Transposable elements and G-quadruplexes Eduard Kejnovsky 1* , Viktor Tokan 1 , and Matej Lexa 2 1 Department of Plant Developmental Genetics, Institute of Biophysics, Academy of Sciences of the Czech Republic, Brno, Czech Republic 2 Faculty of Informatics, Masaryk University, Brno, Czech Republic * To whom correspondence should be addressed: +420 541517203, email: [email protected] Keywords: transposable elements, LTR retrotransposons, DNA and RNA quadruplexes, G- quadruplexes, transcription, recombination, replication Abbreviations: TEs, transposable elements; LTR, long terminal repeat; PQS, potential quadruplex-forming sequence; Abstract A significant part of eukaryotic genomes is formed by transposable elements (TEs) containing not only genes but also regulatory sequences. Some of the regulatory sequences located within TEs can form secondary structures like hairpins or three-stranded (triplex DNA) and four-stranded (quadruplex DNA) conformations. This review focuses on recent evidence showing that G-quadruplex-forming sequences in particular are often present in specific parts of TEs in plants and humans. We discuss the potential role of these structures in the TE life- cycle as well as the impact of G-quadruplexes on replication, transcription, translation, chromatin status and recombination. The aim of this review is to emphasize that TEs may serve as vehicles for the genomic spread of G-quadruplexes. These non-canonical DNA structures and their conformational switches may constitute another regulatory system that, together with small and long non-coding RNA molecules and proteins, contribute to the complex cellular network resulting in the large diversity of eukaryotes. Introduction The currently emerging paradigm presents the genome as a dynamic system generating its own rearrangements. The most dynamic genome components are the transposable elements (TEs) - discrete segments of DNA that can move from one position to another in the genome, either by a “copy and paste” or a “cut and paste” mechanism. Previously presented as “junk DNA” TEs are now considered as important factors of genome dynamics giving the genomes evolutionary potential (Biémont and Vieira 2006). High genome dynamic is more important for sessile organisms like plants and this may explain why TEs are activated by environmental stresses in such species. The composition of various TE families differs in various taxa. For example, long terminal repeat (LTR) retrotransposons are the dominating species of TEs in plants while non-LTR retrotransposons are more abundant in mammals. Regulatory regions of transposable elements The main types of transposable elements differ in sequence but more especially in global structure – the presence and localization of genes and regulatory regions (Wicker et al. 2007). For example, LTR retrotransposons contain GAG and POL genes (and ENV gene in retroviruses) and also several regulatory sequences. They include promoters that are localized in LTRs and the transcription of the whole element starts here. Other regulatory regions are

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Transposable elements and G-quadruplexes
Eduard Kejnovsky1*, Viktor Tokan1, and Matej Lexa2
1Department of Plant Developmental Genetics, Institute of Biophysics, Academy of Sciencesof the Czech Republic, Brno, Czech Republic2Faculty of Informatics, Masaryk University, Brno, Czech Republic
* To whom correspondence should be addressed: +420 541517203, email: [email protected]
Keywords: transposable elements, LTR retrotransposons, DNA and RNA quadruplexes, G-quadruplexes, transcription, recombination, replication
Abbreviations: TEs, transposable elements; LTR, long terminal repeat; PQS, potential quadruplex-forming sequence;
AbstractA significant part of eukaryotic genomes is formed by transposable elements (TEs) containingnot only genes but also regulatory sequences. Some of the regulatory sequences locatedwithin TEs can form secondary structures like hairpins or three-stranded (triplex DNA) andfour-stranded (quadruplex DNA) conformations. This review focuses on recent evidenceshowing that G-quadruplex-forming sequences in particular are often present in specific partsof TEs in plants and humans. We discuss the potential role of these structures in the TE life-cycle as well as the impact of G-quadruplexes on replication, transcription, translation,chromatin status and recombination. The aim of this review is to emphasize that TEs mayserve as vehicles for the genomic spread of G-quadruplexes. These non-canonical DNAstructures and their conformational switches may constitute another regulatory system that,together with small and long non-coding RNA molecules and proteins, contribute to thecomplex cellular network resulting in the large diversity of eukaryotes.
IntroductionThe currently emerging paradigm presents the genome as a dynamic system generating itsown rearrangements. The most dynamic genome components are the transposable elements(TEs) - discrete segments of DNA that can move from one position to another in the genome,either by a “copy and paste” or a “cut and paste” mechanism. Previously presented as “junkDNA” TEs are now considered as important factors of genome dynamics giving the genomesevolutionary potential (Biémont and Vieira 2006). High genome dynamic is more importantfor sessile organisms like plants and this may explain why TEs are activated by environmentalstresses in such species. The composition of various TE families differs in various taxa. Forexample, long terminal repeat (LTR) retrotransposons are the dominating species of TEs inplants while non-LTR retrotransposons are more abundant in mammals.
Regulatory regions of transposable elementsThe main types of transposable elements differ in sequence but more especially in globalstructure – the presence and localization of genes and regulatory regions (Wicker et al. 2007).For example, LTR retrotransposons contain GAG and POL genes (and ENV gene inretroviruses) and also several regulatory sequences. They include promoters that are localizedin LTRs and the transcription of the whole element starts here. Other regulatory regions are

the primer-binding site (PBS) and polypurine tract (PPT) where reverse transcription of thefirst and the second strands , respectively, starts (Sabot and Schulman 2006).
Retroviruses, which are very similar to LTR retrotransposons, have additional signalsequences, e.g. a packaging signal sequence (psi element), located downstream from the PBSsite that is composed of four stem-loop sequences known to facilitate the dimerization of twoRNA genomes and allow packaging of RNA into viral cores (Lever et al. 1989; Lawrence etal. 2003). Moreover, a G-rich sequence encompassing the 5’ end of GAG gene in HIV RNAforms a G-quadruplex that also helps dimerization of retroviral RNA genomes (Sundquist andHeaphy 1993). Plant LTR retrotransposons often contain extra open reading frames(Steinbauerová et al. 2011) and their 3’UTR regions (untranslated region located between polgene and the right LTR) sometimes harbor satellite repeats (Macas et al. 2009). DNAtransposons, either autonomous (e.g. Mariner or Mutator elements) or their non-autonomouscounterparts (such as MITE elements, miniature inverted-repeat transposable elements)contain terminal inverted repeats (TIRs) that also have the potential to form hairpins (Bureauand Wessler 1992). As a consequence, microRNA or siRNA can be derived from MITEelements (Piriyapongsa and Jordan 2008).
Sequences adopting multi-stranded structures found inside transposable elementsThe right-handed double helix of B-DNA has been known since 1953 (Watson and Crick2003) but it has become increasingly clear that DNA can adopt a variety of alternativeconformations depending on its sequence motifs, ionic environment and interactions withproteins. These structures include three-stranded or four-stranded conformations – triplexes orG-quadruplexes (Rich 1993). In addition to Watson-Crick base pairing, Hoogsteen basepairing is involved here. Triplexes are often formed in homopurine/homopyrimidine stretchesand triplets consist of Py-Pu-Py or Pu-Pu-Py while G-quadruplexes are formed in DNA orRNA regions containing four continuous runs of three or more guanines, separated by loops ofvariable length. Guanines form a square planar structure called a guanine tetrad associatedthrough Hoogsteen base pairing and stabilized by cations, especially potassium (Bochman etal. 2012). G-quadruplexes adopt a variety of topologies (Figure 1) depending on theorientation (parallel, antiparallel or 3:1 hybrid) or number of involved strands i.e.intramolecular (one strand) or intermolecular (bi or tetra molecular). It has been recentlyproposed that quadruplexes progress through intermediate states such as G-hairpin and G-triplex (Rajendran et al. 2014, Cerofolini et al. 2014).
The first reports, to our knowledge, demonstrating the presence of G-quadruplexes intransposable elements, come from the 1990s. The first study showed a potassium-dependentblock of primer extension at the G-rich polypurine tract present in the 3’UTR of L1 elements(LINE-1, long interspersed nuclear element) that was assigned to the G-quadruplex formation(Howell and Usdin 1997). Another study demonstrated that the 3’ non-coding region ofdrosophila HeT-A telomeric non-LTR retrotransposon, contains sequences with the propensityto form G-quadruplex DNA in vitro (Abad and Villasante 1999). The G-quadruplex-formingregion of HeT-A may thus structurally behave as the telomeric repeats common to themajority of eukaryotes.
Recent large-scale analyses showed that transposable elements contain sequences with thepotential to form multi-stranded DNA structures (Figure 2). Potential G-quadruplex- andtriplex-forming sequences (PQS and PTS) were found inside plant LTR retrotransposons(Lexa et al. 2014a) and in human HERV, LINE and SVA elements (Savage et al. 2013; Lexa etal. 2014b). G-quadruplexes have been found in all known human papillomaviruses (Tlučková

et al. 2013) and in U3 regions of the HIV-1 virus (Piekna-Przybylska et al. 2014). Moreover,Sireviruses contain four polypurine tracts in their 3’UTRs that could be potential regions oftriplex DNA formation (Bousios et al. 2010).
In plant retrotransposons, potential G-quadruplex sequences (PQS) have been found in longterminal repeats (LTRs) at specific distances from transcription start sites and often formeddoublets separated by about 400 bp (Lexa et al. 2014a). Longer guanine runs that more readilyadopt G-quadruplexes were more often found in recent retrotransposons, suggesting afunction in the TE life cycle. Circular dichroism spectroscopy confirmed that these PQSsreadily adopt parallel-stranded or antiparallel-stranded G-quadruplexes (Figure 1). LTRretrotransposons also contain potential triplex-forming sequences, characterized bypolypurine/polypyrimidine runs and these motifs were most abundant in the 3’UTR ofretrotransposons (Lexa et al. 2014a).
Human SVA elements and especially evolutionarily young SVA elements have the highestcontent of G-quadruplex-forming sequences of all human transposable element families(Savage et al. 2013). PQSs were also found inside LTRs of human endogenous retroviruses(HERV) and in 3’UTRs of LINE-1 elements (Lexa et al. 2014b). Evolutionarily young SVA-Felements contained more PQS than middle-aged SVA-C and these had more PQSs than theoldest SVA-A elements (Figure 3). Moreover, full-length and active LINE-1 elements showedgreater abundance of PQS. Circular dichroism spectroscopy revealed that most of these PQShad adopted monomolecular or bimolecular G-quadruplex structures (Lexa et al. 2014b).
Some TE families populate the genomes with short fragments of their full-length elements. InLTR retrotransposons, short fragments are represented by “solo LTRs” formed by ectopicrecombination between full-length elements (Vicient et al. 1999). In LINEs, target-primedreverse transcription (TPRT) is an inefficient process which often terminates before copyingthe entire element, leading to fragments truncated at their 5’end (Szak et al. 2002; Han 2010).It is remarkable that both LTRs of LTR retrotransposons and 3’ ends of LINE-1 elementscontain the majority of potential G-quadruplex-forming sequences. This could beadvantageous for the host genome with these fragments serving as vehicles for spreading G-quadruplexes (Kejnovsky and Lexa 2014). Formation of G-quadruplexes in TEs and thesubsequent TE-shortening processes may help to seed the genome with PQS.
The role of G-quadruplexes in TE life cycle and in cellular processesThe greater abundance of PQS within recent, full-length and active elements indicates animportant role of G-quadruplexes. Moreover, formation of multi-stranded DNA structures,either hairpins or G-quadruplex DNA, was shown to be involved in dimerization of the HIV-1genomic RNA molecules found in virus particles (Sundquist and Heaphy 1993; Lawrence etal. 2003). Similarly, long polypurine tract (PPT) located in 3’UTR of LINE-1retrotransposons, where reverse transcription of the second cDNA strand starts, can formintrastrand G-quadruplex DNA blocking strand extension (Howell and Usdin 1997). Thesefindings suggest that DNA conformation plays a role in the life cycle of transposable elementsand possibly this affects not only transcription and translation but also reverse transcriptionand integration.
However, G-quadruplex-forming regions inside TEs can also influence genomic and cellularprocesses (for review see Bochman et al. 2012). Four-stranded conformation may be a barrierto replication, a hotspot for recombination, participate in changes in chromatin status orenhance/inhibit transcription. G-quadruplexes formed by RNA molecules can affect

posttranscriptional processes or translation (Figure 4). G-quadruplexes found in an internalribosomal entry site (IRES) of viruses may have a tunable regulatory role in translationinitiation (Morris et al. 2010).
During replication, G-quadruplexes present within TEs pose a barrier to the replicationmachinery but they can be unwound by cellular helicases (e.g. FANCJ, PIF1, BLM, WRN)and the balance between stabilization and destabilization of four-stranded structures canregulate replication speed (Wu et al. 2008; Sanders 2010). Although PQSs have been found tobe significantly enriched in long nucleosome-free regions, regardless of their genomic context(Schwarzbauer et al. 2012), G-quadruplexes can also be bound by proteins like ATRX(belonging to a family of SWI/SNF chromatin modulators) that attract H3.3 histones tonucleosomes leading to changes in chromatin structure (Whitehouse and Owen-Hughes2010). G-quadruplex regions can promote the opening of the chromatin which may influencethe action of DNA methyltransferases and in this way, participate in the formation ofepigenetic marks (Guerrero-Bosagna et al. 2014).
Earlier studies suggested that the effect of G-quadruplexes on transcription is generallyrepressive, perhaps due to transcription arrest (Broxson et al. 2011). In addition, detailedstudy of human cells displaying either Bloom or Werner syndromes (Smestad and Maher2015) with loss-of-function mutations in helicase (from the RecQ family) resolving G-quadruplexes (Bohr 2008), demonstrated that the abundance of PQS in promoters wasgenerally higher in down-regulated genes and lower in up-regulated genes (Smestad andMaher 2015). However, other analyses showed that G-quadruplexes can influence thetranscription either positively or negatively, depending partly on the strand in which the four-stranded structure is formed (Bochman et al. 2012). The high abundance of G-rich motifs inpromoters and enhancers has led to a model proposing that G-quadruplexes facilitate mutualbinding of promoters and enhancers that could also affect transcription (Hegyi 2015).
The G-quadruplexes within TEs, e.g. close to promoters (localized in LTRs) of LTRretrotransposons (Lexa et al. 2014a), can represent a genomic tool regulating TE transcriptiondepending on cellular environment – e.g. potassium concentration. The higher abundance ofG-quadruplexes in recent and active TE is in agreement with the inhibiting effect of four-stranded structures on transcription of transposable element families that represent the highestthreat for the genome.
Transposable elements, like all genomic repeats, are templates for homologous recombinationand TE-born G-quadruplexes can even be recombination hotspots (Boán and Gómez-Márquez2010). Amplification of transposable elements, especially by the “copy and paste”mechanism, helps G-quadruplex-forming sequences to spread in the genome. This process isopposed by G-quadruplex erosion or loss, and the dynamic balance between the two processesdetermines the abundance of these DNA conformational regulators, with possible effect on allthe above processes.
Interactions of TE-derived proteins with G-quadruplexesTransposable elements are often surrounded by microsatellite sequences (Nadir et al. 1996;Kejnovský et al. 2013) that are known to readily form unusual DNA conformation includingleft-handed Z-DNA, triplexes and quadruplexes (Pearson and Sinden 1998; Kejnovská et al.2001). DNA with changed conformation is often nucleosome-free (Huppert andBalasubramanian 2007; Wong and Huppert 2009), and it is known that TEs prefer to beinserted into open chromatin (Liu et al. 2009). Unusual DNA conformations are thus not only

formed inside TEs but can also represent the genomic targets of new TE insertions duringwhich changed DNA structures are recognized by transposase (encoded by DNA transposons)or by integrase (encoded by retrotransposons).
The relationship between unusual conformation and TEs could be further demonstrated byproteins originating from TEs (as in the examples of TE domestication/exaptation), that canretain their affinity to unusual conformations. These proteins preferentially utilize secondarystructure recognition than sequence-specific binding. For example, G-quadruplexes arerecognized and cleaved by the RAG1 protein (Nambiar et al. 2011) that evolved fromtransposase of the Transib family of DNA transposons (Kapitonov and Jurka 2005).Moreover, the p53 tumor suppressor protein that binds sites located inside Alu and L1elements (Cui et al. 2011) can also strongly bind G-quadruplex DNA (Quante et al. 2012).Similarly, the multifunctional protein, Ku70 can bind cDNA of Ty1 yeast retrotransposon andsimultaneously has high affinity to G-quadruplex DNA (Downs and Jackson 1999;Paramasivam et al. 2009). In addition, PQSs located upstream of genes often representexapted fragments of TEs (Volff 2006) that have become a promoter (Huda et al. 2011a) or anenhancer (Huda et al. 2011b), originally representing promoters present insideretrotransposon’s LTRs. G-quadruplexes located in these regulatory regions are bound by anumber of proteins (Brázda et al. 2014).
G-quadruplexes inside TEs in relation to RNA infrastructureThe question still remains about a potential link between regulatory systems based on G-quadruplex formation and small RNA-mediated gene silencing. In gene-derived mRNAs, G-quadruplex-forming sequences are accumulated in 3’UTR regions (Beaudoin and Perreault2013). The 3’UTR regions are simultaneously the targets of regulatory proteins as well asmicroRNA molecules resulting in either translation repression or mRNA degradation (Barrettet al. 2012). The 3’UTR-located G-quadruplexes therefore, possess the ability to interferewith the miRNA regulatory network (Beaudoin and Perreault 2013).
The situation is different in LTR retrotransposons where PQSs are mostly found within longterminal repeats while piRNA molecules target 3’UTRs (McCue et al. 2013). In LINE-1elements, PQSs are accumulated in their 3’UTRs but piRNAs and siRNA target the wholelength of elements by changing DNA or histone methylation (Chen et al. 2012; Pezic et al.2014). For this reason, G-quadruplex formation and small RNA silencing are probablycomplementary modes of transposable element life cycle regulation. To the best of ourknowledge, there is no evidence for direct interactions of piRNAs or siRNAs with G-quadruplex DNA.
Guanine quadruplexes are also formed in various non-genic RNA molecules (Millevoi et al.2012). As the majority of the genome is transcribed (Hangauer et al. 2013), the G-quadruplexes found in TE-derived RNAs make a significant contribution to the cellular poolof four-stranded structures. Potential G-quadruplexes were found in long non-coding RNAmolecules, lnc RNAs (Jayaraj et al. 2012), that take part in many important cellular processes,and are often derived from transposable elements (Kapusta et al. 2013). LncRNA moleculesfrequently contain stem-loop structures (Kapusta et al. 2013) and it is probable that G-quadruplexes present in TEs appear there because formation of secondary structures isprobably important for lncRNA function (Kapusta et al. 2013). Similarly, triplex DNAstructures were found in MALAT1 lnc RNA (Brown et al. 2014).

Taken together, multi-stranded RNA structures, together with the action of various small RNAmolecules, contribute to the control of gene expression of both host genes and transposableelements, especially when the genome is pervasively transcribed.
Conclusions and perspectivesHere, we have reviewed the recent findings of sequence motifs that adopt unusual DNAconformations, especially G-quadruplex DNA inside transposable elements. We havediscussed the potential role of these structures in the transposable element life cycle as well asthe impact of G-quadruplexes on the genome structure and function as a whole. Although anumber of publications have reported the existence of unusual DNA conformations in silicoand in vitro, few have shown their role in vivo and more detailed analyses are required.
We support the view that transposable elements can serve as vehicles for genomic spread ofG-quadruplexes. These represent DNA conformational switches providing in this way anadditional level of genome expression regulation, especially in large and massivelytranscribed genomes. Such DNA and RNA conformational changes, inside and outsidetransposable elements, in addition to small RNA and protein infrastructure sometimes derivedfrom TEs, can shape the evolving cellular regulatory network.
AcknowledgementsThis research was supported by the Czech Science Foundation (grant 15-02891S). We wouldlike to thank Professor Boris Vyskot for a critical reading of this manuscript.
Figure Legends:Figure 1. Topologies of G-quadruplex DNA. (a) A 3-D model of the human telomericquadruplex DNA (from PDB 2JPZ, Dai et al., 2007); (b) atomic details of the planar G4 tetradtypical for G4-quadruplexes. Topologies of G-quadruplexes: intramolecular parallel-stranded(c), intramolecular antiparallel-stranded (d), intramolecular 3:1 hybrid (e), parallelbimolecular (f), antiparallel bimolecular (g) and tetramolecular (h) G-quadruplexes withorientation of strands (indicated by arrows). Guanine tetrads are shown in orange.
Figure 2. Localization of G-quadruplexes within transposable elements. Positions of G-quadruplexes and triplex DNA within plant LTR retrotransposons (a), human LINE-1elements (b) and human SVA elements (c). GAG and POL genes, ORFs, regulatory regionslike long terminal repeats (LTRs) with promoters, region with variable number of tandemrepeats (VNTR), hexamer region (HEX), 5’UTR and 3’UTR regions are shown.
Figure 3. G-quadruplexes within transposable elements of various age and activity. Thenumber of G-quadruplexes within plant LTR retrotransposons (a), human LINE-1 (b), humanSVA (c) and human Alu elements (d) depending on their age (a, c, d) or completeness andactivity (b). Evolutionarily younger plant LTR retrotransposons as well as human SVA andAlu elements contain more PQS than older elements. Full-length and active human LINE-1elements also have more PQS than truncated or inactive LINE-1 elements.
Figure 4. Potential roles of G-quadruplexes in genome. Formation of short variants oftransposable elements containing G-quadruplexes – solo LTRs by ectopic recombination and5’ end truncated LINE-1 by non-complete target-primed reverse transcription, TPRT (middlebox). Effect of G-quadruplexes on DNA replication (a), recombination (b), transcription (c)and chromatin remodeling (d).

References
Abad JP, Villasante A (1999) The 3’ non-coding region of the Drosophila melanogaster HeT-Atelomeric retrotransposon contains sequences with propensity to form G-quadruplex DNA.FEBS Lett 453:59–62
Barrett LW, Fletcher S, Wilton SD (2012) Regulation of eukaryotic gene expression by theuntranslated gene regions and other non-coding elements. Cell Mol Life Sci 69:3613–34
Beaudoin J-D, Perreault J-P (2013) Exploring mRNA 3’-UTR G-quadruplexes: evidence ofroles in both alternative polyadenylation and mRNA shortening. Nucleic Acids Res 41:5898–5911
Biémont C, Vieira C (2006) Genetics: junk DNA as an evolutionary force. Nature 443:521–4
Boán F, Gómez-Márquez J (2010) In vitro recombination mediated by G-quadruplexes.Chembiochem: European Journal of Chemical Biology 11:331–4
Bohr VA (2008) Rising from the RecQ-age: the role of human RecQ helicases in genomemaintenance. Trends Biochem Sci 33:609–20
Bochman ML, Paeschke K, Zakian VA (2012) DNA secondary structures: stability andfunction of G-quadruplex structures. Nature Rev Genet 13:770–80
Bousios A, Darzentas N, Tsaftaris A, Pearce SR (2010) Highly conserved motifs in non-coding regions of Sirevirus retrotransposons: the key for their pattern of distribution withinand across plants? BMC Genomics 11:89
Brázda V, Hároníková L, Liao JCC, Fojta M (2014) DNA and RNA quadruplex-bindingproteins. International J Mol Sci 15:17493–517
Brown JA, Bulkley D, Wang J, et al (2014) Structural insights into the stabilization ofMALAT1 noncoding RNA by a bipartite triple helix. Nature Struct Mol Biol 21:633–40
Broxson C, Beckett J, Tornaletti S (2011) Transcription arrest by a G quadruplex forming-trinucleotide repeat sequence from the human c-myb gene. Biochemistry 50:4162–72
Bureau TE, Wessler SR (1992) Tourist: a large family of small inverted repeat elementsfrequently associated with maize genes. Plant Cell 4:1283–94
Cui F, Sirotin M V, Zhurkin VB (2011) Impact of Alu repeats on the evolution of human p53binding sites. Biology Direct 6:2
Dai J, Carver M, Punchihewa C, Jones RA, Yang D (2007). Structure of the Hybrid-2 typeintramolecular human telomeric G-quadruplex in K+ solution: insights into structurepolymorphism of the human telomeric sequence. Nucleic Acids Res 35:4927-4940
Downs JA, Jackson SP (1999) Involvement of DNA end-binding protein Ku in Ty elementretrotransposition. Mol Cell Biol 19:6260–8
Guerrero-Bosagna C, Weeks S, Skinner MK (2014) Identification of genomic features inenvironmentally induced epigenetic transgenerational inherited sperm epimutations. PloSONE 9:e100194
Han JS (2010) Non-long terminal repeat (non-LTR) retrotransposons: mechanisms, recentdevelopments, and unanswered questions. Mobile DNA 1:15

Hangauer MJ, Vaughn IW, McManus MT (2013) Pervasive transcription of the humangenome produces thousands of previously unidentified long intergenic noncoding RNAs.PLoS Genetics 9:e1003569
Hegyi H (2015) Enhancer-promoter interaction facilitated by transiently forming G-quadruplexes. Scientific Reports 5:9165
Howell R, Usdin K (1997) The ability to form intrastrand tetraplexes is an evolutionarilyconserved feature of the 3’ end of L1 retrotransposons. Mol Biol Evol 14:144–55
Huda A, Bowen NJ, Conley AB, Jordan IK (2011a) Epigenetic regulation of transposableelement derived human gene promoters. Gene 475:39–48
Huda A, Tyagi E, Mariño-Ramírez L, et al (2011b) Prediction of transposable element derivedenhancers using chromatin modification profiles. PloS ONE 6:e27513
Huppert JL, Balasubramanian S (2007) G-quadruplexes in promoters throughout the humangenome. Nucleic Acids Res 35:406–13
Cerofolini L, Amato J, Giachetti A, Limongelli V, Novellino E, Parrinello M et al (2014) G-triplex structure and formation propensity. Nucleic Acids Res 42:13393-13404
Chen L, Dahlstrom JE, Lee S-H, Rangasamy D (2012) Naturally occurring endo-siRNAsilences LINE-1 retrotransposons in human cells through DNA methylation. Epigenetics7:758–71
Jayaraj GG, Pandey S, Scaria V, Maiti S (2012) Potential G-quadruplexes in the human longnon-coding transcriptome. RNA Biol 9:81–6
Kapitonov V V, Jurka J (2005) RAG1 Core and V(D)J Recombination Signal Sequences WereDerived from Transib Transposons. PLoS Biol 3:e181
Kapusta A, Kronenberg Z, Lynch VJ, et al (2013) Transposable elements are majorcontributors to the origin, diversification, and regulation of vertebrate long noncoding RNAs.PLoS Genetics 9:e1003470
Kawakami T, Strakosh SC, Zhen Y, Ungerer MC (2010) Different scales of Ty1/copia-likeretrotransposon proliferation in the genomes of three diploid hybrid sunflower species.Heredity 104:341–50
Kejnovská I, Tůmová M, Vorlícková M (2001) (CGA)(4): parallel, anti-parallel, right-handedand left-handed homoduplexes of a trinucleotide repeat DNA. Biochim Biophys Acta1527:73–80
Kejnovsky E, Lexa M (2014) Quadruplex-forming DNA sequences spread byretrotransposons may serve as genome regulators. Mobile Genet Elements 4:e28084
Kejnovský E, Michalovova M, Steflova P, et al (2013) Expansion of Microsatellites onEvolutionary Young Y Chromosome. PLoS ONE 8:e45519
Lawrence DC, Stover CC, Noznitsky J, et al (2003) Structure of the intact stem and bulge ofHIV-1 Psi-RNA stem-loop SL1. J Mol Biol 326:529–42
Lever A, Gottlinger H, Haseltine W, Sodroski J (1989) Identification of a sequence requiredfor efficient packaging of human immunodeficiency virus type 1 RNA into virions. J Virol63:4085–7
Lexa M, Kejnovský E, Steflová P, et al (2014a) Quadruplex-forming sequences occupydiscrete regions inside plant LTR retrotransposons. Nucleic Acids Res 42:968–78

Lexa M, Steflova P, Martinek T, et al (2014b) Guanine quadruplexes are formed by specificregions of human transposable elements. BMC Genomics 15:1032
Liu S, Yeh C-T, Ji T, et al (2009) Mu transposon insertion sites and meiotic recombinationevents co-localize with epigenetic marks for open chromatin across the maize genome. PLoSGenetics 5:e1000733
Macas J, Koblízková A, Navrátilová A, Neumann P (2009) Hypervariable 3’ UTR region ofplant LTR-retrotransposons as a source of novel satellite repeats. Gene 448:198–206
McCue AD, Nuthikattu S, Slotkin RK (2013) Genome-wide identification of genes regulatedin trans by transposable element small interfering RNAs. RNA Biol 10:1379–95
Millevoi S, Moine H, Vagner S (2012) G-quadruplexes in RNA biology. WileyInterdisciplinary Reviews: RNA 3:495–507
Morris MJ, Negishi Y, Pazsint C, et al (2010) An RNA G-quadruplex is essential for cap-independent translation initiation in human VEGF IRES. J Am Chem Soc 132:17831–9
Nadir E, Margalit H, Gallily T, Ben-Sasson S a (1996) Microsatellite spreading in the humangenome: evolutionary mechanisms and structural implications. Proc Natl Acad Sci USA93:6470–5
Nambiar M, Goldsmith G, Moorthy BT, et al (2011) Formation of a G-quadruplex at theBCL2 major breakpoint region of the t(14;18) translocation in follicular lymphoma. NucleicAcids Res 39:936–48
Paramasivam M, Membrino A, Cogoi S, et al (2009) Protein hnRNP A1 and its derivativeUp1 unfold quadruplex DNA in the human KRAS promoter: implications for transcription.Nucleic Acids Res 37:2841–53
Pearson CE, Sinden RR (1998) Trinucleotide repeat DNA structures: dynamic mutations fromdynamic DNA. Curr Opin Struct Biol 8:321–30
Pezic D, Manakov SA, Sachidanandam R, Aravin AA (2014) piRNA pathway targets activeLINE1 elements to establish the repressive H3K9me3 mark in germ cells. Genes Develop28:1410–28
Piekna-Przybylska D, Sullivan MA, Sharma G, Bambara RA (2014) U3 region in the HIV-1genome adopts a G-quadruplex structure in its RNA and DNA sequence. Biochemistry53:2581–93
Piriyapongsa J, Jordan IK (2008) Dual coding of siRNAs and miRNAs by plant transposableelements. RNA 14:814–21
Quante T, Otto B, Brázdová M, et al (2012) Mutant p53 is a transcriptional co-factor thatbinds to G-rich regulatory regions of active genes and generates transcriptional plasticity. CellCycle 11:3290–303
Rajendran A, Endo M, Hidaka K, Sugiyama H (2014) Direct and single-moleculevisualization of the solution-state structures of G-hairpin and G-triplex intermediates. AngewChem Int Ed 53:4107-4112
Rich A (1993) DNA comes in many forms. Gene 135:99–109
Sabot F, Schulman a H (2006) Parasitism and the retrotransposon life cycle in plants: ahitchhiker’s guide to the genome. Heredity 97:381–8
Sanders CM (2010) Human Pif1 helicase is a G-quadruplex DNA-binding protein with G-quadruplex DNA-unwinding activity. Biochem J 430:119–28

Savage AL, Bubb VJ, Breen G, Quinn JP (2013) Characterisation of the potential function ofSVA retrotransposons to modulate gene expression patterns. BMC Evol Biol 13:101
Schwarzbauer K, Bodenhofer U, Hochreiter S (2012) Genome-wide chromatin remodelingidentified at GC-rich long nucleosome-free regions. PloS ONE 7:e47924
Smestad J, Maher L (2015) Putative G-quadruplex forming sequence signatures in genesdifferentially transcribed upon loss of BLM or WRN helicases. BioRxiv.doi:http://dx.doi.org/10.1101/013664
Steinbauerová V, Neumann P, Novák P, Macas J (2011) A widespread occurrence of extraopen reading frames in plant Ty3/gypsy retrotransposons. Genetica 139:1543–55
Sundquist WI, Heaphy S (1993) Evidence for interstrand quadruplex formation in thedimerization of human immunodeficiency virus 1 genomic RNA. Proc Natl Acad Sci USA90:3393–7
Szak ST, Pickeral OK, Makalowski W, et al (2002) Molecular archeology of L1 insertions inthe human genome. Genome Biol 3: research0052
Tlučková K, Marušič M, Tóthová P, et al (2013) Human papillomavirus G-quadruplexes.Biochemistry 52:7207–16
Vicient C, Suoniemi A, Anamthawat-Jónsson K, et al (1999) Retrotransposon BARE-1 and ItsRole in Genome Evolution in the Genus Hordeum. Plant Cell 11:1769–1784
Volff J-N (2006) Turning junk into gold: domestication of transposable elements and thecreation of new genes in eukaryotes. BioEssays 28:913–22
Watson JD, Crick FHC (2003) A structure for deoxyribose nucleic acid. 1953. Nature421:397–8
Whitehouse I, Owen-Hughes T (2010) ATRX: Put me on repeat. Cell 143:335–6
Wicker T, Sabot F, Hua-Van A, et al (2007) A unified classification system for eukaryotictransposable elements. Nature Rev Genet 8:973–82
Wong HM, Huppert JL (2009) Stable G-quadruplexes are found outside nucleosome-boundregions. Molecular BioSystems 5:1713–9
Wu Y, Shin-ya K, Brosh RM (2008) FANCJ helicase defective in Fanconia anemia and breastcancer unwinds G-quadruplex DNA to defend genomic stability. Mol Cell Biol 28:4116–28
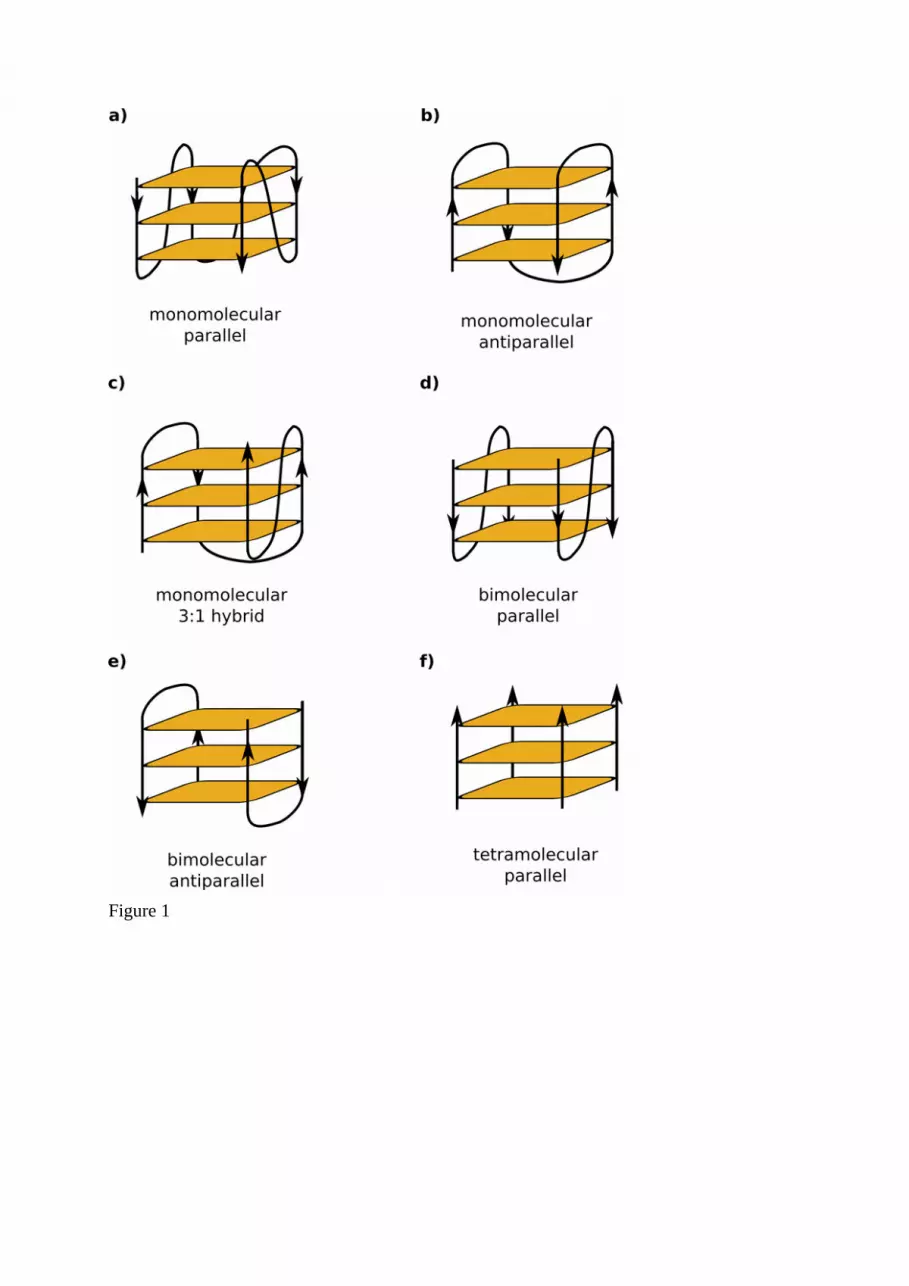
Figure 1

Figure 2
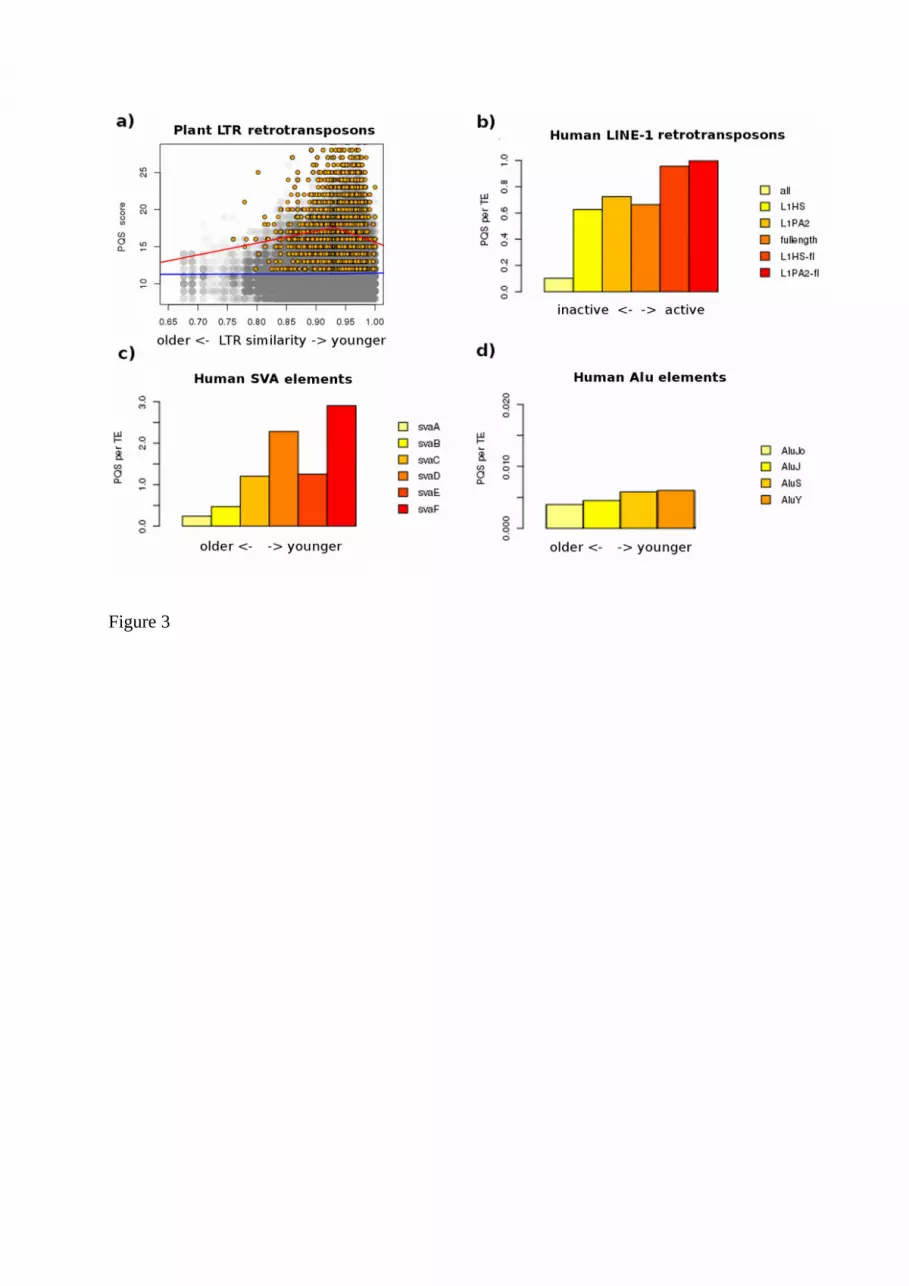
Figure 3

Figure 4
Related Documents











