ORIGINAL PAPER Transposon-based high sequence diversity in Avr-Pita alleles increases the potential for pathogenicity of Magnaporthe oryzae populations P. K. Singh & S. Thakur & R. Rathour & M. Variar & S. K. Prashanthi & A. K. Singh & U. D. Singh & V . Sharma & N. K. Singh & T. R. Sharma Received: 28 November 2013 /Revised: 28 February 2014 /Accepted: 2 March 2014 # Springer-Verlag Berlin Heidelberg 2014 Abstract Magnaporthe oryzae causes rice blast that is one of the most devastating diseases of rice worldwide. Highly var- iable nature of this fungus has evolved itself against major resistance genes in newly released rice varieties. Understanding the population structure of this fungus is es- sential for proper utilization of the rice blast resistance genes in rice crop plants. In the present study, we analyzed 133 isolates of M. oryzae from ten countries to find the allelic variation of Avr-Pita gene that is triggering Pita-mediated resistance in rice plant. The diversity analysis of these alleles showed higher level of nucleotide variation in the coding regions than the noncoding regions. Evolutionary analysis of these alleles indicates that Avr-Pita gene is under purifying selection to favor its major alleles in 133 isolates analyzed in this study. We hypothesize that the selection of favorable Avr- Pita allele in these isolates may occur through a genetic mechanism known as recurrent selective sweeps. A total of 22 functional Avr-Pita protein variants were identified in this study. Insertion of Pot3 transposable element into the promot- er of Avr-Pita gene was identified in virulent isolates and was Electronic supplementary material The online version of this article (doi:10.1007/s10142-014-0369-0) contains supplementary material, which is available to authorized users. P. K. Singh : S. Thakur : N. K. Singh : T. R. Sharma (*) National Research Centre on Plant Biotechnology, IARI, New Delhi 110012, India e-mail: [email protected] T. R. Sharma e-mail: [email protected] P. K. Singh e-mail: [email protected] S. Thakur e-mail: [email protected] N. K. Singh e-mail: [email protected] P. K. Singh : V. Sharma Department of Bioscience and Biotechnology, Banasthali University, Tonk, Rajasthan 304 022, India V. Sharma e-mail: [email protected] R. Rathour Department of Agricultural Biotechnology, CSK Himachal Pradesh Agricultural University, Palampur, Himachal Pradesh 176062, India e-mail: [email protected] M. Variar Central Rainfed Upland Rice Research Station, CRRI, Hazaribagh, Jharkhand 825301, India e-mail: [email protected] S. K. Prashanthi School of Agricultural Biotechnology, University of Agricultural Sciences, Dharwad, Karnataka 58001, India e-mail: [email protected] A. K. Singh : U. D. Singh Indian Agricultural Research Institute, New Delhi 110012, India A. K. Singh e-mail: [email protected] U. D. Singh e-mail: [email protected] Funct Integr Genomics DOI 10.1007/s10142-014-0369-0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Transposon-based high sequence diversity in Avr-Pita allelesincreases the potential for pathogenicity of Magnaportheoryzae populations
P. K. Singh & S. Thakur & R. Rathour & M. Variar &
S. K. Prashanthi & A. K. Singh & U. D. Singh & V. Sharma &
N. K. Singh & T. R. Sharma
Received: 28 November 2013 /Revised: 28 February 2014 /Accepted: 2 March 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Magnaporthe oryzae causes rice blast that is one ofthe most devastating diseases of rice worldwide. Highly var-iable nature of this fungus has evolved itself against majorresistance genes in newly released rice varieties.Understanding the population structure of this fungus is es-sential for proper utilization of the rice blast resistance genesin rice crop plants. In the present study, we analyzed 133isolates of M. oryzae from ten countries to find the allelicvariation of Avr-Pita gene that is triggering Pita-mediatedresistance in rice plant. The diversity analysis of these alleles
showed higher level of nucleotide variation in the codingregions than the noncoding regions. Evolutionary analysis ofthese alleles indicates that Avr-Pita gene is under purifyingselection to favor its major alleles in 133 isolates analyzed inthis study. We hypothesize that the selection of favorable Avr-Pita allele in these isolates may occur through a geneticmechanism known as recurrent selective sweeps. A total of22 functional Avr-Pita protein variants were identified in thisstudy. Insertion of Pot3 transposable element into the promot-er of Avr-Pita gene was identified in virulent isolates and was
Electronic supplementary material The online version of this article(doi:10.1007/s10142-014-0369-0) contains supplementary material,which is available to authorized users.
P. K. Singh : S. Thakur :N. K. Singh : T. R. Sharma (*)National Research Centre on Plant Biotechnology, IARI, NewDelhi 110012, Indiae-mail: [email protected]
T. R. Sharmae-mail: [email protected]
P. K. Singhe-mail: [email protected]
S. Thakure-mail: [email protected]
N. K. Singhe-mail: [email protected]
P. K. Singh :V. SharmaDepartment of Bioscience and Biotechnology, Banasthali University,Tonk, Rajasthan 304 022, India
V. Sharmae-mail: [email protected]
R. RathourDepartment of Agricultural Biotechnology, CSK Himachal PradeshAgricultural University, Palampur, Himachal Pradesh 176062, Indiae-mail: [email protected]
M. VariarCentral Rainfed Upland Rice Research Station, CRRI, Hazaribagh,Jharkhand 825301, Indiae-mail: [email protected]
S. K. PrashanthiSchool of Agricultural Biotechnology, University of AgriculturalSciences, Dharwad, Karnataka 58001, Indiae-mail: [email protected]
A. K. Singh :U. D. SinghIndian Agricultural Research Institute, New Delhi 110012, India
A. K. Singhe-mail: [email protected]
U. D. Singhe-mail: [email protected]
Funct Integr GenomicsDOI 10.1007/s10142-014-0369-0

suggested that mobility of repeat elements in avirulence genesofM. oryzae seems to help in emergence of new virulent typesof the pathogen. Allele-specific markers developed in thisstudy will be helpful to identify a particular type of Avr-Pitaallele fromM. oryzae population which can form the basis forthe deployment of Pita gene in different epidemiologicalregions.
Keywords Avr-Pita .Magnaporthe oryzae . Nucleotidevariation . Pot3 . Race shifting . Haplotype
AbbreviationsAvr AvirulenceInDel Insertion and deletionSNP Single nucleotide polymorphismASM Allele-specific marker
Introduction
The rice blast disease caused by ascomycetous fungus,Magnaporthe oryzae (Couch et al. 2005), is one of the mostdevastating diseases of rice worldwide (Ou 1985). The bestway to manage this menace is by using host resistanceemploying major and/or minor resistance (R) genes integratedwith effective cultural practices. Identification of major Rgenes started in the early decades of nineteenth century bythe discovery of physiological races of the rice blast pathogenthat differ in their ability to cause disease on different ricevarieties (Yamada 1985). The rice varieties containing major Rgenes are effective against the M. oryzae races containing thecorresponding Avr gene (Silue et al. 1992). To date, approxi-mately 100 R genes for blast resistance have been mapped and20 of these genes cloned in rice (Sharma et al. 2012).
The resistance in newly released rice cultivars against therice blast fungal pathogen can be broken down rapidly due tothe high level of changes occurring in the genome of thispathogen. One way to overcome this problem is pyramidingof multiple rice blast resistance genes, each recognizing aparticular set of M. oryzae isolates, into a single rice cultivarthat makes it broad-spectrum resistant. The rapid evolution ofnew races of M. oryzae is attributed to loss of function ofavirulence effector genes that correspond to the major resis-tance genes. New virulent races of the plant pathogen emergeby genetic modifications of Avr genes using mechanisms likepoint mutation (Joosten et al. 1994), insertion of transposons(Kang et al. 2001), deletions (Dodds et al. 2006), and frame-shift mutations (Ridout et al. 2006). Extensive surveys of largenatural populations of fungal pathogens revealed that instabil-ity of Avr genes is a commonmechanism of evolution towardsvirulence (Van de Wouw et al. 2010). Till date, approximately40 different Avr genes of M. oryzae have been identified (Maet al. 2006), of which 9 genes Avr-Pita (Orbach et al. 2000),
Avr1CO39 (Farman and Leong 1998), PWL1 (Kang et al.1995), PWL2 (Sweigard 1995), ACE1 (Fudal et al. 2005),AvrPiz-t (Li et al. 2009), Avr-Pia, Avr-Pii, and Avr-Pik/km/kp
(Yoshida et al. 2009) have been cloned and characterized.The rice blast resistance gene, Pita, has been characterized
and is being used extensively in blast resistance breedingprograms worldwide. The effectiveness of this gene relies onthe ability to recognize the pathogen’s correspondingavirulence gene Avr-Pita. It has been reported that the proc-essed Avr-Pita protein from the rice blast fungus interactsdirectly with the protein of blast resistance gene Pita leadingto high degree of resistance (Bryan et al. 2000; Jia et al. 2000).The original Avr-Pita, renamed as Avr-Pita1, is a member ofthe gene family consisting of two additional members’ viz.Avr-Pita2 and Avr-Pita3 that share 92 and 71 % nucleotidesequence identity, respectively. Avr-Pita2 was a functionalavirulence gene in contrast to Avr-Pita3 (Khang et al. 2008).Avr-Pita gene is unstable in certain fungal strains and pro-duces spontaneous mutants very frequently under laboratory(Valent and Chumley 1991) and field conditions (Dai et al.2010). Molecular analysis implicated deletion events, in theform of point mutations (Orbach et al. 2000), and transposoninsertion (Kang et al. 2001) for the generation of mutant types(Zhou et al. 2007). Further, the telomeric position of the genealso accounts for the instability of the gene (Orbach et al.2000). Apart from that, transposable elements also play amajor role in the evolution of genome subsequently affectingvirulence pattern. A transposable element (Pot3) located at thepromoter region is associated with the virulence in M. oryzaeisolates (Kang et al. 2001). This element has also been iden-tified to disrupt the protease motif of the Avr-Pita allelecausing shift from avirulence to virulence (Zhou et al. 2007;Dai et al. 2010).
For better understanding, the mechanisms of frequent ap-pearance of new races could potentially allow us to surveypathogen populations for their potential for the evolution ofcertain races. Such information in turn will help us to minimizethe incidence of rapid breakdown of resistance by assisting theidentification of proper R genes for breeding and/or engineer-ing, and will also facilitate the deployment of disease resis-tance in a way that minimizes the evolution of new races.Therefore, it is essential to study about the Avr genes and theiralleles in naturally-occurring field isolates in the differentgeographical regions of the world, which may yield valuableinformation to facilitate the proper deployment of R geneswith durable resistance against the rice blast disease. The blastresistance gene, Pita, was well studied in different geograph-ical locations of the world as well as in India (Thakur et al.2013) in contrast toAvr-Pita gene. Therefore, the present workattempts to study the Avr-Pita variants identified fromM. oryzaepopulations to define the polymorphism patterns, molecularevolution, and the selective forces shaping the evolution of theAvr-Pita gene in the populations.
Funct Integr Genomics

Materials and methods
Fungal cultures and growth conditions
The infected leaves of rice genotypes with typical blast diseasesymptoms collected from different parts of the country wereused to establish single spore cultures of the pathogen. A totalof 80 field isolates of M. oryzae were collected from the fourmajor rice blast hot spot regions of India viz., Eastern, North-western, Northern, and Southern India (Table S1). The detailsrelated to these isolates, like geographical parameters,GenBank accession number, and pathogenicity on Pita gene,are given in Table S1. Established cultures were subsequentlymaintained as desiccated fungal colonized filter disks follow-ing the method of Valent et al. (1986). The stored cultureswere revived by culturing on oatmeal agar medium (oatmeal50 g/L, agar 15 g/L) for 10 days at 25°C. For inducingsporulation, the fungal cultures were subcultured onMathur's medium (Mathur et al. 1960) and incubated at25°C under blue and white fluorescence light for 7 days. Forobtaining mycelial growth, the fungal plugs (5 mm) weretransferred to flasks containing liquid yeast extract broth(yeast extract 3 g/L, casamino acid 3 g/L, and glucose 10 g/L) and incubated in dark for 5 days on a rotary shaker at150 rpm. The mycelia were collected onWhatman filter paperand washed with distilled autoclaved water to remove tracesof the media and then aseptically dried on sterilized blottingsheets. The harvested mycelia were stored at −20°C and usedfurther for the isolation of genomic DNA.
Pathogenicity assays
The pathogenicity of M. oryzae isolates was tested on mono-genic rice line IRBLTA-K1, harboring Pita gene and suscep-tible genotype Lijiang Xintuan Heigu (LTH) following stan-dard procedures as described elsewhere (Bonman et al. 1986).About 30–40 mL of the spore suspension (2×105 conidia/ml)containing gelatin (0.1 %) and Tween-20 (0.02 %) was sprayedonto 21-day-old seedlings using a glass atomizer. Inoculatedseedlings were kept in a humidity chamber maintained at 25±1 °C and sprayed three to four times a day with distilled waterto maintain high humidity. Disease reactions were recordedafter 7 days of inoculation by using a 0–5 disease rating scale(Mackill and Bonman 1992). A disease reaction of 0–3 wasconsidered as resistant type, whereas a reaction of 4–5 wasconsidered as susceptible type. The pathogenicity of eachisolate was tested thrice to reconfirm the results.
DNA extraction, PCR amplification, sequencing,and sequence analysis
Genomic DNAwas isolated from frozen mycelia using CTABmethod essentially developed by Murray and Thompson
(1980) with minor modifications in the extraction buffer (ad-dition of 1 % sodium dodecyl sulfate and 4 mM spermidine).Five primer pairs were designed (Table S2) using the sequenceinformation of Avr-Pita (AF207841) for amplification andsequencing of the alleles (Untergrasser et al. 2012). Besides,95 Avr-Pita alleles belonging to ten countries weredownloaded from NCBI (http://blast.ncbi.nlm.nih.gov/) andused in the present study for combined analysis (Table S3). Atotal of 133 Avr-Pita alleles were categorized into sixpopulations based on their country of origin. Less than fivealleles available from different countries were grouped intoothers category. The PCR amplification, sequencing, andsequence analysis methods were similar to that of Thakuret al. (2013). The haplotype network was constructed manu-ally as well as by using TCS1.21 (http://darwin.uvigo.es/;Clement et al. 2000) and NETWORK software (http://www.fluxus-engineering.com/sharenet.htm), respectively. The Avr-Pita3 gene, which is a nonfunctional member of the Avr-Pitagene family, was used as an out-group for this analysis.
Allele-specific marker development
To distinguish the avirulent and virulent Avr-Pita alleles fromdifferent isolates of M. oryzae, allele-specific markers weredesigned based on Pot3 transposon element (AF333034) andthe original Avr-Pita gene (AF207841). Two sets of primer pairs,TRSpot3-1 (F; CATCACTAACATCAGCAAAGTC and R;CTTTTAACGGCGTGATAGCC) and TRSpot3-2 (F; AGGCGTATATCGGATGTTAGT and R; CTTTTAACGGCGTGATAGCC), were used for amplification of Pot3 fragments from theisolates ofM. oryzae using Pfu polymerase (FINNZYMES OY,Keilaranta, Espoo, Finland) with the following thermal cyclingconditions: initial DNAdenaturation at 95 °C for 2min followedby 35 cycles of 95 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min,and final extension at 72 °C for 5 min and hold at 4 °C.
Results
Virulence analysis
The 80 fungal isolates collected from different geographicalregions of India were tested for their virulence on amonogenicline IRBLTA-K1 containing blast resistance gene Pita and asusceptible check LTH. We found that 38 isolates were aviru-lent on Pita gene and the rest were virulent. All the isolates werevirulent on the susceptible check. An example of the virulence/avirulence phenotypes of the isolates is given in Fig. 1.
Nucleotide variation and evolution of Avr-Pita alleles
All the 133 Avr-Pita alleles (95 Avr-Pita alleles downloadedfrom NCBI database plus 38 functional alleles sequenced in
Funct Integr Genomics

this study) were grouped into six populations based on theircountry of origin. In combined analysis, maximum number(160) of SNPs were obtained in the alleles of OTH categoryincluding isolates from Japan, the Philippines, Brazil, BurkinaFaso, and United States of America followed by the popula-tion of Japan (76), while highest number of InDels (175) andrecombination events (8) were found in Indian population.Total SNPs and InDels for the six populations analyzed in thisstudy were 232 and 62, respectively (Table 1).
To study the evolutionary dynamics of the Avr-Pita, pat-terns of nucleotide diversity in 133 Avr-Pita orthologues fromthe six populations of ten countries were examined with fourstatistical parameters, such as D statistics (Tajima's D test), D*and F* statistics (Fu and Li's test), and Fay and Wu's normal-ized H (FW-Hn) test (Table 2). The most statistical parame-ters’ values were negative, except for the populations of Japan
and USAwhich deviated from the neutrality model. Positivevalues of D, D*, and F* tests were obtained in case of thepopulation from Japan for coding and noncoding regions andalso the entire Avr-Pita gene. In case of the population fromUSA, positive values of these tests were obtained in codingregion of this gene. The most significant deviation fromneutrality model was observed in M. oryzae population fromThailand with negative value (−2.64665) of Tajima’s D testobtained at P<0.001 level of significance, while the positivevalue (1.59752) of this test was obtained in M. oryzae popu-lation of Japan. However, negative values for all above men-tioned neutrality statistics atP<0.02 level of significance werefound for the combined population (Table 2). Overall, thenucleotide diversity was higher in the coding regions of 133Avr-Pita alleles as compared to the noncoding regions of thesealleles. Average pairwise values for nucleotide diversity (π)and Watterson’s nucleotide diversity estimator (θw) of all thealleles were 0.01559 and 0.04627, respectively (Table 2).Among the Avr-Pita alleles of different geographical loca-tions, the nucleotide diversity was maximum in the alleles ofJapanese isolates (π=0.06161) followed by the isolates fromothers countries (π=0.04320). Among these six populations,the segregation sites (S) within the Avr-Pita locus were veryhigh (73.91 %) in OTH population category (Table 2).
The synonymous (Pisyn) and nonsynonymous (Pinon)substitutions and their ratios (Pinon/Pisyn) along with Juckesand Cantor’s corrections (Ks, Ka, and Ka/Ks) were calculatedfor 133 Avr-Pita alleles of the six populations (Table 3). Thelowest values for both the ratios, Pinon/Pisyn and Ka/Ks (JC),were obtained in the Indian isolates (0.055592 and 0.55249).The Ka/Ks ratios were more than 1 in case of Chinese andJapanese (1.08000 and 1.12059) populations of M. oryzae.The Ka/Ks value for the whole set of populations was lessthan 1 (0.77229). In all the populations, the value ofnonsynonymous substitutions (Pinon=0.01575) was less thanin synonymous substitutions (Pisyn=0.02025) in the entireregions of Avr-Pita alleles (Table 3).
In the present study, we found 85 haplogroups (H) of 133alleles belonging to six different populations including origi-nal sequences of Avr-Pita and Avr-Pita3 genes (Fig. 2).Maximum (221) connecting haplotypes were missing fromH19 to H85, and non-rice infecting isolates formed separate
Fig. 1 Reaction of different isolates of M. oryzae on monogenic line ofrice having Pita+ and Pita− genes. 1 and 2 reactions ofM. oryzae isolatesei2 and nwi28 virulent on Pita; 3 and 4 reactions of isolates ni4 and si74avirulent onmonogenic rice line IRBLTA-K1 containingPita gene; 5 and6 susceptible check Lijiang Xintuan Heigu (LTH-Pita)
Table 1 SNPs and InDels iden-tified in the Avr-Pita alleles ofM. oryzae populations
IND India; CHN China; JAPJapan; THD Thailand; USA Unit-ed State of America; OTH Othercountries (Philippines, Indonesia,Brazil, Colombia, and BurkinaFaso); ALL all analyzed Avr-Pitaalleles; n number of Avr-Pitaalleles
Population n Transitions Transversions Total SNPs Insertions/Deletions Recombinations
IND 41 22 11 33 175 8
CHN 12 11 1 12 6 1
JAP 7 42 34 76 4 0
THD 30 21 29 50 47 0
USA 32 15 9 24 121 7
OTH 11 98 62 160 17 7
ALL 133 136 96 232 62 16
Funct Integr Genomics
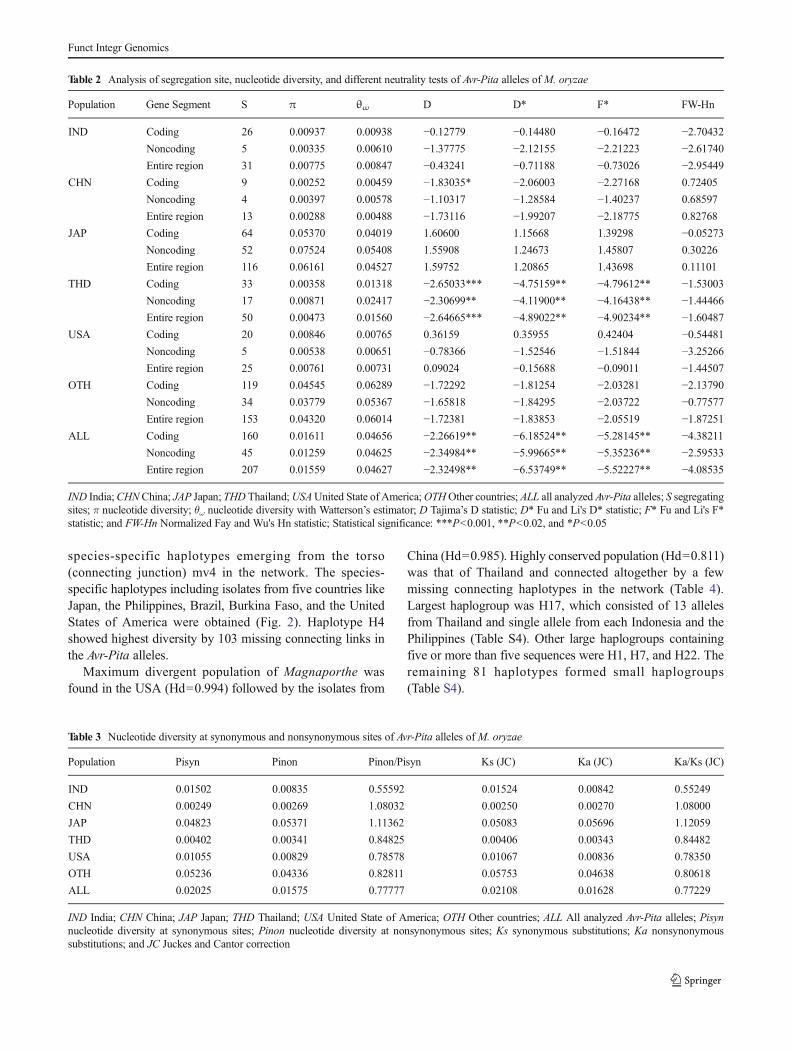
species-specific haplotypes emerging from the torso(connecting junction) mv4 in the network. The species-specific haplotypes including isolates from five countries likeJapan, the Philippines, Brazil, Burkina Faso, and the UnitedStates of America were obtained (Fig. 2). Haplotype H4showed highest diversity by 103 missing connecting links inthe Avr-Pita alleles.
Maximum divergent population of Magnaporthe wasfound in the USA (Hd=0.994) followed by the isolates from
China (Hd=0.985). Highly conserved population (Hd=0.811)was that of Thailand and connected altogether by a fewmissing connecting haplotypes in the network (Table 4).Largest haplogroup was H17, which consisted of 13 allelesfrom Thailand and single allele from each Indonesia and thePhilippines (Table S4). Other large haplogroups containingfive or more than five sequences were H1, H7, and H22. Theremaining 81 haplotypes formed small haplogroups(Table S4).
Table 2 Analysis of segregation site, nucleotide diversity, and different neutrality tests of Avr-Pita alleles ofM. oryzae
Population Gene Segment S π θω D D* F* FW-Hn
IND Coding 26 0.00937 0.00938 −0.12779 −0.14480 −0.16472 −2.70432Noncoding 5 0.00335 0.00610 −1.37775 −2.12155 −2.21223 −2.61740Entire region 31 0.00775 0.00847 −0.43241 −0.71188 −0.73026 −2.95449
CHN Coding 9 0.00252 0.00459 −1.83035* −2.06003 −2.27168 0.72405
Noncoding 4 0.00397 0.00578 −1.10317 −1.28584 −1.40237 0.68597
Entire region 13 0.00288 0.00488 −1.73116 −1.99207 −2.18775 0.82768
JAP Coding 64 0.05370 0.04019 1.60600 1.15668 1.39298 −0.05273Noncoding 52 0.07524 0.05408 1.55908 1.24673 1.45807 0.30226
Entire region 116 0.06161 0.04527 1.59752 1.20865 1.43698 0.11101
THD Coding 33 0.00358 0.01318 −2.65033*** −4.75159** −4.79612** −1.53003Noncoding 17 0.00871 0.02417 −2.30699** −4.11900** −4.16438** −1.44466Entire region 50 0.00473 0.01560 −2.64665*** −4.89022** −4.90234** −1.60487
USA Coding 20 0.00846 0.00765 0.36159 0.35955 0.42404 −0.54481Noncoding 5 0.00538 0.00651 −0.78366 −1.52546 −1.51844 −3.25266Entire region 25 0.00761 0.00731 0.09024 −0.15688 −0.09011 −1.44507
OTH Coding 119 0.04545 0.06289 −1.72292 −1.81254 −2.03281 −2.13790Noncoding 34 0.03779 0.05367 −1.65818 −1.84295 −2.03722 −0.77577Entire region 153 0.04320 0.06014 −1.72381 −1.83853 −2.05519 −1.87251
ALL Coding 160 0.01611 0.04656 −2.26619** −6.18524** −5.28145** −4.38211Noncoding 45 0.01259 0.04625 −2.34984** −5.99665** −5.35236** −2.59533Entire region 207 0.01559 0.04627 −2.32498** −6.53749** −5.52227** −4.08535
IND India;CHNChina; JAP Japan; THD Thailand;USAUnited State of America;OTHOther countries; ALL all analyzed Avr-Pita alleles; S segregatingsites; π nucleotide diversity; θω nucleotide diversity with Watterson’s estimator; D Tajima’s D statistic; D* Fu and Li's D* statistic; F* Fu and Li's F*statistic; and FW-Hn Normalized Fay and Wu's Hn statistic; Statistical significance: ***P<0.001, **P<0.02, and *P<0.05
Table 3 Nucleotide diversity at synonymous and nonsynonymous sites of Avr-Pita alleles of M. oryzae
Population Pisyn Pinon Pinon/Pisyn Ks (JC) Ka (JC) Ka/Ks (JC)
IND 0.01502 0.00835 0.55592 0.01524 0.00842 0.55249
CHN 0.00249 0.00269 1.08032 0.00250 0.00270 1.08000
JAP 0.04823 0.05371 1.11362 0.05083 0.05696 1.12059
THD 0.00402 0.00341 0.84825 0.00406 0.00343 0.84482
USA 0.01055 0.00829 0.78578 0.01067 0.00836 0.78350
OTH 0.05236 0.04336 0.82811 0.05753 0.04638 0.80618
ALL 0.02025 0.01575 0.77777 0.02108 0.01628 0.77229
IND India; CHN China; JAP Japan; THD Thailand; USA United State of America; OTH Other countries; ALL All analyzed Avr-Pita alleles; Pisynnucleotide diversity at synonymous sites; Pinon nucleotide diversity at nonsynonymous sites; Ks synonymous substitutions; Ka nonsynonymoussubstitutions; and JC Juckes and Cantor correction
Funct Integr Genomics

Amino acid diversification of Avr-Pita alleles
Thirty-eight protein sequences translated from the Avr-Pitaalleles of avirulent isolates were used for the amino aciddiversification analysis. Firstly, all the amino acid sequenceswere aligned and then compared with the original Avr-Pitaprotein (AF207841). A total of 22 protein variants wereidentified (Table 5). However, no significant change wasfound in the zinc protease motif (ten amino acid long motif)of all the protein variants except for a single amino acidchange found at 173rd position (valine-isoleucine) in the nineprotein variants.
Pot3 insertion in Avr-Pita allele of virulent isolates
Out of 80 Avr-Pita alleles sequenced in the present study, 42were virulent on blast resistance gene Pita. Of these, 37 alleleshad the code for the proteins identical to that of original Avr-
Fig. 2 Haplotype network of Avr-Pita alleles. Avr-Pita3 was used asoutgroup. Size of the circle reflects proportional number of sequences anddifferent colors indicate different populations of M. oryzae. H haplotype;
mv shows connecting junction. More than 1 missing haplotypes are givenas number on the connecting links
Table 4 Haplotype diversity at Avr-Pita locus in M. oryzae populations
Population Hn Hd V (Hd) SD (Hd)
IND 26 0.971 0.00017 0.013
CHN 11 0.985 0.00162 0.040
JAP 6 0.952 0.00912 0.096
THD 15 0.811 0.00511 0.071
USA 30 0.994 0.00011 0.011
OTH 10 0.982 0.00215 0.046
ALL 85 0.980 0.00003 0.006
IND India; CHN China; JAP Japan; THD Thailand; USA United State ofAmerica; OTH Other countries; ALL All analyzed Avr-Pita alleles plusoriginal Avr-Pita and Avr-Pita3 genes; Hn Number of Haplotypes; HdHaplotype diversity; V (Hd) Variance of Haplotype diversity; and SD(Hd) Standard deviation of Haplotype diversity
Funct Integr Genomics
Pita gene. These 37 isolates showing virulence on resistancegene Pita were analyzed for the possible insertions of Pot3element in their promoter region. Pot3 insertion was identifiedin the promoter region of 29 virulent isolates (Fig. 3a).However, the remaining eight virulent isolates (ei26, ei27,ei40, nwi50, nwi209, ni25, ni26, and si68) did not havePot3 insertions in the promoter region or other regions of thegene.
Development of allele specific functional markers
Two allele-specific markers (ASMs), TRSpot3-1 andTRSpot3-2, were successfully developed and validated inthe field isolates of M. oryzae for detecting allele specificityof Avr-Pita gene (Fig. 3b). The Pot3 fragment of virulent Avr-Pita alleles was amplified (1,560 bp amplicon) from 29 iso-lates ofM. oryzae using the ASMs. The virulence analysis ofM. oryzae isolates with and without Pot3 element in thepromoter region of Avr-Pita gene was observed on the basisof rice blast disease reactions on the Pita containing mono-genic rice line IRBLTA-K1 (Fig. 3c).
Discussion
The rice M. oryzae pathosystem is considered as a model forthe study of host-pathogen interactions (Correll et al. 2000).Many advances have been made in the recent years in the areaof genetics and molecular biology ofM. oryzae (Talbot 1995).The molecular understanding of how the Avr gene triggersresistance in host plant should lead to the development ofnovel strategies for broad-spectrum and durable disease resis-tance in a particular geographical area (Bryan et al. 2000). ThePita gene is one of the most effective R genes deployed forrice blast resistance (Orbach et al. 2000). The effectiveness ofthis gene relies on its ability to recognize the pathogen’scorresponding avirulence gene (Avr-Pita). The Pita-mediatedresistance response is because of the physical and directinteraction between Avr-Pita protein and Pita protein (Bryanet al. 2000; Jia et al. 2000). However, the high genomeplasticity of M. oryzae enables it to rapidly evolve new viru-lent races in the field that curtail the useful life span of newlyreleased resistant cultivars (Skamnioti and Gurr 2009).Several studies demonstrated that partial deletion, complete
Table 5 New functional Avr-Pita protein variants identified in M. oryzae population
Protein variantsa No. of isolateb Position of amino acids
5c 74 81 82 87 88 98 103 127 135 154 158 168 173 191 193
Avr-Pitad n.a. - W N D R L Q K N G P N G V Y W
PV1 6 L R N D R L Q K N G R N G V Y W
PV2 2 L W N D R L Q K N G P K V V Y W
PV3 1 L R N D K L Q N N G P N G I C W
PV4 2 L W N D K S Q K N G R N V V Y W
PV5 2 L W S N K S Q N N E P N G I Y W
PV6 1 L W N D E L Q K N E P N G V Y W
PV7 1 L W S N K L Q N N E P N G I Y W
PV8 1 L W N D K L Q K N G R N G V Y W
PV9 1 L W N D E L R K N G R N F V Y W
PV10 1 L W N D E L Q N D E R N G V Y W
PV11 2 L W S D R L Q N N E P N G I Y W
PV12 2 L W N D K L Q K N G R K R V Y W
PV13 2 L W S N K L Q N N G P N G I C W
PV14 1 L W N D R L Q K N G P N G I C W
PV15 1 L W S N K L Q N N E P N G I C W
PV16 1 L W S N K L Q N N G P N G I Y W
PV17 1 L W N D K S Q K N G P H G V Y W
PV18 1 L W N D K L Q K N G P N G V Y W
PV19 4 L W N D R L Q K N G P N V V Y W
PV20 2 L W N D R L Q K N G P N G V C W
PV21 1 L W S N K L Q N N E P N G I Y R
PV22 2 L W N D R L Q K N G P N G V C W
aGroups of Avr-Pita variants based on amino acid sequences; b Number of isolates from which the respective Avr-Pita protein variant was identified;C Leucine insertion after 5th amino acid; dOriginal Avr-Pita protein; n.a Not applicable
Funct Integr Genomics

deletion, frame-shift mutations, and sequence variation in thepromoter region of the Avr-Pita sequences of M. oryzae iso-lates resulted in the loss of avirulence function of the gene(Dai et al. 2010; Kasetsomboon et al. 2012).
The significant negative values observed for all the neu-trality tests in the combined country-wise analysis (D, D*, F,and FW-Hn) suggested that the Avr-Pita gene has been sub-jected to purifying selection in all the worldwide populationsof Magnaporthe and is still in expansion mode. The value ofD test for Thai isolates deviated at greater extent from theneutrality and suggested that these isolates have been exposedto greater selection pressure in nature to maintain their favor-able Avr-Pita alleles by removing the deleterious mutations.Similar divergence patterns have also been reported for Thaiand Chinese isolates with significant deviation from neutralityat the Avr-Pita locus and suggested that they were possiblyexposed to a higher level of selection pressure in nature(Kasetsomboon et al. 2012).
Species-specific haplotypes formed by non-rice infectingisolates of different countries with maximum torso(connecting junction) linked together show their evolutionaryevidence by following the highest missing connecting haplo-types from Avr-Pita3. Couch et al. (2005) clearly distin-guished the two lineages of Magnaporthe spp. intoM. oryzae andM. grisea using molecular analysis and report-ed host-specific lineages of Magnaporthe spp.
Two predominant hypotheses have been proposed to ex-plain the coevolution of R and Avr genes in different host-pathogen systems, the “arms race” and the “trench warfare”hypotheses (Dai et al. 2010). Many researchers have reportedthe low level of diversity at Pita locus (Bryan et al. 2000; Jia
et al. 2000; Jia et al. 2003; Huang et al. 2008; Lee et al.2009a, b), in contrast to a high allelic diversity observed atAvr-Pita locus (Dai et al. 2010; Chuma et al. 2011;Kasetsomboon et al. 2012). These results clearly supportedthe “trench warfare” hypothesis over the “arms race” theoryto explain the R-Avr coevolution in the rice blast pathosystem.In the present study, we have recorded a high variability at theAvr-Pita locus. In contrast, we have recorded a low level ofpolymorphism at the Pita locus among the rice landracescollected from different geographic regions of India (Thakuret al. 2013). Extensive surveys of rice germplasm includingIndian germplasm collections have indicated a low level ofvariation at Pita locus (Wang et al. 2008; Huang et al. 2008;Lee et al. 2009a). These results also favor trench-warfaretheory to explain the coevolution of Avr-Pita and Pita underIndian conditions. However, Lee et al. (2009a) and Jia (2009)reported that the Pita gene has evolved under extensiveselection pressure during rice crop breeding for resistance tosupport the “arms race” hypothesis. In addition, recent iden-tification of another critical gene Ptr(t) that is required forPita-mediated resistance (Jia and Martin 2008) and presenceof a transposon elements at the promoter region of Pita (Leeet al. 2009a) in resistant rice germplasm has suggesteduncovering of the evolutionary dynamics of Pita and Avr-Pita genes and is solely based on the sequence variation atthese two loci. It is an over simplification and shall requireanalysis of additional cooperating genes. An examination ofphysical interactions among Pita, Avr-Pita, and Ptr(t) mayactually reveal the facts about rice blast disease resistance andthe coevolution pattern of Avr-Pita: Pita in rice-M. oryzaehost-pathogen system.
Fig. 3 Pot3 transposable element insertion in promoter region of Avr-Pita gene; a a graphical representation of Pot3 element insertion innonfunctional Avr-Pita allele; b PCR amplification of Avr-Pita alleleswith Pot3 element insertion in the promoter region from 12 virulentisolates ofM. oryzae using a primer pair Pot3-1. Lanes 1 to 12 represent
amplification products from M. oryzae isolates with Pot3 element andlanes 13 to 16 correspond to isolates without Pot3 insertion. cReaction ofM. oryzae isolates containing Pot3 insertions (1–12) and without Pot3insertions (13–16) on rice line IRBLTA-K1 (Pita+). Rice line LTH(Pita−) used as a susceptible control (17)
Funct Integr Genomics

In the present study, high nucleotide variation was ob-served in the Avr-Pita alleles of M. oryzae isolates. Thededuced amino acid sequences of the 37 alleles from virulentisolates were similar to the original Avr-Pita gene. Since 29 ofthese isolates were shown to have Pot3 insertion in the pro-moter region of their Avr-Pita gene, the loss of function of thepromoter seems to be the most plausible reason for the loss ofAvr-Pita-mediated avirulence. The insertion of Pot3 elementwas, however, not identified in the remaining eight virulentisolates that have Avr-Pita alleles identical to the original Avr-Pita gene. The virulence of these isolates on Pita gene may bedue to some other uncharacterized structural changes or inser-tion of other transposable elements in the promoter region ofAvr-Pita gene. The transposable elements are reported to playan important role in the regulation of gene action. Apart fromPot3, several other transposable elements like Inago1, Inago2,and Pyret (Nakayashiki et al. 2001); MGLR3 (Kang 2001);MGR583 (Hamer et al. 1989; Kachroo et al. 1997); Pot 2(Kachroo et al. 1994a, b); and Occan (Kito et al. 2003) havebeen identified in the flanking regions of Avr-Pita homologs(Chuma et al. 2011). It is believed that the ubiquitous distri-bution of Pot3 elements throughout the genome ofM. oryzaewas responsible for changing the functions of many genes.Similar insertion of Pot3 elements in the promoter of Avr-Pitagene has been identified and suggested to be one of thereasons for change in the virulence spectrum of M. oryzae(Kang et al. 2001). In another study, the Pot3 insertion hasalso been reported in the coding region of the alleles ofvirulent isolate (Dai et al. 2010). Similarly, the genetic chang-es caused by transposons have also been associated with racevariation in M. oryzae (Shull and Hamer 1994).
For effective deployment of Pita-based resistant cultivars,it is imperative to assess the distribution of functional allele ofthe Avr-Pita gene in target M. oryzae populations. Therefore,two allele-specific markers (ASMs) were developed and val-idated on a set of virulent and avirulent isolates ofM. oryzae.These markers were able to distinguish functional as well asnonfunctional Avr-Pita alleles in the M. oryzae populations.Several molecular markers have been successfully utilized inthe M. oryzae populations by different researchers for variedapplications (Lebrun et al. 1990; Borromeo et al. 1993). Thefunctional markers developed in the present study would beimportant to analyze M. oryzae populations in different ricegrowing regions.
We can conclude that a high level of nucleotide sequencepolymorphism was detected especially in the coding regionsof Avr-Pita gene among the worldwide collection ofM. oryzaealleles. We showed that the Avr-Pita gene has been subjectedto a natural purifying directional selection. The high allelicdiversity observed in this study for Avr-Pita is clearly sugges-tive of an ongoing “trench warfare” in rice blast pathosystem.Our results suggest that inactivation of Avr-Pita by Pot3insertion would be one of the mechanisms responsible for
the loss of Pita-mediated resistance in rice. Two allele-specific markers targeting nonfunctional Avr-Pita alleles weredeveloped in the present study and were found useful indistinguishing different Pita compatible isolates of the patho-gen. These markers can be used to assess the distribution offunctional alleles of the Avr-Pita gene in the target M. oryzaepopulations for effective deployment of Pita-mediatedresistance.
Acknowledgments The financial assistance received from NationalAgricultural Innovation Project (NAIP) (C4/C1071), ICAR, by TRSharma is gratefully acknowledged.
Conflict of interests The authors have declared that no conflict ofinterests exists.
Author’s contributions Conceived and designed the experiments andwrote final manuscript: TRS. Performed the experiments and analyzeddata: PKS. Wrote first draft of the manuscript: PKS, RR. Contributedmaterials/analysis tools: ST, MV, SKP, VS, AKS, UD, and NKS.
References
Bonman JM, Vergel De Dios TI, Khin MM (1986) Physiologic special-ization of Pyricularia oryzae in the Philippines. Plant Dis, St Paul70:767–769
Borromeo ES, Nelson RJ, Bonman JM, Leung H (1993) Genetic differ-entiation among isolates of Pyricularia infecting rice and weedhosts. Phytopathology 83:393–399
Bryan BT, Wu KS, Farrall L, Jia Y, Hershey H, McAdams S, Tarchini R,Donaldson G, Faulk K, Valent B (2000) A single amino aciddifference distinguishes resistant and susceptible allele of the riceblast resistance gene Pi-ta. Plant Cell 12:2033–2045
Chuma I, Isobe C, Hotta Y, Ibaragi K, Futamata N, Motoaki K, Yoshida K,Terauchi R, Fujita Y, Nakayashiki H, Valent B, Tosa Y (2011)Multiple translocation of the AVR-Pita1 effector gene among chro-mosomes of the rice blast fungus Magnaporthe oryzae and relatedspecies. PLoS Pathog 7:e1002147. doi:10.1371/journal.ppat.1002147
Clement M, Posada D, Crandall KA (2000) TCS: a computer program toestimate gene genealogies. Mol Ecol 9:1657–1660
Correll JC, Harp TL, Guerber JC, Zeigler RS, Liu B, Cartwright RD, LeeFN (2000) Characterization of Pyriculariagrisea in the UnitedStates using independent genetic and molecular markers.Phytopathology 90:1396–1404
Couch BC, Fudal I, Lebrun M-H, Tharreau D, Valent B, Kim P,Notteghem J-L, Kohn LM (2005) Origins of host-specific popula-tions of the blast pathogenMagnaporthe oryzae in crop domestica-tion with subsequent expansion of pandemic clones on rice andweeds of rice. Genetics 170:613–630
Dai Y, Jia Y, Correll J, Wang X, Wang Y (2010) Diversification andevolution of the avirulence gene AVR-Pita1 in field isolates ofMagnaporthe oryzae. Fungal Genet Biol 37:973–980
Dodds PN, Lawrence GJ, Catanzariti AM, Teh T, Wang CIA, AyliffeMA, Kobe B, Ellis JG (2006) Direct protein interaction underliesgene-for-gene specificity and coevolution of the flax resistancegenes and flax rust avirulence genes. Proc Natl Acad Sci U S A103:8888–8893
FarmanML, Leong SA (1998) Chromosome walking to the AVR1-CO39avirulence gene of Magnaporthe grisea: Discrepancy between thephysical and genetic maps. Genetics 150:1049–1058
Funct Integr Genomics

Fudal I, Bohnert HU, Tharreau D, Lebrun MH (2005) Transposition ofMINE, a composite retrotransposon, in the avirulence gene ACE1 ofthe rice blast fungus Magnaporthe grisea. Fungal Genet Biol 42:761–772
Hamer JE, Farrall L, Orbach MJ, Valent B, Chumley FG (1989) Hostspecies-specific conservation of a family of repeated DNA se-quences in the genome of a fungal plant pathogen. Proc Natl AcadSci U S A 86:9981–9985
Huang CL, Hwang SY, Chiang YC, Lin TP (2008) Molecular evolutionof thePi-ta gene resistant to rice blast inwild rice (Oryza rufipogon).Genetics 179:1527–1538
Jia Y (2009) Artificial introgression of a large fragment around the Pi-tarice blast resistance gene in backcross progenies and several eliterice cultivars. Heredity 103:333–339
Jia Y, Martin R (2008) Identification of a new locus, Ptr(t), required forrice blast resistance gene Pi-ta-mediated resistance. Mol PlantMicrobe Interact 21:396–403
Jia Y, McAdams SA, Bryan GT, Hershey HP, Valent B (2000) Directinteraction of resistance gene and avirulence gene products confersrice blast resistance. EMBO J 19:4004–4014
Jia Y, Bryan GT, Farrall L, Valent B (2003) Natural variation at the Pi-tarice blast resistance locus. Phytopathology 93:1452–1459
Joosten MHAJ, Cozijnsen AJ, de Wit PJGM (1994) Host resistance to afungal tomato pathogen lost by a single base-pair change in anavirulence gene. Nature 367:384–386
Kachroo P, Chattoo BB, Leong SA (1994a) Pot2, an inverted repeattransposon fromMagnaporthe grisea. Mol GenGenet 245:339–348
Kachroo P, Leong SA, Chattoo BB (1994b) Pot2, an inverted repeattransposon from the rice blast fungus Magnaporthe grisea. MolGen Genet 245:339–348
Kachroo P, Ahuja M, Leong SA, Chattoo BB (1997) Organization andmolecular analysis of repeated DNA sequences in the rice blastfungus Magnaporthe grisea. Curr Genet 31:361–369
Kang S (2001) Organization and distribution pattern ofMGLR-3, a novelretrotransposon in the rice blast fungusMagnaporthe grisea. FungalGenet Biol 32:11–19
Kang S, Sweigard JA, Valent B (1995) The PWL host specificity genefamily in the blast fungus Magnaporthe grisea. Mol Plant MicrobeInteract 8:939–948
Kang S, LebrunM-H, Farrall L, Valent B (2001) Gain of virulence causedby insertion of a Pot3 transposon in a Magnaporthe griseaavirulence gene. Mol Plant-Microbe Interact 14:671–674
Kasetsomboon T, Kate-Ngam S, Sriwongchai T, ZhouB, Jantasuriyarat C(2012) Sequence variation of avirulence gene AVR-Pita1 in riceblast fungus, Magnaporthe oryzae. Mycol Prog. doi:10.1007/s11557-012-0867-1
Khang CH, Park S-Y, Lee Y-H, Valent B, Kang S (2008) Genomeorganization and evolution of the AVR-Pita avirulence gene familyin the Magnaporthe grisea species complex. Mol Plant-MicrobeInteract 21:658–670
Kito H, Takahashi Y, Sato J, Fukiya S, Sone T, Tomita F (2003) Occan, anovel transposon in the Fot1 family, is ubiquitously found in severalMagnaporthe grisea isolates. Curr Genet 42:322–331
Lebrun MH, Capy MP, Garcia N, Detertra M, Brygoo Y, Notteghem JL,Vales M (1990) In Proceedings of the Rice Genetics Conference.IRRI, Philippines
Lee S, Costanzo S, Jia Y, Olsen KM, Caicedo AL (2009a)Evolutionary dynamics of the genomic region around the blastresistance gene Pi-ta in AA genome Oryza species. Genetics 183:1315–1325
Lee SK, Song MY, Seo YS, Kim HK, Ko S, Cao PJ, Suh JP, Yi G, RohJH, Lee S, An G, Hahn TR, Wang GL, Ronald P, Jeon JS (2009b)Rice Pi5-mediated resistance to Magnaporthe oryzae requires thepresence of two coiled-coil-nucleotide-binding-leucine-rich repeatgenes. Genetics 181:1627–1638
Li W, Wang B, Wu J, Lu G, Hu Y, Zhang X, Zhang Z, Zhao Q, Feng Q,Zhang H, Wang Z, Wang GL, Han B, Wang Z, Zhou B (2009) TheMagnaporthe oryzae avirulence gene AvrPiz-t encodes a predictedsecreted protein that triggers the immunity in rice mediated by theblast resistance gene Piz-t. Mol Plant Microbe Interact 22:411–420
Ma JF, Tamai K, Yamaji N, Mitani N, Konishi S, Katsuhara M, IshiguroM, Murata Y, Yano M (2006) A silicon transporter in rice. Nature440:688–691
Mackill DJ, Bonman JM (1992) Inheritance of near-isogenic lines of rice.Phytopathology 82:746–749
Mathur RL, Mathure BL, Bhatanagar GC (1960) Blackening of bajra(PennisetumtyphoideaStapf and Habbard) grains in ear headscaused by Curvularialunata (Wakk). Boed. Syn. C. Pennisiti(Mitra). Boed. Proc Natl Acad Sci India 30:323–330
Murray MG, Thompson WF (1980) Rapid isolation of high molecularweight plant DNA. Nucleic Acids Res 8:4321–4325
Nakayashiki H, Matsuo H, Chuma I, Ikeda K, Betsuyaku S et al (2001)Pyret, a Ty3/gypsy retrotransposon inMagnaporthe grisea containsan extra domain between the nucleocapsid and protease domains.Nucleic Acids Res 29:4106–4113
Orbach MJ, Farrall L, Sweigard J, Chumley FG, Valent B (2000) Atelomeric avirulence gene determines efficacy for the rice blastresistance gene Pi-ta. Plant Cell 12:2019–2032
Ou SH (1985) Rice Diseases. CAB International 380 pRidout CJ, Skamnioti P, Porritt O, Sacristan S, Jones JDG, Brown JKM
(2006) Multiple avirulence paralogues in cereal powdery mildewfungi may contribute to parasite fitness and defeat of plant resis-tance. Plant Cell 18:2402–2414
Sharma TR, Rai AK, Gupta SK, Vijayan J, Devanna BN, Ray S (2012)Rice blast management through host resistance: Retrospect andprospects. Agric Sci 1:37–52
Shull V, Hamer JE (1994) Genomic structure and variability inPyriculariagrisea. In: Rice Blast Disease, Zeigler RS, Teng PS,Leong SA (eds) CAB International.Wallingford, England, pp 65–86
Silue D, Notteghem JL, Tharreau D (1992) Evidence for a gene for generelationship in the Oryza sativa Magnaporthe grisea pathosystem.Phytopathology 82:577–582
Skamnioti P, Gurr SJ (2009) Against the grain: Safe guarding rice fromrice blast disease. Trends Biotechnol 27:141–150
Sweigard JA (1995) Identification, cloning, and characterization ofPWL2, a gene for host species specificity in the rice blast fungus.Plant Cell 7:1221–1233
Talbot NJ (1995) Having a blast: Exploring the pathogenicity ofMagnaporthe grisea. Trends Microbiol 3:9–15
Thakur S, Gupta YK, Singh PK, Rathour R, Variar M, Prashanthi SK,Singh AK, Singh UD, Chand D, Rana JC, Singh NK, Sharma TR(2013) Molecular diversity in rice blast resistance gene Pi-ta makesit highly effective against dynamic population of Magnaportheoryzae. Funct Integr Genomics. doi:10.1007/s10142-013-0325-4
Untergrasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, RemmM,Rozen SG (2012) Primer3—New capabilities and interfaces.Nucleic Acids Res 40(15):e115
Valent B, Chumley FG (1991)Molecular genetic analysis of the rice blastfungus, Magnaporthe grisea. Annu Rev Phytopathol l29:443–463
Valent B, Crawford MS, Weaver CG, Chumley FG (1986) Geneticstudies of pathogenicity and fertility of Magnaporthe grisea. IowaState J Res 60:569–594
Van de Wouw AP, Cozijnsen AJ, Hane JK, Brunner PC, McDonald BAet al (2010) Evolution of linked avirulence effectors inLeptosphaeria maculans is affected by genomic environment andexposure to resistance genes in host plants. PLoS Pathog 6:e1001180
WangX, Jia Y, Shu Q,WuD (2008) Haplotype diversity at the Pi-ta locusin cultivated rice and its wild relatives. Phytopathology 98:1305–1311
Funct Integr Genomics

YamadaM (1985) Pathogenic specialization of rice blast fungus in Japan.Jpn Agric Res Q 19:178–183
Yoshida K, Saitoh H, Fujisawa S, Kanzaki H, Matsumura H, Yoshida K,Tosa Y, Chuma L, Takano Y, Win J, Kamoun S, Terauchi R (2009)Association genetics reveals three novel avirulence genes from the
rice blast fungal pathogen Magnaporthe oryzae. Plant Cell 21:1573–1591
Zhou E, Jia Y, Correll J, Lee FN (2007) Instability of the Magnaportheoryzae avirulence geneAVR-Pita alters virulence. Fungal Genet Biol44:1024–1034
Funct Integr Genomics
Related Documents



![circRNAs Are Involved in the Rice-Magnaporthe oryzae … · circRNAs Are Involved in the Rice-Magnaporthe oryzae Interaction1[OPEN] Jing Fan,a,2 Weili Quan,b,2 Guo-Bang Li,a,2 Xiao-Hong](https://static.cupdf.com/doc/110x72/609760baf9f98337ca16efe8/circrnas-are-involved-in-the-rice-magnaporthe-oryzae-circrnas-are-involved-in-the.jpg)







