Proof Delivery Form Please return this form with your proof CUP reference: Date of delivery: Journal and Article number: HYG486 Volume and Issue Number: &(0) Number of colour figures: Number of pages (not including this page): 8 Epidemiology and Infection Here is a proof of your article for publication in the Journal. Please print out the file and check the proofs carefully, make any corrections necessary on a hardcopy, and answer queries on the proofs. Please return the corrected hardcopy proof together with the copyright transfer form and offprint order form as soon as possible (no later than 3 days after receipt) to: Steve Parr 20 Hall Lane Burwell Cambs CB5 0HE UK To avoid delay from overseas, please send the proof by air mail or courier. ’ You are responsible for correcting your proofs. Errors not found may appear in the published journal. ’ The proof is sent to you for correction of typographical errors only. Revision of the substance of the text is not permitted, unless discussed with the editor of the journal. ’ Please answer carefully any queries raised from the typesetter. ’ A new copy of a figure must be provided if correction of anything other than a typographical error introduced by the typesetter is required—please provide this in eps format and print it out and staple it to the proof. ’ If the paper was written in LateX, please do not send corrections as Latex code as we have now translated the Latex to typesetting code. If you have no corrections to make or very few corrections please contact [email protected] email the corrections (quoting page and line number) to save having to return your paper proof. In this case the copyright transfer form (which must be signed) can be sent as an accompanying pdf file. Note to Author : Papers are generally published online within 4 weeks of receipt of the corrected proof. Author queries: AQ1: Check change to ‘‘within the framework’’ is OK. AQ2: Supply address details for Bio-Rad. AQ3: ‘‘IU’’ has been used throughout, not ‘‘UI’’ AQ4: Ref. 27 was not cited in text – it has been added here – OK? AQ5: Supply details of any conflict of interest. If none simply state ‘‘None’’. AQ6: Supply English version of title for ref. 1. Typesetter queries: Non-printed material:

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Proof Delivery Form
Please return this form with your proof
CUP reference:
Date of delivery:
Journal and Article number: HYG486
Volume and Issue Number: &(0)
Number of colour figures:
Number of pages (not including this page): 8
Epidemiology and Infection
Here is a proof of your article for publication in the Journal. Please print out the file and check the proofs carefully, make anycorrections necessary on a hardcopy, and answer queries on the proofs.
Please return the corrected hardcopy proof together with the copyright transfer form and offprint order form as soon as possible(no later than 3 days after receipt) to:
Steve Parr20 Hall LaneBurwellCambsCB5 0HEUK
To avoid delay from overseas, please send the proof by air mail or courier.
’ You are responsible for correcting your proofs. Errors not found may appear in the published journal.’ The proof is sent to you for correction of typographical errors only. Revision of the substance of the text is not permitted,
unless discussed with the editor of the journal.’ Please answer carefully any queries raised from the typesetter.’ A new copy of a figure must be provided if correction of anything other than a typographical error introduced by the
typesetter is required—please provide this in eps format and print it out and staple it to the proof.’ If the paper was written in LateX, please do not send corrections as Latex code as we have now translated the Latex to
typesetting code.
If you have no corrections to make or very few corrections please contact [email protected] email the corrections(quoting page and line number) to save having to return your paper proof. In this case the copyright transfer form(which must be signed) can be sent as an accompanying pdf file.
Note to Author : Papers are generally published online within 4 weeks of receipt of the corrected proof.
Author queries:
AQ1: Check change to ‘‘within the framework’’ is OK.AQ2: Supply address details for Bio-Rad.AQ3: ‘‘IU’’ has been used throughout, not ‘‘UI’’AQ4: Ref. 27 was not cited in text – it has been added here – OK?AQ5: Supply details of any conflict of interest. If none simply state ‘‘None’’.AQ6: Supply English version of title for ref. 1.
Typesetter queries:
Non-printed material:

Abcdefghi
VAT REG N0. GB 823 8476 09
Epidemiology and Infection volume…..no……..
Offprints
25 offprints of each article will be supplied free to each first named author and sent to a single address. Please complete this form and send it to the editor (address below) within 14 days of the date stamped on it. Please give the address to which your offprints should be sent. They will be despatched by surface mail within one month of publication. For an article by more than one author this form is sent to you as the first named. All extra offprints should be ordered by you in consultation with your co-authors Number of offprints required in addition to the 25 free copies
Offprints to be sent to (PRINT IN BLOCK CAPITALS) ……………………………………………………………………………………. …………………………………………………………………………………………………………………………………………………………... ………………………………………………………………………………………………………………….. Post/Zip Code……………… Date…………………………………Author(s)………………………………………………………………………….. Article title……………………………………………………………………………………………………………….. All enquiries about offprints should be addressed to the publisher: Journals Office, Cambridge University Press, Edinburgh Building, Shaftesbury Road, Cambridge. CB2 2BS UK
Charges for extra offprints
Number of copies 25 50 100 150 200 per 50 extra 1-4 pages £27.00 £43.50 £69.50 £95.50 £123.50 £27.00 5-8 pages £43.50 £65.00 £95.50 £128.50 £159.50 £73.50 9-16 pages £48.00 £72.50 £114.00 £152.50 £197.50 £48.00 17-24 pages £52.50 £80.50 £132.50 £180.50 £239.50 £52.50 extra 8 pages £8.00 £12.50 £20.00 £28.00 £41.50 £8.00
Methods of payment VAT at the local rate may he added to the above charges if paid by EU residents not registered for VAT. If registered, please quote your VAT number, or the VAT number of any agency paying on your behalf if registered. ❑ Payment against invoice. The invoice will be sent to you after publication of your article. ❑ Cheques should he made out to Cambridge University Press. ❑ Payment by someone else. Please enclose the official order when returning this form. Or ensure that when the order is sent, it mentions the name of the journal and the article title. Payment may be made by any credit card bearing the Interbank Symbol.
Signature of card holder ………………Amount….. …………Date…………….. Card number……………………………… Card expiry date………………………… Card verification number………………………. The card verification number is a 3 digit number printed on the back of your Visa or Master card, it appears after and to the right of your card number. For American Express the verification number is 4 digits, and printed on the front of your card, after and to the right of your card number.
Please advise if address registered with card company is different from above
For office use only
Reference Sent Acknowledged Price Received
offprint order form

transfer of copyright abcdefghi Please read the notes overleaf and then complete, sign, and return this form to the Journals Office, Cambridge University Press, University Printing House, Shaftesbury Road, Cambridge CB2 2BS, UK as soon as possible.
EPIDEMIOLOGY AND INFECTION In consideration of the publication in EPIDEMIOLOGY AND INFECTION of the contribution entitled:...................................................................................................................................................
..............................................................................................................................................................................................
by (all authors’ names): ..........................................................................................................................................................
..............................................................................................................................................................................................
1 To be filled in if copyright belongs to you Transfer of copyright
I/we hereby assign to Cambridge University Press, full copyright in all formats and media in the said contribution.
I/we warrant that I am/we are the sole owner or co-owners of the material and have full power to make this agreement, and that the material does not contain any scandalous matter or infringe any existing copyright.
I/we further warrant that permission has been obtained from the copyright holder for any material not in my/our copyright and the appropriate acknowledgement made to the original source. I/we attach copies of all permission correspondence.
I/we hereby assert my/our moral rights in accordance with the UK Copyrights Designs and Patents Act (1988).
Signed (tick one) □ the sole author(s)
□ one author authorised to execute this transfer on behalf of all the authors of the above article
Name (block letters) .............................................................................................................................................................
Institution/Company............................................................................................................................................................
Signature: .............................................................. Date: .................................................................................................
(Additional authors should provide this information on a separate sheet.)
2 To be filled in if copyright does not belong to you a Name and address of copyright holder..............................................................................................................................
.........................................................................................................................................................................................
.........................................................................................................................................................................................
.........................................................................................................................................................................................
b The copyright holder hereby grants to Cambridge University Press the non-exclusive right to publish the contribution in the journal and to deal with requests from third parties in the manner specified in paragraphs 4 and 6 overleaf.
(Signature of copyright holder or authorised agent) .........................................................................................................
3 US Government exemption I/we certify that the paper above was written in the course of employment by the United States Government so that no copyright exists.
Signature: .............................................................. Name (Block letters): ........................................................................
4 Requests received by Cambridge University Press for permission to reprint this article should be sent to (see para. 5 overleaf) Name and address (block letters) ...........................................................................................................................................
..............................................................................................................................................................................................

Notes for contributors 1 The information provided on this form will be held in perpetuity for record purposes. The name(s) and address(es) of
the author(s) of the contribution may be reproduced in the journal and provided to print and online indexing and abstracting services and bibliographical databases.
2 The Journal's policy is to acquire copyright in all contributions. There are two reasons for this: (a) ownership of
copyright by one central organisation tends to ensure maximum international protection against unauthorised use; (b) it also ensures that requests by third parties to reprint or reproduce a contribution, or part of it, are handled efficiently and in accordance with a general policy that is sensitive both to any relevant changes in international copyright legislation and to the general desirability of encouraging the dissemination of knowledge.
3 Two ‘moral rights’ were conferred on authors by the UK Copyright Act in 1988. In the UK an author’s ‘right of
paternity’, the right to be properly credited whenever the work is published (or performed or broadcast), requires that this right is asserted in writing.
4 Notwithstanding the assignment of copyright in their contribution, all contributors retain the following
non-transferable rights: • The right to (continue to) post a preprint of the contribution on their personal or departmental web page provided the
first screen contains the statement that the paper has been accepted for publication in Epidemiology and Infection published by Cambridge University Press together with the appropriate copyright notice. On publication the full bibliographical details (volume: issue number (date), page numbers) must be inserted after the journal title.
• Subject to file availability, the right to post the contribution as published on their own or their departmental home page provided the first screen includes full bibliographical details and the appropriate copyright notice.
• The right to make hard copies of the contribution or an adapted version for their own purposes, including the right to make multiple copies for course use by their students, provided no sale is involved.
• The right to reproduce the paper or an adapted version of it in any volume of which they are editor or author. Permission will automatically be given to the publisher of such a volume, subject to normal acknowledgement.
5 We shall use our best endeavours to ensure that any direct request we receive to reproduce your contribution, or a
substantial part of it, in another publication (which may be an electronic publication) is approved by you before permission is given.
6 Cambridge University Press co-operates in various licensing schemes that allow material to be photocopied within agreed
restraints (e.g. the CCC in the USA and the CLA in the UK). Any proceeds received from such licenses, together with any proceeds from sales of subsidiary rights in the Journal, directly support its continuing publication.
7 It is understood that in some cases copyright will be held by the contributor’s employer. If so, Cambridge University
Press requires non-exclusive permission to deal with requests from third parties, on the understanding that any requests it receives from third parties will be handled in accordance with paragraphs 4 and 5 above (note that your approval and not that of your employer will be sought for the proposed use).
8 Permission to include material not in your copyright If your contribution includes textual or illustrative material not in your copyright and not covered by fair use / fair
dealing, permission must be obtained from the relevant copyright owner (usually the publisher or via the publisher) for the non-exclusive right to reproduce the material worldwide in all forms and media, including electronic publication. The relevant permission correspondence should be attached to this form.
If you are in doubt about whether or not permission is required, please consult the Permissions Controller, Cambridge
University Press, The Edinburgh Building, Shaftesbury Road, Cambridge CB2 2RU, UK. Fax: +44 (0)1223 315052. Email: [email protected].
Please make a duplicate of this form for your own records

Trends in antimicrobial resistance phenotypes in non-typhoid
Salmonellae from human and poultry origins in France
J. CAILHOL 1, R. LAILLER 2, P. BOUVET 3, S. LA VIEILLE1, F. GAUCHARD 1,
P. SANDERS 4AND A. BRISABOIS 2*
1 French Agency for Food Safety (AFSSA), Direction de l’evaluation des risques nutritionnels et sanitaires,
Maisons-Alfort, France2 AFSSA Lerqap, Maisons-Alfort, France3 Centre National de Reference des Salmonella (CNRS), Institut Pasteur, Paris, France4 AFSSA, Fougeres, France
(Accepted 13 May 2005)
SUMMARY
A total of 1873 strains from human origin and 4283 strains from non-human origin of Salmonella
enterica serotypes Typhimurium, Enteritidis, Heidelberg, Hadar and Virchow, collected over
three years 1993, 1997 and 2000, were examined in order to determine the rate of antimicrobial
resistance to 12 antimicrobial drugs. The objective of the study was to describe and to compare
the evolution of the main resistance types in human and non-human isolates, focusing on the
poultry sector. The evolution and the rates of antimicrobial resistances for the five serotypes, with
the exception of Virchow, were almost comparable in strains isolated from human and non-
human sources over the period studied. The most striking result concerning single resistance was
the spectacular increase of the resistance frequency to nalidixic acid for the strains belonging to
serotypes Hadar and Virchow, especially in the poultry food sector (14% in 1993 vs. 72% in 2000
for Salmonella Virchow, 4% in 1993 vs. 70% in 2000 for Salmonella Hadar) and also in human
isolates (24% in 1997 vs. 48% in 2000 for S. Virchow, 31% in 1997 vs. 78% in 2000 for
S. Hadar). In addition to the classical resistance to ampicillin, streptomycin, sulphonamide,
chloramphenicol and tetracycline (ASSuCT resistance type), which stabilized between 1997 and
2000, the emergence of a new resistance type was observed.
INTRODUCTION
Salmonellosis is the most frequent foodborne disease
reported in France, often transmitted by foods of
animal origin, especially raw eggs, egg products or
poultry meats, causing more than 300 outbreaks per
year [1]. To address this public health problem,
national surveillance networks to monitor Salmo-
nellae of both human and non-human origins were
set up. These networks are used to reveal trends in
the evolution of different serotypes, monitor their
antimicrobial susceptibility, detect outbreaks, and
contribute to their investigation. Antimicrobial re-
sistance monitoring has become more important since
the appearance and dissemination of multidrug-
resistant Salmonella Typhimurium phage type
DT104, conferring resistance to ampicillin, strepto-
mycin, sulphonamide, chloramphenicol and tetra-
cycline (resistance-type ASSuCT). The genes encoding
for these resistances are clustered on the chromo-
somal genomic island SGI1 and can be transferred
* Author for correspondence : Dr A. Brisabois, AFSSA Lerqap,Maisons-Alfort, France.(Email : [email protected])
Epidemiol. Infect. (2005), &, 1–8. f 2005 Cambridge University Press
doi:10.1017/S0950268805004863 Printed in the United Kingdom

clonally and also horizontally to other serotypes [2, 3].
The increasing evolution of resistance-type ASSuCT
has been reported in several countries since the 1990s
[4–7]. Besides this resistance type, other antimicrobial
resistances, notably to quinolones, appeared initially
in developing countries [8] and then worldwide [9, 10].
Many reports suggest that the main cause of in-
creasing antimicrobial resistance among Salmonella
isolates from animals is the misuse and overuse of
antimicrobials in the breeding sector, and that some
of these strains may be transmitted through food to
humans, thereby contributing to the increase of anti-
microbial resistance among humans [11–13]. These
high antimicrobial resistance levels in zoonotic Salmo-
nella may contribute to an increase of morbidity in
human infections because resistant bacteria may be
more virulent and the treatment less effective [14, 15].
The purpose of this descriptive study was to evalu-
ate trends in antimicrobial resistance in the main
Salmonella enterica serotypes of human and poultry
sources over 8 years in France, by reviewing data
collected annually in 1993, 1997 and 2000. The
following five predominant serotypes were chosen
for the study: Typhimurium, Enteritidis, Hadar,
Heidelberg and Virchow. Observed trends are
discussed with respect to data on veterinary anti-
microbial consumption in France.
MATERIALS AND METHODS
Bacterial isolates
Surveillance of isolates from animal and food origin
has been carried out by the French Agency for Food
Safety (AFSSA) through the ‘Salmonella ’ Network.
Strains were classified in three main sectors : (i) animal
health; meaning poultry, pig, cattle or other farm
animal isolates and nearby environment isolates, rep-
resenting more than 70% of the total collected iso-
lates; (ii) food hygiene; meaning isolates from animal
food products, feedstuffs and slaughter houses, for
25% of isolates ; (iii) ecosystems; meaning water
purification plant, harbour and sewage isolates cor-
responding to <5% of isolates. However, the third
sector did not have a sufficiently large sample size and
was not used in this analysis.
Strains were collected on the basis of voluntary
participation of veterinary and food hygiene labora-
tories, located throughout the national area. All col-
lected strains were sent with information concerning
the origin, animal species, categories of food and
regional localization of the samples. The samples
were mainly collected within the framework AQ1of self-
regulating systems based on legislation requirements
or on random official sampling (eggs) and monitoring
plans (poultry farming), and within the animal
breeding environment.
The French Salmonella Reference Centre (CNRS)
at the Pasteur Institute (Paris, France) has conducted
the surveillance of human isolates by collecting
specification sheets and isolates from public and
private clinical laboratories of analysis for several
decades. The strains have been isolated from clinical
samples (blood or faeces), and sent to the CNRS on
the basis of voluntary participation. Human strains
were selected annually for susceptibility testing, in
respect of the distribution of serotypes.
Serotyping and antimicrobial susceptibility testing
Serotype was determined by the Kauffmann–White
method, using Bio-Rad AQ2(formerly Sanofi Diagnostics
Pasteur) standard antisera and home-made antisera.
This study reports the results observed for the
five serotypes : Typhimurium, Enteritidis, Hadar,
Virchow and Heidelberg, selected on the basis of their
high frequency of isolation both in human and non-
human origins (Table 1). The antimicrobial sus-
ceptibility was tested by standard disk diffusion on
Mueller–Hinton agar plates according to the guide-
lines of the Comite de l’Antibiogramme de la Societe
Francaise de Microbiologie (CASFM).
Zone diameters were automatically read by the
Osiris system (Bio-Rad) using the break-points
defined by the guidelines of CASFM, and have been
interpreted as predictive of treatment effectiveness
classes: resistant (R), intermediate (I), susceptible (S).
Strains classified as intermediate class were con-
sidered in this study as susceptible. E. coli ATCC
25922 strain was used for quality control.
Ten antimicrobial drugs and two combinations of
antimicrobials have been tested on all isolates as
follows (disk content indicated in parentheses) : nali-
dixic acid (30 mg), ampicillin (10 mg) for non-human
isolates, amoxicillin (10 mg) for human isolates,
amoxicillin-clavulanic acid (20 mg+10 mg), strepto-
mycin (10 IU) AQ3, trimethoprim–sulphamethoxazole
(1.25 mg+23.75 mg), trimethoprim (5 mg), sulphon-
amide (200 mg), tetracycline (30 IU), chloramphenicol
(30 mg), gentamycin (10 IU), cefotaxime (30 mg),
ofloxacin (5 mg). These listed antibiomicrobials cor-
respond to those recommended for the surveillance of
2 J. Cailhol and others
antimicrobial susceptibility by Caprioli et al. [16] and
by the European group (Office International des
Epizooties). Sulphonamide was not tested in animal
isolates in 1993. Concerning the ampicillin and
amoxicillin disks tested, we considered that their
cross-susceptibilities and cross-resistances were com-
plete [17]. Table 2 gives the number of tested
strains and fully susceptible strains for each studied
serotype.
Statistical analysis
Statistical analyses were performed using Epi-Info
software (version 6.04b, CDC, Atlanta, GA, USA).
We assessed the emergence of new R-types, possibly
clustered in a gene, by comparing the observed rate of
associated resistance to the predicted rate if these
resistances were occurring independently, using the
McNemar test. The x2 trends test was used for evalu-
ating evolution of resistance to nalidixic acid and
ofloxacin over the 8 years. P values <0.05 were con-
sidered as significant.
RESULTS
Frequency of isolation and global rate of resistance
(Tables 1 and 2)
S. Typhimurium was the second most common sero-
type found in human isolates and the most common
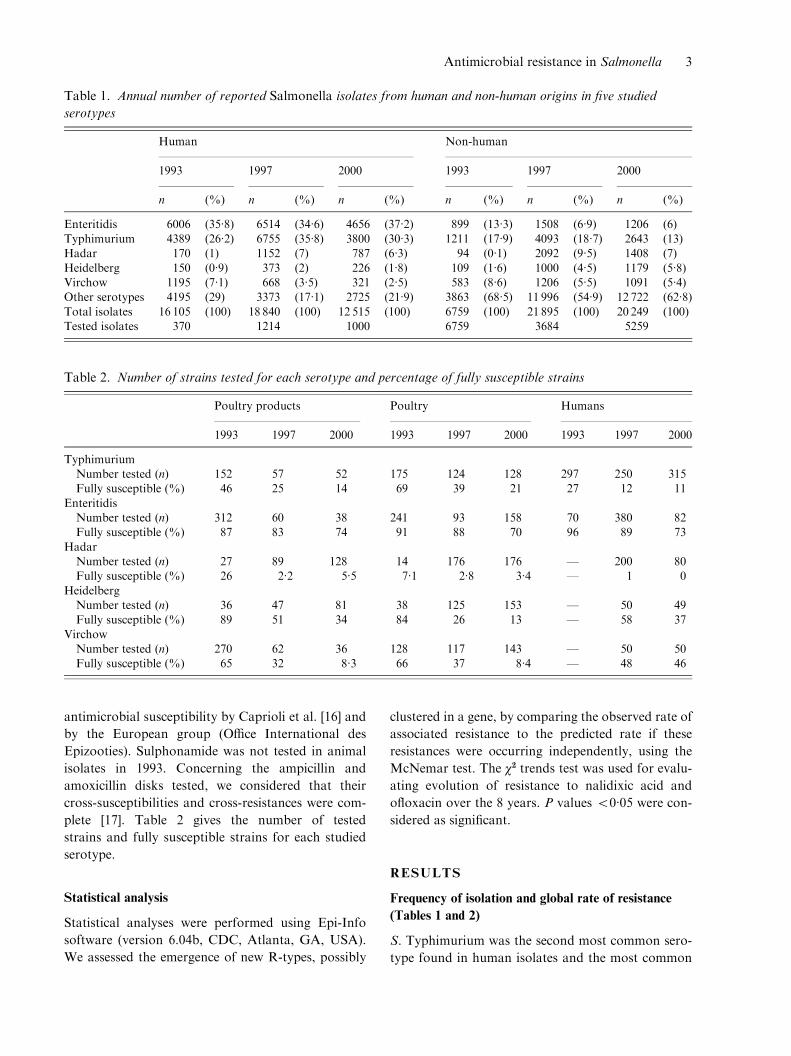
Table 1. Annual number of reported Salmonella isolates from human and non-human origins in five studied
serotypes
Human Non-human
1993 1997 2000 1993 1997 2000
n (%) n (%) n (%) n (%) n (%) n (%)
Enteritidis 6006 (35.8) 6514 (34.6) 4656 (37.2) 899 (13.3) 1508 (6.9) 1206 (6)Typhimurium 4389 (26.2) 6755 (35.8) 3800 (30.3) 1211 (17.9) 4093 (18.7) 2643 (13)Hadar 170 (1) 1152 (7) 787 (6.3) 94 (0.1) 2092 (9.5) 1408 (7)
Heidelberg 150 (0.9) 373 (2) 226 (1.8) 109 (1.6) 1000 (4.5) 1179 (5.8)Virchow 1195 (7.1) 668 (3.5) 321 (2.5) 583 (8.6) 1206 (5.5) 1091 (5.4)Other serotypes 4195 (29) 3373 (17.1) 2725 (21.9) 3863 (68.5) 11 996 (54.9) 12 722 (62.8)
Total isolates 16 105 (100) 18 840 (100) 12 515 (100) 6759 (100) 21 895 (100) 20 249 (100)Tested isolates 370 1214 1000 6759 3684 5259
Table 2. Number of strains tested for each serotype and percentage of fully susceptible strains
Poultry products Poultry Humans
1993 1997 2000 1993 1997 2000 1993 1997 2000
TyphimuriumNumber tested (n) 152 57 52 175 124 128 297 250 315Fully susceptible (%) 46 25 14 69 39 21 27 12 11
EnteritidisNumber tested (n) 312 60 38 241 93 158 70 380 82Fully susceptible (%) 87 83 74 91 88 70 96 89 73
HadarNumber tested (n) 27 89 128 14 176 176 — 200 80Fully susceptible (%) 26 2.2 5.5 7.1 2.8 3.4 — 1 0
HeidelbergNumber tested (n) 36 47 81 38 125 153 — 50 49Fully susceptible (%) 89 51 34 84 26 13 — 58 37
VirchowNumber tested (n) 270 62 36 128 117 143 — 50 50Fully susceptible (%) 65 32 8.3 66 37 8.4 — 48 46
Antimicrobial resistance in Salmonella 3

in animal isolates. The proportion of human S.
Typhimurium susceptible to all the antimicrobials
tested decreased significantly over the 8 years (27% to
11%, P<10x5).
S. Enteritidis was the most frequent serotype in
human isolates among the serotypes tested and
its rank was second or third among non-human iso-
lates. Since 1993, susceptible human, animal and
food isolates have been decreasing; nevertheless it
remained the most susceptible serotype among those
studied.
S. Hadar constituted <1% of all isolates in 1993
but increased rapidly between 1993 and 1997 both in
human and non-human isolates. There were already
few susceptible human isolates in 1997 (1%) and they
totally disappeared in 2000.
Isolation of the serotype S. Heidelberg increased
strongly among non-human strains between 1993 and
2000. The proportion of susceptible strains in human
isolates decreased between 1997 and 2000 from 58%
down to 37%. Although levels of resistance were very
low to all drugs in 1993, the proportion of susceptible
strains has steadily declined. Strains isolated from
animals had generally higher resistance rates than
those from poultry products.
S. Virchow isolation decreased between 1993 and
2000 in both human and non-human sectors. It was
the only serotype in which the proportion of suscep-
tible strains was stable in human isolates (48% of all
isolates in 1997 and 46% in 2000). The proportion of
susceptible isolates from the poultry sector steadily
decreased during the 1993–2000 period.
ASSuCT R-type and new R-types (Table 3)
The frequency of resistance type (R-type) ASSuCT
strains greatly increased among Typhimurium iso-
lates between 1993 and 2000 (P<10x3). The occur-
rence of this R-type was much higher than would be
predicted by the independent occurrence of each of
the antimicrobials separately. In 2000, 40% of human
isolates and 45% of poultry sector isolates harbour-
ing the ASSuCT R-type were also nalidixic acid-
resistant. Moreover, 43% of trimethoprim-resistant
human isolates also presented the ASSuCT R-type.
However, the ASSuCT R-type appears not to have
disseminated among the other serotypes.
Even without any spread of R-type ASSuCT, the
serotype Hadar has shown the most spectacular in-
crease in resistance over the 8 years with the lowest
rate of susceptible strains among the five serotypes.
Resistances to ampicillin or amoxicillin, streptomycin
and tetracycline, with or without the resistance to
nalidixic acid often occurred together but they were
not statistically associated in a particular R-type.
In Heidelberg, human strains showed approxi-
mately the same antimicrobial resistance as those
from poultry products, with an increase in rates
between 1997 and 2000. A novel R-type ASSuT,
emerged strongly from 4% in 1997 to 54% in 2000 for
animal isolates. This R-type was detected in 6% of
human isolates in 1997 and represented 22% in 2000,
commonly with additional resistance to trimetho-
prim. These R-type values were much higher than the
predicted values based on the occurrence of each
Table 3. Percentages of ASSuCT R-type and other association of resistances among the five serotypes
R-type
Poultry products Poultry Humans
1993 1997 2000 1993 1997 2000 1993 1997 2000
ASSuCT Typhimurium 3.9* 39 40 8.6* 28 42 41 61 59Predicted values 0.5* 2.5 4.6 0.1* 1.5 4.9 5.4 17 16ASSuCT Enteritidis 0* 0 0 1.6* 0 0.6 0 0.2 0
ASSuCT Hadar 0* 0 0 0* 0.6 0.6 n.d. 0.1 0ASSuCT Heidelberg 0.3* 0 1.2 2.6* 2.4 1.3 n.d. 6 0ASSuCT Virchow 1.1* 6.4 5.5 2.3* 0.8 1.4 n.d. 0 2AST Hadar 0 48 60 0 61 49 n.d. 68 55
Predicted values — 44 51 — 57 46 — 61 55ASTNal Hadar 0 0.5 0.6 0 0.6 0.4 n.d. 0.7 0.6Predicted values — 0.3 0.4 — 0.4 0.2 — 0.5 0.4
ASSuT Heidelberg 0* 4.3 25 0* 4 54 n.d. 6 22Predicted values — 0.2 2.6 — 1.6 22 — 0.2 2.7
* Sulphonamide was not tested.
4 J. Cailhol and others
antimicrobial separately and confirmed the co-
occurrence of these resistances were not independent.
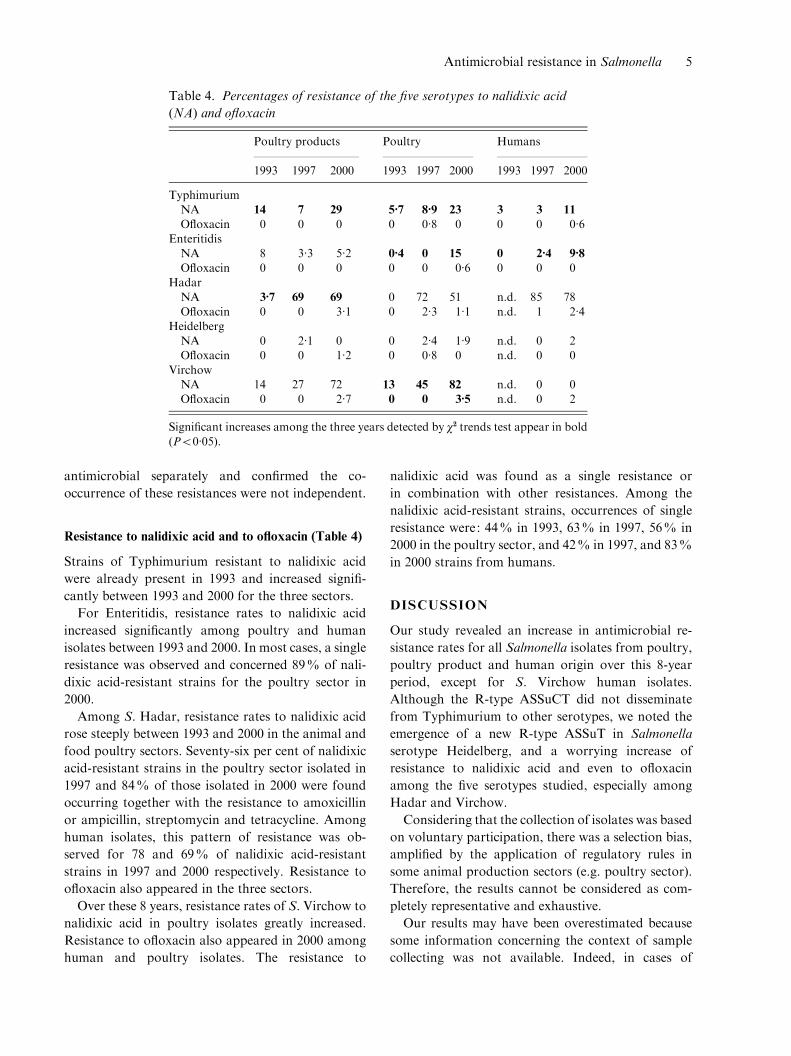
Resistance to nalidixic acid and to ofloxacin (Table 4)
Strains of Typhimurium resistant to nalidixic acid
were already present in 1993 and increased signifi-
cantly between 1993 and 2000 for the three sectors.
For Enteritidis, resistance rates to nalidixic acid
increased significantly among poultry and human
isolates between 1993 and 2000. In most cases, a single
resistance was observed and concerned 89% of nali-
dixic acid-resistant strains for the poultry sector in
2000.
Among S. Hadar, resistance rates to nalidixic acid
rose steeply between 1993 and 2000 in the animal and
food poultry sectors. Seventy-six per cent of nalidixic
acid-resistant strains in the poultry sector isolated in
1997 and 84% of those isolated in 2000 were found
occurring together with the resistance to amoxicillin
or ampicillin, streptomycin and tetracycline. Among
human isolates, this pattern of resistance was ob-
served for 78 and 69% of nalidixic acid-resistant
strains in 1997 and 2000 respectively. Resistance to
ofloxacin also appeared in the three sectors.
Over these 8 years, resistance rates of S. Virchow to
nalidixic acid in poultry isolates greatly increased.
Resistance to ofloxacin also appeared in 2000 among
human and poultry isolates. The resistance to
nalidixic acid was found as a single resistance or
in combination with other resistances. Among the
nalidixic acid-resistant strains, occurrences of single
resistance were: 44% in 1993, 63% in 1997, 56% in
2000 in the poultry sector, and 42% in 1997, and 83%
in 2000 strains from humans.
DISCUSSION
Our study revealed an increase in antimicrobial re-
sistance rates for all Salmonella isolates from poultry,
poultry product and human origin over this 8-year
period, except for S. Virchow human isolates.
Although the R-type ASSuCT did not disseminate
from Typhimurium to other serotypes, we noted the
emergence of a new R-type ASSuT in Salmonella
serotype Heidelberg, and a worrying increase of
resistance to nalidixic acid and even to ofloxacin
among the five serotypes studied, especially among
Hadar and Virchow.
Considering that the collection of isolates was based
on voluntary participation, there was a selection bias,
amplified by the application of regulatory rules in
some animal production sectors (e.g. poultry sector).
Therefore, the results cannot be considered as com-
pletely representative and exhaustive.
Our results may have been overestimated because
some information concerning the context of sample
collecting was not available. Indeed, in cases of
Table 4. Percentages of resistance of the five serotypes to nalidixic acid
(NA) and ofloxacin
Poultry products Poultry Humans
1993 1997 2000 1993 1997 2000 1993 1997 2000
TyphimuriumNA 14 7 29 5.7 8.9 23 3 3 11
Ofloxacin 0 0 0 0 0.8 0 0 0 0.6EnteritidisNA 8 3.3 5.2 0.4 0 15 0 2.4 9.8
Ofloxacin 0 0 0 0 0 0.6 0 0 0HadarNA 3.7 69 69 0 72 51 n.d. 85 78
Ofloxacin 0 0 3.1 0 2.3 1.1 n.d. 1 2.4HeidelbergNA 0 2.1 0 0 2.4 1.9 n.d. 0 2Ofloxacin 0 0 1.2 0 0.8 0 n.d. 0 0
VirchowNA 14 27 72 13 45 82 n.d. 0 0Ofloxacin 0 0 2.7 0 0 3.5 n.d. 0 2
Significant increases among the three years detected by x2 trends test appear in bold
(P<0.05).
Antimicrobial resistance in Salmonella 5

human disease, an antimicrobial treatment could have
been administered before collecting stool samples and
may have selected resistant strains. Nevertheless, the
geographical distribution and the number of samples
were large enough to reveal trends in antimicrobial
resistance during this 8-year period.
Since Salmonella infection is a zoonotic disease,
antimicrobial resistance of strains from animals and
humans have been frequently linked to antimicrobial
use in agriculture [4, 7, 11–13, 18–24]. For this reason,
monitoring of antimicrobial resistance among Salmo-
nella from food-animals and humans on the one hand,
and of antimicrobial use in agriculture, on the other,
has been initiated recently in France [25, 26] and
worldwide (WHO, DANMAP, Euro-surveillance).
The comparison between the evolution of anti-
microbial resistance and antibiotic use in agriculture
is complicated, first because of the lack of statistics in
this sector, and second due to the presence of multiple
interaction factors. Nevertheless, our observations
can be evaluated based on the first study of veterinary
antimicrobials sold in France in 1999 [G. Moulin,
personal communication, 1999]. During that year,
tetracycline, aminoglycoside, ampicillin or amoxicillin
and sulphonamides composed 83% of the antibiotics
used as veterinary drugs in animals. In this study, the
highest rates of resistance were observed among these
antimicrobial families, as shown by the association of
resistance to ampicillin, streptomycin and teracycline
in serotype Hadar, or the emergence of the R-type
ASSuT in serotype Heidelberg. In contrast, no
resistance to chloramphenicol was observed among
serotype Heidelberg, although it acquired the R-type
ASSuT. One hypothesis for this observation is
that Heidelberg was rare before 1994, the year
chloramphenicol was banned. Therefore, Heidelberg
strains have not been exposed to a selective pressure
by this antibiotic. Comparing R-type ASSuT occur-
rences observed with the predictive values, suggests
that genes encoding this R-type may be clustered on
the same genetic element. Molecular analysis should
be conducted in order to assess the organization of
genetic elements associated with this new R-type.
Similarly, the highest levels of ASSuCT resistance
were observed among S. Typhimurium, both in
poultry and in human isolates. This could be ex-
plained by the clonal spread of Typhimurium DT104,
linked to the well-documented pattern of anti-
microbial resistance ASSuCT [2–4, 7, 22, 23]. In our
study, frequency of ASSuCT R-type strains remained
stable for the serotype Typhimurium between 1997
and 2000 but were 10-fold higher than the predictive
values given by the occurrence of each antimicrobial
drug independently. Moreover, no evidence of large
horizontal dissemination of this R-type was observed
even if some previous results showed the presence of
the genomic island structure SGI1 in other serotypes
such as Agona, Paratyphi B and Albany [27].
Furthermore, besides regulation on the use of
specific antimicrobials, policies to eradicate infected
flocks may also have played a role. For instance, since
1998 poultry flocks infested by S. Typhimurium and
S. Enteritidis have been eradicated in accordance with
the CEE zoonosis directive (no. 92/117). The fre-
quency of isolation of S. Enteritidis is stable among
human strains, and low in non-human strains, prob-
ably due to these legislative requirements.
We also observed the emergence of nalidixic acid
resistance among human and poultry isolates in
S. Typhimurium. Resistance to quinolones was
studied in 2001 by Cloeckaert and colleagues regard-
ing the emergence of Typhimurium DT104 after the
introduction of enrofloxacin in veterinary medicine in
1993 [2, 27, 28] AQ4. In 1999 in France, quinolones rep-
resented 1.5% of the total of quantities of anti-
microbial drugs sold, which is relatively low, but the
available data cannot allow us to analyse a correlation
between consumption and resistance. Oral formu-
lations of quinolones and fluoroquinolones have been
authorized for the treatment of E. coli infection in
poultry. When a prophylactic or curative treatment is
given to the whole flock or herd, via mixing with feed
or water, some resistant strains can emerge and
spread through animals not taking antibiotics at a
bactericidal concentration [19, 29].
Cases of treatment failure of ceftriaxone and
decreased susceptibility to ciprofloxacin in human
isolates have been observed [21, 30, 31]. Furthermore,
some studies demonstrate the presence of subpopu-
lations with resistant or decreased susceptibility to
fluoroquinolone among the nalidixic acid-resistant
isolates, which might influence the outcome of fluoro-
quinolone treatment [32]. In 1999, Threlfall drew our
attention to the increase of isolated strains with
decreased susceptibility to ciprofloxacin among
strains belonging to serotypes Virchow and Hadar in
England [33]. In our study, these two serotypes had
the highest rates of resistance to nalidixic acid. The
most striking increase of resistance concerned the
serotype Hadar with the frequent association of nali-
dixic acid resistance to resistance to amoxicillin,
streptomycin and tetracycline. In Finland, between
6 J. Cailhol and others

1995 and 1997, quinolone-resistant strains of S.
Hadar harboured a resistance to streptomycin, tetra-
cycline and sometimes to ampicillin or amoxicillin [9].
Our results are also in accord with Cruchaga et al.,
who observed that the ASTNal resistance pattern
with additional cephalotin resistance was the most
frequent R-type [5]. With regard to our results, it
should be noted that resistance to ofloxacin emerged
in 2000 among S. Hadar human strains.
The results of our analyses show some trends in the
evolution of antimicrobial resistances in Salmonella
strains in France and led us to detect some new
R-type emergence in the five most-studied serotypes.
The resistance monitoring system must be improved
and should be compared with the distribution of
veterinary antimicrobials. Such improved surveillance
will be necessary to analyse the mechanisms and
modes of dissemination, and public health impact of
both the emerging resistances and the pathogens
themselves.
ACKNOWLEDGEMENTS
The authors thank V. Vaillant of the Institut de Veille
Sanitaire (InVS) for expert assistance. Part of this
work has been supported by InVS. We gratefully
acknowledge Mrs Jayne Ireland and Mrs Claire
Evans (AFSSA) for greatly improving the English
grammar and style of the manuscript.
DECLARATION OF INTEREST
&.AQ5
REFERENCES
1. Haeghebaert S, Le Querrec F, Gallay A, Bouvet P,
Gomez M, Vaillant V. Les toxi-infections alimentairescollectives en France en 1999 et 2000. In: SurveillancenationaleAQ6 des maladies infectieuses, 1998–2000. Institut
de Veille Sanitaire, Saint-Maurice, France. Novembre2002: 161–169.
2. Cloeckaert A, Schwarz S. Molecular characterization,
spread and evolution of multidrug resistance inSalmonella enterica typhimuriumDT104. Vet Res 2001;32 : 301–310.
3. Threlfall EJ. Epidemic salmonella typhimurium DT104 – a truly international multiresistant clone.J Antimicrob Chemother 2000; 46 : 7–10.
4. Breuil J, Brisabois A, Casin I, Armand-Lefevre L, Fremy
S, Collatz E. Antibiotic resistance in salmonellae iso-lated from humans and animals in France: comparative
data from 1994 and 1997. J Antimicrob Chemother2000; 46 : 965–971.
5. Cruchaga S, Echeita A, Aladuena A, Garcia-Pena J,
Frias N, Usera MA. Antimicrobial resistance in salmo-nellae from humans, food and animals in Spain in 1998.
J Antimicrob Chemother 2001; 47 : 315–321.6. Leegaard TM, Caugnat DA, Froholm LO, Hoiby EA,
Lassen J. Emerging antibiotic resistance in Salmonellatyphimurium in Norway. Epidemiol Infect 2000; 125 :
473–480.7. Murphy TM, McNamara E, Hill M, et al.
Epidemiological studies of human and animal
Salmonella typhimurium DT104 and DT104b isolatesin Ireland. Epidemiol Infect 2001; 126 : 3–9.
8. Hart CA, Kariuki S. Antimicrobial resistance in
developing countries. BMJ 1998; 317 : 647–650.9. Hakanen A, Siitonen A, Kotilainen P, Huovinen P.
Increasing fluoroquinolone resistance in Salmonella
serotypes in Finland during 1995–1997. J AntimicrobChemother 1999; 43 : 145–148.
10. Threlfall EJ, Ward LR, Rowe B. Resistance to cipro-floxacin in non-typhoidal salmonellas from humans in
England and Wales – the current situation. ClinMicrobiol Infect 1999; 5 : 130–134.
11. Aarestrup FM.Association between the consumption of
antimicrobial agents in animal husbandry and theoccurrence of resistant bacteria among food animals.Int J Antimicrob Agents 1999; 12 : 279–285.
12. McEwen SA, Fedorka-Cray PJ. Antimicrobial useand resistance in animals. Clin Infect Dis 2002; 34
(Suppl 3) : S93–S106.
13. van den Bogaard AE, Stobberingh EE. Epidemiology ofresistance to antibiotics. Links between animals andhumans. Int J Antimicrob Agents 2000; 14 : 327–335.
14. Barza M, Travers K. Excess infections due to anti-
microbial resistance : the ‘Attributable Fraction’. ClinInfect Dis 2002; 34 (Suppl 3) : S126–S130.
15. Travers K, Barza M. Morbidity of infections caused by
antimicrobial-resistant bacteria. Clin Infect Dis 2002;34 (Suppl 3) : S131–S134.
16. Caprioli A, Busani L, Martel JL, Helmuth R.
Monitoring of antibiotic resistance in bacteria ofanimal origin : epidemiological and microbiologicalmethodologies. Int J Antimicrob Agents 2000; 14 :295–301.
17. Fuchs PC, Barry AL, Pfaller MA, Hardy DJ,
McLaughlin JC, Gerlach EH. In vitro activity ofampicillin, amoxicillin, ampicillin-sulbactam, and
amoxicillin-clavulanic acid against consecutive clinicalisolates of Enterobacteriaceae. Diagn Microbiol InfectDis 1993; 17 : 171–175.
18. Franklin A, Acar J, Anthony F, et al. Antimicrobialresistance : harmonisation of national antimicrobialresistance monitoring and surveillance programmes in
animals and in animal-derived food. Rev Sci Tech 2001;20 : 859–870.
19. Bager F, Helmuth R. Epidemiology of resistance toquinolones in Salmonella. Vet Res 2001; 32 : 285–290.
20. Davies RH, Teale CJ, Wray C, et al. Nalidixic acidresistance in salmonellae isolated from turkeys and
Antimicrobial resistance in Salmonella 7

other livestock in Great Britain. Vet Rec 1999; 144 :320–322.
21. Fey PD, Safranek TJ, Rupp ME, et al. Ceftriaxone-resistant salmonella infection acquired by a child fromcattle. N Engl J Med 2000; 342 : 1242–1249.
22. Poppe C, Smart N, Khakhria R, Johnson W, Spika J,
Prescott J. Salmonella typhimurium DT104: a virulentand drug-resistant pathogen. Can Vet J 1998; 39 :559–565.
23. Rabsch W, Tschape H, Baumler AJ. Non-typhoidalsalmonellosis : emerging problems. Microbes Infect2001; 3 : 237–247.
24. White DG, Zhao S, Sudler R, et al. The isolation ofantibiotic-resistant salmonella from retail groundmeats. N Engl J Med 2001; 345 : 1147–1154.
25. Martel JL, Tardy F, Brisabois A, Lailler R, Coudert M,
Chaslus-Dancla E. The French antibiotic resistancemonitoring programs. Int J Antimicrob Agents 2000;
14 : 275–283.26. Martel JL, Tardy F, Sanders P, Boisseau J. New trends
in regulatory rules and surveillance of antimicrobialresistance in bacteria of animal origin. Vet Res 2001;
32 : 381–392.27. Cloeckaert A, Sidi Boumedine K, Flaujac G, Imberechts
H, D’Hooghe I, Chaslus Dancla E. Occurrence of a
Salmonella enterica serotype Typhimurium DT104-likeantibiotic resistance gene cluster including the floR gene
in S. enterica serotype Agona. Antimicrob AgentsChemother 2000; 44 : 1359–1361.
28. Cloeckaert A, Chaslus-Dancla E. Mechanisms ofquinolone resistance in Salmonella. Vet Res 2001; 32 :291–300.
29. Johnston AM. Use of antimicrobial drugs in veterinarypractice. BMJ 1998; 317 : 665–667.
30. Threlfall EJ, Skinner JA, Graham A, Ward LR,
Smith HR. Resistance to ceftriaxone and cefotaxime in
non-typhoidal Salmonella enterica in England andWales, 1998–99. J Antimicrob Chemother 2000; 46 :860–862.
31. Walker RA, Lawson AJ, Lindsay EA, et al. Decreasedsusceptibility to ciprofloxacin in outbreak-associatedmultiresistant Salmonella typhimuriumDT104. Vet Rec
2000; 147 : 395–396.32. Hakanen A, Kotilainen P, Jalava J, Siitonen A,
Huovinen P. Detection of decreased fluoroquinolone
susceptibility in Salmonellas and validation of nalidixicacid screening test. J Clin Microbiol 1999; 37 :3572–3577.
33. Threlfall EJ, Ward LR, Skinner JA, Graham A.
Antimicrobial drug resistance in non-typhoidal salmo-nellas from humans in England and Wales in 1999:decrease in multiple resistance in Salmonella enterica
serotypes Typhimurium, Virchow, and Hadar. MicrobDrug Resist 2000; 6 : 319–325.
8 J. Cailhol and others
Related Documents











