1238 VOLUME 12 NUMBER 12 DECEMBER 2011 NATURE IMMUNOLOGY Cells of the T H 17 subset of helper T cells ‘preferentially’ produce inter- leukin 17A (IL-17A), IL-17F and IL-22 and have an indispensable role in host defense against infection by extracellular pathogens 1 . IL-22 is a member of the IL-10 cytokine family 2 . In addition to being produced by T H 17 cells, IL-22 is produced by a variety of leukocytes, includ- ing CD4 + T cells and CD8 + T cells, natural killer cells and lymphoid tissue–inducer cells 3,4 . IL-17A and IL-22 both enhance the innate immune responses of tissue fibroblasts, epithelial cells and other stromal cells 1 . The syn- ergistic functions of IL-17A and IL-22 are essential not only to host defense against infections but also to the pathogenesis of inflamma- tory and autoimmune diseases. For example, both IL-17A and IL-22 are required for host control of Klebsiella pneumonia infection in the lung 5,6 , but they act synergistically on keratinocytes and participate in the development of psoriasis 7–9 . However, the functions of IL-17A and IL-22 do not overlap com- pletely 10–12 . IL-22 augments epithelial proliferation and healing responses, whereas IL-17A induces much stronger proinflammatory effects and more recruitment of neutrophils 13,14 . In vivo, IL-22 can exert both tissue-protective and proinflammatory functions depend- ing on other environmental factors, such as the cytokine milieu 3,15 . In bleomycin-induced lung inflammation, the tissue-protective versus proinflammatory functions of IL-22 are controlled by the absence or presence of IL-17A, respectively 12 . Consistent with that, only a fraction of IL-17A-producing T cells also express IL-22 (refs. 7,8). In humans, a T H 22 subset of helper T cells, which produce IL-22 but not IL-17, has been identified 16,17 . In addition, the presence of T cells that produce both IL-22 and IL-17 is associated with mucosal healing, whereas the infiltration of T cells that produce only IL-17 in the mucosa has been linked to active colitis in a patient with ulcerative colitis 18 . Such data suggest that under certain physiological condi- tions, T cells may produce IL-22 or IL-17 or a combination of both to initiate the appropriate downstream responses. Understanding the differences in the regulation of IL-22 and IL-17 in T cells should pro- vide insight into the physiological and pathogenic functions of these two cytokines in various diseases. IL-17A and IL-22 are regulated differently in T cells 8 . The combi- nation of IL-6 and transforming growth factor-β (TGF-β) represents the minimal requirement for new T H 17 differentiation of naive mouse T cells and for IL-17A production 19–21 . However, IL-6 alone is suffi- cient to induce IL-22 from naive T cells in vitro in mice and humans 8,16 . Most notable, however, is the disparate regulation of IL-17A and IL-22 by TGF-β. Although it is required for robust induction of IL-17A in naive T cells 19–21 , TGF-β is not only dispensable for IL-22 expression but actually inhibits IL-22 production in mouse and human T cells 8,16 . Several transcription factors, including RORγt, AhR and BATF, are essential for the development of T H 17 cells and the production of IL-17. The same transcription factors also are crucial for IL-22 expres- sion, as T cells deficient in any of these are also considerably impaired in their ability to produce IL-22 (refs. 22–26). However, the under- lying transcriptional mechanisms that account for the differences in the regulation of IL-17 and IL-22 by TGF-β remain elusive. Here we demonstrate that the TGF-β-dependent suppression of IL-22 in T cells was mediated by the transcription factor c-Maf. Originally described as a factor specific for type 2 helper T cells (T H 2 cells) that is involved in the induction of IL-4 expression 27 , c-Maf has since 1 Department of Immunology, Genentech, South San Francisco, California, USA. 2 Bioinformatics & Computational Biology, Genentech, South San Francisco, California, USA. 3 Present address: Target Discovery & Validation, Novo Nordisk Inflammation Research Center, Seattle, Washington, USA. Correspondence should be addressed to W.O. ([email protected]). Received 9 May; accepted 6 September; published online 16 October 2011; doi:10.1038/ni.2134 Transcription factor c-Maf mediates the TGF- b- dependent suppression of IL-22 production in T H 17 cells Sascha Rutz 1 , Rajkumar Noubade 1 , Céline Eidenschenk 1 , Naruhisa Ota 1 , Wenwen Zeng 1 , Yan Zheng 1,3 , Jason Hackney 2 , Jiabing Ding 1 , Harinder Singh 1 & Wenjun Ouyang 1 Interleukin 22 (IL-22), which is produced by cells of the T H 17 subset of helper T cells and other leukocytes, not only enhances proinflammatory innate defense mechanisms in epithelial cells but also provides crucial protection to tissues from damage caused by inflammation and infection. In T H 17 cells, transforming growth factor-b (TGF-b) regulates IL-22 and IL-17 differently. IL-6 alone induces T cells to produce only IL-22, whereas the combination of IL-6 and high concentrations of TGF- b results in the production of IL-17 but not IL-22 by T cells. Here we identify the transcription factor c-Maf, which is induced by TGF- b, as a downstream repressor of Il22. We found that c-Maf bound to the Il22 promoter and was both necessary and sufficient for the TGF-b-dependent suppression of IL-22 production in T H 17 cells. ARTICLES © 2011 Nature America, Inc. All rights reserved. © 2011 Nature America, Inc. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1238 VOLUME 12 NUMBER 12 DECEMBER 2011 nature immunology
Cells of the TH17 subset of helper T cells ‘preferentially’ produce inter-leukin 17A (IL-17A), IL-17F and IL-22 and have an indispensable role in host defense against infection by extracellular pathogens1. IL-22 is a member of the IL-10 cytokine family2. In addition to being produced by TH17 cells, IL-22 is produced by a variety of leukocytes, includ-ing CD4+ T cells and CD8+ T cells, natural killer cells and lymphoid tissue–inducer cells3,4.
IL-17A and IL-22 both enhance the innate immune responses of tissue fibroblasts, epithelial cells and other stromal cells1. The syn-ergistic functions of IL-17A and IL-22 are essential not only to host defense against infections but also to the pathogenesis of inflamma-tory and autoimmune diseases. For example, both IL-17A and IL-22 are required for host control of Klebsiella pneumonia infection in the lung5,6, but they act synergistically on keratinocytes and participate in the development of psoriasis7–9.
However, the functions of IL-17A and IL-22 do not overlap com-pletely10–12. IL-22 augments epithelial proliferation and healing responses, whereas IL-17A induces much stronger proinflammatory effects and more recruitment of neutrophils13,14. In vivo, IL-22 can exert both tissue-protective and proinflammatory functions depend-ing on other environmental factors, such as the cytokine milieu3,15. In bleomycin-induced lung inflammation, the tissue-protective versus proinflammatory functions of IL-22 are controlled by the absence or presence of IL-17A, respectively12. Consistent with that, only a fraction of IL-17A-producing T cells also express IL-22 (refs. 7,8). In humans, a TH22 subset of helper T cells, which produce IL-22 but not IL-17, has been identified16,17. In addition, the presence of T cells that produce both IL-22 and IL-17 is associated with mucosal
healing, whereas the infiltration of T cells that produce only IL-17 in the mucosa has been linked to active colitis in a patient with ulcerative colitis18. Such data suggest that under certain physiological condi-tions, T cells may produce IL-22 or IL-17 or a combination of both to initiate the appropriate downstream responses. Understanding the differences in the regulation of IL-22 and IL-17 in T cells should pro-vide insight into the physiological and pathogenic functions of these two cytokines in various diseases.
IL-17A and IL-22 are regulated differently in T cells8. The combi-nation of IL-6 and transforming growth factor-β (TGF-β) represents the minimal requirement for new TH17 differentiation of naive mouse T cells and for IL-17A production19–21. However, IL-6 alone is suffi-cient to induce IL-22 from naive T cells in vitro in mice and humans8,16. Most notable, however, is the disparate regulation of IL-17A and IL-22 by TGF-β. Although it is required for robust induction of IL-17A in naive T cells19–21, TGF-β is not only dispensable for IL-22 expression but actually inhibits IL-22 production in mouse and human T cells8,16. Several transcription factors, including RORγt, AhR and BATF, are essential for the development of TH17 cells and the production of IL-17. The same transcription factors also are crucial for IL-22 expres-sion, as T cells deficient in any of these are also considerably impaired in their ability to produce IL-22 (refs. 22–26). However, the under-lying transcriptional mechanisms that account for the differences in the regulation of IL-17 and IL-22 by TGF-β remain elusive.
Here we demonstrate that the TGF-β-dependent suppression of IL-22 in T cells was mediated by the transcription factor c-Maf. Originally described as a factor specific for type 2 helper T cells (TH2 cells) that is involved in the induction of IL-4 expression27, c-Maf has since
1Department of Immunology, Genentech, South San Francisco, California, USA. 2Bioinformatics & Computational Biology, Genentech, South San Francisco, California, USA. 3Present address: Target Discovery & Validation, Novo Nordisk Inflammation Research Center, Seattle, Washington, USA. Correspondence should be addressed to W.O. ([email protected]).
Received 9 May; accepted 6 September; published online 16 October 2011; doi:10.1038/ni.2134
Transcription factor c-Maf mediates the TGF-b-dependent suppression of IL-22 production in TH17 cellsSascha Rutz1, Rajkumar Noubade1, Céline Eidenschenk1, Naruhisa Ota1, Wenwen Zeng1, Yan Zheng1,3, Jason Hackney2, Jiabing Ding1, Harinder Singh1 & Wenjun Ouyang1
Interleukin 22 (IL-22), which is produced by cells of the TH17 subset of helper T cells and other leukocytes, not only enhances proinflammatory innate defense mechanisms in epithelial cells but also provides crucial protection to tissues from damage caused by inflammation and infection. In TH17 cells, transforming growth factor-b (TGF-b) regulates IL-22 and IL-17 differently. IL-6 alone induces T cells to produce only IL-22, whereas the combination of IL-6 and high concentrations of TGF-b results in the production of IL-17 but not IL-22 by T cells. Here we identify the transcription factor c-Maf, which is induced by TGF-b, as a downstream repressor of Il22. We found that c-Maf bound to the Il22 promoter and was both necessary and sufficient for the TGF-b-dependent suppression of IL-22 production in TH17 cells.
A rt i c l e s©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.
nature immunology VOLUME 12 NUMBER 12 DECEMBER 2011 1239
been shown to mediate IL-10 expression in T cells28–30 and macro-phages31. Here we identified c-Maf as a repressor of Il22 but not of Il17. It bound to the Il22 promoter and overcame the positive transcriptional regulation of Il22 by other TH17 transcription factors, including RORγt and BATF. Furthermore, we found that AhR was not required for opti-mal IL-22 production in T cells in the absence of TGF-β.
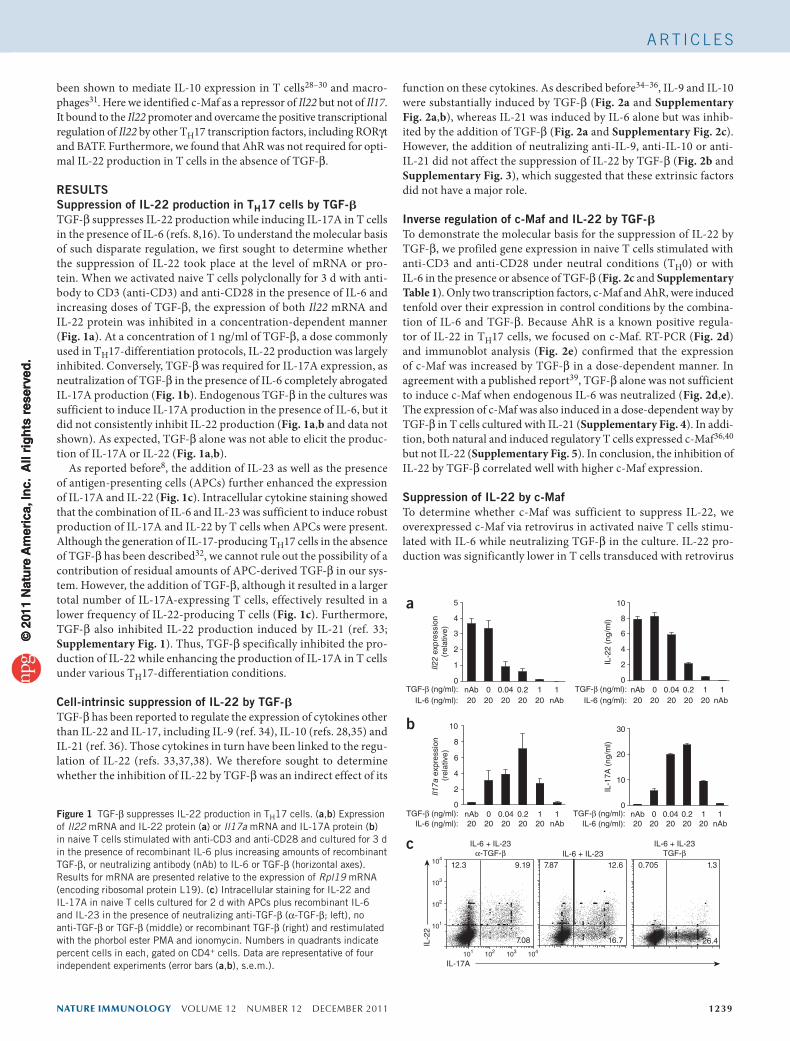
RESULTSSuppression of IL-22 production in TH17 cells by TGF-bTGF-β suppresses IL-22 production while inducing IL-17A in T cells in the presence of IL-6 (refs. 8,16). To understand the molecular basis of such disparate regulation, we first sought to determine whether the suppression of IL-22 took place at the level of mRNA or pro-tein. When we activated naive T cells polyclonally for 3 d with anti-body to CD3 (anti-CD3) and anti-CD28 in the presence of IL-6 and increasing doses of TGF-β, the expression of both Il22 mRNA and IL-22 protein was inhibited in a concentration-dependent manner (Fig. 1a). At a concentration of 1 ng/ml of TGF-β, a dose commonly used in TH17-differentiation protocols, IL-22 production was largely inhibited. Conversely, TGF-β was required for IL-17A expression, as neutralization of TGF-β in the presence of IL-6 completely abrogated IL-17A production (Fig. 1b). Endogenous TGF-β in the cultures was sufficient to induce IL-17A production in the presence of IL-6, but it did not consistently inhibit IL-22 production (Fig. 1a,b and data not shown). As expected, TGF-β alone was not able to elicit the produc-tion of IL-17A or IL-22 (Fig. 1a,b).
As reported before8, the addition of IL-23 as well as the presence of antigen-presenting cells (APCs) further enhanced the expression of IL-17A and IL-22 (Fig. 1c). Intracellular cytokine staining showed that the combination of IL-6 and IL-23 was sufficient to induce robust production of IL-17A and IL-22 by T cells when APCs were present. Although the generation of IL-17-producing TH17 cells in the absence of TGF-β has been described32, we cannot rule out the possibility of a contribution of residual amounts of APC-derived TGF-β in our sys-tem. However, the addition of TGF-β, although it resulted in a larger total number of IL-17A-expressing T cells, effectively resulted in a lower frequency of IL-22-producing T cells (Fig. 1c). Furthermore, TGF-β also inhibited IL-22 production induced by IL-21 (ref. 33; Supplementary Fig. 1). Thus, TGF-β specifically inhibited the pro-duction of IL-22 while enhancing the production of IL-17A in T cells under various TH17-differentiation conditions.
Cell-intrinsic suppression of IL-22 by TGF-bTGF-β has been reported to regulate the expression of cytokines other than IL-22 and IL-17, including IL-9 (ref. 34), IL-10 (refs. 28,35) and IL-21 (ref. 36). Those cytokines in turn have been linked to the regu-lation of IL-22 (refs. 33,37,38). We therefore sought to determine whether the inhibition of IL-22 by TGF-β was an indirect effect of its
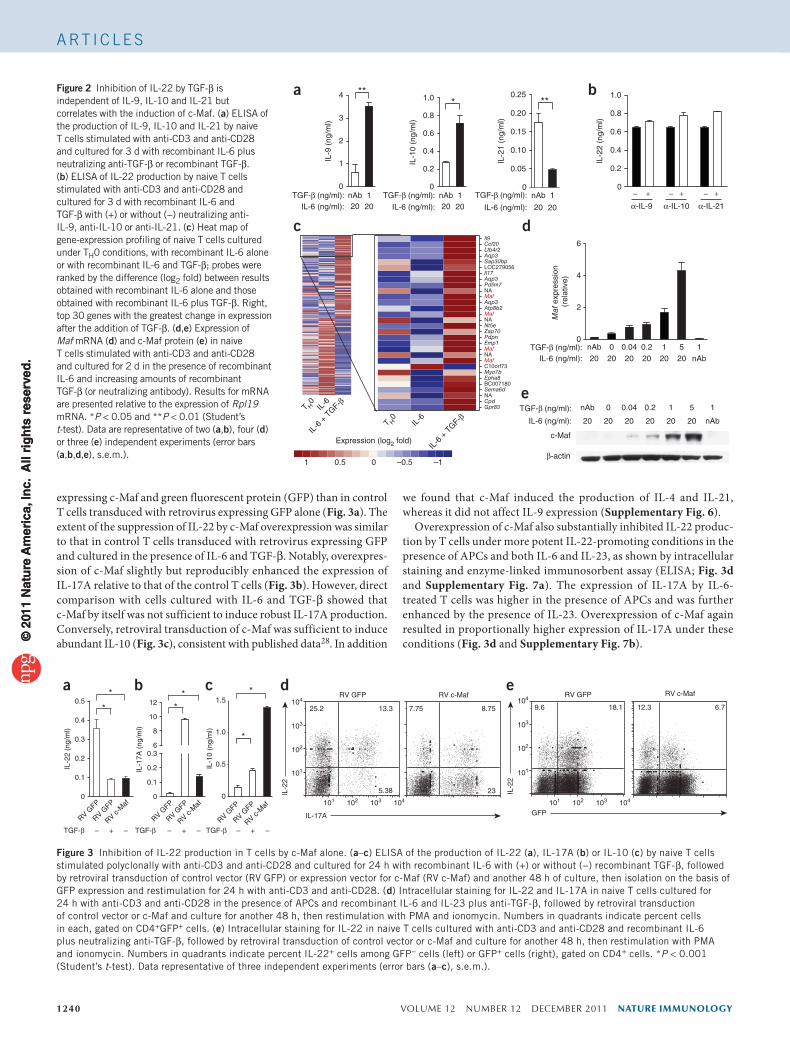
function on these cytokines. As described before34–36, IL-9 and IL-10 were substantially induced by TGF-β (Fig. 2a and Supplementary Fig. 2a,b), whereas IL-21 was induced by IL-6 alone but was inhib-ited by the addition of TGF-β (Fig. 2a and Supplementary Fig. 2c). However, the addition of neutralizing anti-IL-9, anti-IL-10 or anti-IL-21 did not affect the suppression of IL-22 by TGF-β (Fig. 2b and Supplementary Fig. 3), which suggested that these extrinsic factors did not have a major role.
Inverse regulation of c-Maf and IL-22 by TGF-bTo demonstrate the molecular basis for the suppression of IL-22 by TGF-β, we profiled gene expression in naive T cells stimulated with anti-CD3 and anti-CD28 under neutral conditions (TH0) or with IL-6 in the presence or absence of TGF-β (Fig. 2c and Supplementary Table 1). Only two transcription factors, c-Maf and AhR, were induced tenfold over their expression in control conditions by the combina-tion of IL-6 and TGF-β. Because AhR is a known positive regula-tor of IL-22 in TH17 cells, we focused on c-Maf. RT-PCR (Fig. 2d) and immunoblot analysis (Fig. 2e) confirmed that the expression of c-Maf was increased by TGF-β in a dose-dependent manner. In agreement with a published report39, TGF-β alone was not sufficient to induce c-Maf when endogenous IL-6 was neutralized (Fig. 2d,e). The expression of c-Maf was also induced in a dose-dependent way by TGF-β in T cells cultured with IL-21 (Supplementary Fig. 4). In addi-tion, both natural and induced regulatory T cells expressed c-Maf36,40 but not IL-22 (Supplementary Fig. 5). In conclusion, the inhibition of IL-22 by TGF-β correlated well with higher c-Maf expression.
Suppression of IL-22 by c-MafTo determine whether c-Maf was sufficient to suppress IL-22, we overexpressed c-Maf via retrovirus in activated naive T cells stimu-lated with IL-6 while neutralizing TGF-β in the culture. IL-22 pro-duction was significantly lower in T cells transduced with retrovirus
IL-2
2
IL-17A
IL-6 + IL-23TGF-β
IL-6 + IL-23α-TGF-β
c7.87 12.6
16.7
12.3 9.19
7.08
0.705 1.3
26.4
IL-6 + IL-23
101 102 103 104
101
102
103
104
IL-2
2 (n
g/m
l)
Il22
expr
essi
on
(rel
ativ
e)
a
0
2
4
6
8
10
0
1
2
3
4
5
TGF-β (ng/ml):IL-6 (ng/ml):
TGF-β (ng/ml):IL-6 (ng/ml):
TGF-β (ng/ml):IL-6 (ng/ml):
0.04 0.2 1 10nAb20 20 20 20 20 nAb
TGF-β (ng/ml):IL-6 (ng/ml):
0.04 0.2 1 10nAb20 20 20 20 20 nAb
IL-1
7A (
ng/m
l)
b
Il17a
exp
ress
ion
(rel
ativ
e)
0
10
20
30
0
2
4
6
8
10
0.04 0.2 1 10nAb20 20 20 20 20 nAb
0.04 0.2 1 10nAb20 20 20 20 20 nAb
Figure 1 TGF-β suppresses IL-22 production in TH17 cells. (a,b) Expression of Il22 mRNA and IL-22 protein (a) or Il17a mRNA and IL-17A protein (b) in naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 3 d in the presence of recombinant IL-6 plus increasing amounts of recombinant TGF-β, or neutralizing antibody (nAb) to IL-6 or TGF-β (horizontal axes). Results for mRNA are presented relative to the expression of Rpl19 mRNA (encoding ribosomal protein L19). (c) Intracellular staining for IL-22 and IL-17A in naive T cells cultured for 2 d with APCs plus recombinant IL-6 and IL-23 in the presence of neutralizing anti-TGF-β (α-TGF-β; left), no anti-TGF-β or TGF-β (middle) or recombinant TGF-β (right) and restimulated with the phorbol ester PMA and ionomycin. Numbers in quadrants indicate percent cells in each, gated on CD4+ cells. Data are representative of four independent experiments (error bars (a,b), s.e.m.).
A rt i c l e s©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.
1240 VOLUME 12 NUMBER 12 DECEMBER 2011 nature immunology
A rt i c l e s
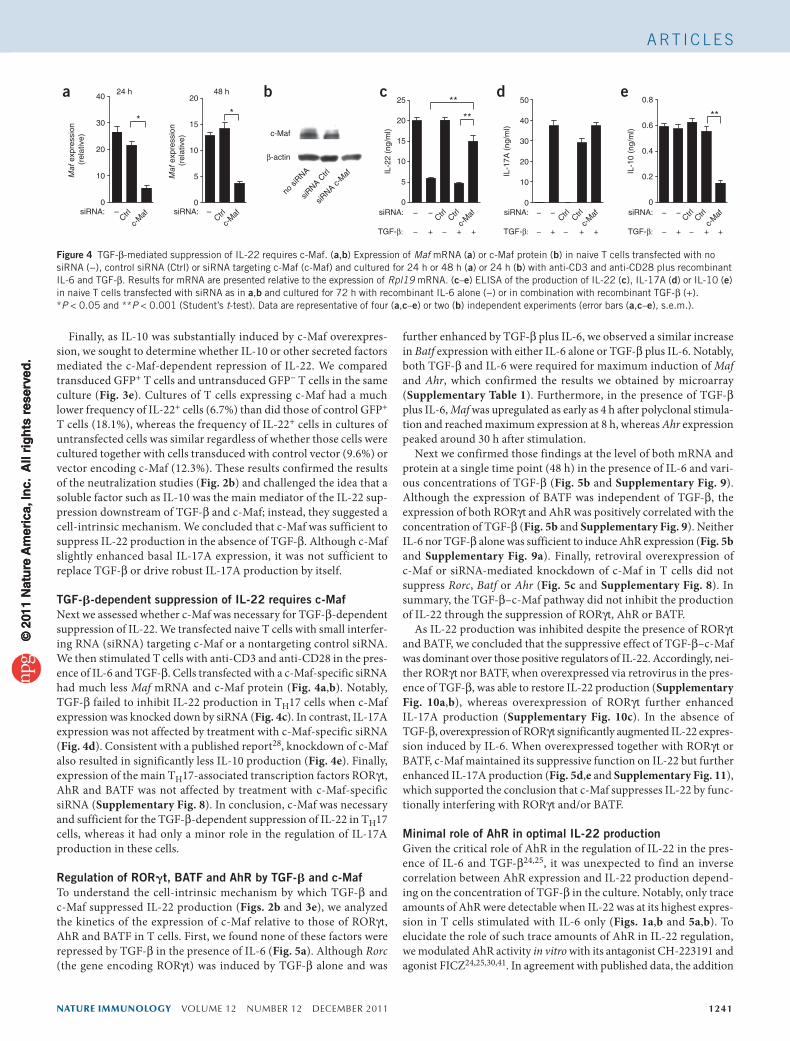
expressing c-Maf and green fluorescent protein (GFP) than in control T cells transduced with retrovirus expressing GFP alone (Fig. 3a). The extent of the suppression of IL-22 by c-Maf overexpression was similar to that in control T cells transduced with retrovirus expressing GFP and cultured in the presence of IL-6 and TGF-β. Notably, overexpres-sion of c-Maf slightly but reproducibly enhanced the expression of IL-17A relative to that of the control T cells (Fig. 3b). However, direct comparison with cells cultured with IL-6 and TGF-β showed that c-Maf by itself was not sufficient to induce robust IL-17A production. Conversely, retroviral transduction of c-Maf was sufficient to induce abundant IL-10 (Fig. 3c), consistent with published data28. In addition
we found that c-Maf induced the production of IL-4 and IL-21, whereas it did not affect IL-9 expression (Supplementary Fig. 6).
Overexpression of c-Maf also substantially inhibited IL-22 produc-tion by T cells under more potent IL-22-promoting conditions in the presence of APCs and both IL-6 and IL-23, as shown by intracellular staining and enzyme-linked immunosorbent assay (ELISA; Fig. 3d and Supplementary Fig. 7a). The expression of IL-17A by IL-6-treated T cells was higher in the presence of APCs and was further enhanced by the presence of IL-23. Overexpression of c-Maf again resulted in proportionally higher expression of IL-17A under these conditions (Fig. 3d and Supplementary Fig. 7b).
a b
c
c-Maf
β-actin
e
T H0
IL-6
IL-6
+ T
GF-β
Maf
Maf
Maf
Maf
T H0
IL-6
IL-6
+ T
GF-β
0
0.2
0.4
0.6
0.8
1.0
IL-2
2 (n
g/m
l)
Il9Ccl20Ltb4r2Aqp3Sap30bpLOC279056Il17Aqp3Pdlim7NA
Aqp3Atp8b2
NANt5eZap70PdpnEmp1
NA
C10orf73Myo7bEpha8BC007180Sema6dNACpdGpr83
Maf
exp
ress
ion
(rel
ativ
e)
α-IL-9
− +α-IL-10
− +α-IL-21
− +
d
***
**
0
1
2
3
4
0
0.2
0.4
0.6
0.8
1.0
0
0.05
0.10
0.15
0.20
0.25
IL-9
(ng
/ml)
IL-1
0 (n
g/m
l)
IL-2
1 (n
g/m
l)
TGF-β (ng/ml): nAb 12020
nAb 12020
nAb 1
2020IL-6 (ng/ml):
0
2
4
6
TGF-β (ng/ml):
IL-6 (ng/ml):
TGF-β (ng/ml):
IL-6 (ng/ml):
TGF-β (ng/ml):
IL-6 (ng/ml):
TGF-β (ng/ml):IL-6 (ng/ml):
0.04 0.2 1 10nAb20 20 20 20 20 nAb
5
0.04 0.2 1 10nAb 5
20
20 20 20 20 20 nAb20
Expression (log2 fold)
1 0 –0.5 –10.5
Figure 2 Inhibition of IL-22 by TGF-β is independent of IL-9, IL-10 and IL-21 but correlates with the induction of c-Maf. (a) ELISA of the production of IL-9, IL-10 and IL-21 by naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 3 d with recombinant IL-6 plus neutralizing anti-TGF-β or recombinant TGF-β. (b) ELISA of IL-22 production by naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 3 d with recombinant IL-6 and TGF-β with (+) or without (−) neutralizing anti-IL-9, anti-IL-10 or anti-IL-21. (c) Heat map of gene-expression profiling of naive T cells cultured under TH0 conditions, with recombinant IL-6 alone or with recombinant IL-6 and TGF-β; probes were ranked by the difference (log2 fold) between results obtained with recombinant IL-6 alone and those obtained with recombinant IL-6 plus TGF-β. Right, top 30 genes with the greatest change in expression after the addition of TGF-β. (d,e) Expression of Maf mRNA (d) and c-Maf protein (e) in naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 2 d in the presence of recombinant IL-6 and increasing amounts of recombinant TGF-β (or neutralizing antibody). Results for mRNA are presented relative to the expression of Rpl19 mRNA. *P < 0.05 and **P < 0.01 (Student’s t-test). Data are representative of two (a,b), four (d) or three (e) independent experiments (error bars (a,b,d,e), s.e.m.).
IL-2
2 (n
g/m
l)
a cb*
IL-1
7A (
ng/m
l)
0
0.1
TGF-β – –+ TGF-β – –+ TGF-β – –+
0.2
0.3
0.4
0.5
0
0.1
0.2
0.36
8
10
12
0
0.5
1.0
1.5*
IL-1
0 (n
g/m
l)
*
*
*
* 25.2 13.3
5.38
7.75 8.75
23IL-2
2
IL-17A
RV GFP RV c-Mafd
101 102 103 104 101 102 103 104
104
103
102
101
104
103
102
101
e9.6 18.1 12.3 6.7
IL-2
2
GFP
RV GFP RV c-Maf
RV GFP
RV c-M
af
RV GFP
RV GFP
RV c-M
af
RV GFP
RV GFP
RV c-M
af
RV GFP
Figure 3 Inhibition of IL-22 production in T cells by c-Maf alone. (a–c) ELISA of the production of IL-22 (a), IL-17A (b) or IL-10 (c) by naive T cells stimulated polyclonally with anti-CD3 and anti-CD28 and cultured for 24 h with recombinant IL-6 with (+) or without (−) recombinant TGF-β, followed by retroviral transduction of control vector (RV GFP) or expression vector for c-Maf (RV c-Maf) and another 48 h of culture, then isolation on the basis of GFP expression and restimulation for 24 h with anti-CD3 and anti-CD28. (d) Intracellular staining for IL-22 and IL-17A in naive T cells cultured for 24 h with anti-CD3 and anti-CD28 in the presence of APCs and recombinant IL-6 and IL-23 plus anti-TGF-β, followed by retroviral transduction of control vector or c-Maf and culture for another 48 h, then restimulation with PMA and ionomycin. Numbers in quadrants indicate percent cells in each, gated on CD4+GFP+ cells. (e) Intracellular staining for IL-22 in naive T cells cultured with anti-CD3 and anti-CD28 and recombinant IL-6 plus neutralizing anti-TGF-β, followed by retroviral transduction of control vector or c-Maf and culture for another 48 h, then restimulation with PMA and ionomycin. Numbers in quadrants indicate percent IL-22+ cells among GFP− cells (left) or GFP+ cells (right), gated on CD4+ cells. *P < 0.001 (Student’s t-test). Data representative of three independent experiments (error bars (a–c), s.e.m.).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature immunology VOLUME 12 NUMBER 12 DECEMBER 2011 1241
A rt i c l e s
Finally, as IL-10 was substantially induced by c-Maf overexpres-sion, we sought to determine whether IL-10 or other secreted factors mediated the c-Maf-dependent repression of IL-22. We compared transduced GFP+ T cells and untransduced GFP− T cells in the same culture (Fig. 3e). Cultures of T cells expressing c-Maf had a much lower frequency of IL-22+ cells (6.7%) than did those of control GFP+ T cells (18.1%), whereas the frequency of IL-22+ cells in cultures of untransfected cells was similar regardless of whether those cells were cultured together with cells transduced with control vector (9.6%) or vector encoding c-Maf (12.3%). These results confirmed the results of the neutralization studies (Fig. 2b) and challenged the idea that a soluble factor such as IL-10 was the main mediator of the IL-22 sup-pression downstream of TGF-β and c-Maf; instead, they suggested a cell-intrinsic mechanism. We concluded that c-Maf was sufficient to suppress IL-22 production in the absence of TGF-β. Although c-Maf slightly enhanced basal IL-17A expression, it was not sufficient to replace TGF-β or drive robust IL-17A production by itself.
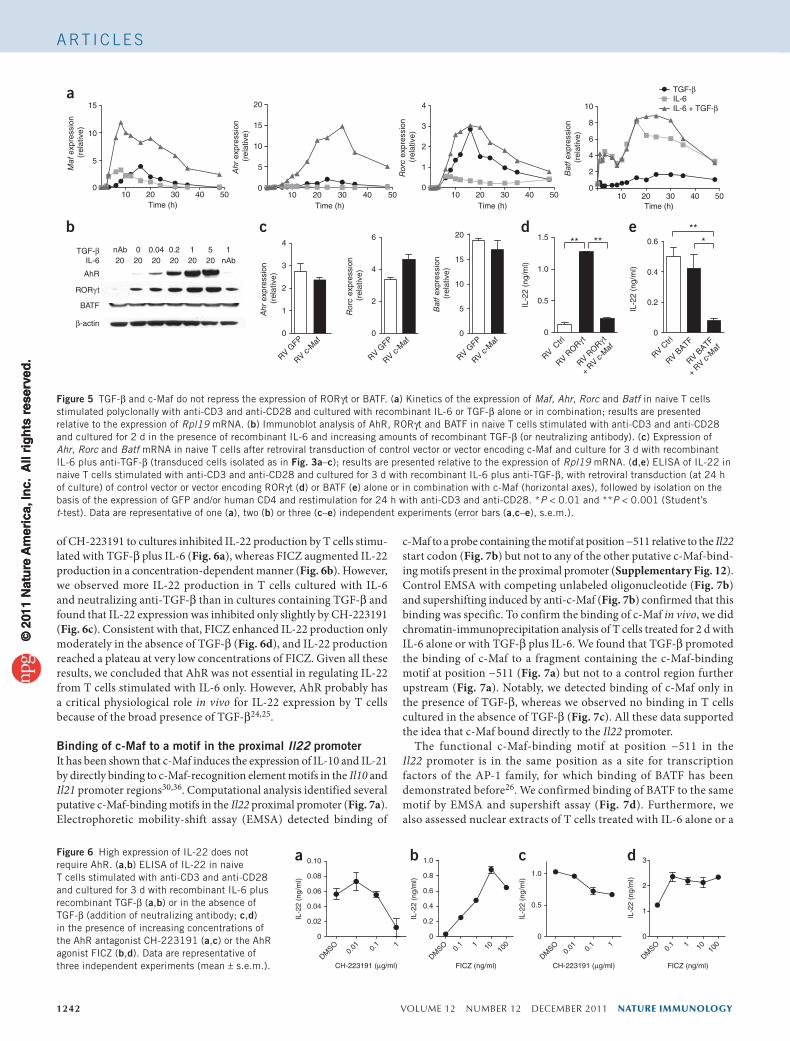
TGF-b-dependent suppression of IL-22 requires c-MafNext we assessed whether c-Maf was necessary for TGF-β-dependent suppression of IL-22. We transfected naive T cells with small interfer-ing RNA (siRNA) targeting c-Maf or a nontargeting control siRNA. We then stimulated T cells with anti-CD3 and anti-CD28 in the pres-ence of IL-6 and TGF-β. Cells transfected with a c-Maf-specific siRNA had much less Maf mRNA and c-Maf protein (Fig. 4a,b). Notably, TGF-β failed to inhibit IL-22 production in TH17 cells when c-Maf expression was knocked down by siRNA (Fig. 4c). In contrast, IL-17A expression was not affected by treatment with c-Maf-specific siRNA (Fig. 4d). Consistent with a published report28, knockdown of c-Maf also resulted in significantly less IL-10 production (Fig. 4e). Finally, expression of the main TH17-associated transcription factors RORγt, AhR and BATF was not affected by treatment with c-Maf-specific siRNA (Supplementary Fig. 8). In conclusion, c-Maf was necessary and sufficient for the TGF-β-dependent suppression of IL-22 in TH17 cells, whereas it had only a minor role in the regulation of IL-17A production in these cells.
Regulation of RORgt, BATF and AhR by TGF-b and c-MafTo understand the cell-intrinsic mechanism by which TGF-β and c-Maf suppressed IL-22 production (Figs. 2b and 3e), we analyzed the kinetics of the expression of c-Maf relative to those of RORγt, AhR and BATF in T cells. First, we found none of these factors were repressed by TGF-β in the presence of IL-6 (Fig. 5a). Although Rorc (the gene encoding RORγt) was induced by TGF-β alone and was
further enhanced by TGF-β plus IL-6, we observed a similar increase in Batf expression with either IL-6 alone or TGF-β plus IL-6. Notably, both TGF-β and IL-6 were required for maximum induction of Maf and Ahr, which confirmed the results we obtained by microarray (Supplementary Table 1). Furthermore, in the presence of TGF-β plus IL-6, Maf was upregulated as early as 4 h after polyclonal stimula-tion and reached maximum expression at 8 h, whereas Ahr expression peaked around 30 h after stimulation.
Next we confirmed those findings at the level of both mRNA and protein at a single time point (48 h) in the presence of IL-6 and vari-ous concentrations of TGF-β (Fig. 5b and Supplementary Fig. 9). Although the expression of BATF was independent of TGF-β, the expression of both RORγt and AhR was positively correlated with the concentration of TGF-β (Fig. 5b and Supplementary Fig. 9). Neither IL-6 nor TGF-β alone was sufficient to induce AhR expression (Fig. 5b and Supplementary Fig. 9a). Finally, retroviral overexpression of c-Maf or siRNA-mediated knockdown of c-Maf in T cells did not suppress Rorc, Batf or Ahr (Fig. 5c and Supplementary Fig. 8). In summary, the TGF-β–c-Maf pathway did not inhibit the production of IL-22 through the suppression of RORγt, AhR or BATF.
As IL-22 production was inhibited despite the presence of RORγt and BATF, we concluded that the suppressive effect of TGF-β–c-Maf was dominant over those positive regulators of IL-22. Accordingly, nei-ther RORγt nor BATF, when overexpressed via retrovirus in the pres-ence of TGF-β, was able to restore IL-22 production (Supplementary Fig. 10a,b), whereas overexpression of RORγt further enhanced IL-17A production (Supplementary Fig. 10c). In the absence of TGF-β, overexpression of RORγt significantly augmented IL-22 expres-sion induced by IL-6. When overexpressed together with RORγt or BATF, c-Maf maintained its suppressive function on IL-22 but further enhanced IL-17A production (Fig. 5d,e and Supplementary Fig. 11), which supported the conclusion that c-Maf suppresses IL-22 by func-tionally interfering with RORγt and/or BATF.
Minimal role of AhR in optimal IL-22 productionGiven the critical role of AhR in the regulation of IL-22 in the pres-ence of IL-6 and TGF-β24,25, it was unexpected to find an inverse correlation between AhR expression and IL-22 production depend-ing on the concentration of TGF-β in the culture. Notably, only trace amounts of AhR were detectable when IL-22 was at its highest expres-sion in T cells stimulated with IL-6 only (Figs. 1a,b and 5a,b). To elucidate the role of such trace amounts of AhR in IL-22 regulation, we modulated AhR activity in vitro with its antagonist CH-223191 and agonist FICZ24,25,30,41. In agreement with published data, the addition
a
0
10
20
30
40
IL-2
2 (n
g/m
l)
siRNA: − − − − −−
IL-1
7A (
ng/m
l)
b
c-Maf
0
5
10
15
2024 h 48 h
β-actin
Maf
exp
ress
ion
(rel
ativ
e)
Maf
exp
ress
ion
(rel
ativ
e)
0
5
10
15
20
25
0
10
20
30
40
50
0
0.2
0.4
0.6
0.8
IL-1
0 (n
g/m
l)
TGF-β: − − − − −−+ + + + + + ++ +
c
siRNA c-
Maf
siRNA C
trl
no si
RNA
c-M
afCtrl–
d e
**
**
** **
Ctrl
Ctrl
c-M
af siRNA:
TGF-β: C
trl C
trl
c-M
af siRNA:
TGF-β: C
trl C
trl
c-M
afsiRNA:
c-M
afCtrl–siRNA:
Figure 4 TGF-β-mediated suppression of IL-22 requires c-Maf. (a,b) Expression of Maf mRNA (a) or c-Maf protein (b) in naive T cells transfected with no siRNA (−), control siRNA (Ctrl) or siRNA targeting c-Maf (c-Maf) and cultured for 24 h or 48 h (a) or 24 h (b) with anti-CD3 and anti-CD28 plus recombinant IL-6 and TGF-β. Results for mRNA are presented relative to the expression of Rpl19 mRNA. (c–e) ELISA of the production of IL-22 (c), IL-17A (d) or IL-10 (e) in naive T cells transfected with siRNA as in a,b and cultured for 72 h with recombinant IL-6 alone (−) or in combination with recombinant TGF-β (+). *P < 0.05 and **P < 0.001 (Student’s t-test). Data are representative of four (a,c–e) or two (b) independent experiments (error bars (a,c–e), s.e.m.).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1242 VOLUME 12 NUMBER 12 DECEMBER 2011 nature immunology
A rt i c l e s
of CH-223191 to cultures inhibited IL-22 production by T cells stimu-lated with TGF-β plus IL-6 (Fig. 6a), whereas FICZ augmented IL-22 production in a concentration-dependent manner (Fig. 6b). However, we observed more IL-22 production in T cells cultured with IL-6 and neutralizing anti-TGF-β than in cultures containing TGF-β and found that IL-22 expression was inhibited only slightly by CH-223191 (Fig. 6c). Consistent with that, FICZ enhanced IL-22 production only moderately in the absence of TGF-β (Fig. 6d), and IL-22 production reached a plateau at very low concentrations of FICZ. Given all these results, we concluded that AhR was not essential in regulating IL-22 from T cells stimulated with IL-6 only. However, AhR probably has a critical physiological role in vivo for IL-22 expression by T cells because of the broad presence of TGF-β24,25.
Binding of c-Maf to a motif in the proximal Il22 promoterIt has been shown that c-Maf induces the expression of IL-10 and IL-21 by directly binding to c-Maf-recognition element motifs in the Il10 and Il21 promoter regions30,36. Computational analysis identified several putative c-Maf-binding motifs in the Il22 proximal promoter (Fig. 7a). Electrophoretic mobility-shift assay (EMSA) detected binding of
c-Maf to a probe containing the motif at position −511 relative to the Il22 start codon (Fig. 7b) but not to any of the other putative c-Maf-bind-ing motifs present in the proximal promoter (Supplementary Fig. 12). Control EMSA with competing unlabeled oligonucleotide (Fig. 7b) and supershifting induced by anti-c-Maf (Fig. 7b) confirmed that this binding was specific. To confirm the binding of c-Maf in vivo, we did chromatin-immunoprecipitation analysis of T cells treated for 2 d with IL-6 alone or with TGF-β plus IL-6. We found that TGF-β promoted the binding of c-Maf to a fragment containing the c-Maf-binding motif at position −511 (Fig. 7a) but not to a control region further upstream (Fig. 7a). Notably, we detected binding of c-Maf only in the presence of TGF-β, whereas we observed no binding in T cells cultured in the absence of TGF-β (Fig. 7c). All these data supported the idea that c-Maf bound directly to the Il22 promoter.
The functional c-Maf-binding motif at position −511 in the Il22 promoter is in the same position as a site for transcription factors of the AP-1 family, for which binding of BATF has been demonstrated before26. We confirmed binding of BATF to the same motif by EMSA and supershift assay (Fig. 7d). Furthermore, we also assessed nuclear extracts of T cells treated with IL-6 alone or a
a
10 20 30 40 500
5
10
15
20
10 20 30 40 500
5
10
15
Maf
exp
ress
ion
(rel
ativ
e)
Ahr
exp
ress
ion
(rel
ativ
e)
TGF-βIL-6IL-6 + TGF-β
Time (h) Time (h)
b
10 20 30 40 500
1
2
3
4
Ror
c ex
pres
sion
(r
elat
ive)
10 20 30 40 500
2
4
6
8
10
Bat
f exp
ress
ion
(rel
ativ
e)
Time (h) Time (h)
AhR
BATF
nAb 0 0.04 0.2 1 5 120 20 20 20 20 20 nAb
β-actin
TGF-βIL-6
RORγ t
0
1
2
3
4
RV GFP
RV c-M
af0
2
4
6
RV GFP
RV c-M
af0
5
10
15
20
RV GFP
RV c-M
af
Ahr
exp
ress
ion
(rel
ativ
e)
Ror
c ex
pres
sion
(r
elat
ive)
Bat
f exp
ress
ion
(rel
ativ
e)
c
0
0.2
0.4
0.6
IL-2
2 (n
g/m
l)
RV Ctrl
RV BATF
RV BATF
+ RV c-
Maf
*
0
0.5
1.0
1.5
IL-2
2 (n
g/m
l)
RV Ctrl
RV RORγ t
RV RORγ t
+
RV c-M
af
** ****d e
Figure 5 TGF-β and c-Maf do not repress the expression of RORγt or BATF. (a) Kinetics of the expression of Maf, Ahr, Rorc and Batf in naive T cells stimulated polyclonally with anti-CD3 and anti-CD28 and cultured with recombinant IL-6 or TGF-β alone or in combination; results are presented relative to the expression of Rpl19 mRNA. (b) Immunoblot analysis of AhR, RORγt and BATF in naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 2 d in the presence of recombinant IL-6 and increasing amounts of recombinant TGF-β (or neutralizing antibody). (c) Expression of Ahr, Rorc and Batf mRNA in naive T cells after retroviral transduction of control vector or vector encoding c-Maf and culture for 3 d with recombinant IL-6 plus anti-TGF-β (transduced cells isolated as in Fig. 3a–c); results are presented relative to the expression of Rpl19 mRNA. (d,e) ELISA of IL-22 in naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 3 d with recombinant IL-6 plus anti-TGF-β, with retroviral transduction (at 24 h of culture) of control vector or vector encoding RORγt (d) or BATF (e) alone or in combination with c-Maf (horizontal axes), followed by isolation on the basis of the expression of GFP and/or human CD4 and restimulation for 24 h with anti-CD3 and anti-CD28. *P < 0.01 and **P < 0.001 (Student’s t-test). Data are representative of one (a), two (b) or three (c–e) independent experiments (error bars (a,c–e), s.e.m.).
ba c
DMSO
0.01 0.
1 1
IL-2
2 (n
g/m
l)
CH-223191 (µg/ml)DM
SO 0.1 1 10 10
0
IL-2
2 (n
g/m
l)
FICZ (ng/ml)DM
SO0.
01 0.1 1
DMSO 0.
1 1 10 100
IL-2
2 (n
g/m
l)
FICZ (ng/ml)
0
1
2
3
0
0.2
0.4
0.6
0.8
1.0
0
0.02
0.04
0.06
0.08
0.10
0
0.5
1.0
IL-2
2 (n
g/m
l)
CH-223191 (µg/ml)
dFigure 6 High expression of IL-22 does not require AhR. (a,b) ELISA of IL-22 in naive T cells stimulated with anti-CD3 and anti-CD28 and cultured for 3 d with recombinant IL-6 plus recombinant TGF-β (a,b) or in the absence of TGF-β (addition of neutralizing antibody; c,d) in the presence of increasing concentrations of the AhR antagonist CH-223191 (a,c) or the AhR agonist FICZ (b,d). Data are representative of three independent experiments (mean ± s.e.m.).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
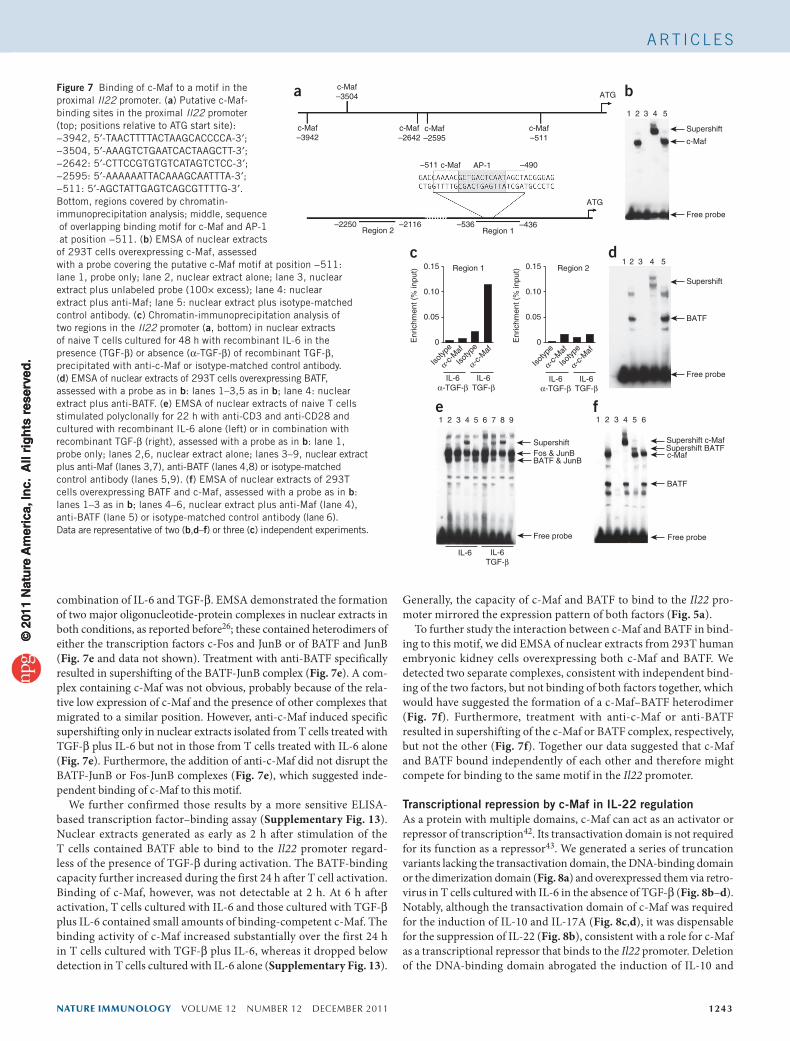
nature immunology VOLUME 12 NUMBER 12 DECEMBER 2011 1243
A rt i c l e s
combination of IL-6 and TGF-β. EMSA demonstrated the formation of two major oligonucleotide-protein complexes in nuclear extracts in both conditions, as reported before26; these contained heterodimers of either the transcription factors c-Fos and JunB or of BATF and JunB (Fig. 7e and data not shown). Treatment with anti-BATF specifically resulted in supershifting of the BATF-JunB complex (Fig. 7e). A com-plex containing c-Maf was not obvious, probably because of the rela-tive low expression of c-Maf and the presence of other complexes that migrated to a similar position. However, anti-c-Maf induced specific supershifting only in nuclear extracts isolated from T cells treated with TGF-β plus IL-6 but not in those from T cells treated with IL-6 alone (Fig. 7e). Furthermore, the addition of anti-c-Maf did not disrupt the BATF-JunB or Fos-JunB complexes (Fig. 7e), which suggested inde-pendent binding of c-Maf to this motif.
We further confirmed those results by a more sensitive ELISA-based transcription factor–binding assay (Supplementary Fig. 13). Nuclear extracts generated as early as 2 h after stimulation of the T cells contained BATF able to bind to the Il22 promoter regard-less of the presence of TGF-β during activation. The BATF-binding capacity further increased during the first 24 h after T cell activation. Binding of c-Maf, however, was not detectable at 2 h. At 6 h after activation, T cells cultured with IL-6 and those cultured with TGF-β plus IL-6 contained small amounts of binding-competent c-Maf. The binding activity of c-Maf increased substantially over the first 24 h in T cells cultured with TGF-β plus IL-6, whereas it dropped below detection in T cells cultured with IL-6 alone (Supplementary Fig. 13).
Generally, the capacity of c-Maf and BATF to bind to the Il22 pro-moter mirrored the expression pattern of both factors (Fig. 5a).
To further study the interaction between c-Maf and BATF in bind-ing to this motif, we did EMSA of nuclear extracts from 293T human embryonic kidney cells overexpressing both c-Maf and BATF. We detected two separate complexes, consistent with independent bind-ing of the two factors, but not binding of both factors together, which would have suggested the formation of a c-Maf–BATF heterodimer (Fig. 7f). Furthermore, treatment with anti-c-Maf or anti-BATF resulted in supershifting of the c-Maf or BATF complex, respectively, but not the other (Fig. 7f). Together our data suggested that c-Maf and BATF bound independently of each other and therefore might compete for binding to the same motif in the Il22 promoter.
Transcriptional repression by c-Maf in IL-22 regulationAs a protein with multiple domains, c-Maf can act as an activator or repressor of transcription42. Its transactivation domain is not required for its function as a repressor43. We generated a series of truncation variants lacking the transactivation domain, the DNA-binding domain or the dimerization domain (Fig. 8a) and overexpressed them via retro-virus in T cells cultured with IL-6 in the absence of TGF-β (Fig. 8b–d). Notably, although the transactivation domain of c-Maf was required for the induction of IL-10 and IL-17A (Fig. 8c,d), it was dispensable for the suppression of IL-22 (Fig. 8b), consistent with a role for c-Maf as a transcriptional repressor that binds to the Il22 promoter. Deletion of the DNA-binding domain abrogated the induction of IL-10 and
c-Maf–3504
c-Maf–3942
c-Maf–2642
c-Maf–2595
–511
–2250 –2116Region 2 Region 1
–536 –436
–490c-Maf AP-1
c-Maf–511
ATG
ATG
a1 2 3 4 5
Supershift
c-Maf
Free probe
b
IL-6TGF-β
IL-6
1 2 3 4 5
1 2 3 4 5 6 1 2 3 4 5 6
Supershift c-MafSupershift BATFc-Maf
BATF
7 8 9
Supershift
SupershiftFos & JunBBATF & JunB
BATF
Free probe
Free probe Free probe
d
e f
0.15 Region 1
Enr
ichm
ent (
% in
put)
Enr
ichm
ent (
% in
put) Region 2
Isotyp
e
Isotyp
e
α-c-M
af
α-c-M
af
Isotyp
e
Isotyp
e
α-c-M
af
α-c-M
af0
IL-6α-TGF-β
IL-6TGF-β
0.05
0.10
0.15
0
0.05
0.10
IL-6α-TGF-β
IL-6TGF-β
c
Figure 7 Binding of c-Maf to a motif in the proximal Il22 promoter. (a) Putative c-Maf- binding sites in the proximal Il22 promoter (top; positions relative to ATG start site): −3942, 5′-TAACTTTTACTAAGCACCCCA-3′; −3504, 5′-AAAGTCTGAATCACTAAGCTT-3′; −2642: 5′-CTTCCGTGTGTCATAGTCTCC-3′; −2595: 5′-AAAAAATTACAAAGCAATTTA-3′; −511: 5′-AGCTATTGAGTCAGCGTTTTG-3′. Bottom, regions covered by chromatin- immunoprecipitation analysis; middle, sequence of overlapping binding motif for c-Maf and AP-1 at position −511. (b) EMSA of nuclear extracts of 293T cells overexpressing c-Maf, assessed with a probe covering the putative c-Maf motif at position −511: lane 1, probe only; lane 2, nuclear extract alone; lane 3, nuclear extract plus unlabeled probe (100× excess); lane 4: nuclear extract plus anti-Maf; lane 5: nuclear extract plus isotype-matched control antibody. (c) Chromatin-immunoprecipitation analysis of two regions in the Il22 promoter (a, bottom) in nuclear extracts of naive T cells cultured for 48 h with recombinant IL-6 in the presence (TGF-β) or absence (α-TGF-β) of recombinant TGF-β, precipitated with anti-c-Maf or isotype-matched control antibody. (d) EMSA of nuclear extracts of 293T cells overexpressing BATF, assessed with a probe as in b: lanes 1–3,5 as in b; lane 4: nuclear extract plus anti-BATF. (e) EMSA of nuclear extracts of naive T cells stimulated polyclonally for 22 h with anti-CD3 and anti-CD28 and cultured with recombinant IL-6 alone (left) or in combination with recombinant TGF-β (right), assessed with a probe as in b: lane 1, probe only; lanes 2,6, nuclear extract alone; lanes 3–9, nuclear extract plus anti-Maf (lanes 3,7), anti-BATF (lanes 4,8) or isotype-matched control antibody (lanes 5,9). (f) EMSA of nuclear extracts of 293T cells overexpressing BATF and c-Maf, assessed with a probe as in b: lanes 1–3 as in b; lanes 4–6, nuclear extract plus anti-Maf (lane 4), anti-BATF (lane 5) or isotype-matched control antibody (lane 6). Data are representative of two (b,d–f) or three (c) independent experiments.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
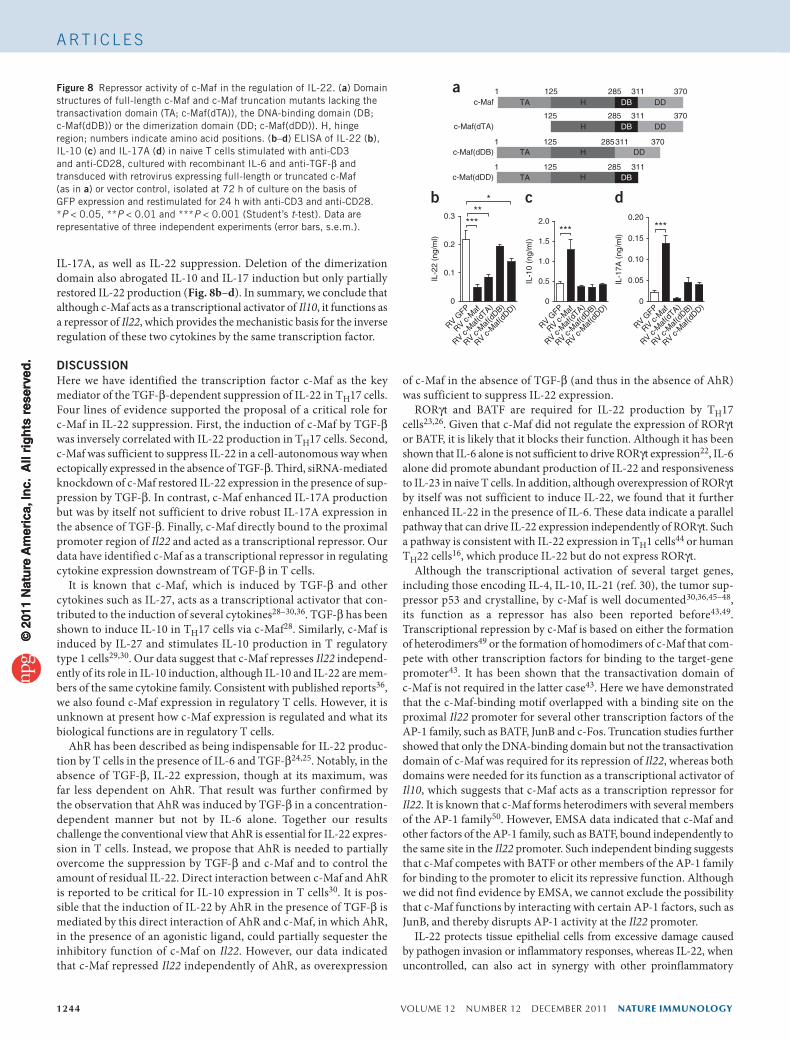
1244 VOLUME 12 NUMBER 12 DECEMBER 2011 nature immunology
A rt i c l e s
IL-17A, as well as IL-22 suppression. Deletion of the dimerization domain also abrogated IL-10 and IL-17 induction but only partially restored IL-22 production (Fig. 8b–d). In summary, we conclude that although c-Maf acts as a transcriptional activator of Il10, it functions as a repressor of Il22, which provides the mechanistic basis for the inverse regulation of these two cytokines by the same transcription factor.
DISCUSSIONHere we have identified the transcription factor c-Maf as the key mediator of the TGF-β-dependent suppression of IL-22 in TH17 cells. Four lines of evidence supported the proposal of a critical role for c-Maf in IL-22 suppression. First, the induction of c-Maf by TGF-β was inversely correlated with IL-22 production in TH17 cells. Second, c-Maf was sufficient to suppress IL-22 in a cell-autonomous way when ectopically expressed in the absence of TGF-β. Third, siRNA-mediated knockdown of c-Maf restored IL-22 expression in the presence of sup-pression by TGF-β. In contrast, c-Maf enhanced IL-17A production but was by itself not sufficient to drive robust IL-17A expression in the absence of TGF-β. Finally, c-Maf directly bound to the proximal promoter region of Il22 and acted as a transcriptional repressor. Our data have identified c-Maf as a transcriptional repressor in regulating cytokine expression downstream of TGF-β in T cells.
It is known that c-Maf, which is induced by TGF-β and other cytokines such as IL-27, acts as a transcriptional activator that con-tributed to the induction of several cytokines28–30,36. TGF-β has been shown to induce IL-10 in TH17 cells via c-Maf28. Similarly, c-Maf is induced by IL-27 and stimulates IL-10 production in T regulatory type 1 cells29,30. Our data suggest that c-Maf represses Il22 independ-ently of its role in IL-10 induction, although IL-10 and IL-22 are mem-bers of the same cytokine family. Consistent with published reports36, we also found c-Maf expression in regulatory T cells. However, it is unknown at present how c-Maf expression is regulated and what its biological functions are in regulatory T cells.
AhR has been described as being indispensable for IL-22 produc-tion by T cells in the presence of IL-6 and TGF-β24,25. Notably, in the absence of TGF-β, IL-22 expression, though at its maximum, was far less dependent on AhR. That result was further confirmed by the observation that AhR was induced by TGF-β in a concentration-dependent manner but not by IL-6 alone. Together our results challenge the conventional view that AhR is essential for IL-22 expres-sion in T cells. Instead, we propose that AhR is needed to partially overcome the suppression by TGF-β and c-Maf and to control the amount of residual IL-22. Direct interaction between c-Maf and AhR is reported to be critical for IL-10 expression in T cells30. It is pos-sible that the induction of IL-22 by AhR in the presence of TGF-β is mediated by this direct interaction of AhR and c-Maf, in which AhR, in the presence of an agonistic ligand, could partially sequester the inhibitory function of c-Maf on Il22. However, our data indicated that c-Maf repressed Il22 independently of AhR, as overexpression
of c-Maf in the absence of TGF-β (and thus in the absence of AhR) was sufficient to suppress IL-22 expression.
RORγt and BATF are required for IL-22 production by TH17 cells23,26. Given that c-Maf did not regulate the expression of RORγt or BATF, it is likely that it blocks their function. Although it has been shown that IL-6 alone is not sufficient to drive RORγt expression22, IL-6 alone did promote abundant production of IL-22 and responsiveness to IL-23 in naive T cells. In addition, although overexpression of RORγt by itself was not sufficient to induce IL-22, we found that it further enhanced IL-22 in the presence of IL-6. These data indicate a parallel pathway that can drive IL-22 expression independently of RORγt. Such a pathway is consistent with IL-22 expression in TH1 cells44 or human TH22 cells16, which produce IL-22 but do not express RORγt.
Although the transcriptional activation of several target genes, including those encoding IL-4, IL-10, IL-21 (ref. 30), the tumor sup-pressor p53 and crystalline, by c-Maf is well documented30,36,45–48, its function as a repressor has also been reported before43,49. Transcriptional repression by c-Maf is based on either the formation of heterodimers49 or the formation of homodimers of c-Maf that com-pete with other transcription factors for binding to the target-gene promoter43. It has been shown that the transactivation domain of c-Maf is not required in the latter case43. Here we have demonstrated that the c-Maf-binding motif overlapped with a binding site on the proximal Il22 promoter for several other transcription factors of the AP-1 family, such as BATF, JunB and c-Fos. Truncation studies further showed that only the DNA-binding domain but not the transactivation domain of c-Maf was required for its repression of Il22, whereas both domains were needed for its function as a transcriptional activator of Il10, which suggests that c-Maf acts as a transcription repressor for Il22. It is known that c-Maf forms heterodimers with several members of the AP-1 family50. However, EMSA data indicated that c-Maf and other factors of the AP-1 family, such as BATF, bound independently to the same site in the Il22 promoter. Such independent binding suggests that c-Maf competes with BATF or other members of the AP-1 family for binding to the promoter to elicit its repressive function. Although we did not find evidence by EMSA, we cannot exclude the possibility that c-Maf functions by interacting with certain AP-1 factors, such as JunB, and thereby disrupts AP-1 activity at the Il22 promoter.
IL-22 protects tissue epithelial cells from excessive damage caused by pathogen invasion or inflammatory responses, whereas IL-22, when uncontrolled, can also act in synergy with other proinflammatory
ac-Maf
3112851 125TA H DB DD
311285125H DB DDc-Maf(dTA)
370
370
c-Maf(dDB)2853111 125
TA H DD370
c-Maf(dDD)3112851 125
TA H DB
b c d
IL-2
2 (n
g/m
l)
***
RV GFP
RV c-M
af
RV c-M
af(d
TA)
RV c-M
af(d
DB)
RV c-M
af(d
DD)0
0.1
0.2
0.3 ***
IL-1
0 (n
g/m
l)
0
0.5
1.0
1.5
2.0
RV GFP
RV c-M
af
RV c-M
af(d
TA)
RV c-M
af(d
DB)
RV c-M
af(d
DD)
***
0
0.05
0.10
0.15
0.20
IL-1
7A (
ng/m
l)
RV GFP
RV c-M
af
RV c-M
af(d
TA)
RV c-M
af(d
DB)
RV c-M
af(d
DD)
***
Figure 8 Repressor activity of c-Maf in the regulation of IL-22. (a) Domain structures of full-length c-Maf and c-Maf truncation mutants lacking the transactivation domain (TA; c-Maf(dTA)), the DNA-binding domain (DB; c-Maf(dDB)) or the dimerization domain (DD; c-Maf(dDD)). H, hinge region; numbers indicate amino acid positions. (b–d) ELISA of IL-22 (b), IL-10 (c) and IL-17A (d) in naive T cells stimulated with anti-CD3 and anti-CD28, cultured with recombinant IL-6 and anti-TGF-β and transduced with retrovirus expressing full-length or truncated c-Maf (as in a) or vector control, isolated at 72 h of culture on the basis of GFP expression and restimulated for 24 h with anti-CD3 and anti-CD28. *P < 0.05, **P < 0.01 and ***P < 0.001 (Student’s t-test). Data are representative of three independent experiments (error bars, s.e.m.).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature immunology VOLUME 12 NUMBER 12 DECEMBER 2011 1245
A rt i c l e s
cytokines, including IL-17A, to amplify proinflammatory responses and cause superfluous tissue remodeling, as has been observed in some autoimmune and inflammatory diseases, such as psoriasis8. The precise regulation of IL-22 production, thus, is essential for maintaining harmony between innate tissue-defense mechanisms and epithelial homeostasis during infection and inflammation in various tissues. For the TGF-β–c-Maf pathway, we have provided a molecular mechanism for the negative regu-lation of IL-22 in TH17 cells. Our data will help to advance the under-standing of the complicated biology of IL-22 in various diseases.
METHODSMethods and any associated references are available in the online version of the paper at http://www.nature.com/natureimmunology/.
Note: Supplementary information is available on the Nature Immunology website.
AUTHOR CONTRIBUTIONSS.R. did most of the experiments and analyzed the data; R.N. contributed to Figures 2, 4 and 5 and Supplementary Figure 4; C.E. contributed to Figures 2 and 5 and Supplementary Figure 1; W.Z. and H.S. contributed to Figure 7; Y.Z. contributed to Figures 1 and 2; N.O. and J.D. cloned c-Maf and RORγt constructs; J.H. analyzed Affymetrix data; W.O. devised and planned the project; and S.R. and W.O. wrote the manuscript.
COMPETING FINANCIAL INTERESTSThe authors declare competing financial interests: details accompany the full-text HTML version of the paper at http://www.nature.com/natureimmunology/.
Published online at http://www.nature.com/natureimmunology/. reprints and permissions information is available online at http://www.nature.com/reprints/index.html.
1. Korn, T., Bettelli, E., Oukka, M. & Kuchroo, V.K. IL-17 and Th17 cells. Annu. Rev. Immunol. 27, 485–517 (2009).
2. Pestka, S. et al. Interleukin-10 and related cytokines and receptors. Annu. Rev. Immunol. 22, 929–979 (2004).
3. Ouyang, W. Distinct roles of IL-22 in human psoriasis and inflammatory bowel disease. Cytokine Growth Factor Rev. 21, 435–441 (2010).
4. Colonna, M. Interleukin-22-producing natural killer cells and lymphoid tissue inducer-like cells in mucosal immunity. Immunity 31, 15–23 (2009).
5. Ye, P. et al. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense. J. Exp. Med. 194, 519–527 (2001).
6. Aujla, S. et al. IL-22 mediates mucosal host defense against Gram-negative bacterial pneumonia. Nat. Med. 14, 275–281 (2008).
7. Liang, S.C. et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 203, 2271–2279 (2006).
8. Zheng, Y. et al. Interleukin-22, a TH17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature 445, 648–651 (2007).
9. Guilloteau, K. et al. Skin inflammation induced by the synergistic action of IL-17A, IL-22, oncostatin M, IL-1α, and TNF-α recapitulates some features of psoriasis. J. Immunol. 184, 5263–5270 (2010).
10. Zenewicz, L.A. et al. Interleukin-22 but not interleukin-17 provides protection to hepatocytes during acute liver inflammation. Immunity 27, 647–659 (2007).
11. Zheng, Y. et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat. Med. 14, 282–289 (2008).
12. Sonnenberg, G.F. et al. Pathological versus protective functions of IL-22 in airway inflammation are regulated by IL-17A. J. Exp. Med. 207, 1293–1305 (2010).
13. Nograles, K.E. et al. Th17 cytokines interleukin (IL)-17 and IL-22 modulate distinct inflammatory and keratinocyte-response pathways. Br. J. Dermatol. 159, 1092–1102 (2008).
14. Ouyang, W., Kolls, J. & Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 28, 454–467 (2008).
15. Sanjabi, S., Zenewicz, L.A., Kamanaka, M. & Flavell, R.A. Anti-inflammatory and pro-inflammatory roles of TGF-β, IL-10, and IL-22 in immunity and autoimmunity. Curr. Opin. Pharmacol. 9, 447–453 (2009).
16. Duhen, T., Geiger, R., Jarrossay, D., Lanzavecchia, A. & Sallusto, F. Production of interleukin 22 but not interleukin 17 by a subset of human skin-homing memory T cells. Nat. Immunol. 10, 857–863 (2009).
17. Trifari, S., Kaplan, C., Tran, E., Crellin, N. & Spits, H. Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from TH-17, TH1 and TH2 cells. Nat. Immunol. 10, 864–871 (2009).
18. Broadhurst, M.J. et al. IL-22+ CD4+ T Cells are associated with therapeutic Trichuris trichiura infection in an ulcerative colitis patient. Science Transl. Med. 2, 60ra88 (2010).
19. Veldhoen, M., Hocking, R.J., Atkins, C.J., Locksley, R.M. & Stockinger, B. TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
20. Bettelli, E. et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441, 235–238 (2006).
21. Mangan, P.R. et al. Transforming growth factor-β induces development of the TH17 lineage. Nature 441, 231–234 (2006).
22. Ivanov, I.I. et al. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 126, 1121–1133 (2006).
23. Yang, X.O. et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors RORα and RORγ. Immunity 28, 29–39 (2008).
24. Veldhoen, M. et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 453, 106–109 (2008).
25. Veldhoen, M., Hirota, K., Christensen, J., O’Garra, A. & Stockinger, B. Natural agonists for aryl hydrocarbon receptor in culture medium are essential for optimal differentiation of Th17 T cells. J. Exp. Med. 206, 43–49 (2009).
26. Schraml, B. et al. The AP-1 transcription factor Batf controls TH17 differentiation. Nature 460, 405–409 (2009).
27. Kim, J.I., Ho, I.C., Grusby, M.J. & Glimcher, L.H. The transcription factor c-Maf controls the production of interleukin-4 but not other Th2 cytokines. Immunity 10, 745–751 (1999).
28. Xu, J. et al. c-Maf regulates IL-10 expression during Th17 polarization. J. Immunol. 182, 6226–6236 (2009).
29. Pot, C. et al. Cutting edge: IL-27 induces the transcription factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote differentiation of IL-10-producing Tr1 cells. J. Immunol. 183, 797–801 (2009).
30. Apetoh, L. et al. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nat. Immunol. 11, 854–861 (2010).
31. Cao, S., Liu, J., Song, L. & Ma, X. The protooncogene c-Maf is an essential transcription factor for IL-10 gene expression in macrophages. J. Immunol. 174, 3484–3492 (2005).
32. Ghoreschi, K. et al. Generation of pathogenic TH17 cells in the absence of TGF-β signalling. Nature 467, 967–971 (2010).
33. Nurieva, R. et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 448, 480–483 (2007).
34. Veldhoen, M. et al. Transforming growth factor-β ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 9, 1341–1346 (2008).
35. McGeachy, M. et al. TGF-β and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain TH-17 cell–mediated pathology. Nat. Immunol. 8, 1390–1397 (2007).
36. Hiramatsu, Y. et al. c-Maf activates the promoter and enhancer of the IL-21 gene, and TGF-β inhibits c-Maf-induced IL-21 production in CD4+ T cells. J. Leukoc. Biol. 87, 703–712 (2009).
37. Gu, Y. et al. Interleukin 10 suppresses Th17 cytokines secreted by macrophages and T cells. Eur. J. Immunol. 38, 1807–1813 (2008).
38. Dumoutier, L., Louahed, J. & Renauld, J.C. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. J. Immunol. 164, 1814–1819 (2000).
39. Yang, Y., Ochando, J., Yopp, A., Bromberg, J.S. & Ding, Y. IL-6 plays a unique role in initiating c-Maf expression during early stage of CD4 T cell activation. J. Immunol. 174, 2720–2729 (2005).
40. Chen, W. et al. Conversion of peripheral CD4+CD25− naive T cells to CD4+CD25+ regulatory T cells by TGF-β induction of transcription factor Foxp3. J. Exp. Med. 198, 1875–1886 (2003).
41. Quintana, F.J. et al. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
42. Yang, Y. & Cvekl, A. Large Maf transcription factors: cousins of AP-1 proteins and important regulators of cellular differentiation. Einstein J. Biol. Med. 23, 2–11 (2007).
43. Dhakshinamoorthy, S. & Jaiswal, A.K. c-Maf negatively regulates ARE-mediated detoxifying enzyme genes expression and anti-oxidant induction. Oncogene 21, 5301–5312 (2002).
44. Gurney, A.L. IL-22, a Th1 cytokine that targets the pancreas and select other peripheral tissues. Int. Immunopharmacol. 4, 669–677 (2004).
45. Ho, I.C., Hodge, M.R., Rooney, J.W. & Glimcher, L.H. The proto-oncogene c-maf is responsible for tissue-specific expression of interleukin-4. Cell 85, 973–983 (1996).
46. Hale, T.K. et al. Maf transcriptionally activates the mouse p53 promoter and causes a p53-dependent cell death. J. Biol. Chem. 275, 17991–17999 (2000).
47. Chauhan, B.K., Yang, Y., Cveklová, K. & Cvekl, A. Functional interactions between alternatively spliced forms of Pax6 in crystallin gene regulation and in haploinsufficiency. Nucleic Acids Res. 32, 1696–1709 (2004).
48. Yang, Y., Chauhan, B.K., Cveklova, K. & Cvekl, A. Transcriptional regulation of mouse αB- and γF-crystallin genes in lens: opposite promoter-specific interactions between Pax6 and large Maf transcription factors. J. Mol. Biol. 344, 351–368 (2004).
49. Hedge, S.P., Kumar, A., Kurschner, C. & Shapiro, L.H. c-Maf interacts with c-Myb to regulate transcription of an early myeloid gene during differentiation. Mol. Cell. Biol. 18, 2729–2737 (1998).
50. Kataoka, K., Noda, M. & Nishizawa, M. Maf nuclear oncoprotein recognizes sequences related to an AP-1 site and forms heterodimers with both Fos and Jun. Mol. Cell. Biol. 14, 700–712 (1994).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature immunology doi:10.1038/ni.2134
ONLINE METHODSMice. BALB/c mice from Jackson Laboratories were housed under specific pathogen– free conditions and used at 8–12 weeks of age. All animal experiments were approved by the Genentech Institutional Animal Care and Use Committee.
Plasmids. Sequence of the full-length c-Maf coding region or fragments encod-ing amino acids 125–370 (lacking the transactivation domain) or 1–311 (lack-ing the dimerization domain) were amplified by PCR from a splenic cDNA library, with the introduction of a Kozak sequence and a start codon (trans-activation mutant) or a stop codon (dimerization mutant) and were cloned into retroviral expression vector pMSCV-IRES-GFP or pMSCV-IRES-hCD4 at BglII and XhoI sites. The c-Maf mutant with deletion of the DNA-binding domain (amino acids 285–311) was generated by site-directed mutagenesis with the QuikChange XL kit according to the manufacturer’s protocol (Agilent Stratagene). The coding regions of Rorc and Batf were amplified by PCR, with the introduction of a Kozak sequence, and the fragments were cloned into retroviral expression vector pMSCV-IRES-GFP or pMSCV-IRES-hCD4 at XhoI–EcoRI or BglII–XhoI sites.
T cell purification. Naive CD4+ T cells were purified by pre-enrichment of CD4+ T cells from spleens and lymph nodes by magnetic-activated cell sort-ing (CD4 MicroBeads; Miltenyi Biotec) and sorting by flow cytometry for CD4+CD62L+CD25− T cells (FACSAria; BD Biosciences) after staining with anti-CD4 (RM4-5), anti-CD62L (MEL-14), anti-CD25 (PC61; all from BD Biosciences). Alternatively, naive T cells were purified by magnetic-activated cell sorting. First, cells from spleens and lymph nodes were stained with biotin-conjugated anti-CD25 (7D4) and fluorescein isothiocyanate–conjugated anti-CD4 (RM4-5; both from BD Biosciences), and samples were depleted of CD25+ cells with Anti-Biotin MicroBeads (Miltenyi Biotec). Next, CD4+ T cells were isolated with Anti-FITC MultiSort MicroBeads (Miltenyi Biotec). After release of beads, CD62L+CD4+ T cells were purified with CD62L MicroBeads (Miltenyi Biotec). Both procedures resulted in a similar purity of >97%. Anti-MHC Class II MicroBeads (Miltenyi Biotec) were used for the isolation of APCs from fractions depleted of CD4+ cells, then APCs were irradiated at 30 Gy. Regulatory T cells were sorted by flow cytometry according to their expression of CD4 and CD25.
T cell cultures. All cultures were incubated in DMEM with 10% (vol/vol) FCS (HyClone), 2 mM l-glutamine, 1 mM sodium pyruvate, 0.1 mM non-essential amino acids, 55 µM β-mercaptoethanol, 100 U/ml of penicillin and 100 µg/ml of streptomycin (all from Invitrogen). For APC cocultures, CD4+CD25−CD62L+ naive T cells from BALB/c mice were stimulated at a ratio of 1:4 with irradiated APCs (positive for major histocompatibility complex class II) in the presence of anti-CD3 (0.5 µg/ml; 145-2C11) and anti-CD28 (1 µg/ml; 37.51; both from BD Biosciences) under IL-17- or IL-22-induc-ing conditions. For ‘plate-bound’ cultures, plates were coated overnight with anti-CD3 (5 µg/ml in PBS). Anti-CD28 (1 µg/ml) was added to naive T cells cultured under the following conditions: IL-17 induction, anti-IL-4 (10 µg/ml; 11B11; BD Biosciences), anti-IFN-γ (10 µg/ml; XMG1.2; BD Biosciences), recombinant IL-6 (20 ng/ml), recombinant IL-23 (10 ng/ml) and recom-binant human TGF-β (1 ng/ml; R&D Systems); IL-22 induction, anti-IL-4 (10 µg/ml), anti-IFN-γ (10 µg/ml), recombinant IL-6 (20 ng/ml), anti-TGF-β (10 µg/ml; 1D11; R&D Systems); induced regulatory T cell polarization, anti-IL-4 (10 µg/ml), anti-IFN-γ (10 µg/ml), recombinant human TGF-β (5 ng/ml) and recombinant IL-2 (2 ng/ml; R&D Systems). FICZ (Enzo Life Sciences) or CH-223191 (Calbiochem) was added to some cultures at various concentra-tions. Neutralizing anti-IL-9 (D9302C12; BD Biosciences), anti-IL-10 (JES5-2A5; BD Biosciences) or anti-IL-21 (FFA21; eBioscience) was added to some cultures at a concentration of 10 µg/ml.
Knockdown via siRNA. Chemically modified siRNA51 was synthesized in-house as follows (where mC, mU, mA, mT or mG indicates a methoxy nucle-otide, and dG, dA, dT or dC indicates a deoxy nucleotide): siRNA specific for cMaf, 5′-mCmUGAUGAAGUUUGAAGUdGdAdAmAmAmA-3′ (sense) or 5′-dTUUCACUUCAAACUUCAUCmAmG-3′ (antisense); and control siRNA, 5′-GmGmAGCGCACCAUCUUCdTdCdAmAmTmT-3′(sense) or 5′-dTUGAGAAGAUGGUGCGCUmCmC-3′ (antisense).
Naive T cells (3 × 106) were transfected with 600 pmol siRNA in mouse T cell nucleofection solution through the use of program X-001 on a Nucleofector (Lonza). Cells were transferred directly into DMEM with 10% (vol/vol) FCS, 2 mM l-glutamine, 1 mM sodium pyruvate, 0.1 mM nonessential amino acids, 55 µM β-mercaptoethanol, 100 U/ml of penicillin and 100 µg/ml of streptomycin, were allowed to ‘rest’ for 2 h and then were cultured and stimulated.
Retroviral transduction. For retroviral transduction of CD4+ T cells, Phoenix E cells were transiently transfected by the calcium-phosphate method with retroviral expression plasmids together with the retroviral packaging plasmid pEco. After 48 h, viral supernatants were collected, filtered through a 0.45-µm filter, supplied with 10 mM HEPES, pH 7.2, and Polybrene (10 µg/ml) and added to T cells that had been stimulated polyclonally and cultured for 24 h. T cells (2.5 × 106) were incubated in 12-well plates with 1.5 ml viral supernatant and were centrifuged for 75 min at 700g at 32 °C. Thereafter, viral supernatant was replaced by conditioned culture medium.
Affymetrix microarray. Samples were hybridized to Affymetrix Mouse Genome 430 2.0 microarrays and processed according to the manufactur-er’s protocols (Affymetrix). Raw data were normalized with the Micrarray Suite 5 algorithm, and then transformed (log2) for further analysis. Probes were ranked by the difference (log2) between expression in cells treated with recombinant IL-6 alone and expression in those treated with recombinant IL-6 and TGF-β, and the probes with the greatest difference were selected for further analysis.
Restimulation and intracellular staining. T cells were stimulated for 4 h with PMA (phorbol 12-myristate 13-acetate; 10 ng/ml; Sigma) and ionomycin (1 µg/ml; Invitrogen). Brefeldin A (eBioscience) was added at a concentration of 5 µg/ml. After fixation with 2% (vol/vol) paraformaldehyde, cells were made permeable in Cytoperm buffer (BD Biosciences) and were stained intracellu-larly for 20 min at 20 °C with anti-IL-22 (1H8PWSR; eBioscience), anti-IL-17 (TC11-18H10; eBioscience) or anti-CD4 (BD Biosciences). Samples were acquired on a FACSCalibur (BD Biosciences), and FlowJo (TreeStar) was used for data analysis.
ELISA. DuoSet ELISA kits were used according to manufacturer’s protocol (R&D Systems) for ELISA of IL-4, IL-10, IL-17 and IL-21, and a Ready-Set-Go kit was used according to the manufacturer’s protocol (eBioscience) for ELISA of IL-9. The ELISA for IL-22 was developed ‘in-house’ and was done as described8.
Real-time RT-PCR. Samples were analyzed by real-time RT-PCR with TaqMan One-Step RT-PCR Master Mix reagents (Applied Biosystems) and the appropriate primers and probes (Supplementary Table 2). Results normalized to those of the control housekeeping gene encoding ribos-omal protein L19 were calculated according to the change in cycling threshold (∆CT) as follows: ∆∆CT = sample ∆CT − reference ∆CT.
Immunoblot analysis. Whole-cell lysates were prepared from 2 × 106 cells in radioimmunoprecipitation buffer: 50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% (vol/vol) Triton-X-100 and 0.25% (vol/vol) deoxycholate with Complete Protease Inhibitor (Roche). Proteins (20 µg per lane, measured by bicinchoninic acid assay (Pierce)) were separated by SDS-PAGE through 10% (vol/vol) Novex Tris-Glycine gels (Invitrogen), transferred to nitrocellulose membranes and analyzed by immunoblot. Membranes were blocked for 30 min at 20 °C in 5% (vol/vol) milk in TBST (10 mM Tris, pH 7.5, 150 mM NaCl, 0.1% (vol/vol) Tween-20) to minimize nonspecific binding, were probed overnight at 4 °C in blocking buffer with anti-c-Maf (M-153; Santa Cruz Biotechnology), rabbit polyclonal anti-AhR (PA1-515; Thermo Scientific), anti-RORγt (B2D; eBioscience), anti-BATF (8A12; Sigma Aldrich) or rabbit polyclonal anti-β-actin (A2066; Sigma-Aldrich), washed extensively in TBST, and incubated at 20 °C with horseradish peroxidase–conjugated anti-rabbit (7074) or anti-mouse (7076; both from Cell Signaling) or anti–rat horseradish peroxidase (sc-2006; Santa Cruz Biotechnology). Proteins were visualized with SuperSignal West Pico chemiluminescent substrate (Pierce).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature immunologydoi:10.1038/ni.2134
Chromatin immunoprecipitation. T cells (2 × 107) were crosslinked for 10 min with 1% (vol/vol) paraformaldehyde. Nuclei were isolated and subjected to sonication with Covaris for 25 min (20% duty cycle; intensity, 5; cycles per burst, 200). DNA fragmentation was monitored by agarose gel electrophoresis and was typically in the range of 0.2–0.5 kilobases. Anti-c-Maf (5 µg; M-153; Santa Cruz Biotechnology) or control immunoglobulin G (sc-2027; Santa Cruz Biotechnology) and Protein A/G Plus UltraLink Resin (immobilized protein A and G; Thermo Scientific) were used for immunoprecipitation. Precipitated DNA was analyzed by 40 cycles of quantitative PCR (primer sets, Supplementary Table 2). Immunoprecipitation efficiency was calculated relative to input.
EMSA. Nuclear extracts of 293T cells or TH17 cells were prepared with the NE-PER Nuclear and Cytoplasmic Extraction Kit (Thermo Scientific). A LightShift Chemiluminescent EMSA kit was used for EMSA according to the manufacturer’s protocol (Thermo Scientific). DNA–nuclear extract binding reactions were done as follows: nuclear extracts (3 µg) were preincubated for 10 min on ice with 1× binding buffer (Thermo Scientific), 50 ng/µl of poly (dI:dC), 2.5% (vol/vol) glycerol, 0.05% (vol/vol) Nonidet P-40, 50 mM KCl and 5 mM MgCl2. In some experiments, antibodies for supershifting (1 µg each) were added to the reactions (anti-c-Maf (NB600-267; Novus Biologicals) and anti-BATF (8A12; Sigma-Aldrich)). In some experiments, a 100-fold molar excess of nonbiotinylated competitor oligonucleotide was then added to each reaction, followed by incubation for 5 min at 20 °C. After that incubation,
biotin-labeled oligonucleotides (Supplementary Table 2) were added and reactions were incubated for 20 min at 20 °C. The 5′-biotinylated sense and antisense oligonucleotides were annealed by heating (of equimolar amounts in 100 mM NaCl and 10 mM Tris, pH 7.4) to 96 °C, followed by slow cooling to 20 °C. Annealed oligonucleotides were purified from a 3% agarose gel with a Gel Extraction kit (Qiagen).
Transcription factor–binding assay. Plates were coated overnight with anti-Maf (NB600-267; Novus Biologicals) or anti-BATF (8A12; Sigma-Aldrich) at a concentration of 5 µg/ml each in PBS. After blockade of nonspecific binding with 1% (wt/vol) BSA in PBS, samples containing 2 µg nuclear extracts, 5 nM biotinylated oligonucleotide (probe IL-22pro-511, which covers the putative c-Maf motif at position −511) and EMSA binding buffer that had been prein-cubated for 30 min at 20 °C, were added, followed by incubation for 2 h. After samples were washed and then incubated for 1 h with streptavidin–horseradish peroxidase, bound oligonucleotides were detected as described above for ELISA.
Statistical analysis. Statistical significance was calculated by the unpaired t-test with the Prism software (GraphPad).
51. Mantei, A. et al. siRNA stabilization prolongs gene knockdown in primary T lymphocytes. Eur. J. Immunol. 38, 2616–2625 (2008).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.©
201
1 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
Related Documents











