Physiologia Plantarum 133: 140–156. 2008 Copyright ª Physiologia Plantarum 2008, ISSN 0031-9317 Transgenic tobacco plants overexpressing polyamine oxidase are not able to cope with oxidative burst generated by abiotic factors Panagiotis N. Moschou, Ioannis D. Delis, Konstantinos A. Paschalidis and Kalliopi A. Roubelakis-Angelakis* Department of Biology, University of Crete, PO Box 2280, 71409 Heraklion Crete, Greece Correspondence *Corresponding author, e-mail: [email protected] Received 17 September 2007; revised 29 November 2007 doi: 10.1111/j.1399-3054.2008.01049.x The molecular and biochemical mechanism(s) of polyamine (PA) action remain largely unknown. Transgenic tobacco plants overexpressing polyamine oxidase (PAO) from Zea mays exhibited dramatically increased expression levels of Mpao and high 1,3-diaminopropane (Dap) content. All fractions of spermidine and spermine decreased significantly in the transgenic lines. Although Dap was concomitantly generated with H 2 O 2 by PAO, the latter was below the detection limits. To show the mode(s) of H 2 O 2 scavenging, the antioxidant machinery of the transgenics was examined. Specific isoforms of peroxidase, superoxide dismutase and catalase were induced in the trans- genics but not in the wild-type (WT), along with increase in activities of additional enzymes contributing to redox homeostasis. One would expect that because the antioxidant machinery was activated, the transgenics would be able to cope with increased H 2 O 2 generated by abiotic stimuli. However, despite the enhanced antioxidant machinery, further increase in the intracellular reactive oxygen species (ROS) by exogenous H 2 O 2 , or addition of methylviologen or menadione to transgenic leaf discs, resulted in oxidative stress as evidenced by the lower quantum yield of PSII, the higher ion leakage, lipid peroxidation and induction of programmed cell death (PCD). These detrimental effects of oxidative burst were as a result of the inability of transgenic cells to further respond as did the WT in which induction of antioxidant enzymes was evident soon following the treatments. Thus, although the higher levels of H 2 O 2 generated by overexpression of Mpao in the transgenics, with altered PA homeostasis, were successfully controlled by the concomitant activation of the antioxidant machinery, further increase in ROS was detrimental to cellular functions and induced the PCD syndrome. Abbreviations – ADC, Arg decarboxylase; APX, ascorbate peroxidase; ASA, ascorbate; CAT, catalase; DAB, diamino benzidine; DAO, diamine oxidase; Dap, 1,3-diaminopropane; DCFH-DA, 2#,7#-dichlorodihydrofluorescein diacetate; DHA, dehydroascorbate; DHAR, dehydroascorbate reductase; ICF, intercellular washing fluid; MDHAR, monodehydroascorbate reductase; MV, methylviologen; NBT, nitroblue tetrazolium; ODC, Orn decarboxylase; PA, polyamine; PAO, polyamine oxidase; PCD, programmed cell death; PCR, polymerase chain reaction; PH-PA, pellet hydrolyzed polyamine; POX, peroxidase; Put, putrescine; ROS, reactive oxygen species; S-PA, soluble polyamine; SAM, S-adenosyl-L-methionine; SAMDC, S-adenosyl-L-methionine decarboxylase; SH-PA, soluble hydrolyzed polyamine; SOD, superoxide dismutase; Spd, spermidine; SPDS, Spd synthase; Spm, spermine; WT, wild-type. 140 Physiol. Plant. 133, 2008

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiologia Plantarum 133: 140–156. 2008 Copyright ª Physiologia Plantarum 2008, ISSN 0031-9317
Transgenic tobacco plants overexpressing polyamineoxidase are not able to cope with oxidative burstgenerated by abiotic factorsPanagiotis N. Moschou, Ioannis D. Delis, Konstantinos A. Paschalidis andKalliopi A. Roubelakis-Angelakis*
Department of Biology, University of Crete, PO Box 2280, 71409 Heraklion Crete, Greece
Correspondence
*Corresponding author,
e-mail: [email protected]
Received 17 September 2007; revised
29 November 2007
doi: 10.1111/j.1399-3054.2008.01049.x
The molecular and biochemical mechanism(s) of polyamine (PA) action
remain largely unknown. Transgenic tobacco plants overexpressing polyamine
oxidase (PAO) from Zea mays exhibited dramatically increased expressionlevels of Mpao and high 1,3-diaminopropane (Dap) content. All fractions of
spermidine and spermine decreased significantly in the transgenic lines.
Although Dap was concomitantly generated with H2O2 by PAO, the latter was
below the detection limits. To show the mode(s) of H2O2 scavenging, the
antioxidant machinery of the transgenics was examined. Specific isoforms of
peroxidase, superoxide dismutase and catalase were induced in the trans-
genics but not in the wild-type (WT), along with increase in activities of
additional enzymes contributing to redox homeostasis. Onewould expect thatbecause the antioxidant machinery was activated, the transgenics would be
able to cope with increased H2O2 generated by abiotic stimuli. However,
despite the enhanced antioxidant machinery, further increase in the
intracellular reactive oxygen species (ROS) by exogenous H2O2, or addition
of methylviologen or menadione to transgenic leaf discs, resulted in oxidative
stress as evidenced by the lower quantum yield of PSII, the higher ion leakage,
lipid peroxidation and induction of programmed cell death (PCD). These
detrimental effects of oxidative burst were as a result of the inability oftransgenic cells to further respond as did the WT in which induction of
antioxidant enzymes was evident soon following the treatments. Thus,
although the higher levels of H2O2 generated by overexpression of Mpao in
the transgenics, with altered PA homeostasis, were successfully controlled by
the concomitant activation of the antioxidant machinery, further increase in
ROS was detrimental to cellular functions and induced the PCD syndrome.
Abbreviations – ADC, Arg decarboxylase; APX, ascorbate peroxidase; ASA, ascorbate; CAT, catalase; DAB, diamino benzidine;
DAO, diamine oxidase; Dap, 1,3-diaminopropane; DCFH-DA, 2#,7#-dichlorodihydrofluorescein diacetate; DHA, dehydroascorbate;
DHAR, dehydroascorbate reductase; ICF, intercellular washing fluid; MDHAR, monodehydroascorbate reductase; MV, methylviologen;
NBT, nitroblue tetrazolium; ODC, Orn decarboxylase; PA, polyamine; PAO, polyamine oxidase; PCD, programmed cell death; PCR,
polymerase chain reaction; PH-PA, pellet hydrolyzed polyamine; POX, peroxidase; Put, putrescine; ROS, reactive oxygen species;
S-PA, soluble polyamine; SAM, S-adenosyl-L-methionine; SAMDC, S-adenosyl-L-methionine decarboxylase; SH-PA, soluble
hydrolyzed polyamine; SOD, superoxide dismutase; Spd, spermidine; SPDS, Spd synthase; Spm, spermine; WT, wild-type.
140 Physiol. Plant. 133, 2008

Introduction
The main cellular polyamines (PAs), the diamine putres-
cine (Put) and the tri- and tetra-amines spermidine (Spd)and spermine (Spm), respectively, are small, ubiquitous
cationic molecules at cytosolic pH. During the past
decade, it has been increasingly accepted that PA
homeostasis affects growth and development and also
stress responses in eukaryotic cells (Arias et al. 2005, Fos
et al. 2003, Kusano et al. 2007 and references therein,
Kuznetsov et al. 2007, Papadakis and Roubelakis-
Angelakis 2005, Paschalidis and Roubelakis-Angelakis2005a, 2005b, Perez-Amador et al. 2002, Takahashi et al.
2004, Tiburcio et al. 1994,Vuosku et al. 2006, Yamaguchi
et al. 2006).
The pathway of PA biosynthesis is ubiquitous in living
organisms. It is a relatively short pathway in terms of
participating enzymes, most of which have been charac-
terized and the corresponding genes/cDNAs cloned from
different sources (Kaur-Sawhney et al. 2003, Kusano et al.2007). Orn decarboxylase (ODC, EC 4.1.1.19) and Arg
decarboxylase (ADC, EC 4.1.1.19) catalyze the removal
of the carboxyl group from Orn and Arg, respectively, to
yield Put. S-adenosyl-L-methionine decarboxylase
(SAMDC, EC 4.1.1.50) introduces S-adenosyl-L-methio-
nine (SAM) into the pathway, which is then used in its
decarboxylated form (dcSAM) as an aminopropyl donor
in the conversion of Put to Spd and Spm (Roje 2006). Theactual transfer of the aminopropyl moiety is catalyzed by
the enzymes Spd synthase (SPDS, EC 2.5.1.16) and Spm
synthase (EC 2.5.1.22) (Hashimoto et al. 1998, Panicot
et al. 2002). The catabolic phase involves deamination of
Put, Spd and Spm by the action of amine oxidases. The
flavoprotein polyamine oxidase (PAO, EC 1.5.3.3)
oxidizes Spd and Spm at their secondary amino groups.
PAO yields D1-pyrroline and 1,5-diazabicyclononanefrom Spd and Spm, respectively, along with 1,3-diami-
nopropane (Dap) and H2O2. In contrast, Put is N-
methylated by Put N-methyltransferase, which catalyzes
the first committed steps in the biosynthesis of nicotine
and tropane alkaloids (Sato et al. 2001).
In tobacco plants, PA titers and biosynthesis follow
a basipetal decrease along the plant axis and negatively
correlate with cell size. The shoot apical meristem is themain site of Spd and Spm biosynthesis, and the root
synthesizesmostly Put. High and low SPDS are correlated
with cell division and expansion, respectively. Put
biosynthetic pathways are differently regulated in hyper-
and hypogeous tobacco tissues: only ADC is responsible
for Put synthesis in old, hypergeous vascular tissues, and
ODC in hypogeous tissues. Furthermore, ODC expres-
sion coincides with early cell divisions in marginalsectors of the lamina, and SPDS strongly correlates with
later cell divisions in the vascular regions (Paschalidis and
Roubelakis-Angelakis 2005b). Expression of diamine
oxidase (DAO, EC 1.4.3.6) and PAO in developing
tobacco tissues precedes and overlaps with nascent
nuclear DNA and also with peroxidases (POXs) and
lignification. In mature and old tobacco tissues, expres-sion of PAO and POX coincides with G2 nuclear phase
and endoreplication. In young vs the older roots,
expression of PAO and POX decreases with parallel
inhibition of G2 advance and endoreplication. DNA
synthesis early in development and the advance in cell
cycle/endocycle are temporally and spatially related to
PA catabolism and vascular development (Paschalidis
and Roubelakis-Angelakis 2005a).PAO, in addition to reducing PAs, is also a H2O2-
producing enzyme, which has been proposed to partic-
ipate in developmental processes and responses to biotic
and abiotic stresses (Cona et al. 2005, 2006, Paschalidis
and Roubelakis-Angelakis 2005a, Paschalidis et al. 2001,
Rea et al. 2004, Yoda et al. 2003). Recently, the first
evidence of a direct DNA oxidative damage by murine
Spmoxidase activitieswas shown (Amendola et al. 2005),and Spm content was proposed to act as a DNA shielding
molecule and a scavenger against radical species (Lovaas
1997).
Plant response to oxidative stress is controlled by
a cascade of molecular networks, which may activate
stress-related mechanisms either to reestablish homeo-
stasis and to protect proteins and membranes leading to
cell survival or to induce susceptibility leading tooxidative damage, such as lipid peroxidation and ion
leakage leading to senescence and cell death (Alvarez
et al. 1998, Papadakis and Roubelakis-Angelakis 2005,
Sasaki-Sekimoto et al. 2005). Plant engineering strategies
for oxidative stress tolerance include the expression of
genes involved in signaling and regulatory pathways,
antioxidant genes (Laloi et al. 2004, Mittler 2006) or
genes encoding for enzymes mediating synthesis ofantioxidant molecules. Genetic engineering for
increased biosynthesis of specific PAs resulted, in several
cases, in plants with modified stress response. Over-
expression of adc induced a significant increment in Put
and a small increase in Spd and Spm levels, ultimately
protecting the plants from drought (Capell et al. 2004).
Overexpression of samdc from yeast in tomato resulted in
higher levels of Spm and Spd and lycopene in fruits,increasing the nutrient value of tomato juice (Mehta et al.
2002). Overexpression of spds cDNA from Cucurbita
ficifolia in Arabidopsis thaliana increased significantly
Spm levels and enhanced tolerance to various stresses
(Kasukabe et al. 2004). Rea et al. (2004) have developed
transgenics overexpressing Mpao. In these transgenics,
high expression levels of the transgene were detected in
Physiol. Plant. 133, 2008 141
the extracellular space and the amount of soluble
polyamines (S-PAs) was lower to that of the wild-type
(WT), while a small but significant change in reactive
oxygen species (ROS)-scavenging capacity was verified.
In this work, we also developed tobacco transgenics
overexpressing the maize pao gene (Fig. 1). Our Mpaooverexpressers were thoroughly analyzed for all fractions
of PAs [free, soluble conjugated and insoluble conjugated
PAs; S-PAs, soluble hydrolyzed polyamines (SH-PAs) and
pellet hydrolyzed polyamines (PH-PAs), respectively],
along with the expression of all genes involved in the
antioxidant machinery and the H2O2 fate, in an effort
to unravel their behavior under normal and oxidative
stress conditions. The transgenics contained significantly
reduced S-Spd and S-Spm, and the increased activity of
PAO resulted in the production of H2O2, which signaled
the expression of specific antioxidant isoenzymes. Thus,no intracellular H2O2 was allowed to be built up. On the
contrary, when exogenous H2O2 or elicitors of H2O2
were supplied, the transgenic cells with the impaired
PA homeostasis were not able to overcome and the
intracellular H2O2 level was beyond the required
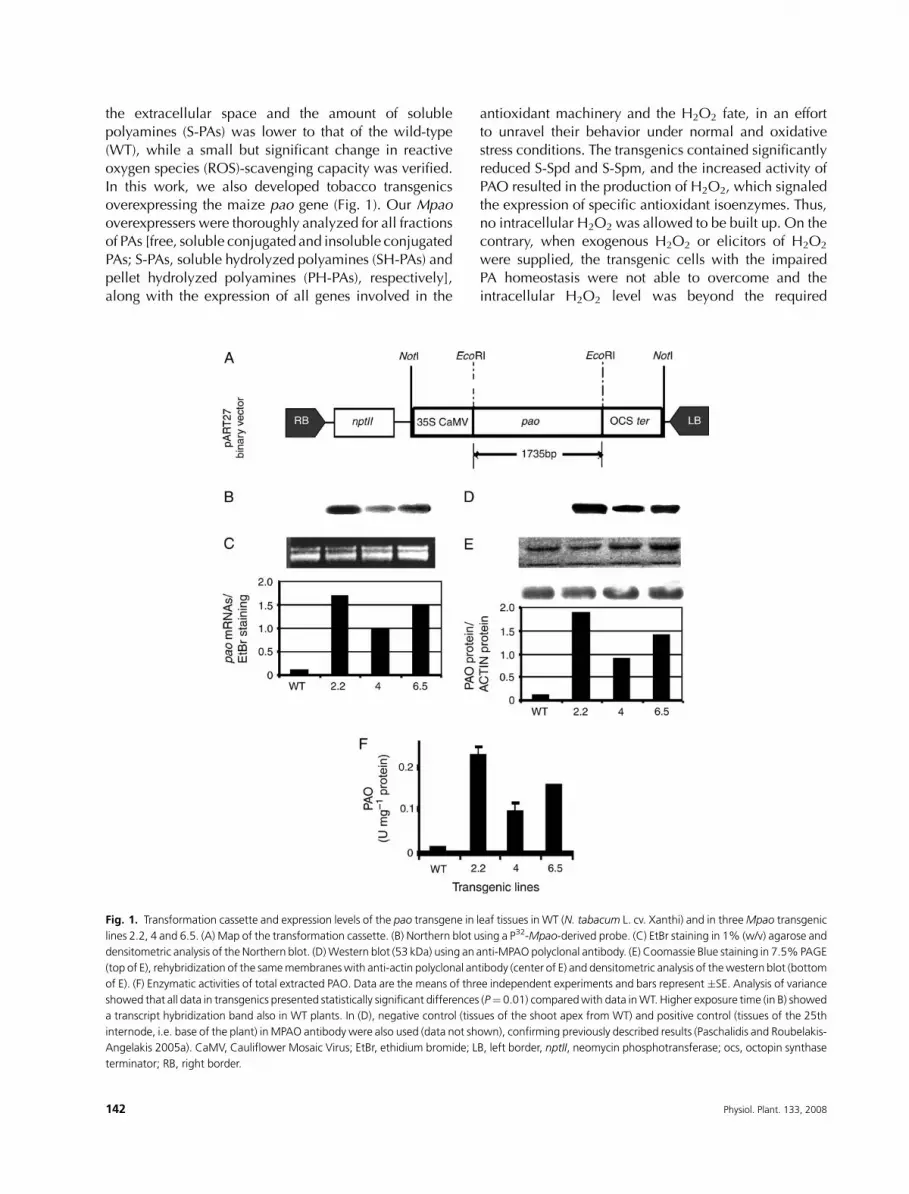
Fig. 1. Transformation cassette and expression levels of the pao transgene in leaf tissues in WT (N. tabacum L. cv. Xanthi) and in threeMpao transgenic
lines 2.2, 4 and 6.5. (A) Map of the transformation cassette. (B) Northern blot using a P32-Mpao-derived probe. (C) EtBr staining in 1% (w/v) agarose and
densitometric analysis of the Northern blot. (D)Western blot (53 kDa) using an anti-MPAO polyclonal antibody. (E) Coomassie Blue staining in 7.5%PAGE
(top of E), rehybridization of the samemembraneswith anti-actin polyclonal antibody (center of E) and densitometric analysis of thewestern blot (bottom
of E). (F) Enzymatic activities of total extracted PAO. Data are the means of three independent experiments and bars represent �SE. Analysis of variance
showed that all data in transgenics presented statistically significant differences (P¼ 0.01) comparedwith data inWT. Higher exposure time (in B) showed
a transcript hybridization band also in WT plants. In (D), negative control (tissues of the shoot apex from WT) and positive control (tissues of the 25th
internode, i.e. base of the plant) inMPAO antibody were also used (data not shown), confirming previously described results (Paschalidis and Roubelakis-
Angelakis 2005a). CaMV, Cauliflower Mosaic Virus; EtBr, ethidium bromide; LB, left border, nptII, neomycin phosphotransferase; ocs, octopin synthase
terminator; RB, right border.
142 Physiol. Plant. 133, 2008

‘signature’ to signal further expression of the antioxidant
genes. Instead, the programmed cell death (PCD) syn-
drome was induced and executed.
Materials and methods
Plant material and growth conditions
Plants of tobacco (Nicotiana tabacum cv. Xanthi) were
grown in a growth chamber with an irradiance ofapproximately 50 mmol m22 s21, temperature of 25 �2�C and 16:8 h photoperiod. Cell suspension cultures
were generated from calli derived from shoot explants.
Localization of PAO: preparation of intercellularwashing fluid and fractions of soluble, extractableand tightly wall-bound PAO
Intercellular washing fluid (ICF) fraction was obtained as
described previously by Rea et al. (2004). After two
additional washings of the apoplastic compartment to
obtain residual intracellularwashing fluids, the tissuewas
grounded and the soluble, extractable and tightly wall-
bound PAO fractions were obtained according to Cona
et al. (2005). To obtain as pure fractions as possible, in the
washing procedures, three subfractions were obtained inwhich PAO activity was also estimated, giving gradually
declining PAO activity, with the third subfraction
exhibiting insignificant levels of PAO activity. PAO
activity in the fractions of cell suspension cultures was
measured as follows. Exponentially growing cells were
harvested and cells pelleted at 800 g at room temperature.
The supernatant corresponds to the ICF fraction. The
pellet was washed with culture medium until no measur-able activity in the supernatant after the centrifugation
step could be obtained. The supernatant derived from the
last wash was concentrated 20 times before the measure-
ment of PAO activity. Approximately 10 washes were
enough to completely eliminate residual PAO activity in
the supernatant. Pellet fractions (soluble, extractable and
tightly wall bound) were obtained as described above for
the leaf.
Vector construction and plant transformation
The maize pao cDNA (Tavladoraki et al. 1998) wassubcloned in pART 7 plasmid vector using EcoRI. The
pART 7 was digested with NotI and the excised fragment
subcloned into the pART27 binary vector, which was used
to transform Agrobacterium tumefaciens strain LBA4404.
Stable transformation was performed using tobacco
leaf discs and transformants were selected against 150
mg l21 kanamycin in Murashige and Skoog medium.
Screening of transgenics
Polymerase chain reaction (PCR) was performed usingspecific primers for the nptII gene with the sequence 5#-GGT TCT CCGGCCGCT TGG-3# for the forward primer
and 5#-TCGGGAGCGGCGATACCG-3# for the reverseprimer. Southern blotting was performed using the
restriction enzyme NotI. DNAwas transferred to a mem-
brane and hybridized to a cDNA maize pao-derived32P-labeled probe.
PA analysis, protein extraction and enzyme assays
Soluble, soluble conjugated and insoluble conjugated PAfractions were determined as previously described, using
an HP 1100 High Performance Liquid Chromatographer
(Hewlett-Packard, Waldbronn, Germany) (Kotzabasis
et al. 1993), and a standard curve for Dap was also
determined.
Total proteins were extracted as already described in
Papadakis and Roubelakis-Angelakis (2005). For PAO
and DAO assays, a spectrophotometric method develo-ped by Federico et al. (1985) was used with minor modi-
fications. Briefly for PAO, tissue was ground to a fine
powder in liquid nitrogen with a mortar and pestle and
proteins were extracted after the addition of 5 volumes
g21 FW of ice-cold K-phosphate buffer pH 6.5, supple-
mented with 10 mM dithiothreitol and 10 mM pyridoxal
phosphate. Samples were left on ice for 20 min and
centrifuged at 10 000 g at 4�C for 20 min. Aliquots of thesupernatant were added in the reaction buffer containing
K-phosphate buffer pH 6.5 and 10mM Spd-3HCl (Sigma-
Aldrich, Greece) to a final volume of 500 ml and incub-
ated for 1 h. The reaction was stopped with the addition
of 63 ml 20% TCA, and subsequently, 1.25 ml 1 M
diaminobenzidine was added. Samples were incubated
for 30min at 4�C, and the absorbancewas read at 460 nm
after a brief centrifugation. One unit of enzyme repre-sents the amount of enzyme that catalyzes the oxidation
of 1 mmol of substrate per minute. A radiometric method
was also used for PAO and DAO assays according to
Paschalidis and Roubelakis-Angelakis (2005a).
Spectrophotometric assays were used for POX enzyme
activities according to Paschalidis and Roubelakis-
Angelakis (2005a). Total superoxide dismutase (SOD)
activity was quantified according to the photochemicalmethod of Beauchamp and Fridovitch (1971). Mono-
dehydroascorbate reductase (MDHAR) and dehydroas-
corbate reductase (DHAR) activities were determined
according to Serrano et al. (1994) and Asada (1984),
respectively. For the determination of ascorbate peroxi-
dase (APX), total proteins were similarly extracted by
further adding 1 mM ascorbate (ASA). APX activity was
Physiol. Plant. 133, 2008 143

determined according to Nakano and Asada (1981).
Catalase (CAT) activity was determined by measuring the
initial rate of H2O2 decomposition at 240 nm (e: 0.036mM21 cm21). One milliliter of the reaction mixture con-
sisted of 50 mM K-phosphate, pH 7.0, and 15 mMH2O2.
Native IEF, activity staining, determination of ASAand protease assay
Native IEF PAGE was used to detect the isoenzymes of
POX by activity staining of protein extracts separated in
gel over a pH gradient 3.0–10.0. Isoenzymes were
visualized by incubating native IEF gels in solution
containing 50 mM K-phosphate buffer, pH 7.4, 0.1 mgml21 4-chloro-1-napthol and 0.16% H2O2.
For the activity staining of APX, 10mMASAwas added
in the IEF electrophoresis buffer, and 10% gels were
prerun for 30 min at 20 mA (Rao et al. 1995). Sub-
sequently, the gelswere incubated in thedark in a solution
containing 50 mM K-phosphate, pH 7.0, and 2 mM ASA;
the gels were incubated in the dark for another 30 min in
50 mM K-phosphate, pH 7.0, 4 mM ASA and 2 mMH2O2.Bands were visualized after the incubation of gels in
coloring solution [50 mM K-phosphate, pH 7.8, 14 mM
tetramethylethylenediamine (TEMED) and 1.2 mM nitro-
blue tetrazolium (NBT)]. For the activity staining of SOD,
a method described in Beauchamp and Fridovitch (1971)
was used. Native gels were stained for CAT activity ac-
cording to Clare et al. (1984).
For the determination of ASA content in leaves,a previously described method of Yasui and Hayashi
(1991) was used. Tissues (1 g FW) were homogenized in
3 ml of cold 5% (w/v) metaphosphoric acid (Yamamoto
et al. 2005). After centrifugation at 12 000 g for 20 min,
the supernatant was used for the determination of ASA
and dehydroascorbate (DHA). ASA and DHA were
separated in anHPLC system (HP1100;Hewlett-Packard)
using 2 mM perchloric acid aqueous solution anddetected by absorbance at 300 nm after reaction with
100 mM NaOH containing 100 mM NaBH4.
For protease activity assay, 20 mg of total protein was
separated in 10% acrylamide gel containing 1 mg ml21
gelatin. After electrophoresis, the resolving gel was
incubated overnight at 37�C in 40 mM Tris–HCl, pH
8.6, and 0.2% Triton X100. The gel was stained in
Coomassie Blue R-250, destained and the proteaseactivity appeared as white bands.
In situ H2O2 and O�22 and epifluorescent detection
of ROS
For in situ detection of H2O2, a modified previously
described method (Yoda et al. 2003) was used. Spd (10
mM) was infiltrated in leaves with a 1-ml syringe without
a needle, and they were incubated at 25�C under con-
tinuous light for the indicated time points. At the end
of incubation time, DAB (1 mg ml21, pH 3.8) was in-
filtrated and leaves were incubated for 1 h. Deposits
formed were visualized after discoloring leaves in boiledabsolute ethanol. For the in situ detection of O�2
2 , the
same procedure as described above for leaf discs was
followed, except that 0.5 mg ml21 NBT in K-phosphate
buffer pH 7.8 was infiltrated in untreated tissues and 0.1
mg ml21 for treated ones.
In situ localization of H2O2 was performed using
the highly sensitive, cell-permeable probe 2#,7#-dichlo-rodihydrofluorescein diacetate (DCFH-DA) (MolecularProbes, Eugene, OR). Cells were harvested, after centri-
fugation at 800 g, and were incubated in 1 ml buffer [20
mM K-phosphate, pH 6.0, supplemented with 50 mM
DCFH-DA and 3mgml21 horseradish peroxidase (Sigma-
Aldrich)] for 10 min at 25�C in darkness. An aliquot of
cells (0.1 ml) was removed, washed in the same buffer
and visualized immediately. Pictures were taken with an
epifluorescence microscope (Nikon Eclipse E800 1, Tokyo,Japan) with excitation filter EX 450-490 and emission
filter BA 520 using a SONY655 SONYDXC-950P (Tokyo,
Japan) camera.
Western blotting, RNA extraction and northernblotting
Total protein extracts were electrophoretically resolved,transferred to membranes and hybridized against an anti-
PAOmaize polyclonal antibody (Angelini et al. 1995), an
anti-CAT tomato polyclonal antibody, an anti-SOD-2 and
SOD-4 polyclonal antibody and an anti-actin polyclonal
antibody (Sigma-Aldrich). Total RNA was extracted with
the optimized hot phenol method according to Iandolino
et al. (2004), transferred to amembrane and hybridized to
a pao cDNA-derived 32P-labeled probe.
Chl a fluorescence, ion leakage, lipid peroxidationand DNA fragmentation
Chl a fluorescence emission was measured with a Handy
PEA instrument (Hansatech Instruments, Norfolk, UK).
Samples were dark adapted for 15 min and illuminated
afterward with continuous light (650 nm peak wavelength1800 mmol photons m22 s21 maximum light intensity) for
5 s provided by an array of three light-emitting diodes
focusedonacircleof 5mmdiameter of the sample surface.
Ion leakage was measured as conductivity using a K610
conductivity meter (mV/cm; Consort, Turnhout, Belgium).
Malondialdehyde contentwas used as indicator of lipid
peroxidation because of increased ROS generation and
144 Physiol. Plant. 133, 2008

was determined by a color reaction with thiobarbituric
acid (Heath and Packer 1968).
DNA fragmentation assay was performed as stated
in Papadakis and Roubelakis-Angelakis (2005). For the
detection of nuclear DNA fragmentation (Tunel assay), cell
cultureswere treatedwith 10mMH2O2 (DNase Iwas usedas a positive control). Untreated cells were the controls.
After an 18-h treatment, the cells were fixed in 4% (w/v)
paraformaldehyde in PBS buffer (pH 7.4), as described
by Asai et al. (2000). Nuclei were stained with 4#,6-diamidino-2-phenylindole (1mgml21) and the free 3#-OH
groups of fragmented DNAmolecules were labeled by the
Tunel (TdT-mediated dUTP nick-end labeling) method,
using the InSituCellDeathDetectionKit (Promega,Madison,WI), as instructed by the manufacturer. DNA was
visualized using a NIKON 800 fluorescence microscope.
Results
Production and screening of Mpao tobaccotransgenic lines: overexpression of Mpao generesulted in increased PAO protein and specificactivity
N. tabacum L. cv. Xanthi plants were transformed via
Agrobacterium-mediated transformation, using the
pART7/pART27 system, carrying the full-length maize
pao cDNAclone, downstreamof the constitutive promoter
35S originated by the Cauliflower Mosaic Virus (Fig. 1A).
Several transgenic lines were obtained by selecting trans-formants on kanamycin. Furthermore, PCR analysis for
the kanamycin gene (nptII) and Southern blotting were
performed to verify the stable transgene insertion into the
plants’ nuclear DNA. Both transcript expression and PAO
protein activity levels were determined in T0, T1 and T2
generations (Fig. 1) with similar results. The transgenics
overexpressing Mpao showed an approximately 20%
decrease in height and internode length (Table 1). Threetransgenic lines 2.2, 4 and 6.5 were used for further
analysis. Leaves (Fig. 1), stems and roots (data not shown)
were molecularly and biochemically characterized. In all
transgenic tissues, the levels of Mpao transcripts were
higher, comparedwithWT (Fig. 1B, C) and correlatedwith
both higher PAO protein levels in the soluble (Fig. 1D, E)
and the insoluble (data not shown) fraction andwith higher
enzymatic activities (Fig. 1F).
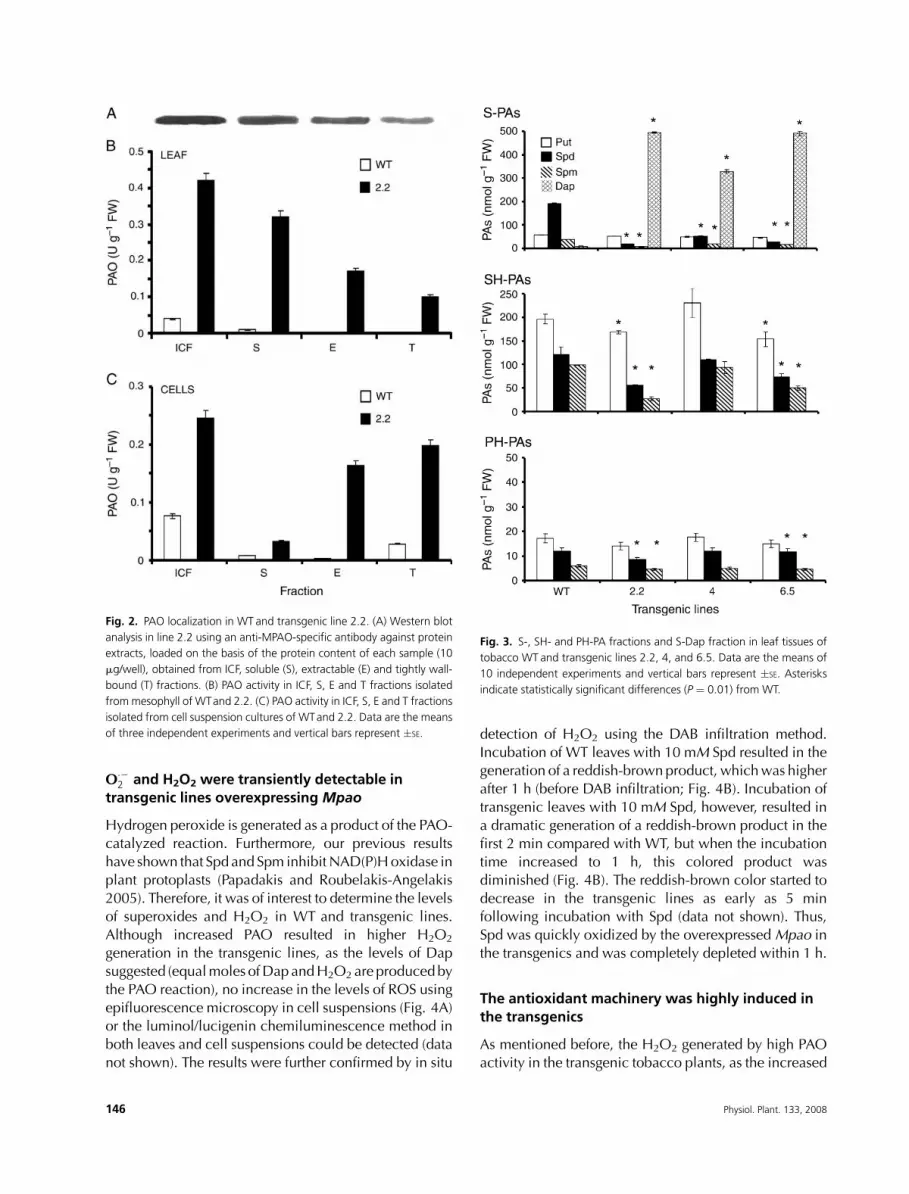
PAO enzymatic activity is localized mainlyextracellularly
The native MPAO protein is mainly localized in the
apoplasticcompartment (Conaetal. 2005). Toexamine the
localization of the recombinant enzyme in the transgenics,
different fractions of ICF were collected from leaf tissues(Fig. 2). Also, the leaf extract was separated in three main
fractions, namely soluble (S), extractable (E) and tightly
wall bound (T) (Fig. 2). Thehighest PAOenzymatic activity
and immunoreactive protein in the transgenics was
localized in the ICF fraction, followed by the S fraction
(Fig. 2A, B). A similar trend was found in the WT plants.
TheE fractionwasobtainedby increasing the ionic strength
of thebuffer toextract the ionically cellwall-boundPAO. Inmature tissues, the E fraction was higher (pellet fraction;
Paschalidis and Roubelakis-Angelakis 2005a), and in
leaves of transgenics, the E fraction accounted for app-
roximately 15–25% of the total PAO activity. In addition,
the T fraction of PAO (obtained after the addition of Triton
X-100 to the pellet) accounted for no more than 8% of the
total PAO activity in transgenics but was not detectable in
the leaves of WT plants.
Overexpression of Mpao reduced significantly theendogenous Spd and Spm levels with concomitanthigh production of Dap
S-PA, SH-PA and PH-PA titers were determined in the first
fully developed leaf ofWTand transgenic lines (Fig. 3). A
general decrease in the titers of soluble higher PAs, andespecially of Spd, was observed in the transgenic lines,
whereas the SH-PA and PH-PA fractions did not decrease
substantially. Put was the more abundant PA and it also
decreased in the transgenics, except in line 4 (Fig. 3). The
decrease in Put levels of transgenic lines 2.2 and 6.5
could be attributed to increase in SAMDC-specific
activities in the leaves, which were 0.58 � 0.03 nmol
CO2mg21 h21, 0.92� 0.06 nmol CO2mg21 h21, 0.57�0.04 nmol CO2 mg21 h21 and 0.81 � 0.04 nmol CO2
mg21 h21 protein in WT, S2.2, S4 and S6.5 lines,
respectively. To exclude the possibility that PAO
could act on Put, or alternatively DAO was upregulated,
the Put oxidizing activity was measured; the transgenics
did not exhibit significant increase in DAO activity (data
not shown). In addition, free Dap levels increased
significantly in all transgenic lines, especially in theleaves (Fig. 3).
Table 1. Biometric characters including height (H), internode length (IL)
and distance of the first fully developed leaf from shoot apex (A) inWTand
transgenic lines 2.2, 4 and 6.5. Values are expressed in mm, and data are
the means of 10 independent plants � SE. Asterisks indicate statistically
significant differences at P � 0.001.
Characteristics WT 2.2 4 6.5
H 62.0 � 1.8 52.6 � 4.3* 57.6 � 2.9 56.2 � 2.4*
IL 11.6 � 0.7 8.5 � 1.1* 8.0 � 0.6* 8.0 � 0.6*
A 21.4 � 2.2 13.0 � 0.6* 17.0 � 1.6 13.2 � 0.6*
Physiol. Plant. 133, 2008 145
O�22 and H2O2 were transiently detectable in
transgenic lines overexpressing Mpao
Hydrogen peroxide is generated as a product of the PAO-
catalyzed reaction. Furthermore, our previous results
have shown that Spd and Spm inhibit NAD(P)Hoxidase in
plant protoplasts (Papadakis and Roubelakis-Angelakis
2005). Therefore, it was of interest to determine the levels
of superoxides and H2O2 in WT and transgenic lines.
Although increased PAO resulted in higher H2O2
generation in the transgenic lines, as the levels of Dap
suggested (equalmoles ofDap andH2O2 are producedby
the PAO reaction), no increase in the levels of ROS using
epifluorescence microscopy in cell suspensions (Fig. 4A)
or the luminol/lucigenin chemiluminescence method in
both leaves and cell suspensions could be detected (data
not shown). The results were further confirmed by in situ
detection of H2O2 using the DAB infiltration method.
Incubation of WT leaves with 10 mM Spd resulted in thegeneration of a reddish-brownproduct, whichwas higher
after 1 h (before DAB infiltration; Fig. 4B). Incubation of
transgenic leaves with 10 mM Spd, however, resulted in
a dramatic generation of a reddish-brown product in the
first 2 min compared with WT, but when the incubation
time increased to 1 h, this colored product was
diminished (Fig. 4B). The reddish-brown color started to
decrease in the transgenic lines as early as 5 minfollowing incubation with Spd (data not shown). Thus,
Spd was quickly oxidized by the overexpressed Mpao in
the transgenics and was completely depleted within 1 h.
The antioxidant machinery was highly induced inthe transgenics
As mentioned before, the H2O2 generated by high PAOactivity in the transgenic tobacco plants, as the increased
Fig. 2. PAO localization in WT and transgenic line 2.2. (A) Western blot
analysis in line 2.2 using an anti-MPAO-specific antibody against protein
extracts, loaded on the basis of the protein content of each sample (10
mg/well), obtained from ICF, soluble (S), extractable (E) and tightly wall-
bound (T) fractions. (B) PAO activity in ICF, S, E and T fractions isolated
frommesophyll of WTand 2.2. (C) PAO activity in ICF, S, E and T fractions
isolated from cell suspension cultures of WTand 2.2. Data are the means
of three independent experiments and vertical bars represent �SE.
Fig. 3. S-, SH- and PH-PA fractions and S-Dap fraction in leaf tissues of
tobacco WT and transgenic lines 2.2, 4, and 6.5. Data are the means of
10 independent experiments and vertical bars represent �SE. Asterisks
indicate statistically significant differences (P ¼ 0.01) from WT.
146 Physiol. Plant. 133, 2008

Dap levels suggested (Fig. 3), could not be detected by
in vitro measurements or by in situ staining but was only
detected when DAB and Spd were infiltrated simulta-
neously or within a 2-min interval (Fig. 4). These results
suggested that the antioxidant machinery should have
been induced promptly in the transgenics. Thus, the
antioxidant enzyme activities were monitored. Increasedspecific activities of PAOwas accompanied by increased
POX (EC 1.11.1.7) activity in the overexpressing lines
(Fig. 5B) and correlated well with the PAO levels (Fig. 1).
The abundance of themost anodic POX isoform 6 and the
cathodic isoform 1 was greater in all transgenic lines and
that of isoforms 4 and 5 was lower, whereas isoforms 2
and 3 were unaffected toMpao overexpression (Fig. 5A).
Isoform 1 could be implicated in ROS detoxification incells, whereas isoform 6 could participate in the cell wall
cross-linking, although a dual role for both POXs cannot
be ruled out. Anodic POX isoforms are considered to be
localized in the cell wall contributing to developmental
processes, such as the regulation of the balance between
cell wall expansion and stiffening (Boeuf et al. 1999).
PAs have been reported to inhibit NADPH oxidase in
both animals and plants (Ogata et al. 1996, Papadakis andRoubelakis-Angelakis 2005) and to exert a protective role
via this inhibition. Therefore, a decrease in PA titers might
have resulted in higher NADPH oxidase activity and O�22
levels, the latter being scavenged by SOD (EC 1.15.1.1).
The specific activity of SOD increased by 28, 44 and 30%
in the transgenic lines 2.2, 4 and 6.5, respectively,
compared withWT (Fig. 6B). Three isoenzymes were de-
tected: the slower migrating in native PAGE MnSODmit,followed by Cu/ZnSODcyt and the faster migrating
FeSODchl (Fig. 6A). The increase in SOD activity was
mainly because of the increased cytosolic isoform, as
indicated by the isoenzymic analysis and immunoreac-
tive SOD protein (Fig. 6C).
In parallel to SOD activity, CAT (EC 1.11.1.6)-specific
activity increased with two main isoforms being induced
(Fig. 6E,D), as did CATprotein levels in the overexpressing
Fig. 4. In vivo ROS epifluorescence in suspension cells and in situ DAB detection of H2O2 (in leaves) inWT tobacco and transgenic lines 2.2, 4 and 6.5. (A)
Epifluorescence ROS detection inWT, 2.2, 4 and 6.5 cell suspension cultures. (B) H2O2 detection using the DAB infiltrationmethod. Leaveswere infiltrated
with 10 mM Spd and incubated for 0 min (syringe spot at the bottom right of the leaf), 1 min (top left), 2 min (bottom left) and 1 h (top right) before the
infiltration of DAB and decoloration, as described in Materials and methods.
Fig. 5. Native IEF of POX and enzymatic specific activity in the leaf soluble
fraction of tobaccoWTand transgenic lines 2.2, 4, and 6.5. (A) IEF at a pH
gradient 3–10. (B) Specific activity. Data are the means of three
independent experiments and bars represent�SE. POX activity presented
statistically significant differences (P¼ 0.01) in lines 4 and 6.5, compared
with WT.
Physiol. Plant. 133, 2008 147

148 Physiol. Plant. 133, 2008

lines (Fig. 6F), and followed similar pattern to PAO levels.
These results could support the posttranslational regula-
tion of CAT, possibly via the action of PAs, as Kim et al.
(2001) and Rhee et al. (2005) have already proposed.
APX (EC 1.11.1.11) is a H2O2-detoxifying enzymewith
low Km for its substrate, acting especially at low substrate
concentrations. At higher concentrations of H2O2, the
main detoxifying effect may be exerted by CAT. The main
cytosolic isoenzyme of APX (Fig. 6H) and APX (Fig. 6I)
activity increased, in a pattern similar to that of SOD and
CAT. It is of interest that line 4, one of the less Mpao
overexpressing lines with lower PAO levels (Fig. 1) and
higher PA levels (Fig. 3), showed the highest expression
levels of POX (Fig. 5), SOD (Fig. 6A, B, C) and APX
(Fig. 6H, I), compared with lines 2.2 and 6.5. Thus,
moderate overexpression of Mpao in line 4 (Fig. 1), and
PA levels (Fig. 3), resulted in stronger induction of the
above-mentioned antioxidant enzymes, whereas higher
Mpao expression levels in line 2.2 (and line 6.5), resulting
to even lower titers of PAs, was associated with lower
induction of the antioxidant enzymes, suggesting that
higherMpao overexpression beyond a specific threshold
results in higher accumulation of ROS and inability of the
antioxidant machinery.
In contrast to animal cells, plant cells synthesize high
amounts of ASA, an additional hydrophilic redox buffer
that provides robust protection against oxidative chal-
lenge (Laloi et al. 2004). The redox state of ASA is
controlled by, among others, two enzymes of Halliwell–
Asada pathway, MDHAR (EC 1.6.54) and DHAR (EC
1.8.5.1), which increased significantly in the transgenic
lines (Fig. 6G). In addition, ASA content decreased in 2.2
and 6.5 transgenic lines, whereas DHA increased
(Table 2). Antioxidants, such as ASA, may induce the
expression of genes associated with abiotic stress pro-
tection and provide information about the cellular redox
homeostasis (Foyer and Noctor 2005), which, in turn,
could result in appropriate induction of events linked to
development/acclimation processes or, alternatively,
execution of cell death programs.
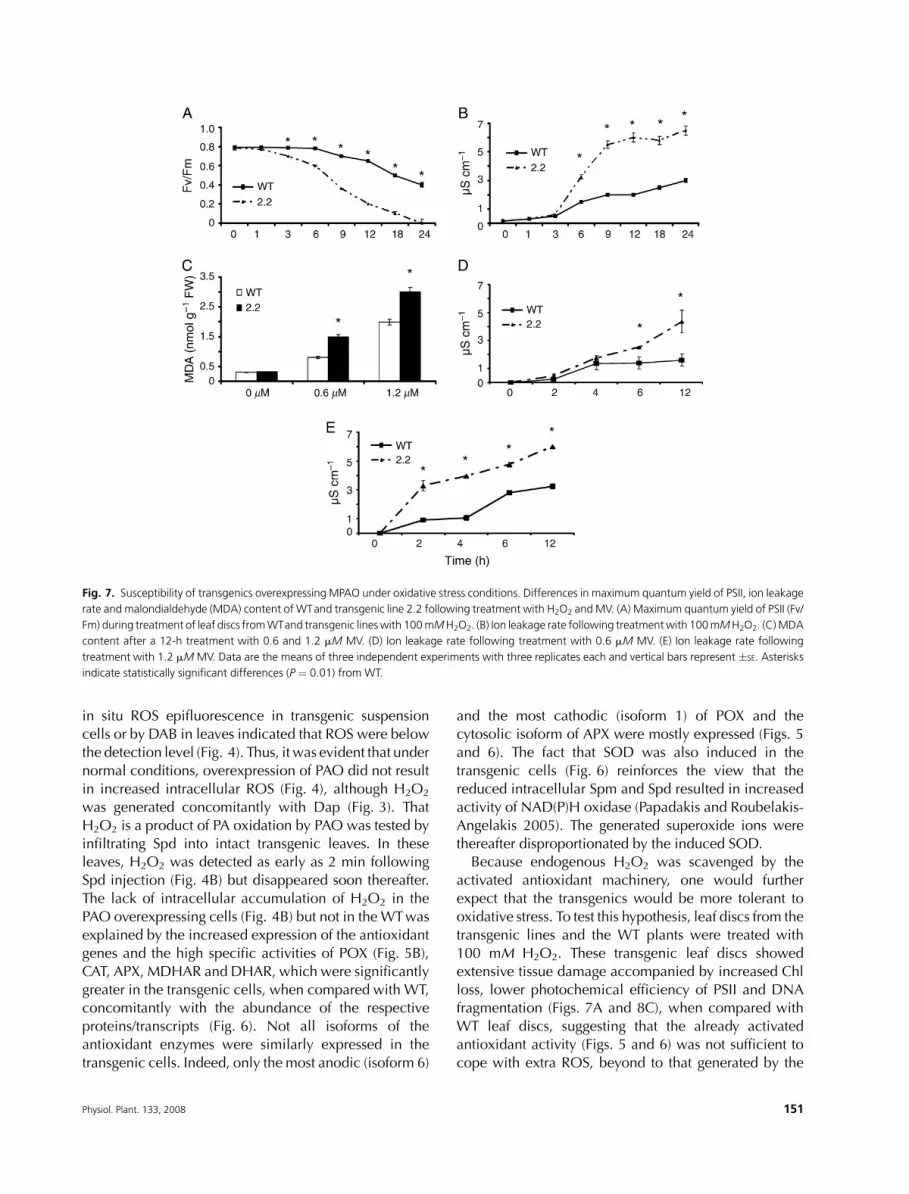
Transgenic lines overexpressing Mpao weresusceptible to induced oxidative stress
Because the transgenic lines exhibited increased steady-
state antioxidant activity, one would expect that they
could also be more tolerant to additional ROS, induced
by abiotic agents. Leaf discs from the transgenics treated
with 100 mM H2O2 showed Chl loss and decreased
photochemical efficiency of PSII (Fv/Fm; Fig. 7A) accom-
panied by ion leakage (Fig. 7B), lipid peroxidation
(Fig. 7C), extensive tissue damage (Fig. 8C) and DNAfragmentation (data not shown). Similar results were
observed using 0.6 and 1.2 mM methylviologen (MV)
under normal photon flux conditions (50 mmol m22 s21)
(Fig. 7D, E).On the contrary, leaf discs from theWTplants
were significantly more tolerant. These data indicate that
major decline in PA levels, namely Spd and Spm, may
well result in increased susceptibility of the transgenics to
oxidative stress, expressed as instability of the photosyn-thetic apparatus and loss of cell membrane integrity.
To examine what was the reason of failure of the
overexpressers to efficiently scavenge the additional
H2O2, the antioxidant enzymes POX, CAT, SOD and
APX were studied after treatment with H2O2 in both WT
and transgenic lines; POX, CAT and SOD increased the
first 6–12 h only in the WT and declined to basal levels
thereafter. Moreover, two isoenzymes of CAT wereinduced during stress, whereas an increment of all SOD
isoenzymes could be observed inWT (Fig. 9B,D). Similar
results were obtained when plants or cell suspension
cultures were treated with MV or menadione, respec-
tively (data not shown). As APX is labile to high H2O2
concentrations, it was quickly inactivated after 6 h; the
inactivation occurred more rapidly in the transgenics and
the same trendwas observed for SOD,which is also labileto high H2O2 (Fig. 9F, G), suggesting that transgenic cells
accumulate higher amounts of ROS or, alternatively, PA
depletion may have a detrimental effect on enzymatic
activities. Moreover, higher lipid peroxidation and ROS
accumulation in the transgenicswere observedwhen leaf
discs or cells were treated with MV or menadione,
Fig. 6. Isoenzymic analyses and specific activities of SOD, CATand APX, and immunoreactive SOD andCAT protein in the soluble fraction of leaves inWT
and 2.2, 4 and 6.5 transgenic lines. (A) Detection of isoenzymes of SODby activity staining of protein extracts separated in native gel. (B) Specific activity of
SOD. (C) Western blot of the cytoplasmic SOD protein (top of C), Coomassie blue staining in 7.5% PAGE (second from top of C), rehybridization of the
same membrane with anti-actin polyclonal antibody (third from top of C) and densitometric analysis of the SOD protein (bottom of C). (D) Detection of
isoenzymes of CAT by activity staining of protein extracts separated in native gel. (E) Specific activity of CAT. (F) Western blot of CAT protein (top of F),
Coomassie Blue staining in 7.5% PAGE (second from top of F), rehybridization of the samemembrane with anti-actin polyclonal antibody (third from top
of F) and densitometric analysis of the CAT protein (bottom of F). (G) Specific activities of MDHAR and DHAR. (H) APX activity staining in protein extracts
separated in native gel. (I) Specific activity of APX. Numerical data are the means of three independent experiments and bars represent �SE. Analysis of
variance showed that data on SOD, CATand APX in transgenic lines were significantly different (P¼ 0.01), comparedwith the corresponding data inWT.
Similarly, DHAR activity was also significantly different in lines 2.2 and 6.5 and MDHAR activity in line 2.2, compared with WT.
Physiol. Plant. 133, 2008 149

respectively (see also below; Figs. 7C and 8B), suggestingthe inability of the transgenic cells to control and retain
low levels of ROS. The reduced PA levels may account for
the higher ROS accumulation, either through their
protective role on the antioxidant mechanism or through
a positive effect on specific antioxidant activities. A direct
role for PAs on ROS scavenging cannot be ruled out. An
interesting finding was the differential induction of
protease activity between WT and transgenics duringstress conditions (Fig. 9H, I).
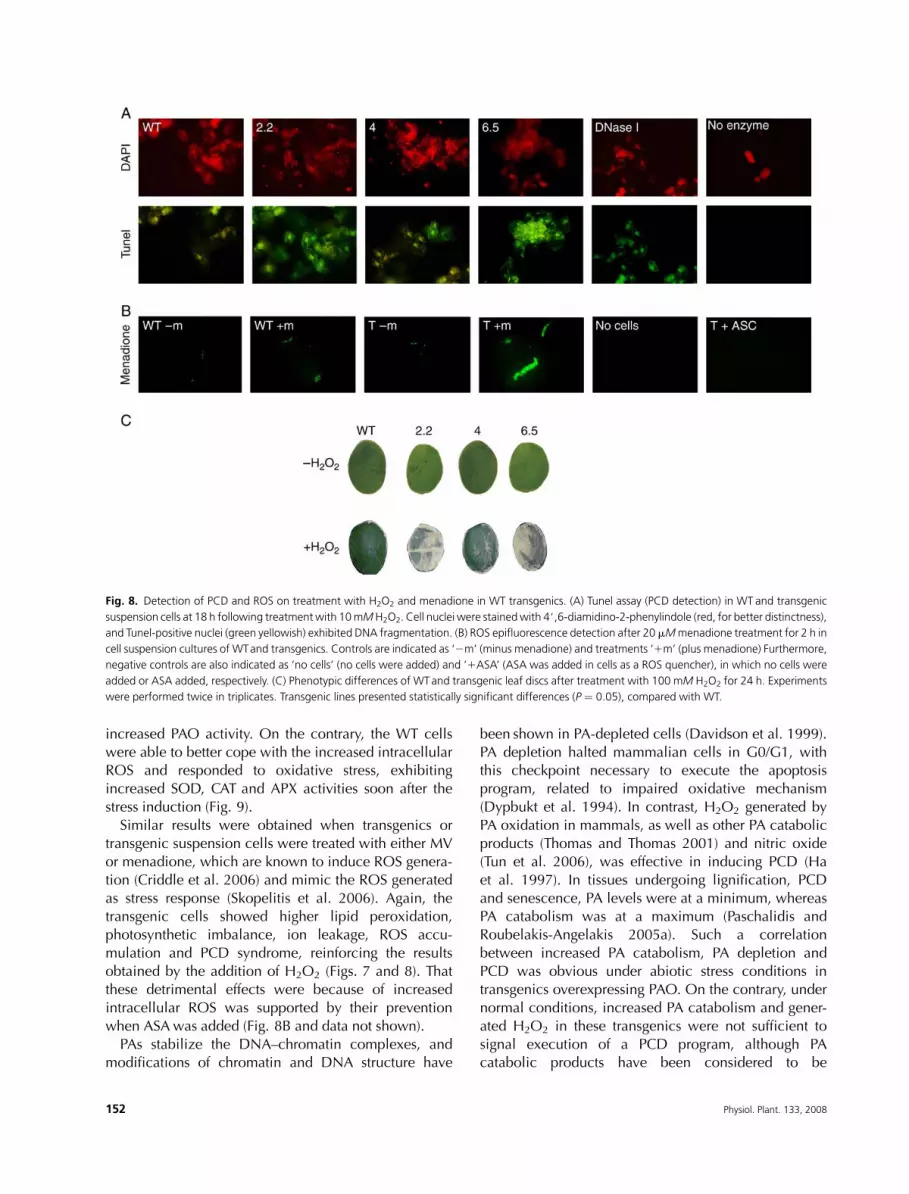
To examinewhether the detrimental effect ofH2O2was
due to induction of the PCD syndrome, nuclear DNA
fragmentation (Tunel) in cells (Fig. 8A) was performed.
Positive Tunel nuclei occurred to a great extent in
transgenic cells, suggesting that the resulting damages
were because of the execution of a PCD program,
possibly triggered by excessive ROS.Menadione was used as an elicitor of ROS (Skopelitis
et al. 2006) to test their differential accumulation in
transgenics during stress conditions. The accumulation of
ROS only in transgenic lines 2 h after the incubation with
10 or 20 mM menadione suggested that the antioxidant
machinery in transgenics is insufficient to sustain ROS
levels within a normal limit even 1 h after the incubation
with the elicitor. The simultaneous incubation of trans-genic cells with ASA reduced ROS levels to almost non-
detectable amounts (Fig. 8B; T 1ASA).
Discussion
Hydrogen peroxide at low concentrations can participate
in the signaling cascade, inducing the expression of
various genes, including the antioxidant ones and/orother stress-responsive genes, whereas at high concen-
trations, H2O2 participates in the induction of the PCD
syndrome (Papadakis and Roubelakis-Angelakis 2005,
Skopelitis et al. 2006, Vacca et al. 2004). Also, H2O2 is
the substrate for the POX-mediated cross-linking of cell
walls (Fry et al. 2000). Thus, the effect of PAs in plant
growth, development and stress responses could be
exerted either directly or via the H2O2 generated by theirPAO-mediated oxidation. Furthermore, Spd and to
a lesser extend Spm inhibit, at least in in vitro plantmodels such as plant protoplasts, NAD(P)H oxidase
which under stress conditions mediates superoxide
generation, whereas Put in the same in vitro model
prevents the execution of the PCD syndrome (Papadakis
and Roubelakis-Angelakis 2005). Therefore, PAs seem to
contribute to the homeostasis of ROS. They could act as
direct radical scavengers; Spm and other PAs have been
reported to act as antioxidants directly or through thechelation of Fe21 that catalyzes the generation of OH
radical via the Fenton reaction (Lovaas 1997) or by
inhibiting the NADPH oxidase mediating the production
of superoxide ions (Papadakis and Roubelakis-Angelakis
2005).
Recently, the studies aiming to show the molecular/
physiological roles of PAs have been facilitated by the
availability of transgenic plants, with altered expressionof the adc gene in rice (Capell et al. 2004), the spds gene
in Arabidopsis (Kasukabe et al. 2004), the samdc gene in
tomato (Mehta et al. 2002) and theMpao gene in tobacco
(Rea et al. 2004). These transgenics have mostly been
assessed for their stress response or phenotype alteration,
that is the adc overexpressors show improved tolerance to
drought (Capell et al. 2004) and the samdcoverexpressors
have greater lycopene content in tomato fruits (Mehtaet al. 2002). In parallel to work by Rea et al. (2004), we
developed tobacco transgenics overexpressing theMpao
gene (Fig. 1). Three transgenic lines were selected that
exhibited more than 10-fold greater PAO-specific activ-
ities compared with the WT, in agreement with the
respective abundance of the Mpao transcripts and PAO
protein (Fig. 1). A statistically significant reduction in the
endogenous Spd and Spmwas found, in analogy with thedramatic increase in the abundance of PAO activity, and
this reduction was accompanied by higher free Dap
content, which, along with H2O2, is a product of the PA
oxidation (Fig. 3).
Because H2O2 is the product of PA oxidation by PAO,
one would expect that there should be a parallel increase
in the endogenous ROS content in the transgenic cells.
Attempts to measure the endogenous H2O2 using thelucigenin/luminol reactions (data not shown) or to detect
Table 2. ASA and DHA contents, ASA pool and redox state. ASA and DHA content, ASA pool (ASA1 DHA) and redox state (ASA/DHA) determined in
tobacco WTand transgenics. Data are the means of two independent experiments � SE. Asterisks indicate statistically significant differences (P ¼ 0.01)
from WT.
Line ASA (mmol g21 FW) DHA (mmol g21 FW) ASA 1 DHA (mmol g21 FW) ASA/DHA
WT 0.898 � 0.014 0.122 � 0.005 1.020 � 0.022 7.33 � 0.08
2.2 0.845 � 0.011* 0.138 � 0.006* 0.982 � 0.019 6.12 � 0.04*
4 0.899 � 0.016 0.127 � 0.011 1.025 � 0.024 7.09 � 0.12
6.5 0.841 � 0.012* 0.154 � 0.012* 0.995 � 0.028 5.47 � 0.14*
150 Physiol. Plant. 133, 2008
in situ ROS epifluorescence in transgenic suspension
cells or by DAB in leaves indicated that ROS were below
the detection level (Fig. 4). Thus, it was evident that under
normal conditions, overexpression of PAO did not result
in increased intracellular ROS (Fig. 4), although H2O2
was generated concomitantly with Dap (Fig. 3). That
H2O2 is a product of PA oxidation by PAO was tested by
infiltrating Spd into intact transgenic leaves. In theseleaves, H2O2 was detected as early as 2 min following
Spd injection (Fig. 4B) but disappeared soon thereafter.
The lack of intracellular accumulation of H2O2 in the
PAO overexpressing cells (Fig. 4B) but not in the WTwas
explained by the increased expression of the antioxidant
genes and the high specific activities of POX (Fig. 5B),
CAT, APX, MDHAR and DHAR, which were significantly
greater in the transgenic cells, when compared with WT,concomitantly with the abundance of the respective
proteins/transcripts (Fig. 6). Not all isoforms of the
antioxidant enzymes were similarly expressed in the
transgenic cells. Indeed, only the most anodic (isoform 6)
and the most cathodic (isoform 1) of POX and the
cytosolic isoform of APX were mostly expressed (Figs. 5
and 6). The fact that SOD was also induced in the
transgenic cells (Fig. 6) reinforces the view that the
reduced intracellular Spm and Spd resulted in increased
activity of NAD(P)H oxidase (Papadakis and Roubelakis-
Angelakis 2005). The generated superoxide ions were
thereafter disproportionated by the induced SOD.Because endogenous H2O2 was scavenged by the
activated antioxidant machinery, one would further
expect that the transgenics would be more tolerant to
oxidative stress. To test this hypothesis, leaf discs from the
transgenic lines and the WT plants were treated with
100 mM H2O2. These transgenic leaf discs showed
extensive tissue damage accompanied by increased Chl
loss, lower photochemical efficiency of PSII and DNAfragmentation (Figs. 7A and 8C), when compared with
WT leaf discs, suggesting that the already activated
antioxidant activity (Figs. 5 and 6) was not sufficient to
cope with extra ROS, beyond to that generated by the
Fig. 7. Susceptibility of transgenics overexpressing MPAO under oxidative stress conditions. Differences in maximum quantum yield of PSII, ion leakage
rate and malondialdehyde (MDA) content of WTand transgenic line 2.2 following treatment with H2O2 andMV. (A) Maximum quantum yield of PSII (Fv/
Fm) during treatment of leaf discs fromWTand transgenic lineswith 100mMH2O2. (B) Ion leakage rate following treatmentwith 100mMH2O2. (C)MDA
content after a 12-h treatment with 0.6 and 1.2 mM MV. (D) Ion leakage rate following treatment with 0.6 mM MV. (E) Ion leakage rate following
treatment with 1.2 mM MV. Data are the means of three independent experiments with three replicates each and vertical bars represent �SE. Asterisks
indicate statistically significant differences (P ¼ 0.01) from WT.
Physiol. Plant. 133, 2008 151
increased PAO activity. On the contrary, the WT cells
were able to better cope with the increased intracellularROS and responded to oxidative stress, exhibiting
increased SOD, CAT and APX activities soon after the
stress induction (Fig. 9).
Similar results were obtained when transgenics or
transgenic suspension cells were treated with either MV
or menadione, which are known to induce ROS genera-
tion (Criddle et al. 2006) and mimic the ROS generated
as stress response (Skopelitis et al. 2006). Again, thetransgenic cells showed higher lipid peroxidation,
photosynthetic imbalance, ion leakage, ROS accu-
mulation and PCD syndrome, reinforcing the results
obtained by the addition of H2O2 (Figs. 7 and 8). That
these detrimental effects were because of increased
intracellular ROS was supported by their prevention
when ASAwas added (Fig. 8B and data not shown).
PAs stabilize the DNA–chromatin complexes, andmodifications of chromatin and DNA structure have
been shown in PA-depleted cells (Davidson et al. 1999).
PA depletion halted mammalian cells in G0/G1, withthis checkpoint necessary to execute the apoptosis
program, related to impaired oxidative mechanism
(Dypbukt et al. 1994). In contrast, H2O2 generated by
PA oxidation in mammals, as well as other PA catabolic
products (Thomas and Thomas 2001) and nitric oxide
(Tun et al. 2006), was effective in inducing PCD (Ha
et al. 1997). In tissues undergoing lignification, PCD
and senescence, PA levels were at a minimum, whereasPA catabolism was at a maximum (Paschalidis and
Roubelakis-Angelakis 2005a). Such a correlation
between increased PA catabolism, PA depletion and
PCD was obvious under abiotic stress conditions in
transgenics overexpressing PAO. On the contrary, under
normal conditions, increased PA catabolism and gener-
ated H2O2 in these transgenics were not sufficient to
signal execution of a PCD program, although PAcatabolic products have been considered to be
Fig. 8. Detection of PCD and ROS on treatment with H2O2 and menadione in WT transgenics. (A) Tunel assay (PCD detection) in WT and transgenic
suspension cells at 18 h following treatmentwith 10mMH2O2. Cell nuclei were stainedwith 4#,6-diamidino-2-phenylindole (red, for better distinctness),
and Tunel-positive nuclei (green yellowish) exhibited DNA fragmentation. (B) ROS epifluorescence detection after 20 mMmenadione treatment for 2 h in
cell suspension cultures of WTand transgenics. Controls are indicated as ‘2m’ (minus menadione) and treatments ‘1m’ (plus menadione) Furthermore,
negative controls are also indicated as ‘no cells’ (no cells were added) and ‘1ASA’ (ASA was added in cells as a ROS quencher), in which no cells were
added or ASA added, respectively. (C) Phenotypic differences of WTand transgenic leaf discs after treatment with 100 mM H2O2 for 24 h. Experiments
were performed twice in triplicates. Transgenic lines presented statistically significant differences (P ¼ 0.05), compared with WT.
152 Physiol. Plant. 133, 2008

pro-apoptotic signals (Thomas and Thomas 2001). High
PA levels have been correlated with cell wall formation,
high redox state and protoplast regeneration (Papadakis
et al. 2005), whereas Spd- and Spm-treated protoplasts
accumulated high levels of intracellular H2O2,
generated by PAO, whose activity increased along withCAT activity (Papadakis and Roubelakis-Angelakis
2005).
As H2O2 has a high diffusion rate inside the cell, a dual
role for H2O2, dependent on specific concentrations, can
be proposed: (1) low augmentation in H2O2 concen-
trations (that may remain in cell walls), caused by PAO
overexpression, successfully acted as signal for the
induction of the antioxidant machinery, leading to cellsurvival/adaptation and normal plant growth and (2)
higher H2O2 concentration (that may also diffuse in the
secretory pathway), caused not only by PAO over-
expression but also by other mechanisms such as
NAD(P)H oxidase/SOD reactions or by exogenous
addition, induced susceptibility and PCD. H2O2 concen-
trations beyond a specific threshold shifted the response
from normal growth to susceptibility and induced thePCD syndrome in the transgenics. This is of interest
because it is now widely accepted that H2O2 is a nodal
point in the signal transduction pathway, which, depend-
ing on its concentration (‘signature’), may target different
subsets of genes. When the antioxidant machinery has
been induced, further increase of ROS do not act as a new
signal for the expression of the antioxidant genes, and this
antioxidantmachinery is no longer active and sufficient toprevent oxidative damage. Instead, the higher ROS levels
induce expression of the PCD syndrome. Whether this
inability for further antioxidant activity is the result and/or
linked to the reduced PA levels and if the homeostasis of
the endogenous PAs contributes to the regulation of the
antioxidant machinery of cells and in doing that
a threshold of endogenous concentration should be
maintained remains an open question.
Acknowledgements – The authors are grateful to Dr P.
Tavladoraki (University Roma Tre, Italy) for the generous gifts
of anti-MPAO polyclonal antibody andMpao cDNA and to Dr
G. Inamine (United States Department of Agriculture, Belts-
ville, MD) for providing the antibody against CAT. The project
was co-funded by the European Social Fund and National
resources, projects Herakleitos and Pythagoras, and was
implemented in the frame of COST858.
References
Alvarez ME, Pennell RI, Meijer PJ, Ishikawa A, Dixon RA,
Lamb C (1998) Reactive oxygen intermediates mediate
a systemic signal network in the establishment of plant
immunity. Cell 92: 773–784
Fig. 9. Expression of POX, CAT, SOD and APX following treatment with
H2O2 in WT and transgenic (2.2) leaf discs. (A) POX activity during
treatmentwith 100mMH2O2. (B) Isoenzymic analysis of CAT in 0, 12 and
24 h. (C) Analysis of CAT activity during treatment with 100 mM H2O2.
(D) Isoenzymic analysis of SOD in 0, 12 and 24 h. (E) Analysis of SOD
activity during treatment with 100 mM H2O2. (F) Isoenzymic analysis of
APX. (G) Analysis of APX activity during treatment with 100 mM H2O2.
(H) Native gel for protease at 0 h and 18 h and (I) Relative protease activity
(densitometry of the native in H) at 0 h and 18 h during treatment with
100 mM H2O2. Data are the means of three independent experiments
with three replicates each and vertical bars represent �SE. Asterisks
indicate statistically significant differences (P ¼ 0.01) from WT.
Physiol. Plant. 133, 2008 153

Amendola R, Bellini A, Cervelli M, Degan P, Marcocci L,
Martini F, Mariottini P (2005) Direct oxidative DNA
damage, apoptosis and radio sensitivity by spermine
oxidase activities in mouse neuroblastoma cells. Biochim
Biophys Acta 1755: 15–24
Angelini R, Federico R, Bonfante P (1995) Maize polyamine
oxidase: antibody production and ultrastructural
localization. J Plant Physiol 145: 686–669
Arias M, Carbonell J, Agusti M (2005) Endogenous free
polyamines and their role in fruit set of low and high
parthenocarpic ability citrus cultivars. J Plant Physiol 162:
845–853
Asada K (1984) Chloroplasts: formation of active oxygen and
its scavenging. Methods Enzymol 105: 422–429
Asai T, Stone JM, Heard JE, Kovtun Y, Yorgey P, Sheen J,
Ausubel FM (2000) Fumonisin B1-induced cell death in
Arabidopsis protoplasts requires jasmonate-, ethylene-,
and salicylate-dependent signalling pathways. Plant Cell
12: 1823–1835
Beauchamp C, Fridovitch I (1971) Superoxide dismutase:
improved assays and an assay applicable to acrylamide
gels. Anal Biochem 44: 276–287
Boeuf G, Legrand B, Rambour S (1999) Influence of light
conditions on development, apoplastic peroxidase
activities and peroxidase isoenzymes in chicory root
explants. Physiol Plant 106: 331–336
Capell T, Bassie L, Christou P (2004) Modulation of the
polyamine biosynthetic pathway in transgenic rice confers
tolerance to drought stress. Proc Natl Acad Sci USA 101:
9909–9914
Clare DA, Duong MN, Darr D, Archibald F, Fridovich I
(1984) Effects of molecular oxygen on detection of
superoxide radical with nitroblue tetrazolium and on
activity stains for catalase. Anal Biochem 140: 532–537
Cona A, Moreno S, Cenci F, Federico R, Angelini R (2005)
Cellular re-distribution of flavin-containing polyamine
oxidase in differentiating root and mesocotyl of Zea mays
L. seedlings. Planta 221: 265–276
Cona A, Rea G, Angelini R, Federico R, Tavladoraki P (2006)
Functions of amine oxidases in plant development and
defence. Trends Plant Sci 11: 80–88
Criddle DN, Gillies S, Baumgartner-Wilson HK, Jaffar M,
Chinje EC, Passmore S, Chvanov M, Barrow S,
Gerasimenko OV, Tepikin AV, Sutto R, Petersen OH (2006)
Menadione-induced reactive oxygen species generation
via redox cycling promotes apoptosis of murine pancreatic
acinar cells. J Biol Chem 281: 40485–40492
Davidson NE, Hahm HA, McCloskey DE, Woster PM, Casero
J (1999) Clinical aspects of cell death in breast cancer: the
polyamine pathway as a new target for treatment. Endocr
Relat Cancer 6: 69–73
Dypbukt JM, Ankarcrona M, Burkitt M, Sjoholm A, Strom K,
Orrenius S, Nicotera P (1994) Different prooxidant
levels stimulate growth, trigger apoptosis, or produce
necrosis of insulin-secreting RINm5F cells. The role
of intracellular polyamines. J Biol Chem 269:
30553–30560
Federico R, Angelini R, Cesta A, Pini C (1985) Determination
of diamine oxidase in lentil seedlings by enzymic
activity and immunoreactivity. Plant Physiol 79:
62–64
Fos M, Proano K, Alabadi D, Nuez F, Carbonell J,
Garcia-Martinez JL (2003) Polyamine metabolism is
altered in unpollinated parthenocarpic pat-2 tomato
ovaries. Plant Physiol 131: 359–366
Foyer CH, Noctor G (2005) Oxidant and antioxidant
signalling in plants: a re-evaluation of the concept of
oxidative stress in a physiological context. Plant Cell Env
28: 1056–1071
Fry SC, Willis SC, Paterson AEJ (2000) Intraprotoplasmic and
wall-localised formation of arabinoxylan-bound
diferulates and larger ferulate coupling-products in maize
cell-suspension cultures. Planta 211: 679–692
Ha HC, Woster PM, Yager JD, Casero RA (1997) The role of
polyamine catabolism in polyamine analogue-induced
programmed cell death. Proc Natl Acad Sci USA 94:
11557–11562
Hashimoto T, Tamaki K, Suzuki K, Yamada Y (1998)
Molecular cloning of plant spermidine synthases. Plant
Cell Physiol 39: 73–79
Heath RL, Packer L (1968) Photoperoxidation in isolated
chloroplast: I. Kinetics and stoichiometry of fatty acid
peroxidation. Arch Biochem Biophys 125: 189–198
Iandolino AB, da Silva FG, Lim H, Choi H, Williams LE, Cook
DR (2004) High-quality RNA, cDNA, and derived EST
libraries from Grapevine (Vitis vinifera L). PMB Rep 22:
269–278
Kasukabe Y, He L, Nada K, Misawa S, Ihara I, Tachibana S
(2004) Over-expression of spermidine synthase enhances
tolerance to multiple environmental stresses and
up-regulates the expression of various stress-regulated
genes in transgenic Arabidopsis thaliana. Plant Cell Physiol
45: 712–722
Kaur-Sawhney R, Tiburcio AF, Atabella T, Galston AW (2003)
Polyamines in plants: an overview. J Cell Mol Biol 2: 1–12
Kim SY, Kwon OJ, Park JW (2001) Inactivation of catalase and
superoxide dismutase by singlet oxygen derived from
photoactivated dye. Biochimie 83: 4372444
Kotzabasis K, Christakis-Hampsas MD, Roubelakis-Angelakis
KA (1993) A narrow-bore HPLC method for the
identification and quantitation of free, conjugated and
bound polyamines. Anal Biochem 214: 484–489
Kusano T, Yamaguchi K, Berberich T, Takahashi Y (2007)
Advances in polyamine research in 2007. J Plant Res 120:
345–350
Kuznetsov V, Shorina M, Aronova E, Stetsenko L, Rakitin V,
Shevyakova N (2007) NaCl- and ethylene-dependent
cadaverine accumulation and its possible protective role in
the adaptation of the common ice plant to salt stress. Plant
Sci 172: 363–370
154 Physiol. Plant. 133, 2008

Laloi C, Apel K, Danon A (2004) Reactive oxygen signalling:
the latest new. Cur Opin Plant Biol 7: 323–328
Lovaas E (1997) Antioxidative and metal-chelating effects of
polyamines. Adv Pharmacol 38: 119–149
Mehta RA, Cassol T, Li N, Ali N, Handa AK, Mattoo AK
(2002) Engineered polyamine accumulation in tomato
enhances phytonutrient content, juice quality, and vine
life. Nat Biotechnol 20: 613–618
Mittler R (2006) Abiotic stress, the field environment and
stress combination. Trends Plant Sci 11: 15–19
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged
by ascorbate-specific peroxidase in spinach chloroplasts.
Plant Cell Physiol 22: 876–880
Ogata K, Nishimoto N, Uhlinger DJ, Igarashi K, Takeshita M
(1996) Spermine suppresses the activation of human
neutrophil NADPH oxidase in cell-free and
semi-recombinant systems. Biochem J 313: 549–554
Panicot M, Minguet EG, Ferrando A, Alcazar R, Blazquez
MA, Carbonell J, Altabella T, Koncz C, Tiburcio AF
(2002) A polyamine metabolon involving aminopropyl
transferase complexes in Arabidopsis. Plant Cell 14:
2539–2551
Papadakis AK, Paschalidis KA, Roubelakis-Angelakis KA
(2005) Biosynthesis profile and endogenous titers of
polyamines differ in totipotent and recalcitrant plant
protoplasts. Physiol Plant 125: 10–20
Papadakis AK, Roubelakis-Angelakis KA (2005) Polyamines
inhibit NADPH oxidase-mediated superoxide generation
and putrescine prevents programmed cell death induced
by polyamine oxidase-generated hydrogen peroxide.
Planta 220: 826–837
Paschalidis KA, Aziz A, Geny L, Primikirios NI,
Roubelakis-Angelakis KA (2001) Polyamines in grapevine.
In: Roubelakis-Angelakis KA (ed) Molecular Biology and
Biotechnology of the Grapevine. Kluwer Academic
Publishers, Dordrecht, The Netherlands, pp 109–152
Paschalidis KA, Roubelakis-Angelakis KA (2005a) Sites and
regulation of polyamine catabolism in the tobacco plant.
Correlations with cell division/expansion, cell cycle
progression, and vascular development. Plant Physiol 138:
2174–2184
Paschalidis KA, Roubelakis-Angelakis KA (2005b) Spatial and
temporal distribution of polyamine levels and polyamine
anabolism in different organs/tissues of the tobacco plant.
Correlations with age, cell division/expansion, and
differentiation. Plant Physiol 138: 142–152
Perez-Amador MA, Leon J, Green PJ, Carbonell J (2002)
Induction of the arginine decarboxylase ADC2 gene provides
evidence for the involvement of polyamines in the wound
response in Arabidopsis. Plant Physiol 130: 1454–1463
Rao M, Hale BA, Ormrod DP (1995) Amelioration of
ozone-induced oxidative damage in wheat plants grown
under high carbon dioxide. Plant Physiol 109: 421–432
Rea G, de Pinto MC, Tavazza R, Biondi S, Gobbi V, Ferrante
P, De Gara L, Federico R, Angelini R, Tavladoraki P (2004)
Ectopic expression of maize polyamine oxidase and pea
copper amine oxidase in the cell wall of tobacco plants.
Plant Physiol 134: 1414–1426
Rhee SG, Yong KS, Kang SW, Woo HA, Chang TS (2005)
Controlled elimination of intracellular H2O2. Regulation
of peroxiredoxin, catalase, and glutathione peroxidase via
post-translational modification. Antioxid Redox Signal 7:
619–626
Roje S (2006) S-Adenosyl-L-methionine: beyond the
universal methyl group donor. Phytochemistry 67:
1686–1698
Sasaki-Sekimoto Y, Taki N, Obayashi T, Aono M, Matsumoto
F, Sakurai N, Suzuki H, Hirai MY, Noji M, Saito K, Masuda
T, Takamiya K-I, Shibata D, Ohta H (2005) Coordinated
activation of metabolic pathways for antioxidants and
defence compounds by jasmonates and their roles in
stress tolerance in Arabidopsis. Plant J 44:
653–668
Sato F, Hashimoto T, Hachiya A, Tamura K, Choi K,
Morishige T, Fujimoto H, Yamada Y (2001) Metabolic
engineering of plant alkaloid biosynthesis. Proc Natl Acad
Sci USA 98: 367–372
Skopelitis DS, Paranychianakis NV, Paschalidis KA, Pliakonis
ED, Delis ID, Yakoumakis DI, Kouvarakis A, Papadakis AK,
Stephanou EG, Roubelakis-Angelakis KA (2006) Abiotic
stress generates ROS that signal expression of anionic
glutamate dehydrogenases to form glutamate for proline
synthesis in tobacco and grapevine. Plant Cell 18:
2767–2781
Serrano A, Cordoba F, Gonzales-Reyes JA, Navas P, Villalba
JM (1994) Purification and characterization of two distinct
NAD(P)H dehydrogenases from onion (Allium cepa L) root
plasma membrane. Plant Physiol 106: 87–96
Takahashi Y, Uehara Y, Berberich T, Ito A, Saitoh H, Miyazaki
A, Terauchi R, Kusano T (2004) A subset of hypersensitive
response marker genes, including HSR203J, is the
downstream target of a spermine signal transduction
pathway in tobacco. Plant J 40: 586–595
Tavladoraki P, Schinina ME, Cecconi F, Di Agostino S,
Manera F, Rea G, Mariottini P, Federico R, Angelini R
(1998) Maize polyamine oxidase: primary structure from
protein and cDNA sequencing. FEBS Lett 426: 62–66
Thomas T, Thomas TJ (2001) Polyamines in cell growth and
cell death: molecular mechanism and therapeutic
applications. Cell Mol Life Sci 58: 244–258
Tiburcio AF, Besford RT, Borrell A (1994) Posttranslational
regulation of arginine decarboxylase synthesis by spermine
in osmotically-stressed oat leaves. Biochem Soc Trans 22:
455S
Tun NN, Santa-Catarina C, Begum T, Silveira V, Handro W,
Floh EIS, Scherer GFE (2006) Polyamines induce rapid
biosynthesis of nitric oxide (NO) in Arabidopsis thaliana
seedlings. Plant Cell Physiol 47: 346–354
Vacca RA, de Pinto MC, Valenti D, Passarella S, Marra E,
Gara DL (2004) Production of Reactive oxygen species,
Physiol. Plant. 133, 2008 155

alteration of cytosolic ascorbate peroxidase, and
impairment of mitochondrial metabolism are early events
in heat shock-induced programmed cell death in tobacco
bright-yellow 2 cells. Plant Physiol 134: 1100–1112
Vuosku J, Jokela A, Laara E, Saaskilahti M, Muilu R, Sutela S,
Altabella T, Sarjala T, Haggman H (2006) Consistency of
polyamine profiles and expression of arginine
decarboxylase in mitosis during zygotic embryogenesis of
Scots pine. Plant Physiol 142: 1027–1038
Yamaguchi K, Takahashi Y, Berberich T, Imai A, Miyazaki A,
Takahashi T, Michael A, Kusano T (2006) The polyamine
spermine protects against high salt stress in Arabidopsis
thaliana. FEBS Lett 22: 6783–6788
Yamamoto A, Bhuiyan MNH, Waditee R, Tanaka Y, Esaka M,
Oba K, Jagendorf AT, Takabe T (2005) Suppressed
expression of the apoplastic ascorbate oxidase gene
increases salt tolerance in tobacco and Arabidopsis plants.
J Exp Bot 56: 1785–1796
Yasui Y, Hayashi M (1991) Simultaneous determination of
ascorbic acid and dehydroascorbic acid by high
performance liquid chromatography. Analytical Science 7:
125–128
Yoda H, Yamaguchi Y, Sano H (2003) Induction of
hypersensitive cell death by hydrogen peroxide produced
through polyamine degradation in tobacco plants. Plant
Physiol 132: 1973–1981
Edited by C. H. Foyer
156 Physiol. Plant. 133, 2008
Related Documents











