A cholesterol free, high-fat diet suppresses gene expression of cholesterol transporters in murine small intestine Heleen M de Vogel-van den Bosch 1,2 , Nicole JW de Wit 1,2 , Guido JEJ Hooiveld 1,2 , Hanneke Vermeulen 1,2 , Jelske N van der Veen 3 , Sander M Houten 4 , Folkert Kuipers 2,3 , Michael Müller 1,2 , Roelof van der Meer 1,2,5 1 Nutrition, Metabolism and Genomics Group, Division of Human Nutrition, Wageningen University, 6703HD Wageningen, the Netherlands; 2 Nutrigenomics Consortium, TI Food and Nutrition, Wageningen, the Netherlands; 3 Center for Liver, Digestive, and Metabolic Diseases, Laboratory of Pediatrics, University-Medical Center Groningen, Groningen, The Netherlands; 4 Department of Pediatrics/Emma Children's Hospital, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; and 5 NIZO Food Research, Ede, The Netherlands Running head: Dietary fat suppresses intestinal cholesterol transporters To whom correspondence should be addressed: Nicole de Wit, PhD, Nutrition, Metabolism & Genomics Group, Division of Human Nutrition, Wageningen University, Bomenweg 2, NL-6703 HD, Wageningen, the Netherlands. Tel: (+31) 317 485791; Fax: (+31) 317 483342; E-mail: [email protected] Page 1 of 37 Articles in PresS. Am J Physiol Gastrointest Liver Physiol (March 20, 2008). doi:10.1152/ajpgi.00360.2007 Copyright © 2008 by the American Physiological Society.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A cholesterol free, high-fat diet suppresses gene expression of cholesterol
transporters in murine small intestine
Heleen M de Vogel-van den Bosch 1,2, Nicole JW de Wit1,2, Guido JEJ Hooiveld1,2, Hanneke
Vermeulen1,2, Jelske N van der Veen3, Sander M Houten4, Folkert Kuipers2,3, Michael Müller1,2, Roelof
van der Meer1,2,5
1Nutrition, Metabolism and Genomics Group, Division of Human Nutrition, Wageningen University,
6703HD Wageningen, the Netherlands; 2Nutrigenomics Consortium, TI Food and Nutrition, Wageningen,
the Netherlands; 3Center for Liver, Digestive, and Metabolic Diseases, Laboratory of Pediatrics,
University-Medical Center Groningen, Groningen, The Netherlands; 4Department of Pediatrics/Emma
Children's Hospital, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands;
and 5NIZO Food Research, Ede, The Netherlands
Running head: Dietary fat suppresses intestinal cholesterol transporters
To whom correspondence should be addressed: Nicole de Wit, PhD, Nutrition, Metabolism & Genomics
Group, Division of Human Nutrition, Wageningen University, Bomenweg 2, NL-6703 HD, Wageningen,
the Netherlands. Tel: (+31) 317 485791; Fax: (+31) 317 483342; E-mail: [email protected]
Page 1 of 37Articles in PresS. Am J Physiol Gastrointest Liver Physiol (March 20, 2008). doi:10.1152/ajpgi.00360.2007
Copyright © 2008 by the American Physiological Society.

2
ABSTRACT
Transporters present in the epithelium of the small intestine determine the efficiency by which dietary and
biliary cholesterol are taken up into the body and thus control whole-body cholesterol balance. Niemann-
Pick C1 Like Protein 1 (Npc1l1) transports cholesterol into the enterocyte, whereas ATP-binding cassette
transporters Abca1 and Abcg5/Abcg8 are presumed to be involved in cholesterol efflux from the
enterocyte towards plasma HDL and back into the intestinal lumen, respectively. Abca1, Abcg5, and
Abcg8 are well-established LXR target genes. We examined the effects of a high-fat diet on expression
and function of cholesterol transporters in the small intestine in mice. Npc1l1, Abca1, Abcg5, and Abcg8
were all down-regulated after 2, 4, and 8 weeks on a cholesterol-free, high-fat diet. The high-fat diet did
not affect biliary cholesterol secretion but diminished fractional cholesterol absorption from 61% to 42%
(p<0.05). In an acute experiment, in which triacylglycerols of unsaturated fatty acids were given by
gavage, we found that this down-regulation occurs within a 6 hours time frame. Studies in LXRα-null
mice, confirmed by in vitro data, showed that fatty acid-induced down-regulation of cholesterol
transporters is LXRα-independent and associated with a post-translational increase in Hmg-CoA
reductase activity that reflects induction of cholesterol biosynthesis as well as with a doubling of neutral
fecal sterol loss. This study highlights the induction of adaptive changes in small intestinal cholesterol
metabolism during exposure to dietary fat.
Supplementary keywords:
Cholesterol absorption, ABC transporters, Npc1l1; diet; fatty acids
Page 2 of 37

3
INTRODUCTION
The epithelium of the small intestine acts as a gatekeeper that controls the efficiency by which dietary and
biliary cholesterol enter the body and thus plays a critical role in the regulation of whole-body cholesterol
balance (20, 39, 46). Intestinal cholesterol absorption has long been considered as a passive process,
however, this view has changed considerably during the last years. Niemann-Pick C1 Like Protein 1
(Npc1l1) has been identified as a cholesterol absorption transporter localized at the apical membrane of
the small intestine (2, 13). ATP-binding cassette transporters Abca1, Abcg5, and Abcg8 are presumed to
facilitate cholesterol efflux from the enterocyte. Abca1 promotes cholesterol efflux at the basolateral
membrane to Apo-AI for HDL formation (15, 27, 28, 36). In contrast, the Abcg5/g8 heterodimer is
responsible for transport of cholesterol from the enterocyte back into the intestinal lumen, a process that
limits overall absorption efficacy (47, 52). Mutations in Abcg5 and Abcg8 contribute to plant sterol
accumulation in plasma that is seen in patients with sitosterolemia (5). Abca1, Abcg5, and Abcg8 are
target genes of the nuclear receptor liver X receptor (LXR), a receptor that is known to be involved in
control of cholesterol-related metabolic processes (34, 45). Oxysterols, which are derivatives of
cholesterol, are natural ligands of LXRs. 27-Hydroxycholesterol, which is synthesized from cholesterol
by Cytochrome P450, family 27, subfamily a (Cyp27a1) (23), is a common oxysterol in the small
intestine (22).
During fat absorption from the small intestinal lumen, cholesterol is required for the formation of
chylomicrons (1, 33, 50). Therefore, as fat intake increases and more chylomicrons have to be formed
(18), more cholesterol is theoretically needed for this process. We hypothesized that on a cholesterol-free,
high-fat diet, cholesterol will be retained in the enterocyte to be available for chylomicron formation to
allow for efficient transport of dietary fat out of the enterocyte. Such an effect could theoretically be
mediated by a down-regulation of cholesterol efflux transporters. To study this hypothesis, we performed
a low- and high-fat diet intervention in mice. We found that in the small intestine gene expression levels
Page 3 of 37

4
of cholesterol efflux transporters were suppressed on the high-fat diet. Surprisingly, this coincided with a
down-regulated gene expression of the cholesterol absorption protein Npc1l1 and with diminished
fractional cholesterol absorption. In additional studies performed to investigate the mechanism behind this
down-regulation, we found that these high-fat induced changes in gene expression of intestinal cholesterol
transporters were LXRα-independent.
Page 4 of 37

5
MATERIALS AND METHODS
Materials
Palmitic acid, stearic acid, oleic acid, linoleic acid, docosahexaenoic acid, fatty acid-free BSA, HMG-
CoA and carboxy methylcellulose were obtained from Sigma-Aldrich (Zwijndrecht, the Netherlands). 27-
Hydroxycholesterol was purchased from Research Plus (Manasquan, NJ, USA). Radiolabeled
[14C]HMG-CoA was obtained from Amersham Biosciences (Diegem, Belgium). Trilinolein, trilinolenin,
tridocosahexaenoin, trieicosapentaenoin, which are synthetic triacylglycerols with three identical acyl
moieties, were from Nu-Chek-Prep, Inc (Elysian, MN, USA), whereas triolein was from Fluka
(Zwijndrecht, the Netherlands). Cholesterol-D7 and cholesterol-D5 were purchased from Cambridge
Isotope Laboratories (Andover, MA, USA). Intralipid was obtained from Fresenius Kabi (Den
Bosch, The Netherlands).
Animals and diets
Male C57BL/6J mice were purchased from Harlan (Horst, The Netherlands). Lxra+/+ and Lxra–/– mice,
generated by Deltagen using standard gene-targeting methods, were bred at the animal facility of the
University Medical Center Groningen, The Netherlands (32). These mice have a mixed background of
C57BL/6J and 129/OlaHsd strains. SV129 mice were originally from Charles-River (Maastricht, The
Netherlands) and bred at the animal facility of Wageningen University, Wageningen, The Netherlands.
All mice were housed in a light- and temperature-controlled facility and had free access to water and
standard laboratory chow (RMH-B, Hope Farms, Woerden, The Netherlands). All experiments were
approved by the Ethical Committee on animal testing of Wageningen University and were performed in
accordance with the national law.
In a first study we investigated the effect of a cholesterol-free, high-fat diet intervention on intestinal gene
expression of C57BL/6J mice. To adapt mice to the diets, all mice were fed the low-fat purified diet for 3
Page 5 of 37

6
weeks. Thereafter, the mice were divided into two groups and the nine week-old mice were fed a
powdered, cholesterol-free, high- or low-fat purified diet (n=6) for 2, 4, and 8 weeks. Low-fat and high-
fat diets have been based on 'Research Diets' formulas: D12450B / D12451, with some adaptations with
regard to fat (Research Diet Services, Wijk bij Duurstede, The Netherlands), to mimic the fatty acid
composition of a human, Western-type diet. The compositions of the diets are given in Table 1 of the
supplemental data. It should be noticed that the energy density of all nutrients, except for fat and
carbohydrate, was made identical in each diet. After 2, 4, and 8 weeks, mice were anaesthetized with a
mixture of isofluorane (1.5%), nitrous oxide (70%) and oxygen (30%). Blood was collected by orbital
puncture. Plasma was obtained by centrifuging at 203xg for 10 minutes and stored at –80 °C. The small
intestines were excised and the remaining fat and pancreatic tissue were carefully removed. The small
intestines were divided in three equal parts (proximal, middle and distal), cut open longitudinally, and
washed with PBS. Small intestinal mucosa was scraped, snap-frozen in liquid nitrogen, and stored at -80
°C until RNA isolation.
In a second study, male C57BL/6J were subjected to a 2-weeks intervention with a high- or low-fat
diet, as was described for the first study (Low-fat: n=6, high-fat: n=8). After 2 weeks, mice received an
intravenous dose of 0.3 mg (0.763 µmol) cholesterol-D7 dissolved in 20% Intralipid® and an oral dose of
0.6 mg (1.535 µmol) cholesterol-D5 dissolved in medium-chain triglyceride oil. Blood spots were
collected from the tail on filter paper before and 3 days after administration of labeled cholesterol for
measurement of fractional cholesterol absorption. At 72 h after administration, mice were anaesthetized
by intraperitoneal injection with Hypnorm (fentanyl/fluanisone, 1 ml/kg) and Diazepam (10 mg/kg), and
the gallbladder was canulated as described (42). Bile was collected for 15 minutes. Body temperature was
stabilized using a humidified incubator. Fractional cholesterol absorption was measured with the dual
isotope method (see below).
In a third study, three months-old male SV129 mice were given an oral gavage with 400 µl of the
synthetic triacylglycerols triolein (~396 mg), trilinolein (~349 mg), trilinolenin (~360 mg),
Page 6 of 37

7
trieicosapentaenoin (~370 mg), and tridocosahexaenoin (~370 mg). An oral gavage with carboxy
methylcellulose was given as a control. Two weeks before the start of the experiment, the mice were
switched to a diet consisting of a modified AIN76A diet, in which corn oil was replaced by olive oil
(Research diet services, Wijk bij Duurstede, The Netherlands). Four hours before the gavage, the mice
were fasted. Six hours after the oral gavage the mice were anaesthetized. The small intestines were
isolated and flushed with ice-cold PBS and the remaining fat and pancreatic tissue was carefully removed.
The small intestines were snap-frozen in liquid nitrogen and stored at -80 °C until RNA isolation.
In a fourth study, 4-5 months-old female LXRα-null mice (32) and corresponding LXRα+/+
littermates (wild-type) were fed the same high- and low-fat diets as in the first study for two weeks. Prior
to this diet intervention, the mice were fed for two weeks the cholesterol-free, low-fat diet, to adapt to the
purified diet. Mice were housed per 2 in a cage and per cage feces were collected at 24 hours, 48 hours
and after 2 weeks of diet intervention. After 2 weeks of diet intervention mice were anaesthetized, blood
was collected via a heart puncture, and plasma was stored at –80 °C. Small intestines were processed the
same way as in the first experiment.
Cell culture
Mouse small intestinal epithelial (MSIE) cells were a generous gift from Robert Whitehead (Vanderbilt
University, Nashville, USA) (49). Cells were grown in RPMI 1640 medium (Cambrex, Verviers,
Belgium) supplemented with 5% fetal bovine serum, 100 U penicillin/ml, 100 µg streptomycin/ml (all
from Cambrex, Verviers, Belgium), and 5 U/ml of murine gamma interferon (IFN-γ) (Invitrogen, Breda,
The Netherlands). The MSIE cells were derived from the transgenic temperature-sensitive simian virus 40
large-T antigen Immortimouse and, as such, were grown at the permissive temperature of 33°C. For all
experiments, cells were maintained in serum-free medium which contained 11 g/l fatty acid free BSA and
was supplemented with additives (5 µM 27-hydroxycholesterol, or 500 µM fatty acids). Before adding to
the cells, fatty acids were bound to BSA by preincubation in the BSA-containing medium for 30 minutes
Page 7 of 37

8
at 37°C. A 50 mmol/L stock solution of each fatty acid was prepared by dissolving the fatty acid in 70
mmol/L KOH. 27-Hydroxycholesterol was dissolved in ethanol.
RNA isolation
Total RNA was isolated using TRIzol reagent (Invitrogen, Breda, The Netherlands) according to the
manufacturer’s instructions. For microarray hybridization the isolated RNA was further column-purified
(SV total RNA isolation system Promega, Leiden, The Netherlands). RNA concentration was measured
on a NanoDrop ND-1000 UV-Vis spectrophotometer (Isogen, Maarssen, The Netherlands) and analyzed
on a bioanalyzer (Agilent Technologies, Amsterdam, the Netherlands) with 6000 Nano Chips according
to the manufacturer’s instructions.
Array hybridization and microarray data analysis
RNA of the proximal segment and the middle segment of the small intestine were pooled per treated
group (n=6) and hybridized to Affymetrix Mouse genome 430 2.0 arrays. Detailed methods for the
labeling and subsequent hybridizations to the arrays are described in the eukaryotic section in the
GeneChip Expression Analysis Technical Manual Rev. 3 from Affymetrix, which is available upon
request (Santa Clara, CA). Arrays were scanned on a GeneChip Scanner 3000 (Affymetrix). Data analysis
was performed in Microarray Analysis Suite version 5.0 (Affymetrix). Array data have been submitted to
the Gene Expression Omnibus, accession number GSE8582.
Real-time PCR analysis
cDNA of the middle segment of the small intestine was synthesized from 1 µg of total RNA using the
Reverse transcription system (Promega, Leiden, The Netherlands) following the supplier’s protocol.
cDNA was PCR amplified with Platinum Taq DNA polymerase (all reagents were from Invitrogen). Most
of the primer sequences were obtained from the PrimerBank of the Harvard University (48), or otherwise
Page 8 of 37

9
constructed with primer3 (Http://primer3.sourceforge.net). Primers were tested for specificity by BLAST
analysis. The sequences of primers used are available on request. PCR was carried out using SYBR green
on a MyIQ thermal cycler (Bio-Rad laboratories BV, Veenendaal, The Netherlands) with the following
thermal cycling conditions: 8 min at 94˚C, followed by 45 cycles of 94˚C for 15 s and 60˚C for 1 min. All
samples were performed in duplicate and normalized to cyclophilin A and 18S expression. Only the
results of the cyclophilin A normalization are shown as they are representative for the results of the 18S
normalization.
Analysis of bile composition and cholesterol absorption
Analytical procedure - Cholesterol was extracted from blood spots with 1 ml of 95% ethanol/acetone (1:1
v/v) for gas chromatography/mass spectrometric (GC/MS) analysis according to Neese et al. (20).
Unesterified cholesterol from blood spots was subsequently derivatized using N,O-bis-
(trimethyl)trifluoroacetamide with 1 % trimethylchlorosilane at room temperature. Biliary lipids were
extracted according to Bligh and Dyer (7). Biliary concentrations of cholesterol and phospholipids were
determined as previously described (31). Bile salts were measured enzymatically.
GC/MS measurements of mass isotopomer distribution - Cholesterol trimethylsilylether derivatives were
separated with a Trace MS plus GC/MS (Interscience, Breda, the Netherlands), using a 20 m × 0.18 mm
(0.18 µm film thickness) DB17 ms column (J&W Scientific, Falson, CA). The oven temperature was
programmed from 140 to 280°C at 20°/min. A splitless injection was applied. Ions monitored were m/z
458-465 corresponding to the m0-m8 mass isotopomers. The fractional isotopomer distribution measured
(m0-m8) was corrected for the fractional distribution due to natural abundance of 13C and 2H by multiple
linear regression as described by Lee et al. (21) to obtain excess fractional distribution of mass
isotopomers (M0-M8) resulting from isotope dilution of administered labeled compounds. In this
approach, M5 represented the fractional contribution of the orally administered label and M7 the fractional
contribution of the i.v. administered label.
Page 9 of 37

10
Fractional cholesterol absorption measurement – Fractional cholesterol absorption was calculated a s
described for the plasma dual isotope ratio method (40). Blood spots obtained at 72 hours after
intravenous and oral administration were used for the calculation of fractional cholesterol absorption (8).
Fractional cholesterol absorption was calculated as the ratio of the fraction orally-administered
cholesterol-D5 and the fraction IV-administered cholesterol-D7 as measured in bloodspots obtained 72
hours after administration, after being corrected for its orally and IV administered dose.
HMGR activity assay
Activity of 3-hydroxy-3-methylglutaryl-Coenzyme A reductase (Hmgcr) was measured in the study
with LXRα-/- and wild-type mice as described by Brown et al. (9) with some modifications. We used
scrapings of the proximal part of the intestine, as the middle part was completely used to study gene
expression. The scrapings were dissolved in HMGR assay buffer containing 100 mM KPi, 200 mM KCl,
5 mM EGTA, 5 mM EDTA, 10 mM DTT, and 10 mg/ml leupeptin (pH 7.1). The tissue was disrupted
using an Ultra-turrax followed by sonication (twice at 8-watt output, 40 J, at room temperature). The
homogenate was centrifuged for 5 minutes at 1000xg. Protein concentration was determined in the
supernatant using the Bradford method. The homogenates were diluted to approximately 0.5 mg/mL.
One-hundred microliters of the resulting homogenate was preincubated for 10 min at 37 °C with 60 µl of
cofactor-mix containing 175 mM glucose-6-phosphate, 6.7 mM NADPH, 16.7 mM EDTA, and 0.7 unit
glucose-6- phosphate dehydrogenase. The enzyme reactions were started with the addition of 1.8 nmol of
[14C]HMG-CoA and 5.6 nmol of HMG-CoA in 40 µl of H2O. After a 30-min incubation period at 37 °C,
reactions were terminated by adding 50 µl of 1.2 N HCl. After 30 min, the product was extracted three
times with 2 ml of ethyl acetate. The extracts were evaporated to dryness and analyzed by silica thin layer
chromatography using a solvent system toluene:acetone (1:1) dried with Na2SO4. The formed product was
quantified by phosphorimaging (Fuji FLA-3000) with the aid of the Aida software package using samples
Page 10 of 37

11
with known amounts of [14C]mevalonate. For each mouse, the activity of HMGR was determined in
duplicate.
Other analytical methods
Feces were lyophilized, weighed, and homogenized. Neutral sterols were analyzed according to Arca et
al. (3). Total bile acids in feces were determined as described previously (17). Pooled plasma samples
from all animals of one group were used for lipoprotein separation by fast protein liquid chromatography
(FPLC) as described previously (42). Total cholesterol was measured using a commercially available kit
(Roche Molecular Biochemicals, Mannheim, Germany).
Statistical analysis
All data are reported as the mean ± SEM. The differences between the mean values were tested for
statistical significance by the two-tailed Student’s t-test. P-values <0.05 were considered to be significant.
Page 11 of 37

12
RESULTS
Cholesterol transporter genes are down-regulated in small intestines of mice on a cholesterol-free,
high-fat diet
In this study we first focused on the expression of genes involved in cholesterol transport in the small
intestine of C57BL/6J wild-type mice fed purified, cholesterol-free, low- or high-fat diets. Microarray
results of cholesterol transporters and related genes at week 2, 4, and 8 of the middle segment of the small
intestine are shown in table 1. For week 2, additional qRT-PCR analyses were performed, which
accurately confirmed our array results (Table 1). Similar gene expression patterns as were found in the
middle part of the small intestine were also seen in the proximal segments. However, as changes were
most prominent in the middle segment, we focused on this part of the small intestine.
An expected up-regulation of microsomal triglyceride transfer protein (Mttp), involved in
chylomicron assembly, was observed as it is known that an elevated fat intake stimulates chylomicron
formation (18, 25, 44). In line with the assumption that more chylomicron formation means an elevated
requirement of cholesterol, we found a decrease in gene expression of cholesterol efflux transporters
Abca1, Abcg5, and Abcg8. Surprisingly, the cholesterol absorption transporter Npc1l1 was also down-
regulated. Additionally, Cyp27a1 was down-regulated, which suggests that the conversion of cholesterol
into 27-hydroxycholesterol is reduced on a high-fat diet. Gene expression of Hmgcr, the key enzyme for
cholesterol synthesis, was not changed. Based on the down-regulation of Npc1l1, we hypothesized that on
a cholesterol-free, high-fat diet, cholesterol absorption is diminished. Moreover, the cholesterol efflux at
the apical and basolateral side via Abcg5, Abcg8, and Abca1 will be reduced, as cholesterol in the
enterocyte is mainly required for chylomicron formation (Figure 1).
A cholesterol-free, high-fat diet decreases fractional cholesterol absorption
Page 12 of 37

13
Fractional cholesterol absorption was measured to check whether the reduced gene expression of the
cholesterol absorption transporter Npc1l1 functionally leads to diminished cholesterol absorption. Figure
2 shows that a cholesterol-free, high-fat diet for two weeks reduced fractional cholesterol absorption from
61% to 42% compared to a cholesterol-free, low-fat diet in C57BL/6J wild-type mice, which is a
reduction of 31%. Additionally, we determined biliary secretion rates of cholesterol and bile acids to
check whether differences in cholesterol absorption could be due to altered bile formation induced on a
high-fat diet. However, we found no differences in these parameters between a low- and high-fat diet fed
mice (Table 2). Since the mice were fed cholesterol-free diets, unchanged biliary cholesterol secretion
implies that the absolute cholesterol absorption was reduced by 31% on a high-fat diet.
Down-regulation of cholesterol transporters is an acute response to dietary triacylglycerols
To investigate whether the suppression of gene expression of the cholesterol transporters reflects an acute
process, we gavaged SV129 mice with 400 µl of unsaturated fatty acids in the form of triacylglycerols.
Six hours after gavage, the small intestine was harvested and gene expression was analyzed by qRT-PCR
(Figure 3). This experiment showed that cholesterol efflux transporters were already down-regulated after
6 hours exposure to all fatty acids studied without marked differences between the different fatty acids.
Furthermore, a pronounced decrease in gene expression of Npc1l1 was found in this short-term study. In
addition, Mttp was induced, with most pronounced induction by C18:1, while Cyp27a1 was equally
down-regulated by all acyl species. In contrast to the long term diet intervention, Hmgcr expression was
induced by C18:1 but not by C20:5 or C22:6 fatty acids. Additionally, this acute study in SV129 mice
showed that down-regulation of cholesterol efflux and absorption transporter gene expression by dietary
fat is mouse strain-independent.
Down-regulation of Abca1 is caused by unsaturated fatty acids and is not due to shortage of the
endogenous LXR ligand 27-hydroxycholesterol
Page 13 of 37

14
Because 27-hydroxycholesterol is the major natural LXR ligand present in the enterocytes (22),
suppression of Cyp27a1 expression might indicate that a reduced expression of LXR target genes Abca1,
Abcg5 and Abcg8 upon high-fat feeding is due to shortage of endogenous LXR ligand (Figure 1). To
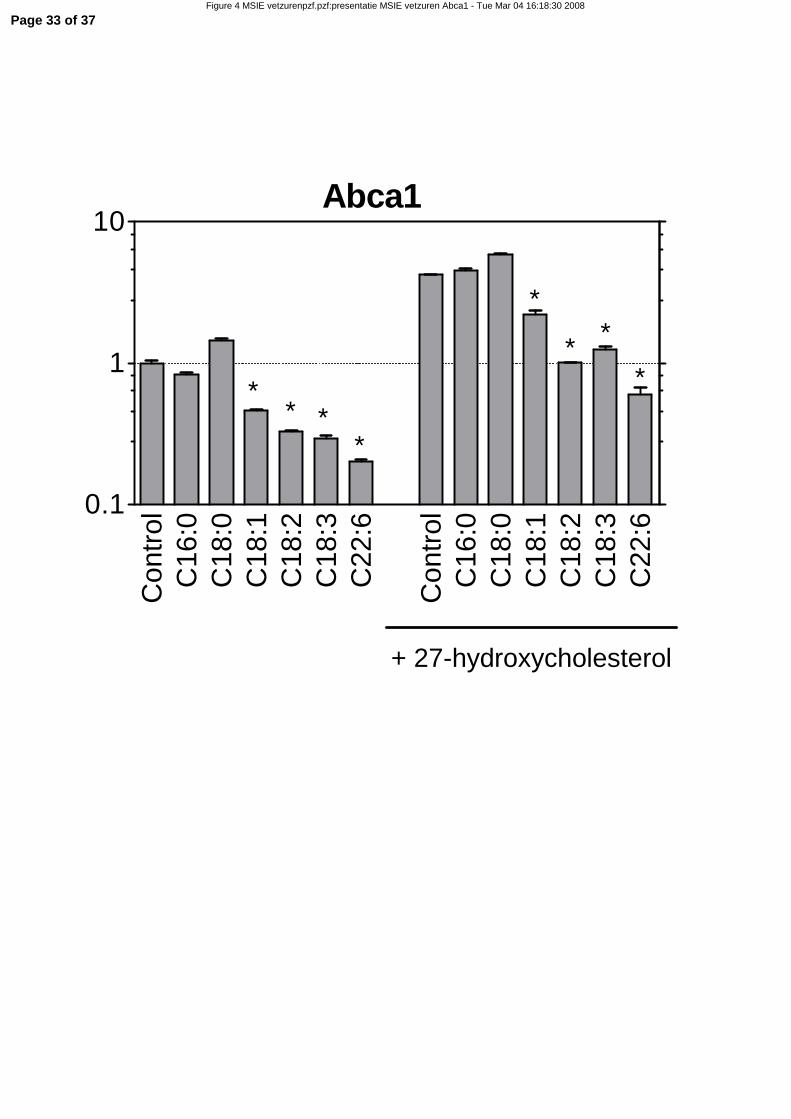
evaluate this possibility, we incubated mouse small intestinal epithelial (MSIE) cells with fatty acids with
and without additional 27-hydroxycholesterol (Figure 4). Because Abcg5, Abcg8, and Npc1l1 were not
expressed in this cell line, only Abca1 expression was used as read-out. As expected, Abca1 was strongly
up-regulated by 27-hydroxycholesterol (5-fold). The unsaturated fatty acids oleic acid, linoleic acid, and
docosahexaenoic acid (DHA) all suppressed expression of Abca1. Even after induction of basal Abca1
expression with 27-hydroxycholesterol, these unsaturated fatty acids were able to down-regulate
expression to a similar extend (Figure 4). The saturated species palmitic and stearic acids did not affect
Abca1 expression. These data indicate that down-regulation of the cholesterol efflux transporter Abca1 is
caused by unsaturated fatty acids. This down-regulation is in contrast to our hypothesis as depicted in
Figure 1, independent on the level of the LXR ligand 27-hydroxycholesterol.
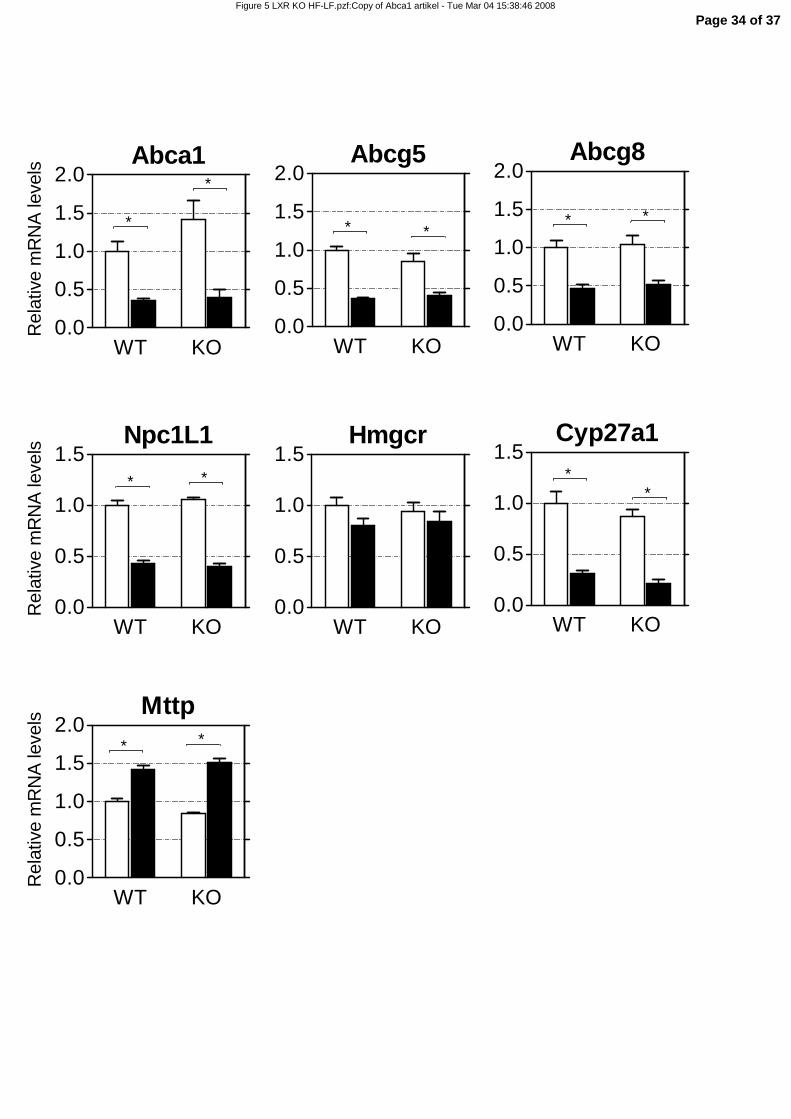
Down-regulation of cholesterol efflux transporters is LXRα-independent
To address whether the fat-induced down-regulation of the ABC transporters is due to interference with
LXRα signaling, we fed LXRα-/- and wild-type mice similar cholesterol-free, high-fat and low-fat diets
for two weeks (Table 1, supplemental data). Gene expression was analyzed with qRT-PCR (Figure 5).
Abca1, Abcg5, and Abcg8 were similarly down-regulated by the high-fat diet in the wild-type mice and
the LXRα-/- mice. Also Npc1l1, Hmgcr, Mttp and Cyp27a1 showed the same differential gene expression
in response to high-fat feeding in wild-type and LXRα-/- mice (Figure 5). This demonstrates that down-
regulation of cholesterol efflux transporters in the small intestine by dietary fat is LXRα-independent.
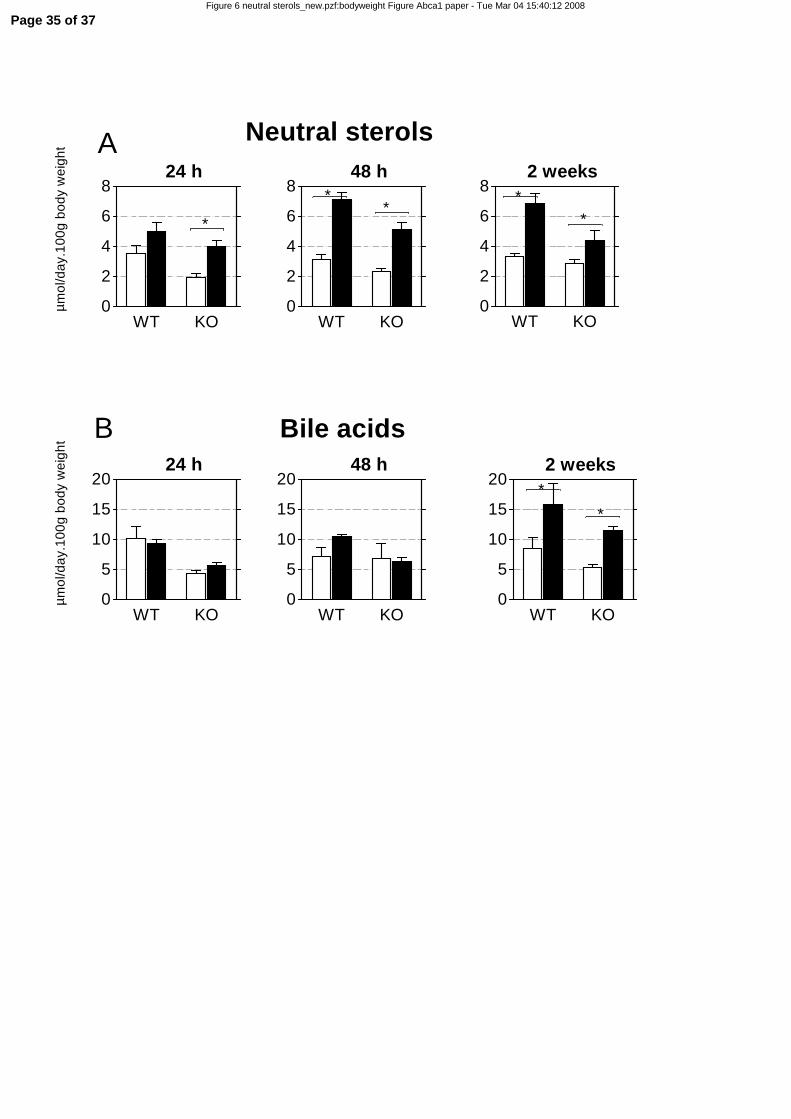
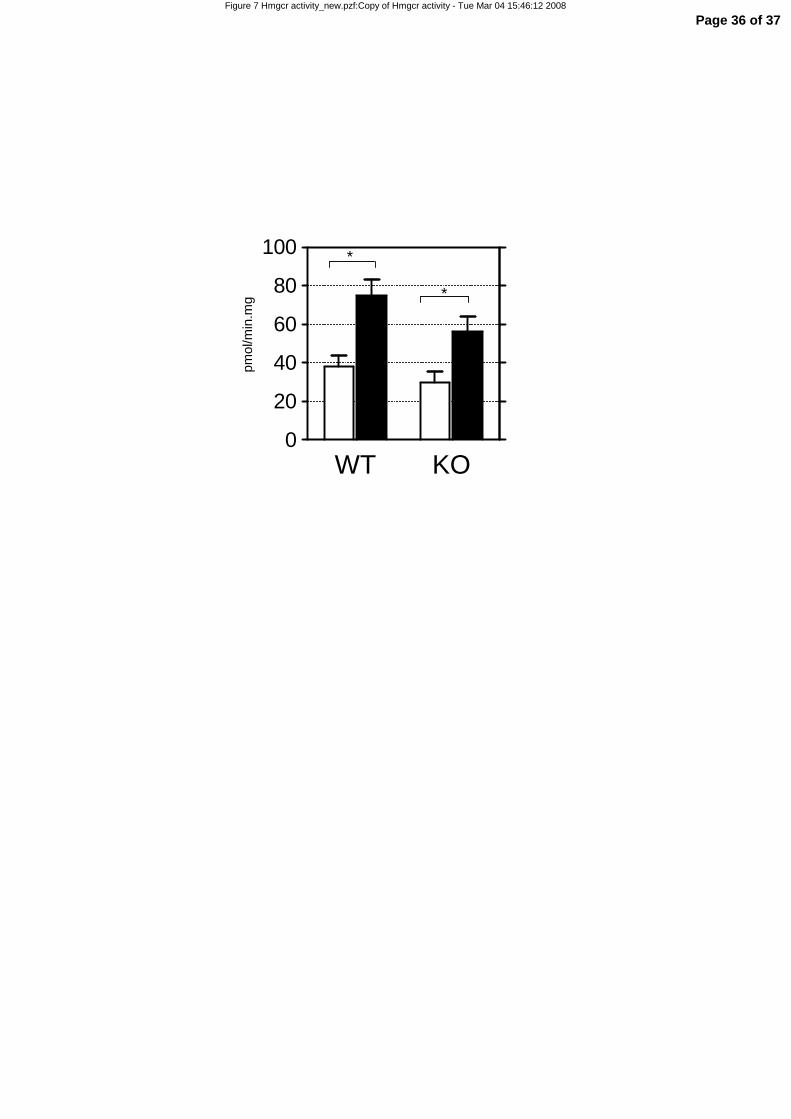
A cholesterol free, high-fat diet increases fecal neutral sterol excretion and small-intestinal
HMGCR activity without affecting plasma cholesterol in mice.
Page 14 of 37

15
As changes in cholesterol absorption generally affects fecal sterol concentrations (11, 12, 24), we
measured neutral sterols in feces of wild-type and LXRα-/- mice fed the low-fat and a high-fat diet for 24
hours, 48 hours, and two weeks (Figure 6). In addition, fecal bile acids were determined. Already on the
first day, the high-fat diet increased the fecal excretion of neutral sterols in wild-type and LXRα-/- mice
(Figure 6A), corroborating the acute down-regulation of Npc1l1, shown in Figure 3. This indicates that
the diminished cholesterol absorption found after 2 weeks on a high fat diet (Figure 2) represents an acute
response to dietary fat. Bile acid excretion, on the other hand, was only elevated after two weeks on a
high-fat diet (Figure 6B).
As no cholesterol was present in the diet and fractional cholesterol absorption was reduced, we
speculated that small intestinal cholesterol synthesis must be increased on a high-fat diet as cholesterol is
needed for chylomicron synthesis. This elevation was not evident at gene expression level (Table 1,
Figure 5), however, it is known that Hmgcr activity is determined by post-transcriptional regulatory
mechanisms (16). Therefore, we measured Hmgcr activity in the small intestine and found it indeed to be
significantly elevated in mice fed a high-fat diet compared to mice fed a low-fat diet for two weeks
(Figure 7).
To determine whether the observed suppression of cholesterol transporter expression is reflected in
plasma cholesterol levels, we measured cholesterol in plasma of FPLC fractions of pooled plasma of
wild-type and LXRα-/- mice fed a low-fat or a high-fat diet for 2 weeks (Figure 8). Separation of plasma
lipoproteins by FPLC revealed that the HDL cholesterol levels were higher in the wild-type than in the
LXRα-/- mice (Figure 8). No changes were found in the HDL-sized fractions in the high-fat fed mice
compared to the low-fat fed mice in both wild-type and LXRα-/- backgrounds.
Page 15 of 37

16
DISCUSSION
The major finding of this study is that, in the murine small intestine, cholesterol efflux transporters
Abca1, Abcg5, and Abcg8 and the cholesterol absorption transporter Npc1l1, as well as the fractional
cholesterol absorption, were all suppressed by feeding a cholesterol-free, high-fat diet to mice. The down-
regulation appeared to be an acute response that is maintained during a chronic diet intervention and is,
surprisingly, not LXRα-dependent as revealed by in-vitro studies and diet intervention in LXRα-null
mice.
We showed with an in-vivo gavage study that down-regulation of cholesterol efflux and absorption
transporters by fatty acids is an acute process. The gavage was given after a fasting period of four hours,
after which the mouse stomach is nearly empty (4). This implies that in this acute experiment only the
gavaged triacylglycerols influence small intestinal gene expression and that the down-regulation of small-
intestinal cholesterol transporters is a fat-induced effect. In these studies we could only apply
triacylglycerols of unsaturated fatty acids, as these are fluid at room temperature. However, saturated and
unsaturated fatty acids were compared by an in-vitro approach, in which only the unsaturated species
showed a down-regulation of Abca1. Based on this observation, we speculate that, also in vivo, only
unsaturated fatty acids induce down-regulation of cholesterol transporters, but this requires further
investigation.
The diminished fractional cholesterol absorption in our study is in line with results reported by
Satchithanandam et al. (35), who found that a high-fat sesame oil diet decreases the lymphatic cholesterol
absorption in the small intestine of rats. In our study, the high-fat diet almost doubled fecal neutral sterol
excretion (Figure 6). However, based on the biliary cholesterol secretion that was not affected by the
high-fat diet (Table 2), we could calculate that the diminished cholesterol absorption should only lead to a
50% increase (i.e. (0.58/0.39-1)*100%) in cholesterol excretion. We suggest that this higher fecal neutral
sterol excretion could be due to a direct cholesterol secretion by the intestinal epithelium, as recently
Page 16 of 37

17
shown by Van der Velde et al. (43). It appears that this novel pathway is stimulated by high-fat diet
feeding in mice. Studies in humans did not show increased neutral sterol levels in feces on a high-fat diet
(10), which might suggest a different effect of a high-fat diet on intestinal cholesterol absorption/secretion
between humans and rodents. Whether this is due to species-dependent differences in fat-induced
regulation of Npc1l1 will be further investigated. As reduced cholesterol absorption on a high-fat intake is
conflicting with the higher need of cholesterol for chylomicron formation, we expected increased
cholesterol synthesis in the enterocyte. However, gene expression data showed that the key enzyme for
cholesterol synthesis, i.e. Hmgcr, was only up-regulated in the acute fatty acid experiment and was not
changed after 2, 4, and 8 weeks of high-fat feeding. Therefore, we additionally measured Hmgcr activity
and it turned out to be increased in the cholesterol-free, high-fat fed mice, implying elevated small
intestinal cholesterol synthesis. This observation is in line with increased intestinal cholesterol synthesis
in rat studies with corn oil by Stange et al. (37, 38). In conjunction with down-regulation of cholesterol
efflux transporters, this increased intestinal cholesterol synthesis probably provides sufficient cholesterol
for chylomicron formation despite the overall reduction in cholesterol absorption.
Fecal neutral sterol levels were higher and fractional cholesterol absorption was lower on a
cholesterol-free, high-fat diet, despite a strongly reduced expression of Abcg5 and Abcg8. As previously
described, deficiency of Abcg5 and Abcg8 leads to no (30) or only mild (51, 53) decrease in fecal neutral
sterol content. Based on these data, we conclude that the effect of down-regulation of cholesterol uptake
(Npc1l1) overrules the effect of down-regulation of the presumed cholesterol efflux transporters (Abcg5
and Abcg8). In the last few years, Npc1l1 has emerged as an important key component of small intestinal
sterol uptake system (2, 13) and here we show that Npc1l1 likely plays a pivotal role in the control of
cholesterol absorption during exposure to a high-fat diet. Future research is needed to investigate the
mechanism of this reduced expression of Npc1l1 during a high-fat diet intervention.
In contrast to the acute elevation of neutral sterols, fecal bile acid secretion was only elevated after 2
weeks of high-fat diet intervention. As a result of a high-fat diet more chylomicrons are formed, causing
Page 17 of 37

18
cholesterol accumulation in the liver (6). Synthesis of bile acids is one of the predominant mechanisms
for the excretion of excess cholesterol from hepatocytes, implying that the observed increase in bile acid
secretion, which reflects hepatic bile acid synthesis, might be a secondary effect of the high-fat diet. This
hypothesis is in line with liver gene expression data that showed that Cyp7a1 was not changed in the
acute experiment but was increased in the long-term high-fat diet intervention (data not shown).
It should be noted that our intervention studies were performed with cholesterol-free, purified low-
and high-fat diets. The in-vitro study showed that the down-regulation of cholesterol transporter genes
was not caused by a lack of cholesterol derivatives as even with addition of 27-hydroxycholesterol
unsaturated fatty acids still could decrease expression of Abca1. To investigate if the down-regulation of
the cholesterol efflux transporters is mediated by LXR we used LXRα-/- mice. As in wild-type and knock-
out mice the same degree of down-regulation was seen, we concluded that the down-regulation is LXRα-
independent. However, it has to be noted that in these LXRα-/- mice LXRβ is still present and might be
able to compensate the loss of LXRα. Although LXR double knock-out mice would be a preferable
model to study LXR involvement, we believe that it is not very likely that LXRβ can completely
compensate for the loss of LXRα. In case of a partial compensation we would have expected a
diminished down-regulation of the cholesterol transporters in the LXRα null mice. So far, compensation
of LXRα by LXRβ in the intestine has not been reported, but it is known that in liver LXRβ is not able to
compensate for the loss of LXRα (29). Moreover, we showed in our in-vitro experiment with a ligand for
both LXRα and LXRβ that unsaturated fatty acids still could down-regulate Abca1.
Duval et al. (14) implied that Npc1l1 is LXR-dependently down-regulated. However, in their study no
LXR-/- mice were included to discern direct or indirect involvement of LXR. Our results indicate that
dietary fat- induced down-regulation of Npc1l1 and the cholesterol efflux transporters in the intestine is
LXRα-independent, which implies that another transcription factor is involved in this process. The study
of Duval et al. (14) furthermore suggests that Npc1l1 is not repressed by PPARα. However, Valasek et al.
Page 18 of 37

19
recently showed that Npc1l1 is PPARα-dependently down-regulated by fenofibrate (41). Furthermore,
from studies in our own lab with wild-type and PPARα-/- mice we know that PPARα activation with
WY14,643 results in reduced levels of Npc1l1 (unpublished data). So, PPARα might be involved in the
fatty acid-dependent down-regulation of Npc1l1 and, thereby, in the control of intestinal cholesterol
absorption on a high-fat diet. On the other hand it is known that Abca1 is PPARα-dependently up-
regulated (19). In addition, Abcg5 and Abcg8 are not known to be regulated by PPARα. This implies that
PPARα is not the common regulator in the fatty acid-induced down-regulation of cholesterol transporters.
This suggests a different mechanism for Npc1l1 and cholesterol efflux transporters. Another potential
candidate regulator is PPARδ, as it is previously described that Npc1l1 is PPARδ-dependently down-
regulated in the murine intestine and CaCo2 cells (42).
In conclusion, our data show that on a cholesterol-free, high-fat diet, fractional cholesterol absorption
is diminished. We propose that, possibly in an attempt to spare intracellular cholesterol for the
chylomicron formation, cholesterol efflux via ABC transporter related pathways is reduced. In addition,
to compensate for reduced uptake of cholesterol in the enterocyte, cholesterol synthesis is induced. This
work shows that the down-regulation of cholesterol transporters is mediated by unsaturated fatty acids.
Studies with LXRα-null mice indicate, surprisingly, that this down-regulation is not dependent on the
presence of LXRα. PPARs might be feasible candidates to regulate cholesterol transporters on a high-fat
diet, but additional studies are required to pinpoint the mechanism by which unsaturated fatty acids down-
regulate cholesterol transporters in the small intestine.
ACKNOWLEDGEMENTS
The authors would like to thank Denise Jonker-Termont, Rick Havinga, Renze Boverhof, and Annelies
Stroeve for excellent technical assistance, Bert Weijers, René Bakker and Juul Baller for skillful
Page 19 of 37

20
biotechnical assistance, Mechteld Grootte Bromhaar and Jenny Jansen for expert microarray
hybridizations, and Bert Groen for the stimulating discussions.
Page 20 of 37

21
REFERENCES
1. Albrink MJ, Glenn WW, Peters JP, and Man EB. The transport of lipids in chyle. J Clin Invest 34: 1467-1475, 1955.2. Altmann SW, Davis HR, Jr., Zhu LJ, Yao X, Hoos LM, Tetzloff G, Iyer SP, Maguire M, Golovko A, Zeng M, Wang L, Murgolo N, and Graziano MP. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 303: 1201-1204., 2004.3. Arca M, Montali A, Ciocca S, Angelico F, and Cantafora A. An improved gas-liquid chromatographic method for the determination of fecal neutral sterols. J Lipid Res 24: 332-335, 1983.4. Barrachina MD, Martinez V, Wang L, Wei JY, and Tache Y. Synergistic interaction between leptin and cholecystokinin to reduce short-term food intake in lean mice. PNAS 94: 10455-10460, 1997.5. Berge KE, Tian H, Graf GA, Yu L, Grishin NV, Schultz J, Kwiterovich P, Shan B, Barnes R, and Hobbs HH. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 290: 1771-1775, 2000.6. Biddinger SB, Almind K, Miyazaki M, Kokkotou E, Ntambi JM, and Kahn CR. Effects of diet and genetic background on sterol regulatory element-binding protein-1c, stearoyl-CoA desaturase 1, and the development of the metabolic syndrome. Diabetes 54: 1314-1323, 2005.7. Bligh EG, and Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37: 911-917, 1959.8. Bosner MS, Lange LG, Stenson WF, and Ostlund RE, Jr. Percent cholesterol absorption in normal women and men quantified with dual stable isotopic tracers and negative ion mass spectrometry. J Lipid Res 40: 302-308, 1999.9. Brown MS, Dana SE, and Goldstein JL. Regulation of 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase Activity in Cultured Human Fibroblasts. Comparison of cells from a normal subject and from a patient with homozygous familial hypercholesterolemia. J Biol Chem 249: 789-796, 1974.10. Brussaard JH, Katan MB, and Hautvast JG. Faecal excretion of bile acids and neutral steroids on diets differing in type and amount of dietary fat in young healthy persons. Eur J Clin Invest 13: 115-122, 1983.11. Chen C-W, and Cheng H-H. A Rice Bran Oil Diet Increases LDL-Receptor and HMG-CoA Reductase mRNA Expressions and Insulin Sensitivity in Rats with Streptozotocin/Nicotinamide-Induced Type 2 Diabetes. J Nutr 136: 1472-1476, 2006.12. Chevallier F, and Lutton C. The intestine is the major site of cholesterol synthesis in the rat. Nat New Biol 242: 61-62, 1973.13. Davis HR, Jr., Zhu L-j, Hoos LM, Tetzloff G, Maguire M, Liu J, Yao X, Iyer SPN, Lam M-H, Lund EG, Detmers PA, Graziano MP, and Altmann SW. Niemann-Pick C1 Like 1 (NPC1L1) Is the Intestinal Phytosterol and Cholesterol Transporter and a Key Modulator of Whole-body Cholesterol Homeostasis. J Biol Chem 279: 33586-33592, 2004.14. Duval C, Touche V, Tailleux A, Fruchart JC, Fievet C, Clavey V, Staels B, and Lestavel S. Niemann-Pick C1 like 1 gene expression is down-regulated by LXR activators in the intestine. Biochem Biophys Res Commun 340: 1259-1263, 2006.
Page 21 of 37

22
15. Francis GA, Knopp RH, and Oram JF. Defective removal of cellular cholesterol and phospholipids by apolipoprotein A-I in Tangier Disease. J Clin Invest 96: 78-87, 1995.16. Goldstein JL, and Brown MS. Regulation of the mevalonate pathway. Nature 343: 425-430, 1990.17. Govers MJ, Termont DS, and Van der Meer R. Mechanism of the antiproliferative effect of milk mineral and other calcium supplements on colonic epithelium. Cancer Res 54: 95-100, 1994.18. Kalopissis AD, Griglio S, and Le Liepvre X. Intestinal very low density lipoprotein secretion in rats fed various amounts of fat. Biochim Biophys Acta 711: 33-39, 1982.19. Knight BL, Patel DD, Humphreys SM, Wiggins D, and Gibbons GF. Inhibition of cholesterol absorption associated with a PPAR alpha-dependent increase in ABC binding cassette transporter A1 in mice. J Lipid Res 44: 2049-2058., 2003.20. Kruit JK, Groen AK, van Berkel TJ, and Kuipers F. Emerging roles of the intestine in control of cholesterol metabolism. World J Gastroenterol 12: 6429-6439, 2006.21. Lee WN, Byerley LO, Bergner EA, and Edmond J. Mass isotopomer analysis: theoretical and practical considerations. Biol Mass Spectrom 20: 451-458, 1991.22. Li T, Chen W, and Chiang JYL. PXR induces CYP27A1 and regulates cholesterol metabolism in the intestine. J Lipid Res 48: 373-384, 2007.23. Lund E, Andersson O, Zhang J, Babiker A, Ahlborg G, Diczfalusy U, Einarsson K, Sjovall J, and Bjorkhem I. Importance of a Novel Oxidative Mechanism for Elimination of Intracellular Cholesterol in Humans. Arterioscler Thromb Vasc Biol 16: 208-212, 1996.24. Lutton C, Magot T, and Chevallier F. Effect of dietary long-chain fatty acids on the rates of cholesterol turnover processes, cholesterol origin and distribution in the rat intestinal lumen. Reprod Nutr Dev 20: 1467-1479, 1980.25. Mero N, Syvanne M, Rosseneu M, Labeur C, Hilden H, and Taskinen MR. Comparison of three fatty meals in healthy normolipidaemic men: high post-prandial retinyl ester response to soybean oil. Eur J Clin Invest 28: 407-415, 1998.26. Mitro N, Mak PA, Vargas L, Godio C, Hampton E, Molteni V, Kreusch A, and Saez E. The nuclear receptor LXR is a glucose sensor. Nature 2006.27. Mulligan JD, Flowers MT, Tebon A, Bitgood JJ, Wellington C, Hayden MR, and Attie AD. ABCA1 Is Essential for Efficient Basolateral Cholesterol Efflux during the Absorption of Dietary Cholesterol in Chickens. J Biol Chem 278: 13356-13366, 2003.28. Murthy S, Born E, Mathur SN, and Field FJ. LXR/RXR activation enhances basolateral efflux of cholesterol in CaCo-2 cells. J Lipid Res 43: 1054-1064, 2002.29. Peet DJ, Turley SD, Ma W, Janowski BA, Lobaccaro JM, Hammer RE, and Mangelsdorf DJ. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 93: 693-704, 1998.30. Plosch T, Bloks VW, Terasawa Y, Berdy S, Siegler K, van der Sluijs F, Kema IP, Groen AK, Shan B, Kuipers F, and Schwartz M. Sitosterolemia in ABC-Transporter G5-deficient mice is aggravated on activation of the liver-X receptor. Gastroenterology 126: 290-300, 2004.31. Plosch T, Kok T, Bloks VW, Smit MJ, Havinga R, Chimini G, Groen AK, and Kuipers F. Increased hepatobiliary and fecal cholesterol excretion upon activation of the liver X receptor is independent of ABCA1. J Biol Chem 277: 33870-33877., 2002.
Page 22 of 37

23
32. Plosch T, van der Veen JN, Havinga R, Huijkman NCA, Bloks VW, and Kuipers F. Abcg5/Abcg8-independent pathways contribute to hepatobiliary cholesterol secretion in mice. Am J Physiol Gastrointest Liver Physiol 291: G414-423, 2006.33. Redgrave TG, Vassiliou GG, and Callow MJ. Cholesterol is necessary for triacylglycerol-phospholipid emulsions to mimic the metabolism of lipoproteins. Biochim Biophys Acta 921: 154-157, 1987.34. Repa JJ, Berge KE, Pomajzl C, Richardson JA, Hobbs H, and Mangelsdorf DJ. Regulation of ATP-binding Cassette Sterol Transporters ABCG5 and ABCG8 by the Liver X Receptors alpha and beta. J Biol Chem 277: 18793-18800, 2002.35. Satchithanandam S, Reicks M, Calvert RJ, Cassidy MM, and Kritchevsky D. Coconut oil and sesame oil affect lymphatic absorption of cholesterol and fatty acids in rats. J Nutr 123: 1852-1858, 1993.36. Schmitz G, and Langmann T. Structure, function and regulation of the ABC1 gene product. Curr Opin Lipidol 12: 129-140, 2001.37. Stange EF, and Dietschy JM. The origin of cholesterol in the mesenteric lymph of the rat. J Lipid Res 26: 175-184, 1985.38. Stange EF, Suckling KE, and Dietschy JM. Synthesis and coenzyme A-dependent esterification of cholesterol in rat intestinal epithelium. Differences in cellular localization and mechanisms of regulation. J Biol Chem 258: 12868-12875, 1983.39. Turley SD, and Dietschy JM. Sterol absorption by the small intestine. Curr Opin Lipidol 14: 233-240, 2003.40. Turley SD, Herndon MW, and Dietschy JM. Reevaluation and application of the dual-isotope plasma ratio method for the measurement of intestinal cholesterol absorption in the hamster. J Lipid Res 35: 328-339, 1994.41. Valasek MA, Clarke SL, and Repa JJ. Fenofibrate reduces intestinal cholesterol absorption via PPARalpha -dependent modulation of NPC1L1 expression in mouse. J Lipid Res 2007.42. van der Veen JN, Kruit JK, Havinga R, Baller JFW, Chimini G, Lestavel S, Staels B, Groot PHE, Groen AK, and Kuipers F. Reduced cholesterol absorption upon PPAR{delta} activation coincides with decreased intestinal expression of NPC1L1. J Lipid Res 46: 526-534, 2005.43. van der Velde AE, Vrins CL, van den Oever K, Kunne C, Oude Elferink RP, Kuipers F, and Groen AK. Direct intestinal cholesterol secretion contributes significantly to total fecal neutral sterol excretion in mice. Gastroenterology 133: 967-975, 2007.44. van Greevenbroek MM, van Meer G, Erkelens DW, and de Bruin TW. Effects of saturated, mono-, and polyunsaturated fatty acids on the secretion of apo B containing lipoproteins by Caco-2 cells. Atherosclerosis 121: 139-150, 1996.45. Venkateswaran A, Laffitte BA, Joseph SB, Mak PA, Wilpitz DC, Edwards PA, and Tontonoz P. Control of cellular cholesterol efflux by the nuclear oxysterol receptor LXRalpha. PNAS 97: 12097-12102, 2000.46. Wang DQ. Regulation of intestinal cholesterol absorption. Annu Rev Physiol 69: 221-248, 2007.47. Wang HH, Patel SB, Carey MC, and Wang DQ. Quantifying anomalous intestinal sterol uptake, lymphatic transport, and biliary secretion in Abcg8(-/-) mice. Hepatology 45: 998-1006, 2007.
Page 23 of 37

24
48. Wang X, and Seed B. A PCR primer bank for quantitative gene expression analysis. Nucleic Acids Res 31: e154, 2003.49. Whitehead RH, VanEeden PE, Noble MD, Ataliotis P, and Jat PS. Establishment of Conditionally Immortalized Epithelial Cell Lines from both Colon and Small Intestine of Adult H-2Kb-tsA58 Transgenic Mice. PNAS 90: 587-591, 1993.50. Wood P, Imaichi K, Knowles J, Michaels G, and Kinsell L. The Lipid Composition Of Human Plasma Chylomicrons. J Lipid Res 5: 225-231, 1964.51. Yu L, Hammer RE, Li-Hawkins J, Von Bergmann K, Lutjohann D, Cohen JC, and Hobbs HH. Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc Natl Acad Sci U S A 99: 16237-16242., 2002.52. Yu L, Li-Hawkins J, Hammer RE, Berge KE, Horton JD, Cohen JC, and Hobbs HH. Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J Clin Invest 110: 671-680, 2002.53. Yu L, York J, von Bergmann K, Lutjohann D, Cohen JC, and Hobbs HH. Stimulation of Cholesterol Excretion by the Liver X Receptor Agonist Requires ATP-binding Cassette Transporters G5 and G8. J Biol Chem 278: 15565-15570, 2003.
Page 24 of 37

25
FIGURE LEGENDS
Figure 1. Hypothesis for the high-fat induced suppression of cholesterol efflux transporters.
The enterocyte cartoon shows a schematic overview of the microarray results. White means not regulated,
light grey means down-regulated, and dark grey means up-regulated. = chylomicron. = 27-
hydroxycholesterol, its concentration was not measured. Based on the microarray data we hypothesized
that on a cholesterol-free, high-fat diet, cholesterol absorption is diminished and that cholesterol in the
enterocyte is mainly used for chylomicron formation to transport fat out of the enterocyte. Therefore, less
cholesterol is converted to the LXR ligand 27-hydroxycholesterol via Cyp27a1, leading to a possible
down-regulation of LXR target genes Abca1, Abcg5, and Abcg8.
Figure 2. Fractional cholesterol absorption in high-fat and low-fat fed C57BL/6J mice.
Cholesterol absorption in high-fat (HF, black bar) and low-fat (LF, white bar) fed mice was measured
using the plasma dual-isotope method (LF: n = 5, HF: n=8). After 2 weeks of treatment, mice received an
intravenous injection of [3H]cholesterol and an oral dose of [14C]cholesterol. Plasma samples obtained 72
h after administration were used for the calculation of fractional cholesterol absorption. Data are
presented as mean ± standard error, n=5 (LF), n=8 (HF) * Significant differences by Student’s t-test (P<
0.05).
Figure 3. Relative mRNA levels in small intestines of SV129 mice gavaged with
triacylglycerols. 4-5 month-old male SV129 mice were given synthetic triacylglycerols via oral gavage.
mRNA expression was measured by qRT-PCR, the control was set to 1. Significance was determined
using an unpaired Student’s t-test, and was for all treatments calculated compared to the control. * P-
value < 0.05. Data are presented as mean ± standard error, n=4.
27-OH
Page 25 of 37

26
Figure 4. Effect of 27-hydroxycholesterol and fatty acids on gene expression of Abca1 in MSIE
cells. Confluent MSIE cells were incubated for 6 hours with 500 µM fatty acids and/or 5µM 27-
hydroxycholesterol. mRNA expression was measured by qRT-PCR. The Y-axes is in a log 10 scale.
Significance was determined using an unpaired Student’s t-test, and was for all treatments calculated
compared to the control. * P-value < 0.05. Data are presented as mean ± standard error, n=3.
Figure 5. Relative mRNA levels in small intestines of wild-type and LXRα-/- mice fed a high-
fat diet for 2 weeks. 4-5 month-old female wild-type (WT) and LXRα-/- (KO) mice were fed a low-fat
(LF) or high-fat (HF), cholesterol-free diet for two weeks. mRNA expression was measured by qRT-PCR.
White bars represent the LF diet group, black bars represent the HF diet group. WT LF values were set to
1. Significance was determined using an unpaired Student’s t-test. * P-value < 0.05. Data are presented as
mean ± standard error, n=6 (WT), n=4 (KO).
Figure 6. Fecal neutral sterol and bile acid excretion in wild-type and LXRα-/- mice fed high-
and low-fat diets. 4-5 month-old female wild-type (WT) and LXRα-/- (KO) mice were fed a low-fat (LF)
or high-fat (HF) for two weeks. Feces were collected after 24 and 48 hours and after two weeks of diet
intervention and analyzed for neutral sterol and bile acid concentrations. White bars represent the LF diet
group, black bars represent the HF diet group. Neutral sterol and bile acid concentrations were normalized
for body weight. Significance was determined using an unpaired Student’s t-test. * P-value < 0.05. Data
are presented as mean ± standard error, n=3 (WT), n=2 (KO). A, neutral sterol concentrations in feces
after 24 hours, 48 hours, and 2 weeks. B, bile acid concentrations in feces after 24 hours, 48 hours, and 2
weeks.
Figure 7. Hmg-CoA reductase activity in intestinal scrapings of high-fat and low-fat fed wild-
type and LXRα-/- mice. 4-5 month-old female wild-type (WT) and LXRα-/- (KO) mice were fed a low-
Page 26 of 37

27
fat (LF) or high-fat (HF) for two weeks. Hmg-CoA reductase activity was measured in scrapings of the
proximal part of the small intestine. White bars represent the LF diet group, black bars represent the HF
diet group. Significance was determined using an unpaired Student’s t-test. * P-value < 0.05. Data are
presented as mean ± standard error, n=6 (WT), n=4 (KO)
Figure 8. Fast-protein liquid chromatography (FPLC) separation of plasma lipoproteins of
high-fat and low-fat fed wild-type and LXRα-/- mice. 4-5 month-old female wild-type (WT) and
LXRα-/- (KO) mice were fed a low-fat (LF) or high-fat (HF) for two weeks. Plasma from all individual
mice per group was pooled and subjected to gel filtration using Superose 6 columns. Cholesterol content
in each fraction was measured. HDL, high-density lipoprotein; LDL, low-density lipoprotein. A, WT
mice; B, KO mice.
Page 27 of 37

Table 1: Microarray results high-fat diet intervention in C57Bl/6J mice
Gene
symbol
Affy probe
set ID
FC wk 2
HF vs LF
FC wk 4
HF vs LF
FC wk 8
HF vs LF
FC qRT-PCR
wk 2 HF vs LF Gene name
Abca1 1421840_at -2.4 -2.0 -2.0 -3.5 ± 0.6* ATP-binding cassette, sub-family A, member 1
Abcg5 1419393_at -1.8 -1.7 -1.6 -2.1 ± 0.2 * ATP-binding cassette, sub-family G, member 5
Abcg8 1420656_at -1.9 -2.0 -1.4 -1.7 ± 0.2 * ATP-binding cassette, sub-family G, member 8
Npc1L1 1438514_at -1.5 -1.5 -1.5 -1.6 ± 0.1 * Niemann-Pick C1 Like 1 protein
Mttp 1419399_at 1.3 1.4 1.3 1.6 ± 0.2 * Microsomal triglyceride transfer protein
Hmgcr 1427229_at NC NC NC -1.2 ± 0.1 3-hydroxy-3-methylglutaryl-Coenzyme A reductase
Cyp27a1 1417590_at -2.9 -2.8 -2.0 -3.9 ± 0.5 * Cytochrome P450, family 27, subfamily a, polypeptide 1
C57BL/6J mice were fed a cholesterol free, low- (LF) or high-fat (HF) diet for 2, 4, and 8
weeks. Microarray analysis was performed on RNA isolated from the middle segment of the
small intestine and fold changes (FC) are given for week (wk) 2, 4, and 8. The fold changes of wk
2 were confirmed by qRT-PCR and presented as mean ± standard error, n=6. NC, not changed;
*Significant difference compared to the LF group (Student’s t-test, P-value< 0.05).
Page 28 of 37

Table 2. Bile flow and biliary secretion rates in high-fat and low-fat fed C57BL/6J mice
Low-fat High-fat
Bile flow (µl/ 15min) 16.3 ± 1.8 17.0 ± 1.3
Bile salts (nmol/ min.100 g body weight) 196.3 ± 45.6 194.3 ± 24.9
Cholesterol (nmol/ min.100 g body weight) 1.4 ± 0.2 1.8 ± 0.2
Phospholipids (nmol/ min.100 g body weight) 24.2 ± 3.1 28.5 ± 3.3
Values are expressed as mean ± standard error. Low-fat: n=5, high-fat: n=8.
Page 29 of 37

Lumen
Lymph
HF diet
Abca1
Abcg5/8
Hmgcr
Cyp27a1
Biliary Cholesterol
Mttp
Lxr
Npc1l1
27-OH
Page 30 of 37

0
25
50
75
100%
abs
orpt
ion
*
Figure 2 cholesterol absorption.pzf:cholesterol absorption - Tue Mar 04 15:48:34 2008
Page 31 of 37

Ab
ca1Control
C18:1
C18:2
C18:3
C20:5
C22:6
0.0
0.5
1.0
1.5A
bcg
5
Control
C18:1
C18:2
C18:3
C20:5
C22:6
0.0
0.5
1.0
1.5A
bcg
8
Control
C18:1
C18:2
C18:3
C20:5
C22:6
0.0
0.5
1.0
1.5
Np
c1L1
Control
C18:1
C18:2
C18:3
C20:5
C22:6
0.0
0.5
1.0
1.5C
yp27a1
ControlC18:1 C18:2 C18:3 C20:5 C22:6
0.0
0.5
1.0
1.5
Hm
gcr
ControlC18:1C18:2C18:3C20:5C22:6
0.00.5
1.0
1.5
2.0
2.5M
ttp
ControlC18:1C18:2C18:3C20:5C22:6
0.0
0.5
1.0
1.5
2.0
2.5
Relative mRNA levelsRelative mRNA levelsRelative mRNA levels
**
**
**
**
**
**
**
*
**
**
**
**
**
**
*
**
**
*
Figure 3 acute.pzf:acute figure A
bca1 artikel - Tue M
ar 04 15:49:42 2008
Page 32 of 37
Abca1C
ontr
olC
16:0
C18
:0C
18:1
C18
:2C
18:3
C22
:6
Con
trol
C16
:0C
18:0
C18
:1C
18:2
C18
:3C
22:6
0.1
1
10
** *
*
*
**
*
+ 27-hydroxycholesterol
Figure 4 MSIE vetzurenpzf.pzf:presentatie MSIE vetzuren Abca1 - Tue Mar 04 16:18:30 2008
Page 33 of 37
Abca1
WT KO0.0
0.5
1.0
1.5
2.0Abcg5
WT KO0.0
0.5
1.0
1.5
2.0Abcg8
WT KO0.0
0.5
1.0
1.5
2.0
Npc1L1
WT KO0.0
0.5
1.0
1.5Hmgcr
WT KO0.0
0.5
1.0
1.5
Mttp
WT KO0.0
0.5
1.0
1.5
2.0
Cyp27a1
WT KO0.0
0.5
1.0
1.5
Rel
ativ
e m
RN
A le
vels
Rel
ativ
e m
RN
A le
vels
Rel
ativ
e m
RN
A le
vels
*
*
* ** *
* *
* *
**
Figure 5 LXR KO HF-LF.pzf:Copy of Abca1 artikel - Tue Mar 04 15:38:46 2008
Page 34 of 37
24 h
WT KO0
2
4
6
8 48 h
WT KO0
2
4
6
8 2 weeks
WT KO0
2
4
6
8
24 h
WT KO0
5
10
15
20 48 h
WT KO0
5
10
15
20 2 weeks
WT KO0
5
10
15
20
A
B
Neutral sterols
Bile acids
*
**
**
µ mol
/day
.100
g bo
dy w
eigh
t
*
*
µ mol
/day
.100
g bo
dy w
eigh
t
Figure 6 neutral sterols_new.pzf:bodyweight Figure Abca1 paper - Tue Mar 04 15:40:12 2008
Page 35 of 37
WT KO0
20
40
60
80
100pm
ol/m
in.m
g *
*
Figure 7 Hmgcr activity_new.pzf:Copy of Hmgcr activity - Tue Mar 04 15:46:12 2008
Page 36 of 37

Wild type
0 10 20 30 40 500.0
0.1
0.2
0.3
0.4
0.5WT LFWT HF
Fraction
Ch
ole
ster
ol c
on
cen
trat
ion
mM
LXRαααα -/-
0 10 20 30 40 500.0
0.1
0.2
0.3
0.4
0.5KO LFKO HF
Fraction
Cho
lest
erol
con
cent
rati
on m
M
HDL
HDL
A B
Figure 8 cholesterol fractions_new.pzf:Figure cholesterol fractions paper - Tue Mar 04 15:47:45 2008
Page 37 of 37
Related Documents







![Heterogeneity across the murine small and large intestine · whereas those in the large intestine drive the synthesis of β-defensins[25] and cathelicidins[26]. As well as differences](https://static.cupdf.com/doc/110x72/5f48175dc3e01178682dfc5e/heterogeneity-across-the-murine-small-and-large-intestine-whereas-those-in-the-large.jpg)



