Transglutaminase Activity Is Involved in Polyamine-Induced Programmed Cell Death Francesco Facchiano,* ,1 Daniela D’Arcangelo,* Antonella Riccomi,* Alessandro Lentini,² Simone Beninati,² and Maurizio C. Capogrossi* *Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, Istituto di Ricovero e Cura a Carattere Scientifico, Rome, Italy; and ²Dipartimento di Biologia, Laboratorio di Biologia Cellulare, Universita ` di Tor Vergata, Rome, Italy Natural polyamines, i.e., putrescine, spermidine, and spermine, are ubiquitous molecules essential for cell proliferation and differentiation. In the present study, the effect of polyamines on primary cultures of bovine aortic endothelial cells (BAECs), rat aortic smooth muscle cells (RASMCs), and a human mela- noma cell line was examined. While in the absence of fetal calf serum (FCS) polyamines had no effect on viability, in the presence of FCS spermidine and spermine, at concentrations close to physiologic lev- els, induced a dose-dependent cell death, whereas pu- trescine was ineffective. RASMCs were significantly more sensitive than other cells. FACS analysis, oligo- nucleosome ELISA, Hoechst nuclear staining, and An- nexin V-FITC quantification showed that cell death was likely due to apoptosis. Cells exposed to spermi- dine showed a marked increase of intracellular trans- glutaminase (TGase) activity (;30-fold over control). Inhibitors of polyamine oxidation or inhibitors of TGase activity prevented polyamine-induced apopto- sis. Moreover, tissue TGase overexpression signifi- cantly increased cell sensitivity to polyamine, suggest- ing that this effect is likely related to enhanced intracellular TGase activity. These data indicate that polyamines may modulate cell viability through a novel TGase-dependent process. © 2001 Academic Press Key Words: angiogenesis; apoptosis; melanoma cell; polyamine; programmed cell death; transglutaminase; vascular cells. INTRODUCTION The polyamines putrescine, spermidine, and sperm- ine are polycations naturally occurring in both pro- karyotic and eukaryotic organisms [1, 2] where they exert a wide range of biological functions, like protein synthesis and cell division control [2], protein and nu- cleic acid structure stabilization, and ion channel con- trol [1, 3]. In addition, synthesis of polyamines is rap- idly induced when cells are stimulated to grow and divide [1, 2]. Therefore, polyamines have been indi- cated as potential targets for therapeutic interventions [2, 4]. In eukaryotic organisms, ornithine decarboxyl- ase catalyzes the key step in polyamine biosynthesis. Polyamine catabolism may occur via three main bio- chemical pathways: acetylation, protein transamida- tion, and oxidative degradation. Acetylation is a rate- limiting step in spermidine and spermine catabolism and occurs through the action of the spermidine/sperm- ine N1-acetyltransferase [5]. Protein transamidation is catalyzed by transglutaminase (TGase, EC 2.3.2.13), which covalently cross-links polyamines to specific glu- tamine residues in proteins and appears to modulate polyamine availability in cells [6, 7]. Oxidative degra- dation of polyamine leads to putrescine from spermine and spermidine [8]. Polyamines content has been mea- sured in human tissues and physiologic fluids and their baseline levels are reported to be elevated in various pathologic conditions. Several studies hypothesized that polyamines play a role in wound healing and promote angiogenesis; however, mechanisms involved are not yet completely elucidated [9 –11]. It has been shown that polyamines, at very high concentration, are toxic in the presence of ruminant serum, but molecular mechanisms underlying this toxicity are still unclear. On the other hand, spermine at high concentration protects thymocytes from apoptosis [12], while its oxi- dation leads to necrosis of leukemia cells [13]. There- fore, further study is needed to clarify the molecular mechanisms responsible for polyamine toxicity and to investigate cell type specificity. Further, polyamine ef- fect on primary cultures of smooth muscle cells (SMCs), i.e., cells directly exposed to serum factors when an endothelial damage occurs, has never been evaluated. In the present study, the effect of three natural polyamines was examined at concentrations close to circulating serum levels on primary cultures of vascular endothelial and SMCs and on a human mel- anoma cell line. It was found that spermidine and spermine, but not putrescine, induced cell death, likely due to apoptosis, in the presence of FCS. A key role for 1 To whom reprint requests should be addressed at Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, via Monti di Creta 104, 00167 Rome, Italy. Fax: 139.06.66462430. E-mail: [email protected]. Experimental Cell Research 271, 118 –129 (2001) doi:10.1006/excr.2001.5356, available online at http://www.idealibrary.com on 118 0014-4827/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Cell Research 271, 118–129 (2001)doi:10.1006/excr.2001.5356, available online at http://www.idealibrary.com on
Transglutaminase Activity Is Involved in Polyamine-InducedProgrammed Cell Death
Francesco Facchiano,*,1 Daniela D’Arcangelo,* Antonella Riccomi,* Alessandro Lentini,†Simone Beninati,† and Maurizio C. Capogrossi*
*Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, Istituto di Ricovero e Cura a Carattere Scientifico, Rome, Italy;and †Dipartimento di Biologia, Laboratorio di Biologia Cellulare, Universita di Tor Vergata, Rome, Italy
Natural polyamines, i.e., putrescine, spermidine,and spermine, are ubiquitous molecules essential forcell proliferation and differentiation. In the presentstudy, the effect of polyamines on primary cultures ofbovine aortic endothelial cells (BAECs), rat aorticsmooth muscle cells (RASMCs), and a human mela-noma cell line was examined. While in the absence offetal calf serum (FCS) polyamines had no effect onviability, in the presence of FCS spermidine andspermine, at concentrations close to physiologic lev-els, induced a dose-dependent cell death, whereas pu-trescine was ineffective. RASMCs were significantlymore sensitive than other cells. FACS analysis, oligo-nucleosome ELISA, Hoechst nuclear staining, and An-nexin V-FITC quantification showed that cell deathwas likely due to apoptosis. Cells exposed to spermi-dine showed a marked increase of intracellular trans-glutaminase (TGase) activity (;30-fold over control).Inhibitors of polyamine oxidation or inhibitors ofTGase activity prevented polyamine-induced apopto-sis. Moreover, tissue TGase overexpression signifi-cantly increased cell sensitivity to polyamine, suggest-ing that this effect is likely related to enhancedintracellular TGase activity. These data indicate thatpolyamines may modulate cell viability through anovel TGase-dependent process. © 2001 Academic Press
Key Words: angiogenesis; apoptosis; melanoma cell;polyamine; programmed cell death; transglutaminase;vascular cells.
INTRODUCTION
The polyamines putrescine, spermidine, and sperm-ine are polycations naturally occurring in both pro-karyotic and eukaryotic organisms [1, 2] where theyexert a wide range of biological functions, like proteinsynthesis and cell division control [2], protein and nu-cleic acid structure stabilization, and ion channel con-
1 To whom reprint requests should be addressed at Laboratorio diPatologia Vascolare, Istituto Dermopatico dell’Immacolata, viaMonti di Creta 104, 00167 Rome, Italy. Fax: 139.06.66462430.E-mail: [email protected].
1180014-4827/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
trol [1, 3]. In addition, synthesis of polyamines is rap-idly induced when cells are stimulated to grow anddivide [1, 2]. Therefore, polyamines have been indi-cated as potential targets for therapeutic interventions[2, 4]. In eukaryotic organisms, ornithine decarboxyl-ase catalyzes the key step in polyamine biosynthesis.Polyamine catabolism may occur via three main bio-chemical pathways: acetylation, protein transamida-tion, and oxidative degradation. Acetylation is a rate-limiting step in spermidine and spermine catabolismand occurs through the action of the spermidine/sperm-ine N1-acetyltransferase [5]. Protein transamidation iscatalyzed by transglutaminase (TGase, EC 2.3.2.13),which covalently cross-links polyamines to specific glu-tamine residues in proteins and appears to modulatepolyamine availability in cells [6, 7]. Oxidative degra-dation of polyamine leads to putrescine from spermineand spermidine [8]. Polyamines content has been mea-sured in human tissues and physiologic fluids and theirbaseline levels are reported to be elevated in variouspathologic conditions. Several studies hypothesizedthat polyamines play a role in wound healing andpromote angiogenesis; however, mechanisms involvedare not yet completely elucidated [9–11]. It has beenshown that polyamines, at very high concentration, aretoxic in the presence of ruminant serum, but molecularmechanisms underlying this toxicity are still unclear.On the other hand, spermine at high concentrationprotects thymocytes from apoptosis [12], while its oxi-dation leads to necrosis of leukemia cells [13]. There-fore, further study is needed to clarify the molecularmechanisms responsible for polyamine toxicity and toinvestigate cell type specificity. Further, polyamine ef-fect on primary cultures of smooth muscle cells(SMCs), i.e., cells directly exposed to serum factorswhen an endothelial damage occurs, has never beenevaluated. In the present study, the effect of threenatural polyamines was examined at concentrationsclose to circulating serum levels on primary cultures ofvascular endothelial and SMCs and on a human mel-anoma cell line. It was found that spermidine andspermine, but not putrescine, induced cell death, likelydue to apoptosis, in the presence of FCS. A key role for

cwpatctaia
awt
100 ml. The reaction was stopped as described [19]. The neutralizing
1eiP[s
Bcwwnm
w
119POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
enhanced intracellular transglutaminase (TGase) ac-tivity, likely induced by an oxidative product of poly-amine degradation, was identified. These findings rep-resent the first evidence that spermidine andspermine, at concentrations within the physiologicrange, induce cell apoptosis in a TGase-dependentmanner. In conclusion, the reported results reveal anovel apoptotic pathway and suggest a function fortissue transglutaminase in apoptotic processes.
MATERIALS AND METHODS
Cells. Primary cultures of bovine aortic endothelial cells (BAECs)were prepared, cultured, and checked for cell culture purity as de-scribed [14]. All experiments were performed with BAECs at pas-sages 2–7, from three different preparations.
Primary rat aorta smooth muscle cells (RASMCs) were prepared,cultured, and checked for cell culture purity as described [15]. Allexperiments were performed with RASMCs at passages 4–6, fromthree different preparations. Additional experiments were carriedout with bovine aortic smooth muscle cells, with human umbilicalvein endothelial cells (HUVECs, Mascia Brunelli), and with a humancutaneous melanoma cell line (Mel 120) kindly supplied by Dr.Michele Maio (Aviano, Italy). The investigation conforms to theGuide for the Care and Use of Laboratory Animals published by theUS National Institutes of Health (NIH Publication 85-23, revised1996).
Cell culture and proliferation assay. Cells were maintained inDulbecco’s modified Eagle’s medium (DMEM) supplemented with10% fetal calf serum (FCS) (Hyclone Inc.), 2 mM L-glutamine, and100 IU/ml penicillin/streptomycin (Gibco BRL) in humidified 5% CO2
atmosphere, at 37°C. Three batches of FCS were used with similarresults FCS for RASMCs was heated for 30 min at 56°C to inactivatethe complement. Cells were passaged by using trypsin/EDTA. Forproliferation assays, BAECs (5 3 104 cells), RASMCs (1 3 105 cells),and Mel 120 (1 3 105 cells) were seeded onto uncoated six-well tissueulture plates. After 24 h, the medium was replaced with DMEMithout serum; 48 h later, medium was replaced with DMEM sup-lemented with 10% FCS containing different concentrations of poly-mines (Sigma Chemical Co.) with or without inhibitors. At differentime points, adherent cells were harvested with trypsin/EDTA andounted with a hemacytometer. Cell viability was evaluated byrypan blue exclusion assay. Polyamine oxidation was inhibited bydding 20 mM pentamidine or 1 mM aminoguanidine (Sigma Chem-cals Co.) to the culture medium. TGase activity was inhibited bydding monodansylcadaverine (MDC, 0–150 mM) (Sigma Chemicals
Co.) or 300 mg/ml RS-10025-002 (Syntex Inc.), a 3-halo-dihydroisox-zole compound specifically developed to inhibit TGase [16]. To checkhether TGase inhibition might interfere with polyamine oxidation,
he rate of degradation of [14C]spermine to spermidine and to pu-trescine, was assayed (1 h at 37°C) in the presence of purified amineoxidase from rat liver [17] and in the presence of MDC at 10 and 100mM. Under these conditions, oxidase activity was unchanged, whilealmost complete inhibition (98%) of polyamine oxidation wasachieved in samples treated with 10 mM MDL-72,527 (not shown).Purified catalase activity was checked according to the manufactur-er’s instructions (Sigma Chemical Co.) and added to the medium (300U/ml final concentration) at the beginning of the 24 h incubation.Heat denaturation of FCS was carried out by heating 15 min at100°C.
TGase activity. Total TGase enzymatic activity was assayed inprotein extracts prepared as described [18]. Aliquots of 300 mg ofproteins were incubated at 37°C for 30 min in assay buffer (10 mMtris-aminomethane, Tris, pH 7.8, 800 mM CaCl2, 14 mM dithiotreitol,2 mM MgCl2, 5 mg/ml dimethylcasein, Sigma Chemicals Co., and 4mCi [3H]spermidine, Amersham Life Science Inc.) in a final volume of
anti-TGase antibody blocking the extracellular TGase activity wasprepared and tested as described [20]. For intracellular TGase ac-tivity (in vivo TGase assay), cells plated on 100-mm petri dishes (1 306) were grown in the presence of 3H-labeled spermidine (3 mCi/ml),ither in the absence or in the presence of spermidine. After 24 hncubation, cells were detached by scraping, washed three times withBS, and proteins were trichloroacetic acid-precipitated as described
19]. Total polyamine content and TGase specific activity were mea-ured as described [21].Flow cytometric cell cycle analysis and evaluation of apoptosis.AECs and RASMCs plated onto six-well plates (1 3 105 cells) were
ultured in the presence of polyamines, trypsinized, washed onceith PBS, and fixed in 70% cold ethanol. Fixed cells were washedith PBS containing 0.1% bovine serum albumin (Santa Cruz Biotech-ology, Inc.), incubated with 50 mg/ml ribonuclease A (RNase A) for 30in at 37°C and 50 mg/ml propidium iodide, according to standard
protocols, and gated out electronically [22]. The relative DNA contentand cell distribution in various cell cycle compartments were deter-mined with a FACScan Becton Dickinson Instrument and theCellQuest software. Quantification of apoptosis in cells was performedby means of photometric enzyme immunoassay to determine cytoplas-mic mono- and oligo-nucleosomes according to manufacturer’s instruc-tions (Cell Death Plus, Boehringer Mannheim). Results were expressedas optical density (A405 nm 2 A490 nm) after blank subtraction. An addi-tional evaluation of apoptosis was performed by means of the AnnexinV-FITC Apoptosis detection kit II (Pharmingen). Cells (1 3 105) wereplated in six-well plates, and exposed for 24 h to polyamines in thepresence of serum, then harvested and stained according to manufac-turer’s instructions. Cells positive for PI, for Annexin V-FITC or for bothwere quantified by flow cytometry. Apoptosis was also evaluated byHoechst 33258 dye nuclear staining, as reported [23].
DNA transfection. Transient transfection was achieved by cal-cium phosphate-DNA coprecipitation, as reported [24]. An expres-sion vector previously described [25] (generous gift of Dr. VittorioGentile, Naples) was used to overexpress tissue TGase in primarycultures of vascular cells. Plasmid was purified as described [26].BAECs and RASMCs (5 3 105) plated in 100-mm petri dishes werecotransfected with 20 mg of purified pSG5-TGase plasmid or emptyvector (mock transfected cells) and with 2 mg of a pNeoEGFP plasmidcontaining the G418 resistance gene (Clontech Laboratories), ex-pressing green fluorescent protein. Transfected cells were culturedfor 3 days with G418 (0.5 mg/ml, Gibco, BRL) to enrich the cellpopulation of transfected cells. Total TGase enzymatic activity wasmeasured in cell extracts by incorporation of labeled polyamine intodimethylcasein, in TGase-plasmid transfected BAECs and RASMCsand it was compared to mock-transfected cells. Transfected BAECsand RASMCs showed a total TGase activity at least 15 or 18 timeshigher than mock transfected, respectively. Enzymatic activity inmock transfected cells was 2.9 and 0.22 pmol of [3H]spermidineincorporated/h/mg of protein, for BAEC and RASMC, respectively.Cotransfection with two vectors, at the reported ratio 10:1, results inthe internalization of both plasmids by the same cell [27]. Therefore,sensitivity to spermidine was evaluated by counting, under UV lightmicroscopy, GFP positive cells only, in order to overcome the poten-tial limitations of low transfection efficiency. Transfection efficiencywas higher than 40%.
Statistical analysis. Data were expressed as mean 6 standarddeviation. Student’s two-tailed t test was performed and a P # 0.05
as considered statistically significant.
RESULTS
Effects of Polyamines on Vascular andMelanoma Cells
The time-dependent effect of 40 mM putrescine,spermidine, and spermine on BAEC, RASMC, and
R
mvpSm
1
zrtf
120 FACCHIANO ET AL.
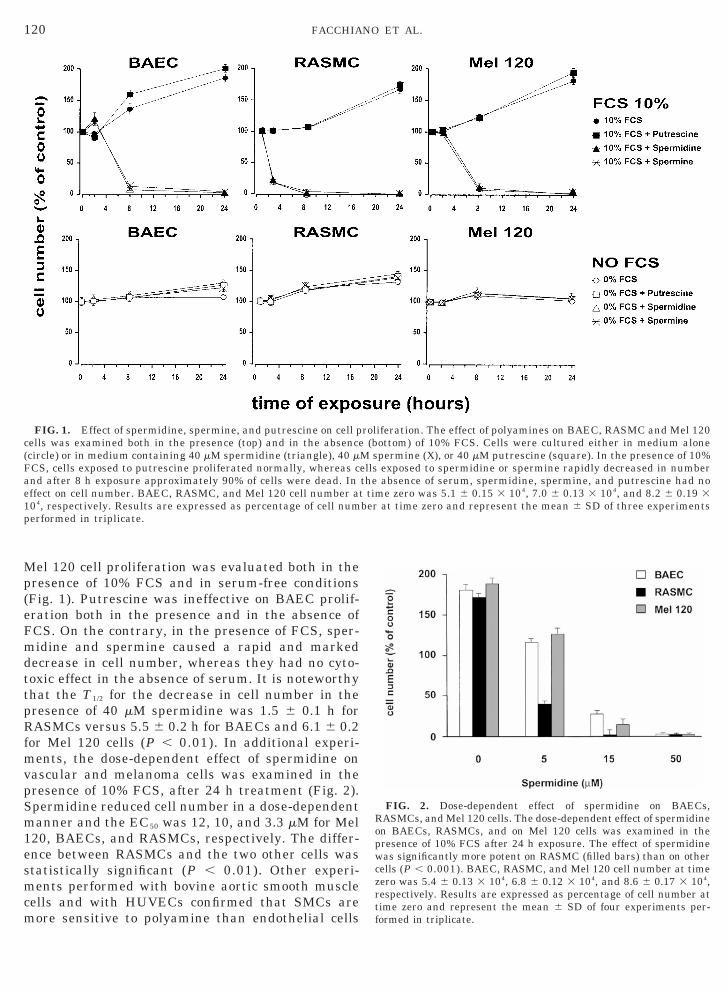
Mel 120 cell proliferation was evaluated both in thepresence of 10% FCS and in serum-free conditions(Fig. 1). Putrescine was ineffective on BAEC prolif-eration both in the presence and in the absence ofFCS. On the contrary, in the presence of FCS, sper-midine and spermine caused a rapid and markeddecrease in cell number, whereas they had no cyto-toxic effect in the absence of serum. It is noteworthythat the T1/2 for the decrease in cell number in thepresence of 40 mM spermidine was 1.5 6 0.1 h for
ASMCs versus 5.5 6 0.2 h for BAECs and 6.1 6 0.2for Mel 120 cells (P , 0.01). In additional experi-
ents, the dose-dependent effect of spermidine onascular and melanoma cells was examined in theresence of 10% FCS, after 24 h treatment (Fig. 2).permidine reduced cell number in a dose-dependentanner and the EC50 was 12, 10, and 3.3 mM for Mel
120, BAECs, and RASMCs, respectively. The differ-ence between RASMCs and the two other cells wasstatistically significant (P , 0.01). Other experi-ments performed with bovine aortic smooth musclecells and with HUVECs confirmed that SMCs aremore sensitive to polyamine than endothelial cells
FIG. 1. Effect of spermidine, spermine, and putrescine on cell pcells was examined both in the presence (top) and in the absence(circle) or in medium containing 40 mM spermidine (triangle), 40 mFCS, cells exposed to putrescine proliferated normally, whereas cand after 8 h exposure approximately 90% of cells were dead. Ineffect on cell number. BAEC, RASMC, and Mel 120 cell number a
04, respectively. Results are expressed as percentage of cell numperformed in triplicate.
roliferation. The effect of polyamines on BAEC, RASMC and Mel 120(bottom) of 10% FCS. Cells were cultured either in medium alone
M spermine (X), or 40 mM putrescine (square). In the presence of 10%ells exposed to spermidine or spermine rapidly decreased in numberthe absence of serum, spermidine, spermine, and putrescine had not time zero was 5.1 6 0.15 3 104, 7.0 6 0.13 3 104, and 8.2 6 0.19 3ber at time zero and represent the mean 6 SD of three experiments
FIG. 2. Dose-dependent effect of spermidine on BAECs,RASMCs, and Mel 120 cells. The dose-dependent effect of spermidineon BAECs, RASMCs, and on Mel 120 cells was examined in thepresence of 10% FCS after 24 h exposure. The effect of spermidinewas significantly more potent on RASMC (filled bars) than on othercells (P , 0.001). BAEC, RASMC, and Mel 120 cell number at timeero was 5.4 6 0.13 3 104, 6.8 6 0.12 3 104, and 8.6 6 0.17 3 104,espectively. Results are expressed as percentage of cell number atime zero and represent the mean 6 SD of four experiments per-ormed in triplicate.
121POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
(not shown). These data indicated that spermidineand spermine, at concentrations close to physiologiclevels, markedly induced cell death in the presence ofFCS, while putrescine was not effective, and thatthis effect was significantly more rapid and potent onRASMCs than on BAECs and Mel 120 cells.
FACS Analysis and DNA Fragmentation Analysis
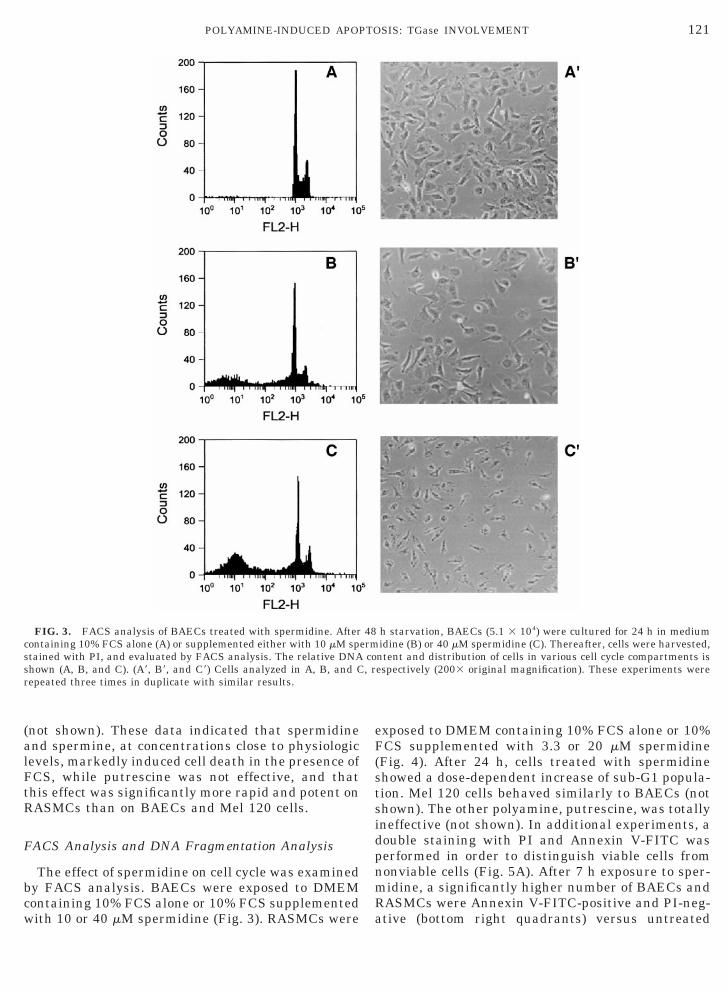
The effect of spermidine on cell cycle was examinedby FACS analysis. BAECs were exposed to DMEMcontaining 10% FCS alone or 10% FCS supplementedwith 10 or 40 mM spermidine (Fig. 3). RASMCs were
FIG. 3. FACS analysis of BAECs treated with spermidine. Aftercontaining 10% FCS alone (A) or supplemented either with 10 mM spstained with PI, and evaluated by FACS analysis. The relative DNAshown (A, B, and C). (A9, B9, and C9) Cells analyzed in A, B, and Crepeated three times in duplicate with similar results.
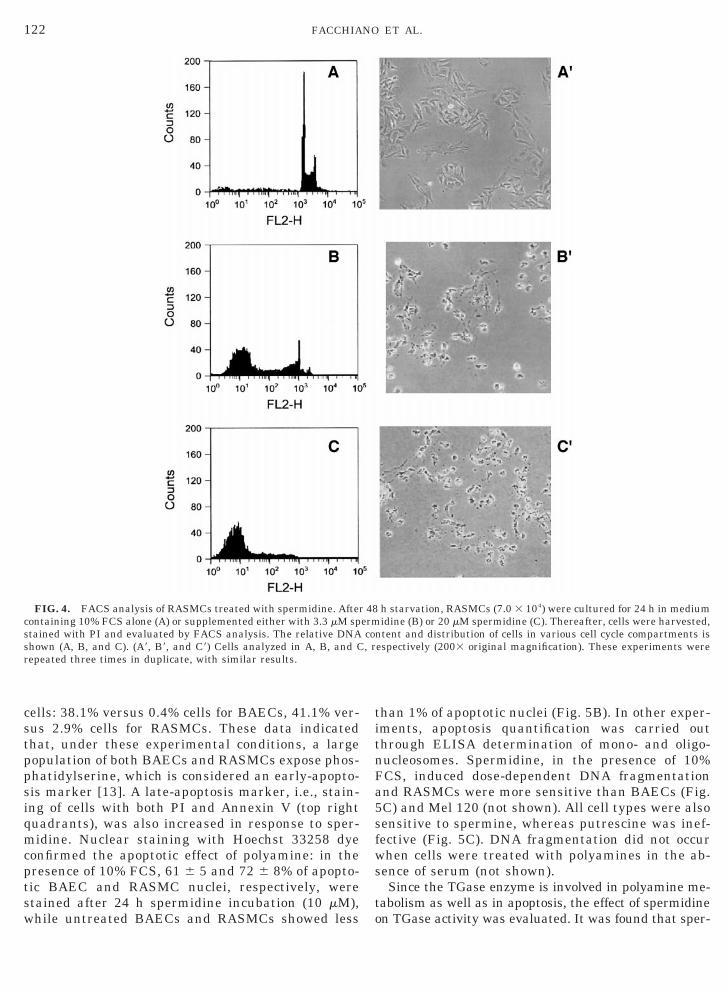
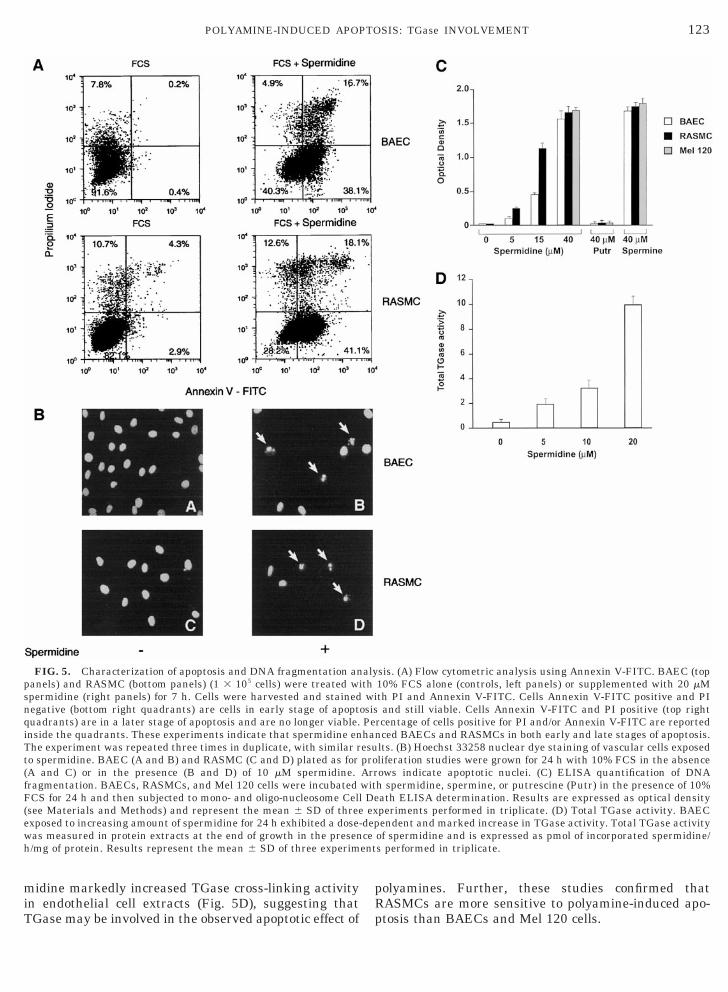
exposed to DMEM containing 10% FCS alone or 10%FCS supplemented with 3.3 or 20 mM spermidine(Fig. 4). After 24 h, cells treated with spermidineshowed a dose-dependent increase of sub-G1 popula-tion. Mel 120 cells behaved similarly to BAECs (notshown). The other polyamine, putrescine, was totallyineffective (not shown). In additional experiments, adouble staining with PI and Annexin V-FITC wasperformed in order to distinguish viable cells fromnonviable cells (Fig. 5A). After 7 h exposure to sper-midine, a significantly higher number of BAECs andRASMCs were Annexin V-FITC-positive and PI-neg-ative (bottom right quadrants) versus untreated
h starvation, BAECs (5.1 3 104) were cultured for 24 h in mediumidine (B) or 40 mM spermidine (C). Thereafter, cells were harvested,ntent and distribution of cells in various cell cycle compartments isespectively (2003 original magnification). These experiments were
48erm
co, r
122 FACCHIANO ET AL.
cells: 38.1% versus 0.4% cells for BAECs, 41.1% ver-sus 2.9% cells for RASMCs. These data indicatedthat, under these experimental conditions, a largepopulation of both BAECs and RASMCs expose phos-phatidylserine, which is considered an early-apopto-sis marker [13]. A late-apoptosis marker, i.e., stain-ing of cells with both PI and Annexin V (top rightquadrants), was also increased in response to sper-midine. Nuclear staining with Hoechst 33258 dyeconfirmed the apoptotic effect of polyamine: in thepresence of 10% FCS, 61 6 5 and 72 6 8% of apopto-tic BAEC and RASMC nuclei, respectively, werestained after 24 h spermidine incubation (10 mM),while untreated BAECs and RASMCs showed less
FIG. 4. FACS analysis of RASMCs treated with spermidine. Aftecontaining 10% FCS alone (A) or supplemented either with 3.3 mM spstained with PI and evaluated by FACS analysis. The relative DNAshown (A, B, and C). (A9, B9, and C9) Cells analyzed in A, B, and Crepeated three times in duplicate, with similar results.
than 1% of apoptotic nuclei (Fig. 5B). In other exper-iments, apoptosis quantification was carried outthrough ELISA determination of mono- and oligo-nucleosomes. Spermidine, in the presence of 10%FCS, induced dose-dependent DNA fragmentationand RASMCs were more sensitive than BAECs (Fig.5C) and Mel 120 (not shown). All cell types were alsosensitive to spermine, whereas putrescine was inef-fective (Fig. 5C). DNA fragmentation did not occurwhen cells were treated with polyamines in the ab-sence of serum (not shown).
Since the TGase enzyme is involved in polyamine me-tabolism as well as in apoptosis, the effect of spermidineon TGase activity was evaluated. It was found that sper-
h starvation, RASMCs (7.0 3 104) were cultured for 24 h in mediumidine (B) or 20 mM spermidine (C). Thereafter, cells were harvested,
ntent and distribution of cells in various cell cycle compartments isespectively (2003 original magnification). These experiments were
r 48ermco, r
snqiTt(fF(
123POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
midine markedly increased TGase cross-linking activityin endothelial cell extracts (Fig. 5D), suggesting thatTGase may be involved in the observed apoptotic effect of
FIG. 5. Characterization of apoptosis and DNA fragmentation anpanels) and RASMC (bottom panels) (1 3 105 cells) were treated wpermidine (right panels) for 7 h. Cells were harvested and stainedegative (bottom right quadrants) are cells in early stage of apoptuadrants) are in a later stage of apoptosis and are no longer viable.nside the quadrants. These experiments indicate that spermidine enhe experiment was repeated three times in duplicate, with similar ro spermidine. BAEC (A and B) and RASMC (C and D) plated as forA and C) or in the presence (B and D) of 10 mM spermidine.ragmentation. BAECs, RASMCs, and Mel 120 cells were incubatedCS for 24 h and then subjected to mono- and oligo-nucleosome Cell
see Materials and Methods) and represent the mean 6 SD of threeexposed to increasing amount of spermidine for 24 h exhibited a dose-was measured in protein extracts at the end of growth in the presenh/mg of protein. Results represent the mean 6 SD of three experim
polyamines. Further, these studies confirmed thatRASMCs are more sensitive to polyamine-induced apo-ptosis than BAECs and Mel 120 cells.
sis. (A) Flow cytometric analysis using Annexin V-FITC. BAEC (top10% FCS alone (controls, left panels) or supplemented with 20 mMth PI and Annexin V-FITC. Cells Annexin V-FITC positive and PIand still viable. Cells Annexin V-FITC and PI positive (top right
rcentage of cells positive for PI and/or Annexin V-FITC are reportednced BAECs and RASMCs in both early and late stages of apoptosis.lts. (B) Hoechst 33258 nuclear dye staining of vascular cells exposedliferation studies were grown for 24 h with 10% FCS in the absenceows indicate apoptotic nuclei. (C) ELISA quantification of DNAh spermidine, spermine, or putrescine (Putr) in the presence of 10%ath ELISA determination. Results are expressed as optical densityperiments performed in triplicate. (D) Total TGase activity. BAECendent and marked increase in TGase activity. Total TGase activity
of spermidine and is expressed as pmol of incorporated spermidine/s performed in triplicate.
alyith
wiosisPehaesuproArrwitDeex
depceent
rfhshoeostnaca3laAiitmi1
oswaearpFc1C(m
tm
124 FACCHIANO ET AL.
Investigating Biochemical Pathways Involved in theApoptotic Effect of Polyamine on Vascular Cells
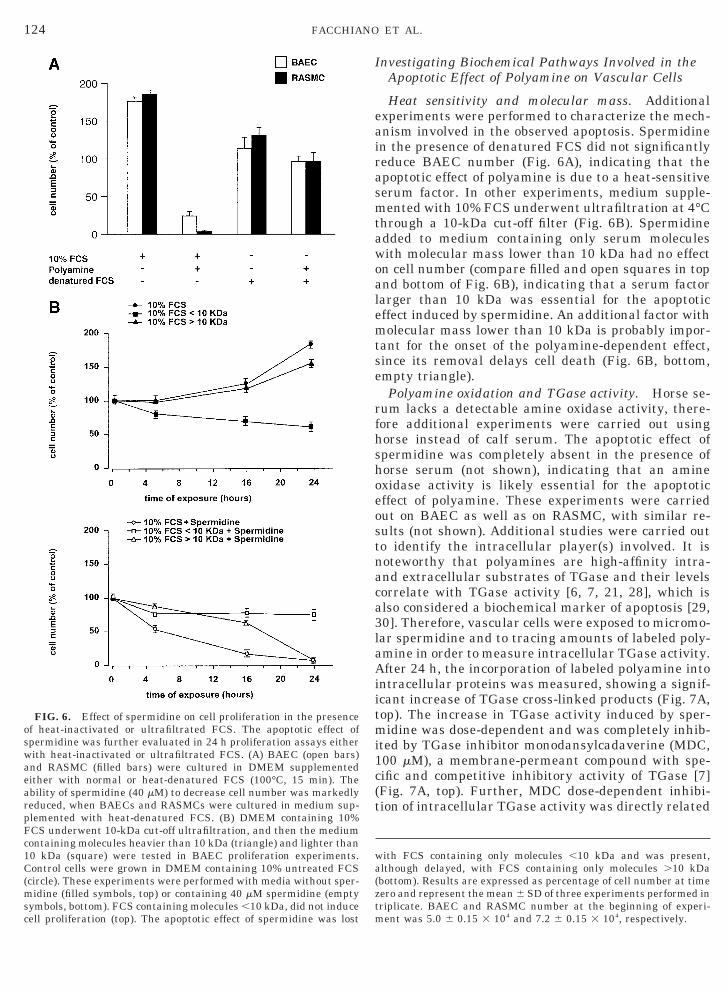
Heat sensitivity and molecular mass. Additionalexperiments were performed to characterize the mech-anism involved in the observed apoptosis. Spermidinein the presence of denatured FCS did not significantlyreduce BAEC number (Fig. 6A), indicating that theapoptotic effect of polyamine is due to a heat-sensitiveserum factor. In other experiments, medium supple-mented with 10% FCS underwent ultrafiltration at 4°Cthrough a 10-kDa cut-off filter (Fig. 6B). Spermidineadded to medium containing only serum moleculeswith molecular mass lower than 10 kDa had no effecton cell number (compare filled and open squares in topand bottom of Fig. 6B), indicating that a serum factorlarger than 10 kDa was essential for the apoptoticeffect induced by spermidine. An additional factor withmolecular mass lower than 10 kDa is probably impor-tant for the onset of the polyamine-dependent effect,since its removal delays cell death (Fig. 6B, bottom,empty triangle).
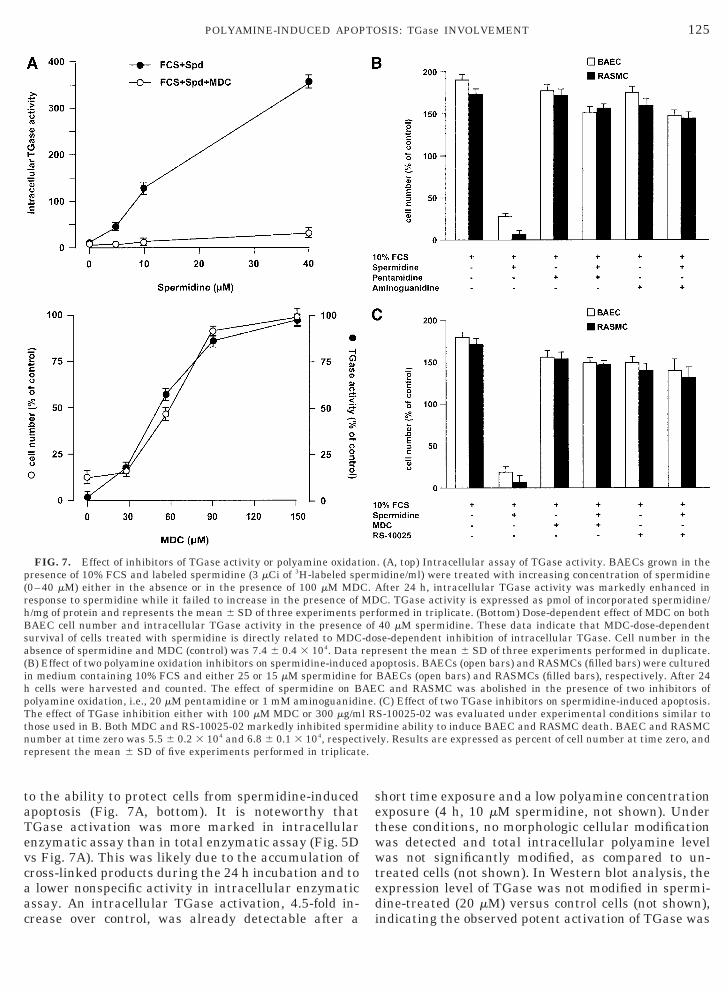
Polyamine oxidation and TGase activity. Horse se-um lacks a detectable amine oxidase activity, there-ore additional experiments were carried out usingorse instead of calf serum. The apoptotic effect ofpermidine was completely absent in the presence oforse serum (not shown), indicating that an aminexidase activity is likely essential for the apoptoticffect of polyamine. These experiments were carriedut on BAEC as well as on RASMC, with similar re-ults (not shown). Additional studies were carried outo identify the intracellular player(s) involved. It isoteworthy that polyamines are high-affinity intra-nd extracellular substrates of TGase and their levelsorrelate with TGase activity [6, 7, 21, 28], which islso considered a biochemical marker of apoptosis [29,0]. Therefore, vascular cells were exposed to micromo-ar spermidine and to tracing amounts of labeled poly-mine in order to measure intracellular TGase activity.fter 24 h, the incorporation of labeled polyamine into
ntracellular proteins was measured, showing a signif-cant increase of TGase cross-linked products (Fig. 7A,op). The increase in TGase activity induced by sper-idine was dose-dependent and was completely inhib-
ted by TGase inhibitor monodansylcadaverine (MDC,00 mM), a membrane-permeant compound with spe-
cific and competitive inhibitory activity of TGase [7](Fig. 7A, top). Further, MDC dose-dependent inhibi-tion of intracellular TGase activity was directly related
with FCS containing only molecules ,10 kDa and was present,although delayed, with FCS containing only molecules .10 kDa(bottom). Results are expressed as percentage of cell number at timezero and represent the mean 6 SD of three experiments performed inriplicate. BAEC and RASMC number at the beginning of experi-ent was 5.0 6 0.15 3 104 and 7.2 6 0.15 3 104, respectively.
FIG. 6. Effect of spermidine on cell proliferation in the presencef heat-inactivated or ultrafiltrated FCS. The apoptotic effect ofpermidine was further evaluated in 24 h proliferation assays eitherith heat-inactivated or ultrafiltrated FCS. (A) BAEC (open bars)nd RASMC (filled bars) were cultured in DMEM supplementedither with normal or heat-denatured FCS (100°C, 15 min). Thebility of spermidine (40 mM) to decrease cell number was markedlyeduced, when BAECs and RASMCs were cultured in medium sup-lemented with heat-denatured FCS. (B) DMEM containing 10%CS underwent 10-kDa cut-off ultrafiltration, and then the mediumontaining molecules heavier than 10 kDa (triangle) and lighter than0 kDa (square) were tested in BAEC proliferation experiments.ontrol cells were grown in DMEM containing 10% untreated FCS
circle). These experiments were performed with media without sper-idine (filled symbols, top) or containing 40 mM spermidine (empty
symbols, bottom). FCS containing molecules ,10 kDa, did not inducecell proliferation (top). The apoptotic effect of spermidine was lost
rhBsa
T
125POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
to the ability to protect cells from spermidine-inducedapoptosis (Fig. 7A, bottom). It is noteworthy thatTGase activation was more marked in intracellularenzymatic assay than in total enzymatic assay (Fig. 5Dvs Fig. 7A). This was likely due to the accumulation ofcross-linked products during the 24 h incubation and toa lower nonspecific activity in intracellular enzymaticassay. An intracellular TGase activation, 4.5-fold in-crease over control, was already detectable after a
FIG. 7. Effect of inhibitors of TGase activity or polyamine oxidatpresence of 10% FCS and labeled spermidine (3 mCi of 3H-labeled sp(0–40 mM) either in the absence or in the presence of 100 mM MDesponse to spermidine while it failed to increase in the presence of/mg of protein and represents the mean 6 SD of three experimentsAEC cell number and intracellular TGase activity in the presenceurvival of cells treated with spermidine is directly related to MDCbsence of spermidine and MDC (control) was 7.4 6 0.4 3 104. Data
(B) Effect of two polyamine oxidation inhibitors on spermidine-inducein medium containing 10% FCS and either 25 or 15 mM spermidineh cells were harvested and counted. The effect of spermidine on Bpolyamine oxidation, i.e., 20 mM pentamidine or 1 mM aminoguanidi
he effect of TGase inhibition either with 100 mM MDC or 300 mg/mthose used in B. Both MDC and RS-10025-02 markedly inhibited spenumber at time zero was 5.5 6 0.2 3 104 and 6.8 6 0.1 3 104, respectrepresent the mean 6 SD of five experiments performed in triplicat
short time exposure and a low polyamine concentrationexposure (4 h, 10 mM spermidine, not shown). Underthese conditions, no morphologic cellular modificationwas detected and total intracellular polyamine levelwas not significantly modified, as compared to un-treated cells (not shown). In Western blot analysis, theexpression level of TGase was not modified in spermi-dine-treated (20 mM) versus control cells (not shown),indicating the observed potent activation of TGase was
. (A, top) Intracellular assay of TGase activity. BAECs grown in theidine/ml) were treated with increasing concentration of spermidinefter 24 h, intracellular TGase activity was markedly enhanced inC. TGase activity is expressed as pmol of incorporated spermidine/
formed in triplicate. (Bottom) Dose-dependent effect of MDC on both40 mM spermidine. These data indicate that MDC-dose-dependentse-dependent inhibition of intracellular TGase. Cell number in theresent the mean 6 SD of three experiments performed in duplicate.poptosis. BAECs (open bars) and RASMCs (filled bars) were culturedBAECs (open bars) and RASMCs (filled bars), respectively. After 24C and RASMC was abolished in the presence of two inhibitors of(C) Effect of two TGase inhibitors on spermidine-induced apoptosis.
S-10025-02 was evaluated under experimental conditions similar todine ability to induce BAEC and RASMC death. BAEC and RASMCly. Results are expressed as percent of cell number at time zero, and
ionermC. AMDper
of-dorepd aforAEne.l Rrmiivee.
cs
et
ishdt
126 FACCHIANO ET AL.
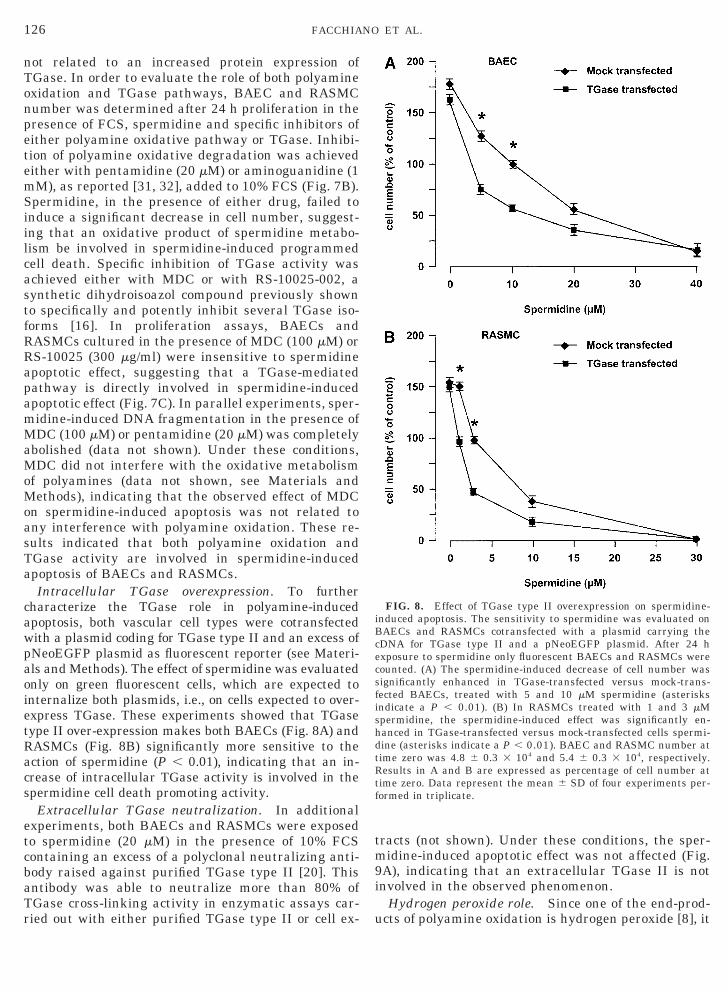
not related to an increased protein expression ofTGase. In order to evaluate the role of both polyamineoxidation and TGase pathways, BAEC and RASMCnumber was determined after 24 h proliferation in thepresence of FCS, spermidine and specific inhibitors ofeither polyamine oxidative pathway or TGase. Inhibi-tion of polyamine oxidative degradation was achievedeither with pentamidine (20 mM) or aminoguanidine (1mM), as reported [31, 32], added to 10% FCS (Fig. 7B).Spermidine, in the presence of either drug, failed toinduce a significant decrease in cell number, suggest-ing that an oxidative product of spermidine metabo-lism be involved in spermidine-induced programmedcell death. Specific inhibition of TGase activity wasachieved either with MDC or with RS-10025-002, asynthetic dihydroisoazol compound previously shownto specifically and potently inhibit several TGase iso-forms [16]. In proliferation assays, BAECs andRASMCs cultured in the presence of MDC (100 mM) orRS-10025 (300 mg/ml) were insensitive to spermidineapoptotic effect, suggesting that a TGase-mediatedpathway is directly involved in spermidine-inducedapoptotic effect (Fig. 7C). In parallel experiments, sper-midine-induced DNA fragmentation in the presence ofMDC (100 mM) or pentamidine (20 mM) was completelyabolished (data not shown). Under these conditions,MDC did not interfere with the oxidative metabolismof polyamines (data not shown, see Materials andMethods), indicating that the observed effect of MDCon spermidine-induced apoptosis was not related toany interference with polyamine oxidation. These re-sults indicated that both polyamine oxidation andTGase activity are involved in spermidine-inducedapoptosis of BAECs and RASMCs.
Intracellular TGase overexpression. To furthercharacterize the TGase role in polyamine-inducedapoptosis, both vascular cell types were cotransfectedwith a plasmid coding for TGase type II and an excess ofpNeoEGFP plasmid as fluorescent reporter (see Materi-als and Methods). The effect of spermidine was evaluatedonly on green fluorescent cells, which are expected tointernalize both plasmids, i.e., on cells expected to over-express TGase. These experiments showed that TGasetype II over-expression makes both BAECs (Fig. 8A) andRASMCs (Fig. 8B) significantly more sensitive to theaction of spermidine (P , 0.01), indicating that an in-rease of intracellular TGase activity is involved in thepermidine cell death promoting activity.
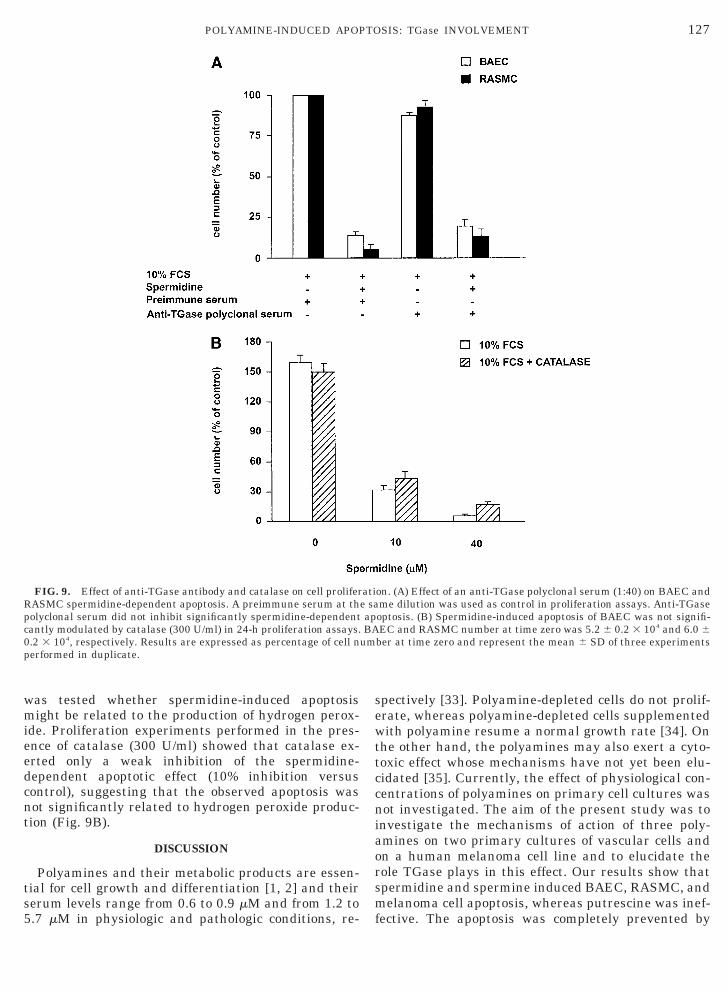
Extracellular TGase neutralization. In additionalxperiments, both BAECs and RASMCs were exposedo spermidine (20 mM) in the presence of 10% FCS
containing an excess of a polyclonal neutralizing anti-body raised against purified TGase type II [20]. Thisantibody was able to neutralize more than 80% ofTGase cross-linking activity in enzymatic assays car-ried out with either purified TGase type II or cell ex-
tracts (not shown). Under these conditions, the sper-midine-induced apoptotic effect was not affected (Fig.9A), indicating that an extracellular TGase II is notinvolved in the observed phenomenon.
Hydrogen peroxide role. Since one of the end-prod-ucts of polyamine oxidation is hydrogen peroxide [8], it
FIG. 8. Effect of TGase type II overexpression on spermidine-induced apoptosis. The sensitivity to spermidine was evaluated onBAECs and RASMCs cotransfected with a plasmid carrying thecDNA for TGase type II and a pNeoEGFP plasmid. After 24 hexposure to spermidine only fluorescent BAECs and RASMCs werecounted. (A) The spermidine-induced decrease of cell number wassignificantly enhanced in TGase-transfected versus mock-trans-fected BAECs, treated with 5 and 10 mM spermidine (asterisksndicate a P , 0.01). (B) In RASMCs treated with 1 and 3 mMpermidine, the spermidine-induced effect was significantly en-anced in TGase-transfected versus mock-transfected cells spermi-ine (asterisks indicate a P , 0.01). BAEC and RASMC number atime zero was 4.8 6 0.3 3 104 and 5.4 6 0.3 3 104, respectively.
Results in A and B are expressed as percentage of cell number attime zero. Data represent the mean 6 SD of four experiments per-formed in triplicate.
127POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
was tested whether spermidine-induced apoptosismight be related to the production of hydrogen perox-ide. Proliferation experiments performed in the pres-ence of catalase (300 U/ml) showed that catalase ex-erted only a weak inhibition of the spermidine-dependent apoptotic effect (10% inhibition versuscontrol), suggesting that the observed apoptosis wasnot significantly related to hydrogen peroxide produc-tion (Fig. 9B).
DISCUSSION
Polyamines and their metabolic products are essen-tial for cell growth and differentiation [1, 2] and theirserum levels range from 0.6 to 0.9 mM and from 1.2 to5.7 mM in physiologic and pathologic conditions, re-
FIG. 9. Effect of anti-TGase antibody and catalase on cell proliferRASMC spermidine-dependent apoptosis. A preimmune serum at thpolyclonal serum did not inhibit significantly spermidine-dependentcantly modulated by catalase (300 U/ml) in 24-h proliferation assays.0.2 3 104, respectively. Results are expressed as percentage of cell nperformed in duplicate.
spectively [33]. Polyamine-depleted cells do not prolif-erate, whereas polyamine-depleted cells supplementedwith polyamine resume a normal growth rate [34]. Onthe other hand, the polyamines may also exert a cyto-toxic effect whose mechanisms have not yet been elu-cidated [35]. Currently, the effect of physiological con-centrations of polyamines on primary cell cultures wasnot investigated. The aim of the present study was toinvestigate the mechanisms of action of three poly-amines on two primary cultures of vascular cells andon a human melanoma cell line and to elucidate therole TGase plays in this effect. Our results show thatspermidine and spermine induced BAEC, RASMC, andmelanoma cell apoptosis, whereas putrescine was inef-fective. The apoptosis was completely prevented by
on. (A) Effect of an anti-TGase polyclonal serum (1:40) on BAEC andme dilution was used as control in proliferation assays. Anti-TGaseoptosis. (B) Spermidine-induced apoptosis of BAEC was not signifi-EC and RASMC number at time zero was 5.2 6 0.2 3 104 and 6.0 6
ber at time zero and represent the mean 6 SD of three experiments
atie saapBA
um

128 FACCHIANO ET AL.
inhibitors of polyamine oxidation. The role of TGasewas evaluated since polyamines are high-affinity sub-strates of this enzyme [6, 7, 21, 28] and since TGaseactivity was shown to play a still unclear role inapoptosis [29, 30, 36]. Exposure to spermidine mark-edly increased the intracellular TGase activity andMDC and RS-10025, two specific inhibitors of severalTGase isoforms [7, 16], abolished the apoptotic effect ofspermidine, and completely inhibited TGase activity,without any effect on oxidative degradation of poly-amines in vitro. TGase type II overexpression madecells significantly more sensitive to polyamine. TheTGase involved in this apoptosis is likely intracellular,since an antibody neutralizing extracellular TGase didnot affect polyamine-induced apoptosis. It is intriguingthat inhibition of both polyamine oxidation and TGaseactivity almost completely abolished polyamine-in-duced apoptosis, leaving open the question of how bothpathways are functionally related to the described ap-optotic effect. One possibility is that polyamine oxida-tion and TGase-dependent pathways are sequential orcross-talking processes; therefore, this apoptosis maybe due to the activation of an intracellular TGase ac-tivity likely induced by a product of an extracellularpolyamine oxidation pathway. This proposed mecha-nism may not apply to apoptotic stimuli other thanpolyamines (e.g., serum deprivation) and further inves-tigation is required to identify the TGase isoform(s)actually involved in apoptotic processes, as recentlysuggested [36]. Our results do not exclude that otherTGase isoforms, different from type II, may be involvedin polyamine-induced apoptosis, since the inhibitorsused in this study are also active on other TGase iso-forms. It should be also taken into account that TGasecan modulate cell functions by acting either as a cross-linking enzyme or as a G-protein (Gah) [37] and thatintracellular TGase is silent under normal conditions[28]. Therefore, TGase activated by polyamines mightmodulate or trigger a signal transduction pathwaydownstream from such G-protein action. The results ofthe present study may have high patho-physiologicalrelevance. In fact, the physiological serum level of poly-amines increases in pathological conditions like cancer[33], diabetes mellitus [38], uremic disease [39], andcerebral ischemia [40]. Such polyamine levels are inthe low micromolar range, i.e., within the range of theEC50 effective in our experiments, suggesting the exis-tence of a homeostatic balance between mitogenic andapoptotic properties of polyamines. One of the mostintriguing findings of the present study is that SMCsare significantly more sensitive to polyamines thanendothelial cells. In fact, EC50 of spermidine onRASMCs was about threefold lower than on BAECs, aswell as the T1/2 on RASMCs was at least threefoldshorter than on BAECs. Since endothelial cells aredirectly exposed to the blood flow, the different sensi-tivity to polyamine suggests that the endothelium may
represent a physiological protective barrier againsttoxic effects of serum polyamines. The interruption ofthis barrier, which occurs for instance in atheroscle-rotic lesions and angioplasty, may expose SMCs toserum polyamines and may be responsible for SMCdysfunction described under such conditions. Further,the impaired angiogenesis observed in diabetes may bedirectly related to the increased levels of polyaminesobserved in diabetic patients [38]. Thus, the reporteddata indicate that polyamines and intracellular TGaseactivation may represent a novel regulatory pathwayof cell viability and apoptosis possibly involved in hu-man diseases characterized by elevated serum levels ofpolyamines.
The authors thank Dr. Antonio Facchiano for critical reading ofthe manuscript and Mrs. Gabriella Ricci for secretarial assistance.The financial support from Federazione Italiana Ricerca sul Cancro(FIRC) and from Telethon-Italy (Grant 1167) is gratefully acknowl-edged.
REFERENCES
1. Tabor, C. W., and Tabor, H. (1976). 1,4-Diaminobutane (pu-trescine), spermidine, and spermine. Annu. Rev. Biochem. 45,285–306.
2. Pegg, A. E. (1988). Polyamine metabolism and its importance inneoplastic growth and a target for chemotherapy. Cancer Res.48, 759–774.
3. Williams, K. (1997). Interactions of polyamines with ion chan-nels. Biochem. J. 325, 289–297.
4. Marton, L. J., and Pegg, A. E. (1995). Polyamines as targets fortherapeutic intervention. Annu. Rev. Pharmacol. 35, 55–91.
5. Casero, R. A., and Pegg, A. E. (1993). Spermidine/spermineN1-acetyltransferase—The turning point in polyamine metab-olism. FASEB J. 7, 653–661.
6. Folk, J. E. (1980). Transglutaminases. Annu. Rev. Biochem. 49,517–531.
7. Lorand, L., and Conrad, S. M. (1984). Transglutaminases. Mol.Cell Biochem. 58, 9–35.
8. Seiler, N. (1995). Polyamine oxidase, properties and functions.Prog. Brain Res. 106, 333–344.
9. Wang, J. Y., and Johnson, L. R. (1992). Role of transglutami-nase and protein cross-linking in the repair of mucosal stresserosions. Am. J. Physiol. 262, G818–G825.
10. Takigawa, M., Enomoto, M., Nishida, Y., Pan, H. O., Kinoshita,A., and Suzuki, F. (1990). Tumor angiogenesis and polyamines:Alpha-difluoromethylornithine, an irreversible inhibitor of or-nithine decarboxylase, inhibits B16 melanoma-induced angio-genesis in ovo and the proliferation of vascular endothelial cellsin vitro. Cancer Res. 50, 4131–4138.
11. Auvinen, M. (1997). Cell transformation, invasion, and angio-genesis: A regulatory role for ornithine decarboxylase and poly-amines? J. Natl. Cancer Inst. 89, 533–537.
12. Brune, B., Hartzell, P., Nicotera, P., and Orrenius, S. (1991).Spermine prevents endonuclease activation and apoptosis inthymocytes. Exp. Cell Res. 195, 323–329.
13. Bonneau, M. J., and Poulin, R. (2000). Spermine oxidation leadsto necrosis with plasma membrane phosphatidylserine redistri-bution in mouse leukemia cells. Exp. Cell. Res. 259, 23–34.
14. D’Arcangelo, D., Facchiano, F., Barlucchi, L. M., Melillo, G., Illi,B., Testolin, L., Gaetano, C., and Capogrossi, M. C. (2000).

Acidosis inhibits endothelial cell apoptosis and function and
1
1
1
1
1
2
2
2
27. Wigler, M., Sweet, R., Sim, G. K., Wold, B., Pellicer, A., Lacy, E.,
2
3
3
129POLYAMINE-INDUCED APOPTOSIS: TGase INVOLVEMENT
induces basic fibroblast growth factor and vascular endothelialgrowth factor expression. Circ. Res. 86, 312–318.
5. Facchiano, A., De Marchis, F., Turchetti, E., Facchiano, F.,Guglielmi, M., Denaro, A., Palumbo, R., Scoccianti, M., andCapogrossi, M. C. (2000). The chemotactic and mitogenic effectsof platelet-derived growth factor-BB on rat aorta smooth mus-cle cells are inhibited by basic fibroblast growth factor. J. CellSci. 113, 2855–2863.
6. Auger, M., McDermott, A. E., Robinson, V., Castelhano, A. L.,Billedeau, R. J., Pliura, D. H., Krantz, A., and Griffin, R. G.(1993). Solid-state 13C NMR study of a transglutaminase-in-hibitor adduct. Biochemistry 32, 3930–3934.
7. Beninati, S., Mantile, G., Desantis, A., and Abbruzzese, A.(1992). Covalent polyamine protein conjugates: Analysis anddistribution. Life Chem. Rep. 10, 29–33.
8. Gentile, V., Porta, R., Chiosi, E., Spina, A., Valente, F., Pezone,R., Davies, P. J., Alaadik, A., and Illiano, G. (1997). tTGase/Galpha h protein expression inhibits adenylate cyclase activity inBalb-C 3T3 fibroblasts membranes. Biochim. Biophys. Acta1357, 115–122.
9. Facchiano, F., and Luini, A. (1992). Tetanus toxin potentlystimulates tissue transglutaminase. A possible mechanism ofneurotoxicity. J. Biol. Chem. 267, 13,267–13,271.
0. Facchiano, F., Benfenati, F., Valtorta, F., and Luini, A. (1993).Covalent modification of synapsin I by a tetanus toxin-activatedtransglutaminase. J. Biol. Chem. 268, 4588–4591.
1. Beninati, S., Piacentini, M., Cocuzzi, E. T., Autuori, F., andFolk, J. (1988). Covalent incorporation of polyamines as gam-ma-glutamyl derivatives into CHO cell protein. Biochim. Bio-phys. Acta 952, 325–333.
2. Telford, W. G., King, L. E., and Fraker, P. J. (1992). Compara-tive evaluation of several DNA binding dyes in the detection ofapoptosis-associated chromatin degradation by flow cytometry.Cytometry 13, 137–143.
23. Bovolenta, C., Testolin, L., Benussi, L., Lievens, P. M., andLiboi, E. (1998). Positive selection of apoptosis-resistant cellscorrelates with activation of dominant-negative STAT5. J. Biol.Chem. 273, 20,779–20,784.
24. Jordan, M., Schallhorn, A., and Wurm, F. M. (1996). Transfect-ing mammalian cells: Optimization of critical parameters af-fecting calcium-phosphate precipitate formation. Nucleic AcidsRes. 24, 596–601.
25. Gentile, V., Thomazy, V., Piacentini, M., Fesus, L., and Davies,P. J. (1992). Expression of tissue transglutaminase in Balb-C3T3 fibroblasts: Effects on cellular morphology and adhesion.J. Cell Biol. 119, 463–474.
26. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). “Molecu-lar Cloning: A Laboratory Manual,” 2nd ed., Cold Spring Har-bor Laboratory, Cold Spring Harbor, NY.
Received January 15, 2001
Maniatis, T., Silverstein, S., and Axel, R. (1979). Transforma-tion of mammalian cells with genes from procaryotes and eu-caryotes. Cell. 16, 777–785.
28. Greenberg, C. S., Birckbichler, P. J., and Rice, R. H. (1991).Transglutaminases: Multifunctional cross-linking enzymesthat stabilize tissues. FASEB J. 5, 3071–3077.
9. Fesus, L. (1993). Biochemical events in naturally occurringforms of cell death. FEBS Lett. 328, 1–5.
0. Melino, G., and Piacentini, M. (1998). ‘Tissue’ transglutaminasein cell death: a downstream or a multifunctional upstreameffector? FEBS Lett. 430, 59–63.
1. Libby, P. R., and Porter, C. W. (1992). Inhibition of enzymes ofpolyamine back-conversion by pentamidine and berenil. Bio-chem. Pharmacol. 44, 830–832.
32. Ivanova, S., Botchkina, G. I., Al-Abed, Y., Meistrell, M., Batli-walla, F., Dubinsk, J. M., Iadecola, C., Wang, H., Gregersen,P. K., Eaton, J. W., and Tracey, K. J. (1998). Cerebral ischemiaenhances polyamine oxidation: identification of enzymaticallyformed 3-aminopropanal as an endogenous mediator of neuro-nal and glial cell death. J. Exp. Med. 188, 327–340.
33. Hospattankar, A. V., Advani, S. H., Vaidya, N. R., Electric-walla, S. E., and Braganca, B. M. (1980). Elevation of serumpolyamines in malignant lymphomas and acute myeloid leuke-mia. Int. J. Cancer 25, 463–466.
34. Pegg, A. E. (1984). The role of polyamine depletion and accu-mulation of decarboxylated S-adenosylmethionine in the inhi-bition of growth of SV-3T3 cells treated with alpha-difluoro-methylornithine. Biochem. J. 224, 29–38.
35. Nairn, L. M., Lindsay, G. S., Woster, P. M., and Wallace, H. M.(2000). Cytotoxicity of novel unsymmetrically substituted in-hibitors of polyamine biosynthesis in human cancer cells.J. Cell Physiol. 182, 209–213.
36. De Laurenzi, V., and Melino, G. (2001). Gene disruption oftissue transglutaminase. Mol. Cell Biol. 21, 148–155.
37. Nakaoka, H., Perez, D. M., Baek, K. J., Das, T., Husain, A.,Misono, K., Im, M. J., and Graham, R. M. (1994). Gh: A GTP-binding protein with transglutaminase activity and receptorsignaling function. Science 264, 1593–1596.
38. Seghieri, G., Anichini, R., Ciuti, M., Gironi, A., Bernardini, F.,and Franconi, F. (1997). Raised erythrocyte polyamine levels innon-insulin-dependent diabetes mellitus with great vessel dis-ease and albuminuria. Diabetes Res. Clin. Pract. 37, 15–20.
39. Saito, A., Takagi, T., Chung, T. G., and Ohta, K. (1983). Serumlevels of polyamines in patients with chronic renal failure.Kidney Int. Suppl. 16, S234–S237.
40. Johnson, T. D. (1998). Polyamines and cerebral ischemia. Prog.Drug Res. 50, 193–258.
Revised version received August 6, 2001Published online October 11, 2001
Related Documents











