RESEARCH ARTICLE M. Gerwig K. Haerter K. Hajjar A. Dimitrova M. Maschke F. P. Kolb A. F. Thilmann E. R. Gizewski D. Timmann Trace eyeblink conditioning in human subjects with cerebellar lesions Received: 22 March 2005 / Accepted: 28 July 2005 / Published online: 18 November 2005 Ó Springer-Verlag 2005 Abstract Trace eyeblink conditioning was investigated in 31 patients with focal cerebellar lesions and 19 age- matched controls. Twelve patients presented with lesions including the territory of the superior cerebellar artery (SCA). In 19 patients lesions were restricted to the ter- ritory of the posterior inferior cerebellar artery (PICA). A 3D magnetic resonance imaging was used to deter- mine the extent of the cortical lesion and possible involvement of cerebellar nuclei. Eyeblink conditioning was performed using a 40 ms tone as conditioned stim- ulus (CS) followed by a stimulus free trace-interval of 400 ms and a 100 ms air-puff as unconditioned stimulus (US). In SCA patients with lesions including parts of the cerebellar interposed nucleus trace eyeblink conditioning was significantly impaired. Pure cortical lesions of the superior cerebellum were not sufficient to reduce acqui- sition of trace conditioned eyeblink responses. PICA patients were not impaired in trace eyeblink condition- ing. Consistent with animal studies the findings of the present human lesion study suggest that, in addition to forebrain areas, the interposed nucleus is of importance in trace eyeblink conditioning. Although cortical cere- bellar areas appear less important in trace compared with delay eyeblink conditioning, the present data strengthen the view that cerebellar structures contribute to different forms of eyeblink conditioning paradigms. Keywords Classical conditioning Cerebellum Interposed nucleus Associative learning Introduction In the last decades classical conditioning of the eyeblink reflex has been used as a simple and successful model in research of associative motor learning and memory in animals and humans (Thompson et al. 1997, 2000; Yeo and Hesslow 1998). Most commonly, delay eyeblink conditioning has been investigated, in which the condi- tioned stimulus (CS) overlaps and coterminates with the unconditioned stimulus (US) (Green and Woodruff-Pak 2000). Trace eyeblink conditioning, a more complex form of associative learning, is characterized by a stim- ulus-free trace interval that separates the CS from the US requiring a ‘‘memory-trace’’ to associate the CS with the later US. It has been suggested that the delay para- digm may represent a basic form of procedural (implicit) learning and that trace eyeblink conditioning may serve as a model of declarative (explicit) memory (for review: Christian and Thompson 2003). CR-acquisition in delay eyeblink conditioning has been shown essentially dependent on neural circuitry within the cerebellum (for review: Thompson et al. 1997; Yeo and Hesslow 1998; Steinmetz 2000), but is not sig- nificantly reduced following hippocampal lesions (Weiss et al. 1999; Tseng et al. 2004) Likewise, several previous studies in cerebellar patients and functional imaging studies in healthy human subjects showed involvement of the cerebellum in delay eyeblink conditioning (Topka et al. 1993; Daum et al. 1993, Woodruff-Pak et al. 1996; Ramnani et al. 2000; Gerwig et al. 2003). On the other hand, animal data provide evidence that trace eyeblink conditioning may require both forebrain as well as cerebellar circuits. Numerous studies have shown that hippocampal lesions significantly reduce the M. Gerwig K. Haerter K. Hajjar A. Dimitrova M. Maschke D. Timmann (&) Departments of Neurology, University of Duisburg-Essen, Hufelandstrasse 55, 45147 Essen, Germany E-mail: [email protected] Tel.: +49-201-7233816 Fax: +49-201-7235901 E. R. Gizewski Department of Neuroradiology, University of Duisburg-Essen, Hufelandstrasse 55, 45147 Essen, Germany F. P. Kolb Department of Physiology, University of Munich, Munich, Germany A. F. Thilmann Department of Neurology, Fachklinik Rhein-Ruhr, Essen, Germany Exp Brain Res (2006) 170: 7–21 DOI 10.1007/s00221-005-0171-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
M. Gerwig Æ K. Haerter Æ K. Hajjar Æ A. Dimitrova
M. Maschke Æ F. P. Kolb Æ A. F. Thilmann
E. R. Gizewski Æ D. Timmann
Trace eyeblink conditioning in human subjects with cerebellar lesions
Received: 22 March 2005 / Accepted: 28 July 2005 / Published online: 18 November 2005� Springer-Verlag 2005
Abstract Trace eyeblink conditioning was investigated in31 patients with focal cerebellar lesions and 19 age-matched controls. Twelve patients presented with lesionsincluding the territory of the superior cerebellar artery(SCA). In 19 patients lesions were restricted to the ter-ritory of the posterior inferior cerebellar artery (PICA).A 3D magnetic resonance imaging was used to deter-mine the extent of the cortical lesion and possibleinvolvement of cerebellar nuclei. Eyeblink conditioningwas performed using a 40 ms tone as conditioned stim-ulus (CS) followed by a stimulus free trace-interval of400 ms and a 100 ms air-puff as unconditioned stimulus(US). In SCA patients with lesions including parts of thecerebellar interposed nucleus trace eyeblink conditioningwas significantly impaired. Pure cortical lesions of thesuperior cerebellum were not sufficient to reduce acqui-sition of trace conditioned eyeblink responses. PICApatients were not impaired in trace eyeblink condition-ing. Consistent with animal studies the findings of thepresent human lesion study suggest that, in addition toforebrain areas, the interposed nucleus is of importancein trace eyeblink conditioning. Although cortical cere-bellar areas appear less important in trace comparedwith delay eyeblink conditioning, the present data
strengthen the view that cerebellar structures contributeto different forms of eyeblink conditioning paradigms.
Keywords Classical conditioning Æ Cerebellum ÆInterposed nucleus Æ Associative learning
Introduction
In the last decades classical conditioning of the eyeblinkreflex has been used as a simple and successful model inresearch of associative motor learning and memory inanimals and humans (Thompson et al. 1997, 2000; Yeoand Hesslow 1998). Most commonly, delay eyeblinkconditioning has been investigated, in which the condi-tioned stimulus (CS) overlaps and coterminates with theunconditioned stimulus (US) (Green and Woodruff-Pak2000). Trace eyeblink conditioning, a more complexform of associative learning, is characterized by a stim-ulus-free trace interval that separates the CS from theUS requiring a ‘‘memory-trace’’ to associate the CS withthe later US. It has been suggested that the delay para-digm may represent a basic form of procedural (implicit)learning and that trace eyeblink conditioning may serveas a model of declarative (explicit) memory (for review:Christian and Thompson 2003).
CR-acquisition in delay eyeblink conditioning hasbeen shown essentially dependent on neural circuitrywithin the cerebellum (for review: Thompson et al. 1997;Yeo and Hesslow 1998; Steinmetz 2000), but is not sig-nificantly reduced following hippocampal lesions (Weisset al. 1999; Tseng et al. 2004) Likewise, several previousstudies in cerebellar patients and functional imagingstudies in healthy human subjects showed involvementof the cerebellum in delay eyeblink conditioning (Topkaet al. 1993; Daum et al. 1993, Woodruff-Pak et al. 1996;Ramnani et al. 2000; Gerwig et al. 2003).
On the other hand, animal data provide evidence thattrace eyeblink conditioning may require both forebrainas well as cerebellar circuits. Numerous studies haveshown that hippocampal lesions significantly reduce the
M. Gerwig Æ K. Haerter Æ K. Hajjar Æ A. DimitrovaM. Maschke Æ D. Timmann (&)Departments of Neurology, University of Duisburg-Essen,Hufelandstrasse 55, 45147 Essen, GermanyE-mail: [email protected].: +49-201-7233816Fax: +49-201-7235901
E. R. GizewskiDepartment of Neuroradiology, University of Duisburg-Essen,Hufelandstrasse 55, 45147 Essen, Germany
F. P. KolbDepartment of Physiology, University of Munich,Munich, Germany
A. F. ThilmannDepartment of Neurology, Fachklinik Rhein-Ruhr,Essen, Germany
Exp Brain Res (2006) 170: 7–21DOI 10.1007/s00221-005-0171-2

acquisition of conditioned responses (CR) in trace eye-blink conditioning (Solomon et al. 1986; Daum et al.1989; Moyer et al. 1990). Animal lesion and recordingstudies suggest an additional role of the cerebellum intrace eyeblink conditioning (Woodruff-Pak et al. 1985;Gruart et al. 2000). Moreover, some studies in hippo-campectomized rabbits and decerebrate guinea pigsshowed that cerebellar structures may be sufficient tomaintain CR-acquisition in a trace eyeblink condition-ing task (Port et al. 1986; James et al. 1987; Kotani et al.2003).
Using longer trace intervals than in animals, traceeyeblink conditioning has been found reduced followinghippocampal lesions in humans (Woodruff-Pak 1993;McGlinchey-Berroth et al. 1997; Clark and Squire 1998).So far, trace eyeblink conditioning has not been inves-tigated in a group of patients with defined cerebellarlesions. One case report only examined trace eyeblinkconditioning in a patient with cerebellar cortical degen-eration (Fortier et al. 2000). In that patient CR-acqui-sition was impaired in a temporal discriminationconditioning task, but preserved in trace as well as delayeyeblink conditioning. The authors concluded that cer-ebellar nuclei may be important in delay as well as traceeyeblink conditioning.
In the present study trace eyeblink conditioning wasinvestigated in a larger group of patients with focalcerebellar lesions. Because findings of a previous delayeyeblink conditioning study had shown that CR-inci-dences were significantly reduced in patients with corti-cal lesions including lobules HVI and/or Crus I and withlesions of the interposed nucleus (Gerwig et al. 2003),patients with lesions including the territory of thesuperior cerebellar artery (SCA) and with lesions withinthe territory of the posterior inferior cerebellar artery(PICA) were compared. In humans lobules HVI andCrus I and the cerebellar nuclei are commonly suppliedby the SCA (Amarenco et al. 1993). The aim of the studywas to examine whether trace eyeblink conditioning wasimpaired in patients with lesions of superior parts of thecerebellar hemisphere including cortical areas and/orcerebellar nuclei and possibly unaffected in patients withlesions restricted to the posterior inferior cerebellum.
Methods
Subjects
A total of 31 patients with focal cerebellar lesions (24males, 7 females; mean age 52.8±11.4 years, age range32–69 years) and 19 age- and sex-matched healthy con-trols (13 males, 6 females; mean age 51.7±12.1 years,age range 30–69 years) were included. Nineteen patientspresented with infarcts restricted to the territory of thePICA (mean age 51.1±11.2 years, age range 34–69 years). In all of them lesions were unilateral. FifteenPICA patients were male, four were female. Mean timesince lesion in PICA patients was 2.0 years (range
between 0.5 months and 6 years). In 12 patients lesionsincluded the territory of the SCA (mean age55.6±11.7 years, age range 32–69 years), lesions wereunilateral in nine of them. Eight SCA patients weremale, four were female. Nine SCA patients suffered fromischemic infarcts, one patient from surgical lesion fol-lowing astrocytoma, one from cerebellitis. One 62-year-old female patient with cerebellar agenesis was includedin the SCA group. Mean time since lesion in theremaining SCA patients was 3.7 years (range between0.5 months and 20 years). Six SCA patients, seven PICApatients, and seven of the controls had participated in aprevious delay eyeblink conditioning study (Gerwiget al. 2003).
The neurological examination according to the ataxiarating scale from Trouillas et al. (1997) revealed mildsigns of ataxia (total ataxia score <10 out of 100) in allof the PICA patients. Seven of the SCA patients showedmild signs of ataxia, two moderate signs (total ataxiascore 10–20), and three SCA patients presented withmore severe signs (total ataxia score >20 out of 100) ofcerebellar ataxia. None of the patients showed brainstemor other extracerebellar neurological signs based uponneurological examination except one SCA patient (cer-28) with intermittent hemiparaesthesia on the right side.This patient was included, because MRI scans showedischemia of the cerebellum, but not the brainstem. Noneof the control subjects presented with a history of neu-rological diseases revealed neurological signs. All cere-bellar patients and control subjects were free from anydrugs possibly influencing nervous system functions.None of the cerebellar patients or control subjectsshowed clinical signs of impaired visual or hearingcapacities on routine neurological examination. In thebeginning of the experiment hearing threshold was as-sessed in each subject. The clinical and lesion data of thepatients are summarized in Table 1. The study was ap-proved by the local ethics committee of the University ofDuisburg-Essen and informed consent was obtainedfrom all subjects.
MR imaging
Methodological details of brain imaging procedureshave been reported previously (Gerwig et al. 2003) andare repeated in brief. Individual three-dimensional (3D)MRI data sets were acquired using a T1-weightedMPRAGE sequence on a Siemens Sonata 1.5 Tesla MRscanner.
Cerebellar lesions were manually traced and saved asregions of interest (ROI) using MRIcro software (Ror-den and Brett 2000). Data sets were spatially normalizedto standard stereotaxic brain space (MNI 152-space)using SPM99 software (Wellcome Department of Cog-nitive Neurology, London) according to the MontrealNeurologic Institute (MNI) protocol and presentedwithin a Talairach grid (Evans et al. 1994). MNI coor-dinates of individual cerebellar lesions were determined
8

in horizontal (x), sagittal (y), and vertical (z) directionsand defined within a vermal (x £ ±10 mm), a paraver-mal (x=11 to 24 mm), and a lateral hemispheral section(x>24 mm). Volumes of cerebellar lesions have beencalculated based on non-normalized ROIs using MRI-cro. The Schmahmann et al. (2000) 3D MRI atlas of thehuman cerebellum was used to identify affected lobulesand the 3D MRI atlas of cerebellar nuclei introduced byDimitrova et al. (2002) to identify affected cerebellarnuclei.
In addition lesions were superimposed on axial MRslices of a healthy subject acquired by a fast low-angleshot (FLASH) sequence which was used to visualizecerebellar nuclei. Data of the patient with cerebellaragenesis were excluded because of problems with nor-malization due to changes in normal anatomy.Throughout Results and Discussion the term affectedcerebellar nuclei is used to indicate damage to the
interposed nucleus. A 3D MRI data sets were availablein all patients. Additional axial and sagittal 2D T2-weighted cranial MR images were acquired and in-spected for possible brainstem involvement or otherextracerebellar lesions. MRI data was acquired at thetime of the experiment, except in the patient with cere-bellar agenesis, three of the other SCA patients and fivePICA patients who had participated in the previousdelay conditioning study. In those patients initial MRIwas performed more than 2 months after the lesion.
Trace eyeblink conditioning
In order to compare results of the present trace para-digm to our previous delay eyeblink conditioning study(Gerwig et al. 2003) a similar short CS–US interstimulusinterval (440 ms) was used. The CS was a 540 ms, 1 kHz
Table 1 Clinical characteristics of cerebellar subjects
Cerebellar disorder Ataxia rating scale
Age(year)
Sex Diseaseduration(year)
Posture(0–34)*
Upper limbs(0–36)
Lower limbs(0–16)
Speech(0–8)
Oculo-motor(0–6)
Total(0–100)
PICA groupcer-01 d PICA l 41 F 2.5 0 0 0 0 0 0cer-02 d PICA r 58 M 2 1 0 0 0 0 1cer-03 PICA r 39 F 1 2 1 1 0 0 4cer-04 PICA l 53 F 2 2 1 0 0 0 3cer-05 d PICA r 65 M 2.5 0 0 0 0 0 0cer-06 PICA r 62 M 3 2 0 0 0 0 2cer-07 d PICA l 52 M 2 1 0 0 0 0 1cer-08 d PICA l 63 M 4 1 2 0 0 0 3cer-09 d PICA l 52 M 2.5 0 0 0 0 0 0cer-10 PICA r 67 M 2.5 2 1 0 0 0 3cer-11 d PICA l 38 M 2 0 0 0 0 0 0cer-12 PICA r 47 M 2.5 1 0 0 0 0 1cer-13 PICA r 40 M 0.5 months 4 2 0 0 0 6cer-14 PICA l 49 M 2.5 0 0 0 0 0 0cer-15 PICA l 34 M 0.5 months 1 0 0 0 0 1cer-16 PICA l 44 M 3 months 2 0 0 0 0 2cer-17 PICA r 69 M 6 1 1 0 0 0 2cer-18 PICA r 60 M 3 months 4 3 0 0 1 8cer-19 PICA r 37 M 3 months 4 1 0 0 0 5SCA groupcer-20 d SCA l 52 F 2 0 1 0 0 0 1cer-21 d SCA l 55 F 5 0 0 0 0 0 0cer-22 SCA r 68 M 2 7 4 1 0 0 12cer-23 SCA l 32 M 4 months 1 4 0 0 0 5cer-24 Astrocyt.r 65 F 20 0 0 0 0 0 0cer-25 d SCA l 47 M 2.5 0 0 0 0 0 0cer-26 d SCA l 69 M 5 2 0 0 0 0 2cer-27 d SCA r 58 M 1.5 3 3 2 0 0 8cer-28 SCA l 50 M 1 10 6 2 0 1 19cer-29 SCA b 68 M 1 4 3 0 0 0 7cer-30 PML b 41 M 0.5 months 4 5 2 2 0 13cer-31 d Cer. Agenesis 62 F 62 7 7 4 3 1 22
Ataxia score based on Cooperative Ataxia Rating Scale (Trouillas et al. 1997); *range of ataxia score with zero indicating normalperformanceDuration of disease = time since lesion, M male, F female, cer-No. cerebellar patient, d participation in previous delay conditioning study(Gerwig et al. 2003), r right, l left, b bilateral, PICA infarct of posterior inferior cerebellar artery, SCA infarct/lesion within the territory ofthe superior cerebellar artery, PML progressive multifocal leukencephalopathy (pure cerebellar lesion at time of testing), Astrocyt.Astrocytoma
9

tone in the earlier delay study and consisted of a tonewith a duration of 40 ms and a frequency of 2 kHz inthe present trace study. This was followed by a stimulus-free trace interval of 400 ms. An air-puff (duration100 ms; intensity 400 KPa at source, 110 KPa at nozzle)served as the unconditioned stimulus (US) and was di-rected through a nozzle near the outer canthus of the eyeat a distance of about 10 mm. The tone was presentedunilaterally and ipsilaterally to the US. At the beginningof the experiment ten CS-alone trials and ten US-alonetrials were presented in an unpaired and random order,followed by 100 paired CS–US trials. Ten CS-alonetrials were presented at the end of the experiment asextinction trials. Throughout the experiment the inter-trial interval varied randomly between 20 and 35 s.Surface EMG recordings were taken from orbicularisoculi muscles bilaterally with electrodes fixed to thelower eyelid and to the nasion. Signals were fed to EMGamplifiers (sampling rate 1,000 Hz, band pass filter fre-quency between 100 Hz and 2 KHz), full wave rectifiedand further filtered offline (100 Hz). The right and theleft side were tested on two separate days which were7 days apart, that is CS and US were first applied to oneside and then changed to the contralateral side in thesecond session. The sequence of affected and unaffectedside was alternated between individual patients.
The frequency of spontaneous blinks was measuredby recording eyeblink responses within 1 min both at thebeginning and the end of the experiment.
EMG recordings were analyzed on a trial basis byusing a commercial software (Axograph 4.9, AxonInstruments Inc., USA).
Analysis of unconditioned responses in unpaired trials
Unconditioned eyeblink responses (UR) in unpairedtrials were analyzed on the side of the stimulus andcontralateral of the stimulus. The air-puff evoked URrepresents the late bilateral response (R2 component) ofthe blink reflex, which is associated with clinical visibleblinking (Ongerboer de Visser 1983a, b). The air-puffcommonly evokes the R2, but not the R1 component,which possibly requires higher stimulation intensities(Peshori et al. 2001).
Timing parameters were assessed. Onset, time topeak, and duration were compared using analysis ofvariance with repeated measures with stimulation side(stimulated versus not stimulated) and side (affectedversus unaffected in patients with unilateral lesions; af-fected matched versus unaffected matched in controls) aswithin subject factor and group (controls versus SCA,PICA) as between subject factor.
Analysis of conditioned and extinction responses
Conditioned eyeblink responses in paired and extinctiontrials within the CS–US interval were visually identified
where EMG activity significantly deviated from the pre-CS EMG baseline level. EMG activity lasting at least50 ms or (in paired trials) merging into superimposedunconditioned response of at least twice the amplitudeof mean EMG baseline activity and clear rising slopewas defined as CR. The time window for detection ofCRs was from 150 ms after CS-onset to the onset of theUS. Responses occurring within the 150 ms intervalafter CS-onset were considered as reflexive responses tothe tone (i.e. alpha-responses) and not conditioned re-sponses (Woodruff-Pak et al. 1996). Trials with alpha-responses and spontaneous blinks occurring prior to CS-onset were excluded from the analysis (Bracha et al.2000).
The number of conditioned responses in paired trialswas expressed as the percentage of trials containing re-sponses with respect to each block of ten trials (per-centage CR-incidence) and the total number of trials(total percentage CR-incidence). In extinction trials theabsolute number of CRs are presented.
Depending on the order of the testing (affected orunaffected side first) affected side in patients with uni-lateral lesions was matched to the first or second testedside in controls. These will be referred to the affectedmatched side and unaffected matched side in controls.
In the majority of patients lesions were unilateral andthe contralateral side was defined as unaffected side. Inthree of the SCA patients lesions were bilateral (cer-29,cer-30, and cer-31; see Table 2 below). In these patientsboth sides were affected. The side, which was more af-fected based on MRI (cer-29) or the side of the domi-nant hand (cer-30 and cer-31) were set as ‘‘affected’’ andentered into group analysis of the affected side. Com-paring the affected and unaffected sides results of pa-tients with unilateral lesions only were considered.Results of the second affected side in the three patientswith bilateral lesions are not shown.
In statistical analysis CR responses were included onthe side of the stimulus. CRs were expressed bilaterallyin the majority of cases [mean ± SD; control group:92±9.11% (matched affected)/89.7±13.3% (matchedunaffected); PICA group 88.5 SD 14.3% (affected)/90.9±13.1% (unaffected); SCA group 79.0±24.8%(affected)/90.6±9.2% (unaffected)]. Percentage ofbilateral CRs did not differ significantly between con-trols and patients (all p values >0.1; oneway ANOVA),except on the affected side in the SCA group comparedto controls (p=0.045). In the SCA group, however,unilateral CRs were more frequent on the side of thestimulus [affected side: 12.5±20.5% (stimulated)/8.4±10.9% (not stimulated); unaffected: 7.95±7.2%(stimulated)/1.44±2.7% (not stimulated)].
In paired trials, analysis of variance (ANOVA withrepeated measures) was calculated with percentage CR-incidence as dependent variable, block (1–10) as withinsubject factor, and group (control versus cerebellar,SCA patients and PICA patients) as between subjectfactor. In patients with unilateral lesions side effectswere analyzed also (affected versus unaffected side in
10

cerebellar patients; affected matched versus unaffectedmatched side in controls). For post hoc testing, ANOVAwas calculated separately within each group.
In extinction trials the absolute number of CRs wascompared between groups using unpaired t-tests.
Level of significance was set at p<0.05. For all ef-fects, the degrees of freedom were adjusted, if appro-priate, according to Greenhouse and Geisser.
Awareness
To assess awareness during conditioning the subjectsanswered a seven item questionnaire after the condi-tioning session. First, the participants were asked toidentify and describe the stimuli used in this experiment(tone, air-puff) and second, whether there was a possiblecontingency of both stimuli. If yes, they should decide if
Table 2 Affected cerebellar lobules and deep nuclei in cerebellar patients as revealed by stereotaxically normalized 3D MRI
Subject Vermis Hemisphere Nuclei Volume
Paravermal(x=10–24 mm)
Lateral hemispheral(x>24 mm)
PICA groupcer-01 r: VIIAt, VIIB l: Crus II, VIIB, VIIIA l: Crus II, VIIB, VIIIA n.a. 7.6 cc
l: VIIAt, VIIBcer-02 r: VIIB, VIIIA/B r: Crus II, VIIB, VIIIA/B r: Crus II, VIIB n.a. 19.9 cccer-03 r: IX, VIIIA/B, VIIB, VIIAt r: Crus II, VIIB, VIIIA/B n.a. 4.5 cccer-04 l: Crus I/II n.a. 0.9 cccer-05 r: VIIB, VIIIA/B, IX r: VIIB, VIIIA/B, IX r: VIIIA n.a. 2.9 cccer-06 r: Crus II, VIIB, VIIIA/B, IX r: Crus II, VIIB, VIIIA/B n.a. 9.2 cccer-07 l: VIIAt , VIIB, VIIIA/B, IX l: Crus I/II, VIIB, VIIIA/B l: Crus I/II, VIIB, VIIIA n.a. 11.5 cccer-08 r: VIIAt, VIIB, VIIIA/B, IX l:Crus II, VIIB, VIIIA/B, IX l: VIIB, VIIIA/B l: dentate 20.5 cc
l: VIIAt, VIIAf, VIIB, VIIIA/B, IXcer-09 l: VIIB, VIIIA/B, IX l: Crus I/II, VIIB, VIIIA/B, IX l: Crus I/II, VIIB, VIIIA/B l: dentate 32.0 cccer-10 l: IX r: Crus I/II, VIIB, VIIIA/B, IX r: Crus I/II, VIIB, VIIIA/B r: dentate 32.7 cc
r: VIIAt, VIIAf, VIIB, VIIIA/B, IXcer-11 l: VIIAt, VIIB, VIIIA/B, IX l: Crus I/II, VIIB, VIIIA/B l: Crus I/II, VIIB, VIIIA/B l: dentate 13.3 cccer-12 l: VIIIA/B, IX r: Crus II, VIIB, VIIIA/B r: dentate 7.0 cc
r: VIIAt, VIIB, VIIIA/B,IXcer-13 l: VIIB, VIIIA/B, IX r: Crus I/II, VIIB, VIIIA/B, IX r: Crus I/II, VIIB,VIIIA/B r: dentate 32.2 cc
r: VIIAt, VIIAf, VIIB, VIIIA/B, IX, Xcer-14 l: VIIAt, VIIB, VIIIA/B l: Crus II, VIIB, VIIIA/B l: dentate 3.8 cccer-15 r: VIIIB, IX, X l: Crus I/II, VIIB, VIIIA/B, l: dentate 37.8 cc
l: VIIAt , VIIIA/B, IX, Xcer-16 r: VIIIB, IX l: Crus II, VIIB, VIIIA/B, IX l: Crus II, VIIB, VIIIA/B l: dentate 11.8 cc
l: VIIB, VIIIA/B, IXcer-17 l: VIIIA/B, IX r: Crus II, VIIB, VIIIA/B, IX r: Crus II, VIIB, VIIIA/B r: dentate 22.7 cc
r: VIIIA/B, IXcer-18 l: VIIAt, VIIB, VIIIA r: Crus I/II, VIIB, VIIIA/B, IX r: Crus I/II, VIIB, VIIIA r: dentate 19.8 cc
r: Crus I/II, VIIB, VIIIA/Bcer-19 r: VIIB, VIIIA/B r: Crus II, VIIB, VIIIA/ B r: Crus II, VIIIA r: dentate 9.8 ccSCA groupcer-20 l: IV, V, VI, Crus I l: VI, Crus I n.a. 10.7 cccer-21 l: VI, V l: VI n.a. 3.1 cccer-22 r: V, VI r: V, VI n.a. 4.4 cccer-23 l: IV, V, VI l: V, VI n.a. 2.1 cccer-24 r: VII At, VIIB, VIIIA/B r: VI, Crus I/II, VIIB r: VI, Crus I/II r: dentate 12.8 cccer-25 l: III l: IV, V, VI l: VI l: interposed 1.4 cccer-26 l: III, IV l: III, IV, V, VI l: V, VI l: interposed 6.4 cccer-27 r: IV, V, VI r: III, IV, V, VI, Crus I r: IV, V, VI, Crus I r: interposed 10.1 cc
r: dentatecer-28 l: IV, V l: V, VI l: VI l: interposed 3.1 cc
l: dentatecer-29 r: V, VI r: VI, Crus I r: interposed 8.4 cc
l: Crus I r: dentatecer-30 r: IV,V, VI; l: IV r: IV, V, VI r: VI, Crus I b: interposed 5.8 cc
l: V, VI l: VI r: dentatecer-31 Cerebellar agenesis
All patients presented with ischemic infarction except cer-24 (postsurgical lesion), cer-30 (cerebellitis) and cer-31 (cerebellar agenesis).Cerebellar lobules are named according to Schmahmann et al. (2000). Individual cerebellar lesions are determined within vermal, paravermaland lateral hemispheral sectionsPICA infarction of posterior inferior cerebellar artery, SCA lesion within the territory of the superior cerebellar artery, r right, l left, bbilateral, n.a. not affected, interposed interposed nucleus, dentate dentate nucleus, volume volume of the lesion in cc cubic centimetres (seeMethods for details)
11
there was a specified order of the stimuli within themain, i.e. the middle part of the study (presentation ofpaired stimuli). Finally, subjects were asked if the fol-lowing statements regarding order of the stimuli weretrue or false: (1) tone and air-puff were usually simul-taneously in time, (2) tone came usually before the air-puff, (3) tone came usually after the air-puff and (4) theorder of tone and air-puff changed.
It was assessed if patients and controls recognizedstimuli (item 1). Subjects who gave wrong answers initems 2–7 were defined as ‘‘unaware’’. Comparison ofmean CR-incidences in subjects with and withoutawareness was compared using unpaired t-tests in thecontrol, PICA, and SCA groups.
Results
MR findings
Table 2 summarizes the extent of individual lesions, thathas affected cerebellar lobules within the vermal, par-avermal, and lateral hemispheral sections and whetherthe deep cerebellar nuclei were affected or not.
In nine out of 12 SCA patients the lesion was re-stricted to the SCA-territory involving Crus I andabove and extended into the common PICA-territory,that is Crus II and below in three of the SCA patients.In seven SCA patients including the patient with cere-bellar agenesis (cer-25, cer-26, cer-27, cer-28, cer-29,cer-30, and cer-31) lesions affected at least part of the
interposed nucleus. In five SCA patients the interposednucleus was not affected. In five SCA patients (cer-25,cer-26, cer-27, cer-28, and cer-29) the interposednucleus was affected unilaterally, in addition thedentate nucleus was involved in three SCA patients(cer-27, cer-28, and cer-29).
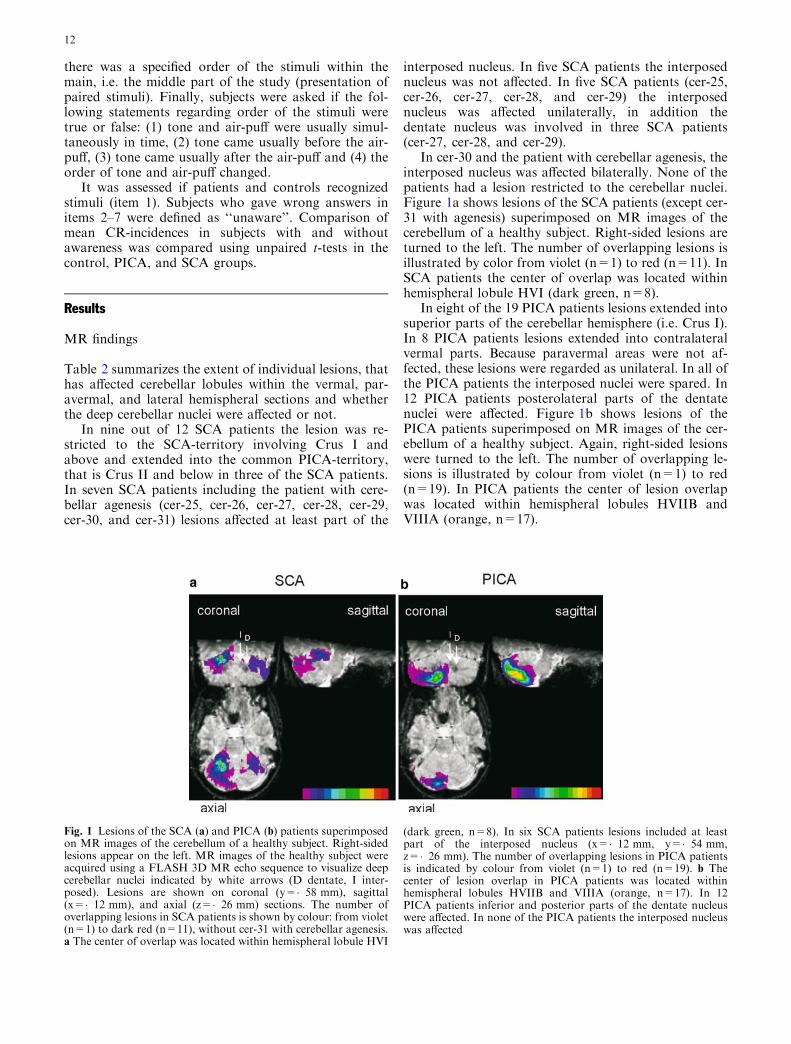
In cer-30 and the patient with cerebellar agenesis, theinterposed nucleus was affected bilaterally. None of thepatients had a lesion restricted to the cerebellar nuclei.Figure 1a shows lesions of the SCA patients (except cer-31 with agenesis) superimposed on MR images of thecerebellum of a healthy subject. Right-sided lesions areturned to the left. The number of overlapping lesions isillustrated by color from violet (n=1) to red (n=11). InSCA patients the center of overlap was located withinhemispheral lobule HVI (dark green, n=8).
In eight of the 19 PICA patients lesions extended intosuperior parts of the cerebellar hemisphere (i.e. Crus I).In 8 PICA patients lesions extended into contralateralvermal parts. Because paravermal areas were not af-fected, these lesions were regarded as unilateral. In all ofthe PICA patients the interposed nuclei were spared. In12 PICA patients posterolateral parts of the dentatenuclei were affected. Figure 1b shows lesions of thePICA patients superimposed on MR images of the cer-ebellum of a healthy subject. Again, right-sided lesionswere turned to the left. The number of overlapping le-sions is illustrated by colour from violet (n=1) to red(n=19). In PICA patients the center of lesion overlapwas located within hemispheral lobules HVIIB andVIIIA (orange, n=17).
Fig. 1 Lesions of the SCA (a) and PICA (b) patients superimposedon MR images of the cerebellum of a healthy subject. Right-sidedlesions appear on the left. MR images of the healthy subject wereacquired using a FLASH 3D MR echo sequence to visualize deepcerebellar nuclei indicated by white arrows (D dentate, I inter-posed). Lesions are shown on coronal (y=�58 mm), sagittal(x=�12 mm), and axial (z=�26 mm) sections. The number ofoverlapping lesions in SCA patients is shown by colour: from violet(n=1) to dark red (n=11), without cer-31 with cerebellar agenesis.a The center of overlap was located within hemispheral lobule HVI
(dark green, n=8). In six SCA patients lesions included at leastpart of the interposed nucleus (x=�12 mm, y=�54 mm,z=�26 mm). The number of overlapping lesions in PICA patientsis indicated by colour from violet (n=1) to red (n=19). b Thecenter of lesion overlap in PICA patients was located withinhemispheral lobules HVIIB and VIIIA (orange, n=17). In 12PICA patients inferior and posterior parts of the dentate nucleuswere affected. In none of the PICA patients the interposed nucleuswas affected
12
Unconditioned responses in unpaired trials
There was no significant difference comparing UR onset,peaktime, and duration in unpaired trials between con-trols, PICA, and SCA patients (group effect: all p values>0.4, ANOVA with repeated measures; Table 3), ex-cept a tendency of duration to be larger on the affectedand stimulated side in SCA patients compared to con-trols (side by stimulation by group interaction:p=0.063). Increased UR duration following lesions ofthe superior cerebellum is expected (for detaileddescription of amplitude changes see Gerwig et al. 2004).
In both patients and controls UR onset and peaktimeoccurred later on the side contralateral to the air-puffstimulus compared to the side of the stimulus. This wastrue on the affected and unaffected side in patients andon the affected matched and unaffected matched side in
controls (Table 3). Effects of stimulation side were sig-nificant (onset: p values <0.001; peaktime: PICA versuscontrols p<0.001) or reached significance (peaktime:SCA versus controls p=0.069; ANOVA with repeatedmeasures). There were no significant group by stimula-tion, group by side (affected versus unaffected), andgroup by side by stimulation interaction effects (all pvalues >0.2).
Trace conditioning
CR-incidences in all cerebellar patients and controlsubjects
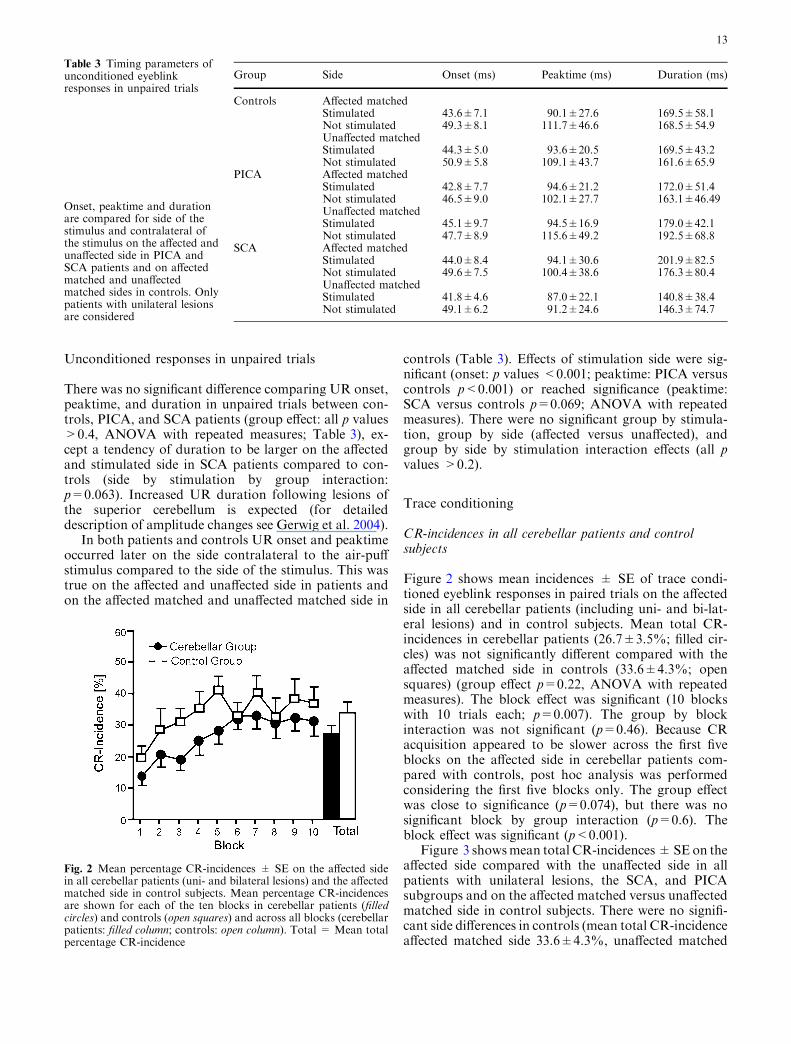
Figure 2 shows mean incidences ± SE of trace condi-tioned eyeblink responses in paired trials on the affectedside in all cerebellar patients (including uni- and bi-lat-eral lesions) and in control subjects. Mean total CR-incidences in cerebellar patients (26.7±3.5%; filled cir-cles) was not significantly different compared with theaffected matched side in controls (33.6±4.3%; opensquares) (group effect p=0.22, ANOVA with repeatedmeasures). The block effect was significant (10 blockswith 10 trials each; p=0.007). The group by blockinteraction was not significant (p=0.46). Because CRacquisition appeared to be slower across the first fiveblocks on the affected side in cerebellar patients com-pared with controls, post hoc analysis was performedconsidering the first five blocks only. The group effectwas close to significance (p=0.074), but there was nosignificant block by group interaction (p=0.6). Theblock effect was significant (p<0.001).
Figure 3 showsmean total CR-incidences±SE on theaffected side compared with the unaffected side in allpatients with unilateral lesions, the SCA, and PICAsubgroups and on the affected matched versus unaffectedmatched side in control subjects. There were no signifi-cant side differences in controls (mean total CR-incidenceaffected matched side 33.6±4.3%, unaffected matched
Table 3 Timing parameters ofunconditioned eyeblinkresponses in unpaired trials
Onset, peaktime and durationare compared for side of thestimulus and contralateral ofthe stimulus on the affected andunaffected side in PICA andSCA patients and on affectedmatched and unaffectedmatched sides in controls. Onlypatients with unilateral lesionsare considered
Group Side Onset (ms) Peaktime (ms) Duration (ms)
Controls Affected matchedStimulated 43.6±7.1 90.1±27.6 169.5±58.1Not stimulated 49.3±8.1 111.7±46.6 168.5±54.9Unaffected matchedStimulated 44.3±5.0 93.6±20.5 169.5±43.2Not stimulated 50.9±5.8 109.1±43.7 161.6±65.9
PICA Affected matchedStimulated 42.8±7.7 94.6±21.2 172.0±51.4Not stimulated 46.5±9.0 102.1±27.7 163.1±46.49Unaffected matchedStimulated 45.1±9.7 94.5±16.9 179.0±42.1Not stimulated 47.7±8.9 115.6±49.2 192.5±68.8
SCA Affected matchedStimulated 44.0±8.4 94.1±30.6 201.9±82.5Not stimulated 49.6±7.5 100.4±38.6 176.3±80.4Unaffected matchedStimulated 41.8±4.6 87.0±22.1 140.8±38.4Not stimulated 49.1±6.2 91.2±24.6 146.3±74.7
Fig. 2 Mean percentage CR-incidences ± SE on the affected sidein all cerebellar patients (uni- and bilateral lesions) and the affectedmatched side in control subjects. Mean percentage CR-incidencesare shown for each of the ten blocks in cerebellar patients (filledcircles) and controls (open squares) and across all blocks (cerebellarpatients: filled column; controls: open column). Total = Mean totalpercentage CR-incidence
13
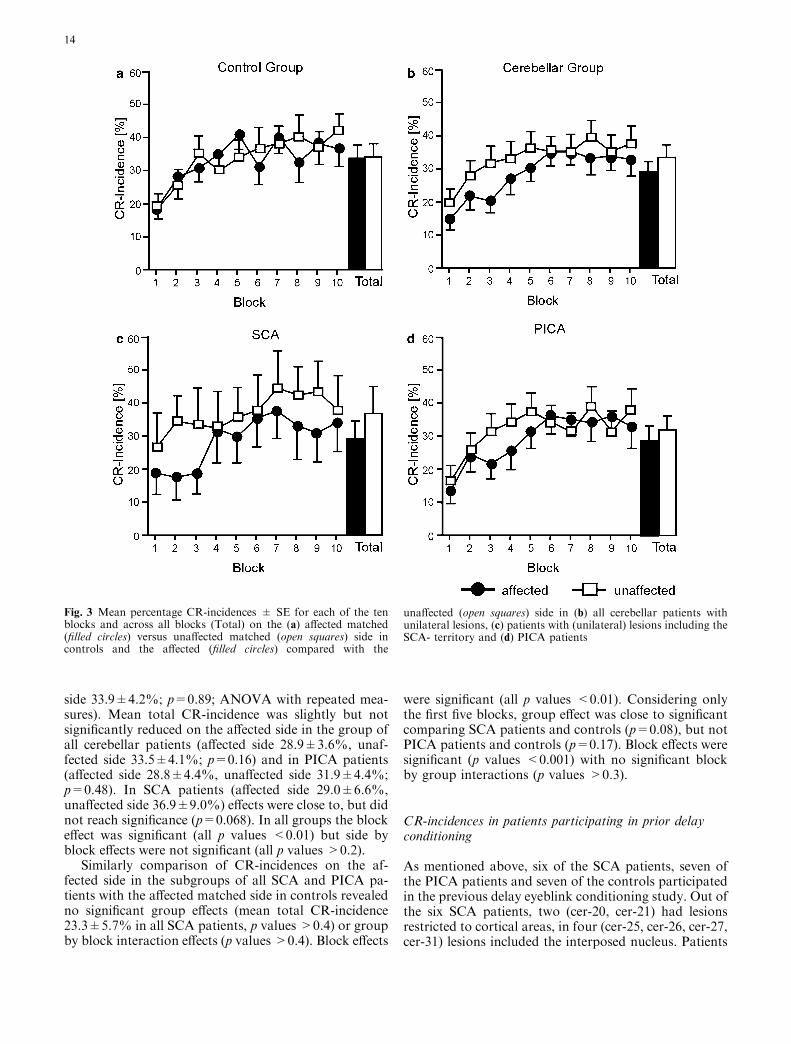
side 33.9±4.2%; p=0.89; ANOVA with repeated mea-sures). Mean total CR-incidence was slightly but notsignificantly reduced on the affected side in the group ofall cerebellar patients (affected side 28.9±3.6%, unaf-fected side 33.5±4.1%; p=0.16) and in PICA patients(affected side 28.8±4.4%, unaffected side 31.9±4.4%;p=0.48). In SCA patients (affected side 29.0±6.6%,unaffected side 36.9±9.0%) effects were close to, but didnot reach significance (p=0.068). In all groups the blockeffect was significant (all p values <0.01) but side byblock effects were not significant (all p values >0.2).
Similarly comparison of CR-incidences on the af-fected side in the subgroups of all SCA and PICA pa-tients with the affected matched side in controls revealedno significant group effects (mean total CR-incidence23.3±5.7% in all SCA patients, p values >0.4) or groupby block interaction effects (p values >0.4). Block effects
were significant (all p values <0.01). Considering onlythe first five blocks, group effect was close to significantcomparing SCA patients and controls (p=0.08), but notPICA patients and controls (p=0.17). Block effects weresignificant (p values <0.001) with no significant blockby group interactions (p values >0.3).
CR-incidences in patients participating in prior delayconditioning
As mentioned above, six of the SCA patients, seven ofthe PICA patients and seven of the controls participatedin the previous delay eyeblink conditioning study. Out ofthe six SCA patients, two (cer-20, cer-21) had lesionsrestricted to cortical areas, in four (cer-25, cer-26, cer-27,cer-31) lesions included the interposed nucleus. Patients
Fig. 3 Mean percentage CR-incidences ± SE for each of the tenblocks and across all blocks (Total) on the (a) affected matched(filled circles) versus unaffected matched (open squares) side incontrols and the affected (filled circles) compared with the
unaffected (open squares) side in (b) all cerebellar patients withunilateral lesions, (c) patients with (unilateral) lesions including theSCA- territory and (d) PICA patients
14
who underwent previous delay conditioning are indi-cated by ‘‘d’’ in Table 1.
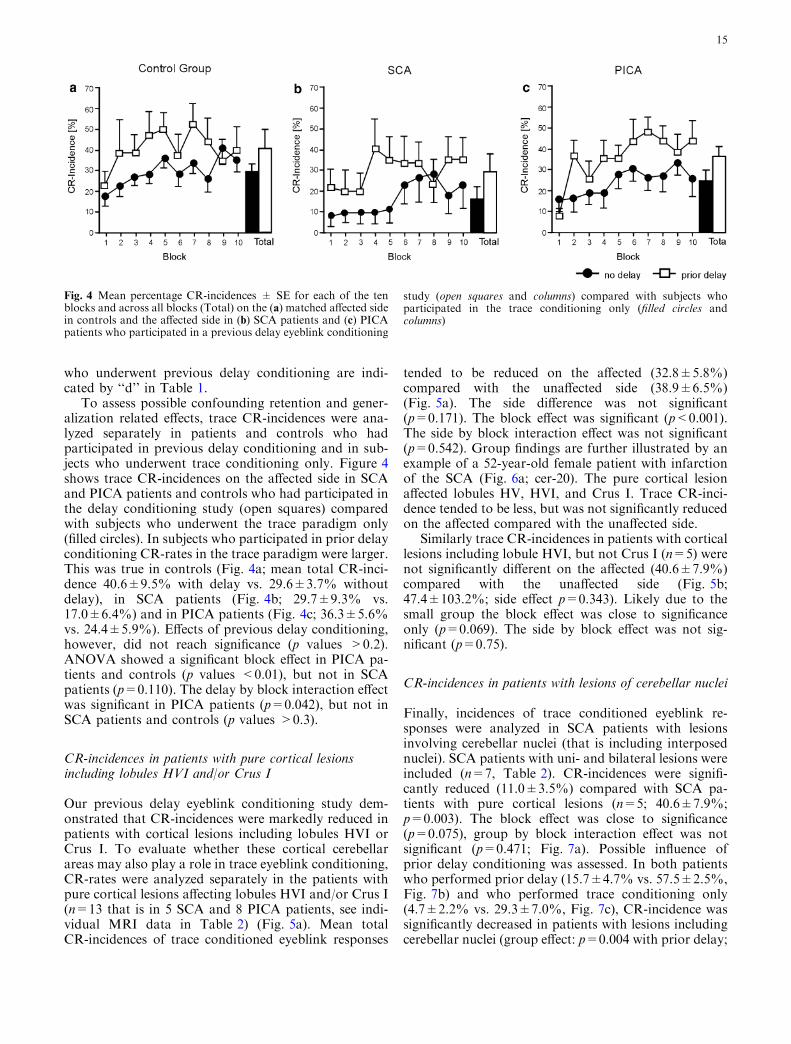
To assess possible confounding retention and gener-alization related effects, trace CR-incidences were ana-lyzed separately in patients and controls who hadparticipated in previous delay conditioning and in sub-jects who underwent trace conditioning only. Figure 4shows trace CR-incidences on the affected side in SCAand PICA patients and controls who had participated inthe delay conditioning study (open squares) comparedwith subjects who underwent the trace paradigm only(filled circles). In subjects who participated in prior delayconditioning CR-rates in the trace paradigm were larger.This was true in controls (Fig. 4a; mean total CR-inci-dence 40.6±9.5% with delay vs. 29.6±3.7% withoutdelay), in SCA patients (Fig. 4b; 29.7±9.3% vs.17.0±6.4%) and in PICA patients (Fig. 4c; 36.3±5.6%vs. 24.4±5.9%). Effects of previous delay conditioning,however, did not reach significance (p values >0.2).ANOVA showed a significant block effect in PICA pa-tients and controls (p values <0.01), but not in SCApatients (p=0.110). The delay by block interaction effectwas significant in PICA patients (p=0.042), but not inSCA patients and controls (p values >0.3).
CR-incidences in patients with pure cortical lesionsincluding lobules HVI and/or Crus I
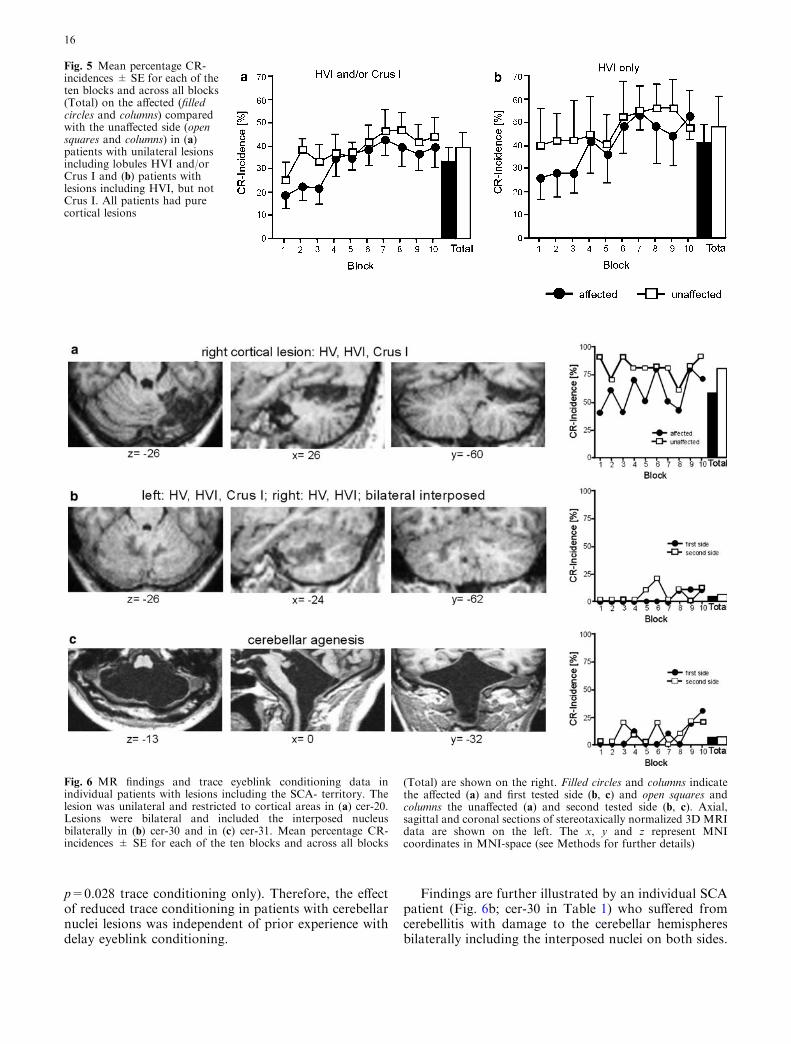
Our previous delay eyeblink conditioning study dem-onstrated that CR-incidences were markedly reduced inpatients with cortical lesions including lobules HVI orCrus I. To evaluate whether these cortical cerebellarareas may also play a role in trace eyeblink conditioning,CR-rates were analyzed separately in the patients withpure cortical lesions affecting lobules HVI and/or Crus I(n=13 that is in 5 SCA and 8 PICA patients, see indi-vidual MRI data in Table 2) (Fig. 5a). Mean totalCR-incidences of trace conditioned eyeblink responses
tended to be reduced on the affected (32.8±5.8%)compared with the unaffected side (38.9±6.5%)(Fig. 5a). The side difference was not significant(p=0.171). The block effect was significant (p<0.001).The side by block interaction effect was not significant(p=0.542). Group findings are further illustrated by anexample of a 52-year-old female patient with infarctionof the SCA (Fig. 6a; cer-20). The pure cortical lesionaffected lobules HV, HVI, and Crus I. Trace CR-inci-dence tended to be less, but was not significantly reducedon the affected compared with the unaffected side.
Similarly trace CR-incidences in patients with corticallesions including lobule HVI, but not Crus I (n=5) werenot significantly different on the affected (40.6±7.9%)compared with the unaffected side (Fig. 5b;47.4±103.2%; side effect p=0.343). Likely due to thesmall group the block effect was close to significanceonly (p=0.069). The side by block effect was not sig-nificant (p=0.75).
CR-incidences in patients with lesions of cerebellar nuclei
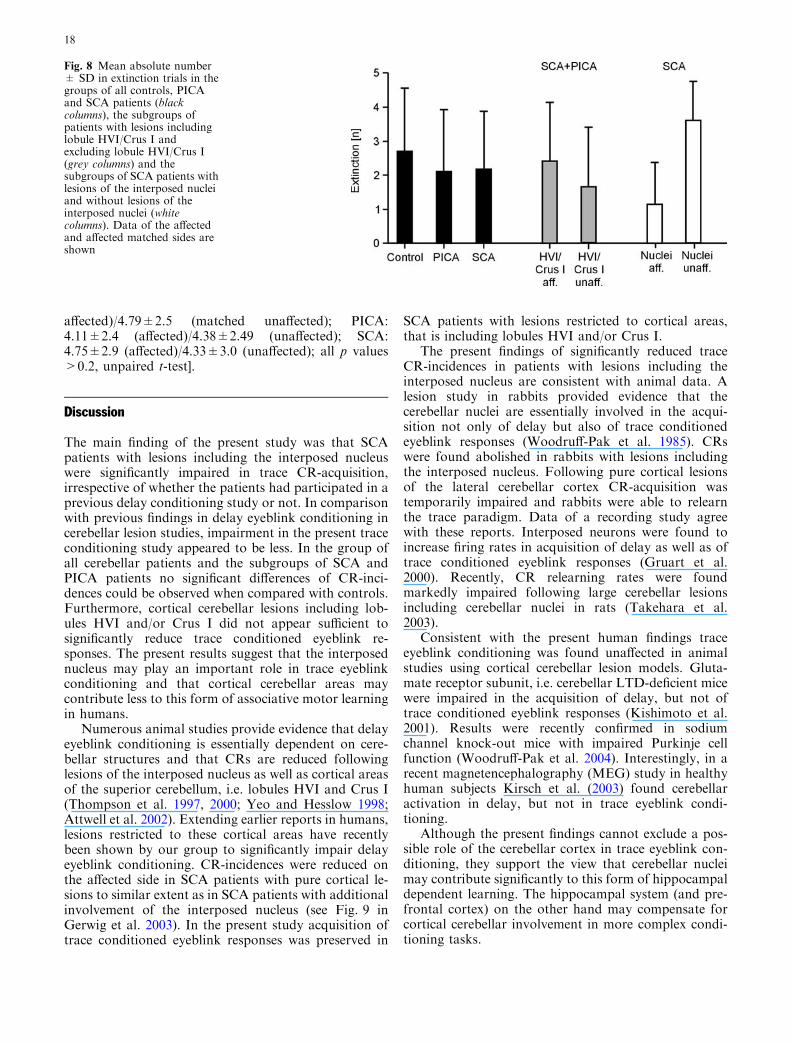
Finally, incidences of trace conditioned eyeblink re-sponses were analyzed in SCA patients with lesionsinvolving cerebellar nuclei (that is including interposednuclei). SCA patients with uni- and bilateral lesions wereincluded (n=7, Table 2). CR-incidences were signifi-cantly reduced (11.0±3.5%) compared with SCA pa-tients with pure cortical lesions (n=5; 40.6±7.9%;p=0.003). The block effect was close to significance(p=0.075), group by block interaction effect was notsignificant (p=0.471; Fig. 7a). Possible influence ofprior delay conditioning was assessed. In both patientswho performed prior delay (15.7±4.7% vs. 57.5±2.5%,Fig. 7b) and who performed trace conditioning only(4.7±2.2% vs. 29.3±7.0%, Fig. 7c), CR-incidence wassignificantly decreased in patients with lesions includingcerebellar nuclei (group effect: p=0.004 with prior delay;
Fig. 4 Mean percentage CR-incidences ± SE for each of the tenblocks and across all blocks (Total) on the (a) matched affected sidein controls and the affected side in (b) SCA patients and (c) PICApatients who participated in a previous delay eyeblink conditioning
study (open squares and columns) compared with subjects whoparticipated in the trace conditioning only (filled circles andcolumns)
15
p=0.028 trace conditioning only). Therefore, the effectof reduced trace conditioning in patients with cerebellarnuclei lesions was independent of prior experience withdelay eyeblink conditioning.
Findings are further illustrated by an individual SCApatient (Fig. 6b; cer-30 in Table 1) who suffered fromcerebellitis with damage to the cerebellar hemispheresbilaterally including the interposed nuclei on both sides.
Fig. 5 Mean percentage CR-incidences ± SE for each of theten blocks and across all blocks(Total) on the affected (filledcircles and columns) comparedwith the unaffected side (opensquares and columns) in (a)patients with unilateral lesionsincluding lobules HVI and/orCrus I and (b) patients withlesions including HVI, but notCrus I. All patients had purecortical lesions
Fig. 6 MR findings and trace eyeblink conditioning data inindividual patients with lesions including the SCA- territory. Thelesion was unilateral and restricted to cortical areas in (a) cer-20.Lesions were bilateral and included the interposed nucleusbilaterally in (b) cer-30 and in (c) cer-31. Mean percentage CR-incidences ± SE for each of the ten blocks and across all blocks
(Total) are shown on the right. Filled circles and columns indicatethe affected (a) and first tested side (b, c) and open squares andcolumns the unaffected (a) and second tested side (b, c). Axial,sagittal and coronal sections of stereotaxically normalized 3D MRIdata are shown on the left. The x, y and z represent MNIcoordinates in MNI-space (see Methods for further details)
16
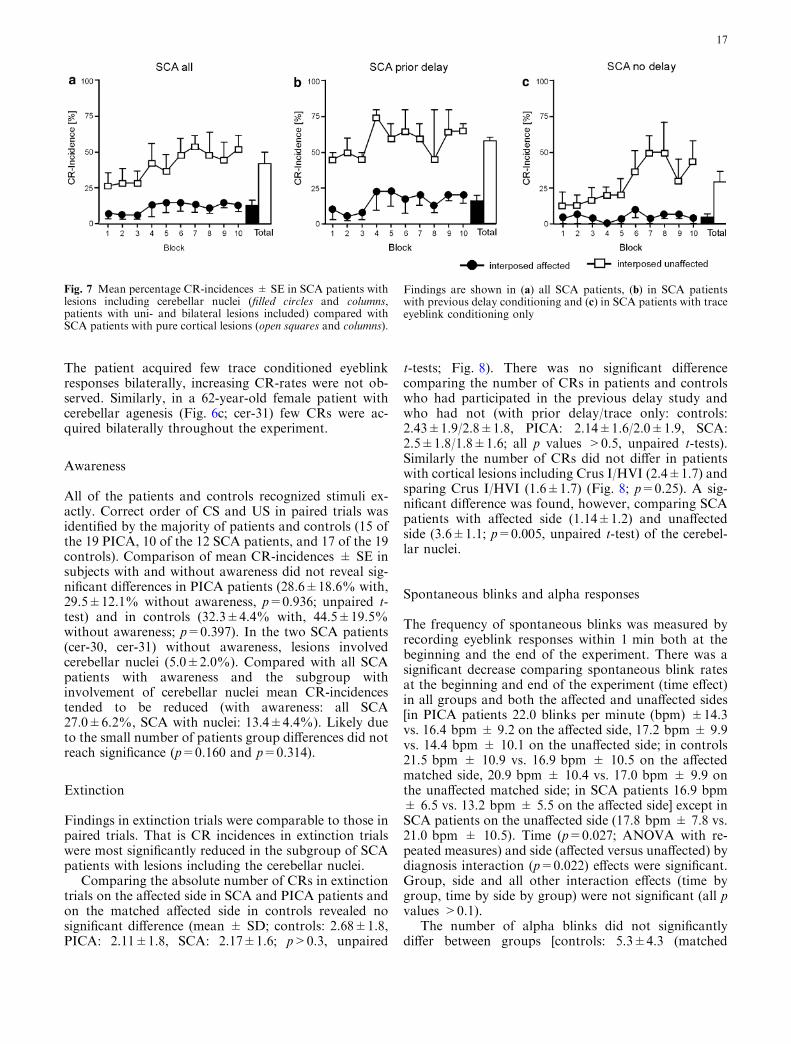
The patient acquired few trace conditioned eyeblinkresponses bilaterally, increasing CR-rates were not ob-served. Similarly, in a 62-year-old female patient withcerebellar agenesis (Fig. 6c; cer-31) few CRs were ac-quired bilaterally throughout the experiment.
Awareness
All of the patients and controls recognized stimuli ex-actly. Correct order of CS and US in paired trials wasidentified by the majority of patients and controls (15 ofthe 19 PICA, 10 of the 12 SCA patients, and 17 of the 19controls). Comparison of mean CR-incidences ± SE insubjects with and without awareness did not reveal sig-nificant differences in PICA patients (28.6±18.6% with,29.5±12.1% without awareness, p=0.936; unpaired t-test) and in controls (32.3±4.4% with, 44.5±19.5%without awareness; p=0.397). In the two SCA patients(cer-30, cer-31) without awareness, lesions involvedcerebellar nuclei (5.0±2.0%). Compared with all SCApatients with awareness and the subgroup withinvolvement of cerebellar nuclei mean CR-incidencestended to be reduced (with awareness: all SCA27.0±6.2%, SCA with nuclei: 13.4±4.4%). Likely dueto the small number of patients group differences did notreach significance (p=0.160 and p=0.314).
Extinction
Findings in extinction trials were comparable to those inpaired trials. That is CR incidences in extinction trialswere most significantly reduced in the subgroup of SCApatients with lesions including the cerebellar nuclei.
Comparing the absolute number of CRs in extinctiontrials on the affected side in SCA and PICA patients andon the matched affected side in controls revealed nosignificant difference (mean ± SD; controls: 2.68±1.8,PICA: 2.11±1.8, SCA: 2.17±1.6; p>0.3, unpaired
t-tests; Fig. 8). There was no significant differencecomparing the number of CRs in patients and controlswho had participated in the previous delay study andwho had not (with prior delay/trace only: controls:2.43±1.9/2.8±1.8, PICA: 2.14±1.6/2.0±1.9, SCA:2.5±1.8/1.8±1.6; all p values >0.5, unpaired t-tests).Similarly the number of CRs did not differ in patientswith cortical lesions including Crus I/HVI (2.4±1.7) andsparing Crus I/HVI (1.6±1.7) (Fig. 8; p=0.25). A sig-nificant difference was found, however, comparing SCApatients with affected side (1.14±1.2) and unaffectedside (3.6±1.1; p=0.005, unpaired t-test) of the cerebel-lar nuclei.
Spontaneous blinks and alpha responses
The frequency of spontaneous blinks was measured byrecording eyeblink responses within 1 min both at thebeginning and the end of the experiment. There was asignificant decrease comparing spontaneous blink ratesat the beginning and end of the experiment (time effect)in all groups and both the affected and unaffected sides[in PICA patients 22.0 blinks per minute (bpm) ±14.3vs. 16.4 bpm ± 9.2 on the affected side, 17.2 bpm ± 9.9vs. 14.4 bpm ± 10.1 on the unaffected side; in controls21.5 bpm ± 10.9 vs. 16.9 bpm ± 10.5 on the affectedmatched side, 20.9 bpm ± 10.4 vs. 17.0 bpm ± 9.9 onthe unaffected matched side; in SCA patients 16.9 bpm± 6.5 vs. 13.2 bpm ± 5.5 on the affected side] except inSCA patients on the unaffected side (17.8 bpm ± 7.8 vs.21.0 bpm ± 10.5). Time (p=0.027; ANOVA with re-peated measures) and side (affected versus unaffected) bydiagnosis interaction (p=0.022) effects were significant.Group, side and all other interaction effects (time bygroup, time by side by group) were not significant (all pvalues >0.1).
The number of alpha blinks did not significantlydiffer between groups [controls: 5.3±4.3 (matched
Fig. 7 Mean percentage CR-incidences ± SE in SCA patients withlesions including cerebellar nuclei (filled circles and columns,patients with uni- and bilateral lesions included) compared withSCA patients with pure cortical lesions (open squares and columns).
Findings are shown in (a) all SCA patients, (b) in SCA patientswith previous delay conditioning and (c) in SCA patients with traceeyeblink conditioning only
17
affected)/4.79±2.5 (matched unaffected); PICA:4.11±2.4 (affected)/4.38±2.49 (unaffected); SCA:4.75±2.9 (affected)/4.33±3.0 (unaffected); all p values>0.2, unpaired t-test].
Discussion
The main finding of the present study was that SCApatients with lesions including the interposed nucleuswere significantly impaired in trace CR-acquisition,irrespective of whether the patients had participated in aprevious delay conditioning study or not. In comparisonwith previous findings in delay eyeblink conditioning incerebellar lesion studies, impairment in the present traceconditioning study appeared to be less. In the group ofall cerebellar patients and the subgroups of SCA andPICA patients no significant differences of CR-inci-dences could be observed when compared with controls.Furthermore, cortical cerebellar lesions including lob-ules HVI and/or Crus I did not appear sufficient tosignificantly reduce trace conditioned eyeblink re-sponses. The present results suggest that the interposednucleus may play an important role in trace eyeblinkconditioning and that cortical cerebellar areas maycontribute less to this form of associative motor learningin humans.
Numerous animal studies provide evidence that delayeyeblink conditioning is essentially dependent on cere-bellar structures and that CRs are reduced followinglesions of the interposed nucleus as well as cortical areasof the superior cerebellum, i.e. lobules HVI and Crus I(Thompson et al. 1997, 2000; Yeo and Hesslow 1998;Attwell et al. 2002). Extending earlier reports in humans,lesions restricted to these cortical areas have recentlybeen shown by our group to significantly impair delayeyeblink conditioning. CR-incidences were reduced onthe affected side in SCA patients with pure cortical le-sions to similar extent as in SCA patients with additionalinvolvement of the interposed nucleus (see Fig. 9 inGerwig et al. 2003). In the present study acquisition oftrace conditioned eyeblink responses was preserved in
SCA patients with lesions restricted to cortical areas,that is including lobules HVI and/or Crus I.
The present findings of significantly reduced traceCR-incidences in patients with lesions including theinterposed nucleus are consistent with animal data. Alesion study in rabbits provided evidence that thecerebellar nuclei are essentially involved in the acqui-sition not only of delay but also of trace conditionedeyeblink responses (Woodruff-Pak et al. 1985). CRswere found abolished in rabbits with lesions includingthe interposed nucleus. Following pure cortical lesionsof the lateral cerebellar cortex CR-acquisition wastemporarily impaired and rabbits were able to relearnthe trace paradigm. Data of a recording study agreewith these reports. Interposed neurons were found toincrease firing rates in acquisition of delay as well as oftrace conditioned eyeblink responses (Gruart et al.2000). Recently, CR relearning rates were foundmarkedly impaired following large cerebellar lesionsincluding cerebellar nuclei in rats (Takehara et al.2003).
Consistent with the present human findings traceeyeblink conditioning was found unaffected in animalstudies using cortical cerebellar lesion models. Gluta-mate receptor subunit, i.e. cerebellar LTD-deficient micewere impaired in the acquisition of delay, but not oftrace conditioned eyeblink responses (Kishimoto et al.2001). Results were recently confirmed in sodiumchannel knock-out mice with impaired Purkinje cellfunction (Woodruff-Pak et al. 2004). Interestingly, in arecent magnetencephalography (MEG) study in healthyhuman subjects Kirsch et al. (2003) found cerebellaractivation in delay, but not in trace eyeblink condi-tioning.
Although the present findings cannot exclude a pos-sible role of the cerebellar cortex in trace eyeblink con-ditioning, they support the view that cerebellar nucleimay contribute significantly to this form of hippocampaldependent learning. The hippocampal system (and pre-frontal cortex) on the other hand may compensate forcortical cerebellar involvement in more complex condi-tioning tasks.
Fig. 8 Mean absolute number± SD in extinction trials in thegroups of all controls, PICAand SCA patients (blackcolumns), the subgroups ofpatients with lesions includinglobule HVI/Crus I andexcluding lobule HVI/Crus I(grey columns) and thesubgroups of SCA patients withlesions of the interposed nucleiand without lesions of theinterposed nuclei (whitecolumns). Data of the affectedand affected matched sides areshown
18

Based on extensive animal research hippocampal le-sions were found to significantly impair trace CR-acquisition in rabbits (Solomon et al. 1986; Moyer et al.1990), in rats (Weiss et al. 1999), and in mice (Tsenget al. 2004). Moreover, recording studies demonstratedthat hippocampal neuronal activity increases in traceeyeblink conditioning (McEchron and Disterhoft 1997),but hippocampal involvement appears to be temporarilylimited (Kim et al. 1995; Takehara et al. 2002). Themedial prefrontal cortex has also been shown criticallyinvolved in trace eyeblink conditioning (Kronforst-Collins and Disterhoft 1998; Weible et al. 2000).
Transneuronal tracing experiments revealed evidencethat both the cerebellar nuclei and the hippocampus areconnected to the prefrontal cortex through thalamicpathways (Yamamoto et al. 1992; Middleton and Strick1994). Furthermore, hippocampal conditioning-relatedactivity has been found disrupted following lesions ofthe interposed nucleus (Clark et al. 1984; Sears andSteinmetz 1990). The connection, however, does notseem to be made via the ventrolateral thalamus or thered nucleus (Sears et al. 1996).
It has been suggested that the cerebellum in generalmay play a role in the simple association between the CSand US and may provide a common pathway in con-ditioning paradigms of different complexity (Green andWoodruff-Pak 2000). Hippocampal structures may beinvolved in more complex conditioning paradigmsrequiring the formation of associations in addition to thesimple CS–US association. The maintainance of thestimulus trace may be provided by the hippocampus tothe cerebellum, which may generate the simple stimulusassociation (Clark et al. 2002).
It has been shown that the involvement of the hip-pocampus in trace eyeblink conditioning differs betweenspecies and depends on the length of the interstimulusinterval. Using trace intervals of 500 ms CR-acquisitionwas reduced following hippocampal lesions in rabbits(Solomon et al. 1986; Moyer et al. 1990). In amnesicpatients with hippocampal lesions, however, trace con-ditioning was mildly reduced using a 500 ms traceinterval, but markedly impaired in case of long traceintervals of 1,000 ms (McGlinchey-Berroth et al. 1997;Clark and Squire 1998). Since a short trace interval wasused in the present study, hippocampal structures maynot be required. Therefore, findings in cerebellar pa-tients have to be confirmed using a longer stimulus-freeinterval.
Lack of awareness did not explain group differencesin the present experiment. Although two of the SCApatients with lesions involving cerebellar nuclei were notaware of the CS–US order, the same was true in fourPICA patients and two controls. Eyeblink conditioningwas not significantly reduced in the PICA patients andcontrols who were not aware of the stimulus order.
It may be argued that lack of conditioning deficits onthe affected side in patients with lesions restricted to the
cerebellar cortex may have resulted from overtraining.The experience of previous delay eyeblink conditioningtended to enhance trace CR-incidences in the patientswith focal cerebellar lesions as well as in controls, al-though effects were not of significance. This may be dueto effects of retention and generalization although adifferent tone was used as conditioned stimulus. Incontrast, in patients with degenerative cerebellar disor-ders we did not find an improvement of conditionedresponse rates across three sessions of delay eyeblinkconditioning in a 1 year follow-up study (Timmannet al. 2004). Deficits in conditioning may be compen-sated in patients with focal lesions, but not in patientswith slowly progressive and more diffuse cerebellar dis-ease.
In addition, less impairment in trace compared withdelay eyeblink conditioning in SCA patients with purecortical lesions may in part have resulted from ordereffects. In the present study testing of the affected andunaffected side was alternated between patients. Inprevious delay studies (Woodruff-Pak et al. 1996; Ger-wig et al. 2003) the affected side was always investigatedfirst in all of the patients and effects of retention andgeneralization might have improved learning on theunaffected side.
There was only one patient (cer-23) with a pure cor-tical lesion who did not participate in earlier delayconditioning and who was tested on the affected sidefirst. Total CR-incidence in this patient was less on theaffected (43%) compared to the unaffected side (54%).Reduction, however, was small on the affected side andit is difficult to draw clear conclusions.
To determine a possible role of the cerebellar cortexin trace conditioning, future studies need to be per-formed in degenerative cerebellar disorders and a largergroup of conditioning-naıve patients with focal cere-bellar lesions.
In conclusion, the present results provide evidencethat cerebellar structures contribute to trace eyeblinkconditioning in humans. Extending previous findings indelay conditioning, data show that the human cerebel-lum takes part in different forms of associative motorlearning. The present findings support a role of theinterposed nucleus in trace conditioning. An additionalrole of the cerebellar cortex, however, cannot be ex-cluded.
Acknowledgments The authors like to thank Beate Brol for help ineyeblink data analysis and preparation of the figures. The studywas supported by a grant of the Deutsche Forschungsgemeinschaft(Ti 239/7-1).
References
Amarenco P, Rosengart A, DeWitt LD, Pessin MS, Caplan LR(1993) Anterior inferior cerebellar artery territory infarcts.Mechanisms and clinical features. Arch Neurol 59:154–161
19

Attwell PJ, Ivarsson M, Millar L, Yeo CH (2002) Cerebellarmechanisms in eyeblink conditioning. Ann NY Acad Sci978:79–92
Bracha V, Zhao L, Irwin KB, Bloedel JR (2000) The human cer-ebellum and associative learning: dissociation between theacquisition, retention and extinction of conditioned eyeblinks.Brain Res 860:87–94
Christian KM, Thompson RF (2003) Neural substrates of eyeblinkconditioning: acquistion and retention. Learn Mem 11:427–455
Clark GA, McCormick DA, Lavond DG, Thompson RF (1984)Effects of lesions of cerebellar nuclei on conditioned behavioraland hippocampal neuronal responses. Brain Res 291:125–136
Clark RE, Squire LR (1998) Classical conditioning and brain sys-tems: the role of awareness. Science 280:77–81
Clark RE, Manns JR, Squire LR (2002) Classical conditioning,awareness, and brain systems. Trends Cogn Sci 6:524–531
Daum I, Channon S, Canavan AG (1989) Classical conditioning inpatients with severe memory problems. J Neurol NeurosurgPsychiatry 52:47–51
Daum I, Schugens MM, Ackermann H, Lutzenberger W, DichgansJ, Birbaumer N (1993) Classical conditioning after cerebellarlesions in humans. Behav Neurosci 107:748–756
Dimitrova A, Weber J, Redies C, Kindsvater K, Maschke M, KolbFP, Forsting M, Diener HC, Timmann D (2002) MRI atlas ofthe human cerebellar nuclei. Neuroimage 17:240–255
Evans AC, Kamber M, Collins DL, MacDonald D (1994) AnMRI-based probablistic atlas of neuroanatomy. In: Shorvon S,Fish D, Andermann F, Bydder GM, Stefan H (eds) Magneticresonance scanning and epilepsy, Plenum, NY, pp 263–274
Fortier CB, Disterhoft JF, McGlinchey-Berroth R (2000) Cere-bellar cortical degeneration disrupts discrimination learning butnot delay or trace classical eyeblink conditioning. Neuropsy-chology 14:537–550
Gerwig M, Dimitrova A, Kolb FP, Maschke M, Brol B, Kunnel A,Boring D, Thilmann AF, Forsting M, Diener HC, Timmann D(2003) Comparison of eyeblink conditioning in patients withsuperior and posterior inferior cerebellar lesions. Brain 126:71–94
Gerwig M, Dimitrova A, Maschke M, Kolb FP, Forsting M,Timmann D (2004) Amplitude changes of unconditioned eye-blink responses in patients with cerebellar lesions. Exp BrainRes 155:341–351
Green JT, Woodruff-Pak DS (2000) Eyeblink classical condition-ing: hippocampal formation is for neutral stimulus associationsas cerebellum is for association-response. Psychol Bull 126:138–158
Gruart A, Guillazo-Blanch G, Fernandez-Mas R, Jimenez-Diaz L,Delgado-Garcia JM (2000) Cerebellar posterior interpositusnucleus as an enhancer of classically conditioned eyelid re-sponses in alert cats. J Neurophysiol 84:2680–2690
James GO, Hardiman MJ, Yeo CH (1987) Hippocampal lesionsand trace conditioning in the rabbit. Behav Brain Res 23:109–116
Kim JJ, Clark RE, Thompson RF (1995) Hippocampectomy im-pairs the memory of recently, but not remotely, acquired traceeyeblink conditioned responses. Behav Neurosci 109:195–203
Kirsch P, Achenbach C, Kirsch M, Heinzmann M, Schienle A,Vaitl D (2003) Cerebellar and hippocampal activation duringeyeblink conditioning depends on the experimental paradigm: aMEG study. Neural Plasticity 10:291–301
Kishimoto Y, Kawahara S, Suzuki M, Mori H, Mishina M, KirinoY (2001) Classical eyeblink conditioning in glutamate receptorsubunit delta 2 mutant mice is impaired in the delay paradigmbut not in the trace paradigm. Eur J Neurosci 13:1249–1253
Kotani S, Kawahara S, Kirino Y (2003) Trace eyeblink condi-tioning in decerebrate guinea pigs. Eur J Neurosci 17:1445–1454
Kronforst-Collins MA, Disterhoft JF (1998) Lesions of the caudalarea of rabbit medial prefrontal cortex impair trace eyeblinkconditioning. Neurobiol Learn Mem 69:147–162
McEchron MD, Disterhoft JF (1997) Sequence of single neuronchanges inCA1hippocampusof rabbits duringacquisitionof traceeyeblink conditioned responses. J Neurophysiol 78:1030–1044
McGlinchey-Berroth R, Carrillo MC, Gabrieli JD, Brawn CM,Disterhoft JF (1997) Impaired trace eyeblink conditioning inbilateral, medial-temporal lobe amnesia. Behav Neurosci111:873–882
Middleton FA, Strick PL (1994) Anatomical evidence for cerebellarand basal ganglia involvement in higher cognitive function.Science 266:458–461
Moyer JR Jr, Deyo RA, Disterhoft JF (1990) Hippocampectomydisrupts trace eye-blink conditioning in rabbits. Behav Neurosci104:243–252
Ongerboer de Visser BW (1983a) Anatomical and functionalorganization of reflexes involving the trigeminal system in man:jaw reflex, blink reflex, corneal reflex, and exteroceptive sup-pression. In: Desmedt JE (ed) Advances in Neurology, RavenPress, NY, pp 727–738
Ongerboer de Visser BW (1983b) Comparative study of cornealand blink reflex latencies in patients with segmental or withcerbral lesions. In: Desmedt JE (ed) Advances in Neurology,Raven Press, NY, pp 757–772
Peshori KR, Schicatano EJ, Gopalaswamy R, Sahay E, Evinger C(2001) Aging of the trigeminal blink system. Exp Brain Res136:351–363
Port RL, Romano AG, Steinmetz JE, Mikhail AA, Patterson MM(1986) Retention and acquisition of classical trace conditionedresponses by rabbits with hippocampal lesions. Behav Neurosci100:745–752
Ramnani N, Toni I, Josephs O, Ashburner J, Passingham RE(2000) Learning- and expectation-related changes in the humanbrain during motor learning. J Neurophysiol 84:3026–3035
Rorden C, Brett M (2000) Stereotaxic display of brain lesions.Behav Neurol 12:191–200
Schmahmann JD, Dojon J, Toga AW, Petrides M, Evans AC(2000) MRI atlas of the human cerebellum. Academic Press,San Diego, CA
Sears LL, Steinmetz JE (1990) Acquisition of classically condi-tioned-related activity in the hippocampus is affected by lesionsof the cerebellar interpositus nucleus. Behav Neurosci 104:681–692
Sears LL, Logue SF, Steinmetz JE (1996) Involvement of theventrolateral thalamic nucleus in rabbit classical eyeblink con-ditioning. Behav Brain Res 74:105–117
Solomon PR, Van der Schaaf ER, Thompson RF, Weisz DJ (1986)Hippocampus and trace conditioning of the rabbit’s classicallyconditioned nictitating membrane response. Behav Neurosci100:729–744
Steinmetz JE (2000) Brain substrates of classical eyeblink condi-tioning: a highly localized but also distributes system. BehavBrain Res 110:13–24
Takehara K, Kawahara S, Takatsuki K, Kirino Y (2002) Time-limited role of the hippocampus in the memory for trace eye-blink conditioning in mice. Brain Res 951:183–190
Takehara K, Kawahara S, Kirino Y (2003) Time-dependent reor-ganization of the brain components underlying memory reten-tion in trace eyeblink conditioning. J Neurosci 23:9897–9905
Thompson RF, Bao S, Chen L, Cipriano BD, Grethe JS, Kim JJ,Thompson JK, Tracy JA, Weninger MS, Krupa DJ (1997)Associative learning. Int Rev Neurobiol 41:151–189
Thompson RF, Swain R, Clark R, Shinkman P (2000) Intracere-bellar conditioning-Brogden and Gantt revisited. Behav BrainRes 110:3–11
Timmann D, Gerwig M, Frings M, Maschke M, Kolb FP (2004)Eyeblink conditioning in patients with hereditary ataxia: a1 year follow-up study. Exp Brain Res 162:332–345
Topka H, Valls-Sole J, Massaquoi SG, Hallett M (1993) Deficit inclassical conditioning in patients with cerebellar degeneration.Brain 116:961–969
Trouillas P, Takayanagi T, Hallett M, Currier RD, SubramonySH, Wessel K, Bryer A, Diener HC, Massaquoi S, GomezCM, Coutinho P, Ben Hamida M, Campanella G, Filla A,Schut L, Timmann D, Honnorat J, Nighoghossian N, Man-yam B (1997) International cooperative ataxia rating scale for
20

pharmacological assessment of the cerebellar syndrome. TheAtaxia Neuropharmacology Committee of the World Feder-ation of Neurology. J Neurol Sci 145:205–211
Tseng W, Guan R, Disterhoft JF, Weiss C (2004) Trace eyeblinkconditioning is hippocampally dependent in mice. Hippocam-pus 14:58–65
Weible AP, McEchron MD, Disterhoft JF (2000) Corticalinvolvement in acquisition and extinction of trace eyeblinkconditioning. Behav Neurosci 114:1058–1067
Weiss C, Bouwmeester H, Power JM, Disterhoft JF (1999) Hip-pocampal lesions prevent trace eyeblink conditioning in thefreely moving rat. Behav Brain Res 99:123–132
Woodruff-Pak DS, Lavond DG, Thompson RF (1985) Traceconditioning: abolished by cerebellar nuclear lesions but notlateral cerebellar cortex aspirations. Brain Res 348:249–260
Woodruff-Pak DS (1993) Eyeblink classical conditioning in H. M.:delay and trace paradigms. Behav Neurosci 107:911–925
Woodruff-Pak DS, Papka M, Ivry RB (1996) Cerebellar involve-ment in eyeblink classical conditioning in humans. Neuropsy-chology 10:443–458
Woodruff-Pak DS, Levin SI, Chae M, Kim SE, Meisler MH (2004)Delay classical eyeblink conditioning and morris water mazeare impaired in mice with conditional inactivation of SCN8A(Nav1.6) in cerebellar Purkinje cells. Program No. 325.16,Abstract Viewer/Itinerary Planner. Society for Neuroscience,Online, Washington, DC
Yamamoto T, Yoshida K, Yoshikawa H, Kishimoto Y, Oka H(1992) The medial dorsal nucleus is one of the thalamic relays ofthe cerebellocerebral responses to the frontal association cortexin the monkey: horseradish peroxidase and fluorescent dyedouble staining study. Brain Res 579:315–320
Yeo CH, Hesslow G (1998) Cerebellum and conditioned reflexes.Trends Cogn Sci 2:322–330
21
Related Documents











