Please cite this article in press as: Börschig, C., et al. Traits of butterfly communities change from specialist to generalist characteristics with increasing land-use intensity. Basic and Applied Ecology (2013), http://dx.doi.org/10.1016/j.baae.2013.09.002 ARTICLE IN PRESS BAAE-50733; No. of Pages 8 Basic and Applied Ecology xxx (2013) xxx–xxx Traits of butterfly communities change from specialist to generalist characteristics with increasing land-use intensity Carmen Börschig a,∗ , Alexandra-Maria Klein b,c , Henrik von Wehrden b,d,e , Jochen Krauss f a Agroecology, University of Göttingen, Grisebachstrasse 6, 37075 Göttingen, Germany b Institute of Ecology, Leuphana University of Lüneburg, Scharnhorstrasse 1, 21335 Lüneburg, Germany c Institute of Earth and Environmental Sciences, Chair of Nature Conservation and Landscape Ecology, University of Freiburg, Tennenbach- erstrasse 4, 79106 Freiburg, Germany d Centre for Methods, Leuphana University of Lüneburg, Scharnhorstrasse 1, 21335 Lüneburg, Germany e Research Institute of Wildlife Ecology, Savoyen Strasse 1, Vienna 1160, Austria f Department of Animal Ecology and Tropical Biology, Biocentre, University of Würzburg, Am Hubland, 97074 Würzburg, Germany Received 29 April 2013; accepted 2 September 2013 Abstract Land-use intensification leads to species loss and shifts in community composition, but only few studies examine how these dynamics affect ecological and life-history traits. We thus investigated whether ecological and life-history traits differ between butterfly communities of grasslands with different land-use intensity. We conducted butterfly transect surveys in 137 grassland sites in three regions of Germany and compiled 10 species-specific ecological and life-history traits from the literature. These traits are associated with food plant specialisation, dispersal, distribution, reproduction and development. We calculated a land- use intensity gradient based on the amount of fertilise mowing frequency and grazing intensity. We analysed differences of traits characteristics between butterfly communities along the land-use intensity gradient in a fourth-corner analysis, thus considering correlations between traits. Six ecological and life-history traits changed from characteristics associated with specialists to such associated with generalists with increasing land-use intensity. These traits characteristics in intensified grasslands were: high dispersal propensity, large distribution range, low population density, more than one generation per year, hibernation in a more advanced developmental stage and a long flight period. The functional homogenisation of the butterfly communities with changes from specialist to generalist trait characteristics with increasing land-use intensity may have severe consequences for ecosystem functioning and services. Zusammenfassung Landnutzungsintensivierung führt zu Artenverlust und zu Verschiebungen der Artenzusammensetzung innerhalb von Lebens- gemeinschaften, aber nur wenige Studien berücksichtigen biologische und ökologische Merkmale in Bezug auf diese Prozesse. Wir untersuchten daher, ob sich Schmetterlingslebensgemeinschaften auf Grünlandflächen unterschiedlicher Landnutzungsin- tensität hinsichtlich ihrer biologischen und ökologischen Merkmale unterscheiden. Wir sind in drei Regionen Deutschlands auf 137 Grünlandflächen Transekte zum Zählen von Schmetterlingen gelaufen. Wir stellten Angaben zur Nahrungsspezia- lisierung, Ausbreitung, Verbreitung, Reproduktion und Entwicklung der beobachteten Schmetterlingsarten aus der Literatur ∗ Corresponding author. Tel.: +49 551 393739; fax: +49 551 398806. E-mail addresses: [email protected], [email protected] (C. Börschig). 1439-1791/$ – see front matter © 2013 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved. http://dx.doi.org/10.1016/j.baae.2013.09.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

B
Tc
C
a
b
c
ed
e
f
R
A
dbstuccshmce
Z
gWtal
1h
ARTICLE IN PRESSAAE-50733; No. of Pages 8
Basic and Applied Ecology xxx (2013) xxx–xxx
raits of butterfly communities change from specialist to generalistharacteristics with increasing land-use intensity
armen Börschiga,∗, Alexandra-Maria Kleinb,c, Henrik von Wehrdenb,d,e, Jochen Kraussf
Agroecology, University of Göttingen, Grisebachstrasse 6, 37075 Göttingen, GermanyInstitute of Ecology, Leuphana University of Lüneburg, Scharnhorstrasse 1, 21335 Lüneburg, GermanyInstitute of Earth and Environmental Sciences, Chair of Nature Conservation and Landscape Ecology, University of Freiburg, Tennenbach-rstrasse 4, 79106 Freiburg, GermanyCentre for Methods, Leuphana University of Lüneburg, Scharnhorstrasse 1, 21335 Lüneburg, GermanyResearch Institute of Wildlife Ecology, Savoyen Strasse 1, Vienna 1160, AustriaDepartment of Animal Ecology and Tropical Biology, Biocentre, University of Würzburg, Am Hubland, 97074 Würzburg, Germany
eceived 29 April 2013; accepted 2 September 2013
bstract
Land-use intensification leads to species loss and shifts in community composition, but only few studies examine how theseynamics affect ecological and life-history traits. We thus investigated whether ecological and life-history traits differ betweenutterfly communities of grasslands with different land-use intensity. We conducted butterfly transect surveys in 137 grasslandites in three regions of Germany and compiled 10 species-specific ecological and life-history traits from the literature. Theseraits are associated with food plant specialisation, dispersal, distribution, reproduction and development. We calculated a land-se intensity gradient based on the amount of fertilise mowing frequency and grazing intensity. We analysed differences of traitsharacteristics between butterfly communities along the land-use intensity gradient in a fourth-corner analysis, thus consideringorrelations between traits. Six ecological and life-history traits changed from characteristics associated with specialists touch associated with generalists with increasing land-use intensity. These traits characteristics in intensified grasslands were:igh dispersal propensity, large distribution range, low population density, more than one generation per year, hibernation in aore advanced developmental stage and a long flight period. The functional homogenisation of the butterfly communities with
hanges from specialist to generalist trait characteristics with increasing land-use intensity may have severe consequences forcosystem functioning and services.
usammenfassung
Landnutzungsintensivierung führt zu Artenverlust und zu Verschiebungen der Artenzusammensetzung innerhalb von Lebens-
Please cite this article in press as: Börschig, C., et al. Traits of butterfly communities change from specialist to generalist characteristicswith increasing land-use intensity. Basic and Applied Ecology (2013), http://dx.doi.org/10.1016/j.baae.2013.09.002
emeinschaften, aber nur wenige Studien berücksichtigen biologische und ökologische Merkmale in Bezug auf diese Prozesse.ir untersuchten daher, ob sich Schmetterlingslebensgemeinschaften auf Grünlandflächen unterschiedlicher Landnutzungsin-
ensität hinsichtlich ihrer biologischen und ökologischen Merkmale unterscheiden. Wir sind in drei Regionen Deutschlandsuf 137 Grünlandflächen Transekte zum Zählen von Schmetterlingen gelaufen. Wir stellten Angaben zur Nahrungsspezia-isierung, Ausbreitung, Verbreitung, Reproduktion und Entwicklung der beobachteten Schmetterlingsarten aus der Literatur
∗Corresponding author. Tel.: +49 551 393739; fax: +49 551 398806.E-mail addresses: [email protected], [email protected] (C. Börschig).
439-1791/$ – see front matter © 2013 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.baae.2013.09.002

B
2
zMdKmzglms©
K
I
a&otttKwltithotettT2tmnta2p&
BldTltotm
ARTICLE IN PRESSAAE-50733; No. of Pages 8
C. Börschig et al. / Basic and Applied Ecology xxx (2013) xxx–xxx
usammen. Wir berechneten einen Landnutzungsintensitätsgradienten basierend auf der Menge des genutzten Düngers, derahdhäufigkeit, und der Beweidungsintensität. Wir errechneten die Unterschiede der Ausprägungen der Merkmale zwischen
en Schmetterlingslebensgemeinschaften entlang des Landnutzungsgradienten mit Hilfe einer Fourth Corner Analyse, dieorrelationen zwischen den Merkmalen berücksichtigt. Die biologischen und ökologischen Merkmale der Schmetterlingsge-einschaften verschoben sich mit zunehmender Landnutzungsintensität von Merkmalen, die mit Spezialisten assoziiert sind,
u solchen, die für Generalisten charakteristisch sind. Dazu gehörten: hohes Dispersionsverhalten, großes Verbreitungsareal,eringe Populationsdichte, Produktion von mehr als einer Generation pro Jahr, Überwinterung in einem höheren Entwick-ungsstadium und eine ausgedehntere Flugperiode. Die funktionelle Homogenisierung von Schmetterlingslebensgemeinschaften
it zunehmender Landnutzungsintensität kann schwerwiegende Konsequenzen für Ökosystemfunktionen und Ökosystemdien-tleistungen haben.
2013 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.
rasslan
sh(tlcsgbfSbspw(wpica&wteaFcfsera
eywords: Development; Dispersal; Distribution; Flight period; G
ntroduction
Almost 40% of the terrestrial ice-free land is covered bygroecosystems (Ellis, Klein Goldewijk, Siebert, Lightman,
Ramankutty 2010) and increasing land-use intensity is onef the main drivers of biodiversity loss (Sala et al., 2000). Dueo different ecological and life-history traits (hereafter termedraits), which evolved under distinct environmental condi-ions, some species might suffer disadvantages (Kotiaho,aitala, Komonen, & Päivinen 2005; Öckinger et al., 2010)hile other species might even benefit from increasing
and-use intensity (McKinney & Lockwood, 1999). The iden-ification of disadvantageous traits associated with land-usentensity allows predictions about (1) community responseso future intensification and (2) identification of species with aigh extinction risk (Kotiaho et al., 2005). The consequencesf increasing land-use intensity for ecological and life-historyraits are relatively well studied for bees (e.g. Williamst al. 2010), but the majority of butterfly studies on speciesraits have focused only on the effects of habitat fragmen-ation, isolation and habitat area loss (Steffan-Dewenter &scharntke 2000; Brückmann, Krauss, & Steffan-Dewenter010; Öckinger et al. 2010). Studies on multiple butterflyraits along a land-use intensity gradient in grasslands areissing to date. This is surprising given that butterflies are
ot only a relatively large and taxonomically well knownaxa, but also indicators of environmental change and over-ll biodiversity (Thomas 2005; van Swaay, Warren, & Loïs006). Additionally, comprehensive information about Euro-ean butterfly species traits is available (Settele, Feldmann,
Reinhardt 1999; Thomas 2005).Grasslands are vital habitats for many arthropods (Diacon-
olli, Dalang, Holderegger, & Bürgi 2012), but increasingand-use intensity in grasslands leads to decreasing speciesiversity (Tscharntke, Klein, Kruess, Steffan-Dewenter, &hies 2005). Arthropods in extensively managed grasslands
ive in diverse and often stable habitats with low perturba-ion. Arthropods in intensively managed grasslands on the
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
ther hand have access to only few host plants, and haveo cope with high habitat disturbance frequencies due toowing or grazing (Kruess & Tscharntke 2002). Habitat
(
d; Life-history traits; Population density; Reproduction; Voltinism
tability in space and time favours specialised species, whileabitats with high disturbances favour generalist speciesClavel, Julliard, & Devictor 2010). We therefore assumehat traits of butterfly communities change with increasingand-use intensity from characteristics associated with spe-ialists to characteristics associated with generalists. In ourtudy we chose traits relevant for survival and competition inrasslands. Generally butterfly specialists are characterisedy a narrow feeding niche whereas generalists show a broadeeding niche (e.g. Steffan-Dewenter & Tscharntke 2000).pecialised butterfly larvae can feed on plants with high her-ivore defence mechanisms, which assures a food sourcehared with only few competing species in highly com-etitive extensive grasslands. A broad feeding niche ratherarrants food plant availability in highly disturbed grasslands
Öckinger et al. 2010). Other traits characteristics associatedith generalists are a high distribution range and high dis-ersal ability, which enables butterflies to reach new habitatsn disturbed landscapes (Tscharntke et al. 2005). Specialistsan achieve high population densities, while generalists usu-lly show low population densities (Verberk, van der Welde,
Esselink 2010). Further traits characteristics associatedith generalists are high reproductive potential (e.g. mul-
iple generations per year) and fast development (e.g. fastgg maturation) compensating frequent habitat disturbancend food removal in intensive grasslands (Kitahara, Sei, &ujii 2000). We therefore expect that the selected traits willhange with increasing land-use intensity towards a broadeeding niche, high dispersal ability, high dispersal propen-ity, a large distribution range, low population density, highgg number, multiple generations per year, fast egg matu-ation, hibernation in a more advanced developmental stagend a long flight period.
Our predictions are:
(a) Butterfly diversity decreases with increasing land-useintensity.
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
b) Traits of butterfly communities change from charac-teristics associated with specialists to characteristicsassociated with generalists.

ARTICLE IN PRESSBAAE-50733; No. of Pages 8
pplied
rcbus
M
S
amTaoTatgccimietuaea
v(laisbaigaosentwua
L
t2awFaB
Bt
agvtWtforabaF
fls(Sd(ppt(o
S
wsdia
cas
C. Börschig et al. / Basic and A
Since traits within species communities are usually cor-elated we used a fourth-corner analysis to conduct oneomprehensive trait analysis, accounting for biases generatedy highly correlated traits (Dray & Legendre 2008). Anothernique aspect of our study is the large number of sampledtudy sites in three independent study regions.
aterials and methods
tudy regions and land-use intensity
The effect of land-use intensity on butterfly ecologicalnd life-history traits was studied in three regions in Ger-any within the project ‘Biodiversity Exploratories’ in 2008.he regions are separated by 350 km and differ in climatend agricultural structure, but every region shows a rangef grasslands covering a wide land-use intensity gradient.he three selected regions are UNESCO protected areasnd have proportionally high amounts of semi-natural habi-ats compared to many others regions in Germany. Due toeological conditions the region Schorfheide has no cal-areous grasslands in contrast to the other two regions. Wehose to sample three distinct regions aiming to researchf butterfly traits change similarly with grassland manage-
ent intensity across distinct but similar regions. For furthernformation about the regions see Fischer et al. (2010). Inach region we chose a minimum of 44 grasslands (137 inotal) spanning a gradient from extensive to intensive landse. Grasslands were mown one to three times per yearnd/or grazed by sheep, cattle or horse. Grasslands wereither unfertilised or different amounts of fertilisers werepplied.
We used a land-use intensity index (LUI), a continuousariable, to quantify variation of multiple land-use typesBlüthgen et al. 2012). The index does not suffer informationoss due to categorisation and makes different types of man-gement, such as mowing and grazing, comparable. The LUIs useful when an overall measurement for land-use inten-ity is needed with heterogeneous management strategies,ut effects cannot be deduced to single management vari-bles such as grazing or fertilisation. We obtained detailednformation on the land-use practises from a questionnaireiven to all land owners and land users. The LUI integratesmount of fertiliser (F), mowing frequency (M) and intensityf grazing (G). Grazing livestock was translated into live-tock units weighted for their impact on grasslands (Blüthgent al. 2012). For each study site i each individual LUI compo-ent (Fi, Mi, Gi) was standardised relative to its mean withinhe corresponding region R (FR, MR, GR). Regional meansere used, as the different agricultural regions may lead tonbalanced contributions of the three LUI components to the
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
ggregate index.
UIi = Fi
FR
+ Mi
MR
+ Gi
GR
cw2S
Ecology xxx (2013) xxx–xxx 3
The LUI was calculated for each grassland study site forhe study year 2008 and the previous two years 2006 and007 and the average of these three years was taken tochieve a better estimate of land-use intensity. Similar indicesere developed and used previously (Herzog et al. 2006).or details on differences between indices and on meansnd ranges of the different land-use categories we refer tolüthgen et al. (2012).
utterfly surveys and ecological and life-historyraits
We surveyed butterflies from 7 May to 17 August 2008long fixed transects randomly chosen in each of the 137rassland study sites. We counted all species and their indi-idual numbers within 2.5 m either side and 5 m in front ofhe scientists on transects of 300 m length within 30 min.
e sampled butterflies by sweep net and either releasedhem after identification or collected them if necessary forurther identification by dissection of genitalia in the lab-ratory. We sampled all 137 study sites three times in aandomised sequence within the regions approximately once
month, but only when weather conditions were suitable forutterfly activity (Pollard 1977). For butterfly identificationnd nomenclature we followed Settele, Steiner, Reinhardt,eldmann, and Herrmann (2009).We compiled 10 ecological and life-history traits of butter-
ies within three ecologically relevant groups: (1) Food plantpecialisation (feeding niche); (2) Dispersal and distributionwing length as a proxy for dispersal ability (Sekar 2012;tevens, Trochet, Van Dyck, Clobert, & Baguette 2012),ispersal propensity, distribution index, population density);3) Reproduction and development (egg number, generationser year, egg maturation time, hibernation stage and flighteriod). We extracted the trait data from the following litera-ure: Higgins and Riley (1978), Settele et al. (1999), Kudrna2002), Settele et al. (2009). For more detailed informationn the traits see Appendix A: Tables 1 and 2.
tatistical methods
We calculated species diversity using the Shannon Index,hich accounts for relative abundance and the number of
pecies within a site. We used a linear model to test if butterflyiversity differs between regions and changes with land-usentensity (LUI) and tested also the interaction between regionnd LUI.
We tested for changes of the 10 traits between butterflyommunities with increasing land-use intensity. Trait vari-bles without natural numbers were given values to enabletatistical analysis, e.g. the different hibernation stages were
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
oded from 1 to 7 (egg, three larval stages, pupa, adult, over-intering in warmer regions) (see Appendix A: Tables 1 and). This approach was also applied in previous studies (e.g.tevens et al. 2012). We tested the relationship between traits
ARTICLE IN PRESSBAAE-50733; No. of Pages 8
4 C. Börschig et al. / Basic and Applied Ecology xxx (2013) xxx–xxx
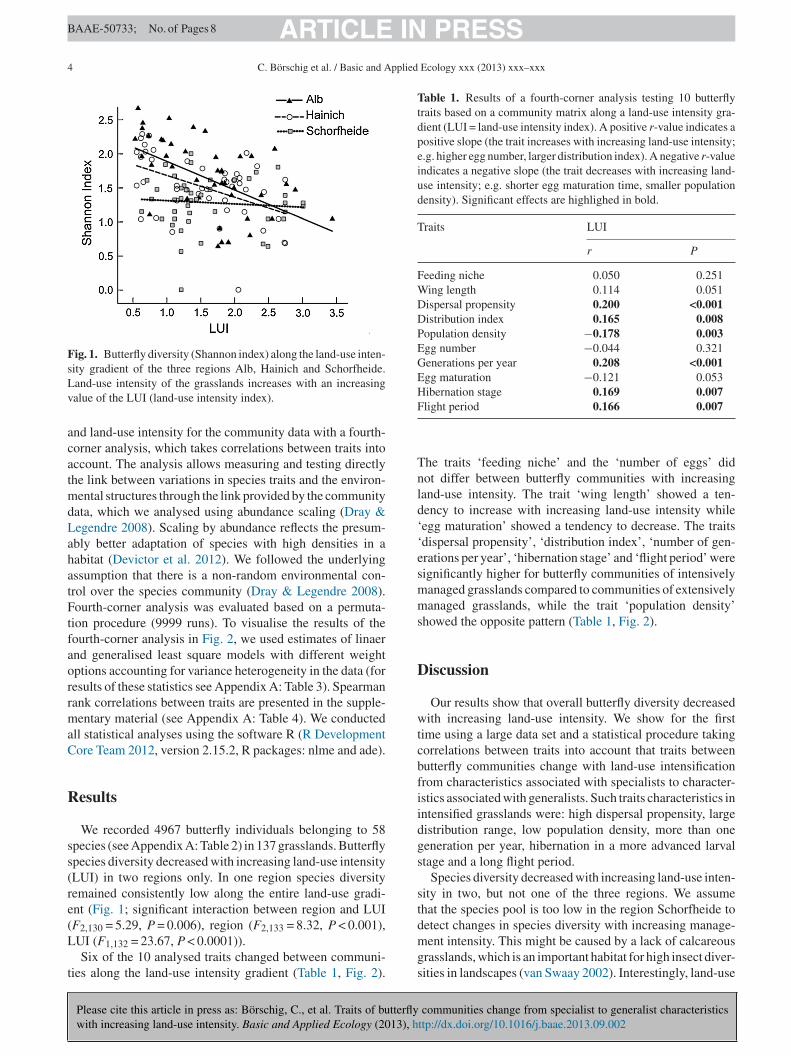
Fig. 1. Butterfly diversity (Shannon index) along the land-use inten-sity gradient of the three regions Alb, Hainich and Schorfheide.Lv
acatmdLahatFtfaorrmaC
R
ss(re(L
t
Table 1. Results of a fourth-corner analysis testing 10 butterflytraits based on a community matrix along a land-use intensity gra-dient (LUI = land-use intensity index). A positive r-value indicates apositive slope (the trait increases with increasing land-use intensity;e.g. higher egg number, larger distribution index). A negative r-valueindicates a negative slope (the trait decreases with increasing land-use intensity; e.g. shorter egg maturation time, smaller populationdensity). Significant effects are highlighed in bold.
Traits LUI
r P
Feeding niche 0.050 0.251Wing length 0.114 0.051Dispersal propensity 0.200 <0.001Distribution index 0.165 0.008Population density −0.178 0.003Egg number −0.044 0.321Generations per year 0.208 <0.001Egg maturation −0.121 0.053Hibernation stage 0.169 0.007F
Tnld‘‘esmms
D
wtcbfiidgs
st
and-use intensity of the grasslands increases with an increasingalue of the LUI (land-use intensity index).
nd land-use intensity for the community data with a fourth-orner analysis, which takes correlations between traits intoccount. The analysis allows measuring and testing directlyhe link between variations in species traits and the environ-
ental structures through the link provided by the communityata, which we analysed using abundance scaling (Dray &egendre 2008). Scaling by abundance reflects the presum-bly better adaptation of species with high densities in aabitat (Devictor et al. 2012). We followed the underlyingssumption that there is a non-random environmental con-rol over the species community (Dray & Legendre 2008).ourth-corner analysis was evaluated based on a permuta-
ion procedure (9999 runs). To visualise the results of theourth-corner analysis in Fig. 2, we used estimates of linaernd generalised least square models with different weightptions accounting for variance heterogeneity in the data (foresults of these statistics see Appendix A: Table 3). Spearmanank correlations between traits are presented in the supple-entary material (see Appendix A: Table 4). We conducted
ll statistical analyses using the software R (R Developmentore Team 2012, version 2.15.2, R packages: nlme and ade).
esults
We recorded 4967 butterfly individuals belonging to 58pecies (see Appendix A: Table 2) in 137 grasslands. Butterflypecies diversity decreased with increasing land-use intensityLUI) in two regions only. In one region species diversityemained consistently low along the entire land-use gradi-nt (Fig. 1; significant interaction between region and LUI
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
F2,130 = 5.29, P = 0.006), region (F2,133 = 8.32, P < 0.001),UI (F1,132 = 23.67, P < 0.0001)).Six of the 10 analysed traits changed between communi-
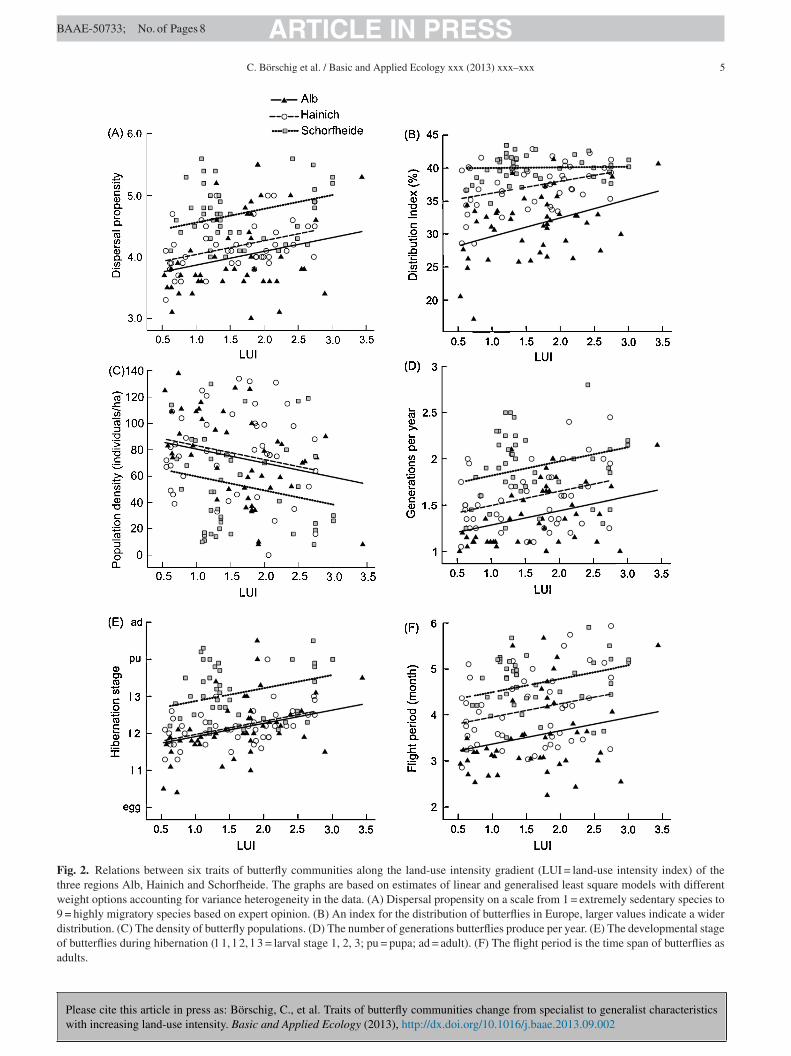
ies along the land-use intensity gradient (Table 1, Fig. 2).
dmgs
light period 0.166 0.007
he traits ‘feeding niche’ and the ‘number of eggs’ didot differ between butterfly communities with increasingand-use intensity. The trait ‘wing length’ showed a ten-ency to increase with increasing land-use intensity whileegg maturation’ showed a tendency to decrease. The traitsdispersal propensity’, ‘distribution index’, ‘number of gen-rations per year’, ‘hibernation stage’ and ‘flight period’ wereignificantly higher for butterfly communities of intensivelyanaged grasslands compared to communities of extensivelyanaged grasslands, while the trait ‘population density’
howed the opposite pattern (Table 1, Fig. 2).
iscussion
Our results show that overall butterfly diversity decreasedith increasing land-use intensity. We show for the first
ime using a large data set and a statistical procedure takingorrelations between traits into account that traits betweenutterfly communities change with land-use intensificationrom characteristics associated with specialists to character-stics associated with generalists. Such traits characteristics inntensified grasslands were: high dispersal propensity, largeistribution range, low population density, more than oneeneration per year, hibernation in a more advanced larvaltage and a long flight period.
Species diversity decreased with increasing land-use inten-ity in two, but not one of the three regions. We assumehat the species pool is too low in the region Schorfheide toetect changes in species diversity with increasing manage-
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
ent intensity. This might be caused by a lack of calcareousrasslands, which is an important habitat for high insect diver-ities in landscapes (van Swaay 2002). Interestingly, land-use
Please cite this article in press as: Börschig, C., et al. Traits of butterfly communities change from specialist to generalist characteristicswith increasing land-use intensity. Basic and Applied Ecology (2013), http://dx.doi.org/10.1016/j.baae.2013.09.002
ARTICLE IN PRESSBAAE-50733; No. of Pages 8
C. Börschig et al. / Basic and Applied Ecology xxx (2013) xxx–xxx 5
Fig. 2. Relations between six traits of butterfly communities along the land-use intensity gradient (LUI = land-use intensity index) of thethree regions Alb, Hainich and Schorfheide. The graphs are based on estimates of linear and generalised least square models with differentweight options accounting for variance heterogeneity in the data. (A) Dispersal propensity on a scale from 1 = extremely sedentary species to9 = highly migratory species based on expert opinion. (B) An index for the distribution of butterflies in Europe, larger values indicate a widerdistribution. (C) The density of butterfly populations. (D) The number of generations butterflies produce per year. (E) The developmental stageof butterflies during hibernation (l 1, l 2, l 3 = larval stage 1, 2, 3; pu = pupa; ad = adult). (F) The flight period is the time span of butterflies asadults.

ARTICLE IN PRESSBAAE-50733; No. of Pages 8
6 pplied
iwrcg
faHf“bin(
iimffidhhbmdtbgrgtGaowflC
us(cpmpmcraprbh
ssCt2dv&dtu2(ud
lqdflttgwsfibsgasafnbmmpDip(bea(ytiady
C. Börschig et al. / Basic and A
ntensity affects species diversity differently between regions,hereas effects on most traits were constant across all three
egions (see Fig. 2 and Appendix A: Table 3). Therefore traitsan be used as additional indicators for land-use intensity inrasslands (Pfestorf et al. 2013).
We expected butterfly communities to show broadereeding niches in intensively rather than in extensively man-ged grasslands, as shown in previous studies (e.g. Dennis,odgson, Grenyer, Shreeve, & Roy 2004). However, we
ound no significant effect of land-use intensity on the traitfeeding niche” with the fourth-corner analysis, which coulde explained by correlations between traits. For example, anncreasing diversity in the diet of butterflies increases theumber of generations per year and extends the flight periodAltermatt 2010).
We have shown that dispersal propensity and distributionncreased in butterfly communities with increasing land-usentensity, while population density decreased. Butterfly com-unities in intensively managed grasslands are faced with
requent habitat disturbances and unreliable and low diverseood resources. Therefore such communities can be dom-nated by species with a high dispersal ability and a highispersal propensity, enabling them to (re-) colonise disturbedabitats (Stevens, Turlure, & Baguette 2010). In contrast,abitat stability of extensively managed grasslands favoursutterfly communities with low emigration rates, due to highortality risk of between-habitat dispersal and high costs of
ispersal in terms of time and energy which could be allocatedo reproduction (Hovestadt & Nieminen 2009). A large distri-ution range of butterfly communities in intensively managedrasslands can be explained by several traits of butterflies cor-elating with distribution. High dispersal propensity, manyenerations per year and a long flight period allow butterflieso explore and colonise distant habitats (Lester, Ruttenberg,aines, & Kinlan 2007). The habitat area of extensively man-
ged grasslands dropped drastically while the habitat areaf intensively managed grasslands has increased in Europe,hich reduced the distribution range of specialised butter-y species (van Swaay et al. 2006; Polus, Vandewoestijne,houtt, & Baguette 2007).We found that population densities decreased with land-
se intensity. In intensively managed grasslands usually thepecies richness, but also the (relative) abundance is lowTscharntke et al. 2005). High mowing and grazing frequen-ies causing unpredictable food resources might decreaseopulation densities of butterfly communities in intensivelyanaged grasslands supported by the associated high dis-
ersal propensity of such communities. In stable extensivelyanaged habitats with a low perturbation, butterflies can
omplete their life cycle and achieve high densities. Thearity of extensively managed grassland also might inducen increase of population densities, as other suitable habitat
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
atches are difficult to reach (Päivinen et al. 2005). Cor-espondingly, Dennis et al. (2004) found low densities ofutterfly species feeding on plants characterising intensifiedabitats. Also Kotiaho et al. (2005) showed that threatened
egtW
Ecology xxx (2013) xxx–xxx
pecies, usually inhabiting extensively managed grasslands,how higher population densities than unthreatened species.ontrary to the expectation that species with broad dis-
ribution ranges are also locally abundant (Gaston et al.000), we found a negative relation between populationensity and distribution. These findings are in line with pre-ious studies (Päivinen et al. 2005; Komonen, Päivinen,
Kotiaho 2009) doubting the generality of the positiveensity-distribution concept (Johnson 1998). One reason forhis discrepancy might be differences in the spatial scalesed for calculating densities in the studies (Cowley et al.001; Hartley & Kunin 2003) or a bias in sampling effortsKomonen et al. 2009). However, interpretations remain spec-lative and further investigations into the generality of theistribution–density relationship for insects is needed.
A high reproductive potential and fast development withong flight periods can compensate high disturbance fre-uency, frequent food resource removal and low plantiversity in a perturbed intensively managed habitat. Butter-y communities with a high average egg load could increase
he number of potential offspring surviving and the produc-ion of more generations per year may accelerate populationrowth (Cizek, Fric, & Konvicka 2006). A high egg loadas observed for butterfly species feeding on plants of inten-
ively managed habitats (Dennis et al. 2004). However, ourndings did not show a relation between egg numbers inutterfly communities and changing land land-use inten-ity. Similarly, butterfly communities of intensively managedrasslands did not show the expected faster egg maturationfter adult emergence than butterfly communities of exten-ively managed grasslands. Immediate egg maturation afterdult emergence could avoid untimely food removal in arequently disturbed habitat. Contrasting, our data show aegative tendency. The egg maturation time is higher forutterfly communities in extensively rather than intensivelyanaged grasslands. Fast egg maturation and a high egg loadight be explained by high competition and a potentially high
redation risk in extensively managed grasslands (Steffan-ewenter & Tscharntke 2002). Butterfly communities in
ntensively managed grasslands produce more generationser year, which is highly correlated with a long flight periodAltermatt 2010). Intensively managed grasslands show few,ut very abundant plant species. These plants have long veg-tation periods, flowering early in the year and regrowingfter recurrent disturbance events such as grazing or mowingGarnier et al. 2007). This may allow more generations perear for butterflies and a longer flight period, as comparedo extensively managed grasslands. Butterfly communitiesn intensively managed grasslands hibernate on average in
more advanced developmental stage. A more advancedevelopmental stage allows emergence of adults early in theear, which is beneficial, because management starts usually
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
arly on highly intensified grasslands. Mowing and intenserazing destroy larval habitats and also can be detrimen-al to immobile larvae (van Noordwijk, Flierman, Remke,
allisDeWries, & Berg 2012).

ARTICLE IN PRESSBAAE-50733; No. of Pages 8
pplied
tild(DiHfce
A
rfFLaAtamIWdggF‘FeB
A
fj
R
A
B
B
C
C
C
D
D
D
D
E
E
F
G
G
H
H
H
C. Börschig et al. / Basic and A
In conclusion we showed that traits of butterfly communi-ies change from specialist to generalist characteristics withncreasing land-use intensity. This is in line with findings onandscape homogenisation, habitat loss and fragmentationisadvantaging butterflies with traits typical for specialistsSteffan-Dewenter & Tscharntke 2000; Öckinger et al. 2010;iacon-Bolli et al. 2012). This leads to communities increas-
ngly dominated by few generalist butterfly species (Ekroos,eliölä, & Kuussaari 2010; Öckinger et al. 2010). Such
unctional homogenisation of communities can have severeonsequences for ecosystem functioning and services (Clavelt al. 2010).
cknowledgements
We are grateful to Elke Vockenhuber, Vesna Gagic, threeeviewers and the editor Klaus Hövemeyer for very help-ul comments on the manuscript. Elke Vockenhuber, Jochenründ and Christoph Scherber provided statistical support.aszló Somay identified some butterflies. We thank the man-gers of the three exploratories, Swen Renner, Sonja Gockel,ndreas Hemp and Martin Gorke and Simone Pfeiffer for
heir work in maintaining the plot and project infrastructure,nd Markus Fischer, the late Elisabeth Kalko, Eduard Linsen-air, Dominik Hessenmöller, Jens Nieschulze, Daniel Prati,
ngo Schöning, Francois Buscot, Ernst-Detlef Schulze andolfgang W. Weisser for their role in setting up the Bio-
iversity Exploratories project. We thank the farmers foriving permission to conduct our butterfly transects in theirrasslands. This work has been funded by the Deutscheorschungsgemeinschaft (DFG) Priority Programme 1374Infrastructure-Biodiversity-Exploratories’ (KL 1849/3-1).ieldwork permits were provided by the responsible statenvironmental offices of Baden-Württemberg, Thüringen andrandenburg (according to § 72 BbgNatSchG).
ppendix A. Supplementary data
Supplementary material related to this article can beound, in the online version, at http://dx.doi.org/10.1016/.baae.2013.09.002.
eferences
ltermatt, F. (2010). Tell me what you eat and I’ll tell you when youfly: Diet can predict phenological changes in response to climatechange. Ecology Letters, 13, 1475–1484.
lüthgen, N., Dormann, C. F., Prati, D., Klaus, V. H., Kleinebecker,T., Hölzel, N., et al. (2012). A quantitative index of land-use
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
intensity in grasslands: Integrating mowing, grazing and fertil-ization. Basic and Applied Ecology, 13, 207–220.
rückmann, S. V., Krauss, J., & Steffan-Dewenter, I. (2010). But-terfly and plant specialists suffer from reduced connectivity
H
Ecology xxx (2013) xxx–xxx 7
in fragmented landscapes. Journal of Applied Ecology, 47,799–809.
izek, L., Fric, Z., & Konvicka, M. (2006). Host plant defences andvoltinism in European butterflies. Ecological Entomology, 31,337–344.
lavel, J., Julliard, R., & Devictor, V. (2010). Worldwide declineof specialist species: Toward a global functional homog-enization? Frontiers in Ecology and the Environment, 9,222–228.
owley, M. J. R., Thomas, C. D., Roy, D. B., Wilson, R. J., León-Cortés, J. L., Gutiérrez, D., et al. (2001). Density–distributionrelationships in British butterflies. I. The effect of mobil-ity and spatial scale. Journal of Animal Ecology, 70,410–425.
ennis, R. L. H., Hodgson, J. G., Grenyer, R., Shreeve, T. G., &Roy, D. B. (2004). Host plants and butterfly biology. Do host-plant strategies drive butterfly status? Ecological Entomology,29, 12–26.
evictor, V., van Swaay, C., Brereton, T., Brotons, L., Chamberlain,D., Heliölä, J., et al. (2012). Differences in the climatic debtsof birds and butterflies at a continental scale. Nature ClimateChange, 2, 121–124.
iacon-Bolli, J., Dalang, T., Holderegger, R., & Bürgi, M. (2012).Heterogeneity fosters biodiversity: Linking history and ecologyof dry calcareous grasslands. Basic and Applied Ecology, 13,641–653.
ray, S., & Legendre, P. (2008). Testing the species traits-environment relationships: The fourth-corner problem revisited.Ecology, 89, 3400–3412.
kroos, J., Heliölä, J., & Kuussaari, M. (2010). Homogenization oflepidopteran communities in intensively cultivated agriculturallandscapes. Journal of Applied Ecology, 47, 459–467.
llis, E. C., Klein Goldewijk, K., Siebert, S., Lightman, D., &Ramankutty, N. (2010). Anthropogenic transformation of thebiomes, 1700 to 2000. Global Ecology and Biogeography, 19,589–606.
ischer, M., Bossdorf, O., Gockel, S., Hänsel, F., Hemp, A., Hessen-möller, D., et al. (2010). Implementing large-scale and long-termfunctional biodiversity research: The Biodiversity Exploratories.Basic and Applied Ecology, 11, 473–485.
arnier, E., Lavorel, S., Ansquer, P., Castro, H., Cruz, P., Dolezal,J., et al. (2007). Assessing the effects of land-use change on planttraits, communities and ecosystem functioning: A standardizedmethodology and lessons from an application to 11 Europeansites. Annals of Botany, 99, 967–985.
aston, K. J., Blackburn, T. M., Greenwood, J. J. D., Gregory, R. D.,Quinn, R. M., & Lawton, J. H. (2000). Abundance–occupancyrelationships. Journal of Applied Ecology, 37, 39–59.
artley, S., & Kunin, W. E. (2003). Scale dependency of rarity,extinction risk, and conservation priority. Conservation Biology,17, 1559–1570.
erzog, F., Steiner, B., Bailey, D., Baudry, J., Billeter, R., Bukácek,R., et al. (2006). Assessing the intensity of temperate Euro-pean agriculture at the landscape scale. European Journal ofAgronomy, 24, 165–181.
iggins, L. G., & Riley, N. D. (1978). Die Tagfalter Europas undNordwestafrikas. Singhofen: Verlag Paul Parey.
ovestadt, T., & Nieminen, M. (2009). Costs and benefits of dis-
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
persal in butterflies. In J. Settele, T. Shreeve, M. Konvicka, & H.Van Dyck (Eds.), Ecology of butterflies in Europe (pp. 97–106).Cambridge: Cambridge University Press.

ARTICLE IN PRESSBAAE-50733; No. of Pages 8
8 pplied
J
K
K
K
K
K
L
M
Ö
P
P
P
P
R
S
S
S
S
S
S
S
S
T
T
v
v
v
V
C. Börschig et al. / Basic and A
ohnson, C. N. (1998). Species extinction and the relationshipbetween distribution and abundance. Nature, 394, 272–274.
itahara, M., Sei, K., & Fujii, K. (2000). Patterns in the structureof grassland butterfly communities along a gradient of humandisturbance: Further analysis based on the generalist/specialistconcept. Population Ecology, 42, 135–144.
omonen, A., Päivinen, J., & Kotiaho, J. S. (2009). Missingthe rarest: Is the positive interspecific abundance–distributionrelationship a truly general macroecological pattern? BiologyLetters, 5, 492–494.
otiaho, J. S., Kaitala, V., Komonen, A., & Päivinen, J. (2005).Predicting the risk of extinction from shared ecological charac-teristics. Proceedings of the National Academy of Sciences of theUnited States of America, 102, 1963–1967.
ruess, A., & Tscharntke, T. (2002). Grazing and diversity ofgrasshoppers, butterflies, and trap-nesting bees and wasps. Con-servation Biology, 16, 1570–1580.
udrna, O. (2002). The distribution atlas of European butterflies.Oedippus 20, Vester Skerninge: Apollo Books.
ester, S. E., Ruttenberg, B. I., Gaines, S. D., & Kinlan, B. P. (2007).The relationship between dispersal ability and geographic rangesize. Ecology Letters, 10, 745–758.
cKinney, M. L., & Lockwood, J. L. (1999). Biotic homog-enization: A few winners replacing many losers in thenext mass extinction. Trends in Ecology & Evolution, 14,450–453.
ckinger, E., Schweiger, O., Crist, T. O., Debinski, D. M., Krauss,J., Kuussaari, M., et al. (2010). Life-history traits predict speciesresponses to habitat area and isolation: A cross-continental syn-thesis. Ecology Letters, 13, 969–979.
äivinen, J., Grapputo, A., Kaitala, V., Komonen, A., Kotiaho, J.,Saarinen, K., et al. (2005). Negative density–distribution rela-tionship in butterflies. BMC Biology, 3, 5.
festorf, H., Weiß, L., Müller, J., Boch, S., Socher, S. A., Prati, D.,et al. (2013). Community mean traits as additional indicators tomonitor effects of land-use intensity on grassland plant diversity.Perspectives in Plant Ecology, Evolution and Systematics, 15,1–11.
ollard, E. (1977). Method for assessing changes in abundance ofbutterflies. Biological Conservation, 12, 115–134.
olus, E., Vandewoestijne, S., Choutt, J., & Baguette, M. (2007).Tracking the effects of one century of habitat loss and fragmenta-tion on calcareous grassland butterfly communities. Biodiversityand Conservation, 16, 3423–3436.
Please cite this article in press as: Börschig, C., et al. Traits of butterflywith increasing land-use intensity. Basic and Applied Ecology (2013), h
Development Core Team. (2012). R: A language andenvironment for statistical computing. Vienna, Austria: R Foun-dation for Statistical Computing. ISBN: 3-900051-07-0, URL:http://www.R-project.org
W
Available online at www.s
ScienceD
Ecology xxx (2013) xxx–xxx
ala, O. E., Chapin, F. S., Armesto, J. J., Berlow, E., Bloomfield,J., Dirzo, R., et al. (2000). Global biodiversity scenarios for theyear 2100. Science, 287, 1770–1774.
ekar, S. (2012). A meta-analysis of the traits affecting dispersalability in butterflies: Can wingspan be used as a proxy? Journalof Animal Ecology, 81, 174–184.
ettele, J., Feldmann, R., & Reinhardt, R. (1999). Die TagfalterDeutschlands. Stuttgart: Ulmer Verlag.
ettele, J., Steiner, R., Reinhardt, R., Feldmann, R., & Herrmann, G.(2009). Schmetterlinge. Die Tagfalter Deutschlands. Stuttgart:Ulmer Verlag.
teffan-Dewenter, I., & Tscharntke, T. (2000). Butterfly communitystructure in fragmented habitats. Ecology Letters, 3, 449–456.
teffan-Dewenter, I., & Tscharntke, T. (2002). Insect communitiesand biotic interactions on fragmented calcareous grasslands – Amini review. Biological Conservation, 104, 275–284.
tevens, V. M., Trochet, A., Van Dyck, H., Clobert, J., & Baguette,M. (2012). How is dispersal integrated in life histories: A quan-titative analysis using butterflies. Ecology Letters, 15, 74–86.
tevens, V. M., Turlure, C., & Baguette, M. (2010). A meta-analysisof dispersal in butterflies. Biological Reviews, 85, 625–642.
homas, J. A. (2005). Monitoring change in the abundance and dis-tribution of insects using butterflies and other indicator groups.Philosophical Transactions of the Royal Society B: BiologicalSciences, 360, 339–357.
scharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I., &Thies, C. (2005). Landscape perspectives on agricultural inten-sification and biodiversity – Ecosystem service management.Ecology Letters, 8, 857–874.
an Noordwijk, C. G. E., Flierman, D. E., Remke, E., Wallis-DeWries, M. F., & Berg, M. P. (2012). Impact of grazingmanagement on hibernating caterpillars of the butterfly Melitaeacinxia in calcareous grasslands. Journal of Insect Conservation,16, 909–920.
an Swaay, C. A. M. (2002). The importance of calcareous grass-lands for butterflies in Europe. Biological Conservation, 104,315–318.
an Swaay, C. A. M., Warren, M., & Loïs, G. (2006). Biotope use andtrends of European butterflies. Journal of Insect Conservation,10, 189–209.
erberk, W. C. E. P., van der Welde, G., & Esselink, H. (2010).Explaining abundance–occupany relationships in specialists andgeneralists: A case study on aquatic macroinvertebrates in stand-ing waters. Journal of Animal Ecology, 79, 589–601.
illiams, N. M., Crone, E. E., Roulston, T. H., Minckley, R. L.,
communities change from specialist to generalist characteristicsttp://dx.doi.org/10.1016/j.baae.2013.09.002
Packer, L., & Potts, S. G. (2010). Ecological and life-history traitspredict bee species responses to environmental disturbances.Biological Conservation, 143, 2280–2291.
ciencedirect.com
irect
Related Documents











