RESEARCH ARTICLE Trait means and reaction norms: the consequences of climate change/invasion interactions at the organism level Charlene Janion • Hans Petter Leinaas • John S. Terblanche • Steven L. Chown Received: 20 January 2010 / Accepted: 28 June 2010 / Published online: 15 July 2010 Ó Springer Science+Business Media B.V. 2010 Abstract How the impacts of climate change on biological invasions will play out at the mechanistic level is not well understood. Two major hypotheses have been proposed: invasive species have a suite of traits that enhance their performance relative to indigenous ones over a reasonably wide set of circumstances; invasive species have greater phenotypic plasticity than their indigenous counterparts and will be better able to retain performance under altered conditions. Thus, two possibly independent, but complementary mechanistic perspectives can be adopted: based on trait means and on reaction norms. Here, to dem- onstrate how this approach might be applied to understand interactions between climate change and invasion, we investigate variation in the egg development times and their sensitivity to temperature amongst indigenous and introduced springtail species in a cool temperate ecosystem (Marion Island, 46°54 0 S 37°54 0 E) that is undergoing significant cli- mate change. Generalized linear model analyses of the linear part of the development rate curves revealed significantly higher mean trait values in the invasive species compared to indigenous species, but no significant interactions were found when comparing the thermal reaction norms. In addition, the invasive species had a higher hatching success than the indigenous species at high temperatures. This work demonstrates the value of explicitly examining variation in trait means and reaction norms among indigenous and invasive species to understand the mechanistic basis of variable responses to climate change among these groups. Electronic supplementary material The online version of this article (doi:10.1007/s10682-010-9405-2) contains supplementary material, which is available to authorized users. C. Janion (&) S. L. Chown Centre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University, Private Bag X1, Matieland 7602, South Africa e-mail: [email protected] H. P. Leinaas Department of Biology, University of Oslo, Blindern, P.O. Box 1066, 0316 Oslo, Norway J. S. Terblanche Department of Conservation Ecology and Entomology, Stellenbosch University, Private Bag X1, Matieland 7602, South Africa 123 Evol Ecol (2010) 24:1365–1380 DOI 10.1007/s10682-010-9405-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
Trait means and reaction norms: the consequencesof climate change/invasion interactionsat the organism level
Charlene Janion • Hans Petter Leinaas • John S. Terblanche •
Steven L. Chown
Received: 20 January 2010 / Accepted: 28 June 2010 / Published online: 15 July 2010� Springer Science+Business Media B.V. 2010
Abstract How the impacts of climate change on biological invasions will play out at the
mechanistic level is not well understood. Two major hypotheses have been proposed:
invasive species have a suite of traits that enhance their performance relative to indigenous
ones over a reasonably wide set of circumstances; invasive species have greater phenotypic
plasticity than their indigenous counterparts and will be better able to retain performance
under altered conditions. Thus, two possibly independent, but complementary mechanistic
perspectives can be adopted: based on trait means and on reaction norms. Here, to dem-
onstrate how this approach might be applied to understand interactions between climate
change and invasion, we investigate variation in the egg development times and their
sensitivity to temperature amongst indigenous and introduced springtail species in a cool
temperate ecosystem (Marion Island, 46�540S 37�540E) that is undergoing significant cli-
mate change. Generalized linear model analyses of the linear part of the development rate
curves revealed significantly higher mean trait values in the invasive species compared to
indigenous species, but no significant interactions were found when comparing the thermal
reaction norms. In addition, the invasive species had a higher hatching success than the
indigenous species at high temperatures. This work demonstrates the value of explicitly
examining variation in trait means and reaction norms among indigenous and invasive
species to understand the mechanistic basis of variable responses to climate change among
these groups.
Electronic supplementary material The online version of this article (doi:10.1007/s10682-010-9405-2)contains supplementary material, which is available to authorized users.
C. Janion (&) � S. L. ChownCentre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University,Private Bag X1, Matieland 7602, South Africae-mail: [email protected]
H. P. LeinaasDepartment of Biology, University of Oslo, Blindern, P.O. Box 1066, 0316 Oslo, Norway
J. S. TerblancheDepartment of Conservation Ecology and Entomology, Stellenbosch University, Private Bag X1,Matieland 7602, South Africa
123
Evol Ecol (2010) 24:1365–1380DOI 10.1007/s10682-010-9405-2

Keywords Biological invasions � Climate change � Development rate �Phenotypic plasticity � Soil arthropod
Introduction
The major drivers of biodiversity change are well known. Habitat alteration, over-
exploitation, biological invasions and co-extinctions were identified as an ‘evil quartet’
nearly two decades ago (Diamond 1989). Together with climate change, their present and
likely future effects on biodiversity and human well-being remain among the most sig-
nificant problems presently facing humanity (Sala et al. 2000; Chown and Gaston 2008).
Although much is known about the mechanistic basis of each of these drivers, how they are
likely to interact has not been comprehensively investigated across the full range of
interactions (Brook et al. 2008). For example, understanding of the nature and outcome of
interactions among habitat alteration and invasion is growing rapidly, and it is clear that the
‘driver-passenger’ relationship is often context dependent (Daehler 2003; Didham et al.
2007). Similarly, appreciation of how habitat alteration, patchiness, climate change and
short-term evolutionary responses act in isolation and in concert to affect species distri-
butions across the landscape is also growing rapidly (Warren et al. 2001; Carroll and Fox
2007; de Mazancourt et al. 2008). By contrast, interactions between climate change and
invasion are poorly understood.
It is widely thought that climate change will exacerbate the extent and impact of
biological invasions, affecting each of the stages in the invasion process (Cannon 1998;
Hobbs and Mooney 2005; Theoharides and Dukes 2007), and, where non-indigenous
species have established, often favouring them over indigenous species (Dukes and
Mooney 1999). Whilst evidence is accumulating to support some of these conjectures (e.g.,
Holzapfel and Vinebrooke 2005; Chown et al. 2007), the number of empirical studies of
interactions between climate change and invasion remains relatively limited (Walther et al.
2009). Indeed, several recent reviews have concluded that the information available is
insufficient for empirical generalization (Ward and Masters 2007; Chown and Gaston
2008; Brook 2008). This is especially true for how such interactions might play out at the
mechanistic level (Agrawal 2001; Stachowicz et al. 2002). Nonetheless, two major
hypotheses have been proposed.
First, invasive species have a suite of traits that enhance their performance relative to
indigenous ones over a reasonably wide set of circumstances. In consequence, climate
change will continue to mean that these species perform better than their indigenous
counterparts. The foundation of this idea stretches back at least to Elton (1958) and has
formed the basis of a large literature attempting to identify the species-level characteristics
that might promote invasion. For example, in plants, invasive species are often charac-
terized by large stature, high seed mass, rapid and profuse seedling emergence, and self-
fertilization (Richardson and Pysek 2006; van Kleunen and Johnson 2007; van Kleunen
et al. 2008, but see also Davis and Shaw 2001; Daehler 2003). In consequence, these
species are able to take advantage of a reasonably broad range of conditions, especially
those that are considered favourable for plant performance generally, such as warm, moist,
nutrient rich circumstances (Stohlgren et al. 2008). This idea has also been extended to
many other groups. For example, invasive invertebrate species are apparently often small-
bodied and have high-growth rates, so promoting their success in novel environments
(Lawton and Brown 1986; Gaston et al. 2001). In birds, behavioural flexibility, broad
1366 Evol Ecol (2010) 24:1365–1380
123

environmental tolerances and wide resource use are all thought to favour invasion (Duncan
et al. 2003). Thus, if invasive species have a suite of characteristics that enable them to
succeed under a reasonably wide range of conditions they should perform better under
warmer conditions too. In ectotherms, the generally positive relationship between growth
rate and temperature (Chown and Nicolson 2004; Ward and Masters 2007) means that this
will be especially likely as long as the temperature increase does not exceed organismal
thermal safety margins (see also Deutsch et al. 2008; Frazier et al. 2008), and mismatches
in phenology do not limit resource availability (Parmesan 2007). However, this hypothesis
provides no grounds for expecting that climate change will increasingly favour invasive
species.
Second, invasive species have greater phenotypic plasticity than their indigenous
counterparts. In consequence, they will be better able to retain performance under the
altered circumstances precipitated by climate change. This hypothesis is a component of
the idea first proposed by Baker (1965) of a ‘‘general purpose genotype’’, and has garnered
some support both for plants (Daehler 2003; Richardson and Pysek 2006) and animals
(Trussell and Smith 2000; Rosecchi et al. 2001; Duncan et al. 2003; Dzialowski et al. 2003,
but see also Lee et al. 2007). In the context of climate change, Stachowicz et al. (2002)
demonstrated that invasive ascidians had steeper growth rate–temperature relationships
than an indigenous species, and argued that this has favoured the introduced species under
warming, an idea supported by several years of demographic data. By contrast, Chown
et al. (2007) found no difference in the extent of phenotypic plasticity for desiccation
tolerance in indigenous and invasive springtails. However, they showed that the form of
plasticity differed between these two groups. Invasive species are favoured following
exposure to warm conditions, whilst indigenous species perform better following a low
temperature treatment.
Clearly, plasticity may evolve independently of or in concert with the trait mean, and
indeed how this interaction should be viewed and modelled has been the subject of much
discussion (see Scheiner 1993; de Jong 1995, 2005; Via et al. 1995; Ghalambor et al.
2007). Nonetheless, as a matter of convenience the mean and plasticity can be considered
relatively independent (see Ghalambor et al. 2007: 396). Thus, the mechanistic, organis-
mal-level exploration of interactions between climate change and invasion can be
approached from complementary perspectives: the way that differences in mean trait
values among indigenous and invasive species are likely to affect performance or survival
under a new set of conditions, and the way that variation in reaction norms among these
two groups of species may affect fitness under altered conditions (Lee 2002; Lee et al.
2003; Dybdahl and Kane 2005). This does not mean that mean trait values and plasticity
are necessarily independent (see de Jong 1995, 2005; van Kleunen and Fischer 2005;
Ghalambor et al. 2007), but does provide a framework within which the historical per-
spectives of differences in trait values and in plasticity among indigenous and invasive
species can be interpreted.
Here, we adopt this approach to investigate the likely impacts of interactions between
climate change and invasion, mediated through egg development rate and its response to
temperature, in a springtail assemblage from a cool, temperate ecosystem that is under-
going significant climate change (Chown et al. 2007; see Dybdahl and Kane 2005 for a
similar approach). Springtails were chosen as the study organisms because they are a
globally significant group of soil invertebrates that play major roles in ecosystem func-
tioning (Rusek 1998), including in the system we investigate (Gabriel et al. 2001; Hugo
et al. 2004). Egg development rate (or time, given that rate = 1/development time) was
selected because it is an important component of fitness in springtails (as is development
Evol Ecol (2010) 24:1365–1380 1367
123

rate more generally—Sibly and Calow 1986; Roff 2002), and the egg development period
encompasses a substantial portion of the life cycle of an individual springtail (van Straalen
1994; Birkemoe and Leinaas 2000).
Specifically, we first test the hypothesis that over the linear portion of the rate–
temperature curve (this portion of the rate–temperature curves is most often the subject of
investigation—see e.g., Trudgill et al. 2005; de Jong and van der Have 2008), mean egg
development should be faster in the invasive than in the indigenous species. Next we
evaluate the prediction of the reaction norm hypothesis that the invasive species should
have steeper rate–temperature relationships than the indigenous species (which has its basis
not only in the findings of Stachowicz et al. (2002) for ascidians, but also in terms of the
same hypothesis proposed for insects from the same region (Chown et al. 2002)). The
outcomes of these two tests are subsequently explored in the context of what is known
about the population responses of these species to current climates and climate change, to
further knowledge of the likely effects of rapid climate change on this system (see le Roux
and McGeoch 2008).
Methods
Study site and species
All species used in this study were collected from sub-Antarctic Marion Island (46�540S37�540E), which has a cool, wet, windy climate that has shown a substantial change over
the last 50 years, including more than a 1.3�C increase in mean annual temperature
(Chown and Froneman 2008). Sixteen springtail species occur on the island, of which six
are invasive, four endemic and six have a broader, sub-Antarctic distribution (Gabriel et al.
2001; Frenot et al. 2005). The invasive springtails are mostly of European origin (Chown
et al. 2002) and are thought to have been introduced by humans with animal fodder
provided for sheep that were kept on the island in the 1950s (Cooper and Condy 1988).
Thus they have been on the island for at least 40 years (Deharveng 1981). In Europe,
original records of the species are mostly from Scandinavia and northern areas of Germany
(Fjellberg 1998; Potapov 2001). Low mitochondrial cytochrome oxidase I (COI) haplotype
numbers (typically a single haplotype) in the invasive springtail species are indicative of
colonization by a few individuals, whilst the indigenous species are characterized by
considerable haplotype diversity (Myburgh et al. 2007).
For this study, the seven most common Poduromorpha and Entomobryomorpha species
on the island were investigated. Cryptopygus dubius Deharveng (Isotomidae), C. tricuspisEnderlein and C. antarcticus travei Deharveng are indigenous, whilst Ceratophyselladenticulata Bagnall (Hypogastruridae), Parisotoma notabilis Schaeffer (Isotomidae),Isotomurus cf. palustris Muller (Isotomidae) and Pogonognathellus flavescens Tullberg
(Tomoceridae) are invasive (Gabriel et al. 2001). The species also differ in depth distri-
bution within the soil profile: Isotomurus cf. palustris, C. antarcticus travei and Pogon-ognathellus flavescens are epedaphic or soil-surface dwellers, Cryptopygus dubius and
Ceratophysella denticulata hemi-edaphic or litter dwellers, and C. tricuspis and Pariso-toma notabilis euedaphic or soil dwellers (see van Straalen 1994 and Hopkin 1997 for
discussion of depth distributions in springtails).
Several hundred adult specimens of each species were collected in the field (below
100 m above sea level) using an aspirator and placed into 30 ml vials with moist Plaster-
of-Paris substrates and small amounts of detritus for shelter and food, and transported back
1368 Evol Ecol (2010) 24:1365–1380
123

to the laboratory within 5 h of collection. In the laboratory, specimens were kept in 40 ml
containers, with moist Plaster-of-Paris substrates and small amounts of detritus and algae
as food, in incubators (LABCON, Johannesburg, South Africa, accurate to ±1�C) at 5�C
with a 12L: 12D photoperiod. The animals were then transferred to Oslo, Norway, where
most work was undertaken except for a few additional trials with Cryptopygus antarcticustravei and Isotomurus cf. palustris, which were done in Stellenbosch South Africa. In both
places animals were maintained under identical conditions in the same make of incubators
(Sanyo MIR 153, Osaka, Japan, accurate to ±0.5�C). All individuals used for experiments
were acclimated at 15�C (or 10�C in the case of Cryptopygus antarcticus travei) for at least
14 days before the start of the experiment, as egg production was low at 5�C. Distilled
water was added once a week to maintain high humidity and springtails were provided with
excess algae as a food source (replaced once to twice a week to prevent growth of mould).
Embryonic development rates
Previous studies have demonstrated low survival when springtail individuals are kept
separately (e.g., Birkemoe and Leinaas 2000). Thus, 10–20 adults from the stock cultures
were randomly placed in culture boxes (diameter = 3.4 cm, depth = 3 cm) lined with
Plaster-of-Paris mixed with charcoal powder (ratio of 9:1). For optimum egg production,
and to reduce maternal effects, culture boxes were kept at 15�C for one generation, or, for
Cryptopygus antarcticus travei only, at 10�C because mortality in this species was high at
15�C. Egg batches (Cfive eggs) were then collected daily from culture boxes of each
species (except P. flavescens which laid single eggs) and assigned to a particular tem-
perature treatment (see below). For each species and each treatment, eggs were distributed
at random among at least four culture vials to avoid container effects. The vials were then
placed into the incubators (Sanyo MIR) at a set temperature and 12L:12D light regime.
Eggs were inspected daily until hatching, and at this time containers were randomized
among shelves to avoid shelf effects. Egg development times (determined as the number of
days between egg laying and hatching (Birkemoe and Leinaas 2000)) were obtained at
c. 5�C intervals from 5 to 28.8�C to provide complete performance curves (see Angilletta
2006; Deere and Chown 2006). Due to technical difficulties (incubator failure) no egg
development time measurements were obtained for P. flavescens above 25�C. Cryptopygusantarcticus travei eggs failed to hatch at 20�C and above.
Hatching success is often traded off against egg development rate (Stearns 1992;
Stillwell and Fox 2005; Geister et al. 2009). This means that for a given species, a high egg
development rate may not necessarily also mean high survival under those conditions,
resulting in low fitness at high temperatures. Therefore, we also examined hatching success
at 15�C and at 25�C for all species except C. antarcticus travei which had zero hatching
success at 20 and 25�C. Specifically, we determined whether hatching success declined in
the indigenous vs. invasive species (irrespective of soil depth category) at the higher
temperatures. Hatching success was recorded as the number of eggs that hatched from the
total number of eggs in the batch. This value was converted to a percentage value (number
of hatched eggs/total number eggs in batch 9 100).
Analyses
First, whilst recognizing that rate–temperature relationships typically have a non-linear
form (Stearns 1992; Angilletta 2006), and that a variety of analytical approaches are
available to deal with this non-linearity (e.g. David et al. 1997; Izem and Kingsolver 2005)
Evol Ecol (2010) 24:1365–1380 1369
123

only the linear portion of the reaction norm (5–20�C in this case, hereafter rate–temper-
ature or R–T curve) was analyzed. Such analyses are typical of investigations of rate–
temperature relationships (see Trudgill et al. 2005; Ikemoto 2005; de Jong and van der
Have 2008). Moreover, the aim here was to consider the sensitivity to development over
the linear part of the temperature range as the reaction norm, and the mean across these
temperatures as the mean trait value.
For the analysis of mean trait values and their reaction norms over the linear portion of
the R–T curve, general linear models, implemented in Statistica v.8 (Statsoft, Tulsa, OK,
USA), were used. In all cases, mean egg development rate per egg batch was used as the
independent data because individual eggs within batches are unlikely to represent inde-
pendent data (with the exception of Pogonognathellus flavescens which laid eggs singly).
The first hypothesis tested was that mean log10 egg development rate, holding temperature
constant, differs among the invasive and indigenous species and among the soil depth
categories. Thus, the model included temperature as a covariate to estimate the least-
squares mean trait value. However, because the rate–temperature relationship might differ
among species, a general linear model specifically testing for whether interactions are
present between temperature and status (indigenous vs. invasive), and between temperature
and depth category was initially implemented (see Quinn and Keough 2002: 349, the test
was implemented in Statistica v. 8). No slope heterogeneity was found, and in effect this
formed the test of whether the linear reaction norms differed among the species.
Because each status by depth group was typically only represented by a single species
(except for the ‘epedaphic invasive’ group which included Isotomurus cf. palustris and
Pogonognathellus flavescens), the power of the above tests to assess the extent to which
any differences among the status and depth groups might be considered general, could be
thought of as limited. Therefore, further to examine the extent to which these differences
might be biologically significant we also undertook a randomization analysis where
development rate data were randomized among the same number of status and depth
groups. Here we used a resample-without-replacement protocol for both status and depth
categorical variables (see Parr et al. 2005). Then, we calculated mean log10 egg devel-
opment rate for 100 random re-samples (run in Microsoft Excel) and computed their 95%
confidence limits. This allowed us to undertake a comparison of randomized depth/inva-
sive status samples relative to observed egg development rate for each category. We
reasoned that if particular status and depth group means were different to a random
selection of the data, additional confidence could be placed in their biological significance.
Because phylogenetic relatedness may have played a major role in influencing the
patterns in trait variation found among these species (given that the data set for the
indigenous species included only the genus Cryptopygus) we undertook a second set of
analyses using phylogenetic generalized least squares (PGLS). The PGLS and generalized
estimating equation (GEE) analyses were implemented in the APE module (Paradis et al.
2004) of the R language open-source software package (following methods outlined in
Halsey et al. 2006). The species level phylogeny was constructed using mitochondrial COI
sequence data (Supplementary material S3). The tree concurred with previously published
analyses (Stevens et al. 2006), with the exception of Cryptopygus tricuspis clustering with
Ceratophysella denticulata and Pogonognathellus flavescens, as opposed to with other
Cryptopygus species (C. antarcticus travei and C. dubius). No reason exists to suppose that
Cryptopygus is paraphyletic (based on limited 28S data and on Stevens et al. 2006). Rather
the outcome was likely due to hemiplasy (Avise and Robinson 2008). Therefore, to
determine branch lengths, the evolutionary tree was constrained to group C. tricuspiswithin the other Cryptopygus species. Branch lengths were determined in PAUP*
1370 Evol Ecol (2010) 24:1365–1380
123

(Swofford 2001) using maximum likelihood analyses and plotted on the tree. Thereafter,
PGLS, using these estimated branch lengths in units of mutational change, was used to
assess the strength of the effect of the hypothetical phylogeny on the dependent variable by
estimating lambda (k). k is an estimate of the phylogenetic covariance in the data and lies
between 0 and 1; k = 1 indicates a strong influence of phylogeny while k = 0 indicates
little influence of phylogeny (see Halsey et al. 2006). In the GEE models significance
levels were calculated from phylogenetically adjusted degrees of freedom. The PGLS and
GEE analyses were used to investigate variation among the least squares means obtained
from the General Linear Model.
For analyses of hatching success variation among the indigenous and invasive species at
the higher temperatures (15 and 25�C), a generalised linear model analysis, assuming a
binomial distribution and a logit link function, was undertaken in SAS (V 9.1) to assess
effects of temperature variation, status (indigenous vs. invasive species) and interactions
between temperature and species.
Owing to the substantial emphasis that has been placed recently on identifying how the
temperature optimum (Topt), rate at the temperature optimum (umax) from performance
curves might differ among species, environments and treatments (e.g., Gilchrist 1996;
Kingsolver and Huey 1998; Angilletta et al. 2002, 2003; Deere and Chown 2006; Deutsch
et al. 2008) these traits were estimated for each species. This was done using a modification
of the method proposed by Angilletta (2006). For each species, the mean rate per batch at
each temperature was calculated and a curve was fitted to these data using TableCurve 2D
(SYSTAT Inc, 2002, San Jose, California, USA). The curve with the largest coefficient of
determination was selected, irrespective of the number of terms, because the aim was to
find the best-fitting curve so that Topt and umax could be determined from the fitted curve (in
the latter case using Gilchrist’s (1996) approach with 0.1�C temperature increments).
Typically, the best fits were provided by the Exponentially Modified Gaussian, and Half
Gaussian Modified Gaussian curves (Supplementary material S1). In the case of Crypto-pygus antarcticus travei, individual level data were used because mean data were available
only at four temperatures so precluding use of the peaked curve equations of TableCurve
2D. In this case a LogNormal4 curve provided the best fit (Supplementary material S1).
Values for Topt and umax were compared among the indigenous and invasive groups using
a single classification ANOVA.
Results
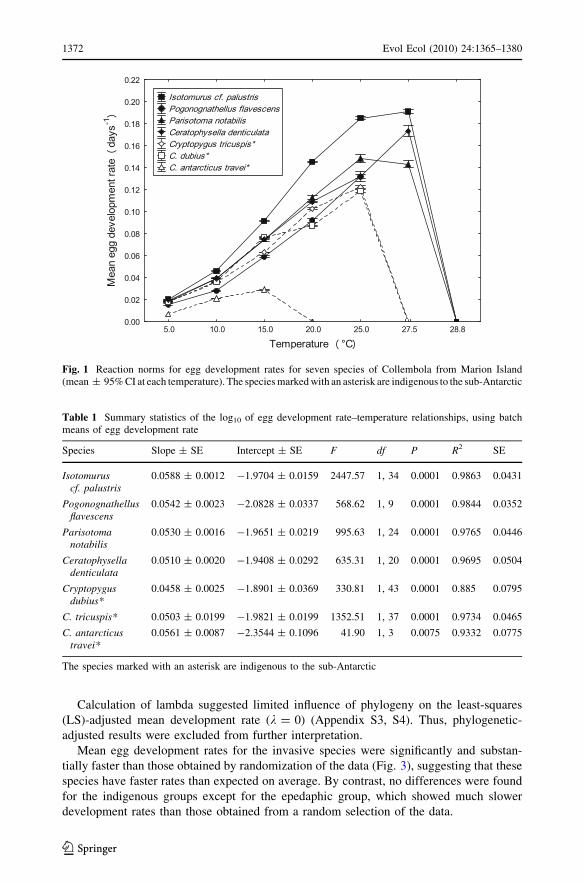
Temperature had the expected effect on development rate (Fig. 1; Table 1), with maximum
development temperatures found at 27.5�C in the invasive species Isotomurus cf. palustris,
Ceratophysella denticulata and Parisotoma notabilis, and typically at lower temperatures
for the indigenous ones (see Appendix S2 for mean data values). In the generalized linear
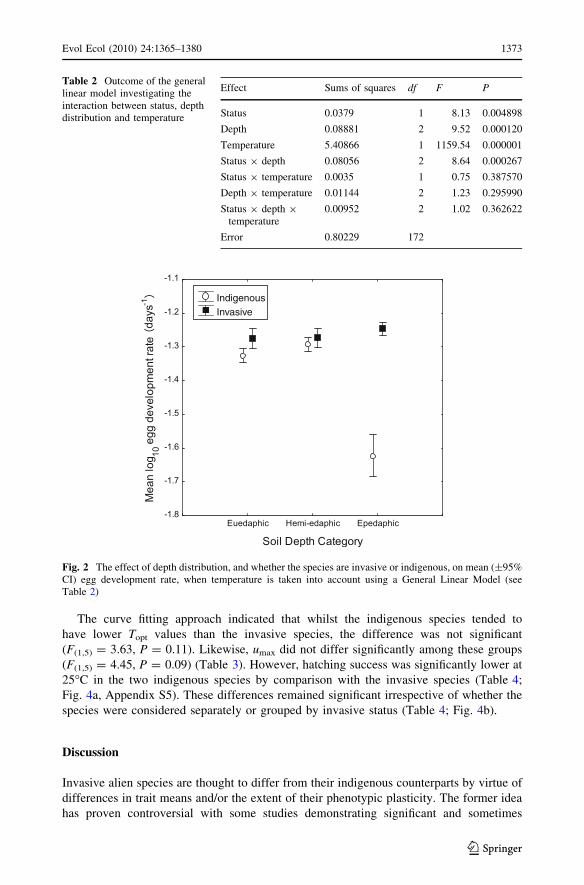
model analyses, the homogeneity of slopes model revealed that depth and status had a
significant effect on the mean egg development rate, but no slope heterogeneity was
identified among the groups (Table 2). Means and confidence intervals for each of the
groups indicated that invasive species typically had faster mean egg development rates
than the indigenous species except in the hemi-edaphic depth group (Fig. 2). Because the
slopes of the rate–temperature relationships did not differ significantly among the groups
(including no 3-way interaction), it is clear that the reaction norms, or form and extent of
plasticity, do not differ among them.
Evol Ecol (2010) 24:1365–1380 1371
123
Calculation of lambda suggested limited influence of phylogeny on the least-squares
(LS)-adjusted mean development rate (k = 0) (Appendix S3, S4). Thus, phylogenetic-
adjusted results were excluded from further interpretation.
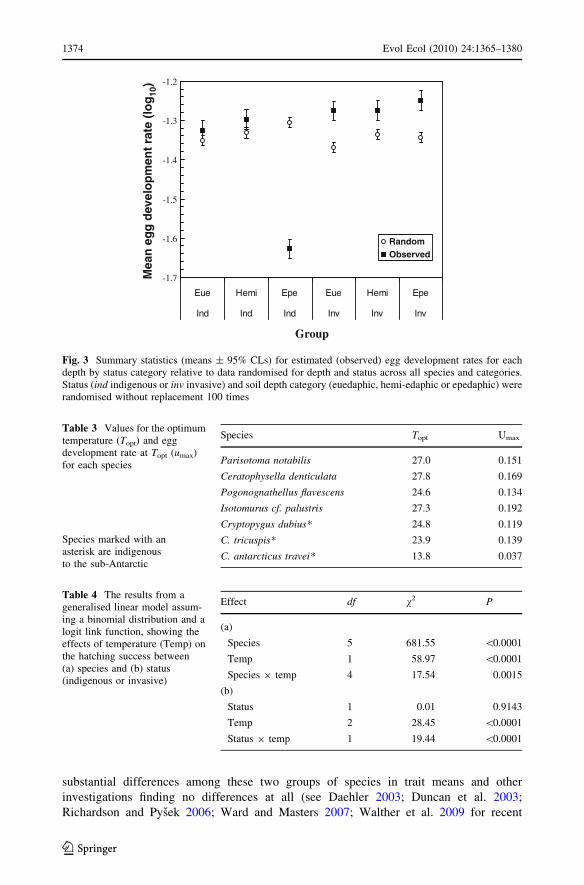
Mean egg development rates for the invasive species were significantly and substan-
tially faster than those obtained by randomization of the data (Fig. 3), suggesting that these
species have faster rates than expected on average. By contrast, no differences were found
for the indigenous groups except for the epedaphic group, which showed much slower
development rates than those obtained from a random selection of the data.
Fig. 1 Reaction norms for egg development rates for seven species of Collembola from Marion Island(mean ± 95% CI at each temperature). The species marked with an asterisk are indigenous to the sub-Antarctic
Table 1 Summary statistics of the log10 of egg development rate–temperature relationships, using batchmeans of egg development rate
Species Slope ± SE Intercept ± SE F df P R2 SE
Isotomuruscf. palustris
0.0588 ± 0.0012 -1.9704 ± 0.0159 2447.57 1, 34 0.0001 0.9863 0.0431
Pogonognathellusflavescens
0.0542 ± 0.0023 -2.0828 ± 0.0337 568.62 1, 9 0.0001 0.9844 0.0352
Parisotomanotabilis
0.0530 ± 0.0016 -1.9651 ± 0.0219 995.63 1, 24 0.0001 0.9765 0.0446
Ceratophyselladenticulata
0.0510 ± 0.0020 -1.9408 ± 0.0292 635.31 1, 20 0.0001 0.9695 0.0504
Cryptopygusdubius*
0.0458 ± 0.0025 -1.8901 ± 0.0369 330.81 1, 43 0.0001 0.885 0.0795
C. tricuspis* 0.0503 ± 0.0199 -1.9821 ± 0.0199 1352.51 1, 37 0.0001 0.9734 0.0465
C. antarcticustravei*
0.0561 ± 0.0087 -2.3544 ± 0.1096 41.90 1, 3 0.0075 0.9332 0.0775
The species marked with an asterisk are indigenous to the sub-Antarctic
1372 Evol Ecol (2010) 24:1365–1380
123
The curve fitting approach indicated that whilst the indigenous species tended to
have lower Topt values than the invasive species, the difference was not significant
(F(1,5) = 3.63, P = 0.11). Likewise, umax did not differ significantly among these groups
(F(1,5) = 4.45, P = 0.09) (Table 3). However, hatching success was significantly lower at
25�C in the two indigenous species by comparison with the invasive species (Table 4;
Fig. 4a, Appendix S5). These differences remained significant irrespective of whether the
species were considered separately or grouped by invasive status (Table 4; Fig. 4b).
Discussion
Invasive alien species are thought to differ from their indigenous counterparts by virtue of
differences in trait means and/or the extent of their phenotypic plasticity. The former idea
has proven controversial with some studies demonstrating significant and sometimes
Table 2 Outcome of the generallinear model investigating theinteraction between status, depthdistribution and temperature
Effect Sums of squares df F P
Status 0.0379 1 8.13 0.004898
Depth 0.08881 2 9.52 0.000120
Temperature 5.40866 1 1159.54 0.000001
Status 9 depth 0.08056 2 8.64 0.000267
Status 9 temperature 0.0035 1 0.75 0.387570
Depth 9 temperature 0.01144 2 1.23 0.295990
Status 9 depth 9temperature
0.00952 2 1.02 0.362622
Error 0.80229 172
Indigenous Invasive
Fig. 2 The effect of depth distribution, and whether the species are invasive or indigenous, on mean (±95%CI) egg development rate, when temperature is taken into account using a General Linear Model (seeTable 2)
Evol Ecol (2010) 24:1365–1380 1373
123
substantial differences among these two groups of species in trait means and other
investigations finding no differences at all (see Daehler 2003; Duncan et al. 2003;
Richardson and Pysek 2006; Ward and Masters 2007; Walther et al. 2009 for recent
-1.7
-1.6
-1.5
-1.4
-1.3
-1.2
Eue Hemi Epe Eue Hemi Epe
Ind Ind Ind Inv Inv Inv
Group
Mea
n e
gg
dev
elo
pm
ent
rate
(lo
g10
)
RandomObserved
Fig. 3 Summary statistics (means ± 95% CLs) for estimated (observed) egg development rates for eachdepth by status category relative to data randomised for depth and status across all species and categories.Status (ind indigenous or inv invasive) and soil depth category (euedaphic, hemi-edaphic or epedaphic) wererandomised without replacement 100 times
Table 3 Values for the optimumtemperature (Topt) and eggdevelopment rate at Topt (umax)for each species
Species marked with anasterisk are indigenousto the sub-Antarctic
Species Topt Umax
Parisotoma notabilis 27.0 0.151
Ceratophysella denticulata 27.8 0.169
Pogonognathellus flavescens 24.6 0.134
Isotomurus cf. palustris 27.3 0.192
Cryptopygus dubius* 24.8 0.119
C. tricuspis* 23.9 0.139
C. antarcticus travei* 13.8 0.037
Table 4 The results from ageneralised linear model assum-ing a binomial distribution and alogit link function, showing theeffects of temperature (Temp) onthe hatching success between(a) species and (b) status(indigenous or invasive)
Effect df v2 P
(a)
Species 5 681.55 \0.0001
Temp 1 58.97 \0.0001
Species 9 temp 4 17.54 0.0015
(b)
Status 1 0.01 0.9143
Temp 2 28.45 \0.0001
Status 9 temp 1 19.44 \0.0001
1374 Evol Ecol (2010) 24:1365–1380
123
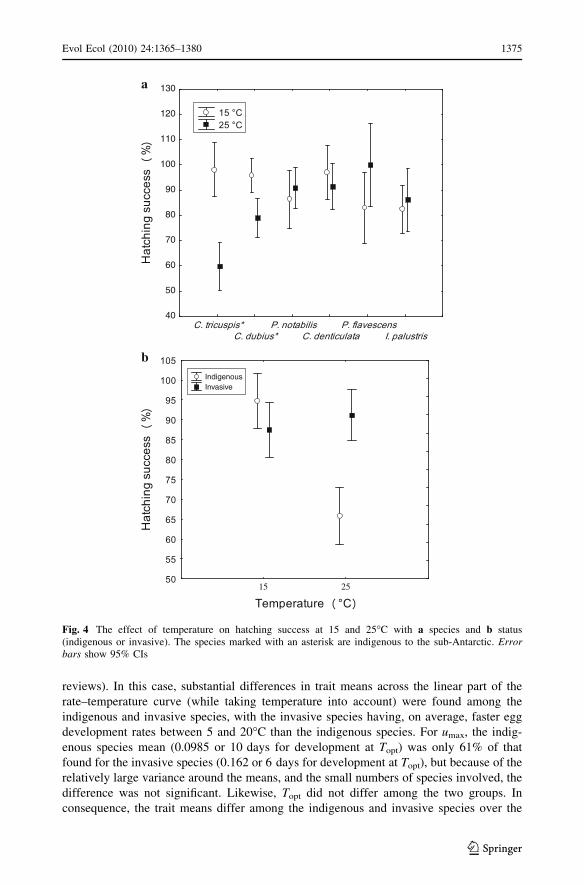
reviews). In this case, substantial differences in trait means across the linear part of the
rate–temperature curve (while taking temperature into account) were found among the
indigenous and invasive species, with the invasive species having, on average, faster egg
development rates between 5 and 20�C than the indigenous species. For umax, the indig-
enous species mean (0.0985 or 10 days for development at Topt) was only 61% of that
found for the invasive species (0.162 or 6 days for development at Topt), but because of the
relatively large variance around the means, and the small numbers of species involved, the
difference was not significant. Likewise, Topt did not differ among the two groups. In
consequence, the trait means differ among the indigenous and invasive species over the
40
50
60
70
80
90
100
110
120
130
Indigenous Invasive
15 25
a
b
Fig. 4 The effect of temperature on hatching success at 15 and 25�C with a species and b status(indigenous or invasive). The species marked with an asterisk are indigenous to the sub-Antarctic. Errorbars show 95% CIs
Evol Ecol (2010) 24:1365–1380 1375
123

5–20�C part of the reaction norm, and possibly at its optimum. Importantly, however,
hatching success was lower in the indigenous species at 25�C (zero in C. antarcticus above
20�C) than in the invasive species, although such differences were not apparent at 15�C.
Thus, the invasive species appear to be at an advantage in terms of egg development rate
and hatching success at high temperature compared with the indigenous species in most
groups, except perhaps for the hemi-edaphic species where the difference was not
significant.
Given the fact that each status x depth category was typically only represented by a
single species (a consequence of the nature of the springtail assemblages at the site we
investigated—see Gabriel et al. 2001), it might be argued that the differences among the
groups are simply an artefact of the species investigated. The randomization procedure
suggested that, perhaps with the exception of the slow developing C. antarcticus, the
indigenous species are no different to what might be expected for a random assortment of
egg development rates. However, the invasive species always had faster rates than might
be expected from a random re-allocation of the data. Thus, further support is provided for
the idea that the invasive species typically have faster development rates than the indig-
enous ones. The limited influence of phylogeny (k = 0) also showed that the result is not
confounded by a strong phylogenetic signal in the data. In consequence, at least for this
assemblage of springtails, it does appear that the invasive species typically have much
faster development rates than the indigenous ones, and a lower susceptibility to hatching
failure at higher temperatures.
This outcome of a significant difference in trait means is in keeping with what has been
found for adult lower and upper lethal temperatures for a similar group of species from
Marion Island (including many of those studied here), where the indigenous species tended
to have lower freezing points (=lower lethal temperature in this case) and lower, high
temperature tolerances than the invasive species (Slabber et al. 2007). By contrast, duration
of survival of dry conditions (75% RH) did not differ among the indigenous and invasive
species for the adults, although acclimation to high temperature certainly increased sur-
vival time in the invasive species, whilst often having the converse effect on the indigenous
ones (Chown et al. 2007). In consequence, it appears that conclusions about whole
organismal responses cannot simply be generalized. Rather, the responses likely vary from
trait to trait, and in concert will determine the performance outcome for the organism
concerned (see also van Kleunen and Johnson 2007). Nonetheless, overall it appears that
warmer and drier conditions will certainly benefit invasive springtail species on Marion
Island relative to their indigenous counterparts.
Across the 5–20�C part of the reaction norm, the slopes of the R–T relationships did not
differ among the species. This outcome is in keeping with that found for other traits in the
same species, notably survival time under desiccating conditions and lower and upper
critical thermal limits (Chown et al. 2007; Slabber et al. 2007), and with other investigations
where no differences in plasticity have been found (see Daehler 2003; Lee et al. 2007).
However, it differs from a range of studies where significant and often substantial differ-
ences in phenotypic plasticity among indigenous and invasive species within the same taxa
have been found (e.g., Trussell and Smith 2000; Rosecchi et al. 2001; Stachowicz et al.
2002; Duncan et al. 2003; Dzialowski et al. 2003; Richardson and Pysek 2006).
From a climate change perspective the present data, in the context of what is known
about means and reaction norms of other traits in these species (Chown et al. 2007; Slabber
et al. 2007), suggest that the invasive species should be predominant in low elevation warm
areas of Marion Island, and should continue to be favoured by ongoing warming and
drying. This is indeed what has been found to date. Ecological surveys of the island have
1376 Evol Ecol (2010) 24:1365–1380
123

found exceptionally high densities and dominance of the invasive species at low elevations
across the island, but an absence of these species at higher, cold, dry elevations (Gabriel
et al. 2001). Moreover, field experiments that have simulated predicted warming and
drying on the island (see le Roux and McGeoch 2008) resulted in substantial declines in the
abundances of the indigenous species and little change in the abundance of an invasive
species (Chown et al. 2007). The outcomes of this study also support previous predictions,
made on the grounds of qualitative comparisons of insect life history data, that invasive
species will be favoured by climate change at the island given their short development
times (Chown et al. 2002). However, they contradict the idea that steeper rate–temperature
relationships in invasive than in indigenous species will further benefit the former under
warmer conditions (Chown et al. 2002). Nonetheless, further evidence that climate change
is benefiting the invasive springtails would be provided by demonstrations of an altitudinal
range expansion. This work is presently underway.
In conclusion, the present study has demonstrated that mean egg development rate and
hatching success at high temperature, but not the thermal reaction norms of development
rate, differ consistently between indigenous and invasive springtails on Marion Island, and
that these differences are likely to have substantial implications for assemblage structure as
the climate continues to change (see also Janion et al. 2009). In so doing it demonstrates
how, by considering mean and plasticity of traits separately (see Ghalambor et al. 2007),
insight into the mechanistic basis of interactions among climate change and invasion may
be obtained. It also provides further evidence that the interactions among climate change
and invasion are likely to be synergistic (see also Stachowicz et al. 2002) and that such
interactions need urgent further investigation (Brook 2008). Whether our results for
springtails can be generalized more broadly to this group remains to be determined through
further study of assemblages of indigenous and invasive species elsewhere. Nonetheless, it
supports a growing body of work showing that the traits of indigenous and invasive species
differ considerably, probably owing to the characteristics that ensure survival and estab-
lishment in the first place (Blackburn et al. 2009).
Acknowledgments We thank Erika Nortje, Heidi Sjursen Konestabo and various members of the MarionIsland relief teams for assistance in the field. Bettine Jansen van Vuuren and Angela McGaughran assistedwith the COI sequence data and phylogenetic analysis. Janne Bengtsson, Melodie McGeoch and twoanonymous referees provided useful comments on a previous version of the ms. The South African NationalAntarctic Programme provided logistic support. This work was funded partly by a SA-Norway scienceliaison grant awarded jointly to HPL and SLC. The work forms a contribution to the SCAR EBAProgramme.
References
Agrawal AA (2001) Phenotypic plasticity in the interactions and evolution of species. Science 294:321–326.doi:10.1126/science.1060701
Angilletta MJ (2006) Estimating and comparing thermal performance curves. J Thermal Biol 31:541–545.doi:10.1016/j.jtherbio.2006.06.002
Angilletta MJ, Niewiarowski PH, Navas CA (2002) The evolution of thermal physiology in ectotherms.J Thermal Biol 27:249–268. doi:10.1016/S0306-4565(01)00094-8
Angilletta MJ, Wilson RS, Navas CA et al (2003) Tradeoffs and the evolution of thermal reaction norms.Trends Ecol Evol 18:234–240. doi:10.1016/S0169-5347(03)00087-9
Avise JC, Robinson TJ (2008) Hemiplasy: a new term in the lexicon of phylogenetics. Syst Biol57:503–507. doi:10.1080/10635150802164587
Evol Ecol (2010) 24:1365–1380 1377
123

Baker HG (1965) Characteristics and modes of origin of weeds. In: Baker HG, Stebbins GL (eds) Thegenetics of colonizing species. Academic Press, New York, pp 147–168
Birkemoe T, Leinaas HP (2000) Effects of temperature on the development of an Arctic Collembola(Hypogastrura tullbergi). Funct Ecol 14:693–700. doi:10.1046/j.1365-2435.2000.00478.x
Blackburn TM, Lockwood JL, Cassey P (2009) Avian invasions. The ecology and evolution of exotic birds.Oxford University Press, Oxford
Brook BW (2008) Synergies between climate change, extinctions and invasive invertebrates. Wildl Res35:249–252. doi:10.1071/WR07116
Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers and global change. TrendsEcol Evol 23:453–460. doi:10.1016/j.tree.2008.03.011
Cannon RJC (1998) The implications of predicted climate change for insect pests in the UK, with emphasison non-indigenous species. Global Change Biol 4:785–796. doi:10.1046/j.1365-2486.1998.00190.x
Carroll SP, Fox CW (2007) Dissecting the evolutionary impacts of plant invasions: bugs and beetles asnative guides. Global Change Biol 13:1644–1657. doi:10.1111/j.1365-2486.2007.01403.x
Chown SL, Froneman PW (2008) The Prince Edward Islands. Land–sea interactions in a changing climate.African Sun Media, Stellenbosch
Chown SL, Gaston KJ (2008) Macrophysiology for a changing world. Proc R Soc B 275:1469–1478. doi:10.1098/rspb.2008.0137
Chown SL, Nicolson SW (2004) Insect physiological ecology. Mechanisms and patterns. Oxford UniversityPress, Oxford
Chown SL, McGeoch MA, Marshall DJ (2002) Diversity and conservation of invertebrates on the sub-Antarctic Prince Edward Islands. Afr Entomol 10:67–82
Chown SL, Slabber S, McGeoch MA et al (2007) Phenotypic plasticity mediates climate change responsesamong invasive and indigenous arthropods. Proc R Soc B 274:2661–2667. doi:10.1098/rspb.2007.0772
Cooper J, Condy PR (1988) Environmental conservation at the sub-Antarctic Prince Edward Islands: areview and recommendations. Environ Conserv 15:317–326
Daehler CC (2003) Performance comparisons of co-occurring native and alien invasive plants: implicationsfor conservation and restoration. Ann Rev Ecol Evol Syst 34:183–211. doi:10.1146/annurev.ecolsys.34.011802.132403
David JR, Gibert P, Gravot E et al (1997) Phenotypic plasticity and developmental temperature in Dro-sophila: analysis and significance of reaction norms of morphometrical traits. J Thermal Biol22:441–451. doi:10.1016/S0306-4565(97)00063-6
Davis MB, Shaw RG (2001) Range shifts and adaptive responses to quaternary climate change. Science292:673–679. doi:10.1126/science.292.5517.673
de Jong G (1995) Phenotypic plasticity as a product of selection in a variable environment. Am Nat145:493–512. doi:10.1086/285752
de Jong G (2005) Evolution of phenotypic plasticity: patterns of plasticity and the emergence of ecotypes.New Phytol 166:101–118
de Jong G, van der Have TM (2008) Temperature dependence of development rate, growth rate and size:from biophysics to adaptation. In: Whitman DW, Ananthakrishnan TN (eds) Phenotypic plasticity ofinsects: mechanisms and consequence. Science, Enfield, pp 461–526
de Mazancourt C, Johnson E, Barraclough TG (2008) Biodiversity inhibits species’ evolutionary responsesto changing environments. Ecol Lett 11:380–388. doi:10.1111/j.1461-0248.2008.01152.x
Deere JA, Chown SL (2006) Testing the beneficial acclimation hypothesis and its alternatives for locomotorperformance. Am Nat 168:630–644. doi:10.1086/508026
Deharveng L (1981) Collemboles des iles subantarctiques de l’Ocean Indien Mission J. Trave 1972–1973.Comite National Francaise des Recherches Antarctiques 48:33–108
Deutsch CA, Tewksbury JJ, Huey RB et al (2008) Impacts of climate warming on terrestrial ectothermsacross latitude. Proc Natl Acad Sci USA 105:6668–6672. doi:10.1073/pnas.0709472105
Diamond JM (1989) Overview of recent extinctions. In: Western D, Pearl MC (eds) Conservation for thetwenty-first century. Oxford University Press, Oxford, pp 37–41
Didham RK, Tylianakis JM, Gemmell NJ et al (2007) Interactive effects of habitat modification and speciesinvasion on native species decline. Trends Ecol Evol 22:489–496. doi:10.1016/j.tree.2007.07.001
Dukes J, Mooney HA (1999) Does global change increase the success of biological invaders? Trends EcolEvol 14:135–139. doi:10.1016/S0169-5347(98)01554-7
Duncan RP, Blackburn TM, Sol D (2003) The ecology of bird introductions. Annu Rev Ecol Evol Syst34:71–98. doi:10.1146/annurev.ecolsys.34.011802.132353
Dybdahl MF, Kane SL (2005) Adaptation vs. phenotypic plasticity in the success of a clonal invader.Ecology 86:1592–1601. doi:10.1890/04-0898
1378 Evol Ecol (2010) 24:1365–1380
123

Dzialowski AR, Lennon JT, O’Brien WJ et al (2003) Predator-induced phenotypic plasticity in the exoticcladoceran Daphnia lumholtzi. Freshw Biol 48:1593–1602. doi:10.1046/j.1365-2427.2003.01111.x
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, LondonFjellberg A (1998) Fauna Entomologica Scandinavica Volume 35. The Collembola of Fennoscandia and
Denmark. Part I: Poduromorpha. Brill, LeidenFrazier MR, Huey RB, Berrigan D (2008) Thermodynamics constrains the evolution of insect population
growth rates: ‘‘warmer is better’’. Am Nat 168:512–520. doi:10.1086/285797Frenot Y, Chown SL, Whinam J et al (2005) Biological invasions in the Antarctic: extent, impacts and
implications. Biol Rev 80:45–72. doi:10.1017/S1464793104006542Gabriel AGA, Chown SL, Barendse J et al (2001) Biological invasions on Southern Ocean islands: the
Collembola of Marion Island as a test of generalities. Ecography 24:421–430. doi:10.1111/j.1600-0587.2001.tb00477.x
Gaston KJ, Chown SL, Mercer RD (2001) The animal species-body size distribution of Marion Island. ProcNatl Acad Sci USA 98:14493–14496. doi:10.1073/pnas.251332098
Geister TL, Lorenz MW, Hoffmann KH et al (2009) Energetics of embryonic development: effects oftemperature in egg and hatchling composition in a butterfly. J Comp Physiol B 179:87–98. doi:10.1007/s00360-008-0293-5
Ghalambor CK, McKay JK, Carroll SP et al (2007) Adaptive versus non-adaptive phenotypic plasticity andthe potential for contemporary adaptation in new environments. Funct Ecol 21:394–407. doi:10.1111/j.1365-2435.2007.01283.x
Gilchrist GW (1996) A quantitative genetic analysis of thermal sensitivity in the locomotor performancecurve of Aphidius ervi. Evolution 50:1560–1572
Halsey LG, Butler PJ, Blackburn TM (2006) A phylogenetic analysis of the allometry of diving. Am Nat167:276–287. doi:10.1086/499439
Hobbs RJ, Mooney HA (2005) Invasive species in a changing world: the interactions between global changeand invasives. In: Mooney HJ, Mack RN, McNeely JA, Neville LE, Schei PJ, Waage JK (eds) Invasivealien species. A new synthesis. Island Press, Washington, pp 310–331
Holzapfel AM, Vinebrooke RD (2005) Environmental warming increases invasion potential of alpine lakecommunities by imported species. Global Change Biol 11:2009–2015. doi:10.1111/j.1365-2486.2005.001057.x
Hopkin S (1997) Biology of the springtails. Insecta: Collembola. Oxford University Press, OxfordHugo EA, McGeoch MA, Marshall DJ et al (2004) Fine scale variation in microarthropod communities
inhabiting the keystone species Azorella selago on Marion Island. Polar Biol 27:446–473. doi:10.1007/s00300-004-0614-4
Ikemoto T (2005) Intrinsic optimum temperature for development of insects and mites. Environ Entomol34:1377–1387. doi:10.1603/0046-225X-34.6.1377
Izem R, Kingsolver JG (2005) Variation in continuous reaction norms: quantifying directions of biologicalinterest. Am Nat 166:277–289. doi:10.1086/431314
Janion C, Worland MR, Chown SL (2009) Assemblage level variation in lower lethal temperature: the roleof invasive species on sub-Antarctic Marion Island. Physiol Entomol 34:284–291. doi:10.1111/j.1365-3032.2009.00689.x
Kingsolver JG, Huey RB (1998) Evolutionary analyses of morphological and physiological plasticity inthermally variable environments. Am Zool 38:545–560. doi:10.1093/icb/38.3.545
Lawton JH, Brown KC (1986) The population and community ecology of invading insects. Phil Trans R SocB 314:607–617
le Roux PC, McGeoch MA (2008) Changes in climate extremes, variability and signature on sub-AntarcticMarion Island. Clim Change 86:309–329. doi:10.1007/s10584-007-9259-y
Lee CE (2002) Evolutionary genetics of invasive species. Trends Ecol Evol 17:386–391. doi:10.1016/S0169-5347(01)02405-3
Lee CE, Remfert JL, Gelembiuk GW (2003) Evolution of physiological tolerance and performance duringfreshwater invasions. Integr Comp Biol 43:439–449. doi:10.1093/icb/43.3.439
Lee C, Remfert J, Chang Y-M (2007) Response to selection and evolvability of invasive populations.Genetica 129:179–192. doi:10.1016/S0169-5347(01)02405-3
Myburgh M, Chown SL, Daniels SR et al (2007) Population structure, propagule pressure, and conservationbiogeography in the sub-Antarctic: lessons from indigenous and invasive springtails. Divers Distr13:143–154. doi:10.1111/j.1472-4642.2007.00319.x
Paradis E, Claude J, Strimmer K (2004) APE: analysis of phylogenetics and evolution in R language.Bioinformatics 20:289–290
Parmesan C (2007) Influences of species, latitudes and methodologies on estimates of phenological responseto global warming. Global Change Biol 13:1860–1872. doi:10.1111/j.1365-2486.2007.01404.x
Evol Ecol (2010) 24:1365–1380 1379
123

Parr CL, Sinclair BJ, Andersen AN et al (2005) Constraint and competition in assemblages: a cross-continental and modeling approach for ants. Am Nat 165:481–494
Potapov M (2001) Synopses on Palaearctic Collembola, volume 3, Isotomidae. In: Dunger W (ed) Staa-tliches Museum fur Naturkunde, Gorlitz
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge UniversityPress, Cambridge
Richardson DM, Pysek P (2006) Plant invasions: merging the concepts of species invasiveness and com-munity invasibility. Progress Phys Geogr 30:409–431. doi:10.1191/0309133306pp490pr2006
Roff DA (2002) Life history evolution. Sinauer Associates, SunderlandRosecchi E, Thomas F, Crivelli AJ (2001) Can life-history traits predict the fate of introduced species? A
case study on two cyprinid fish in southern France. Freshw Biol 46:845–853. doi:10.1046/j.1365-2427.2001.00715.x
Rusek J (1998) Biodiversity of Collembola and their functional role in the ecosystem. Biodiv Conserv7:1207–1219
Sala OE, Chapin FS, Armesto JJ et al (2000) Global biodiversity scenarios for the year 2100. Science287:1770–1774. doi:10.1126/science.287.5459.1770
Scheiner SM (1993) Genetics and evolution of phenotypic plasticity. Ann Rev Ecol Syst 24:35–68. doi:10.1146/annurev.es.24.110193.000343
Sibly RM, Calow P (1986) Physiological ecology of animals. An evolutionary approach. Blackwell Sci-entific Publications, Oxford
Slabber S, Worland MR, Leinaas HP et al (2007) Acclimation effects on thermal tolerances of springtailsfrom sub-Antarctic Marion Island: indigenous and invasive species. J Insect Physiol 53:113–125. doi:10.1016/j.jinsphys.2006.10.010
Stachowicz JJ, Terwin JR, Whitlatch RB et al (2002) Linking climate change and biological invasions:ocean warming facilitates nonindigenous species invasions. Proc Natl Acad Sci USA 99:15497–15500.doi:10.1073/pnas.242437499
Stearns SC (1992) The evolution of life histories. Oxford University Press, OxfordStevens M, Greenslade P, Hogg ID et al (2006) Southern Hemisphere springtails: could any have survived
glaciation of Antarctica? Mol Biol Evol 23:574–882. doi:10.1093/molbev/msj073Stillwell RC, Fox CW (2005) Complex patterns of phenotypic plasticity: interactive effects of temperature
during rearing and oviposition. Ecology 86:924–934. doi:10.1890/04-0547Stohlgren TJ, Barnett DT, Jarnevich CS et al (2008) The myth of plant species saturation. Ecol Lett
11:313–322. doi:10.1111/j.1461-0248.2008.01153.xSwofford DL (2001) PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods). Sinauer
Associates, SunderlandTheoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous
species success during four stages of invasion. New Phytol 176:256–273. doi:10.1111/j.1469-8137.2007.02207.x
Trudgill DL, Honek A, van Straalen NM (2005) Thermal time: concepts and utility. Ann Appl Biol146:1–14. doi:10.1111/j.1744-7348.2005.04088.x
Trussell GC, Smith LD (2000) Induced defenses in response to an invading crab predator: an explanation ofhistorical and geographic phenotypic change. Proc Natl Acad Sci USA 97:2123–2127. doi:10.1073/pnas.040423397
van Kleunen M, Fischer M (2005) Constraints on the evolution of adaptive phenotypic plasticity in plants.New Phytol 166:49–60. doi:10.1111/j.1469-8137.2004.01296.x
van Kleunen M, Johnson SD (2007) South African Iridaceae with rapid and profuse seedling emergence aremore likely to become naturalized in other regions. J Ecol 95:674–681. doi:10.1111/j.1365-2745.2007.01250.x
van Kleunen M, Manning JC, Pasqualetto V et al (2008) Phylogenetically independent associations betweenautonomous self fertilization and plant invasiveness. Am Nat 171:195–201. doi:10.1086/525057
van Straalen NM (1994) Adaptive significance of temperature responses in Collembola. Acta Zool Fennica195:135–142
Via S, Gomulkiewicz R, DeJong G et al (1995) Adaptive phenotypic plasticity: consensus and controversy.Trends Ecol Evol 10:212–217. doi:10.1016/S0169-5347(00)89061-8
Walther G-R, Roques A, Hulme PE et al (2009) Alien species in a warmer world: risks and opportunities.Trends Ecol Evol 24:686–693. doi:10.1016/j.tree.2009.06.008
Ward NL, Masters GJ (2007) Linking climate change and species invasion: an illustration using insectherbivores. Global Change Biol 13:1605–1615. doi:10.1111/j.1365-2486.2007.01399.x
Warren MS, Hill JK, Thomas JA et al (2001) Rapid responses of British butterflies to opposing forces ofclimate and habitat change. Nature 414:65–69. doi:10.1038/35102054
1380 Evol Ecol (2010) 24:1365–1380
123
Related Documents











