Transcriptional Activation of Prion Protein Gene in Growth-Arrested and Differentiated Mouse Erythroleukemia and Human Neoplastic Cells Dimitrios D. Gougoumas, Ioannis S. Vizirianakis, and Asterios S. Tsiftsoglou 1 Laboratory of Pharmacology, Department of Pharmaceutical Sciences, Aristotle University of Thessaloniki, GR-54006 Thessaloniki, Greece The prion protein (PrP C ) is a GPI-anchored sialogly- coprotein that has attracted worldwide attention over the years due to its involvement in the pathogenesis of transmissible spongiform encephalopathies in sheep (scrapie), cattle (BSE), and humans (CJD). To under- stand the precise role of the Prn-p gene in cell growth and differentiation we investigated the expression pattern of the Prn-p gene in proliferating cells and in cells arrested in growth either by confluency or by induction of terminal differentiation. Viral-trans- formed mouse spleen hematopoietic cells named mu- rine erythroleukemia (MEL) and other types of induc- ible cells (human neuroectodermal RD/TE-671, myoid RD cells) were employed. Cells grown exponentially, at confluency, or irreversibly arrested in growth at terminal differentiation state were analyzed by fluo- rescence cell sorting and Northern blot hybridization to estimate the steady-state level of PrP mRNA at dif- ferent phases of the cell cycle. MEL cells that failed to differentiate from treatment with N 6 -methyladenosine (N 6 mAdo), an inhibitor of differentiation, were also analyzed for PrP mRNA level. Our results indicate the following: (a) growth arrest of cells at G 1 phase by confluency or by induction of terminal differentiation led to increased accumulation of PrP mRNA tran- scripts, an event observed also in differentiated MEL, RD/TE-671, and RD cells independent of the inducer used; (b) treatment of MEL cells with N 6 mAdo pre- vented early activation of the Prn-p gene in cells treated with the inducer; and (c) cell-free nuclear run- off studies showed enhanced expression of the Prn-p gene due to transcriptional activation. These findings indicate, for the first time, that the Prn-p gene, which is thought to be a housekeeping gene, is transcription- ally activated in G 1 phase in confluent and terminally differentiated cells. This information may be valuable in understanding the overaccumulation of PrP in some differentiated tissues and may let us repress Prn-p gene activation by novel agents. © 2001 Academic Press Key Words: prion protein; Prn-p gene; growth; differ- entiation; inducers; neoplastic cells; MEL; mRNA; transcripts; gene activation. INTRODUCTION The prion protein PrP C is a sialoglycoprotein bound to the plasma membrane of cells by a glycophosphati- dyl inositol anchor [1]. Abnormal aggregation of a mod- ified isoform of PrP C , termed scrapie prion protein (PrP Sc ), has been implicated in several transmissible spongiform encephalopathies such as scrapie in sheep, BSE in cattle, and Creutzfeldt–Jakob disease in hu- mans [2]. Despite the plethora of information accumu- lated thus far with respect to the physicochemical properties of PrP C , very little is known of the regula- tion of expression of the Prn-p gene, which encodes PrP C in tissues. This question is essential, since the degree of expression of the Prn-p gene regulates the level of PrP C accumulated in the cell and the extent of expression of this gene may be closely related to the developmental or degenerative state of the cells. Al- though existing evidence suggests that PrP C is in- volved in normal synaptic function [3], mitogenic re- sponse of lymphocytes [4], Ca 21 metabolism [5], Cu 21 uptake [6], and oxidative stress [7], the precise biolog- ical role remains unknown. Moreover, differential de- velopmental regulation of the Prn-p gene during brain maturation in rats and hamsters predisposes for a possible role in cell growth and differentiation [8, 9]. PrP is encoded by the third exon of the Prn-p gene, which is located on chromosome 2 in mice [10] and on chromosome 20 in humans [11]. PrP C was first found in neurons of the adult central nervous system [12], but now it is well known that it is expressed in other tissues, including muscle [13] and cells of hematopoietic origin [14]. Evidence suggesting a possible role in transmission of the prion agent through lymphohematopoietic cells [15] prompted us to investigate the expression patterns of the Prn-p gene in hematopoietic progenitor cells, like murine erythro- leukemia (MEL) cells. In a preliminary study reported 1 To whom correspondence and reprint requests should be ad- dressed at the Laboratory of Pharmacology, Department of Pharma- ceutical Sciences, Aristotle University of Thessaloniki, GR-54006 Thessaloniki, Greece. Fax: 0030 31 997618. E-mail: tsif@ pharm.auth.gr. 408 0014-4827/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved. Experimental Cell Research 264, 408 – 417 (2001) doi:10.1006/excr.2001.5151, available online at http://www.idealibrary.com on

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ctt(sapcifriRatrtfd
af
vtogiia
Experimental Cell Research 264, 408–417 (2001)doi:10.1006/excr.2001.5151, available online at http://www.idealibrary.com on
Transcriptional Activation of Prion Protein Gene in Growth-Arrestedand Differentiated Mouse Erythroleukemia and Human Neoplastic Cells
Dimitrios D. Gougoumas, Ioannis S. Vizirianakis, and Asterios S. Tsiftsoglou1
Laboratory of Pharmacology, Department of Pharmaceutical Sciences, Aristotle University of Thessaloniki, GR-54006 Thessaloniki, Greece
et
The prion protein (PrPC) is a GPI-anchored sialogly-oprotein that has attracted worldwide attention overhe years due to its involvement in the pathogenesis ofransmissible spongiform encephalopathies in sheepscrapie), cattle (BSE), and humans (CJD). To under-tand the precise role of the Prn-p gene in cell growthnd differentiation we investigated the expressionattern of the Prn-p gene in proliferating cells and inells arrested in growth either by confluency or bynduction of terminal differentiation. Viral-trans-ormed mouse spleen hematopoietic cells named mu-ine erythroleukemia (MEL) and other types of induc-ble cells (human neuroectodermal RD/TE-671, myoidD cells) were employed. Cells grown exponentially,t confluency, or irreversibly arrested in growth aterminal differentiation state were analyzed by fluo-escence cell sorting and Northern blot hybridizationo estimate the steady-state level of PrP mRNA at dif-erent phases of the cell cycle. MEL cells that failed toifferentiate from treatment with N6-methyladenosine
(N6mAdo), an inhibitor of differentiation, were alsonalyzed for PrP mRNA level. Our results indicate theollowing: (a) growth arrest of cells at G1 phase by
confluency or by induction of terminal differentiationled to increased accumulation of PrP mRNA tran-scripts, an event observed also in differentiated MEL,RD/TE-671, and RD cells independent of the inducerused; (b) treatment of MEL cells with N6mAdo pre-
ented early activation of the Prn-p gene in cellsreated with the inducer; and (c) cell-free nuclear run-ff studies showed enhanced expression of the Prn-pene due to transcriptional activation. These findingsndicate, for the first time, that the Prn-p gene, whichs thought to be a housekeeping gene, is transcription-lly activated in G1 phase in confluent and terminally
differentiated cells. This information may be valuablein understanding the overaccumulation of PrP insome differentiated tissues and may let us repressPrn-p gene activation by novel agents. © 2001 Academic
Press
1 To whom correspondence and reprint requests should be ad-dressed at the Laboratory of Pharmacology, Department of Pharma-ceutical Sciences, Aristotle University of Thessaloniki, GR-54006Thessaloniki, Greece. Fax: 0030 31 997618. E-mail: tsif@
pharm.auth.gr.4080014-4827/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
Key Words: prion protein; Prn-p gene; growth; differ-ntiation; inducers; neoplastic cells; MEL; mRNA;ranscripts; gene activation.
INTRODUCTION
The prion protein PrPC is a sialoglycoprotein boundto the plasma membrane of cells by a glycophosphati-dyl inositol anchor [1]. Abnormal aggregation of a mod-ified isoform of PrPC, termed scrapie prion protein(PrPSc), has been implicated in several transmissiblespongiform encephalopathies such as scrapie in sheep,BSE in cattle, and Creutzfeldt–Jakob disease in hu-mans [2]. Despite the plethora of information accumu-lated thus far with respect to the physicochemicalproperties of PrPC, very little is known of the regula-tion of expression of the Prn-p gene, which encodesPrPC in tissues. This question is essential, since thedegree of expression of the Prn-p gene regulates thelevel of PrPC accumulated in the cell and the extent ofexpression of this gene may be closely related to thedevelopmental or degenerative state of the cells. Al-though existing evidence suggests that PrPC is in-volved in normal synaptic function [3], mitogenic re-sponse of lymphocytes [4], Ca21 metabolism [5], Cu21
uptake [6], and oxidative stress [7], the precise biolog-ical role remains unknown. Moreover, differential de-velopmental regulation of the Prn-p gene during brainmaturation in rats and hamsters predisposes for apossible role in cell growth and differentiation [8, 9].PrP is encoded by the third exon of the Prn-p gene,which is located on chromosome 2 in mice [10] and onchromosome 20 in humans [11].
PrPC was first found in neurons of the adult centralnervous system [12], but now it is well known that it isexpressed in other tissues, including muscle [13] andcells of hematopoietic origin [14]. Evidence suggestinga possible role in transmission of the prion agentthrough lymphohematopoietic cells [15] prompted us toinvestigate the expression patterns of the Prn-p genein hematopoietic progenitor cells, like murine erythro-
leukemia (MEL) cells. In a preliminary study reported
otPsm
(l(F
emipCG
i
aihRar
bg
409Prn-p GENE EXPRESSION IN NEOPLASTIC CELLS
elsewhere [16], we have found that the Prn-p gene isexpressed in MEL cells.
In order to investigate how the Prn-p gene is regu-lated in growing and terminally differentiated cells, weemployed MEL, human neuroectodermal RD/TE-671,and myoid RD cells, all of which can be induced todifferentiate in culture [17–19]. Terminally differenti-ated cells are irreversibly arrested in G1 phase andhave lost their ability to proliferate. In this study, wehave investigated whether growth of MEL cells fromlow density to confluency, as well as differentiation ofMEL, RD/TE-671, and RD cells, causes cell cycle- anddifferentiation-dependent changes in the expression ofthe Prn-p gene. Cells grown in cultures with or withoutthe inducers DMSO and UDP-4 (a member of the ure-ido-derivatives of pyridine) [20] were assessed for dif-ferentiation in culture. In addition, cells were sub-jected to Northern blot hybridization analysis with theuse of mouse and human 32P-labeled DNA PrP probes,in order to measure the level of PrP mRNA accumu-lated at different stages of cell growth and differentia-tion. Cells were examined by fluorescence-activatedcell sorting analysis (FACS) for cell cycling.
Here, we report that confluent and terminally differ-entiated MEL cells mainly arrested at G1 phase of thecell cycle express relatively higher levels of PrP RNAtranscripts in contrast to untreated, nondifferentiated,and exponentially growing cells, in which the Prn-pgene is expressed in hardly detectable levels. Similaraccumulation of Prn-p transcripts was observed inther cell types (RD/TE-671, RD) undergoing differen-iation. Interestingly, the higher expression of thern-p gene in G1-arrested cells is attributed to tran-criptional activation as confirmed by run-off experi-ents. That activation of Prn-p gene expression is
indeed a part of differentiation process was furthersupported by experiments with MEL cells blocked fromdifferentiating by treatment with N6–methyladenosine(N6mAdo). Such treatment did not allow activation ofthe Prn-p gene to occur. Similarly, experiments withMEL subcloned cells resistant to inducers (cells unableto differentiate and undergo growth arrest due to ter-minal differentiation) have revealed that the pattern ofPrn-p gene expression is quite similar to that of controluntreated cells.
MATERIALS AND METHODS
Cell cultures. Cell lines employed throughout this study were: (a)highly inducible murine erythroleukemia cells (MEL-745PC-4), (b) asubclone of DMSO-resistant MEL-745 cells (MEL-8R), (c) humanrhabdomyosarcoma cells (RD), and (d) human neuroectodermal cells(RD/TE-671). All these cell lines were maintained in Dulbecco’s mod-ified Eagle’s medium (DMEM) containing 10% (v/v) fetal calf serum(Gibco, Long Island, NY) and antibiotics (penicillin and streptomycin100 mg/ml). Cells were incubated at 37°C in a humidified atmosphere
containing 5% (v/v) CO2.Chemicals. Dimethyl sulfoxide (DMSO), benzidine dihydrochlo-ride, vanadyl ribonucleotide complexes, and proteinase K were pur-chased from Boehringer Mannheim (Mannheim, Germany). UDP-4,a potent inducer of differentiation, was synthesized in our labora-tory, as described earlier [20]. N6mAdo and diethyl pyrocarbonate(DEPC) were purchased from Sigma (St. Louis, MO). [a-32P]dCTPRedivue, 10 mCi/ml, ;3000 Ci/mmol) and the multiprime DNAabeling system kit (RPN 1600 Y) were obtained from AmershamUK). DNA-Prep Reagents Kit was purchased from Coulter (Miami,L).Probes and primers. A genomic DNA fragment (2.15 kb, EcoRI)
ncoding the mouse Prn-p gene was derived from a recombinant plas-id purchased from ATCC (Cat. No. 63010). PCR products correspond-
ng to the entire open reading frame of the mouse Prn-p gene (senserimer, 59-CCGCTCGAGAAAAGAGAGGCTGAAGCTAAAAAGCGG-CA-39, and antisense primer, 59-GGAATTCTTAGTGGTGGTGGTG-TGGTGGGATCTTCTCCCGTCGTAATA-39), as well as the human
one (sense primer, 59-CCGCTCGAGAAAAGAGAGGCTGAAGCTAAG-AAGCGCCCGAAGCCTGGA-39, and antisense primer, 59-GAATTC-TTAGTGGTGGTGGTGGTGGTGCGATCCTCTCTGGTAATAGGC-39),were synthesized and used as hybridization probes. A cDNA hamsterPrP fragment (974 bp, EcoRI–HindIII) was derived from the PEA 974plasmid [21]. A cDNA encoding the rat GAPDH gene (426 bp, ApaI–PstI) was used as an internal control [22]. Finally, a 224-bp PCRfragment encoding exon 2 of the mouse bmajor globin gene was availablen our laboratory and used as a differentiation marker for MEL cells.
Isolation of total cytoplasmic RNA and Northern blot hybridizationnalysis. Total cytoplasmic RNA was prepared from control andnducer-treated cells at different times during incubation. Cells werearvested from culture by trypsinization (for the monolayer culturesD and RD/TE-671), collected by centrifugation at 150g for 5 min,nd washed three times with ice-cold PBS (pH 7.4). The pellet wasesuspended in 0.2 ml lysis buffer (0.14 M NaCl, 1.5 mM MgCl2, 10
mM Tris–Cl (pH 6.8), 0.1% NP-40, and 10 mM vanadyl ribonucle-otide complexes), vortexed for 10 s, overlaid on an equal volume oflysis buffer containing 24% w/v sucrose and 0.2% w/v NP-40, andcentrifuged at 12,000g for 15 min at 4°C. The supernatant was thencollected, digested with proteinase K (200 mg/ml) for 30 min at 37°C,extracted with phenol/chloroform, precipitated with ice-cold ethanolin the presence of 0.3 M sodium acetate (pH 5.0), dehydrated, andfinally dissolved in DEPC-treated H2O. Equal samples of purifiedRNA (15 mg) were electrophoretically separated on a denatured 1.0%w/v agarose gel in the presence of 2.2 M formaldehyde, transferredonto nylon membranes (Hybond; Amersham), and hybridized withrandom primed 32P-labeled DNA probes (described above). Mem-branes were washed and autoradiographed using Kodak X-Omat ARfilm.
Induction and assessment of differentiation. MEL, RD, and RD/TE-671 cells were exposed to inducers as follows: MEL cells wereincubated with (i) the inducer DMSO, (ii) DMSO in the presence orabsence of N6mAdo (an inhibitor of commitment to differentiation),or (iii) UDP-4. At certain timed intervals as indicated in the text, theproportion of differentiated cells was assessed cytochemically withbenzidine–H2O2 solution, which stains cells producing hemoglobin[23]. RD and RD/TE-671 cells were exposed to 1 mM UDP-4 and thenexamined microscopically for morphological maturation, as de-scribed by Pappas et al. [19].
Fluorescence-activated cell sorting analysis. MEL cells treatedwith or without DMSO (1.5% v/v) were harvested at certain timeintervals as indicated in the text (see Results), washed twice withice-cold PBS, centrifuged at 500g, resuspended in cold “Saline GM”uffer (glucose 1.1 g/L, NaCl 8 g/L, KCl 0.4 g/L, Na2HPO4 z H2O 0.39/L, KH2PO4 0.15 g/L, EDTA 0.5 mM), and fixed with 95% ethanol.
Then, samples were prepared, stained with propidium iodide (DNA-Prep Reagents Kit), and analyzed on an Epics XL-MCL fluorescence
cell sorting apparatus [24].
tNMN8lpd
icqcatta(vdupaiwsdfibt((msw
410 GOUGOUMAS, VIZIRIANAKIS, AND TSIFTSOGLOU
In vitro transcription assay (run-off). Intact nuclei were isolatedand purified according to “Current Protocols in Molecular Biology”[25]. In brief, cells were harvested, centrifuged at 500g, and washedwice with ice-cold PBS. The cell pellet was lysed by addition of 8 mlP-40 lysis buffer (10 mM Tris–HCl, pH 7.4, 10 mM NaCl, 3 mMgCl2, 0.5% v/v NP-40) and centrifuged at 500g at 4°C for 5 min.uclei were stored in 400 ml glycerol buffer (50 mM Tris–HCl, pH.3, 40% v/v glycerol, 5 mM MgCl2, 0.1 mM EDTA) and frozen iniquid nitrogen or processed accordingly. Run-off transcription ex-eriments were performed in a final assay volume of 400 ml, asescribed earlier [25], by using [32P]UTP (10 mCi/ml, ;3000 Ci/
mmol) as precursor and 1 3 108 nuclei per reaction. Then, 32P-labeledRNA was purified and hybridized to 5 mg of PrP, GAPDH, or bmajor
DNA probes immobilized on nitrocellulose filters. Finally the mem-branes were exposed to X-ray film.
RESULTS
The Steady-State Level of PrP mRNA Increasesas MEL Cells Progress from Exponentialto Confluent State
As a first step to explore the biological role of thePrn-p gene in cell growth, we examined whether it isregulated during the cell cycle in growing and conflu-ent cells. We approached this question by performingtwo sets of experiments: First, highly inducible MELcells were grown from low to high cell density andassessed for their capacity to produce PrP RNA tran-scripts by Northern blot hybridization analysis. Sec-ond, MEL cells were kept continuously proliferating bydilution and feeding of the cultures with fresh medium.After 72 h exponential growth, the culture was splitand one portion was diluted and fed with fresh mediumto maintain exponential growth conditions, while theother was kept until cells reached confluency (growtharrest). This experimental design aimed to resolvewhether the Prn-p gene is differentially expressed atvarious states of cell growth, that is, during the expo-nential and the confluent state. After 96 h of incuba-tion, one culture contained mainly cells arrested atstationary phase, while the majority of cells in theother culture were still proliferating, as shown by thegrowth kinetics curves in Fig. 1A. Cell cycle analysis byFACS has indeed shown that cells growing from low tohigh density (at confluency) contained more (39.9%)cells in G1 phase and fewer (20.4%) in S phase, asshown in Figs. 1B–1D. A small number of apoptoticcells was detected after 96 h incubation (14.6%). Underthese conditions, the accumulation of PrP RNA tran-scripts has gradually increased from very low levelseen in proliferating cells to high level in confluentcells (Fig. 2). Knowing that a large portion of the cellsat confluency are growth arrested at G1 phase as men-tioned above, we concluded that the Prn-p gene, whichis hardly expressed in exponentially proliferating cells,gradually produces PrP mRNA at confluent cells (seeFig. 2A). Additional experiments in which confluent
cells were diluted with fresh medium at lower celldensity and converted into exponentially growing cellsconfirmed the activation of the Prn-p gene from a low toa high level. As shown in Fig. 2 (experiment II), con-fluent cells after 120 and 144 h continued to expresshigh levels of PrP mRNA, in contrast to those cells thatwere diluted to low density and failed to accumulatePrP mRNA transcripts upon switch from confluent toexponential phase of growth. These data suggest thatthe expression of the Prn-p gene increases as cellsprogress from the exponential phase of growth to con-fluency and vice versa, that is, Prn-p gene expression isgradually repressed when confluent cells are stimu-lated by fresh medium with FCS to enter the exponen-tial phase of growth. Therefore, activation of the Prn-pgene and accumulation of RNA transcripts are consis-tent with growth arrest at G1 phase of the cell cycle.
The Prn-p Gene Is Accumulated in TerminallyDifferentiated and Growth-Arrested MEL Cells
The observation that expression of the Prn-p gene isncreased when exponentially growing MEL cells be-ome growth arrested prompted us to investigate theuestion whether accumulation of Prn-p transcriptsould also occur in terminally differentiated cells whichre also arrested in growth due to terminal matura-ion. MEL cells were induced to differentiate and ma-ure by treatment with the chemical inducers DMSOnd UDP-4. Cells were treated with the inducer DMSO1.5% v/v) for 24 h. At this time, the culture was di-ided. One portion was diluted (1:10) with fresh me-ium including the inducer, while the other was keptntil cells reached confluency (Fig. 3D) also in theresence of DMSO. Assessment of erythroid differenti-tion in all cultures (diluted with fresh medium or notn the presence of DMSO) indicated that both culturesere grown quite similarly and contained about the
ame proportion of hemoglobin-containing cells (benzi-ine-positive cells) at 72, 96, and 120 h (Fig. 3E). Thisnding indicated that in both cultures the cells hadeen committed already and matured to the same ex-ent. The similar growth pattern observed in both casesFig. 3D), taken together with the differentiation dataFig. 3E), indicates that despite the addition of fresh
edium upon dilution, further growth was not ob-erved. Substantial proportion of DMSO-treated cellsere committed and arrested in G1 phase (36.0%) as
expected, although a lot of cells were still proliferating(S phase, 50.0%) even after 96 h incubation (Figs. 3A,3B, and 3C). This cell-cycle kinetics is attributed toasynchronous cell population, since a portion of cellsneeded time to progress from S to G1 phase in thepresence of DMSO. Under these conditions, the expres-sion of the Prn-p gene was increased in committed cellscompared to control exponentially growing cells that
produced hardly detectable levels of PrP mRNA (Fig.
acadaosh
Pcp
incubation, as the arrow shows, the culture was separated in two
411Prn-p GENE EXPRESSION IN NEOPLASTIC CELLS
4). In particular, the relative steady-state level of PrPmRNA reached a peak value in 48 h in both undilutedand diluted cultures and then declined although itremained at a higher level compared to that of controlcells. This suggests that the Prn-p gene is activated at
critical time when the majority of benzidine-positiveells reach a value of .50% and declines a little there-fter when differentiation progresses at 72 h. If thisecline is attributed to accumulation of apoptotic cellst later times (13.7%) is not known. As far as the levelf GADPH is concerned, this declined only at latetages in terminally differentiated MEL cells, as weave seen many times in our experiments [26].Overall, the findings presented thus far indicate that
rn-p gene expression accumulates in growth-arrestedells, regardless of whether they became confluent orermanently arrested at G1 phase of the cell cycle due
to terminal differentiation.
The Expression Pattern of the Prn-p Gene in MELCells Blocked from Differentiating by N6mAdo andin DMSO-Resistant MEL Cells Is Similar to Thatof Control Untreated Cells
To further demonstrate whether accumulation ofPrn-p transcripts results from growth arrest at G1
phase, as we mentioned above, we applied a reverseapproach. We exposed inducible cells to both DMSOand N6mAdo (an agent that blocks initiation of com-mitment in these cells) and in addition used MEL cellsresistant to differentiation by DMSO.
In MEL cells blocked from differentiating byN6mAdo as shown earlier [27], the expression patternof the Prn-p gene (Fig. 5) was similar to that of controluntreated cells (Fig. 2) and not to that of DMSO-treated cells (Fig. 4). That is, N6mAdo prevented theearly activation of the Prn-p gene by DMSO and failedto do so for the activation of the Prn-p gene observed athigher cell densities (confluency), like in the controluntreated cells reaching confluency. Similarly, whenDMSO-resistant MEL cells were exposed to DMSO(1.5% v/v) and assessed for the steady-state level of PrPmRNA, we observed that such cells gradually accumu-lated increased levels of PrP mRNA only at confluency
parts. One portion was diluted with freshly prepared medium con-taining fetal bovine serum to sustain continuous proliferation, whilethe other remained as it was (undiluted). These two cultures exhib-ited markedly different rates of growth. Cells in the undiluted cul-ture reached confluency after 96 to 120 h as expected, while cells inthe diluted culture continued to grow. (B, C, and D) Cells incubatedand grown, as described for A, were analyzed by FACS analysis (seedetails under Materials and Methods) with the use of propidiumiodide after 0, 48, and 96 h of incubation. Proportion of cells invarious cell cycle phases (G1, G2, S, and A, proportion of apoptotic
FIG. 1. Growth kinetics and FACS analysis of cells growing fromexponential to confluent state. (A) MEL cells were seeded (1 3 105
cells/ml) in DMEM containing 10% FCS and antibiotics. After 72 h
cells) is depicted in the insets.
MMaulPemi
aAttpe
RN
412 GOUGOUMAS, VIZIRIANAKIS, AND TSIFTSOGLOU
(after 72 h in culture) and not during the exponentialphase of growth (Fig. 6). Upon dilution, DMSO-resis-tant cells continued to grow exponentially in the pres-ence of DMSO, but failed to produce substantial levelsof PrP mRNA, in contrast to DMSO-sensitive MELcells (Figs. 3 and 4). Therefore, we conclude that accu-mulation of PrP RNA transcripts occurs in fact duringcell cycle arrest, regardless of whether this resultsfrom confluency, or irreversible maturation, or evenloss of cell proliferation.
Differentiation of MEL, RD, and RD/TE-671 Cellswith UDP-4 Leads to Increased Expressionof the Prn-p Gene
To rule out the possibility that the accumulation ofPrn-p transcripts is not specific to the inducer, DMSO,or to the cell line employed (MEL cells) in the previousexperiments and that it rather represents a more gen-eral phenomenon, we exposed MEL and other two celllines of human origin (RD/TE-671, RD) to UDP-4 (an-other potent inducer of differentiation) for 48 and 96 h.Under these conditions, these cells were terminallydifferentiated as shown earlier by our laboratory [19,20]. When such UDP-4-treated cells were assessed forthe level of PrP mRNA by Northern blot hybridization
FIG. 2. Accumulation of PrP RNA transcripts in MEL cells growintervals, total cytoplasmic RNA was purified from MEL cells and andescribed under Material and Methods. GAPDH and 28S rRNA werestaining the membrane with methylene blue as indicated by an arrowin cells growing from exponential to confluent state (undiluted cultuthe diluted culture (Fig. 1A, Œ) by the symbol (1) after 96, 120, andmounts of moPrP and/or GAPDH RNA transcripts compared to thatnalyzer (version 3.0, Media Cybernetics, U.S.A.). The results obtai
ranscript shown is normalized by the amount of 28S rRNA presenthe original culture was divided and only one portion was diluted wresented above with the only exception that cells grown for longxperiment I for 96 h before the culture was divided in two parts: o
fresh medium keeping cells at lower density (1). At 120 and 144 hhybridization for the steady-state level of PrP (moPrP) and GAPDH
analysis using labeled PCR probes constructed with
primers specific to human and mouse Prn-p genes (seeaterials and Methods), we observed the following:EL cells expressed high levels of PrP mRNA after 48
nd 96 h incubation with UDP-4 compared to controlntreated cells (0 h) (Fig. 7). Similarly, two other cell
ines (RD/TE-671, RD) have shown an increase in therP mRNA (huPrP) level at 96 h, although to a lesserxtent. Mouse probe (moPrP) did not detect any PrPRNA in RD/TE-671 and RD human cells. The bottom
llustrates the level of GAPDH and bmajor in MEL andother cell lines. In contrast to nonhematopoietic RD/TE-671 and RD cells, only MEL cells synthesized sub-stantial bmajor globin mRNA during the differentiationprocess, as expected. These data leave no doubt thatinduction of differentiation leads to activation of thePrn-p gene not only in MEL cells, but also in RD andRD/TE-671 cells of different histological origin thatrespond to the same inducer, UDP-4. Therefore, weconclude that Prn-p gene expression is increased aspart of the terminal maturation process in differentcell types.
Transcriptional Activation in MEL Cells Due toConfluency or Differentiation
The data described above have indicated that the
from exponential to confluent state. Experiment I. At various timeed by Northern blot hybridization using a mouse genomic PrP probeed as internal markers. The position of 28S rRNA was visualized byhe steady-state accumulation level of PrP RNA transcripts (moPrP)in Fig. 1A, F) is shown by the symbol (2), while that in the cells ofh incubation. Quantitative scanning analysis to assess the relative
28S rRNA as shown in the blots was performed by using the Gel-Proare shown in A and B. Note that the relative mRNA level for eachach lane. The arrows in the A and B indicate the point (72 h) whenfresh medium. Experiment II. This experiment was similar to thateriods were assessed. In particular, MEL cells were grown as ineaving cells undiluted at confluency (2) and the other diluted withotal cytoplasmic RNA was purified and analyzed by Northern blot
A transcripts as described for experiment I.
ingalyzus. T
re144of
nedin either pne l, t
expression of the Prn-p gene is regulated via the cell

c
sl
adsgmtavpm
wpa
413Prn-p GENE EXPRESSION IN NEOPLASTIC CELLS
cycle phases in growing and differentiating cells. Todistinguish whether the changes seen in the level ofPrP mRNA are due to transcriptional rather than post-transcriptional activation of Prn-p gene, we performedcell-free nuclear run-off transcriptional studies. In par-ticular, we performed run-off studies with intact nucleiprepared from control (untreated) and DMSO-treatedcells, as well as confluent cultured cells arrested at G1
phase. As shown in Fig. 8, the level of PrP RNA tran-scripts in confluent and DMSO-treated cells is rela-tively higher than that seen in rapidly growing cells.Although the level of newly synthesized mRNA wasstill lower than that of bmajor globin mRNA, a gene thatis selectively activated during erythroid maturation inMEL cells, the relative level of PrP RNA transcript washigher in G1-arrested cells. The expression level ofGAPDH, a gene used as an internal control, was un-changed in growing, arrested, and terminally differen-tiated cell nuclei. These data indicate that the Prn-pgene is transcriptionally activated in DMSO-treatedand confluent MEL cells arrested at G1 phase of the cell
FIG. 3. Kinetics of cell growth, accumulation of hemoglobin-proith DMSO. (A, B, and C) FACS analysis was performed in DMSropidium iodide (see Materials and Methods). The proportion ofpoptotic cells) is depicted in the insets. (D) Highly inducible MEL
v/v). After 24 h incubation shown by the arrow, the culture was sother was diluted (1:10) with freshly prepared medium containinalmost identical patterns of growth in contrast to that seen in Fig(E) Accumulation of hemoglobin-producing cells (benzidine-positivfor D (F, undiluted; Œ, diluted cultures).
ycle.
DISCUSSION
The precise mechanism(s) via which PrPC is synthe-ized in mammalian cells and is converted to its patho-ogical isoform PrPSc responsible for degenerative
transmissible encephalopathies is still under intensiveinvestigation [28]. Although a vast amount of workwith transgenic animals bearing mutations in thePrn-p gene has added valuable information on the ab-normal aggregation of PrPSc [29], very little is knownwith respect to the physiological role of the Prn-p genend its transcriptional regulation during cell growth,ifferentiation, and even cell death. Complete under-tanding of the regulation of expression of the Prn-pene in several species would provide valuable infor-ation on how PrP is produced and accumulated inside
he animal cells. Moreover, new insights on the mech-nistic aspects of Prn-p gene regulation could providealuable information on how one can design and ex-loit novel agents that could selectively prevent accu-ulation of Prn-p gene products.
cing cells, and FACS analysis of MEL cells induced to differentiatereated MEL cells after 0, 48, and 96 h incubation with the use ofls in each phase of the cell cycle (G1, G2, S, and A, proportion oflls were seeded at 1 3 105 cells/ml in the presence of DMSO (1.5%in two portions. One was kept as it was (undiluted, F) while the
he same inducer (diluted culture, Œ). The two cultures exhibiteddue to induction of commitment that restricts cell proliferation.
ells) at 24-h intervals in cultures exposed to DMSO as described
duO-tcelce
plitg t. 1Ae c
In this study, we attempted to ask simple but fun-

grldtaescaesta
ie
sosc
414 GOUGOUMAS, VIZIRIANAKIS, AND TSIFTSOGLOU
damental questions concerning the regulation of Prn-pene expression in highly proliferating, growth-ar-ested, and terminally differentiated cells. Since aarge number of genes are repressed during MEL cellifferentiation [17], we were surprised to observe thathe Prn-p gene was transcriptionally activated in cellsrrested at G1 phase of the cell cycle either by conflu-ncy or by terminal maturation. Moreover, we ob-erved that promotion of cell proliferation by switchingonfluent cells into exponential-phase cells by dilutionnd feeding with FCS led to decreased Prn-p genexpression, suggesting that during proliferation of cellsignals are transduced to repress the Prn-p gene at theranscriptional level. Finally, we observed that growthrrest of cells in G1 phase in differentiating and non-
differentiating cells leads to activation of Prn-p geneexpression. This result is in agreement with previousobservations that correlate activation of the Prn-p genen PC12 cells and neuronal precursor cell lines differ-ntiated by growth factors [30, 31].Activation of Prn-p gene expression in G1 phase cells
reported here, for the first time, is likely to be part ofthe cell-cycle events occurring in several differentiat-ing cell types, including hematopoietic MEL, RD/TE-
FIG. 4. Accumulation of PrP RNA transcripts (moPrP) in termi-nally differentiated MEL cells. Northern blot hybridization analysisof the steady-state accumulation of PrP mRNA in cells from diluted(1) and undiluted (2) cultures growing in the presence of 1.5% v/vDMSO. The positions of 28S rRNA and GAPDH (used as internalmarkers) are indicated. A mouse genomic DNA probe encoding PrPwas used as probe. Quantitative scanning analysis for PrP (A) andfor GAPDH (B) mRNAs was performed as indicated in Fig. 2. Thearrows in A and B show the point in time (24 h) when the originalculture was split apart.
671, or RD cells. This suggests that transcription of the m
Prn-p gene can be regulated either by external growthstimuli or by differentiation agents. Transcriptionalactivation of the Prn-p gene in growth-arrested cellsand in cells unable to proliferate or even undergoingdegeneration can be of enormous significance for therole of the Prn-p gene in degenerative disorders, sincetranscription of the Prn-p gene may be modulated bydifferent external stimuli.
Unfortunately, the use of various polyclonal and onemonoclonal antibody raised against PrP did not enableus to detect prion protein even in cells expressing rel-atively high levels of PrP RNA transcripts. We antici-pated that such failure in detection is attributed tovery low and hardly detectable levels of prion proteinin MEL cells as reported by previous observations [14,32]. PrP protein has been detected in monocytes, lym-phocytes, and CD341 bone marrow stem cells, but not
FIG. 5. Effect of N6mAdo on cytoplasmic accumulation of PrPRNA transcripts in DMSO-treated MEL cells. MEL-745PC-4A cellswere exposed simultaneously to DMSO (1.5% v/v) and N6mAdo (0.5mM). At specific time intervals cells were removed from culture andtotal cytoplasmic RNA was isolated (note that time 0 means thatRNA was isolated 1 h after exposure to DMSO and N6mAdo). 15 mgof total cytoplasmic RNA per sample was electrophoretically sepa-rated on a 1% agarose gel, transferred onto a nylon membrane, andhybridized at 65°C with 32P-labeled DNA fragments coding for ham-ter PrP mRNA or rat GAPDH (described under Material and Meth-ds). Ethidium bromide staining pattern of electrophoresed RNAamples is shown, with the positions of 18S and 28S rRNAs indi-ated. Quantitative scanning analysis for PrP (A) and GAPDH (B)
RNAs was performed as indicated in Fig. 2.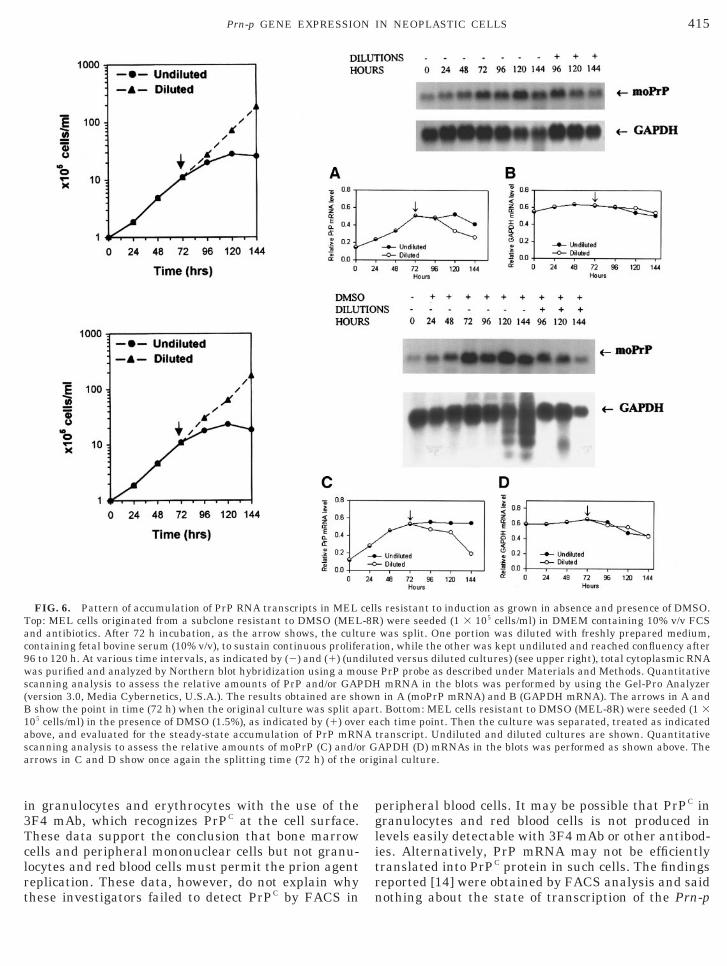
Tclrt
orig
415Prn-p GENE EXPRESSION IN NEOPLASTIC CELLS
in granulocytes and erythrocytes with the use of the3F4 mAb, which recognizes PrPC at the cell surface.
hese data support the conclusion that bone marrowells and peripheral mononuclear cells but not granu-ocytes and red blood cells must permit the prion agenteplication. These data, however, do not explain why
FIG. 6. Pattern of accumulation of PrP RNA transcripts in MELTop: MEL cells originated from a subclone resistant to DMSO (MELand antibiotics. After 72 h incubation, as the arrow shows, the cultcontaining fetal bovine serum (10% v/v), to sustain continuous prolife96 to 120 h. At various time intervals, as indicated by (2) and (1) (unwas purified and analyzed by Northern blot hybridization using a moscanning analysis to assess the relative amounts of PrP and/or GAP(version 3.0, Media Cybernetics, U.S.A.). The results obtained are shB show the point in time (72 h) when the original culture was split ap105 cells/ml) in the presence of DMSO (1.5%), as indicated by (1) oveabove, and evaluated for the steady-state accumulation of PrP mRNscanning analysis to assess the relative amounts of moPrP (C) and/oarrows in C and D show once again the splitting time (72 h) of the
hese investigators failed to detect PrPC by FACS in
peripheral blood cells. It may be possible that PrPC ingranulocytes and red blood cells is not produced inlevels easily detectable with 3F4 mAb or other antibod-ies. Alternatively, PrP mRNA may not be efficientlytranslated into PrPC protein in such cells. The findingsreported [14] were obtained by FACS analysis and said
s resistant to induction as grown in absence and presence of DMSO.) were seeded (1 3 105 cells/ml) in DMEM containing 10% v/v FCSwas split. One portion was diluted with freshly prepared medium,
ion, while the other was kept undiluted and reached confluency afterted versus diluted cultures) (see upper right), total cytoplasmic RNAPrP probe as described under Materials and Methods. QuantitativemRNA in the blots was performed by using the Gel-Pro Analyzerin A (moPrP mRNA) and B (GAPDH mRNA). The arrows in A and
t. Bottom: MEL cells resistant to DMSO (MEL-8R) were seeded (1 3ch time point. Then the culture was separated, treated as indicated
transcript. Undiluted and diluted cultures are shown. QuantitativeAPDH (D) mRNAs in the blots was performed as shown above. Theinal culture.
cell-8R
ureratdiluuseDH
ownar
r eaAr G
nothing about the state of transcription of the Prn-p

cadnf
(hccm9hoel
416 GOUGOUMAS, VIZIRIANAKIS, AND TSIFTSOGLOU
gene in hematopoietic cells. Regardless of whatevermechanism(s) underlies this discrepancy in bone mar-row cells and peripheral blood cells, our observationthat endogenous Prn-p gene is transcriptionally acti-vated in growth-arrested transformed hematopoieticcells undergoing erythroid differentiation like MELcells is intriguing and consistent with the ability ofsome differentiated hematopoietic cells to producePrPC protein.
One can argue that whatever findings presentedhere with the use of transformed hemopoietic MELcells may not apply directly to what really occurs inneurons or other cell types undergoing differentiation,growth arrest at G1 phase, or apoptosis. It is, however,important to remember that cells of hematopoietic or-igin are involved in transmission of prion infectiousagents toward CNS, as mentioned earlier [15], andthat RD/TE-671, the neuroectodermal cell line usedhere, is differentiated into neuron-like cells undergoingmaturation and cell growth arrest [19]. Although it isquite premature to draw conclusions and invent work-ing hypotheses to explain to what extent transcrip-tional activation of the Prn- p gene in several cell types
FIG. 7. Steady-state accumulation of human (huPrP) and mousemoPrP) PrP RNA transcripts in murine erythroleukemia (MEL),uman neuroectodermal (RD/TE-671), and human rhabdomyosar-oma (RD) cells differentiated by UDP-4. MEL, RD/TE-671, and RDells were grown for 96 h in the presence of the inducer UDP-4 (1M). Total cytoplasmic RNA was isolated from cells at 0, 48, and
6 h and Northern blot hybridization analysis was carried out withuman and mouse PCR-derived PrP probes (see Materials and Meth-ds), to assess the steady-state level of PrP RNA transcripts. Thethidium bromide pattern of 28S rRNA and the steady-state mRNAevels of GAPDH were used as internal markers. A bmajor globin DNA
fragment was used as a biochemical marker of erythroid differenti-ation in MEL cells (see Materials and Methods). The extent of dif-ferentiation in RD/TE-671 and RD cells was estimated morphologi-
cally as described elsewhere [19].ontributes to biosynthesis and accumulation of largemounts of PrP inside the cells and possible tissueegeneration, we consider the findings presented hereovel and provocative, since they may lead us to searchor novel agents able to repress Prn-p gene activation.
We thank Dr. T. Sklaviadis (Laboratory of Pharmacology, Depart-ment of Pharmaceutical Sciences, Aristotle University of Thessa-loniki), for his helpful discussion during the early phase of this study,as well as Dr. F. Kanakoudi-Tsakalidou and Mrs. A. Taparkou (Lab-oratory of Clinical Immunology, First Department of Pediatrics,Aristotle University of Thessaloniki), for FACS analysis.
REFERENCES
1. Stahl, N., Borchelt, D. R., Hsiao, K., and Prusiner, S. B. (1987).Scrapie prion protein contains a phosphatidylinositol glycolipid.Cell 51, 229–240.
2. Belay, E. D. (1999). Transmissible spongiform encephalopa-thies in humans. Annu. Rev. Microbiol. 53, 283–314.
3. Collinge, J., Whittington, M. A., Sidle, K. C., Smith, C. J.,Palmer, M. S., Clarke, A. R., and Jeffreys, J. G. (1994). Prionprotein is necessary for normal synaptic function. Nature 370,295–297.
4. Mabbot, N. A., Brown, K. L., Manson, J., and Bruce, M. E.(1997). T-lymphocyte activation and the cellular form of theprion protein. Immunology 92, 161–165.
5. Whatley, S. A., Powel, J. F., Politopoulou, G., Campbell, I. C.,Brammer, M. J., and Percy, N. S. (1995). Regulation of intra-cellular free calcium levels by the cellular prion protein.NeuroReport 6, 2333–2337.
6. Hornshaw, M. P., McDermott, J. R., and Candy, J. M. (1995).Copper binding to the N-terminal tandem repeat regions ofmammalian and avian prion protein. Biochem. Biophys. Res.Commun. 207, 621–629, doi:10.1006/bbrc.1995.1233.
7. Brown, D. R., Schulz-Schaeffer, W. J., Schmidt, B., andKretzschmar, H. A. (1997). Prion protein-deficient cells show al-tered response to oxidative stress due to decreased SOD-1 activity.Exp. Neurol. 146, 104–112, doi:10.1006/exnr.1997.6511.
8. Lieberburg, L. (1987). Developmental expression and regionaldistribution of the scrapie-associated protein mRNA in the ratcentral nervous system. Brain Res. 417, 363–366.
9. Mobley, W. C., Neve, R. L., Prusiner, S. B., and McKinley, M. P.
FIG. 8. Run-off transcription assay of Prn-p gene expression inMEL cell-free nuclei. Intact nuclei were isolated from MEL cellsgrown with DMSO for 72 h (A), arrested at confluent state for 120 hwith no addition in culture (B), and grown exponentially (C). Subse-quently, the nuclei were subjected to in vitro run-off transcriptionassay by hybridizing the newly synthesized and labeled RNA with aPCR-amplified PrP probe. GAPDH and bmajor globin DNA probesserved as internal markers.
(1988). Nerve growth factor increases mRNA levels for the prion

1
1
1
1
1
1
1
1
1
1
417Prn-p GENE EXPRESSION IN NEOPLASTIC CELLS
protein and the a-amyloid protein precursor in developing ham-ster brain. Proc. Natl. Acad. Sci. USA 85, 9811–9815.
0. Westaway, D., Goodman, P. A., Mirenda, C. A., McKinley,M. P., Carlson, G. A., and Prusiner, S. B. (1987). Distinct prionproteins in short and long scrapie incubation period mice. Cell51, 651–662.
1. Liao, Yu-Cheng, J., Lebo, R. V., Clawson, G. A., and Smuckler,E. A. (1986). Human prion protein cDNA: Molecular cloning,chromosomal mapping, and biological implications. Science233, 364–367.
2. Kretzschmar, H. A., Prusiner, S. B., Stowring, L. E., and DeArmond, S. J. (1986). Scrapie prion proteins are synthesized inneurons. Am. J. Pathol. 122, 1–5.
3. Horiuchi, M., Yamazaki, N., Ikeda, T., Ishiguro, N., and Shina-gawa, M. (1995). A cellular form of prion protein (PrPC) exists inmany non-neuronal tissues of sheep. J. Gen. Virol. 76, 2583–2587.
4. Dodelet, V. C., and Cashman N. R. (1998). Prion protein expres-sion in human leukocyte differentiation. Blood 91, 1556–1561.
5. Klein, M. A., Frigg, R., Flechsig, E., Raeber, A. J., Kalinke, U.,Bluethmann, H., Bootz, F., Suter, M., Zinkernagel, R. M., andAguzzi, A. (1997). A crucial role for B cells in neuroinvasivescrapie. Nature 390, 687–690.
6. Vizirianakis, I. S., Tsiftsoglou, A. S., and Sklaviadis, T. (1994).Developments in slow virus research: Implication of PrP pro-tein involvement in cell growth and differentiation. In “Trans-missible Spongiform Encephalopathies” (R. Bradley and B.Marchant, Eds.), pp. 421–429, Commission of the EuropeanCommunities, Brussels.
7. Tsiftsoglou, A. S., and Robinson, S. H. (1992). Differentiation ofmurine erythroleukemia cells and human HL-60 cell lines. In“Concise Reviews in Clinical and Experimental Hematology”(M. J. Murphy, Ed.), pp. 295–306, Alphamed Press, Dayton,OH.
8. Marks, P. A., Sheffery, M., and Rifkin, R. A. (1987). Induction oftransformed cells to terminal differentiation and the modula-tion of gene expression. Cancer Res. 47, 659–666.
9. Pappas, I., Sophianos, D., Tzartos, S., and Tsiftsoglou, A. S.(1996). Expression of memory, differentiation, and repression ofc-myc and p53 genes in human RD/TE-671 cells induced by aureido-derivative of pyridine (UDP-4). Cell Growth Differ. 7,797–809.
20. Pappas, I. S., Niopas, I., and Tsiftsoglou, A. S. (1992). Ureidoderivatives of pyridine: A new class of inducers of murine eryth-roleukemia cell differentiation. Anticancer Drug Des. 7, 153–
161.21. Robakis, N. K., Sawh, P. S., Wolfe, G. C., Rubenstein, R., Carp,R. I., and Innis, R. (1986). Isolation of a cDNA clone encodingthe leader peptide of prion protein and expression of the homol-ogous gene in various tissues. Proc. Natl. Acad. Sci. USA 83,6377–6381.
22. Fort, P., Marty, L., Piechaczy, K., el Sabrouty, S., Dani, C.,Jeanteur, P., and Blanchard, J. M. (1986). Various rat adulttissues express only one major mRNA species from glyceralde-hyde-3-phosphate dehydrogenase multigenic family. NucleicAcids Res. 13, 1431–1442.
23. Orkin, S. H., Harosi, F. I., and Leder, P. (1975). Differentiationin erythroleukemic cells and their somatic hybrids. Proc. Natl.Acad. Sci. USA 72, 98–102.
24. Crissman, H. A., and Steinkamp, J. A. (1973). Rapid, simulta-neous measurement of DNA, protein, and cell volume in singlecells from large mammalian cell population. J. Cell Biol. 59,766–771.
25. Greenberg, M. E., and Bender, T. P. (1994). Identification ofnewly transcribed RNA. In “Current Protocols in MolecularBiology,” unit 4.10.1–4.10.4, Wiley, New York.
26. Vizirianakis, I. S., Pappas, I. S., Gougoumas, D., and Tsiftso-glou, A. S. (1999). Expression of ribosomal protein S5 clonedgene during differentiation and apoptosis in murine erythroleu-kemia (MEL) cells. Oncol. Res. 11, 409–419.
27. Vizirianakis, I. S., and Tsiftsoglou, A. S. (1995). N6-Methylad-enosine inhibits murine erythroleukemia cell maturation byblocking methylation of RNA and memory via conversion toS-(N6-methyl)-adenosylhomocysteine. Biochem. Pharmacol. 50,1807–1814.
28. Beleay, E. D. (1999). Transmissible spongiform encephalopa-thies in humans. Annu. Rev. Microbiol. 53, 283–314.
29. Raeber, A. J., Brandner, S., Klein, M. A., Benninger, Y.,Musahl, C., Frigg, R., Roeckl, C., Fischer, M. B., Weissmann, C.,and Aguzzi, A. (1998). Transgenic and knockout mice in re-search on prion diseases. Brain Pathol. 8, 715–733.
30. Lazarini, F., Castelnau, P., Chermann, J.-F., Deslys, J.-P., andDormont, D. (1994). Modulation of prion protein gene expres-sion by growth factors in cultured mouse astrocytes and PC-12cells. Mol. Brain Res. 22, 268–274.
31. Kuwahara, C., Kubosaki, A., Nishimura, T., Nasu, Y., Naka-mura, Y., Saeki, K., Matsumoto, Y., and Onodera T. (2000).Enhanced expression of cellular prion protein gene by insulin ornerve growth factor in immortalized mouse neuronal precursorcell lines. Biochem. Biophys. Res. Commun. 269, 763–766.
32. Scott, M. R. D., Butler, D. A., Bredesen, D. E., Walchli, M.,Hsiao, K. K., and Prusiner, S. B. (1988). Prion protein expres-
sion in cultured cells. Protein Eng. 2, 69–76.Received July 25, 2000Revised version received December 5, 2000Published online March 1, 2001
Related Documents











