clarify current status of the genus Xylotrechus Chevrolat, 1860 in Turkey.
Subfamily CERAMBYCINAE Latreille, 1802 Tribe CLYTINI Mulsant, 1839 Type genus: Clytus Laicharting, 1784 Genus XYLOTRECHUS Chevrolat, 1860 Type species: Clytus sartorii Chevrolat, 1860
According to Sama & Löbl in Löbl & Smetana (2010), the genus Xylotrechus Chevrolat, 1860 is represented by eighty-five species of four subgenera in Palaearctic Region. The subgenus Kostiniclytus Danilevsky, 2009 (type sp.: Xylotrechus zaisanicus Plavilstshikov, 1940) has three species that are distributed in Kazakhstan and Mongolia. So, it is not represented in Turkey. The subgenus Ootora Niisato & Wakejima, 2008 (type sp.: Clytus villioni Villard, 1892) has four species that are distributed in China, Japan, Far East Russia and Oriental Region. So, it is not represented in Turkey. The subgenus Xyloclytus Reitter, 1913 (type sp. : Clytus chinensis Chevrolat, 1852) has three species that are distributed from European Russia to Kazakhstan, Mongolia, Korea, China, Japan, East Siberia and Far East Russia. So, it also is not represented in Turkey. The nominotypical subgenus Xylotrechus Chevrolat, 1860 (type sp. : Clytus sartorii Chevrolat, 1860) is only represented in Turkey. The subgenus has seventy-five species in Palaearctic Region. These species are distributed from W Europe and Mediterranean Region to East Palaearctic Region and Oriental Region. They occur mostly in East Palaearctic Region. Thirteen species are distributed also in Oriental Region. The genus has over 180 species in the World fauna now.
In Turkey, the genus Xylotrechus has been represented only by two species of the nominotypical subgenus as X. antilope (Schoenherr, 1817) and X. arvicola (Olivier, 1795) until now. X. stebbingi Gahan, 1906 is recorded for the first time from Turkey with the present work.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 277
The species X. capricornus (Gebler, 1830) and X. ibex (Gebler, 1825) have been reported only by Lodos (1998) for Turkey in his unrealistic list without any exact locality. According to known distribution areas of these species, they are impossible for Turkey. The known distribution areas are Austria, Poland, Slovakia, Ukraine, European Russia, European Kazakhstan, West Siberia for X. Capricornus and Finland, Poland, European Russia, West and East Siberia, Far East Russia, Kazakhstan, Mongolia, China for X. ibex. So, they are not confirmed for Turkey.
Moreover, according to Sama & Löbl in Löbl & Smetana (2010), Xylotrechus rusticus (Linnaeus, 1758) and Xylotrechus sieversi (Ganglbauer, 1890) that are represented in Turkey, were placed in the genera Rusticoclytus Vives, 1977 and Turanoclytus Sama, 1994 respectively. So, Rusticoclytus rusticus (Linnaeus, 1758) and Turanoclytus sieversi (Ganglbauer, 1890) are not evaluated in the present paper.
Consequently, the Turkish Xylotrechus comprises of only three species. They are presented as follows: Subgenus XYLOTRECHUS Chevrolat, 1860 Type species: Clytus sartorii Chevrolat, 1860
Xylotrechus antilope (Schoenherr, 1817)
Orig. comb.: Clytus antilope Schoenherr, 1817 Subspecies: Xylotrechus antilope antilope (Schoenherr, 1817) Xylotrechus antilope obliquefasciatus (Pic, 1890) Other names: Clytus hieroglyphicus Drapiez, 1819; Xylotrechus antilope var. sekerai Podaný, 1970 (invalid name); Xylotrechus antilope ab. lentoi Paulian, 1979 (invalid name); Xylotrechus antilope sekerai (Podaný) Paulian, 1986 (invalid name). This species has two subspecies. The subspecies Xylotrechus antilope obliquefasciatus (Pic, 1890) occurs only in North Africa (Algeria, Morocco, Tunisia). So the species is represented by the nominotypical subspecies in Turkey. It is distributed in NW and W Turkey for Turkey. Records in Turkey: Ġstanbul prov.: Belgrad Forest (Acatay, 1943); Ġstanbul prov.: Alem Mountain (Schimitschek, 1944); Ġstanbul prov.: Polonez village (Demelt & Alkan, 1962; Demelt, 1963); Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama, 2002); Bursa prov.: Uludağ (Adlbauer, 1992); European Turkey (Althoff & Danilevsky, 1997); Manisa prov.: Muradiye (Tezcan & Rejzek, 2002); Kırklareli prov.: Demirköy (pers. comm.); Black Sea Region (Özdikmen, 2007); Marmara and Aegean Regions (Özdikmen, 2008a,b) (Fig. 2). Distribution: Europe (Portugal, Spain, France, Italy, Slovenia, Croatia, Bosnia-Herzegovina, Albania, Greece, Bulgaria, European Turkey, Romania, Hungary, Austria, Switzerland, Germany, Czechia, Slovakia, Netherland, Norway, Poland, Sweden, Belorussiya, Ukraine, Crimea, Moldavia, European Russia), Caucasus (Armenia, Azerbaijan, Georgia), Turkey, Cyprus, Iran, North Africa (Algeria, Morocco, Tunisia). Chorotype: W-Palaearctic.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 278
Xylotrechus arvicola (Olivier, 1795) Orig. comb.: Callidium arvicola Olivier, 1795 Other names: Clytus heydeni Stierlin, 1864; Clytus kraatzi Lederer, 1864; Clytus tridentatus Bleuse, 1905; Xylotrechus inbasalis Pic, 1911; Xylotrechus herzegovinus Pic, 1913; Xylotrechus subangulosus Pic, 1924; Xylotrechus arvicola ab. basifasciatus Plavilstshikov, 1940; Xylotrechus arvicola ab. degradatus Plavilstshikov, 1940; Xylotrechus abbreviatus Schmidt, 1951; Xylotrechus arvicola var. layrei Villiers, 1970. This species probably distributes rather widely in Turkey. It is the first record for Tekirdağ province. Material examined: Tekirdağ prov.: 01.06.1976, on Asphodelus sp., 1 specimen, det. Holzschuh. Material housed in the collection of LEMT (Lodos Entomological Museum, Turkey) of Plant Protection Department, Faculty of Agriculture, Ege University, Bornova, Ġzmir, Turkey. Records in Turkey: Giresun prov.: Tirebolu (Karadua Forests, Yağlıdere village) (Sekendiz, 1981); Turkey (Danilevsky & Miroshnikov, 1985; Lodos, 1998; Sama, 2002); Hatay prov.: Ġskenderun (Amanos Mountains) (Öymen, 1987); Adana prov.: Kozan (Feke) (Adlbauer, 1992); European Turkey (Althoff & Danilevsky, 1997); Trabzon prov.: Maçka (Alkan, 2000); Kırklareli prov.: Demirköy (pers. comm.); Black Sea Region (Özdikmen, 2007); Marmara (Özdikmen, 2008a) (Fig. 3). Distribution: Europe (Portugal, Spain, France, Corsica, Italy, Sicily, Sardinia, Albania, Slovenia, Croatia, Bosnia-Herzegovina, Serbia, Macedonia, Greece, Bulgaria, European Turkey, Romania, Hungary, Austria, Switzerland, Belgium, Netherlands, Germany, Luxembourg, Czechia, Slovakia, Poland, Estonia, Latvia, Lithuania, Belorussia, Ukraine, Crimea, Moldavia, European Russia), Caucasus (Armenia, Azerbaijan, Georgia), Turkey, Syria, North Africa (Algeria, Morocco). Chorotype: W-Palaearctic.
Xylotrechus stebbingi Gahan, 1906 (Fig.1)
Orig. comb.: Callidium arvicola Olivier, 1795 Other names: Clytus heydeni Stierlin, 1864; Clytus kraatzi Lederer, 1864; Clytus tridentatus Bleuse, 1905; Xylotrechus inbasalis Pic, 1911; Xylotrechus herzegovinus Pic, 1913; Xylotrechus subangulosus Pic, 1924; Xylotrechus arvicola ab. basifasciatus Plavilstshikov, 1940; Xylotrechus arvicola ab. degradatus Plavilstshikov, 1940; Xylotrechus abbreviatus Schmidt, 1951; Xylotrechus arvicola var. layrei Villiers, 1970. This species distributes at least in Western Anatolia for Turkey. It is the first record for Turkey. The original description of Gahan (1906) as follows:
Xylotrechus stebhingi sp. n. Brown; head and prothorax clothed with grey pubescence; the prothorax
with four small brown spots in a transverse row across the middle—two dorsal and two lateral. Elytra subglabrous, testaceous brown, narrowly

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 279
covered with grey pubescence at the base, marked with some small spots of ashy-grey pubescence which form three interrupted bands—one near the base, another just before the middle, the third midway between it and the apex; the apex also narrowly bordered with ashy grey. Body beneath covered with grey pubescence, a rather large posterior spot on each of the metathoracic episterna ashy white. Head with the lateral carinae oblique, slightly curved, extending below almost to a level with the lower margin of the eyes; front narrowed between the eyes, furnished with two prominent convergent carinae, which are united below. Antennae less than half the length of the body; third joint slightly longer than the first. Prothorax widest behind the middle, very slightly narrowed in front, strongly narrowed towards the base; disc with a median asperate carina, which is broader and more strongly raised behind than in front. Femora rather strongly thickened; the hind pair extending a little past the apex of the elytra. First joint of hind tarsi twice as long as the second and third united.
Length 12-18; breadth 3.1/4-5 mm. Hab. North-West Himalayas: Bashahr State (E. P. Stebbing); Tibet.
Material examined: Ġzmir prov.: Bornova, N 38º 27‘ 20‘‘ / E 27º 13‘ 37‘‘, 34 m, 09.08.2010, 14 specimens, 11.08.2010, 34 specimens, 13.08.2010, 58 specimens, 16.08.2010, 47 specimens, 18.08.2010, 34 specimens, 20.08.2010, 44 specimens, 23.08.2010, 48 specimens, 25.08.2010, 31 specimens, 27.08.2010, 35 specimens, 01.09.2010, 20 specimens, 03.09.2010, 2 specimens, 07.09.2010, 10 specimens, 13.09.2010, 12 specimens, 15.09.2010, 18 specimens, 17.09.2010, 5 specimens, 22.09.2010, 1 specimen. Totally 413 specimens. Adults reared from died branches of white mulberry (Morus alba Linnaeus) (Rosales: Moraceae). Material housed in the collection of LEMT (Lodos Entomological Museum, Turkey) of Plant Protection Department, Faculty of Agriculture, Ege University, Bornova, Ġzmir, Turkey and in the collection of Zoology Museum of Gazi University (ZMGU), Ankara, Turkey. Records in Turkey: Absent (Fig. 4). Distribution: Europe (France, Italy, Slovenia, Greece, Switzerland), Afghanistan, Bhutan, Nepal, Pakistan, India, Tajikistan, China, Oriental Region, Turkey, Israel, North Africa (Tunisia). Chorotype: Mediterraneo-Sindian + Oriental.
LITERATURE CITED Acatay, A. 1943. Ġstanbul çevresi ve bilhassa Belgrad ormanındaki zararlı orman böcekleri, mücadeleleri ve iĢletme üzerine tesirleri. T. C. Ziraat Vekâleti Yüksek Ziraat Enstitüsü ÇalıĢmaları, Ankara, 142: 163 pp. Adlbauer, K. 1992. Zur Faunistik und Taxonomie der Bockkäferfauna der Türkei II (Coleoptera, Cerambycidae). Entomofauna, 13 (30): 485-509. Alkan, H. 2000. Türkiye orman Cerambycidae (Insecta, Coleoptera)‘lerinin tanıtımı ve Doğu Karadeniz Bölgesindeki türlerin araĢtırılması. Yüksek Lisans Tezi. Karadeniz Teknik Üniversitesi Fen Bilimleri Enstitüsü, Trabzon, 227 pp. Althoff, J. & Danilevsky, M. L. 1997. A Check-List of Longicorn Beetles (Coleoptera, Cerambycoidea) of Europe. Slovensko Entomološko Društvo Štefana Michielija. Ljubljana, 64 pp. Danilevsky, M. L. & Miroshnikov A. I. 1985. Timber-Beetles of Caucasus (Coleoptera, Cerambycidae). The Key. Krasnodar, 419 pp. Demelt, C. V. 1963. Beitrag zur Kenntnis der Cerambycidenfauna Kleinasiens und 13. Beitrag zur Biologie palaearkt. Cerambyciden, sowie Beschreibung einer neuen Oberea-Art. Entomologische Blatter 59 (3) : 132-151.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 280
Demelt, C. V. & Alkan, B. 1962. Short information of Cerambycidae Fauna of Turkey. Bitki Koruma Bülteni 2 (10): 49-56. Gahan, C. J. 1906. The fauna of British India, including Ceylon and Burma. Coleoptera Vol.-I- (Cerambycidae). Taylor & Francis, Red Lion Court, Fleet Street, London, 329 pp [244]. Lodos, N. 1998. Entomology of Turkey VI (General, Aplied and Faunistic). Ege Ü. Ziraat Fak. Yayınları No: 529, E. Ü. Faculty of Agriculture Press, Ġzmir, 300 pp. Öymen, T. 1987. The Forest Cerambycidae of Turkey. Ġ. Ü. Forest Faculty, Ġstanbul, 146 pp. Özdikmen, H. 2007. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part I - Black Sea Region. Munis Entomology & Zoology, 2 (2): 179-422. Özdikmen, H. 2008a. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part II – Marmara Region. Munis Entomology & Zoology, 3 (1): 7-152. Özdikmen, H. 2008b. The Longicorn Beetles of Turkey (Coleoptera: Cerambycidae) Part III – Aegean Region. Munis Entomology & Zoology, 3 (1): 355-436. Sama, G. 2002. Atlas of the Cerambycidae of Europe and the Mediterranean Area, Volume I, Kabourek, Zlin, 173 pp. Sama, G. & Löbl, I. 2010. Cerambycinae. In Löbl, I. & Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera. Vol. 6, Chrysomeloidea, pp. 143-207 [180-183]. Schmitschek, E. 1944. Forstinsekten der Türkei und Ihre Umwelt Grundlagen der türkischen Forstentomologie, Volk und Reich Verlag Prag, 125-141 pp. Sekendiz, O. A. 1981. Doğu Karadeniz bölümünün önemli teknik hayvansal zararlıları üzerine araĢtırmalar. K. T. Ü. Orman Fakültesi Yayınları no: 12, Trabzon, 114 pp. Tezcan, S. & Rejzek, M. 2002. Longhorn beetles (Coleoptera: Cerambycidae) recorded in cherry orchards in Western Turkey. Zoology in the Middle East, 27: 91-100.
Figure 1. Xylotrechus stebbingi Gahan, 1906 on Morus alba.
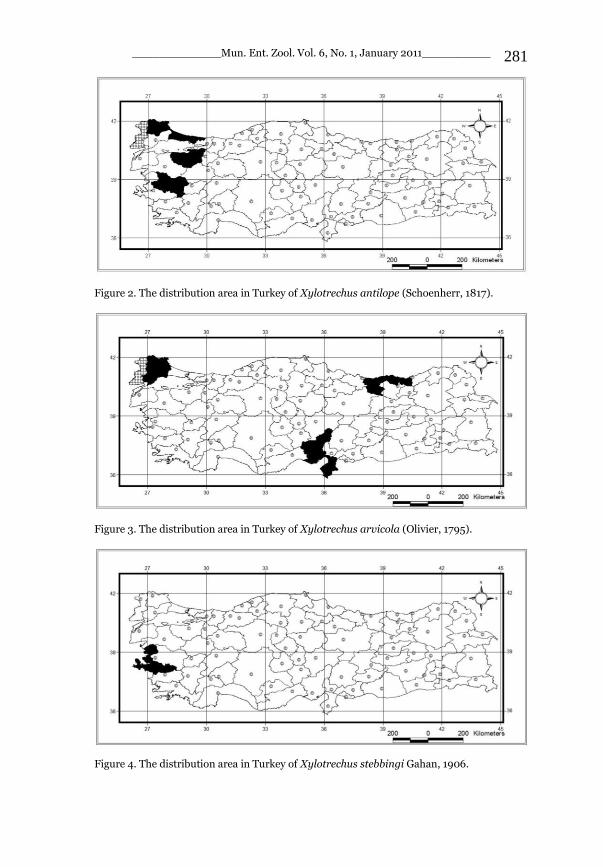
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 281
Figure 2. The distribution area in Turkey of Xylotrechus antilope (Schoenherr, 1817).
Figure 3. The distribution area in Turkey of Xylotrechus arvicola (Olivier, 1795).
Figure 4. The distribution area in Turkey of Xylotrechus stebbingi Gahan, 1906.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 282
FUMIGANT TOXICITY OF CARUM COPTICUM L. OIL AGAINST TRIBOLIUM CONFUSUM DU VAL, RHYZOPERTHA
DOMINICA F. AND ORYZAPHILUS SURINAMENSIS L.
Afsaneh Shokri Habashi*, Mohammad Hassan Safaralizadeh and Seyed Ali Safavi
*Department of Plant Protection, Faculty of agriculture, Urmia University, Urmia, West Azerbaijan, P.O. Box 57135-165, IRAN. E-mail: [email protected] [Habashi, A. S., Safaralizadeh, M. H. & Safavi, S. A. 2011. Fumigant toxicity of Carum copticum L. oil against Tribolium confusum du Val, Rhyzopertha dominica F. and Oryzaphilus surinamensis L.. Munis Entomology & Zoology, 6 (1): 282-289] ABSTRACT: Plant secondary metabolites play an important role in the plant-insect interactions. Some compounds extracted from plants have insecticidal activity against insects. The chemical composition of the essential oil from seeds of Carum copticum grown in mashhad, Iran, was studied by gas chromatography mass spectrometry (GC-MS). Thymol (43%), γ-terpinen (15.85%) and β-cymene (21.67%) were found to be the major constituents of the oil. Fumigant toxicity test of above-mentioned plant essential oil against adults of Tribolium confusum du Val, Rhyzopertha dominica F. and Oryzaephilus surinamensis L. were carried out at 27±2°C and 60±5% RH. Results demonstrated that O. surinamensis (LC50 = 1.69 μl/l air) was significantly more susceptible than R. dominica (LC50 = 19.01 μl/l air) and T. confusum (LC50 = 58.70 μl/l air) at 24 h length of exposure to this essential oil. In all cases, considerable differences were observed in mortality of insects with essential oil vapor in different concentrations and times. As expected, mortality was increased by increasing of doses and exposure time after 72 h fumigation. KEY WORDS: Carum copticum, Essential oil, Tribolium confusum, Rhyzopertha dominica, Oryzaephilus surinamensis, fumigant toxicity.
In many storage systems, fumigants are the most economical and convenient tools for managing stored-grain insect pests, not only because of their ability to kill a broad spectrum of pests but also because of their easy penetration into the commodity while leaving minimal residues (Mueller, 1990). Currently, methyl bromide and phosphine fumigants are widely used for insect pest control in stored products. However, because of its ozone depletion potential, methyl bromide is being phased out the world over. Additionally, it has been reported that some stored-product insects are found to have developed resistance to phosphine in many countries (Subramanyam & Hagstrum, 1995). Hence, there is a need to develop new types of selective insect-control alternatives with fumigant action. In fact, management of stored product pests, using substances of natural origin, is nowadays the subject of many studies (Isman, 2006).
Carum copticum is popularly known as ajowan; it is an annual herbaceous plant bearing feathery leaves and white flowers grow in compound umbels. When the seeds are ripe, they are dried and threshed. Ajowan is widely distributed plant throughout the world and especially in India (Thangam & Dhananjayan., 2003). It is indigenous to southern India and is cultivated in various areas such as Europe, Egypt, Pakistan, Afghanistan and Iran (Gersbach & Reddy, 2002). Ajowan fruits are an important commercial product for the food/flavoring industry and they accumulate up to 5% essential oil in compartments referred to as canals or vita (Minija & Thoppil, 2002). Composition of the essential oil has been analyzed extensively (Gersbach & Reddy, 2002; Minija & Thoppil., 2002). The essential oil is considered for its antimicrobial and insecticidal activity. Rani

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 283
and Khullar (2004) showed antimicrobial activity of ajowan essential oil against multi-drug resistant Salmonella typhi (Schroeter).
In the present study, chemical constituents of the essential oil from C. copticum were analyzed by GC and GC-MS. Moreover, fumigant toxicity of the experimented material was assessed against Confused flour beetle T. confusum du val, Lesser grain borer R. dominica and Saw-toothed grain Beetle O. surinamensis.
MATERIALS AND METHODS Insects
Preliminary population was obtained from laboratory stock cultures maintained at the Entomology Department, Urmia University, Iran. T. confusum, R. dominica and O. surinamensis were reared in glass containers (1 litter) containing wheat flour mixed with yeast (10:1 w/w), whole wheat grains and raisin, respectively, that was covered by a fine mesh cloth for ventilation. The cultures were maintained in the darkness in a growth chamber set at 27±2°C and 60%±5% RH.
Adult insects, 1-7 days old, were used for fumigant toxicity tests. All experimental procedures were carried out under the same environmental conditions as the cultures. Extraction of the essential oil
Seeds of C. copticum were collected from mashhad, Iran. Essential oil was extracted from dried seeds with a Clevenger-type apparatus to hydrodistillation. For extraction of mentioned essential oil, 100 g of air-dried and grinded seeds were put in the Clevenger then some sterile water was added. After 2 h distillation, extracted essential oil was collected. Analysis of the essential oil
The oil composition was analyzed by gas chromatography-mass spectrometry (GC–MS). GC–MS analysis was performed using a Thermofinnigan Trace GC 2000 equipped with a MS fused silica capillary column (30 m × 0.25 mmi.d, film thickness 0.25 μm). For GC–MS detection, an electron ionization system with ionization energy of 200 eV was used. Carrier gas was helium at a flow rate of 1.5 ml/min; injection speed 1ml/min; split flow 15 ml/min. The oven temperature was programmed from 120 to 260°C at 5°C/min, then held isothermal for 12 min and finally raised to 350°C at 10°C/min. A relative percentages of the oil constituents was expressed as percentages by peak area normalization. The identification of individual compounds of essential oils was based on comparison of their relative retention index with those of authentic samples on DB-225 capillary column, and by matching of their mass spectra and peaks with those obtained from authentic samples. Fumigant toxicity
To determine the fumigant toxicity of the C. copticum oil, filter papers (Whatman No. 1, cut into 2 cm diameter pieces) were impregnated with oil at doses calculated to give equivalent fumigant concentrations. Then the impregnated filter paper was attached to the under-surface of the screw cap of a plastic tube (35 mL). The caps were screwed tightly on the plastic tube containing twenty adults (1-7 days old) of each insect, separately. Each concentration and control was replicated three times. Mortality was determined 24, 48 and 72 h

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 284
after exposure. Those insects that did not move when lightly probed or shaken in the light and mild heat were considered dead. Statistical analysis
The experiment was arranged by factorial experiment and the data were analyzed with analysis of variance (ANOVA) by using the SPSS 16.0 software. Differences between means were tested using Tukey test and values with p < 0.05 were considered significantly different. Probit analysis was used to estimate LC50 and LC95 values with their fiducial limits by SPSS 16.0 software package.
RESULTS Chemical constituents of essential oil
Nine compounds were identified in the oil of C. copticum representing 96.1% of the total oil, with thymol (43%), γ-terpinen (15.85%) and β-cymene (21.67%) as the major constituents (Table 1). Fumigant toxicity
Carum copticum oil showed potential fumigant activity against adults of O. surinamensis, R. dominica and T. confusum at different concentrations and exposure times. LC50 values were obtained 1.69, 19.01 and 58.70 μl/l air after 24 h exposure for O. surinamensis, R. dominica and T. confusum, respectively. LC50
values were resulted 0.80, 15.12 and 51.96 μl/l air, for O. surinamensis, R. dominica and T. confusum, respectively after 48 h of exposure. These values were obtained for O. surinamensis, R. dominica and T. confusum 0.43 μl/l , 12.83 μl/l and 47.05 μl/l air, respectively after 72 h length of exposure (Table. 2 A).As expected, in all of three tested insects results showed that the increasing of exposure time was caused to decreasing LC50 and LC95. Results of LC50 and LC95 values demonstrated that O. surinamensis is more susceptible than R. dominica and T. confusum. Therefore, T. confusum is more resistance than two experimented pest in three times of experiments.
Table. 2 B showed that LT50 and LT95 values for each three experimented insect. For obtaining LT50 values highest concentration of essential oil was used in each insect. The LT50 value was obtained 9.74, 11.85 and 5.19 h and LT95 was resulted 43.81, 33.8 and 62.25 h for O. surinamensis R. dominica and T. confusum, respectively.
Figure A demonstrated the mortality percentages of five different concentrations in three times on T. cofusum. Figure B showed the percentage of mortality in five concentrations of essential oil for R. dominica in three times. Also figure C showed these values for O. surinamensis.
DISCUSSION
Results of this study showed that essential oil of C. copticum had potent fumigant toxicity against adults of O. surinamensis, R. dominica and T. confusum. The insecticidal activity varied with insect species, concentrations of the oil and extension of exposure times. To the best of our knowledge, no studies have been reported previously concerning the activity of C. copticum as fumigant on O. surinamensis, R. dominica and T. confusum.
The insecticidal constituents of many plant extracts and essential oils are monoterpenoids. Due to their high volatility they have fumigant action that might be of great importance for stored product insects (Lee et al., 1997; Ahn et al.,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 285
1998). Thymol, a monoterpenoid is the major component in C. copticum essential oil. There are numerous reports on insecticidal activity of thymol. Regnault-Roger and Hamraoui (1995) tested 22 essential oils for their fumigant toxicity to the bean weevil, Acanthoscelides obtectus (Say), and found that thymol, carvacrol and terpineol to be effective in inhibiting beetle reproduction. Erler (2005) reported the fumigant activity of thymol against adults and eggs of Tribolium confusum Jacquelin Du Val, and larvae and eggs of Ephestia kuehniella Zeller. β-cymene (21.67%), γ-terpinene (15.85%), α-pinene (4.86%), β-pinene (3.62%), 4- terpineol (1.65%) are the other components of C. copticum oil that have insecticidal activity. For instance, among the major monoterpenoids of essential oil, it was reported that γ-terpinene and thymol were the most active constituents against adults of the rice weevil, S. oryzae (Erler, 2007). α-pinene was reported to be toxic to T. confusum (Ojimelukwe & Alder, 1999). Suya et al. (1998) showed the fumigant activity of thymol against Rhyzopertha dominica after its isolation from the essential oil of Elsholtzia spp. So the toxic effects of C. copticum oil could be attributed to thymol and other components.
As major constituents of C. copticum are monoterpenoids, they are typically volatile and can penetrate into insect commodity rapidly (Lee et al., 2003). There are several reports indicating that Tribolium sp. is relatively tolerant to essential oils of different plants (Xie et al., 1995; Huang et al., 1997; Liu & Ho, 1999). Sahaf et al. (2007) studied chemical constituents and fumigant toxicity of essential oil from C. copticum against S. oryzae and T. castaneum. They observed that S. oryzae (LC50 = 0.91 μL/L) were significantly susceptible than T. castaneum (LC50 = 33.14 μL/L). Obtained LC50 and LC95 values of current study showed that these results is more different from than Sahaf's et al. results. Probably, these differences is due to condition of hydrodistillation, plant species, the used plant part in these studies, the season and the method of harvesting of target plant and geographical zone and pedological conditions for the plant growth.
Our observations indicated that T. confusum is more tolerant than R. dominica and O. surinamensis.
The insecticidal activity of some essential oils from Apiaceae has been evaluated against a number of stored product insects. Also, the essential oil of C. copticum had activity against growth stages of Indian meal moth, Plodia interpunctella (Hubner) (Lepidoptera: Pyralidae) (Shojaaddini et al., 2008). Chaubey (2007) investigated insecticidal activity of Trachyspermum ammi (Umbelliferae), Anethum graveolens (Umbelliferae) and Nigella sativa (Ranunculaceae) essential oils against stored product beetle T. castaneum. The death of adults of T. castaneum was caused by fumigation with these essential oils. In an other experiment using extracts of Foeniculum vulgare Gaetner (Apiaceae) fruit, over 90% mortality was achieved in adults of S. oryzae and C. chinensis at 3 or 4 days after treatment (Kim et al., 2003). Ogendo et al. (2008) evaluated bioactivity of Ocimum gratissimum L. oil and two of its constituents against five stored product pests. The oil caused 98%, 99% and 100% mortality of R. dominica, O. surinamensis and C. chinensis, respectively, 24 h after treatment, whereas eugenol achieved 79%, 61% and 100% mortality of the same insects except for T. castaneum which was more tolerant. Kordali et al. (2008) tested insecticidal properties of essential oil isolated from Turkish Origanum acutidens on two stored product pests. O. acutidens oil caused 68.3% and 36.7% mortality of S. granarius and T. confusum adults, respectively, after 96 h of exposure. Results showed that the oil was more toxic against S. granarius as compared with its toxicity against T. confusum. Ebadollahi et al. (2010) tested toxicity of essential oil of Agastache foeniculum (Pursh) Kuntze to Oryzaephilus surinamensis L. and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 286
Lasioderma sericorne F. LC50 data's for adults showed that O. surinamensis was more susceptible than L. serricorne at the exposure time 24 h. Our experiments demonstrated that O. surinamensis adults were more susceptible than R. dominica and T. confusum. Results of the present and earlier studies demonstrated that the essential oil from C. copticum have had toxic properties on the insect pests. The use of plant materials in pest control could become important supplements or alternatives to imported synthetic pesticides. Therefore, it is important that appropriate technology is developed to promote a direct preparation of traditional pesticides at the extensive level for those poor farmers who have no access to commercial pesticides or cannot afford.
ACKNOWLEDGEMENTS
I would like to acknowledge the financial support provided to this research by the University of Urmia in Iran and biotechnology institute of Urmia. Also first author expressed best regards to my colleagues Arman Abdolmaleki, Iman Sharifian, Jafar Hoseinzadeh and Ramin Tandorost for their useful guidance.
LITERATURE CITED Ahn, Y. J. Lee, S. B., Lee, H. S. & Kim, G. H. 1998. Insecticidal and acaricidal activity of caravacrol and b-thujaplicine derived from Thujopsis dolabrata var. hondai sawdust. Journal of Chemical Ecology, 24: 1–90. Chaubey, M. K. 2007. Insecticidal activity of Trachyspermum ammi (Umbelliferae), Anethum graveolens (Umbelliferae) and Nigella sativa (Ranunculaceae) essential oils against storedproduct beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). African Journal of Agricultural Research, 2 (11): 596–600. Ebadollahi, A. Safaralizadeh, M. H. Pourmirza, A. A. & Gheibi, S. A. 2010. toxicity of essential oil of Agastache foeniculum (Pursh) Kuntze to Oryzaephilus surinamensis L. and Lasioderma sericorne F. Journal of Plant Protection Research, 50 (2): 215-219. Erler, F. 2005. Fumigant activity of six monoterpenoids from aromatic plants in Turkey against the two stored-product pests confused flour beetle, Tribolium confusum, and Mediterranean flour moth, Ephestia kuehniella. Journal of Plant Disease Protection, 112: 602-611. Erler, F. 2007. Fumigant activity of monoterpenoides against the rice weevil, Sitophilus oryzae. In: S. Navarro, C. Adler, J. Riudavets, V. Stejskal, (eds.) ―Integrated Protection of Stored Products‖ IOBC/WPRS, Prague, Czech Republic, Bull, 30 (2). pp: 281–285. Gersbach, P. V. & Reddy, N. 2002. Non-invasive localization of thymol accumulation in Carum copticum (Apiaceae) fruits by chemical shift selective magnetic resonance imaging. Ann. Bot, 90: 253–257. Huang, Y. Tan, J. M. W. L. Kini, R. M. & Ho, S. H. 1997. Toxic and antifeedant action of nutmeg oil against Tribolium castaneum (Herbst) and Sitophilus zeamais Motsch. Journal of Stored Products Research, 33: 289-298. Isman, M. B. 2006. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annual Review of Entomology, 51: 45-66. Kim, S. I. Roh, J. Y. Kim, D. H. Lee, H. S. & Ahn, Y. J. 2003. Insecticidal activities of aromatic plant extracts and essential oils against Sitophilus oryzae and Callosobruchus chinensis. Journal of Stored Products Research, 39: 293-303. Kordali, S. Cakir, A. Ozer, H. Cakmakci, R. Kesdek, M. & Mete, E. 2008. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresource Technology, 99: 8788–8795.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 287
Lee, S., Tsao, R., Peterson, C. & Coats, J. R. 1997. Insecticidal activity of monoterpenoids to western corn rootworm (Coleoptera: Chrysomelidae), twospotted spidermite (Acari: Tetranychidae), and house fly (Diptera: Muscidae). Ecotoxicology, 90(4): 883–892. Lee, S., Peterson, C. J. & Coats, J. R. 2003. Fumigation toxicity of monoterpenoids to several stored product insects. Journal of Stored Products Research, 39: 77-85. Liu, Z. L. & Ho, S. H. 1999. Bioactivity of the essential oil extracted from Evodia rutaecarpa Hook f. et Thomas against the grain storage insects, Sitophilus zeamais Motsch. And Tribolium castaneum (Herbst). Journal of Stored Products Research, 35: 317-328. Minija, J. & Thoppil, J. E. 2002. Essential oil composition of Trachyspermum ammi (L) Sprague from South India. Indian Journal of Pharmaceutical Sciences, 64: 250–251. Mueller, D. K. 1990. Fumigation, Handbook of Pest Control , Franzak and Foster Co., Cleveland, Ohio. pp: 901-939. Ogendo, J. O., Kostyukovsky, M., Ravid, U., Matasyoh, J. C., Deng, A. L., Omolo, E. O., Kariuki, S. T. & Shaaya, E. 2008. Bioactivity of Ocimum gratissimum L. oil and two of its constituents against five insect pests attacking stored food products. Journal of Stored Products Research, 44: 328–334. Ojimelukwe, P. C. & Alder, C. 1999. Potential of zimtaldehyde, 4-allyl-anisol, linalool, terpineol and other phytochemicals for the control of the confused flour beetle (Tribolium confusum J. d. V.) (Col: Tenebrionidae). Journal of Pesticide Science, 72: 81-86. Rani, P. & Khullar, N. 2004. Antimicrobial evaluation of some medicinal plants for their antienteric potential against multi-drug resistant Salmonella typhi. Phytotherapy Research, 18: 670–673. Regnault-Roger, C. & Hamraoui, A .1995. Fumigant toxic activity and reproductive inhibition induced by monoterpenes on Acanthoscelides obtectus (Say) (Coleoptera), a bruchid of kidney bean (Phaseolus vulgaris L.). Journal of Stored Products Research, 31: 291–299. Sahaf, B. Z., Moharramipour, S. & Meshkatalsadat, M. H. 2008. Fumigant toxicity of essential oil from Vitex pseudo-negundo against Tribolium castaneum (Herbst) and Sitophilus oryzae (L.). Journal of Asia-Pacific Entomology, 11 (4): 175-179. Shojaaddini, M. Moharramipour, S. & Sahaf, B. Z. 2008. Fumigant toxicity of essential oil from Carum copticum against Indian meal moth, Plodia interpunctella. Journal of Plant Protection Research, 48 (4): 411-419. Subramanyam, B. & Hagstrum, D. W. 1995. Resistance measurement and management. Integrated Management of Insects in Stored Products. Marcel Dekker, New York. pp: 331-397. Suya, W., Yongjia, J., Chunming, L. & Yingjie, Z. 1998. Studies of population reproductive inhibiting effect on volatile oil from Elsholtiza sp. against stored-product insects. In: Zuxun, J., Quan, L., Yongsheng, L., Xianchang, T., Lianghua, G. (Eds.), Proceedings of the Seventh International Conference on Stored-product Protection, 14–19 October 1998, Beijing, China, vol. 1. Sichuan Publishing House of Science and Technology, Chengdu, P.R. China, 858–865. Thangam, C. & Dhananjayan, R. 2003. Antiinflammatory potential of the seeds of Carum copticum Linn. Indian Journal of Pharmacology, 35: 388–391. Xie, Y. S., Fields, P. G., Isman, M. B., Chen, W. K. & Zhang, X. 1995. Insecticidal activity of Melia toosendan extracts and toosendanin against three stored product insects. Journal of Stored Products Research, 31: 259-265.
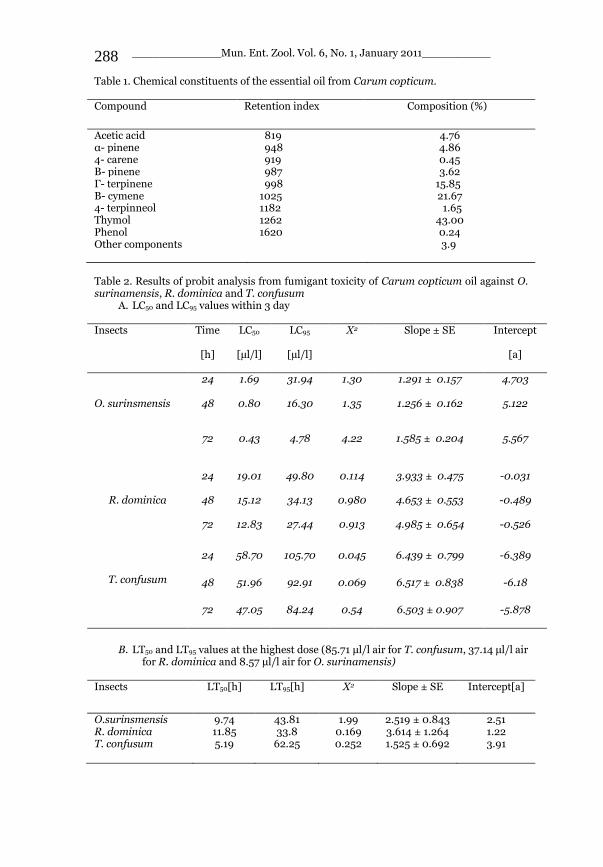
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 288
Table 1. Chemical constituents of the essential oil from Carum copticum. Compound Retention index Composition (%)
Acetic acid 819 4.76 α- pinene 948 4.86 4- carene 919 0.45 Β- pinene 987 3.62 Γ- terpinene 998 15.85 Β- cymene 1025 21.67 4- terpinneol 1182 1.65 Thymol 1262 43.00 Phenol 1620 0.24 Other components 3.9
Table 2. Results of probit analysis from fumigant toxicity of Carum copticum oil against O. surinamensis, R. dominica and T. confusum
A. LC50 and LC95 values within 3 day
Insects Time
[h]
LC50
[µl/l]
LC95
[µl/l]
Χ2 Slope ± SE
Intercept
[a]
O. surinsmensis
24 1.69 31.94 1.30 1.291 ± 0.157 4.703
48 0.80 16.30 1.35 1.256 ± 0.162 5.122
72 0.43 4.78 4.22 1.585 ± 0.204 5.567
R. dominica
24 19.01 49.80 0.114 3.933 ± 0.475 -0.031
48 15.12 34.13 0.980 4.653 ± 0.553 -0.489
72 12.83 27.44 0.913 4.985 ± 0.654 -0.526
T. confusum
24 58.70 105.70 0.045 6.439 ± 0.799 -6.389
48 51.96 92.91 0.069 6.517 ± 0.838 -6.18
72 47.05 84.24 0.54 6.503 ± 0.907 -5.878
B. LT50 and LT95 values at the highest dose (85.71 μl/l air for T. confusum, 37.14 μl/l air
for R. dominica and 8.57 μl/l air for O. surinamensis)
Insects LT50[h] LT95[h] Χ2 Slope ± SE
Intercept[a]
O.surinsmensis 9.74 43.81 1.99 2.519 ± 0.843 2.51 R. dominica 11.85 33.8 0.169 3.614 ± 1.264 1.22 T. confusum 5.19 62.25 0.252 1.525 ± 0.692 3.91
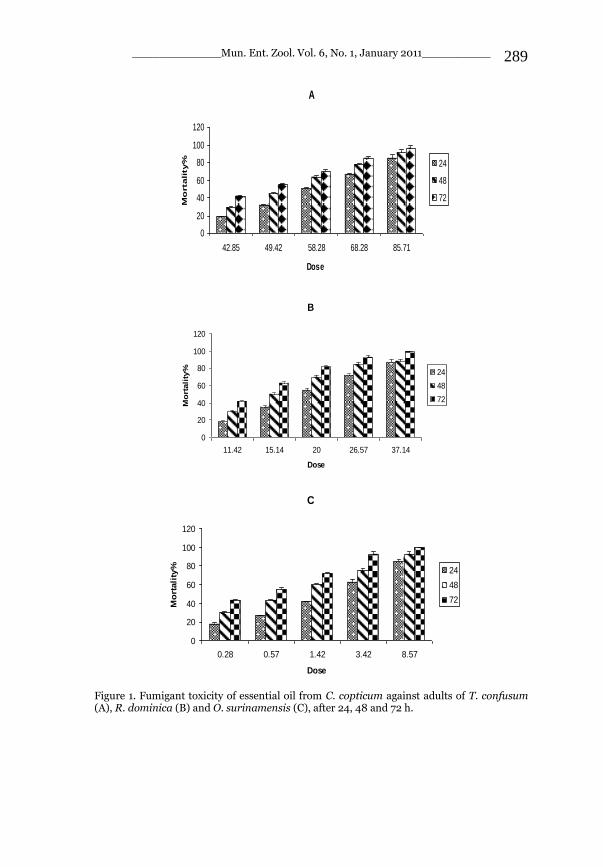
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 289
A
0
20
40
60
80
100
120
42.85 49.42 58.28 68.28 85.71
Dose
Mo
rtality% 24
48
72
B
0
20
40
60
80
100
120
11.42 15.14 20 26.57 37.14
Dose
Mo
rtali
ty% 24
48
72
C
0
20
40
60
80
100
120
0.28 0.57 1.42 3.42 8.57
Dose
Mo
rtali
ty% 24
48
72
Figure 1. Fumigant toxicity of essential oil from C. copticum against adults of T. confusum (A), R. dominica (B) and O. surinamensis (C), after 24, 48 and 72 h.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 290
IN VIVO USE OF STREPTOMYCIN SULFATE FOR BACTERIAL DISEASE CONTROL IN ANTHERAEA ASSAMENSIS HELFER
THROUGH LEAF FRESHNESS TECHNIQUE
Himangshu Barman*
* Central Muga Eri Research and Training Institute, Central Silk Board, Lahdoigarh, Jorhat – 785700, Assam, INDIA. E-mail: [email protected] [Barman, H. 2011. In vivo use of streptomycin sulfate for bacterial disease control in Antheraea assamensis Helfer through leaf freshness technique. Munis Entomology & Zoology, 6 (1): 290-296] ABSTRACT: Among all biotic and abiotic constrains in Muga silkworm culture, disease is the most consistent and devastating one. Bacterial flacherie is such a silkworm disease that inflects more than 42% crop loss. Antibacterial drugs like streptomycin sulfate can be used successfully through leaf freshness technique useable in indoor rearing of Muga culture. Drug solution is mixed with leaf sap as absorbent by Som plant twigs that larvae feed on and, targeted disease causing bacteria in larval gut are exposed to streptomycin action. In vivo use of streptomycin sulfate resulted complete control of flacherie disease caused by Bacillus bacteria up to fourth instars. Being bacteria free in gut, larval feeding was more that was directly reflected in larval body weight and indirectly increases vurvivality of larvae. In this experiment with in vivo use of streptomycin sulfate 42% more survivality up to fifth instars of Muga larvae was achieved over 92% mortality in control rearing. KEY WORDS: Flacherie, streptomycin sulfate, leaf freshness, survivality, body weight, mortality, in vivo.
Antheraea assamensis Helfer, the golden-yellow silk producer silk moth, is semi-domesticated sericigenous insect species endemic to North East India particularly Brahmaputra valley of Assam. This silk moth is semi-domesticated owing to the fact that only cocooning and grainage operations are conducted indoor and, reared on outdoor host plant trees. It is polyphagous, multivoltine reared in six different seasons throughout the year. Out of these six seasons two seasons viz., May-June and October-November are commercial crop season, whereas other seasons are seed crop season. Again, the seed crops during December-January and June –July are called pre-seed crop. Thus, each commercial crop is preceded by one pre-seed crop and one seed crop. Since this pattern of Muga silkworm cultivation has been an age old practice, it is obviously environment controlled and the rearing performance is quite different in each season. Being outdoor in nature Muga silkworm are exposed to various natural vagaries and thus prone to numbers of diseases and pests leading to heavy crop loss (Choudhury, 1981; Samson, 1987 and Thangavelu et al. 1988). Of all constrains, silkworm diseases being the most important that inflect heavy loss to the crop. The ‗Flacherie‘ disease caused by bacteria is most important, causing serious damage to the Muga silkworm (Chakravorty et al. 2007). The chief disease affecting silkworm is flacherie caused by Bacillus bacteria. One casual bacillus of silkworm flacherie is B. thuringiensis which is widely distributed facultative entomogenous bacterium with as many as 34 varieties. It is a garm-positive spore forming bacterium widely distributed in the soils of various regions of the world. The endotoxin of B. thuringiensis is known to destroy the gut lining, causing paralysis and death in many insect species belonging to orders, Diptera and Lepidoptera including economically important insects such as silkworm, Bombyx mori ( Aizawa, 1971; Nataraju et al., 1991). B. thuringiensis infected

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 291
larvae lost appetite and became sluggish from 5 to 6 hours of infection, larvae vomited the green fluid, excreta were soft and stick to the rearing bed. During molting, skin was not shed properly. The infection also led to diarrhea. As the disease advanced, the larvae became extremely sluggish, showed irritability to touch as if in pain. Later the colour of the larvae started to change into dark colour and larvae became almost inactive and unable to spin cocoons. The larval body started to shrink and the larvae became completely paralyzed. Finally, larvae completely turned to brown colour. Within 30 minutes after the larvae ingested the spores of B. thuringiensis, the mid gut epithelial cells became disorganized compared to healthy larvae. Some of the epithelial cells became detached from the wall of the mid gut. There exist 34 different varieties of B. thuringiensis. Among the 23 varieties of Bt tested, only eight were reported to be pathogenic to silkworms and the rest as non-pathogenic (Selvakumar et al. 1999). Disease incidence may be due to the lowering of pH by the introduction of bacteria, which provides congeniality and could lead to degeneration of peritropic membrane, which blocks the absorption of nutrients that is reflected by cessation of feeding. From flacherie infected mulberry silkworm, Sridhar et al. (2000) isolated bacteria belonging to genus Streptococcus.
The biological defense against pathogens in insects included the innate physical barriers viz., integument and intestinal wall and humeral responses such as the activation of prophenol oxidase cascade and induction of immune proteins namely, lysozymes, lectins, antibacterial proteins and antifungal proteins primarily by the fat bodies. Intestine harbor a great diversity of native microbes which promote gut maturation, and integrity, antagonism against pathogens by producing antimicrobial proteins and immune modulation (Girishkumar et al., 2005). An countable number of researchers made effort to control ‗Flacherie‘ in Muga silkworm by various biological and non-biological agents as spray, including streptomycin, an antibacterial drug, and reported as effective. Streptomycin sulfate is a bactericidal antibiotic and is a water-soluble amino glycoside derived from Streptomyces griseus. It is marketed as the sulfate salt of Streptomycin. The chemical name of Streptomycin sulfate is D-Strep amine, O - 2 - dioxin - 2 - (methyl amino) - α - L - glucopyranosyl - (1→2) - O - 5 - dioxin - 3 - C - formyl - α - L - lyxofuranosyl - (1→4) - N,N1-bis(aminoiminomethyl)-,sulfate (2:3) (salt). The molecular formula for Streptomycin Sulfate is (C21H39N7O12)2 -3H2SO4 and the molecular weight is 1457.41. It acts by interfering with normal protein synthesis. Streptomycin is considered a second-line agent for the treatment of gram-negative bacillary bacteria. In this present context of study, effort has been made to test the efficacy of this drug in vivo in controlling bacterial diseases under indoor reared Muga silkworm through a new leaf freshness technique.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 292
MATERIALS AND METHOD
Eggs of Muga silkworm dfls were placed in plastic container covered with wet foam pad below the lid. The foam pad was wetted and squeezed two times in a day to maintain optimum humidity (80 – 85%) inside the container. Tender leafy twigs of Som plant (Persea bombycina Kost) were collected in bucket with water. Collected twigs were treated in clean water mixed with certain chemical solutions for three hours for freshness (proofed to keep twigs fresh). Clean and blank water was poured in two bottles in one of which, two treated twigs were inserted (C). In the other bottle, a solution of streptomycin sulfate @ 5% / liter was added to make the water in the bottle 0.5% strength (T). Another two Som plant twigs already treated were inserted into the second bottle. Now, these two bottles were placed in plastic buckets separately so as to collect dead larvae as well as litters. On 02-02-2010 day, 50 nos. hatched out larvae were brushed in each treatment (T & C), which were allowed to grow inside room. Closed watch was maintained not to crawl out from these. Each day dead larvae and litters were collected for record. Leafy twigs were replaced with newly treated one as and when necessary. Fresh solution in T bottle and water in C bottle were added as the leafs absorbed it. Collected litters of each instar were weighted and recorded. Data were tabulated for discussion.
RESULTS AND DISCUSSION
Data of the experiment in contrast between treatment and control over different parameters related to silkworm rearing have been shown in Table-1 and statistically analyzed. All these parameters are related to the growth and development of Muga silkworm which in turn, influence directly or indirectly by bacterial diseases. In the T rearing, symptoms of flacherie disease were not seen up to IVth instars to cause larval death. The death of larvae was found to be due to some other causes like failure to molt out into next instar, loss of griping power resulting fall down from bait, stunted growth with slow feeding rate and ultimate death. However, in fifth instar two cases of dead larvae hanging from twig branch were recorded indicating flacherie incidence. Contrary to T rearing, most of the larval death occurred in C rearing due to bacterial flacherie exhibiting the typical symptom hanging with its head downward from twig‘s branch and body turning into shrinkage to black-brown colour. Data in the table-1 profoundly indicate low mortality and high survivality throughout all the instars in T rearing than recorded in case of C rearing (Fig.1). Trend value of survivality in T is much higher (408) than C (298). This firmly establishes a changing positive trend in larval survivality as result of flacherie control by streptomycin sulfate. This also supported high mortality trend (156.64) among C larvae without streptomycin treatment in contrast to lower mortality trend (61.02) in T. At the end of fifth instar, 50% well grown larvae survived for spinning in T rearing against only 8% larval survivality in control rearing (C). A difference of 42% was found in aggregate larval mortality of all instars in these two treatments. Thus, bacterial flacherie caused 42% larval mortality as calculated from the mortality data of T and C rearing. Larval growth measured as larval weight (body tissue growth), although indicates differences between the treatments, the differences were within the scope of limitation and not distant. Therefore, body tissue growth in T rearing and C rearing, also statistically analyzed as trend value 14.5868 and 13.531 respectively, was not significantly different. Earlier worker like Venkatachalamurthi et al. (1951) reported that streptomycin significantly

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 293
increased the body weight of silkworm larvae. The litter weights of each instar in these two rearing, exhibited a marked differences in size of population that indirectly support the changed survivality of Muga larvae through in vivo use of antibacterial agent (streptomycin sulfate) in treatment rearing (T).
Streptomycin sulfate solution in the bottle did find its way into the leaves through twig‘s stem of which lower ends were kept immersed in the solution. So, as a physiological principle the solution of streptomycin was pulled by leaves triggered by transpiration. The leaves were ingested by the larvae in T rearing. Streptomycin sulfate along with leaf feed entered into the gut of larvae, and then into gut wall tissue system. A fall in gut pH (8.0-8.2 to 7.0-7.2) due to bacterial infection has been reported (Ono et al., 1968; Chitra et al., 1974). Inside the targeted bacterial cells present in tissue system and gut, streptomycin sulfate interfere in protein synthetic mechanism. The mode of action how streptomycin inhibits or kills the bacteria has been explained by Villanova (1990) and Stenfors Arnesen et al., 2008. As this antibacterial drug entered inside the bodies of larvae in T rearing, there was almost no dead report due to typical flacherie disease. Whereas in control rearing (C) through leaf freshness technique, 42% mortality was recorded due to bacterial flacherie out of 92% total larval mortality. Aruga et al., 1971 reported that bacterial flacherie is the most important disease affecting silkworm, causing 70% crop loss in Japan. Thus, the antibacterial action of streptomycin sulfate inside larval body has been proofed against flacherie disease causing bacteria in Muga silkworm, A. assamensis Helfer. Similar result was also reported by Savithri et al. (2003b). They reported that antibiotics streptomycin @500 ppm reduced satisfactorily disease incidence in the worms. Dutta et al. (2010) reported in vitro use of streptomycin sulfate @1000 ppm and successfully controlled bacterial diseases in Muga silkworm up to 52.37%. Besides streptomycin sulfate, other antibacterial agents have also been reported to control bacterial diseases in silkworm. Intestinal microbial environment can be fortified by supplementing probiotic microbes. This could lead to higher survival, fewer incidences of diseases and reduced mortality along with low feed conversion ratio and increased production (Tannock, 1997). The inhibitory potential of Ampicillin, Amoxicillin, Chloramphenicol and Tetracycline in different concentration has been reported positive (in vitro) against Bacillus coagulans, a bacterium that cause silkworm disease (Savithri et al., 2003a,b).
Bacterial flacherie has been found to be a major constrain in Muga cultivation in Assam that cause heavy loss in each season. The prevention or management of this disease is a vital component for successful rearing for higher yield and quality cocoons. Thus, in vivo application of streptomycin sulfate through leaf freshness technology may through ray of hope in controlling bacterial flacherie in Muga silkworm indoor cultivation for bumper harvest and quality cocoons.
LITERATURE CITED Aizawa, K. 1971. Strain improvement and preservation of virulence of pathogens. In. Microbial control of insects and mites, H. D. Burges and N. W. Hussy (Eds.), Academic press London. pp. 655-672. Aruga, H. & Tanada, Y. 1971. The cytoplasm Polyhedrosis virus of the silkworm. University of Tokyo Press, Japan, p. 234. Chakravorty, R., Das, R., Neog, K., Das, K. & Sahu, M. 2007. A diagnostic manual for diseases and pest of Muga silkworm and their host plants. Published by CMER&TI, Central Silk Board, Lahdoigarh, Jorhat, Assam. pp 1-47.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 294
Chitra, C., Karanth, N. G. K. & Vasantharajan, V. N. 1974. Studies on ‗Sappe‘ disease of the silkworm Bombyx mori L. II. Effect of age of larva on the manifestation of the disease. J. Invertebr. Pathol., 24; 218-232. Choudhury, S. N. 1981. Disease, pest and parasites. In: Choudhury, S. N. (Eds.). Muga silk industry. Directorate of Sericulture and Weaving. Govt. of Assam, Guwahati, India. pp. 74-81. Dutta, P., Neog, K., Das, R., Das, K., Handique, P. K. & Chakravorty, R. 2010. Evaluation of some botanicals, antibiotics, carbon source and carrier against the bacterial disease of Muga silkworm, Antheraea assamensis. Scrocologia, 50 (1); 113-122. Girishkumar, C. P., Thangam Menon & Anitha Devi, C. P. 2005. Bacteria for breakfast: Probiotics for good health. Advanced Biotech., March, pp. 15-20. Nataraju, B., Balavenkatasubbaiah, M., Baig, M., Singh, D. B. & Sengupta, K. 1991. A report on the distribution of Bacillus thuringiensis in Seri cultural areas of Karnataka. Indian Jn. Seric., 30 (1): 56-58. Ono, M. & Kato, S. 1968. Studies on the dissolution of peritropic membrane of the silkworm Bombyx mori. Scric. Expt. Stn. Jpn., 23: 9-16. Samson, M. V. 1987. Bacterial diseases of silkworm and their control. Ph. D thesis, CSR&TI, Mysore; 1-137. Savithri, G. & Murali Mohan, P. 2003a. Pathogenocity of the bacterium Bacillus coagulans in silkworm Bombyx mori L. Indian J. Seric., 42 (1): 4-8. Savithri, G. & Murali Mohan, P. 2003b. In vitro screening of antibiotics against Bacillus coagulans. Indian J. Seric., 4231: 25-28. Selvakumar, T., Nataraju, B. & Datta, R. K. 1999. Characterization of Bacillus thuringiensis varieties in relation to pathogenicity to silkworm, Bombyx mori. Indian J. Seric., 1: 75-78. Sridhar, R., Subramanian, A. & Chandramohan, N. 2000. Efficacy of two antibiotics against bacterial flacherie of silkworm, Bombyx mori L., 39 (2): 176-177. Stenfors Arnesen, L. P. & Fagerlund, A. 2008. From soil to gut: Bacillus cereus and its food poisoning toxins. Granum PEFEMS Microbiol Rev. Jul, 32 (4): 579-606. E-pub 2008 Apr 15. Tannock, G. W. 1997. Modification of the normal micro biota by diet, stress, antimicrobial agents and probiotics. In. Microbiology Series, Mackie R. L., B. A. Issacson, (Eds.), Chapman and Hall, International Thomson publishing, New York, pp. 435-465. Thangavelu, K., Chakravorty, A. K.., Bhagowati, A. K. & Isa, Md. 1988. Handbook of muga culture, Central Silk Board, Bangalore. Venkatachalamurthi, M. R., Shankarappa, B. S. & Sreenivasaya, M. 1951. Influence of antibiotics on the growth of silkworm Bombyx mori L.. Curr. Sci., 20: 269-270. Villanova, P. A. 1990. National Committee for Clinical Laboratory Standards. Performance Standards for Antimicrobial Disk Susceptibility Tests-Fourth Edition. Approved Standard NCCLS Document M2-A4.Vol.10, No.7, NCCLS.
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 295
Table 1. Tabulation of data showing comparison between treatment and control on different parameters and their statistical analysis.
0
50
100
150
200
250
300
T C T C T C T C T C
I instar II instar III instar IV instar V instar
Instars wise data of T & C
Varia
ble
s
Growth
Mortality
Survivality
Litters Wt.
Figure 1. Graphical presentation of data of treatment and control on larval growth, mortality, survivality and litter weight.
Larval Instars
Nos. of larvae
brushed
Treatment with streptomycin (T) Control without treatment (C)
T C Larval weight in g (5 nos.)
Mortality (%)
Survivability (%)
Litters Weight (g)
Larval weight in g (5 nos.)
Mortality (%)
Survivability (%)
Litters weight (g)
I instar 50
50 0.0332 0 100 1.41 0.0218 4 96 1.409
II instar 0.0906
0 100 5.907 0.0892 6.25 90 5.893
III instar 0.414 12 88 20.972 0.4 28.89 64 17.851
IV instar 2.229 20.45 70 42.299 2.153 37.5 40 30.14
V instar 11.82 28.57 50 256.8 10.867 80 8 81.645
TREND 14.5868 61.02 408 327.388 13.531 156.64 298 136.938

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 296
Figure 2. Photographs of Control rearing (A, B), Treatment rearing (C, D) and Treatment of leaf for freshness (E) before use as feed.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 297
THE OCCURRENCE OF GONATOCERUS LITORALIS (HALIDAY, 1833)(CHALCIDOIDEA: MYMARIDAE)
IN IRAN, WITH A NEW HOST RECORD
Majid Fallahzadeh* and John T. Huber**
*Islamic Azad University, Jahrom Branch, Department of Entomology, Fars, IRAN. E-mail: [email protected] ** Natural Resources Canada c/o AAFC, 960 Carling Ave., Ottawa, ON, K1A 0C6, CANADA.
[Fallahzadeh, M. & Huber, J. T. 2011. The occurrence of Gonatocerus litoralis (Haliday, 1833) (Chalcidoidea, Mymaridae) in Iran, with a new host record. Munis Entomology & Zoology, 6 (1): 297-300] ABSTRACT: Gonatocerus litoralis (Haliday, 1833) (Hymenoptera, Chalcidoidea, Mymaridae), reared from field-collected eggs of Zyginidia sohrab Zachvatkin, 1947 (Hemiptera, Cicadellidae) on wheat in Fars province, is added the faunal list for Mymaridae in Iran. Nine species of Mymaridae reported so far from Iran are listed and their distribution within the country is given. KEY WORDS: New host record, Hymenoptera, Mymaridae, Gonatocerus litoralis, Iran.
Fairyflies (Hymenoptera: Chalcidoidea: Mymaridae) are important, worldwide egg parasitoids of other insects. More than 1400 species and 100 genera are known (Noyes, 2009). Because fairyflies are so small, averaging less than 1 mm in length, they are rarely collected despite their abundance and thus, not surprisingly, little is known about them. Despite being a large country with various geographical regions the fauna of Iranian Mymaridae is very poorly known. Huber et al. (2009) listed the species recorded from Iran but no studies have focused specifically on that country. Here, we provide information on the distribution of Mymaridae in Iran, add one species to the published records, and provide a new host record. The mymarid genera of Iran will be similar to those found in UAE (Huber et al., 2009), but because Iran is a much bigger country with a greater diversity of habitat and climate more genera than the 13 found in UAE probably occur there. We expect that a large number of species remain to be discovered though many will already be described. Mymaridae have been used successfully in biological control (Huber, 1986) so further study of their occurrence, distribution and biology in Iran should be encouraged.
MATERIALS AND METHODS
Specimens representing two genera and species of Mymaridae, Anagrus sp. and Gonatocerus litoralis Haliday were reared from parasitized eggs of Zyginidia sohrab Zachvatkin, 1947 (Hemiptera, Cicadellidae) in a wheat field and preserved in ethanol. The single Anagrus specimen lacked both antennae so could not be identified to species. The specimens of Gonatocerus were critical point dried and one specimen was slide mounted in Canada balsam for identification using Mathews (1986). Voucher specimens are deposited in the Canadian National Collection of Insects, Ottawa, Ontario, Canada (CNC). Voucher specimens of the leafhopper host are deposited in the Department of Entomology, Faculty of Agriculture, Jahrom Islamic Azad University, Fars, Iran.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 298
Gonatocerus litoralis Haliday, 1833 Material examined: Fars province, Marvdasht, 4♀♀, 2♂♂ ex Zyginidia sohrab Zachvatkin, 1947 (Hemiptera: Cicadellidae) on Titicum aestivum L., 2. V. 2008, M. Fallahzadeh. Previous host records for G. litoralis are: Acocephalus sp. (Hemiptera: Cicadellidae) (Trjapitzin 1978 [as Lymaenon effusi Bakkendorf]) and Cicadula sexnotata (Hemiptera: Cicadellidae) (Thompson 1958, Baquero & Jordana 2003). Zyginidia sohrab is a new host record. Gonatocerus Nees, 1834 contains over 250 nominal species worldwide, 47 of which have been recorded in the Palearctic region (Huber, 1986; Noyes 2009). Only one species of Gonatocerus, G. ovicenatus, which belongs to the ater species group, was previously recorded from Iran (Huber 1988). Gonatocerus litoralis is the first species of the litoralis species group to be recorded for Iran.
List and distribution of Mymaridae in Iran
Anagrus (Anagrus) atomus (Linnaeus, 1767)
Iranian records. Khorasan Province (Triapitsyn, 1998), Tehran Province, Karaj (Walker et al., 1997) Fars Province, Kavar [incorrectly given as Kavar Province in Triapitsyn & Berezovskiy (2004)], Isfahan Province (Hesami, 2004, 2009). Hosts: Arboridia kermanshah Dlabola (Hemiptera: Cicadellidae) on grape (Triapitsyn, 1998), Circulifer tenellus (Baker, 1895) (Hemiptera: Cicadellidae) in sugar beet field (Walker et al., 1997), Progeny of, 4th generation, University of California, Riverside, USA, quarantine colony on eggs of Circulifer tenellus (Baker, 1895) (Hemiptera: Cicadellidae) (Triapitsyn & Berezovskiy, 2004), Arboridia kermanshah Dlabola (Hemiptera: Cicadellidae) on grape, and Edwardsiana rosae (L.) on rose, sweetbriar and blackberry (Hesami et al., 2004, 2009).
Anagrus (Anagrus) avalae Soyka, 1956 Iranian records. Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn & Berezovskiy (2004)]. Host: Unknown.
Anagrus (Anagrus) nigriceps (Smits van Burgst, 1914) Iranian records. Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn & Berezovskiy (2004)]. Host: Unknown.
Erythmelus (Erythmelus) flavovarius (Walker, 1846) Iranian records. Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn (2003)]. Host: Unknown.
Erythmelus (Erythmelus) israelensis Viggiani & Jesu, 1985 Iranian records. Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn (2003)]. Host: Unknown.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 299
Erythmelus (Parallelaptera) panis (Enock, 1909) Iranian records. West Azerbaijan Province, Orumieh (Akbarzadeh Shoukat, 1998), Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn (2003)]. Host: Stephanitis pyri (Fabricius, 1775) (Hemiptera, Tingidae) in apple orchard (Akbarzadeh Shoukat, 1998).
Erythmelus (Parallelaptera) rex (Girault, 1911) Iranian records. Tehran Province, Karaj [incorrectly given as Markazi Province in Triapitsyn (2003)]. Hosts [in North America]: Adelphocoris sp., Lygus hesperus Knight (Miridae).
Gonatocerus ovicenatus Leonard & Crosby, 1915 Iranian records. 30 km W. Zanjan (on Tabriz road), sweeping in poplar grove, 9.vi.1978, J.T. Huber (8♀♀, CNC), Bostanabad, 8.vi.1978, J.T. Huber (1♀, CNC) (Huber 1988). Host [in North America]: Rhytidodus decimasquartus (Schrank) (Cicadellidae).
Stephanodes reduvioli (Perkins, 1905) Iranian records. Tehran Province, Karaj, Shahdasht (Huber & Fidalgo, 1997). Hosts [in Hawaiian Islands]: Nabis spp. (Nasbidae) (Huber & Fidalgo 1997).
ACKNOWLEDGEMENTS
We wish to thank M. Taghizadeh (Agricultural and Natural Resources Center of Fars, Iran) for identification of the leafhopper specimens. The research was supported by Islamic Azad University, Jahrom Branch, Fars, Iran.
LITERATURE CITED Akbarzadeh Shoukat, G. 1998. The first report on the occurrence of the egg parasitoid of pear lace bug in Iran. Applied Entomology and Phytopathology, 66 (1/2): 44. Baquero, E. & Jordana, R. 2003. The genus Gonatocerus Nees (Hymenoptera Chalcidoidea Mymaridae) in corn fields of Navarra, North Spain. Redia, 85: 1-19. Hesami, S., Seyedoleslami, H. & Ebadi, R. 2004. Biology of Anagrus atomus (Hym., Mymaridae), an egg parasitoid of the grape leafhopper Arboridia kermanshah (Hom., Cicadellidae). Entomological Science, 7: 271-276. Hesami, S., Seyedoleslami, H. & Hatami, B. 2009. Impact of overwintering refugia of Anagrus atomus on egg parasitism of grape leafhopper, Arboridia kermanshah. Plant Protection Journal, Islamic Azad University, Shiraz Branch, 1 (1): 94-106. Huber, J. T. 1986. Systematics, biology, and hosts of the Mymaridae and Mymarommatidae (Insecta: Hymenoptera): 1758-1984. Entomography, 4: 185-243. Huber, J. T. 1988. The species groups of Gonatocerus Nees in North America with a revision of the sulphuripes and ater groups (Hymenoptera: Mymaridae). Memoirs of the Entomological Society of Canada, 141: 1-109. Huber, J. T. & Fidalgo, P. 1997. Review of the genus Stephanodes (Hymenoptera: Mymaridae). Proceedings of the Entomological Society of Ontario, 128: 27-63. Huber, J. T., Viggiani, G. & Jesu, R. 2009. Order Hymenoptera, family Mymaridae. In: van Harten, A. (Ed.), Arthropod Fauna of the UAE. Volume 2. Dar Al Ummah Printing, Adu Dhabi, UAE. 270–297. Matthews, M. J. 1986. The British species of Gonatocerus Nees (Hymenoptera: Mymaridae), egg parasitoids of Homoptera. Systematic Entomology, 11: 213-229.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 300
Noyes, J. S. 2009. Universal Chalcidoidea Database. Available from: www.nhm.ac.uk/entomology/chalcidoids/index.html (accessed 28 May 2009). Thompson, W. R. 1958. A catalogue of the parasites and predators of insect pests. Section 2. Host parasite catalogue, Part 5. Commonwealth Agricultural Bureaux, Commonwealth Institute of Biological Control, Ottawa, Ontario, Canada, 571 pp. Triapitsyn, S. V. 1998. Anagrus (Hymenoptera: Mymaridae) egg parasitoids of Erythroneura spp. and other leafhoppers (Homoptera: Cicadellidae) in North American vineyards and orchards: a taxonomic review. Transactions of the American Entomological Society, 124: 77-112. Triapitsyn, S. V. 2003. Review of the Mymaridae (Hymenoptera, Chalcidoidea) of Primorskii Krai: genus Erythmelus Enock, with taxonomic notes on some extralimital species. Far Eastern Entomologist, 126: 1-44. Triapitsyn, S. V. & Berezovskiy, V. V. 2004. Review of the genus Anagrus Haliday, 1833 (Hymenoptera: Mymaridae) in Russia, with notes on some extralimital species. Far Eastern Entomologist, 139: 1-36. Trjapitzin, V. A. 1978. 18. Mymaridae (Mymarids). In: Medvedev, G.S. (Ed.) Keys to the Insects of the European Part of the USSR. Hymenoptera. Vol. III. Part II. Academy of Sciences of the USSR. Institute of Zoology. 942-982. [1987 English translation by Amerind Publishing, New Dehli]. Walker, G. P., Zareh, N., Bayoun, I. M., & Triapitsyn, S. V. 1997. Introduction of western Asian egg parasitoids into California for biological control of beet leafhopper, Circulifer tenellus. Pan-Pacific Entomologist, 73 (4): 236-242.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 301
A NEW SPECIES OF PROCAMALLANUS (DENTICAMALLANUS) (NEMATODA: CAMALLANIDAE)
IN THORACOCHARAX STELLATUS (PISCES: GASTEROPELECIDAE) FROM ARGENTINA
Geraldine Ramallo*
* Instituto de Invertebrados, Fundación Miguel Lillo. Miguel Lillo 251. (4000) San Miguel de Tucumán, ARGENTINA. E-mail: [email protected] [Ramallo, G. 2011. A new species of Procamallanus (Denticamallanus) (Nematoda: Camallanidae) in Thoracocharax stellatus (Pisces: Gasteropelecidae) from Argentina. Munis Entomology & Zoology, 6 (1): 301-305] ABSTRACT: A new species, Procamallanus (Denticamallanus) ana n. sp., from Thoracocharax stellatus (Kner, 1858) intestine is described and illustrated. It represents the first reported species of Argentina and the second one of South America. It differs from its congener mainly because the presence of saw-like spiral thickenings of buccal capsule and outstanding basal ring in both sexes; females with 5-7 saw-like spiral thickenings in the inner surface of posterior half of capsule without tooth-like structure; males with two pairs of postanal papillae and one saw-like spiral thickening in the inner surface of capsule although with two single and conical teeth at its posterior half. KEY WORDS: Procamallanus (Denticamallanus), Nematodes parasites, freshwater fishes, Argentina.
Gasteropelecids members are widespread from Panama to the southern ending of South America. These fishes are able to perform short flights over water surface, usually as an escape strategy from aquatic predators. As far as know, they are the unique flying fishes caused by a propulsive force due to their greatly enlarged pectoral muscles (Monasterio de Gonzo, 2003).
Gasteropelecidae family encompasses three genus (Carnegiella Eigenmann, 1909, Gasteropelecus Eigenmann, 1909 and Thoracocharax Fowler, 1907) and nine species. Thoracocharax genus occurs in Argentina with only one species, T. stellatus (Kner 1858) (Monasterio de Gonzo, 2003; López et al., 2003).
Procamallanus Baylis, 1923 (Nematoda: Camallanidae) include several species described from freshwater and marine fishes. Many current authors consider those specimens with spiral ridges in the buccal capsule as distinctive genus: Spirocamallanus Olsen, 1952 (Moravec et al., 2000).
From freshwater fishes in the Neotropical Realm three Procamallanus subgenus were recorded: Procamallanus Baylis, 1923, Spirocamallanus Olsen, 1952 y Denticamallanus Moravec & Thatcher, 1997. The subgenus Procamallanus (Denticamallanus) comprise only one valid species, Procamallanus (Denticamallanus) dentatus Moravec & Thatcher, 1997 parasitic of Bryconops alburnoides (Cypriniformes) recorded in Brazil (Moravec, 1998).
The aim of this paper was to describe the first species of the subgenus Procamallanus (Denticamallanus) in Thoracocharax stellatus from Argentina and the second species from South America.
MATERIAL AND METHODS
In November 2006, 29 specimens of Thoracocharax stellatus (sex not specified; Lst = 3.5-5 cm) from Río Teuquito, (24º12'0.5‖ S; 62º54'13‖ W), Salta, Argentina were collected and examined for helminths. Fishes were 10% formalin

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 302
fixed before preservation in 70% ethanol. Body cavity was opened by a mid-ventral incision and the digestive tract was removed. Nematodes were cleared in lactophenol and examined under light microscope. Drawings were made with the aid of LEICA microscope. Quantitative descriptors of parasite populations were estimated based on definitions of Bush et al. (1997). Measurements are given in millimeters unless otherwise stated. For SEM (scanning electronic microscopy) examination specimens were dehydrated throughout an ethanol series, acetone and ether, coated with gold and examined in a Jeol JSM-35CF SEM.
Type specimens were deposited in the Colección Helmintológica Fundación Miguel Lillo (CH-N-FML), Miguel Lillo 251, (4000) San Miguel de Tucumán, Argentina. The fishes were deposited in the Colección Ictiológica Fundación Miguel Lillo (CI-FML).
RESULTS
Camallanidae Railliet & Henry, 1915 Procamallanus (Denticamallanus) ana sp. nov.
(Figures 1-14) General: Medium sized nematodes with transversaly striated cuticle. Mouth opening circular, surrounded six elevations, provide with eigth cephalic papillae arranged in two circlets and two lateral amphids (Figs. 3, 11). Orange-brown buccal capsule almost spherical in both sexes, with well development basal ring, with 5-7 in females and one saw-like spiral thickening but provided with two conical teeth at its posterior half in males. Buccal capsule surrounded by outer, colours-less hyaline layer (Figs. 4, 5). Muscular esophagus is considerably shorter than glandular one. Glandular esophagus length is two-fold in males and three-fold in females in relation to esophagus muscular length. Minute deirids situated just below nerve ring level (Figs. 2, 13); excretory pore somewhat posterior to nerve ring level (Fig. 1). Spicules small and equal (Fig. 10). Conical shaped tail with digitiform ending process for both sexes. Diagnosis. Males and females with saw-like spiral thickenings of bucal capsule (Fig. 12) and outstanding basal ring. Females with 5-7 saw-like spiral thickenings in the inner surface of posterior half of capsule without tooth-like structure; males with two pairs of postanal papillae and one saw-like spiral thickening in the inner surface of capsule although with two single and conical teeth at its posterior half. Male (n=12) measurements of holotype in parentheses. Body 2.80-4.00 (3.68) long, 0.13-0.18 (0.18) wide. Buccal capsule with basal ring; capsule 0.030-0.033 (0.033) long, 0.030-0.045 (0.036) wide; basal ring 0.003-0.004 (0.033) long, 0.020-0.030 (0.020) wide. Muscular esophagus 0.16-0.20 (0.18) long, 0.05-0.07 (0.06) wide; glandular esophagus 0.33-0.50 (0.49) long, 0.05-0.10 (0.08) wide. Length ratio of muscular and glandular esophagus 1:2.5 (2.7). Esophagus and buccal capsule length represents 19-24% (19%) of total body length. Deirids, nerve ring and excretory pore 0.10-0.16 (0.12), 0.10-0.13 (0.12) and 0.20-0.31 (0.20), respectively, from anterior extremity. Caudal alae absent. Caudal papillae: 2-3 pairs preanal, 1 pair adanal and 2 pairs postanal (Fig. 9). Spicules well sclerotized, short, equally long 0.040-0.046 (0.046). Gubernaculum absent. Phasmids not observed. Tail 0.13-0.18 (0.14) long with bluntly pointed tip. Female (n=12 gravid specimens) measurements of alllotype in parentheses. Body 17.00-23.40 (20.2) long, 0.31-0.53 (0.48) wide. Buccal capsule with basal ring;

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 303
capsule 0.037-0.050 (0.037) long, 0.043-0.063 (0.043) wide; basal ring 0.006 (0.006) long, 0.027-0.040 (0.033) wide. Muscular esophagus 0.25-0.32 (0.26) long, 0.10-0.12 (0.12) wide; glandular esophagus 0.93-1.15 (0.95) long, 0.09-0.20 (0.10) wide. Length ratio of muscular and glandular esophagus 1:3.1-4.1 (3.6). Esophagus and buccal capsule length represents 6-8% (6.2%) of total body length. Distance from anterior end of deirids, nerve ring, and excretory pore 0.14-0.20 (0.17), 0.12-0.17 (0.13) and 0.22-0.30 (0.22), respectively. Vulva near body centre, 8.00-11.00 (9.00) from posterior end (Fig. 6). Uterus containing numerous larvae 0.34-0.37 (0.36) long, 0.02 (0.02) wide (Fig. 7). Short tail bluntly pointed appendage. Tail length including caudal appendage about 0.14-0.19 (0.17), appendage 0.06-0.10 (0.07) long (Figs. 8, 14). Phasmids not observed. Type materials: Holotype: male CH-N-FML # 07475; allotype: female CH-N-FML # 07476, and paratypes (2 males, and 2 females) CH-N-FML # 07477. Type host: Thoracocharax stellatus (Kner, 1858) (Pisces, Gasteropelecidae), CI-FML # 4926, collected in 2006. Type locality: Río Teuquito (24º12'0.5‖S; 62º54'13‖W), Rivadavia Deparment, Province of Salta, Argentina. Prevalence: 86% (25/29). Thoracocharax stellatus harbored 42 specimens of Procamallanus (Spirocamallanus) ana sp. nov. Site of infection: Intestine. Mean intensity: 1.7 nematodes per fish. Etymology: The specific name is given in honor to Ana Valdez Fanjul, Elisa Fanjul´s first daugther, collegue, friend, and bird ecologist. Remarks: Procamallanus (Denticamallanus) dentatus specimens are larger than those of P. (D.) ana n. sp.. Main differences are observed in total length (22.78-35.77 vs. 17.00-23.40) and spicules length (0.078-0.110 vs. 0.043).
There are difficulties to observed vulva in both species, likewise Moravec (1998) vulva of only one specimen of P. (D.) dentatus was observed. A cause of this would be great uterus size for the presence of high quantity of embryos in different development stage (until first-stage larvae).
Procamallanus (Denticamallanus) dentatus and P. (D.) ana n. sp. differ for basal ring of buccal capsule (indistinct vs. well development), cuticle morphology (almost smooth vs. transversaly striated), postanal papillae number (three pairs vs. two pairs), adanal papillae (absent vs. one pair), spiral thickenings of bucal capsule (smooth vs. saw-like), inner surface of female capsule provided with (4-5 spiral tickening vs. 5-7 spiral tickening), low tooth-like structures (present vs. absent), inner surface of male capsule provided with (ten conical teeth vs. 2 teeth) and (without spiral tickening vs. with one spiral thickening).
In this paper we describe the first species of subgenus Procamallanus (Denticamallanus) from Argentina and the second species of the subgenus from South America.
ACKNOWLEDGEMENTS
Thanks to the Fundación Miguel Lillo for providing financial support for fish collections, particularly to Fabiana Cancino ichthyologist for providing the Thoracocharax stellatus specimens; to Andrés Ortiz for his technical support in figures digitalization; and to the Laboratory of Electron Microscopy staff, especially to Alberto Andrada Barone for their assistance.
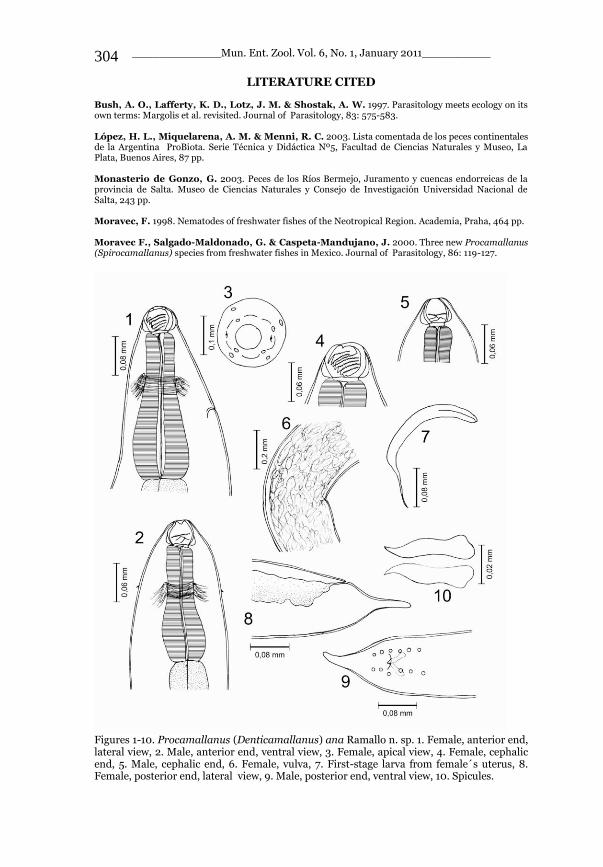
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 304
LITERATURE CITED Bush, A. O., Lafferty, K. D., Lotz, J. M. & Shostak, A. W. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. Journal of Parasitology, 83: 575-583. López, H. L., Miquelarena, A. M. & Menni, R. C. 2003. Lista comentada de los peces continentales de la Argentina ProBiota. Serie Técnica y Didáctica Nº5, Facultad de Ciencias Naturales y Museo, La Plata, Buenos Aires, 87 pp. Monasterio de Gonzo, G. 2003. Peces de los Ríos Bermejo, Juramento y cuencas endorreicas de la provincia de Salta. Museo de Ciencias Naturales y Consejo de Investigación Universidad Nacional de Salta, 243 pp. Moravec, F. 1998. Nematodes of freshwater fishes of the Neotropical Region. Academia, Praha, 464 pp. Moravec F., Salgado-Maldonado, G. & Caspeta-Mandujano, J. 2000. Three new Procamallanus (Spirocamallanus) species from freshwater fishes in Mexico. Journal of Parasitology, 86: 119-127.
Figures 1-10. Procamallanus (Denticamallanus) ana Ramallo n. sp. 1. Female, anterior end, lateral view, 2. Male, anterior end, ventral view, 3. Female, apical view, 4. Female, cephalic end, 5. Male, cephalic end, 6. Female, vulva, 7. First-stage larva from female´s uterus, 8. Female, posterior end, lateral view, 9. Male, posterior end, ventral view, 10. Spicules.
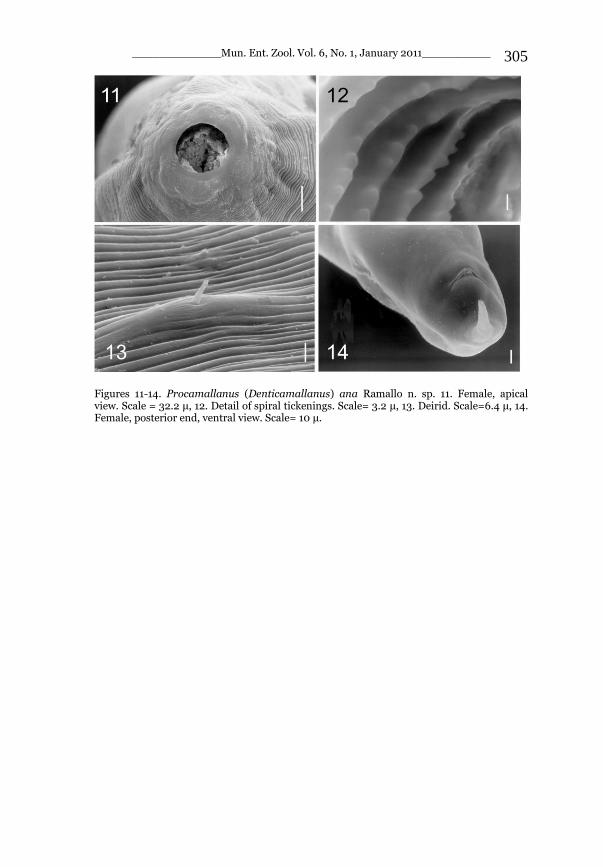
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 305
Figures 11-14. Procamallanus (Denticamallanus) ana Ramallo n. sp. 11. Female, apical view. Scale = 32.2 μ, 12. Detail of spiral tickenings. Scale= 3.2 μ, 13. Deirid. Scale=6.4 μ, 14. Female, posterior end, ventral view. Scale= 10 μ.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 306
COMPARISON OF THE EFFECT OF CUTTING OFF STAMENS OF POMEGRANATES AND RELEASING TRICHOGRAMMA
EMBRYOPHAGUM WITH ECTOMYELOIS CERATONIAE (LEP.: PYRALIDAE) IN SAVEH REGION, IRAN
Esmail Karami*, Abbas Mirabzadeh**,
Zahra Rafiei Karahroudi* and Saideh loni*
* Islamic Azad University, Arak branch, Young Researchers club, Arak, IRAN. ** Iran Scientific & Industrial Research Institution, IRAN. [Karami, E., Mirabzadeh, A., Karahroudi, Z. R. & Ioni, S. 2011. Comparison of the effect of cutting off stamens of pomegranates and releasing Trichogramma embryophagum with Ectomyelois ceratoniae (Lep. pyrallidae) in Saveh region, Iran. Munis Entomology & Zoology, 6 (1): 306-311] ABSTRACT: Pomegranate (Punica granatum) is one of the most important crops in Iran. Markazi province has a lot of wide Pomegranate‘s orchards. However, there are some pests that can damage these crops. Carob moth, Ectomyelois ceratoniae Zell. (Lep.: Pyralidae) is the most important pest, especially in Iran. The damage by this pest can at times reach up to 80%. To control it, three methods are used in Saveh region: a) Releasing the eggs of T. embryophagum for biological control of this pest.b) Cutting off stamens of pomegranate. c) Using the two methods together.The purpose of this research was to compare the efficiency of these three methods .To achieve this objective, a one-hectare-orchard was chosen for each treatment. The containing treatments: Releasing T. embryophagum, cutting off stamens of pomegranate, using the two methods together and control. Cutting off the stamens of pomegranate was done two times and the release of T. embryophagum was done10 times and once in every 10 days. Cutting off the stamens of pomegranate was done two times. 400 pomegranates were randomly picked out of each treatment (four repetitions of 100) and were kept for 2 months in the warehouse. The results of research showed that there was a significant difference between the treatments. Reduction of injury was observed in all of the treatments in comparison to the control. Infection amounts were 30% in control, 10.5% in releasing of T. embryophagum, 5.4% in cutting off the stamens of pomegranate and 4.5% in using the two methods together. There was no significant difference between cutting off the stamens of pomegranate and using the two methods together in the warehouse. KEY WORDS: Pomegranate, Cutting off stamens, Biological control.
Pomegranate is a native Iranian fruit and Iran is the first producer of this product in the world. This crop suffers from some pests such as Ectomyelois ceratoniae, Euzophera bigella and some mites that decrease its production (Sadat Akhavi, 2004). Carob moth (Ectomeylois ceratoniae) has been reported on many crops (Farzaneh, 1987; Mehrnejad, 1992). It is the most important pomegranate pest in Iran (Shakerai & Sadat Akhavi, 2003). The appearance of black spots on the fruit is the first symptom of infection and the begging of rotting and rancidity process of the fruit. The Damage caused by the pest varies remarkably depending on the species of the crops and the time of the year. The damage can at times reach up to 80%. This pest hibernates in different larve instars in the remaining fruits on the trees, fruits under the trees, under the bark of the trees and on other hosts (Ahmadian, 1997) .In early spring, the larvae gradually change into pupa and because pomegranates don‘t blossom at this time, the insects move to other hosts or die. Moths start appearing simultaneous with blossoming and fruiting and prolong until the end of June or beginning of July (Shakeri & Sadat Akhavi, 2003). Moths mate and lay on anthers, filaments and at times on the sepals (Farzaneh, 1987). First instar larvae hatches out from the eggs and stays in the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 307
crown of pomegranate. This pest penetrates into the fruit at its late second or third larve instar (Farzane, 1987), lowering the market value as a result of fungi attraction.
However, there are some methods that are used to control this pest in the country. According to Shojai, Esmaili and Najafi (1995) collecting infected pomegranates on and under the trees is the best way to control this pest. Noori et al. (1993), Nasrollahi (1998) and Mirkarimi (1999) contend that releasing T. embryophagum is a very effective method for controlling this pest. Shakeri and Sadat Akhavi (2004) consider using resistant species as the best method for controlling the pest. Mirkarimi (1999) states that filling the crown of the pomegranate with mud to prevent the pests from laying in the crown is an economic and safe method for controlling the pest.
Insecticides are not used against the insect in Iran because: Spraying with insecticides kills the natural enemies. The population of Tenuipalpus species increase after using insecticides. Pomegranate trees are very sensitive to poisons. Three methods are used to control this pest in Saveh including a) releasing T.
embryophagum, b) Cutting off the stamens of pomegranate and c) using the two methods together. However, orchardists don‘t know which method is more effective. In this research, the co-efficiency of these three methods will be tested in the warehouse.
MATERIAL AND METHOD
Four orchards, approximately each one about one hectare, were chosen in Saveh. Saveh is one of the cities of Markazi province in the central part of Iran. The orchards were all similar (in terms of orchards age, species of pomegranate trees, soil tissue and etc). In Saveh cultivated species are often Malas (Mirjalili, 2003). Therefore, we chose orchards in which sweet-and-sour species are cultivated.
The containing treatments: releasing T. embryophagum, cutting off stamens of pomegranate, using the two methods together. For control group the release of T. embryophagum was done10 times and once in every 10 days (Ahmadian, 1996; Mirkarimi, 1999). The wasps were purchased from insectariums of Plants Protection Office and transported to the orchards. Trichocards were kept dangling in the shades of the trees in the evening because sun rays decrease hatching eggs of parasitoid wasps. The wasps were put on the trichocards. They are kept at a temperature of 2-4 centigrade and transported to orchards at the time of release. Cutting off stamens of pomegranate was done two times. Pomegranate product flowers 3 times in a year. The first and the second time of flowering are more important. Since this method is expensive, cutting off the stamens of pomegranate is done only for the first and the second flowering. The machine used for cutting off the stamens was an electromotor with a brush on the tip, which rotates by a battery. When cutting off the stamens of pomegranate, the tip of the electromotor is put into pomegranate crowns. The rotating brush cuts out the anthers, destroying the haven for the insect to lay eggs. In the meantime, these two methods are used together in a separate garden.
For the comparison of the effect of the treatments in the warehouse, 400 pomegranates were randomly picked out of each treatment (four repetitions of 100) and were kept for 2 months in the warehouse). During this time period, the eggs of Carob moth hatch and penetrate into the pomegranates and infect the fruits. Infection symptoms usually appear in a month. After a month, infected

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 308
fruits are counted and separated from other fruits. After two months the infected fruits are counted again. Finally, the infection percentage for each treatment in each replication is determined.
SAS software version 6.12 was used for data analysis. Also, the test do in form of complete randomization design. The comparison of averages do dankan test.
RESULTS AND DISCUSSION
The analysis of the data shows that there are significant differences between the treatments. (F= 398.69; df= 3, 12; P<0.001).
The highest level of infection was observed in control treatment with 30%.There is a significant difference between control and other treatments.
The treatment of releasing T. embryophagum showed a 10.5% infection, while in comparison the control showed a 19.5% drop in infection. Therefore, releasing T. embryophagum showed lower infection in the warehouse. The treatment of releasing T. embryophagum showed a significant difference with control and using the two methods together at 0.05. However, no significant difference was observed in the treatment of cutting off stamens of pomegranate.
The treatment of cutting off stamens of pomegranate with 5.5% infection showed a 24.5% infection reduction in comparison to control. No significant difference was found between cutting off stamens of pomegranate and using the two methods together. But a significant difference was found between this treatment with control and releasing T. embryophagum. It showed that using the two methods together decreases the infection more. But there wasn‘t a significant difference in the reduction of injury in comparison to using the two methods together. Finally, using the two methods together showed a 4.75% infection which means that this method was the best method in lowering the infection.
There wasn‘t any treatment in the warehouse. The analysis of the warehouse
data showed that using the two methods together was the best one in lowering the infection. The second effective method was cutting off stamens of pomegranate and releasing T. embryophagum. Therefore, the three methods of control studied in the research were effective in lowering the injury. Comparison effect of time and treatments in control of Carob moth in the warehouse:
Pomegranates were counted two times in the warehouse and the data was analyzed using factorial design with randomization. The results of the analysis showed that there are significant differences between the treatments in two times. (F= 481.17,847.45,241.79; df= 3, 1,3; P<0.001).
The highest infection percentage 10.31% was observed at the end of October (first time of pomegranates counting). We have seen 2.25% infection at the end of November that showed 8.06% infection reduction in comparison to first time of pomegranates counting. There was a significant difference between the two times.
The comparison of the two time periods showed the highest level of infection in the first time of data collection in all the treatments and lower percent of infection in the second time of data collection. So, most insect appearance was observed a month after warehousing. The minimum level of infection was observed in using the two methods together and the second time of data collection. Most of the eggs hatch in a month and damages the pomegranates in the warehouse.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 309
According to Kashkooli and Eghtedar (1975), the time span of egg incubation is 8-9 days and the time span of larvae appearance is 8-20 days, altogether making 16-29 days. For this reason, it takes a month for insects to appear.
The temperature in the warehouse is usually 25 5. Therefore, the maximum, minimum and optimum temperatures are 30, 20 and 25 respectively. When larvae grow, they penetrate into the fruit and feed on the core of the pomegranate, inside part of the pomegranate and outside the fruit bark. In a few days, some spots start to appear on the fruits, which are the symptoms of infection (Kashkuli and Eghtedar, p.54). Therefore, when symptoms appear on the fruit, it means that the larvae are in their third or fourth instar.
Based on Yusefi‘s (1381) study concerning the effect of temperature on life steps and act of the insect, in 25C°(the average of temperature in the warehouse),the average of eggs incubation is 6.7 days and the duration of larve stage is 33.4 days. As a result, this insect needs 40.1 days to complete incubation of eggs and larva stages. As already mentioned, when symptoms appear on the fruit, the larvae are in their third or fourth instar. Since no larve fifth instar has been reported in the references, subtracting larve fifth instar from 40.1, the symptoms can be observed in more than 30 days and less than 40 days. Also because eggs laid on stamens happen at different times, like hatching and the appearance of larvae, eggs laid earlier in the garden, hatch in the first month of keeping in the warehouse. Naturally in eggs laid later, larvae appear in the second month. Only the eggs that were laid some days before being kept in the warehouse showed symptoms of damage in the second month. Therefore, most of the eggs hatch in first month after being kept in the warehouse.
Farmers and exporters must sell pomegranates in a month. If they wish to keep them more than a month, they should keep them in a cold place because low temperature delays hatching of eggs and larvae appearance stages.
ACKNOWLEDGEMENTS
This research supported by the Iran Scientific & Industrial Research Institution.Autors thank Eng. Ghahremani for all of his helps in Saveh.
LITERATURE CITED Ahmadian, H. 1993. Final report from efficiency investigation Trichogramma spp. against Ectomyelois ceratoniae. Agriculture Researches Center of Yazd (plant diseases and pests part). 136 pp. Ahmadian, H. 1996. Final report from economic investigation biological control against Ectomyelois ceratoniae by Trichogramma embryophagum. Agriculture researches center(pests and plant disease part). 25 pp. Ahmadian, H. 1998. Wide application of Trichogramma against Ectomyelois ceratoniae in Yazd Province. Agriculture Researches Center of Yazd (Plant Diseases and Pests part). 27 pp. Farzaneh, A. 1987. Ectomyelois ceratoniae in Iran. Papers of First symposium of pomegranate problems investigation in Iran. University Jehad of Agriculture College and Natural Resource of Tehran. 1: 17-19. Mehrnejad, M. 1992.Biology of Carob moth (new pest on pistachio) in Rafsanjan. Plant desease and pests publication, (1,2). Mirabzadeh, A. 2006. First year report integrated control Ectomyielois ceratoniae in Yazd. 128 pp. Mirjalili, A. 2002. Knowing of pomegrana. Publication of Agriculture Education. 235 pp.
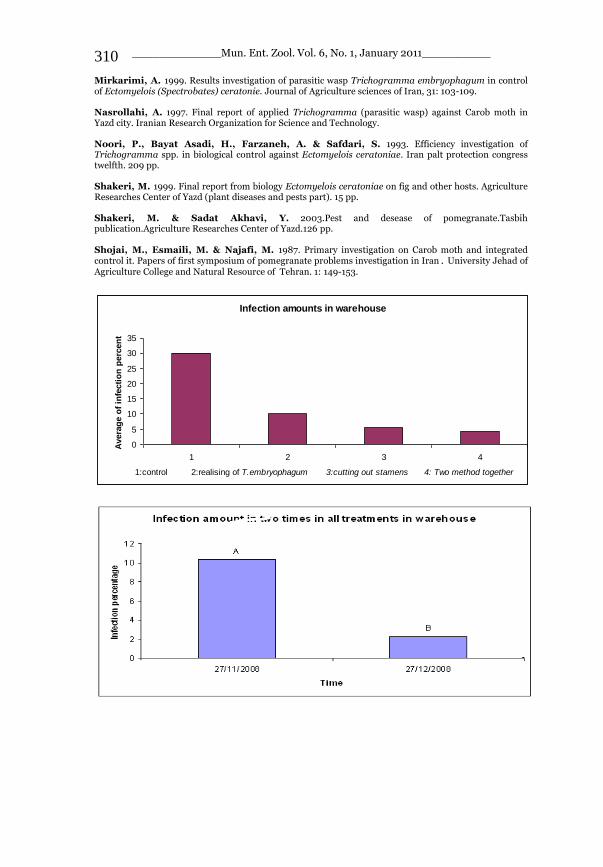
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 310
Mirkarimi, A. 1999. Results investigation of parasitic wasp Trichogramma embryophagum in control of Ectomyelois (Spectrobates) ceratonie. Journal of Agriculture sciences of Iran, 31: 103-109. Nasrollahi, A. 1997. Final report of applied Trichogramma (parasitic wasp) against Carob moth in Yazd city. Iranian Research Organization for Science and Technology. Noori, P., Bayat Asadi, H., Farzaneh, A. & Safdari, S. 1993. Efficiency investigation of Trichogramma spp. in biological control against Ectomyelois ceratoniae. Iran palt protection congress twelfth. 209 pp. Shakeri, M. 1999. Final report from biology Ectomyelois ceratoniae on fig and other hosts. Agriculture Researches Center of Yazd (plant diseases and pests part). 15 pp. Shakeri, M. & Sadat Akhavi, Y. 2003.Pest and desease of pomegranate.Tasbih publication.Agriculture Researches Center of Yazd.126 pp. Shojai, M., Esmaili, M. & Najafi, M. 1987. Primary investigation on Carob moth and integrated control it. Papers of first symposium of pomegranate problems investigation in Iran University Jehad of Agriculture College and Natural Resource of Tehran. 1: 149-153.
Infection amounts in warehouse
0
5
10
15
20
25
30
35
1 2 3 4
Avera
ge o
f in
fecti
on
perc
en
t
1:control 2:realising of T.embryophagum 3:cutting out stamens 4: Two method together
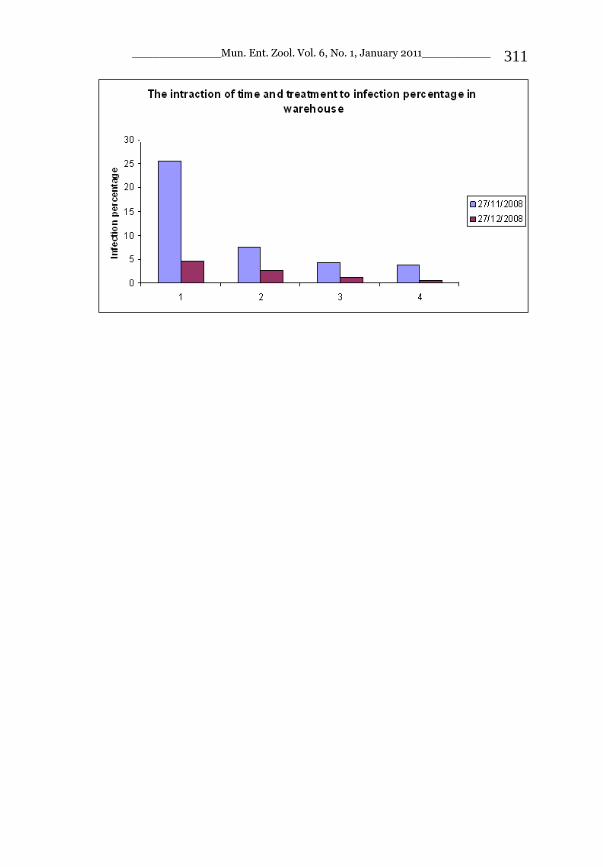
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 311

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 312
BEAUVERIA BASSIANA (HYPHOMYCETES: MONILIALES) INFECTION DURING ECDYSIS OF SILKWORM BOMBYX
MORI (LEPIDOPTERA: BOMBYCIDAE)
Kuniyil Chandrasekharan* and Byrappa Nataraju * Silkworm Pathology Section, Central Sericultural Research and Training Institute, Mysore, Karnataka, INDIA. [Chandrasekharan, K. & Nataraju, B. 2011. Beauveria Bassiana (Hyphomycetes: Moniliales) Infection During Ecdysis Of Silkworm Bombyx Mori (Lepidoptera: Bombycidae). Munis Entomology & Zoology, 6 (1): 312-316] ABSTRACT: Mulberry silkworm, Bombyx mori is the most commercially exploited insect which is domesticated worldwide for the production of silk cocoons. Domestication and constant rearing for the past several thousands of years made the silkworm highly susceptible to different pathogens. Different micro-organisms such as viruses, bacteria, fungi and microsporidia cause infectious diseases in silkworm. White muscardine caused by the fungal pathogen Beauveria bassiana is one of the most common and devastating diseases of silkworm. All the life stages of silkworm are susceptible to B. bassiana and its infection to silkworm during ecdysis (moulting) is studied and discussed in the present paper. The silkworms were inoculated with the pathogen at 1 × 105 conidia/ml dose at different durations (18, 12 and 6 hours) before and after moult. Inoculation was also done during the moult. Mortality was 100% when the inoculation was done 12 hours before and 6 hours after moulting. Mortality was significantly less when the larvae were inoculated during moulting or 6 hours before moult. The present findings indicate the inability of B. bassiana to invade the silkworm during its moulting period. Skin casting and related processes during ecdysis may be preventing the fungus to enter inside the haemocoel of silkworm to cause infection and mortality. KEY WORDS: Beauveria bassiana, silkworm, Bombyx mori, moulting.
Among the diseases of silkworm, white muscardine caused by B. bassiana inflicts heavy economic loss to the sericulturists in India. The climatic condition in the tropics is congenial for survival, infection and spread of this disease. All the life stages of the silkworm, viz. egg, larvae, cocoon, pupae and moth are found susceptible to the fungal pathogen, Beauveria bassiana (Chandrasekharan, 2008) The degree of susceptibility varied based on the age of individual stages as well as the dosage of the pathogen.
All the larval instars are susceptible to B. bassiana and the susceptibility varied among the instars. B. bassiana infects cutaneously and when the silkworm grows the integument becomes comparatively more resistant as far as the fungal pathogen is concerned. The integument of insects by itself is a strong barrier for many microbes such as viruses, bacteria and protozoa. It is not only a water proof physical barrier but also contains chemicals that inhibit the growth and penetration of micro organisms (David, 1967). The waxy epicuticular layer of silkworm integument contains free medium chain saturated fatty acids, presumably caprylic, capric and linoleic acids that inhibit the invasion of the fungi. Higher amount of free medium chain fatty acids is observed in the cuticle of silkworm strains resistant to Aspergillosis (Koidsumi, 1957). Lipids in the epicuticle of silkworm, Bombyx mori inhibit the invasion of B. bassiana and Paecilomyces fumosa-roseus. The living integument and exuviae of resistant Chinese silkworm race had a greater anti-fungal action than that from susceptible Japanese strain (Koidsumi & Wada, 1955).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 313
Fifth instar larvae are reported to be more susceptible than other instar larvae (Steinhaus, 1949; Reddy, 1978). Studies of Mukerji (1919) indicated that moulting larvae were more susceptible than other stages of instar. Susceptibility varies according to different stage of development of silkworm. Within the same instar, newly ecdysed larvae are more susceptible than those approaching the moult. In fifth instar, the infection decreases as the silkworm grows but ripe larvae and early pupa again shows increased susceptibility.
The most common route of host invasion is through the external integument, although infection through the digestive tract is possible (Gabriel, 1959). Yanagita (1987) studied on the oral infection of silkworm with B. bassiana and Yanagita & Iwashita (1987) revealed the histology of silkworm inoculated orally with the fungus. Usually the conidia attach to the cuticle, germinate and penetrate the cuticle (Boucias et al., 1988; Lefebvre, 1934). The surface ultra structure of B. bassiana infecting silkworm was studied by Vineet Kumar et al. (1994) and the infection process such as germination, penetration, and invasion was also studied further by them (Vineet Kumar et al., 1999). Once in haemocoel, the mycelium ramifies throughout the host, forming yeast like hyphal bodies often referred to as blastospores (Masera, 1952).
MATERIALS AND METHODS
Infection of B. bassiana during ecdysis was studied during all the four moults of CSR2 CSR4 silkworm hybrid larvae. B. bassiana conidia were harvested from the culture plates by using sterilized loop and suspended in sterilized distilled water which formed the stock inoculum. The concentration of conidia in the stock inoculum was determined using Neubauer haemocytometer. The required conidial suspensions i.e. 1 × 105 conidia/ml was prepared by diluting the original stock suspension in sterilized distilled water. The silkworms were inoculated 18, 12 and 6 hours before moult (HBM) and 18, 12 and 6 hours after moult (HAM). Inoculation was also done during the moult. Hundred larvae were kept in each replication and three replications for each treatment were maintained. Mortality due to white muscardine was recorded daily and the progressive mortality was calculated.
RESULTS AND DISCUSSION
Some insects are susceptible to diseases only at certain stages of the life cycle. In the case of fungal infections, the reports are varied. Certain fungi infect all stages of host insects, but in some cases the egg and pupal stages are more resistant than the larval and adult stages. The larval stages may show maturation immunity. The critical period in the development of the larvae to infectious diseases is shortly after moulting. When its' newly regenerated gut epithelium and newly formed integument are more susceptible to the fungal penetration (Muller Kogler, 1967). The amino acid composition in the larval surfaces varies among instars and with the time after moulting. Amines and peptides are also present but do not inhibit the germination of fungi. The fungal cell wall components are also reported to activate immune response in silkworm (Bidochka & Hajek, 1998).
Studies on the infection during the moulting period of larvae indicated some interesting results. Just prior to and just after moulting the larvae succumbed heavily to B. bassiana but during moulting the larvae were found less infected. The progressive mortality due to white muscardine is provided in Table 1. 100% mortality was recorded when the larvae were inoculated more than 12 hours

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 314
before moulting and more than 6 hours after moulting. The mortality after inoculating B. bassiana during moulting time was 1.00, 9.33, 30.33 and 39.33% in the fourth, third, second and first moults respectively. Inoculation during 6 hours before the third and fourth moult resulted in significantly less mortality of 60.67 and 31.33%, respectively (Figures 1a -1d).
Our results are in contrary to the studies of Mukerji (1919) who indicated moulting larvae as more susceptible than other stages of instar. He also reported that the susceptibility varied according to different stages of the silkworm. Within the same instar, newly ecdysed larvae are more susceptible than those approaching the moult. In fifth instar, the infection decreases as the silkworm grows but ripe larvae and early pupae again show increased susceptibility. The present finding indicates the inability of the fungus to invade the integument during moulting as it would be in the process of skin casting. The germinated conidia on the outer old skin may be inactivated during the process of ecdysis and ultimately they might be pushed off from the vicinity of silkworm skin. During moulting, the multiplication and development of inclusion bodies of NPV or CPV viruses were nearly inhibited until after moulting. Early instars silkworms are comparatively more susceptible to infection by microbial pathogen and it decreased with larval ageing from first to fourth instars (Aruga & Watanabe, 1964; Kobara et. al., 1967).
ACKNOWLEDGEMENTS
The first author wishes to express his gratitude to the Director, Central Sericultural Research and Training Institute, Mysore for providing laboratory facilities. The authors are also thankful to the colleagues in Silkworm Pathology section of the Institute.
LITERATURE CITED Aruga, H. & Watanabe, H. 1964. Resistance to per os infection with cytoplasmic polyhedrosis virus in the silkworm, B. mori L. Journal of Insect Pathology, 6: 387-394. Bidochka, M. J. & Hajek, A. E. 1998. A nonpermissive entomophthoralean fungal infection increases activation of insect prophenoloxidase. Journal of Invertebrate Pathology, 72 (3): 231-238. Boucias D. G., Pendland J. C. & Latge, J. P. 1988. Nonspecific factors involved in attachment of entomopathogenic Deuteromycetes to host insect cuticle. Applied and Environmental Microbiology, 54: 1795-1805. Chandrasekharan, K. 2008. Studies on the management of white muscardine disease in the silkworm, Bombyx mori L. Ph.D Thesis, University of Mysore, Mysore p. 313. David, W. A. L. 1967. The physiology of insect integument in relation to the invasion of pathogens. In: Insects and Physiology. J. W. L. Beament and J. E. Treherne (eds), Oliver and Boyd. London. pp. 17-35. Gabriel, B. P. 1959. Fungus infection of insects via the alimentary canal. Journal of Insect Pathology, 1: 319-330. Kobara, R., Aruga, H. & Watanabe, H. 1967. Effect of larval growth on the susceptibility of silkworm Bombyx mori L. to the cytoplasmic polyhedrosis virus. Journal of Sericultural Science Japan, 36: 165-168. Koidsumi, K. & Wada, Y. 1955. Studies on the antimicrobial function of insect lipids. IV. Racial differences in the antifungal activity in the silkworm integument. Japanese Journal of Applied Zoology, 20: 184-190. Koidsumi, K. 1957. Antifungal action of cuticular lipids in insects. Journal of Insect Physiology, 1: 40-51.
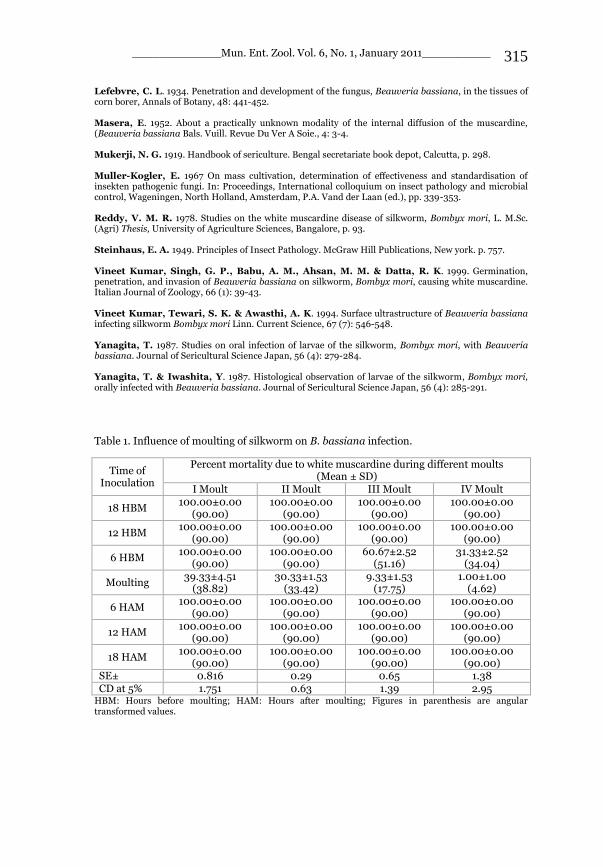
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 315
Lefebvre, C. L. 1934. Penetration and development of the fungus, Beauveria bassiana, in the tissues of corn borer, Annals of Botany, 48: 441-452. Masera, E. 1952. About a practically unknown modality of the internal diffusion of the muscardine, (Beauveria bassiana Bals. Vuill. Revue Du Ver A Soie., 4: 3-4. Mukerji, N. G. 1919. Handbook of sericulture. Bengal secretariate book depot, Calcutta, p. 298. Muller-Kogler, E. 1967 On mass cultivation, determination of effectiveness and standardisation of insekten pathogenic fungi. In: Proceedings, International colloquium on insect pathology and microbial control, Wageningen, North Holland, Amsterdam, P.A. Vand der Laan (ed.), pp. 339-353. Reddy, V. M. R. 1978. Studies on the white muscardine disease of silkworm, Bombyx mori, L. M.Sc. (Agri) Thesis, University of Agriculture Sciences, Bangalore, p. 93. Steinhaus, E. A. 1949. Principles of Insect Pathology. McGraw Hill Publications, New york. p. 757. Vineet Kumar, Singh, G. P., Babu, A. M., Ahsan, M. M. & Datta, R. K. 1999. Germination, penetration, and invasion of Beauveria bassiana on silkworm, Bombyx mori, causing white muscardine. Italian Journal of Zoology, 66 (1): 39-43. Vineet Kumar, Tewari, S. K. & Awasthi, A. K. 1994. Surface ultrastructure of Beauveria bassiana infecting silkworm Bombyx mori Linn. Current Science, 67 (7): 546-548. Yanagita, T. 1987. Studies on oral infection of larvae of the silkworm, Bombyx mori, with Beauveria bassiana. Journal of Sericultural Science Japan, 56 (4): 279-284. Yanagita, T. & Iwashita, Y. 1987. Histological observation of larvae of the silkworm, Bombyx mori, orally infected with Beauveria bassiana. Journal of Sericultural Science Japan, 56 (4): 285-291.
Table 1. Influence of moulting of silkworm on B. bassiana infection.
Time of Inoculation
Percent mortality due to white muscardine during different moults (Mean ± SD)
I Moult II Moult III Moult IV Moult
18 HBM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00)
12 HBM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00)
6 HBM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 60.67±2.52
(51.16) 31.33±2.52
(34.04)
Moulting 39.33±4.51
(38.82) 30.33±1.53
(33.42) 9.33±1.53
(17.75) 1.00±1.00
(4.62)
6 HAM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00)
12 HAM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00)
18 HAM 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) 100.00±0.00
(90.00) SE± 0.816 0.29 0.65 1.38 CD at 5% 1.751 0.63 1.39 2.95
HBM: Hours before moulting; HAM: Hours after moulting; Figures in parenthesis are angular transformed values.
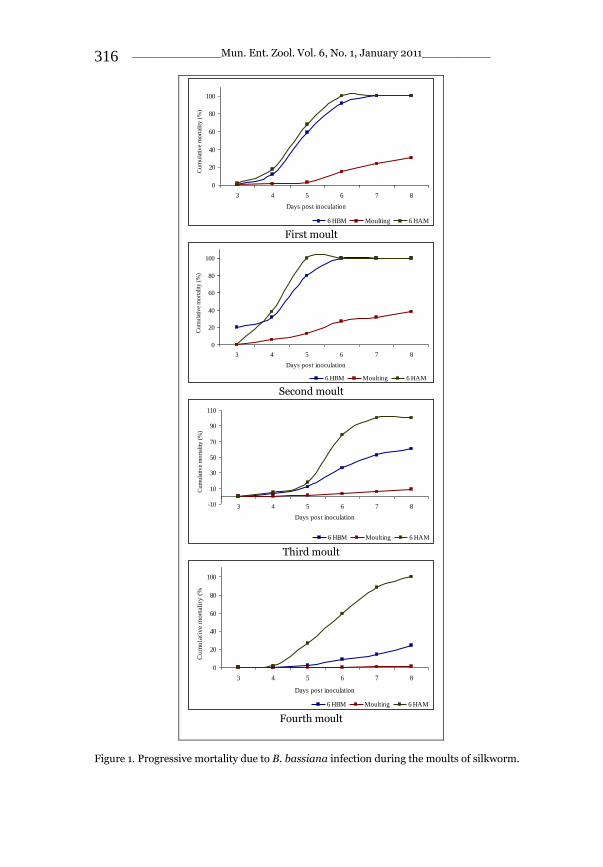
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 316
0
20
40
60
80
100
3 4 5 6 7 8
Days post inoculationC
um
ula
tive
mort
ality (
%)
6 HBM Moulting 6 HAM
First moult
0
20
40
60
80
100
3 4 5 6 7 8
Days post inoculation
Cum
ula
tiv
e m
ort
ality
(%
)
6 HBM Moulting 6 HAM Second moult
-10
10
30
50
70
90
110
3 4 5 6 7 8
Days post inoculation
Cum
ula
tiv
e m
ort
ality
(%
)
6 HBM Moulting 6 HAM
Third moult
0
20
40
60
80
100
3 4 5 6 7 8
Days post inoculation
Cu
mu
lati
ve m
ort
ali
ty (
%)
6 HBM Moulting 6 HAM
Fourth moult
Figure 1. Progressive mortality due to B. bassiana infection during the moults of silkworm.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 317
A GENUS WITH TWO SPECIES AS NEW RECORDS FOR IRANIAN HOVER FLY FAUNA (DIPTERA: SYRPHIDAE)
Samad Khaghaninia*, Reza Farshbaf Pour Abad*
and Süleyman Saribiyik**
* Dept. of Plant Protection, Faculty of Agriculture, University of Tabriz, 51664, Tabriz, IRAN. E-mail: [email protected] ** Education Faculty, Kastamonu University, Kastamonu, TURKEY. [Khaghaninia, S., Pour Abad, R. F. & Sarıbıyık, S. 2011. A genus with two species as new records for Iranian hover fly fauna (Diptera: Syrphidae). Munis Entomology & Zoology, 6 (1): 317-320] ABSTRACT: The genus Anasimyia Schiner, 1864 is recorded for the first time from Iran. Nine and six specimens were identified as Anasimyia contracta Claussen & Trop, 1980 and Anasimyia transfuga (Linnaeus, 1758) respectively. The related key besides their diagnosis characters and photos are provided. KEY WORDS: Anasimyia, Anasimyia contracta, Anasimyia transfuga, Syrphidae, Qurigol lake, Iran.
The genus Anasimyia was described by Schiner (1864) with Musca transfuga Linnaeus, 1758 designated as the type species (Peck, 1988). This genus belonged to Milesiinae subfamily and Eristalini tribe which almost break down into two groups: the Helophilus group with a distinctly striped thorax (including Anasmyia, Lejops and Parhelophilus) and the remaining genera without strips on the thorax. However, in Anasimyia the build is narrow and the appearance is dark, the thoracic stripes being narrow (occasionally absent) and the grey or yellowish marking on the abdomen being restricted (Stubbs & Fulk, 2002).
Anasimyia may be treated as a subgenus of Lejops in the future, but there are opposing opinions. Certainly the two genera are close in a number of characteristics such as the narrow build, the double-banded hind tibia and the heavily dusted sternites. The genus has been revised by Claussen & Torp (1980) and the position in Ireland and Great Britain reviewed by Speight (1981) and Stubbs (1981) respectively. Adults will be found at their aquatic breeding sites, often in association with tall emergent vegetation.
Recently, the check list of Iran hover flies was reviewed by Dousti & Hayat (2006) which shown no record of this genus from Iran.
MATERIAL AND METHODS
The specimens were collected from west of Qurigol lake on reed beds and bulrush using common entomological net (Fig. 1). Qurigol is a small, about 200 hectares expanse, fresh to brackish lake with associated marshes in the steppe uplands of northwestern Iran. There are extensive areas of reed beds. It is situated about 40 km East-Southeast of Tabriz city. The surrounding area is semi-arid, and there is wheat cultivation on the west and damp grasslands on the southwest. The geographical coordinates are 37° 55' N; 46° 42' to 46° 44' E. The specimens were identified based on valid keys such as Claussen & Torp (1980), Bei- Bienko (1988), Stubbs & Falk (2002) and Speight (2010).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 318
RESULTS
Anasimyia Schiner, 1864 Diagnostic characters: Hind tibia with tow dark bands (sometimes faint), yellow in between. Antennae reddish-orange, at least in part. Sternites densely grey-dusted, unlike parhelophilus (Fig. 2).
Anasimyia contracta Claussen & Trop, 1980 (Claussen et Torp Pedersen, 1980): Mitt. Zool. Mus. Univ. Kiel, 1(4): 4 (Anasimyia). Type-locality: ―Denmark, WJS: Graerup Langso vest‖. Material examined: 9 specimens (5♂♂, 4♀♀): West of Qurigol lake; 37°55' N 46°41' E, 1911 m , 4 Aug. 2009 (Collected by S. Khaghaninia, Deposited at Insect Museum of Tabriz University). Diagnostic characters: This is a dark species which is especially slender in the male. The species name is derived from the contracted margins on tergite 2 which are quite distinct. The markings are narrow and strongly angled hooks, but the outer limb can be reduced leaving the inner portions as small colored spots. Wing length 5- 7.25 mm (Fig. 3). Flowers visited: white umbellifers; Myosotis, Potentilla palustris, Ranunculus, Sorbus aucuparia. Note: Larvae are of the ‗long-tailed‘ type, living in ponds and ditches where decaying vegetation is abundant, especially in fen (Imhof, 1979). There is a very close association with bulrush at the margin of ditches, ponds and lakes. The adults rarely more than a few meters from margins of ponds, where they fly low over the water and settle in the sun, usually on foliage of large-leaved emergent vegetation, such as Menyanthes; they also fly in and out of stands of sedges and reeds, making their movements very difficult to follow (Stubbs, 1981; Speight, 2010). Distribution: Europe Sweden, Finland, Great Britain, Denmark, Netherlands, Belgium, German Federal Republic, Poland, Hungary, Ireland; Scandinavia south to the Mediterranean basin; USSR: Estonian, Latvian, Byelorussian, Ukrainian (Peck, 1988; Ball & Morris, 2000; Speight, 2010).
Anasimyia transfuga (Linnaeus, 1758) (Linnaeus, 1758): Syst., Ed. 10, 1: 594 (Musca). Type-locality: ―Svecia‖ (Sweden). Material examined: 6 specimens (4♂♂, 2♀♀): West of Qurigol lake; 37°55' N 46°41' E, 1911 m , 4 Aug. 2009 (Collected by S. Khaghaninia, Deposited at Insect Museum of Tabriz University). Diagnostic characters: This dark hoverfly has steeply inclined hooked marking, like a tick, with the inner limb strongly oblique. Similar marking are found in contracta, but transfuga has the lateral margins of tergite 2 straight. Wing length 6.5- 7 mm (Fig. 4). Flowers visited: white umbellifers, Caltha palustris, Ranunculus repens, Sonchus arvensis, Sorbus aucuparia, Taraxacum. Note: Accumulated data now show it to be less frequent than contracta and, though it can occur alongside that species, it shows no obvious association with bulrush, preferring the pollen of other tall emergent pond-side vegetation such as bur-reeds (Sparganium sp.) and sweet-grass, the adults, settling on emergent stems etc., often where the water is up to 1m deep (Imhof, 1979; Speight, 2010). Distribution: Europe: Sweden, Finland, Great Britain, Denmark, Netherlands, Belgium, Germany, Poland, Hungary, Norway, Ireland, Czechoslovakia, Austria, France, Yugoslavia, Romania, Turkey, USSR: Estonian, Latvian, Lithuanian, Russian, Byelorussian, Ukrainian, Moldavian, Georgian, Azerbaijan, Armenian, as far as Southern parts of Central Siberia (Tuva) (Peck, 1988; Ball & Morris, 2000; Reemer & Smit, 2007; Sarıbıyık, 2008; Speight, 2010).
ACKNOWLEDGEMENTS
We would like to thank University of Tabriz for supporting this work under the Research Grant Contract No. 27.557.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 319
LITERATURE CITED Ball, S. G. & Morris, R. K. A. 2000. Provisional atlas of British hoverflies (Diptera, Syrphidae). Biological Record Centre, CEH Monks Wood. 167 pp. Bei- Bienko, G. 1988. Keys to the insects of the European part of the USSR. Volume V. Diptera and Siphonaptera. Part II. Smithonian Institution Libraries and the National Science Foundation Washington, D.C. 10- 148. Claussen, L. & Trop, E. 1980. Untersuchungen uber vir europaische Arten der Gattung Anasimyia Schiner, 1864 (Insecta, Diptera, Syrphidae). Mitt. Zool. Mus. Kiel., 1: 1- 11. Dousti, A. F. & Hayat, R. 2006. A catalogue of the Syrphidae (Insecta: Diptera) of Iran. J. Entomol. Res. Soc., 8 (3): 5-38. Imhof, G. 1979. Arthropod communities connected with phragmites. Monographia Biol., 37: 389-405. Peck, L. V. 1988. Family Syrphidae. PP. 11- 230 in Soos, A. (Ed.) Catalogue of Palearctic Diptera. Vol. 8, 363 PP. Akademiai Kiado, Budapest. Reemer, M. & Smit, J. T. 2007. Some hoverfly records from Turkey (C: Syrphidae), Volucella, 8: 135-146. Speight, M. C. D. 1981. The Irish Anasimyia species including a key and first records of A. contracta (Diptera: Syrphidae). Ir. Nat. J., 20: 229- 234. Speight, M. C. D. 2010. Species accounts of European Syrphidae (Diptera) 2010. In: Speight, M. C. D., Castella, E., Sarthou, J.-P. and Monteil, C. (eds.). Syrph the Net, the database of European Syrphidae, vol. 59, 285 pp., Syrph the Net publications, Dublin. Stubbs, A. E. 1981. Anasimyia contracta Claussen & Trop, 1980 and A. interpuncta (Harris, 1776) (Diptera: Syrphidae) in Britain. Proc. Trans. Br. Ent. Nat. Hist. Soc., 14: 10-14. Sarıbıyık, S. 2008. Contributions to the Syrphidae fauna of Turkey (Diptera: Syrphidae). Entomological News, 119 (5): 501-508. Stubbs, A. E. & Falk, S. J. 2002. British hover flies. An illustrated identification guide. Pub. The british Entomology and Natural History Sosiety, Reading, UK.
Figure 1. Location of sampling points on satellite image (SPOT) of Qurigol lake environment.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 320
Figure 2. Anasimyia Schiner, 1864, a: hind leg, b: abdomen; sternal view.
Figure 3. Anasimyia contracta, a: male; dorsal view, b: female; the same view, c and d: the head; female and male at lateral view.
Figure 4. Anasimyia transfuga, a: male; dorsal view, b: female; the same view, c and d: the head; female and male at lateral view.
a b
a b
c
d
a b
c
d

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 321
A NEW SPECIES OF THE GENUS CYRTARACHNE (ARANEAE: ARANEIDAE) FROM WESTERN GHATS, INDIA
K. Sunil Jose*
* Department of Zoology, Deva Matha College, Kuravilangad, Kerala, INDIA-686633. E-mail: [email protected] [Jose, K. S. 2011. A new species of the genus Cyrtarachne (Araneae: Araneidae) from Western Ghats, India. Munis Entomology & Zoology, 6 (1): 321-324] ABSTRACT: A new species namely Cyrtarachne keralensis sp. nov. (Araneidae), is recorded from Western Ghats of Kerala, India. Detailed morphological characters and illustrations of body and copulatory organs of the species are presented. KEY WORDS: Cyrtarachne keralensis sp. nov., new species, Kerala, India.
The Western Ghats, also known as the Sahyadri Hills, is well known for its rich and unique assemblage of flora and fauna. Norman Myers included the Western Ghats amongst the 25 biodiversity hot-spots identified in the world (Myers et al., 2000). However, the diversity of the invertebrate fauna is poorly studied from these regions.
Cyrtarachne is a genus of nocturnal orb weaving spiders common in South Asia, distinguished from other araneids by its shell like abdomen with numerous sigilla on the dorsum. They build large webs with few radii and with visible viscid droplets. 55 species are recorded from the world so far (Platnick, 2010).
The Western Ghats of Kerala is rich in the diversity of spiders and a total of 431 species of spiders (Jose, 2009) are presently recorded from here. Several new species are discovered from here. In the present paper a new species belonging to the genus Cyrtarachne (Araneidae) is described and illustrated.
MATERIALS AND METHODS
The present species were collected during a study on the diversity of spiders in Western Ghats of Kerala. Spiders were collected by methods described by Tikader (1987). Collected spiders were preserved in 70% alcohol; internal genitalia were dissected out with the help of sharp scalpel. After dissection, the epigynum was cleared in 10% KOH or clove oil. It was then studied with the help of Stereomicroscope Leica MS-5. Drawings were made with the aid of a drawing tube. All measurements are in mm taken with an ocular micrometer. Measurements of legs are shown as: femur, patella, tibia, metatarsus, tarsus: Total length.
The type material is preserved in 80% alcohol and deposited in a reference collection housed at the Department of Zoology, Deva Matha College, Kuravilangad, Kottayam, Kerala, India. (Voucher No AR00102). Abbreviations used are as follows: AME = Anterior median eyes ALE = Anterior lateral eyes PME = Posterior median eyes PLE = Posterior lateral eyes
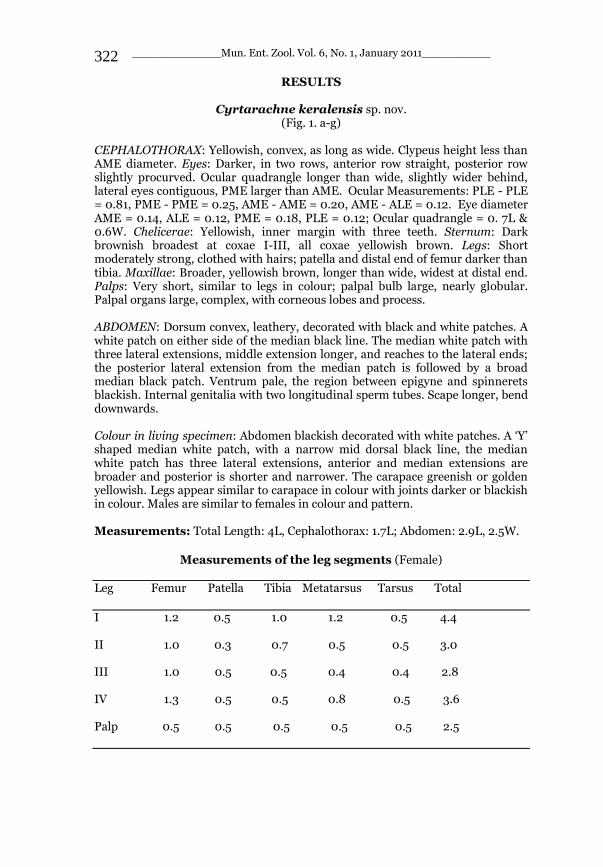
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 322
RESULTS
Cyrtarachne keralensis sp. nov. (Fig. 1. a-g)
CEPHALOTHORAX: Yellowish, convex, as long as wide. Clypeus height less than AME diameter. Eyes: Darker, in two rows, anterior row straight, posterior row slightly procurved. Ocular quadrangle longer than wide, slightly wider behind, lateral eyes contiguous, PME larger than AME. Ocular Measurements: PLE - PLE = 0.81, PME - PME = 0.25, AME - AME = 0.20, AME - ALE = 0.12. Eye diameter AME = 0.14, ALE = 0.12, PME = 0.18, PLE = 0.12; Ocular quadrangle = 0. 7L & 0.6W. Chelicerae: Yellowish, inner margin with three teeth. Sternum: Dark brownish broadest at coxae I-III, all coxae yellowish brown. Legs: Short moderately strong, clothed with hairs; patella and distal end of femur darker than tibia. Maxillae: Broader, yellowish brown, longer than wide, widest at distal end. Palps: Very short, similar to legs in colour; palpal bulb large, nearly globular. Palpal organs large, complex, with corneous lobes and process. ABDOMEN: Dorsum convex, leathery, decorated with black and white patches. A white patch on either side of the median black line. The median white patch with three lateral extensions, middle extension longer, and reaches to the lateral ends; the posterior lateral extension from the median patch is followed by a broad median black patch. Ventrum pale, the region between epigyne and spinnerets blackish. Internal genitalia with two longitudinal sperm tubes. Scape longer, bend downwards. Colour in living specimen: Abdomen blackish decorated with white patches. A ‗Y‘ shaped median white patch, with a narrow mid dorsal black line, the median white patch has three lateral extensions, anterior and median extensions are broader and posterior is shorter and narrower. The carapace greenish or golden yellowish. Legs appear similar to carapace in colour with joints darker or blackish in colour. Males are similar to females in colour and pattern. Measurements: Total Length: 4L, Cephalothorax: 1.7L; Abdomen: 2.9L, 2.5W.
Measurements of the leg segments (Female)
Leg Femur Patella Tibia Metatarsus Tarsus Total
I 1.2 0.5 1.0 1.2 0.5 4.4
II 1.0 0.3 0.7 0.5 0.5 3.0
III 1.0 0.5 0.5 0.4 0.4 2.8
IV 1.3 0.5 0.5 0.8 0.5 3.6
Palp 0.5 0.5 0.5 0.5 0.5 2.5

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 323
Etymology: Named after the state of Kerala. Holotype: Female: INDIA: Kerala, Ernakulam, Edamalayar, 10°13'18"N, 76°42'21"E, coll. Sunil Jose K., 16.x.2002 Paratypes: 2 Females, 1 Male, Ernakulam, 16.x.2002, Coll. Sunil Jose K. Type deposition: Department of Zoology, Deva Matha College, Kuravilangad, Kerala, India. Distribution: India: Ernakulam, Edamalayar (KL). DIAGNOSIS: Closely similar to C. gravelyi Tikader, 1960 but can be separated by following differences:
1. Sigilla present on the abdomen in C. gravelyi Tikader, 1960 while sigilla are absent in C. keralensis sp. nov.
2. Structure of Internal genitalia and epigyne shows difference in both
species.
3. Inner margin of chelicerae with three teeth in C. keralensis sp. nov.
ACKNOWLEDGEMENTS
Financial assistance from Kerala State Council for Science, Technology and Environment (KSCSTE) is gratefully acknowledged.
LITERATURE CITED
Ferguson. 1906. Travancore State Manual, Govt. of Kerala. I: 160. Myers, N., Mittermeier, R. A., Mittermeier, C. G., Fonseca Gab, D. A. & Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature, 403: 853–858. Platnick, N. I. 2010. The world spider catalog, version 10.5. American Museum of Natural History, online http://research.amnh.org/entomology/spiders/catalog/index.html Pocock, R. I. 1900. The fauna of British India, including Ceylon and Burma. Arachnida. London, pp. 1-279. Subrahmanyam, T. V. 1941. Some Indian spiders: Their season of prosperity. Jour. Bombay nat. Hist. Soc., 42: 217-219. Subrahmanyam, T. V. 1954. Habits and Habitats of some common spiders found in western India. Jour. Bombay nat. Hist. Soc., 52: 874-885. Subrahmanyam, T. V. 1968. An Introduction to the study of Indian spiders. Jour. Bombay nat. Hist. Soc., 65 (2): 453-461. Jose, K. S. 2005. A Faunistic Survey of Spiders ; (Araneae: Arachnida) in Kerala, Ph.D Thesis, M.G. University, Kottayam. Online at http://www.mgutheses.org/
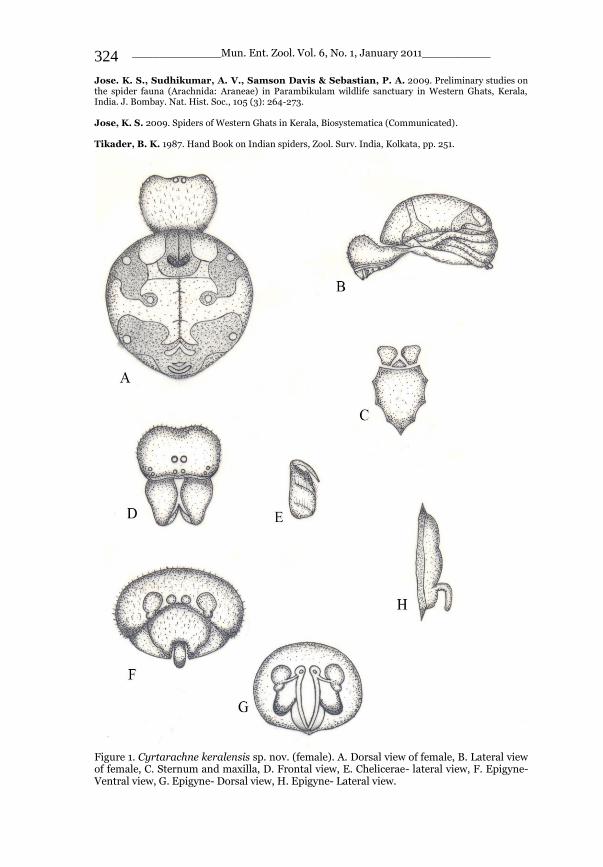
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 324
Jose. K. S., Sudhikumar, A. V., Samson Davis & Sebastian, P. A. 2009. Preliminary studies on the spider fauna (Arachnida: Araneae) in Parambikulam wildlife sanctuary in Western Ghats, Kerala, India. J. Bombay. Nat. Hist. Soc., 105 (3): 264-273. Jose, K. S. 2009. Spiders of Western Ghats in Kerala, Biosystematica (Communicated). Tikader, B. K. 1987. Hand Book on Indian spiders, Zool. Surv. India, Kolkata, pp. 251.
Figure 1. Cyrtarachne keralensis sp. nov. (female). A. Dorsal view of female, B. Lateral view of female, C. Sternum and maxilla, D. Frontal view, E. Chelicerae- lateral view, F. Epigyne- Ventral view, G. Epigyne- Dorsal view, H. Epigyne- Lateral view.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 325
AN INVESTIGATION ON THRIPS FAUNA OF GUILAN PROVINCE, NORTH OF IRAN (INSECTA: THYSANOPTERA)
Jalal Jalali Sendi*, Idin Zibaee** and Kambiz Minaee***
* Department of Plant Protection, College of Agriculture, University of Guilan, Rasht, IRAN. E-mail: [email protected] ** Plant Protection Department, Faculty of Agriculture, University of Tehran, Karaj, IRAN. E-mails: [email protected] *** Department of Plant Protection, College of Agriculture, University of Shiraz, Shiraz, IRAN. [Sendi, J. J., Zibaee, I. & Minaee, K. 2011. An investigation on Thrips fauna of Guilan province, North of Iran (Insecta: Thysanoptera). Munis Entomology & Zoology, 6 (1): 325-329] ABSTRACT: Seventeen species of thrips (Insecta: Thysanoptera) were collected in the present survey from various plants in Guilan province. A fungus feeding genus, Hoplothrips was recorded from Iran for the first time. Ecological information about all these 17 species is provided. Three species, Franklineilla intonsa, Microcephalothrips abdominalis and Thrips tabaci are considered as Tospovirus vectors in Guilan provience. KEY WORDS: Thrips, Thysanoptera, Guilan provience, fauna, Hoplothrips.
Approximately 5800 species are currently recognised in the insect order Thysanoptera (Mound, 2010), and about one hundred of these species have been considered as crop pests, causing damage by feeding or by transmitting virus diseases to growing crops (Lewis, 1997). Most pest Thysanoptera are members of the family Thripidae (Mound, 1997). The significance of thrips as crop pests has been reviewed by Lewis (1997). Thrips that are serious crop pests are usually highly adaptable and polyphagous species. This adaptability is reflected not only in their capacity to feed on various sources, but also in variation in length of larval life, body size at pupation, pupation site, and threshold temperatures for development (Morse & Hoddle, 2006).
Very little is known about the Thysanoptera fauna of Iran. Particularly the information related to the species composition of thrips and their significance as potentially phytophagous or carnivorous is lacking (Minaei et al., 2000). The objectives of the present work were to identify the thrips fauna of Guilan province, their distribution and hosts.
MATERIAL AND METHODS
Thrips specimens have been collected into 60% ethyl alcohol. The specimens were beaten from flowers and leaves. A small plastic beating tray was used and speciments picked off with a small brush into the collecting fluid in plastic ependorf tubes contained a pencil written label of plant, locality and date.
A microscopic slide mount using Hoyers Mountant was prepared using a form of the protocol given in world Thyasanoptera (http://anic.ento.csiro.au/ thrips/field_lab/index.html).
RESULT AND DISCUSION
In this survey, 17 species of Thysanoptera belonging to 3 families (Aeolothripidae, Phlaeothripidae and Thripidae) in Guilan province are reported

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 326
(Table 1). In this list the genus Hoplothrips is recorded in Iran for the first time. In this study two females of this genus were collected so it is not possible to recognize that at species level with the materials. The species of this genus are living under bark of trees, on fungi or in turf (Priesner, 1965).
All of Thyasoptera recorded here are associated with green plants, although two species, A. intermedius and A. collaris are persumbly faculatative predators. It is demonstrated that ―host records‖ based on winged adults that have dispersed from their breeding site is unreliable (Mound, 2005). In Iran there are many host plant records for thrips that may be not true and Minaei et al. (2007) gave some examples of these misunderstanding. So it is not possible to assume that there is a real association with thrips and plants which recorded here (Table 1). Three species recorded here including F. intonsa, Microcephalothrips abdominalis and T. tabaci are reported as vector of tospoviruses around the world (Ullman et al., 1997). Tomato spotted wilt virus (TSWV) has been recorded in Iran, and transmission of cineraria (Senecio sp.) isolate of TSWV has been confirmed by Thrips tabaci (Rasoulpour & Izadpanah, 2003). In addition a new Tospovirus species infecting tomato namely Tomato fruit yellow ring virus has been recorded in Iran and Microcephalothrips abdominalis is recorded as the vector (Ghotbi et al., 2003; Ghotbi et al., 2005). So they can be considered as important pest thrips. Chirothrips manicatus is widely reported as a pest of grasses (Minaei & Mound, 2010). In U.S.A., infestation of Bent Grass (Agrostis sp.) by this species was estimated at 32% (Rao & Alderman, 2005).
Pseudodendrothrips mori is reported as pest of mulbery in Guilan province (Etebari et al., 1999). Taeniothrips inconsequens (Uzel, 1895) and Tenothrips frici are resposbile for damage to flowers in some part of world (Lewis, 1997) and Iran (Esmaeili, 1983). Thrips flavus is the first thrips recorded in Iran by Afshar (1938) as a cotton pest however this species is polyphagous (Zur Strassen, 2003). Thrips meridionalis (Priesner) is collecting on various Rosaceae and Fabaceae (Zur Strassen, 2003) and in this study this host relashinship was confirmed.
Thrips nigropilosus, lives on Asteraceae flowers and widespread in Europe, also in North America, and in Kenya reported as a pest of Pyrethrum (Mound et al., 1976). Haplothrips aculeatus is widespread from Europe to Japan on various Poaceae, and has been found commonly on such plants in north and south Iran (Minaei & Mound, 2008). The species is recorded from various parts of Iran (Bhatti et al., 2007). Haplothrips reuteri is considered to be both common and widespread from south-eastern Europe to India particularly in flowers of various Asteraceae (Minaei & Mound, 2008). It has been recorded from several localities in Iran (Bhatti et al., 2007) and considered as a dominant species in Shiraz region (Minaei & Alichi, 2001).
ACKNOWLEDGEMENTS
Authors wishes to express their deep gratitude to University of Guilan for financial support. We also express our thanks to H. Asaadi for identification of hosts.
LITERATURE CITED Afshar, J. 1938. Pests of summer crops, vegetables, industrial plants, and pastures in Iran and their control. 124 pp. General office of agriculture, Tehran [in Persian]. Bhatti, J. S., Alavi, J., Zur Strassen, R. & Telmadarraiy, Z. 2009. Thysanoptera in Iran 1938–2007. An Overview. Part 1. Thrips, 7: 1–172.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 327
Esmaili, M. 1983. Pests of Fruit Trees, sepehr publication, Tehran, 578 pp. Etebari, K., Jalali, J. & Taksokhan, M. 1999. First record of mulberry thrips Pseudodendrothrips mori NIWA (Thy: Thripidae) from of mulberry orchards of north iran. Journal of entomological society of Iran, Vol.18, No.1-2. Ghotbi, T., Gilasian, E. & Shahraeen, N. 2003. Detection of tospoviruses in individual thrips by ELISA from ornamental plants in Tehran and Markazi provinces. Applied Entomology and Phytopathology, 70: 33-34. Ghotbi, T., Shahraeen, N. & Winter, S. 2005. Occurrence of tospoviruses in ornamental and weed species in Markazi and Tehran provinces in Iran. Plant Disease, 89: 425-429. Gilasian, E., Moharramipour, S. & Alavi, J. 2000. One genera and five species of Thysanoptera as new records for Iran fauna. Proc. 14th Iranian Plant Prot. Congr., p. 341. Lewis, T. 1997. Pest thrips in perspective. In: Lewis T, editor. Thrips as crop pests. Wallingford (UK): CAB International, p. 1–13. Minaei, K., Azemayeshfard, P. & Mound, L. A. 2007. The Thrips genus-group (Thysanoptera: Thripidae) in Iran. Journal of Entomological Society of Iran, 27: 29-36 (in English with Persian Abstract). Minaei, K. & Mound, L. A. 2010. Grass-flower thrips of the genus Chirothrips (Thysanoptera: Thripidae), with a key to species from Iran. Zootaxa, 2411: 33-43. Minaei, K. & Mound, L. A. 2008. The Thysanoptera Haplothripini (Phlaeothripidae) of Iran. Journal of Natural History, 42: 2617-2658. Minaei, K. & Alichi, M. 2001. Thrips of the genus Haplothrips (Thysanoptera: Phlaeothripidae) in Shiraz region. Journal of Entomological Society of Iran, 20: 33-45 (in Persian with English Abstract). Minaei, K., Alichi, M. & Ahmadi, A. A. 2001. The thrips family Aeolothripidae (Thysanoptera: Terebrentia) in the Fars province. Iran Agircultural Rearch, 20: 53-66 (in English with Persian Abstract). Morse, J. G. & Hoddle, M. S. 2006. Invation biology of thrips. Annual. Review of Entomology, 51: 67–89. Mound, L. A., Morison, G. D., Pitkin, B. R. & Palmer, J. M. 1976. Thysanoptera. Handbooks for the Identification of British Insects, 1 (11): 1–79. Priesner, H. 1965. A monograph of the Thysanoptera of the Egyptian deserts. Publ. Inst. Désert Egypte, 13: 1-549. Rao, S. & Alderman, S. C. 2005. Infestation of Bent Grass by a New Seed Pest, Chirothrips manicatus (Thysanoptera: Thripidae), in oregon. Journal of the entomological society of british Columbia, 102: 77-78. Rasoulpour, R. & Izadpanah, K. 2003. Transmission of cineraria isolate of tomato spotted wilt virus by onion thrips in Shiraz. Iranian Journal of Plant Pathology, 39: 28. Ullman, D. E., Sherwood, J. L. & German T. L. 1997. Thrips as Vectors of Plant Pathogens In: Lewis T, editor. Thrips as crop pests. Wallingford (UK): CAB International, p. 539-566. Zur Strassen, R. 2003. Die terebranten Thysanopteren Europas und des Mittelmeer-Gebietes. Die Tierwelt Deutschlands, 74: 1-277.
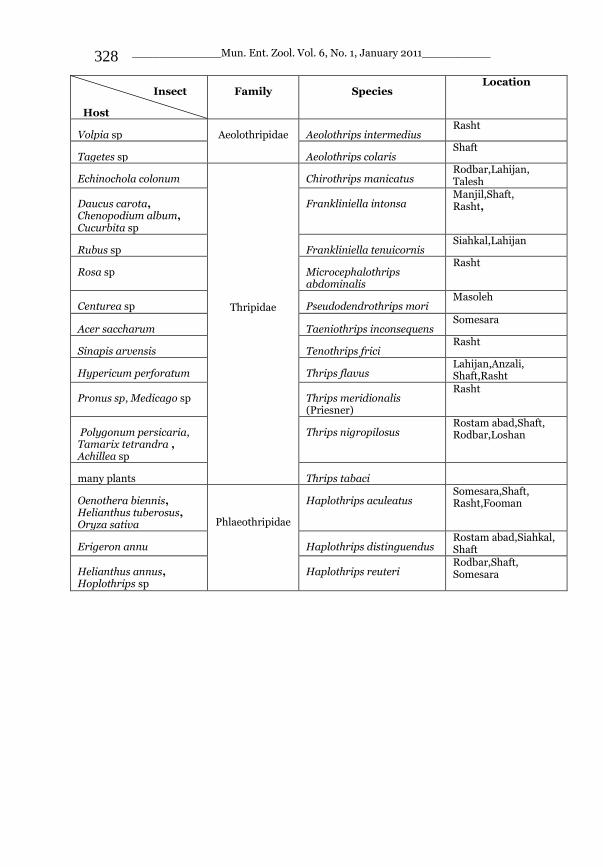
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 328
Insect
Host
Family Species Location
Volpia sp Aeolothripidae Aeolothrips intermedius Rasht
Tagetes sp Aeolothrips colaris Shaft
Echinochola colonum
Thripidae
Chirothrips manicatus Rodbar,Lahijan, Talesh
Daucus carota, Chenopodium album, Cucurbita sp
Frankliniella intonsa Manjil,Shaft, Rasht,
Rubus sp Frankliniella tenuicornis Siahkal,Lahijan
Rosa sp Microcephalothrips abdominalis
Rasht
Centurea sp Pseudodendrothrips mori Masoleh
Acer saccharum Taeniothrips inconsequens Somesara
Sinapis arvensis Tenothrips frici Rasht
Hypericum perforatum Thrips flavus Lahijan,Anzali, Shaft,Rasht
Pronus sp, Medicago sp Thrips meridionalis (Priesner)
Rasht
Polygonum persicaria, Tamarix tetrandra , Achillea sp
Thrips nigropilosus Rostam abad,Shaft, Rodbar,Loshan
many plants Thrips tabaci
Oenothera biennis, Helianthus tuberosus, Oryza sativa
Phlaeothripidae
Haplothrips aculeatus Somesara,Shaft, Rasht,Fooman
Erigeron annu Haplothrips distinguendus Rostam abad,Siahkal, Shaft
Helianthus annus, Hoplothrips sp
Haplothrips reuteri Rodbar,Shaft, Somesara
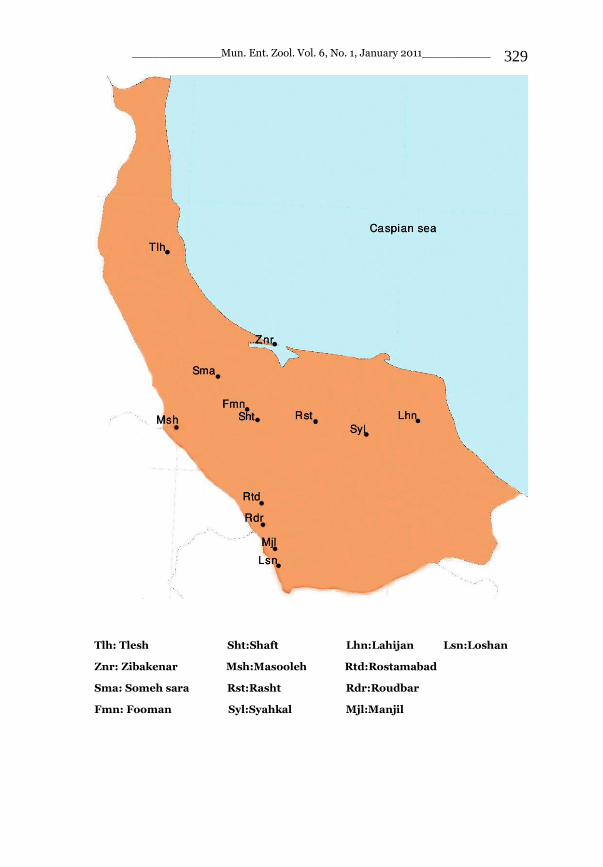
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 329
Tlh: Tlesh Sht:Shaft Lhn:Lahijan Lsn:Loshan
Znr: Zibakenar Msh:Masooleh Rtd:Rostamabad
Sma: Someh sara Rst:Rasht Rdr:Roudbar
Fmn: Fooman Syl:Syahkal Mjl:Manjil

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 330
EFFECT OF TEMPERATURE AND DENSITY ON LARVAL PUPATION SITE PREFERENCE IN SIBLING SPECIES OF
DROSOPHILA (DROSOPHILIDAE: DIPTERA)
N. B. Vandal, G.S. Siddalingamurthy and N. Shivanna* * Department of Studies in Zoology, Karnatak University, Dharwad 580003, INDIA. E-mail: [email protected] [Vandal, N. B., Siddalingamurthy, G. S. & Shivanna, N. 2011. Effect of temperature and density on larval pupation site preference in sibling species of Drosophila (Drosophilidae: Diptera). Munis Entomology & Zoology, 6 (1): 330-338] ABSTRACT: Larval pupation site preference (PSP) has been studied in sibling species of D. melanogaster and D. simulans at different density and temperature. It revealed significant variation between altered and controlled experiments. D. melanogaster larvae prefer to pupate less on glass and more in media when temperature and density altered than control. D. simulans larvae prefers glass lesser than control at 30ºC and higher than control at 200 and 250 larvae/vial at 15, 20 and 25ºC, whereas the media pupation is contrasts to the glass pupation in D. simulans. The media pupation is more at 30ºC and less at other temperature at all type of density compared to control. D. melanogaster larvae does not prefer to pupate on cotton at 30ºC, the larvae of D. simulans does not prefer o pupate on cotton at all density and temperatures. Two-way ANOVA revealed that variation of PSP in both the species at varied temperature and density at different sites was found to be significant. KEY WORDS: Drosophila, pupation site preference, density, temperature, larva, habitat choice.
The larval pupation site preference (PSP) is an important event in Drosophila preadult development, because the place selected by the larva can have decisive influence on their subsequent survival as pupae (Sameoto & Miller, 1968). The larval PSP has been analyzed by two types of phenotypic character one is the pupation height and the other is pupation site choice. The pupation height studies have been made using different factors such as moisture (Mensua, 1967; Sameoto & Miller, 1968), light (Pandey & Singh, 1993), temperature (Sokal et al., 1960; Schnebel & Grossfield, 1986; 1992), density (Singh & Pandey, 1993a), sex (Bauer, 1984), larval developmental time (Markow, 1979), selection for high and low pupation height and its genetic control (Casares & Carracedo, 1987; Singh & Pandey, 1993b). The larval PSP has also been studied by analyzing the percentage of pupae pupated at different sites viz; cotton, glass and medium in the culture. The studies of Barker (1971), Shirk et al. (1988), Shivanna et al. (1996) showed that under normal condition most of the species prefer to pupate maximum on media, and reported that the PSP has been related to the quantity of larval salivary gland protein. It is not influenced by increased water in the media and larval density, whereas the larval locomotory path length, pattern and substrates play a role in the preference of the sites for pupation (Vandal et al., 2003; Vandal & Shivanna, 2004; 2005a; 2005b).
The effect of temperature has been studied in different species of Drosophila on both adult and preadult characters. Compared to individuals reared at the standard temperature of about 25°C, rearing at a colder temperature (16 to 17°C) results in increased egg size (Delpuech et al., 1995), larval and pupal duration (Gebhardt & Stearns, 1988; French et al., 1998), larval and adult size at eclosion (Gebhardt & Stearns, 1992; David et al., 1994), larger wing size (Noach et al., 1996; Imasheva et al., 1997), larval critical weight (de Moed et al., 1999;

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 331
Bochdanovits & De Jong, 2003), life span, life time fecundity and progeny production but reduces daily fecundity ( Partridge et al., 1995).
Increased larval crowding of cultures results in a decrease of food availability over time and increase in metabolic waste levels especially ammonia (Borash et al., 1998). The major phenotypic effects of rearing larvae at high density (several hundred per vial) versus moderate (50 to 100 per vial) density has increased larval and pupal mortality (Joshi & Mueller, 1993; Roper et al., 1996), larval development time (Mueller et al., 1993; Chippindale et al., 1997; Prasad et al., 2001), pupation height (Joshi & Mueller, 1993; Pandey & Singh, 1993) and adult life span (Joshi & Mueller, 1997; Zwaan et al., 1991) whereas it reduced the adult size (Santos et al., 1997) and fecundity (Chippindale et al., 1993). De Souza et al. (1970) reported that the simple genetic control for pupation site choice in D. willistoni under high-density conditions clearly implicates a different type of behaviour than the pupation height measured for D. melanogaster and D. simulans. Larval adaptations are less specific than oviposition in response to temperature stress and it is more pronounced for puparia than adults in Drosophila (Kaneshiro et al., 1973; Coyene et al., 1983). Joshi (1997) reviewed the obvious effects of density on adaptations of both larva and adult, the food medium is rapidly becomes very moist and soggy, and individuals pupating on or close to the surface of the medium have an increased chance of being dislodged and drowned in the medium. The information about the effect of varied density in relation to temperature on larval PSP has not been available. In view of this it is planned to study the larval PSP using combined factors such as density and temperature. For the present investigation the sibling species, D. melanogaster and D. simulans belongs to melanogaster species group occupying different sites for pupation at constant conditions were taken.
MATERIALS AND METHODS
Sibling species, D. melanogaster and D. simulans were taken to study the effect of temperatures with varying larval densities on PSP (Bock & Wheeler, 1972; Ehrman, 1978). D. melanogaster and D. simulans primarily tropical but have expanded to temperate zones. These Drosophila species were collected from the Drosophila stock centre, University of Mysore, Mysore, India maintained since 20 years.
In order to maintain uniformity with regard to the density and age of the larvae the eggs were collected every 6 hours using modified technique of Delcour described by Ramachandra & Ranganath, (1988) and allowed to hatch. First instar larvae about 50 100, 150, 200, and 250 from the cultures were isolated and transferred to vial (10 X 3.8cm) containing equal quantity of wheat cream agar medium (Shivanna et al., 1996). About 50μl of dilute yeast was added everyday to feed the larva and to maintain the moisture content of the food medium. The culture vials with varied densities were kept at four different temperatures viz; 15C°, 20C°, (control 22 1 C) 25C° and 30°C with RH 80%.
Ten replicates were carried out for each experiment. The mean values as well as percentage of pupation was calculated based on the number of larvae pupated at different sites viz; cotton, glass and medium. The primary data (number of pupae on different sites) was subjected to two-way ANOVA (SPSS software was used).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 332
RESULTS
Table 1 shows the mean percentage of glass pupation at different larval density and temperature in D. melanogaster and D. simulans. It reveals that D. melanogaster prefer to pupate maximum on glass at different density and temperature. Compared to control, the glass pupation was decreased at all the temperatures and larval density except in 150 and 200 larvae/vial at 25ºC. The larvae of D. simulans prefer to pupate minimum on glass compared to control; it was increased in 150 larvae/vial at 20ºC and 25ºC followed by 200 and 250 larvae/vial at 15ºC, 20ºC and 25ºC respectively.
Table 2 reveals that D. melanogaster larvae prefer to pupate lesser on media than glass at different density and temperature. Compared to control the media pupation was increased in all larval densities and temperatures. The larvae of D. simulans prefer to pupate higher on media than glass at different density and temperature. Compared to control the media pupation was decreased in all larval density and temperatures except at 30ºC in 100 to 250 larvae /vial.
Cotton pupation was found to be less at all temperature and density in D. melanogaster. The larvae of D. melanogaster do not prefer to pupate on cotton in control and 30°C. The cotton preference was increased in 250 larvae / vial at 15°C, 200 and 250 larvae / vial at 20°C and 25°C. The larvae of D. simulans do not prefer to pupate on cotton both in control and treatment experiment (Table 3).
The larval mortality was more in all density and temperature compared to control in D.melanogaster and similar in D.simulans except at 200 and 250 larvae/vial at 30ºC and 250 larvae/vial at 25ºC compared to control (Table 4). The combined effect of temperatures and densities at three different sites viz; cotton, glass and media pupation was analyzed by two-way ANOVA (temperature X density X sites), it revealed that the variation of larval pupation site preference in both the species is highly significant (Table 5).
DISCUSSION
In Drosophila, temperature is the most important environmental factor, which affects all biological process at the molecular, cellular and organismic levels (David et al., 1983). Temperature involves limitations on Drosophila behavioural alterations, which are expressed only at or above a critical temperature. Different species shows different optimum temperature for developing and some cannot be grown above certain temperature, the flies were moved to cooler place above 41°C (Grossfield, 1978). High temperature and dry periods for several days may act as strong selective force on developing pupae and the pupal survivorship decreases at lower temperature than the higher temperature in D. melanica (Tonzetich & Ward, 1972). D. melanogaster at control and different temperature with varied density preferred to pupate maximum on glass and minimum on media and cotton, whereas its sibling species D. simulans preferred to pupate maximum in/on media and minimum on glass.
Any larva pupating on the surface of the medium is likely to be buried if there is still an actively feeding larval population which suggests that the fitness of genotypes which vary in pupation height may be a function of larval density (Mueller & Sweet, 1986). Sokal et al. (1960) investigated genetic and environmental factors that govern the selection of pupation sites by D. melanogaster. It reveals that densities below 52 eggs / vial do not affect pupation site. At high-density level considered, there appears no relation between pupation

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 333
site and density (Sokal et al., 1960). In contrast, Pandey & Singh (1993) and Joshi (1997) reported that there is increase in pupation height with increased larval density in D. melanogaster, D. ananassae, D. bipectinata, D. malerkotliana and D. biarmipes. Present study reveals that percentage of glass pupation was decreased and media pupation was increased at all density and temperature in D. melanogaster, whereas D. simulans larvae showed contrasts results to D. melanogaster. Pupation on cotton was found at highest density and lowest temperature in D. melanogaster. Joshi & Mueller (1996) reported that at high larval density when there is a shortage of food to the larvae it switches from feeding to pupation.
Highest larval mortality was found in D. melanogaster at all density and temperatures in D. simulans highest mortality was observed with 150 larvae / vial at 15ºC and 30ºC, 200 and 250 larvae / vial at 20ºC and 25ºC compared to control (Table 4). In cultures of high density of larvae or adults, the food medium rapidly becomes very moist and soggy. In such cultures, individuals pupating on or close to the surface of the medium have an increased chance of being dislodged and drowned in the medium (Joshi & Mueller, 1993). Earlier studies also reported highest pupal mortality above the food surface on vials even at low densities (Joshi & Mueller, 1993; Borash et al., 1998). Present study highest mortality was found at high temperature and density in D. melanogaster and D. simulans.
Vandal & Shivanna (2005a) showed no effect of water content of the medium and larval density on PSP in closely related sibling, sympatric, virilis and repleta group species when reared separately and reported that the (control) basic nature of PSP of the species has not been changed even though there is lack of space due to increased density. Among the species analyzed, D. melanogaster and D. simulans are closely related sibling species belonging to the melanogaster subgroup. It reveals that D. melanogaster and D. simulans prefer to pupate maximum extent on glass and medium at different density and temperature respectively. When the larvae were exposed to altered and combined temperature and density affected the PSP on all the sites in both the species. Two-way ANOVA revealed that variation of pupation site preference both in D. melanogaster and D. simulans at different sites with combined effect of density and temperature is found significant (Table 5).
The effect of larval density on PSP at constant temperature reveals that the larval PSP does not changed with increased density and they preferred to pupate as similar to control (Vandal & Shivanna, 2005a). The effect of temperature on larval PSP at constant larval density (50 larva / vial) revealed that at lowest temperature of 15ºC the glass pupation was increased and decreased at highest temperature of 30ºC (Vandal & Shivanna, 2007). The above studies indicate that density has no role in PSP whereas temperature plays major role in PSP. The present study reveals that when the larvae were exposed to both temperature and density simultaneously the PSP varies significantly. Therefore it is concluded that in presence of temperature, density also affects the PSP. Temperature and density together affect the larval behaviour, which causes the differential pupation site preference in sibling species of Drosophila.
ACKNOWLEDGEMENTS
Thanks are due to Professor H.A.Ranganath Drosophila Stock Centre Department of Zoology, University of Mysore, ‗Manasagangotri‘ Mysore, for Drosophila stocks and UGC minor research project sanctioned to Dr. N. Shivanna F No. 31-234/2005 dated 31/3/2006 for financial assistance.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 334
LITERATURE CITED Barker, J. S. F. 1971. Ecological differences and competitive interaction between D. melanogaster and D. simulans in small laboratory populations, Oecologia, 8: 139. Bauer, S. J. 1984. Sex differences in pupation site choice in Drosophila melanogaster. Dros. Inf. Serv., 60, 58. Bochdanovits, Z. & De Jong, G. 2003. Temperature dependent larval resource allocation shaping adult body size in Drosophila melanogaster. J. Evol. Biol., 16: 1159-1167. Bock, I. R. & Wheelar, M. R., 1972. The Drosophila melanogaster species Group, Univ Tex, Publ No 7213, 1-102. Borash, D. J., Gibbs, A. G., Joshi, A. & Mueller, L. D. 1998. A genetic polymorphism maintained by natural selection in temporally varying environment. Am. Nat., 151: 148-156. Casares, P. & Carracedo, M. C. 1987. Pupation height in Drosophila: Sex differences and influence of larval developmental time. Behav. Genet., 17: 523-535. Chippindale, A. K., Lerio, A. M., Kim, S. B. & Rose, M. R. 1993. Phenotypic plasticity and selection in Drosophila life history evoltion. I. Nutrition and the cost of reproduction. J. Evol. Biol., 6: 171-193. Chippindale, A. K., Alipaz, J. A., Chen, H. W. & Rose, M. R. 1997. Experimental evolution of accelerated development in Drosophila. 1. Developmental speed and larval survival. Evolution, 51: 1536-1551. Coyen, J. A., Bundgaard, J. & Prout, T. 1983. Geographic variation of tolerance to environmental stress in Drosophila subobscura. Am. Nat., 122: 478-488. David, J. R., Allemand, R., Van Herreweg, J. & Cohe, Y. 1983. Ecophysiology: abiotic factors. In The Genetics and Biology of Drosophila (edited by, Ashburner, M., Carson, H.L & Thompson J.N) vol 3B. Pp. 106-109. Acad. Press. London. David, J. R., Moretea, B., Gautier, J. R., Stockel, A. & Imasheva, A. G. 1994. Reactions norms of size characters in relation to growth temperature Drosophila melanogaster: an isofemale line analysis. Genet. Select. Evol., 26: 229-251. De Moed, G. H., de Jong, G. & Scharloo, W. 1999. The energetics growth of Drosophila melanogaster. Effect of temperature and food conditions. Neth. J. Zool., 48: 169-188. De Souza, H. L., da Cunha, A. B. & dos Santos, E. P. 1970. Adaptive polymorphism of behaviour developed in laboratory population of Drosophila willistoni. Am. Nat., 102: 583. Delpuech, J. M., Moreteau, B., Chicke, J., Pla, E., Vouidibio, J. & David, J. R. 1995. Phenotypic plasticity and reaction norms in temperate and tropical population of Drosophila melanogaster: ovarion size and development temperature. Evolution, 49: 670-675. French, V., Feast, M. & Partridge, L. 1998. Body size and cell size in Drosophila: the developmental responses to temperature. J. Insect. Physiol., 44: 1081-1089. Gebhardt, M. B. & Stearns, S. C. 1988. Reaction norms for developmental time and weight at eclosion in Drosophila mercatorum. J. Evol. Biol., 1: 335-354. Gebhardt, M. B. & Stearns, S. C. 1992. Phenotypic plasticity for life history characters in Drosophila melanogaster. III. Effect of the environment on genetic parameters. Genet. Res,, 60: 87-101. Grossfield, J. 1978. Non-sexual behaviour of Drosophila, in The genetics & biology of Drosophila Vol 2a, edited by M Ashburner & TRF Wright. (Academic Press, New York), 1-179. Imasheva, A. G., Loeschcke, V., Zhivatovsky, L. A. & Lazebny, O. E. 1997. Effects of extreme temperature on phenotypic variation and developmental stability in Drosophila melanogaster and Drosophila buzzatti. Biol. J. Linn. Soc., 6: 117-126. Joshi, A. 1997. Laboratory studies of density-dependent selection: Adaptation to crowding in Drosophila melanogaster. Curr. Sci., 72: 555-562.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 335
Joshi, A. & Mueller, L. D. 1988 Evolution of higher feeding rate in Drosophila due to density-dependent natural selection. Evolution, 42: 1090-1093. Joshi, A. & Mueller, L. D. 1993. Directional and stabilizing density-dependent natural selection for pupation height in Drosophila melanogaster. Evolution, 47: 176-184. Joshi, A. & Mueller, L. D. 1996. Density-dependent natural selection in Drosophila: trade-offs between larval food acquisition and utilization. Evol.Ecol., 10: 463-474. Joshi, A. & Mueller, L. D. 1997. Adult crowding effects on longevity in Drosophila melanogaster increase in age independent mortality. Curr. Sci., 72: 255-260. Kaneshiro, K. Y., Carson, H. l., Fowler, K. & Heed, W. D. 1973. Niche separation in pair of homosequential Drosophila species from the island of Hawaii. Amer.Nat., 197: 766-774. Markow, T. A. 1979. Survey of intra and inter specific variation for pupation height in Drosophila. Behav. Genet., 9: 209. Mensua, J. 1967. Some factors affecting pupation height of Drosophila. Dros. Inf. Serv., 42: 76. Mueller, L. D. & Sweet, V. F. 1986. Density- dependent natural selection in Drosophila: evolution of pupation height. Evolution, 40: 1354-1356. Mueller, L. D., Graves, J. L. Jr. & Rose, M. R. 1993. Interaction between density-dependent and age specific selection in D. melanogaster. Func. Ecol., 7: 469-479. Noach, E. J. K., de Jong, G. & Scharloo, W. 1996. Phenotypic plasticity in morphological traits in Drosophila melanogaster. J. Evol. Biol., 9: 831-844. Pandey, M. B. & Singh, B. N. 1993. Effect of biotic and abiotic factors on pupation height in four species of Drosophila. Ind. J .Exp. Biol., 31: 912-916. Partridge, L., Barrie, B., Fowler, K. & French, V. 1995. Evolution and development of body size and cell size in Drosophila melanogaster in response to temperature. Evolution, 48: 126-1276. Prasad, N. G., Mallikarjun, Shakarad, Amit, D., Rajamani, M. & Joshi, A. 2001. Correlated responses to selection in Drosophila: the evolution of larval trait. Evolution, 556: 1363-1372. Ramachandra, N. B. & Ranganath, H. A. 1988. Estimation of population fitness of parental races Drosophila nasuta nasuta and Drosophila nasuta albomicana and of the newly evolved Cytoraces I and II. Genome, 30: 58. Roper, C., Pignatelli, P. & Partridge, L. 1996. Evolutionary responses of Drosophila melanogaster life history to differences in larval density. J. Evol. Biol., 9: 609-622. Sameoto, D. D. & Miller, R. S. 1968. Selection of pupation site by Drosophila melanogaster, Drosophila simulans. Ecology, 49: 177. Santos, M., Borash, D. J., Bounlutay, N. & Mueller, L. D. 1997. Density-dependent natural selection in Drosophila: evolution of growth rate and body size. Evolution, 51: 420-432. Schnebel, E. M. & Grossfield, J. 1986. Pupation- temperature range in 12 species Drosophila species from different ecological backgrounds. Experientia, 42: 600-604. Schnebel, E. M. & Grossfield, J. 1992. Temperature effects on pupation height response in four Drosophila species traids. J. Insect. Physiol., 38: 727-732. Shirk, P. D., Roberts, P. A. & Harn, C. H. 1988. Synthesis and secretion of salivary gland proteins in Drosophila gibberosa during larval and prepupal development. Roux‘s Arch. Dev. Biol., 197: 66. Shivanna, N., Siddalinga Murthy, G. S. & Ramesh, S .R. 1996. Larval pupation site preference and its relationship to the glue proteins in a few species of Drosophila. Genome, 39: 105-111. Singh, B. N. & Pandey, M. B. 1993a. Selection for high and low pupation height in Drosophila ananassae. Behav. Genet., 23: 239.
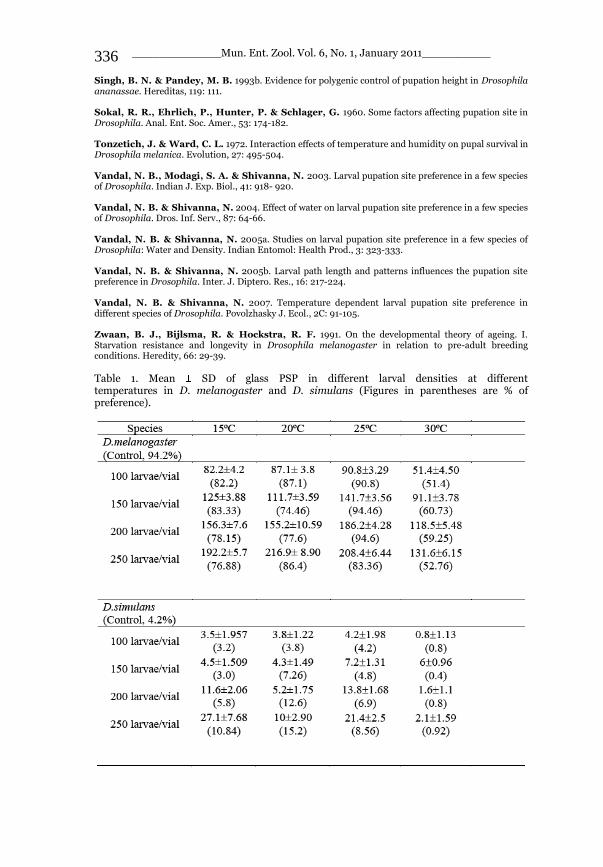
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 336
Singh, B. N. & Pandey, M. B. 1993b. Evidence for polygenic control of pupation height in Drosophila ananassae. Hereditas, 119: 111. Sokal, R. R., Ehrlich, P., Hunter, P. & Schlager, G. 1960. Some factors affecting pupation site in Drosophila. Anal. Ent. Soc. Amer., 53: 174-182. Tonzetich, J. & Ward, C. L. 1972. Interaction effects of temperature and humidity on pupal survival in Drosophila melanica. Evolution, 27: 495-504. Vandal, N. B., Modagi, S. A. & Shivanna, N. 2003. Larval pupation site preference in a few species of Drosophila. Indian J. Exp. Biol., 41: 918- 920. Vandal, N. B. & Shivanna, N. 2004. Effect of water on larval pupation site preference in a few species of Drosophila. Dros. Inf. Serv., 87: 64-66. Vandal, N. B. & Shivanna, N. 2005a. Studies on larval pupation site preference in a few species of Drosophila: Water and Density. Indian Entomol: Health Prod., 3: 323-333. Vandal, N. B. & Shivanna, N. 2005b. Larval path length and patterns influences the pupation site preference in Drosophila. Inter. J. Diptero. Res., 16: 217-224. Vandal, N. B. & Shivanna, N. 2007. Temperature dependent larval pupation site preference in different species of Drosophila. Povolzhasky J. Ecol., 2C: 91-105. Zwaan, B. J., Bijlsma, R. & Hockstra, R. F. 1991. On the developmental theory of ageing. I. Starvation resistance and longevity in Drosophila melanogaster in relation to pre-adult breeding conditions. Heredity, 66: 29-39.
Table 1. Mean SD of glass PSP in different larval densities at different temperatures in D. melanogaster and D. simulans (Figures in parentheses are % of preference).
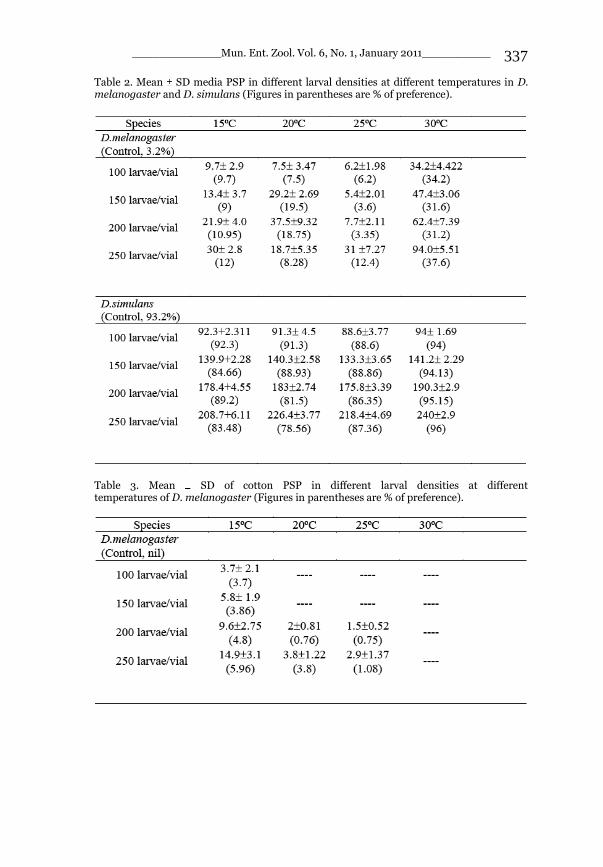
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 337
Table 2. Mean SD media PSP in different larval densities at different temperatures in D. melanogaster and D. simulans (Figures in parentheses are % of preference).
Table 3. Mean SD of cotton PSP in different larval densities at different temperatures of D. melanogaster (Figures in parentheses are % of preference).
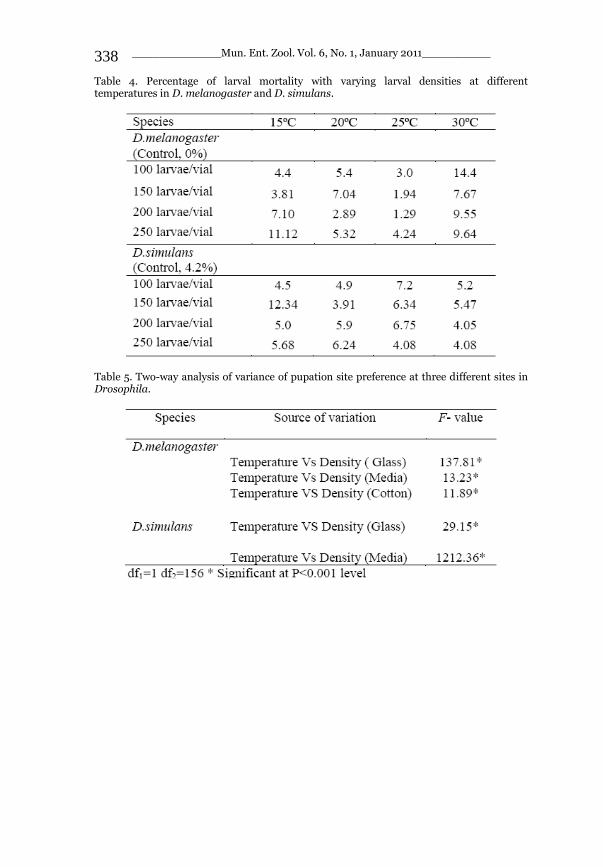
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 338
Table 4. Percentage of larval mortality with varying larval densities at different temperatures in D. melanogaster and D. simulans.
Table 5. Two-way analysis of variance of pupation site preference at three different sites in Drosophila.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 339
INSECTICIDAL EFFECT OF SIX NATIVE MEDICINAL PLANTS ESSENTIAL OIL ON INDIAN MEAL MOTH,
PLODIA INTERPUNCTELLA HÜBNER (LEP.: PYRALIDAE)
Zahra Rafiei-Karahroodi*, Saied Moharramipour**, Hossein Farazmand*** and Javad Karimzadeh-Esfahani****
* Department of Entomology, College of Agriculture, Islamic Azad University Arak branch, Arak, IRAN. E-mail: [email protected] ** Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, IRAN. E-mail: [email protected] *** Agricultural Entomological Research Department, Iranian Research Institute of Plant Protection, Tehran, IRAN. E-mail: [email protected] **** Agricultural and Natural Resources Research Center, Esfahan, IRAN. [Rafiei-Karahroodi, Z., Moharramipour, S., Farazmand, H. & Karimzadeh-Esfahani, J. 2011. Insecticidal effect of six native medicinal plants essential oil on Indian meal moth, Plodia interpunctella Hübner (Lep.: Pyralidae). Munis Entomology & Zoology, 6 (1): 339-345] ABSTRACT: The essential oils are considered powerful source of natural derivatives useful against stored product pests; they are as new control strategies. In this research it was investigated fumigant toxicity of six essential oils from Melissa officinalis L., Mentha piperata L., Petroselinum sativum Hoffmann, Lavandula angustifolia Mill., Ziziphora clinopodioides Lam., and Artemisia dracunculus L., on first instar larvae and eggs, ovipositional deterrency of Plodia interpunctella Hübner. LC50 values were determined. Ovicidal activity was assessed in Petri dishes on one-day old eggs that exposures three concentrations as 3, 12 and 24 micro liter essential oil per liter air. For studying effect of essential oils on oviposition deterrence one pair of new emergence adult were exposured by 2 µl essential oil. LC50s were between 5 to 10 µl/l and Z. clinopodioides had most ovicidal activity. Increasing concentrations increased activity of essential oils. The least oviposited eggs have been observed in L. angustifolia, M. piperata and Z. clinopodioides. These results showed that L. angustifolia, and Z. clinopodioides had more ovicide and egg deterrence effect for this pest. The essential oils investigated in this study are used as pharmaceuticals and in flavoring. They are considered because they are less harmful to humans than most synthetic insecticides. KEY WORDS: Essential oil, Plodia interpunctella, Insecticidal effect.
The Indian meal moth (IMM), Plodia interpunctella Hübner (Lep.: Pyralidae) is a major economic insect pest of stored products. The insect is a pest of grain, grain-based crops, and more than 20 different nuts, fruits, and candies from the agricultural system (Hamlin et al., 1931; Johnson et al., 1992). It prefers to feed on broken grains and more especially on milled cereal, dried fruits and almonds, pistachios and walnuts and groundnuts. It is found in warehouses (Perez-Mendoza & Aguilera-Pena, 2004). The larvae spin a silky web inside and on top of the food surface and feed in it. it contains larval excreta and gives an unpleasant odor to the infested food (Phillips et al., 2000). The loss of chemical insecticides and resistance to synthetic insecticides, new laws and interpretations of those laws, economic costs of pesticide regulations, consumer preferences and expectations, have important consequences for the management of IMM and other stored-product insects. The impending loss of the fumigant methyl bromide through compliance with the Montreal Protocol (Anonymous, 2004) will undoubtedly further affect management programs for IMM, accelerating the demand for new control strategies (Zettler, 1973; Phillips et al., 2000; Lee et al.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 340
2001). There is therefore the need to develop safer alternatives to conventional fumigants. Several natural products, including essential oils from many species of spices, herbs and medicinal plants, are known to have a range of useful biological properties against insects (Regnault-Roger, 1997; Shaaya et al. 1997; Obeng-Ofori & Reichmuth, 1997; Isman, 2000; Tripathi et al., 2002; Mahfuz, 2007). The essential oils are useful to control stored product pests and their insecticidal activities are considerable (Lee et al., 2001).
Although many studies have demonstrated the fumigant activity of various essential oils and their constituents on adults and larval stages of stored grain insects, relatively little attention has been given to the egg stage(Obeng-Ofori & Reichmuth, 1997; Tripathi et al., 2002). Essential oils have a low toxicity to vertebrate, high volatility, and toxicity to stored-grain insect pests. Most of these oils are environmentally non-persistent and non-toxic to humans (with some exceptions), while being effective against several pest species. (Regnault-Roger, 1997; Shaaya et al. 1997; Isman, 2000; Tripathi et al., 2002; Mahfuz, 2007).
The biologically active materials derived from plant sources can act as larvicide, ovicide and oviposition deterrence. In recent years, essential oils have received much attention as potentially useful bioactive compounds against insects. Essential oils of Myrtus communis L., Origanum syriacum L., Lavendula stoechas L. have been documented for larvicidal activities towards Culex pipens molestus (Pushpanathan et al., 2006; Raja & William, 2008).Very little consideration has been given to the egg stage and only recently have some authors described toxicity of essential oils against eggs of stored product insects (Obeng-Ofori & Reichmuth, 1997) and they should have the ability to kill all stages of insects. (Yunc et al., 2000; Ngamo et al., 2007; Regnault-Roger, 1997).
In the present work, larvicidal and Ovicidal activity in addition to oviposition detterency of essential oils from six medicinal plants have been studied against IMM.
MATERIAL AND METHODS
Plant materials included Lavandula angustifolia Mill. (Leaves), Artemisia dracunculus L. (leaves), Ziziphora clinopodioides Lam. (leaves), Melissa officinalis L. (leaves), Petroselinum sativum Hoffmann (seeds) and Mentha piperata L. (leaves), were collected in Markazi province of Iran, from Agricultural Research Center. Plants were dried in dark, cut in pieces and hydrodistilled. The extraction of essential oils was carried out in a Clevenger-type.
The IMM was reared in growth chamber sets at 28±1 ºС, 60±5% r.h., photoperiod of 11:13 (L: D) h. Larvae were reared in an artificial diet containing: wheat bran (800 g), brewers yeast (160 g), honey (200 ml), Glycerin (200 ml), Methyl-parabon (1 g), Choloramphenicol (1 g) (Sait et al., 1997). Adults after immergence were transferred to plastic funnel that covered with a net cloth for gathering eggs.
The experiment was carried out in four replications with first instar larvae. For each replication, essential oil solutions were poured in the inner surface of petri dish‘s door. After 20 minutes, ten larvae with one gram were placed in petri dishes and protected with parafilm. The mortality was recorded after 24 h. Data were analyzed by Probit analysis using the SAS 6.12.
To assess ovicidal activity of essential oils, each petri dish has divided to 80 cells by tangle-foot glue and has placed a one-day old egg in each cell. The eggs were treated with three concentrations of essential oils (3, 12 and 24 µl/l air) in

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 341
three replications. The larval emergences were determined after 4 days. Percent of egg hatching was calculated by formula (Bruce et al., 2004).
B
AEH
100%
%EH= percent of egg hatching A= number of hatched eggs B= number of all eggs Mortality in treatments was corrected by Abbott‘s formula (Abbott, 1925).
C
CTMortality
100
100)(%
T=Mortality in treatment C= Mortality in control
Oviposition deterrency was studied with one pair of new adults treated with 2 µl essential oils. After four days, fecundity of moths was recorded and in all treatments were calculated percent of inhibitory of egg releasing comparison with control by formula (Chaubey, 2008).
Nc
NtI
100100%
I= Inhibitory of egg releasing Nt= Number of eggs in treatment Nc= Number of eggs in control
All data was analyzed as completed randomized design SAS 6.12 (ANOVA). All means have been compared by Turkey‘s test at 95% probability.
RESULTS Larvicidal activity of volatile oils against P. interpunctella: The results of larvicidal effects showed that all essential oils have toxicant effect on first instar larvae of Indian meal moth. LC50 of essential oils has been showed in table 1. Most and least LC50 of essential oils were observed in lemon balm and lavender about 5.57 and 9.11 µl/l, respectively. All essential oils were very toxicant for first instar larvae of P. interpunctella and LC50 of them were less than 10 µl/l. Ovicidal activity of volatile oils against P. interpunctella: The results of this study revealed that the essential oils exhibited significant ovicidal activity against P. interpunctella. There are significant differences between essential oils at 1% (F=33.12; df= 5, 54; P<0.01). Also there are significant differences between concentrations at 1% (F=322.16; df= 2, 54; P<0.01).
The results showed that by increasing concentration of essential oils, percent of egg mortality increases. Eggs mortality has been recorded in 3, 12 and 24 µl/l concentrations about 29, 42 and 75.38 percent, respectively. This experiment also showed there are significant differences between egg mortality that induced by essential oils. Ziziphora clinopodioides and L. angustifolia had more ovicidal effect about 68.73 and 53.78 percent, respectively. Between other essential oils there are no significant differences, but they have Ovicidal effect about 40-46% (Table 2).
This experiment also shows there is significant difference between egg mortality induced by essential oils with different concentrations (Table 2). Essential oils showed variable toxicity to eggs of P. interpunctella. L. angustifolia, Z. clinopodioides and A. dracunculus essential oils at 24 micro liters per liter concentration had most ovicidal effect about 85.98, 84.24 and 82.75, respectively.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 342
The lowest ovicidal effect has been induced by A. dracunculus, P. sativum and M. officinalis at 3 µl/l. It is observed that in all concentrations Z. clinopodioides had most Ovicidal effect. Effect of essential oils on oviposition of P. interpunctella adults: The results of analyses of variance effect of essential oils on oviposition of P. interpunctella adults showed that there was significant difference at 1% between percent of egg released in different treatments (essential oils) (F=30.97; df= 5, 18; P<0.01). It is observed the least inhibitory of egg released in A. dracunculus treatment. Releasing eggs were inhibited about 36.17%. There were not any significant differences between other essential oils and egg releasing in adults were inhibited in P. sativum, M. officinalis, L. angustifolia, M. piperata and Z. clinopodioides about 82.17, 96.01, 97.50, 98.83 and 99.67 percent, respectively (Figure 1). These results showed except A. dracunculus all essential oils had high deterrence of egg oviposition effect on Indian meal adult as fumigant pesticide.
DISCUSSION
In this study the efficacy of the oils followed in the order Parsley, Lavender, Tarragon, Z. clinopodioides, Lemmon balm, and Peppermint. The LD50 values in case of 24 h after treatment were 9.46, 9.21, 7.21, 6.48, 5.57, and 8.09 on first instar larvae of Indian meal moth, respectively. Regnault-Roger (1997) found LC50 of Lavender and Peppermint on Acanthoscelides obtectus was 12.6 and 22.4. The differences between our results with another researches is about the differences between insects. Mirkazemi (2007) showed LC50 of five essential oils on Tribolium confusum and Callosobruchus maculatus. LC50 of A. dracunculus and L. verra has been achieved about 26.24 and 50.33 on C. maculatus and 9.07 and 19.44 on T. confusum, respectively. In our results LC50 were less than it, it may be the cause of different of insects in two experiments. First instar larvae of P. interpunctella are more susceptible to essential oils.
The results of Raja & William (2008) revealed that the essential oils exhibited significant ovicidal activity against the cowpea beetle. Among the treatment categories citrodora oil showed 88.43% protection followed by citronella oil 58.01% protection. The oils tested showed moderate effect on the eggs of C. maculatus. Bioactive compounds of plant origin having insecticidal and ovicidal properties are being used as grain protection agents against beetles in storage (Raja & William, 2008).
Risha et al., (1990) have reported oil vapors have strong latent effect toxic to adults and eggs of C. pahseoli. Singh et al., (1995) reported plant volatile oils have strong odor that will block the tracheal respiration by S. oryzae in stored sorghum, which ultimately kill the insect (Raja & William, 2008). The highest mortality in the egg chorion and embryo died due to decrease of oxygen for respiration (Raja & William, 2008).
The difference observed among the mortality due to these oils is due to their active volatiles mostly monoterpenes which are very active on insects. The amount of eggs laid by treated adults is significantly lower than control. Some essential oils are able to halt the egg incubation or to kill the emerging larvae. The present work gives evidence on the negative impact of the application of these essential oils on adult‘s oviposition. Lethal concentrations of essential oil need important quantity of material for their extraction (Ngamo et al., 2007). Many environmental factors affect the breakdown of essential oils, most importantly, temperature and light. Limited residual toxicity is an important advantage for these pesticides (Isman, 2000).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 343
The essential oils investigated in this study are used as pharmaceuticals and in flavoring and are therefore considered less harmful to humans than most conventional insecticides and they can use as safe fumigants for controlling P. interpunctella.
LITERATURE CITED Abbott, W. S. 1925. A method for computing the effectiveness of an insecticide. Journal of Economic Entomology, 18: 265-267. Anonymous. 2004. Notice of Proposed Rulemaking- Protection of Stratospheric Ozone: Process for Exempting Critical uses From the Phaseout of Methyl Bromide. Federal Register, 69: 52365-52402. Bruce, T. J. A., Birkett, M. A., Blande, J., Hooper, A. M., Martin, J. L., Bhupinder, K., Prosser, I., Smart, L. E. & Wadhams, L. J. 2005. Response of economically important aphids to components of Hemizygia petiolata essential oil. Pest Management Science, 61: 1115-1121. Chaubey, M. K. 2008. Fumigant toxicity of essential oils from some common spices against pulse beetle, Collosobruchus chinensis (Coleoptera: Bruchidae). Journal of Oleo Science, 57: 171-179. Hamlin, J. C., Reed, W. D. & Phillips, M. E. 1931. Biology of the Indian meal moth on dried fruits in California. USDA Technical Bulletin, 242: 27 pp. Isman, M. B. 2000. Plant essential oils for pest and disease management. Crop Protection, 19: 603-608. Johnson, J. A., Wofford, P. L. & Whithehand, L. C. 1992. Effect of diet and temperature on development rates, survival, and reproduction of the Indian meal moth (Lepidoptera: Pyralidae). Journal of Economic Entomology, 85: 561-566. Lee, S., Lee, B., Choi, W., Park, B., Kim, J. & Campbell, B. 2001. Fumigant toxicity of volatile natural products from Korean spices and medicinal plants towards the rice weevil, Sitophilus oryzae (L). Pest Management Science, 57: 548-553. Mahfuz, I. & Khalequzzaman, M. 2007. Contact and fumigant toxicity of essential oils against Callosobruchus maculates. Journal of Zoology Rajshahi University, 26: 63-66. Mirkazemi, F. 2007. Insecticidal Effects of Essential Oils from Lavandula verra, Rosmarinus officinalis, Foeniculum vulga, Satureja hortensiss, Artemisia dracunculus on Some Stored Product Insects, MSC tesis, Islamic Azad University Arak Branch, 103 pp. Ngamo Tinkeu, L. S., Goudum, A., Ngassoum, M. B., Mapongmestsem, Lognay, G., Malaisse, F. & Hance, T. 2007. Chronic toxicity of essential oils of 3 local aromatic plants towards Sitophilus zeamais Motsch. (Coleoptera: Curculionidae). African Journal of Agricultural Research, 2: 164-167. Obeng-Ofori, D. & Reichmuth, C. 1997. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. International Journal of Pest Management, 43: 89-94. Perez-Mendoza, J. & Aguilera-Pena, M. 2004. Development, reproduction, and control of the Indian meal moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in stored seed garlic in Mexico. Journal of Stored Products Research, 40: 409-421. Phillips, T. W., Berbert, R. C. & Cuperus, G. W. 2000. Post-harvest integrated pest management. In: Francis, F. J. (Ed.), Encyclopedia of Food Science and Technology. 2nd ed. Wiley Inc., New York, pp. 2690–2701. Pushpanathan, T., Jebanesan, A. & Govindarajan, M. 2006. Larvacidal, ovicidal and repellent activities of Cymbopogon citratus Stapf (Graminae) essential oil against the filarial mosquito Culex quinquefasciatus (Say) (Diptera: Culicidae). Tropical biomedicine, 23: 208-212. Raja, M., & William, S. 2008. Impact of volatile oils of plants against the cowpea beetle Callosobruchus maculates (FAB.) (Coleoptera: Bruchidae). International Journal of Integrative Biology, 2: 62-64.
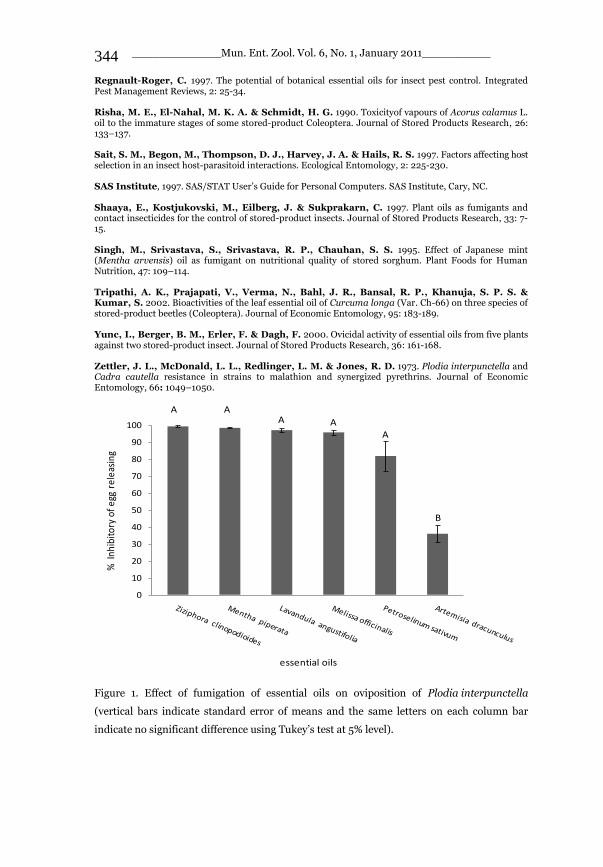
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 344
Regnault-Roger, C. 1997. The potential of botanical essential oils for insect pest control. Integrated Pest Management Reviews, 2: 25-34. Risha, M. E., El-Nahal, M. K. A. & Schmidt, H. G. 1990. Toxicityof vapours of Acorus calamus L. oil to the immature stages of some stored-product Coleoptera. Journal of Stored Products Research, 26: 133–137. Sait, S. M., Begon, M., Thompson, D. J., Harvey, J. A. & Hails, R. S. 1997. Factors affecting host selection in an insect host-parasitoid interactions. Ecological Entomology, 2: 225-230. SAS Institute, 1997. SAS/STAT User‘s Guide for Personal Computers. SAS Institute, Cary, NC. Shaaya, E., Kostjukovski, M., Eilberg, J. & Sukprakarn, C. 1997. Plant oils as fumigants and contact insecticides for the control of stored-product insects. Journal of Stored Products Research, 33: 7-15. Singh, M., Srivastava, S., Srivastava, R. P., Chauhan, S. S. 1995. Effect of Japanese mint (Mentha arvensis) oil as fumigant on nutritional quality of stored sorghum. Plant Foods for Human Nutrition, 47: 109–114. Tripathi, A. K., Prajapati, V., Verma, N., Bahl, J. R., Bansal, R. P., Khanuja, S. P. S. & Kumar, S. 2002. Bioactivities of the leaf essential oil of Curcuma longa (Var. Ch-66) on three species of stored-product beetles (Coleoptera). Journal of Economic Entomology, 95: 183-189. Yunc, I., Berger, B. M., Erler, F. & Dagh, F. 2000. Ovicidal activity of essential oils from five plants against two stored-product insect. Journal of Stored Products Research, 36: 161-168. Zettler, J. L., McDonald, L. L., Redlinger, L. M. & Jones, R. D. 1973. Plodia interpunctella and Cadra cautella resistance in strains to malathion and synergized pyrethrins. Journal of Economic Entomology, 66: 1049–1050.
Figure 1. Effect of fumigation of essential oils on oviposition of Plodia interpunctella
(vertical bars indicate standard error of means and the same letters on each column bar
indicate no significant difference using Tukey‘s test at 5% level).
A AA A
A
B
0
10
20
30
40
50
60
70
80
90
100
% I
nhib
itory
of e
gg r
elea
sing
essential oils
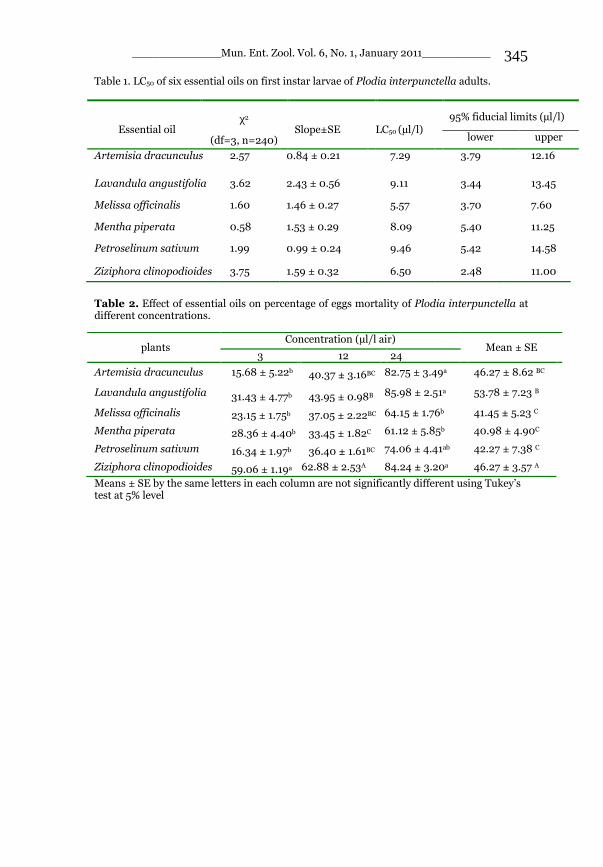
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 345
Table 1. LC50 of six essential oils on first instar larvae of Plodia interpunctella adults.
Essential oil χ2
(df=3, n=240) Slope±SE LC50 (µl/l)
95% fiducial limits (µl/l)
lower upper
Artemisia dracunculus 2.57 0.84 ± 0.21 7.29 3.79 12.16
Lavandula angustifolia 3.62 2.43 ± 0.56 9.11 3.44 13.45
Melissa officinalis 1.60 1.46 ± 0.27 5.57 3.70 7.60
Mentha piperata 0.58 1.53 ± 0.29 8.09 5.40 11.25
Petroselinum sativum 1.99 0.99 ± 0.24 9.46 5.42 14.58
Ziziphora clinopodioides 3.75 1.59 ± 0.32 6.50 2.48 11.00
Table 2. Effect of essential oils on percentage of eggs mortality of Plodia interpunctella at different concentrations.
Means ± SE by the same letters in each column are not significantly different using Tukey‘s test at 5% level
plants Concentration (µl/l air)
Mean ± SE 3 12 24
Artemisia dracunculus 15.68 ± 5.22b 40.37 ± 3.16BC
82.75 ± 3.49a 46.27 ± 8.62 BC
Lavandula angustifolia 31.43 ± 4.77b
43.95 ± 0.98B
85.98 ± 2.51a 53.78 ± 7.23 B
Melissa officinalis 23.15 ± 1.75b
37.05 ± 2.22BC
64.15 ± 1.76b 41.45 ± 5.23 C
Mentha piperata 28.36 ± 4.40b
33.45 ± 1.82C
61.12 ± 5.85b 40.98 ± 4.90C
Petroselinum sativum 16.34 ± 1.97b
36.40 ± 1.61BC
74.06 ± 4.41ab 42.27 ± 7.38 C
Ziziphora clinopodioides 59.06 ± 1.19a
62.88 ± 2.53A 84.24 ± 3.20a 46.27 ± 3.57 A

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 346
IDENTIFICATION OF APTEROUS VIVIPAROUS OF CEREAL APHIDS IN EGYPT
(HEMIPTERA: STERNORRHYNCHA: APHIDIDAE)
A. Helmi* * Department of Plant Protection, Faculty of Agriculture, Ain Shams University, EGYPT. E-mail: [email protected] [Helmi, A. 2011. Identification of apterous viviparous of cereal aphids in Egypt (Hemiptera: Sternorrhyncha: Aphididae). Munis Entomology & Zoology, 6 (1): 346-357] ABSTRACT: Thirteen cereal aphid species represent five subfamilies from different cereal plants in different localities of Egypt were surveyed, described and illustrated. Usual hosts as well as diagnostic characters for both fresh and microscopic mounted materials of each species were provided. Dichotomous and pictorial keys for apterous viviparous females of these aphids were included to aid personnel charged with detetion, identification and control of aphids associated with cereals in Egypt. KEY WORDS: Apterous, viviparous females, cereal aphids, Aphididae, taxonomic key, pictorial key, identification.
Cereal crops of the family Gramineae are the main source of the carbohydrates in the human diet and feed throughout the world. Grains are account for over half of the world's food energy, or even more counting grain consumed indirectly as animal feed. Global agricultural production outpaced the doubling of the world's population occurred between 1950 and 1990.
When aphids' populations are large, they can greatly diminish plant‘s vigor or even kills it through mechanical injury by removal of sap during feeding. Aphids also have the capability of transmitting persistent, semi-persistant and non persistent viruses among different plants that would not otherwise be considered hosts. Aphids also produce a sticky substance (honeydew) during feeding. This substance may be problematic when it fouls the corn tassel and interferes with pollination and encourages fungal growth (Chan et al., 1991 and Clifford et al., 2004). The previous key of aphids in Egypt was presented by Habib & El-Kady, 1961 was based on alate viviparous females only. There are no published keys that include apterous forms, althought these forms are the most settled on the plants and easy to collect, so the objectives of this study were to develop keys and main characters illustrations of apterous viviparous females of cereal aphids to aid personnel charged with detetion, identification and control of aphids associated with cereals in Egypt.
MATERIALS AND METHODS Collecting and preservation:
Apterous viviparous females of different cereal aphid species were collected from different cereal host plants (crops and weeds) from different localities of Egypt throughout two successive years (2007 and 2008). Aphid samples were brought to the laboratory alive on a part of the host plant in paper bags. Aphid colonies often consist of mainly immature individuals, and the proportion of adults in the sample can be increased by keeping the aphids alive for a few days in a cool place before preserving them. Possible collection data were note down at the time of collection, including host plant, locality and date. Also some biological information were noted such as colour of alive aphid individuals, position of

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 347
colony on host plant, and any changes of the host plants that may be due to aphid infestation. Some aphid species were successfully reared on their host plant in the laboratory in Chimeny glass cages for maintenance and mounting. Aphid samples were preserved in tightly stoppered tubes fitted with two volume of 70% lacto-alcohol (ethyl alcohol added to one volume of 75% w/w lactic acid) and few drops of glycerin. Samples mounting:
Aphid samples were washed in 5% glacial acetic acid then embedded in 10% sodium hydroxide over night at room temperature. The second day sodium hydroxide was decanted and add 1cm depth of glacial acetic acid for 5 minutes, then samples were dehydrated by gradually series of ethyl alcohol concentrations. Samples were cleared using chloral-phenol solution (1 vol. chloral hydrate: 1 vol. phenol) and left in this solution overnight at the room temperature. Samples then were ready to be mounted in the swan's gum chloral media on cleaned micro slides and carefully covered by a clean cover slip to avoid any air bubbles, and then the slides left horizontally in a hot plate at 50 oC for 5 days to dry. Labeling and slide storage:
Square pieces of cardboard about 0.5 – 1.0 mm thick are glued to either end of the slide On the left side of the label the scientific name of the aphid, the form and the name of the person who identified the aphid are indicated. The label on the right hand side lists the host plant, locality, date, name of collector and the number of collection. Slides are stored in boxes or drawers in a horizontal position, and the genera are arranged in alphabetical sequence. The species are also arranged alphabetically within the genera. Samples drafting:
Morphological diagnostic characters of apterous viviparous females of different cereal aphid species were drafted using the drawing eye-piece. Some measurements of these characters were made using micrometric slide and lens.
Aphids on Cereals in Egypt I. Aphids on cereal roots:
Two aphids' species were found to infest cereal roots only during this work; Anoecia corni (Fabricius) and Tetraneura africana Van der Goot.
Subfamily Anoeciinae
Anoecia corni (Fabricius, 1775) Dog Wood Aphid
The first record of this aphid species in Egypt by Theobald, 1922 on the cereal weed species Cyperus longus. It was recorded also on different cereal weed plants at Giza and Sharqiya governorates by Elnagar et al., 1978, Amin, 1979 and Megahed, 2000. Also, this aphid species was recorded by Habib & El-Kady, 1961 on wheat plants in El-Gezeirh region. Diagnostic characters: Fresh materials: Tortoise-looks like in shape. Pale greenish-grey with sclerotized parts black. Mounted materials: Frontal tubercles not developed; unguis as twice as the basal part, apical rostral segment acute, cauda circular and siphunculi on hairy cones. Materials: Twenty specimens of this species were collected during this work. Five of them on roots of barley at Sakha, Kafr El-Sheikh Governorate, November 2007 and five specimens on roots of wheat at Shalakan, Qalubiya Governorate, December 2008 and the rest ten specimens were recorded on wheat plants at Giza Governorate, March, 2008.
Subfamily Eriosomatinae
Tetraneura africana Van der Goot, 1912 This aphid species was recorded for the first time in Egypt on Cynodon dactylon roots
by Theobald, 1922. It was recorded after that at Mansoura, Zagazig, Cairo, Giza and Fayoum

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 348
Governorate (Hassan, 1958). It was recorded also at Giza Governorate by other authors (Habib & El-Kady, 1961; Elnagar et al., 1978; Amin, 1979). Diagnostic characters: Fresh materials: Body globose large in size (3 mm), buff colored to pale brown with a dark brown head and prothorax. Dorsum with long spine-like hairs. Mounted materials: Frontal tubercles not developed unguis too short less than the basal part, apical rostral segment acute, cauda broadly rounded and siphunculi ring-like. Materials: Five specimens of this species were collected from roots of Cynodon dactylon at Giza Governorate, February, 2008.
II. Aphids on cereal aerial parts: A- Infesting cereal weeds only:
Three aphids' species were found to infest aerial parts of cereal weeds only during this work; Hyalapterus pruni (Geoffroy) on Arundo donax only. While the other two species, Saltusaphis scirpus Theobald and Schizaphis minuta (Van der Goot) on Scirpus, Cyperus rotundus only.
Subfamily Aphidinae
Hyalapterus pruni (Geoffroy, 1762) Mealy Plum aphid
This aphid species was recorded for the first time in Egypt by Willcocks, 1916 on both Arundo donax and Phragmites comnunis and then recorded at Giza Governorate by Elnagar et al., 1978 and Amin, 1979. Diagnostic characters: Fresh materials: Body elongate, green or pink, covered with white powdery wax. Mounted materials: Frontal tubercles not well developed; unguis twice as long as the basal part, apical rostral segment acute, cauda elongate with five long hairs, siphunculi cylindrical and shorter than the cauda. Materials: Fourty specimens of this species were collected all on Arundo donax from Qalyabiya, Giza, Sharqiya and Behaira Governorates during March and April, 2008.
Schizaphis minuta (Van der Goot, 1917)
Cyperus Aphid This species was recorded for the first time in Egypt by Habib and El-Kady, 1961 using a
light trap. There wasn‘t any record about it until it was observed after thirty nine years on Cyperus rotundus by El-Fatih, 2000 and 2006. Diagnostic characters: Fresh materials: Very small in size (1.2mm: 1.5mm). Dark-olive in colour. Legs yellow with black distal parts. Mounted materials: Frontal tubercles not well developed, unguis five times as long as the basal part, apical rostral segment acute, cauda elongate with about 6 long hairs, siphunculi cylindrical about twice as long as the cauda. Materials: Six specimens were collected during this work all on Cyperus, C. rotundus from Giza and Qalyabiya Governorates in December, 2007 and January, 2008.
Subfamily Saltusaphidinae Saltusaphis scirpus Theobald, 1915
It was recorded for the first time in Egypt by Willcocks, 1910 on Sedge ―Scirpus sp.‖. Also it was caught on light trap in 28 Dec. 1958 at Kobba Palace and four specimens were recorded on Cyperus sp. 7 Dec. 1944 at Dokki region by Habib and El-Kady, 1961. Then it was recoded at Zagazig, Sharkiea Governorate (Megahed, 2000) and at Giza Governorate on Cyperus rotundus by El-Fatih 2000 and 2006. Diagnostic characters: Fresh materials: Pale grey-brown with dark brown inter-segmental markings legs, peculiarly characterized by the Cat-like setting when rest. Considered the fastest cereal aphid species whereas it springs off the plant on the slightest alarm.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 349
Mounted materials: Frontal tubercles not developed Antennae almost reaching body length, unigus as 1.5 long as the basal part, apical rostral segment short and blunt, cauda knobbed and anal plate bilobed and siphunculi short on hairy cones. Materials: Five specimens were collected during this work all on Scirpus, C. rotundus from Qalyabiya in January, 2008 and from Giza, March, 2008.
B. Infesting cereal crops and weeds: Eight cereal aphid species were found to infest both cereal crops and
weeds during this work.
Subfamily Aphidinae Diuraphis noxia (Mordvilko, 1914)
Russian Wheat Aphid It was mentioned that the first record of this aphid species in Egypt was recorded by
Attia, 1988. But during this revision study it was noticed that Diuraphis noxia was the same species which was found on the leaves of wheat in March 1957 at Koubba Palace by (Habib and El-Kady, 1961) and they defined it as Cuernavaca noxia (Mordvilko. It was recorded after that by many authors on wheat and barley plants at both of Beni-Suef (El-Lathy, 1999); Ismailia (Noaman et al., 1992); Sinai (Abd El-Salam, 1999). Also it was recorded on B. catharticus and A. fatua cereal weed plants by El-Fatih, 2000. Diagnostic characters: Fresh materials: Small in size, spindle-shaped, pale yellow green or grey-green covered with white wax powder. Mounted materials: Frontal tubercles not well developed, unigus twice as long as the basal part, apical rostral segment acute, supra-caudal process present on the eighth abdominal tergum, that about as long as the cauda, siphunculi short and cube shaped. Materials: Fifteen specimens were collected during this work. Six of them were on wheat plants at Giza Governorate. Five of them were on Bromus catharticus and four were on Avena fatua.
Metopolophium dirhodum (Walker, 1849) Rose-grass aphid, Rose-grain aphid
This aphid species was recorded at the first time in Egypt on wheat plants at Beni-Suef Governorate (El-Lathy, 1999) and on barley and different cereal weed plants at Giza Governorate (El-Fatih, 2000, 2006). Diagnostic characters: Fresh materials: Body green to yellow green with dark green longitudinal stripe along the middle of the dorsum. Small to medium sized, body length 1.7-2.7 mm, elongate, antennae green with dark bands on apices of antennal segments 3, 4 and the base of 6. Cauda and siphunculi pale. Mounted materials: Frontal tubercles well developed (Diverging); unigus 4 times as long as the basal part, apical rostral segment acute, Cauda elongate with about 5 pairs of long setae, siphunculi cylindrical about twice as long as the cauda Materials: Thirty specimens were collected during this work. Ten of them were on barley plants at Giza Governorate in February, 2007. Five of them were on Lolium temulentum January, 2008. Six of them were on Phalaris minor January, 2008. Eight specimens were on Avena fatua April, 2008.
Rhopalosiphum maidis (Fitch, 1856) Corn-leaf aphid
The first record of this aphid species in Egypt was on the cereal weed plant, Andropogon halepensis (Willcocks, 1922). Then it was recorded on cereal weeds at both of Giza Governorate (Megahed et al., 1978, Elnagar et al., 1978, Amin, 1979, Abdel-Wahab, 1998, El-Fatih 2000 & 2006); Zagazig region, Sharkiea Governorate (Megahed, 2000) and Kharga and Dakhla Oases (Hassan, 1957).
It was recorded also on wheat plants at both of Kharga & Dakhla Oases (Hassan, 1957); Beni-Suef, Assiut & Sohag (Mohamed, 1992); Assiut (Slman, 1993 & 1997); Sinai (Abd El-Rahman, 1997) and Beni-Suef (Abd El-Salam, 1999 and El-Lathy, 1999).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 350
It was recorded also on barley plants at Middle Egypt (El-Hariry, 1979); Giza (Ibrahim, 1990, Abd El-Salam, 1999 and El-Fatih, 2000 & 2006); Sohag (Slman & Ahmed, 2005); Ismailia (Noaman et al., 1992) and both of Nubaria, Borg El-Arab and Marsa Mattrouh (El-Sayed et al., 1995).
It was recorded also on maize plants (Willcocks, 1922 & Hall, 1926). Then it was recorded at Kharga & Dakhlia Oases (Hassan, 1957); Qalubia (Yossef, 1990); Giza (Abd El-Salam, 1999) and Gharbiah (El-Khouly et al., 1994). Diagnostic characters: Fresh materials: Body rather elongate (1.9-2.6 mm) with short antennae and siphunculi, color blue green to olive green with reddish-purple areas around siphunculi bases. Siphunculi and cauda dark. Mounted materials: Frontal tubercles not well developed. Antenna 6 segmented; unigus twice as long as the basal part. Apical rostral segment acute. Cauda elongate with 2 pairs of lateral setae. Siphunculi slightly swollen, 4.5 times as long as the length of cauda. Materials: Forty specimens were collected during this work. Ten of them were on barley plants at Giza Governorate in December, 2007. Ten of them were on Avena fatua during February, 2008. Seven of them were on Lolium temulentum in March, 2008. Thirteen specimens were on Phalaris moinor May, 2008 at Giza Governorate.
Rhopalosiphum padi (Linnaeus, 1758) Bird Cherry-Oat Aphid
The first record of this aphid species in Egypt was on the cereal weed plants, Panicum colonum and Phalaris sp. (Hassan, 1958). It was recorded also on another different cereal weed plants at Giza Governorate by many authors (Megahed et al., 1978, Elnagar et al., 1978, Amin, 1979, Abdel-Wahab, 1998, El-Fatih 2000 & 2006), while it was recorded at Zagazig region, Sharqiya Governorate (Megahed, 2000). The first record of this aphid species on wheat plants in Egypt was at Koubba palace (Habib & El-Kady, 1961). Then it was recorded at Beni-Suef, Assiut & Sohag (Mohamed, 1992); Assiut (Slman, 1993 & 1997); Sinai (Abd El-Rahman, 1997); Beni-Suef (Abd El-Salam, 1999 and El-Lathy, 1999).
It was recorded also on barley plants at Middle Egypt (El-Hariry, 1979); Giza (Ibrahim, 1990; El-Fatih, 2000); Abd El-Salam, 1999 and El-Fatih 2000 & 2006; Beni-Suef, Assiut and Sohag (Mohamed, 1992), Sohag (Slman & Ahmed, 2005); Ismailia (Noaman et al., 1992) and both of Nubaria, Borg El-Arab and Marsa Mattrouh (El-Sayed et al., 1995), Sinai (Abd El-Salam, 1999). It was recorded also on maize plants at Qalubia Governorate (Yossef, 1990) and at Sinai (Abd El-Salam, 1999). Diagnostic characters: Fresh materials: body broadly rounded, small to medium sized (2: 2.4 mm), colour yellowish to bluish green with orange patches around base of siphunculi. Siphunculi and cauda dark Mounted materials: frontal tubercles not well developed. Antennae 6 segments; unigus four times as long as the basal part, apical rostral segment blunt. Cauda elongate with four lateral setae. Siphunculi cylindrical, twice as long as the length of cauda. Materials: Forty specimens were collected during this work. Thirteen specimens on Avena fatua from Giza and Qalyabiya Governorates in January, 2007 and twenty specimens were on barley in March, 2008. Seven specimens were on wheat plants from Beni-Suif Governorate, April, 2008.
Schizaphis graminum (Rondani), 1852. Green Bug
The first record of this aphid species in Egypt was on the cereal weed plant, (Willcocks, 1922). After that it was recorded on other cereal weeds (Megahed et al., 1978, Elnagar et al., 1978, Amin, 1979, Abdel-Wahab, 1998, El-Fatih 2000 & 2006, while it was recorded at Zagazig region, Sharqiya Governorate by Megahed, 2000. Also, it was recorded on barley plants by El-Hariry, 1979, Ibrahim and Afifi, 1991, Abd El-Salam, 1999 and El-Fatih 2000 & 2006. Diagnostic characters: Fresh materials: Body elongate oval shaped; small in size (1.5 to 2 mm), yellowish to bluish green with a darker spinal stripe on the dorsum of abdomen, head and prothorax yellowish green. Siphunculi pale with slightly flared and darkened tips.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 351
Mounted materials: Frontal tubercles not well developed. Antennae 6 segmented; unigus 3 times as long as the basal part. Apical rostral segment acute. Cauda elongate with 4 long lateral setae. Siphunculi cylindrical, about 1.6 as long as the length of cauda. Materials: Thirty five specimens were collected during this work. Fourteen specimens were recorded on Avena fatua from Giza and Beni-Suef Governorates in February, 2007 and sixteen specimens were on Cynodon dactylon in January, 2008.
Schizaphis rotundiventris Signoret, 1860 Oil pulm aphis
This aphid species was recorded as a first time in Egypt by Theobald, 1922 as a new species under the name Aphis acori on Sedge, Cyperus longus. Then it was recorded by Hall, 1926 under the name Toxoptera acori. After approximately seventy four years it was observed at Giza Governorate on different cereal weed plants by El-Fatih 2000, Abdel-Wahab, 2004 and El-Fatih 2006 under the name Schizaphis cyperi.
Borner and Heinze (1957) stated that Schizaphis cyperi (Van der Goot) (Ainslie) is a junior synonym of S. rotundiventris Borner and Heinze list (Ainslie) after Van der Goot as the describer, evidently synonymizing the two species. Diagnostic characters: Fresh materials: Body pear shaped; small in size (1.3 to 1.8 mm). Dark-red, antennae and legs yellow with black bases and tips. Mainly infesting cereal weeds while rarely on economic crops Mounted materials: Frontal tubercles not well developed. Antennae 6 segmented; unigus 5 times as long as the basal part. Apical rostral segment blunt. Cauda elongate with 4 long lateral setae. Siphunculi cylindrical, about 1.6 as long as the length of cauda. Materials: Twenty five specimens were collected during this work. Eight specimens on Digitaria sanguinalis from Giza Governorates in November, 2007 and five specimens were on Cynodon dactylon in March, 2008 from Giza Governorates. Seven specimens were on Echinocloa colonum plants and five of them were on Sorghum virgatum from Beni-Suif Governorate, April, 2008.
Sitobion avenae (Fabricius, 1775) English Grain Aphid
The first record of this aphid species in Egypt was recorded on barley plants by Willcocks, 1922. Then it was recorded also by Hall, 1926. After that it was recorded at Giza region by both of El-Hariry, 1979; Ibrahim & Afifi, 1991, El-Fatih 2000 and 2006. It was recorded at Sinai by Abd El-Salam, 1999.
It was recorded also on different cereal weed plants at Giza Governorate by many authers (Hall, 1926; Megahed et al., 1978; Elnagar et al., 1978, El-Fatih, 2000 and 2006, while it was recorded at both of Zagazig region, Sharqiya Governorate by Megahed, 2000.
The first record of this aphid species on wheat plants was at Koubba palace (Habib & El-Kady, 1961). Then it was recorded at Beni-Suef, Assiut & Sohag (Mohamed, 1992), Assiut (Slman, 1993 & 1997); Sinai (Abd El-Rahman, 1997); Beni-Suef (Abd El-Salam, 1999 and El-Lathy, 1999). Diagnostic characters: Fresh materials: Body broadly spindle-shaped about 2.2-3.5 mm in length; antennae, legs and siphunculi long. Dorsum bright green to red brown in colour, with ten faint inter-segmental markings. Leg segments black towards ends while antennae and siphunculi uniformly black. Mainly infesting economic cereal crops Mounted materials: Frontal tubercles well developed (Diverging). Antennae 6 segmented; unigus 3 times as long as the basal part. Apical rostral segment acute. Cauda elongate with 6 of lateral setae and 1-2 pre-apical setae. Siphunculi cylindrical much longer than cauda and with a sub-apical polygonal reticulated apex. Materials: Thirty two specimens were collected during this work. Fifteen of them were on barley plants at Giza and Qalyabiya Governorate in December, 2007. Six of them were on Avena fatua in March, 2007. Seven of them were on Lolium temulentum in March, 2008. Four specimens were on S. virgatum in May, 2008 at Giza Governorate.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 352
Subfamily Chaitophorinae Sipha (Rungsia) maydis Passerini, 1860
It was recorded before as a first time in Egypt at Giza Governorate on Aegilops sp. (El-Hariry, 1991). Then it was recorded on other different cereal weed plants (El-Fatih 2000). Diagnostic characters: Fresh materials: Body pear-shaped, somewhat dorsoventrally flattened, rather small (1.3-1.6mm). Shining dark-brown to almost black on dorsum; which fully sclerotized and armored with long spine-like hairs. Live on upper sides of leaf blades near the bases. Mounted materials: Frontal tubercles not well developed. Antennae 6 segmented; unigus 5 times as long as the basal part. Apical rostral segment acute. Cauda broadly rounded with 5 long hairs. Siphunculi ring-like. Materials: Eleven specimens were collected during this work. Four specimens on Bromus catharticus from Giza Governorates in April, 2007. Seven specimens were on Sorghum virgatum in February, 2008 from Qalyabiya Governorates. Key of cereal aphids in Egypt based on fresh materials:
1. Infesting cereal roots only…………………………………………………………………………. 2 - Infesting aerial parts…………………………………………………………………………………. 3 2(1). Tortoise-looks like in shape. Pale greenish-grey with sclerotized parts black.
Dorsum hairs inconspicuous. Infesting cereal weeds and crops…….…A. corni
- Globose in shape. Buff colored to pale brown with dark brown head and prothorax. Dorsum with long spine like hairs. Infesting cereal weeds and rarely on cereal crops…………………………………………….………..….....T. africana
3. Infesting cereal weeds only………………………………………………………………………... 4 - Infesting both cereal weeds and crops………………………………………………………... 6 4(3). Body elongate, green or pink, covered with white powdery wax. Found
infesting Arundo donax only……………………….………………………..…….H. pruni
- Body not covered with any wax. Found infesting Cyperus rotundus only………. 5 5(4). Pale grey-brown with dark brown inter-segmental markings legs, peculiarly
characterized by the Cat-like setting when rest. Considered the fastest cereal aphid species whereas it springs off the plant on the slightest alarm……………………………………………………………………………………….S. scirpus
- Dark-olive in colour. Legs yellow with black distal parts…........……..S. minuta 6(3). Tip of abdomen with double "tail" (supra caudal process)……………..D. noxia - Tip of abdomen with cauda only……………………………………………………….………... 7 7(6). Body fully sclerotized and armored with long spine like hairs…….…S. maydis - Body not sclerotized, dorsum hairs short and inconspicuous……………………….... 8 8(7). Mainly on plant ears and rarely on leaves………………………………………..………….. 9 - Infesting leaves only……………………………………………………………………………….…. 10 9(8). Bright-green, antennae and legs completely black. Mainly infesting economic
cereal crops…………………………………..………………………………………..…S. avenae
- Dark-red, antennae and legs yellow with black bases and tips. Mainly infesting cereal weeds while rarely on economic crops……………...….S. rotundiventrus
10(9). Dorsum with distinct longitudinal mid-dorsal stripe…………………………………… 11 - Dorsum without any stripe…………………………………………………………………………. 12 11(10). Light green, antennae uniformly black….....................................S. graminum - Yellow green, antenna green with dark bands on apices of antennal segments
3, 4 and the base of 6……………………………………………………....…..M. dirhodum
12(10). Olive-green, antennae and siphunculi with dark tips. Siphunculi base with reddish-orange spot……………………………………………………………………....R. padi
- Bright-green, antennae and siphunculi completely black. Siphunculi base without spots………………………………………………………………………...…R. maidis
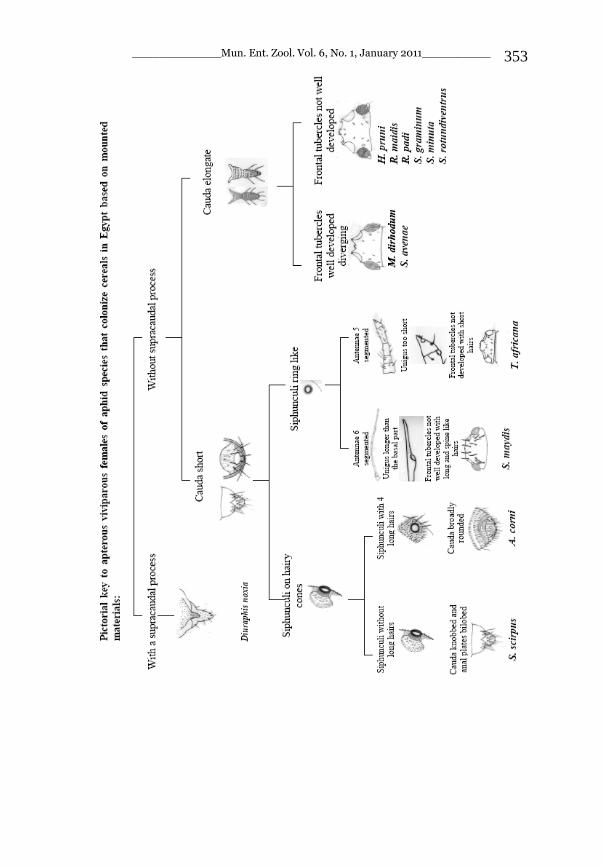
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 353
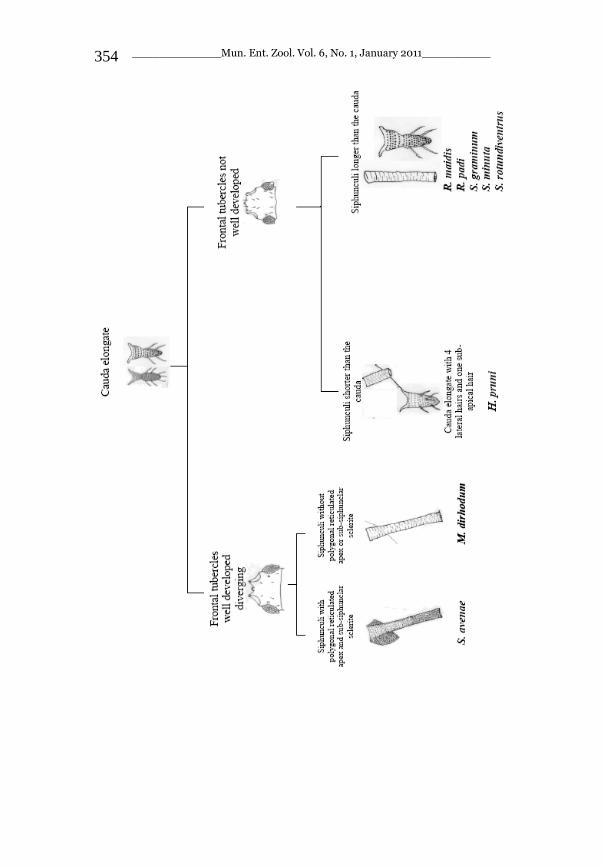
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 354
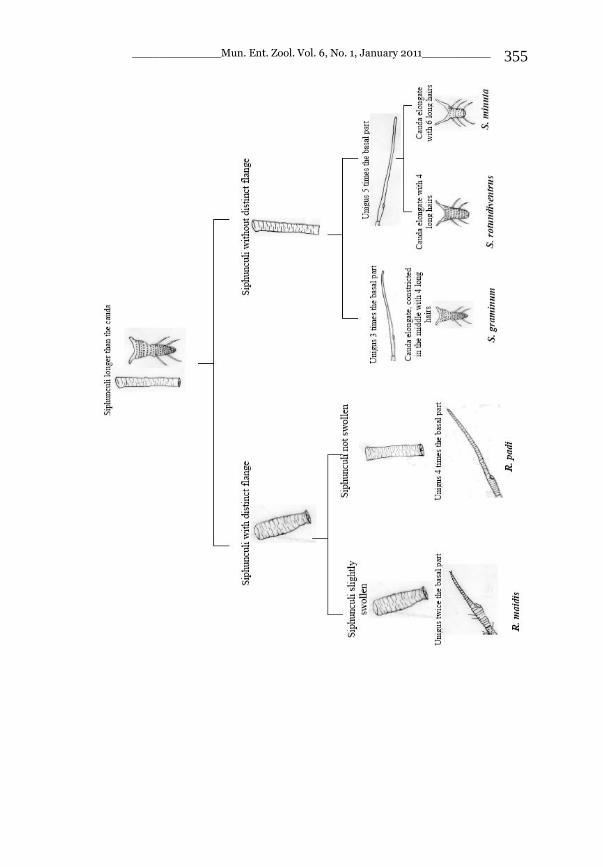
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 355

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 356
LITERATURE CITED Abd El-Rahman, M. A. A. 1997. Biological and ecological studies on cereal aphids and their control in Upper Egypt. M. Sc. Thesis, Fac. Agric., Assiut Univ., Egypt. 231 pp. Abd El-Salam, S. A. 1999. Studies on the aphid fauna of Sinai Governorates. Ph. D. Thesis, Fac. Agric., Cairo Univ., Egypt. 222 pp. Abdel-Wahab, A. S. E. 1998. Aphid species and aphid-borne viruses associated with faba bean in Egypt. M. Sc. Thesis, Fac. Agric., Cairo Univ., Egypt.147 pp. Abdel-Wahab, A. S. E. 2004. Insects and Insect-Norne viruses associated with alliaceae crops in Egypt. Ph. D. Thesis, Fac. Agric. Cairo Univ., Egypt, 250 pp. Amin, A. H. 1979. Studies on the aphid fauna of wild plants in Egypt with special reference to aphid-borne viruses. M. Sc. Thesis, Fac. Agric., Cairo Univ., Egypt. Attia, A. A. 1988. Diuraphis noxia (Mordvilko) (Homoptera: Aphididae): A recent addition to the aphid fauna of Egypt. Bull. Soc. Entomol. Egypt, 68: 267-237. Borner, C. & Heinze, K. 1957. Aphidina-Aphidoidea. pp. 1-402. In H. Blunck, Ed. Teirische Schadlinge an Nutzpflanzen. 2. Teil. 4. Lieferung. P. Parey, Berlin. El-Fatih, M. M. 2000. Cereal aphids in Egypt and their impact on wheat. M. Sc. Thesis, Fac. Agric., Cairo Univ., Egypt., 146 pp. El-Fatih, M. M. 2006. Seasonal abundance and certain biological aspects of cereal aphids on barley in Egypt (Giza region). Ph.D. Thesis, Fac. Agric., Cairo Univ., Egypt, 146 pp. El-Hariry, M. A. 1979. Biological and ecological studies on aphids attacking corn and wheat in Egypt. M. Sc. Thesis, Fac. Agric., Ain-Shams Univ., Egypt. 187 pp. El-Hariry, M. A. 1991. Record of Sipha maydis (Passerini) on Aegilops in Egypt. Fourth Arab Congress of plant protection, Cairo 1-5 Dec. El-khouly, A. S., Ali, M. A., Ibrahim, I. & Naga, S. A. 1994. Effect of inter cropping maize and cowpea on their susceptibility to infestation with aphids. Bull. Soc. Entomol. Egypt, 72 (220). El-Lathy, K. H. 1999. Integrated management of aphids on wheat crop. Ph. D. Thesis, Environmental Studies and Research Institute, Ain-Shams Univ., Egypt. 123 pp. Elnagar, S., Megahed, M. M. & Amin, A. H. 1978. The aphid fauna of wild plants in Giza, Egypt. Bull. Soc. Entomol., Egypt, 62: 219-225. El-Sayed, A. A., Bishara, S. T., Noaman, M. M. & El-Defrawi, G. M. 1995. Survey and field screening of aphids affecting barley. Egypt. J. Agric. Res., 73 (4). Habib, A. & El-kady, E. A. 1961. The Aphididae of Egypt. Bull. Soc. Entomol. Egypt, 45:1-137 pp. Hall, W. J. 1926. Notes on the Aphididae of Egypt. (Tech. and Sci. Serv.), Min. Agric. Egypt, Bull., 68: 1-62. Hassan, M. S. 1957. Studies on the damage and control of A. maidis in Egypt. Bull. Entomol. Soc. Egypt, XLI: 213-230. Hassan, M. S. 1958. Root aphids of Egypt. Ph. D. Thesis Fac. Agric., Cairo Univ., Egypt. Ibrahim, A. M. A. 1990. Corn leaf aphid, Rhopalosiphum maidis (F) (Hom., Aphididae) on wheat and associated primary parasitoids and hyperparasitoids. Bulletin de la Societe Entomologique d'Egypt, 69: 149-157. Ibrahim, A. M. A. & Afifi, A. I. 1991. The relationship between the cereal aphids and aphidophagous syrphids, coccinellids and chrysopids on wheat and barley in Egypt. Bull. Fac. Agric., Cairo Univ., 42 (1): 151-166. Megahed, H. E. A. 2000. Studies on aphids. Ph. D. Thesis, Fac. Agric., Zagazig Univ., Egypt, 229 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 357
Megahed, M. M., Elnagar, S. & Amin, A. H. 1978. Seasonal abundance of four cereal aphids on wild plants in Giza, Egypt. Bull. Soc. Entomol. Egypt., 62: 227-230. Mohamed, M. A. 1992. Ecological and biological studies on wheat insect pests in Egypt. Ph. D. Thesis, Fac. Agric. Al-Azhar Univ. Egypt. 144 pp. Noaman, M. M., Bishara, S. I., El-Sayed, A. A, El-Hariry, M. A. & Miller, R. H. 1992. A field survey of aphids infesting barley in Egypt with results of field and laboratory screening for aphid resistance. Assiut J. Agric. Sci., 23 (1): 303-309. Rondani, C. 1852. Aphis graminum n. sp. Nuove Ann. Sci. Nat. Bologna. 6: 9-11. Slman, F. A. A. 1993. Studies on some aphid species infesting wheat plants and their natural enemies in Upper Egypt. M. Sc. Fac. Agric., Minia Univ., Egypt. 85 pp. Slman, F. A. A. 1997. Studies on certain factors affecting the distribution of cereal aphids in wheat fields in Upper Egypt, Ph. D. Thesis, Fac. Agric., Minia Univ., Egypt. 178 pp. Slman, F. A. A. & Ahmed, M. A. 2005. Seasonal abundance of cereal aphids and ladybird beetle, Coccinella undecimpunctata (L.) on four cereal crops in South Egypt. Assiut Journal of Agricultral Science, 36 (4): 205-215. Theobald, F. V. 1922. New Aphididae found in Egypt. Bull. Entomol. Res., VII.: 39. Willcocks, F. C. 1916. Some notes on the mealy plum aphid: Hyalopterus pruni Fabricius. Bull. Soc. Roy. Entomol. Egypt, IV, p.33. Willcocks, F. C. 1922. A survey of the more important economic insects and mites in Egypt. Sult. Agric. Soc., Cairo. Willcocks, F. C. 1925. The insect and related pests of Egypt, Vol (2), Insects and mites feeding on gramineous crops and products in the field, granary and mill. Sult. Agric. Soc., Cairo. Youssef, E. Y. 1990. Eological and biological studies on maize aphid insects. M. Sc. Thesis, Fac. Agric., Ain-Shams Univ. Egypt. 249 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 358
STUDY OF THE GENUS UROPHORA ROBINEAU-DESVOIDY, 1830 (DIPTERA: TEPHRITIDAE) IN ECEBSIR REGION WITH
TWO SPECIES AS NEW RECORDS FOR IRAN
Yaser Gharajedaghi*, Samad Khaghaninia*, Reza Farshbaf Pour Abad* and Murat Kütük**
* Dept. of Plant Protection, Faculty of Agriculture, University of Tabriz, IRAN. E-mail: [email protected] ** University of Gaziantep, Faculty of Science and Art, Department of Biology, 27310, Gaziantep, TURKEY. [Gharajedaghi, Y., Khaghaninia, S., Pour Abad, R. P. & Kütük, M. 2011. Study of the genus Urophora Robineau-Desvoidy, 1830 (Diptera: Tephritidae) in Ecebsir region with two species as new records for Iran. Munis Entomology & Zoology, 6 (1): 358-362] ABSTRACT: Based on specimens collected from Ecebshir area during 2009-2010, five species of Urophora were recognized of which two species Urophora affinis and U. solstitialis are new records for the Iran insect fauna. Identification key, the locality, host plants as well as photos of verified species are provided. KEY WORDS: Tephritidae, Fruit flies, Myopitinae, Urophora, Ecebsir, Iran.
Ecebsir region is located in south western of East Azerbayjan province, close to eastern beach of the Urumiyeh Lake with UTM (Universal Transfer Mercator) coordinate system, X from 572964.47 to 599802.25 E; Y from 4147773.18 to 4161843.04 N and varying latitude from 1744 m to 2113 m. This area has rich grass lands with various species of Astraceae, Umblifera, Legominaceae and Ronunculaceae.
Tephritidae (true fruit flies) is a large family of the order Diptera with more than 4400 described species around the world. Considering their damage on fruit plantations, they are important insects from the agricultural point of view as well as forest entomology (Merz, 2001).
The genus Urophora includes about 100 species, sometimes known as gall flies, most of which are Palaearctic. However, the genus also includes seven Nearctic species, thirty-two Neotropical species, five Afrotropical species and two Oriental species (White & Korneyev, 1989).
The genus was first named by Meigen (1800) as Euribia, to include eighteen unlisted species Later, Urophora was described by Robineau Desvoidy (1830), who included Musca cardui Linnaeus, which was subsequently designated as the type species by Westwood (1840) Between 1835 and 1855 (White & Korneyev, 1989).
Morphologically, proboscis elongated with narrow labella, posteromarginal wing cells (alar cells) not tapering toward apex; M1+2 ending in alar margin near apex (Richter, 1970).
Some Urophora species are practical or potential agents for the biological control of composite (Asteraceae) plants of Palaearctic origin which have become noxious to weeds. The species which have been successfully established are U. affims and U. quadnfasaata for Centaurea maculosa and C. diffusa, U. cardui for Cirsium arvense, and U. stylata for Cirsium vulgar. Two additional species are currently being considered as potential biocontrol agents, namely U. urunaseva for Centaurea solstitiahs, and U. solstitialis for Carduus acanthoides and C. nutans (Harris 1984a,b; Harris & Wilkinson, 1984; White & Clement, 1987 and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 359
White & Korneyev, 1989). Since the Urophora species in Ecebsir region have not been investigated thus it subjected for the present study.
MATERIALS AND METHODS
Adult specimens were swept weekly on flowers head of Asteraceus plants in twenty-three localities which situated through the working area during 2009- 2010 (Fig. 1). The samples were killed in a killing jar containing potassium cyanide and the voucher specimens were deposited at Insect Museum of Tabriz University. The species were identified based on valid keys (Freidberg & Kugler, 1989; Freidberg & Mathis, 1986; Richter, 1970).
RESULTS
Five species of the genus Urophora were identified which infested the head of Astraceus plants. Two of them marked with an asterisk are being newly reported for the Iran insect fauna and all of them listed as follows: *Urophora affinis, U. impicta, U. jaceana, U. quadrifasciata and *U. solstitialis. Key to studied species of the genus Urophora:
1. Wings with 3 or 4 distinct crossband (Fig. 2; a, b, c, d)………………………………………2 - Wings without distinct crossband (Fig. 2; e).……………….……………………..….U. impicta 2. Wings with 3 distinct crossband (Fig. 2; a)……………………...…….……………..…U. affinis - Wings with 4 distinct crossband (Fig. 2; b, c, d)………...……………………....…….………….3 3. Aculeus with subapical steps (Fig. 3; a, b)…………..…………….………………………………...4 - Aculeus without subapical steps (Fig. 3; c)……….……………….…………U. quadrifasciata 4. Femora yellow, whit black stripes …………………...……………….……………….U. solstitialis - Femora yellow, whitout black stripes……………......…………………………….……U. jaceana
Species Urophora Robineau-Desvoidy, 1830 in Ecebsir:
Urophora affinis Frauenfeld, 1857 Material examined: 5 specimens (3♂♂, 2♀♀): Gunbed; 37°30' N 46°01' E, 1437 m, 13 February 2010 (collected by Y. Gharajedaghi, Depasited at Insect Museum of Tabriz University). Host plants: Centaurea spp. (Korneyev & With, 1993). Distribution: Central Europe, east to Afghanistan; introduced to western North America (White & Korneyev, 1989). New record for the Iran fauna. Diagnostic characters: U. affinis affinis; Head. first flagellomere orange; palpi yellow, darkening to orange api-cally; palpi shape similar to U.cardui; gena about 0.3-0.4 times eye height; labellum about 1.5 times length of first flagellomere. Thorax. scutum with a dense tomentum which obscures the underlying cuticle; postpronotal lobe largely yellow; basal scutellar seta on or near margin of central yellow and marginal black area; femora orange. Wing. base hyaline; subbasal crossband absent, or represented by a faint mark extending between veins Ri and Ai; discal crossband sometimes broken into spots; discal and preapical crossbands separate; hyaline area between preapical and discal crossbands 3-4 times as broad along vein R4+5 as breadth of preapical crossband on vein R4+5; preapical and apical crossbands separate (Fig. 2; a). Urophora impicta Hering, 1942 Material examined: 1 specimens (1♀). Host plants: Cousinia eryngioides (Korneyev & White, 1993). Distribution: Russia, Turkmenistan, Afghanistan (Thompson, 1998).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 360
Urophora jaceana Hering, 1935 Material examined: 9 specimens (7♂♂, 2♀♀). Host plants: Centaurea jacea L., and C. nigra L. (White & Korneyev, 1989). Distribution: Most of Europe; from Ireland and France in the West and Norway and Northern Russia (Kola Peninsula) in the North to the Pyrenees, Alps, Caucasus, and Turkey in the South and Ural Mountains in the East; introduced into Eastern America (Korneyev & White, 1999; Kutuk, 2003). Urophora quadrifasciata Meigen, 1826 Material examined: 2 specimens (1♂, 1♀). Host plants: The larvae develop in flowerhead galls on Centaurea iberica Trev. ex Spreng, C. solstitialis L., C. aspera L., C. sterilis L., C. breviceps Trev. ex Spreng., C. nicaeensis Trev. ex Spreng., C. procurrens Sieb., C. jacea L., C. maculosa Lam., C. splendens Sieb., C. nigrescens L., C. cyanus L, C. calpitropa L., C. nigra L., and Serratula tinctoria L. (Giray, 1979; White, 1988; White & Korneyev, 1989; Freidberg & Kugler, 1989; Merz, 1994; Korneyev & White, 2000; Kutuk, 2003). Distribution: Most of Europe, Turkey, Kazakhstan, Israel, Iran, East Palaearctic and Nearctic region, and North Africa (Kutuk & Ozgur, 2003; Merz & Korneyev, 2004). Urophora solstitialis Linnaeus, 1758 Material examined: 1 specimens (1♀): Zaviyeh; 37°29' N 45°52' E, 1320 m, 24 March 2010 (collected by Y. Gharajedaghi, Depasited at Insect Museum of Tabriz University). Hostplants: Carduus defloratus L., C. nutans L., C. personata (L.) Jacq., C. acanthoides L., C. crispus L., Cirsium vulgare (Savi) Ten., and C. heterophyllum (L.) Hill. (White, 1988; Merz, 1994; Kutuk, 2003). Distribution: Most of Europe, except Spain and Mediterranean region (Merz and Korneyev, 2004), North Caucasus, Armenia, Turkey, Kazakhstan, Western China (Xingjian), Far East Russia (Magadan) (Korneyev and White, 1999), North America, Australia, and New Zealand (Norrbom et al., 1999). New record for the Iran fauna. Diagnostic characters: Head. Height of gena 0.3-0.5 height of eye. Thorax. Bases of b scut at border of yellow, medial and lateral black areas of scutellum, only in some montane spms. on black background. Femora yellow, with black stripes or spots, except some montane spms. with almost entirely black femora; in most spms. only femora with black stripe and often without it. Wing. Bands: subbasal band distinct, black, extending from anterior margin of wing to A\, slightly posterior to it; from discal band isolated at C by yellow interval; hyaline interval between discal and preapical bands approximately twice as wide as preapical band at level of R4+5; preapical and apical bands fused to Rj+i or isolated (Fig. 2; b). Terminalia. Apex of aculeus as in (Fig. 3; a), Distance between primary and secondary processes of blade considerably greater than 1/14 (1/8-1/12) total length of blade and greater than width of apex of blade distal of primary steps or (rarely) as long as width of apex of blade. WL ♀ = 3.8±0.34 (3.0-4.6), AL = 2.8+0.42 (1.9-3.6), AL/WL = 0.71+0.11 (0.49-0.92) (n = 64). In spms. from N Russia (probably associated with Carduus crispus only) blade of ovipositor somewhat shorter than average (AL = 2.3+0.25, AL/WL = 0.58±0.04) and in populations from S regions (associated with different species of Carduus, including C. nutans aggr. with large inflorescence), slightly longer (AL = 3.1±0.32, AL/WL = 0.80±0.04), (Korneyev & White, 1993).
LITERATURE CITED Freidberg, A. & Kugler, J. 1989. Fauna Palaestina Insecta IV. Diptera: Tephritidae. Israel at Keterpress Enterprises, Jerusalem, 212 pp. Freidberg, A. & Mathis, W. N. 1986. Studies of Terelliinae (Diptera: Tephritidae) A Revision of the Genus Neaspilota Osten Sacken. Smithsonian Contributions to Zoology, No. 439, 75 p. Giray, H. 1979. TUrkiye Trypetidae (Diptera) faunasma ait ilk liste. Tiirkiye Bitki Koruma Dergisi, 3 (1): 35-46. Harris, P. 1984a. Current approaches to biological control of weeds. In: Kelleher, J.S. & Hulme, M.A. (eds), Biological Control Programmes against insects and weeds in Canada 1969-1980, pp. 95-104.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 361
Harris. P. 1984b. Carduus nutans L., nodding thistle and C. acanthoides L., plumeless thistle (Compositae). In: Kelleher, J.S. & Hulme, M.A. (eds), Biological Control Programmes against insects and weeds in Canada 1969—1980, pp. 95-104. Harris, P. & Wilkinson, A. T. S. 1984. Cirsium vulgare (Savi) Ten., bull thistle (Compositae). In: Kelleher, J. S. & Hulme, M. A. (eds), Biological Control Programmes against insects and weeds in Canada 1969-1980, pp. 147-154. Korneyev, V. A. & White, I. M. 1993. Fruit flies of the genus Urophora R.-D. (Diptera, Tephritidae) of east Palaearctic. II. Review of species of the subgenus Urophora s. str. Communication 2. – Entomologicheskoe obozenic, 72: 232-247. Korneyev, V. A. & White, I. M. 1999. Fruit flies of genus Urophora R-D. (Diptera: Tephritidae) of east palaearctic. lII.key to species. Ent, Obozr., 78 (2): 464-482. Korneyev, V. A. & White, I. M. 2000. Fruit-flies of the genus Urophora R.-D (Diptera, Tephritidae) of East Palaearctic. IV. Conclusion. Ent. obozr. 79: 239-253. [In Russian; English summary; English translation Entomol. Rev., Wash., 79: 296-309. Kutuk, M. 2003. Doğu Akdeniz Bolgesi Urophora Robineau-Desvoidy (Diptera: Tephritidae) faunası ve sistemati.i uzerine arafltırmalar. Turkiye Entomoloji Dergisi, 27: 149-160. Kutuk, M. & Ozgur, A. F. 2003. Faunistical and systematical studies on the genus Tephritis Latreille, 1804 (Diptera: Tephritidae) in the South West of Turkey along with new records. Turk. Entomol. Derg., 27: 243-252. Merz, B. 1994. Insecte Helvetica Fauna 10. Herausgegeben von der Schweizeischen Entomologischen Gesellschaft. Diptera Tephritidae, Hge. Geneve, 198 pp. Merz, B. 2001. Faunistics of the Tephritidae (Diptera) of the Iberian Peninsula and the Baleares. Bulletin de la Societe Entomologique Suisse, 74: 91-98. Merz, B. & Korneyev, V. A. 2004. Fauna Europea: Tephritidae In Pape, T. (ed.) (2004) Fauna Europea: Diptera, Brachycera. Fauna Europea version 1.1, http://www.faunaeur.org. Norrbom, A. L., Carroll, L. E., Thompson, F. C., White, I. M. & Freidberg, A. 1999. Norrbom Systematic Database of Names // Fruit Fly Expert Identification System and Systematic Information Database / Ed. F. C. Thompson. Myia, 1999. p. 65–299. Richter, V. A. 1970. [Family Tephritidae (Trypetidae)], In: Key to the insects of the European part of the USSR. Zool. Ins. Acad. Nauk. SSSR, Leningrad, p. 132-172. Thompson, F. C. 1998. Fruit Fly Expert Identification System and Systematic Information Database. North American Dipterists Society, Backhuys Publishers, Leiden, the Netherlands. White, I. M. 1988. Tephritid flies: Handbooks for the identification of British insects. Vol. 10, part 5a. London: Royal Entomological Society of London. 134 pp. White, I. M. & Clement, S. L. 1987. Systematic notes on Urophora (Diptera, Tephritidae) species associated with Centaurea solstitialis (Asteraceae, Cardueae) and other Palaearctic weeds adventive in North America. Proceedings of the Entomological Society of Washington, 89: 571-580. White, I. M., Headrick, D. H., Norrbom, A. L. & Carroll, L. E. 1999. Glossary. p. 881–924. In: Aluja, M. & Norrbom, A. L. (Eds.), Fruit flies (Tephritidae): Phylogeny and evolution of behavior. CRC Press, Boca Raton. [16] + 944 p. White, I. M. & Komeyev, V. A. 1989. A revisionof the western paIaearctic species of Urophora Robineau-Desvoidy (Diptera: Tephritidae). Systematic Entomology, 14: 327-374.
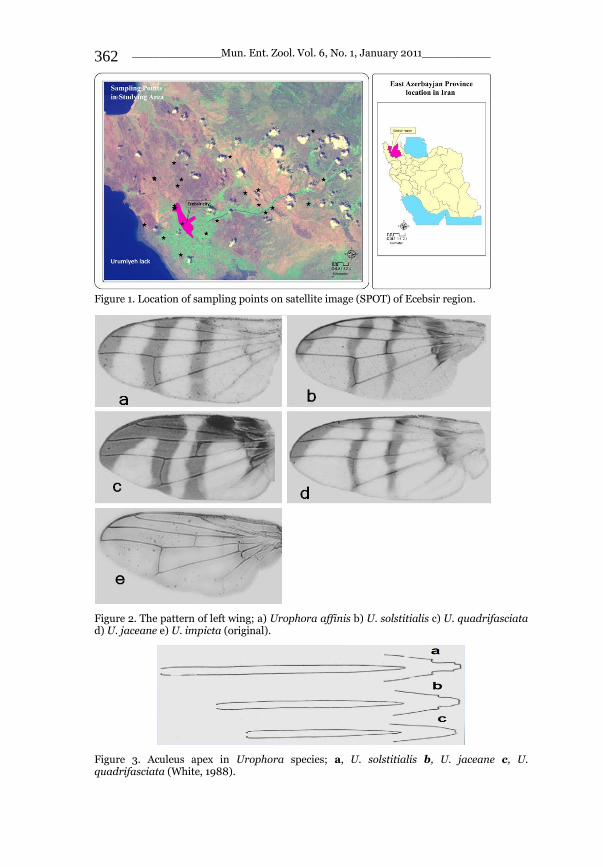
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 362
Figure 1. Location of sampling points on satellite image (SPOT) of Ecebsir region.
Figure 2. The pattern of left wing; a) Urophora affinis b) U. solstitialis c) U. quadrifasciata d) U. jaceane e) U. impicta (original).
Figure 3. Aculeus apex in Urophora species; a, U. solstitialis b, U. jaceane c, U. quadrifasciata (White, 1988).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 363
MOLECULAR FINGERPRINTING OF CERTAIN CEREAL APHIDS IN EGYPT (HEMIPTERA: STERNORRHYNCHA:
APHIDIDAE) USING RAPD AND ISSR MARKERS
A. Helmi*, A. F. Khafaga** and M. M. El-Fatih***
* Plant Protection Department, Fac. of Agric., Ain Shams Univ., PB. 68 hadak Shobra El Kheima, Cairo, EGYPT. E-mail: [email protected] ** National Center for Radiation Research and Technology, Atomic Energy Authority, Nasr City, Cairo, EGYPT. *** Plant Protection Research Institute, Agriculture Research Center, Dokki, Giza, EGYPT. [Helmi, A., Khafaga, A. F. & El-Fatih, M. M. 2011. Molecular Fingerprinting of certain cereal aphids in Egypt (Hemiptera: Sternorrhyncha: Aphididae) using RAPD and ISSR markers. Munis Entomology & Zoology, 6 (1): 363-376] ABSTRACT: Cereal aphids are one of the most important insect pests limiting cereal production worldwide. Classical morphological criteria for aphid species identification may be affected by environmental factors such as climatic conditions and physiological status of the host plant. So, two modern molecular techniques; Random Amplified Polymorphic DNA (RAPD) and Inter Simple Sequence Repeats (ISSRs) were used to find diagnostic markers for fingerprinting eleven cereal aphids those collected from different cereal plants and from different localities in Egypt. Eight RAPD and five ISSRs primers were successively proven 97 and 69 markers that could be used to differentiate the eleven different cereal aphid species. Also these molecular techniques with 26 diagnostic morphological characters were used to find the Phylogenetic relationship among the different collected species; that divided into two clusters with similarity matrix percentages of 73% and 82%. From these results it could concluded that these techniques could be used successively to fingerprint and identify these aphid species and differentiate among them. KEY WORDS: Cereal aphids, Fingerprinting, RAPD, ISSRs, Identification, Markers, Microsatellites, Phylogeny.
Aphids are one of the most wide spread groups of pests in agricultural systems and may cause crop losses in forest trees, field crops and horticultural crops. Plants in these production systems may be affected directly or indirectly as a result of the presence of aphid populations (Minks & Harrewijn, 1989). There are approximately 5,000 described species of aphids (Remaudiere & Remaudiere, 1997). Cereal aphids are one of the most important insect pests limiting cereal production worldwide (Vickerman & Wratten, 1979; Dixon, 1987; D‘Arcy & Mayo, 1997). Proper identification of agronomical important species of aphids is necessary in order to establish properly their diversity and population dynamics in a crop. Classical morphological criteria for aphid species identification suffer from at least two drawbacks: they depend on adult individuals since in many cases nymphal instars do not lend themselves to an accurate determination, and they may be affected by environmental factors such as climatic conditions and physiological status of the host plant (Cenis et al., 1993; Figueroa et al., 1999). Thus, identification based on morphological traits has been problematic for several closely related species (Loxdale & Brookes, 1989). Random Amplified Polymorphic DNA (RAPD) has proven to be invaluable source of markers for many sap-sucking insect pests identification such as aphid species (Cenis et al., 1993; Lushai et al., 1997; Figueroa et al., 2005 and Shahadi-Fatima et al., 2007). Another genetic technique is Simple Sequence Repeats (ISSRs or ‗microsatellites‘) which has been used by plant biologists for a variety of applications (Wolfe & Liston, 1998) and rarely used in animals (Kostia et al.,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 364
2000; Reddy et al., 1999). This technique used for population-level studies in two species of cyclically parthenogenetic aphids, Acyrthosiphon pisum and Pemphigus obesinymphae (Abbot, 2001). Also used to identify different biotypes of greenbug, Schizaphis graminum (Weng et al., 2007). The ISSRs technique is used here for the first time to differentiate among different separate insect species.
This study aimed to differentiate among eleven cereal aphid species found in Egypt, and also to establish molecular genetic fingerprint for these species using RAPD and ISSRs polymorphism and elucidate relationships among these species.
MATERIALS AND METHODS Survey of certain cereal aphid species:
Field survey of aphid species was carried out during the period 2008-2009 in different localities of Egypt. Eleven cereal aphid species were collected from different localities allover the years. The species are listed alphabetically by scientific name in Table (1).
Samples from the infested plants were transferred to the laboratory and alate individuals of aphid were mounted on slides for identification by using available keys.
Identification of these species was conducted according to the key of Fathi and El-Fatih (2009). The eleven species were reared on their host separately, where one apterous adult was transferred by a brush to a healthy plant. Nymphs were leaved to feed and developed to adults. This method was repeated for two off-springs. The adults of the second offspring of each species were put in tubes and preserved at -20 ºC until use in the biochemical and molecular studies. RAPD-PCR Analysis a. DNA Extraction
DNA were extracted from eleven different species of adult Aphid.Animal tissues were ground under liquid nitrogen to a fine powder, then bulked DNA extraction was performed using DNeasy plant Mini Kit (QIAGEN). b. Polymerase Chain Reaction (PCR)
PCR amplification was performed using eight random 10 mer arbitrary primers synthesized by (Operon biotechnologies, Inc.Germany).
Amplification was conducted in 25 µl reaction volume containing the following reagents: 2.5 µl of dNTPs (2.5 mM), 2.5 µl MgCl2 (2.5 mM), and 2.5 µl of 10 x buffer, 3.0 µl of primer (10 pmol), 3.0 µl of template DNA (25 ng / µl), 1 µl of Taq polymerase (1U/ µl) and 10.5 µl of sterile dd H2O. The DNA amplifications were performed in an automated thermal cycle (model Techno 512) programmed for one cycle at 94º C for 4 min followed by 45 cycles of 1 min at 94º C, 1 min at 36º C, and 2 min at 72º C. the reaction was finally stored at 72º C for 10 min. Amplified products were size-fractioned using ladder marker (100bp +1.5 Kbp) by electrophoresis in 1.5 % agarose gels in TBE buffer at 120 V for 1 h. the bands were visualized by ethidium bromide under UV florescence and photographed. ISSR-PCR Analysis a. DNA Extraction
Eleven different species of adult Aphids samples were collected and extracted DNA from them. Animal tissues were ground under liquid nitrogen to a fine powder, and then bulked DNA extraction was performed using DNeasy plant Mini Kit (QIAGEN).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 365
b. Polymerase Chain Reaction (PCR) PCR amplification was performed using five Inter Simple Sequence Repeat
(ISSR) Table, 3. Amplification was conducted in 25 µl reaction volume containing the
following reagents: 2.5 µl of dNTPs (2.5 mM), 2.5 µl Mgcl2 (2.5 mM), and 2.5 µl of 10 x buffer, 3.0 µl of primer (10 pmol), 3.0 µl of template DNA (25 ng / µl), 1 µl of Taq polymerase (1U/ µl) and 10.5 µl of sterile dd H2O. The DNA amplifications were performed in an automated thermal cycle (model Techno 512) The PCRs were programmed for one cycle at 94º C for 4 min followed by 45 cycles of 1 min at 94 ºC, 1 min at 57 ºC, and 2 min at 72 ºC. The reaction was finally stored at 72 ºC for 10 min. Amplified products were size-fractioned using ladder marker100bp (1000, 900, 800, 700, 60, 500, 400, 300, 200 and 100bp) by electrophoresis in 1.5 % agarose gels in TBE buffer at 120 V for 30 min. the bands were visualized by ethidium bromide under UV florescence and photographed. Densitometry Scanning and Analysis:
All gels resulted from DNA fingerprints, were scanned using Bio-Rad GelDoc2000 to calculate the pair-wise differences matrix and plot the dendrogram among different aphid species. Phylogenetic relationship among the eleven species:
To study the Phylogenetic relationship among different aphid species based on three different criteria; eight RAPD primers, five ISSR primers and twenty-six diagnostic morphological characters were used. In this method all employed characters were regarded as being of equal importance. The morphological characters were selected to be of most taxonomic importance according to Fathi and El Fatih 2009. The analysis data of the three criteria were taken as basis for assessing the relationships between each species and the others. Statistical soft ware program (SPSS program) was used to calculate the similarity matrix. The degree of similarity was shown as numbers between 0 and 1 for the most dissimilar and the most similar pair of species, respectively. The performing and drawing of the phylogenetic tree was done by Statistical Package for Social Science (SPSS ver. 10) computer program.
RESULTS Random Amplified Polymorphic DNA (RAPD):
Eight RAPD primers were tested against the eleven cereal aphid species to find markers for identification of these species. These primers generated 167 fragments, 97 bands of them were considered as markers for different species (58.1%), the highest number of markers was 20 bands generated by OPC-01 and OPC-13, while the lowest number of markers was 4 bands generated by OPD-07. the highest number of markers was 12 bands detected for S. graminum, while the lowest number of markers was 5 bands detected for M. dirhodum (Table, 5). Thirty-seven common bands (22.2%) were detected among the eleven species found by the eight tested primers. OPC-01 primer:
This primer generated 23 bands with molecular weight ranged from 903 to 111bp. and the average number of bands generated in different species ranged from 8 bands in S. avenae and 4 bands in M. dirhodum. This primer showed 87.1% polymorphism whereas three common bands among the eleven species were detected (784, 378 and 250bp.). Twenty marker bands were detected for the eleven species; the highest number of markers was three bands for S. graminum

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 366
and S avenae, while the lowest number was one band in R. maidis, S. minuta, M. dirhodum and H. pruni. While the other five species, each of them has two marker bands. OPC-13 primer:
This primer generated 30 bands with molecular weight ranged from 1360 to 105bp. and the average number of bands generated in different species ranged from 11 bands in H. pruni and 4 bands R. maidis. This primer showed 90% polymorphism whereas three common bands were detected among the eleven species with molecular weights 764, 431 and 355bp. Twenty marker bands were detected for the eleven species, the highest number of markers was five bands for H. pruni, while the lowest number was one band for six species; A. corni, T. africana, R. maidis, S. graminum, S. avenae and S. scirpus. Three marker bands were detected for two species; S. rotundiventrus and S. minuta, two marker bands for R. padi and M. dirhodum. OPD-07 primer:
This primer generated 16 bands with molecular weight ranged from 1231 to 223bp. and the average number of bands generated in different species ranged from 10 bands in six species; R. maidis, S. rotundiventrus, S. avenae, M. dirhodum, H. pruni and S. scirpus to 5 bands in S. graminum and S. minuta. This primer showed 68.75% polymorphism whereas five common bands (1231, 750, 520, 392 and 328bp.) among the eleven species were detected. Only four marker bands; 886, 639, 795 and 935bp. were detected for A. corni, R. maidis, R. padi and M. dirhodum, respectively. OPE-03 primer:
This primer generated 24 bands with molecular weight ranged from 1186 to 208bp. and the average number of bands generated in different species ranged from 12 bands in A. corni to 5 bands in S. rotundiventrus. This primer showed 83.3% polymorphism whereas four common bands (737, 387, 356 and 208bp) among the eleven species were detected. Thirteen marker bands were detected for eight species, three marker bands for S. scirpus; two marker bands for three species; A. corni, T. africana and S. avenae. While only one marker band was detected for four species; R. maidis, R. padi, S. minuta and H. pruni. OPE-06 primer:
This primer generated 19 bands with molecular weight ranged from 1132 to 190bp. and the average number of bands generated in different species ranged from 8 bands in A. corni to 4 bands in S. graminum. This primer showed 79 % polymorphism whereas four common bands (421, 332, 222 and 190bp) among the eleven species were detected. Ten marker bands were detected for seven species, two marker bands for three species; A. corni, S. minuta and S. scirpus. While one marker band (monomorphic) was detected for four species; R. padi, S. rotundiventrus, S. avenae and H. pruni. OPI-17 primer:
This primer generated 21 bands with molecular weight ranged from 1343 to 162bp. and the average number of bands generated in different species ranged from 10 bands in T. africana and S. graminum to 7 bands in two species; A. corni and R. padi. This primer showed 66.7% polymorphism whereas seven common bands (988, 841, 656, 557, 486, 419 and 343bp.) among the eleven species were detected. fourteen marker bands (monomorphic) were detected for nine species, the highest number of markers was five bands detected for S. graminum, while the lowest number was one band was detected for seven; A. corni, R. maidis, S. minuta, S. rotundiventrus, S. avenae, M. dirhodum and H. pruni. While two

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 367
marker bands were detected for T. africana. No marker bands for two species; R. padi and S. scirpus. OPL-20 primer:
This prime generated 15 different fragments with molecular weight ranged from 1240 to 202bp, the average number of bands generated in different species ranged from 9 bands in R. maidis to 7 bands in S. rotundiventrus, H. pruni and S. scirpus; while the other eight species have 8 bands. This primer showed 60% polymorphism whereas, six common bands were detected by this primer with molecular weights 1240, 731, 662, 507, 400 and 202bp. Six marker bands in six different species were detected (one marker band for each species); A. corni, T. africana, R. maidis, R. padi, S. rotundiventrus and S. avenae. No marker bands for the other six species. OPQ-15 primer:
This primer generated 19 bands with molecular weight ranged from 1162 to 202bp. and the average number of bands generated in different species ranged from 8 bands in three species; T. african, R. maidis, R. padi, to 5 bands in M. dirhodum. This primer showed 73.7% polymorphism whereas five common bands among the eleven species were detected (683, 410, 341, 238 and 202bp.). Ten marker bands (monomorphic) were detected for nine species; the highest number of markers was two bands detected for S. graminum. While just one marker band was detected for eight species; A. corni, T. africana, R. maidis, R. padi, S. rotundiventrus, S. minuta, H. pruni and S. scirpus. While no marker bands were detected for S. avenae and M. dirhodium. Inter simple sequence repeats (ISSRs):
Five ISSRs primers were tested against the eleven cereal aphid species to find markers for identification of these species. These primers generated 122 fragments, 69 bands of these fragments were considered as markers for different species (56.6%), the highest number of markers was 15 bands generated by two primers; HP-09 and HP-13, while the lowest number of markers was 12 bands generated by HP-14 primer. A. corni had the highest number of marker bands (10 bands), while the lowest number of markers was 3 bands detected for S. scirpus (Table, 6). Twenty-one common bands (17.2%) were detected among the eleven species by these five tested primers. HP-09 Primer:
This primer generated 23 bands with molecular weight ranged from 1109 to 117bp. and the average number of bands generated in different species ranged from 8 bands in A. corni to 5 bands in R. padi and S. avenae. This primer showed 82.6% polymorphism whereas 4 common bands (422, 349, 254 and 172bp) were detected among the eleven species. Fifteen marker bands (monomorphic) were detected for ten species; the highest number of markers was three bands were detected for A. corni, while the lowest number was one marker band was detected for six species; R. padi, S. graminum, S. minuta, S. avenae, H. pruni and S. scirpus. While no marker bands were detected for M. dirhodium. HP-11 Primer:
This primer generated 30 bands with molecular weight ranged from 1301 to 124bp. This primer showed 73.3 % polymorphism, where eight common bands (930, 780, 685, 516, 436, 288, 242 and 174bp) were detected among the eleven species. The average number of bands generated in different species ranged from 15 bands in S. rotundiventrus to 10 bands in S. scirpus. Fourteen marker bands were detected for the eleven species; the highest number of markers was two bands were detected for three species; A. corni, S. graminum and S.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 368
rotundiventrus, while the lowest number was one marker band was detected for the other eight species. HP-12 Primer:
This primer generated 22 bands with molecular weight ranged from 842 to 95bp. This primer showed 90.9% polymorphism, where two common bands (323 and 230bp) were detected among the eleven species. The average number of bands generated in different species ranged from 9 bands in S. avenae to 2 bands in T. africana. Thirteen marker bands were detected for eight species; the highest number of markers was three bands were detected for S. avenae, while the lowest number was one marker band was detected for four species; A. corni, R. maidis, R. padi and S. rotundiventrus. Two marker bands were detected for three species; S. graminum, M. dirodium and H. pruni. While no marker bands were detected in T. africana, S. minuta and S. scirpus. HP-13 Primer:
This primer generated 25 bands with molecular weight ranged from 1016 to 123bp. This primer showed 88% polymorphism, where three common bands (640, 398 and 174bp) were detected among the eleven species. The average number of bands generated in different species ranged from 10 bands in S. avenae to 5 bands in H. pruni. Fifteen marker bands were detected for the eleven species; the highest number of markers was two bands were detected for four species; A. corni, T. africana, R. maidis and S. graminum. While the lowest number was one marker band was detected for the other seven species. HP-14 Primer:
This primer generated 22 bands with molecular weight ranged from 963 to 32bp. This primer showed 81.8% polymorphism, where four common bands (261, 192, 153 and 101bp) were detected among the eleven species. The average number of bands generated in different species ranged from 8 bands in S. avenae, H. pruni and S. scirpus to 5 bands in S. minuta and M. dirhodium. Twelve marker bands were detected for nine species; the highest number of markers was two bands were detected for three species; A. corni, S. rotundiventrus and S. avenae. While the lowest number was one marker band was detected for six species; T. africana, R. maidis, R. padi, S. graminum, S. minuta and M. dirhodium. While no marker bands were detected for two species; H. pruni and S. scirpus. Phylogenetic relationship among the eleven species:
Genetic similarities and genetic relatedness amongst the eleven cereal aphid species were based on data obtained of three different criteria; eight RAPD primers and five ISSRs primers as molecular markers and as well as twenty-six diagnostic morphological characters. These data were subjected to using SPSS computer program to support the existence of high level of genetic relatedness amongst the eleven cereal aphid species. The relatedness dendrogram was indicated two main clusters. The first cluster included ten aphid species while, the second included A. corni only with similarity matrix percentage of 73%. The first cluster was divided into two sub-clusters with similarity matrix percentage of 82%; the first one included six aphid species with similarity matrix percentage of 88%; H. pruni, S. scirpus, S. avenae, M. dirhodum, S. rotundiventrus and S. minuta. While the second sub-cluster contained four aphid species with similarity matrix percentage of 84%; S. graminum, R. maidis, R. padi and T. africana. The highest similarity matrix percentage in this phylogenetic relationship was 98% between H. pruni and S. scirpus, while the lowest similarity matrix percentage found between A. corni and H. pruni (73%) (Fig. 3). These results indicated the success of these techniques with morphological characters to draw the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 369
Phylogenetic relationship of these aphids' species whereas the species those belonging to the same genus are closest to each other.
DISCUSSION
Some of the insect species are easy to identify and categorize, while for others, such as aphids species are difficult because of their small size and morphological similarity. Moreover, it is further difficult to identify morphological variation due to environmental factors by available traditional methods (Loxdale and Brookes, 1989; Cenis et al., 1993; Figueroa et al., 1999). To overcome these problems, the advanced molecular techniques, viz., randomly amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) and simple sequence repeats (ISSRs) have been useful tools in assessing insect genetic diversity (Black et al., 1992; Cenis, 2003; Gobbi et al., 2003; Sartor et al., 2008; Sharma et al., 2008; Perumal et al., 2009; Qiu et al., 2009). RAPD markers have become the most common yardsticks for measuring genetic differences between individuals, within and between related species or population (Jain et al. 2010). This technique also used to distinguish different geographical and/or host associated populations of some cryptic complex species (Zitoudi et al., 2001; Bulman et al., 2005; Helmi, 2011). In Egypt RAPD-PCR technique is successively used to fingerprint of some sap-sucking insect species belonging into the same taxonomic category such as fingerprinting of ten aphids species belonging to Genus Aphis (Shahadi-Fatima et al., 2007) and also used to fingerprint fifteen whitefly species (Amin et al., 2009).
Inter simple sequence repeats (ISSRs) is a valuable addition to the inventory of PCR-based methods for rapid, large-scale screening of genetic variations. The vagaries of PCR and the chosen method of band detection limit any PCR-based marker (Wolfe & Liston, 1998). But ISSRs markers are typically highly reproducible, due to stringent annealing temperatures, long primers, and low primer-template mismatch (that is, the primers are not ‗arbitrary‘, but designed a priori to anchor onto anonymous SSR loci; Wolfe et al., 1998) and ISSRs can reveal polymorphisms without more elaborate detection protocols (Esselman et al., 1999).
ISSRs method has shown much promise for the study of the population biology of plants (Clausing et al., 2000; Hess et al., 2000), but rarely used in animals (Reddy et al., 1999; Kostia et al., 2000 and Abbot, 2001). There are two available known studies in use of ISSRs in aphids; one of them was on population-level studies in two species of cyclically parthenogenetic aphids; Acyrthosiphon pisum and Pemphigus obesinymphae (Abbot et al., 2001) who reported that IISSRs are suitable for invertebrate populations those have small size bodies and low levels of within-population variation. While the other study was on host-associated genetic differences and regional differences among the green bug, Schizaphis graminum biotypes (Weng et al., 2007) who cited that the use of ISSRs would be useful for aphid genetic, ecological, and evolutionary studies and can potentially shorten the time and cost for biotype identification.
CONCLUSION
Molecular fingerprinting of eleven cereal aphid species collected from Egypt were carried out using two modern genetic techniques; RAPD-PCR and ISSRs. These techniques successively generated many molecular markers for different studied species, those could be used to identify these aphid species and differentiate among them. Also these two techniques in addition to some

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 370
diagnostic morphological characters were used to determine the Phylogenetic relationship among the eleven aphid species.
LITERATURE CITED Abbot, P. 2001. Individual and population variation in invertebrates revealed by Inter-simple Sequence Repeats (ISSRs). 3 pp. Journal of Insect Science , 1.8. Available online: insectscience.org/1.8 Abbot, P., Withgott, J. H. & Moran, N. A. 2001. Genetic conflict and conditional altruism in social aphid colonies. Proc. Nat. Acad. Sci. USA. 9; 98 (21): 11839-41. Amin, A. H., Helmi, A. & Emam, K. A. 2009. Genetic fingerprinting of fifteen whitefly species in Egypt. (Hemiptera: Aleyrodidae). J. Biol. Chem. Environ. Sci., 4 (4): 473-490. Black, W. C., DuTeau, N. M. & Puterka, G. J. 1992. Use of the random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) to detect DNA polymorphisms in aphids (Homoptera: Aphididae). Bull. Entomol. Res., 82: 51-159. Bulman, S. R., Stufkens, M. A. W., Eastop, V. F. & Teulon, D. A. J. 2005. Rhopalosiphum aphids in New Zealand. II. DNA sequences reveal two incompletely described species. New Zealand Journal of Zoology, 32 (1): 37-45. Cenis, J. L. 2003. Markers for the identification of biotypes of Bemisia tabaci: the Mediterranean basin as a case study. 3rd International Bemisia Workshop Barcelona 17-20 March, 2003. Cenis, J. L., Perez, P. & Fereres, A. 1993. Identification of Aphid (Homoptera: Aphididae) species and clones by Random Amplified Polymorphic DNA. Ent. Soc. Amer., 86: 545-550. Clausing, G., Vickers, K. & Kadereit, J. W. 2000. Historical biogeography in a linear system: genetic variation of Sea Rocket (Cakilemaritima) and Sea Holly (Eryngium maritimum) along European coasts. Mol. Ecol., 9: 1823-1833. D’Arcy, C. J. & Mayo, M. 1997. Proposals for changes in luteovirus taxonomy and nomenclature. Arch. Virol., 142: 1285-1287. Dixon, A. F. G. 1987. Cereal aphids as an applied problem. Agr. Zool. Rev., 2: 1-57. Esselman, E. J., Jianqiang, L., Crawford, D. J., Windus, G. & Wolfe, A. D. 1999.Clonal diversity in the rare Calamagrostis porteri ssp. insperata (Poaceae): comparative results for allozymes and random amplified polymorphic DNA (RAPD) and intersimple sequence repeat (ISSR) markers. Mol. Ecol., 8: 443-451. Fathi, A. H. & El Fatih, M. M. 2009. Identification of apterous viviparous of cereal aphids in Egypt (Hemiptera: Sternorrhyncha: Aphidoidea). Bull. Ent. Soc. Egypt, 86: 307-325. Figueroa, C. C., Simon, J. C., Gallic, J. F. & Niemeyer, H. M. 1999. Molecular markers to differentiate two morphologically-close species of the genus Sitobion. Ent. exp. & appl., 92: 217-225. Gobbi, A., Pascual, S., Avilés, M., Beitia, F., Hernández-Suárez, E. & Carnero, A. 2003. RAPD-PCR characterization of Bemisia tabaci (Gennadius) populations in the Canary Islands. 3rd International Bemisia Workshop, Barcelona: 17-20 March, 2003. Goldstein, D. B. & Schlotterer, C. 1999. Microsatellites: Evolution and applications, Oxford: Oxford University Press. Helmi, A. 2011. Host-associated population variations of Bemisia tabaci (Genn.) (Hemiptera: Sternorrhyncha: Aleyrodidae) characterized with random DNA markers. Int. J. Zool. Res., 7: 77-84. Hess, J., Kadereit, J. W. & Vargas, P. 2000. The colonization history of Olea europaea L. in Macaronesia based on internal transcribed spacer 1 (ITS-1) sequences, randomly amplified polymorphic DNAs (RAPD), and intersimple sequence repeats (ISSR). Mol. Ecol., 9: 857-868. Jain, S. K., Neekhra, B., Pandey, D. & Jain, K. 2010. RAPD marker system in insect study: A review, Indian Journal of Biotechnology, 9: 7-12.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 371
Kostia, S., Ruohonen-Lehto, M., Vainola, R. & Varvio, S. L. 2000. Phylogenetic information in inter-SINE and inter-SSR fingerprints of the Artiodactyla and evolution of the BovtA SINE. Heredity, 84: 37-45. Lopes-da-Silva, M. & Vieira, L. G. E. 2007. Analysis of the genetic diversity in Metopolophium dirhodum (Walker) (Hemiptera, Aphididae) by RAPD markers. Revista Brasileira de Entomologia, 51 (1): 54-57. Loxdale, H. D. & Brookes, C. P. 1989. Separation of three Rubus-feeding species of aphid Sitobion fragariae (Wlk.), Macrosiphum funestum (Macch.) and Amphorophora rubi (Kalt.) (Hemiptera: Aphididae) by electrophoresis. Ann. Appl. Biol., 115: 399–404. Lozier, J. D., Foottit, R. G., Miller, G. L., Mills, N. J. & Roderick, G. K. 2008. Molecular and morphological evaluation of the aphid genus Hyalopterus Koch (Insecta: Hemiptera: Aphididae), with a description of a new species. Zootaxa, 1688: 1-19. McGregor, C. E., Lambert, C. A., Greyling, M. M., Louw, J. H. & Warnich, L. 2000. A comparative assessment of DNA fingerprinting techniques (RAPD, ISSR, AFLP and SSR) in tetraploid potato (Solanum tuberosum L.) germplasm. Euphytica, 113: 135-144. Minks, A. K. & Harrewijn, P. 1989. World crop pests, 2C, Aphids their biology, Natural enemies and Control. Volume C. Elsevier. Netherland. 312 pp. Parsons, B. J., Newbury, H. J., Jackson, M. T. & Ford-Lloyd, B. V. 1997. Contrasting genetic diversity relationships are revealed in rice (Oryza sativa L.) using different marker types. Molecular Breeding, 3: 115-125. Perumal, Y., Marimuthu, M., Salim, A. P. & Ponnusamy, B. 2009. Host Plant Mediated Population Variations of Cotton Whitefly Bemisia tabaci (Gennadius) (Aleyrodidae: Homoptera) Characterized with Random DNA Markers. Am. J. Bioch. and Biotech., 5: 40-46. Provan, J., Powell, W. & Waugh, R. 1996. Analysis of cultivated potato (Solanum tuberosum) Molecular Systematics of Plants II: DNA Sequencing, pp. 43-86. New York: Kluwer. Qiu, B., Chen, Y., Peng, L. L. W., Li, X., Ahmed, M. Z., Mathur, V., Duandand, Y. & Ren, S. 2009. Identification of three major Bemisia tabaci biotypes in China based on morphological and DNA polymorphisms. Progress in Natural Science. 19: 713-718. Reddy, K. D., Nagaraju, J. & Abraham, E. G. 1999. Genetic characterization of the silkworm Bombyx mori by simple sequence repeat (SSR)-anchored PCR. Heredity, 83: 681-687. Remaudière, G. & Remaudière, M. 1997. Catalogue des Aphididae du monde. Institut National de la Recherche Agronomique, Paris. 473 p. Sandstrom, J. & Moran, N. 1999. How nutritionally imbalanced is phloem sap for aphids? Entomologia Experimentalis et Applicata, 91: 203-210. Sartor, C., Demichelis, S., Cenis, J. L., Coulibaly, A. K. & Bosco, D. 2008. Genetic variability of Bemisia tabaci in the Mediterranean and Sahel Regions. Bull. Insectology, 61: 161-162. Shahadi-Fatima, H., Amin, A. H. & Soliman, Kh. A. 2007. Genetic fingerprints and phylogenetic relationships of ten Aphis species found in Egypt (Hemiptera: Aphididae). J. Biol. Chem. Environ. Sci., 2 (4): 173-188. Sharma, R. K., Gupta, V. K., Jindal, J. & Dilawari, V. K. 2008. Host associated variations in whitefly, Bemisia tabaci (Genn.). Indian J. Biotech., 7: 366-370. Vickerman, G. P. & Wratten, S. D. 1979. The biology and pest status of cereal aphids (Hemiptera: Aphididae) in Europe: a review. Bull. Entomol. Res., 69: 1-32. Weng, Y., Azhaguvel, P., Michels, G. J. & Rudd, J. C. 2007. Cross-species transferability of microsatellite markers from six aphid (Hemiptera: Aphididae) species and their use for evaluating biotypic diversity in two cereal aphids. Insect Mol. Biol., 16 (5): 613 622. Wolfe, A. D. & Liston, A. 1998. Contributions of PCR-based methods to plant systematics and evolutionary biology. In: Soltis DE, Soltis PS, Doyle JJ, editors. Wolfe AD, Xiang Q-Y, Kephart SR. 1998.
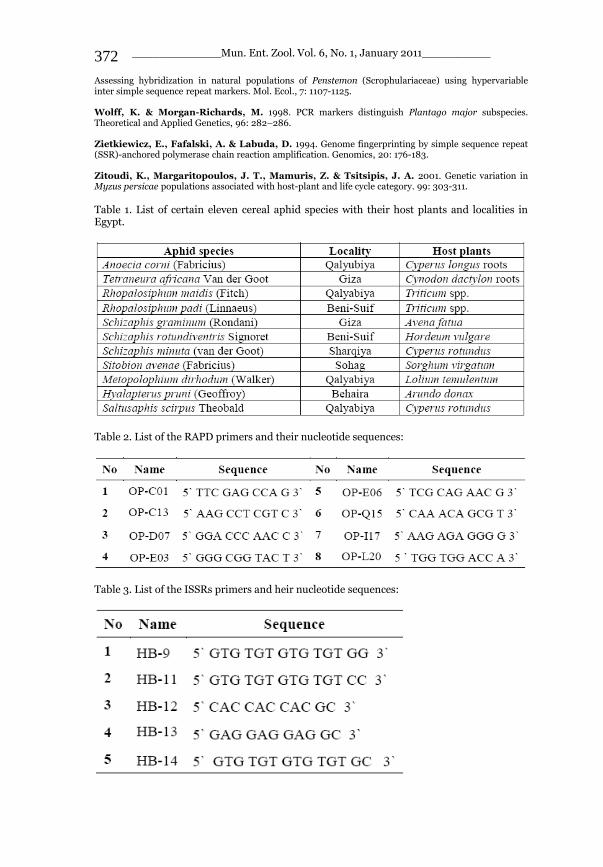
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 372
Assessing hybridization in natural populations of Penstemon (Scrophulariaceae) using hypervariable inter simple sequence repeat markers. Mol. Ecol., 7: 1107-1125. Wolff, K. & Morgan-Richards, M. 1998. PCR markers distinguish Plantago major subspecies. Theoretical and Applied Genetics, 96: 282–286. Zietkiewicz, E., Fafalski, A. & Labuda, D. 1994. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20: 176-183. Zitoudi, K., Margaritopoulos, J. T., Mamuris, Z. & Tsitsipis, J. A. 2001. Genetic variation in Myzus persicae populations associated with host-plant and life cycle category. 99: 303-311.
Table 1. List of certain eleven cereal aphid species with their host plants and localities in Egypt.
Table 2. List of the RAPD primers and their nucleotide sequences:
Table 3. List of the ISSRs primers and heir nucleotide sequences:
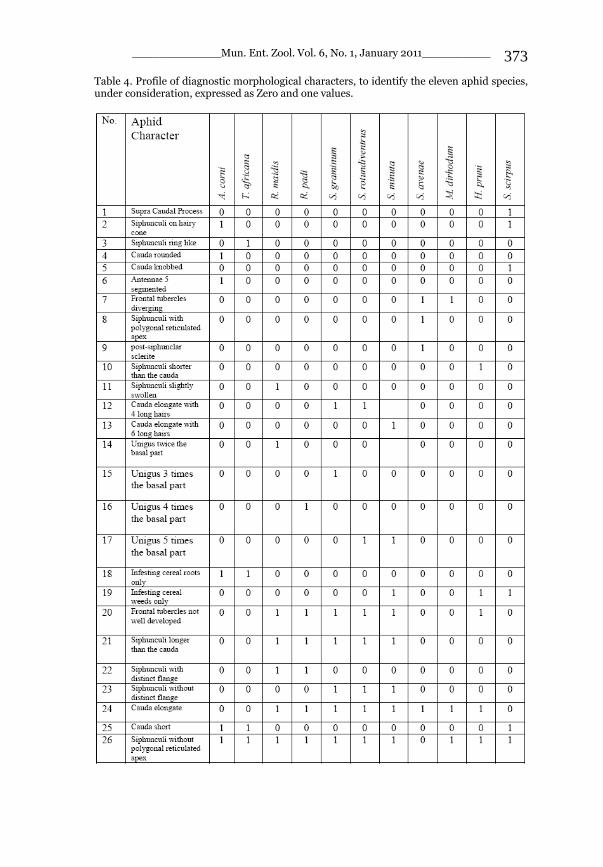
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 373
Table 4. Profile of diagnostic morphological characters, to identify the eleven aphid species, under consideration, expressed as Zero and one values.
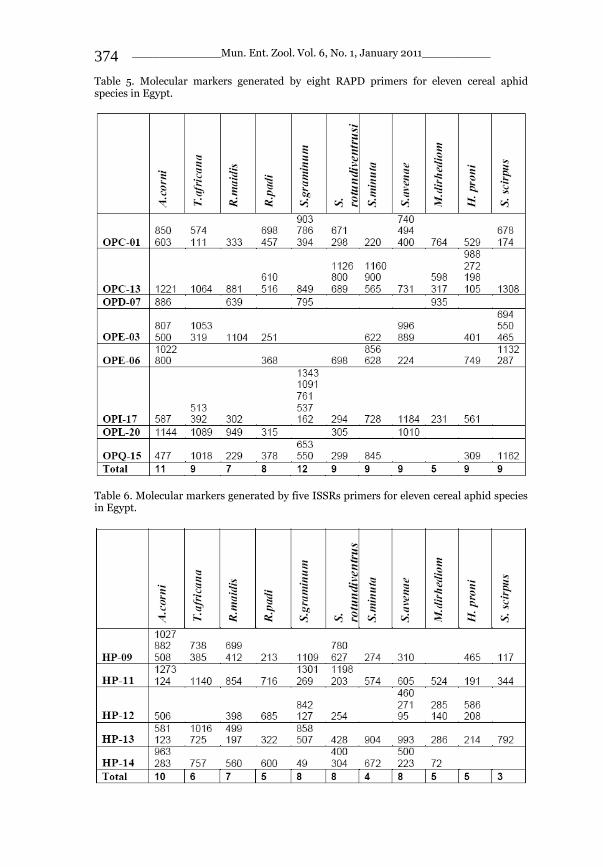
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 374
Table 5. Molecular markers generated by eight RAPD primers for eleven cereal aphid species in Egypt.
Table 6. Molecular markers generated by five ISSRs primers for eleven cereal aphid species in Egypt.
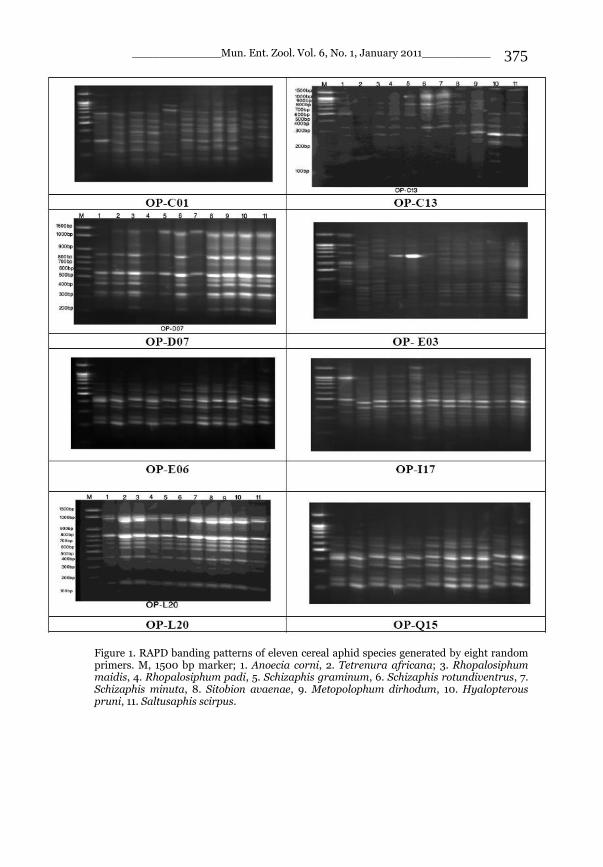
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 375
Figure 1. RAPD banding patterns of eleven cereal aphid species generated by eight random primers. M, 1500 bp marker; 1. Anoecia corni, 2. Tetrenura africana; 3. Rhopalosiphum maidis, 4. Rhopalosiphum padi, 5. Schizaphis graminum, 6. Schizaphis rotundiventrus, 7. Schizaphis minuta, 8. Sitobion avaenae, 9. Metopolophum dirhodum, 10. Hyalopterous pruni, 11. Saltusaphis scirpus.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 376
Figure 2. ISSR banding patterns of eleven cereal aphid species generated by five primers. M, 1000 bp marker; 1. Anoecia corni, 2. Tetrenura africana; 3. Rhopalosiphum maidis, 4. Rhopalosiphum padi, 5. Schizaphis graminum, 6. Schizaphis rotundiventrus, 7. Schizaphis minuta, 8. Sitobion avaenae, 9. Metopolophum dirhodum, 10. Hyalopterous pruni, 11. Saltusaphis scirpus.
Figure 3. Phylogenetic dendrogram among eleven cereal aphid species based on three criteria; RAPD markers, ISSRs markers and certain diagnostic morphological characters.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 377
THE EFFECT OF REGULATING COMPOUNDS ON THE GROWTH OF TRIBOLIUM CONFUSUM DU VAL
(COLEOPTERA: TENEBRIONIDAE)
Saeideh loni*, Maryam Moarefi**, Hossain Farazmand*** and Esmaeile Karami*
* Department of Entomology, Islamic Azad University, Arak Branch, Young Researchersclub of Arak, IRAN. E-mail: [email protected] ** Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Karaj Branch, Eram Blvd, Azadi St., Mehrshahr, P.O. Box 31876-44511, Karaj, IRAN. *** Assistant Professor, Iranian Research Institute of Plant Protection, Tehran, IRAN. [Ioni, S., Moarefi, M., Farazmand, H. & Karami, E. 2011. The effect of regulating compounds on the growth of Tribolium confusum du Val (Coleoptera: Tenebrionidae). Munis Entomology & Zoology, 6 (1): 377-385] ABSTRACT: Confused flour beetle, Tribolium confusum du Val (Col.: Tenebrionidae) is one of the most important stored product pests of flour in Iran. In this research the effects of insect growth regulators (IGRs): teflubenzuron, hexaflumuron and pyriproxyfen on oviposition rate, emergence of adults, repellency effects of 17 days-larvae and adults of confused flour beetle have been studied. The results showed that, the highest and lowest of inhibitory reproduction were observed in teflubenzuron (88.5%) and pyriproxyfen (77.4%), respectively. All IGRs significantly were reduced the oviposition rate of T. confusum.The maximum and minimum of inhibitory emergences of adults were observed in Teflubenzuron (86.26%) and pyriproxyfen (75.73%), respectively. Results showed that, reproduction and adult emergences inhibitories were directly correlated with the IGRs concentrations. Repellency effects of IGRs on adults indicated that, the highest and lowest were recorded for teflubenzuron (48.12%) and pyriproxyfen (39.37%). This effect‘s experiment on 17-days old larvae showed that, maximum and minimum repellency were in teflubenzuron (47.5%) and pyriproxyfen (36.87%), respectively. These results suggested that IGRs could be used as safe insecticides to control of confused flour beetle. KEY WORDS: insect growth regulator, teflubenzuron, hexaflumuron, pyriproxyfen, oviposition rate, emergence of adults, repellency of adults and larvae.
In Iran, an average of 10 to 20 percent of agricultural products, stored up in warehouses is wasted by pests (Moharamipour et al., 2003). Tribolium confusum is considered as one of the most important storage pests and can cause serious damage to store up products. This pest is more damaging in areas with hot and semisoft climates (Hollingsworth et al., 2002; Songa Reno, 1998). These insects not only cause a lot of damages while feeding, but also infest stored products with their larva layers and excrements and consequently lower the quality of stored products greatly. Adult insects and larva also feed on broken up grains (Bagheri Zenouz, 1995). At present, one of the most common methods of controlling stored up products is using the fumigant compounds such as methylbromide and phosphin. These two pesticides are very toxic for humans and have very severe side effects, therefore they are not used much anymore and are becoming limited (Bell & Wilson, 1995; Daglish & Collins, 1999). Furthermore it has been reported from many countries that the stored up products pests have become resistant against phosphine poison (Marouf et al., 2008). Presently extensive research is being done to find new compounds which cause less damage to the environment. Insect growth regulators like teflubenzuron, hexaflumuron (chitinase inhibitor) and pyriproxyfen (semi juvenile hormone) are among such compounds, which are less poisonous for human and other vertebrates. These compounds are cheap and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 378
can be easily produced on an industrial scale. In addition to these, low resistance of insects to such products and their complete degridation in short time are the most specific characters, (Mc Gregor and Kramer, 1977). Insect growth regulators are suitable for controlling stored up pests (Loschiavo, 1976). Furthermore application of diflobenzuron, chitin inhibitor, can cause 100% decrease in the first generation of adult Tribuliom confusum (Mc Gregor & Kramer, 1977). Since Mrize (1986) has shown that using doses higher than 600 ppm of R-20458 compounds in diet of Sitophilus oryzae cause infertility of S. oryzae species and stop the generation of F1.
In this research, the effect of the two chitin inhibitor, including, teflubenzuron and hexaflumuron and also pyriproxyfen semi juvenile hormone on oviposition and adult emergence and repelling effect in the larva stage of Tribolium confusum was studied to find the best compound affecting Tribolium confusum.
MATERIALS AND METHODS
In this study the effect of teflubenzuron, pyriproxyfen and hexaflumuron compounds were tested on 15 day larva and adult Tribolium confusum insects at agricultural college of Islamic Azad University–Arak branch in 2009-2010. Rearing of Tribolium confusum
An experiment was done by Arak university entomology group at its laboratory to test the Tribolium confusum‘s population. Mass rearing of this insect was maintained at the growth chamber set at 27 ± 2 °C and 75 ± 5% relative humidity on foodstuff in dark condition containing flour mix with yeast (17:1, w/w) in cylindrical container with height of 18 centimeter and diameter of 8 centimeters. The rate of inhibitory reproduction (IR)
To examine the effect of toxins on inhibitory reproduction of adults, on seven day adult insects were tested. First, paper filters with concentration of 4, 8, 16, 32 and 64 ppm were floated in compounds (with acetone solvent) and then were transferred to testing containers of 18 centimeter height and with diameter of 8 centimeter. After 30 minutes, to evaporate the acetone with the help on an aspirator, 20 seven day adult insects were released to the containers with define volume, having five grams of wheat flour and yeast (17:1), after 5 days oviposition on flours, the insects were softly eliminated from the containers. The containers were kept in testing conditions of (27 ± 2 °C and 75 ± 5% RH), and the larvae were counted after 20 days.The inhibitory rate was calculated by the following formula (Nazem Rafie et al., 2003).
IR=CK
TCK100
CK: The number of larvae in observer T: The number of larvae in under care Effect of the compounds on emergence of adults 7day adult insects used to measure the emergence rate. In this experiment, 5 different concentrations of compounds including 1, 2, 4, 8 and 16 ppm were used. Also in checking the experiment, only acetone solvent was used. One milliliter of each concentration was sprayed on 5 grams of flour and mixed with flour. 20, 7day adult insect were selected and transferred to dishes containing flour and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 379
permitted to feed and lay eggs. After 4 days adults were removed from dishes and were kept in 27 ± 2 °C and 75 ± 5 % RH in incubator. After 40 days the number of adult insects in each dish was counted. Repellency rate In this experiment Wattman forty paper filters with the diameter of 8 cm was cut into half from the middle. One half of the paper was floated in specific concentration of insecticide compound (containing three concentration of 65, 250, and 1000 ppm), and the other half was floated only in acetone solvent. After 20 minutes when paper filters were completely dried, paper filter halves which were treated with solvent and compounds were stick together. 20, seven day adult were selected and placed in the center of the paper filter. This experiment was also carried out with 20 and 17 day larvae in a separate experiment; after 24 hours, adult and 17 day larvae of T.confusum were counted on each half of paper filters. Repellency percentage of different concentration compounds were calculated using Talukder & Howse formula, 1993 and 1995. (Nazem Rafie et al., 2003).
PR=2(C – 50)
C: Percentage of insects on half of the paper observer filter PR: Repellency Percentage Analytical analysis Each of the experiments executed and repeated under completely randomized design in 4 replications. The data statistics were derived by SAS software and the average of experimental treatments were compared by with Duncan‘s range test were at a 5 % probability level.
RESULTS The effect of growth regulating compounds on inhibitory reproduction rate of T. confusum The results of the analysis proved that there is a meaningful relation between studied concentration and inhibitory reproduction. According to the results obtained, teflubenzuron and pyriproxyfen compounds had respectively the most and the least effect on the inhibitory reproduction of Adult Tribolium confusum in such a way that with the concentration of 65% ppm the inhibitory reproduction rate is recorded to be 100% and 93.28 % respectively. Based on the results, in 65% ppm and 32% ppm concentrations the teflubenzuron compound, and in the concentration of 65 ppm the hexaflumuron compound had the 100 % inhibitory reproduction. According to the data obtained the least amount of inhibitory reproduction is from pyriproxyfen compound showing 58.95% inhibitory reproduction at concentration of 4 ppm .It was also observed that teflubenzuron compound did not make a significant difference at concentration ranges of 16, 32 and 56 ppm. Also, at concentrations of 32 and 65 pyriproxyfen did not show any significant difference either. The experiment showed that there weren‘t any significant differences between 8 ppm Teflubenzuron, 8ppm Hexaflumuron and 16 ppm pyriproxyfen compounds. The obtained data also showed that there was not any significant difference between 4 ppm compounds tested.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 380
The effect of growth regulatory compounds on inhibitory emergence of adult of T. confusum Analysis results showed that the highest inhibitory rate on T. confusum was caused by teflubenzuron compound, so that this compound at 16 ppm concentration made the inhibitory emergence rate of adult insects increase to 98.66 percent and the lowest inhibitory rate related to pyriproxyfen compound, so that this compound at 16 ppm increased the inhibitory rate to 93.3 percent. According to data obtained, the inhibitory rate of 16 ppm hexaflumuron was 96 % which showed a significant difference compared to 1, 2 and 4 ppm. Results showed that the inhibitory rates of teflubenzuron, hexaflumorun and pyriproxyfen compounds at 16 ppm were 98.66 and 93.33 %, respectively, which did not show a significant difference. The inhibitory rate of 4 ppm teflubenzuron was 90% which did not have a significant difference compared to 8 ppm teflubenzuron. Also the inhibitory rate of this compound at 2 ppm was found 80% which did not show any significant difference compared to 4 ppm hexaflumorun and pyriproxyfen compound with inhibitory rate of 84 and 81.33 %, respectively. At 1 ppm concentration hexaflumorun and pyriproxyfen inhibitory emergence rates were 50.66 and 46 percent respectively which showed that there wasn‘t any significant difference between these two compounds at this density. The comparison percentage of repellency effect of compounds on 7 day adult of T. confusum The results of the repellency test of compounds on adult T.confusum insects showed that teflubenzuron compound had the highest repellency percentage, compared to hexaflumuron and pyriproxyfen compounds .Furthermore the pyriproxyfen compound showed the lowest repellency percentage. The variance analysis results showed that the teflubenzuron repellency percentage shows a significant difference compared to the other two compounds but there wasn‘t any significant difference between hexaflumuron and pyriproxyfen compounds, so have been put in the teflubenzuron compound showed 95 recent repelling at 1000 ppm (Fig. 1). Pyriproxyfen compound showed 35% repellency at 65 ppm was the lowest rate. In a comparative study, at 250 ppm, teflubenzuron compound had the highest effect on 7 day adult T. confusum insects compared to hexaflumuron and pyriproxyfen compounds, with the rate 65%. Also at this concentration hexaflumuron and pyriproxyfen compounds, both showed 52.5% repellency, which overall all three compounds didn‘t show a significant difference at this concentration. Variance analysis of repellence percentage of compounds showed that compound mutual effect concentrations are significant. Results showed that teflubenzuron compound at 1000 ppm have a higher repellency compared to other compounds tested, but at 1000 ppm, hexaflumuron and pyriproxyfen compounds didn‘t show a significant difference at level of 5 percent. Also at 65 ppm hexaflumuron, teflubenzuron and pyriproxyfen at level of 5% didn‘t show any repellency percentage. Comparison of repellency percentage compounds on old Tribolium confusum larvae The result of this repellency percentage test on T. confusum larvae showed that Teflubenzuron compound had the highest and pyriproxyfen compound had the lowest repellency percentage. According to statistical analysis results, there was no significant difference between hexaflumuron and pyriproxyfen compounds at 5 percent level and they were placed in one group. Variance percentage analysis of repellency percentage of compounds being studied showed

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 381
that the mutual effects of compounds and concentration were significant. This study showed that teflubenzuron compound at 1000 ppm had the highest repellency percentage compared to other who compounds and increased repellency rate to 98.5 % and this compound at this concentration showed a significant difference in repellency; this repellency percentage was higher than those in hexaflumuron and pyriproxyfen compounds, which shows its higher strength in repellency. At 250 ppm concentration teflubenzuron compound with 67.5% repellency did not show significant difference compared to the repellency percentage of hexaflumuron and pyriproxyfen compounds at 1000 ppm concentration regarding the T. confusum larvae. At 1000, 250 and 65 ppm concentration of hexaflumuron and pyriproxyfen there was no significant difference in their repellency percentage (Fig. 2). Also hexaflumuron, teflubenzuron and pyriproxyfen compounds being tested did not show a significant difference at 65 ppm.
DISCUSSION According to the studies on inhibitory reproduction, it was observed that as the concentration increases, the inhibitory reproduction increases the oviposition as well, this is in agreement with previous results obtained (Loschiavo, 1976), pointing out that with the increasing the concentration of inhibitory growth compounds being used, prevents adult oviposition. Also according to the studies done, it was observed that inhibitory compounds for chitin synthesis in high concentrations, stop oviposition hundred percent in the performed experiments. These results is also in agreement with obtained results (Nawrot et al., 1987) showed that using teflubenzuron decreases T. confusum oviposition hundred percent. According to these studies insect growth inhibitory compounds play a role in oviposition inhibitory which agrees with the results obtained by Leschiavo in 1976. Results relate to the inhibitory effect of anti-chitin synthesis compound is more than that of be a Babu et al. (1991), which reported that triflumuoron and flucydoxuroun compounds at 20 ppm concentration reduce Callosobruchus chinensis oviposition at the rate of 75 and 80% respectively, While teflubenzuron and hexaflumuron inhibitory oviposition at 16 ppm concentration where 93.93% and 86.86% respectively. The results of this experiment also indicated that anti-chitin synthesis compounds have a high effect on inhibitory of the oviposition compared to synthetic juvenile hormone compounds which is in agreement with the results obtained (Eisa et al., 1986) pointing out the high effect of inhibitory chitin synthesis toxins in T. castaneum . Results from this study, agree with the effects of triflumuron on T. castaneum oviposition rate (Eisa et al., 1984). In these experiments mutual effect and concentration where not found significant which leads to the conclusion that regardless of the kind of the compound, concentration increase, leads to higher percentage of inhibitory rate .Results are obtained from the effect of inhibitory corrupt compounds on inhibitory growth rate of adult insects show that teflubenzuron and pyriproxyfen have the highest and lowest effect on emergence rates of adult T. cofusum insect respectively, so that at 16 ppm they show 96.66% and 93.33% inhibitory rate, respectively. Methoprene is highly effective in reducing adult Plodia interpunctella emergence after larval exposure (Jenson et al., 2009). Also similar experiment by Mulla and Main (1982) showed that using triflumuron compound at five ppm concentration cause 92.5% reduction of the first generation for T. castaneum, for Oryzaephilus surinamensis 97.5% and for R. dominica it was 100%. This research showed that in low concentrations of

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 382
teflubenzuron the inhibitory emergence rates of adult insect was more than that of RH-5849 and triflumuron. Since Ammar (1988) has shown that chlorfluazuron, flufenozuron and 473 compounds with concentration of 0.5-25%, on adult Sitophylus oryzae, caused 91% - 98% reduction of the first generation within eight months after. Also five month after treatment, the reduction of first-generation of S. oryzae was 76/54 – 100%, while this research showed that the two anti-chitin synthesis, Teflubenzuron and Hexaflumuron compound and Pyriproxyfen, synthetic juvenile hormone, will cause a 93%- 98% reduction after four and five months, respectively. The results of the experiments performed, showed that increasing the concentration of growth inhibitory compounds in the insect diets, decreases the emergence rates of healthy insects, in such a way that the reduction trend continued up to 2 ppm concentration. In Teflubenzuron compound concentrations higher than 16 ppm and in Hexaflumuron compounds higher than 32 ppm no adult emerged. Since Eisa et al. (1986) has shown that chlorofluazuron and flufenoxuron compounds at densities of 1 and 10 ppm where tested on T. castaneum and no adults emerge. Pyriproxyfen compound at densities higher than 8 ppm a percentage of adult emerged with the incomplete formation. At densities from 2 to 16 ppm, besides the emergence of complete insects which appear to have no unnatural characteristics in their limbs, a percentage emerged as unnatural insects as well. In these insects unnatural characteristics were seen more in the front wings, in such a way that we are sure to deform and twisted in odd figures (asymmetry on left and right wings) were observed. Furthermore, sometime the formations were so severe that insects could not even walk. Sometimes the wings and feet were deformed in such an extent that the emergence of the insect out of its pupa is almost impossible half way through which causes the insect to die inside. Also, the internal and external organs of existing pupa, in a specific concentrations, have been deformed in such a way that the larvae dies.For example the similarity of pupa and larva's head. The general conclusion of the discussion clarifies the point that anti-chitin synthesis teflubenzuron and hexaflumuron compounds are more effective than pyriproxyfen synthetic juvenile hormone compounds on emergence of adults and also along with the increase in diet concentration compounds, which larva and pupa periods go through it, the rate of complete emergence would reduce to that extent that it higher concentrations than 16 ppm there is no adult emergence which can be due to prolonged larval growth as well as interference with metamorphosis or the death of pupa. In this research the effect of insects growth preventing compounds on adult insect exit rate was studied and it was shown that teflubenzuron had the highest repelling rate in such a way that in 1000 ppm this rate was recorded to be 98.7% and 97.4% on adult Tribolium confusum larvae and adult insect, respectively .In a similar experiment by Liu & Ho (1999) the Evodia rutaecrpa herbal oil had the highest repelling factor with 88.5 %. In this experiment the lowest percentage was for pyriproxyfen in such a way that in 65 ppm concentration this rate was 47.9 % and 48.5 % for Tribolium confusum larvae and adult insects respectively. The difference between these repelling compounds can be due to the fact that the fumigant activity of pyriproxyfen is much less than teflubenzuron compound and as a consequence its repelling property is low as well or the teflubenzuron compound odor is more repelling in comparison to that of the pyriproxyfen. The other reason that can be accounted for teflubenzuron more repelling property is that probably the effective compound can strongly repel the insect. This is in agreement with the results obtained (Xie et al., 1996), pointed out that Aphanamixis polystachya could strongly repel T. castaneum. The repellency

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 383
effect of Melia toosendan which can strongly repel adult T. castaneum could be attributed to its major constituent, tousendanin (Nazem Rafie et al., 2003). Studies conducted to test the repellency of pea, Pisum sativum (L.), products against Tribolium castaneum (Herbst.), Sitophilus oryzae (L.) and Rhyzopertha dominica (F.) showed that paddy grains treated with 1% concentration were repellent to adults. Tribolium castaneum was repelled most quickly followed by S. oryzae and R. dominica (Pretheep Kumar et al., 2004).
ACKNOWLEDGEMENTS We gratefully acknowledge the financial support of Islamic Azad University, through this research program.
LITERATURE CITED Ammar, I. M. A. 1988. Residual bioactivity of insect growth regulators against Sitophilus oryzae in wheat grain. Anzeiger fur Schadlingskde Pflanzenschutz Umweltschutz, 61 (3): 56-60. Babu, R. T., Hussaini, S. H., Sriramulu, M. & Siddiqui, M. K. H. 1991. Effect of inert clay and insect growth regulators on the development of Callosobruchus chinensis L. and the germination of mungbean seed [Vigna radiate (L.) Wilczek]. Tropical Science, 31: 217-221. Bagheri-Zenouz, E. 1995. Stored products pests and its control methods. 2nd ed. 309 pp. Sepehr Publishing. Bell, C. H. & Wilson, S. M. 1995. Phosphine tolerance and resistance in Trogoderma granarium (Everts.) (Coleoptersa: Dermestidae). Journal of Stored Products Research, 31: 199-205. Daglish, G. J. & Collins, P. J. 1999. Improving the relervance of assays for Phpsphine resistance. In: Stored product protection, Eds. Jin, X., Liang, Q. Liang, Y. S., Tan, X.C. and Guan, L. H., pp. 584-593. Eisa, A. A., Ammar, I. M. A. & EL-Sheikh, A. E. A. 1986. Effect of two insect growth regulators and the ovicide isoxathion on the red flour beetle Tribolium castaneum when admixed with wheat flour and egg treatment. Annals of Agricultural Science, Moshtohor, 24 (7): 1689-1696. Eisa, A. A., Ammar, I. M. A., Radwan, H. S. A. & EL-Nabawi, A. 1984. Larvicidal activity and development inhibition of the red flour beetle Tribolium castaneum fed insect growth regulator treated wheat flour. Minufiya Journal of Agricultural Research, 8: 405-414. Hollingsworth, C. S., Coil, W. M., Murray, K. D. & Ferro, D. N. 2002. Intergrated Pest Management for Northeast Schools. Natural Resource, Agriculture and Engineering Service. NRAES-152, pp.60. Jenson, E. A., Arthur, F. H. & Nechols, J. R. 2009. Efficacy of an esfenvalerate plus methoprene aerosol for the control of eggs and fifth instars of Plodia interpunctella (Lepidoptera: Pyralidae). Insect Science. 17 (1): 21-28. Liu, Z. I. & Ho, S. H. 1999. Bioactivity of the essential oil extracted from Evodia rutaecarpa Hook against the grain storage insects, Sitophilus zeamais Motsch. And Tribolium castaneum Herbst. Journal of Stored Products Research, 35: 317-328. Loschiavo, S. R. 1976. Effects of the synthetic insect growth regulators methoprene and hydroprene on survival, development or reproduction of six species of stored products insects. Journal of Economic Entomology, 69 (3): 395-399. Marouf, A., Sangari, S., & Jabbari, L. 2008. An investigation on fumigant effect of the extract of Origanum vulgare (Lamiales: Lamiaceae) for control of two stored-product beetles. Journal of Entomological Society of Iran. 27 (2): 29-41. McGregor, H. E. & Kramer, K. J. 1977. Activity of Dimilin (TH 6040) against Coleoptera in stored wheat and corn. Journal of Economic Entomology, 69: 479.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 384
Mian, L. S. & Mulla, M. S. 1982. Biological activity of insect growth regulators against four stored product coleopterans. Journal of Economic Entomology, 75 (1): 80-85. Moharramipour, S., Nazemirafieh, J., Morovati, M., Talebi, A. A. & Fathipour, Y. 2003. Effectiveness of exreacts of Nerium oleander, Lavandula officinalis and Ferula assafoeda on nutritional indices of Tribolium castaneum adults. Journal of Entomological Society of Iran. 23 (1): 69-89. Nawrot, J., Czaplicki, E. & Sobotka, W. 1987. Effectiveness of new chitin synthesis inhibitors against some stored product pests. pp. 583-590. In: Proceedings of the Fourth International Working Conference on Stored Product Protection, Tel Aviv, Israel, September 1987. Nazemi-Rafie, J., Moharamipour, S. & Marouti, M. 2003. Insecticidal activities of etanoli extera Nerium oleander, Lavandula offcinalis and metanoli extera Ferula assafoetida against Ephestia kuehniella. Proceeding of 15th Iranian plant protection congress, P.144. Pretheep Kumar, P., Mohan, S. & Balasubramanian, G. 2004. Effect of whole-pea flour and a protein-rich fraction as repellents against stored-product insects. Journal of Stored Products Research, 40 (2004): 547-552. Songa, J. & Rono, W. 1998. Indigenous methods for bruchid beetle (Coleoptera: Bruchidae) control in stored beans (Phaseolus vulgaris L.). Internatioal Journal of Management, 44 (1): 1-4. Xie, Y. S., Bodnaryk, R. P. & Fields, P. G. 1996. A rapid and simple flour disk bioassay for testing natural substances active against stored-product insects. Canadian Entomologist, 128: 865-875.
Table 1. Shows the effect of regulating compounds with different concentrations on the inhibitory reproduction of adult T.confusium under the laboratory simulated conditions.
Pyriproxyfen Hexaflumuron Teflubenzuron concentration
f 3.41 58.95 f 2.58 64.92 ef 58 2 67.91 4 ppm
ef 5.92 68.65 ed 2.58 76.11 cd 4.66 79.10 8 ppm
cd 17.09 79.10 ab 5.17 90.29 ab 7.75 95.52 16 ppm
cb 4.66 87.31 ab 6.46 94.20 a 0.00 100 32 ppm
ab 5.92 93.28 a 0.00 100 a 0.00 100 65 ppm
1- Means ± SE 2- Means within column followed by the same letter not found significant (P<0.05, DMRT) Table 2. The effect of growth regulatory compounds with various concentrations on inhibitory emergence of adult of T .confusum in laboratory conditions.
1- Means ± SE 2- Means within column followed by the same letter not found significant (P<0.05, DMRT)
Pyriproxyfen Hexaflumuron Teflubenzuron Concentration
f 4.00 46.4 f 9.01 50.66 e 5.77 66.66 1 ppm
e 2.00 68 e 1.15 70.66 d 4.00 80 2 ppm
d 1.15 81.32 cd 3.46 84 bc 0.00 90 4 ppm
bc 2.00 90 b 3.05 90.66 ab 2 96 8 ppm
ab 1.15 93.33 ab 2.00 96 a 2.3 98.66 16 ppm
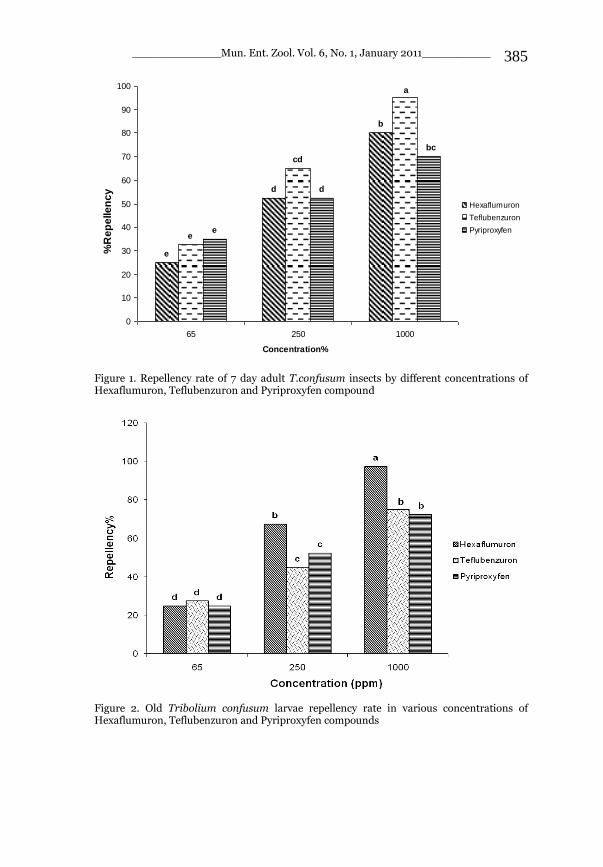
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 385
b
d
e
a
cd
e
bc
d
e
0
10
20
30
40
50
60
70
80
90
100
65 250 1000
Concentration%
%R
ep
elle
nc
y
Hexaflumuron
Teflubenzuron
Pyriproxyfen
Figure 1. Repellency rate of 7 day adult T.confusum insects by different concentrations of Hexaflumuron, Teflubenzuron and Pyriproxyfen compound
Figure 2. Old Tribolium confusum larvae repellency rate in various concentrations of Hexaflumuron, Teflubenzuron and Pyriproxyfen compounds

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 386
A LIST OF POMPILIDAE (HYMENOPTERA) OF GÖLCÜK NATURAL PARK, ISPARTA, TURKEY
George Japoshvili*, İsmail Karaca**, Raymond Wahis***
* Entomology and Biocontrol Research Centre, Ilia State University, Tbilisi, GEORGIA. E-mail: [email protected] ** Department of Plant Protection, Suleyman Demirel University, Isparta, TURKEY. E-mail: [email protected] *** Gembloux Agro bio tech, Université de Liège, Entomologie fonctionnelle et évolutive, Gembloux, BELGIQUE. E-mail: [email protected] [Japoshvili, G., Karaca, İ. & Wahis, R. 2011. A list of Pompilidae (Hymenoptera) 0f Gölcük Natural Park, Isparta, Turkey. Munis Entomology & Zoology, 6 (1): 386-388] ABSTRACT: Pompilidae species were collected during 2008-2009 in Gölcük Natural Park, which belongs to Isparta province. Two methodologies for collecting were used: pitfall and malaise trapping. Thirty-three species belonging to three subfamilies and 14 genera were recorded during this survey. One species is new record for Turkey: Priocnemis pogonioides (Costa, 1883). Twenty-eight more species are new records for Isparta province. KEY WORDS: Pompilidae, Gölcük, Isparta, Turkey.
The family Pompilidae, known as the ―spider wasps‖ is almost cosmopolitane. There are 4000-4500 species of pompilids worldwide and more then 200 species in Turkey. The aim of this study is to survey Pompilidae distributed in Golcuk Natural Park (GNP). GNP is an arid area located eight km southwest of city Isparta and has a total area of 5,925 ha (Gül et al., 2005). With its diverse vegetation and wildlife, geomorphological structure, aesthetically pleasing landscape, and recreational opportunities, GNP is one of the most important areas of the Lakes District in Turkey.
Passive sampling took place in GNP at altitudes from 1227 m to 1611 m. Insect samples were collected from March to October, 2008 and 2009, using pitfall and Malaise trapping methods. At all sampling sites 10 pitfall traps were set. The pitfall traps consisted of circular pots, each 11 cm in diameter and 11 cm deep, and were dug into the soil with the opening at the soil surface. At the beginning of the study, dry traps were used, but a month later, it was necessary to slightly modify the methodology. We added 2% formaldehyde to the traps to avoid sample damage due to the insects themselves and other animals such as mice, shrews, and lizards. Pitfall traps were checked weekly. Malaise traps were set in three locations: 1) Pilav Tepe (1520 m), 2) Acacia reforested area near the main entrance of GNP (1414 m), and 3) an area which was previously used for agricultural cultivation (1452 m). Malaise traps were checked and material was taken every 10 days. After collecting the captured insects, they were dried and pinned promptly or were transferred to jars of 75% alcohol and mounted at a later time. Material was identified by third author.
The species list is arranged alphabetically for each family. After the valid name of the relevant taxon, the records (if available) of the authors with the locality, date, and collector follow. New taxa for the Isparta Province are marked with an asterix (*), and Turkish fauna are marked with two asterixes (**). All distributional information about species included in our list, except new records, is given in following papers: Ozbek et al., 1999, 2000; Smissen, 2003; Anlas et al., 2005; Wahis, 2006a,b; Kirpik, 2009.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 387
The following institutional abbreviations are used: GNP – Gölcük Natural Park; MT – Malaise trap; PT – pitfall trap. Voucher specimens are deposited in the entomological collections of Agricultural Faculty of Suleyman Demirel University, Isparta, Turkey. Subfamily Ceropalinae: Ceropales helvetica Tournier, 1889* Subfamily Pepsinae: Cryptocheilus notatus (Rossius, 1792)* Cryptocheilus setiger Wolf, 1999* Dipogon variegatus (Linné, 1758)* Priocnemis pogonioides (Costa, 1883)** Priocnemis vulgaris (Dufour, 1841) Subfamily Pompilinae: Agenioideus excisus (Morawitz, 1890)* Agenioideus nubecula (Costa, 1874)* Agenioideus ruficeps (Eversmann, 1886)* Agenioideus sericeus (Vander Linden, 1827)* Anoplius viaticus (Linné, 1758) Aporinellus moestus (Klug, 1834)* Aporinellus sexmaculatus (Spinola, 1805)* Arachnospila anceps (Wesmael, 1851)* Arachnospila consobrina (Dahlbom, 1843)* Arachnospila fumipennis (Zetterstedt, 1838)* Arachnospila fuscomarginata (Thomson, 1870)* Arachnospila minutula (Dahlbom, 1842)* Arachnospila sogdianoides (Wolf, 1964) Arachnotheutes leucurus (Morawitz, 1891)* Arachnotheutes rufithorax (Costa, 1834)* Batozonellus lacerticida (Pallas, 1771)* Eoferreola manticata (Pallas, 1771)* Episyron albonotatum (Vander Linden, 1827)* Episyron gallicum (Tournier, 1889)* Evagetes anatolicus V. d. Smissen, 2003* Evagetes crassicornis (Shuckard, 1837) Evagetes elongatus (Lepeletier, 1845)* Evagetes pectinipes (Linné, 1758)* Evagetes siculus (Lepeletier, 1845)* Evagetes trispinosus (Kohl, 1886)* Evagetes tumidosus (Tourniser, 1890)* Tachyagetes furvescens Wahis, 1970*
As a result of sampling over the time of two years, we identified 33 species of Pompilidae belonging to three subfamilies and 14 genera. Ceropalinae is represented by one genus and one species and Pepsinae by three genera and five species. The highest number of species recorded from Pompilinae, with 10 genera and 27 species. One species is new records for Turkey: Priocnemis pogonioides (Costa, 1883). Twenty-eight of the 33 species are new records for Isparta Province.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 388
ACKNOWLEDGEMENTS
We would like to thank to Elizabeth Murray, Department of Entomology, University of California, Riverside, CA, USA, who kindly helped to check English and gave advice to improve this paper.
LITERATURE CITED Anlas, S., Wahis, R. & Tezcan, S. 2005. Some additional notes on Pompilidae (Hymenoptera) fauna of Turkey. Journal of Turkish Entomology, 29 (4): 255-258. Gul, A., Orucu, O. & Karaca, O. 2005. Determination of Potential Regions by Using Recreation Suitabilty Analaysis (Reference Gölcük Reserve). Symposium of Protected Natural Areas. 8-10 September 2005, S.D.Ü., Isparta, Turkey. [In Turkish]. Kirpik, M. 2009. Faunistic study on Pompilidae (Insecta: Hymenoptera) family in Ankara, Kırıkkale, and Çankırı provinces. African Journal of Biotechnology, 8 (2): 316-328. Ozbek, H., Yildirim, E., Wolf, H. & Wahis, R. 1999. The Pompilidae (Hymenoptera, Aculeata) fauna of Turkey. Part 1: Ceropalinae and Pepsinae. Zoology in the Middle East, 18: 91-105. Ozbek, H., Yildirim, E., Wolf, H. & Wahis, R. 2000. The Pompilidae (Hymenoptera, Aculeata) fauna of Turkey. Part II: Pompilinae. Zoology in the Middle East, 21: 109-128. Smissen, J. van der. 2003. Revision der europaïschen und türkischen Arten der Gattung Evagetes Lepeletier 1845 unter Berücksichtigung der Geäderbweichungen. Mit zweisprachigem Schlüssel zur Determination (Hymenoptera: Pompilidae). Verhandlungen des Vereins für Naturwissenschaftliche Heimatforschung zu Hamburg e., 42: 1-253. Wahis, R. 2006a. Notules taxinomiques sur les Pompilides I (1-24) (Hymenoptera Pompilidae). Notes fauniques de Gembloux, 59 (3): 129-138. Wahis, R. 2006b. Mise à jour du Catalogue systématique des Hyménoptères Pompilides de la région ouest-européenne. Additions et Corrections. Notes fauniques de Gembloux, 59 (3): 31-36.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 389
FAUNISTIC STUDY ON HEMIPTERA IN ARASBARAN AREA (EAST AZARBAIJAN PROVINCE, IRAN)
Mostafa Nikdel*, Ali Asgar Dordaei* and Armand Matocq**
* Agricultural and Natural Resource Research Center of East Azarbaijan, Tabriz-IRAN. E-mail: [email protected] ** LNPV, Station d'Entomologie, Supagro bâtiment 16, 2, Place Viala, F-34060 Montpellier cedex 01, FRANCE. [Nikdel, M., Dordaei, A. A. & Matocq, A. 2011. Faunistic study on Hemiptera in Arasbaran area (East Azarbaijan province, Iran). Munis Entomology & Zoology, 6 (1): 389-395] ABSTRACT: In the course of studing insect fauna in Arasbaran area (in East Azarbaijan Province), a survay was counducted on Hemiptera in townships of Kaleibar, Khod Afarin, Varzeghan and Horand during 1998-2001. The specimens were collected on trees, shrubs and herbaceous plants and were identified as 50 species belonged to 43 genera, 13 families. Among the determined specimens the following five genera (are specified with an asterisk) and 10 species are newly recorded from Iran as *Aquarius lacustris, *Blepharidopterus angulatus, Cyllecoris histrionius, Dicranocephalus agilis, Dicranocephalus medius, Heterogaster affinis, *Peirates hybridus, Picromerus bidens, Sehirus luctuosus, *Trichocorixa sp., *Hesperocorixa scabricula. KEY WORDS: Arasbaran area, Hemiptera, fauna, new record, Iran.
The Heteroptera, or ―true bugs‖, are among the most diverse groups within the Hemiptera, and are usually identified by the strong odor produced by the metathoracic scent glands. The Hemiptera is the fifth largest group of insects with at least 80.000 described species with the Heteroptera being the largest group with more than 35.000 species (Forero, 2006; Gullan & Cranston, 1994).
The Heteroptera insects feed on plant juices or live as predators and parasites. Many of such insects that feed on the plant are known as serious plant pests (Safavi, 1993). The damage caused by the insect as a result of sucking sap from food plants, is often increased by the salivary enzymes, which may considerably alter the quality of plant products. On the other hand, many predators, catch other insects and Acarina, and very beneficial from agricultural point of view (Hassazadeh et al., 2009). Terrestrial species may be very important agricultural pests, pest controlling predators, or vectors of disease. Aquatic species help control nuisance insects, some are used for human or pet food, some are bitting nuisances, some may feed on hatchery fish, only corixids are normally fed upon by fish (Slater, 1981).
The study of the true bugs fauna of Iran was started by M. Safavi in the 1940s and later by other researchers has continued. They were studied mainly on visiable and big size hemipterans (Safavi, 1991; Safavi, 1993; Linnavuori & Modarres Awal, 1998; Modarres Awal, 2002; Yarmand et al., 2004; Sedghian et al., 2004; Askari, 2009; Hassazadeh et al., 2009 and Khaghaninia et al., 2010), and published some lists of the Hemiptera species which had been described or recorded from Iran, with identification key for 27 families. Even so, the true bugs that occur in Iran are poorly known especially the aqueous bugs. The data for many groups is sparse and no comprehensive account or checklist exists for the country. The number of recorded true bugs species in Iran is not more than 700 according to published papers.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 390
Arasbaran is one of the important forest areas in Iran. The area is characterized by special climatic characteristics, high biodiversity, presence of rare fauna and flora and vegetation elements associated with various climates. This has placed the area as a biosphere reserve under the UNESCO's MAB program.
The aim of this paper is to improve our understanding and to provide more information about the fauna and distribution of the true bugs of Arasbaran forest and rangelands (townships of Kaleibar, Khodafarin, Varzeghan and Horand) in Azarbaijan province, North West of Iran.
MATERIAL AND METHODS
Hemipterans were sampled with irregular surveys at different locations in the Arasbaran area on grasslands and forest trees during 1998- 2001. Most of the examined materials were collected by using a short-handled net, by sweeping back and forth among herbaceous vegetation, aspirator and light trap, the net can also be placed underside the plant and shaken. In aquatic collecting, nets were used to sweep the top of the water. In the laboratory, true bugs specimens were mounted and pinned for identification soon after collection using the usual methods. All of the specimens were identified and confirmed by third author. The examined material of the hemiperan species were deposited at Agricultural and Natural Resource Research Center of East Azarbaijan, Plant Pests and Diseases Research Institute, Tehran, Iran.
RESULTS
In this study the faunistic information, geographical distribution and locality data including altitude and GPS cordination (Latlon = lathitude & longitude) for each species for 50 species of Arasbaran area belonging to 43 genera, 13 families from Hemiptera are presented. Of which five genera (Aquarius, Blepharidopterus, Peirates, Trichocorixa and Hesperocorixa) and 10 species are newly recorded from Iran based on the records scattered in the literature (Safavi, 1991; Safavi, 1993; Linnavuori, 1997; Linnavuori & Modarres Awal, 1998; Modarres Awal, 2002; Yarmand et al., 2004; Sedghian et al., 2004; Hassazadeh et al., 2009 and Khaghaninia et al., 2010). The newly reported genera and species are marked with one and two asterisks, respectively. Family Acanthosomatidae Signoret, 1864 (Shield Bugs) Acanthosoma haemorrhoidale (Linnaeus, 1758) Material examined: 8 specimens, Khoda Afarin, Kalaleh, 1250 m. GPS cordination: N38 40.57 E46 32.05 Note: This species has been collected on different forest trees especially Crataegus meyeri and Quercus spp from Iran (Sadaghian et al., 2004).
Family Coreidae Leach, 1815 (Leaffooted Bugs) Ceraleptus gracilicornis (Herrich-Schäffer, 1835) Material examined: 8 specimens, Kaleibar, Hejrandoost, 1350 m. GPS cordination: N38 52.551 E46 57.54 Coreus marginatus (Linnaeus, 1758) Material examined: 15 specimens, Khoda Afarin, Choulan, 800 m. GPS cordination: N38 48.28 E46 32.16

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 391
Gonocerus acuteangulus (Goeze, 1778) Material examined: 17 specimens, Khoda Afarin, Choulan, 750 m. GPS cordination: N38 48.28 E46 32.16 Syromastus rhombeus (Linnaeus, 1767) Material examined: 5 specimens, Khoda Afarin, Research Forest, 1350 m. GPS coordination: N38 53.27 E46 46.50 Camptopus lateralis (Germar 1817) Material examined: 2 specimens, Khoda Afarin, Choulan, 810 m. GPS cordination: N38 48.28 E46 32.16
Family Cydnidae Billberg, 1820 (Burrowing Bugs) Sehirus luctuosus** Mulsant & Rey, 1866 Material examined: 2 specimens, Kaleibar, Makidy village, 1600 m. GPS cordination: N39 00.44 E46 41.33 Note: This species similar to Pentatomidae but with strong spines on tibiae and burrow in the ground where it feed on plant roots.
Family Lygaeidae Schilling, 1829 (Seed Bugs) Lygaeus saxatilis (Scopoli, 1763) Material examined: 6 specimens, Khoda Afarin, Kalaleh village, 1350 m. GPS cordination: N38 40.57 E46 32.05 Lygaeus pandarus (Scopoli, 1763) Material examined: 20 specimens, Kaleibar, Naposhteh village, 1510 m. GPS cordination: N38 51.715 E46 56.14 Lygaeus equestris (Linnaeus, 1758) Material examined: 11 specimens, Khoda Afarin, Research Forest, 1350 m. GPS cordination: N38 53.27 E46 46.50 Eremocoris sp. Material examined: 3 specimens, Khoda Afarin, Research Forest, 1350 m. GPS cordination: N38 53.29 E46 46.53 Heterogaster affinis** Herrich-Schäffer, 1835 Material examined: 1 specimen, Khoda Afarin, Kalaleh, 1360 m. GPS cordination: N38 56.33 E46 45.60 Lasiocoris amomalus (Kolenati, 1845) Material examined: 2 specimens, Khoda Afarin, Forest Nursery, 1350 m. GPS cordination: N38 56.33 E46 45.60 Rhyparochromus sp. Material examined: 6 specimens, Kaleibar, Makidy, 1580 m and Varzegan, Tazekand, 1142 m. GPS cordination: N39 00.44 E46 41.33 and N38 52.39 E46 49.88 Tropidothorax leucopterus (Goeze, 1778) Material examined: 2 specimens, Kaleibar, Makidy village, 1700 m. GPS cordination: N39 00.44 E46 41.33

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 392
Family Miridae Hahn, 1831 (Leaf Bugs) Rhabdomiris striatellus striatellus (Fabricius, 1794) Material examined: 5 specimens, Kaleibar, Makidy village, 1750 m. GPS cordination: N39 02.43 E46 41.36 Note: The species has been collected for the first time from Arasbaran forests, East Azarbaijan province in Iran (Yarmand et al., 2004). Cyllecoris histrionius** (Linnaeus, 1767) Material examined: 2 specimens, Kaleibar, Shojae Abad, 1780 m. GPS coordination: N38 55.53 E 46 46.21 Lygus rugulipennis Poppius, 1911 Material examined: 2 specimens, Kaleibar, Makidy village, 1750 m. GPS coordination: N39 02.43 E46 41.36 Deraeocoris lutescens Schilling, 1837 Material examined: 3 specimens, Khoda Afarin, Kalaleh, 1350 m. GPS cordination: N38 40.57 E46 32.05 Adelphocoris seticornis (Fabricius 1775) Material examined: 2 specimens, Khoda Afarin, Kalaleh Sofla, 1300 m. GPS coordination: N38 56.33 E46 45.60 Blepharidopterus*angulatus** Fallen, 1807 Material examined: 3 specimens, Kaleibar, Aliabad, 1650 m. GPS coordination: N38 53.77 E46 62.49 Adelphocoris vandalicus (Rossi, 1790) Material examined: 4 specimens, Khoda Afarin, Kalaleh Sofla, 850 m. GPS coordination: N38 56.31 E46 44.62 Lygus gemellatus gemellatus (Herrich-Schaeffer, 1835) Material examined: 2 specimens, Kaleibar, Aliabad, 1550 m. GPS coordination: N38 52.79 E46 61.48 Oncotylus viridiflavus viridiflavus (Goeze, 1778) Material examined: 4 specimens, Kaleibar, Shojae Abad, 1780 m. GPS coordination: N38 55.53 E 46 46.21 Note: This species is commonly distributed in Iran on Tamarix spp (Modarres Awal, 2002).
Family Nabidae Costa, 1852 (Damsel Bugs) Nabis sp. Material examined: 8 specimens, Kaleibar, Shojae Abad, 1780 m. GPS coordination: N38 55.53 E 46 46.21 Prostemma guttula (Fabricius, 1787) Material examined: 5 specimens, Kaleibar, Makidy village, 1750 m ,1 specimensKhoda Afarin, Kalaleh Sofla, 850 m. GPS coordination: N39 02.43 E46 41.36, N38 56.31 E46 44.62
Family Reduviidae Latreille, 1807 (Assassin Bugs) Peirates* hybridus** (Scopoli, 1763) Material examined: 13 specimens, Khoda Afarin, Kalaleh Sofla, 800 m. GPS coordination: N38 56.32 E46 45.63

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 393
Rhinocoris sp. Material examined: 25 specimens, Khoda Afarin, Kalaleh Olia, 1350 m. GPS coordination: N38 40.57 E46 32.05
Family Rhopalidae Amyot & Serville, 1845 (Scentless plant Bugs) Corizus hyosciami (Linnaeus, 1758) Material examined: 2 specimens, Kaleibar, Naposhteh village, 1410 m. GPS coordination: N39 00.69 E46 50.19 Maccevethus sp. Material examined: 3 specimens, Kaleibar, Naposhteh, 1000 m. GPS coordination: N38 52.542 E46 57.53 Rhopalus parumpunctatus Schilling, 1829 Material examined: 30 specimens, Khoda Afarin, Research Forest, 1250 m. GPS coordination: N38 41.34 E46 34.07 Maccevethus caucasicus (Kolenati, 1845) Material examined: 6 specimens, Khoda Afarin, Research Forest, 1350 m. GPS coordination: N38 57.36 E46 45.62
Family Pentatomidae Leach, 1815 (Stink Bugs) Carpocoris mediterraneus Tamanini, 1958 Material examined: 30 specimens, Kaleibar, Makidi, 1550 m. GPS coordination: N39 00.44 E46 41.33 Palomena prasina (Linnaeus, 1761) Material examined: 9 specimens, Khoda Afarin, Research Forest, 1350 m. GPS coordination: N38 57.36 E46 45.62 Note: This species is commonly called Green Shield Bug. Piezodorus litearatus (Fabricius, 1794) Material examined: 14 specimens, Khoda Afarin, Kalaleh Sofla, 800 m. GPS coordination: N38 56.33 E46 45.60 Pentatoma rufipes (Linnaeus, 1758) Material examined: 15 specimens, Varzeghann, Khoinarood, 1495 m. GPS coordination: N38 48.37 E46 46.16 Note: This species is commonly distributed on forest area and called Forest Bug. Eurydema ornatum Linnaeus, 1758 Material examined: 10 specimens, Khoda Afarin, Kalaleh Sofla, 850 m., 10 specimens, Kaleibar, Vinag, 1350 m. GPS coordination: N38 56.33 E46 45.60, N39 00.69 E46 50.19 Eurydema putoni Jakovlev, 1877 Material examined: 6 specimens, Horand, Dehrood, 850 m., 10 specimens, Kaleibar, Gandomnan, 1350 m. GPS coordination: N38 52.158 E46 49.310, N38 59.53 E46 Note: The two above mentioned species has been collected from different regions of Iran on many kinds of cultural plants (Modarres Awal, 2002). Rhaphigaster nebolosa (Poda, 1761) Material examined: 2 specimens, Kaleibar, Garmanab, 1650 m. GPS coordination: N38 55.66 E46 46.82

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 394
Mustha sp. Material examined: 30 specimens, Khoda Afarin, Ainalou, 1450 m. GPS coordination: N38 53.27 E46 46.50 Picromerus bidens ** (Linnaeus, 1758) Material examined: 2 specimens, Varzeghan, Mardanaghom, 1020 m. GPS coordination: N38 51.40 E46 49.46 Note: This species is predator of Lymantria dispar larvae in the forest areas of Iran. Dolycoris baccarum (Linnaeus, 1758) Material examined: 15 specimens, Khoda Afarin, Garmanab, 1300 m. GPS coordination: N38 55.66 E46 46.82
Family Scutelleridae Leach, 1815 (Shield-Backed Bugs) Graphosoma italicum (Linnaeus, 1758) Material examined: 18 specimens, Khoda Afarin, Kharil, 1350 m. GPS coordination: N38 48.37 E46 46.16 Eurygaster integriceps Puton, 1881 Material examined: 3 specimens, Kaleibar, Makidi, 1550 m. GPS coordination: N 38 48.37 E46 51.58 Ancyrosoma leucogrammes (Gmelin, 1790) Material examined: 2 specimens, Horand, Tambakulo village, 1850 m. GPS coordination: N38 56.053 E46 49.30
Family Stenocephalidae Dallas, 1852 (Spurgebugs) Dicranocephalus medius** (Mulsant et Rey, 1870) Material examined: 3 specimens, Horand, Khankandi village, 1400 m. GPS coordination: N38 531.40 E46 37.410 Dicranocephalus agilis** (Scopoli, 1763) Material examined: 4 specimens, Khoda Afarin, Tazehkand, 1300 m. GPS coordination: N38 54.39 E46 48.06
Family Corixidae Leach, 1815 (Water Boatmen) Trichocorixa* sp. Material examined: 2 specimens, Khoda Afarin, Ashegloo village, Aras river, 450 m. GPS coordination: N38 57.282 E45 41.173 Hesperocorixa* scabricula** (Walley, 1936) Material examined: 2 specimens, Varzeghan, Khoinarood, 1750m. GPS coordination: N38 71.02 E46 76.33
Family Gerridae Leach, 1815 (Water Striders) Aquarius* (=Gerris) lacustris** (Linnaeus 1758) Material examined: 3 specimens, Varzeghan, Haft Cheshmeh, 2120. GPS coordination: N38 55.20 E 46 46.45
ACKNOWLEDGEMENTS
We are indebted to Eng. B. Sadaghian and Eng. M.H. Pezeshki for them helping in field works and preparing specimnces for identification.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 395
LITERATURE CITED Askari, O., Pourabad, R. F. & Khaganinia, S. 2009. Faunistic study of Heteroptera of Zanjanroud region in Zanjan province of Iran. Munis Entomology & Zoology, 4 (2): 560-563. Forero, D. 2006. New Heteroptera (insecta: Hemiptera) records from Colombia. Caldasia, 28 (1): 125-128. Gullan, P. J. & Cranston, P. S. 2005. The Insects: An Outline of Entomology (third edition). Blackwell Publishing, 505 pp. Hassazadeh, M., Pourabad, R. F., Gharaat, M. A. & Beykpor, A. R. 2009. A study on the Heteroptera fauna of Shend Abad region and environ (Iran). Munis Entomology & Zoology, 4 (2): 527-530. Khaghaninia, S., Pour Abad, R. F. & Askari, O. 2010. First record for seven species of leaf bugs (Hemiptera, Miridae) for Iran insect fauna from East Azarbayjan province. Munis Entomology & Zoology, 5 (1): 309-310. Linnavuori, R. E. & Modarres Awal, M. 1998. Studies on the Heteroptera of the Khorasan province in N.E. Iran. Nepomorpha, Gerromorpha, Cimicomorpha (Nabidae, Anthocoridae), and Pentatomorpha (Coreidae). Entomology Fennica, 9: 237-241. Linnavuori, R. E. 1997.Taxonomic Studies on Miridae (Heteroptera) of Yemen and Iran. Acta Universiatis Carolinae Biologica, 40 (1996): 301-320. Modarres Awal, M. 2002. List of agricultural pests and their natural enemies in Iran. III. Ed, Univ. Ferdowsi, Mashhad, 429 pp. Safavi, M. 1991. Key to the Iranian hemipters (15), Family Nepidae. Journal of Entomological Society of Iran, 11: 45-49. Safavi, M. 1993. Key to the Iranian hemipters (16), Family Acanthosomatidae. Journal of Entomological Society of Iran, 12, 13: 105-107. Sadaghian, B., Dordaei, A. A. & Nikdel, M. 2004. An investigation on some Heteroptera in arasbaran forests. 16th Iranian plant protection congress, 1: 128. Slater, J. A. 1981. How to Know the True Bugs (Hemiptera-Heteroptera) William Brown Publishers, Iowa. Yarmand H., Sadeghi, S. E., Asgari, H., Mehrabi, A. & Matocq, A. 2004. Diversity of some Miridae (Heteroptera) species associated with forests and rangelands of Iran, 16th Iranian plant protection congress, 1: 154.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 396
IMPACT OF MOBILE PHONES ON THE DENSITY OF HONEY BEES
S. Sainudeen Sahib*
* PG & Research Dept. of Zoology, S. N. College, Kollam-691001, Kerala, INDIA. E-mail: [email protected] [Sainudeen Sahib, S. 2011. Impact of mobile phone on the density of Honey Bees. Munis Entomology & Zoology, 6 (1): 396-399] ABSTRACT: Apiculture has developed in to an important industry in India as honey and bee-wax have become common products. Recently a sharp decline in population of honey bees has been observed in Kerala. Although the bees are susceptible to diseases and attacked by natural enemies like wasps, ants and wax moth, constant vigilance on the part of the bee keepers can overcome these adverse conditions. The present plunge in population (< 0.01) was not due to these reasons. It was caused by man due to unscientific proliferation of towers and mobile phones. KEY WORDS: Electromagnetic radiation, Apiculture, Colony collapse disorder.
Bees and other insects have survived and evolved complex immune system on this planet over a span of millions of years. It is not logical that they would now suddenly die out now due to diseases and natural parasites. This suggests another factor has been introduced to their environment that disrupts their immune system. This man made factor is the mobile towers and mobile phones.
The public is not being informed of the threat due to deliberate attempts on the part of mobile phone makers to mask the direct causal relationship. Over the past several months a cadre of scientists, funded by the deep pockets of the mobile phone industry, has suggested viruses, bacteria, and pesticides are to blame for the unprecedented honey bee decline. Rather than critically assessing the problem, the industry is dealing with it as a politics and public relation problems thus manipulating perception of the appropriate remedy. Sadly, this deceptive practice is business as usual for the mobile phone industry.
If the reason behind the population decrease were biological or chemical there would be a pattern of epidemic spread. Observers would be able to trace the spread of bee disappearance from a source similar to the spread of SARS a few years ago. This pattern did not occur, however mobile towers and mobile phones meet the criterion.
New experiments suggest a strong correlation between population decline and cellular equipment. The massive amount of radiation produced by towers and mobile phones is actually frying the navigational skills of the honey bees and preventing them from returning back to their hives. The thriving hives suddenly left with only queens, eggs and hive bound immature worker bees. Thus electromagnetic radiation exposure provides a better explanation for Colony Collapse Disorder (CCD) than other theories. The path of CCD in India has followed the rapid development of cell phone towers, which cause atmospheric electromagnetic radiation.
Insects and other small animals would naturally be the first to obviously be affected by this increase in ambient radiation since naturally they have smaller bodies and hence less flesh to be penetrated by exposure to microwaves. The behavioral pattern of bees alters when they are in close proximity to mobile phones and towers. The vanished bees are never found, but thought to die singly far from home. Bee keepers told that several hives have been abruptly abandoned.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 397
If towers and mobile phones increase the honey bees might be wiped out in ten years. Radiation of 900 MHz is highly bioactive, causing significant alternation in the physiological function of living organisms 7.
MATERIALS AND METHODS
Six colonies of honeybees (Apis mellifera) were selected. Three colonies were selected as test colonies (T1, T2 & T3) and the rest were as control (C1, C2 & C3). The test colonies were provided with mobile phones in working conditions with frequency of 900 MHz for 10 minutes for a short period of ten days. EMF (Electromotive field) power density was measured with the help of RF Power density meter. The control colonies had not provided with mobile phones. Queen prolificacy was calculated in terms of egg laying rate of the queen 1. Flight activity and returning ability were measured as number of worker bees leaving and returning respectively to the hives per minute: before exposure, during exposure and after exposure.
RESULTS
The results of the studies are presented in Table. The present study showed
that after ten days the worker bees never returned hives in the test colonies. The massive amount of radiation produced by mobile phones and towers is actually frying the navigational skills of the honey bees and preventing them from returning back to their hives 1-4. It was shown that the total bee strength was significantly higher in the control colonies being nine comb frames as compared to one in the test colony at the end of the experiment. The thriving hives suddenly left with only queens, eggs and hive bound immature worker bees. The queens in the test colonies produced fewer eggs/day (100) compared to the control (350). It has previously been reported that there is low egg laying rate in queens exposed to high voltage transmission lines 5 or exposure of the queen bees to cell phone radiation stimulated her to produce only drones 6. Thus electromagnetic radiation (EMR) exposure provides a better explanation for Colony Collapse Disorder (CCD) than other theories. The path of CCD in India has followed the rapid development of cell phone towers and cell phones, which cause atmospheric electromagnetic radiation.
DISCUSSION
Some countries have sought to limit the proliferation of mobile towers with strict rules. But in India no such rules have been formulated or implemented. Given the proliferation of mobile phone towers and their vital role in communications, solutions to the problem will not be as simple as eliminating the towers. One possibility is shielding the bee hives with EMR resistant materials.
Another solution would be granting local communities the ability to control whether or not to install mobile towers. On one hand, community members would be able to exert some control over their environment and determine whether the benefits outweigh the costs and risks. On the other, it is highly susceptible to manipulation by powerful influences, especially since the bee keepers have significantly less influence, power and wealth than the mobile phone companies.
However, Indians could risk losing even this right to self determination if the cellular providers can impose a country wide mandate prohibiting regulation against them, similar to the Telecommunications Act of 1996 in the United States.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 398
The Act prohibited local governments from making sitting decisions based on the perceived health impacts of wireless facilities. Indian advocates are concerned that such regulations might be upheld in India as they were in the United States in order to ―eliminate service gapes in its cellular telephone service area.‖
In Kerala there are about 600,000 beehives and over 100,000 workers are engaged in Apiculture. A single hive may yield 4-5 kg of honey. Moreover, the destruction of bee hives could be a major environmental disaster. Honeybees are responsible for pollinating over 100 commonly eaten fruit and vegetable crops and without bees the food system would be in serious trouble. Rural village dependent on locally grown foods would be most vulnerable. The need of the hour is to check unscientific proliferations of mobile phone towers. More research is essential on how to protect the bee hives from the electromagnetic exposure, but perhaps more to study the impacts on humans.
All mobile phone towers emit microwave radiations, which is in the radio frequency radiation (RFR), part of the spectrum of electromagnetic waves. Though RFR, like Ultra-violet (UV) and Infra-red light, is a source of non-ionizing radiation, these radiations, together with ionizing electromagnetic radiations such as X- rays, gamma rays make up the electromagnetic spectrum. Radio frequency of the electromagnetic waves ranged from 100 kilo hertz (KHz) to 300 Giga hertz (GHz). Radio frequency radiation is a source of thermal energy and in adequate doses, has all the known effects of heating on biological systems 7.
Despite a growing number of warnings from scientists, like me, the Government has done nothing to protect people and the environment. Steps must be taken to control the installations of mobile phone towers by imposing restrictions. Installation of towers should be regulated near thickly populated areas, educational institutions, hospitals etc. Sharing of towers by different companies should be encouraged, if not mandated. To prevent overlapping high radiations fields, new towers should not be permitted within a radius of one kilometer of existing towers.
More must also be done to compensate individuals and communities put at risk. Insurance covering diseases related to towers, such as cancer, should be provided for free to people living in 1 km radius around the tower. Independent monitoring of radiation levels and overall health of the community and nature surrounding towers is necessary to identify hazards early. Communities need to be given the opportunity to reject cell towers and national governments need to consider ways of growing their cellular networks without constantly exposing people to radiation.
LITERATURE CITED
Aday, W. R. 1975. ―Introduction: Effects of electromagnetic radiation on the nervous system‖. Annals NY Acad. Sci., 247: 15-20. Brandes, C. & Frish, B. 1986. Production of mutant drones by treatment of honeybees with X-rays. Apidologie, 17 (4): 356-358. Greenberg, B., Bindokas, V. P. & Gauger, J. R. 1981. Biological effects of a 765 kV transmission line: exposure and thresholds in honeybee colony. Bioelectromagnetics, 2 (4): 315-328. Sharma, P. L. 1958. Brood rearing activity of Apis indica F. and egg laying capacity of its queen. Indian Bee J., 20: 166-173. Warnke, U. 1975. Bienen unter Hochspannung (Bees under high voltage). Umschau., 13: 416-417. Warnke, U. 1976. Effect of electrical charges on honey bees. Bee World, 57 (2): 50-56.
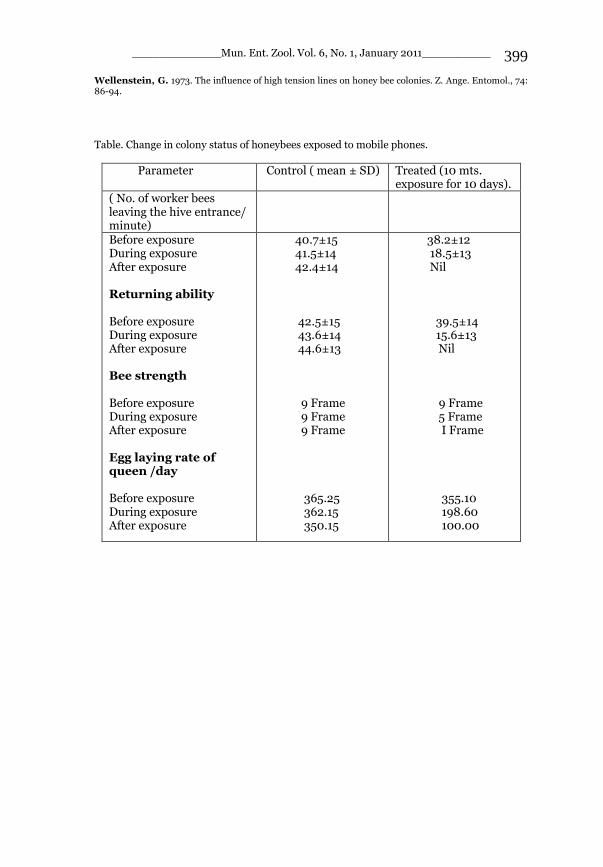
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 399
Wellenstein, G. 1973. The influence of high tension lines on honey bee colonies. Z. Ange. Entomol., 74: 86-94.
Table. Change in colony status of honeybees exposed to mobile phones.
Parameter Control ( mean ± SD) Treated (10 mts. exposure for 10 days).
( No. of worker bees leaving the hive entrance/ minute)
Before exposure During exposure After exposure Returning ability Before exposure During exposure After exposure Bee strength Before exposure During exposure After exposure Egg laying rate of queen /day Before exposure During exposure After exposure
40.7±15 41.5±14 42.4±14 42.5±15 43.6±14 44.6±13 9 Frame 9 Frame 9 Frame 365.25 362.15 350.15
38.2±12 18.5±13 Nil 39.5±14 15.6±13 Nil 9 Frame 5 Frame I Frame 355.10 198.60 100.00

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 400
SESAMIA BOTANEPHAGA TAMS & BOWDEN, 1953 AUCT. IN IRAN, READ SESAMIA NONAGRIOIDES (LEFÈBVRE, 1827)
(LEPIDOPTERA: NOCTUIDAE)
Mehdi Esfandiari*, Mohammad Saeed Mossadegh and Parviz Shishehbor
* Dept. of Plant Protection, College of Agriculture, Shahid Chamran University, Ahvaz, IRAN. E-mail: [email protected] [Esfandiari, M., Mossadegh, M. S. & Shishehbor, P. 2011. Sesamia botanephaga Tams & Bowden, 1953 auct. in Iran, read Sesamia nonagrioides (Lefèbvre, 1827) (Lepidoptera: Noctuidae). Munis Entomology & Zoology, 6 (1): 400-403] ABSTRACT: Sesamia botanephaga Tams & Bowden, 1953 was reported in Iran, for the first time, from sugarcane fields of Khuzestan province, SW Iran. The accuracy of this report was dubitable, because S. botanephaga is a native species to Africa. In this investigation, samples were taken from agricultural fields of Khuzestan and Fars provinces in 2007. Taxonomic study revealed that all samples were Sesamia nonagrioides (Lefèbvre, 1827); thus African S. botanephaga (or S. nonagrioides botanephaga) does not occur in Iran. KEY WORDS: Sesamia botanephaga, Sesamia nonagrioides botanephaga, Sesamia nonagrioides, sugarcane stem borer, Iran.
Apart from Sesamia cretica Lederer, another Sesamia species occurs in SW Iran, as a stem borer pest. For the first time, Ali Pazuki from Iranian Research Institute of Plant Protection identified this stem borer as Sesamia botanephaga Tams & Bowden, 1953 which was collected from sugarcane fields of Haft Tappeh in Khuzestan province, SW Iran (Daniali, 1985). Other authors have followed this identification according to Pazuki (see e.g. Daniali, 1986; Jemsi & Kamali, 1992; Fazeli, 1992; Ranjbar Aghdam, 1999; Mossadegh & Kocheili, 2003). However, as stated by Allsopp & Sallam (2001), the accuracy of reports on S. botanephaga from Iran was dubitable. It should be noted that Nye (1960) sank this species described by Tams & Bowden to the rank of subspecies: Sesamia nonagrioides botanephaga.
The above mentioned stem borer is an important pest of sugarcane, maize and rice in Khuzestan province and has 4-5 annual generations. It attacks several cultivated and non-cultivated graminaceous species (Daniali, 1985; Jemsi & Kamali, 1992; Mossadegh & Kocheili, 2003). It also damages maize and rice in some parts of Fars province, where it has 3-4 annual generations (Fazeli, 1992). A geometric morphometrics study on S. nonagrioides populations in SW Iran by Esfandiari et al. (2010) revealed that the centroid size of both internal and external genitalia in population from rice farms was significantly smaller than the populations from maize and sugarcane farms. This was probably due to smaller stem diameter of rice, as an environmental factor, compared to maize and sugarcane stems.
MATERIAL AND METHODS
Population samples were collected in South and Southwest Iran in 2007: In Khuzestan province: on sugarcane in the following agro-industries:
Amirkabir (31º03´N 48º14´E), Farabi (30º06´N 48º36´E), Imam Khomeini (31º46´N 48º44´E) and Karoon (32º10´N 48º36´E). On maize in the cities of Shushtar (32º07´N 48º44´E) and Behbahan (30º38´N 50º12´E).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 401
In Fars province: on rice in Firoozabad city (29º00´N 52º30´E) and on maize in Noorabad city (30º05´N 51º30´E).
The infested plants were collected and the larvae were reared to adulthood at the laboratory. Adult specimens were checked according to their genitalia structures.
The specimens were deposited in Insect and Mite Collection of Ahvaz (IMCA), Plant Protection Department, Shahid Chamran University of Ahvaz, Iran.
RESULTS
Taxonomic study of genitalia structures of the collected specimens revealed that the discussed stem borer is the same as the Mediterranean corn borer, Sesamia nonagrioides (Lefèbvre, 1827), which occurs in Mediterranean countries. Diagnosis. Male antenna long bipectinate with feather-like lamellae, female antenna ciliate. Head, thorax and ground colour of forewing from pale ochreous and beige to light brown. Straw-coloured forewing has a dark terminal area and sometimes with a few black dots; stigmata absent; Fringe white. Hind wing whitish with a light brown terminal line. Male genitalia (Fig. 1): Uncus small, narrowing at about halfway, elongated valvae with narrower membranous cucullus and thorn like acute costal extension which has a small tooth before apex, sacculus slightly broadened at apex with rows of short stout spines, basal half of valval costa heavily sclerotised, concave and broad; long and narrow juxta without median process, manica without spine; aedeagus short with long caecum, strongly sclerotised in the tip, carina with small spines, vesica with prominent subbasal diverticulum and cristate sclerotised subbasal plate. Female genitalia (Fig. 2): Ovipositor lobes conical, elongated and acute, ductus bursa long and narrow with little chitinized plate near ostium, corpus bursa narrow.
S. cretica, which also occurs in Southwestern Iran, is sometimes very similar externally to S. nonagrioides, but the males can be separated by the much shorter pectination of the antenna in S. cretica. Precise identification of females requires dissection of the genitalia. In S. cretica corpus bursa is broad and ductus bursa is short and broad without chitinized zones. S. cretica larvae are distinguishable from those of S. nonagrioides by the angle between the setae D2, D1 and SD1 on the ninth abdominal segment, which is less than 120 degrees in S. cretica, while in S. nonagrioides are almost in line.
DISCUSSION
Some sugarcane cuttings were imported from Egypt to Iran about 1937 (Anonymous, 1976). Pazuki believes that the mentioned stem borer is the African S. botanephaga, because it was most probably imported from Egypt to Iran by the imported sugarcane cuttings (Personal communication).
We can not be agreeing with Pazuki, because S. botanephaga occurs in West, Central and East Africa, but not in Egypt (Meijerman & Ulenberg, 1996;

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 402
Holloway, 1998; Allsopp & Sallam, 2001). Typical S. nonagrioides occurs in Iran, as stated by Holloway (1998), thus the record of S. botanephaga for Iran should be deleted.
It seems that S. nonagrioides is native to SW Iran rather than an exotic pest and have adopted sugarcane as a host consequent to its cultivation in this region since about 70 years ago. Most of recorded host plant species of S. nonagrioides (Rao & Nagaraja, 1969) occur in SW Iran. Furthermore, Daniali (1985) reported an egg parasitoid of S. nonagrioides, from SW Iran, which nowadays is effective in biological control of this pest. It probably indicates the long-lasting relationship between the pest and its parasitoid in SW Iran.
ACKNOWLEDGEMENTS
We thank Dr. Laszlo Ronkay (Hungarian Natural History Museum, Budapest, Hungary), Dr. Pascal Moyal (IRD/CNRS, France) and Eng. Ali Pazuki (Iranian Research Institute of Plant Protection, Tehran, Iran) for useful discussions. This work was supported by the grant from Shahid Chamran University of Ahvaz.
LITERATURE CITED Allsopp, P. G. & Sallam, M. S. 2001. Sesamia incursion management plan, Ver.1. BSES Publication, Project Report PR01002: 95 pp. Anonymous. 1976. An introduction to sugarcane cultivation and activities of Haft Tappeh agro-industry. Ministry of Agriculture and Natural resources of Iran, Haft Tappeh agro-industry, 36 pp. Daniali, M. 1985. Effect of biological, cultural and chemical control measures against sugarcane stem borers Sesamia spp. (Lep. Noctuidae) in Haft Tappeh, Khuzestan. MSc thesis, Shahid Chamran University, Ahvaz, Iran, 114 pp. Daniali, M. 1986. Role of cultural practices in control of the sugarcane stem borer in the region of Haft Tappeh, Khuzestan. Entomologie et Phytopathologie Appliquees, 53 (1-2): 3-4. Esfandiari, M., Mossadegh, M. S., Shishehbor, P. & Hodjat, S. H. 2010. Male genital size variation in Sesamia nonagrioides (Lef.) populations of south and southwest Iran. 19th Iranian Plant Protection Congress, 31 Jul.- 3 Aug., Tehran, Iran. p. 114. Fazeli, M. J. 1992. Biology and control of Sesamia nonagrioides botanephaga in the Fars province. Applied Entomology and Phytopathology, 59 (1&2): 13-14. Holloway, J. D. 1998. Noctuidae: introduction. In: A. Polaszek (Eds.), African cereal stem borers, economic importance, taxonomy, natural enemies and control, CAB International, Wallingford, pp. 79-86. Jemsi, Gh. & Kamali, K. 1992. Biology, host range and damage of corn stem borer in Khuzestan. Applied Entomology and Phytopathology, 59 (1&2): 31-33. Meijerman, L. & Ulenberg, S. A. 1996. Identification of African stem borer larvae (Lepidoptera: Noctuidae, Pyralidae) based on morphology. Bulletin of Entomological Research, 86: 567-578. Mossadegh, M. S. & Kocheili, F. 2003. A semi descriptive checklist of identified species of arthropods (agricultural, medical, …) and other pests from Khuzestan, Iran. Shahid Chamran University Press. 475 pp. Nye, I. W. B. 1960. The insect pests of graminaceous crops in East Africa. Her Majesty‘s Stationery Office, London. 42 pp. Ranjbar Aghdam, H. 1999. Possibility of in vivo rearing of Platytelenomus hylas Nixon in pink stem borers Sesamia spp. biocontrol. MSc thesis, Shahid Chamran University, Ahvaz, Iran, 116 pp. Rao, V. P. & Nagaraja, H. 1969. Sesamia species as pests of sugarcane. In: J.R. Williams, J.R. Metcalfe, R.W. Mungomery & R. Mathes (Eds.), Pests of Sugar Cane. Elsevier, Amsterdam, 208-223 pp.
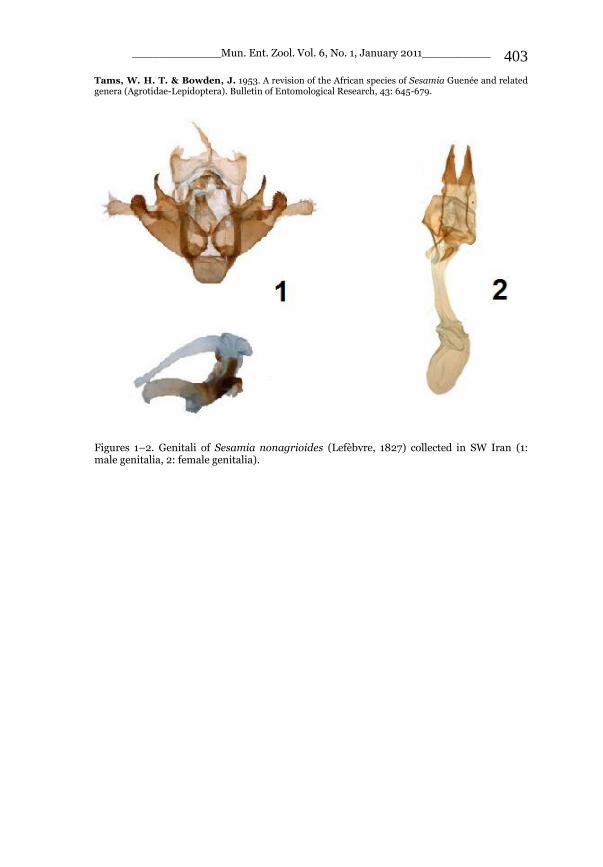
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 403
Tams, W. H. T. & Bowden, J. 1953. A revision of the African species of Sesamia Guenée and related genera (Agrotidae-Lepidoptera). Bulletin of Entomological Research, 43: 645-679.
Figures 1–2. Genitali of Sesamia nonagrioides (Lefèbvre, 1827) collected in SW Iran (1: male genitalia, 2: female genitalia).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 404
PCR-RFLP PATTERN ANALYSIS OF ENTOMOPATHOGENIC NEMATODES ISOLATED FROM AGRO-ECOSYSTEM
FOR IMPLICATING THEIR GENETIC DIVERSITY
M. Razia*, K. Padmanaban*, R. Karthik Raja*, P. Chellapandi** and S. Sivaramakrishnan*
* Department of Biotechnology, School of Life Sciences Bharathidasan University, Tiruchirappalli - 620 024, Tamil Nadu, INDIA. E-mail: [email protected] ** Department of Bioinformatics, School of Life Sciences Bharathidasan University, Tiruchirappalli - 620 024, Tamil Nadu, INDIA. [Razia, M., Padmanaban, K., Karthik Raja, R., Chellapandi, P. & Sivaramakrishnan, S. 2011. PCR-RFLP pattern analysis of entomopathogenic nematodes isolated from agro-ecosystem for implicating their genetic diversity.Munis Entomology & Zoology, 6 (1): 404-411] ABSTRACT: Entomopathogenic nematodes (EPNs) (Rhabditida: Steinernematidae and Heterorhabditidae) were isolated from cultivated lands in Karur region of Tamil Nadu, India. The ITS (internal transcribed spacer) rDNA sequence similarity of isolated EPNs revealed that the nematode strains Steinernema siamkayai, S. pakistanense and Heterorhabditis indica were more prevalent in cultivated eco-systems of that region. PCR-RFLP patterns obtained from 10 different restriction enzymes have shown that these strains were phylogenetically are more or less closely related within species, but they are genetically diverse from location to location in that region. Among 25 nematode strains, 19 strains were phylogenetically related to S. siamkayai (strain CM2) isolated from Belgium. Three strains of Steinernema sp. were related to S. pakistanense strain isolated from USA and remaining three Heterorhabdus sp. similar to H. indica strain cohen 21 from USA. Thus, intra-specific relationships among strains of these nematodes would emphasis that geographic distribution and environmental conditions may be associated to bring such genetic variations and divergence in the genomes of the nematodes. KEY WORDS: Steinernema, Heterorhabdus, Cultivated lands, ITS region, PCR-RFLP, Genetic variation.
Entomopathogenic nematodes (EPNs) belonging to the genera Heterorhabditis carry symbiotic of Photorhabdus luminescens with different subspecies and Steinernema carry symbiotic of Xenorhabdus spp. possess bioinsecticides, antimicrobial and antiviral activities are the curious potential characters. However, genetic variations in these nematodes are one of the determinants for survival of endosymbiotic and non-symbiotic bacteria in the respective hosts. Different nematode species and strains exhibit differences in survival, infectivity and efficacy against particular insect pest. A phylogenetic sketch is a necessary component of the comparative method in evolutionary biology and provides to study gene flow, population structure, biogeography, co-evolution, co-adaptation, co-speciation, and historical ecology (Hominick et al., 1997). Strengthening biosystematics research is extremely essential for the accurate identification of the species and knowing their genetic diversity, which forms the basis for exploiting the use of vast biodiversity resources, as well as their management.
A number of molecular techniques have been used for EPNs identification, including isoenzyme patterns (Akhurst, 1987), total protein patterns (Poinar & Kozodoi, 1988) and immunological techniques (Jackson, 1965). Restriction fragment length polymorphism (RFLP) detection within total genomic DNA is a reliable taxonomic tool that can be used for the identification of single nematode

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 405
(Curran & Webster, 1989; Smits et al., 1991; Reid & Hominick, 1993). The internal transcribed spacer (ITS) region of the nuclear ribosomal DNA (Hillis et al., 1991; Powers et al., 1997) has already reported to be a consistent marker for identifying species within each genus of EPNs. It could provide the requisite heritable characteristic for a thorough phylogenetic analysis (Vrain et al., 1992; Joyce et al., 1994; Cherry et al., 1997). The advantages and taxonomic suitability of this marker include the potential for PCR amplification and sequencing by universal primers, forced uniformity of paralogues via rapid determined evolution, variation due principally to point mutations, apparent independence of variable sites, and phylogenetic information appropriate for species level investigations (Adams, 1998). Moreover, molecular approaches can be enhancement by morphological, morphometric and cross breeding techniques to confirm the identification of reputed new species (Hominick et al., 1997).
The genetic differentiation among species is the result of the interaction of random chance, migration, and natural selection. Various field isolates are often different from one another at different degrees. Therefore, the present work was aimed to identify the genetic differences of EPN strains isolated from different locations of cultivated lands in Karur region of Tamil Nadu, India using PCR-RFLP profiling.
MATERIALS AND METHODS EPN populations
A total of 25 populations of the Heterorhabditis and Steinernema species were used for this study (Table 1). The samples were obtained from cultivated lands of Karur region of Tamil Nadu, India. EPNs were isolated from Galleria mellonella trapping method according to Bedding and Akhurst (1975). Each isolate was given a code as follows KAR (for Karur) followed by serial number. DNA extraction and PCR amplification of the ITS-rDNA
Genomic DNA was extracted by grinding the IJs in a mortar and pester with liquid nitrogen until a fine white powder was produced. The powder was collected and 1 ml of extraction buffer (10 mM Tris-Cl pH = 8, 1% SDS, 0.4 M NaCl, 5 mM EDTA) was added. Then 50 µl proteinase K solution (0.01 g/ml) was added and the mixture was incubated for 30 min at 37 ºC and subsequently for 60 min at 65 ºC. The solution was twice extracted with equal volumes phenol-chloroform- isoamyl alcohol (25:24:1) and finally with chloroform- isoamyl alcohol (24:l). DNA was precipitated from the final aqueous phase by adding 2 volumes of 96 % ethanol and 0.1 volume of 3M ammonium acetate and placing the mixture at - 70ºC for 30 min. The precipitated DNA was centrifuged in a micro centrifuge and dried under vacuum at room temperature. The pellet was resuspended in 50 µl of TE-buffer (0.01 M Tris-Cl pH - 7.5, 0.001 M EDTA) and stored at 4ºC. The nucleic acid concentration was measured in a spectrophotometer at 260 nm. The DNA sample was run by agarose gel electrophoresis using 1 kb markers of lambda DNA as a standard.
The ITS region of rDNA sequence in each strain was amplified by PCR (polymerase chain reaction) and the primers used this study were designed according to Joyce et al. (1994b). PCR reaction were carried out, 100ng Genomic DNA, 1µl of dNTP mix (2.5mM each), Forward primer and Reverse Primer 10 pmole, 5 µl of Taq buffer (10X), 3U of Taq DNA polymerase enzyme and nuclease

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 406
free water was added make up the volume 5µl. PCR running condition was set as 35 cycles of 94°C for 1 min, 55°C for 1min, and 72°C for 2 min. PCR-RFLP pattern analysis
PCR products (5μl) of every isolates were digested with 0.5µl of restriction endonucleases PstI, PvuII, Sau3AI, SalI, AluI, HpaII, HhaI, HindIII, HinfI and XhaI according to the manufacturer‘s instructions with the total volume of 20µl. The digested products were run on a 2% agarose gel, at 100V for 2 hours and photographed using GeneTech gel documentation. A genetic relationship between each pair of strains was valuated as presence or absence of DNA restriction fragment.
RESULTS
The samples collected from different cultivated lands showed positive for the presence of the EPNs (25 strains) of the genus Steinernema and Heterorhabditis. The PCR products of each strain has composed of a part of the ITS region and a part of 18S, rDNA gene and they were yielded a single fragment with a length of approximately 776-784 bp. Blast similarity search results showed that ribosomal DNA (ITS region of nematodes) sequences of each isolate have a maximum sequence similarity with closely related with the sequences of EPNs belonged to the same genera. It reflected to confirm these isolates were belonged to Steinernema and Heterorhabditis sp. Phylogenetic bootstrapping values of scaled in the phylogenetic tree was supported the correspondence of every isolates with their species and promising their distinctiveness as separate species (data not shown). As the results of ITS rDNA sequence similarities and phylogenetic inference, 19 isolates were grouped under the members of S. siamkayai, the 3 isolates were the members of S. pakistanense and the remaining 3 Heterorhabditis isolates were belonged to H. indica. PCR-RFLP pattern analysis
The PCR-RFLP profiles of the isolated EPNs strains were performed with 10 restriction enzymes. The restriction patterns obtained with PstI; PvuII; Sau3AI; SalI; AluI; HpaII; HhaI; HindIII; HinfI and XhaI are represented in Table 2 and Figure 1. The counting numbers of bands generated from each restriction enzyme digestion (PCR-RFLP) was chosen for genetic variation analysis in which more distinct band and numbers of RFLP fragments indicated strain specific variations in the ITS sequences of strains isolated from this region. The genetic variants of these strains isolated from different location of Karur region so that this criterion would be more appropriate to distinguish the strains. PCR-RFLP variant analysis of each strain revealed a little or no discernible intra- specific variability was observed within the species of Steinernema. The isolate S. siamkayai isolated from Krishnarayapuram, Kulithalai and Aravakurichi locations was same strains, while isolate S. pakistanense was existed in Kulithalai and Karur similarly. The strain H. indica was found in Krishnarayapuram and Karur are closely resembles. Although these strains have been identified by morphology and genetic variants in ITS regions of rDNA, they have some genetic polymorphic differences in these genomes. Thus, a polymorphically distinct character of these strains was analyzed by RFLP fragments.
Sau3AI and HhaI digested fragments showed that the strains KAR01, KAR04 and KAR05 have the same genetic polymorphism in ITS sequences. More number of fragments has been generated by restriction enzymes PstI, AluI and XhaI from

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 407
the strains KAR01 and KAR05 as shown in Table 2. SalI digested fragments of ITS sequences were highly varied from strains to strains indicated its genetic variability. Strain KAR04 has a low number of RFLP fragments and same genetic polymorphisms were found in the fragments generated by Sau3AI and HhaI. However, XhaI digested of KAR36 and KAR518 fragments showed a high variability. Other RFLP fragments of ITS sequences of H. indica were highly varied even those strains isolated from same locality of Karur region.
Apart from the genetic variations of these strains, the highly homologous genetic polymorphism in S. siamkayai have been detected in the RFLP fragments resulted from the digestion of Sau3A (KAR12, KAR140, KAR723, KAR240 and KAR571), HhaI (KAR12, KAR140, KAR723, KAR240, KAR571 and KAR06), AluI (KAR12, KAR140, KAR723 and KAR240), HindIII (KAR723, KAR240, KAR571 and KAR06), Hinf I (KAR723, KAR240 and KAR571). The fragments of SalI and HhaI showed the strains specific similarity between KAR 723 and KAR 240, respectively. All the strains significantly varied in the fragments of PstI and HhaI whereas S. pakistanense and H. indica have also genetically differed in the fragments of PstI, HhaI, HpaII and XhaI. A high possibility of polymorphic variation detected in S. siamkayai was in the strain KAR06. Results of RFLP pattern analysis revealed that the nematode H. indica, S. siamkayai and S. pakistanense were highly variable in their genetic diversity that was dependent on the location of isolation.
DISCUSSION
In the present investigation the geographical origin and habitat can influence the morphometric characteristics of the isolates can change respective to those provided in the bibliography based on abiotic factors and host availability (Stock et al., 2000; Poinar, 1992). These morphometric differences are resulted due to intra-specific variability (Stock et al., 2000; Stock & Reid, 2004). Soil nematodes in general are morphologically very conservative so that the higher level taxonomy in this phylum is hampered by a lack of suitable characters and morphology is unreliable as an indicator of genetic differentiation of species. Systematic of EPNs is still in a state of instability and transition. DNA sequencing has become one of the most utilized molecular approaches for inferring phylogenetic history. In our studies, primers have been used to amplify ITS region, including the partial 18S rDNA gene which provides the insights necessary to develop a strong, morphologically based taxonomy according to Hominick et al. (1996). Thus, phylogenetic analysis revealed that 19 isolates were grouped under S. siamkayai, 3 isolates were S. pakistanense and the remaining 3 isolates were belonged to H. indica.
RFLP analysis of PCR amplified products from specific regions of the genome is a powerful taxonomic tool that can be used for identification of single nematodes. Amplified products of the ITS region have, upon restriction enzyme digestion, yielded many RFLP fragments for several rare strains that can be used for species identification of Steinernema isolates (Reid, 1994). This PCR-RFLP profiling reported the genetic divergence of 19 S. siamkayai, 3 S. pakistanense, and 3 H. indica populations. It resulted that the strains H. indica, S. siamkayai and S. pakistanense were highly variable in the RFLP patterns analyzed from the ITS sequences, which were dependent on the location of isolation. This may be resulted due to intrinsic and extrinsic factors, host, genome dynamics and horizontal gene transfer and evolutionary process. These factors could be influenced to bring such genetic variations among EPNs strains isolated from the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 408
same location. The present attempt is useful for identification of EPNs at species level, and to know the species-specific genetic variation to that region. This will provide a new vision to select a species or strain for bio-control testing for controlling the pest in a selected zone.
ACKNOWLEDGEMENTS
The corresponding author is thankful to Department of Science and Technology-Young Scientist Scheme, (Sanction No. SR/FT/L-133/2005), New Delhi, India, for their financial assistance.
LITERATURE CITED Adam, B. J. 1998. Species concepts and the evolutionary paradigm in modern nematology. J. Nematol., 30: 1-21. Akhurst, R. J. 1987. Use of starch gel electrophoresis in the taxonomy of the genus Heterorhabditis (Nematoda: Heterorhabditidae). Nematologica, 33: 1-9. Bedding, R. A. & Akhurst, R. J. 1975. A simple technique for the detection of insect parasitic rhabditid nematodes in soil. Nematologica, 21: 109-110. Cherry, T., Szalanski, A. L., Todd, T. C. & Powers, T. O. 1997. The internal transcribed spacer region of Belonolaimus (Nemata; Belonolaimidae). J. Nematol., 29: 23-29. Curran, J. & Webster, J. M. 1989. Genotypic analysis of Heterorhabditis isolates from North Carolina. J. Nematol., 21: 140-145. Hills, D. M., Moritz, C., Porter, C. A. & Baker, R. J. 1991. Evidence for biased gene conversion in concerted evolution of ribosomal DNA. Science, 251: 308-310. Hominick, W. M., Briscoe, B. R., del Pino, F. G., Heng, J., Hunt, D. J., Kozodoy, E., Mracek, Z., Nguyen, K. B., Reid, A. P., Spiridonov, S., Stock, P., Sturhan, D., Waturu, C. & Yoshida, M. 1997. Biosystematics of entomopathogenic nematodes: current status, protocols and definitions. J. Helminthol., 71: 271-298. Hominick, W. M., Reid, A. P., Bohan, D. A. & Briscoe, B. R. 1996. Entomopathogenic nematodes: biodiversity, geographical distribution and the Convention on Biological Diversity. Biocont. Sci. Technol., 6: 317-332. Jackson, T. J., Wang, H., Nugent, M. J., Griyn, C. T., Burnell, A. M. & Dowds, B. C. A. 1995. Isolation of insect pathogenic bacteria, Providencia rettgeri, from Heterorhabditis spp. J. Appl. Bacteriol., 78: 237-244. Joyce, S. A., Burnell, A. M. & Powers, T. O. 1994a. Characterization of Heterorhabditis isolates by PCR amplification of segments of mtDNA and rDNA genes. J. Nematol., 26: 260-270. Joyce, S. A., Griffin, C. T. & Burnell, A. M. 1994b. The use of isoelectric focusing and polyacrylamide gel electrophoresis of soluble proteins in the taxonomy of the genus Heterorhabditis (Nematoda, Heterorhabditidae). Nematologica, 40: 601-612. Poinar, G. O. & Kozodai, E. M. 1988. Neoaplectana glaseri and N. anomali: sibling species or parallelism? Revue de Nematologie, 11: 13-19. Poinar, G. O., Karunakar, G. K. & Hastings, D. 1992. Heterorhabditis indicus sp. (Rhabditida: Nematoda) from India: Separation of Heterorhabditis sp. by infective juveniles. Fund. Appl. Nematol., 15: 467-472. Powers, T. O., Todd, T. G., Burnell, A. M., Murray, P. C., Flemming, C. C., Szalanski, A. L., Adams, B. J. & Harris, T. S. 1997. The rDNA internal Transcribed spacer as a taxonomic marker for nematodes. J. Nematol., 4: 441-450.
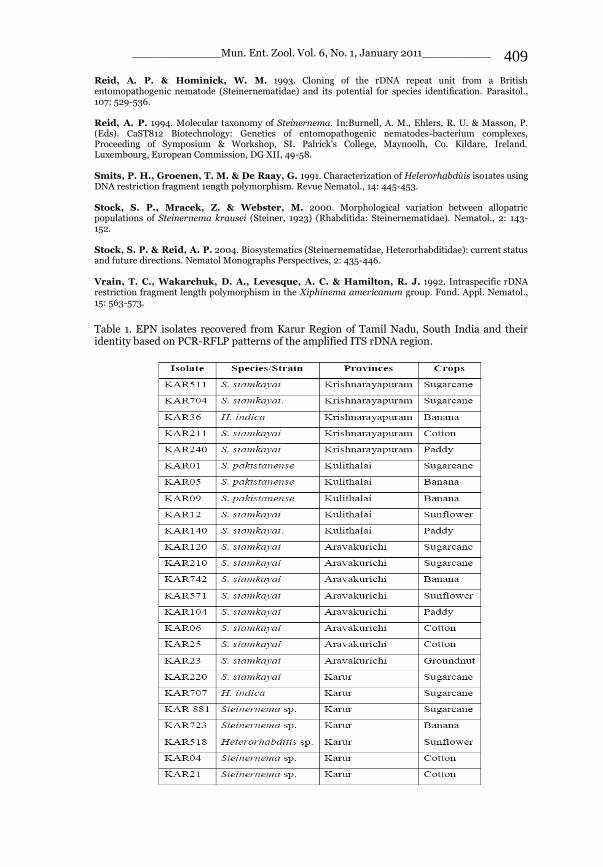
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 409
Reid, A. P. & Hominick, W. M. 1993. Cloning of the rDNA repeat unit from a British entomopathogenic nematode (Steinernematidae) and its potential for species identification. Parasitol., 107: 529-536. Reid, A. P. 1994. Molecular taxonomy of Steinernema. In:Burnell, A. M., Ehlers, R. U. & Masson, P. (Eds). CaST812 Biotechnology: Genetics of entomopathogenic nematodes-bacterium complexes, Proceeding of Symposium & Workshop, SI. Palrick's College, Maynoolh, Co. Kildare, Ireland. Luxembourg, European Commission, DG XII, 49-58. Smits, P. H., Groenen, T. M. & De Raay, G. 1991. Characterization of Helerorhabdùis iso1ates using DNA restriction fragment 1ength polymorphism. Revue Nematol., 14: 445-453. Stock, S. P., Mracek, Z. & Webster, M. 2000. Morphological variation between allopatric populations of Steinernema krausei (Steiner, 1923) (Rhabditida: Steinernematidae). Nematol., 2: 143-152. Stock, S. P. & Reid, A. P. 2004. Biosystematics (Steinernematidae, Heterorhabditidae): current status and future directions. Nematol Monographs Perspectives, 2: 435-446. Vrain, T. C., Wakarchuk, D. A., Levesque, A. C. & Hamilton, R. J. 1992. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fund. Appl. Nematol., 15: 563-573.
Table 1. EPN isolates recovered from Karur Region of Tamil Nadu, South India and their identity based on PCR-RFLP patterns of the amplified ITS rDNA region.
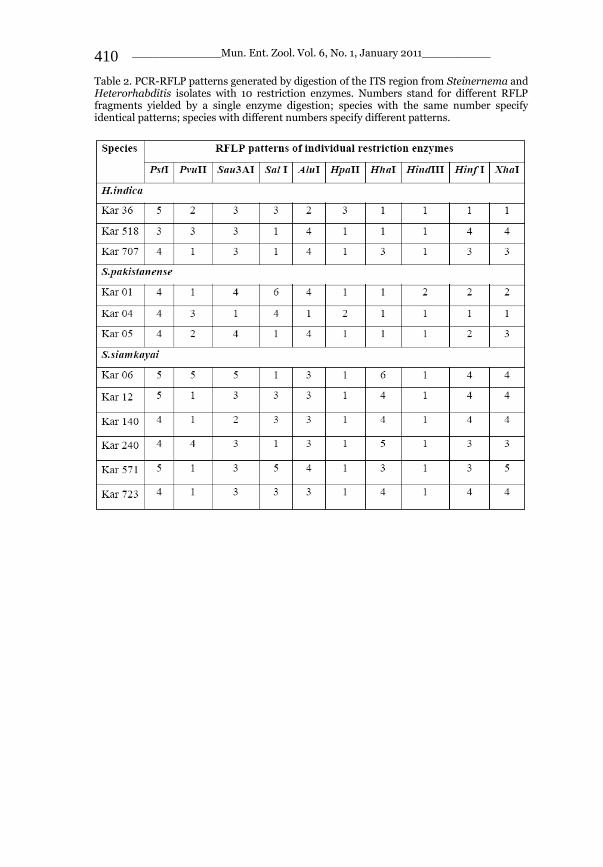
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 410
Table 2. PCR-RFLP patterns generated by digestion of the ITS region from Steinernema and Heterorhabditis isolates with 10 restriction enzymes. Numbers stand for different RFLP fragments yielded by a single enzyme digestion; species with the same number specify identical patterns; species with different numbers specify different patterns.
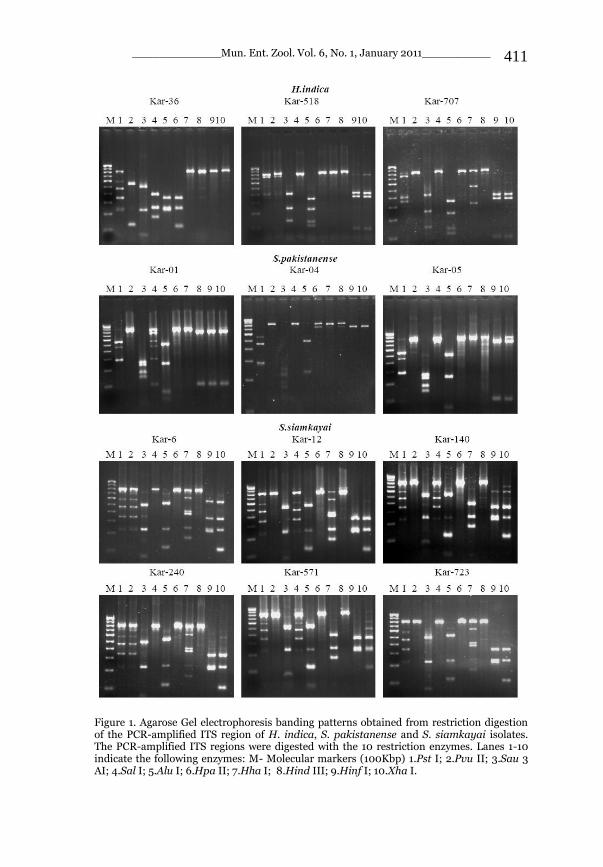
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 411
Figure 1. Agarose Gel electrophoresis banding patterns obtained from restriction digestion of the PCR-amplified ITS region of H. indica, S. pakistanense and S. siamkayai isolates. The PCR-amplified ITS regions were digested with the 10 restriction enzymes. Lanes 1-10 indicate the following enzymes: M- Molecular markers (100Kbp) 1.Pst I; 2.Pvu II; 3.Sau 3 AI; 4.Sal I; 5.Alu I; 6.Hpa II; 7.Hha I; 8.Hind III; 9.Hinf I; 10.Xha I.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 412
EFFECT OF DIET, PHOTOPERIOD AND COLD ON EURYGASTER INTEGRICEPS PUTON DIAPAUSE
(HEMIPTERA: SCUTELLERIDAE)
Azam Amiri*, Ali Reza Bandani and Ali Moslemi-Mehni * Plant Protection Department, College of Agriculture and Natural Resources, University of Tehran, Karaj, IRAN. E-mail: [email protected] [Amiri, A., Bandani, A. R. & Moslemi-Mehni, A. 2011. Effect of diet, photoperiod and cold on Eurygaster integriceps Puton diapause (Hemiptera: Scutelleridae). Munis Entomology & Zoology, 6 (1): 412-421] ABSTRACT: Sunn pest (Eurygaster integriceps Put.) (Hemiptera: Scutelleridae), is a serious pest of cereals causing severe quantitative and qualitative by feeding on leaves, stems and grains. Aim of the current study was to evaluate the effect of photoperiod and diet on the Sunn pest diapause. Thus, the effect of two photoperiod conditions including short day (8 L: 16D) and long day (16L: 8D) and two diet including wheat grains and growing wheat plants on stadium duration, gonad size and haemolymph protein concentration was examined. Also, to extend our studies the effect of cold on gonad size and haemolymph protein concentration was studied. Results showed that stadium duration in short day photoperiod condition (31.43 days) was shorter than long day photoperiod condition (38.58 days) (P<0.05) i.e. when the nymphs were placed in short day conditions and fed on wheat grains their development time (time taken to adult) was shortened about 15–16 days in compare with those nymphs that placed in long day conditions and fed on wheat plants. Short day photoperiod condition and wheat grain diet caused larger ovary size (4.08±0.48 mm2) than long day conditions and wheat plant diet (3.12±0.04 mm2). Cold exposed females and males had lower haemolymph protein concentration than control females and males. The results showed that although photoperiod and diet cause significant changes on the stadium duration, ovaries and testis size and haemolymph electrophoretic bands, their distinct effects on diapause needs to be elaborated more. KEY WORDS: Sunn pest, Diapause, Diet, photoperiod, cold.
Sunn pest (Eurygaster integriceps Put.) (Hemiptera: Scutelleridae), is a serious pest of cereals in the wide area of the globe from Near and Middle East to East and South Europe and North Africa (Radjabi, 2000). The insect causes severe quantitative and qualitative (destruction of gluten protein) damage to crops (sometime up to 100%) by feeding on leaves, stems and grains.
E. integriceps has a monovoltine life cycle, with diapause occurring at the adult stage. The life cycle develops in two different phases in which growth and development take place in wheat, whereas diapause (aestivation and hibernation) occurs in a different habitat, such as oak-forest litter in Europe, or in bushes of Artemisia spp. or Astragalus spp. in the mountains of Asia (Paulian & Popov, 1980). The long diapause period is divided into two distinct steps, aestivation and hibernation (Paulian & Popov, 1980; Radjabi, 2000). So it can be said that the insect is about nine months in diapausing state and it is active only in spring for about three months.
Diapause is an adaptive mechanism for dormancy during periods of unfavorable environmental conditions (Tauber et al., 1986). Diapause is defined as physiologically controlled suppression of growth, development or reproduction. Diapause and non-diapause individuals differ in developmental, physiological, morphological and behavioral characteristics (Tauber et al., 1986; Danks, 1987). One of the most important features of adult diapause is a cessation of reproduction. Thus, when insects have adult diapauses, vitellogenesis does not

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 413
take place after emergence (Danks, 1987). As a result immature ovaries have been used as the criterion of adult diapauses and in males which experience the diapause the testes are reduced in size, but in other cases the testes remain well developed (Denlinger, 2000; Numata & Hidaka, 1983).
Many insects use environmental cues such as photoperiod or temperature and respond with specific physiological, behavioural and morphological modifications that enable them to survive adverse conditions in a state of reduced metabolism (Saunders, 1982; Tauber et al., 1986). Photoperiod plays a major role in diapause induction in many species of insects, particularly those from temperate regions (Beck, 1980; Saunders, 1982).
Quality and quantity of food affect incidence of diapauses in insects (Tauber et al., 1986; Danks, 1987; Overmeer et al., 1989; Fielding, 1990). A restricted intake of nutrients can indirectly lead to the induction of diapause due to a delay in development (Clay & Venard, 1972; Saunders & Bradley, 1984).
Some coleopteran insects such as Colorado potato beetle (Leptinotars decemlineata) and hemipteran insects such as Sunn pest (E. integriceps) enter diapause in the adult stage. In these insects, ovarian development is arrested in diapausing females, and the diapause is called a reproductive diapause (Okuda & Chinzei, 1988).
Since little is known about factors affecting diapause in Sunn pest this research was undertaken to study the effect of two different diet including wheat kernels (grains) and growing wheat plants in two different photoperiods including long day photoperiod (16L: 8D) and short day photoperiod (8L: 16D) on the induction of adult Sunn pest reproductive diapause.
In the current study differences in reproductive organ size and haemolymph protein concentration under long- and short day photoperiods and two different diets were used in the determination of diapause induction.
Also, developmental duration of E. integriceps from egg to adult emergence and the fertility of the females (egg laying activity) were determined under long- and short day photoperiods and two different diets.
The gained knowledge is essential for understanding the seasonal biology of the Sunn pest, and such information is required for the development of effective pest management strategies of the species.
MATERIALS AND METHODS Maintenance of insects' colony
E. integriceps adults have been collected from the Karaj wheat field, Tehran Province, Iran. Stock colony of E. integriceps was maintained in the laboratory under 16L: 8D photoperiod at 26 ±1°C and 55 ± 5% RH on soaked wheat kernels as described by Bandani et al (2009) and Allahyari et al. (2010). Photoperiod and diet treatment
To determine the effect of photoperiod and diet on diapause, eggs (5 day old) were placed under either long-day photoperiod (16L: 8D) or short day photoperiod (8L: 16D) each in two diet regimens including wheat grains and growing wheat plants prior to their ear-bearing stage at 26±1oC and 55% RH. Each treatment was repeated three times each replication with 80 eggs. The growing plants were replaced when wilted or yellowed.
The following parameters including development time to the adulthood, gonad size using drawing tube, haemolymph protein concentration were examined in adult females and males at 1 day and 7day post emergence.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 414
Cold treatment To test the effect of cold treatment on diapause, adult females and males (<24-
h-old) were collected from the stock colony and were transferred to 4oC one day post-emergence. During the cold experiment, the insects did not receive food and water.
Also some diapausing adults (≈45-day-old) collected from the natural habitat where they were in diapausing state and their gonad size was determined.
Control insects (<24-h-old) were collected from the stock colony and were maintained at normal rearing conditions (in laboratory under 16L: 8D photoperiod at 26 ±1°C and 55 ± 5% RH).
Adults (controls and cold exposed adults) were dissected in distilled water at 15, 30 and 45 days post-exposure and gonad size was measured. Individual adult insect after cold-exposed time (15, 30, 45 days) was transferred to normal rearing condition (26 ±1°C) in order to feed and then after three days their haemolymph protein was measured. Transferring to normal rearing condition was done since taking blood from starved insect was problematic. Determination of haemolymph protein concentration
Protein concentration was measured according to the method of Bradford (1976), using bovine serum albumin (Bio-Rad, Munchen, Germany) as a standard. For protein determination, haemolymph from the control and treated adults (females and males) was collected in a chilled calibrated micro capillary pipette through amputated forelegs and diluted (1:1) with anticoagulant buffer (41 mM citric acid, 1.7 mM EDTA, 98 mM NaoH and 186 mM Nacl; pH 4.5). The samples were centrifuged at 10000g for 10 min at 4°C to remove haemocytes and other tissue fragments. The resulting supernatants were stored at -20°C for further analyses. Electrophoresis
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was conducted on 10% slab gels according to the Laemmli (1970). Samples were diluted (1:1) in sample buffer (0.5 M Tris–HCl, pH 6.8, 10% SDS, Glycerol, 2-mercaptoethanol), boiled for 5 min and loaded into the gel along with bromophenol blue as tracking dye. Gels were run in Tris–glycine buffer (Tris base, SDS, glycine, pH 8.3). Following electrophoresis, gels were stained in 0.1% coomassie brilliant blue R-250 in 40% methanol and 10% acetic acid at room temperature. Gels were then destained in 40% methanol and 10% acetic acid until bands appeared. Statistical Analysis
Data were compared by one-way and two-way analysis of variance (ANOVA), followed by LSD multiple range test when significant differences were found at P ≤ 0.05. Software SAS was used for all statistical analyses.
RESULTS Effect of photoperiod and diet treatment on stadium duration
Photoperiod condition caused significant difference in development time of nymphs when reared on both wheat grains and growing wheat plant (P<0.05).
Stadium duration in short day photoperiod condition (31.43 days) was shorter than long day photoperiod condition (38.58 days) (P<0.05) i.e. When the nymphs were placed in short day conditions (8 L: 16D) and fed on wheat grains their

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 415
development time (time taken to adult) was shortened about 15–16 days in compare with those nymphs that placed in long day conditions and fed on wheat plants.
Short day photoperiod condition and wheat grain diet caused a significant decrease in the developmental duration of E. integriceps (28.59 days) compared to the other regimes (P<0.05). Development time to adulthood in short day photoperiod condition and wheat plant diet with long day photoperiod condition and wheat grain diet was almost similar (≈ 34 days). Long day photoperiod condition and wheat plant diet caused longer developmental duration (43.04 days) (Fig 1). Effect of photoperiod and diet treatment on gonad size
As early as the first day of imaginal life, the size of the reproductive organ was different in examined regimes (Table 1). In both sexes, adults promote development of their reproductive organs under wheat grain diet. However, there was not significant difference in ovarian length among regimes. The mean width of ovary in long day condition and wheat grain diet was higher than other regimes (P<0.05). Short day condition and wheat grain diet caused larger ovary size (Size = Width X Length) than long day conditions and wheat plant diet (P<0.05). The mean width, length and size of testis in growing wheat plant reared Sunn pest was smaller than wheat grain reared insects (P<0.05) (Table 1). Effect of photoperiod and diet treatment on haemolymph protein concentration
There was not significant difference in haemolymph protein concentration between genders (Fig. 2). For example, protein concentration in insects reared on short day- and long day photoperiod on wheat grains on first day was 0.12 and 0.14 mg ml-1, respectively. However, total protein concentrations increased over time (P<0.01). Protein concentration in long day photoperiod on wheat grains at first and seven days was 0.12 and 0.25 mg ml-1, respectively. Effect of cold treatment on gonad size
There was not significant difference in ovarian width and length among insects in different days in different treatments i.e. 15-, 30- and 45 days cold exposed adult. However, ovarian length was increased in control insects (at normal rearing conditions (in laboratory under 16L: 8D photoperiod at 26 ±1°C and 55 ± 5% RH) at 45 days after adult emergence (P<0.05) (Table 2). Cold exposed females had larger ovary than control (P<0.05). Ovary size in females collected from natural habitat was similar to control insects (laboratory reared insects).
Testis size increased in control males over time, but its development did not occur in cold exposed males (P<0.01). Also there was not significant difference in gonad size between laboratory reared and insects collected from natural habitat (Table 2). Effect of cold treatment on haemolymph protein concentration
Total protein concentration declined in cold treated insects. Cold exposed females and males had lower haemolymph protein concentration (0.185±0.01 and 0.197±0.005 mg/ml respectively) than control females and males (0.293±0.007 and 0.277±0.048 mg/ml respectively) (P<0.01). There was not significant difference in haemolymph protein concentration between genders in control. However, cold exposed females had lower haemolymph protein than males

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 416
(P<0.01) (Fig.3). Some protein bands of control haemolymph did not observe in cold exposed insects haemolymph (Fig. 4).
DISCUSSION
Photoperiod is a reliable signal for insects to indicate coming winter season. So, insects of the temperate zones often use photoperiod to predict unfavorable conditions. In the current study the effect of photoperiod conditions such as long and short day on the insect stadium duration and future adult reproductive diapause was assessed. Results showed that short day photoperiod condition reduced nymphal duration by almost 7 days indicating that insect use the short day signals to predict unfavorable condition as a result they speed up their growth. However, it did not significantly affect the insect reproductive diapause.
Short day photoperiod induced reproductive diapause in Scotinophara lurida (Hemiptera: Pentatomidae) (Cho et al., 2008) and another pentatomid, Graphosoma lineatum (Nakamura et al., 1996). Solbreck (1979) showed that when laboratory cultures of third or fourth instar nymphs of Neacoryphus bicrucis (Say) (Lygaeidae) were moved from a 16L: 8D to 12L: 12D photoperiod condition, diapause occurred in 66% of adult females reared from third instar nymphs and in 43% of adult females reared from fourth instar nymphs.
Some insects that have summer diapause they use long day lengths instead of short day lengths for entering diapause (Masaki, 1980). The brown stink bug, Euschistus heros showed better reproductive performance at the long day photoperiod and enters reproductive diapause at short day photoperiod (Mourao & Panizzi, 2002). Pyrrhocoris apterus shows ovarian diapause in short day photoperiod; whilst normal reproduction occurs in long day photoperiod (Hodkova, 1975).
Food has an influence on many aspects of diapause (Tauber et al., 1986). Food presence (Fielding, 1990) and food quality (Overmeer et al., 1989) are the factors that contribute to the diapause decision. In the current study it was found that food type (wheat grain and growing wheat plant) did not affect diapause induction significantly but it affected stadium duration (lower nymphal development time).
Interestingly, ovary and testis size was affected by photoperiod and diet. Results showed that when the insects placed in short day conditions and on wheat grains their ovary size increased indicating that wheat grain had more nutrients and perhaps it prevented induction of diapause in the adult. However, it did not affect significantly the insect diapause. Female adults of Protophormia terraenovae produced normal ovaries under non-diapausing conditions, whereas their ovarian diapause occurred under diapause-inducing conditions (Shiga & Numata, 2001). Short day length and low temperature induced female diapause in Oncopeltes fasciatus. However, this condition did not induce diapause in the male (Caldwell & Dingle, 1967; Dingle, 1974).
Environmental temperature is a key factor in determination of physiological process on ectotherm species (Honek & Kocourek, 1990; Chapman, 1998). Temperature determines the biological performances of insects, such as development time of pre-imaginal stages, life expectation and reproductive parameters of the adults, induction and termination of diapause, having by this way a differential contribution of each single individual for the population growth (Kontodimas & Stathas, 2005; Cabral et al., 2006). The rate of diapause development is often influenced by temperature (Tauber et al., 1986).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 417
It has been postulated that in a heteropteran bug, Pyrrhocoris apterus, termination of diapause was stimulated by cold (Kostal et al., 2008).
During cold exposure and forced starvation, total amino acid levels often increase in insects (Hanzal & Jegorov, 1991) and is believed to play a major role in cold- hardening. In our experiment protein concentration and the number of protein bands in the cold exposed females and males of the Sunn pest were less than those of controls. Thus, these insects could not resist cold treatment and died during the experiment.
In conclusion it should be said that diapause is a state of markedly decreased metabolism and activity, as well as reproductive arrest, is induced by changes in photoperiod and temperature, and is orchestrated hormonally (Denlinger, 1983). So the photoperiod changes did not affect diapause induction in the Sunn pest. Although, it has been said that cold affect Sunn pest diapause, current study showed that cold did not affect considerably diapause induction in the Sunn pest.
ACKNOWLEDGEMENTS
This work was funded partly by a grant from University of Tehran.
LITERATURE CITED Allahyari, M., Bandani, A. R. & Rezaei, M. H. 2010. Subcellular fractionation of midgut cells of the sunn pest Eurygaster integriceps (Hemiptera: Scutelleridae): Enzyme markers of microvillar and perimicrovillar membranes. Insect Physiology, 56 (7): 710-717. Bandani, A. R., Kazzazi, M. & Mehrabadi, M. 2009. Purification and characterization of midgut a -amylases of Eurygaster integriceps. Entomological Science, 12: 25-32. Beck, S. D. 1980. Insect Photoperiodism. 2th ed. Academic Press, New York. Bradford, M. M. 1976. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72: 248-254. Cabral, S., Soares, A. O., Moura, R. & Garcia, P. 2006. Suitability of Aphis fabae, Myzus persicae (Homoptera: Aphididae) and Aleyrodes proletella (Homoptera: Aleyrodidae) as prey for Coccinella undecimpunctata (Coleoptera: Coccinellidae). Biological Control, 39: 434-440. Caldwell, R. L. & Dingle, H. 1967 . The regulation of cyclic reproductive and feeding activity in the milkweed bug Oncopeltus by temperature and photoperiod. Biological Bulletin, 133: 510-525. Chapman, R. F. 1998. The insects: structure and function. 4th ed. Cambridge University Press, UK. 770 pp. Cho, J. R., Lee, M., Kima, H. S. & Boo, K. S. 2008. Effect of photoperiod and temperature on reproductive diapause of Scotinophara lurida (Burmeister) (Heteroptera: Pentatomidae). Journal of Asia-Pacific Entomology, 11: 53-57. Clay, M. E. & Venard, C. E. 1972. Larval diapause in the mosquito Aedes triseriatus: effects of diet and temperature on photoperiodic induction. Journal of Insect Physiology, 18: 1441 -1446. Danks, H. V. 1987. Insect Dormancy: An Ecological Perspective. Biological Survey of Canada, Ottawa. Denlinger, D. L. 2000. Molecular Regulation of Insect Diapause. Environmental Stressors and Gene Responses. Edited by K.B. Storey and J. Storey. Denlinger, D. L. 1983. Hormonal control of diapause. In: Kerkut, G. A. & Gilbert, L. I. (Eds), Comprehensive Insect Physiology, Biochemistry, and Pharmacology, Vol. 9,. Pergamon, Oxford. pp. 591-620. Dingle, H. 1974. Diapause in a migrant insect, the milkweed bug Oncopeltus fasciatus (Dallas)(Hemiptera: Lygaeidae). Oecologia, 17: 1-10.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 418
Fielding, D. J. 1990. Photoperiod and food regulate termination of diapause in the squash bug, Anasa tristis. Entomologia Experimentalis et Applicata, 55: 119-124. Hanzal, R. & Jegorov, A. 1991. Changes in free amino acid composition in hemolymph of larvae of the wax moth, Galleria mellonella L. during cold acclimation. Comparative Biochemistry and Physiology, A 100: 957- 962. Hodkova, M. & Socha, R. 1995. Effect of temperature on photoperiodic response in a selected ‗non-diapause‘ strain of Pyrrhocoris apterus (Heteroptera). Physiological Entomology, 20: 303-308. Honek, A. & Kocourek, F. 1990. Temperature and development time in insects: a general relationship between thermal constants. Zool Jb Syst, 117: 401-439. Kontodimas, D. C. & Stathas, G. J. 2005. Phenology, fecundity and life table parameters of the predator Hippodamia variegata reared on Dysaphis crataegi. BioControl, 50: 223-233. Kostal, V., Tollarová, M. & Doležel, D. 2008. Dynamism in physiology and gene transcription during reproductive diapause in a heteropteran bug, Pyrrhocoris apterus, Journal of Insect Physiology, 55: 77-88. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 221: 680-685. Masaki, S. 1980. Summer diapause. Annual Review of Entomology, 25: 1-25. Mourao, A. P. M. & Panizzi, A.R. 2002. Photophase influence on the reproductive diapause, seasonal morphs, and feeding activity of Euschistus heros (Fabr., 1798) (Hemiptera: Pentatomidae). Brazilian Journal of Biology, 62 (2): 231-238. Nakamura, K., Hodek, I. & Hodkova, M. 1996. Recurrent photoperiod response in Graphosoma lineatum (L.). Europian Journal of Entomology, 63: 518-523. Numata, H. & Hidaka, T. 1983. Compound eyes as the photoreceptors in the bean bug. Experientia, 39: 868-869. Okuda, T. & Chinzey, Y. 1988. Vitellogenesis in a lady beetle, Coccinella septempunctata in relation to the aestivation-diapause. Journal of insect physiology, 34 (5): 393-401. Overmeer, W. P. J., Nelis, H. J. C. F., de Leenheer, A. P., Calis, J. N. M. & Veerman, A. 1989. Effect of diet on the photoperiodic induction of diapause in three species of predatory mite, Amblyseius potentillae, Amblyseius cucumeris and Typhlodromus pyri. Experimental and Applied Acarology, 7: 281-287. Paulian, F. & Popov, C. 1980. Sunn pest or cereal bug. Wheat Technical Monograph. Ciba-Geigy Ltd, Basel, Switzerland. pp. 69-74. Radjabi, G. H. 2000. Ecology of Cereal's Sunn pests in Iran. Agricultural Research Education and Extension Organisation Press, Iran Saunders, D. S. 1982. Insect Clocks. In Tauber, M. J., Tauber, C. A., Masaki, S., 1986. Pergamon, Oxford. Seasonal Adaptations of Insects. Oxford University Press. Shiga, S. & Numata, H. 2001. Anatomy and functions of the brain neurosecretory neurons with regard to reproductive diapause in the blow fly Protophormia terraenovae. In: Denlinger, D. L., Giebultowicz, J. & Saunders, D. S. (Eds), Insect Timing: Circadian Rhythmicity to Seasonality. Solbreck, C. 1979. Induction of diapause in a migratory seed bug, Neacoryphus bicrucis (Say) (Heteroptera: Lygaeidae). Oecologia (Berlin) 43: 41-49. Tauber, M. J., Tauber, C. A. & Masaki, S. 1986. Seasonal Adaptations. Oxford University Press, New York.
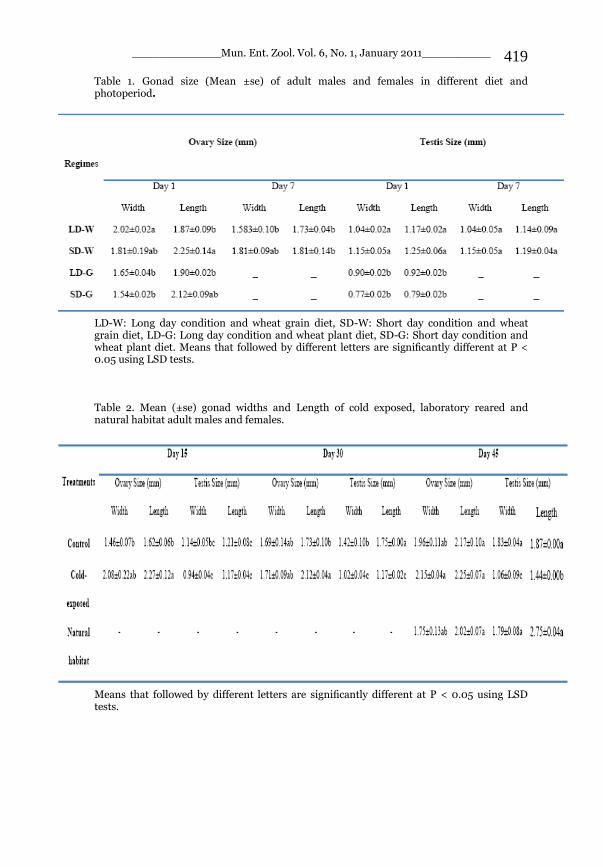
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 419
Table 1. Gonad size (Mean ±se) of adult males and females in different diet and photoperiod.
LD-W: Long day condition and wheat grain diet, SD-W: Short day condition and wheat grain diet, LD-G: Long day condition and wheat plant diet, SD-G: Short day condition and wheat plant diet. Means that followed by different letters are significantly different at P < 0.05 using LSD tests. Table 2. Mean (±se) gonad widths and Length of cold exposed, laboratory reared and natural habitat adult males and females.
Means that followed by different letters are significantly different at P < 0.05 using LSD tests.
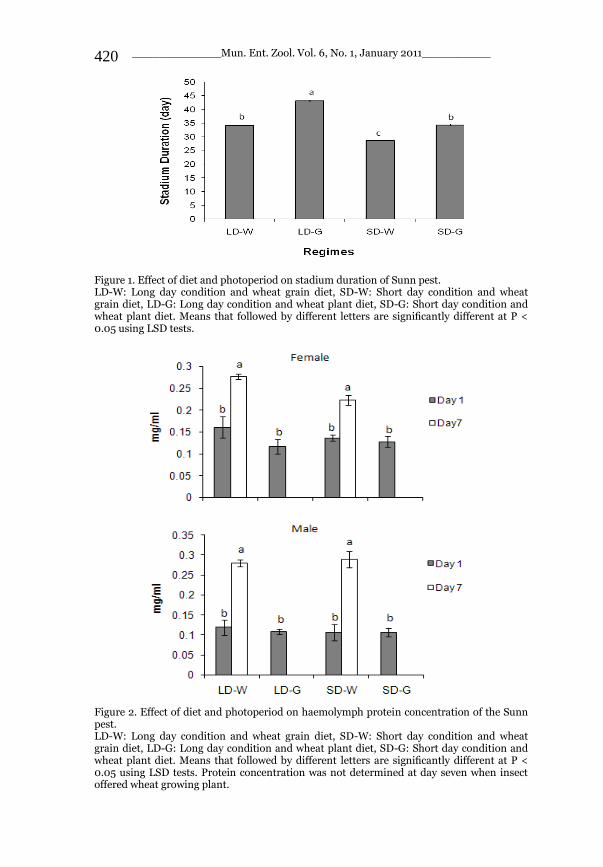
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 420
Figure 1. Effect of diet and photoperiod on stadium duration of Sunn pest. LD-W: Long day condition and wheat grain diet, SD-W: Short day condition and wheat grain diet, LD-G: Long day condition and wheat plant diet, SD-G: Short day condition and wheat plant diet. Means that followed by different letters are significantly different at P < 0.05 using LSD tests.
Figure 2. Effect of diet and photoperiod on haemolymph protein concentration of the Sunn pest. LD-W: Long day condition and wheat grain diet, SD-W: Short day condition and wheat grain diet, LD-G: Long day condition and wheat plant diet, SD-G: Short day condition and wheat plant diet. Means that followed by different letters are significantly different at P < 0.05 using LSD tests. Protein concentration was not determined at day seven when insect offered wheat growing plant.
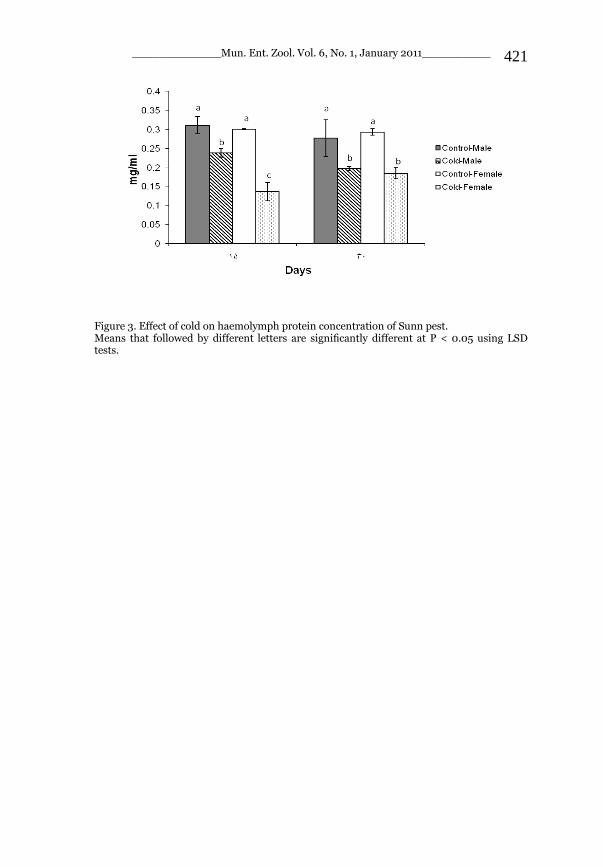
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 421
Figure 3. Effect of cold on haemolymph protein concentration of Sunn pest. Means that followed by different letters are significantly different at P < 0.05 using LSD tests.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 422
A NEW RECORD FOR FLOWER FLIES FAUNA OF TURKEY, CHRYSOTOXUM LESSONAE (GIGLIO-TOS, 1890)
(DIPTERA: SYRPHIDAE)
Süleyman Sarıbıyık* * Kastamonu Üniversitesi, Eğitim Fakültesi, 37200, Kastamonu, TÜRKĠYE. E-mail: [email protected] [Sarıbıyık, S. 2011. A new record for flower flies fauna of Turkey, Chrysotoxum lessonae (Giglio-Tos, 1890) (Diptera: Syrphidae). Munis Entomology & Zoology, 6 (1): 422-424] ABSTRACT: Chrysotoxum lessonae (Giglio-Tos, 1890) is recorded for the first time from Turkey in Bolu, Çankırı and Kastamonu provinces. KEY WORDS: Syrphidae, New record, Turkey.
Hoverflies (Syrphidea) are distributed worldwide, with the greatest species diversity in the New World Tropics. More than 6000 species have been described so far, but probably about 14000 species could belong to the family. Many books are available for the identification of adult stages, at least for Western Palaearctic taxa. The genus Chrysotoxum has been recognised from the beginning of XIX century as Antiopa. While most authors agree in placing the genus within the Syrphini the taxonomic ranks of several entities it comprises are contested. More than 110 Chrysotoxum species have been described so far and, according to Peck (1988), 19 are present in Europe (Caucasus excluded). Several of these species have been described on the basis of small differences in abdomen or leg colour and in other subtle morphological traits (Masetti et. al., 2006).
Until now, ten species of the genus Chrysotoxum have been recorded from Turkey, by Turkish and foreign researchers (Table 1).
MATERIAL AND METHODS
The specimens were collected from yellow flowered compositae found near the small water flows in the Abies sp. and Pinus sp. (Pinaceae) forests in Ilgaz Mountains (Çankırı). With the present work, five specimens were collected from Bolu (Abant), Çankırı (Ilgaz Mountains) and Kastamonu (Küre, Masruf pass). Flies were prepared and labelled following standart protocols for museum material. The following works were used for species identification: Sack, (1932), Violovich, (1974) and Bradescu, 1991. The samples were collected by the author. The specimens are preserved in the personal collection of the author.
RESULTS
Chrysotoxum lessonae (Giglio-Tos, 1890) Material examined: 1 ♂, Bolu, Abant, 1120m., 19.VII.1995,. 1 ♂, Çankırı, Ilgaz Mountain National Park, 1900 m., 08.VIII.1997, 2 ♂♂, Çankırı, Ilgaz Mountain National Park, 1900 m., 18.VIII.2004, 1♀, Kastamonu, Küre Masruf pass, 1250 m., 30.VIII.1996. Flower visited: Yellow flowered Compositae. Range: Italy, Roumania (Peck, 1988). This is a new record for the Turkish fauna.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 423
Table 1. The list is included known Turkish Chrysotoxum species.
Genus Species References pertaining
to Turkey C
hry
so
tox
um
1. Chrysotoxum bicinctum (L., 1758) Dirickx (1994), Sarıbıyık (2000, 2003a,b,d), Sarıbıyık & Hasbenli (2006), Speight (2008)
2. Chrysotoxum cautum (Haris, 1776)
Tuatay et. al. (1967, 1972), Dirickx (1994), Hurkmans et al. (1997), Sarıbıyık (1999, 2003a, b, d), Sarıbıyık & Hasbenli (2006), Reemer & Smit (2007), Speight (2008)
3. Chrysotoxum elegans Loew, 1841 Dirickx (1994), Speight (2008) 4. Chrysotoxum fasciolatum (de Geer, 1776) Sarıbıyık (2000, 2003b, d)
5. Chrysotoxum festivum (L., 1758)
Bischof (1902), Hayat & Alaoğlu (1990), Dirickx (1994), Sarıbıyık & AktaĢ (1996), Hurkmans & Hayat (1997), Hurkmans et al. (1997), Sarıbıyık (1999, 2000, 2003a,b,d), Sarıbıyık & Hasbenli (2006), Reemer & Smit (2007), Speight (2008)
6. Chrysotoxum impressum Becker, 1921 Dirickx (1994)
7. Chrysotoxum intermedium Meigen, 1822
Bischof (1902), Sack (1932), Gadeau De Kerville (1939), Özgür (1987), Claussen & Lucas (1988), Dirickx (1994), Sarıbıyık & AktaĢ (1996), Sarıbıyık (1999, 2000, 2003a,b,c,d), Sarıbıyık & Hasbenli (2006), Reemer & Smit (2007)
8. Chrysotoxum octomaculatum Curtis, 1837 Hurkmans et al. (1997)
9. Chrysotoxum parmense Rondani, 1845
Sack (1932), Violovich, (1974),Özgür (1987), Claussen & Lucas (1988), Dirickx (1994), Sarıbıyık & AktaĢ (1996), Hurkmans et al. (1997), Sarıbıyık (1999, 2000, 2003a,b,d), Sarıbıyık & Hasbenli (2006), Speight (2008)
10. Chrysotoxum vernale Loew, 1841 Hurkmans et al. (1997)
LITERATURE CITED
Bischof, J. 1902. Ergebnisse einer naturwissenschaftlichen Reise zum Erdschias Dagh (Kleinasien). Annales des k. naturhistorischen Hofmuseums, 20: 1-9. Bradescu, V. 1991. Les syrphides de Roumanie (Diptera: Syrphidae) cles de determinationet repartition. Trav. Mus. Hist. nat. ―Grigore Antipa―, 31: 7-83. Claussen, C. & Lucas, Jan, A. W. 1988. Zur Kenntnis der Schwebfliegenfauna der Insel Kreta mit der Beschreibung von Eumerus minotaurus sp. n. (Diptera: Syrphidae). Zeitschrift Entomologie, 9: 133-168. Dirickx, H. G. 1994. Atlas des Diptéres syrphides de la région méditerranéenne. Studiedocumenten van het Koninklijk Belgisch Instituut voor Natuurwetenschappen, 75: 1-317. Gadeau De Kerville, H. 1939. Voyage Zoologique Liste méthodique des invertébrés et des vertébrés récoltés en Asie-Mineure (avril-mai 1912). Paul le chevalier, 1-148, Paris.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 424
Hayat, R. & Alaoğlu, Ö. 1990. Fauna of Syrphidae (Diptera) in the vicinity of Erzurum (Part I) Syrphinae. Türkiye Entomoloji Dergisi, 14 (3): 173-182. Hurkmans, W., Hayat, R. & Özbek, H. 1997. Insect and plants of a marsh at Küçükgeçit, AĢkale, Erzurum, Turkey: A preview on preservation. Türkiye Entomoloji Dergisi, 21 (2): 95-108. Hurkmans, W. & Hayat, R. 1997. Ethology and ecology of Merodon (Diptera: Syrphidae) in Turkey II: descriptions of new species and notes on other syrphid flies. Dipterists Digest., 3: 62-79. Masetti A., Luchetti A., Sommaggio D., Burgio G. & Mantovani B. 2006. Phylogeny of Chrysotoxum species (Diptera: Syrphidae) inferred from morphological and molecular characters. Eur. J. Entomol., 103: 459-467. Özgür, A. F. 1987. Akdeniz Bölgesi kıyı Ģeridi Syrphidae (Diptera) faunası II. Altfamilya: Cheilosiinae, Sphegininae, Pelecocerinae, Milesiinae, Chysotoxinae, Volucellinae, Çukurova Üniversitesi, Ziraat Fakültesi Yayınları, 2 (3): 67-81. Reemer, M. & Smit, J. 2007. Some hoverfly records from Turkey (Diptera: Syrphidae), Volucella, 8: 135-146. Sack, P. 1928-1932. Syrphidae. In, Lindner, E. Die Fliegen der paläarktischen Region, IV (6). Stuttgart, Germany. +451 pp. 18 pls. Sarıbıyık, S. 1999. Syrphinae Fauna of the West Blacksea Region, (Diptera: Syrphidae), Gazi Üniversitesi Kastamonu Eğitim Dergisi, 7 (1): 185-194. Sarıbıyık, S. 2000. Fauna of Syrphidae in Ilgaz and IĢık Mountains and their vicinity (Diptera: Syrphinae). Gazi Üniversitesi Fen Bilimleri Dergisi, 13 (1): 55-70. Sarıbıyık, S. 2003a. Fauna of Syrphinae and Milesiinae (Diptera: Syrphidae) Around Tuz Lake. Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (2): 439-450. Sarıbıyık, S. 2003b. The Evaluation of the Works on Syrphidae (Diptera) Fauna in the Western Blacksea Region. Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (2): 461-466. Sarıbıyık, S. 2003c. Syrphinae Fauna of the KahramanmaraĢ‘s Andırın Town (Diptera: Syrphidae). Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (1): 193-198. Sarıbıyık, S. 2003d. Kastamonunun Syrphid Sinekleri (Diptera: Syrphidae). II. Kastamonu Kültür Sempozyumu (18-20 Eylül 2003), 707-719. Sarıbıyık, S. & Aktas, M. 1996. Contribution to the Syrphidae fauna of Turkey (Diptera: Syrphidae) (I). Gazi Üniversitesi Fen Bilimleri Dergisi, 9 (1): 1-13. Sarıbıyık, S. & Hasbenli, A. 2006. Fauna of Syrphinae and Milesiinae (Diptera, Syrphidae) in the Region of South-West Anatolia. Journal of the Entomological Research Society, 8 (1): 43-52. Speight M. C. D. 2008. Species accounts of European Syrphidae (Diptera) 2008. In: Speight, M. C. D., Castella, E., Sarthou, J. P. & Monteil, C. (eds). Syrph the Net, the database of European Syrphidae, 55: 262 pp. Tuatay, N., Gül, S., Demirtola, A., Kalkandelen A. & Çagatay, N. 1967. Nebat Koruma Müzesi Böcek Kataloğu (1961-1966). Zirai Mücadele ve Zirai Karantina Genel Müdürlüğü Yayınları, Mesleki Kitaplar Serisi, Ankara 66 pp. Tuatay, N., Kalkandelen, A. & Aysev (Çağatay), N. 1972. Nebat Koruma Müzesi Böcek Kataloğu (1961-1971). Zirai Mücadele ve Zirai Karantina Genel Müdürlüğü Yayınları Mesleki Kitaplar Serisi, Ankara p. 119. Violovich, N. A. 1974. A reviev of the palearctic species of the genus Chrysotoxum Mg. (Diptera: Syrphidae). Ent. Review., 53 (1): 140-153.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 425
INVESTIGATION ON PREY PREFERENCE AND SWITCHING BEHAVIOR OF THE PREDATORY BUG, ORIUS NIGER WOLFF UNDER LABORATORY CONDITIONS (HET.:ANTHOCORIDAE)
Fereshteh Salehi*, Valiollah Baniameri**, Ahad Sahragard*
and Jalil Hajizadeh* * Department of Plant Protection, College of Agriculture University of Guilan, IRAN. E-mail: [email protected] ** Iranian Research Institute of Plant Protection (IRIPP); Tehran, IRAN. [Salehi, F., Baniameri, V., Sahragard, A. & Hajizadeh, J. 2011. Investigation on prey preference and switching behavior of the predatory bug, Orius niger Wolff under laboratory conditions (Het.: Anthocoridae).Munis Entomology & Zoology, 6 (1): 425-432] ABSTRACT: Prey preference and switching behavior of Orius niger on three preys, Thrips tabaci, Aphis gosssypii and Tetranychus urticae was examined. Ephestia kuehniella Zeller eggs, corn pollen and green bean pods were used as food and oviposition sites were used, respectively. Experiment was carried out under laboratory condition (5±1°C and 65±5% RH, with a 16:8 L: D photoperiod). Four to six daysold females of the predatory bug was used in experiment. We monitored the prey of the preference of the predatory bug was examined on cucumber leaf disks containing: 10 second instar larvae of Thrips tabaci , 10 second or third instar nymph of Aphis gossypii and 10 adults of Tetranychus urticae for 12 hour in 6 replicates. Number of prey consumed was recorded the preference indexes of T. tabaci, A. gossypii and T urticae were 0.54, 0.35 and 0.089, respectively. For each experiment, a piece of host plant, determined densities (10-40, 25-25, 40-10) of both prey and one predator were placed in a cage Each experiment was lasted for 12h and repeated six times. Switching behavior of predatory bug was examined. The results of host preference showed that there was no significant difference between the mean number stage consumed T. tabaci and A. gossypii (p<0.05) and there was significant difference between the mean number of consumed T. tabaci and A. gossypii with T. urticae (p<0.05). In Switching behavior number of prey consumed were recorded: In the first with increasing of relative prey density, preference rate was decreased (negative preference), near equal density, decreasing of relative prey density, preference rate was increased (positive preference). KEY WORDS: Host preference, switching behavior, Orius niger, Thrips tabaci, Aphis gosssypii, Tetranychus urticae.
In recent years, the use of different species of predatory bugs of genus, Orius as biological control agents has greatly increased in many house vegtables and ornamental crops (Van den Meiracker & Ramakers, 1991; Van de Veire & Degheele, 1992; Cocuzza et al., 1997; Baniameri et al., 2005). This genus consists of 67 species (Yasunaga, 1997). The use of different species of predatory bugs Orius spp. as efficient predators has been taken into consideration and they are used widely as predators of thrips (Murdoch et al., 1985), mites and Aphid (Akramovskaya, 1978) and eggs of pentatomids (Pericart, 1972) and aphids (Yasunaga, 1997) in green house cultures. Orius laevigatus was able to distinguish plants infested with either of the two preys and showed a switch for plant infested with spider mite over plant with thrips, plant infested with thrips contained 20-40 larvae per plant, while plants with spider mites had 800-1000 females per plant. The prediction is that inclusion of the inferior prey always depends on the density of the superior prey, and inferior prey (Venzon et al., 2000). According to Venzon (1999) predatory bug Orius laevigatus preffered patches with high density of inferior prey and intermediate density of superior prey. Different prey types often occure in separate patches, so that generalist

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 426
predator has to make an ‗either/or‘ decision. This decision will not only depend on the quality per prey, but also on the quantity of prey per patch (Kindlmann & Dixon. 1999).prey preference of Orius laevigatus for two prey offered Amblyseius degenerance and Amblyseius cucumertis was different due to behavioral differences between preys, Amblyseius degenerance was more mobile than Amblyseius cucumertis and predator have greater success capturing the less active species (Wittmann & Leather, 1997). Both Orius laevigatus (Wittmann & Leather, 1997) and Orius majusculus (Brodsgaard & Enkegaard, 2005) show clear preference for thrips when given a choice between F. occidentalis larvae and A. degenerans. According to Chow et al (2008) prey preference of anthocorid bugs for phytoseiid mites and Frankliniella occidentalis has been examind among different combination of species and prey sizes (age), Orius insidiosus has little difficulty handling adult A. degenerans but adult thrips were especially difficult to capture and kill. Rogers et al. (2000) showed in Coleomegilla maculate the highest level of predation on small prey, the weight- consumption rate was highest for prey of intermediate sized prey, they founded that level of predation by Orius insidiosus was lowest on large prey, adult Frankliniella occidentalis, but similar for intermediate sized prey, adult A. degenerans. Prey switching in Orius insidiosus showed that this anthocorid always switches to the most abundant prey whether foraging on flowers or foliage (Chow et al., 2008).
For biological control measures, it is necessary to determine the efficacy of natural enemy against pests. Cornel (1976) proposed that switching would be most likely in mobile predators which forage among many patches where prey species occure in different proportions, Sutch conditions are well met in orius species which are good fliers, active foragers and polyphagous predators (Malais & Ravensberg. 2003). The objectives of the current study were to determine prey preference and switching behavior of the predatory bug, Orius niger for three prey: Thrips tabaci, Aphis gosssypii and Tetranychus urticae.
MATERIALS AND METHODS
Predator rearing: Predatory bugs used in the experiments were collected on marguerite flowers Chrysanthemum frutescens L. (Asteracea) in Tehran, Iran. O. niger was separated and identified (Baniameri et al., 2005). The predatory bugs were kept in plastic jars (7×10 cm) whit a hole (Ø 2.5 cm) on the top of the jar for ventilation and covered with antithrips net. They were reared on eggs of E. kuehniella as food at 25±1°C and 65±5% RH, with a 16:8 L: D photoperiod. Green bean pods and corn pollen were supplied as oviposition sites and supplementary food, respectively. The jars were lined with crumpled wipes papers to provide hiding place to reduce cannibalsm (Baniameri et al., 2005).
Thrips tabaci rearing: T. tabaci was reared on cucumber leaves of the variety of Negin at 25±1°C with a 16:8 L: D photoperiod. One or two infested cucumber leaves were placed on the green bean pods in plastic jars (7×10 cm) for infestation.
After 7-8 days second instar larvae of T. tabaci were available on the green bean pods and in this way, a cohort larva was maintained Tetranychus urticae rearing: T. urticae was reared on cucumber plant of variety of Negin at the same condition as above. One or two infested leaves were placed on the clean cucumber plant for infestation. After three-four days, plants were heavily infested by the mite.
Aphis gossypii rearing: A. gossypii were reared on cucumber plant of variety of Negin at the same condition as above. Clean cucumber leaf discs (Ø 5 cm) were placed in the center of a plastic Petri dish contaning moisten cotton wool. One or

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 427
two infested leaves were placed on leaf discs (Ø 5 cm) for infestation. After 3 days, plants were heavily infested with second and fourth nymphal instar of A. gossypii.
Prey preference experiment: to examin prey preference of predatory bug, O. niger plexi glass cages, with 25cm diameter and 10 cm height were used. To facilitate ventilation top of the cage was perforated in five places and five holes (Ø 2.5cm) were made and covered with antithrips net. A central hole (Ø 1 cm) was also made on the top of the cage and was filled with cotton. Three plastic petri dishes, (5cm in diameter) were placed in each cage and a cucumber disk leaf 5cm in diameter was placed on clean moistens filter paper. Petry dishes were designed in equal distances from each other and cage center (an incidental designment). In each petri dish 10 second instar larvae of T. tabaci, 10 second and fourth instar nymph of A. gossypii and 10 adults of T. urticae were placed on leaf disk of cucumber. In each cage four to six-day old females of the predatory bug were released through the central hole of the cage. Females were starved for 24h before release. Each experiment lasted 12h. and replicated 6 times. Number of prey eaten by each predator was recorded at the end of experiment. Laboratory condition was the same as before. Consumed prey was eaten not replaced, which means that in the course of the experiment the relative abundance between prey species changed due to the predation activity of O. niger. Prey preference was also measured using Manly´s index (Manly, 1974; Krebs, 1989; Chesson, 1983):
n
i j
i
p
p
1log
log
where α = Manly´s alpha for prey type p
i= proportion of prey i remaining at the end of the experiment relative to the
original input (i=1,2,3,4,…m); pj = proportion of all prey types together remaining at the end of the experiment relative to the original input (j= 1,2,3,4,…m) m= number of prey type. The α indices can take values between zero and one and the values of the different prey types always sum up to one. In the case of a three prey combination the threshold value is 0.33. Thus α expresses the deviation of the predators diet from a random sample of the available prey. SAS (1998) was used to analyse data. Switching behavior of the predator on figures were drawn To examin the Switching behavior of predatory bug O. niger, on two preys were studied in a transparent cage of plastic (25cm in diameter, 10cm in hight). For proper ventilation the top of the cage 5 also small holes places were coverd with antithrips net. A central hole that is 1cm in diameter was made on the top and filled with cotton. Two Petri-dishes were placed in the cage with 7cm distance and a cucumber leaf disk was placed on the moistened paper. Different combinations of two preys (T. tabaci and A. gossypii) selected as 10-40, 25-25, and 40-10 respectively. Each prey combination was repeated to describe five times. Petri-dishes were placed equidistance from each other and cage center. The condition was as before. At each experiment 4-5 day old females and starved for 24h. Three preys species thst were released in to cages through central hole on the top, are shown in Table 1. Each experiment lasted 12h and then the number of prey eaten was recorded. Murdoch no switch model (1969) was used to describe the data:

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 428
CXX
CXY
100
100
x = percent age of prey type1 C = normalization constant in equal density of prey type1and 2 as:
parn
parnC
2
1
n1 par= average number of prey type 1 eaten. n2 par= average number of prey type 2 eaten. Y = proportion of prey type1 or (type 2) eaten.
RESULTS In preference experiments, consumed prey was not replaced due to the Manly‘s index was used and results were presented in table 1. The results showed that there was no significant differences among the mean number stage consumed T. tabaci and A. gossypii (F=24.9, df =2,15,17 and p<0.0001) number of prey consumed among T. tabaci, A. gossypii and T. urticae (p<0.05). O. niger preferred T. tabaci over A. gossypii and T. urticae Switching behavior experiment of O. niger on T. tabaci and A. gossypii showed that T. tabaci is preferred equal densities to A. gossypii (Fig. 1). The feeding ratio of two preys (T. tabaci and A. gossypii) and C value in equal densities are shown in Table 3.
DISCUSSION The results showed that there was no significant difference between the mean number prey stage consumed T. tabaci (αi=0.54) and A. gossypii (αi=0.35) and there was significant difference among the mean number of T. tabaci, A. gossypii and T. urticae (αi=0.089) eaten (p<0.05, F=24.9). Generalist predators often attack a variety of prey which differs in energetic value and both capture and handling costs (Chow et al., 2008). Optimal foraging theory predicts that predators should utilize large prey to maximize energy gain (Stephans & Krebs, 1986) Additional information on biomass and energetic values for experimental prey of different size (age) would provide a better understanding of differential utilization of prey by O. niger. Manly‘s α index can take values between zero and one and the values of the different prey types always sum up to one (Manly, 1974). O. niger preference experiments with three prey combinations with equal numbers showed a great preference to T. tabaci and A. gossypii against T. urticae. The more phylogenic similarity between predator and prey the more prediction valuablity or prey comparative value for predator has increased. In other words, the better predator can find its need of amino acid and carbohydrates from a prey that has closed phylognetic relationship with itself (Bilde & Toft, 1994). The intrinsic growth rates were higher when predatory bugs Orius laevigatus fed with diets containing thrips than when they were fed spider mites (Venzon et al., 2002). Spider mites produce web that may serve as a refuge from predation (Sabelis & Bakker, 1992). The results showed that there was a positive preference to T. tabaci and negative one for A. gossypii. It was also shown that T. tabaci was preferred by the predator, when its desity was increased

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 429
(a Switching behavior was observed). It has been shown that predatory bugs, Orius laevigatus preferred plants with the inferior prey species (spider mites) when plants with moderate densities of thrips were the alternative. This preference disappeared when thrips density was increased (Venzon et al., 2002). Chow et al. (2008) showed that total and relative predation of thrips and/or mites by O. insidiosus depended on the types of prey available, O. insidiosus tended to switch to the most abundant type of prey. Predators that switch from feeding on one prey species to feeding on another species suddenly change from being harmless to dangerous for this new prey species, while becoming harmless for the old prey (Venzon et al., 2000). Results showed that when two preys are offered in equal proportions (25:25), the predator showed more preference for T. tabaci (C = 1.19). The curves in Fig.2 show the expected proportion of the two species with switch, there is therefore, a distinct positive switching behaviour of the predator was shown for Thrips tabaci. In the first by increasing the relative prey density, preference rate was decreased (negative switching), as the prey density increased a positive switching was observed.
LITERATURE CITED Akramovskaya, E. G. 1978. The biology of some predatory bugs of the family Anthocoridae in the conditions of the Ararat valley in Armenia. Bio. Zh. Armenii., 31 (9): 959-964. Baniameri, V., Soleiman-Nejadian, E. & Mohaghegh, J. 2005. Life table and age – dependent reproduction of the predatory bug, Orius niger Wolff (Heteroptera: Anthocoridae) at three constant temperatures: a demographic analysis. Appl. Entomol. Zool., 40 (4): 545-550. Bilde, T. & Toft, S. 1994. Prey preference and egg production of the carabid Beetlle Agonum dorsal. Entomol. Exp. Appl., 73: 151-156. Chesson, J. 1983. The estimation and analysis of preference and its relationship to foraging models. Ecology, 70: 1227-1235. Coccuzza, G. E., Declercq, P. E., Izzio, S. L., Van de Veire, M., Tirry, L., Degheele, D. & Vacante, V. 1997. Life tables and predation activity of Orius laevigatus (Fieber) and O. albidipennis (Reuter) at three costant temperature. Entomol. Exp. et Appl., 85: 189-198. Chow. A., Chau, A. & Heinz, K. M. 2008. Compatibility of Orius insidiosus (Hemiptera: Anthocoridae) with Amblyseius (Iphiseius) degenerans (Acari: Phytoseiidae) for control of Frankliniella occidentalis (Thysanoptera: thripidae) on greenhouse roses. Biological control, 44: 259-270. Cornell, H. 1976. Stertegies and the adaptive significance of switching in some generalist predators. The American Naturalist, 110: 317-320. Holling, C. S. 1959. Some characteristics of simple types of predation and parasitism. Can. Ent., 91: 385-398. Kindlmann, P. & Dixon, F. G. 1999. Strategis of aphidophagus predators: lessons for modelling insect predator-prey dynamics. J. Appl.Ent., 123: 397-399. Krebs. C. J. 1989. Optimal foraging: theory and experiment. Nature 268: 181-190. Manly, B. 1974. A model for certain types of selection experiments. Biometrics, 30: 281-294. Meiacker, R. A., van den, F. & Ramakers, P. M. J. 1991. Biological control of wesern flower thrips Frakliniella occidentalis, in sweet pepper, with the anthocorid predator, Orius insidiosus. Med. Fac. Landbouwn. Rijksuniv. Gent., 56: 241-249. Malais, M. H. & Ravensberg, W. J. 2003. Knowing and recognizing. The biology of glasshouse pests and their natural enemies. Koppert B.V., Berkel en Rodenrijs, The Netherlands. Murdoch, W. W. 1969. Switching in general predators: experiments on predator specificity and stability of prey populations. Ecological Monographs., 39: 335-354.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 430
Murdoch, W. W., Chesson, J. & Chesson, P. L. 1985. Biological control in theory and practice. Am. Nat., 125: 344-366. Pericart, J. 1972. Hemipteres, Anthocoridae, Cimicidae et Microphysidae de Louest–Palearctique. Faune Eur Bassin Mediter., 7 (1): 398-402. Ramakers, P. M. J. 1978. Possibilities for biological control of Thrips tabaci Lind. (Thysanoptera: Thripidae) in glasshouse. Med. Fac. Landbouww. Rijksuniv. 43: 463-469. Reid, C. D. 1991. Ability of Orius insidiosus (Het.; Anthocoridae) to search for , find and attack European corn earworm eggs on corn. Entomol., 84 (1): 78-79. Rogers, C., Coderre, D. & Boivin, G. 2000. Differential prey utilization by the generalist predator Colemegilla maculata lengi according to prey size and species. Entomologia Experimentalis et Applicata, 94: 3-13. Rutledge, C. E. & O,Neil, R. J. 2005. Orius insidiosus (Say) as a predator of the soybean aphid, Aphis glycines Matsumura. Biological Control., 33: 56-64. Salas Aguilar, J. & Ehler, L. E. 1977. Feeding habits of Orius tristicolor (White). Ann. Entomol. Soc. Am., 70 (1): 60-62. Sabelis, M. W. & Bakker, F. M. 1992. How predatory mites cope with the web of their tetranychid prey: a functional view on dorsal chaetotaxy in the phytoseiidae. Exp. Appl. Acarol., 16: 203-225. SAS Institute Inc. 1989. SAS/STAT User's Guide. ver. 6, 4th edn, vol. 1,2. SAS Institute, Cary, Nc. Stephens, D. W. & Krebs, J. R. 1986. Foraging Theory. Princeton University Press, Princeton. Tavella, L., Arzone, A. & Alma, A. 1991. Researches on Orius laevigatus, a predator of Frankliniella occidentalis in greenhouses a preliminary note. IOBC/ WPRS Bull., 4: 65-72. Tommasini, M. G. & Nicoli, G. 1994. Adult activity of four Orius species reared on two preys. IOBC/ WPRS Bull., 16 (2): 181-184. Van Rijn, P. C. J. 2002. The impact of supplementary food on a prey- predator interaction. Ph.D Thesis, University of Amsterdam, The Netherlands, 251 pp. Van de Veire, M. & Degheele, D. 1992. Biological control of the Western flower Thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in glasshouse sweet peppers with Orius spp. (Het.; Anthocoridae). A cmparative study between O. niger (Wolff) and O. insidiosus (Say). Biocont. Sci.Techn., 2: 281-283. Van den Meiracker, R. A. F. & Ramakers, P. M. J. 1991. Biological control of the western flower Thrips framkliniella occidentalis, in sweet pepper, with the anthocorid predator Orius insidiosus. Med. Fac. Landksuww. Rijksuniv. Gent, 56/2a. Venzon, M., Janssen, A. & Sabelis, M. W. 1999. Attraction of a generalist predator towards herbivore-infested plants. Entomologia Experimentalis et Applicata, 93: 303-313. Venzon, M. 2000. Food webs on plants: the role of a generalist predator. Ph.D thesis, University of Amsterdam, The Netherlands, 91 pp. Venzon, M., Janssen, A. & Sabelis, M. W. 2002. Prey preference and reproductive success of the generalist predator Orius laevigatus. Oikos, 97 (1): 116-124. Wittmann, E. J. & Leather, S. R. 1997. Compatibility of Orius laevigatus Fiber (Hemiptera: Anthocoridae) with Neoseiulus (Amblyseius) cucumeris Oudemans (Acari: Phytoseiidae) and Iphiseius (Amblyseius) degenerans Berlese (Acari: Phytoseiidae) in the biocontrol of Frankliniella occidentalis Pergande (Thysanoptera: Thripidae). Experimental and Applied Acarology, 21: 523-538. Yasunaga, T. 1997. The flower bug genus Orius Wolff (Het.; Anthocoridae) from Japan and Taiwan, Part I. Appl. Entomol. Zool., 32 (2): 355-364.
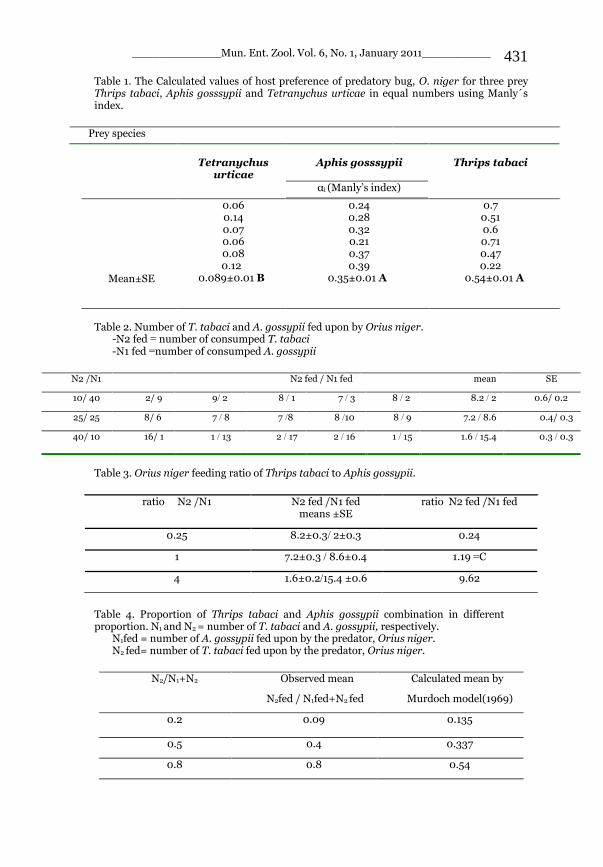
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 431
Table 1. The Calculated values of host preference of predatory bug, O. niger for three prey Thrips tabaci, Aphis gosssypii and Tetranychus urticae in equal numbers using Manly´s index.
Table 2. Number of T. tabaci and A. gossypii fed upon by Orius niger. -N2 fed number of consumped T. tabaci -N1 fed number of consumped A. gossypii
SE mean N2 fed / N1 fed N2 /N1
0.6/ 0.2 8.2 2 8 2 7 3 8 1 9 2 2/ 9 10/ 40
0.4/ 0.3 7.2 8.6 8 9 8 10 7 8 7 8 8/ 6 25/ 25
0.3 0.3 1.6 15.4 1 15 2 16 2 17 1 13 16/ 1 40/ 10
Table 3. Orius niger feeding ratio of Thrips tabaci to Aphis gossypii.
ratio N2 fed /N1 fed N2 fed /N1 fed
means ±SE ratio N2 /N1
0.24 8.2±0.3 2±0.3 0.25
1.19 C 7.2±0.3 8.6±0.4 1
9.62 1.6±0.2 15.4 ±0.6 4
Table 4. Proportion of Thrips tabaci and Aphis gossypii combination in different proportion. N1 and N2 = number of T. tabaci and A. gossypii, respectively. N1fed = number of A. gossypii fed upon by the predator, Orius niger. N2 fed= number of T. tabaci fed upon by the predator, Orius niger.
Calculated mean by
Murdoch model(1969)
Observed mean
N2fed / N1fed+N2 fed
N2/N1+N2
0.135 0.09 0.2
0.337 0.4 0.5
0.54 0.8 0.8
Prey species
Thrips tabaci
Aphis gosssypii
αi (Manly‘s index)
Tetranychus urticae
0.7 0.51 0.6 0.71 0.47 0.22
0.54±0.01 A
0.24 0.28 0.32 0.21 0.37 0.39
0.35±0.01 A
0.06 0.14 0.07 0.06 0.08 0.12
0.089±0.01 B
Mean±SE
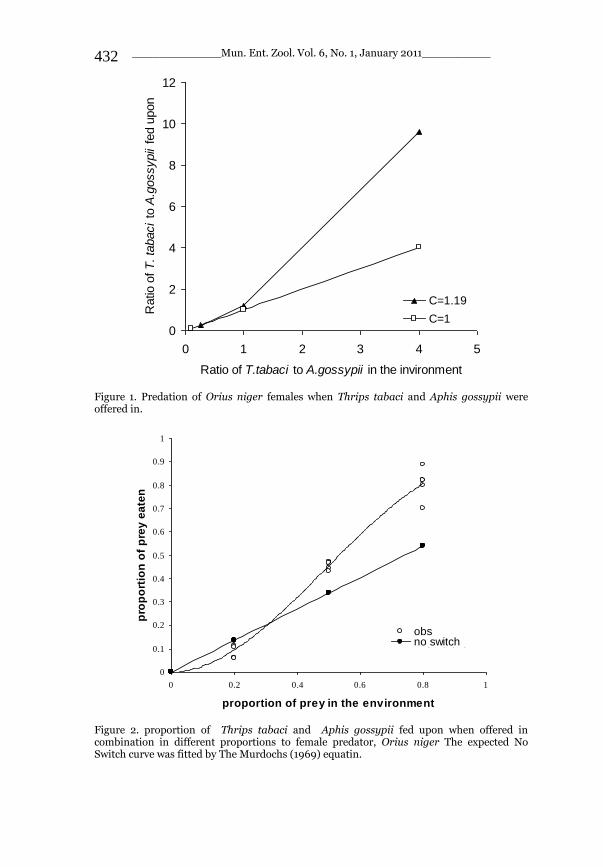
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 432
0
2
4
6
8
10
12
0 1 2 3 4 5
Ratio of T.tabaci to A.gossypii in the invironment
Ratio
of T
. ta
baci
to A
.gossypii fed u
pon
C=1.19
C=1
Figure 1. Predation of Orius niger females when Thrips tabaci and Aphis gossypii were offered in.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 0.2 0.4 0.6 0.8 1
proportion of prey in the environment
pro
po
rtio
n o
f p
rey
ea
ten
obsno switchPoly. (obs)
Figure 2. proportion of Thrips tabaci and Aphis gossypii fed upon when offered in combination in different proportions to female predator, Orius niger The expected No Switch curve was fitted by The Murdochs (1969) equatin.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 433
AN INVESTIGATION ON TURKISH SPHEGINA MEIGEN, 1822 (DIPTERA: SYRPHIDAE)
Süleyman Sarıbıyık*
* Kastamonu Üniversitesi, Eğitim Fakültesi, 37200, Kastamonu, TÜRKĠYE. E-mail: [email protected] [Sarıbıyık, S. 2011. An investigation on Turkish Sphegina Meigen, 1822 (Diptera: Syrphidae). Munis Entomology & Zoology, 6 (1): 433-435] ABSTRACT: Materials of the genera Sphegina sampled from KahramanmaraĢ, Kastamonu and Bolu between 1993 and 2008 are presented. Four species of Sphegina were found to occur in Turkey. Sphegina clavata (Scopoli, 1763) is a new record for the fauna of Turkey. KEY WORDS: Diptera, Syrphidae, Sphegina, new record, Turkey.
Members of the genus Sphegina can be found visiting white Apiaceae and a wide scale of other flowers in shaded, deciduous and coniferous woodland ecosystems. Larvae were usually observed in wet situations under the bark of trees or in sap run (Rotheray, 1993).
Until now four species of the genus Sphegina have been recorded from Turkey, by Turkish and foreign researchers. Sphegina alaoglui Hayat, 1997 (Hayat, 1997) Sphegina clunipes (Fallén, 1816) (Aktas & Saribiyik 1996; Hayat, 1997; Sarıbıyık, 2009) Sphegina elegans Schummel, 1843 (Speight, 2008) and Sphegina sibirica Stackelberg, 1953 (Reemer & Smit, 2007) were hitherto reported for the Turkish fauna.
MATERIAL AND METHODS
Additional data on a further four species of Sphegina has been obtained from material collected by the author between 1993 and 2008 from the districts Kastamonu and Bolu of the Turkish Blacksea region. The specimens have been identified by C. Claussen (Flensburg/ Germany) and S. Sarıbıyık.
The specimens are preserved in the private collection of the author.
RESULTS
As a result of this study, four species of Sphegina were found to occur in Turkey. Sphegina clavata (Scopoli, 1763) is a new record for fauna of Turkey. Distribution of the species in Turkey and rest of world was stated.
Sphegina clavata (Scopoli, 1763) Material examined: 1 ♀, Kastamonu, Karadere village, 1050 m., 07.VIII.1999. Distribution in Turkey: This is a new record for the Turkish fauna. Range: Netherlands, Belgium, France (Paris basin, Vosges, Alps); the Pyrenees and mountainous parts of Central Europe; Italy, Montenegro, Serbia, Slovenia, Greece (Speight, 2008). Flowers visited: White umbellifers.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 434
Sphegina clunipes (Fallén, 1816) Material examined: 2 ♂♂, Bolu, Yedigöller, main road 22. km., Gurbet taĢı locality, 1400 m., 09.VII.1996, 1 ♂, 1 ♀, Kastamonu, Ilgaz Mountain National Park, 1875 m., 03.VII.1999, 1 ♀, 1600 m., 06.VIII.1999, 1 ♂, Yukarı Ġsmailli village, 1400 m., 08.VII.1999, 2 ♂♂, 3 ♀♀, Ilgaz Mountain National Park, Tüfekçi village, Diphan locality, 1500 m., 10.VI.2000, 8 ♂♂, Tosya, Dipsiz göl, 1000 m., 24.VI.2000, 2 ♂♂, Ilgaz Mountain National Park, Tüfekçi village vicinity, 1700 m., 25.VI.2000, 2 ♂♂, 1♀, Kastamonu, Daday, Ballıdağ, 1700 m., 22.V.2007, 4 ♂♂, 5♀, Kastamonu, Daday, Ballıdağ, 1700 m., 09.VI.2007, 2 ♂♂, 3 ♀♀, KahramanmaraĢ, Andırın, Akifiye village, 1600m., 10.VII.2008, 4 ♂♂, 2 ♀♀, KahramanmaraĢ, Andırın, Akifiye village, 1600m., 07.VIII.2008. Distribution in Turkey: Kastamonu and Kayseri (Aktas & Saribiyik, 1996), Artvin (Hayat, 1997), Kastamonu (Sarıbıyık, 2003a,b, 2009). Range: From Fennoscandia south to Iberia; from Ireland eastwards through most of Europe into European parts of Russia and the Caucasus; through Siberia to the Pacific coast; Japan (Speight, 2008). Flowers visited: White umbellifers; Ranunculus.
Sphegina elegans Schummel, 1843 Material examined: 1 ♀, Kastamonu, Karadere village, 1050 m., 07.VIII.1999. Remarks: The specimen agrees morphologically completely with dark females of S. elegans from Central Europe. It differs from the female of the closely related S. alaoglui which gender was shortly described by Mutin (1998) under the name S. pontica, by the yellowish apical tarsomere of the front and middle legs. Distribution in Turkey: Turkey (Speight, 2008). Range: From Fennoscandia south to the Pyrenees and Spain; from Ireland eastwards through central and southern Europe (northern Italy, the former Yugoslavia, and Northern Greece) into European parts of Russia to the Caucasian mountains. It also known from Samos Island in the Mediterranean near the coast of Turkey (Speight, 2008). Flowers visited: White umbellifers.
Sphegina sibirica Stackelberg, 1953 Material examined: 1 ♂, Kastamonu, Bostan village, 23.VI.1999, 1400 m., 2 ♂♂, 1200 m., 03.VII.1999, 1 ♀, Ilgaz Mountain National Park,1875 m., 03.VII.1999, 1 ♂, 1♀, Kastamonu, Daday, Ballıdağ, 1700 m., 22.V.2007, 2 ♂♂, 2♀, Kastamonu, Daday, Ballıdağ, 1700 m., 09.VI.2007. Distribution in Turkey: Trabzon (Reemer & Smit, 2007). Range: From northern Fennoscandia south to central France; from Britain (Scotland to southern England) eastwards through central Europe and European parts of Russia to the Caucasus and into Siberia as far as Kamchatka and Khabarovsk. S.sibirica seems to be spreading rapidly in Europe (Speight 2008). Flowers visited: White umbellifers.
ACKNOWLEDGEMENTS
I would like to thank C. Claussen for his identification of some Sphegina specimens.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 435
LITERATURE CITED Aktas, M. & Sarıbıyık, S. 1996. Contribution to the Syrphidae fauna of Turkey (Diptera: Syrphidae) (II), Milesiinae. Gazi Üniversitesi Fen Bilimleri Dergisi, 9 (1): 15-27. Hayat, R. 1997. Sphegina (Sphegina) alaoglui, a new hover-fly from north-eastern Turkey (Diptera: Syrphidae). Zoology in the Middle East, 14: 109-113. Reemer, M. & Smit, J. 2007. Some hoverfly records from Turkey (Diptera: Syrphidae), Volucella, 8: 135-146. Rotheray, G. E. 1993. Colour guide to hoverfly larvae (Diptera, Syrphidae). Dipterists Digest, 9: 1-156. Sarıbıyık, S. 2003a. The evaluation of the works on Syrphidae (Diptera) fauna in the Western Blacksea Region. Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (2): 461-466. Sarıbıyık, S. 2003b. Kastamonunun Syrphid Sinekleri (Diptera: Syrphidae) II. Kastamonu Kültür Sempozyumu (18-20 Eylül 2003), 707-719. Sarıbıyık, S. 2009. Syrphidae Fauna of Ballidağ (Diptera). Kastamonu Üniversitesi Kastamonu Eğitim Dergisi, 17 (2): 707-714. Speight, M. C. D. 2008. Species accounts of European Syrphidae (Diptera) 2008. In: Speight, M. C. D., Castella, E., Sarthou, J. P. & Monteil, C. (eds). Syrph the Net, the database of European Syrphidae, 55: 262 pp.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 436
FLOWER FLIES OF ISPEREKHAN VALLEY WITH THE FIRST RECORD OF CHALCOSYRPHUS CURRAN, 1925 FROM IRAN
(DIPTERA: SYRPHIDAE)
Samad Khaghaninia* * Dept. of Plant Protection, Faculty of Agriculture, University of Tabriz, 51664, Tabriz, IRAN. E-mail: [email protected] [Khaghaninia, S. 2011. Flower flies of Isperekhan Valley with the first record of Chalcosyrphus Curran, 1925 from Iran (Diptera: Syrphidae). Munis Entomology & Zoology, 6 (1): 436-445] ABSTRACT: Syrphid fauna of Isperekhan valley in Iran was studied during 2008- 2010. The specimens were collected using common entomological net in nineteen localities. In total 315 collected specimens, 37 species belonged to 19 genera and two subfamilies were verified that all of them are as new records for studied region and a species, Chalcosyrphus nemorum (Fabricius), 1805, is as new record for the Iran insect fauna. KEY WORDS: Fauna, Syrphidae, Isperekhan Valley, Iran.
Isperekhan valley is located in northern west of Sahand chain Mountains, the highest as well as biggest chain mountains in Azerbayjan, in East Azerbayjan province-Iran. This area is expanded in about 4.82 km long and UTM (Universal Transfer Mercator) coordinate system, X from 623648.34 to 625550.33 E; Y from 4178983.37 to 4181710.75 N and varying latitude from 2170 m to 2768 m. Ligvan River follows through this region which including various species of Umbelliferea, Astraceae and scattered fields of grain and alfalfa.
Syrphidae is one of the largest and most diverse families of the order Diptera with more than 6000 described species over the world. Among many interesting attributes is their famous precision at hovering. They have the ability to keep the body motionless in the air for quite a period of time during flight. This is the most significant character of these flies, coupled usually with their yellow banded abdomen which resembling bees or wasps. This group consists of small to medium flies 6- 18 mm long which can be distinguished by the special venation of the wing (spurious vein) (Kevan & Baker, 1983). Many species are important pollinators of flowering plants (Faegri & van der Pijl, 1979; Sarıbıyık, 2003). In addition, the immatures of numerous species are predators of destructive aphids and other pests (Gilbert, 1981). The Iran Syrphids have been studied in various localities by some taxonomists (Modarres Awal, 1997; Khiaban et al., 1998; Dousti, 1999; Gharali et al., 2000; Alichi et al., 2002; Gharali et al., 2002; Goldasteh et al., 2002; Sadeghi et al., 2002; Golmohammadi & Khiaban, 2004; Gilasian, 2005; Khaghaninia, 2010; Ehteshamnia et al., 2010; Gharali & Reemer, 2010; Khaghaninia et al., 2010a,b,c). Checklists of Iranian hover flies were listed by Peck (1988) and Dousti & Hayat (2006). The Syrphid fauna of Sahand‘s mountainsides particularly this area have not yet been studied.
MATERIAL AND METHODS
Studied specimens were collected monthly, during 2008- 2010. Flies were caught using sweeping entomological net in nineteen localities which are situated through all the working area (Fig. 1).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 437
After killing the collected specimens in cyanid bottle, they were placed in a desiccator (having water at its bottom) for about 24 h in order to soak and soften then pinned using 000, 00, 0, 1 and 2 mounted pins and their wings and legs set on appropriate setting boards to facilitate morphological studies and the others were put into tubes filled with 70% alcohol. For identification, the materials were examined under a Nikon (SMZ 1000) binocular microscope. The identification was done upto the specific level with the help of valid keys such as Vockeroth and Thompson (1987), Bei-Bienko (1988), Stubbs & Falk (2002), Lyneborg & Barkemeyer (2005) and Speight (2010). The distribution of the species was mostly given using Speight (2010).
RESULTS
Thirty seven species belonged to 19 genera and two subfamilies were
yielded by present study. All of the verified species are as new records for the studied area and a species which marked by an asterisk newly introduced to Iran fauna and totally listed as follows: Subfamily Syrphinae Chalcosyrphus Curran, 1925 Diagnostic characters: Metasternum hairy, male hind femur not arched. Tergites strongly transverse. Hind femur distinctly broader one third from tip than one third from base. *Chalcosyrphus nemorum (Fabricius, 1805) Syst. antl.: 192 (Milesia). Type-locality: ―Austria‖. Material examined: 1 specimen (1♂): Isperekhan valley; 37°46' N 46°24' E, 2504 m , 8 Aug. 2009 (Collected by S. Khaghaninia, Deposited at Insect Museum of Tabriz University). Diagnostic characters: having two pairs of pale spots on the abdomen, both sexes are very short-bodied, the abdomen being little longer than the thorax, the tergites are strongly transverse. In the males, the orange spots are square, separated by a thick black cross whose limbs are of equal length. The female markings are generally less sharp and normally grayish-orange or grey (Hippa, 1978; Chandler, 1998) (Fig. 2). Distribution: southern Finland, Ireland, Britain and the Atlantic seaboard of Europe from Denmark to the Pyrenees and northern Spain, Also in Switzerland, in central Europe, Turkey (Saribiyik & Ozgur, 2000). New record for the Iran insect fauna. Flowers visited: white umbellifers; Anemone nemorosa, Caltha, Euphorbia amygdaloides, Potentilla erecta, Ranunculus, Rubus idaeus, Sorbus aucuparia, Taraxacum (Speight, 2010). Chrysotoxum elegans Loew, 1841 Stettin. ent. Ztg, 2: 140 (Chrysotoxum). Type locality: ―Wien‖ (Austria). Material examined: 3 specimens (2♂♂, 1♀). Iranian Records: Oromieh (Khiaban et al., 1998; Khiaban & Parchami, 2001), Iran (Amirimoghadam & Sirjani, 2004). Distribution: Fennoscandia south to Iberia and the Mediterranean; through central and southern Europe into European parts of Russia as far as the Caucasus Mountains and into Turkey, Iran. Chrysotoxum festivum (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 593 (Musca). Type locality: ―Europa‖. Material examined: 3 specimens (1♂, 2♀♀). Distribution: Fennoscandia south to Iberia and the Mediterranean, including N Africa; from Ireland eastwards through much of Europe, Great Britain, Norway, Sweden, Finland, Spain, Italy, Yugoslavia, Bulgaria, into Turkey and European parts of Russia; through Siberia to the Pacific coast; Japan; northern India, Iran.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 438
Chrysotoxum vernale Loew, 1841 Stettin. ent. Ztg, 2: 138 (Chrysotoxum).Type-locality: not given (―hier gefangen‖) [= environs of Poznan] (Poland). Material examined: 7 specimens (2♂♂, 5♀♀). Distribution: Fennoscandia south to the Pyrenees; from Britain (southern England) eastwards through most of Europe into Asia almost to the Pacific; Iran. Chrysotoxum verralli (Collin, 1940) Entomologist‘s mon. Mag., 76: 155 (Chrysotoxum, for Chrysotoxum octomaculatum: Verrall, 1901: British flies, 8: 647, not Curtis, 1837; misidentiflcation). Type localities ―at Harpenden (Herts.), from Timworth (Suffolk), Chippenham Fen and Fleam Dyke (Cambs.), and Fowl Mere near Wretham (Norfolk)‖ (Great Britain). Material examined: 1 specimen (1♀). Distribution: Denmark south to central France; Britain (Wales and central/southern England) eastwards through central Europe into European parts of Russia to the Caucasus and on into eastern Siberia, Iran. Episyrphus balteatus (De Geer, 1776) Mém. Ins., 6: 116 (Musca). Type locality: not given (Sweden). Material examined: 11 specimens (3♂♂, 8♀♀). Distribution: Fennoscandia to the Mediterranean; Canary Isles, Azores and N Africa; Ireland through Eurasia to the Pacific coast; south through the Oriental region to Sri Lanka; Australia. This is an extremely migratory species and records from offshore islands of northern Europe, such as the Faroes (Jensen, 2001) are assumed to be due to annual immigration, rather than indicative of the occurrence of resident populations, Iran. Eupeodes corollae (Fabricius, 1794) Entom. Syst., 4: 306 (Syrphus). Type locality: Kilia [=Kiel] [Germany]. Material examined: 3 specimens (3♀♀). Distribution: from Iceland, Fennoscandia and the Faroes (Jensen, 2001) south to Iberia, the Mediterranean, Madeira, the Canary Isles and N Africa; coastal States of Africa down to and including S Africa; Mauritius; from Ireland eastwards through most of Europe into European parts of Russia; through Siberia from the Urals to the Pacific coast; Japan; China; Formosa, Iran. Eupeodes latifasciatus (Macquart, 1829) Mém. Soc. Sci, Agric. Lille,1827-1828: 242 and Ins. Dipt., 4: 94 (Syrphus). Type locality: ‘‘Arras‘‘ (France). Material examined: 1 specimen (1♂). Distribution: From Iceland and Fennoscandia south to Iberia, the Mediterranean (including Cyprus), N Africa and Turkey; from Ireland eastwards through most of Europe into European parts of Russia; through Siberia from the Urals to the Pacific coast (Sakhalin and Kuril Isles); India; in N America from Alaska south to California and Texas, Iran. Eupeodes nuba (Wiedemann, 1830) Aussereurop. Zweifl. Insekt., 2: 136 (Syrphus). Type locality: ‗‘Nubien‘‘ (Sudan). Material examined: 2 specimens (1♂, 1♀). Distribution: Canary Isles, Mediterranean basin, from southern France to Italy (Sicily) and parts of the former Yugoslavia, Crete, Cyprus, Lebanon, Israel, Egypt and Morocco; Switzerland in central Europe, Roumania; Transcausasus and south-western parts of Asia (Uzbekistan, Kirghizistan, Tajikistan) to Afghanistan and Mongolia. In eastern parts of the Afrotropical region from Ethiopia south to S Africa (inclusive), Iran. Melanostoma mellinum (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 593 (Musca). Type-locality: Svecia (Sweden). Material examined: 39 specimens (15♂♂, 24♀♀). Distribution: From Iceland and Fennoscandia south to Iberia, the Mediterranean and North Africa; from Ireland eastwards through most of Europe into European parts of Russia;

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 439
Siberia from the Urals to the Pacific coast; North America from Alaska to Quebec and south to Washington, Iran. Melanostoma scalare (Fabricius, 1794) Entom. Syst., 4: 308 (Syrphus). Type- locality: ―Kiliae‖ [= Kiel] [D]. Material examined: 5 specimens (3♂♂, 2♀♀). Distribution: From Iceland and Fennoscandia south to Iberia, the Mediterranean and North Africa; from Ireland eastwards through most of Europe into European parts of Russia; in Siberia from the Urals to the Pacific coast (Kuril Isles); in eastern parts of the Afrotropical region south to Zimbabwe; throughout the Oriental region to New Guinea, Iran. Paragus bicolor (Fabricius, 1794) Entom. Syst., 4: 297 (Syrphus). Type locality: ―Barbariae‖ [= NW Africa]. Material examined: 4 specimens (1♂, 3♀♀). Distribution: from southern Sweden and Denmark (extinct in Belgium) south to the Mediterranean and North Africa; from France eastwards through central and southern Europe to Mongolia; Iran and Afghanistan; North America, Iran. Paragus compeditus Wiedemann, 1830 Aussereurop. Zweifl. Insekt., 2:89 (Paragus). Type locality: ―Egypten‖ (Egypt). Material examined: 5 specimens (2♂♂, 3♀♀). Distribution: Italy, USSR-South European territory, Transcaucasus, Kazakhstan, Soviet Middle Asia, Afghanistan, North China, Egypt (Peck, 1988) and Turkey (Hayat and Claussen, 1997) to North Africa and the Afrotropical region, Iran. Paragus tibialis (Fallén, 1817) Syrphici Sveciae: 60 (Pipiza). Type locality: in Vestrogothia; in arvis montosis Scaniae [=prov. Vastergotland and prov. Skane] (Sweden). Material examined: 5 specimens (2♂♂, 3♀♀). Distribution: Uncertain at present, due to confusion with other species until recently; apparently occurs from southern Norway, Sweden and Denmark south to the Mediterranean coast of Europe, North Africa and the Canary Isles; from Britain (southern England) eastwards through central and southern Europe to the former Yugoslavia, Turkey, Israel, Nearctic and Oriental Regions, Iran. Scaeva pyrastri (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 594 (Musca). Type locality: Svecia (Sweden). Material examined: 5 specimens (3♂♂, 2♀♀). Distribution: Fennoscandia south to Iberia, the Mediterranean, Canary Isles and North Africa; from Ireland east through much of Europe and Asia Minor into European Russia; through Siberia from the Urals to the Pacific coast (Kuril Isles); India; China; North America from Alaska to California and New Mexico, Iran. Scaeva albomaculata (Macquart, 1842) Mém. Soc. Sci. Agric. Lille, 1841(1): 146 and Dipt. exot., 2(2): 86 (Syrphus). Type localities:‘‘Mont-sinai‘‘ (Egypt).‘‘Alger‘‘ (Algeria). Material examined: 4 specimens (1♂, 3♀♀). Distribution: Iberian peninsula and round the Mediterranean basin to Morocco; Canary Islands; eastward through southern Russia, the Caucasus and southern Siberia to the far east and northern China; Afghanistan, Mongolia; highly migratory and occasionally reaches as far north as Britain, Iran. Sphaerophoria rueppelli (Wiedemann, 1830) Aussereurop. zweifl. Insekt., 2: 141 (Syrphus). Type locality: Nubien; Abyssinia (lectotype des. Vockeroth, 1971:1633). Material examined: 34 specimens (16♂♂, 18♀♀). Distribution: from southern Norway and Sweden south to N Africa and the Canary Isles; from Ireland east through central and southern Europe, including Greece, Turkey and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 440
Mediterranean islands into Asia Minor, Russia and Afghanistan and on to the Pacific coast, China and Korea; in eastern parts of the Afrotropical region south to Kenya, Iran. Sphaerophoria scripta (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 594 (Musca). Type- locality: ‗‘Svecia‘‘ (Sweden); ‗‘Uppsala, Sweden‘‘ (lectoype des. Vockeroth, 1971: 1633). Material examined: 11 specimens (5♂♂, 6♀♀). Distribution: A highly migratory species; southwest Greenland, Iceland and Fennoscandia south to the Mediterranean, the Canary Isles and N Africa; from Ireland eastwards through much of the Palaearctic to the Pacific coast of Asia; Kashmir and Nepal, Iran. Sphaerophoria turkmenica Bankowska, 1964 Annls zool., Warsz., 22(15): 345 (Sphaerophoria). Type locality: ‗‘Turkmenische SSR, West Kopet Dag, Berg Siunt‘‘. Material examined: 4 specimens (2♂♂, 2♀♀). Distribution: parts of European Russia; the Caucasus (Armenia, Azerbaijan); Arabian peninsula (Oman),Turkmenistan; Kazakhstan; Turkey (Hayat & Alaoglu, 1990); Iran. Syrphus ribesii (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 593 (Musca). Type locality: Svecia. (Sweden). Material examined: 7 specimens (4♂, 3♀♀). Distribution: From Iceland and Fennoscandia south to Iberia and the Mediterranean; Canary Isles; from Ireland eastwards through most of Europe into Turkey, European parts of Russia and Afghanistan; from the Urals to the Pacific coast (Kuril Isles); Japan; North America from Alaska south to central parts of the USA, Iran. Xanthogramma pedissequum (Harris, 1776) Expos. Eng. Ins.: 61, tab. XV, fig. 19 (Musca). Type locality: not given (England). Material examined: 4 specimens (2♂♂, 2♀♀). Distribution: Uncertain, due to confusion with related species, but from from Britain and Atlantic seabord countries south to the Paris basin and into central Europe to the Alps (France, Switzerland); Iran.
Subfamily Milesiinae Chrysogaster basalis Loew, 1857 Wien. Ent. Mschr., 1: 5 (Chrysogaster). Type-locality: ―Deutschland‖ (Germany). Material examined: 3 specimens (1♂, 2♀♀). Distribution: Europe: from northern France and southern Germany south to Spain and Portugal and in N Africa; Switzerland; Roumania and much of the Balkan Peninsula (Bosnia-Herzegovina, Croatia, Greece, Macedonia, Serbia). Assuming musatovi is the same species as basalis, then C.basalis also occurs in the Ukraine, the Caucasus and on into Kazakhstan and Tajikistan; Iran (Khaghaninia et al., 2010a). Chrysogaster viduata (Linnaeus, 1758) Syse. Nat., Ed. 10, 1: 598 (Musca). Type-locality: ―Europa‖. Material examined: 3 specimens (1♂, 2♀♀). Distribution: Norway, Sweden, Finland, Denmark, The Netherlands, Belgium, Luxembourg, Germany, Poland, Czech Republic and Slovakia, France, Austria, Hungary, Spain, Italy, the former Yugoslavia, Bulgaria, USSR-Central and South European territories, Transcaucasus, Iran. Cheilosia aerea Dufour, 1848 Material examined: 4 specimens (3♂♂, 1♀). Distribution: Poland south to the Mediterranean; from the Netherlands eastwards through much of central and southern Europe into European parts of Russia as far as the Transcaucasus; Iran (Khaghaninia et al., 2010a).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 441
Cheilosia proxima (Zetterstedt, 1843) Dipt. Scand., 2:792 (Eristalis). Type- locality: ―in Ostrogothia …; ad Haradshammar‖ (Sweden). Material examined: 8 specimens (5♂♂, 3♀♀). Distribution: Fennoscandia south to Pyrenees and mountainous parts of Spain; Britain eastwards through much of Europe into Turkey and European parts of Russia; in Siberia from the Urals to Kamchatka; Iran. Eristalinus aeneus (Scopoli, 1763) Ent. Carniolica: 356 sex?; (Conops).Type locality: Idria (Yugoslavia). Material examined: 19 specimens (8♂♂, 11♀♀). Distribution: Cosmopolitan; southern Sweden south to N Africa and the Canary Isles; on into the Afrotropical region south to Kenya and Tanzania; from Ireland eastwards through central and southern Europe and on through Russia and China to the Pacific and south into the Oriental region; Mauritius; in North America from Minnesota and Ontario south to California and Texas; Hawaii, Australia and the Gilbert and Ellis islands in Australasia; Bermuda, Iran. Eristalis arbustorum (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 591 (Musca).Type locality: Europa. Material examined: 29 specimens (12♂♂, 17♀♀). Distribution: Throughout the Palaearctic region, including North Africa; North America from Wisconsin to Labrador and south to Kansas and South Carolina; reaches the Oriental region in northern India; Iran. Eristalis tenax (Linnaeus, 1758) Syst. Nat., Ed. 10, 1: 591 (Musca).Type locality: Svecia (Sweden). Material examined: 4 specimens (1♂, 3♀♀). Distribution: highly migratory; cosmopolitan; the most widely distributed syrphid species in the world, known from all regions except the Antarctic; found throughout Europe except in the far north. It occasionally reaches offshore islands of northern Europe, such as the Faroes; Iran. Eumerus sogdianus Stackelberg, 1952 Trudy zoll Inst., 12: 390 (Eumerus). Type–locality: Tajikistan: Stalinabad [=Dushanbe] calley of the r. Kafernighan. Material examined: 3 specimens (1♂, 2♀♀). Distribution: Denmark south to southern Spain; from Belgium eastwards through central and southern Europe into European parts of Russia and on into central Asia (Kazakhstan, Tajikistan, Uzbekistan, Mongolia); China. The presence of this species in Western Europe has only been recognised recently, Iran. Eumerus strigatus (Fallén, 1817) Syrphici Sveciae: 61 (Pipiza). Type locality: Vestrogothia; et in Scania [=prov. Vastergotland and prov. Skane] (Sweden). Material examined: 1 specimen (1♂). Distribution: Fennoscandia south to Iberia and the Mediterranean; much of Europe through into Turkey and Russia; from the Urals to the Pacific coast (Sakhalin); Japan; introduced to North America and recorded from both Canada and the USA; introduced to both Australia and New Zealand; Iran. Merodon aberrans Egger, 1860 Verh. Zool. – bot. Ges. Wien, 10: 664 (Merodon). Type locality: ―Austria‖. Material examined: 2 specimens (1♂, 1♀). Distribution: Through central Europe from Germany, the Czech Republic and the Alps (France, Switzerland, Austria) to Hungary and Roumania and on to the Ukraine and southern Russia; in southern Europe from mountainous parts of Portugal and Spain eastwards to Italy, Albania, the former Yugoslavia and Greece and on to Turkey and round

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 442
the Mediterranean (Lebanon) into N Africa (Morocco), including Mediterranean islands e.g. Crete.; Iran (Khaghaninia et al., 2010a). Merodon warnckei Hurkmans, 1993 Tijdschrift voor Entomologie, 136: 147-234. Type loclity: ―Hakkari and Van‖ (Turkey). Material examined: 1 specimen (1♀). Distribution: Turkey, Iran. Pipizella caucasica Skufjin, 1976 Ent. Obozr., 55(4): 932 (Pipizella).Type locality: plateau Lagonaki, North Caucasus. Material examined: 14 specimens (6♂♂, 8♀♀). Distribution: Caucasus (Georgia) and north-east Turkey; Iran. Pipizella divicoi (Goeldlin de Tiefenau, 1974) Mitt. Schweiz. Ent. Ges., 47 (3- 4): 238 (Heringia). Type locality: ―Roschenz (Jura BE)‖ (Switzerland). Material examined: 18 specimens (5♂♂, 13♀♀). Distribution: From the Netherlands south to the Mediterranean coast of Spain; from Belgium and northern France (Rhine valley) eastwards through central and southern Europe (Italy, the former Yugoslavia) to Turkey and European parts of Russia and on through Siberia to the Pacific coast; Mongolia. This species disappears with introduction of irrigation to its dry grassland habitat; Iran (Khaghaninia et al., 2010a). Syritta flaviventris Macquart, 1842 Mém. Soc.Sci. Agric. Lille, 1841 (1): 135 and Dipt. Exot., 2 (2): 75 (Syritta). Type locality: Senegal (W Africa). Material examined: 9 specimens (7♂♂, 2♀♀). Distribution: Portugal, Spain and round the Mediterranean from the southern coast of France to Turkey and on to north Africa; in eastern parts of the Afrotropical region to the southern tip of Africa and in Madagascar. Known from various Mediterranean islands: Corsica, Sardinia, Sicily, Crete, plus Cape Verde. Also cited from Argentina, Brazil and Chile in the Neotropical region, Mexico and Texas (USA) in the Nearctic and Easter Island in Oceania (to which it has supposedly been introduced); Iran. Syritta pipiens (Linnaeus, 1758) Syst. Nat., Ed.10, 1: 594 (Musca).Type locality: Europa. Material examined: 20 specimens (8♂♂, 12♀♀). Distribution: Becoming cosmopolitan; known from most of the Palaearctic, including North Africa, most of North America, South America and the Oriental region. But records from the Afrotropical region are apparently erroneous; Iran. Spazigaster ambulans (Fabricius, 1798) Suppl. Entom. Syst.: 562 (Syrphus). Type-locality: ―Italia‖. Material examined: 18 specimens (11♂♂, 7♀♀). Distribution: Schwarzwald (Germany), Alps, Apennines, Hungary, northern parts of the former Yugoslavia, the Carpathians, the Caucasus, Turkey, Iran (Gharali and Reemer, 2010 and Khaghaninia et al., 2010d).
ACKNOWLEDGEMENTS
This work acknowledged of Tabriz University for financial supporting by the Research Grant Contract No. 27.557. The author greatly appreciates kind assistance of Dr. Suleyman Saribiyik (Education Faculty, Kastamonu University, Turkey) for determination the part of studied material.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 443
LITERATURE CITED Alichi, M., Asadi, G. H. & Gharali, B. 2002. Aphidophagus syrphids of Fars province. Proceedings of 14th Iranian Plant Protection Congress, p. 181. Bei- Bienko, G. 1988. Keys to the insects of the European part of the USSR. Volume V. Diptera and Siphonaptera. Part II. Smithonian Institution Libraries and the National Science Foundation Washington, D.C. 10- 148. Chandler, P. J. 1998. Checklists of insects of the British Isles (New Series) part I: Diptera. Hand book for Identification of British Insects, 12 (1): 1-234. Dousti, A. F. 1999. Fauna and Diversity of Syrphid flies in Ahwaz region. M.S. Thesis, Shahid-Chamran University, 129 pp. Dousti, A. F. & Hayat, R. 2006. A catalogue of the Syrphidae (Insecta: Diptera) of Iran. J. Entomol. Res. Soc., 8 (3): 5-38. Ehteshamnia, N., Khaghaninia, S. & Farshbaf Pour Abad, R. 2010. Some hover flies of subfamily Syrphinae of Qurigol fauna in East Azerbayjan province, Iran. Mun. Ent. Zool., 5 (2): 499-505. Faegri, K. & van der Pijl, L. 1979. The principles of pollination ecology. Pergamon, Oxford, England. Gharali, B. & Reemer, M. 2010. Extension of the known range of the genus Spazigaster (Diptera: Syrphidae) to Iran. North-Western Journal of Zoology, 6 (1): 140-143. Gharali, B., Alichi, M. & Radjabi, G. R. 2000. The new records of syrphid flies (Dip.: Syrphidae). Proceeding of the 14th Iranian Plant Protection Congress, p. 348. Gilasian, E. 2005. New record of one genus and six species of Syrphidae (Diptera) from Iran. Journal of Entomological Society of Iran, 25 (1): 75-76. Gilbert, F. S. 1981. Foraging ecology of hoverflies: Morphology of the mouthparts in relation to feeding on nectar and pollen in some common urban species. Ecol. Entomol., 6: 245-262. Goldasteh, Sh., Bayat Asadi, H., Shojaee, M. & Baniameri, V. A. 2002. A faunistic survey of Syrphidae (Diptera) in Gorgan region. Proceeding of the 15th Iranian Plant Protection Congress, p. 168. Golmohammadi, Gh. & Khiaban, N. 2004. Hoverflies (Diptera: Syrphidae) fauna of wheat fields in Sistan region. Proceedings of 16th Iranian Plant Protection Congress, p. 132. Hayat, R. & Alaoðlu, O. 1990. Fauna of Syrphidae (Diptera) in the vicinity of Erzurum (Part I). Syrphinae. Türk Entomoloji Dergisi, 14 (3): 173-182. Hayat, R. & Claussen, C. 1997. A new species and new records of the genus Paragus Latreille, 1804 from Turkey (Diptera: Syrphidae). Zoology in the Middle East, 14: 99-108. Hippa, H. 1978. Classification of Xylotini (Diptera, Syrphidae). Acta. Zool. Fenn., 156: 1-153. Kevan, P. G. & Baker, H. G. 1983. Insects as flower visitors and pollinators. Annu. Rev. Entomol., 28: 407-453. Khaghaninia, S. 2010. Faunistic study on flower flies of Zunuz region in East Azerbayjan province- Iran. Mun. Ent. Zool., 5 (2): 586-593. Khaghaninia, S., Farshbaf Pour Abad, R. & Hayat, R. 2010a. Seven species as new records for haver flies fauna of Iran (Diptera; Syrphidae) from Qaradag forests. Mun. Ent. Zool., 5 (1): 307-308. Khaghaninia, S., Farshbaf Pour Abad, R. & Ehteshaminia, N. 2010b. Some of hover flies fauna of subfamily Milesinae (Diptera: Syrphidae) of Qurigol in East Azerbaijan Province, Northwest Iran. Munis Entomology and Zoology, 5: 911-916. Khaghaninia, S., Jafarlu, M., Khiaban, N. G. & Askari, O. 2010c. Introduction to hover flies (Diptera; Syrphidae) of sunflower and pumpkin fields in West Azerbayjan province- Iran. Mun. Ent. Zool., 5 (1): 270- 277.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 444
Khaghaninia, S., Pour Abad, R. & Saribiyik, S. 2010d. Four genera and seven species as new records for Iran hover flies fauna (Diptera: Syrphidae) from East Azarbayjan province. Proceeding of IXth European Congress of Entomology, pp: 232. Khiaban, N. G., Hayat, R., Safaralizadeh, M. & Parchami, M. 1998. A faunistic survey of Syrphidae in Uromieh region. Proceeding of the 13th Iranian Plant Protection Congress, p. 231. Lyneborg, L. & Barkemeyer, W. 2005. The genus Syritta: A world revision of the genus Syritta. Volume 15. Apollo Books Pub. Modarres Awal, M. 1997. Syrphidae; pp. 253-254. In: List of agricultural pests and their natural enemies in Iran. Ferdowsi Univ. Press, 429 pp. Peck, L. V. 1988. Family Syrphidae. PP. 11- 230 in Soos, A. (Ed.) Catalogue of Palearctic Diptera. Vol. 8, 363 PP. Akademiai Kiado, Budapest. Sadeghi, H., Kayvanfar, N. & Mojtahedzadeh, K. 2002. Hover flies (Dip.: Syrphidae) fauna of Mashhad region. Proceeding of the 15th Iranian Plant Protection Congress, p. 169. Saribiyik, S. 2003. Fauna of Syrphinae and Milesinae (Diptera: Syrphidae) around Tuz lake. Kastamonu Education Journal., 11 (2): 439- 450. Saribiyik, S. & Ozgur, A. F. 2000. New records of Melesiinae (Diptera: Syrphidae) from Turkey. J. Ent. Res. Soc., 2 (3): 5-13. Speight, M. C. D. 2010. Species accounts of European Syrphidae (Diptera) 2010. In: Speight, M.C.D., Castella, E., Sarthou, J.-P. and Monteil, C. (eds.). Syrph the Net, the database of European Syrphidae, vol. 59, 285 pp., Syrph the Net publications, Dublin. Stubbs, A. E. & Falk, S. J. 2002. British hover flies. An illustrated identification guide. Pub. The british Entomology and Natural History Society, Reading, UK. Vockeroth, J. R. & Tompson, F. C. 1987. Syrphidae in: Manual of Nearctic Diptera. Biosystematic Research Center. Ottawa- Ontario, 2: 713- 742.
Figure 1. Location of sampling points on satellite image (SPOT) of Isperekhan valley.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 445
(a)
(b)
Figure 2. Chalcosyrphus nemorum, male, a: dorsal view, b: lateral view.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 446
A NEW RECORD FOR FLOWER FLIES FAUNA OF TURKEY MYOLEPTA VARA (PANZER, 1798) (DIPTERA: SYRPHIDAE)
Süleyman Sarıbıyık*
* Kastamonu Üniversitesi, Eğitim Fakültesi, 37200, Kastamonu, TÜRKĠYE. E-mail: [email protected] [Sarıbıyık, S. 2011. A new record for flower flies fauna of Turkey, Myolepta vara (Panzer, 1798) (Diptera: Syrphidae). Munis Entomology & Zoology, 6 (1): 446-448] ABSTRACT: Myolepta vara (Panzer), 1798, is recorded for the first time from Bolu and Turkey. One specimen was collected from Abant Lake Natural Park of Bolu, Turkey. KEY WORDS: Syrphidae, new record, fauna, Bolu.
Myolepta Newman, 1838 is a small genus of hoverflies with representatives in the Palaearctic, Nearctic, Neotropical, Afrotropical and Oriental regions. For the Palaearctic region 10 species of Myolepta are listed by Peck (1988), but there has remained confusion about the status and identity of some of these species. Besides, the differences between some species have been inadequately described in most literature (Reemer et al., 2004).
Peck (1988) lists seven European species of Myolepta. In the revision of Reemer et al. (2004) one of them, M. helvetica (Wainwright) is shown to be a synonym of M. vara. In the same publication M. trojana is described from south-western parts of Turkey (Speight, 2008).
Adults of Myolepta usually occur in mature deciduous or evergreen forests where suitable habits for the larvae (i.e. rotten and dead wood) are easily accessible. The only comprehensive review of the genus in the palaearctic region was out of date and included only four species from which M. ruficornis Zetterstedt, 1843 belongs to the genus Lejota Rondani, 1857. Reemer et al. (2004) revised the genus in the western part of the palaearctic region and described three new species namely M. carthaginiensis, M. mada and M. trojana. They also synonymised two species and designated a neotype for M. potens (Babak and Reemer, 2008).
The first record of Myolepta vara from Turkey is based on specimen which was collected from Bolu province (1♀ west of Bolu city, Abant Gölü Tabiat Parkı (Abant Lake Natural Park). Biology: Myolepta vara occurs in deciduous forests (Quercus) with over-mature trees, including ailuvial hardwood forest and oak-hornbeam (Quercus/Carpinus) forest. The adults feed on flowers ofvarious shrubs and trees. They have been observed sitting in patches of sunlight on shady, low places like bare ground, cow pats, tree trunks, leaves and even on asphalt. This behaviour, in combination with its cryptic colours probably contributes to the elusive reputation of the species. The larva has been found in decaying organic matter at the bottom of a small cavity in a recently felled Quercus trunk. The pupal stage of a reared specimen lasted for 18 days Flight period found from April l5th until June 23rd. The species is univoltine (Reemer et al., 2004).
Until now, four species of the genus Myolepta have been recorded from Turkey, by Turkish and foreign researchers. Myolepta dubia, M. nigritarsis, M. potens, M. trojana (Özgür, 1987; Sarıbıyık, 1999, 2001, 2003a,b,c,d, Reemer et al., 2005; Speight, 2008).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 447
MATERIAL AND METHODS
This specimen was collected from white flowered Umbelliferae found near the small water flows in the Fagus sp. (Fagaceae) Abies sp. and Pinus sp. (Pinaceae) forests in Abant Lake Natural Park of Bolu by the author (Leg. S. Sarıbıyık). The collected specimen was prepared and labelled following standart protocols for museum material. The following works were used for species identification: Sack, (1932), Bańkowska (1963) and Reemer et al. (2004).
The specimen is preserved in the private collection of the author. Two photos belonging to the specimen are given (Figs.1, 2).
RESULTS
Myolepta vara (Panzer, 1798) Material examined: 1 ♀, Bolu, Abant Lake Natural Park (Abant Gölü Tabiat Parkı), 1120 m., 19.VII.1995. Flower visited: white flowered Umbelliferae. Range: the Netherlands south through Belguim (extinct?) and France south to the Mediterranean and central Spain; through central and South Europe to Switzerland, Austria and Roumania and on to the Balkans and the Caucasus (Azerbaijan), also, supposedly, from the Pacific coastal region of Asiatic Russia. This species appears to be scarce over most of its European range (Speight, 2008). This is a new record for the Turkish fauna.
LITERATURE CITED Bańkowska, R. 1963. Klucze do oznaczania owadów Polski. Czesc XXVII. Muchówki– Diptera, zeszyt 34 – Syrphidae, 236 pp. Gharali, B. & Reemer, M. 2008. The first record of the genus Myolepta Newman, 1838 (Diptera: Syrphidae) from Iran. North-Western Journal of Zoology, 4 (2): 331-334. Özgür, A. F. 1987. Akdeniz Bölgesi kıyı Ģeridi Syrphidae (Diptera) faunası II. Altfamilya: Cheilosiinae, Sphegininae, Pelecocerinae, Milesiinae, Chysotoxinae, Volucellinae, Çukurova Universitesi. Ziraat. Fakültesi Yayinları, 2 (3): 67-81. Peck, L. V. 1988. Family Syrphidae, pp. 11-230. In, Soos, A and Papp, L. (Editors). Catalogue of Palaeartic Diptera, 8: 363 pp. Reemer, M., Hauser, M. & Speight, M. C. D. 2004. The genus Myolepta Newman in the West-Palaearctic region. Studia Dipterologica, 11 (2): 553-580. Sack, P. 1928-1932. Syrphidae. In, Lindner, E. Die Fliegen der paläarktischen Region, IV (6). Stuttgart, Germany. +451 pp. 18 pls. Sarıbıyık, S. 1999. Milesiinae Fauna of the West Blacksea Region (Diptera: Syrphidae). Kastamonu Education Journal, 7 (1): 195-204. Sarıbıyık, S. 2001. New Records of the Subfamily Milesiinae (Diptera: Syrphidae) from Turkey. Journal of the Entomological Research Society, 3 (3): 43-51. Sarıbıyık, S. 2003a. Fauna of Syrphinae and Milesiinae (Diptera: Syrphidae) Around Tuz Lake. Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (2): 439-450. Sarıbıyık, S. 2003b. The Evaluation of the Works on Syrphidae (Diptera) Fauna in the Western Blacksea Region. Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (2): 461-466.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 448
Sarıbıyık, S. 2003c. Milesiinae Fauna of the KahramanmaraĢ‘s Andırın Town (Diptera: Syrphidae). Gazi Üniversitesi Kastamonu Eğitim Dergisi, 11 (1): 159-164. Sarıbıyık, S. 2003d. Kastamonunun Syrphid Sinekleri (Diptera: Syrphidae). II. Kastamonu Kültür Sempozyumu (18-20 Eylül 2003), 707-719. Speight, M. C. D. 2008. Species accounts of European Syrphidae (Diptera) 2008. in: Speight M. C. D., Castella E., Sarthou J. P., Monteil, C. (eds). Syrph the Net, the database of European Syrphidae, 55: 262 pp., Syrph the Net publications, Dublin.
Figure 1. Myolepta vara; female adult at dorsal view.
Figure 2. Myolepta vara; female adult at lateral view.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 449
TRITROPHIC INTERACTIONS OF NINETEEN CANOLA CULTIVARS- CHROMATOMYIA HORTICOLA –
PARASITOIDS IN ARDABIL REGION
Seyed Ali Asghar Fathi* * Department of Plant Protection, University of Mohaghegh Ardabili, Ardabil, IRAN. E-mails: [email protected]; [email protected] [Fathi, S. A. A. 2011. Tritrophic interactions of nineteen canola cultivars- Chromatomyia horticola - parasitoids in Ardabil region. Munis Entomology & Zoology, 6 (1): 449-454] ABSTRACT: The leafminer, Chromatomyia horticola (Goureau) (Diptera: Agromyzidae), is one of the most important pest of canola, Brassica napus L., crop in Ardabil region. In this research, the parasitoid species of the leafminer larvae were collected and identified in the experimental field of canola in the University of Mohaghegh Ardabili during 2008 and 2009. Then the percentage of abundance of each of parasitoid species was determined in the experimental field. In 2008 and 2009, Diglyphus isaea (Walker) had the highest abundance (48.3% and 42.8%, respectively) and Chrysocharis Pubichornis (Zetterstedt), Dacnusa sibirica Telenga and Pediobius sp. had lower abundance, respectively. Also, the density of larvae and the percentage of parasitized larvae were determined on nineteen canola cultivars in the experimental field. The density of larvae was lower on Opera, Hyola401 and RGS003 than on the other tested cultivars during 2008 and 2009. The percentage of parasitized larvae on Opera, hyola401 and RGS003 was significantly higher than on the other tested cultivars. Therefore, it could be concluded that using of Opera, Hyola401 and RGS003 cultivars might be increased the efficiency of parasitoids in control of the larvae of C. horticola. KEY WORDS: Chromatomyia horticola, parasitoid, identification, canola cultivars, interaction, Ardabil.
The leafminer, Chromatomyia horticola (Goureau) (Diptera: Agromyzidae) is an important pest of canola, Brassica napus L., crop in Ardabil region, Iran (Saljoqi et al., 2006; Fathi, 2010). This insect is polyphagous with worldwide distribution (Gençer, 2005). The leafminer females create leaf punctures by ovipositior for feeding or oviposition. Adult females insert their eggs into the leaf tissue. The larva consumed mesophyll parenchyma tissue and creates continuous mines in leaves. In addition to reducing the level of photosynthesis in the infested plant, leafminers have also been shown to transmit plant pathogens (Parrella et al., 1985; Parrella, 1987; Capinera, 2008).
The parasitoid species of C. horticola have been reported in previous studies (Gençer, 2005). The leafminer parasitoids mainly belong to Braconidae, Eulophidae and Pteromalidae. Eulophids include more than 80% of leafminer parasitoids (Gençer, 2004, 2005; LaSalle & Parrella, 1991). Use of biocontrol agents has an important role in the IPM of leafminers. But, the control level by biocontrol agents is not sufficient to achieve adequate control of leafminers (Capinera, 2008). Therefore, further research is required to investigate the potential of using resistant cultivars in combination with biocontrol agents in the integrated management of leafminers. Therefore, study of interactions of canola cultivars - C. horticola - parasitoids can potentially improve pest management programs.
Canola cultivars, namely Talayh, Jewel, PF/7045/91, Licord, SIM043, SIM046, Elvis, Zarfam, RGS003, Opera, Okapi, Ebonite, Elite, Orient,Option500, Hyola60, Hyola308 Hyola401, and Adder have recently been introduced in Ardabil. Currently, Zarfam, Okapi and Ebonite are being planted on

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 450
vast acreage compared with other cultivars tested in this research. In an earlier research (Fathi, 2010) the resistance of some canola cultivars was evaluated to C. horticola. But, our literature review indicated that no study have been done on interactions of canola cultivars - C. horticola – parasitoids. Therefore, the purpose of this research is (a) identify of the parasitoid species of C. horticola in canola field in Ardabil region and (b) study of tritrophic interactions of C. horticola – parasitoids on studied canola cultivars.
MATERIAL AND METHODS Preparation of the experimental field
The experimental field in a 1000 m2 was selected in the Agricultural Research Station of the University of Mohaghegh Ardabili, Ardabil, Iran (elevation of 1332 m; longitude 48º 17' E; latitude 38º 15' N) in 2008 and 2009. Seeds of cultivars, namely Talayh, Jewel, PF/7045/91, Licord, SIM043, SIM046, Elvis, Zarfam, RGS003, Opera, Okapi, Ebonite, Elite, Orient, Option500, Hyola60, Hyola308, Hyola401, and Adder were obtained from the Seed and Plant Improvement Institute of Karaj (Iran). All tested cultivars were planted in October-2007 and 2008 to vernalize during winter in a field, following a randomized complete block design with four blocks for each cultivar. The distance of 2 m between blocks was selected for sampling. The row to row and plant to plant distances were maintained at 75 cm and 20 cm, respectively. The field was managed according to the local practice with weekly flood irrigation and hand weeding. No insecticides were applied to the plants. Dithane fungicide (Mancozeb, Rohm and Haas Co., Philadelphia, PA) was used to prevent foliar disease before inflorescence emergence. Also, nitrogen fertilizer (100 kg ha-1) was used in rosette stage in may-2008 and 2009. Identification of parasitoid species and their abundance
In experimental canola field, the leaves infested to leafminer larvae observed on plants of each cultivar in each block were collected and transferred to the laboratory. These larvae were maintained in the growth chamber at 23°C±1 and 50±5 RH and 14:10 h (L:D) until adult emergence of wasps or leafminer. The species of parasitoid of leafminer larvae in each sample were separated and identified under stereomicroscope by their morphological characters (Darvas et al., 1999; Noyes, 2003; Gençer, 2004, 2005, 2009). Then the number of each of parasitoid species in each sample was recorded. These data was used for calculation of the percentage of abundance of each species. The larval densities and parasitism rates
The density of C. horticola larvae and the percentage of parasitized larvae of leafminer were investigated on 19 cultivars of canola in the experimental canola field. Sampling on each cultivar was carried out every three days starting from the stem extension stage and continued until the late seed development stage between 10:00 and 11:00. On each sampling date, 4 plants from each of the 19 cultivars per block were selected randomly and the number of mines contains leafminer larvae per leaf were recorded using a 20X hand lens. The leaves infested to leafminer larvae observed on 4 plants of each cultivar in each block were collected and transferred to the laboratory. These larvae were maintained in the growth chamber at 23°C±1 and 50±5 RH and 14:10 h (L:D) until adult emergence of wasps or leafminer. The numbers of adult C. horticola and parasitoid species were recorded to determine the percentage of parasitized larvae in the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 451
experimental field. Also, mines without leafminer larvae were not account in calculation of the percentage of parasitized larvae. One of the suitable indicators for evaluating the performance of parasitoids on different plant cultivars is comparison of rate of parasitized larvae of leafminer on different cultivars of canola (Rauf et al., 2000; Civelek, 2002; Chen et al., 2003; Gençer, 2004, 2005, 2009). Data analysis
Prior to analysis, data were log-transformed to correct the heterogeneity of variance whereas data on parasitism rate were arcsine-transformed. Data on the abundance of each of parasitoid species were analyzed using one-way ANOVA. Data on larval densities and rates of parasitism in two years were analyzed by a combined analysis, completely randomized block design in two years (PROC GLM, SAS Institute, 2005). The differences among treatment means were compared using Tukey's HSD test (PROC ANOVA, SAS Institute, 2005).
RESULTS Identification of parasitoid species and their abundance
In this research, Diglyphus isaea (Walker), Chrysocharis pubicornis (Zetterstedt) and Pediobius sp. (Hym.: Eulophidae), and Dacnusa sibirica Telenga (Hym.: Braconidae) were identified as parasitoid of C. horticola larvae in canola fields of Ardabil region. The percentage of abundance of each of parasitoid species was significantly different compared with each other in two years (df=3, 108; F=3.36; P=0.0026) (Table 1). In 2008 and 2009, D. isaea had the highest abundance (48.3% and 42.8%, respectively) among the parasitoid species of C. horticola in Ardabil region. The abundance of other parasitoid species decreased in the following order: Chrysocharis pubicornis, Dacnusa sibirica and Pediobius sp. in both years (Table 1). The larval densities and parasitism rates
The density of C. horticola larvae differed significantly among the 19 canola cultivars for both study years (df=18, 108; F=3.62; P=0.0001) (Fig. 1). The density of larvae was lower on Opera, Hyola401 and RGS003 than on the other tested cultivars during 2008 and 2009. For other cultivars the density of larvae decreased in the following order: Talayh, Elvis, Ebonite, Hyola60, Okapi, Licord, PF/7045/91, Adder, Hyola308, SIM043, Jewel, Elite, Orient, SIM046, Option500 and Zarfam in 2008; and Elvis, Adder, Talayh, Hyola60, Licord, PF/7045/91, Hyola308, Jewel, Ebonite, Zarfam, Option500, SIM046, Okapi, Orient, SIM043 and Elite in 2009 (Fig. 1).
The percentage of parasitized larvae of C. horticola was significantly different among the 19 canola cultivars tested (df=18, 108; F=5.81; P=0.0001) (Fig. 2). In 2008, the parasitism rate was higher on Opera, Hyola401 and RGS003 than on other tested cultivars; however parasitism rates decreased in the following order for other cultivars: Okapi, Ebonite, Elvis, Zarfam, Licord, Option500, Orient, Talayh, Elite, Jewel, SIM046, Hyola60, Hyola308, Adder, SIM043 and PF/7045/91 (Fig. 2). In 2009, the parasitism rates decreased in the following order: Opera, RGS003, Okapi, Hyola401, SIM043, Elite, Ebonite, Elvis, Zarfam, Orient, Hyola308, Option500, SIM046, Licord, Hyola60, Talayh, Adder, Jewel and PF/7045/91 (Fig. 2).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 452
DISCUSSION
The Diglyphus isaea had highest abundance among the parasitoid species of Chromatomyia horticola in canola fields in Ardabil region. This parasitoid species has been reported as the dominant parasitoid of C. horticola larvae in Turkey (Uygun et al., 1995; Gençer, 2004, 2005; Cikman et al., 2006; Gençer, 2009).
The results of field experiments indicated that the density of C. horticola larvae was lower on Opera, RGS003 and Hyola401 among the 19 canola cultivars tested during 2008 and 2009. Also, the parasitism rate of C. horticola larvae was higher on Opera, Hyola401 and RGS003 respectively among the tested cultivars in 2008 and 2009. The low density of larvae and high parasitism rate on Opera, Hyola401 and RGS003 cultivars should result in lower pest damage and higher crop yield. These results indicated that different levels of resistance in cultivars have different effects on level of parasitism. Difference in leaf trichome density that could be related to these interactions was not discernible between 19 studied cultivars of canola. In most cases, the difference in the rate of parasitized larvae on different host species was associated with the cuticle thickness and volatile profiles of the tested species (Fagoonee & Toory, 1983; Parrella et al., 1983; Knodel-Montz et al., 1985; Minkenberg & Fredrix, 1989; Minkenberg & Ottenheim, 1990; Carolina et al., 1992; Erb et al., 1993; Wei et al., 2000). Our findings are consistent with previous studies. Fathi et al. (2010) concluded that Opera, RGS003 and Hyola401 exhibited antibiosis and reduced developmental rate, fecundity and survival of C. horticola. Plants responsible for antibiosis are known to have an indirect effect by increasing the exposure of the insect to its native natural enemies as a result of prolonged developmental time.
Our findings suggesting that using of resistant cultivars in integrated with biocontrol agents could play an important role in the IPM of C. horticola. The results of this study indicated that Opera, RGS003 and Hyola401 cultivars had lower larval density and higher parasitism rate by parasitoids. Therefore, these cultivars could increase the efficacy of D. isaea in control of C. horticola larvae. These results can be useful in decreasing the current dependence on using of chemical insecticide. The present findings in combination with our earlier results suggest that using Opera, RGS003 and Hyola401 integrated with D. isaea should result in increased efficacy of this parasitoid and lead to effective and more sustainable management of C. horticola in the region.
ACKNOWLEDGEMENTS
The Research Council of Mohaghegh Ardabili University (Iran) is gratefully acknowledged for financial support of this research.
LITERATURE CITED Capinera, J. L. 2008. American serpentine leafminer, Liriomyza trifolii (Burgess) (Insecta: Diptera: Agromyzidae). University of Florida IFAS Extension. Carolina, J., Herr, C. & Johnson, M. W. 1992. Host plant preference of Liriomyza sativae (Diptera: Agromyzidae) populations infesting green onion in Hawaii. Environmental Entomology, 21: 1097-1102. Chen, X., Lang, F., Xu, Z., He, J. & Ma, Y. 2003. The occurrence of leafminers and their parasitoids on vegetables and weeds in Hangzhou area, Sout heast China. BioControl, 48: 515–527. Cikman, E., Beyarslan, A. & Civelek, H. S. 2006. Parasitoids of leafminers (Diptera: Agromyzidae) from southeast Turkey with 3 new records. Turkish Journal of Zoology, 30: 167-173.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 453
Civelek, H. S. 2002. A new record for the parasitoid fauna of Chromatomyia horticola (Goureau, 1851) (Diptera: Agromyzidae) Pediobius metallicus (Nees, 1934) (Hymenoptera: Eulophidae). Turkish Journal of Entomology, 26: 155–159. Darvas, B., Andersen, A. & Thuroczy, C. S. 1999. Generalist hymenopteran parasitoids of the leaf-miner Chromatomyia fuscula (Zett.) (Diptera: Agromyzidae). Journal of Natural History, 33: 1089–1105. Erb, A. W., Lindquist, R. K., Flickinger, J. N. & Casey, M. L. 1993. Resistance of selected interspecific Lycopersicon hybrids to Liriomyza trifolli (Diptera: Agromyzidae). Journal of Economic Entomology, 86: 100-109. Fagoonee, I. & Toory, V. 1983. Preliminary investigations of host selection mechanisms by the leafminer Liriomyza trifolli. Insect Science and its Application, 4: 337-341. Fathi, S. A. A. 2010. Host preference and life cycle parameters of Chromatomya horticola Gourea (Diptera: Agromyzidae) on canola cultivars. Munis Entomology and Zoology, 5: 247-252. Gençer, L. 2004. A study on the chalcidoid (Hym.: Chalcidoidae) parasitoids of leafminers (Dip.: Agromyzidae) in Ankara province. Turkish Journal of Zoology, 28: 119-122. Gençer, L. 2005. Chalcidoid parasitoids of Chromatomyia horticola (Gour.) (Dip. Agromyzidae) in Sivas province, Turkey. Journal of Pest Science, 78: 41-43. Gençer, L. 2009. Contribution of the knowledge of the chalcid parasitoid complex (Hymenoptera: Chalcidoidea) of agromyzid leafminers (Diptera: Agromyzidae) from Turkey, with new hosts and records. Journal of Plant Protection Research, 49: 158-161. Knodel-Montz, J. J., Lyons, R. E. & Poe, S. L. 1985. Photoperiod affects chrysanthemum host plant selection by leafminers (Diptera: Agromyzidae). HortScince, 20: 708-710. LaSalle, J. & Parrella, M. P. 1991. The chalcidoid parasites (Hymenoptera: Chalcidoidea) of economically important Liriomyza species (Dip. Agromyzidae) in North America. Proceedings of the Entomological Society of Washington, 93: 571-591. Minkenberg, O. P. J. M. & Fredrix, M. J. J. 1989. Preference and performance of an herbivorous fly, Liriomyza trifolli (Diptera: Agromyzidae), on tomato plants differing in leaf nitrogen. Annals of the Entomological Society of America, 82: 350-354. Minkenberg, O. P. J. M. & Ottenheim, J. J. G. W. 1990. Effect of leaf nitrogen content of tomato plants on preference and performance of a leafmining fly. Oecologia, 83: 291-298. Noyes, J. S. 2003. Universal Chalcidoidea database, http://www.nhm.ac.uk/entomology/chalcidoids. Parrella, M. P. 1987. Biology of Liriomyza. Annual Review of Entomology, 32: 201-224. Parrella, M. P., Robb, K. L. & Bethke, J. 1983. Influence of selected host plants on the biology of Liriomyza trifolii (Diptera: Agromyzidae). Annals of the Entomological Society of America, 76: 112-115. Parrella, M. P., Jones, V. P., Youngman, R. R. & Lebeck, L. M. 1985. Effect of leaf mining and leaf stippling of Liriomyza spp. on photosynthetic rates of chrysanthemum. Annals of the Entomological Society of America, 78: 90-93. Rauf, A., Shepard, B. M. & Marshall, M. W. 2000. Leafminers in vegetables, ornamental plants and weeds in Indonesia: surveys of host crops, species composition and parasitoids. International Journal of Pest Management, 46: 257-266. Saljoqi, A. U. R., Hussain, S. R. N. & Khan, S. A. 2006. Insect pests of canola crop (other than aphid). Journal of Agricultural and Biological Science, 1: 19-21. SAS Institute. 2005. SAS/Stat users guide. SAS Institute, Cary, NC, USA. Uygun, N., Polatoz, Z. & Baspinar, H. 1995. Faunistic studies on Agromyzidae (Diptera) in the southeast Mediterranean region of Turkey. Turkish Journal of Entomology, 19: 123–136. Wei, J., Zou, L., Kuang R. & He, L. 2000. Influence of leaf tissue structure on host feeding selection by pea leafminer Liriomyza huidobrensis (Diptera: Agromyzidae). Zoological Studies, 39: 295-300.
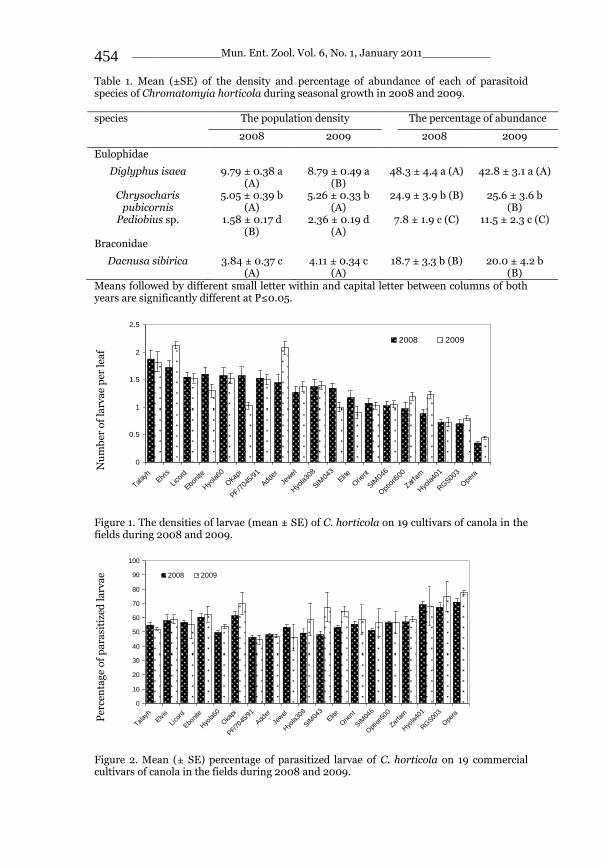
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 454
Table 1. Mean (±SE) of the density and percentage of abundance of each of parasitoid species of Chromatomyia horticola during seasonal growth in 2008 and 2009. species The population density The percentage of abundance
2008 2009 2008 2009
Eulophidae
Diglyphus isaea 9.79 ± 0.38 a (A)
8.79 ± 0.49 a (B)
48.3 ± 4.4 a (A) 42.8 ± 3.1 a (A)
Chrysocharis pubicornis
5.05 ± 0.39 b (A)
5.26 ± 0.33 b (A)
24.9 ± 3.9 b (B) 25.6 ± 3.6 b (B)
Pediobius sp. 1.58 ± 0.17 d (B)
2.36 ± 0.19 d (A)
7.8 ± 1.9 c (C) 11.5 ± 2.3 c (C)
Braconidae
Dacnusa sibirica 3.84 ± 0.37 c (A)
4.11 ± 0.34 c (A)
18.7 ± 3.3 b (B) 20.0 ± 4.2 b (B)
Means followed by different small letter within and capital letter between columns of both years are significantly different at P≤0.05.
Nu
mb
er o
f la
rva
e p
er l
eaf
0
0.5
1
1.5
2
2.5
Talay
hElv
is
Lico
rd
Eboni
te
Hyo
la60
Oka
pi
PF/704
5/91
Adder
Jewel
Hyo
la30
8
SIM04
3
Elite
Orie
nt
SIM04
6
Opt
ion5
00
Zarfa
m
Hyo
la40
1
RG
S003
Ope
ra
2008 2009
Figure 1. The densities of larvae (mean ± SE) of C. horticola on 19 cultivars of canola in the fields during 2008 and 2009.
Per
cen
tag
e o
f p
ara
siti
zed
la
rva
e
0
10
20
30
40
50
60
70
80
90
100
Talayh
Elvis
Lico
rd
Ebonite
Hyo
la60
Oka
pi
PF/70
45/9
1
Adder
Jewel
Hyo
la30
8
SIM04
3
Elite
Orie
nt
SIM04
6
Opt
ion5
00
Zarfam
Hyo
la40
1
RG
S003
Ope
ra
2008 2009
Figure 2. Mean (± SE) percentage of parasitized larvae of C. horticola on 19 commercial cultivars of canola in the fields during 2008 and 2009.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 455
ON THE DIAGNOSTICS OF MEALYBUG OCCURRING ON COTTON, SPHERICAL MEALYBUG NIPAECOCCUS
VIRIDIS (NEWSTEAD) (HEMIPTERA: PSEUDOCOCCIDAE)
Asha Thomas* and V. V. Ramamurthy* * Division of Entomology, Indian Agricultural Research Institute, New Delhi 110 012, INDIA. E-mails: [email protected]; [email protected] [Thomas, A. & Ramamurthy, V. V. 2011. On the diagnostics of mealybug occurring on cotton, spherical mealybug Nipaecoccus viridis (Newstead) (Hemiptera: Pseudococcidae). Munis Entomology & Zoology, 6 (1): 455-459] ABSTRACT: Detailed studies on the mealybugs collected from different parts of India on cotton and other hosts revealed the abundance of Nipaecoccus viridis (Newstead) (Hemiptera: Pseudococcidae), and established that it is emerging as next most widely distributed mealybug after Phenacoccus solenopsis Tinsley. This mealybug can be distinguished by its even segmented antennae, distribution of setae similar to cerariian setae on dorsal surface of the body and number of cerarii discernible with difficulty. The need to differentiate this from other important mealybugs occurring in cotton agroecosystems has been taken care of through making its diagnostics as simple as possible. The examination of populations from different regions, simplifying its diagnostics through illustration of taxonomic characters significant at all taxonomic levels and explaining their terminology has also been achieved. KEY WORDS: Coccoidea, taxonomic characters, illustrated diagnostics.
Cotton is an economically important crop and pests and pathogens are the major threats to its productivity. Among these mealybugs of the family Pseudococcidae are currently being reported as serious pests. Of the twenty three species of mealybugs reported worldwide, incidence of seven species had been recorded so far from India. These are Phenacoccus solenopsis Tinsley, Nipaecoccus viridis (Newstead), Maconellicoccus hirsutus (Green), Rastrococcus iceryoides Green, Ferrisia virgata Newstead and Paracoccus marginatus Williams and Granara de Willink. Among these P. solenopsis is most widely distributed and its diagnostics and complexities in taxonomic characters had been already illustrated (Thomas & Ramamurthy, 2008). Studies undertaken under the auspices of a project of Technology Mission on Cotton at the Indian Agricultural Research Institute revealed that N. viridis (Hemiptera: Pseudococcidae) is emerging as the next most injurious mealybug, in particular, in parts of Andhra Pradesh, Delhi, Maharashtra and Orissa. The perusal of taxonomic information available on this species reveals that there had been confusions on its species identity due to misidentifications (Ayyar, 1921; Ghose, 1961; Green, 1908; Kirkalady, 1902) and synonymy (Ali, 1970; De Lotto, 1958; Green, 1922; Zimmerman, 1948). Also, detailed illustrated diagnostics as available for P. solenopsis needs to be developed and made available to facilitate its diagnostics. Hence, the present study analyses N. viridis (Hemiptera: Pseudococcidae) at all taxonomic levels and provides a simplified diagnostics to enable its authentic identification.
MATERIALS AND METHODS
Adult female mealybugs collected from cotton during 200-2010 from Andhra
Pradesh, Delhi, Maharashtra and Orissa. 273 specimens were processed using

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 456
standard procedure (Hodges & Hodges, 2004). The terminologies followed are those of Ferris (1950), Downie & Gullan (2004) and Willams (2004).
RESULTS
Nipaecoccus viridis (Newstead) (Hemiptera: Pseudococcidae) is distinguished at the family Pseudococcidae level by the characters namely, broadly oval shaped body, (Fig. 1 & 2) 3.080±0.507 x 2.292±0.216 mm; antennae seven segmented (Fig. 1a & Fig. 2a), 0.210±0.130 mm long, its apical segment with 9-11 hair like setae, 4 fleshy and 2 stiff setae near apex, preapical segment with 2-4 hair like setae and also with a fleshy seta; clypeolabral shield (Fig. 2b) about 0.150±.003 mm long; labium (Fig. 2c) about 0.120±.003 mm long with 11 pairs of setae; spiracles (Fig. 2d,e): width of anterior peritremes 0.040±0.007 mm, posterior peritreme 0.055±0.005 mm; legs well developed (Fig. 1f & Fig. 2f), metathoracic leg: coxa 0.124±.002 mm, trochanter 0.073±0.014 mm, femur 0.136±0.006 mm, tibia 0.100±0.008 mm, tarsus 0.067±0.005 mm, claw 0.028±0.008 mm; translucent pores present on anterior and posterior surface of hind coxa and posterior surface of hind tibia (Fig. 1h & Fig. 2g,h); tarsal digitules both knobbed (Fig. 1i & Fig. 2i); claw digitule 0.044±0.001 mm long, claw without denticle; tubular ducts of varied size present (Fig. 1k & 2k), 0.009±0.001 x 0.002±0.0005 mm; ostioles moderately developed, one pair each present anteriorly and posteriorly (Fig. 1m & Fig. 2l,m); trilocular pores present on dorsal and ventral surface (Fig. 1n & Fig. 2n), about 0.004 mm wide, frequent throughout and fairly evenly distributed; dorsal surface with lanceolate setae, similar to those of cerariian but shorter and variable (Figs. 1o & 2o), 0.021±0.001 mm long, with their collar 0.005±0.001 mm; ventral surface with flagellate setae, those on head 0.040±0.007 mm long and those on abdomen 0.020±0.013 mm long, other setae minute, 0.011 mm long, with their collar 0.003 mm wide; discoidal pores about 0.002 mm, simple, distributed sparsely; anal ring 0.105±0.006 x 0.098±0.009 mm at the apex, with 2 rows of cells and 6 slender setae, each setae about 0.030±0.060 mm long (Figs. 1s & 2s); anal lobes moderately developed and less sclerotized; number of cerarii (Fig. 1p & 2p) difficult to verify precisely, anterior cerarii on abdomen, mesothorax and metathorax discernible with paired lanceolate setae, 0.019±0.002 mm long with their collar 0.008±.0005 mm wide and 2-6 trilocular pores, distance between the lanceolate setae increasing towards thoracic region; anal lobe cerarii with two broadly lanceolate setae which are 0.018±0.007 mm long, with 6-10 trilocular pores situated on their margins; circulus, between III and IV segments (Fig. 1q & 2q), round to quadrate 0.081±0.016 x 0.100±0.010 mm; and multilocular disc pores present ventrally (Fig. 1r & 2r), 0.007±0.0009 mm wide on abdomen and 0.007±0.001 mm wide on thorax. At the subfamily Pseudococcinae level it is defined by the seven segmented, 0.210±0.130 mm long antennae; tarsal digitules knobbed, claw without denticle; and quinquelocular pores absent. At the tribe Trabutinini level it is defined by its bluish black appearance, and presence of elongated dorsal setae which are 0.021±0.001 mm long. At the generic level Nipaecoccus is defined by the broadly ovoid body, 3.080±0.507 x 2.292±0.216 mm; antennae seven segmented and 0.210±0.130 mm long; legs well developed, claws stout, 0.028±0.008 mm long, without denticle, tarsal digitules knobbed; hind coxa and hind tibia with translucent pores; venter with oral collar tubular ducts, 0.009 x 0.001 mm long; cerarii 1-17 pairs; dorsal setae, 0.021±0.001 mm long, similar to cerariian setae; and venter with its multilocular disc pores 0.007±0.009 mm wide. At species level viridis is distinguished by seven segmented, 0.210±0.130

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 457
mm long antennae; multilocular disc pores ventrally, 0.007±0.009 mm wide; and oral collar tubular ducts 0.009 x 0.001 mm long, numerous, reaching thorax and present medially and in marginal zone.
DISCUSSION
Authentic diagnostics of N. viridis is possible with the combination of seven segmented antennae, claw without denticle, tarsal digitule knobbed, dorsal setae which is often similar in size and shape to the cerariian setae all over the body, multilocular disc pores and oral collar tubular ducts present medially and to the marginal zone throughout. The peculiarity of difficulty in distinguishing the number of cerarii is also one of the important characters of this species. Normally bluish black when preserved in spirit and sometimes difficult to prepare and stain on the microslides are its other distinct differentiating characters. On the contrary, P. solenopsis has nine segmented antennae, claws with denticle, tarsal digitule setose, 18 pairs of cerarii and multilocular disc pores concentrated medially on the abdominal segments VI, VII and VIII, and marginally on 1 to IV.
There had been misidentifications of N. viridis as N. nipae due to similarity in characters and the difficulties in processing due to its soft body. According to Williams (2004), nipae differs from viridis in its characteristics of multilocular disc pores on abdomen only and presence of oral collar tubular ducts medially and submedially; also N. viridis is longer (upto 4 mm) than nipae (2.3 mm).
Analysis of mealybug samples obtained through consistent sampling during 2007-2010 reveal that 86% comprises P. solenopsis, 13% of N. viridis and 0.7% of M. hirsutus and 0.3% of P. marginatus; these also, reveals the presence of P.solenopsis throughout the country while N. viridis is only in the states namely Andhra Pradesh, Delhi, Maharashtra, and Orissa; M. hirustus has been observed at Delhi, Maharashtra, and Tamil Nadu. These findings reveal that N. viridis is emerging as another important injurious mealybug after P.solenopsis in the cotton agroecosystems of India. Also enough attention is required to consider all the taxonomic characters together for flawless species identification.
LITERATURE CITED Ali, S. M. 1970. A catalogue of Oriental Coccoidea. Part IV (Insecta: Homoptera: Coccoidea). Indian Museum Bulletin, 5: 71-150. Ayyar, T. V. R. 1921. A checklist of Coccidae of the Indian origin. Proceedings of Entomological meeting, IV, Pusa, pp. 336-362. Downie, D. A. & Gullan, P. J. 2004. Phylogenetic analyses of mealybugs (Hemiptera: Coccoidea: Pseudococcidae) based on DNA sequences from three nuclear genes, and review of the higher classification. Systematic Entomology, 29: 238-259. De Lotto, G. 1958. The Pseudococcidae (Homoptera: Pseudococcidae). Bulletin of the British Museum Entomology, 7: 77-120. Ferris, G. F. 1950. Atlas of the scale insects of North America. V, The Pseudococcidae (Part 1). Standard University Press, 278 pp. Green, E. E. 1908. Remarks on Indian scale insects (Coccidae). Part III. Memoirs of the Department of Agriculture in India, Entomology Series, 2: 5-46. Green, E. E. 1922. The Coccidae of Ceylon. Part V. London, Dulau & Co. pp. 105-169. Ghose, S. K. 1961. Studies on some coccids of economic importance of West Bengal. Indian Agriculture, 5 (1): 57-78.
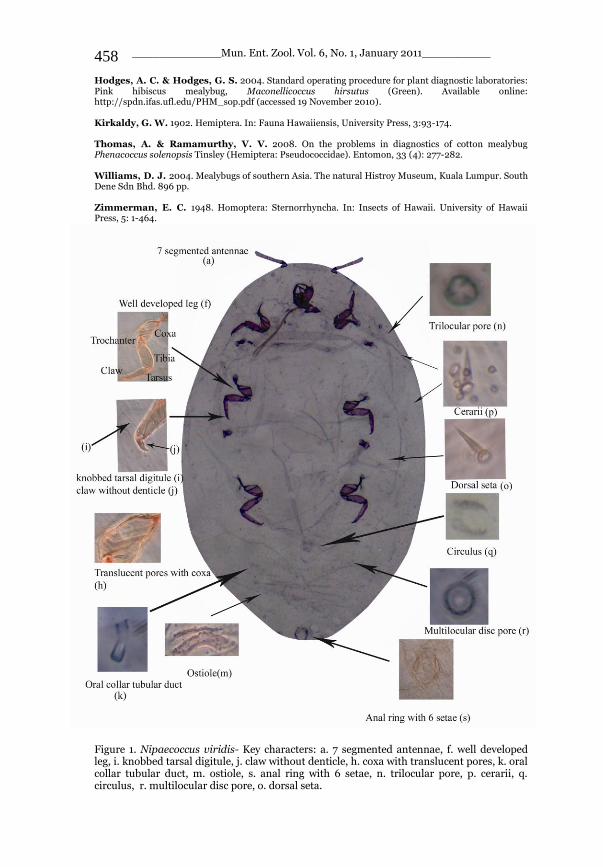
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 458
Hodges, A. C. & Hodges, G. S. 2004. Standard operating procedure for plant diagnostic laboratories: Pink hibiscus mealybug, Maconellicoccus hirsutus (Green). Available online: http://spdn.ifas.ufl.edu/PHM_sop.pdf (accessed 19 November 2010). Kirkaldy, G. W. 1902. Hemiptera. In: Fauna Hawaiiensis, University Press, 3:93-174. Thomas, A. & Ramamurthy, V. V. 2008. On the problems in diagnostics of cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Entomon, 33 (4): 277-282. Williams, D. J. 2004. Mealybugs of southern Asia. The natural Histroy Museum, Kuala Lumpur. South Dene Sdn Bhd. 896 pp. Zimmerman, E. C. 1948. Homoptera: Sternorrhyncha. In: Insects of Hawaii. University of Hawaii Press, 5: 1-464.
Figure 1. Nipaecoccus viridis- Key characters: a. 7 segmented antennae, f. well developed leg, i. knobbed tarsal digitule, j. claw without denticle, h. coxa with translucent pores, k. oral collar tubular duct, m. ostiole, s. anal ring with 6 setae, n. trilocular pore, p. cerarii, q. circulus, r. multilocular disc pore, o. dorsal seta.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 459
Figure 2. Nipaecoccus viridis- key characters: Diagrammatic view. A-dorsal, B-ventral. a. 7 segmented antennae, b. clypeolabral shield, c. labium, d. anterior spiracle, e. posterior spiracle, f. well developed leg, g. tibia with translucent pores, h. coxa with translucent pores, i. knobbed tarsal digitule, j. claw without denticle, k. oral collar tubular duct, l. anterior ostiole, m. posterior ostiole, n. trilocular pore, o. dorsal seta, p. cerarii, q. circulus, r. multilocular disc pore, s. anal ring with 6 setae.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 460
STUDY ON THE COMBINED IMPACT OF CITOWETT AND COMMERCIAL NEEM EXTRACT AGAINST
PIERIS BRASSICAE L. UNDER LABORATORY CONDITIONS (LEPIDOPTERA: PIERIDAE)
Vine Shirzad*, Bernadet Sahak*, Ali Asghar Pourmirza
and Youbert Ghosta * Department of Plant Protection, Agricultural Faculty, Urmia University, West Azarbaijan, IRAN. E-mails: [email protected]; [email protected] [Shirzad, V., Sahak, B., Pourmirza, A. A. & Ghosta, Y. 2011. Study on the combined impact of citowett and commercial neem extract against Pieris brassicae L. under laboratory conditions (Lepidoptera: Pieridae). Munis Entomology & Zoology, 6 (1): 460-463] ABSTRACT: To determine the impact of citowett oil and commercial neem extract mixture against the Pieris brassica these experiments were under taken. Citowett as a wetting agent was used in conjunction with neem extract against the larval stage of the pest in question. The LC50 values of neem extract and combination with citowett oil for second and third instars larvae were estimated, 4.40, 3.89 and 3.38, 2.16 ppm, respectively. The combination of neem extract with citowett increased the larval mortality rate up to 15%. This implies that the application of the mixture is a sound measure for enhancing lethality impacts. Based on collected data it could be concluded that combination of these two less hazardous chemicals is merit to be considered as a candidate control agent against Pieris brassica. KEY WORDS: Compatibility, botanical origin compound, wetting agent, Pieris brassica.
Cabbage (Brassica oleracea L. var. capitata) is an important vegetable crop grown in many countries in the world (Ţnidarčič et al., 2008). Several Lepidopterian pests attack cabbage vegetables and the most serious damage is caused mainly by the larvae of several species such as: small white butterfly (Pieris rapae L.), large white butterfly (Pieris brassicae L.), cabbage moth (Mamestra brassicae L.) and diamondback moth (Plutella xylostella L.). Pieris brassicae L., is a cosmopolitan insect, and is found wherever cruciferous plants are grown. Sometimes massive outbreaks of this pest may occur and injury on cabbage crops could be extensive (Metaspalu et al., 2009). The economic damage occurs when the percent destroyed leaf area reach within 3.97-10.41 in early cabbage, 10.75-13.76 in the late cabbage (Ashfag et al., 2006). The damage notably affects the value of this crop because its consumption and sale happen when it is still fresh (Cartea et al., 2009).
Azadirachtin, a chemical complex found in seeds of neem, Azadirachta indica A. Juss, is the main component responsible for the toxic, repellent, antifeedant, growth-inhibiting, oviposition-inhibiting and sterilizing effects in insects ( Sukontason et al ., 2008). Neem extracts has also been shown to have direct detrimental and histopathological effects on many insect tissues, for example, muscles, fat body, and gut epithelial cells (Jogar et al., 2008). It is usually safe for beneficial organisms, such as bees, predators and parasitoids, mammals, and for the environment. This plant is an interesting option for integrated pest management programs, since it is selective, presents a less negative impact on the ecosystems and works in association with biological control organisms ( Sukontason et al ., 2008).
Citowett as a wetting agent has been used in conjunction with neem extract, when it is added to a spray mixture, foliage wettability is greatly improved on the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 461
surface of either "waxy" or "hairy" leaf surfaces or buds (Anonymus. 2008). The combination of two or more control options may minimize risk and costs of chemicals, reduce resistance development against the treatments and increase effectiveness of the treatments (Yuya et al., 2009).
With retrospect, the objectives of this study were to determine the combined effects of Citowett oil and Neem extracts against the Large White Butterfly, on cabbage, and to determine the minimum effective rate(s) of the combinations that can provide adequate protection to cabbage against Pieris brassicae.
MATERIALS AND METHODS
The eggs of cabbage butterfly were collected from the fields of Urmia. Newly hatched larvae were fed with fresh cabbage leaves. Larvae were reared under laboratory conditions 21 ± 2ºC and relative humidity 75±10 percent on fresh cabbage leaves. The combined effect of neem extract with Citowett oil on the second and third instars larvae of P. brassicae was investigated. To evaluate the synergistic effect of neem extract and citowett, we used a solution which contained 75 and 25% of nemm extract and citowit, respectively. To this end, concentrations of 1, 2, 4, 8, 16% of neem with 1% of citowett alongside untreated leaves were employed. Each experiment was replicated three times on different days. Freshly cut leaves of cabbage were immersed in a prepared insecticide solution for 10 s and hung vertically to air dry for 30 minutes. Control leaves were treated similarly with tap water. Filter paper was placed inside a plastic petri-dish and treated leaf was placed on top of the filter paper. Fifteen randomly selected second and third instars larvae were released in each petri dish, separately and held under laboratory conditions.
RESULTS
Antifeedant effects of neem extracts insecticides were indicated by patterns of larval development and foliage consumption. We observed that the color of the larvae became darker a few days after they fed on compounds-treated leaves, and turned almost complete black when they died. When second instars larvae were introduced on treated leaf disks, they ceased feeding, slowly dying, and did not cause noticeable foliage damage. When third instars larvae were introduced on treated leaf disks, they normally initiated feeding for a certain period of time, and eventually stopped feeding. Therefore, some degree of feeding damage occurred on the treated leaves, although significantly less foliage damage was caused by the younger and smaller larvae than that caused by the larger and older larvae (Table 1 and 2).
DISCUSSION
Botanical insecticides are an important group of naturally occurring, often slow-acting crop protectants that are usually safer to humans and the environment than conventional insecticides. Moreover, thanks to the fact that botanical insecticides contain mixtures of biologically active substances, no resistance is developed in pests and pathogens. Therefore the use of plant insecticides has been recommended ever more as a suitable alternative of plant protection with minimum negative risks (Pavela, R.2009). The results showed that neem was effective against P. brassicae, significantly reducing the survival of larvae feeding on cabbage leaves treated with the extract. In our tests, the extract

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 462
did not produce rapid mortality. Following ingestion of treated plant material, larvae became lethargic and exhibited slow locomotion and reactions. These compounds exhibited a significant lethal effect on second and third larval instars, although the effect was slow and varied among the larval instars. Therefore, the compound should be applied as early as possible when the larvae are in second instars of developmental stage. Neem might be enhanced the management measures of Lepidopterous pests in the vegetable agroecosystems because they do not persist in the environment, have unique modes of action, low mammalian toxicity, and may be potentially compatible with natural enemies.
ACKNOWLEDGEMENTS
We would like acknowledge the financial support provided to this research by the Urmia University.
LITERATURE CITTED Anonymus. 2008. Citowett Plus.http://pr-rp.pmra-arla. gc. ca/PR-SOL/pr-web.ve1?p-ukid=9862. Ashfag, M., Gill, T. I. & Ali, A. 2006. Consumption and utilization of various plants by parasitized and unparasitized larvae of cabbage butterfly (Pieris brassicae L). Pak. Entomol., 28: 1-4. Atawodi, S. E. & Atawodi J. C. 2009. Azadirachta indica (neem): a plant of multiple biological and pharmacological activities. Phytochem. Rev., 8: 601-620. Calvo, D. & Molina, J. M. 2003. Effects of a commercial Neem (Azadirachta indica) extract on streblote panda larvae. Phytoparasitica, 31 (4): 365-370. Cartea, M. E., Padilla, G., Vilar, M. & Velasco, P. 2009. Incidence of the major Brassica pests in Northwestern Spain. Plant Resistance. 767-773. Greenberg, S. M., Showler, A. T. & Liu, T. 2005. Effects of neem-based insecticides on beet armyworm (Lepidoptera: Noctuidae). Insect Science, 12: 17-23. Jogar, K., Kuusik, A., Metspalu, L., Hiiesaar, K., Grishakova, M. & Luik, A. 2008. Effects of Neem EC on gas exchange, tracheal ventilation, and water loss in diapausing pupae of Pieris brassicae.Entomologial Experimentalis et Applicata, 126: 165-173. Jankwska, B. 2006. The occurrence of some Lepidoptera pests on different cabbage vegetables. Journal of Plant Protection Research, 46: 181-190. Metaspalu, L., Hiiesaar, K., Jogar, K., Svilponis, E., Ploomi, A., Kivimagi, I., Luik, A. & Menshikova, N. 2009. Oviposition preference of Pieris brassicae (L) on different Brassica oleracea var. Capitatal. Cultivars. Agronomy Research, 7: 456-411. Pavela, R. 2009. Effectiveness of some botanical insecticides against Spodoptera littoralis Boisduvala (Lepidoptera: Noctudiae), Myzus persicae Sulzer (Hemiptera: Aphididae) and Tetranychus urticae Koch (Acari: Tetranychidae). Plant Protection Science, 45: 161-167. Sukontason, K. L., Olson, J. K., Chailapakul, O. & Sukontason, K. 2008. Efficacy of neem extract against the blowfly and housefly. Parasitol Res., 103: 535-544. Younas, M., Naeem, M., Raqib, A. & Masud, S. 2004. Population dynamics of cabbage butterfly (Pieris brassicae) and cabbage aphids (Brevicoryne brassicae) on five cultivars of cauliflower at Peshawar. Asian Journal of Plant Sciences, 3 (3): 391-393. Yuya, A. I., Tadesse, A. Azerefegne, F., & Tefera, T. 2009. Efficacy of combining Niger seed oil with malathion 5% dust formulation on maize against the maize weevil, Sitophilus zeamais (Coleoptera: Curculionidae). Journal of Stored Products Research, 45: 67-70. Zabel, A., Manojlovic, B., Rajkovic, S., Stankovic, S. & Kostic, M. 2002. Effect of Neem extract on Lymantria dispar L. (Lepidoptera: Lymantriidae) and Leptinotarba decemlineata Say. (Coloptera: Chrysomelidae). J. Pest Science, 75: 19-25.
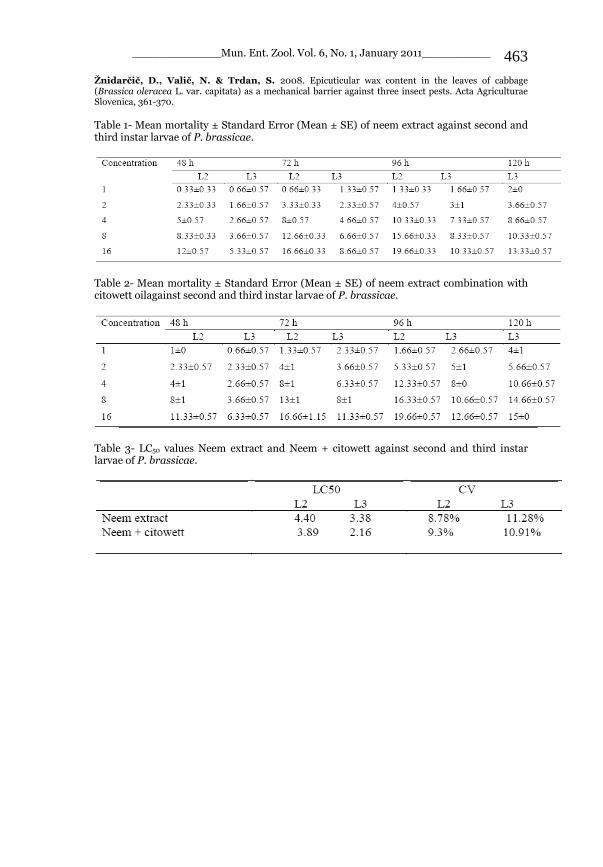
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 463
Žnidarčič, D., Valič, N. & Trdan, S. 2008. Epicuticular wax content in the leaves of cabbage (Brassica oleracea L. var. capitata) as a mechanical barrier against three insect pests. Acta Agriculturae Slovenica, 361-370.
Table 1- Mean mortality ± Standard Error (Mean ± SE) of neem extract against second and third instar larvae of P. brassicae.
Table 2- Mean mortality ± Standard Error (Mean ± SE) of neem extract combination with citowett oilagainst second and third instar larvae of P. brassicae.
Table 3- LC50 values Neem extract and Neem + citowett against second and third instar larvae of P. brassicae.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 464
REDESCRIPTION OF BRACHYAULAX CYANEOVITTA (WALKER) (HETEROPTERA: SCUTELLERIDAE)
Shama Parveen*, Sucheta Khokhar**,
Md. Kamil Usmani*** and V. V. Ramamurthy*
* Network Project on Insect Biosystematics, Division of Entomology, Indian Agricultural Research Institute, New Delhi – 110012, INDIA. E-mail: [email protected] ** Zoology Department, CCS Haryana Agricultural University, Hisar, Haryana 125004, INDIA. E-mail: [email protected] *** Zoology Department, Aligarh Muslim University, Aligarh, Uttar Pradesh 202002, INDIA. E-mail: [email protected] [Parveen, S., Khokhar, S., Usmani, Md. K. & Ramamurthy, V. V. 2011. Redescription of Brachyaulax cyaneovitta (Walker) (Heteroptera: Scutelleridae). Munis Entomology & Zoology, 6 (1): 464-468] ABSTRACT: Brachyaulax cyaneovitta (Scutelleridae: Scutellerinae) was described from China by Walker (1867), and this genus is known so far from five species. It is distinguished by its small (almost 0.5x as Scutellera spp.) odoriferous orifice and abdominal furrow reaching only up to second abdominal segment. The present study describes the female of B. cyaneovitta along with its genitalia and the morphometrics of the essential taxonomic characters. B. cyaneovitta can be distinguished by head, pronotum, metathoracic scent gland, abdomen etc., which are illustrated herein. KEY WORDS: Brachyaulax cyaneovitta, Scutelleridae, redescription, female genitalia.
Brachyaulax (Heteroptera: Scutelleridae) was first described by Stal (1871) with its type species as B. rufomaculata from Philippines. This genus is closely related to Scutellera Lamarck but can be distinguished in its small sized (almost 0.5x as that of Scutellera spp.) odoriferous orifice and ventral abdominal furrow reaching only upto second abdominal segment. This genus presently has five species viz., cyaneovitta (Walker), kukenthali Breddin, majuscula Distant, oblonga (Westwood) and rufomaculata Stal among which only B. cyaneovitta and B. oblonga are recorded from India. These are very similar, difficult to distinguish leading to confusions in their species identities. B. cyaneovitta was first described from China by Walker (1867) as Scutellera cyaneovitta; in 1899, Distant synonymyzed it with B. oblonga (described from Java). Distant (1902) in his monograph on the Hemiptera of erstwhile British India including Ceylon and Burma, recognized B. oblonga as the lone widely distributed species in South and Southeast Asia, and recorded its occurrence from several localities in India, continental Southeast Asia and the Malay Archipelago. Esaki (1926) considered the synonymy proposed by Distant (1899) as erroneous; also Hoffman (1935a,b) treated B. cyaneovitta as a distinct species. Finally, Tsai and Redei (2010) after examining a syntype of B. oblonga, and holotype of B. cyaneovitta concluded that these are distinct.
Despite these conclusions, the distinction of B. cyaneovitta and its description remains inadequate, especially in terms of morphometrics of essential taxonomic characters and female genitalia. Hence an attempt was made to fulfill this gap and define the species in its entirety to enable its authentic identification and the details are presented herein.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 465
MATERIALS AND METHODS
Material for the study is from the National Pusa Collection (NPC), Division of Entomology, IARI, New Delhi. The external morphology was studied and line diagrams prepared with Nikon SMZ 10 stereozoom microscope, fitted with an ocular and stage micrometer. The genitalia were studied after treatment of abdomen in 10% KOH for 30 min to soften it, opening the same with fine needle on the lateral sides, extracting the genitalic structure, and boiling in 10% KOH for 5 min at 100º C. These genitalic structures were studied under Nikon MZ 10 and Leica MZ 16 A stereozoom microscopes. The terminology is after Pendergrast (1957), Scudder (1959) and Mc Donald (1966). The measurements are in mm, head breadth measured across eyes, and that of scutellum and abdomen at base. Abbreviation used in figures: Dsd: Distal spermathecal duct; Dsf: Distal flange; Evp: Evaporatorium; Ey: Eye; Fr: Furrow; Gnx: Gonocoxa; Oc: Ocellus; Ost: Ostiole; Pr: Peritreme; Prf: Proximal flange; Prt: Paratergite; Psd: Proximal spermathecal duct; Sd: Spermathecal duct; Spb: Spermathecal bulb; scales of magnifications as in illustrations, each scale shown equals to 1.0 mm.
RESULTS
Brachyaulax cyaneovitta (Walker, 1867) (Fig. 1)
Scutellera cyaneovitta Walker, 1867: 16; Distant 1899:35 Tectocoris oblonga Westwood, 1837: Distant 1902: 52 Scutellera amethystine Germar, 1839: Ho, 2003: 195
Redescription General Colour (Fig. 1) greenish or violaceous blue with black spots dorsally; antenna, sternum, femora, tibia and all tarsal segments black; labium, coxa and trochanter brown, lateral margins of pronotum, lateral margins and a central basal patch to the ventral abdomen irregularly reddish ochraceous. Head (Fig. 2A) declivent, length (3.5) and breadth (3.7) subequal, lateral margins sinuate, tylus surpasses jugal lobes, preocular region 2.2x as longer post ocular; interocellar distance 2.1x to interocular while ocellus placed 0.8 apart from eye; a black spot from apex to base in the central lobe and a spot around each eye present, and eyes protruding laterally. Antennae (Fig. 2B) five segmented, attached ventrally, near to eyes, first segment (1.0) never reach beyond apex of the head and it is subequal to third (1.05), second smallest amongst all and also less than 0.5x as the first; fourth 1.2x as third while fifth longest and almost 3.5x as long as second and total length 5.3. Labium (Fig. 2C) four segmented, extends upto second abdominal segment, first smallest (0.9), second largest (1.95) and almost 2.2x to previous one, third and fourth measure 1.05 and 1.25, respectively, and total length 5.15. Thorax with pronotum (Fig. 2D) having its anterior margin straight, breadth across anterior angles subequal to that at middle (3.8) while that at posterior angles 1.53x as that at the middle, and its anterior angles subquadrate while posterior ones obtuse. Surface before discal area depressed, large and deep punctures at anterior margin and in depressed area, six black spots arranged in two transverse series, the posterior one largest and small hairs present near to

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 466
marginal area laterally. Scutellum (Fig. 2E) covers whole of the abdomen, 1.55x long as breadth at base; ten spots on the surface, three basal, middle one linear and elongated, two before middle, sometimes attached to the lateral margins and sometimes connected, two small and lateral, sometimes connected with the preceding, two a little before apex, sometimes connected and one apical with apex rounded. Metathoracic scent gland (Fig. 2F) orifice spherical, peritreme transverse, slightly curved medially, grooved throughout the length, evaporatorial surface sulcated or wrinkled, and widely extending to mesopleuron. Anterior margin of prosternum concave medially, and with a deep groove resting labium. Legs (Fig. 2H, 2I and 2J) have no taxonomic information, its measurements of length of fore, middle and hind femora 2.1, 2.8 and 3.2, respectively, while that of fore, middle and hind tibia 2.5, 2.8 and 3.6, respectively, tarsi with three segments, second smallest, first and third subequal and almost 2x as long the second. Abdomen (Fig. 2G) 1.25x as long as broad, median furrow upto second segment to hold the labium in resting condition, and with a greenish or bluish-black subquadrate spot in each segment more towards lateral area. Female genitalia (Fig. 2K and 2L) with ovipositor having eighth paratergite triangular and of moderate size, its postero- lateral angle projecting posteriorly, ninth small and lobe like, first gonocaxa large, its posterior margin medially sinuted, and bear small hairs on its posterior margin. Spermathecal bulb elongate with apical end rounded, proximal and distal flanges vasiform, pump region short, distal duct 0.5x as thick as proximal, its dilation large and balloon shaped. Male genitalia already described by Kumar (1964). Material examined: INDIA: South India, Valparai, 1♀, 5.vi.1998, G. Ravindra coll. Distribution: India (Assam), Myanmar.
ACKNOWLEDGEMENTS
We would like to thank Indian Council of Agricultural Research, New Delhi for providing financial assistance to Network Project on Insect Biosystematics.
LITERATURE CITED Distant, W. L. 1899. Rhynchotal notes. Heteroptera: Scutellerinae and Graphosominae. Annals and Magazine of Natural History, (7) 4: 29-52. Distant, W. L. 1902. The Fauna of British India including Ceylon and Burma Vol. I. Taylor and Francis, London. 438 pp. Esaki, T. 1926. Verzeichniss der Hemiptera-Heteroptera der Insel Formosa. Annales historico- naturales Musei nationalis Hungarici, 24: 136-189. Germar, E. F. 1839. Beitrage zu einer monographie der Schildwanzen. Zeitschrift für die Entomologie, 1 (1):1-146. Ho, J. Z. 2003. Illustrated handbook of true bugs of Taiwan. Kiss Nature Series, 2, 1-280. Chin- Chin Publication, Taipei.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 467
Hoffmann, W. E. 1935a. An abridged catalogue of certain Scutelleroidea (Plataspidae, Scutelleridae and Pentatomidae) of China, Indo-China, and Taiwan. Lignan University Science Bulletin, 7: 1-294. Hoffmann, W. E. 1935b. Notes on eight pentatomids not hitherto recorded from Formosa. Transactions of the Natural History Society of Formosa, 25 (136-139): 57-62. Kumar, R. 1964. On the structure and function of the so-called ejaculatory reservoir in Pentatomoidea (Hemiptera: Heteroptera). Proceedings of the Royal Society of Queensland, 75 (8): 51-65. McDonald, F. J. D. 1966. The genitalia of North American Pentatomoidea (Hemiptera: Heteroptera). Quaestiones Entomologicae, 2: 7-150. Pendergrast, J. G. 1957. Studies on the reproductive organs of the Heteroptera with a consideration of their bearing on classification. Transaction of the Royal Entomological Society of London, 109: 1-63. Scudder, G. G. E. 1959. The female genitalia of the Heteroptera: Morphology and bearing on classification. Transaction of the Royal Entomological Society of London, 111: 405-467. Stal, C. 1871. Hemiptera insularum Philippinarum. Bidrag till Philippinska oarnes Hemipter-fauna. Ofversigt af Kongliga Vetenskaps-Akademiens Forhandlinger, 27 (7): 607-776. Tsai, J. F. & Redei, D. 2010. Taxonomic notes on Oriental and Pacific jewel bugs (Hemiptera: Heteroptera: Scutelleridae). Zootaxa, 2572: 25-47. Walker, F. 1867. Catalogue of the specimens of Hemiptera Heteroptera in the collection of the British Museum. E. Newman, London, Part I: 1-240. Westwood, J. O. 1837. A catalogue of Hemiptera in the collection of the Rev. F. W. Hope, M. A. with short Latin diagnoses of the new species. London, Part 1: 46 pp.
Figure 1. Brachyaulax cyaneovitta (Walker, 1867)
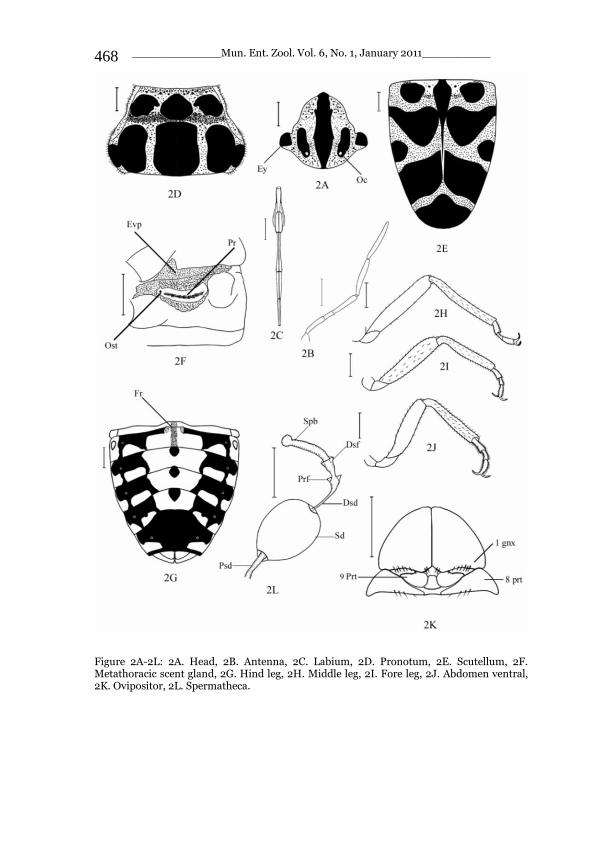
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 468
Figure 2A-2L: 2A. Head, 2B. Antenna, 2C. Labium, 2D. Pronotum, 2E. Scutellum, 2F. Metathoracic scent gland, 2G. Hind leg, 2H. Middle leg, 2I. Fore leg, 2J. Abdomen ventral, 2K. Ovipositor, 2L. Spermatheca.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 469
SUSCEPTIBILITIES OF THE CABAGE APHID, BREVICORYNE BRASSICAE (LINNAEUS) TO SELECTED
ORGANOPHOSPOROUS INSECTICIDES
A. Moslemi*, S. Ravan*, B. Mehrabi* and A. R. Bandani** * Plant Protection Department, Faculty of Agriculture, University of Zabol, Zabol, IRAN. ** Plant Protection Department, College of Agriculture and Natural Resources, University of Tehran, Karaj, IRAN. E-mail: [email protected] [Moslemi, A., Ravan, S., Mehrabi, B. & Bandani, A. R. 2011. Susceptibilities of the cabage aphid, Brevicoryne brassicae (Linnaeus) to selected organophosporous insecticides. Munis Entomology & Zoology, 6 (1): 469-474] ABSTRACT: In this study the effect of three insecticides including Trichlorophen, chlorpyriphos-methyl and Oxydemton-methyl on Brevicoryne brassicae in a leaf dip assay was investigated. At the highest and the lowest concentrations of Oxydemton-methyl 91.16 and 25 % mortality was achieved, respectively. Fifty percent mortality (LC50) was recorded at 19.62 ppm with fudicial limit of 11.41 and 30.73 ppm. Also, LC10 and LC90 were estimated to be 1.08 and 356.27 ppm. When different concentrations of Trichlorophen was used at the lowest and the highest concentrations about 21.27 and 87.23 % mortality was achieved, respectively which correspond to 30 and 480 ppm of the insecticide doses. Lethal concentration of 10, 50 and 90% was calculated using probit analysis and estimated LC10, LC50 and LC90 were 14.30, 95.96 and 643.89 ppms, respectively. Estimated lethal concentration of 10, 50 and 90 for chlorpyriphos-methyl were equal to 14.06, 59.65 and 253.10 ppm, respectively. Results showed that the most effective insecticide was Oxydemton-methyl with the LC50 value of 19.62 ppm followed by chlorpyriphos methyl with the LC50 value of 59.65 ppm and Trichlorophen with the LC50 value of 95.96 ppm. Therefore, Oxydemton-methyl act on insect through contact and systemic action that is major reason why this insecticide has low LC50 and is more effective than the other two insecticides which are contact and stomach insecticides. KEY WORDS: Cabbage aphid, Oxydemton-methyl, Chlorpyriphos methyl, Trichlorophen.
The cabbage aphid (Brevicoryne brassicae L.) is a widely distributed and severe pest of horticultural and oil-seed brassica crops. The aphid first indentified in Europe, but now its distribution is worldwide especially in most countries with a temperate climate (Ellis & Singh, 1993; Singh & Ellis, 1993).
This aphid feeds exclusively on all cultivated and wild cruciferous plants. The aphid major plant hosts include: broccoli, Brussels sprouts, cauliflower, and head cabbage. It also feeds on the other species of the family Cruciferae, however, damage is usually less severe than on cabbage. Cabbage aphid populations, if not controlled, often build to very high densities. Heavily infested plants acquire a grayish appearance due to the mass of aphid bodies on the foliage.
Aphids feed by sucking sap from their hosts. Infested seedlings may become stunted and distorted. Continued feeding on mature plants causes wilting, yellowing and general stunting of the plants. Honeydew and sooty mold are often evident. High densities also cause the leaves to wrinkle and curl, usually cupping downward. Under dry conditions, aphids cause the plants to wilt, and leaf tissue that has been fed upon may turn yellow. Cabbage aphid prefers the youngest tissue and highest portions of the plant, but may occur on both the upper and lower surface of foliage. Flower heads of seed crops may be attacked, reducing the setting of seed. Contamination of the plants with honeydew and aphids can cause considerable loss (Trumble, 1982; Pickel et al., 1983).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 470
In addition to the direct effects of feeding by aphids on plant growth, and the damage caused by aphid contamination of foliage, cabbage aphid can also be a vector of plant viruses. Over 30 viruses are known to be transmitted by B. brassica. Cauliflower mosaic and cabbage ring spot virus transmission were studied by Broadbent (1954). Cabbage aphid transmits cauliflower mosaic more effectively because this virus concentrates in the young tissue of the plant, which is the preferred feeding site of the aphid.
In warm climates aphid colonies consist completely of females. Reproduction does not involve mating or egg laying. Females give birth to live female nymphs. In temperate climates, aphids reproduce as above during the warmer periods of the year. In the fall, the reproduction changes. Males are produced in response to a decrease in photoperiod or temperature (Blackman & Eastop, 1984). Mating occurs and females lay eggs. Aphids overwinter in the egg stage. Life cycle duration ranges from 16 - 50 days and is greatly influenced by temperature. The life cycle is shortened at higher temperatures.
Naturally occurring parasites and predators are important factors in regulating population densities. Syrphid fly maggots and lady beetles are efficient predators of aphids. There are many insecticides that are effective against this aphid. Because of the waxy nature of the pest and crop, care must be taken that sprays provide good wetting of the crop. Proper rates of surfactants in combination with well adjusted spray equipment are important to achieve control with minimum effort. So, aim of the current study was to evaluate susceptibilities of cabbage aphid population to insecticides used for control of this aphid species in Iran. These insecticides include Trichlorophen, oxydemeton- methyl, and chlorpyriphos methyl. So, the efficacy of these insecticides on Brevicoryne brassicae was investigated to determine these insecticides effectiveness on the aphid. Bioassays using treated leaf disks were used to determine dose response curves for all insecticides used.
MATERIALS AND METHODS Insect rearing
Brevicoryne brassicae used in these experiments were collected from rape field of Saveh and transferred to laboratory. The aphid was reared on rape var. Havila in greenhouse at 22 ± 2 °C, a 12 h light: 12 h dark cycle, and relative humidity of 70 ± 5%. These colonies were kept on each rearing plants for several generations. Every week (5-7 days) plants were replaced with new ones in order to keep colonies alive. Apterous adults from these colonies were used in this study. Seedlings used for aphids culturing as well as producing leaf disks for insecticide bioassays were grown in plastic pots in above mentioned conditions. Toxicity bioassay
Three insecticides used in this experiment were Trichlorophen (Diptrex®), chlorpyriphos (Reldan®) and Oxydemton-methyl (Metasystox-R®) 25% EC (German's Bayer Company). Leaf dip assays were performed according to the procedures described by Lowery et al. (2005). Initially, for each insecticide on each population, bracketing test was done to determine doses that produce satisfactory range (10% - 90% mortality). The concentrations used for Trichlorophen were 0 (control), 30, 60, 120, 240, and 480 ppm. The concentrations used for Oxydemton-methyl were 0 (control), 3, 10, 30, 100, and 300 ppm and for chlorpyriphos were 0 (control), 20, 40, 80, 160 and 320 p pm.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 471
All three insecticides were diluted with distilled water and each assay consisted of 25 apterous adult per treatment (each dose) and each treatment replicated 5 times. Plant leaf was cut (three weeks old seedlings leaf) and dipped into insecticide solution for 10 seconds and allowed to dry for 30 minutes before exposing the insects to it. For controls plant leaves were treated with distilled water alone.
Mortality was assessed after 24 hours. Mortality data were corrected with Abbott‘s formula (Abbot, 1925). Data Analysis
In these experiments concentration-mortality regression for the adult from each bioassay was evaluated statistically using probit analysis (Polo-PC Probit and Logit analysis; LeOra Software 1997) to determine the lethal concentrations (LC50s). Differences in toxicity were considered significant when 95 % Fiducial Limit (FL) did not overlap (Adams et al., 1990).
RESULTS Toxicity of Oxydemton-methyl
In this experiment 5 doses of pesticide including 0 (control), 3, 10, 30, 100 and 300 ppm were used. At the highest and the lowest concentrations 91.16 and 25 % mortality was achieved, respectively. Fifty percent mortality (LC50) was recorded at 19.62 ppm which fudicial limit was 11.41 and 30.73. Also, LC10 and LC90 were estimated to be 1.08 and 356.27 ppm, respectively (Table 1). Figure 1 shows regression line of effect of Oxydemton-methyl against B. brassicae. Toxicity of Trichlorophen
Six concentrations including 0 (control), 30, 60, 120, 240 and 480 ppm of this insecticide were used in bioassay against B. brassicae. The lowest and the highest mortality obtained were 21.27 and 87.23 %, respectively which correspond to 30 and 480 ppm used doses of the insecticide.
Lethal concentration of 10, 50 and 90% was calculated using probit analysis and estimated LC10, LC50 and LC90 were 14.30, 95.96 and 643.89 ppms, respectively (Table 1).
Figure 2 shows regression line of effect of Trichlorophen against B. brassicae. Toxicity of chlorpyriphos methyl
In the bioassay of chlorpyriphos methyl against B. brassicae six concentrations including 0 (control), 20, 40, 80, 160, and 320 ppm were used. The highest and the lowest mortality obtained were 20.83 and 93.75 % which correspond to 20 and 320 ppm insecticide concentrations.
Estimated lethal concentration of 10, 50 and 90 were equal to 14.06, 59.65 and 253.10 ppm (Table 1). Figure 3 shows regression line of chlorpyriphos methyl against B. brassicae.
DISCUSSIONS
In this study the effect of three insecticides including Trichlorophen, chlorpyriphos methyl and Oxydemton-methyl on B. brassicae in a leaf dip assay was investigated. Results showed that the most effective insecticide was Oxydemton-methyl with the LC50 value of 19.62 ppm followed by chlorpyriphos methyl with the LC50 value of 59.65 ppm and Trichlorophen with the LC50 value of

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 472
95.96 ppm. Also there are significant differences between their toxicity toward the aphid since there are not overlap between upper and lower limit of LC50s.
Although all these insecticide are from organsophosphate group that inhibits acetylcholinesterase enzyme so that disrupt nervous system, they have different toxicity toward cabbage aphid. Oxydemton-methyl is used for effective control of pest by contact and systemic action. It is used to control many destructive pests that attack certain vegetable, fruit, and field crops as well as ornamental flowers, shrubs, and trees. Primarily it is effective against aphids, mites, thrips, and sawflies.
Chlorpyrifos methyl is an organsophosphate insecticide, too and its popular trade names are Dursban, Empire and Lorsban. Chlorpyrifos is moderately toxic and chronic exposure has been linked to neurological effects, developmental disorders, and autoimmune disorders.
Chlorpyrifos methyl is used for control of pest in homes and gardens. However, since 2000 Chlorpyrifos methyl has been banned for use in homes and home garden because of health concerns in the area where it was being extensively used (Milne, 1995).
Chlorpyrifos methyl is "one of the most widely used organophosphate insecticides," according to the United States Environmental Protection Agency (EPA). The crops with the most intense chlorpyrifos methyl use are cotton, corn, almonds, and fruit trees including oranges and apples.
Trichlorfon is an organophosphate insecticide used to work both by contact and stomach poison action. It is used for vegetable, fruit and field crops. It is also used for control of insect on livestock, ornamental and forestry plantings (Simon, 2008).
Interestingly, when compare these insecticides slope it can be observed that Oxydemton-methyl, Trichlorfon and chlorpyrifos methyl have slope of equal to 1.01, 1.55 and 2.04, respectively.
Therefore, Oxydemton-methyl act on insect through contact and systemic action that is major reason why this insecticide has low LC50 and is more effective than the other two insecticides which are contact and stomach insecticides.
LITERATURE CITED Amin, A. H. & ElDefray, G. M. 1981. The Biology of the Cabbage Aphid, Brevicoryne brassicae (L.), in Egypt. Bull. Soc. Ent. Egypte, (63): 111-118. Ellis, P. R. & Singh, R. 1993. A review of the host plants of the cabbage aphid, Brevicoryne brassicae (Homoptera, Aphididae). IOBC/WPRS bulletin, 16 (5): 192-201. Ellis, S. A., Oakley, J. N., Parker, W. E. & Raw, K. 1999. The development of an action threshold for cabbage aphid (Brevicoryne brassicae) in oilseed rape in the UK. Ann. app/. Bid. (1999), 134: 153-162. Elmer, H. S. & Brawner, O. L. 1975. Control of Brown Soft Scale in Central Valley. Citrograph, 60 (11): 402-403. Flint, M. L. 1985. Cabbage Aphid, Brevicoryne brassicae. pp. 36-39. In Integrated Pest Management for Cole Crops and Lettuce. University of California Publication 3307. 112 pages. Hill, D. S. 1983. Brevicoryne brassicae (L.). pp. 154-155. In Agricultural Insect Pests of the Tropics and Their Control, 2nd Edition. Cambridge University Press, Cambridge, London, New York, New Rochelle, Melbourne, Sydney. 746 pages. Milne, G. W. A. 1995. CRC- Handbook of Pesticides. CRC-press, London, UK. Shohey, H. H. 1963. Differential Toxicity of Insecticides to the Cabbage Aphid and Two Associated Entomophagous Insect Species. Journal of Economic Entomology, 56 (6): 844-847.
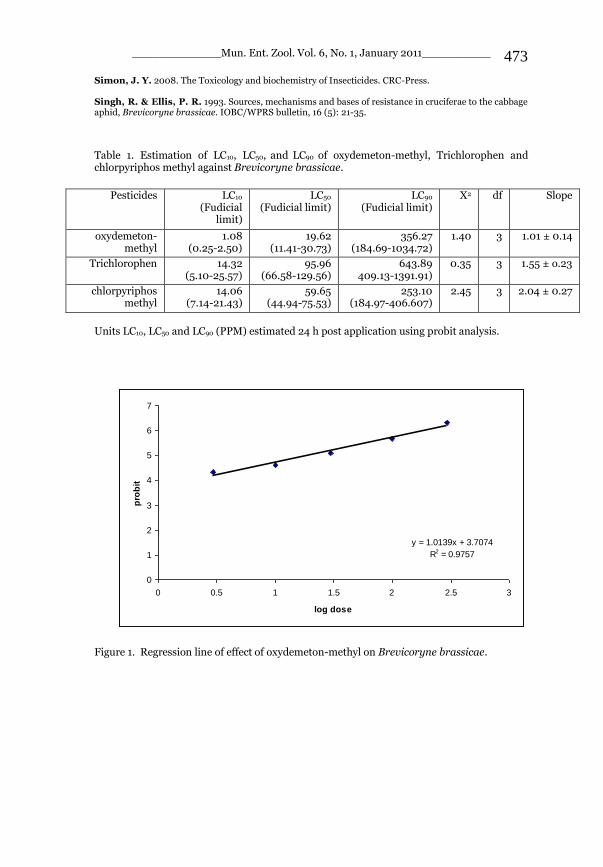
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 473
Simon, J. Y. 2008. The Toxicology and biochemistry of Insecticides. CRC-Press. Singh, R. & Ellis, P. R. 1993. Sources, mechanisms and bases of resistance in cruciferae to the cabbage aphid, Brevicoryne brassicae. IOBC/WPRS bulletin, 16 (5): 21-35.
Table 1. Estimation of LC10, LC50, and LC90 of oxydemeton-methyl, Trichlorophen and chlorpyriphos methyl against Brevicoryne brassicae.
Pesticides LC10
(Fudicial limit)
LC50
(Fudicial limit) LC90
(Fudicial limit) X2 df Slope
oxydemeton- methyl
1.08 (0.25-2.50)
19.62 (11.41-30.73)
356.27 (184.69-1034.72)
1.40 3 1.01 ± 0.14
Trichlorophen 14.32 (5.10-25.57)
95.96 (66.58-129.56)
643.89 409.13-1391.91)
0.35 3 1.55 ± o.23
chlorpyriphos methyl
14.06 (7.14-21.43)
59.65 (44.94-75.53)
253.10 (184.97-406.607)
2.45 3 2.04 ± 0.27
Units LC10, LC50 and LC90 (PPM) estimated 24 h post application using probit analysis.
y = 1.0139x + 3.7074
R2 = 0.9757
0
1
2
3
4
5
6
7
0 0.5 1 1.5 2 2.5 3
log dose
pro
bit
Figure 1. Regression line of effect of oxydemeton-methyl on Brevicoryne brassicae.
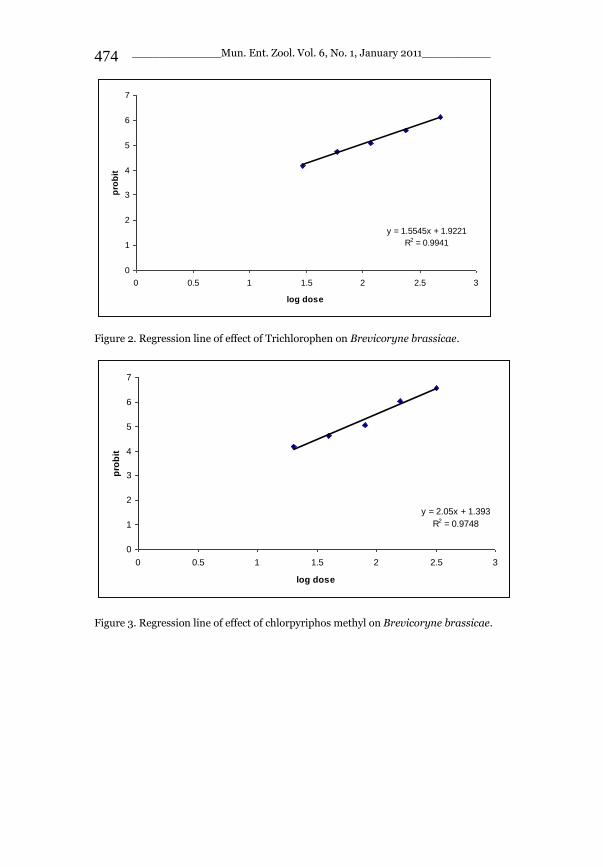
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 474
y = 1.5545x + 1.9221
R2 = 0.9941
0
1
2
3
4
5
6
7
0 0.5 1 1.5 2 2.5 3
log dose
pro
bit
Figure 2. Regression line of effect of Trichlorophen on Brevicoryne brassicae.
y = 2.05x + 1.393
R2 = 0.9748
0
1
2
3
4
5
6
7
0 0.5 1 1.5 2 2.5 3
log dose
pro
bit
Figure 3. Regression line of effect of chlorpyriphos methyl on Brevicoryne brassicae.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 475
SUBSTITUTE NAMES FOR TWO HEMIPTERAN GENERA NAMES PREOCCUPIED BY TRILOBITES GENERA
(HEMIPTERA)
Hüseyin Özdikmen* * Gazi Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü, 06500 Ankara / TÜRKĠYE, e-mail: [email protected] [Özdikmen, H. 2011. Substitute names for two hemipteran genera names preoccupied by trilobites genera (Hemiptera). Munis Entomology & Zoology, 6 (1): 475-476] ABSTRACT: Two junior homonym genus group names were detected among the hemipteran genus group names. So, the following replacement names are herein proposed: Koteya nom. nov. for Keithia Koteja, 2000 and Youngus nom. nov. for Yunnanaspis Young, 1986. Accordingly, new combinations are herein proposed for the species currently included in these genus group names. Koteya luzzii (Koteja, 2000) comb. nov. and Youngus rubus (Young, 1986) comb. nov.. KEY WORDS: nomenclatural change, homonymy, replacement name, Hemiptera.
Two previously proposed genus group names in Hemiptera are nomenclaturally invalid, as the genus group names have already been used by a different authors in Trilobita. In accordance with Article 60 of the International Code of Zoological Nomenclature, I propose substitute names for these genus names.
Family ERIOCOCCIDAE Genus KOTEYA nom. nov.
Keithia Koteja, 2000. Studies on fossils in amber, with particular reference to the Cretaceous of New Jersey. Backhuys Publishers Leiden: 183. (Insecta: Hemiptera: Eriococcidae). Preoccupied by Keithia Raymond, 1924. Proc. Boston Soc. nat. Hist., 37, 451. (Trilobita: Asaphida, Ptychaspididae).
Remarks on nomenclatural change: The name Keithia was initially introduced by Raymond, 1924 as a genus of trilobites with the type species Keithia schucherti Raymond, 1924 from Gorge Fm, Vermont, USA. Later Koteja (2000) described a scale insect genus under the same name (with the type species Keithia luzzii Koteja, 2000 by original designation from New Jersey, USA). Thus, the genus name Keithia Koteja, 2000 is a junior homonym of the genus Keithia Raymond, 1924. So I propose a new replacement name Koteya nom. nov. for the genus name Keithia Koteja, 2000. Etymology: This genus name is dedicated to J. Koteja who is current author of the preexisting genus Keithia. I preferred the word ―Koteya‖ instead of ―Koteja‖ for latinization as generic name. Summary of nomenclatural changes: Koteya nom. nov.
pro Keithia Koteja, 2000 (non Raymond, 1924) Koteya luzzii (Koteja, 2000) comb. nov.
from Keithia luzzii Koteja, 2000

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 476
Family DIASPIDIDAE Genus YOUNGUS nom. nov.
Yunnanaspis Young, 1986. Contributions from Shanghai Institute of Entomology 6: 109. (Insecta: Hemiptera: Diaspididae). Preoccupied by Yunnanaspis Zhang, 1966. Acta palaeont.sin. 14 (2): 159,169. (Trilobita: Redlichiida: Yinitidae).
Remarks on nomenclatural change: The armored scale genus Yunnanaspis Young, 1986 was established for a genus with the type species Yunnanaspis rubus Young, 1986 by original designation from Jing-dong, Yunnan, China in the family Diaspididae. Nevertheless the name Yunnanaspis is already occupied. Zhang (1966) described a trilobite genus Yunnanaspis with the type species Yunnanaspis bilongispinus Zhang, 1966 from Tsanglangpu Fm, Yunnan, China in Trilobita. Thus the genus Yunnanaspis Young, 1986 is a junior homonym of Yunnanaspis Zhang, 1966. So I suggest here that Yunnanaspis Young, 1986 should be replaced with new name Youngus as a replacement name. Etymology: This genus name is dedicated to B. L. Young who is current author of the preexisting genus Yunnanaspis. Summary of nomenclatural changes: Youngus nom. nov.
pro Yunnanaspis Young, 1986 (non Zhang, 1966) Youngus rubus (Young, 1986) comb. nov.
from Yunnanaspis rubus Young, 1986
LITERATURE CITED
International Comission of Zoological Nomenclature. 1999. International Code of Zoological Nomenclature. Fourth Edition. The International Trust for Zoological Nomenclature, London. Koteja, J. 2000b. Scale insects (Homoptera, Coccinea) from upper Cretaceous new Jersey amber. In: Grimaldi, D. A. (Ed.) Studies on fossils in amber, with particular reference to the Cretaceous of New Jersey. Backhuys Publishers, Leiden, The netherlands (498 pp.), p.: 147-229. Raymond, P. E. 1924. New Upper Cambrian and Lower Ordovician trilobites from Vermont. Proceedings of the Boston Society of Natural History, 37: 389–466. Young, B. L. 1986 New genera and species of {Diaspididae} (Coccoidea) from Yunnan and Guizhou. Contributions of the Shanghai Institute of Entomology, 6: 109. Zhang, W. 1966. [On the classification of Redlichiacea, with description of new families and new genera]. Acta Palaeontologica Sinica, 14: 135-184.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 477
DESIGN AND ASSESSMENT OF PRIMERS TO DETECT INSECT PESTS OF BRASSICA CROPS IN THE GUT CONTENTS OF
ARTHROPOD PREDATORS
Reza Hosseini*, Michael A. Keller**, Otto Schmidt** and Li Dong-Mei***
* Department of Plant Protection, College of Agriculture, University of Guilan, Rasht, IRAN. E-mail: [email protected]; [email protected] ** Plant and Pest Science, School of Agriculture, Food and Wine, The University of Adelaide, SA 5005, AUSTRALIA. *** CSIRO Molecular & Health Technologies, 11 Julius Avenue, North Ryde, NSW 2113, AUSTRALIA. [Hosseini, R., Keller, M. A., Schmidt, O. & Dong-Mei, L. 2011. Design and assessment of primers to detect insect pests of Brassica crops in the gut contents of arthropod predators. Munis Entomology & Zoology, 6 (1): 477-492] ABSTRACT: Techniques based on the polymerase chain reaction (PCR) have been shown as powerful tools for ecological studies of predator-prey interactions. Species-specific primers, which are designed to amplify target prey DNA from the gut contents of generalist predators, can potentially be used to develop highly specific and sensitive assays. We developed species-specific primers from the cytochrome oxidase subunit I (COI) gene for six insect pests of Brassica crops (Plutella xylostella, Pieris rapae, Hellula hydralis, Helicoverpa punctigera, Brevicoryne brassicae, and Myzus persicae). Specificity tests confirmed each primer pairs specifically amplifies prey DNA without cross-reactivity to predators or other non-target species, which are commonly found in the same habitats. These molecular markers also allow amplification of a very small amount of target DNA in the presence of substantially greater amounts of predator DNA. Although multiplexing of primers could potentially be used to detect the presence of multiple prey species in a single assay, the sensitivity of it compare with singleplex PCR was lower. Here we show that these primers are specific and sensitive and can be applied in ecological studies of predator-prey interactions in the field. KEY WORDS: Molecular markers, Brassica pests, PCR, cytochrome oxidase subunit I, predation.
One of the most difficult aspects of studying invertebrate predators is assessing their feeding behaviour under natural conditions. Knowledge of prey items and rates of predation are important in determining the effects of predators on prey populations and are especially critical in evaluating the effectiveness of a predator as a biological control agent (Hayes & Lockley, 1990). Unfortunately, detection of predation is technically difficult because prey and predator are relatively small and cryptic in most cases (Greenstone, 1996; Naranjo & Hagler, 1998). In order to overcome these difficulties, biochemical techniques have been developed and used to identify prey species in predators‘ diets. Monoclonal and polyclonal antibodies (Symondson, 2002) and enzyme-electrophoresis (Traugott, 2003) can be used to determine which prey has been consumed by a predator. However, amplification of specific prey DNA using the polymerase chain reaction has proven to be more specific and sensitive in detecting prey remains. It can be used to specifically detect small amounts of prey DNA in the gut contents of invertebrate predators (Hoogendoorn & Heimpel, 2001; Symondson, 2002; Agusti et al., 2003a; Harper et al., 2005; Juen & Traugott, 2005).
PCR-based techniques are rapidly replacing other molecular techniques because molecular biology facilities are widely available and prey-specific primers

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 478
can be used in different contexts once they have been designed. This technique has been successfully used for detection of a variety of prey remains in predators‘ gut contents (Zaidi et al., 1999; Chen et al., 2000; Hoogendoorn & Heimpel, 2001; Agusti et al., 2003a; Agusti et al., 2003b; Harper et al., 2005; Juen & Traugott, 2005; Read et al., 2006; Hosseini et al., 2006b, 2008).
Brassica vegetables and Oilseeds are economically important crops; approximately 3.1 and 26.1 million ha respectively were grown worldwide in 2004 (Food and Agriculture Organization of the United Nations, 2007). These crops are associated several destructive and widespread insect pests. Total damage caused by these pests is substantial; for example management costs for P. xylostella alone were estimated at US$ 1 billion annually in 1997 (Shelton et al., 1997). In Australia, the pests of Brassica crops include the lepidopterans Plutella xylostella (L.), Pieris rapae (L.), Hellula hydralis Guenee and Helicoverpa punctigera Wallengren, and two aphids, Brevicoryne brassicae (L.) and Myzus persicae (Sulzer). Integrated pest management (IPM) systems and the use of biological control methods are preferred approaches to controlling these pests over insecticides due to the prevalence of insecticide resistance in diamondback moth (Shelton et al., 1997).
Generalist predators can play a major role in the control of agricultural pests (Symondson et al., 2002). Studies showed Brassica crops have a rich fauna of predators. Shelton et al. (1983) reported on extensive ground-dwelling predators in cabbage fields in central New York State, and Schmaedick & Shelton (2000) have documented a list of predators associated with P. rapae in cabbage fields of New York State. Hosseini et al. (2006a) reported a range of predators associated with Brassica pests in South Australia. The impact of predators on pests of Brassica crops has not been thoroughly studied and therefore their potential in suppression of major pests of Brassica has not been elucidated. Hooks et al. (2003), found broccoli plants protected by birds and spiders as predators sustained less damage from caterpillars and the plants had greater productivity compared to control plants. In cage exclusion experiments, between 7% and 81% of immature stages of P. xylostella were lost due to predation (Wang et al., 2004). Subsequent extensive research (Furlong et al., 2004) confirmed estimated losses of P. xylostella due to predation of between 2%-85%. Hence, for this one insect, predators are known to cause considerable mortality at times. It is essential to develop a reliable technique to evaluate the diets of key predatory species in order to understand their role in suppressing pests like P. xylostella.
Ma et al. (2005) developed a species-specific marker for P. xylostella based on the internal transcribed spacer (ITS-1) of the ribosomal gene. This specific primer pair was used to detect prey in the gut contents of two polyphagous predators, Nabis kinbergii and Trochosa expolita (reported as Lycosa sp.). In this pilot study of predation of diamondback moth on cauliflower and broccoli farms near Virginia, South Australia, the remains of P. xylostella were detected in the gut contents of both species of field-collected predators.
As a mitochondrial gene, it occurs as multiple copies in each cell (Hoy, 2003), which increases the likelihood of successful amplification of prey residues in the predators‘ gut contents. This gene is a protein-coding gene that has a high level of interspecific variability (Zhang & Hewitt, 1996), which allows closely related species to be separated. Therefore, in the current study, cytochrome oxidase subunit I (COI) was selected as a potentially diagnostic gene.
The aims of the present study were: 1) to develop species-specific primers for each of the six major pests of Brassica crops; 2) to test their specificity and sensitivity against non-target species and other common predators (A false

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 479
positive results can affect the interpretation of predation, especially when generalist predators can feed on a wide range of prey species (Sunderland, 1975); thus the specificity and sensitivity of each primer set are critical and have to be tested before any application in field studies); and 3) to test the potential of multiplexing of two primer pairs to assess for simultaneous detection of DNA from two different target prey species.
MATERIALS AND METHODS Sample collection
Six species of crop pests of Brassica spp. and predators and non-predatory arthropods were collected from different farms in South Australia, but mainly from Pitchford‘s broccoli farm (Currency Creek, South Australia) during 2004 and 2005 [Table 1; e.g. P. xylostella, P. rapae, H. hydralis, H. punctigera, B. brassicae, M. persicae, Nabis kinbergii (Reuter), Hippodamia variegate (Goeze), Trochosa expolita (L.Koch) and Oechalia schellenbergii (Guerin-Meneville)]. In addition, P. xylostella and P. rapae were obtained from a laboratory culture. Collection was done by hand, vacuum sampler or insect net for day active arthropods and with a headlamp at night for wolf spiders (Wallace, 1937). Identification of species (pests and predators) was done morphologically following the most current relevant taxonomic keys and confirmed by comparison to identified specimens in the Waite Insect and Nematode Collection (Waite Campus, The University of Adelaide, South Australia). Wolf spider samples were identified by Volker W. Framenau (Department of Terrestrial Invertebrates, Western Australian Museum, Perth, WA). Generally, collected samples were preserved either in 70% ethanol or stored at -20ºC for future molecular work. Some live predators were kept individually in a plastic cup (7.5 cm diam.× 4.5 cm) provided with a piece of wet cotton wool in the laboratory at room temperature for subsequent feeding experiments. DNA extraction
DNA of individual specimens was extracted from legs in order to avoid contamination with gut contents but DNA of predators used for feeding trails was extracted from the whole body. Each sample was homogenised in 1.5 ml microcentrifuge tubes using a clean and sterile plastic pestle in 400 µl homogenisation buffer (10mM Tris-HCl pH 8.0, 10mM EDTA pH 8.0, 1% SDS). After adding 5 µl proteinase K (20 µg/µl) (Sigma), samples were incubated at 56ºC for 15 min. For removal of RNA, 0.5 µl RNase-A (10mg/ml) (Sigma) was added and the tube was incubated at 37ºC for 15 min. Extraction was done once with Phenol/Chloroform. DNA was precipitated by adding two volumes of cold ethanol (kept at -20ºC overnight) and 0.2 volume of 5 M sodium chloride and then stored at -20ºC for 15 min. After centrifugation at 13000 g for 15 min, the pellet was washed with 70% ethanol and then dried at 65ºC for 5 min. Pellets were resuspended in 200 µl of TE (10mM Tris-HCl pH 8.0, 1mM EDTA) and resulting DNA suspensions were stored at -20ºC. DNA of six species of Brassica plants was extracted by the same Phenol/Chloroform method from the leaves (Table 1).
Preliminary studies showed DNA extracted by Phenol/Chloroform method from the whole bodies of spiders could not be reliably amplified by PCR. Therefore DNA from spiders was extracted using a method that incorporated silica (Hosseini et al., 2007).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 480
DNA concentration was measured for all DNA extracts by NanoDrop (ND-1000 spectrophotometer; NanoDrop technologies Inc., Wilmington, DE, USA) according to the manufacturer‘s manual. PCR and sequencing
Two universal primers, C1-J-1718 as forward and C1-N-2191 as reverse primer which they have proved to work very well with all insects tested (Simon et al., 1994), were used to amplify a portion of the mitochondrial cytochrome oxidase subunit I (COI) gene of the six species of Brassica pests. Amplification was performed in 50 µl total volume of reaction buffer containing 150 µM dNTPs (Fisher Scientific Inc., USA), 2 mM MgCl2, 0.4 µM each primer, 1 U of Taq DNA polymerase (Biotech international Ltd., Australia) and 8 µl of DNA template (20-40 ng). The reaction mix was put into a 0.2 ml PCR tube and amplification was performed in a PTC-200 thermocycler (MJ Research, MA, USA) with the following temperature profile: 95ºC for 2 min followed by 35 cycles of 94ºC for 1 min, 56ºC for 1 min and 72ºC for 1 min and a final elongation at 72ºC for 5 min.
PCR products were separated by electrophoresis on a 1.8% agarose gel in TAE or TBE containing ethidium bromide for DNA staining. COI specific fragments of each species were sliced from the agarose gel and purified using a Perfectprep® Gel Cleanup kit (Eppendorf AG, Hamburg, Germany), following the manufacturer‘s instructions. Sequencing reactions were carried out via the manufacturer‘s instructions using ABI PRISM® Big-Dye Terminator Cycle Sequencing ready reaction mix version 3 (Applied Biosystems, Foster City, California, USA) in both forward and reverse orientations. Sequence alignment and primer design
COI fragments were sequenced from two individuals per species in both forward and reverse directions. Sequencing results were reviewed by the CHROMAS program (version 1.45)(http://www.technelysium.com.au) and SeqEd Version 1.3 (Macintosh) and edited manually for each species separately. The Basic Local Alignment Search Tool (BLAST) was used to compare the similarity of nucleotide sequences with sequences present in GeneBank database (http://www.ncbi.nlm.nih.gov/blast/). All edited sequences were aligned using GENEDOC (http://www.psc.edu/biomed/genedoc). Pairs of primers were designed for each of the six species according to their sequence variations, especially in regions that were unique to each species. Primer design guidelines proposed for the design of efficient and specific primers by Innis & Gelfand (1990) and Saiki (1990) were followed. The primer-primer interactions were analysed using the program ―Oligonucleotide Properties Calculator‖ (http://www.basic.northwestern.edu/biotools/oligocalc.html). Primers were synthesized by Geneworks, Adelaide, South Australia. For optimisation of each primer pair, a gradient PCR program was performed by using gradient thermocycler (PTC-200) with the following temperature profile: 35 cycles at 94ºC for 30 sec, 53ºC as the lower temperature and 65ºC as the higher temperature for 30 sec, 72ºC for 60 sec. A first cycle of denaturation was carried out at 95ºC for 2 min and a last cycle of extension was performed at 72ºC for 5 min. Singleplex and multiplexing of PCR primers
Singleplex PCR was performed in 25 µl total volume of reaction buffer containing 150µM dNTPs (Fisher Scientific Inc., USA), 2 mM MgCl2 , 0.4 µM each of primer pairs for each species (Table 2), 1 U of Taq DNA polymerase (Biotech international Ltd., Australia), 1× of reaction buffer provided by manufacturer and

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 481
4 µl of DNA template. To evaluate the ability of multiplex PCR for simultaneous detection of two target prey DNA, a multiplex PCR procedure was developed and optimised to multiplex the primer pairs DBM-F-2/DBM-R1-1 and HH-F-1/HH-R-1 (Table 2) for P. xylostella and H. hydralis, respectively. These primers have the same annealing temperature but each pair can amplify fragments of different size. Based on a 25 µl reaction volume, the multiplex PCR reaction mix consisted of same materials and concentrations as singleplex, where 0.4 µM each of the P. xylostella primers, 0.2 µM each of the H. hydralis primers and 0.8 µg/µl BSA (Promega, Madison, USA) were used in multiplexing reaction. The cycling conditions for singleplex PCR were 95ºC for 2 min followed by 35 cycles of 94ºC for 30 sec, 30 sec at the specific annealing temperature for each specific primer pairs (Table 2), and 72ºC for 1 min and final extension at 72ºC for 5 min.
For multiplex PCR, annealing temperature used in cycling program was 58ºC for 30 sec and the rest of conditions were the same as singleplex. Primer specificity and sensitivity
The specificity of the primer pairs was tested separately for each of the designed primers by attempting to amplify target DNA (at least 10 individuals) from six Brassica pests and other arthropods, including a representative sample of most abundant Diptera, Hymenoptera, and selected arthropod predators as well as an aphid and a lepidopteran, and six species of Brassica crops collected from the field (Table 1). In order to test if the designed primers would amplify specific DNA in the presence of predator DNA even in high amounts, extracted DNA of each species was diluted and mixed with DNA of a selected predator (T. expolita). The concentration of the predator‘s DNA was 20 ng and constant in all mixtures, while the ratio of predator DNA to target species DNA mixtures varied (Table 3). Specificity and Sensitivity of Multiplex PCR
Apart from testing specificity and sensitivity of primers in the singleplex PCR, separate assays were performed to test specificity and sensitivity of multiplex PCR. Primer pairs DBM-F-2/DBM-R1-1 and HH-F-1/HH-R-1 were used for multiplexing. Four different DNA template mixtures were tested in multiplex PCR: 1) a serial dilution of P. xylostella DNA mixture with DNA of predator, 2) a serial dilution of P. xylostella DNA mixture with a 0.8 ng constant DNA of H. hydralis and DNA of predator 3) a serial dilution of H. hydralis DNA mixture with a DNA of predator, and 4) a serial dilution of H. hydralis DNA mixture with a 1 ng constant DNA of P. xylostella and DNA of predator. The concentrations of serial dilutions for P. xylostella and H. hydralis DNA ranged from 16.8 ng to 2 pg and 11.6 ng to 0.02 pg, respectively and the concentration of the predator‘s DNA (T. expolita) was 20 ng and constant in all mixtures. Detection of prey DNA in gut contents of predators
Four experiments were performed to test the ability to detect DNA from five different preys‘ species in the gut contents of predators. In the first experiment, three species of field-collected predators (5 specimens each of Trochosa expolita, Nabis kinbergii and Hippodamia variegata) were starved at room temperature for at least seven days. After this period each predator was fed a 2nd or 4th instar of P. xylostella. In the second experiment 5 starved T. expolita were fed on a 2nd instar of P. rapae. In the third assay 6 starved T. expolita were fed on a 2nd –3rd instar of H. hydralis and in the fourth experiment which has been done in two separate assay, starved H. variegata (5-6 specimens for each assay) were fed on

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 482
5-10 nymphs of M. persicae or B. brassicae. Predators were frozen at –20 ºC zero to two hour after consuming their prey for subsequent molecular assay. Field samples
In order to test the ability of primers to detect predation in the field, adult of an abundant predator ―Oechalia schellenbergii” (n=82) were collected at random from a broccoli field on a commercial vegetable farm at Currency Creek, South Australia (35º 41´ S, 138º 75´ E). All collected specimens were kept chilled on ice until they were transferred to the laboratory and placed at –80 ºC overnight prior to DNA extraction and PCR using methods and conditions described before.
RESULTS Primer design and specificity and sensitivity
The readable fragments sequence from the COI gene of each of these six species varied in length (P. xylostella 481 bp, P. rapae 478 bp, B. brassicae 502 bp, H. punctigera 514 bp, M. persicae 478 bp and H. hydralia 492 bp). Sequences were submitted to the National Centre for Biotechnology Information Genebank (http://www.ncbi.nlm.nih.gov; Table 2). The sequences of these species were aligned and, on the basis of diagnostic differences among sequences, one species-specific pair of primers was designed for each species. Optimised annealing temperatures ranged from 58ºC to 64ºC for each primer pair (Table 2). The target sequences amplified by the six primers pairs range from 200 to 307bp (Table 2, Fig. 1). Each primer pairs proved to be highly specific against non-target DNA and could amplify the expected fragment size only in the presence of the respective target species DNA (Fig. 1).
Singleplex PCR showed that the highest and lowest detection sensitivity among the designed primers belong to H. hydralis with 0.02 pg and H. punctigera with 27.3 pg (Table 3), where the key pest species ―P. xylostella” could be detected with as little as 2 pg (Fig. 3). Specificity and sensitivity of Multiplex PCR
Multiplex PCR with DBM-F-2/DBM-R1-1 and HH-F-1/HH-R-1 demonstrated the possibility of simultaneous detection of P. xylostella and H. hydralis (Fig. 4) and specificity against other non-target species.
Multiplex PCR showed less sensitivity compare with singleplex PCR. In the multiplex PCR assay, detection sensitivity of H. hydralis primers in the mixture with predator DNA was 8-fold lower while in the mixture with P. xylostella DNA showed a 140-fold lower sensitivity compared to singleplex PCR. Likewise, P. xylostella primer pairs in the mixture with predator DNA had the same sensitivity as singleplex PCR, however their sensitivity in the mixture with H. hydralis DNA was 8.2-fold lower than with singleplex PCR (Table 4). Detection of prey DNA in the gut contents of predators
DNA of target pests P. xylostella, P. rapae, H. hydralis, M. persicae and B. brassicae (Fig. 2A) were detected in the gut contents of T. expolita, N. kinbergii and H. variegata (Fig. 2B). Field samples
Results showed all designed primers can be effectively used for detection of target prey DNA in collected predators from the field. DNA from all six pests was found in the gut contents of O. schellenbergii, which indicates that, this species is

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 483
a polyphagous predator (Fig. 5). Fig.6 as an example shows detection of remains of four preys including P. xylostella, H. Hydralis, P. rapae and B. brassicae in 10 predator specimens collected from the field.
DISCUSSION
Species-specific molecular markers were developed for six Brassica pests. The designed primer pairs were tested against many possible non-target invertebrate species found in broccoli fields as well as Brassica plants with no amplification detected for non-target species, which indicates that these primer pairs were highly specific for the target species. In the only published study of molecular detection of Brassica crop pests in predators‘ gut contents, Ma et al. (2005) showed that using a primer set based on the ITS-1 region not only amplified target DNA but also larger non-specific fragments from Brassica plants.
It has been known that prey DNA in a predators‘ gut is fragmented by digestion enzymes and as a result the detection time of prey DNA depends on the length of the amplification product (Hoogendoorn & Heimpel, 2001; Agusti et al., 2003a). Larger fragments become undetectable in the gut more rapidly than smaller ones (Zaidi et al., 1999; Agusti et al., 2000). However, some studies showed there is no difference in detection rate within certain range of fragment lengths. For example Chen et al. (2000) found no difference in the detection rates of S. avenae fragments shorter than 246bp. In similar cases, studies showed there was not difference in detection rates of DNA fragments between 175 and 387 bp in feeding experiments with cockchafer prey (Juen & Traugott, 2005), and detection for a 127 bp fragment was not significantly higher than for a 463 bp fragment of the A. solstitiale (Juen & Traugott, 2006). For this reason in this study we aimed to identify specific sequences that are 300 bp or smaller to ensure reasonable detection times.
The sensitivity thresholds of species-specific primers in prey detection achieved by PCR is an important issue because it indicates whether a single prey consumed by a predator is sufficient for detection (e.g. Zaidi et al., 1999; Chen et al., 2000; Admassu et al., 2006). In our study a concentration-response trial of prey DNA in the presence of a constant concentration of predator DNA was performed for all six primers pairs. Detection limits ranged from 0.02 pg to 27.3 pg (Table 3). This result is comparable to the highest detection sensitivity levels achieved in a study on used to detect parasitism; Traugott et al. (2006) reported detection limits ranging between 0.6 pg and 46.8 pg of DNA from a parasitoid of P. xylostella. Our results showed that the presence of a second non-target DNA in the singleplex PCR does not influence the detectability of target DNA in all primers pairs examined. For example H. hydralis DNA was detectable in the presence of 106 times the concentration of T. expolita DNA. This enables us to use the whole body of a predator for DNA extraction without any dissection of the gut or its contents. This is a big advantage in field studies and makes the analysis of samples with small predators easier and quicker. In a study of earthworm residues in predators‘ gut contents by Admassu et al. (2006), sensitivity of designed primers was determined to be 0.15 ng/µl of earthworm DNA in the presence of 243 times the amount of predators‘ DNA. Likewise, Sheppard et al. (2004) showed that, despite the presence of large amounts of predator tissue, there was no evidence that non-target material could mask the detection of very small amounts of prey DNA.
Some workers used another approach to determine the sensitivity threshold of primers. For example, Agusti et al. (2003b) in their study were defined the

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 484
sensitivity of designed primers at 10-5 dilution of a target sample and Chen et al. (2000) detected 10-7 aphid equivalents of DNA. However, concentration of the extracted DNA depends on the size of sample and extraction method used therefore results will vary with smaller or larger specimens.
Our results revealed the possibility of applying multiplex PCR for simultaneous detection of at least two prey species in one reaction. This is a big advantage in increasing the efficiency of PCR amplification. However, the result indicated that the ratio of concentration among DNA of target prey is an important factor in detection of each prey in multiplexing system. On the other hand, if the concentration of H. hydralis is higher than P. xylostella, the latter prey may either not be detected or only observed as a very faint band in the gel electrophoresis. A possible explanation may be the competition among DNA of target species. For this reason sensitivity of primers in multiplex was less than singleplex PCR. This was obvious when we mixed DNA of H. hydralis with a constant concentration of P. xylostella and vice versa (Table 4). In a multiplex PCR assay with primers for P. xylostella and one of its parasitoids, detection sensitivity of the parasitoid primers was determined to be 4-fold lower compared to singleplex PCR, but the sensitivity of P. xylostella primers was not determined in the multiplex PCR reaction (Traugott et al., 2006). Harper et al. (2005) developed a multiplex PCR incorporating fluorescent markers to detect mitochondrial DNA fragments from more than 10 prey species simultaneously in the gut contents of generalist predators. However, the higher cost of techniques and equipment used is a limiting factor in the application of this method in ecological investigations. Moreover the sensitivity of the multiplex systems was not tested with varying ratios of prey DNA to check the effect on detection of each species.
Competition between DNA from different target species is a known phenomenon in multiplex PCR (Markoulatos et al., 2002). For example, in a study on simultaneous detection of four bacterial pathogens (Stralin et al., 2005), when there was a clear dominant band in the gel electrophoresis from one pathogen, a relatively weaker PCR band was identified for the pathogen with the lower concentration. Consequently a strong band for one pathogen in the gel electrophoresis may be associated with a decreased sensitivity for detection of other species by multiplex PCR. Therefore, we suggest further investigation should be done to check the real sensitivity of each primer in multiplex PCR before any application in the ecological studies.
This study has demonstrated that species-specific primers targeting six pests of Brassica crops can be used for ecological studies of Brassica pests and to screen field-caught predators for prey species consumption. This study showed that Oechalia schellenbergii collected from the field is a polyphagous predator and could be considered as a potential predator of pests in Brassica crops. This fact has previously been reported in other crops systems that this species feed on a wide range of immature stages of moths, beetles, sawflies and weevils (Mensah, 1999; Copper, 1981; Cordingly, 1981; Awan, 1985a,b; 1988; 1990) and other coleopteran pests (Edwards & Suckling, 1980). Multiplexing of primers has potential for application in ecological investigations but further studied is needed to establish the sensitivity of the method in each case.
ACKNOWLEDGEMENTS This study was supported in part by a grant from Horticulture Australia Ltd.
R. Hosseini was supported by a scholarship from the University of Guilan, Rasht, Iran. We wish to thank Volker W. Framenau for identification of wolf spiders.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 485
LITERATURE CITED Admassu, B., Juen, A. & Traugott, M. 2006. Earthworm primers for DNA-based gut content analysis and their cross-reactivity in a multi-species system. Soil Biology and Biochemistry, 38: 1308-1315. Agusti, N., De Vicente, M. C., & Gabarra, R. 2000. Developing SCAR markers to study predation on Trialeurodes vaporariorum. Insect Molecular Biology, 9: 263-268. Agusti, N., Shayler, S. P., Harwood, J. D., Vaughan, I. P., Sunderland, K. D., & Symondson, W. O. C. 2003a. Collembola as alternative prey sustaining spiders in arable ecosystems: prey detection within predators using molecular markers. Molecular Ecology, 12: 3467-3475. Agusti, N., Unruh, T. R., & Welter, S. C. 2003b. Detecting Cacopsylla pyricola (Hemiptera: Psyllidae) in predator guts by using COI mitochondrial markers. Bulletin of Entomological Research, 93: 179-185. Awan, M. S. 1985a. Anti-predator plays of Heliothis punctigera (Lep: Noctuidae) caterpillar against the predator Oechalia schellenbergii (Hem.: Pentatomidae). Australian Journal of Zoology, 33: 885-890. Awan, M. S. 1985b. Studies on the food preferences of immature stages of predacious pentatomids, Oechalia schellenbergii (Guerin-Meneville) and Cermatulus nasalis (Westwood). Sindh University Research Journal (Sci-Ser.), 17: 69-74. Awan, M. S. 1988. Development and mating behaviour of Oechalia schellenbergii (Guérin-Méneville) and Cermatulus nasalis (Westwood) (Hemiptera: Pentatomidae). Journal of the Australian Entomological Society, 27: 183-187. Awan, M. S. 1990. Predation by three hemipterans: Tropiconabis nigrolineatus, Oechalia schellenbergii and Cermatulus nasalis, on Heliothis punctiger larvae in two types of searching arenas. Entomophaga, 35: 203-210. Chen, Y., Giles, K. L., Payton, M. E., & Greenston, M. H. 2000. Identifying key cereal aphid predators by molecular gut analysis. Molecular Ecology, 9: 1887-1898. Cooper, D. J. 1981. The role of predatory Hemiptera in disseminating a nuclear polyherosis virus of Heliothis punctiger. Journal of the Australian Entomological Society, 20: 145-150. Cordingly, C. L. 1981. Factors causing the mortality of the grape vine moth, Phalaenoides glycine Lewin (Lepidoptera: Agaristidae). Journal of the Australian Entomological Society, 20: 33-38. Edwards, R. B., & Suckling, D. M. 1980. Cermatulus nasalis and Oechalia schellenbergii (Hemiptera: Pentatomidae) as predators of eucalyptus tortoise beetle larvae, Paropsis charybdis (Coleoptera: Chrysomelidae), in New Zealand. New Zealand Entomologist, 7: 158-164. Food and Agriculture Organization of the United Nations. 2007. FAOSTAT: Core production data. faostat.fao.org [accessed 6 February 2007]. Furlong, M. J., Zuhua, S., Shijan, G., Yinquan, L., Sheng, L. S. & Zalucki, M. P. 2004. Quantitative evaluation of the biotic mortality factors affecting diamondback moth in south-east Queensland, Australia. pp. 185-193 in Endersby NM & Ridland PM (Eds.) The management of diamondback moth and other crucifer pests: Proceedings of the 4th International Workshop, Melbourne, Australia, 26-29 November 2001. Greenstone, M. H. 1996. Serological analysis of arthropod predation: past, present and future. In Symondson WOC (Ed.) The Ecology of Agricultural Pests: Biochemical Approaches. Chapman & Hall, London. Harper, G. L., King, R. A., Dodd, C. S., Harwood, J. D., Glen, D. M., Bruford, M. W. & Symondson, W. O. C. 2005. Rapid screening of invertebrate predators for multiple prey DNA targets. Molecular Ecology, 14: 819-827. Hayes, J. L. & Lockley, T. C. 1990. Prey and nocturnal activity of wolf spiders (Aranea:Lycosidae) in cotton fields in the Delta region of Mississippi. Environmental Entomology, 19: 1512-1518.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 486
Hoogendoorn, M. & Heimpel, G. E. 2001. PCR-based gut content analysis of insect predators: using ribosomal ITS-1 fragments from prey to estimate predation frequency. Molecular Ecology, 10: 2059-2067. Hooks, C. R., Pandey, R. R. & Johnson, W. M. 2003. Impact of avian and arthropod predation on lepidopteran caterpillar densities and plant productivity in an ephemeral agroecosystem. Ecological Entomology, 28: 522-532. Hosseini, R., Keller, M. A., Schmidt, O. & Baker, G. J. 2006a. Predators associated with crop pests of Brassica spp. in South Australia. 5th International Workshop on management of the Diamondback Moth and Other Insect Pests. Beijing, China Oct 24-27, 2006. Pp. 338-344. Hosseini, R., Keller, M. A., Schmidt, O. & Dong-Mei, L. 2006b. Molecular detection of common pests of Brassica crops in the guts of predators. 5th International Workshop on management of the Diamondback Moth and Other Insect Pests. Beijing, China Oct 24-27. Hosseini, R., Keller, M. A., Schmidt, O. & Framenau, V. W. 2007. Molecular identification of wolf spiders (Araneae: Lycosidae) by multiplex polymerase chain reaction. Biological Control, 40: 128-135. Hosseini, R., Keller, M. A. & Schmidt, O. 2008. Molecular detection of Plutella xylostella (L.) (Lepidoptera; Plutellidae) in the wolf spider gut using COI mitochondrial markers. 18th Plant Protection Congress. Hamedan University, Iran 24-28 Aug. Hoy, M. A. 2003. Insect Molecular Genetics, an Introduction to Principles and Applications, 2nd edn. Academic Press, San Diego. Innis, M. A. & Gelfand, D. H. 1990. Optimization of PCRs. pp. 3-12. in Innis, M. A., Gelfand, D. H., Sninsky, J. J. & White, T. J. (Eds) PCR Protocols. San Diego, Academic Press. Juen, A. & Traugottm M. 2005. Detecting predation and scavenging by DNA gut content analysis: a case study using a soil insect predator-prey system. Oecologia, 142: 344-352. Juen, A. & Traugott, M. 2006. Amplification facilitators and multiplex PCR: Tools to overcome PCR-inhibition in DNA-gut-content analysis of soil-living invertebrates. Soil Biology and Biochemistry, 38: 1872-1879. Ma, J., Li, D., Keller, M., Schmidt, O. & Feng, X. 2005. A DNA marker to identify predation of Plutella xylostella (Lep., Plutellidae) by Nabis kinbergii (Hem., Nabidae) and Lycosa sp. (Aranaea, Lycosidae). Journal of Applied Entomology, 129: 330-335. Markoulatos, P., Siafakas, N. & Moncany, M. 2002. Multiplex Polymerase Chain Reaction: A Practical Approach. Journal of Clinical Laboratory Analysis, 16: 47-51. Naranjo, S. E. & Hagler, J. R. 1998. Characterizing and estimating the impact of heteropteran predation. pp. 170-197 in Colle, M. & Ruberson, J (Eds) Predatory Heteroptera: their ecology and use in biological control. Entomological Society of America, Lanham, Maryland. Read, D. S., Sheppard, S. K., Bruford, M. W., Glen, D. M. & Symondson, W. O. C. 2006. Molecular detection of predation by soil micro-arthropods on nematodes. Molecular Ecology, 15: 1963-1972. Saiki, R. K. 1990. Amplification of genomic DNA. In PCR Protocols. Pp. 13-20 in Innis, M.A., Gelfand, D.H., Sninsky, J.J. & White, T.J. (Eds) PCR Protocols. San Diego, Academic Press. Schmaedick, M. A., & Shelton, A. M. 2000. Arthropod predators in cabbage (Cruciferae) and their potential as naturally occurring biological control agents for Pieris rapae (Lepidoptera: Pieridae). The Canadian Entomologist, 132: 655-675. Shelton, A. M., Andarolo, J. T. & Hoy, C. W. 1983. Survey of ground-dwelling predaceous and parasitic arthropods in cabbage fields in upstate New York. Environmental Entomology, 12: 1026-1030. Shelton, A. M., Perez, C. J., Tang, J. D. & Vandenberg, J. D. 1997. Prospects for novel approaches towards management of the diamondback moth. pp 7-20 in Sivapragasm A, Loke WH, Hussan, AK & Lim GS (Eds.) Proceedings of the Third International Workshop on the Management of Diamondback Moth and Other Crucifer Pests. Kuala Lumpur.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 487
Sheppard, S. K., Henneman, M. L., Memmott, J. & Symondson, W. O. C. 2004. Infiltration of alien predators into invertebrate food webs in Hawaii: a molecular approach. Molecular Ecology, 13: 2077-2088. Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H. & Flook, P. 1994. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction prime. Annals of the Entomological Society of America, 87: 651-701. Stralin, K., Backman, A., Holmberg, H., Fredlund, H. & Olcen, P. 2005. Design of a multiplex PCR for Streptococcus pneumoniae, Haemophilus influenzae, Mycoplasma pneumoniae and Chlamydophila pneumoniae to be used on sputum samples. Acta Pathologica, Microbiologica et Immunologica Scandinavica, 113: 99-111. Sunderland, K. D. 1975. The diet of some predatory arthropods in cereal crops. Journal of Applied Ecology, 12: 507-515. Symondson, W. O. C. 2002. Molecular identification of prey in predator diets. Molecular Ecology, 11: 627-641. Symondson, W. O. C., Sunderland, K. D. & Greenstone, M. 2002. Can generalist predators be effective biocontrol agents? Annual Review of Entomology, 47: 561-595. Traugott, M. 2003. The prey spectrum of Cantharis-beetle larvae and adults in the field: an electrophoretic approach. Pedobiologia, 47: 161-169. Traugott, M., Zangerl, P., Juen, A., Schallhart, N. & Pfiffner, L. 2006. Detecting key parasitoids of lepidopteran pests by multiplex PCR. Biological Control, 39: 39-46. Wallace, H. K. 1937. The use of the headlight in collecting nocturnal spiders. Entomological News, 48: 107-111. Wang, X. G., Duff, J., Keller, M. A., Zalucki, M. P., Liu, S. S. & Bailey, P. 2004. Role of Diadegma semiclausum (Hymenoptera: Ichneumonidae) in controlling Plutella xylostella (Lepidoptera: Plutellidae): Cage exclusion experiments and direct observation. Biocontrol Science and Technology, 14: 571-586. Zaidi, R. H., Jaal, Z. & Hawkes, N. J. 1999. Can multiple-copy sequences of prey DNA be detected amongst the gut content of invertebrate predators? Molecular Ecology, 8: 2081-2087. Zhang, D. X. & Hewitt, G. M. 1996. Nuclear integrations: challenges for mitochondrial DNA markers. Trends in Ecology and Evolution, 11: 247-251.
Figure 1. Agarose gel electrophoresis of polymerase chain reaction (PCR) amplified DNA using all six pest specific primer pairs for the target species, lane 1, 100 bp molecular marker, lanes 2-3 P. xylostella (293 bp), lanes 4-5, H. hydralis (200 bp), lanes 6-7, B. brassicae (307 bp), lanes 8-9, H. punctigera (270 bp), lanes 10-11, P. rapae (222 bp), lanes 12-13, M. persicae (247 bp), lane14, negative control (Predator‘s DNA).
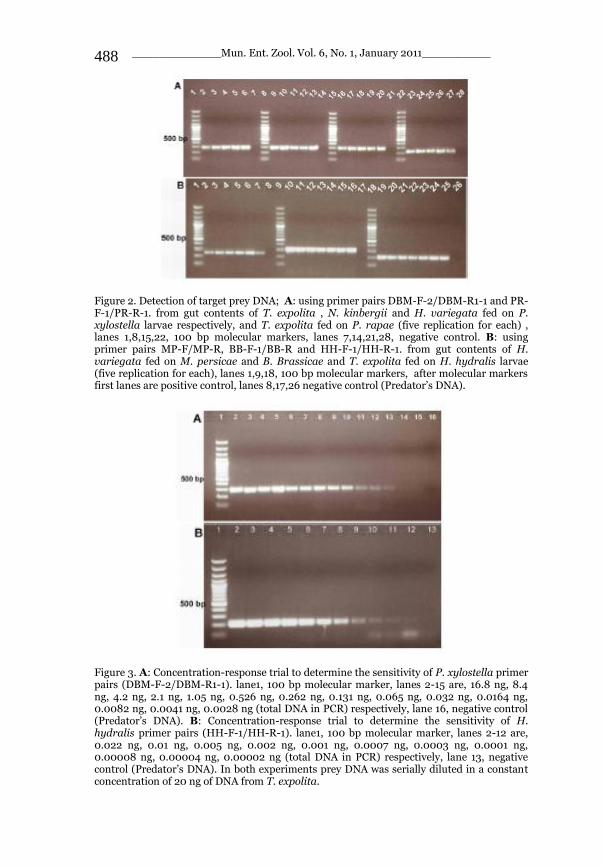
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 488
Figure 2. Detection of target prey DNA; A: using primer pairs DBM-F-2/DBM-R1-1 and PR-F-1/PR-R-1. from gut contents of T. expolita , N. kinbergii and H. variegata fed on P. xylostella larvae respectively, and T. expolita fed on P. rapae (five replication for each) , lanes 1,8,15,22, 100 bp molecular markers, lanes 7,14,21,28, negative control. B: using primer pairs MP-F/MP-R, BB-F-1/BB-R and HH-F-1/HH-R-1. from gut contents of H. variegata fed on M. persicae and B. Brassicae and T. expolita fed on H. hydralis larvae (five replication for each), lanes 1,9,18, 100 bp molecular markers, after molecular markers first lanes are positive control, lanes 8,17,26 negative control (Predator‘s DNA).
Figure 3. A: Concentration-response trial to determine the sensitivity of P. xylostella primer pairs (DBM-F-2/DBM-R1-1). lane1, 100 bp molecular marker, lanes 2-15 are, 16.8 ng, 8.4 ng, 4.2 ng, 2.1 ng, 1.05 ng, 0.526 ng, 0.262 ng, 0.131 ng, 0.065 ng, 0.032 ng, 0.0164 ng, 0.0082 ng, 0.0041 ng, 0.0028 ng (total DNA in PCR) respectively, lane 16, negative control (Predator‘s DNA). B: Concentration-response trial to determine the sensitivity of H. hydralis primer pairs (HH-F-1/HH-R-1). lane1, 100 bp molecular marker, lanes 2-12 are, 0.022 ng, 0.01 ng, 0.005 ng, 0.002 ng, 0.001 ng, 0.0007 ng, 0.0003 ng, 0.0001 ng, 0.00008 ng, 0.00004 ng, 0.00002 ng (total DNA in PCR) respectively, lane 13, negative control (Predator‘s DNA). In both experiments prey DNA was serially diluted in a constant concentration of 20 ng of DNA from T. expolita.
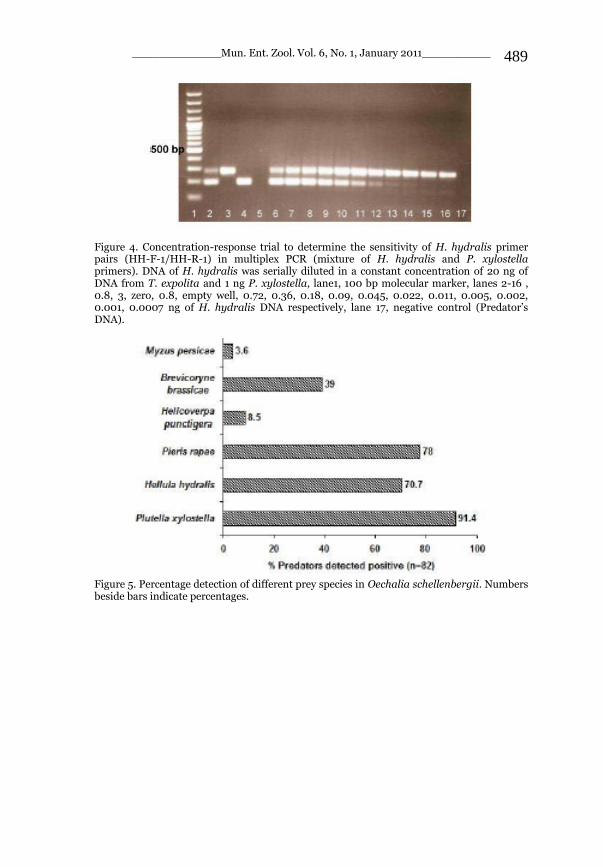
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 489
Figure 4. Concentration-response trial to determine the sensitivity of H. hydralis primer pairs (HH-F-1/HH-R-1) in multiplex PCR (mixture of H. hydralis and P. xylostella primers). DNA of H. hydralis was serially diluted in a constant concentration of 20 ng of DNA from T. expolita and 1 ng P. xylostella, lane1, 100 bp molecular marker, lanes 2-16 , 0.8, 3, zero, 0.8, empty well, 0.72, 0.36, 0.18, 0.09, 0.045, 0.022, 0.011, 0.005, 0.002, 0.001, 0.0007 ng of H. hydralis DNA respectively, lane 17, negative control (Predator‘s DNA).
Figure 5. Percentage detection of different prey species in Oechalia schellenbergii. Numbers beside bars indicate percentages.
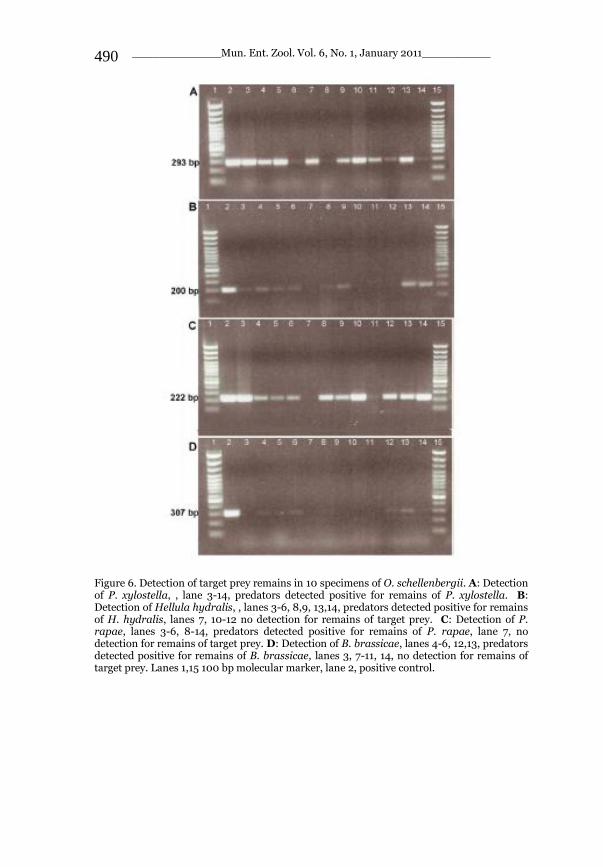
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 490
Figure 6. Detection of target prey remains in 10 specimens of O. schellenbergii. A: Detection of P. xylostella, , lane 3-14, predators detected positive for remains of P. xylostella. B: Detection of Hellula hydralis, , lanes 3-6, 8,9, 13,14, predators detected positive for remains of H. hydralis, lanes 7, 10-12 no detection for remains of target prey. C: Detection of P. rapae, lanes 3-6, 8-14, predators detected positive for remains of P. rapae, lane 7, no detection for remains of target prey. D: Detection of B. brassicae, lanes 4-6, 12,13, predators detected positive for remains of B. brassicae, lanes 3, 7-11, 14, no detection for remains of target prey. Lanes 1,15 100 bp molecular marker, lane 2, positive control.
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 491
Table 1. List of arthropods and Brassica plants used to test PCR primer specificity.
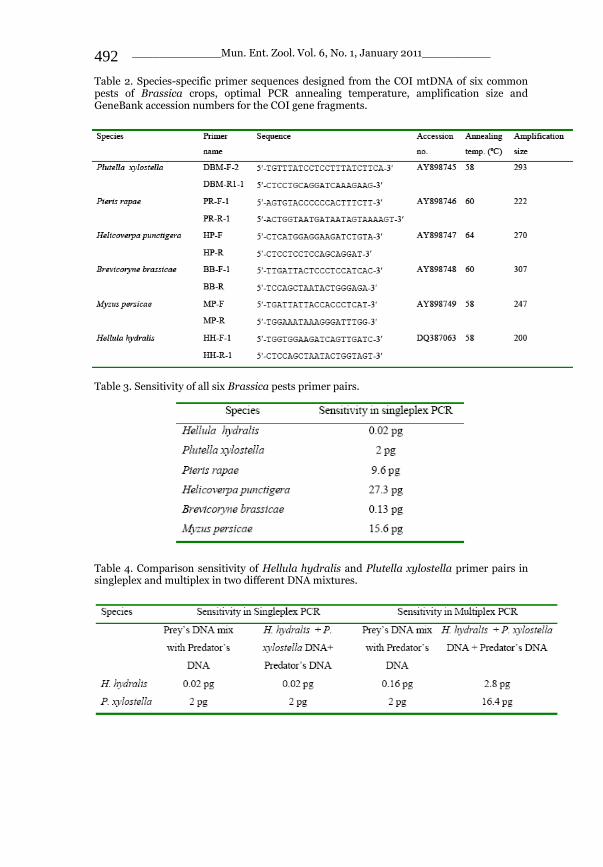
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 492
Table 2. Species-specific primer sequences designed from the COI mtDNA of six common pests of Brassica crops, optimal PCR annealing temperature, amplification size and GeneBank accession numbers for the COI gene fragments.
Table 3. Sensitivity of all six Brassica pests primer pairs.
Table 4. Comparison sensitivity of Hellula hydralis and Plutella xylostella primer pairs in singleplex and multiplex in two different DNA mixtures.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 493
NOTES ON THE NOCTUIDAE FAUNA (LEPIDOPTERA) COLLECTED BY BAIT TRAPS IN ORGANIC VINEYARD AND
ORCHARDS OF KEMALPAŞA (IZMIR) PROVINCE OF TURKEY
Rukiye Tanyeri*, Ahu Üzüm*, Zuhal Okyar**, Serdar Tezcan*** and Nilay Gülperçin****
* Biology Department, Faculty of Science, Ege University, 35100 Bornova, Izmir, TURKEY. E-mail: [email protected] ** Biology Department, Faculty of Science, Trakya University, 22030, Edirne, TURKEY. E-mail: [email protected] *** Plant Protection Department, Faculty of Agriculture, Ege University, 35100 Bornova, Izmir, TURKEY. E-mail: [email protected] **** Natural History Application and Research Center, Ege University, 35100 Bornova, Izmir, TURKEY. E-mail: [email protected] [Tanyeri, R., Üzüm, A., Okyar, Z., Tezcan, S. & Gülperçin, N. 2011. Notes on the Noctuidae fauna (Lepidoptera) collected by bait traps in organic vineyard and orchards of KemalpaĢa (Izmir) province of Turkey. Munis Entomology & Zoology, 6 (1): 493-498] ABSTRACT: In this study, information is given on 40 species belonging to Noctuidae fauna (Lepidoptera) collected by bait traps in organic vineyard and plum, pear and peach orchards in KemalpaĢa (Izmir) province of Western Turkey. KEY WORDS: Organic agriculture, Noctuidae, Fauna, Bait trap.
To prevent the side effects of conventional agriculture to human health and environment, organic or ecological agriculture applications have been initiated all over the world. In the last two decades, its‘ importance improved in Turkey and the application of some ecological production methods has been studied in production areas of Western Turkey. This area has a special importance in production of agricultural crops namely grapes (Vitis vinifera Linnaeus, 1758), plums (Prunus domestica Linnaeus, 1753), pears (Pyrus communis Linnaeus, 1758) and peaches [Prunus persica (Linnaeus) Batsch, 1801]. Generally, there is not detailed information on agrobiodiversity of such areas in Turkey (Tanyeri et al., 2010). In order to supply an additional information in this field, material belonging to Noctuidae (Lepidoptera) fauna collected in organic vineyard (V. vinifera) and plum (P. domestica), pear (P. communis) and peach (P. persica) orchards in KemalpaĢa (Izmir) province of Western Turkey were evaluated in this paper.
MATERIAL AND METHODS
This study was conducted in organic vineyard (V. vinifera) and plum (P. domestica), pear (P. communis) and peach (P. persica) orchards in KemalpaĢa (Izmir) province of Western Turkey during the months of June-October in 2007. Noctuids were collected by bait traps. In each area a total of four bait traps containing 100 ml wine, 900 ml water, 25 gr sugar and 25 ml vinegar per litre were hung for monitoring the adults of noctuids (Lepidoptera: Noctuidae). The traps were emptied at two-week intervals from June to October in 2007. This paper deals with the noctuids which were captured as a by-catch. The material was collected determined by the third author, and is deposited in the Museum of Biology Department, Faculty of Arts and Sciences, Trakya University, Edirne, and the Prof. Dr. Niyazi Lodos Museum (LEMT) of Plant Protection Department,

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 494
Faculty of Agriculture, Ege University, Ġzmir, Turkey. As some of the material was damaged in the liquid of the bait trap, which resulted in the loss of some taxonomic characters, identification was based on genital characteristics. This was done using papers by Pierce, 1967, 1978; Fibiger, 1997; Hacker, 1989; Hacker et al., 2002; Ronkay et al., 2001; Goater et al., 2003; Fibiger et al., 2007.
RESULTS
Material evaluated in this study is listed below in alphabetical order, by subfamilies and by genera within each subfamily.
Acontiinae
Acontia lucida (Hufnagel, 1766) Material examined: Prunus persicae: 14.09.2007 (6). Totally 6 specimens.
Acronictinae
Acronicta megacephala ([Denis & Schiffermüller], 1775) Material examined: Pyrus communis: 19.07.2007 (1); 14.08.2007 (1). Totally 2 specimens.
Acronicta rumicis (Linnaeus, 1758) Material examined: Vitis vinifera: 14.08.2007 (1). P. communis: 30.07.2007 (3); 14.08.2007 (1); 14.09.2007 (2); 05.10.2007 (1). P. persicae: 14.09.2007 (1); 05.10.2007 (3). Totally 12 specimens.
Acronicta tridens ([Denis & Schiffermüller], 1775) Material examined: P. communis: 14.08.2007 (1); 05.10.2007 (1). Totally 2 specimens.
Amphipyrinae
Actinotia hyperici ([Denis & Schiffermüller], 1775) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 30.07.2007 (3); 14.09.2007 (1). Totally 5 specimens.
Hoplodrina ambigua ([Denis & Schiffermüller], 1775) Material examined: P. persicae: 05.10.2007 (1). Totally 1 specimen.
Catocalinae
Anua lunaris ([Denis & Schiffermüller], 1775) Material examined: P. communis: 14.08.2007 (1). Totally 1 specimen.
Catocala conjuncta (Esper, 1787) Material examined: P. communis: 14.08.2007 (1). Prunus domestica: 14.08.2007 (1). Totally 2 specimens.
Catocala conversa (Esper, 1787) Material examined: P. communis: 29.06.2007 (3); 19.07.2007 (1); 30.07.2007 (2). P. persicae: 29.06.2007 (1); 19.07.2007 (2). Totally 9 specimens.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 495
Dysogonia algira (Linnaeus, 1767) Material examined: P. communis: 19.07.2007 (1); 30.07.2007 (1); 14.08.2007 (6); 14.09.2007 (2); 05.10.2007 (3). P. domestica: 30.07.2007 (1). P. persicae: 30.07.2007 (2); 14.09.2007 (3); 05.10.2007 (2). Totally 21 specimens.
Ophiusa tirhaca (Cramer, 1777) Material examined: P. communis: 14.08.2007 (2). Totally 2 specimens.
Prodotis stolida (Fabricius, 1775) Material examined: V. vinifera: 30.07.2007 (1); 14.08.2007 (1); 14.09.2007 (8); 05.10.2007 (20). P. communis: 29.06.2007 (1); 19.07.2007 (1); 30.07.2007 (9); 14.08.2007 (6); 31.08.2007 (1); 14.09.2007 (45); 05.10.2007 (42). P. domestica: 19.07.2007 (1); 30.07.2007 (1); 14.09.2007 (7); 05.10.2007 (16). P. persicae: 29.06.2007 (1); 30.07.2007 (2); 14.09.2007 (13); 05.10.2007 (16). Totally 192 specimens.
Hadeninae
Anapoma riparia (Rambur, 1829) Material examined: P. communis: 05.10.2007 (1). Totally 1 specimen.
Hadula trifolii (Hufnagel, 1766) Material examined: V. vinifera: 19.07.2007 (2); 05.10.2007 (98). P. communis: 30.07.2007 (2); 14.08.2007 (4); 14.09.2007 (29); 05.10.2007 (44). P. domestica: 30.07.2007 (1); 05.10.2007 (9). P. persicae: 19.07.2007 (1); 30.07.2007 (1); 14.09.2007 (8); 05.10.2007 (35). Totally 234 specimens.
Hecatera bicolorata (Hufnagel, 1766) Material examined: P. persicae: 19.07.2007 (1). Totally 1 specimen.
Hecatera dysodea ([Denis & Schiffermüller], 1775) Material examined: P. communis: 14.08.2007 (5); 05.10.2007 (1). P. persicae: 30.07.2007 (1); 14.09.2007 (1); 05.10.2007 (7). Totally 15 specimens.
Laconobia oleracea (Linnaeus, 1758) Material examined: P. communis: 14.08.2007 (1). P. persicae: 05.10.2007 (1). Totally 2 specimens.
Leucania loreyi (Duponchel, 1827) Material examined: P. communis: 05.10.2007 (1). P. persicae: 05.10.2007 (1). Totally 2 specimens.
Leucania putrescens (Hübner, [1824]) Material examined: V. vinifera: 05.10.2007 (4). P. communis: 14.09.2007 (1). Totally 5 specimens.
Mythimna congrua (Hübner, [1817]) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 30.07.2007 (1); 14.09.2007 (1); 05.10.2007 (3). P. persicae: 05.10.2007 (1). Totally 7 specimens.
Mythimna languida (Walker, 1858) Material examined: V. vinifera: 05.10.2007 (3). P. communis: 14.08.2007 (1); 14.09.2007 (2); 05.10.2007 (3). P. domestica: 30.07.2007 (1); 14.09.2007 (3); 05.10.2007 (1). P. persicae: 30.07.2007 (1); 14.09.2007 (1); 05.10.2007 (15). Totally 31 specimens.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 496
Mythimna sicula (Treitschke, 1835) Material examined: V. vinifera: 30.07.2007 (1); 05.10.2007 (1). P. communis: 14.08.2007 (1); 14.09.2007 (5); 05.10.2007 (2). P. persicae: 05.10.2007 (3). Totally 13 specimens.
Mythimna unipuncta (Haworh, 1809) Material examined: V. vinifera: 30.07.2007 (2); 05.10.2007 (11). P. communis: 19.07.2007 (6); 30.07.2007 (8); 14.08.2007 (22); 14.09.2007 (11); 05.10.2007 (95). P. domestica: 29.06.2007 (2); 30.07.2007 (10); 14.08.2007 (13); 14.09.2007 (7); 05.10.2007 (56). P. persicae: 19.07.2007 (2); 30.07.2007 (26); 14.08.2007 (6); 14.09.2007 (12); 05.10.2007 (232). Totally 521 specimens.
Mythimna vitellina (Hübner, 1808) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 14.08.2007 (2). P. persicae: 19.07.2007 (1). Totally 4 specimens.
Sideritis reticulata (Goeze, 1781) Material examined: P. persicae: 05.10.2007 (2). Totally 2 specimens.
Heliothinae
Helicoverpa armigera (Hübner, [1808]) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 30.07.2007 (1); 14.09.2007 (2). P. domestica: 05.10.2007 (2). P. persicae: 14.09.2007 (2). Totally 8 specimens.
Noctuinae
Agrotis atrux (Pinker, 1971) Material examined: P. communis: 14.08.2007 (6); 05.10.2007 (19). P. domestica: 05.10.2007 (12). Totally 37 specimens.
Agrotis puta (Hübner, 1803) Material examined: V. vinifera: 29.06.2007 (1); 19.07.2007 (1); 30.07.2007 (6); 05.10.2007 (214). P. domestica: 14.09.2007 (2); 05.10.2007 (87). P. persicae: 29.06.2007 (5); 19.07.2007 (6); 30.07.2007 (1); 14.09.2007 (12); 05.10.2007 (133). P. communis: 30.07.2007 (6); 14.08.2007 (10); 14.09.2007 (29); 05.10.2007 (251). Totally 764 specimens.
Agrotis segetum ([Denis & Schiffermüller], 1775) Material examined: V. vinifera: 29.06.2007 (1); 19.07.2007 (7); 30.07.2007 (4); 05.10.2007 (4). P. domestica: 19.06.2007 (12); 29.06.2007 (14); 19.07.2007 (34); 30.07.2007 (21); 14.08.2007 (1); 14.09.2007 (2); 05.10.2007 (10). P. persicae: 29.06.2007 (46); 19.07.2007 (49); 30.07.2007 (8); 14.08.2007 (1); 14.09.2007 (11); 05.10.2007 (6). P. communis: 29.06.2007 (62); 19.07.2007 (59); 30.07.2007 (15); 14.08.2007 (15); 14.09.2007 (10); 05.10.2007 (14). Totally 406 specimens.
Euxoa tritici (Linnaeus, 1761) Material examined: V. vinifera: 05.10.2007 (2). P. persicae: 05.10.2007 (2). Totally 4 specimens.
Noctua comes Hübner, 1813 Material examined: P. communis: 19.07.2007 (5). P. domestica: 19.06.2007 (1); 19.07.2007 (3). P. persicae: 29.06.2007 (1); 19.07.2007 (1). Totally 11 specimens.
Noctua fimbriata (Schreber, 1759) Material examined: P. communis: 29.06.2007 (1). Totally 1 specimen.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 497
Noctua janthina ([Denis & Schiffermüller], 1775) Material examined: P. communis: 30.07.2007 (1); 14.08.2007 (1). Totally 2 specimens.
Noctua orbona (Hufnagel, 1766) Material examined: V. vinifera: 19.07.2007 (1). P. communis: 29.06.2007 (14); 14.08.2007 (1). P. domestica: 30.07.2007 (3); 14.08.2007 (1). P. persicae: 05.10.2007 (2). Totally 22 specimens.
Noctua pronuba (Linnaeus, 1758) Material examined: V. vinifera: 19.07.2007 (2). P. persicae: 19.07.2007 (1). Totally 3 specimens.
Peridroma saucia (Hübner, [1803-1808]) Material examined: V. vinifera: 30.07.2007 (1); 05.10.2007 (1). P. communis: 29.06.2007 (3); 30.07.2007 (4); 14.09.2007 (1). P. domestica: 29.06.2007 (1); 30.07.2007 (4). P. persicae: 30.07.2007 (7). Totally 22 specimens.
Xestia xanthographa ([Denis & Schiffermüller], 1775) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 05.10.2007 (2). P. domestica: 05.10.2007 (2). Totally 5 specimens.
Ophiderinae
Aedia funesta (Esper, [1786]) Material examined: V. vinifera: 14.08.2007 (1). P. communis: 19.07.2007 (1); 30.07.2007 (1); 14.08.2007 (1); 14.09.2007 (2); 05.10.2007 (5). P. domestica: 19.07.2007 (1); 30.07.2007 (1). P. persicae: 29.06.2007 (1); 19.07.2007 (1); 30.07.2007 (3); 14.08.2007 (2); 14.09.2007 (4); 05.10.2007 (6). Totally 30 specimens.
Catephia alchymista ([Denis & Schiffermüller], 1775) Material examined: P. communis: 30.07.2007 (7). Totally 7 specimens.
Tyta luctuosa ([Denis & Schiffermüller], 1775) Material examined: V. vinifera: 05.10.2007 (1). P. communis: 30.07.2007 (1); 14.09.2007 (1); 05.10.2007 (2). P. persicae: 14.09.2007 (1); 05.10.2007 (2). Totally 8 specimens.
DISCUSSION
The dominant species in the collected materials was Agrotis puta (764 caught individuals) and the relative abundance of this species is 31.53%. Mythimna unipuncta (521), Agrotis segetum (406), Hadula trifolii (234), and Prodotis stolida (192) followed by the relative abundances of 21.50; 16.76; 9.66; 7.92 percent, respectively.
Among those Acronicta megacephala, Acronicta rumicis, Acronicta tridens, Actinotia hyperici, Hoplodrina ambigua, Catocala conjuncta, Catocala conversa, Dysogonia algira, Ophiusa tirhaca, Prodotis stolida, Hecatera dysodea, Laconobia oleracea, Leucania putrescens, Mythimna vitellina, Agrotis segetum, Noctua comes, Noctua fimbriata, Noctua orbona, Noctua pronuba, Peridroma saucia, Xestia xanthographa, Aedia funesta, Catephia alchymista and Tyta luctuosa were reported from organic cherry orchards by Okyar and Tezcan (2001). In the light of previous works (Hacker, 1989) Acronicta rumicis and A.tridens are known as pests of rosaceous plants.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 498
In organic vineyard the number of recorded species was 20, in plum orchard 15, in peach orchard 27, in pear orchard 34. A total of 405 specimens (16.71%) were recorded in organic vineyard as well as 948 specimens (39.13%) from pear orchard, 726 specimens (29.96%) from peach orchard and 344 specimens (14.20%) from plum orchard.
In the following studies, it is hoped that further studies on the feeding habits of species and their roles in organic vineyards and orchards will be realised.
LITERATURE CITED Fibiger, M. 1997. Noctuidae Europaeae, Genitalia And Supplement To Noctuinae III, Vol. 3:. 418 pp., Entomological Press. Fibiger, M. & Hacker, H. 2007. Noctuidae Europaeae Amphipyrinae, Condicinae, Eriopinae, Xyleninae, Vol. 9, 410 pp., Entomological Press. Goater, B., Ronkay, L.& Fibiger, M. 2003. Noctuidae Europaeae, Catocalinae & Plusiinae. Vol. 10: 452 pp, Entomological Press. Hacker, H. 1989. Die Noctuidae Griechenlands. Mit einer Übersicht über die fauna des Balkanraumes (Lepidoptera, Noctuidae). Herbipoliana, Marktleuthen, Deutschland, 2:1-589 pp. Hacker, H., Ronkay, L. & Hreblay, M. 2002. Noctuidae Europaeae, Hadeninae I. Vol. 4, 419 pp., Entomological Press. Okyar, Z. & Tezcan, S. 2001. On the noctuid fauna (Lepidoptera: Noctuidae) of ecological cherry orchards in western Turkey. Zoology in the Middle East, 22: 95-102. Pierce, F. N. 1967. The Genitalia of The Group Noctuidae of Lepidoptera of the British Island. Hon. Librarion of the Lancanshire and Cheshire Entemological Society; Vice-President of the Liverpool Microscopical Society, 88 pp. Pierce, F. N. 1978. The Genitalia of The Group Noctuidae of the Lepidoptera of the British Islands. An accaont of morphology of the female reproductory organs. Faringdon, Oxon, 62 pp. Ronkay, L., Yela, J. L. & Hreblay, M. 2001. Noctuidae Europaeae, Hadeninae II. Vol. 5: 454 pp., Entomological Press. Tanyeri, R., Üzüm, A., Tezcan, S., Keskin, B.& Gülperçin, N. 2010. Notes on pitfall trap collected Tenebrionidae (Coleoptera) species in organic vineyard and orchards of KemalpaĢa (Izmir) province of western Turkey. Munis Entomology & Zoology, 5, Supplement, 917-919.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 499
DESCRIPTIONS OF THREE NEW SPECIES OF PETALOCEPHALA STÅL, 1853 FROM CHINA
(HEMIPTERA: CICADELLIDAE: LEDRINAE)
Yu-Jian Li* and Zi-Zhong Li**
* Institute of Entomology, Guizhou University, Guiyang, Guizhou Province, 550025, CHINA. E-mail: [email protected] ** Guizhou Key Laboratory for Plant Pest Management of Mountainous Region, Guizhou University, Guiyang, Guizhou Province, 550025, China. E-mail: [email protected] [Li, Y-J. & Li, Z-Z. 2011. Descriptions of three new species of Petalocephala Stål, 1853 from China (Hemiptera: Cicadellidae: Ledrinae). Munis Entomology & Zoology, 6 (1): 499-503] ABSTRACT: The paper deals with three new species of the genus Petalocephala Stål, 1853 (Hemiptera: Cicadellidae: Ledrinae), Petalocephala kuankuoensis sp. nov., Petalocephala dicondylica sp. nov. and Petalocephala gongshanensis sp. nov., from Guizhou and Yunnan, China are described. The type specimens of new species are deposited in the Institute of Entomology, Guizhou University (GUGC). KEY WORDS: Homoptera, Auchenorrhyncha, leafhopper, morphology, new species.
The subfamily Ledrinae is a fairly large group of the 50 major subfamilies of leafhoppers (Dietrich, 2005). Ledrinae represent a relatively small subfamily within the very large and diverse leafhopper family Cicadellidae with over 450 species in 73 genera worldwide (Oman et al., 1990). The subfamily distributed worldwide, but primarily in Australia, Africa, and the South East Asia region, usually on trees and shrubs. This newly revised classification strengthens hypotheses that the Ledrinae is a very old group within Cicadellidae. As recently report, the subfamily includes 5 tribes with a total of 38 genera and approximately 300 species (Jones et al., 2009). Petalocephala Stål is the largest genus (87 described species) in Ledrinae (Jones et al., 2009). These taxa, Petalocephala kuankuoensis sp. nov., Petalocephala dicondylica sp. nov. and Petalocephala gongshanensis sp. nov., are described and illustrated in this paper.
MATERIALS AND METHODS The genital segments of the examined specimens were macerated in 10% NaOH and drawn from preparations in glycerin using a light microscope. Figures of the specimens were made using Leica MZ12.5. The type specimens are deposited in the Institut of Entomology of Guizhou University, Guiyang, China (GUGC).
Petalocephala kuankuoensis sp. nov. Figs 1, 4, 7-11
Description. Body large. Length (including forewing): ♂, 12.5 mm. Body virescent, head and face usually stained red along margin (Figs 1, 4). Eyes dark brown; ocelli translucent (Fig. 1). Forewing semitransparent; an infuscate spot on disc. Pygofer side in lateral aspect tapering posteriorly, end curved dorsally, with a process extending like a oblate hook from middle ventral margin inside (Figs 9, 10). Style long, anterior portion shorter than caudal portion, caudal portion

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 500
obviously robust and tapering posteriorly, apex strongly recurved like a head of duck, with some macrosetae near the recurved area (Figs 7, 8). Connective T-shaped with high dorsomedial keel (Figs 7, 8). Aedeagus with a pair of lateral processes, the processes elongate at both ends, anterior apex flat straight and posterior apex contorted like a hook; shaft tapering unconspicuous, slight curved dorsally in lateral view (Figs 7, 8); gonopore apical. Other characteristics are as shown in Figs 1, 4, 7-11. Type Material. Holotype ♂, CHINA: Guizhou, Suiyang, Kuankuoshui, 6-9 June 2010, coll. Zhang Bin. Host plant. Unknown. Etymology. The species name is derived from the locality of the types. Remarks. This new species is superficially similar to Petalocephala sanguineomarginata Kuoh, but can be distinguished from the latter by: (1) The shape of style is different; (2) pygofer side process extending from middle ventral margin not caudal; (3) The shape of lateral process of aedeagal shaft is different.
Petalocephala dicondylica sp. nov. Figs 2, 5, 12-16
Description. Body large. Length (including forewing): ♂, 11.5 mm. Body virescent, head and face usually stained orange red along margin (Figs 2, 5). Apex of vertex angular forward (Figs 2, 5). Eyes dark brown; ocelli translucent (Fig. 2). Forewing semitransparent, end margin stained yellow; an infuscate spot on disc. Pygofer side in lateral aspect triangular, end curved dorsally, with a process extending like a gracile hook from middle ventral margin inside (Figs 14, 15). Style long, anterior portion shorter than caudal portion, caudal portion obviously robust, apex strongly recurved, with some macrosetae near the recurved area (Figs 12, 13). Connective T-shaped with dorsomedial keel (Figs 12, 13). Aedeagus with a pair of lateral processes, the processes elongate at both ends, thick in the center and tapered at each end; shaft tapering posteriorly and slight curved dorsally in lateral view (Fig. 13); gonopore apical. Other characteristics are as shown in Figs 2, 5, 12-16. Type Material. Holotype ♂, CHINA: Suiyang, Kuankuoshui, 4 June 2010, coll. Zheng Yanli. Host plant. Unknown. Etymology. This species is named from its aedeagus with a vimineous process on each side, the processes elongate at both ends, thick in the center and tapered at each end. Remarks. This new species is superficially similar to Petalocephala kuankuoensis sp. nov., but can be distinguished from the latter by: (1) apex of vertex angular forward; (2) pygofer side inside process very gracile; (3) lateral process of aedeagal shaft tapered at each end, uncontorted.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 501
Petalocephala gongshanensis sp. nov. Figs 3, 6, 17-21
Description. Body large. Length (including forewing): ♂, 13.0-13.1 mm. Body virescent, head and face usually stained brown red along margin (Figs 3, 6). Eyes dark brown; ocelli translucent (Fig. 3). Forewing semitransparent, end margin stained yellow, veins green; an infuscate spot on disc. Pygofer side in lateral aspect trapezoidal, end curved dorsally, with a process extending like a hook from caudal ventral margin inside (Figs 19, 20). Style long, anterior portion shorter than caudal portion, caudal portion obviously robust and tapering posteriorly, apex recurved, with some macrosetae near the recurved area (Figs 17, 18). Connective T-shaped with high dorsomedial keel (Figs 17, 18). Aedeagus with a pair of lateral processes, the processes elongate at both ends, anterior apex straight tapered and posterior apex contorted; shaft burly, tapering posteriorly and slight curved dorsally in lateral view (Fig. 18); gonopore apical. Other characteristics are as shown in Figs 3, 6, 17-21. Type Material. Holotype ♂, CHINA: Yunnan, Gongshan, 7 June 2009, coll. Zhu Daxian. Paratype: 1♂, same data as holotype. Host plant. Unknown. Etymology. The species name is derived from the locality of the types. Remarks. This new species is superficially similar to Petalocephala kuankuoensis sp. nov., but can be distinguished from the latter by: (1) length of vertex longer than half head width (including eyes), but P. kuankuoensis sp. nov. about same as it; (2) pygofer side process extending from caudal ventral margin not middle; (3) lateral process of aedeagal shaft anterior apex straight tapered and posterior apex contorted not like a hook.
ACKNOWLEDGEMENTS We thanks Zhang Bin, College of Life Sciences and Technology, Inner Mongolia Normal University, China, and Zheng Yanli, Institute of Entomology, Guizhou University, China, gave some helps for this study.
LITERATURE CITED Cai, P. & Ge, Z. L. 1992. Three new species of the genus Petalocephala from China (Homoptera: Cicadellidae; Ledrinae). Zoological Research, 13 (2): 117-121. Cen, Y. W. & Cai, P. 2000. Two new species of genus Petalocephala Stål (Homoptera: Cicadellidae; Ledrinae) from China. Entomotaxonomia, 22 (4): 247-250. Dietrich, C. H. 2005. Keys to the families of Cicadomorpha and Subfamilies and Tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Florida Entomologist, 88: 10-15. Kato, M. 1931. Japanese Ledridae. Dobutsugaku Zasshi, 43, 431-440. Jones, J. R. & Deitz L. L. 2009. Phylogeny and systematics of the leafhopper subfamily Ledrinae (Hemiptera: Cicadellidae). Zootaxa, 2186: 1–120. Kuoh, C. L. 1984. Six new species of genus Petalocephala (Homoptera: Ledrinae). Entomotaxonomia, 6 (4): 271-278.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 502
Oman, P. W., Knight W. J. & Nielson M. W. 1990. Leafhoppers (Cicadellidae): a bibliography, generic Check-list and index to the world literature 1956-1985. CAB International Institute of Entomology, Wallingford, U.K. 1-368. Stål, C. 1853. Nya genera bland Hemiptera. Öfversigt af Kongliga Svenska Vetenskaps–Akadamiens Förhandlingar, 10: 231–255.
Figures 1-8. 1. Petalocephala kuankuoensis sp. nov., Head and thorax, dorsal view; 2. Petalocephala dicondylica sp. nov., Head and thorax, dorsal view; 3. Petalocephala gongshanensis sp. nov., Head and thorax, dorsal view; 4. Petalocephala kuankuoensis sp. nov., Face; 5. Petalocephala dicondylica sp. nov., Face; 6. Petalocephala gongshanensis sp. nov., Face.
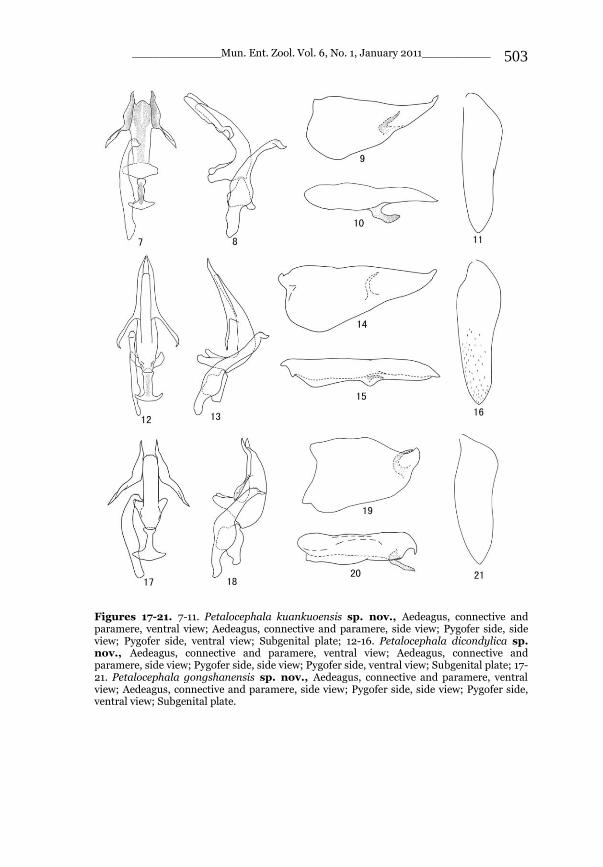
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 503
Figures 17-21. 7-11. Petalocephala kuankuoensis sp. nov., Aedeagus, connective and paramere, ventral view; Aedeagus, connective and paramere, side view; Pygofer side, side view; Pygofer side, ventral view; Subgenital plate; 12-16. Petalocephala dicondylica sp. nov., Aedeagus, connective and paramere, ventral view; Aedeagus, connective and paramere, side view; Pygofer side, side view; Pygofer side, ventral view; Subgenital plate; 17-21. Petalocephala gongshanensis sp. nov., Aedeagus, connective and paramere, ventral view; Aedeagus, connective and paramere, side view; Pygofer side, side view; Pygofer side, ventral view; Subgenital plate.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 504
TWO RARE SPECIES FOR TURKEY OF THE GENUS VADONIA MULSANT, 1863
WITH THEIR PHOTOS OF HABITUS AND MALE GENITALIAE (COLEOPTERA: CERAMBYCIDAE: LEPTURINAE)
Hüseyin Özdikmen*, Eşref Yüksel*
and Atılay Yağmur Okutaner* * Gazi Üniversitesi, Fen-Edebiyat Fakültesi, Biyoloji Bölümü, 06500 Ankara / Türkiye. Email: [email protected] [Özdikmen, H., Yüksel, E. & Okutaner, A. Y. 2011. Two rare species for Turkey of the genus Vadonia Mulsant, 1863 with their photos of habitus and male genitaliae (Coleoptera: Cerambycidae: Lepturinae). Munis Entomology & Zoology, 6 (1): 504-507] ABSTRACT: In the genus Vadonia Mulsant, 1863, two rare species for Turkey are evaluated. New data of these species are also given in the text. KEY WORDS: Vadonia, Lepturinae, Lepturini, Cerambycidae, Turkey.
As commonly accepted that chiefly Palaearctic genus Vadonia Mulsant, 1863 (except the orientalic species V. eckweileri Holzschuh, 1989 from Pakistan) is represented by twenty-three species (with sixteen subspecies) in the whole world. Fourteen species are endemic to different countries. In Turkey, it is represented by fifteen species. Seven species of them are endemic to Turkey as V. bolognai Sama, 1982; V. ciliciensis Daniel & Daniel, 1891; V. danielorum Holzschuh, 1984; V. frater Holzschuh, 1981; V. instigmata Pic, 1889; V. ispirensis Holzschuh, 1993 and V. soror Holzschuh, 1981 (Özdikmen & Turgut, 2009).
The evaluated species in the present paper, Vadonia bicolor (Redtenbacher, 1850) and Vadonia moesiaca (Daniel & Daniel, 1891) are rare species for Turkey. Genus VADONIA Mulsant, 1863
Syn. Neovadonia Kaszab, 1938 Type species: Leptura unipunctata Fabricius, 1787
Vadonia bicolor (Redtenbacher, 1850)
(Fig. 1) This species was described by Redtenbacher (1850) from Fars prov.: Schiraz in SW Iran with the original combination Leptura bicolor Redtenbacher, 1850. Leptura türki that was described by Heyden (1878) from Gorgan prov.: Astrabad in N Iran, is a synonym of V. bicolor. It probably is distributed only in N and E Anatolia for Turkey. The present record is the most west point for the distribution areas of the species. So the species is recorded for the first time to Ankara province in Turkey. Photos of the male genitalia are presented in figure 2. Material examined: Ankara prov.: Kızılcahamam, IĢık Mountain, 26.V.2010, 1 specimen. The material is deposited in Gazi University, Ankara, Turkey. Records in Turkey: Erzurum prov. as 2 females (Daniel & Daniel, 1891); Erzurum prov., Van prov. (Plavilstshikov, 1936); Turkey (Lobanov et al., 1981; Sama & Löbl in Löbl & Smetana, 2010); N Turkey (Danilevsky & Miroshnikov, 1985) (Fig. 3). Range: Caucasus (Azerbaijan), Turkey, Iran.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 505
Chorotype: SW-Asiatic (Anatolo-Caucasian + Irano-Caucasian + Irano-Anatolian).
Vadonia moesiaca (Daniel & Daniel, 1891) (Fig. 4)
This species was described by Daniel & Daniel (1891) from Serbia, Turkey and Balkan with the original combination Leptura moesiaca Daniel & Daniel, 1891. V. bipunctata Kraatz, 1859 and V. beali Pic, 1926 are synonyms of V. moesiaca. It probably is distributed only in Western half of Turkey. It has been known only from Antalya province in SW Turkey as the exact locality. So, the species is recorded for the first time to Ankara province in Turkey. Photos of the male genitalia are presented in figure 5. Material examined: Ankara prov.: Beypazarı, Ġnözü Valley, 02.VI.2009, 1 specimen; Kızılcahamam, IĢık Mountain, 26.V.2010, 1 specimen. Records in Turkey: Turkey (Daniel & Daniel, 1891; Winkler, 1924-1932; Lodos, 1998); Antalya: TaĢağıl (Adlbauer, 1988); European and Asian Turkey (Sama & Löbl in Löbl & Smetana, 2010) (Fig. 6). Range: Europe (Serbia & Montenegro, Macedonia, Albania, Bulgaria, Greece, European Turkey), Asian Turkey. Chorotype: Turano-Mediterranean (Balkano-Anatolian).
LITERATURE CITED Adlbauer, K. 1988. Neues zur Taxonomie und Faunistik der Bockkäferfauna der Türkei (Coleoptera, Cerambycidae). Entomofauna, 9 (12): 257-297. Daniel, K. & Daniel, J. 1891. Revision der mit Leptura unipunctata F. und fulva Deg. verwandten Arten. Coleopteren-Studien, 1: 1-40. Danilevsky, M. L. & Miroshnikov A. I. 1985. Timber-Beetles of Caucasus (Coleoptera, Cerambycidae). The Key. Krasnodar, 419 pp. Heyden, L. F. J. D. 1878. [new taxa]. In: Schneider, O. & Leder, H.: Beiträge zur Kenntniss der kaukasischen Käferfauna. Brünn: W. Burkart, 358 pp. [note: separate issue from Verhandlungen des Naturforschenden Vereins in Brünn, 16 [1877]: 3-258) and 17 [1878]: 3-104]. Lobanov, A. L., Danilevsky, M. L. & Murzin, S. V. 1981. [Systematic list of Longicorn beetles (Coleoptera, Cerambycidae) of the USSR. 1]. Revue d‘Entomologie, 60(4): 784-803 (in Russian). Lodos, N. 1998. Entomology of Turkey VI (General, Aplied and Faunistic). Ege Ü. Ziraat Fak. Yayınları No: 529, E. Ü. Faculty of Agriculture Press, Ġzmir, 300 pp. Özdikmen, H. & Turgut, S. 2009. A review on the genera Pseudovadonia Lobanov et al., 1981 and Vadonia Mulsant, 1863 (Coleoptera: Cerambycidae: Lepturinae). Munis Entomology & Zoology, 4 (1): 29-52. Plavilstshikov, N. N. 1936. Cerambycidae (P.1). In: Faune de l‘URSS. Insects Coléptères. V.21. Moscou, Leningrad: 612 pp. Redtenbacher, L. 1850. Über den charakter der Insekten- Fauna von Südpersien. Denkschr. Acad. Wiss. Wien, 1: 42-53. Sama, G. & Löbl, I. 2010. Cerambycidae (others). In Löbl, I. & Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera, Volume 6, Chrysomeloidea. Apollo Books, Stenstrup. Winkler, A. 1924-1932. Catalogus Coleopterorum regionis palaearcticae. Verlag von Albert Winkler, 1135-1226.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 506
Figure 1. Habitus of Vadonia bicolor (Redtenbacher, 1850)
Figure 2. Male genitalia of Vadonia bicolor (Redtenbacher, 1850).
Figure 3. Known distribution of Vadonia bicolor in Turkey.
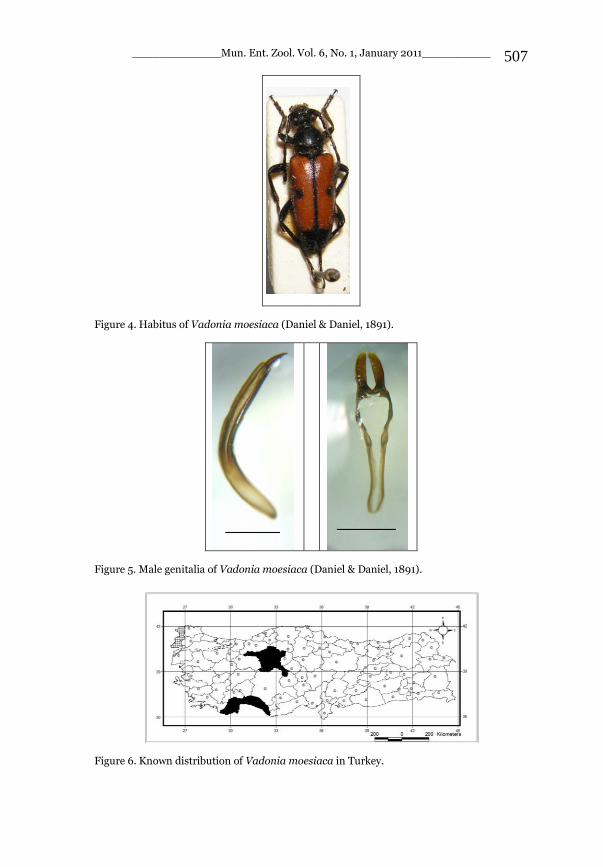
_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 507
Figure 4. Habitus of Vadonia moesiaca (Daniel & Daniel, 1891).
Figure 5. Male genitalia of Vadonia moesiaca (Daniel & Daniel, 1891).
Figure 6. Known distribution of Vadonia moesiaca in Turkey.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 508
A FIRST CASE OF GYNANDROMORPHISM IN STENOPTERUS ATER (LINNAEUS, 1767) (COLEOPTERA, CERAMBYCIDAE)
Moreno Dutto *, Mattia Penna** and Gianfranco Sama ***
* Servizio Igiene e Sanità Pubblica, ASL CN1, via del Follone, 4, 12037 Saluzzo (CN), ITALY. E-mail: [email protected] ** Via Hope 14, 14015 San Damiano d'Asti (AT), ITALY. E-mail: [email protected] *** Via Raffaello Sanzio, 84, 47521 Cesena (FC), ITALY. E-mail: [email protected] [Dutto, M., Penna M. & Sama, G. 2011. A first case of gynandromorphism in Stenopterus ater (Linnaeus, 1767) (Coleoptera, Cerambycidae). Munis Entomology & Zoology, 6 (1): 508-511] ABSTRACT: We describe here the first case of gynandromorphism in Stenopterus ater (Linnaeus, 1767). The case refers to one specimen bearing typically male and female features on its right and left sides respectively, therefore representing a case of complete lateral gynandromorphism. KEY WORDS: Coleoptera, Cerambycidae, Stenopterus ater, chimeric, gynandromorph, Italy.
Stenopterus Illiger, 1804 is a western Palaearctic genus (mostly circum-Mediterranean), including nine species distributed from Turkmenia and northern Iran to Portugal and from Central Europe (chiefly xerothermic localities) to Jordan and south-western Morocco. Three species are known to occur in Italy: S. flavicornis Küster, 1846, S. rufus rufus (Linnaeus, 1767) and S. ater (Linnaeus, 1767); the latter may be easily recognized by the first segment of antennae longitudinally furrowed on the outer side, antennae and legs partly or entirely black and the different shape of male genitalia (Sama, 1995). S. ater is a Western-Mediterranean species, widespread and chiefly common from North Africa to southern France and Italy; local in central Europe (Slovakia) and Balkans, southward to the southern Greece (Peloponnese); unknown in Bulgaria, its occurrence in Hungary, Romania and Crimea needs confirmation (Sama, 2002). In Italy it is recorded from all regions except Trentino-Alto Adige (Sama, 1988; Pesarini & Sabbadini, 1994; Hellrigl, 2010). Its larval development occurs in dead dry wood of branches of nearly all broadleaf trees and bushes. In Italy it is recorded on Carpinus, Ostrya, Pistacia, Ceratonia, Robinia, Ficus, Ulmus and others (Sama, 1988, 2002). Adults feed on various flowers, frequently on Umbellifers, from May to September. Morphologically they reach 6-14 mm in length, with pronounced sexual dichroism: elytra are tipically yellow-brown in male and completely black in female. Please refer to Sama (1985) for the revision of the genus.
We present here the discovery of a gynandromorphous specimen of Stenopterus ater Linnaeus, 1767), taking into consideration that within the family Cerambycidae only three cases of gynandromorphism have ever been documented (Narita et al., 2010).
MATERIALS AND METHODS
One specimen of the longhorn beetle Stenopterus ater (Cerambycidae) with evident gynandromorphous features was collected by one of the authors (M.P.) in San Damiano d'Asti (44°50'06,54''N, 8°04'02,66''E) (Piedmont, Italy) on 4 April 1996 on the inflorescence of an umbellifer. The previous identification, carried

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 509
out with a stereomicroscope (Motic SMZ-168) using Pesarini‘s identification keys (Pesarini & Sabbadini, 1994) was definitively confirmed by the third author. The specimen has partly been damaged during collection and dissection: the left antenna has the last seven joints glued, the two segments missing, the median and the posterior legs, broken during dissection, have been glued.
DISCUSSION
The specimen, which is 8 mm in length and has female genitalia, is characterized by complete lateral gynandromorphism (Fig. 1-3), bearing typically male and female features on its right and left sides respectively. The right part of body shows longer antenna, elytron yellow-ochraceous and shorter than the last abdominal tergite, legs ochraceous except the clubs of all femora and apices of tibiae and tarsi. The left side of body has shorter antenna (the left antenna is two last segments missing), elytron uniformly black and about as long as the last abdominal tergite, femora and tibiae completely black.
Gynandromorphism is an infrequently-occurring abnormality in which phenotypically male and female parts are present in a single individual. It occurs during the embryonic phase of development and can be expressed either as bilateral asymmetry or mosaic areas. There are several causes of individual gynandromorphism: loss or damage to sexual chromosomes, binucleated egg, and infections due to Wolbachia (a frequent endosymbiont in insects) (Narita et al., 2010).
Cases of gynandromorphism are found in various insect groups and are particularly evident in species with pronounced sexual dimorphism and dichroism (Narita et al. 2010). Within the family Cerambycidae examples of gynandromorphism have been reported in Ergates faber (Linnaeus, 1760), Stictoleptura rubra (Linnaeus, 1758) and Rhagium (Megarhagium) mordax (De Geer, 1775) which were described by Balazuc (1952), Weber (1913) and Starzyk (1984) respectively.
Considering that our specimen is laterally gynandromorphous, it is plausible that this abnormality might be due to abnormal embryonic development through the retention of the polar body inside the egg, resulting in two nuclei that were subsequently both fertilized.
LITERATURE CITED Balazuc, J. 1952. Un Ergates faber L. gynandromorphe (Col. Cerambycidae). Bulletin de la Société entomologique de France, 3: 34-38. Hellrigl, K. 2010. Faunistik der Bockkäfer von Südtirol (Coleoptera: Cerambycidae). Forest Observer, 5: 31-152. Narita, S., Pereira, R. A. S., Kjellberg, F. & Kageyama, D. 2010. Gynandromorph and intersex: potential to understand the mechanism of sex determination in arthropods. Terrestrial Arthropods Reviews, 3: 63-96. Pesarini, C. & Sabbadini A. 1994. Insetti della fauna europea. Coleotteri Cerambicidi. Natura, 85 (1-2): 3-132. Sama, G. 1995. Studi sugli Stenopterini. Il genere Stenopterus Illiger, 1804. Bulletin de la Société entomologique de France, 4: 385-410. Sama, G. 1988. Coleoptera Cerambycidae. Fauna d'Italia vol. XXVI. Edizioni Calderini, Bologna, Italia, p. XXXVI + 216.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 510
Sama, G. 1995. Studi sugli Stenopterini. Il genere Stenopterus Illiger, 1804 - Bull. Soc. ent. France, 100 (4): 385-410. Sama, G. 2002. Atlas of the Cerambycidae of Europe and Mediterranean Area. 1: Northern, Western, Central and Eastern Europe. British Isles and Continental Europe from France (excl. Corsica) to Scandinavia and Urals. V. Kabourek, Zlin, p. 173. Starzyk, J. 1984. Rare case of bilateral gynandromorphism in Rhagium mordax (Deg.) (Col., Cerambycidae). Entomologist's Monthly Magazine: 120. Weber, L. 1913. Ein gynandromorphes exemplar von Leptura rubra L. Entomologische Blätter Berlin, 9: 12-15.
Figure 1. Gynandromorphous specimen of Stenopterus ater (Linnaeus, 1767), current status; photograph taken after dissection.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 511
Figure 2. Gynandromorphous specimen of Stenopterus ater (Linnaeus, 1767), photograph taken before dissection (dorsa) view) (photo M. Dutto).
Figure 3. Gynandromorphous specimen of Stenopterus ater (Linnaeus, 1767) , photograph taken before dissection (lateral view) (photo M. Dutto).

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 512
SCIENTIFIC NOTES
MAMMALIAN PESTS OF MUGA SILKWORM CROP
Himangshu Barman*
* Central Muga Eri Research and Training Institute, Central Silk Board, Lahdoigarh, Jorhat – 785700, Assam, INDIA. E-mail: [email protected] [Barman, H. 2011. Mammalian pests of Muga Silkworm Crop. Munis Entomology & Zoology, 6 (1): 512-515]
Muga Silkworm Antheraea assamensis Helfer is not only known for its unique golden coloured cloth fabrics, but also for its delicious test and nutrient values. The pupa of this semi domesticated silkworm variety has been using in different traditional delicious food item preparations by the indigenous tribes of north eastern region of India. These preparations are popular for their test and flavor among the peoples since time immemorial. In present development of science and technology also establishes its high nutritive and medicinal values for certain physiological disorders including diabetes. Silkworms are the most nutritious food with high source of proteins, calcium, magnesium, sodium, iron and vitamin B1, B2 and B3, besides having low content of fat. Silkworm contains 54% protein (%kcal), 43% fat (%kcal), 0.5% calcium (mg/kcal), 0.6% phosphorus (mg/kcal) which are comparatively higher than other insects. Thus, naturally the silkworm invites a comparatively large numbers of pests and predators that feed on them causing a considerable loss to the silkworm crop. To protect themselves silkworm have no such devices that can protect from the pests and predators. They cannot jump, escape or hide, and also slow moving. Since silkworm has no sharp jaws or legs, they cannot bite or harm to others. Their bodies are also soft facilitating easier to shallow and digest.
So naturally due to high nutritious value silkworms are the preferred feed for so many pests and predators of which some are mammalian predators. These mammalian predators in north east India conditions cause great havoc to the silkworm crop production. Sometimes depending upon localities they are solely responsible for crop failure.
Squirrels are very commonly occurring mammalian pest of a good number of agricultural crops including coconut, areca nut, maize, banana etc. Depending upon localities with unavailability‘s of their food and also human communities as their flesh is used as food by certain communities, their population is found from very rare to highly thick in North Eastern India. This small animal belongs to order Rodentia, family Sciuridae of vertebrate. Squirrels are quick and fast moving animal, expert in climbing trees generally live in trees, tree trunk holes. The most common to this region is three striped tree squirrel (Funambulus palmarum) which is diurinal. They are omnivorous in food habits, cannot digest cellulose compelling on foods rich in protein, carbohydrates and fats. Muga silkworms at 4th and 5th larval stages are very good nutritious supplement in their diet. In muga growing areas, they are found in considerable numbers facilitated by other plant canopy nearby the silkworm food tree plantation. Squirrel control in muga crop is still not achieved as any effective physical or mechanical or chemical measure still not coming into force.
Another important mammalian silkworm pest is wild rabbit (Sylvilagus brasiliensis) living in nearby forest vegetation. They belong to Leporidae family of

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 513
order Lagomorpha. These small wild rabbit although herbivorous in food habit they prefer to eat healthy muga silkworm larvae. They consume crawling larvae coming down and moving on soil due to different reasons. Since rabbit cannot climb the Som or Soalu plant they do not harm larvae that are in trees. During the time of cocoon formation when ripen larvae came down the trees (‗joom‘ assembling) in search of suitable place for cocooning, rabbit cause maximum loss even in night time if no measures are taken. This pest is found in almost all muga culturing localities of Assam.
Bats are very destructive mammalian silkworm pest widely occurring in most localities of Assam. In Assam generally two types of Bat are found of which one smaller size is known as insect eating placed in suborder Microchiroptera and, other is bigger size of fruit eating under Megachiroptera suborder. Under these two suborders species like Sphaerias blanford, Pteropus sp., Rousettus leschenvelti, and Cynopterus sphinx are found in Assam. The smaller sized bats habitat in house, unused houses, tree trunk holes, granaries, commonly dark undisturbed places where hiding scopes are available. The bigger sized bats generally live in group of undisturbed dark remote localities preferably in abandon houses, godowns, big trees, stone caves etc. But during night they come out for search of food and fly to long distances. In villages and rural areas, insect eating bats are invariably found flitting around cow shads and field. Bats are migratory installing a unit after they leave will insure they won‘t use it again. Throughout the world there are many species of bats. In fact, bat comprises the largest segment of mammals in the world. It is estimated that over 25% of the worlds mammals are bat. Although bat has strong teeth, they will not use them for chewing through material. In food habit bats are carnivorous. Since bats eat diverse group of insect including the insect pests they are known to be beneficial. Further, bats have the ability to keep mosquito population down. But in muga silkworm cropping, bats are of serious concern as pest. They chew the juice of the matured larvae with multiple dissections. To get ride of bat loss farmers use nylon net of bigger size over the food plantations that restrict the bats in reaching muga larvae. This mechanical control measure is of costly that every farmer cannot afford it. One electronic device of ultrasound can be used as bat repellant since ultrasound has been found to work well at repelling birds, rats, mice and bats. The sounds emitted are that of an animal in distress which in turn alarms the nesting or roosting colony. The theory is that the colony should than become wary of the nest sight and move out. Ultrasound has no penetrating capabilities and can only reach about 25 feet away from the transducers effectively. Once targets are more than 30 feet away they will acknowledge the distress sound but will not generally move away.
Foxes are common mammalian animal found in almost thin forest like vegetation throughout the state of Assam. Foxes are completely carnivorous lived on preying other smaller sized domestic as well as wild animals. They also feed on dead bodies of all kinds. Foxes eat live insects of different groups. During the time of joom assembling by ripen larvae at the tree base, foxes use to eat the ripen larvae if continuous watching and scaring way is not taken to the muga crop. Since muga cropping is practiced in outdoor Som or Soalu plantations it becomes easy for the foxes living nearby forest like vegetation. Although different types of foxes are found all over the world, generally two types of foxes are noticed in Assam. One is of smaller in size with grey coloration. This type of fox commonly live hiding underground nearby riversides, lone areas. The other type is a little bit bigger in size like medium sized dog and reddish-brown in color. They live nearby

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 514
thin forest like vegetation away from human habitat. Fox belongs to the family Canidae under order Carnivora and genus Vulpes.
Rats are very thickly populated mammalian pest of a good number of agricultural crops. Besides, rats are also responsible for spreading some of important diseases and causes damages to household materials including cloths, plastic-wooden utensils, fire from electricity, flood by weakening the river dams by intensive burrowing, derailing by weakening the railway track etc. They live most commonly in human habitat, sewages, jungles, railway tracks, crop fields, cannel dams etc. They belong to Rodentia family of vertebrata. Rodents are of different types in their body size, color, nature, habitat, and body characters. In Assam six different species of rodents are found. Out of these Rattus rattus (Indian mole rat), Rattus norvegicus (sewage rat) and Mus mascullus (house mouse) are found to live in house where foods are easily exposed for their consumption. The other three species are Bandicota bengalensis, B. indica and Mus mus (field mouse). Rats are omnivorous animal. Although their main food is staple food grains, they eat meat of other organisms. The species B. indica lives in forest, horticultural gardens, and jungles of nearby crop fields. Being nutritious the muga silkworm larvae and pupa are eaten by rats. Rats are good climber and can climb Som or Soalu plant for their food. Being nocturnal it is difficult to protect muga silkworm larvae from the rats, particularly when there is scarcity in availability of other food items in their habitat.
Members of Primates are different species and traits of commonly called monkey. In Assam different species of this Primate group are abundantly found in different habitats conditions starting from dense forest to horticulture gardens to temple premises. They are different in size, body colour, appearance, habits etc. Monkey range in size from the Pygmy Marmoset at 140 to 160 millimeters(5-6 inch) long(plus tail) and 120 to 140 grams in weight, to the male Mandrill, almost 1 meter (3.3 ft) long and weighting 35 kg. Diets differ among the various species but may contain any of the following – fruit, leaves, seeds, nuts, flowers, eggs and small animals including insects and spiders. Muga larvae of 4th and 5th instars when become flashy are most favorite diet of the common monkey. Monkey being intelligent than any other animals, apply improve technique to collect and carry muga larvae from the crop field. They come to the muga crop on Som or Soalu trees and carry the larvae by keeping in large number on their bodies. Since monkey is cleaver animal, farmers face great problem in controlling their infestation. Moreover, there is no known controlling method for monkey.
LITERATURE CITED "Squirrel", 2010. Online Wikipedia, the free encyclopedia. Retrieved on 2010-07-13. “Squirrel, Rodents” 2010. Online Indianetzone; Flora & Fauna. Retrieved on 2010-07-13. Nameer, P. O. & Molur, S. 2008. Funambulus palmarum. In: IUCN 2008. IUCN Red List of Threatened Species. Downloaded on 27th November, 2009. Human Ageing Genomic Resources, 2010. AnAge database, retrieved 2010-07-09. AnAge entry for Funambulus palmarum. "Rabbit"2007. Encyclopædia Britannica (Standard Edition ed.). Chicago: Encyclopædia Britannica, Inc.. 2007. Wikipedia 2010. Wikipedia, the free encyclopedia. Retrieved on 2010-07-13. http://www.indianetzone.com/43/bats.htp. Retrieved on 2010-07-13.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 515
http://www.batcon.orgindex.php/media-and-info/bats-archives.htm/?task=vie4wArticle&magArticleID=707. Retrieved on 2010-07-13.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 516
SCIENTIFIC NOTES
NEW REPORT OF COCCINELLID PREDATORS, (COL.: COCCINELLIDAE) FEEDING ON APHID, SCHIZONEURAPHIS
HIMALAYENSIS (GHOSH & RAYCHAUDHURI) (HEM.: APHIDIDAE) ON MUGA FOOD PLANT SOM, PERSEA
BOMBYCINA KOST IN ASSAM, INDIA
S. G. Eswarareddy* and R. K. Rajan * Central Muga Eri Research & Training Institute, Central Silk Board, Ministry of Textiles, Govt. of India, Lahdoigarh, Jorhat-785 700, Assam, INDIA. E-mail: [email protected] [Eswarareddy, S. G. & Rajan, R. K. 2011. New report of coccinellid predators, (Col.: Coccinellidae) feeding on aphid, Schizoneuraphis himalayensis (Ghosh & Raychaudhuri) (Hem.: Aphididae) on Muga Food Plant Som, Persea Bombycina Kost in Assam, India. Munis Entomology & Zoology, 6 (1): 516-517]
Som, Persea bombycina Kost is the primary food/host plant of muga silkworm, Antheraea assamensis (Helfer) (Lepidoptera: Saturniidae) for the production of golden yellow silk in Assam (India). It is attacked by borers, beetles, gall insects, sucking pests. Among sucking pests, recently small green aphids infested the som plants after pruning particularly during new flush. Both nymphs and adults were found in the colonies on tender buds, shoots, lower surface of leaves and suck the sap which results in curling and reduction in the size of the leaves. Aphids also secrete honeydew on tender parts of the plant and develop black sooty mould which affects the photosynthesis activity, vigour of the plant and deterioration of the quality of the leaves which becomes unfit for rearing of A. assamensis. Presence of ants on the plants indicates the incidence of aphids.
During the incidence of aphids on P. bombycina, the number of predatory coccinellid nymphs and adults of Coleophora bowringi Crotch, Coleophora saucia Mulsant, Harmonia dimidiata (Fabricius), Phrynocaria unicolor (Fabricius) (Coleoptera: Coccinellidae) were observed feeding (Fig 1 to 7) on the nymphs and adults of aphid, Schizoneuraphis himalayensis on P. bombycina during the month of June 2010 at Farm No.1, Central Muga Eri Research & Training Institute, Lahdoigarh, Jorhat, Assam (India). This is the first record of above coccinellid predators feeding on aphids. There fore conservation of these predators in som plantations is need of the hour to reduce the aphid population naturally without reaching to the Economic Injury Level (EIL). The above predators can be used as a potential bio control agents for the management of aphids in the primary host plant (som) of muga silkworm.
ACKNOWLEDGEMENTS
Authors thankful to Dr. J. Poorni, Principal Scientist and Dr. Sunil Joshi, Senior Scientist, National Bureau of Agriculturally Important Insects, Indian Council of Agricultural Research, Govt. of India, Bengaluru for identification of predatory coccinellids and aphids respectively.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 517
1 2 Figure. 1 Coccinellid grubs feeding on aphids. Figure 2. Phrynocaria unicolor adults feeding on aphids.
3 4 5 Figure 3. Phrynocaria unicolor. Figure 4. Phrynocaria unicolor Figure 5. Phrynocaria unicolor. adult feeding on nymph of aphid. 6 7 Figure 6. Coleophora bowringi. Figure 7. Hormonia dimidiata.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 518
SCIENTIFIC NOTES NEW REPORT OF XYLOTRUPES GIDEON (LINNAEUS) (COL.:
SCARABAEIDAE) ON SOM, PERSEA BOMBYCINA KOST, A FOOD PLANT OF MUGA SILKWORM, ANTHERAEA
ASSAMENSIS (HELFER) (LEP.: SATURNIIDAE) IN ASSAM, INDIA
S. G. Eswarareddy* and R. K. Rajan
* Central Muga Eri Research & Training Institute, Central Silk Board, Ministry of Textiles, Govt. of India, Lahdoigarh, Jorhat-785 700, Assam, INDIA. E-mail: [email protected] [Eswarareddy, S. G. & Rajan, R. K. 2011. New report of Xylotrupes Gideon (Linnaeus) (Col.: Scarabaeidae) on Som, Persea Bombycina Kost, a food plant of Muga Silkworm, Antheraea assamensis (Helfer) (Lep.: Saturniidae) in Assam, India. Munis Entomology & Zoology, 6 (1): 518-519]
Som, Persea bombycina Kost is the primary food plant for rearing of muga silkworm, Antheraea assamensis (Helfer) (Lepidoptera: Saturniidae) in out door conditions (Som plantations) for the production of golden yellow silk in Assam (India). This perennial tree is attacked by the number of insect pests (caterpillars, borers, beetles, gall insects, sucking pests) and diseases their by affecting the quality of the leaves, reduction in the leaf yield which indirectly influences the production of muga silk. During frequent visits to the institute farms, the new beetles were observed feeding by boring on the stem of som tree, the collected beetles got identified by International taxonomist as Xylotrupes gideon (Linnaeus). This beetle is reported first time in som plantations of Farm No.1 & 3 at Lahdoigarh, Jorhat, Assam during May-October 2010.
These beetles are reddish brown to black and have long horn projecting dorsally from the head in male and in female, the horn is short. Adult beetle responsible for causing damage to som plants and seen at the base of the stem and tree trunks. These beetles mainly bore at the base of the stem and also on tree trunks and feed internal tissues/contents by making big hole from outside without entering inside the stem. During boring, fibrous woody material was comes out from the bored hole. Repeated attack of beetle reduces the growth of the plant. Gummy exudation was observed from bored holes of the stem, which in turn attracts flies.
This beetle is also reported on coconut (in Malaysia, Solomon Islands), oil palm (in Malaysia), Poinciana, Delonex regia (in Indonesia, British Solomons, New Herbides), Caco tree (in New Britain) etc. The biology of X. gideon is yet to be studied on som in Assam conditions. As per the earlier reports on coconut and other host plants in different countries, female lay eggs in manure/compost pits, dead and decaying organic matter, undisturbed heaps. After hatching, the grubs which feed on the decaying matter and adults bore on stem. Average duration of the developmental stages of eggs (21 days), total larval period (188 days), pre pupa (14 days), pupa (2 days), adult female (102 days) and female (90 days) (Bedford, 1975). Management: Collection and destruction of beetles present in the stem, trapping of beetles with attractive breeding material mixed with insecticide, installation of sex pheromone traps to attract female beetles, application of neem

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 519
based granules (Avana @ 1.5 kg/Ac.) or Carbaryl 75 WP (2g/l) to the soil to control the grubs breeding in the soil to be followed to reduce the incidence of beetle in the plantations.
ACKNOWLEDGEMENTS
Authors thankful to Dr. C.A. Viraktamath, Emeritus Professor and Principal Investigator, Network Project on Biosystematics, Department of Agricultural Entomology, University of Agricultural Sciences, GKVK, Bengaluru, Karnataka (India) for identification of insect specimens.
LITERATURE CITED Bedford, G. O. 1975. Observations on the biology of Xylotrupes gideon (Coleoptera: Scarabaeidae) in Melanesia. Journal of Australian Entomological Society, 14: 213-216.
Male Female Figure 1. Adult beetles of Xylotrupes gideon.
Figure 2. Adult X. gideon feeding and boring on the stem of Som, Persea bombycina. Figure 3. Bored hole on stem due to feeding by X. Gideon and gummy substance coming out from the bored hole.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 520
PROF. DR. ALİ DEMİRSOY
He came to the world in 1945 as the son of Mehmet Sadık Demirsoy, a peasant–farmer from Kemaliye, Erzincan prov., Turkey. Demirsoy attended his village Primary School in 1956; Kemaliye Secondary School in 1959; Ankara Gazi High School in 1962. In 1966, he graduated from the department of Natural Sciences of the Faculty of Science, Ankara University. He became an Assistant at the
Department of Biology, Atatürk University in the same year. With a doctorate thesis on Orthoptera Fauna of Erzurum and its environs, Demirsoy became a doctorate degree holder in 1971. In the same year, with a scholarship from DAAD, he went to Germany for language education. On completion of his language course, he won Humboldt bursary to study both at Hamburg University in Paris and in Research Institudes in London. After a thesis on Turkey‘s Caelifera Fauna, Demirsoy made a Habilitasion in 1974. Again, within this period, with finance provided by the United Nations, he participated actively in the study of Oceanography, fishes and sea current in deep sea scientific research at the pole, Grönland and around Iceland. Also in 1984, he won the Alexander von Humboldt bursary again to continue a research on Turkey‘s Fauna at the Zoology Institude of Hamburg University.
He was appointed to Hacettpe University in 1978. In 1980-1981, Prof.
Dr. Demirsoy was the Head of Department of Zoology; 1981–1982 - the Dean, Faculty of Arts and Sciences, Hacettepe University.
Prof. Dr. Demirsoy is continuing his work especially on Orthoptera
whose scientific studies he started together with the other insect groups and invertebrates. It is on record that, his scientific books concerning vertebrate and invertebrate organisms have been the first and comprehensive works on the biodiversity of Turkey. At the end of his studies, he had identified 20 new species for the scientific world in addition to the other 16 new species identified by other scientists and Demirsoy‘s name given. Beside Turkish Fauna, Environment, Zoogeography and Biodiversity, he is also into Nature Conservation and Biology Education.
More than 10 series of lecture books under the name ‗‘The Basic Laws
of Life‘‘ written by Prof. Dr. Ali Demirsoy are still being used as reference sources at the department of Biology in Turkish Universities. Apart from

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 521
lecture books, research, experiment and science, there are other popular works done by Demirsoy which have played the leading roles in the introduction and the development of Biological Sciences in our country.
Prof. Dr. Ali Demirsoy has supervised tens of hundreds of both
Masters and Doctorate students on areas yet to be studied in Biology in Turkey and they are today, distinguished and successful scientists in the world in their different fields of endeavour.
With the efforts shown by Demirsoy toward the development of
Biological Sciences and Education, he is known to be one of the distinguished scientists in Turkey. His work discipline, productivity and personality are worth emulation by young scientists. He has emphasised that starting from human relations to economy, it is necessary that all professional groups should know Biology and has analysed the connections between social relationship and Biogical life. Far from being dogmatic, it is mentioned that a real scientist should have analytic thinking quality. In the development of societies, it is necessary that science and its flow should be in the forefront and within this period, Biological Science has always played the locomotive role.
Presently continuing with his works at the Faculty of Sciences,
Department of Biology of Hacettepe University, Prof. Dr. Ali Demirsoy is not only shining his Biological Science lights illuminating his environment but also, he is enlightening the whole universe. By
Assoc. Prof. Dr. Aydın AKBULUT

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 522
EDITORIAL NOTE BY DR. HÜSEYĠN ÖZDĠKMEN
TO THE MEMORY OF SENIOR RESEARCH SCIENTIST
DR. DMITRY VLADIMIROVICH OBYDOV
I was received a message from Prof. Pierfranco Cavazzuti (Italy) in 13.XII.2010. He informed me about very bad news that my friend Dr. Dmitry Obydov (Russia) dead. So, I dedicated to the loving memory of Dr. Obydov. I have copied the information about him from the Web-Site http://www.zin.ru/ animalia/coleoptera/eng/obydov.htm. I wish to condelence to the Family and Scientific World.
Dmitry V. Obydov - the expert of a world level on taxonomy of the
tribe Carabini. Dmitry - the graduate of the Moscow medical academy named after I. M. Setchenov. His scientific interests - taxonomy of one of large and practically important family of the beetles - the ground beetles (Carabidae). Dmitry - the big enthusiast of scientific work and the fine field researcher who has collected a unique material on Carabini of fauna of Russia and the adjacent lands, and also Oriental Region. Dmitry - the author of 75 articles and several monographies. He have brought especially important contribution to development of taxonomy of such "difficult" groups as genus Callisthenes and subgeneric group "Megodontus" of the genus Carabus.
Dmitry for a long time supports communications with the Zoological
Institute of the Russian Academy of Sciences, actively uses a collection of the beetles in the scientific researches, constantly transfer in the collection of ZIN the paratypes of taxa described by him. Dmitry participates in a number of joint publications with the employees of ZIN, in particular, he participated in a writing of the new edition of the Red Data Book of the Russia Federation (2001).
A. L. Lobanov, June 2008

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 523
EDITORIAL NOTE BY DR. HÜSEYĠN ÖZDĠKMEN
DR. MIKHAIL LEONTIEVITCH DANILEVSKY
My friend Dr. Danilevsky (Russia) is most prominent specialist on the fauna and taxonomy of Cerambycidae of Russia and of the territory of the former USSR. From 1966 up to now, many scientific works by him (including monographs), and many new taxa were described and published by him. Special attention was payed by Danilevsky to the tribe Dorcadiini. His collection trips have been focused to whole Russian territorries, Eastern Europe, Central Asia, Caucasus, Iran. He is also known as one of the best specialists on Cerambycidae larval morphology. Larvae of several dozens of Cerambycidae species
were originally described by him. Danilevsky is responsible for computor versions of annotated
Cerambycidae Checklists of Europe and of the territory of the former USSR (see http://www.cerambycidae.net/).
Danilevsky was born in Moscow in 1948. He began to collect beetles in
final years of middle level school (1965-66), when entered" the biology circle "VOOP" directed by P.P. Smolin (attached to All-Union Society of Nature Protection). In 1971 Danilevsky graduated Biological Faculty of Moscow State University as entomologist. His diploma was devoted to Caucasian Cerambycidae larvae. Just after University he began to study wood-boring Coleopterous larvae in the Institute of Evolutionary Morphology and Ecology of Animals of A.N. Severtsov. In 1977 in the same Institute (now A.N. Severtsov Institute of Ecology and Evolution) Danilevsky defended Ph. D. dissertation on the evolution of Cerambycidae larval morphology with phylogenetic conclusions to the the structure of the family.
Presently continuing with his works at the A. N. Severtsov Institute of
Ecology and Evolution Russian Academy of Sciences, Group of ecology and morphology of xylobionts as a major research scientist in Moscow, Russia. * This note was prepared by H. Özdikmen on the base of the note of A. L. Lobanov, January 2002 in the Website http://www.zin.ru/animalia/ coleoptera/eng/danilevs.htm.

_____________Mun. Ent. Zool. Vol. 6, No. 1, January 2011__________ 524
EDITORIAL NOTE BY DR. HÜSEYĠN ÖZDĠKMEN
GIANFRANCO SAMA
My friend Gianfranco Sama (Italy) is one of the best specialists on Cerambycidae in Holarctic and African zooregions.
He was born in Cesena in 1944. He became interested in entomology while very young, collecting Scarabaeidae, Cerambycidae and Carabidae. In 1973 he met the Austrian entomologist Peter Schurmann, who became his master and who shared with him his whole experience of the research and rearing of immature stages of Cerambycidae. Since the he is carrying on surveys on
phytophagous beetles (expecially Cerambycidae and Buprestidae) from the Mediterranean area. After a first trip to Turkey (1974), he carried out several expeditions to southern Italy, Spain and Balcan Peninsula, often together with Schurmann.
From 1965 to 1969, at the University of Bologna (Fac. Agraria, Istituto di Entomologia) as technician of laboratory and researcher.
From 1971, Collaborator of the Museo di Storia Naturale di Verona and the University of Rome (Istituto di Zoologia) with research on Cerambycidae in Algeria, Morocco, Greece and Turkey.
From 1988, collaborator of the C.N.R. (Consiglio Nazionale delle Ricerche) with pubblication of the volume XXVI regarding Coleoptera Cerambycidae (Fauna d'Italia) and volume 59 Coleoptera Polyphaga XIV (Cerambycidae) of ―Checklist delle specie della fauna italiana‖.
Further researches in Turkey, Spain, Israel, Jordan, Greece, Canary Isl., Tunisia, Lebanon, Syria, Iran, Sardinia, Morocco, Malta, Qatar, Sinai, Egypt, southern Italy, Sicily, UAE, Albania etc.
All materials collected during these trips have been published in about 130 scientific magazines or books, dealing with taxonomy, systematics and faunistic mostly of Cerambycidae, but also Carabidae and Buprestidae and including the description of about 150 new taxa covering Palaearctic, African and Nearctic zooregions. The Cerambycidae are also being illustrated in a Monograph dealing with the Fauna of Europe and the Mediterranean area (including Caucasus and Iran).He is currently engaged with the publication of two volumes dealing with North Africa and the Atlantic Islands.
I have copied the information about him by Guido Sabatinelli, 2008
from the Web-Site http://www.glaphyridae.com/Research/Collaboratori. html#sama.
Related Documents











