Transcriptional Down-regulation of Epidermal Growth Factor (EGF) Receptors by Nerve Growth Factor (NGF) in PC12 Cells Gadi Cohen & Keren Ettinger & Shimon Lecht & Peter I. Lelkes & Philip Lazarovici Received: 8 July 2014 /Accepted: 22 July 2014 # Springer Science+Business Media New York 2014 Abstract Nerve growth factor (NGF) treatment causes a pro- found down-regulation of epidermal growth factor (EGF) receptors (EGFR) during the neuronal differentiation of PC12 cells. This process was characterized by a progressive decrease in EGFR level, as measured by 125 I-EGF binding and Scatchard analysis, tyrosine phosphorylation, Western blot- ting, and bio-imaging using EGF-labeled with a near-infrared probe. Differentiation of the cells with NGF for 5–7 days produces a 95 % reduction in the amount of 35 S-methionine- labeled EGFR. This down-regulation does not occur in PC12- nnr5 cells, which lack the TrkA NGF receptor but is reconstituted in these cells upon their stable transfection with TrkA. The process of NGF-induced EGFR down-regulation was inhibited by K252a, a TrkA antagonist and by anti-TrkA antibodies but not by Thx-B, a blocker of the interaction of NGF with p75 NTR receptors. NGF-induced (heterologous) down-regulation, but not EGF-induced (homologous) down- regulation of EGFR, was blocked in Ras-deficient PC12 cells. NGF treatment for 5–7 days of PC12 cells, grown in suspen- sion or in 3D collagen gels, induces down-regulation of EGFR independent of neurite outgrowth. The messenger RNA (mRNA) for EGFR decreased in a comparable fashion. This process was correlated temporally with a decrease in the transcription of the EGFR gene. Treatment with NGF also increased the cellular content of GCF 2 , a putative inhibitory transcription factor of the EGFR gene. The temporal increase in GCF 2 , like the decrease in the EGFR mRNA, was not seen in TrkA deficient PC12 cells nor in cells expressing dominant- negative Ras. The results suggest that NGF-induced down- regulation of the EGFR is under transcriptional control, is TrkA and Ras-dependent, may involve transcriptional repres- sion by GCF 2 , and independent of mechanisms that lead to NGF-induced neurite outgrowth in PC12cells. This heterolo- gous down-regulation of EGFR would appear to be an effi- cient mean of desensitizing the neuron to proliferative stimuli, thereby representing a safety latch for initiating and sustaining NGF-induced neuronal differentiation. Keywords NGF . EGFR . Down-regulation . TrkA . Ras . PC12 cells . Proliferation . Differentiation . Neurite outgrowth . 2D . 3D . Suspension Introduction Neuropeptides, neurotrophins, cytokines, chemokines, and different growth factors play a key role in the mechanism of action of growth factors regulating the development of the nervous system (Sieber-Blum 1998; Johnson 2001; Wang and Zoghbi 2001; Bernd 2008). However, the interactions and cross talk between these diverse factors are still poorly under- stood (Hall and Ekanayake 1991; Sieber-Blum 1998; Fargali et al. 2012; Williamson and Bilbo 2013). Physiological neu- ronal development requires a proper delicate balance between cell proliferation, differentiation, and apoptosis (de la Rosa and de Pablo 2000; Liu and Greene 2001; Becker and Bonni 2005). Neurogenesis, neuronal differentiation, neuroprotec- tion, and repair are usually controlled by the neurotrophin family of growth factors, which includes nerve growth factor (NGF) (Sofroniew et al. 2001). Mini Review for the special issue: From Molecular Biology to Neurobiology in memory of Uriel Littauer. G. Cohen : K. Ettinger : S. Lecht : P. Lazarovici (*) Faculty of Medicine, School of Pharmacy Institute for Drug Research, The Hebrew University of Jerusalem, Jerusalem 91120, Israel e-mail: [email protected] S. Lecht : P. I. Lelkes : P. Lazarovici Department of Bioengineering and Temple Institute for Regenerative Medicine and Engineering, Temple University, Philadelphia, PA 19122, USA J Mol Neurosci DOI 10.1007/s12031-014-0388-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Transcriptional Down-regulation of Epidermal Growth Factor(EGF) Receptors by Nerve Growth Factor (NGF) in PC12 Cells
Gadi Cohen & Keren Ettinger & Shimon Lecht &Peter I. Lelkes & Philip Lazarovici
Received: 8 July 2014 /Accepted: 22 July 2014# Springer Science+Business Media New York 2014
Abstract Nerve growth factor (NGF) treatment causes a pro-found down-regulation of epidermal growth factor (EGF)receptors (EGFR) during the neuronal differentiation ofPC12 cells. This process was characterized by a progressivedecrease in EGFR level, as measured by 125I-EGF binding andScatchard analysis, tyrosine phosphorylation, Western blot-ting, and bio-imaging using EGF-labeled with a near-infraredprobe. Differentiation of the cells with NGF for 5–7 daysproduces a 95 % reduction in the amount of 35S-methionine-labeled EGFR. This down-regulation does not occur in PC12-nnr5 cells, which lack the TrkA NGF receptor but isreconstituted in these cells upon their stable transfection withTrkA. The process of NGF-induced EGFR down-regulationwas inhibited by K252a, a TrkA antagonist and by anti-TrkAantibodies but not by Thx-B, a blocker of the interaction ofNGF with p75NTR receptors. NGF-induced (heterologous)down-regulation, but not EGF-induced (homologous) down-regulation of EGFR, was blocked in Ras-deficient PC12 cells.NGF treatment for 5–7 days of PC12 cells, grown in suspen-sion or in 3D collagen gels, induces down-regulation of EGFRindependent of neurite outgrowth. The messenger RNA(mRNA) for EGFR decreased in a comparable fashion. Thisprocess was correlated temporally with a decrease in thetranscription of the EGFR gene. Treatment with NGF also
increased the cellular content of GCF2, a putative inhibitorytranscription factor of the EGFR gene. The temporal increasein GCF2, like the decrease in the EGFR mRNA, was not seenin TrkA deficient PC12 cells nor in cells expressing dominant-negative Ras. The results suggest that NGF-induced down-regulation of the EGFR is under transcriptional control, isTrkA and Ras-dependent, may involve transcriptional repres-sion by GCF2, and independent of mechanisms that lead toNGF-induced neurite outgrowth in PC12cells. This heterolo-gous down-regulation of EGFR would appear to be an effi-cient mean of desensitizing the neuron to proliferative stimuli,thereby representing a safety latch for initiating and sustainingNGF-induced neuronal differentiation.
Keywords NGF . EGFR . Down-regulation . TrkA . Ras .
PC12 cells . Proliferation . Differentiation . Neuriteoutgrowth . 2D . 3D . Suspension
Introduction
Neuropeptides, neurotrophins, cytokines, chemokines, anddifferent growth factors play a key role in the mechanism ofaction of growth factors regulating the development of thenervous system (Sieber-Blum 1998; Johnson 2001; Wang andZoghbi 2001; Bernd 2008). However, the interactions andcross talk between these diverse factors are still poorly under-stood (Hall and Ekanayake 1991; Sieber-Blum 1998; Fargaliet al. 2012; Williamson and Bilbo 2013). Physiological neu-ronal development requires a proper delicate balance betweencell proliferation, differentiation, and apoptosis (de la Rosaand de Pablo 2000; Liu and Greene 2001; Becker and Bonni2005). Neurogenesis, neuronal differentiation, neuroprotec-tion, and repair are usually controlled by the neurotrophinfamily of growth factors, which includes nerve growth factor(NGF) (Sofroniew et al. 2001).
Mini Review for the special issue: From Molecular Biology toNeurobiology in memory of Uriel Littauer.
G. Cohen :K. Ettinger : S. Lecht : P. Lazarovici (*)Faculty of Medicine, School of Pharmacy Institute for DrugResearch, The Hebrew University of Jerusalem,Jerusalem 91120, Israele-mail: [email protected]
S. Lecht : P. I. Lelkes : P. LazaroviciDepartment of Bioengineering and Temple Institute for RegenerativeMedicine and Engineering, Temple University, Philadelphia,PA 19122, USA
J Mol NeurosciDOI 10.1007/s12031-014-0388-2

NGF is a 13-kD protein that influences the growth andsurvival of sympathetic neurons and chromaffin cells, affectscholinergic neuronal pathways in the brain frontal cortex,stimulates proliferation and differentiation of neuronal stemcells (Ip et al. 1994; Bath and Lee 2010), and contributes tothe maintenance of the differentiated neuronal phenotypein vitro and in vivo (Manni et al. 2013). Embryonic andadult nervous tissues alike also contain a variety of mito-gens, including insulin-like growth factors-I and -II(Devaskar 1991), platelet-derived growth factor (Yeh et al.1991), basic fibroblast growth factor (Abe and Saito 2001),and epidermal growth factor (Plata-Salaman 1991). In theirabsence, neuronal cells are growth-arrested and turn intopost-mitotic neurons.
Epidermal growth factor (EGF) and its receptors are widelyexpressed in both the central and peripheral nervous systems,including in dopaminergic neurons in the basal ganglia (Piaoet al. 2005; Abe et al. 2009) and play an important role in theproliferation, migration, and survival of numerous cell popu-lations in the embryonic brain (Kornblum et al. 1997). Micelacking epidermal growth factor receptors (EGFRs) exhibitbrain defects including progressive neurodegeneration in thefrontal cortex, olfactory bulb, and thalamus, as characterizedby massive apoptosis (Sibilia et al. 1998). These observationssuggest that in EGFR levels, signaling and regulation mayplay an important role during the development of the nervoussystem.
The rat pheochromocytoma cell line (PC12) is an exten-sively used model to study neuronal differentiation (Fujitaet al. 1989; Ravni et al. 2006). Stimulation of PC12 cells withNGF induces a neuronal differentiation program: The cellsexit the cell cycle, develop an elaborate network of branchingneurites (neuritogenesis), and become electrically excitable(Greene and Tischler 1976). Acquisition of a sympatheticneuronal phenotype entails a complex cascade oftranscription-dependent and -independent biological process-es, which are tightly coordinated topographically and tempo-rally (Greene and Angelastro 2005; van Dijkmans et al. 2008;Mullenbrock et al. 2011). The initial signaling steps triggeredby NGF stimulation are well characterized (Kaplan andMiller2000). After NGF binding to its receptors p75NTR and thetropomyosin-related kinase A (TrkA) receptor, TrkA receptorsdimerize and subsequently autophosphorylate followed byrecruitment of complex of signaling molecules initiating in-tracellular signaling cascades involving, among others, PI3K-Akt, PLCγ-IP3, and Ras-mitogen activated protein kinase(MAPK). NGF-induced neuronal differentiation requires ac-tivation of the TrkA receptor and proceeds downstreamthrough the Ras/MAPK pathway (Vaudry et al. 2002). Onceactivated and translocated to the nucleus, phosphorylatedtranscription factors such as Elk-1, CREB, and many othersstart the transcriptional reprogramming of the cell by turning-on differentiation-related and turning-off proliferation-related
genes (van Dijkmans et al. 2008). A critical regulatory pre-requisite step for NGF-induced differentiation of PC12 cells isthe arrest of the cell cycle, as measured by cessation of DNAsynthesis and cell division (Greene and Tischler 1976;Gunning et al. 1981; Ignatius et al. 1985; Buchkovich andZiff 1994). NGF decreases the growth rate of PC12 cells and,in the short term, causes synchronized PC12 cells to accumu-late in the G1 phase of the cell cycle with a decrease in DNAsynthesis. Acute exposure to NGF arrests the PC12 cell pop-ulation in G1 with an increased number in the G2/M phase(Rudkin et al. 1989). Chronic treatment of PC12 cells withNGF promotes terminal differentiation, in which the terminalphenotype resembles that of sympathetic neurons, as inferredfrom the cessation of cell division, increased adherence toextracellular matrix proteins, neurite extension, and shift fromdopamine to norepinephrine synthesis. Although the mecha-nism by which NGF arrests PC12 cells in the G1 phase of thecell cycle is not yet fully understood, there seems to be a directcorrelation between the differential regulation of specific cellcycle regulatory components and NGF-induced neuronal dif-ferentiation (Hughes et al. 2000). Furthermore, NGF seems tomediate down-regulation of EGFR during the differentiationof PC12 cells therefore making the cells refractory to EGFmitogen signaling (Lazarovici et al. 1987, 1997a). The mo-lecular mechanism responsible for this process is addressed inthe present review, hopefully helping to understand in moredetail the role of down-regulation of EGFR in neuronaldifferentiation.
The Process of NGF-induced HeterologousDown-regulation of EGFR in PC12 Cells
The capacity of NGF-differentiated PC12 cells to interact with125I-EGF (Fig. 1a, b) was correlated to the NGF-inducedneuronal differentiated as inferred from changes in cell mor-phology (Fig. 1a-inset). Treatment of the cells with 50 ng/mlNGF significantly reduced the binding capacity of the cells forEGF after a latency period of 1–2 days. After 5–7 days ofNGF treatment, a near-total disappearance of EGF-bindingcapacity was observed (Fig. 1a). The decrease in EGF bindingcapacity correlated well with the NGF-induced morphologicalchanges, e.g. cell hypertrophy, neurite outgrowth formation,and elongation (Fig. 1a-inset), properties typical of terminallydifferentiated neurons (Fujita et al. 1989). Reduction in thebinding of 125I-EGF to NGF-differentiated PC12 cells may bedue to either reduction in the affinity or in the number ofEGFR. To distinguish between these two possibilities, bindingexperiments were performed with both undifferentiated andNGF-differentiated PC12 cultures and the data was evaluatedby Scatchard analysis (Fig. 1b). While the slope of the curves(affinity) did not change, a 50 and 95 % reduction in thenumber of binding sites was measured after 3 and 6 days of
J Mol Neurosci
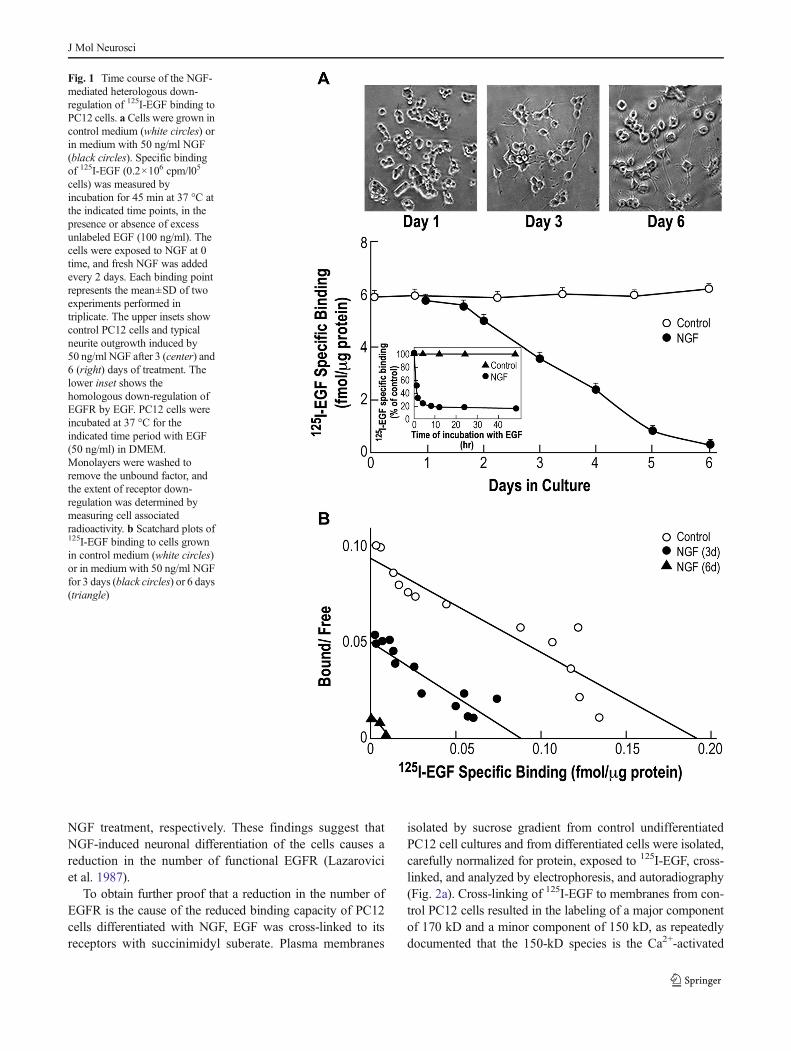
NGF treatment, respectively. These findings suggest thatNGF-induced neuronal differentiation of the cells causes areduction in the number of functional EGFR (Lazaroviciet al. 1987).
To obtain further proof that a reduction in the number ofEGFR is the cause of the reduced binding capacity of PC12cells differentiated with NGF, EGF was cross-linked to itsreceptors with succinimidyl suberate. Plasma membranes
isolated by sucrose gradient from control undifferentiatedPC12 cell cultures and from differentiated cells were isolated,carefully normalized for protein, exposed to 125I-EGF, cross-linked, and analyzed by electrophoresis, and autoradiography(Fig. 2a). Cross-linking of 125I-EGF to membranes from con-trol PC12 cells resulted in the labeling of a major componentof 170 kD and a minor component of 150 kD, as repeatedlydocumented that the 150-kD species is the Ca2+-activated
Fig. 1 Time course of the NGF-mediated heterologous down-regulation of 125I-EGF binding toPC12 cells. a Cells were grown incontrol medium (white circles) orin medium with 50 ng/ml NGF(black circles). Specific bindingof 125I-EGF (0.2×106 cpm/l05
cells) was measured byincubation for 45 min at 37 °C atthe indicated time points, in thepresence or absence of excessunlabeled EGF (100 ng/ml). Thecells were exposed to NGF at 0time, and fresh NGF was addedevery 2 days. Each binding pointrepresents the mean±SD of twoexperiments performed intriplicate. The upper insets showcontrol PC12 cells and typicalneurite outgrowth induced by50 ng/ml NGF after 3 (center) and6 (right) days of treatment. Thelower inset shows thehomologous down-regulation ofEGFR by EGF. PC12 cells wereincubated at 37 °C for theindicated time period with EGF(50 ng/ml) in DMEM.Monolayers were washed toremove the unbound factor, andthe extent of receptor down-regulation was determined bymeasuring cell associatedradioactivity. b Scatchard plots of125I-EGF binding to cells grownin control medium (white circles)or in medium with 50 ng/ml NGFfor 3 days (black circles) or 6 days(triangle)
J Mol Neurosci

protein product of the 170-kD EGFR (Seger et al. 1988).Analysis of the cross-linked EGFR bands in PC12 cells treat-ed for 6 days with NGF indicate complete disappearance ofthe labeling of these bands. This decrease in labeling is con-sistent with a lower number of EGFR in NGF-differentiatedPC12 cells. Another indication for an NGF-induced decreasein the number of EGFR would be a decrease in EGF-stimulated tyrosine kinase activity in differentiated PC12cells. As seen in a typical experiment (Fig. 2b), plasma mem-branes from PC12 cells treated for 6 days with NGF show a95 % reduction in the autophosphorylation intensity of the170-kD band, when compared with membranes from controlundifferentiated cells. A possible mechanism by which NGFmight decrease EGFR number is by decreased synthesis. Thebiosynthesis of the EGFR was studied by labeling control andNGF-differentiated PC12 cells with 35S-methionine for 18 h(Lazarovici et al. 1987). The radioactive cell membranes weresolubilized with detergent, and the EGFR was enriched byimmunoprecipitation with anti-EGFR antibody (Fig. 2c). Thelevels of 35S-methionine-labeled EGFR in the membranes ofNGF-differentiated cells decreased by more than 90 % bycomparison to membranes isolated from control, untreatedcells (Fig. 2c). This result is in accordance with the disappear-ance of autophosphorylated receptor (Fig. 2b). We concludethat the decrease in EGF-binding capacity of the NGF-differentiated PC12 cells (Fig. 1) is due to a reduction in thenumber of cell surface (plasma membrane) receptors, which,in turn, is caused by decreased receptor biosynthesis (Fig. 2).Thus, the NGF-induced loss of EGFR from PC12 cells fulfillsthe criteria for “heterologous down-regulation” (Sibley andLefkowitz 1985).
The process of NGF-induced heterologous down-regulationof EGFR can be visualized by a novel bio-imagingmethod thatuses EGF labeled with a near-infrared dye (EGF-NIR). TheEGF molecule can be conjugated with IRDye800CW at theamino-terminal of the protein due to a free amino groupavailable (Cohen et al. 2012). After reaction with theIRDye800CW, the EGF-NIR was purified by gradient HPLC
chromatography (Fig. 3a), yielding a homogenous single peakas confirmed by SDS-PAGE electrophoresis and NIR visuali-zation (Fig. 3a-inset). EGF-NIR specifically and selectivelybound to EGFR-expressing A431 cells (Fig. 3b). The intensityof the EGF-NIR signal reflected EGFR levels (Cohen et al.2012, 2013; Cohen and Lazarovici 2013a). Using NGF-differentiated PC12 cultures, we demonstrated a significant85 % reduction in EGF-NIR specific binding to the differenti-ated cells (Fig. 3c), indicative of a significant heterologousdown-regulation of EGFR measured by optical imaging(Cohen and Lazarovici 2013b).
In view of the above observation that the NGF-inducedheterologous down-regulation of EGFR is temporally corre-lated with NGF-induced neurite outgrowth, we cultured in thepresence or absence of 50 ng/ml NGF, enhanced green fluo-rescence protein (GFP) expressing PC12 cells for 7 days eitherin conventional two-dimensional (2D) cultures, PC12 cellsembedded in a three-dimensional collagen hydrogel (3D cul-tures) (Arien-Zakay et al. 2009) and PC12 cells grown assuspension cultures (Lazarovici et al. 1997a). Under 3D con-ditions using collagen hydrogels as scaffolds, GFP-PC12 cellsproliferated, formed aggregates, and respond to NGF by exu-berant outgrowth of neurites of different lengths and complex-ity, very similar to wild-type PC12 cells (Arien-Zakay et al.2009). In 2D and 3D cultures, the 50 ng/ml NGF-treated PC12cells extend neurites, and in suspension, they aggregate but donot grow neurites (Fig. 4). Following 7 days of culture underthe various conditions mentioned above, the cells were har-vested and analyzed by Western blotting using an anti-EGFRantibody to measure EGFR down-regulation and an anti-neurofilament-M antibody (NF-M) to measure the increasedexpression of this cytoskeleton protein as marker for theneuronal differentiation of the cells. As seen in Fig. 4, NGF-induced heterologous down-regulation of EGFR under allthree different growth conditions occurred independently ofneurite outgrowth, which was well developed under 2D and3D conditions but absent in cells growing in suspensionculture. PC12 cells grown in suspension in a spinner flask
Fig. 2 Cross linking of 125I-EGF (a), tyrosine autophosphorylation (b)and 35S-methionine incorporation (c) into EGFR in control PC12 cellsand cells after 6 days of differentiation with 50 ng/ml NGF. aMembranesof PC12 cells and A431 cells were incubated for 60 min with 5 nM 125I-EGF on ice in the presence of 0.5 μM disuccinimidyl suberate. b Mem-branes of PC12 cells were incubated for 15 min at 40 °C in a kinasereaction buffer and thereafter incubated for 2 min at 4 °C with 2 μCi of
γ-32P-ATP 50μMunlabeledATP. c PC12 cells were incubated for 18 h inmethionine-free DMEM containing 60 μCi/ml L-35S-methionine, andplasma membranes were prepared. In all experiments, the membraneswere solubilized with RIPA buffer, and EGRRwere immune-precipitatedand analyzed by 7.5 % SDS-PAGE. The autoradiograms of the fixeddried gels are presented. A431 carcinoma cells were used as control.EGFR is represented by the band of 170 kD
J Mol Neurosci

aggregated into macroscopic organoids as evident bymorpho-logical and histological examinations (Fig. 4), as previouslydocumented using different types of bioreactors (Manley andLelkes 2006). In contrast, under all conditions, NGF increasedthe level of the NF-M (Fig. 4) and neuronal tubulin andmicrotubule-associated proteins (data not shown), indicatingNGF-triggered cytoskeleton differentiation under all growthconditions. These findings suggest that NGF-induced heterol-ogous down-regulation of EGFR is a molecular event accom-panying neuronal differentiation and that this effect is inde-pendent of PC12 cell adhesion and the organization of theextracellular matrix (2D vs 3D) and can occur in the absenceof neurite outgrowth (Lazarovici et al. 1997a).
NGF-induced Heterologous Down-regulation of EGFRis TrkA and Ras Dependent
Wild-type (wt) PC12 cells express two types of receptors forNGF: TrkA and p75NTR. In PC12-nnr5 cells, a PC12 variantthat expresses p75NTR but very little TrkA, NGF does notinduce neurite outgrowth (Fig. 5). When human TrkA wasoverexpressed in PC12 cells generating PC12-6.24 stabletransfectants (Katzir et al. 2002) or transfected into PC12-nnr5 cells, generating the PC12-nnr5-TrkA-61 stabletransfectant clone (Lazarovici et al. 1997a), NGF-inducedneurite outgrowth was accelerated (Fig. 5). To determine ifNGF-induced heterologous down-regulation of EGFR is me-diated by TrkA receptors, we assessed byWestern blotting thelevels of EGFR in these several PC12 cell variants upontreatment with NGF. In PC12-nnr5 cells, the level of EGFRwas not altered, even after 7 days of NGF treatment (Fig. 5).By contrast, clear indications for EGFR down-regulation wereseen in parental PC12wt cells as well as in the TrkA overex-pressing PC12-6.24 cells and PC12-nnr5-TrkA-61 cells.Therefore, it seems likely that NGF-induced heterologousdown-regulation of EGFR is mediated by TrkA. Support forthis suggestion was also provided by experiments withK-252a (Fig. 6). K-252a, a relative specific inhibitor ofNGF-induced TrkA tyrosine kinase activity in whole PC12cells (Lazarovici et al. 1997b), prevented the down-regulationof EGFR by NGF in PC12wt cells (Fig. 6a) and PC12 TrkA-overexpressing cells (Lazarovici et al. 1997a). Similarly, aneutralizing antibody of TrkA efficiently inhibited NGF-induced heterologous down-regulation of EGFR (Fig. 6b).Thx-B derivatives, a small non-peptidic antagonists ofp75NTR (Bai et al. 2010) did not affected NGF-induced heter-ologous down-regulation of EGFR (Fig. 6c).
To test whether NGF-induced down-regulation of EGFR isdependent upon Ras activation, we used a PC12 cell variantstably overexpressing the dominant-negative mutant RasN17(PC12-M-M17-26). Expression of the dominant-negative Rasprotein is increased in these cells by treatment with
Fig. 3 Preparation and isolation of EGF-NIR probe and its use forvisualization of NGF-induced heterologous down-regulation of theEGFR in PC12 cells. a HPLC separation of EGF-NIR. Solid line repre-sents absorbance at 226 nm and dotted line indicates the acetonitrilegradient. Inset shows 12 % SDS-PAGE analysis of 10 μg of EGF-NIRscanned with Odyssey near-infrared scanner and of unmodified EGFstained with Coomassie blue. b The specificity and selectivity ofEGF-NIR probe binding measured by NIR imaging. A431 carci-noma cells were incubated for 15 min at 37 °C with 7 ng/mlEGF-NIR in the presence (EGF/EGF-NIR) or absence (EGF-NIR)of 100 ng/ml EGF. Competition experiments with 500 ng/mlNeuroregulin 1 (NRG1/EGF-NIR) were also conducted. In controlexperiments (NIR-Dye), the cultures were incubated with 7 ng/mlNIR-Dye to evaluate non-specific labeling of the cells by com-parison to untreated cultures (untreated). The NIR intensity at800 nm was estimated under identical conditions for all cultures,and the mean±SD (n=9) is presented. Upper insets, NIR scans;lower insets, phase-contrast photomicrographs of the cultures,*p<0.05 versus NIR-Dye; **p<0.05 versus EGF-NIR. c Charac-terization of EGF-NIR binding in control and NGF-differentiatedPC12 cells measured by NIR imaging. Untreated PC12 cells andPC12 cells treated for 7 days with NGF (50 ng/ml) were treatedwith 7 ng/ml of EGF-NIR in the presence (gray bars) or absence(black bars) of 100 ng/ml unlabeled EGF. The NIR intensity at800 nm was estimated under identical conditions for all cultures,and the mean±SD (n=9) is presented. Insets, NIR scans
J Mol Neurosci

dexamethasone (Lazarovici et al. 1997a). Under these condi-tions, NGF-induced heterologous down-regulation of EGFRis completely blocked (Fig. 5 outermost right column).
Taken together, the lack of NGF-induced heterologousdown-regulation of EGFR in PC12-nnr5 cells, the partialreconstitution of the heterologous down-regulation in PC12-
Fig. 4 The level of EGFR in control and NGF-treated PC12 cultures in2D, 3D, and suspension growing conditions. Top: The cultures weregrown in regular 2D conditions (control) in which, upon treatment for7 days with 50 ng/ml NGF (NGF), the cells form a well-differentiatedculture; Culturing under 3D conditions using sandwiched 3D type Icollagen gels (1.5 mg/ml) generated clusters of cells (control) which,upon addition of 50 ng/ml NGF for 7 days, extended long neurites
(NGF). In suspension for 7 days, PC12 cells generated small aggregates(control). Treatment with 50 ng/ml NGF for 7 days generated macro-scopic organoids composed of many cells which failed to extend neurites(NGF).Bottom: Western blotting analysis of EGFR and neurofilament M(NF-M) cytoskeletal marker in PC12 cell cultures under the conditionsdescribed above
Fig. 5 NGF-induced heterologous down-regulation of EGFR in wild-type PC12 (PC12wt), TrkA overexpressor PC12 (PC12-6.24), TrkAdeficient PC12 (PC12-nnr5), TrkA-transfected PC12-nnr5 (PC12-nnr5-TrkA-61 clone) and Ras-deficient PC12 (PC12-M-M17-26) cells. Top:
Western blotting analysis of EGFR; control-untreated cells; NGF-culturestreated for 6 days with 50 ng/ml NGF. Bottom: Phase contrast micro-graphs (×320 original magnification) of the cultures left untreated (con-trol) or treated for 6 days with 50 ng/ml NGF (NGF)
J Mol Neurosci

nnr5-TrkA-61 transfectants, the ability of K-252a and a neu-tralizing antibody to TrkA to block NGF-induced EGFRdown-regulation and the lack of effect of Thx-B support theconcept that this effect is mainly mediated by the TrkA NGFreceptor but not by p75NTR. This conclusion is also supportedby the ability of the Ras dominant-negative protein to blockthis cellular process, since the Ras-Erk signal transductionpathway is initiated by binding of NGF to TrkA receptorsbut not by binding of NGF to p75NTR (Vaudry et al. 2002).This data is consistent with the signal transductionmechanismof TrkA receptors and strongly suggest the involvement of aseries of NGF-induced, neurite outgrowth-independent,TrkA-Ras phosphorylation events in the heterologous down-regulation of EGFR in PC12 cells.
NGF-induced Heterologous Down-regulation of EGFR isTranscriptional Regulated
Figure 7 shows the levels of EGFR messenger RNA (mRNA)in PC12wt cells during NGF-induced differentiation as deter-mined independently both by Northern blotting analysis(Fig. 7a) and by competitive RT-PCR (Fig. 7b). The timepoints are similar to those presented in Fig. 1a. Both ap-proaches indicate a progressive reduction in EGFR mRNAlevels during NGF treatment. In focusing on the major(9.6 kb) EGFR encoding mRNA species, Northern blottinganalysis (Fig. 7a) show that this mRNA species decreased inthe course of NGF treatment and that this decrease precededthe reduction in the protein (Fig. 1a). Competitive RT-PCR, aquantitative procedure for analyzing mRNA levels that ismore sensitive than RNA blot techniques, such as Northernblotting, which provide only semi-quantitative results, shows
a very similar decrease in EGFR mRNA (Shibutani et al.1998). The region selected as a target fragment for PCR
Fig. 6 Time course of the NGF-mediated heterologous down-regulationof 125I-EGF binding to PC12 cells in the presence or absence of neutral-izing anti-TrkA antibody. a Cells were grown in control medium (blacktriangles) or in mediumwith 50 ng/ml NGF (circles) or NGF and 1μg/mlanti-TrkA antibody (white triangles). Specific binding of 125I-EGF (0.2×106 cpm/105 cells) was measured by incubation for 45 min at 37 °C at theindicated time points, in the presence or absence of excess unlabeled100 ng/ml EGF or 1 μg/ml anti-TrkA antibody. The cells were exposed toNGF at 0 time, and fresh NGF was added every 2 days. Each bindingpoint represents the mean±SD of two experiments performed in tripli-cate. b Partial reduction in NGF-induced down-regulation of EGFR in
cultures treated with anti-TrkA, in the presence or absence of NGF-induced differentiation. The level of EGFR was estimated by Westernblotting. PBS-treated cells; anti-TrkA—daily treatment of the culture with1 μg/ml anti-TrkA antibody for 6 days. NGF-differentiated culture for6 days with 50 ng/ml NGF. c The lack of effect of Thx-B, a blocker of theinteraction of NGF with p75NTR receptors. A on NGF-induced heterolo-gous down-regulation of EGFR in PC12 cells. Control-untreated cells;NGF-cultures treated for 6 days with 50 ng/ml NGF. DMSO-control;Thx-B-cultures treated with 10 μMThx-B, a blocker of the interaction ofNGF with p75NTR receptors
Fig. 7 Changes in EGFR mRNA levels induced by NGF differentiationof PC12 cells and evaluated by Northern blotting (a) and by RT-PCR (b).aNorthern blots of poly(A+) RNA of PC12 cells untreated or treated with50 ng/ml NGF for the time points indicated. EGFR and GAPDH controlmRNAs were separated and visualized by 1.8 % Nusieve agarose gelstained with ethidium bromide. b Cells were grown in control medium(triangles) or in medium with 2×10−9 M NGF (circles). Values of EGFRmajor mRNA transcript band (9.6 kb) were estimated by competitive RT-PCR, evaluated at each time point and were calculated as percentages ofuntreated control cells (0 h) and represent the means±S.D. of quadrupli-cate experiments. p<0.05 compared to 0 time
J Mol Neurosci

amplification was almost identical to the sequence of theEGFR DNA fragment used as a probe for Northern blotanalysis. The decrease in EGFR mRNA measured by com-petitive RT-PCR began in the first 12 h of NGF treatment andreached 85% reduction after 5 days of NGF treatment, a resultvery similar to that obtained by Northern blot analysis. For theRT-PCR analysis, this reduction became statistically signifi-cant after 1 day of treatment. GAPDH mRNA levels, theinternal control, were constant during NGF treatment(Fig. 7) (Shibutani et al. 1998).
The evidence that this regulation is exerted at the transcrip-tional level seems quite persuasive. There is a decrease inEGFR mRNA levels comparable to the decrease seen in thereceptor protein itself. This decrease was clearly not caused byany difference in mRNA stability (Shibutani et al. 1998).Direct proof was also obtained by nuclear runoff assay inwhich about 55 % reduction of the nascent mRNA for theEGFR was observed as a result of long-term treatment of thecells with 50 ng/ml NGF (Shibutani et al. 1998). Furtherevidence to support the notion of a transcriptional regulationwas provided by transfecting EGFR promoter constructslinked to luciferase into control, undifferentiated PC12 cellsand NGF-differentiated cells. The decreased luciferaseactivity in the NGF-treated cells, together with the directmeasurement of luciferase mRNA to confirm that thedecrease was at the transcriptional level, provides conclu-sive proof of the transcriptional regulation of this expres-sion of EGFR in PC12 cells (Shibutani et al. 1998; Liu et al.2000, 2001).
GCF2 is a transcriptional repressor of the EGFR gene,which migrates as a 160-kD protein on polyacrylamide gels(Johnson et al. 1992; Reed et al. 1998). The expression levelof this protein in PC12 cells during NGF treatment wasanalyzed by Western blotting. As shown in Fig. 8a, a GSTfusion protein expressed in a bacterial system was recognizedby an antibody raised against GCF2. The level of GCF2 wasclearly increased by up to threefold by NGF-induced celldifferentiation, in a time-dependent manner. To explore therelationship between the increase in GCF2 and the decrease inthe EGFR expression, the NGF-induced increase in GCF2 wasinvestigated for its dependency on Ras. A PC12 cell variantstably overexpressing the dominant-negative mutant RasN17(PC12-M-M17-26) was treated with NGF for 5 days, andGCF2 and EGFR expressions were analyzed by Westernblotting. Figure 8b shows the data from these variants,PC12wt cells as well as from PC12-nnr5 cells. Unlike in thewild type, there was neither an upregulation of GCF2 levelsnor a decrease in EGFR levels in these variant cell lines uponNGF treatment (see also Fig. 5). Of note are the very highbasal levels of GCF2 in the Ras-negative cells. GCF2 wasidentified by differential hybridization and library screeningusing a cDNA for GCF1 (Kageyama and Pastan 1989) andshown to repress transcription from the EGFR promoter
(Beguinot et al. 1995; Rikiyama et al. 2003). While the datapresented here do not prove conclusively that GCF2 is in-volved in the decreased expression of the EGFR in NGF-differentiated PC12 cells, the temporal correlation of the in-creased expression of GCF2 with the decreased expression ofthe EGFR and the fact that both events are dependent on Rasare consistent with that possibility.
EGFR Down-regulation in Tumor Developmentand Therapy
ErbB, EGFR receptor family provides a highly conservedsignaling network that regulates the proliferation of eukaryoticcells. Therefore, dysregulation of EGFR signaling by overex-pression, overphosphorylation, or mutational activation maycause tumor development in different tissues (Yarden andSliwkowski 2001). For this reason, EGFR become a highlyvalidated target for anticancer therapies. Two therapeutic ap-proaches are targeting the EGFR pathway: (i) inhibition of theEGFR intracellular tyrosine kinase domain using small mole-cules such as Erlotinib, Gefitinib, etc. and monoclonal anti-bodies such as Cetuximab or Panitumumab (Visentin et al.2010). Even though the clinical results obtained with thesedrugs are acceptable, the emerging resistance to these drugs iscalling for novel approaches for silencing EGFR signaling
Fig. 8 Changes in GCF2 expression induced by NGF treatment of PC12cells and the effect of TrkA and Ras on the NGF-induced heterologousdown-regulation of EGFR and GCF2 levels in PC12 cells. a PC12 cellswere treated with 50 ng/ml of NGF and harvested at the time pointsindicated. Whole cell lysates (160 μg of protein from each sample) weresubjected to 7.5 % SDS-PAGE electrophoresis and immunoblotted withanti-GCF2 antibody. GST-GCF2 is a fusion protein containing GCF2 usedas a positive control. b PC12 wild type (PC12 wt), TrkA deficient PC12(PC12-nnr5), and Ras-deficient PC12 (PC12-M-M17-26) cells were leftuntreated (control) or treated with 50 ng/ml of NGF (NGF) for 5 days.Whole cell lysates were analyzed by Western blotting using 0.6 μg/mlanti-EGFR antibody and 0.1 μg/ml anti-GCF2 antibody followed bysecondary antibody conjugated to horseradish peroxidase
J Mol Neurosci

activity. It has been reported that treatment with combinationof non-competitive monoclonal antibodies can induce EGFRclustering leading to its down-regulation by molecular mech-anisms involving inhibition of recycling (Spangler et al.2010). Another potential mechanism of dysregulation ofEGFR signaling may involve modulation of its degradation(Kirisits et al. 2007). This aspect has been disregarded so far asa therapeutic approach in modulating EGFR signaling intumors (Grandal and Madshus 2008; Roepstorff et al. 2008).A better understanding of EGFR down-regulation may alsoenhance our diagnostic knowledge and may provide a newinsight in ongoing rational designs of EGFR-targetedtherapeutics.
Interestingly, an earlier study performed with PC12 cellshas shown that NGF can downregulate ErbB2 receptor, an-other member of ErbB receptor family, which is very impor-tant in tumor development (Yarden 2001). Several studieshave reported the expression of NGF and other neurotrophinsin the stroma of and presence of its receptors TrkA and/or p75in the epithelial cells of tumors (Pflug et al. 1995). Thesefindings support the concept of a paracrine regulation in thehuman prostate, breast, thyroid, salivary gland via a stromalNGF-epithelial TrkA receptor interaction (Hondermarck2012; Sortino et al. 2000; Davidson et al. 2001) leading totumor development. The possibility that this paracrine regu-lation may induce EGFR heterologous down-regulation wasnot yet addressed. This hypothesis deserves further investiga-tion and may enhance our understanding on the cross talkbetween NGF and EGFR signaling activity in tumor develop-ment and therapy.
Conclusions
Both acute (minutes to hours) and chronic (days) heterologousdown-regulation of EGFR has been demonstrated in manycell types. This regulation process is characterized by a changein either the affinity or the number of EGFR. For example, adecreased affinity of EGFR was measured upon treatment ofskeletal muscle cells, mouse fibroblasts, SV40-transformedmurine cells with platelet-derived growth factor (Olwin andHauschka 1988), vasopressin (Rozengurt et al. 1981), inter-leukin1 (Bird and Saklatvala 1990), and phorbol esters(Shoyab et al. 1979 ). A decrease in the number of EGFRwas observed in response to thyroid hormone (Mukku 1984),norepinephrine (Cruise et al. 1986), or after infection withadenovirus (Boulukos and Ziff 1993) or Rous sarcoma virus(Wasilenko et al. 1987). The cellular signaling pathways uti-lized by G protein-coupled receptors, tyrosine kinase recep-tors, viruses, or drugs to regulate EGFR involved phosphory-lations on different serine and threonine residues in the intra-cellular domains of these receptors. Therefore, kinase activity-related mechanisms have been proposed for the acute NGF-
induced down-regulation of EGFR in PC12 cells (Brown andCarpenter 1991; Mothe et al. 1993). On the other hand, thesignaling pathway(s) utilized by NGF for the chronic down-regulation of EGFR has not yet been resolved. The chronic,NGF-induced down-regulation of EGFR in PC12 cells isdefinitely different in terms of mechanism from the acutedown-regulation. The chronic, NGF-induced down-regulationof EGFR in PC12 cells is characterized by a progressivedecrease in the number of EGFR, is a transcriptional-regulated event, selectively induced by NGF-TrkA receptorsand mediated by the Ras-MAPK phosphorylation pathway(Fig. 9). The regulation of EGFR expression in normal and
Fig. 9 Mechanistic scheme of NGF-induced heterologous down-regula-tion of EGFR in PC12 cells. Occupancy of TrkA receptors by NGFresults in the stimulation of Ras and B-RAF, which, in turn, stimulatesthe MAPK cascade resulting in the activation of Erk 1/2. Erk 1/2 phos-phorylates in the cytosol or upon its translocation into the nucleus thetranscription factor GCF2. Phosphorylated GCF2 (GCF2-Pi) binds to GCmotif in the EGFR gene promoter and suppresses its transcription. As aresult, the level of mRNA for EGFR is decreased, fewer EGFR aresynthesized and leads to the down-regulation of EGFR. Other potentialsuppressors and activators of EGFR gene promoter, which potentiallymay be modulated by NGF, are also indicated
J Mol Neurosci

tumor cells is complex, involving at least five stimulatorytranscription factors, Sp1 (Kageyama et al. 1988a), ETF(Kageyama et al. 1989), TCF (Johnson et al. 1988), RPF-1(Hudson et al. 1990), and p53 (Deb et al. 1994; Ludes-Meyerset al. 1996; Hughes et al. 2000), and four inhibitory transcrip-tion factors, GCF1 (Kageyama and Pastan 1989; Kageyamaet al. 1988b), GCF2, ETR (Hou et al. 1994), and Wilms tumorfactor, WT1 (Englert et al. 1995). In our hands, NGF-inducedheterologous down-regulation of EGFR was temporally andconsistently correlated with the upregulation of the GCF2protein suggesting that NGF-induced phosphorylation of Erk1/2 may phosphorylate, either in the cytosol or in the nucleus(Groot et al. 2000), the GCF2 protein(s) which will lead to theactivation of its EGFR promoter suppression activity,resulting, in turn, in the reduction in EGFR mRNA levelsand, eventually, EGFR protein expression (Fig. 9).
Clarification of the molecular mechanisms involved inNGF-induced heterologous down-regulation of EGFR willhelp us to understand the regulation of proliferation anddifferentiation in PC12 cells and in neuronal stem cells(Ayuso-Sacido et al. 2010; Doetsch et al. 2002). Because theexpression of EGFR family members are involved in thecontrol of the growth of a large variety of tumors, the inves-tigation of NGF-induced heterologous down-regulation ofEGFR in tumors (Castellon and Mirkin 2003) would alsoseem to be worth pursuing.
Acknowledgments PL holds the Jacob Gitlin Chair in Physiology andis affiliated with and supported by the David R. Bloom Center forPharmacy and the Dr. Adolf and Klara Brettler Center for Research inMolecular Pharmacology and Therapeutics at the Hebrew University ofJerusalem, Israel. PIL is the Laura H Carnell Professor of Bioengineering,Temple University College of Engineering, and is grateful for support bygrants (to PIL) from the Craig H. Neilson Foundation and the Louis andBessie Stein Foundation. GC acknowledges an “Eshkol” fellowship fromthe Israeli Ministry of Science Technology and Space, Israel. This reviewis dedicated to Prof. Uriel Littauer for his scientific contribution toneuroscience, kind friendship, and humble humanity. We are grateful toDr. Davidit Shavit, Dr. Itzhak Katzir, Dr. Gordon Guroff, Dr. MakotoShibutani, Dr. Yasuhiro Katagiri, Dr. Xu-Wen Lin for the excellentcollaboration in performing the studies reviewed and Ms. Zehava Cohenfor the help with the art work.
References
Abe K, Saito H (2001) Effects of basic fibroblast growth factor on centralnervous system functions. Pharmacol Res 43:307–312
Abe Y, Namba H, Zheng Y, Nawa H (2009) In situ hybridization revealsdevelopmental regulation of ErbB1-4 mRNA expression in mousemidbrain: implication of ErbB receptors for dopaminergic neurons.Neuroscience 161:95–110
Arien-Zakay H, Lecht S, Perets A, Roszell B, Lelkes PI, Lazarovici P(2009) Quantitative assessment of neuronal differentiation in three-dimensional collagen gels using enhanced green fluorescence pro-tein expressing PC12 pheochromocytoma cells. J Mol Neurosci 37:225–237
Ayuso-Sacido A, Moliterno JA, Kratovac S et al (2010) Activated EGFRsignaling increases proliferation, survival, and migration and blocksneuronal differentiation in post-natal neural stem cells. J Neurooncol97:323–337
Bai Y, Dergham P, Nedev H et al (2010) Chronic and acute models ofretinal neurodegeneration TrkA activity are neuroprotective whereasp75NTR activity is neurotoxic through a paracrine mechanism. J BiolChem 285:39392–39400
Bath KG, Lee FS (2010) Neurotrophic factor control of adult SVZneurogenesis. Dev Neurobiol 70:339–349
Becker EB, Bonni A (2005) Beyond proliferation-cell cycle control ofneuronal survival and differentiation in the developing mammalianbrain. Semin Cell Dev Biol 16:439–448
Beguinot L, Yamazaki H, Pastan I, Johnson AC (1995) Biochemicalcharacterization of human GCF transcription factor in tumor cells.Cell Growth Differ 6:699–706
Bernd P (2008) The role of neurotrophins during early development.Gene Expr 14:241–250
Bird TA, Saklatvala J (1990) Down-modulation of epidermal growthfactor receptor affinity in fibroblasts treated with interleukin 1 ortumor necrosis factor is associated with phosphorylation at a siteother than threonine 654. J Biol Chem 265:235–240
Boulukos KE, Ziff EB (1993) Adenovirus 5 E1A proteins disrupt theneuronal phenotype and growth factor responsiveness of PC12 cellsby a conserved region 1-dependent mechanism. Oncogene 8:237–248
BrownAB, Carpenter G (1991) Acute regulation of the epidermal growthfactor receptor in response to nerve growth factor. J Neurochem 57:1740–1749
Buchkovich K, Ziff EB (1994) Nerve growth factor regulates the expres-sion and activity of p33cdk2 and p34cdc2 kinases in PC12 pheo-chromocytoma cells. Mol Biol Cell 5:1226–1241
Castellon R, Mirkin BL (2003) Nerve growth factor regulatesneuroectodermal tumor cell responses to mitogenic growth factors.J Neurosci Res 72:239–249
Cohen G, Lazarovici P (2013a) Molecular Imaging of tumor receptor andsignaling molecules using near-infrared nano-approches. In: GovilJN, Sinha S, Navani NK (eds) Nanotechnology: Bioimaging, vol 12.Studium Press LCC, Houston, pp 243–265
Cohen G, Lazarovici P (2013b) Near infrared imaging of EGF receptorsdown-regulation during NGF-induced PC12 cells differentiation.Israel Society for Neuroscience annual meeting. Eilat, Israel
Cohen G, Lecht S, Arien-Zakay H et al (2012) Bio-imaging of colorectalcancer models using near infrared labeled epidermal growth factor.PLoS ONE 7:e48803
Cohen G, Lecht S, Oron-Herman M, Momic T, Nissan A, Lazarovici P(2013) Near infrared optical visualization of epidermal growth factorreceptors levels in COLO205 colorectal cell line, orthotopic tumorin mice and human biopsies. Int J Mol Sci 14:14669–14688
Cruise JL, Coteccina S, Michalopoulos G (1986) Norepinephrine de-creases EGF binding in primary rat hepatocyte cultures. J CellPhysiol 127:39–44
Davidson B, Lazarovici P, Ezersky A et al (2001) Expression levels of thenerve growth factor receptors TrkA and p75 in effusions and solidtumors of serous ovarian carcinoma patients. Clin Cancer Res 7:3457–3464
de la Rosa EJ, de Pablo F (2000) Cell death in early neural development:beyond the neurotrophic theory. Trends Neurosci 23:454–458
Deb SP, Munoz RM, Brown DR, Subler MA, Deb S (1994) Wild-typehuman p53 activates the human epidermal growth factor receptorpromoter. Oncogene 9:1341–1349
Devaskar SU (1991) A review of insulin/insulin-like peptide in the centralnervous system. Adv Exp Med Biol 293:385–396
Doetsch F, Petreanu L, Caille I, Garcia-Verdugo JM, Alvarez-Buylla A(2002) EGF converts transit-amplifying neurogenic precursors inthe adult brain into multipotent stem cells. Neuron 36:1021–1034
J Mol Neurosci

Englert C, Hou X, Maheswaran S et al (1995) WT1 suppresses synthesisof the epidermal growth factor receptor and induces apoptosis.EMBO J 14:4662–4675
Fargali S, Sadahiro M, Jiang C et al (2012) Role of neurotrophins in thedevelopment and function of neural circuits that regulate energyhomeostasis.J. Mol Neurosci 48:654–659
Fujita K, Lazarovici P, Guroff G (1989) Regulation of the differentiationof PC12 pheochromocytoma cells. In: Jaetten AM (ed) Regulationof differentiation in eukaryotic cells: Environmental HealthPerspectives, vol. 80. National Institutes of Health, Bethesda, MD,pp 127–142
Grandal MV, Madshus IH (2008) Epidermal growth factor receptor andcancer: control of oncogenic signalling by endocytosis.J. Cell MolMed 12:1527–1534
Greene LA, Angelastro JM (2005) You can’t go home again: transcrip-tionally driven alteration of cell signaling by NGF. Neurochem Res30:1347–1352
Greene LA, Tischler AS (1976) Establishment of a noradrenergic clonalline of rat adrenal pheochromocytoma cells which respond to nervegrowth factor. Proc Natl Acad Sci U S A 73:2424–2428
Groot M, Boxer LM, Thiel G (2000) Nerve growth factor- and epidermalgrowth factor-regulated gene transcription in PC12 pheochromocy-toma and INS-1 insulinoma cells. Eur J Cell Biol 79:924–935
Gunning PW, Landreth GE, Bothwell MA, Shooter EM (1981)Differential and synergistic actions of nerve growth factor and cyclicAMP in PC12 cells. J Cell Biol 89:240–245
Hall BK, Ekanayake S (1991) Effects of growth factors on the differen-tiation of neural crest cells and neural crest cell-derivatives. Int J DevBiol 35:367–387
Hondermarck H (2012) Neurotrophins and their receptors in breast can-cer. Cytokine Growth Factor Rev 23:357–365
Hou X, Johnson AC, Rosner MR (1994) Induction of epidermal growthfactor receptor gene transcription by transforming growth factor beta1: association with loss of protein binding to a negative regulatoryelement. Cell Growth Differ 5:801–809
Hudson LG, Thompson KL, Xu J, Gill GN (1990) Identificationand characterization of a regulated promoter element in theepidermal growth factor receptor gene. Proc Natl Acad SciUSA 87:7536–7540
Hughes AL, Gollapudi L, Sladek TL, Neet KE (2000)Mediation of nervegrowth factor-driven cell cycle arrest in pc12 cells by p53: simulta-neous differentiation and proliferation subsequent to p53 functionalinactivation. J Biol Chem 275:37829–37837
Ignatius MJ, Chandler CR, Shooter EM (1985) Nerve growth factor-treated, neurite-bearing PC12 cells continue to synthesize DNA. JNeurosci 5:343–351
Ip NY, Boulton TG, Li Yet al (1994) CNTF, FGF, and NGF collaborate todrive the terminal differentiation of MAH cells into postmitoticneurons. Neuron 13:443–455
Johnson MH (2001) Functional brain development in humans. Nat RevNeurosci 2:475–483
Johnson AC, Jinno Y, Merlino GT (1988) Modulation of epidermalgrowth factor receptor proto-oncogene transcription by a promotersite sensitive to S1 nuclease. Mol Cell Biol 8:4174–4184
Johnson AC, Kageyama R, Popescu NC, Pastan I (1992) Expression andchromosomal localization of the gene for the human transcriptionalrepressor GCF. J Biol Chem 267:1689–1694
Kageyama R, Pastan I (1989) Molecular cloning and characterization of ahuman DNA binding factor that represses transcription. Cell 59:815–825
Kageyama R, Merlino GT, Pastan I (1988a) Epidermal growth factor(EGF) receptor gene transcription. Requirement for Sp1 and an EGFreceptor-specific factor. J Biol Chem 263:6329–6336
Kageyama R, Merlino GT, Pastan I (1988b) A transcription factor activeon the epidermal growth factor receptor gene. Proc Natl Acad Sci US A 85:5016–5120
Kageyama R, Merlino GT, Pastan I (1989) Nuclear factor ETF specifi-cally stimulates transcription from promoters without a TATA box. JBiol Chem 264:15508–15514
Kaplan DR, Miller FD (2000) Neurotrophin signal transduction in thenervous system. Curr Opin Neurobiol 10:381–391
Katzir I, Shani J, Regev K, Shabashov D, Lazarovici P (2002) A quan-titative bioassay for nerve growth factor, using PC12 clones express-ing different levels of trkA receptors. J Mol Neurosci 18:251–264
Kirisits A, Pils D, Krainer M (2007) Epidermal growth factor receptordegradation: an alternative view of oncogenic pathways. Int JBiochem Cell Biol 39:2173–2182
Kornblum HI, Hussain RJ, Bronstein JM, Gall CM, Lee DC, Kim B(1997) Seroogy. Prenatal ontogeny of the epidermal growth factorreceptor and its ligand, transforming growth factor alpha, in the ratbrain. J Comp Neurol 380:243–261
Lazarovici P, Dickens G, Kuzuya H, Guroff G (1987) Long-term, heter-ologous down-regulation of the epidermal growth factor receptor inPC12 cells by nerve growth factor. J Cell Biol 104:1611–1621
Lazarovici P, Oshima M, Shavit D et al (1997a) Down-regulation ofepidermal growth factor receptors by nerve growth factor in PC12cells is p140(Trk)-, Ras-, and Src-dependent. J Biol Chem 272:11026–11034
Lazarovici P, Matsuda Y, Kaplan D, Guroff G (1997b) The protein kinaseinhibitors K-252a and staurophin receptors signal transduction. In:Gutman Y, Lazarovici P (eds) Cellular & Molecular Mechanisms ofToxin Action: Toxins and signal transduction, vol. 1. HarwoodAcademic Publishers, Amsterdam, pp 69–74
Liu DX, Greene LA (2001) Neuronal apoptosis at the G1/S cell cyclecheckpoint. Cell Tissue Res 305:217–228
Liu XW, Katagiri Y, Jiang H et al (2000) Cloning and characterization ofthe promoter region of the rat epidermal growth factor receptor geneand its transcriptional regulation by nerve growth factor in PC12cells. J Biol Chem 275:7280–7288
Liu XW, Gong LJ, Guo LYet al (2001) The Wilms’ tumor gene productWT1 mediates the down-regulation of the rat epidermal growthfactor receptor by nerve growth factor in PC12 cells. J Biol Chem276:5068–5073
Ludes-Meyers JH, Subler MA, Shivakumar CV et al (1996)Transcriptional activation of the human epidermal growth factorreceptor promoter by human p53. Mol Cell Biol 16:6009–6019
Manley P, Lelkes PI (2006) A novel real-time system to monitor cellaggregation and trajectories in rotating wall vessel bioreactors. JBiotechnol 125:416–424
Manni L, Rocco ML, Bianchi P et al (2013) Nerve growth factor: basicstudies and possible therapeutic applications. Growth Factors 31:115–122
Mothe I, Ballotti R, Tartare S, Kowalski-Chauvel A, Van Obberghen E(1993) Cross talk among tyrosine kinase receptors in PC12 cells:desensitization of mitogenic epidermal growth factor receptors bythe neurotrophic factors, nerve growth factor and basic fibroblastgrowth factor. Mol Biol Cell 4:737–746
Mukku VR (1984) Regulation of epidermal growth factor receptor levelsby thyroid hormone. J Biol Chem 259:6543–6547
Mullenbrock S, Shah J, Cooper GM (2011) Global expression analysisidentified a preferentially nerve growth factor-induced transcription-al program regulated by sustained mitogen-activated protein kinase/extracellular signal-regulated kinase (ERK) and AP-1 protein acti-vation during PC12 cell differentiation. J Biol Chem 286:45131–45145
Olwin BB, Hauschka SD (1988) Cell surface fibroblast growth factor andepidermal growth factor receptors are permanently lost during skel-etal muscle terminal differentiation in culture. J Cell Biol 107:761–769
Pflug BR, Dionne C, Kaplan DR, Lynch J, Djakiew D (1995) Expressionof a Trk high affinity nerve growth factor receptor in the humanprostate. Endocrinology 136:262–268
J Mol Neurosci

Piao YS, Iwakura Y, Takei N, Nawa H (2005) Differential distribu-tions of peptides in the epidermal growth factor family andphosphorylation of ErbB 1 receptor in adult rat brain. NeurosciLett 390:21–24
Plata-Salaman CR (1991) Epidermal growth factor and the nervoussystem. Peptides 12:653–663
Ravni A, Bourgault S, Lebon A et al (2006) The neurotrophic effects ofPACAP in PC12 cells: control by multiple transduction pathways. JNeurochem 98:321–3219
Reed AL, Yamazaki H, Kaufman JD, Rubinstein Y, Murphy B, JohnsonAC (1998) Molecular cloning and characterization of a transcriptionregulator with homology to GC-binding factor. J Biol Chem 273:21594–21602
Rikiyama T, Curtis J, Oikawa M et al (2003) GCF2: expression andmolecular analysis of repression. Biochim Biophys Acta 1629:15–25
Roepstorff K, Grovdal L, Grandal M, Lerdrup M, van Deurs B (2008)Endocytic down-regulation of ErbB receptors: mechanisms andrelevance in cancer. Histochem Cell Biol 129:563–578
Rozengurt E, Brown KD, Pettican P (1981) Vasopressin inhibition ofepidermal growth factor binding to cultured mouse cells. J BiolChem 256:716–722
Rudkin BB, Lazarovici P, Levi BZ, Abe Y, Fujita K, Guroff G(1989) Cell cycle-specific action of nerve growth factor inPC12 cells: differentiation without proliferation. EMBO J 8:3319–3325
Seger R, Yarden Y, Kashles O, Goldblatt D, Schlessinger J,Shaltiel S (1988) The epidermal growth factor receptor as asubstrate for a kinase-splitting membranal proteinase. J BiolChem 263:3496–500
Shibutani M, Lazarovici P, Johnson AC, Katagiri Y, Guroff G (1998)Transcriptional down-regulation of epidermal growth factor recep-tors by nerve growth factor treatment of PC12 cells. J Biol Chem273:6878–6884
Shoyab M, DeLarco JE, Todaro J (1979) Biologically active phorbolesters specifically alter affinity of epidermal growth factor mem-brane receptors. Nature 279:387–391
Sibilia M, Steinbach J, Sting PL, Aguzzi A, Wagner EF (1998) A strain-independent postnatal neurodegeneration in mice lacking the EGFreceptor. EMBO J 17:719–731
Sibley DR, Lefkowitz RJ (1985) Molecular mechanisms of receptordesensitization using the 13-adrenergic receptor-coupled adenylatecyclase system as a model. Nature 317:124–129
Sieber-Blum M (1998) Growth factor synergism and antagonism in earlyneural crest development. Biochem Cell Biol 76:1039–1050
Sofroniew MV, Howe CL, Mobley WC (2001) Nerve growth factorsignaling, neuroprotection, and neural repair. Annu Rev Neurosci24:1217–1281
Sortino MA, Condorelli F, Vancheri C et al (2000) Mitogenic effect ofnerve growth factor (NGF) in LNCaP prostate adenocarcinomacells: role of the high- and low-affinity NGF receptors. MolEndocrinol 14:124–136
Spangler JB, Neil JR, Abramovitch S et al (2010) Combination antibodytreatment down-regulates epidermal growth factor receptor byinhibiting endosomal recycling. Proc Natl Acad Sci U S A 107:13252–13257
van Dijkmans TF, Hooijdonk LW, Schouten TG et al (2008) Temporaland functional dynamics of the transcriptome during nerve growthfactor-induced differentiation. J Neurochem 105:2388–2403
Vaudry D, Stork PJ, Lazarovici P, Eiden L (2002) Signaling pathways forPC12 cell differentiation: making the right connections. Science296:1648–1649
Visentin M, Biason P, Toffoli G (2010) Drug interactions among theepidermal growth factor receptor inhibitors, other biologics andcytotoxic agents. Pharmacol Ther 128:82–90
Wang VY, Zoghbi HY (2001) Genetic regulation of cerebellar develop-ment. Nat Rev Neurosci 2:484–491
WasilenkoWJ, Shawrer LR,WeberMJ (1987)Down-modulation of EGFreceptors in cells transformed by the src oncogene.J. Cell Physiol131:450–457
Williamson LL, Bilbo SD (2013) Chemokines and the hippocampus: anew perspective on hippocampal plasticity and vulnerability. BrainBehav Immun 30:186–194
Yarden Y (2001) Biology of HER2 and its importance in breast cancer.Oncology 61:1–13
Yarden Y, Sliwkowski MX (2001) Untangling the ErbB signalling net-work. Nat Rev Mol Cell Biol 2:127–137
Yeh HJ, Ruit KG, Wang YX, Parks WC, Snider WD, Deuel TF (1991)PDGF A-chain gene is expressed by mammalian neurons duringdevelopment and in maturity. Cell 64:209–216
J Mol Neurosci
Related Documents











