Current Pharmaceutical Design, 2007, 13, 2935-2950 2935 1381-6128/07 $50.00+.00 © 2007 Bentham Science Publishers Ltd. Trends in Snakebite Envenomation Therapy: Scientific, Technological and Public Health Considerations José María Gutiérrez * , Bruno Lomonte, Guillermo León, Alexandra Rucavado, Fernando Chaves and Yamileth Angulo Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica Abstract: The therapy of snakebite envenomation has been based on the parenteral administration of animal-derived antivenoms. Despite the success of this treatment at reducing the impact of snakebite mortality and morbidity, mostly due to their capacity to neutralize sys- temically-acting toxins, antivenoms are of relatively low efficacy in the prevention of venom-induced local tissue damage, which often leads to permanent disability. The issue of safety also remains a concern, particularly for some antivenoms which induce a relatively high incidence of adverse reactions. Consequently, there is a need to improve the therapy of snakebite envenomations on the following lines: (a) the technologies to produce antivenoms require improvements aimed at obtaining more refined preparations of higher efficacy and safety, while being affordable for the public health systems of developing countries. (b) The growing knowledge on the biochemistry and toxicology of snake venoms should pave the way for the identification of natural and synthetic inhibitors of venom toxins, particularly of those involved in local tissue pathology. Such inhibitors might become a highly effective therapeutic tool for the abrogation of venom- induced local tissue damage. (c) A better knowledge of the inflammatory events secondary to venom actions may open the possibility of modulating such response, in order to prevent further tissue damage and to promote successful tissue repair and regeneration. A global partnership, involving many participants and combining scientific, technological and public health actions, is required to achieve a leap forward in the treatment of snakebite envenomations world-wide. Key Words: Snake venom, antivenom, metalloproteinase inhibitors, phospholipase A 2 inhibitors, necrosis, hemorrhage, inflammation. INTRODUCTION: THE GLOBAL PROBLEM OF SNAKE- BITE ENVENOMATION Envenomations induced by snakebites represent a relevant pub- lic health issue in many regions of the world, particularly in tropical and subtropical countries of Africa, Asia, Latin America and Oce- ania. Although reliable epidemiological information on the actual magnitude of this problem world-wide is scarce, the number of envenomated people every year is likely to be over two million, with tens of thousands of fatalities [1, 2]. There are specific regions in Africa, Asia and Latin America where the incidence and death rates of these envenomations are very high [2-4]. In addition, a percentage of envenomated people that do not die develop severe local tissue damage leading to permanent tissue loss and disability [5, 6]. The species responsible for the highest load of bites and mortality are classified within the families Viperidae and Elapidae, such as the saw-scaled vipers (genus Echis) in Africa [7], the lance- headed pit vipers (Bothrops atrox and B. asper) in South and Cen- tral America [8, 9], and the cobras (genus Naja) and kraits (genus Bungarus) in Asia [6]. Envenomations by viperid snakes induce prominent local patho- logical alterations, i.e. edema, hemorrhage, blistering and necrosis, and a complex pattern of systemic pathophysiological effects asso- ciated with hemorrhage, hemostatic disturbances, hemodynamic perturbations leading to cardiovascular shock, and acute renal fail- ure [8-10]. However, some species, such as the rattlesnakes Crota- lus durissus terrificus and C. scutulatus, do not induce local tissue damage and inflammation but instead promote neurotoxicity, sys- temic myotoxicity, coagulopathy and acute renal failure [11]. En- venomations by elapid species (cobras, kraits, mambas, Australa- sian species and sea snakes) are characterized by neurotoxicity owing to the effect of pre- and post-synaptically-acting neurotoxins which target neuromuscular junctions [6]. The venoms of some cobras cause local necrosis [6, 7] and various elapid venoms, in- cluding those of sea snakes, also induce rhabdomyolysis [12]. Nev- ertheless, beyond these general pathophysiological patterns, there is *Address correspondence to this author at the Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica; Tel: 506-2293135; Fax: 506-2920485; E mail: [email protected] a great variation in the clinical manifestations of snakebite enveno- mations due to the complexity and diversity of snake venom pro- teomes. This review focuses on the treatment of these envenoma- tions, emphasizing on emerging alternatives to improve the therapy of this important disease. IMMUNOTHERAPY OF ENVENOMATIONS: ACHIEVE- MENTS AND LIMITATIONS The parenteral administration of heterologous antivenoms, pre- pared from the plasma of animals, mostly horses or sheep, immu- nized with venoms, constitutes the mainstay in the therapy of snakebite envenomations, since the development of the first an- tivenoms in 1894 [13, 14]. Currently, antivenoms are manufactured in many laboratories in every continent [15] and are composed of either whole IgG molecules, divalent F(ab’) 2 fragments or monova- lent Fab fragments [16]. Most producers manufacture F(ab’) 2 prepa- rations by using a protocol based on pepsin digestion of IgG and further purification of F(ab’) 2 fragments by various techniques, among which ammonium sulphate precipitation is the most com- mon [17], following the general principles of the methodology de- veloped by Pope [18]. Innovations to this general protocol have been introduced over the years, including ion-exchange chromatog- raphy [19, 20], affinity chromatography [21], caprylic acid stabili- zation of IgG [22, 23], and introduction of viral-inactivation steps, such as pasteurization [19]. In general, horses or sheep are immu- nized with whole venoms from either individual species (to obtain ‘monospecific antivenoms’) or various species (to obtain ‘poly- specific antivenoms’) [16]. In terms of presentation, most antiven- oms are liquid, i.e. requiring storage at 4-10 °C and therefore de- manding a cold chain, whereas other products are freeze-dried, thus allowing transportation and storage at higher temperatures and hav- ing a more expanded shelf-life than liquid products. Therapeutic Efficacy of Antivenoms The introduction of antivenoms more than a century ago greatly reduced the mortality of snakebite envenomations. When adminis- tered on time, antivenoms are highly effective in the neutralization of toxins responsible for the systemic effects associated with en- venomations, such as hemorrhage, coagulopathy, and hemodynamic and renal alterations in viperid snakebites, and neurotoxicity caused

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Current Pharmaceutical Design, 2007, 13, 2935-2950 2935
1381-6128/07 $50.00+.00 © 2007 Bentham Science Publishers Ltd.
Trends in Snakebite Envenomation Therapy: Scientific, Technological and Public Health Considerations
José María Gutiérrez*, Bruno Lomonte, Guillermo León, Alexandra Rucavado, Fernando Chaves and Yamileth Angulo
Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica
Abstract: The therapy of snakebite envenomation has been based on the parenteral administration of animal-derived antivenoms. Despite the success of this treatment at reducing the impact of snakebite mortality and morbidity, mostly due to their capacity to neutralize sys-temically-acting toxins, antivenoms are of relatively low efficacy in the prevention of venom-induced local tissue damage, which often leads to permanent disability. The issue of safety also remains a concern, particularly for some antivenoms which induce a relatively high incidence of adverse reactions. Consequently, there is a need to improve the therapy of snakebite envenomations on the following lines: (a) the technologies to produce antivenoms require improvements aimed at obtaining more refined preparations of higher efficacy and safety, while being affordable for the public health systems of developing countries. (b) The growing knowledge on the biochemistry and toxicology of snake venoms should pave the way for the identification of natural and synthetic inhibitors of venom toxins, particularly of those involved in local tissue pathology. Such inhibitors might become a highly effective therapeutic tool for the abrogation of venom-induced local tissue damage. (c) A better knowledge of the inflammatory events secondary to venom actions may open the possibility of modulating such response, in order to prevent further tissue damage and to promote successful tissue repair and regeneration. A global partnership, involving many participants and combining scientific, technological and public health actions, is required to achieve a leap forward in the treatment of snakebite envenomations world-wide.
Key Words: Snake venom, antivenom, metalloproteinase inhibitors, phospholipase A2 inhibitors, necrosis, hemorrhage, inflammation.
INTRODUCTION: THE GLOBAL PROBLEM OF SNAKE-
BITE ENVENOMATION
Envenomations induced by snakebites represent a relevant pub-lic health issue in many regions of the world, particularly in tropical and subtropical countries of Africa, Asia, Latin America and Oce-ania. Although reliable epidemiological information on the actual magnitude of this problem world-wide is scarce, the number of envenomated people every year is likely to be over two million, with tens of thousands of fatalities [1, 2]. There are specific regions in Africa, Asia and Latin America where the incidence and death rates of these envenomations are very high [2-4]. In addition, a percentage of envenomated people that do not die develop severe local tissue damage leading to permanent tissue loss and disability [5, 6]. The species responsible for the highest load of bites and mortality are classified within the families Viperidae and Elapidae, such as the saw-scaled vipers (genus Echis) in Africa [7], the lance-headed pit vipers (Bothrops atrox and B. asper) in South and Cen-tral America [8, 9], and the cobras (genus Naja) and kraits (genus Bungarus) in Asia [6].
Envenomations by viperid snakes induce prominent local patho-logical alterations, i.e. edema, hemorrhage, blistering and necrosis, and a complex pattern of systemic pathophysiological effects asso-ciated with hemorrhage, hemostatic disturbances, hemodynamic perturbations leading to cardiovascular shock, and acute renal fail-ure [8-10]. However, some species, such as the rattlesnakes Crota-lus durissus terrificus and C. scutulatus, do not induce local tissue damage and inflammation but instead promote neurotoxicity, sys-temic myotoxicity, coagulopathy and acute renal failure [11]. En-venomations by elapid species (cobras, kraits, mambas, Australa-sian species and sea snakes) are characterized by neurotoxicity owing to the effect of pre- and post-synaptically-acting neurotoxins which target neuromuscular junctions [6]. The venoms of some cobras cause local necrosis [6, 7] and various elapid venoms, in-cluding those of sea snakes, also induce rhabdomyolysis [12]. Nev-ertheless, beyond these general pathophysiological patterns, there is
*Address correspondence to this author at the Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica; Tel: 506-2293135; Fax: 506-2920485; E mail: [email protected]
a great variation in the clinical manifestations of snakebite enveno-mations due to the complexity and diversity of snake venom pro-teomes. This review focuses on the treatment of these envenoma-tions, emphasizing on emerging alternatives to improve the therapy of this important disease.
IMMUNOTHERAPY OF ENVENOMATIONS: ACHIEVE-
MENTS AND LIMITATIONS
The parenteral administration of heterologous antivenoms, pre-pared from the plasma of animals, mostly horses or sheep, immu-nized with venoms, constitutes the mainstay in the therapy of snakebite envenomations, since the development of the first an-tivenoms in 1894 [13, 14]. Currently, antivenoms are manufactured in many laboratories in every continent [15] and are composed of either whole IgG molecules, divalent F(ab’)2 fragments or monova-lent Fab fragments [16]. Most producers manufacture F(ab’)2 prepa-rations by using a protocol based on pepsin digestion of IgG and further purification of F(ab’)2 fragments by various techniques, among which ammonium sulphate precipitation is the most com-mon [17], following the general principles of the methodology de-veloped by Pope [18]. Innovations to this general protocol have been introduced over the years, including ion-exchange chromatog-raphy [19, 20], affinity chromatography [21], caprylic acid stabili-zation of IgG [22, 23], and introduction of viral-inactivation steps, such as pasteurization [19]. In general, horses or sheep are immu-nized with whole venoms from either individual species (to obtain ‘monospecific antivenoms’) or various species (to obtain ‘poly-specific antivenoms’) [16]. In terms of presentation, most antiven-oms are liquid, i.e. requiring storage at 4-10 °C and therefore de-manding a cold chain, whereas other products are freeze-dried, thus allowing transportation and storage at higher temperatures and hav-ing a more expanded shelf-life than liquid products.
Therapeutic Efficacy of Antivenoms
The introduction of antivenoms more than a century ago greatly reduced the mortality of snakebite envenomations. When adminis-tered on time, antivenoms are highly effective in the neutralization of toxins responsible for the systemic effects associated with en-venomations, such as hemorrhage, coagulopathy, and hemodynamic and renal alterations in viperid snakebites, and neurotoxicity caused

2936 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
by post-synaptically acting short-chain neurotoxins in elapid snake-bites [5]. Moreover, antivenom-reversal of neurotoxic effects in-duced by post-synaptically acting neurotoxins has been demon-strated [5]. In the case of viperid envenomations, the administration of an adequate initial antivenom dose achieves the cease of bleed-ing within 30 min to 6 hr, and corrects coagulation parameters within 6-24 hr [24-26], these being the most relevant clinical and laboratory parameters to judge antivenom efficacy.
The molecular basis of neutralization of toxins by antibodies and their fragments is the blockade of the molecular regions of the toxins responsible for their actions. Such blockade may occur by various mechanisms: (a) the ‘toxic site’ of the toxin might be the epitope recognized by the antibody, (b) antibodies may recognize an epitope located nearby the ‘toxic site’, neutralization being ac-complished by steric hindrance, (c) antibodies may recognize an epitope distant from the ‘toxic site’, inducing conformational changes in the toxin which greatly reduce its affinity to the target, and (d) antibodies may form immunocomplexes with toxins, thus promoting their elimination by phagocytic cells [27, 28]. The affin-ity of antivenom antibodies to their toxins greatly determines their therapeutic efficacy, since antibodies of high affinity are more likely to form stable imunocomplexes with toxins.
Therapeutic Limitations of Antivenoms
In contrast with the efficacy of antivenoms to control most of the systemic manifestations of snakebite envenomations, they have proven partially ineffective in the neutralization of toxins responsi-ble for the local pathological effects induced by most viperid and some elapid snake venoms [5, 29]. The basis for this poor therapeu-tic success is not due to the absence of antibodies against locally-acting toxins in antivenoms, since neutralization assays performed by incubating venom and antivenom prior to testing in laboratory animals demonstrate adequate neutralization [29]. Instead, such lack of neutralization when antivenom is administered after en-venomation, as occurs in actual snakebites, is mostly due to the extremely rapid development of local pathological alterations, which may attain a significant extent within minutes [29-31]. In these circumstances, antibodies or their fragments reach the af-fected tissues only after damage has occurred, and venom-induced microvascular alterations further preclude antivenom distribution to affected tissue [29, 32, 33]. This therapeutic failure is also evident in the case of local edema, which continues to develop despite the administration of antivenom [34]. Edema is mediated by a variety of endogenous inflammatory mediators synthesized and released in the tissues as a consequence of envenomation [35, 36]; thus, ad-ministration of antivenom cannot control such cascades once they are triggered. Moreover, antivenoms have been rather ineffective in reverting neurotoxicity induced by presynaptically-acting neuro-toxic phospholipases A2 (PLA2) [5]. This is due to the fact that such neurotoxins damage the integrity of the nerve terminal, thus pro-longing the effect until muscle reinnervation takes place [37].
A phenomenon related with the poor neutralization of locally-acting toxins and presynaptic neurotoxins is the mismatch between the toxicokinetics of venom myotoxins and neurotoxins and the pharmacokinetics of antibodies in antivenoms [28, 38]. Owing to the low molecular mass of many snake venom components, upon injection they diffuse rapidly into a large volume of distribution, whereas antibodies or their fragments reach a more reduced volume of distribution [28, 38, 39]. Nevertheless, it has been proposed that binding of antibodies to toxins occurs predominantly in the central compartment, i.e. in plasma, and that such toxin sequestration by antibodies promotes a redistribution of tissue-located toxins to the central compartment where they are complexed with antibodies [40, 41]. Thus, the administration of an adequate dose of antivenom and the persistence of antibodies or their fragments in the circulation for a relatively prolonged time must assure the complexation and neu-tralization of toxins present in the bloodstream, and the consequent redistribution of toxins from tissues to the central compartment.
Nevertheless, such redistribution would be of limited therapeutic efficacy when toxins have already induced prominent tissue altera-tions, as in the case of toxins inducing hemorrhage, edema, myone-crosis and neuromuscular junction degeneration.
The Issue of Antivenom Specificity
One of the characteristics inherent to antivenom therapy is the need of an adequate specificity to the venoms being neutralized. This is particularly relevant owing to the great extent of inter- and intraspecific variation in venom composition [42], which demands antivenoms to be produced using venoms representative of the spectrum of antigenic variability from a particular region. Such antigenic complexity requires systematic preclinical studies on the efficacy of antivenoms against the most relevant venoms in each region, as performed in various Latin American countries (see [43] for a review). Results indicate that some polyspecific antivenoms have a wide spectrum of efficacy and, therefore, could be used across various countries for the treatment of viperid envenomations [44, 45]. In contrast, other antivenoms are not effective against venoms of snakes not included in the immunizing mixture [46, 47]. The lack of therapeutic efficacy of some antivenoms, when used in the treatment of envenomations induced by heterologous venoms, has been documented [48], thus reinforcing the relevance of rigor-ous preclinical and clinical assessments of antivenom efficacy be-fore their introduction and adoption for general use in a given coun-try.
Adverse Reactions to Antivenom Administration
Administration of antivenoms is associated, in a proportion of patients, with early and late adverse reactions [49]. Early adverse reactions (EAR) occur within the first hours of the onset of treat-ment and usually consist of urticaria, generalized rash, chills, nau-sea, colics, vomiting and, less often, bronchospasm, angioneurotic edema and hypotension [6, 16, 25]. Very few cases, in which prior exposure to horse or sheep IgGs has occurred, are associated with true anaphylactic shock; instead, most of the cases represent de novo adverse manifestations [6, 49]. The safety profiles of diverse antivenoms differ significantly; some products have a very good profile, inducing a low incidence of EARs, most of them mild [50]. In contrast, other products are associated with a very high incidence of EARs [51, 52]. Late adverse reactions may develop within 4-20 days after antivenom infusion and correspond to the typical ‘serum sickness’ [53], a type III hypersensitivity reaction in which the pa-tient develops antibodies against antivenom proteins [49]. The ac-tual incidence of this type of adverse reaction is unknown for most antivenoms, owing to the inherent difficulties in documenting such reactions once the patients have left health centers; it depends on several factors, among which the total load of antivenom protein injected is likely to predominate. In general terms, EAR are treated with antihistamines, steroids or adrenaline [6, 49]. Pretreatment with these drugs, before the infusion of antivenom, has been advo-cated as a way to reduce the incidence of EAR, although only the administration of adrenaline has shown positive results in con-trolled trials [49, 54, 55].
The mechanisms behind EAR have not been conclusively estab-lished. Complement activation by antivenom proteins, not necessar-ily by immune complexes, has been advocated as a possible cause [49, 56, 57]. Indeed, antivenom proteins are able to directly activate complement in vitro [56, 58-61]. Nevertheless, reduction of an-tivenom-induced complement activation by the addition of -propiolactone, during the manufacture of a whole IgG equine an-tivenom [61], was not associated with a significant drop in the inci-dence of EAR [62], thus raising doubts on the role played by com-plement activation in these reactions. For long time it has been assumed that removal of the Fc fragment of horse IgGs by pepsin digestion in the preparation of antivenoms eliminates complement activation and, consequently, decreases the incidence of EAR [63]. However, F(ab’)2 antivenoms retain complement-activating activity

Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2937
[59, 60]. The Fc region of IgG plays a role in complement activa-tion mediated by immune complexes. However, in the case of an-tivenom administration, complement activation occurs de novo, more likely through the action of protein aggregates present in an-tivenoms [56], and not necessarily by the formation of venom-antivenom complexes. Such reaction has to do more with the over-all physicochemical quality of antivenoms than with the presence of Fc in whole IgG molecules [25]. This is supported by the observa-tion that caprylic acid-fractionated whole IgG antivenoms, which present good physicochemical properties, induce a lower incidence of EARs than many F(ab’)2 antivenoms, devoid of whole IgGs but containing protein aggregates [62, 64]. Therefore, the introduction of steps, in the fractionation protocols, aimed at improving the physicochemical quality of antivenoms by obtaining more purified preparations and reducing the total protein concentration of these immunobiologicals, will contribute to the improvement of an-tivenom safety profile.
The Problem of Antivenom Accessibility in Many Countries
The success of antivenom in the therapy of snakebite enveno-mations is hampered by another factor that belongs to the public health realm, i.e. the crisis in the production, deployment and ac-cessibility of antivenoms in many parts of the world. This is par-ticularly serious in regions where the incidence of snakebites is very high, such as in sub-Saharan Africa and some countries in Asia and Latin America [65-67]. This complex problem has several causes: (a) some pharmaceutical companies, both public and pri-vate, have ceased or decreased their antivenom production, espe-cially for developing countries, due in part to issues of rentability [68]. (b) Some public antivenom production centers have been pri-vatized, with the consequent increment in the prices of antivenoms, some of which are unaffordable for the public health systems of developing countries [66]. (c) There is a crisis in the public health systems of many countries; consequently, owing to their high cost and problems of distribution, antivenoms are frequently not avail-able at rural health posts and clinics where they are mostly needed [69]. As a consequence, many people affected by snakebites do not attend these health facilities, or reach them very late [69], and in-stead often rely on empirical treatments which are ineffective, re-tard access to health centers and, in many instances, provoke delete-rious effects [4, 70]. (d) There is a lack of uniform and validated antivenom treatment protocols in many regions where snakebites constitute a relevant public health problem. This causes an inade-quate use of this precious drug in regions where it is badly needed. Such lack of standard treatment protocols is closely related to an inadequate training of physicians and nurses in the subject of snakebite envenomation therapy [69, 71].
AVENUES FOR IMPROVING THE QUALITY AND AC-
CESSIBILITY OF ANTIVENOMS
Towards Better and Safer Antivenoms
There is an urgent need to develop ambitious technological research agendas aimed at improving the quality and increasing the supply of antivenoms, while keeping the production costs low, in order to increase their effectiveness and safety, as well as to assure their distribution to countries and regions where they are mostly needed. Several issues in antivenom production demand a renewed attention:
Immunization Protocols
The growing body of knowledge on the complexity of venom proteomes should be used to identify their most medically-relevant toxins [72]. This may open the way to use immunizing mixtures comprising these relevant toxins instead of crude venoms, which contain abundant irrelevant components from the toxicological standpoint. Such toxins may be isolated from the venoms by chro-matographic procedures and may be also produced by recombinant DNA technology. A highly promising alternative is the DNA im-munization technology, based on the administration of plasmids
containing cDNA coding for relevant toxins [73]. Such strategy has been successfully used, at the experimental level, in the case of the hemorrhagic metalloproteinase jararhagin [74]. Other areas that need to be explored are the use of novel adjuvants and immuno-modulatory molecules which can enhance the immune response to venom components, as well as the development of more effective immunization protocols based on an in depth understanding of the mechanisms operating in the development of immune response in horses, sheep and other animals used in antivenom preparation.
Methods for Antivenom Purification
A wide range of protein purification technologies have been developed in the last decades, and they offer invaluable possibilities for application in antivenom production. In addition to the tradi-tional enzymatic and salting-out procedures currently employed in antivenom manufacturing, there is a possibility of using ion-exchange chromatography [20, 75], affinity chromatography [21], and novel chromatographic matrices [76], among other alternatives to obtain more refined antivenoms. Such methodologies will con-tribute to the production of antivenoms of improved physicochemi-cal quality, devoid of protein aggregates, with a reduced total pro-tein concentration and of high stability. In addition, the issue of viral and prion safety of antivenoms is receiving growing attention, and various steps have to be introduced in the plasma fractionation protocols (caprylic acid treatment, pasteurization, nanofiltration) to assure the removal of both enveloped and non-enveloped viruses, as well as of prions, that might be present in horse or sheep plasma [77].
Search for Other Animal Species for Antivenom Production
The possibility has been raised of preparing antivenoms com-posed of IgY isolated from the egg yolks of hens immunized with venoms [78]. Recently, the use of camels for immunization with venoms, particularly in Africa and the Middle East, has been sug-gested [79, 80]. Interestingly, camels have two types of IgGs, a classical four-chain IgG and a special IgG devoid of light chains [81]. Camel IgG may be less likely to induce adverse reactions than horse or sheep IgG, as suggested by various in vitro and in vivo assays utilized to estimate the potential for such reactions [82].
Antivenoms Containing Monoclonal Antibodies, Recombinant
Antibodies or Low Molecular Mass Antibody Fragments
The possibility of producing murine or ‘humanized’ mono-clonal antibodies specific against relevant venom toxins has been explored [83]. Some monoclonal antibodies show a good neutraliz-ing profile against particular toxins [27]. However, the fact that snake venoms comprise many different toxins reduces the feasibil-ity of developing antivenoms made exclusively of monoclonal anti-bodies. Nevertheless, the alternative of enriching antivenoms with monoclonal antibodies against particularly toxic venom proteins remains open. On the other hand, a conspicuous mismatch between venom toxicokinetics and antivenom pharmacokinetics has been described (see [28] for a review), particularly with low molecular mass toxins, such as elapid neurotoxins, which have a much larger volume of distribution that antibodies and which distribute rapidly to the tissues [38]. In addition, the pharmacokinetic characteristics of antivenoms have pharmacodynamic implications regarding the neutralization of locally-acting toxins [28, 29]. Thus, the possibility of using small antibody fragments, such as Fab (50 kDa) or the even smaller recombinant scFv has been considered [83-85]. An-other exciting possibility is the use of the small recombinant ‘single domain antibodies’ comprised by the variable region of the camelid IgGs constituted by the heavy chains only [86, 87]. Hence, antibody engineering offers plenty of possibilities for the design of small antibody fragments of potential application in snakebite envenoma-tion [88], provided the production costs can be kept at reasonable levels. All these fragments have a large volume of distribution and reach tissue compartment at a faster rate than IgG or F(ab’)2; never-theless, they are rapidly eliminated [28, 39], thus favoring the recur-rence of envenomation, as observed in the case of Fav antivenoms

2938 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
[89, 90]. Consequently, the strategy of producing antivenoms made exclusively of small antibody fragments is likely to have a low therapeutic efficacy. An alternative, however, is the preparation of antivenoms containing both high molecular mass neutralizing prin-ciples, such as IgG or F(ab’)2, and smaller fragments, such as Fab or scFv [28]. Such combination would assure a rapid distribution to tissue compartments of the small fragments, and the permanence of high levels of antibodies in the circulation to neutralize toxins reaching the bloodstream during a prolonged time span. In turn, circulating antivenom IgG or F(ab’)2 are able to promote, upon binding to circulating toxins, a redistribution of the latter from tis-sue sites to the central compartment [40], thus contributing to the removal of tissue-bound toxins (Fig. 1).
The Need of Meticulous Preclinical and Clinical Testing of An-
tivenoms
The development of novel antivenoms, and the introduction of established antivenoms to countries or regions where they have not been previously used, demand sustained and collaborative efforts for the preclinical testing of their effectiveness using methodologies already described and validated [see for example [43, 91, 92]. Fur-thermore, antivenoms have to be assessed in controlled clinical trials aimed at demonstrating their effectiveness and safety [67]. This demands concerted research efforts and training of local medi-cal staff in issues of bioethics and clinical research, especially in developing countries.
Public Health Issues: the Need for a Global Partnership to In-
crease Production and Accessibility of Antivenoms
The poor access to antivenoms in many countries in Africa, Asia and Latin America demands a concerted international partner-ship to increase their production, particularly in regions with a high incidence of snakebites, and to assure their accessibility. The gath-ering of accurate epidemiological information on the impact and characteristics of snakebites in these regions is a basic requirement to design effective distribution strategies [69, 71]. The undertaking of innovative international cooperation programs, involving public-private partnerships and the participation of the World Health Or-
ganization and its regional offices, is urgently needed in the design of cost-effective programs of antivenom deployment on a world-wide basis [65, 71]. The ambitious international proposals designed to confront diseases such as malaria and AIDS, based on massive donor contributions [93, 94], could be adapted on a smaller scale to address the problem of poor antivenom supply in many countries. In addition, transfer of antivenom-producing technologies to coun-tries having a high incidence of snakebites is another task that has to be urgently considered. Technology transfer programs should be concomitant with short-term training fellowships for technical and professional staff to attend consolidated antivenom-producing labo-ratories, thus fostering novel ways to promote north-south and south-south cooperative efforts. All these possibilities demand the design of innovative strategies and the political will of scientists, antivenom manufacturers, public health administrators and politi-cians alike.
THE CHALLENGE OF VENOM-INDUCED LOCAL TISSUE
DAMAGE
Understanding the Pathogenesis of Local Alterations in Snake-
bite Envenomation: A Necessary Step Towards the Develop-ment of Novel Therapeutic Strategies
Envenomations by species of the family Viperidae are, with some exceptions, characterized by a rapid development of local pathological alterations, among which edema, hemorrhage, necrosis and blistering are the most notorious [6, 95]. In addition, the ven-oms of several Asiatic and African cobras (family Elapidae) also induce local necrosis [6, 7]. The pathogenesis of viperid-induced local tissue damage has received a great deal of attention, and in-volves the direct action of toxins and the effects induced by diverse endogenous mediators that are synthesized and released as a conse-quence of venom entrance into the tissues.
Local Hemorrhage
Local hemorrhage is due to the action of venom metalloprotein-ases [96]. Snake venom metalloproteinases (SVMPs) are zinc-dependent endopeptidases that belong to the family of ‘metzincins’,
Fig. (1). Effect of antivenom administration on the distribution and inhibition of venom toxins in snakebite envenomations. (A) Venom is delivered in a tissue compartment where some toxins cause local effects, and also become absorbed into the blood (central compartment). Then, toxins are distributed, according to their toxicokinetic properties, to diverse tissue compartments in the body. Toxins may exert their deleterious effects in the tissues as well as in blood. (B) An antivenom composed of a mixture of high molecular mass neutralizing molecules (IgG or F(ab’)2; HMA) and low molecular mass neutralizing molecules (Fab or scFV; LMA) is administered by the intravenous route. Continuous arrows represent the distribution of antivenom and discontinuous arrows the distribution of toxins after antivenom administration. LMA are rapidly distributed into a large volume which includes the blood and the extracellular interstitial fluid in tissue compartments; at these locations, LMA bind and neutralize toxins present in the blood and the tissues. LMA have a very short half-life and, conse-quently, their levels in the central compartment drop rapidly. The access of LMA to the tissue compartment where venom was injected (*) is affected in the case of venoms that disrupt the microvasculature. On the other hand, HMA remain mostly in the central compartment, for a relatively prolonged time span, where they bind toxins reaching the circulation later in the course of envenomation. Binding to toxins in blood, in turn, promotes a redistribution of non-neutralized toxins from the tissues to blood, where they become neutralized. A small proportion of HMA may also reach tissue compartments.
Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2939
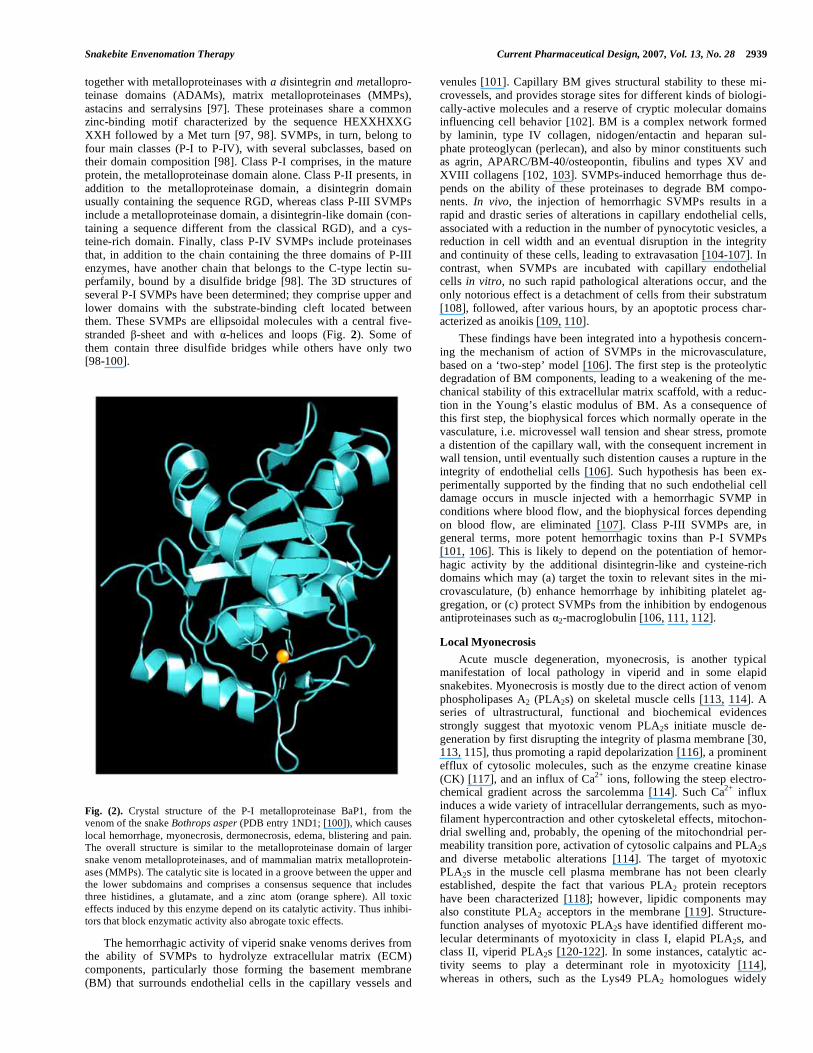
together with metalloproteinases with a disintegrin and metallopro-teinase domains (ADAMs), matrix metalloproteinases (MMPs), astacins and serralysins [97]. These proteinases share a common zinc-binding motif characterized by the sequence HEXXHXXG XXH followed by a Met turn [97, 98]. SVMPs, in turn, belong to four main classes (P-I to P-IV), with several subclasses, based on their domain composition [98]. Class P-I comprises, in the mature protein, the metalloproteinase domain alone. Class P-II presents, in addition to the metalloproteinase domain, a disintegrin domain usually containing the sequence RGD, whereas class P-III SVMPs include a metalloproteinase domain, a disintegrin-like domain (con-taining a sequence different from the classical RGD), and a cys-teine-rich domain. Finally, class P-IV SVMPs include proteinases that, in addition to the chain containing the three domains of P-III enzymes, have another chain that belongs to the C-type lectin su-perfamily, bound by a disulfide bridge [98]. The 3D structures of several P-I SVMPs have been determined; they comprise upper and lower domains with the substrate-binding cleft located between them. These SVMPs are ellipsoidal molecules with a central five-stranded -sheet and with -helices and loops (Fig. 2). Some of them contain three disulfide bridges while others have only two [98-100].
Fig. (2). Crystal structure of the P-I metalloproteinase BaP1, from the venom of the snake Bothrops asper (PDB entry 1ND1; [100]), which causes local hemorrhage, myonecrosis, dermonecrosis, edema, blistering and pain. The overall structure is similar to the metalloproteinase domain of larger snake venom metalloproteinases, and of mammalian matrix metalloprotein-ases (MMPs). The catalytic site is located in a groove between the upper and the lower subdomains and comprises a consensus sequence that includes three histidines, a glutamate, and a zinc atom (orange sphere). All toxic effects induced by this enzyme depend on its catalytic activity. Thus inhibi-tors that block enzymatic activity also abrogate toxic effects.
The hemorrhagic activity of viperid snake venoms derives from the ability of SVMPs to hydrolyze extracellular matrix (ECM) components, particularly those forming the basement membrane (BM) that surrounds endothelial cells in the capillary vessels and
venules [101]. Capillary BM gives structural stability to these mi-crovessels, and provides storage sites for different kinds of biologi-cally-active molecules and a reserve of cryptic molecular domains influencing cell behavior [102]. BM is a complex network formed by laminin, type IV collagen, nidogen/entactin and heparan sul-phate proteoglycan (perlecan), and also by minor constituents such as agrin, APARC/BM-40/osteopontin, fibulins and types XV and XVIII collagens [102, 103]. SVMPs-induced hemorrhage thus de-pends on the ability of these proteinases to degrade BM compo-nents. In vivo, the injection of hemorrhagic SVMPs results in a rapid and drastic series of alterations in capillary endothelial cells, associated with a reduction in the number of pynocytotic vesicles, a reduction in cell width and an eventual disruption in the integrity and continuity of these cells, leading to extravasation [104-107]. In contrast, when SVMPs are incubated with capillary endothelial cells in vitro, no such rapid pathological alterations occur, and the only notorious effect is a detachment of cells from their substratum [108], followed, after various hours, by an apoptotic process char-acterized as anoikis [109, 110].
These findings have been integrated into a hypothesis concern-ing the mechanism of action of SVMPs in the microvasculature, based on a ‘two-step’ model [106]. The first step is the proteolytic degradation of BM components, leading to a weakening of the me-chanical stability of this extracellular matrix scaffold, with a reduc-tion in the Young’s elastic modulus of BM. As a consequence of this first step, the biophysical forces which normally operate in the vasculature, i.e. microvessel wall tension and shear stress, promote a distention of the capillary wall, with the consequent increment in wall tension, until eventually such distention causes a rupture in the integrity of endothelial cells [106]. Such hypothesis has been ex-perimentally supported by the finding that no such endothelial cell damage occurs in muscle injected with a hemorrhagic SVMP in conditions where blood flow, and the biophysical forces depending on blood flow, are eliminated [107]. Class P-III SVMPs are, in general terms, more potent hemorrhagic toxins than P-I SVMPs [101, 106]. This is likely to depend on the potentiation of hemor- hagic activity by the additional disintegrin-like and cysteine-rich domains which may (a) target the toxin to relevant sites in the mi-crovasculature, (b) enhance hemorrhage by inhibiting platelet ag-gregation, or (c) protect SVMPs from the inhibition by endogenous antiproteinases such as 2-macroglobulin [106, 111, 112].
Local Myonecrosis
Acute muscle degeneration, myonecrosis, is another typical manifestation of local pathology in viperid and in some elapid snakebites. Myonecrosis is mostly due to the direct action of venom phospholipases A2 (PLA2s) on skeletal muscle cells [113, 114]. A series of ultrastructural, functional and biochemical evidences strongly suggest that myotoxic venom PLA2s initiate muscle de-generation by first disrupting the integrity of plasma membrane [30, 113, 115], thus promoting a rapid depolarization [116], a prominent efflux of cytosolic molecules, such as the enzyme creatine kinase (CK) [117], and an influx of Ca2+ ions, following the steep electro-chemical gradient across the sarcolemma [114]. Such Ca2+ influx induces a wide variety of intracellular derrangements, such as myo-filament hypercontraction and other cytoskeletal effects, mitochon-drial swelling and, probably, the opening of the mitochondrial per-meability transition pore, activation of cytosolic calpains and PLA2s and diverse metabolic alterations [114]. The target of myotoxic PLA2s in the muscle cell plasma membrane has not been clearly established, despite the fact that various PLA2 protein receptors have been characterized [118]; however, lipidic components may also constitute PLA2 acceptors in the membrane [119]. Structure-function analyses of myotoxic PLA2s have identified different mo-lecular determinants of myotoxicity in class I, elapid PLA2s, and class II, viperid PLA2s [120-122]. In some instances, catalytic ac-tivity seems to play a determinant role in myotoxicity [114], whereas in others, such as the Lys49 PLA2 homologues widely

2940 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
distributed in crotaline species, catalysis is not required [121]. In addition to PLA2s, local myonecrosis is also promoted by the action of hemorrhagic SVMPs, which cause ischemia secondary to mi-crovessel disruption and blood flow interruption [123]. Further-more, the increment in the muscle intracompartmental pressures reduces perfusion, causing ischemia and necrosis [10, 124].
Blistering, Dermonecrosis and Widespread Extracellular Ma-
trix Degradation
The formation of blisters and skin necrosis are typical features in the local pathology of viperid snakebite envenomation [6, 7, 24]. These effects have been reproduced in mice by injecting a hemor-rhagic SVMP, thus suggesting that hydrolysis of proteins at the dermal-epidermal junction may be a key step in the process of blis-tering [125]. In addition, the epithelial cells of the epidermis are drastically affected as part of this process. Moreover, SVMPs are able to degrade ECM components in addition to BM [101, 126], thus promoting a disorganization of the tissue extracellular scaffold, causing perturbations in the cellular spatial organization. ECM degradation is also promoted by the action of hyaluronidase, a widely distributed component in snake venoms. Such widespread degradation of ECM may also contribute to the spreading of venom toxins, as has been demonstrated in the case of a SVMP [127] and hyaluronidase [128, 129]. Moreover, ECM degradation may be mediated by the action of endogenous MMPs, which can be acti-vated by venom enzymes [130].
Beyond the Direct Pathological Actions of Venom Components:
the Role of Inflammation in Tissue Damage
The entrance of venom components into the tissues and the direct pathological effects of toxins inducing necrosis and hemor-rhage promote a prominent and complex inflammatory reaction associated with the synthesis and release of inflammatory mediators followed by the recruitment of leucocytes [95, 131]. Quantification of inflammatory mediators in envenomated tissues has evidenced increments in the levels of eicosanoids, cytokines, MMPs, kinins, nitric oxide, PAF and complement anaphylatoxins, among others [131-138]. Some of these mediators are involved in the onset of increased vascular permeability, edema [35, 36, 135] and pain [139, 140], as well as in the recruitment of an abundant inflammatory infiltrate [135, 136, 141].
There is ample evidence that PLA2s, SVMPs and probably other venom components, at sub-cytotoxic concentrations, are able to stimulate resident tissue cells, such as fibroblasts and macro-phages, to express a complex set of cellular responses associated with up-regulation of genes involved in inflammation and apoptosis in fibroblasts [142] and peritoneal adherent cells [143], and with enhanced phagocytic and pro-inflammatory activities in macro-phages [144]. SVMPs induce apoptosis in endothelial cells [109, 110] and histamine release by mast cells [145]. Moreover, a myo-toxic Lys49 PLA2 homologue is able to induce necrosis, apoptosis and cell proliferation in a lymphoblastoid cell line [146], thus sug-gesting that cells in envenomated tissues may display a wide variety of responses to toxins, ranging from necrotic or apoptotic cell death to activation and proliferation, depending on toxin concentration and on the susceptibility of specific cell types to different venom components. The scenario that emerges from these considerations is one in which the direct pathological effects of toxins combine with the activation of inflammatory and other cellular responses which, in turn, may contribute to further tissue damage, or may enhance the ability of the tissue to cope with the action of the toxins and to start effective repair and regenerative processes, as described in various models of muscle damage [147]. Moreover, recruited in-flammatory cells also contribute with a plethora of mediators which may affect the overall outcome of local tissue alterations. Under-standing such intricate host responses to different venoms and iso-lated toxins is a relevant task, in order to devise more effective ways to manipulate these processes aimed at reducing the impact of tissue damage and promoting effective regenerative responses.
Do Inflammatory Mediators Contribute to Local Tissue
Damage?
The description of a wide range of mediators and inflammatory cells in tissues injected with snake venoms and toxins prompted the hypothesis that such exacerbated inflammatory response might contribute to the local pathological alterations associated with en-venomations. This concept has been reinforced by the findings that mediators such as TNF- and other cytokines, nitric oxide, MMPs, free radicals and polymorphonuclear neutrophils, among others, play key roles in a variety of pathological conditions [147-150]. However, despite the widely held assumption that inflammatory mediators contribute to venom-induced local pathology, this hy-pothesis requires corroboration in experimental models where one or several of these mediators are inhibited.
When the acute local pathological events, i.e. hemorrhage and myonecrosis, are examined in a mouse model of local envenoma-tion by the venom of the viperid snake Bothrops asper, no role has been demonstrated in the pathogenesis of these effects for neutro-phils [151], TNF- , IL-6, IL-1 [152], nitric oxide [138] and MMPs [133]. Similar observations were performed in the case of neutro-phils for the venom of B. jararaca [153] and complement in the case of a SVMP from B. atrox venom, where the extent of hemor-rhage was not reduced by previous complement depletion [154]. These studies do not support the hypothesis that inflammatory me-diators and cells play a relevant role in these acute local pathologi-cal effects. On the other hand, however, TNF- and IL-6 have been shown to participate in the pathogenesis of local dermonecrosis after intradermal injection of a hemorrhagic SVMP in mice [155, 156], whereas nitric oxide, TNF- , IL-1 or IL-6 are not involved in the acute local hemorrhagic lesion induced by this toxin [156]. Moreover, nitric oxide participates in the local myotoxicity induced in rats by crotoxin, a myotoxic and neurotoxic PLA2 from Crotalus durissus terrificus venom [157], whereas no such a role occurs when experiments are carried out in mice (F. Chaves, B. Lomonte and J.M. Gutiérrez, unpublished results), thus reflecting a con-spicuous interspecies variation in the involvement of this mediator.
When all these observations are taken together, an emerging conclusion is that inflammatory mediators and cells do not play a determinant role in some of the acute local pathological alterations, i.e. those that develop within the first few hours of envenomation (mostly hemorrhage and myonecrosis), but may participate in other aspects of venom-induced local pathology. For instance, pathologi-cal effects that develop over several days, such as local dermone-crosis, may involve a significant participation of inflammatory agents [156]. The understanding of these complex phenomena de-mands a precise characterization of the effects being evaluated and the time frame of their development, in order to discern which cells/mediators are involved in which pathological effects. Thus, the possibility of modulating local inflammation as a way to reduce the extent of tissue damage in snakebite envenomation must await a more complete understanding of the role of inflammatory mediators in the pathogenesis of tissue damage. (Fig. 3) summarizes our cur-rent view of the pathogenesis of venom-induced local tissue altera-tions.
Growth Factors and Inflammatory Mediators and Cells Play A
Role in Tissue Reparative and Regenerative Processes
The involvement of inflammatory cells and mediators in snake venom-induced local alterations goes beyond their role in the pathogenesis of tissue damage. Early alterations induced by the action of venom components in the tissues are followed by an in-flammatory response which paves the way for reparative and regen-erative stages. Skeletal muscle regeneration, which occurs through the activation and differentiation of myogenic satellite cells, is de-pendent on the concerted actions of many cells and growth factors [147, 158, 159]. Thus, therapeutic interventions designed to control the extent of inflammation, as a way to prevent further tissue dam-age, must take into consideration that the mediators and cells that

Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2941
are being inhibited may play a beneficial role in the repair and re-generation of the tissues. For instance, elimination of neutrophils in mice does not reduce the extent of hemorrhage, myonecrosis and edema induced by B. asper venom, but significantly impairs skele-tal muscle regeneration, thus evidencing that neutrophils play a key role in muscle regeneration, presumably by removing necrotic ma-terial and stimulating the recruitment of macrophages [151]. More studies are required to gain a comprehensive understanding on the role of inflammation in the reparative and regenerative processes occurring after venom-induced tissue damage.
NOVEL TRENDS IN THE INHIBITION OF TOXINS THAT
CAUSE LOCAL TISSUE DAMAGE
Can Small Antibody Fragments Neutralize More Effectively
PLA2s and SVMPs in the Tissues?
The extremely rapid action of myotoxic PLA2s and SVMPs at the site of venom injection makes it very difficult for intravenously-administered antivenom IgG or F(ab’)2 to reach such tissue com-partments soon enough to achieve neutralization [29]. It has been speculated that the use of antivenoms containing smaller antibody fragments, such as Fab, recombinant Fv or the even smaller recom-binant single domain camelid antibody fragments would improve neutralization of snake venoms [80]. Despite the theoretical appeal of such proposal, an experimental study showed that an ovine Fab antivenom was not more effective than a whole IgG antivenom in the neutralization of local myonecrosis, hemorrhage and edema induced by the venom of B. asper [160]. Similar results were ob-tained in comparing two antivenoms made of Fab and F(ab')2, re-spectively, in the neutralization of hemorrhage induced by Vipera berus venom [161]. Evidently, more studies with different venom-antivenom systems are required to address the question of whether a lower molecular mass of the neutralizing molecules effectively translates into a more efficient performance of these antidotes. Nevertheless, the rapid action of toxins at the site of injection, and the concomitant disruption of the microvasculature by hemorrhagic SVMPs, make it extremely difficult for intravenously-administered antivenoms, regardless of the molecular mass of their active princi-ples, to reach envenomated tissues on time to achieve a significant neutralization of tissue-damaging toxins. An alternative approach is
to administer low molecular mass antibody fragments, of a higher diffusion rate than IgG or F(ab’)2, at the site of venom injection [162], a possibility that requires further experimental assessment.
The Use of Enzyme Inhibitors as an Alternative to Confront Venom-Induced Local Tissue Damage
Studies in biochemistry and experimental pathology indicate that local tissue damage in snakebite envenomation is promoted, essentially, by the action of PLA2s and SVMPs [96, 114], with the contribution of hyaluronidases [163]. In spite to their antigenic diversity, evidenced by the failure of some antivenoms to neutralize these hydrolases in a number of heterologous venoms, the structural and functional features of the catalytic centers in each of these en-zyme types are highly conserved [97, 98], thus opening the possi-bility that enzyme inhibitors might have a wide spectrum of inhibi-tion against toxins from different snake taxa. This possibility is further strengthened by the observation that venom hydrolases are homologous to mammalian endogenous PLA2s and metalloprotein-ases (MMPs and ADAMs), which are involved in many kinds of diseases and for which pharmaceutical research has developed a large arsenal of inhibitory molecules. Thus, an excellent window of opportunity exists by searching for enzyme inhibitors that could halt the progress of local tissue damage upon application at the site of venom injection. This possibility has received solid experimental support in the case of inhibitors of metalloproteinases [164, 165], PLA2s [166] and hyaluronidases [163].
Metalloproteinase Inhibitors
The search for MMP inhibitors has been stimulated by the ob-servation that these endogenous metalloproteinases play a relevant role in local tumor growth, angiogenesis and metastasis [167, 168]. A large number of synthetic MMP inhibitors have been designed, most of which are based on a peptidomimetic moiety, simulating the sequence of the substrate that is cleaved by MMPs, linked to a chelating ligand, which can be thiols, carboxyl alkyls, phosphonic acids, phosphonamides and, predominantly, hydroxamate groups (see [169] for a review). In addition, a number of tetracyclines and chemically-modified tetracyclines are also inhibitors of MMPs [170]. The design of MMP inhibitors has been supported by crystal-
Fig. (3). Schematic view of the pathogenesis of local tissue damage in snakebite envenomation after intramuscular venom injection. Myotoxic phospholipases A2 (PLA2s), hemorrhagic and non-hemorrhagic metalloproteinases (SVMPs), hyaluronidases and other venom components cause direct tissue damage, i.e. myonecrosis, hemorrhage and extracellular matrix degradation. Simultaneously, a complex and pronounced inflammatory response ensues, secondary to tissue damage and also due to the activation of resident tissue cells (fibroblasts, macrophages, endothelial cells) by subcytotoxic concentrations of venom compo-nents and by the action of tissue-derived factors. Such inflammatory response, which causes edema and pain, is also characterized by the recruitment of an abundant cellular infiltrate composed predominantly of neutrophils and macrophages. This complex inflammatory response may promote further tissue dam-age and, at the same time, may contribute to the processes of tissue repair and regeneration.

2942 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
lographic studies and by computer-aided docking analysis of inter-actions between MMPs and inhibitors [171]. The first generation of MMP peptidomimetic hydroxamate inhibitors, such as batimastat and marimastat had a general inhibitory spectrum against MMPs [169]. Subsequent generations of inhibitors were designed to be more specific for some MMPs, based on structural differences in MMPs, especially at the deep S1’ hydrophobic pocket [169].
Clinical trials of MMP inhibitors as anticancer agents have been disappointing for several reasons [169, 172]. However, such syn-thetic compounds are highly effective in the inhibition of several SVMPs [164, 173]. In contrast to the complex processes that medi-ate tumor growth and metastasis, the use of MMP inhibitors in snakebite envenomation should be more simple, since the task is to rapidly block SVMPs in the tissues upon local injection of the in-hibitors. Thus, the pharmacokinetic concerns that have affected the progression of some MMP inhibitors might not be so relevant for SVMP inhibition. Furthermore, the musculoskeletal pain and in-flammation that develop as adverse effects of MMP inhibitor ad-ministration in clinical trials [174] depend on a repeated administra-tion schedule, whereas inhibitors in snakebite envenomation should be administered only once, rapidly after envenomation. Finally, some of the first generation MMP inhibitors had the problem of being too general, thus having a low specificity for certain MMPs [169]. However, in the case of snakebite envenomation, such lack of specificity may constitute an advantage, since such inhibitors would be effective against various SVMPs which may present slight variations in the structure of their catalytic site, particularly in the depth of the S1’ hydrophobic pocket [99, 100].
Therefore, even the MMP inhibitors that have failed as antican-cer agents may be excellent candidates against SVMPs. A good example is the peptidomimetic hydroxamate inhibitor ‘batimastat’, which was capable not only of abrogating the hemorrhagic activity of the SVMP BaP1, from the venom of B. asper [164], but also that of the rest of hemorrhagic SVMPs present in this venom [165]. Importantly, such inhibition was achieved not only when batimastat and SVMPs were incubated prior to injection, but also when the inhibitor was administered after envenomation [164, 165]. The structural interaction of batimastat and other inhibitors with a SVMP from the venom of Crotalus atrox has been studied [175, 176]. The case of batimastat clearly illustrates that MMP inhibitors are likely to be effective in the inhibition of SVMPs, and urges a systematic exploration of the potential of MMP inhibitors devel-oped by the pharmaceutical industry to block the action of venom enzymes at the clinical level. On the other hand, the expanding knowledge on the structures of SVMPs opens the possibility of computer-aided design of inhibitors of SVMPs.
Peptidomimetic inhibitors can also be designed on the basis of the structure of small peptides present in the venoms of viperid snakes which are inhibitors of SVMPs in the venom gland [177-179]. These peptides have relatively low affinity for SVMPs, but nevertheless can be used as lead compounds in the design of inhibi-tors of high affinity [173]. In addition, plants constitute a rich source of SVMP inhibitors, although most of the studies describing inhibitory effects against hemorrhagic activity of snake venoms have been performed with crude extracts [180]. Another source of natural inhibitors of SVMPs is the blood plasma of various mam-mals, as well as of venomous and non-venomous snakes [181]. These belong to the immunoglobulin superfamily, the ficolin/ opsonin P35 lectin family, and the cystatin superfamily [181]. The mechanism of inhibition of some of these proteins involves the formation of non-covalent complexes with SVMPs [182]. The iden-tification of the molecular region(s) involved in enzyme inhibition may pave the way for the design of synthetic or recombinant inhibi-tory peptides of low molecular mass, high diffusion in the tissues and high affinity for SVMPs.
Experimental studies strongly support the hypothesis that local administration, at the site of venom injection, of high-affinity SVMP inhibitors may become a powerful therapeutic approach to
abrogate or significantly reduce local tissue damage induced by SVMPs, i.e. hemorrhage, blistering, dermonecrosis, and myonecro-sis [164, 165]. Local inhibition of SVMPs may also halt their effect in the spreading of the venom, thus decreasing the extent of sys-temic alterations [127]. In addition, it is likely that inhibition of SVMPs at the site of injection may greatly contribute to the reduc-tion of systemic effects in a more direct way, since SVMPs play a protagonic role in the pathogenesis of systemic hemorrhage, hemo-dynamic disturbances and coagulopathy [183-185].
Phospholipase A2 Inhibitors
PLA2 comprises a growing and complex family of enzymes classified in at least 19 groups [186] which play diverse physiologi-cal and pathological roles [187]. They have received special atten-tion from the standpoint of inhibitor design once their relevance in inflammation was disclosed, based on the release of arachidonic acid, the precursor for the biosynthesis of eicosanoids [188]. As in the case of SVMPs, venom PLA2s have strong structural similari-ties with some mammalian secretory PLA2s, especially regarding the catalytic site and the calcium-binding loop [189]. Therefore, the efforts carried out in the search for synthetic and natural inhibitors to mammalian PLA2s, especially of those of group IIA involved in inflammatory and other pathophysiological conditions [190-192], may be of great benefit in finding potent inhibitors of venom PLA2s. However, in contrast with SVMPs, which largely depend on their catalytic activity to induce tissue damage [96], structure-function studies on venom PLA2s have demonstrated that toxic activities are often not related to enzymatic phospholipid hydroly-sis, but instead to ‘pharmacological regions’ in the surface of these molecules which allow them to interact with targets in cells [193, 194] (Fig. 4). Thus, inhibiton of catalytic activity does not necessar-ily imply inhibition of toxicity in many PLA2s [195, 196]. Moreo-ver, there is a growing group of PLA2 homologues having a Lys substituting the usually conservative Asp at position 49 [121]. This substitution, together with others at the calcium-binding loop, abol-ishes catalytic activity, whereas the toxins exert a variety of phar-macological effects, including myotoxicity [121]. This implies that the search for inhibitors against venom PLA2s must be performed by assessing not only the inhibition of enzymatic activity, but most importantly the inhibition of toxic effects.
Natural products represent rich sources of PLA2 inhibitors of different sorts. Inhibitors of venom PLA2s isolated from plants include proteins [197], flavonoids [180, 198], a coumestan known as wedelolactone [199], pterocarpans [200], the nitro compound aristolochic acid [201], other phenolic compounds such as rosma-rinic acid [202] and tea melanin [203], terpenoids such as 4-nerolidylcatechol [204] and manoalide [205], and polysaccharides such as fucoidan [206]. In addition, the glycosaminoglycan heparin, and small fragments derived from heparin, inhibit myotoxic PLA2s from Bothrops sp venoms [207, 208]. The heparin binding site of a Lys49 PLA2 homologue from B. asper venom was mapped to a stretch of basic residues located at the C-terminus [207]. Potent PLA2 inhibitors are also present in the plasma of various animals, most notably snakes [209, 210]. Snake PLA2 inhibitors are classified within three groups: alpha-inhibitors, belonging to the C-type lectin family; beta-inhibitors, having homology with proteins characterized by a leucine-rich repeat; and gamma-inhibitors, pre-senting the three-finger motif characteristic of diverse proteins of the Ly-6 superfamily, urokinase-type plasminogen activator recep-tor and snake venom -neurotoxins [210, 211]. These proteins vary in their inhibitory spectrum towards different types of PLA2s [210]. The structural determinant responsible for PLA2 inhibition in a gamma-type receptor from the serum of Phyton reticulatus was identified [212]. In addition, a potent myotoxin inhibitor, belonging to the immunoglobulin superfamily, was isolated from the serum of the opossum Didelphis marsupialis [213]. It neutralizes myotoxic-ity, but not enzymatic activity, of B. asper PLA2s [213]. Despite this wide spectrum of venom PLA2 inhibitors, few studies have
Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2943
addressed neutralization in experiments involving the administra-tion of the inhibitors after venom has been injected. Therefore, it is necessary to further explore natural products for PLA2 inhibitors of high affinity and potential to halt the toxic action of PLA2s after venom injection.
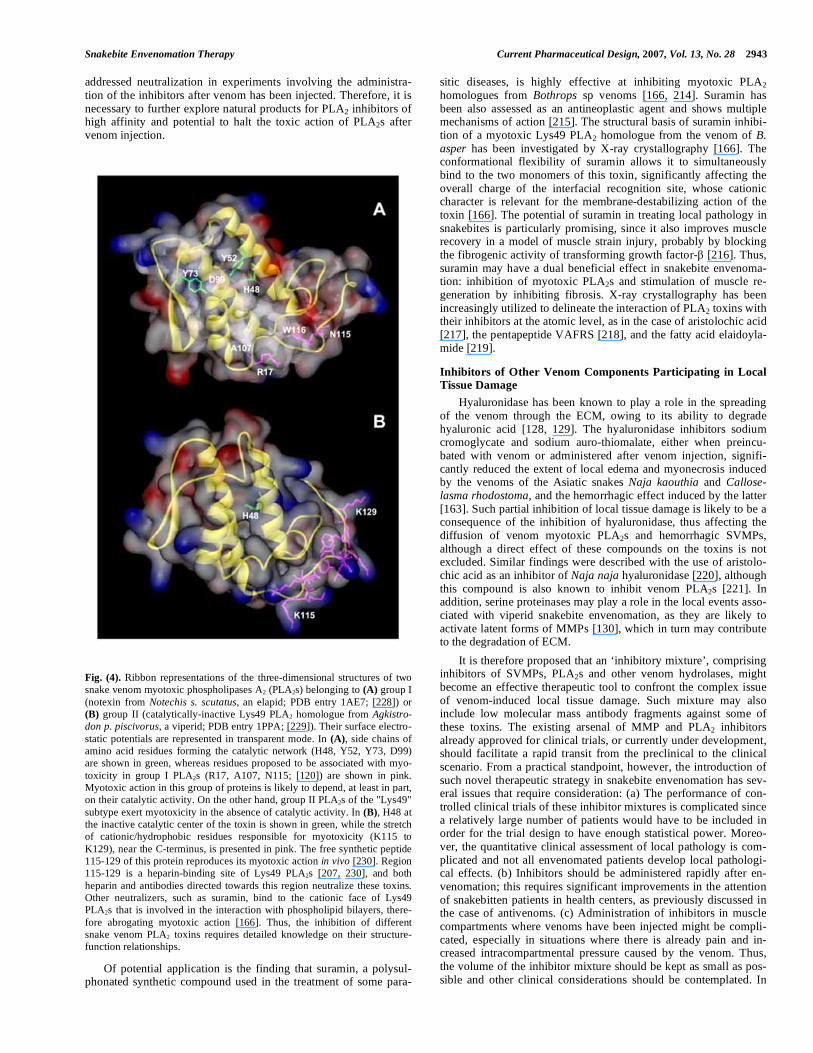
Fig. (4). Ribbon representations of the three-dimensional structures of two snake venom myotoxic phospholipases A2 (PLA2s) belonging to (A) group I (notexin from Notechis s. scutatus, an elapid; PDB entry 1AE7; [228]) or (B) group II (catalytically-inactive Lys49 PLA2 homologue from Agkistro-
don p. piscivorus, a viperid; PDB entry 1PPA; [229]). Their surface electro-static potentials are represented in transparent mode. In (A), side chains of amino acid residues forming the catalytic network (H48, Y52, Y73, D99) are shown in green, whereas residues proposed to be associated with myo-toxicity in group I PLA2s (R17, A107, N115; [120]) are shown in pink. Myotoxic action in this group of proteins is likely to depend, at least in part, on their catalytic activity. On the other hand, group II PLA2s of the "Lys49" subtype exert myotoxicity in the absence of catalytic activity. In (B), H48 at the inactive catalytic center of the toxin is shown in green, while the stretch of cationic/hydrophobic residues responsible for myotoxicity (K115 to K129), near the C-terminus, is presented in pink. The free synthetic peptide 115-129 of this protein reproduces its myotoxic action in vivo [230]. Region 115-129 is a heparin-binding site of Lys49 PLA2s [207, 230], and both heparin and antibodies directed towards this region neutralize these toxins. Other neutralizers, such as suramin, bind to the cationic face of Lys49 PLA2s that is involved in the interaction with phospholipid bilayers, there-fore abrogating myotoxic action [166]. Thus, the inhibition of different snake venom PLA2 toxins requires detailed knowledge on their structure-function relationships.
Of potential application is the finding that suramin, a polysul-phonated synthetic compound used in the treatment of some para-
sitic diseases, is highly effective at inhibiting myotoxic PLA2 homologues from Bothrops sp venoms [166, 214]. Suramin has been also assessed as an antineoplastic agent and shows multiple mechanisms of action [215]. The structural basis of suramin inhibi-tion of a myotoxic Lys49 PLA2 homologue from the venom of B. asper has been investigated by X-ray crystallography [166]. The conformational flexibility of suramin allows it to simultaneously bind to the two monomers of this toxin, significantly affecting the overall charge of the interfacial recognition site, whose cationic character is relevant for the membrane-destabilizing action of the toxin [166]. The potential of suramin in treating local pathology in snakebites is particularly promising, since it also improves muscle recovery in a model of muscle strain injury, probably by blocking the fibrogenic activity of transforming growth factor- [216]. Thus, suramin may have a dual beneficial effect in snakebite envenoma-tion: inhibition of myotoxic PLA2s and stimulation of muscle re-generation by inhibiting fibrosis. X-ray crystallography has been increasingly utilized to delineate the interaction of PLA2 toxins with their inhibitors at the atomic level, as in the case of aristolochic acid [217], the pentapeptide VAFRS [218], and the fatty acid elaidoyla-mide [219].
Inhibitors of Other Venom Components Participating in Local Tissue Damage
Hyaluronidase has been known to play a role in the spreading of the venom through the ECM, owing to its ability to degrade hyaluronic acid [128, 129]. The hyaluronidase inhibitors sodium cromoglycate and sodium auro-thiomalate, either when preincu-bated with venom or administered after venom injection, signifi-cantly reduced the extent of local edema and myonecrosis induced by the venoms of the Asiatic snakes Naja kaouthia and Callose-lasma rhodostoma, and the hemorrhagic effect induced by the latter [163]. Such partial inhibition of local tissue damage is likely to be a consequence of the inhibition of hyaluronidase, thus affecting the diffusion of venom myotoxic PLA2s and hemorrhagic SVMPs, although a direct effect of these compounds on the toxins is not excluded. Similar findings were described with the use of aristolo-chic acid as an inhibitor of Naja naja hyaluronidase [220], although this compound is also known to inhibit venom PLA2s [221]. In addition, serine proteinases may play a role in the local events asso-ciated with viperid snakebite envenomation, as they are likely to activate latent forms of MMPs [130], which in turn may contribute to the degradation of ECM.
It is therefore proposed that an ‘inhibitory mixture’, comprising inhibitors of SVMPs, PLA2s and other venom hydrolases, might become an effective therapeutic tool to confront the complex issue of venom-induced local tissue damage. Such mixture may also include low molecular mass antibody fragments against some of these toxins. The existing arsenal of MMP and PLA2 inhibitors already approved for clinical trials, or currently under development, should facilitate a rapid transit from the preclinical to the clinical scenario. From a practical standpoint, however, the introduction of such novel therapeutic strategy in snakebite envenomation has sev-eral issues that require consideration: (a) The performance of con-trolled clinical trials of these inhibitor mixtures is complicated since a relatively large number of patients would have to be included in order for the trial design to have enough statistical power. Moreo-ver, the quantitative clinical assessment of local pathology is com-plicated and not all envenomated patients develop local pathologi-cal effects. (b) Inhibitors should be administered rapidly after en-venomation; this requires significant improvements in the attention of snakebitten patients in health centers, as previously discussed in the case of antivenoms. (c) Administration of inhibitors in muscle compartments where venoms have been injected might be compli-cated, especially in situations where there is already pain and in-creased intracompartmental pressure caused by the venom. Thus, the volume of the inhibitor mixture should be kept as small as pos-sible and other clinical considerations should be contemplated. In

2944 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
spite of these potential difficulties, the idea of administering a mix-ture of potent inhibitors of SVMPs, PLA2s and other venom com-ponents in situ, rapidly after the event of a snakebite, probably con-stitutes the most promising innovation in the management of these envenomations in many decades.
THE SEARCH FOR DRUGS TO COUNTERACT SYSTEMI-
CALLY-ACTING TOXINS
As discussed above, antivenoms are highly effective in the neu-tralization of snake venom toxins responsible for the systemic ef-fects [5]. Nevertheless, the search for novel inhibitory compounds to counteract systemically-acting toxins may improve the therapy of these envenomations, overcoming the problem of lack of immu-nological specificity of some antivenoms against heterologous ven-oms. Since SVMPs play a central role in hemorrhagic, defibrino-genating and hemodynamic effects induced by viperid envenoma-tions, inhibitors of SVMPs have a great potential to counteract these effects [see the case of B. asper [183]. Moreover, serine proteinase inhibitors may contribute to control the hemostatic manifestations secondary to the action of venom thrombin-like serine proteinases. In addition, the search for novel inhibitors of venom PLA2s may become of high relevance in confronting the potent PLA2s that cause systemic neurotoxicity and myotoxicity in envenomations by many elapid and some viperid venoms. In the case of elapid neuro-toxic venoms containing post-synaptically acting neurotoxins, ex-perimental and clinical studies have demonstrated the usefulness of the acetylcholinesterase inhibitor neostigmine, administered to-gether with atropine, at reducing the paralytic manifestations char-acteristic of these envenomations [6, 222]. Neostigmine, however, is not beneficial in envenomations in which neurotoxicity depends on pre-synaptically acting neurotoxic PLA2s.
The search for novel therapeutic agents that would complement antivenoms in the treatment of snakebite envenomations should be closely linked with the efforts to gain a deeper understanding on the pathophysiology of envenomations. For instance, the role of in-flammatory mediators, such as cytokines, complement and nitric oxide, in the systemic hemodynamic disturbances characteristic of severe viperid envenomations, has not been thoroughly investi-gated, although clinical and experimental studies document an in-crement in the systemic levels of some of these mediators [223-
225]. Antagonism of TNF- reduces the hemodynamic manifesta-tions and the cardiac toxicity induced by the venom of Vipera aspis in a rat model [226, 227]. Undoubtedly, as we advance in our un-derstanding of the pathophysiology of snakebite envenomation, novel therapeutic alternatives will be developed on the basis of this knowledge.
CONCLUDING REMARKS
Animal-derived antivenoms are likely to remain as the central element in the treatment of snakebite envenomation for many years to come, and efforts should be carried out to improve their efficacy and safety, and to promote public health interventions aimed at a widespread deployment and distribution of antivenoms, at afford-able prices, to the regions where they are mostly needed. Such im-provement of antivenoms has to be closely associated with a more complete understanding of the biochemical and toxicological com-plexity of the medically most relevant snake venoms on a world-wide basis. In addition, there is a need for a systematic effort to test inhibitors of SVMPs, PLA2s, hyaluronidases, serine proteinases and other relevant venom components for their potential to abrogate local effects induced by many snake venoms, as well as some of the systemic manifestations of envenomations. The future scenario in the management of snakebite envenomations is foreseen as involv-ing a rapid in situ administration of a mixture of enzyme and toxin inhibitors, followed by an intravenous infusion of effective and safe antivenoms, and the modulation, through a variety of pharmacol-ogical interventions, of the most relevant endogenous mediators that might contribute to the local and systemic manifestations of envenomation. Such interventions should be complemented by various ancillary therapies aimed at controlling the complications that develop as a consequence of envenomation (Fig. 5). There are plenty of scientific, technological and public health challenges in the road to accomplish such scenario.
ACKNOWLEDGEMENTS
Many observations discussed in this review correspond to re-search projects supported by Universidad de Costa Rica, the Inter-national Foundation for Science (IFS), and NeTropica. The collabo-ration of many students and colleagues throughout many years is greatly appreciated.
Fig. (5). Summarized view of the possible future scenario of the therapy of viperid snakebite envenomations that cause local and systemic effects. Continuous arrows represent the direct and indirect effects induced by envenomation, and discontinuous arrows represent the therapeutic actions. The first intervention is an in situ injection of a mixture of inhibitors of venom toxins, which may also include low molecular mass antibody fragments against relevant venom compo-nents. Such intervention is aimed at a rapid inhibition of toxins in the tissue, thus preventing local tissue damage and reducing further systemic pathophysi-ological effects. Then, after transportation of the patient to a health center, intravenous administration of antivenom accomplishes the neutralization of toxins reaching the bloodstream and causing systemic effects, and favors the redistribution of toxins present in the tissues to the bloodstream. In addition, the phar-macological modulation of the inflammatory response to envenomation may prevent further tissue damage while supporting processes involved in tissue repair and regeneration. Finally, ancillary interventions of various sorts directed at controlling some of the complications of envenomation complete this scenario.

Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2945
REFERENCES
[1] Swaroop S, Grab B. Snakebite mortality in the world. Bull WHO 1954; 10: 35-76.
[2] Chippaux JP. Snake-bites: appraisal of the global situation. Bull WHO 1998; 76: 515-24.
[3] Warrell DA, Arnett C. The importance of bites by the saw-scaled or carpet viper (Echis carinatus). Epidemiological studies in Nige-ria and a review of the world literature. Acta Trop 1976; 33: 307-41.
[4] Sharma SK, Chappuis F, Jha N, Bovier PA, Loutan L, Koirala S. Impact of snake bites and determinants of fatal outcomes in south-eastern Nepal. Am J Trop Med Hyg 2004; 71: 234-8.
[5] Warrell DA. In: Gopalakrishnakone P, Tan CK Eds. Recent Ad-vances in Toxinology Research. Singapore, National University of Singapore 1992; 121-53.
[6] Warrell DA. In: Meier J, White J Eds. Handbook of Clinical Toxi-cology of Animal Venoms and Poisons. Florida, CRC Press 1995; 493-594.
[7] Warrell DA. In: Meier J, White J Eds. Handbook of Clinical Toxi-cology of Animal Venoms and Poisons. Florida, CRC Press 1995; 433-92.
[8] Fan HW, Cardoso JL. Clinical toxicology of snake bites in South America. In: Meier J, White J Eds. Handbook of Clinical Toxicol-ogy of Animal Venoms and Poisons. Florida, CRC Press 1995; 667-88.
[9] Gutiérrez JM. In: Meier J, White J Eds. Handbook of Clinical Toxicology of Animal Venoms and Poisons. Florida, CRC Press 1995; 645-65.
[10] Warrell DA. In: Bon C, Goyffon M Eds. Envenomings and Their Treatments. Lyon, Fondation Marcel Mérieux 1996; 63-76.
[11] Azevedo-Marques MM, Cupo P, Coimbra TM, Hering SE, Rossi MA, Laure CJ. Myonecrosis, myoglobinuria and acute renal failure induced by South American rattlesnake (Crotalus durissus terrifi-
cus) envenomation in Brazil. Toxicon 1985; 23: 631-6. [12] White J. In: Meier J, White J Eds. Handbook of Clinical Toxicol-
ogy of Animal Venoms and Poisons. Florida, CRC Press 1995; 595-617.
[13] Bon C. In: Bon C, Goyffon M Eds. Envenomings and Their Treat-ments. Lyon, Fondation Marcel Mérieux 1996; 3-9.
[14] Vital Brazil O. History of the primordia of snake-bite accident serotherapy. Mem Inst Butantan 1987; 49: 7-20.
[15] Meier J. In: Meier J, White J Eds. Handbook of Clinical Toxicol-ogy of Animal Venoms and Poisons. Florida, CRC Press 1995; 689-721.
[16] Lalloo D, Theakston RDG. Snake antivenoms. J Toxicol Clin Toxicol 2003; 41: 277-90.
[17] Raw I, Guidolin R, Higashi HG, Kelen EMA. In: Tu AT Ed. Hand-book of Natural Toxins, Vol 5, Reptile Venoms and Toxins. New York, Marcel Dekker, Inc 1991; 557-81.
[18] Pope CG. The action of proteolytic enzymes on the antitoxins and proteins in immune sera. II. Heat denaturation after partial enzyme action. Br J Exp Pathol 1939; 20: 201-12.
[19] Grandgeorge M, Véron JL, Lutsch C, Makula MF, Riffard P, Pépin S, et al. In: Bon C, Goyffon M Eds. Envenomings and Their Treat-ments. Lyon, Fondation Marcel Mérieux 1996; 161-72.
[20] Jones RG, Landon J. A protocol for ‘enhanced pepsin digestion’: a step by step method for obtaining pure antibody fragments in high yield from serum. J Immunol Methods 2003; 275: 239-50.
[21] Smith DC, Reddi KR, Laing G, Theakston RG, Landon J. An affin-ity purified ovine antivenom for he treatment of Vipera berus en-venoming. Toxicon 1992; 30: 865-71.
[22] dos Santos MC, D’Imperio Lima MR, Furtado GC, Colletto GMDD, Kipnis TL, Dias da Silva W. Purification of F(ab’)2 anti-snake venom by caprylic acid: a fast method for obtaining IgG fragments with large neutralization activity, purity and yield. Toxi-con 1989; 27: 297-303.
[23] Rojas G, Jiménez JM, Gutiérrez JM. Caprylic acid fractionation of hyperimmune horse plasma: description of a simple procedure for antivenom production. Toxicon 1994; 32: 351-63.
[24] Cardoso JLC, Fan HW, Franca FOS, Jorge MT, Leite RP, Nishioka SA, et al. Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bothrops jara-
raca) in Sao Paulo, Brazil. Q J Med 1993; 86: 315-25.
[25] Otero R, Gutiérrez JM, Rojas G, Núñez V, Díaz A, Miranda E, et
al. A randomized blinded clinical trial of two antivenoms, prepared by caprylic acid or ammonium sulphate fractionation of IgG, in Bothrops and Porthidium snake bites in Colombia. Correlation be-tween safety and biochemical characteristics of antivenoms. Toxi-con 1999; 37: 895-908.
[26] Smalligan R, Cole J, Brito N, Laing GD, Mertz BL, Manock S, et
al. Crotaline snake bite in the Ecuadorian Amazon: randomised double blind comparative trial of three South American poly-specific antivenoms. BMJ 2004; 329: 1129-35.
[27] Ménez A. In: Harvey AL Ed. Snake Toxins. New York, Pergamon Press 1991; 35-90.
[28] Gutiérrez JM, León G, Lomonte B. Pharmacokinetic-pharmaco- dynamic relationships of immunoglobulin therapy for envenoma-tion. Clin Pharmacokinet 2003; 42: 721-41.
[29] Gutiérrez JM, León G, Rojas G, Lomonte B, Rucavado A, Chaves F. Neutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon 1998; 36: 1529-39.
[30] Gutiérrez JM, Ownby CL, Odell GV. Pathogenesis of myonecrosis induced by crude venom and a myotoxin of Bothrops asper. Exp Mol Pathol 1984; 40: 367-79.
[31] Moreira L, Gutiérrez JM, Borkow G, Ovadia M. Ultrastructural alterations in mouse capillary blood vessels after experimental in-jection of venom from the snake Bothrops asper (terciopelo). Exp Mol Pathol 1992; 57: 124-33.
[32] Lomonte B, Lundgren J, Johansson B, Bagge U. The dynamics of local tissue damage induced by Bothrops asper snake venom and myotoxin II on the mouse cremaster muscle: an intravital and elec-tron microscopic study. Toxicon 1994; 32: 41-55.
[33] Battellino C, Piazza R, da Silva AMM, Cury Y, Farsky SHP. As-sessment of efficacy on bothropic antivenom therapy on microcir-culatory effects induced by Bothrops jararaca snake venom. Toxi-con 2003; 41: 583-93.
[34] Otero R, Gutiérrez JM, Núñez V, Robles A, Estrada R, Segura E, et
al. Moreno ME & the regional group on antivenom therapy re-search. A randomized double-blind clinical trial of two antivenoms in patients bitten by Bothrops atrox in Colombia. Trans Royal Soc Trop Med Hyg 1996; 90: 696-700.
[35] Trebien HA, Calixto JB. Pharmacological evaluation of rat paw oedema induced by Bothrops jararaca venom. Agents Actions 1989; 26: 292-300.
[36] Chaves F, Barboza M, Gutiérrez JM. Pharmacological study of edema induced by venom of the snake Bothrops asper (terciopelo). Toxicon 1995; 33: 31-9.
[37] Prasarnpun S, Walsh J, Awad SS, Harris JB. Envenoming bites by kraits: the biological basis of treatment-resistant neuromuscular pa-ralysis. Brain 2005; 128: 2987-2996.
[38] Ismail M, Abd-Elsalam MA, Al-Ahaidib MS. Pharmacokinetics of 125I-labelled Walterinnesia aegyptia venom and its specific antiven-ins: flash absorption and distribution of the venom and its toxin versus slow absorption and distribution of IgG, F(ab’)2 and Fab of the antivenin. Toxicon 1998; 36: 93-114.
[39] Scherrmann JM. Antibody treatment of toxin poisoning-Recent advances. Clin Toxicol 1994; 32: 363-75.
[40] Riviere G, Choumet V, Audebert F, Sabouraud A, Debray M, Scherrmann JM, et al. Effect of antivenom on venom pharmacoki-netics in experimentally envenomed rabbits: toward an optimiza-tion of antivenom therapy. J Pharmacol Exp Ther 1997; 281: 1-8.
[41] Bon C. Pharmacokinetics of venom toxins and their modification by antivenom therapy. J Toxicol Toxin Rev 2003; 22: 129-38.
[42] Chippaux JP, Williams V, White J. Snake venom variability: meth-ods of study, results and interpretation. Toxicon 1991; 29: 1279-303.
[43] Gutiérrez JM, Rojas G, Bogarín G, Lomonte B. In: Bon C, Goyffon M Eds. Envenomings and Their Treatments. Lyon, Fondation Mar-cel Mérieux 1996; 223-31.
[44] Otero R, Núñez V, Osorio RG, Gutiérrez JM, Giraldo CA, Posada LE. Ability of six Latin American antivenoms to neutralize the venom of mapaná equis (Bothrops atrox) from Antioquia and Chocó (Colombia). Toxicon 1995; 33: 809-15.
[45] Bogarín G, Morais JF, Yamaguchi IK, Stephano MA, Marcelino JR, Nishikawa AK, et al. Neutralization of crotaline snake venoms from Central and South America by antivenoms produced in Brazil and Costa Rica. Toxicon 2000; 38: 1429-41.

2946 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
[46] Theakston RDG, Laing GD, Fielding CM, Freire Lascano A, Touzet JM, Vallejo F, et al. Treatment of snake bites by Bothrops species and Lachesis muta in Ecuador: laboratory screening of can-didate antivenoms. Trans R Soc Trop Med Hyg 1995; 89: 550-4.
[47] Saravia P, Rojas E, Arce V, Guevara C, López JC, Chaves E, et al. Geographic and ontogenic variability in the venom of the neotropi-cal rattlesnake Crotalus durissus: Pathophysiological and therapeu-tic implications. Rev Biol Trop 2002; 50: 337-46.
[48] Theakston RDG. In: Bon C, Goyffon M Eds. Envenomings and Their Treatments. Lyon, Fondation Marcel Mérieux 1996; 117-26.
[49] Fan HW. In: Cardoso JLC, Franca FOS, Fan HW, Málaque CMS, Haddad V. Animais Peconhentos no Brasil. Biologia, Clínica e Terapeutica. Sao Paulo, Sarvier 2003; 380-93.
[50] Chippaux JP. Short report: Treatment of snake envenomations by a new polyvalent antivenom composed of highly purified F(ab’)2: Results of a clinical trial in northern Cameroon. Am J Trop Med Hyg 1999; 61: 1017-18.
[51] Moran NF, Newman WJ, Theakston RDG, Warrell DA, Wilkinson D. High incidence of early anaphylactoid reactions to SAIMR polyvalent snake antivenom. Trans R Soc Trop Med Hyg 1998; 92: 69-70.
[52] Gawarammana IB, Kularatne SA, Dissanayabe WP, Kumarasiri RP, Senanayake N, Ariyasena H. Parallel infusion of hydrocorti-sone +/- chlorpheniramine bolus injection to prevent acute adverse reactions to antivenom for snakebites. Med J Aust 2004; 180: 20-3.
[53] Jurkovich GJ, Luterman A, McCullar K, Ramenofsky ML, Curreri PW. Complications of Crotalidae antivenin therapy. J Trauma 1988; 28: 1032-7.
[54] Fan HW, Marcopito LF, Cardoso JL, Franca FO, Malaque CM, Ferrari RA, et al. Sequential randomised and double blind trial of promethazine prophylaxis against early anaphylactic reactions to antivenom for Bothrops snakebites. BMJ 1999; 318: 1451-2.
[55] Premawardhena AP, de Silva CE, Fonseka MMD, Gunatilake SB, de Silva HJ. Low dose subcutaneous adrenaline to prevent acute adverse reactions to antivenom serum in people bitten by snakes: randomized, placebo controlled trial. BMJ 1999; 318: 1041-3.
[56] Sutherland SK. Serum reactions. An analysis of commercial an-tivenoms and the possible role of anticomplementary. Med J Aust 1977; 1: 613-15.
[57] Malasit P, Warrell DA, Chantavanich P, Viravan C, Mongkilsapaya J, Singhthong B, et al. Prediction, prevention and mechanism of early (anahylactic) antivenom ractions in victims of snake bites. BMJ 1986; 292: 17-20.
[58] Montero J, Trejos M, Lomonte B. Efecto del suero antiofídico sobre la actividad hemolítica del complemento humano (in vitro) y de conejo (in vitro e in vivo). Rev Costarric Cienc Méd 1989; 10: 1-10.
[59] Morais JF, de Freitas MCW, Yamaguchi IK, dos Santos MC, Dias da Silva W. Snake antivenoms from hyperimmunized horses: com-parison of the antivenom activity and biological properties of their whole IgG and F(ab’)2 fragments. Toxicon 1994; 32: 725-34.
[60] León G, Monge M, Rojas E, Lomonte B, Gutiérrez JM. Compari-son between IgG and F(ab’)2 polyvalent antivenoms: neutralization of systemic effects induced by Bothrops asper venom in mice, ex-travasation to muscle tissue, and potential of adverse reactions. Toxicon 2001; 39: 793-801.
[61] León G, Lomonte B, Gutiérrez JM. Anticomplementary activity of equine whole IgG antivenoms: comparison of three fractionation protocols. Toxicon 2005; 45: 123-8.
[62] Otero R, León G, Gutiérrez JM, Rojas G, Toro MF, Barona J, et al. Efficacy and safety of two whole IgG polyvalent antivenoms, re-fined by caprylic acid fractionation with or without -propiolactone, in the treatment of Bothrops asper bites in Colom-bia. Trans R Soc Trop Med Hyg 2006; 100: 1173-82.
[63] Chippaux JP. The development and use of immunotherapy in Af-rica. Toxicon 1998; 36: 1503-6.
[64] Otero-Patiño R, Cardoso JLC, Higashi HG, Núñez V, Diaz A, Toro MF, et al. A randomized, blinded, comparative trial of one pepsin-digested and two whole IgG antivenoms for Bothrops snake bites in Urabá, Colombia. Am J Trop Med Hyg 1998; 58: 183-9.
[65] Wilde H, Thipkong P, Sitprija V, Chaiyabutr N. Heterologous antisera and antivenins are essential biologicals: perspectives on a worldwide crisis. Ann Intern Med 1996; 125: 233-6.
[66] Theakston RDG, Warrell DA. Crisis in snake antivenom supply for Africa. Lancet 2000; 356: 2104.
[67] Theakston RDG, Warrell DA, Griffiths E. Report of a WHO work-shop on the standardization and control of antivenoms. Toxicon 2003; 41: 541-57.
[68] Laing GD, Harrison RA, Theakston RDG, Renjifo JM, Nasidi A, Gutiérrez JM, et al. Polyspecific snake antivenom may help in an-tivenom crisis. BMJ 2003; 326: 447-8.
[69] Chippaux JP. In: Ménez A Ed, Perspectives in Molecular Toxinol-ogy. New York, John Wiley & Sons Co. 2002; 457-72.
[70] Looareesuwan S, Viravan C, Warrell DA. Factors contributing to fatal snake bite in the rural tropics: analysis of 46 cases in Thai-land. Trans R Soc Trop Med Hyg 1988; 82: 930-4.
[71] Gutiérrez JM, Theakston RDG, Warrell DA. Confronting the ne-glected problem of snake bite envenoming: the need for a global partnership. PLoS Med 2006; 3: 727-31.
[72] Serrano SMT, Shannon JD, Wand D, Camargo ACM, Fox JW. A multifaceted analysis of viperid snake venoms by two-dimensional gel electrophoresis: an approach to understanding venom pro-teomics. Proteomics 2005; 5: 501-10.
[73] Harrison RA. Development of venom toxin-specific antibodies by DNA immunization: rationale and strategies to improve therapy of viper envenoming. Vaccine 2004; 22: 1648-55.
[74] Harrison RA, Moura-da-Silva AM, Laing GD, Wu Y, Richards A, Broadhead A, et al. Antibody from mice immunized with DNA en-coding the carboxyl-disintegrin and cysteine-rich domain (JD9) of the hemorrhagic metalloproteinase, jararhagin, inhibits the main le-thal component of viper venom. Clin Exp Immunol 2000; 121: 358-63.
[75] Raweerith R, Ratanabanangkoon K. Fractionation of equine an-tivenom using caprylic acid precipitation in combination with cati-onic ion-exchange chromatography. J Immunol Methods 2003; 282: 63-72.
[76] Newcombe AR, Cresswell C, Davies S, Watson K, Harris G, O’Donovan K, et al. Optimised affinity purification of polyclonal antibodies from hyper immunized ovine serum using a synthetic Protein A adsorbent, MAbsorbentR A2P. J Chromatogr B 2005; 814: 209-15.
[77] Burnouf T, Griffiths E, Padilla A, Seddik S, Stephano MA, Gutiér-rez JM. Assessment of the viral safety of antivenoms fractionated from equine plasma. Biologicals 2004; 32: 115-28.
[78] Thalley BT, Carroll SB. Rattlesnake and scorpion antivenoms from the egg yolks of immunized hens. Biotechnology 1990; 8: 934-8.
[79] Meddeb-Mouelhi F, Bouhaouala-Zahar B, Benlasfar Z, Hammadi M, Mejri T, Moslah M, et al. Immunized camel sera and derived immunoglobulin subclasses neutralizing Androctonus australis
hector scorpion toxins. Toxicon 2003; 42: 785-91. [80] Harrison RA, Hasson SS, Harmsen M, Laing GD, Conrath K,
Theakston RDG. Neutralisation of venom-induced haemorrhage by IgG from camels and llamas immunized with viper venom and also by endogenous, non-IgG components in camelid sera. Toxicon 2006; 47: 364-8.
[81] Hammers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hamers C, Songa EB, et al. Naturally occurring antibodies de-void of light chains. Nature 1993; 363: 446-8.
[82] Herrera M, León G, Segura A, Meneses F, Lomonte B, Chippaux JP, et al. Factors associated with adverse reactions induced by ca-prylic acid-fractionated whole IgG preparations: comparison be-tween horse, sheep and camel IgGs. Toxicon 2005; 46: 775-81.
[83] Cardoso DF, Yamaguchi IK, Moura da Silva AM. In: Cardoso JLC, Franca FOS, Fan HW, Málaque CMS, Haddad V. Animais Pecon-hentos no Brasil. Biologia, Clínica e Terapeutica. Sao Paulo, Sarvier 2003; 367-79.
[84] Lafaye P, Choumet V, Demangel C, Bon C, Mazie JC. Biologically active human anticrotoxin scFv isolated from a semisynthetic phage library. Immunotechnology 1997; 3: 117-25.
[85] Vaughan TJ, Williams AJ, Pritchard K, Osbourn JK, Pope AR, Earnshaw JC, et al. Human antibodies with sub-nanomolar affini-ties isolated from a large non-immunized phage display library. Nat Biothechnol 1996; 14: 309-14.
[86] Lauwereys M, Ghahroudi MA, Desmyter A, Kinne J, Holzer W, De Genst E, et al. Potent enzyme inhibitors derived from drome-dary heavy-chain antibodies. EMBO J 1998; 17: 3512-20.
[87] Omidfar K, Rasaee MJ, Modjtahedi H, Forouzandeh M, Taghik-hani M, Goldmakani N. Production of a novel camel single-domain antibody specific for the type III mutant EGFR. Tumor Biol 2004; 25: 296-305.

Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2947
[88] Presta L. Antibody engineering for therapeutics. Curr Opin Struct Biol 2003; 13: 519-25.
[89] Meyer WP, Habib AG, Onayade AA, Yakubu A, Smith DC, Nasidi A, et al. First clinical experiences with a new ovine Fab Echis ocel-
latus snake bite antivenom in Nigeria: randomized comparative trial with Institute Pasteur Serum (Ipser) Africa antivenom. Am J Trop Med Hyg 1997; 56: 291-300.
[90] Dart RC, Seifert SA, Boyer LV, Clark RF, Hall E, McKinney P, et
al. A randomized multicenter trial of crotalinae polyvalent immune Fab (ovine) antivenom for the treatment for crotaline snakebite in the United States. Arch Intern Med 2001; 161: 2030-6.
[91] Theakston RDG, Reid HA. Development of simple standard assay procedures for the characterization of snake venoms. Bull WHO 1983; 61: 949-56.
[92] Gutiérrez JM, Rojas G, Lomonte B, Gené JA, Chaves F, Alvarado J, et al. Standardization of assays for testing the neutralizing ability of antivenoms. Toxicon 1990; 28: 1127-9.
[93] Attaran A, Sachs J. Defining and refining international donor sup-port for combating the AIDS pandemic. Lancet 2001; 357: 57-61.
[94] Sachs J. A new global commitment to disease control in Africa. Nat Med 2001; 7: 521-3.
[95] Gutiérrez JM, Lomonte B. In: Cardoso JLC, Franca FOS, Fan HW, Málaque CMS, Haddad V. Animais Peconhentos no Brasil. Biolo-gia, Clínica e Terapeutica. Sao Paulo, Sarvier 2003; 310-23.
[96] Gutiérrez J, Rucavado A. Snake venom metalloproteinases: their role in the pathogenesis of local tissue damage. Biochimie 2000; 82: 841-59.
[97] Bode W, Gomis-Ruth FX, Stockler W. Astacins, serralysins, snake venom and matrix metalloproteinases exhibit identical zinc-binding environments (HEXXHXXGXXH and Met-turn) and topologies an should be grouped into a common family, the ‘metzincins’. FEBS Lett 1993; 331: 134-40.
[98] Fox JW, Serrano SMT. Structural considerations of the snake venom metalloproteinases, key members of the M12 reprolysin family of metalloproteinases. Toxicon 2005; 45: 969-85.
[99] Gomis-Ruth FX, Kress LF, Kellerman J, Mayr I, Lee X, Huber R, et al. Refined 2 Å X-ray crystal structure of the snake venom zinc-endopeptidase adamalysin II. Primary and tertiary structure deter-mination, refinement, molecular structure and comparison with as-tacin, collagenase and thermolysin. J Mol Biol 1994; 239: 513-44.
[100] Watanabe L, Shannon JD, Valente RH, Rucavado A, Alape-Girón A, Kamiguti AS, et al. Amino acid sequence and crystal structure of BaP1, a metalloproteinase from Bothrops asper snake venom that exerts multiple tissue-damaging activities. Protein Sci 2003; 12: 2273-81.
[101] Bjarnason JB, Fox JW. Hemorrhagic metalloproteinases from snake venoms. Pharmacol Ther 1994; 62: 325-72.
[102] Kallury R. Basement membranes: structure, assembly and role in tumor angiogenesis. Nat Rev Cancer 2003; 3: 422-33.
[103] Hudson BG, Reeders ST, Tryggvason K. Type IV collagen: struc-ture, gene organization, and role in human diseases. Molecular ba-sis of Goodpasture and Alport syndromes and diffuse leiomyoma-tosis. J Biol Chem 1993; 268: 26033-6.
[104] Ownby CL, Bjarnason JB, Tu AT. Hemorrhagic toxins from rattle-snake (Crotalus atrox) venom. Pathogenesis of hemorrhage in-duced by three purified toxins. Am J Pathol 1978; 93: 201-18.
[105] Moreira L, Borkow G, Ovadia M, Gutiérrez JM. Pathological changes induced by BaH1, a hemorrhagic proteinase isolated from Bothrops asper (terciopelo) snake venom, on mouse capillary blood vessels. Toxicon 1994; 32: 977-87.
[106] Gutiérrez JM, Rucavado A, Escalante T, Díaz C. Hemorrhage induced by snake venom metalloproteinases: biochemical and bio-physical mechanisms involved in microvessel damage. Toxicon 2005; 45: 997-1011.
[107] Gutiérrez JM, Núñez J, Escalante T, Rucavado A. Blood flow is required for rapid endothelial cell damage induced by a snake venom hemorrhagic metalloproteinase. Microvasc Res 2006; 71: 55-63.
[108] Lomonte B, Gutiérrez JM, Borkow G, Ovadia M, Tarkowski A, Hanson LÅ. Activity of hemorrhagic metalloproteinase BaH-1 and myotoxin II from Bothrops asper snake venom on capillary endo-thelial cells in vitro. Toxicon 1994; 32: 505-10.
[109] Díaz C, Valverde L, Brenes O, Rucavado A, Gutiérrez JM. Charac-terization of events associated with apoptosis/anoikis induced by
snake venom metalloproteinase BaP1 on human endothelial cells. J Cell Biochem 2005; 94: 520-8.
[110] Tanjoni I, Weinlich R, Della-Casa MS, Clissa PB, Saldanha-Gama RF, de Freitas MS, et al. Jararhagin, a snake venom metalloprote-inase, induces a specialized form of apoptosis (anoikis) selective to endothelial cells. Apoptosis 2005; 10: 851-61.
[111] Baramova EN, Shannon JD, Bjarnason JB, Gonias SL, Fox JW. Interaction of hemorrhagic metalloproteinases with human -2 macroglobulin. Biochemistry 1990; 29: 1069-74.
[112] Kamiguti AS, Hay CRM, Zuzel M. Inhibition of collagen-induced platelet aggregation as the result of cleavage of 2 1-integrin by the snake venom metalloproteinase jararhagin. Biochem J 1996; 320: 635-41.
[113] Harris JB, Cullen MJ. Muscle necrosis caused by snake venoms and toxins. Electron Microsc Rev 1990; 3: 183-211.
[114] Gutiérrez JM, Ownby CL. Skeletal muscle degeneration induced by venom phospholipases A2: insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003; 42: 915-31.
[115] Dixon RW, Harris JB. Myotoxic activity of the toxic phospholi-pase, notexin, from the venom of the Australian tiger snake. J Neu-ropathol Exp Neurol 1996; 55: 1230-7.
[116] Melo PA, Burns CF, Blankemeyer JT, Ownby CL. Membrane depolarization is the initial action of crotoxin on isolated murine skeletal muscle. Toxicon 2004; 43: 111-9.
[117] Gutiérrez JM, Núñez J, Díaz C, Cintra ACO, Homsi-Brandeburgo MI, Giglio JR. Skaletal muscle degeneration and regeneration after injection of bothropstoxin-II, a phospholipase A2 isolated from the venom of the snake Bothrops jararacussu. Exp Mol Pathol 1991; 55: 217-29.
[118] Lambeau G, Lazdunski M. Receptors for a growing family of se-creted phospholipases A2. Trends Pharmacol Sci 1999; 20: 162-70.
[119] Díaz C, León G, Rucavado A, Rojas N, Schroit AJ, Gutiérrez JM. Modulation of the susceptibility of human erythrocytes to snake venom myotoxic phospholipases A2: role of negatively charged phospholipids as potential membrane binding sites. Arch Biochem Biophys 2001; 391: 56-64.
[120] Alape-Girón A, Persson B, Cederlund E, Flores-Díaz M, Gutiérrez JM, Thelestam M, et al. Elapid venom toxins: multiple recruit-ments of ancient scaffolds. Eur J Biochem 1999; 259: 225-34.
[121] Lomonte B, Angulo Y, Calderón L. An overview of lysine-49 phospholipase A2 myotoxins from crotalid snake venoms and their structural determinants of myotoxic action. Toxicon 2003; 42: 885-901.
[122] Chioato L, Ward RJ. Mapping structural determinants of biological activities in snake venom phospholipases A2 by sequence analysis and site directed mutagenesis. Toxicon 2003; 42: 869-83.
[123] Gutiérrez JM, Romero M, Núñez J, Chaves F, Borkow G, Ovadia M. Skeletal muscle necrosis and regeneration after injection of BaH1, a hemorrhagic metalloproteinase isolated from the venom of the snake Bothrops asper (terciopelo). Exp Mol Pathol 1995; 62: 28-41.
[124] Franca FOS, Málaque CMS. In: Cardoso JLC, Franca FOS, Fan HW, Málaque CMS, Haddad V. Animais Peconhentos no Brasil. Biologia, Clínica e Terapeutica. Sao Paulo, Sarvier 2003; 72-86.
[125] Rucavado A, Núñez J, Gutiérrez JM. Blister formation and skin damage induced by BaP1, a hemorrhagic metalloproteinase from the venom of the snake Bothrops asper. Int J Exp Pathol 1998; 79: 245-54.
[126] Rucavado A, Lomonte B, Ovadia M, Gutiérrez JM. Local tissue damage induced by BaP1, a metalloproteinase isolated from Bothrops asper (terciopelo) snake venom. Exp Mol Pathol 1995; 63: 186-99.
[127] Anai K, Sugiki M, Yoshida E, Maruyama M. Neutralization of a snake venom hemorrhagic metalloproteinase prevents coagulopa-thy after subcutaneous injection of Bothrops jararaca venom in rats. Toxicon 2002; 40: 63-8.
[128] Tu AT, Hendon RR. Characterization of lizard venom hyaluroni-dase and evidence for its action as a spreading factor. Comp Bio-chem Physiol B 1983; 76: 377-83.
[129] Girish KS, Shashidharamurthy R, Nagaraju S, Gowda TV, Kem-paraju K. Isolation and characterization of hyaluronidase, a "spreading factor" from Indian cobra (Naja naja) venom. Bio-chimie 2004; 86: 193-202.

2948 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
[130] Saravia-Otten P, Frisan T, Thelestam M, Gutiérrez JM. Membrane independent activation of fibroblast proMMP-2 by snake venom: novel roles for venom proteinases. Toxicon 2004; 44: 749-64.
[131] Farsky SHP, Antunes E, Mello SBV. Pro and anti-inflammatory properties of toxins from animal venoms. Curr Drug Targets In-flamm Allergy 2005; 4: 401-11.
[132] Lomonte B, Tarkowski A, Hanson LÅ. Host response to Bothrops
asper snake venom: analysis of edema formation, inflammatory cells, and cytokine release in a mouse model. Inflammation 1993; 17: 93-105.
[133] Rucavado A, Escalante T, Teixeira CFP, Fernandes CM, Díaz C, Gutiérrez JM. Increments in cytokines and matrix metalloprotein-ases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asper. Mediat Inflamm 2002; 11: 121-8.
[134] Teixeira CFP, Landucci ECT, Antunes E, Chacur M, Cury Y. In-flammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003; 42: 947-62.
[135] Zamunér SR, Gutiérrez JM, Muscará MN, Teixeira SA, Teixeira CFP. Bothrops asper and Bothrops jararaca snake venoms trigger microbicidal functions of peritoneal leukocytes in vivo. Toxicon 2001; 39: 1505-13.
[136] Zamunér SR, Zuliani JP, Fernandes CM, Gutiérrez JM, Teixeira CFP. Inflammation induced by Bothrops asper venom: release of proinflammatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon 2005; 46: 806-13.
[137] Zuliani JP, Fernandes CM, Zamunér SR, Gutiérrez JM, Teixeira CFP. Inflammatory events induced by Lys-49 and Asp-49 phos-pholipases A2 isolated fom Bothrops asper snake venom: role of catalytic activity. Toxicon 2005; 45: 335-46.
[138] Chaves F, Teixeira CFP, Gutiérrez JM. Role of nitric oxide in the local and systemic pathophysiological effects induced by Bothrops
asper snake venom in mice. Inflamm Res 2006; 55: 245-53. [139] Chacur M, Picolo G, Gutiérrez JM, Teixeira CFP, Cury Y. Phar-
macological modulation of hyperalgesia induced by Bothrops asper (terciopelo) snake venom. Toxicon 2001; 39: 1173-81.
[140] Chacur M, Longo I, Picolo G, Gutiérrez JM, Lomonte B, Guerra JL, et al. Hyperalgesia induced by Asp49 and Lys49 phospholi-pases A2 from Bothrops asper snake venom: pharmacological me-diation and molecular determinants. Toxicon 2003; 41: 667-78.
[141] Farsky SHP, Walber J, Costa-Cruz M, Cury Y, Teixeira CFP. Leu-kocyte response induced by Bothrops jararaca crude venom: in
vivo and in vitro studies. Toxicon 1997; 35: 185-93. [142] Gallagher P, Bao Y, Serrano SMT, Laing GD, Theakston RDG,
Gutiérrez JM, et al. Role of the snake venom toxin jararhagin in proinflammatory pathogenesis: in vitro and in vivo gene expression analysis of the effects of the toxin. Arch Biochem Biophys 2005; 441: 1-15.
[143] Clissa PB, Laing GD, Theakston RDG, Mota I, Taylor MJ, Moura-da-Silva AM. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on proinflammatory cytokines released by murine peritoneal adherent cells. Toxicon 2001; 39: 1567-73.
[144] Zuliani JP, Gutiérrez JM, Casais e Silva LL, Sampaio SC, Lomonte B, Teixeira CFP. Activation of cellular functions in macrophages by venom secretory Asp-49 and Lys-49 phopholipases A2. Toxicon 2005; 46: 523-32.
[145] Wei JF, Mo YZ, Quiao LY, Wei XL, Chen HQ, Xie H, et al. Potent histamine-releasing activity of atrahagin, a novel snake venom metalloproteinase. Int J Biochem Cell Biol 2006; 38: 510-20.
[146] Mora R, Valverde B, Díaz C, Lomonte B, Gutiérrez JM. A Lys-49 phospholipase A2 homologue from Bothrops asper snake venom induces proliferation, apoptosis and necrosis in a lymphoblastoid cell line. Toxicon 2005; 45: 651-60.
[147] Tidball JG. Inflammatory processes in muscle injury and repair. Am J Physiol Regul Integr Comp Physiol 2005; 288: R345-53.
[148] Korthuis RJ, Grisham MB, Granger DN. Leukocyte depletion attenuates vascular injury in postischemic skeletal muscle. Am J Physiol 1988; 254: H823-7.
[149] Moilanen E, Whittle B, Moncada S. In: Gallin JI, Snyderman R Eds. Inflammation. Basic Principles and Correlates. Philadelphia, Lippincot 1999; 787-800.
[150] Yong VW, Power C, Edwards DR. Metalloproteinases in biology and pathology of the nervous system. Nat Rev Neurosci 2001; 2: 502-11.
[151] Teixeira CFP, Zamuner SR, Zuliani JP, Fernandes CM, Cruz-Hofling MA, Fernandes I, et al. Neutrophils do not contribute to local tissue damage, but play a key role in skeletal muscle regener-arion, in mice injected with Bothrops asper snake venom. Muscle Nerve 2003; 28: 449-59.
[152] Chaves F, Teixeira CFP, Gutiérrez JM. Role of TNF- , IL-1 and IL-6 in the local tissue damage induced by Bothrops asper snake venom: an experimental assessment in mice. Toxicon 2005; 45: 171-8.
[153] Teixeira CFP, Chaves F, Zamunér SR, Fernandes CM, Zuliani JP, Cruz-Hofling MA, et al. Effects of neutrophil depletion in the local pathological alterations and muscle regeneration in mice injected with Bothrops jararaca snake venom. Int J Exp Pathol 2005; 86: 107-15.
[154] Rodrigues FG, Petretski JH, Kanashiro MM, Lemos L, Dias da Silva W, Kipnis TL. The complement system is involved in acute inflammation but not in the hemorrhage produced by a Bothrops
atrox snake venom low molecular mass proteinase. Mol Immunol 2004; 40: 1149-56.
[155] Moura-da-Silva AM, Laing GD, Paine MJI, Dennison JMTJ, Politi V, Crampton JM, et al. Processing of pro-tumor necrosis factor- by venom metalloproteinases: a hypothesis explaining local tissue damage following snake bite. Eur J Immunol 1996; 26: 2000-5.
[156] Laing GD, Clissa PB, Theakston RDG, Moura-da-Silva AM, Tay-lor MJ. Inflammatory pathogenesis of snake venom metalloprotein-ase-induced skin necrosis. Eur J Immunol 2003; 33: 3458-63.
[157] Miyabara EH, Tostes RC, Selistre-de-Araújo HS, Aoki MS, Moris-cot AS. Role of nitric oxide in myotoxic activity induced by cro-toxin in vivo. Toxicon 2004; 43: 425-32.
[158] Grounds M. Towards understanding skeletal muscle regeneration. Path Res Pract 1991; 187: 1-22.
[159] Hawke J, Garry DJ. Myogenic satellite cells: physiology to mo-lecular biology. J Appl Physiol 2001; 91: 534-51.
[160] León G, Valverde JM, Rojas G, Lomonte B, Gutiérrez JM. Com-parative study on the ability of IgG and Fab sheep antivenoms to neutralize local hemorrhage, edema and myonecrosis induced by Bothrops asper (terciopelo) snake venom. Toxicon 2000; 38: 233-44.
[161] Lomonte B, León G, Hanson LÅ. Similar effectiveness of Fab and F(ab')2 antivenoms in the neutralization of hemorrhagic activity of Vipera berus snake venom in mice. Toxicon 1996; 34: 1197-202.
[162] Chaves F, Loría GD, Salazar A, Gutiérrez JM. Intramuscular ad-ministration of antivenoms in experimental envenomation by Bothrops asper: comparison between Fab and IgG. Toxicon 2003; 41: 237-44.
[163] Yingprasertchai S, Bunyasrisawat S, Ratanabanangkoon K. Hyaluronidase inhibitors (sodium chromoglycate and sodium auro-thiomalate) reduce the local tissue damage and prolong the survival time of mice injected with Naja kaouthia and Calloselasma
rhodostoma venoms. Toxicon 2003; 42: 635-46. [164] Escalante T, Franceschi A, Rucavado A, Gutiérrez JM. Effective-
ness of batimastat, a synthetic inhibitor of matrix metalloprotein-ases, in neutralizing local tissue damage induced by BaP1, a hem-orrhagic metalloproteinase from the venom of the snake Bothrops
asper. Biochem Pharmacol 2000; 60: 269-74. [165] Rucavado A, Escalante T, Franceschi A, Chaves F, León G, Cury
Y, et al. Inhibition of local hemorrhage and dermonecrosis induced by Bothrops asper snake venom: effectiveness of early in situ ad-ministration of the peptidomimetic metalloproteinase inhibitor ba-timastat and the chelating agent CaNa2EDTA. Am J Trop Med Hyg 2000; 63: 313-9.
[166] Murakami MT, Arruda EZ, Melo PA, Martinez AB, Calil-Eliás S, Tomaz MA, et al. Inhibition of myotoxic activity of Bothrops asper myotoxin II by the anti-trypanosomal drug suramin. J Mol Biol 2005; 350: 416-26.
[167] Nagase H, Woessner JF. Matrix metalloproteinases. J Biol Chem 1999; 274: 21491-4.
[168] Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol 2001; 17: 463-516.
[169] Rao BG. Recent developments in the design of specific matrix metalloproteinase inhibitors aided by structural and computational studies. Curr Pharm Des 2005; 11: 295-322.
[170] Acharya MR, Venitz J, Figg WD, Sparreboom A. Chemically modified tetracyclines as inhibitors of matrix metalloproteinases. Drug Resist Updat 2004; 7: 195-208.

Snakebite Envenomation Therapy Current Pharmaceutical Design, 2007, Vol. 13, No. 28 2949
[171] Hanessian S, Moitessier N, Therrien E. A comparative docking study and the design of potentially selective MMP inhibitors. J Comput Aided Mol Des 2001; 15: 873-81.
[172] Coussens LM, Fingleton B, Matrisian LM. Matrix metalloprotein-ase inhibitors and cancer: trials and tribulations. Science 2002; 295: 2387-92.
[173] D’Alessio S, Gallina C, Gavuzzo E, Giordano C, Gorini B, Mazza F, et al. Inhibition of adamalysin II and MMPs by phosphonate analogues of snake venom peptides. Bioorg Med Chem 1999; 7: 389-94.
[174] Brown PD. Ongoing trials with matrix metalloproteinase inhibitors. Exp Opin Invest Drugs 2000; 9: 2167-77.
[175] Zhang D, Botos I, Gomis-Ruth FX, Doll R, Blood C, Njoroge FG, et al. Structural interaction of natural and synthetic inhibitors with the venom metalloproteinase, atrolysin C (form D). Proc Natl Acad Sci USA 1994; 91: 8447-51.
[176] Botos I, Scapozza L, Zhang D, Liotta LA, Meyer EF. Batimastat, a potent matrix metalloproteinase inhibitor, exhibits an unexpected mode of binding. Proc Natl Acad Sci USA 1996; 93: 2749-54.
[177] Francis B, Kaiser II. Inhibition of metalloproteinases in Bothrops
asper venom by endogenous peptides. Toxicon 1993; 31: 889-99. [178] Huang KF, Chiou SH, Ko TP, Wang AHJ. Determinants of the
inhibition of a Taiwan habu venom metalloproteinase by its en-dogenous inhibitors revealed by X-ray crystallography and syn-thetic inhibitor analogues. Eur J Biochem 2002; 269: 3047-56.
[179] Munekiyo SM, Mackessy SP. Presence of peptide inhibitors in rattlesnake venoms and their effects on endogenous metalloprote-ases. Toxicon 2005; 45: 255-63.
[180] Soares AM, Ticli FK, Marcussi S, Lourenco MV, Januário AH, Sampaio SV, et al. Medicinal plants with inhibitory properties against snake venoms. Curr Med Chem 2005; 12: 2625-41.
[181] Perales J, Neves-Ferreira AGC, Valente RH, Domont GB. Natural inhibitors of snake venom hemorrhagic metalloproteinases. Toxi-con 2005; 45: 1013-120.
[182] Valente RH, Dragulev B, Perales J, Fox JW, Domont GB. Cloning and characterization of BJ46a, a snake venom metalloproteinase inhibitor from Bothrops jararaca serum. Eur J Biochem 2001; 268: 3042-52.
[183] Rucavado A, Escalante T, Gutiérrez JM. Effect of the metalloprote-inase inhibitor batimastat in the systemic toxicity induced by Bothrops asper snake venom: understanding the role of metallopro-teinases in envenomation. Toxicon 2004; 43: 417-24.
[184] Kamiguti AS, Hay CRM, Theakston RDG, Zuzel M. Insights into the mechanism of haemorrhage caused by snake venom metallo-proteinases. Toxicon 1996; 34: 627-42.
[185] Kamiguti AS. Platelets as targets of snake venom metalloprotein-ases. Toxicon 2005; 45: 1041-9.
[186] Kudo I, Murakami M. Phospholipase A2 enzymes. Prostaglandins Other Lipid Mediat 2002; 68-69: 3-58.
[187] Valentin E, Lambeau G. Increasing molecular diversity of secreted phospholipases A2 and their receptors and binding proteins. Bio-chim Biophys Acta 2000; 1488: 59-70.
[188] Dennis EA. Diversity of group types, regulation, and function of phospholipase A2. J Biol Chem 1994; 269: 13057-60.
[189] Arni RK, Ward RJ. Phospholipase A2 -a structural review. Toxicon 1996; 34: 827-41.
[190] Lehr M. Phospholipase A2 inhibitors in inflammation. Expert Opin Ther Pat 2001; 11: 1123-36.
[191] Hansford KA, Reid RC, Clark CI, Tyndall JDA, Whitehouse MW, Guthrie T, et al. D-tyrosine as a chiral precusor to potent inhibitors of human nonpancreatic secretory phospholipase A2 (IIa) with anti-inflammatory activity. Chembiochem 2003; 4: 181-5.
[192] Woodruff TM, Arumugam TV, Shiels IA, Newman ML, Ross PA, Reid RC, et al. A potent and selective inhibitor of group IIa secre-tory phospholipase A2 protects rats from TNBS-induced colitis. Int Immunopharmacol 2005; 5: 883-92.
[193] Kini RM, Evans HJ. A model to explain the pharmacological ef-fects of snake venom phospholipases A2. Toxicon 1989; 27: 613-35.
[194] Kini RM. In: Kini RM Ed. Venom Phospholipase A2 Enzymes: Structure, Function and Mechanisms. Chichester, Wiley 1997; 1-28.
[195] Kini RM. Excitement ahead: structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003; 42: 827-40.
[196] Rosenberg P. In: Kini RM Ed. Venom Phospholipase A2 Enzymes: Structure, Function and Mechanisms. Chichester, Wiley 1997; 155-83.
[197] Deepa M, Gowda TV. Purification and characterization of a glyco-protein inhibitor of toxic phospholipase from Withania somnifera. Arch Biochem Biophys 2002; 408: 42-50.
[198] Iglesias CV, Aparicio R, Rodrigues-Simioni L, Camargo EA, An-tunes E, Marangoni S, et al. Effects of morin on snake venom phospholipase A2 (PLA2). Toxicon 2005; 46: 751-8.
[199] Melo PA, Ownby CL. Ability of wedelolactone, heparin, and para-bromophenacyl bromide to antagonize the myotoxic effects of two crotaline venoms and their PLA2 myotoxins. Toxicon 1999; 37: 199-215.
[200] Da Silva AJM, Coelho AL, Sima ABC, Moraes RAM, Pinheiro DA, Fernandes FFA, et al. Synthesis and pharmacological evalua-tion of prenylated and benzylated pterocarpans against snake ven-oms. Bioorg Med Chem Lett 2004; 14: 431-5.
[201] Vishwanath BS, Kini RM, Gowda TV. Characterization of three edema-inducing phospholipase A2 enzymes from habu (Trimeresu-
rus flavoviridis) venom and their interaction with the alkaloid aris-tolochic acid. Toxicon 1987; 25: 501-15.
[202] Ticli FK, Hage LIS, Cambraia RS, Pereira PS, Magro AJ, Fontes MRM, et al. Rosmarinic acid, a new snake venom phospholipase A2 inhibitor from Cordia verbenacea (Boraginaceae): antiserum ac-tion potentiation and molecular interaction. Toxicon 2005; 46: 318-27.
[203] Hung YC, Sava V, Hong MY, Huang GS. Inhibitory effects on phospholipase A2 and antivenin activity of melanin extracted from Thea sinensis Linn. Life Sci 2004; 74: 2037-47.
[204] Núñez V, Castro V, Murillo R, Ponce-Soto LA, Merfort I, Lomonte B. Inhibitory effects of Piper umbellatum and Piper peltatum ex-tracts towards phospholipases A2 from Bothrops snake venoms: isolation of 4-narolidylcatechol as active principle. Phytochemistry 2005; 66: 1017-25.
[205] Reynolds LJ, Mihelich ED, Dennis EA. Inhibition of venom phos-pholipases A2 by manoalide and manoalogue. Stoichiometry of in-corporation. J Biol Chem 1991; 266: 16512-7.
[206] Angulo Y, Lomonte B. Inhibitory effects of fucoidan on the activi-ties of crotaline snake venom myotoxic phospholipases A2. Bio-chem Pharmacol 2003; 66: 1993-2000.
[207] Lomonte B, Moreno E, Tarkowski A, Hanson LÅ, Maccarana M. Neutralizing interaction between heparins and myotoxin II, a Lys-49 phospholipase A2 from Bothrops asper snake venom. Identifica-tion of a heparin-binding and cytolytic toxin region by the use of synthetic peptides and molecular modeling. J Biol Chem 1994; 269: 29867-73.
[208] Melo PA, Homsi-Brandeburgo MI, Giglio JR, Suárez-Kurtz G. Antagonism of the myotoxic effects of Bothrops jararacussu venom and bothropstoxin by polyanions. Toxicon 1993; 31: 285-91.
[209] Faure G. Natural inhibitors of toxic phospholipases A2. Biochimie 2000; 82: 833-40.
[210] Lizano S, Domont G, Perales J. Natural phospholipase A2 myo-toxin inhibitor proteins from snakes, mammals and plants. Toxicon 2003; 42: 963-77.
[211] Ohkura N, Ohkuhara H, Inoue S, Ikeda K, Hayashi K. Purification and characterization of three distinct types of phospholipase A2 in-hibitors from the blood plasma of the Chinese mamushi, Agkistro-
don blomhoffi siniticus. Biochem J 1997; 325: 527-31. [212] Thwin MM, Satish RL, Chan STF, Gopalakrishnakone P. Func-
tional site of endogenous phospholipase A2 inhibitor from phyton serum. Phospholipase A2 binding and antiinflammatory activity. Eur J Biochem 2002; 269: 719-27.
[213] Rocha SLG, Lomonte B, Neves-Ferreira AGC, Trugilho MRO, Junqueira-de-Azevedo LM, Ho PL, et al. Functional analysis of DM64, an antimyotoxic protein with immunoglobulin-like struc-ture from Didelphis marsupialis serum. Eur J Biochem 2002; 269: 6052-62.
[214] de Oliveira M, Cavalcante WLG, Arruda EZ, Melo PA, Silva MD, Gallacci M. Antagonism of myotoxic and paralyzing activities of bothropstoxin-I by suramin. Toxicon 2003; 42: 373-9.
[215] Stein CA. Suramin: a novel antineoplastic agent with multiple potential mechanisms of action. Cancer Res 1993; 53: 2239-48.

2950 Current Pharmaceutical Design, 2007, Vol. 13, No. 28 Gutiérrez et al.
[216] Chan YS, Li Y, Foster W, Fu FH, Huard J. The use of suramin, an antifibrotic agent, to improve muscle recovery after strain injury. Am J Sports Med 2005; 33: 43-51.
[217] Chandra V, Jasti J, Kaur P, Srinivasan A, Betzel C, Singh TP. Structural basis of phospholipase A2 inhibition for the synthesis of prostaglandins by the plant alkaloid aristolochic acid from a 1.7 Å crystal structure. Biochemistry 2002; 41: 10914-9.
[218] Singh RK, Vikram P, Makker J, Jabeen T, Sharma S, Dey S, et al. Design of specific peptide inhibitors for group I phospholipase A2: structure of a complex formed between phospholipase A2 from Naja naja sagittifera (group I) and a designed peptide inhibitor Val-Ala-Phe-Arg-Ser (VAFRS) at 1.9 Å resolution reveals unique features. Biochemistry 2003; 42: 11701-6.
[219] Georgieva DN, Rypniewski W, Gabdoulkhakov A, Genov N, Bet-zel C. Asp49 phospholipase A2–elaidoylamide complex: a new mode of inhibition. Biochem Biophys Res Commun 2004; 319: 1314-21.
[220] Girish KS, Kemparaju K. Inhibition of Naja naja venom hyaluron-idase: role in the management of poisonous bite. Life Sci 2006; 78: 1433-40.
[221] Gowda TV. In: Kini RM Ed. Venom Phospholipase A2 Enzymes: Structure, Function and Mechanisms. Chichester, Wiley 1997; 205-21.
[222] Vital Brazil O. Coral snake venoms: mode of action and patho-physiology of experimental envenomation. Rev Inst Med Trop Sao Paulo 1987; 29: 119-26.
[223] Barros SF, Friedlanskaia I, Petricevich VL, Kipnis TL. Local in-flammation, lethality and cytokine release in mice injected with Bothrops atrox venom. Mediat Inflamm 1998; 7: 339-46.
[224] Avila-Aguero ML, París MM, Peterson PK, Gutiérrez JM, Lo-monte B, Faingezicht I. Systemic cytokine response in children bit-ten by snakes in Costa Rica. Pediatr Emerg Care 2001; 17: 425-9.
[225] Petricevich VL, Teixeira CFP, Tambourgi DV, Gutiérrez JM. In-crements in serum cytokine and nitric oxide levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxi-con 2000; 38: 1253-66.
[226] Szold O, Ben-Abraham R, Weinbroum AA, Englender TE, Ovadia D, Sorkine M, et al. Antagonization of TNF attenuates systemic hemodynamic manifestations of envenomation in a rat model of Vipera aspis snakebite. Intensive Care Med 2001; 27: 884-8.
[227] Szold O, Ben-Abraham R, Frolkis I, Sorkine M, Sorkine P. Tumor necrosis factor as a mediator of cardiac toxicity following snake envenomation. Crit Care Med 2003; 31: 1449-53.
[228] Westerlund B, Nordlund P, Uhlin U, Eaker D, Eklund H. The three-dimensional structure of notexin, a presynaptic neurotoxic phospholipase A2 at 2.0 Å resolution. FEBS Lett 1992; 301: 159-64.
[229] Holland DR, Clancy LL, Muchmore SW, Ryde TJ, Einspahr HM, Finzel BC, et al. The crystal structure of a lysine 49 phospholipase A2 from the venom of the cottonmouth snake at 2.0-Å resolution. J Biol Chem 1990; 265: 17649-56.
[230] Núñez CE, Angulo Y, Lomonte B. Identification of the myotoxic site of the Lys49 phospholipase A2 from Agkistrodon piscivorus
piscivorus snake venom: synthetic C-terminal peptides from Lys49, but not from Asp49 myotoxins, exert membrane-damaging activi-ties. Toxicon 2001; 39: 1587-94.
Related Documents











